
Nocturnal exploratory flights, departure time, and direction in a migratory songbird

Diurnal and nocturnal ranging behaviour of Stone-curlews Burhinus oedicnemus nesting in river habitat
CHIARA CACCAMO, ENRICA POLLONARA, NATALE EMILIO BALDACCINI & DIMITRI GIUNCHI*
Department of Biology, Ethology Unit, University of Pisa, Via Volta 6, I-56126 Pisa, Italy
Studying a species under a range of conditions is essential for fully understanding its ecol-ogy and for predicting its response to human impacts on the environment. We investi-gated the spatial behaviour and the habitat characteristics of foraging areas of EurasianStone-curlews breeding in an important but poorly investigated habitat, gravel riverbed,throughout the full 24-h cycle. The data collected for 17 radiotagged birds nesting in theTaro River Regional Park (Parma, Italy) showed a clear split between diurnal and noctur-nal spatial behaviour. Almost all diurnal fixes and about two-thirds of nocturnal oneswere located in the gravel riverbed, which not only provided suitable breeding territoriesbut probably part of the food resources needed for reproduction. Nocturnal excursionsfrom breeding sites to feeding areas (mostly farmland), sometimes of a few kilometres,indicated that these resources do not cover all of the birds’ needs. Night spotlight countsof foraging birds in the agricultural area indicated that Stone-curlews preferred recentlyharvested crops (mainly forage and wheat) and piles of farmyard manure. These habitatpreferences are closely linked to the predominant agricultural activity of the study area,which is characterized by a high density of dairy farms for the production of ParmigianoReggiano cheese. It seems likely that the proximity of gravel riverbed and farmland habi-tats is one of the main causes of the high breeding density recorded in the study area.We propose that the conservation of Stone-curlews at this site could potentially beachieved only by a synergistic management of both natural and agricultural habitats.
Keywords: agriculture, Italy, manure, nocturnal foraging.
Studying an avian species in different contexts isessential for understanding its ecology and for pre-dicting its response to various human impacts onthe environment. Bird resource requirements mayvary both temporally and spatially (e.g. Cody1985, Millspaugh & Marzluff 2001, Gillings et al.2005 and references therein). This variabilityshould be taken into account fully, as reliable infor-mation on species’ needs are crucial for avian con-servation (Marzluff & Sallabanks 1998).
The Eurasian Stone-curlew Burhinus oedicnemus(Charadriiformes, Burhinidae) is a steppe speciesof conservation concern in Europe (Tucker & Evans1997, Burfield 2005) due to its population decline(> 30%), which began in the second half of the19th century and accelerated after the Second
World War, when the species disappeared from theNetherlands and western Germany (BirdLife Inter-national 2004). Between 1970 and 1990, decreasesbecame evident throughout almost the entireEuropean range as suitable breeding habitats werelost due to agricultural intensification (Delanyet al. 2009, Wilson et al. 2009). In Italy the specieshas an unfavourable conservation status (Calvarioet al. 1999), even though for some populations apositive trend has recently been recorded (Tinarelliet al. 2009).
In spite of this conservation status, detailedinformation on the ecology and behaviour of thisspecies is currently quite scarce, mainly due toits secretive and crepuscular habits (Cramp &Simmons 1983, Vaughan & Vaughan Jennings2005). Most published papers concern Englishpopulations, which are on the northern edge of thespecies European range and chiefly use heathlands
*Corresponding author.Email: [email protected]
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Ibis (2011), 153, 707–720

and stony pastures for breeding and foraging(Green & Griffiths 1994, Green & Taylor 1995,Green et al. 2000). These habitats, however, repre-sent only a relatively small proportion of the habi-tats used by Stone-curlew throughout Europe,which include dry grasslands, steppes, sand dunes,gravel riverbeds and semi-deserts, as well as agri-cultural habitats (Cramp & Simmons 1983,Vaughan & Vaughan Jennings 2005, Triolo et al.2011). In particular, studies of populations nestingin bars or banks along rivers where the flow is sea-sonally reduced are completely lacking, eventhough this habitat holds important numbers ofbreeding pairs in Italy (Brichetti & Fracasso 2004,Tinarelli et al. 2009).
The aims of this study were: (1) to investigatethe spatial behaviour of Stone-curlews breeding ina gravel riverbed by means of radiotracking; and(2) to analyse the habitat preferences of birds for-aging in farmland areas located along the riversideby means of nocturnal spotlight counts.
METHODS
Study area
Our study was carried out during 1999–2001 and2007–2008 in the Taro River Regional Park, Parma,Italy (44.74�N, 10.17�E; total area = 25 km2;Fig. 1), which hosts one of the largest populations
in continental Italy (Giunchi et al. 2009), mainlybreeding in the dry gravel riverbed. In this area,Stone-curlew abundance has increased by around30% in the last 10 years and the density of breed-ing pairs is among the highest ever reported forthe species in Europe (Giunchi et al. 2009).Although some changes have occurred (e.g. con-struction of buildings, factories and road infrastruc-tures), the landscape outside the riverbed did notsubstantially change between the two periods ofthe analysis and was dominated by agriculturalland and to a lesser extent by riparian woods, quar-ries, food processing factories, dairy farms andtowns.
Radiotracking
During 1999–2001 a total of 17 incubating Stone-curlews (four in 1999, seven in 2000 and six in2001) were captured on nests using a fall trap. Ourdata indicated that nest disturbance caused by thistrapping method was tolerated by the birds (Giun-chi & Pollonara 2007). Once caught, birds werefitted with radio-transmitters (TW-4 ⁄ Ag357;Biotrack, Wareham, Dorset, UK) weighing 4.5 gand configured to give 4.9 months of expected life.Each tag was glued using cyanoacrylate to thecalamus of the second rectrix and then tied withnylon threads to the calami of both the second andthird rectrices in order to spread the tag’s weight
Figure 1. Map of the study area.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
708 C. Caccamo et al.

over two feathers; the antenna was then tied andglued along the shaft of the second rectrix (moredetails in Kenward 2001). This method of attach-ment did not allow for long tracking times (seeResults) because the radio-transmitters were lostwhen the birds moulted the tail-feathers (tail-moult period June–September, Vaughan &Vaughan Jennings 2005), but minimized distur-bance, as tags fell off before the start of autumnmigration. As transmitters were about 1–1.3% of abird’s body weight, it seems unlikely that taggingsignificantly affected their behaviour (Whittingham1996). Birds were tracked using AVM LA-12receivers (AVM Instrument Company, Champaign,IL, USA) and hand-held four-element Yagi anten-nas (Televilt, Stockholm, Sweden). Fixes were col-lected both during the day (D-fixes: between30 min before sunrise and 30 min after sunset)and night (N-fixes) under a rotational scheme,with a minimum period of 12 h between succes-sive relocations on any one individual to reduceautocorrelation (van Winkle 1975). Two observerstook two bearings in quick succession (< 5 min)from two stations a few hundred metres apart.More bearings (usually one or two) were thentaken by moving to different stations, further awayfrom the lines of the first two and often closer tothe bird (maximum tracking distance, c. 200 m).Given that Stone-curlews are mostly active duringthe night (Cramp & Simmons 1983, Green et al.2000, Vaughan & Vaughan Jennings 2005), relocat-ing all tagged birds required about 1–2 h duringthe day and 4–6 h at night.
Bird activity was estimated as the number ofchanges in the signal strength, caused by alterationin the position of the tag relative to the receiver,per minute (Nch). Birds were classified as ‘inac-tive’ when at least in two bearings Nch was £ 1and ‘active’ otherwise, i.e. only birds showingprolonged bouts of activity, which spanned overmost of the tracking period, were consideredactive. Tag position was determined by triangula-tion (Heezen & Tester 1967, Kenward 2001). Fixeswere calculated using the Lenth method (White &Garrott 1990), which derives a maximum likeli-hood estimation of an animal’s location by assum-ing that bearings are distributed according to acircular normal distribution (von Mises distribu-tion, Mardia 1972). This technique also gives thetelemetry error by using the placement of thetelemetry stations, the number of bearings, and thebearing error, i.e. the deviation of the bearing from
the estimated location, which can be estimated onseries of bearings taken from the same stations andthus having a similar bearing angle. As the estima-tor is assumed to be normally distributed in twodimensions, rather than getting 95% confidencelimits, we get 95% confidence areas (ellipses). Inour case, 95% confidence ellipses were estimatedfor a sample of 100 D- and 100 N-fixes for whichwe have multiple bearings (two to four) taken bydifferent observers from each station. The obtainedarea (mean ± se) was 3750 ± 203 m2 during theday and 11 293 ± 605 m2 at night. The error ofN-fixes was larger because, during the night: (1) itwas only possible to obtain data from stationslocated on marginal roads as access to farmlandswas restricted; and (2) birds were often mobile,which introduced more error into the locationestimates (White & Garrott 1990). The size ofthese error ellipses was considered in the choice ofthe precision rate of the habitat selection analysis(see below).
Data analysis
Given the strong and expected differences in birds’spatial behaviour during the day and at night (seeResults), D- and N-fixes were analysed separately.We calculated diurnal and nocturnal home-rangesusing adaptive 95% local convex hulls (a-LoCoH;Getz et al. 2007). To calculate hulls, the a-LoCoHmethod was used which chooses the neighbouringpoints within an adaptive sphere of influence,rather than considering a fixed number of neigh-bours, as in the original formulation of the LoCoHmethod (k-LoCoH; Getz et al. 2007). Accordingto Getz et al. (2007), a-LoCoH performs betterthan k-LoCoH and simulations suggest that thismethod is robust to deviations from the optimalvalue of a. As a rule of thumb, Getz et al. (2007)suggested that the maximum distance between anytwo points in the dataset (MaxD) provides a goodapproximation of a. We selected different a-valuesfor each individual ⁄ time of the day combinationby means of graphical checks of a vs. area plotsconstructed considering different values of a cen-tred on MaxD. As LoCoH tended to producesmaller estimates than other home-range estima-tors (Huck et al. 2008), we also provided home-range size estimated with 95% minimum convexpolygons (MCP, Mohr 1947). An incremental areaanalysis method calculated using birds ⁄ period(day–night) combinations with more than 40 fixes
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 709

indicated that in three (diurnal and nocturnalLoCoH and diurnal MCP) of four cases between40 and 50 locations were required to describehome-ranges adequately (Fig. 2). To avoid a drasticreduction in sample size, we considered only birdswith more than 40 fixes when trying to give a reli-able estimate of the home-range size. For thisreason, nocturnal MCP might be underestimated.Data regarding the breeding stage of all markedbirds were not available and were thus not consid-ered in the analysis.
By means of photo-interpretation of 1 : 10 000aerial photos collected in winter 1999, integratedand updated with terrestrial surveys with GPS(GPS 12 CX; Garmin, Olathe, KS, USA), we cre-ated a raster map of the study area (cellsize = 60 · 60 m) using Quantum GIS 1.5.0,including six habitat macrocategories: river, farm-land, dry grassland, wood (chiefly Populus, Robiniaand Salix), open water (artificial ponds) and urban-ized areas (towns and stone quarries). Habitatselection was analysed using only N-fixes, as morethan 95% of D-fixes were collected from the samehabitat, and only for birds with more than 40 relo-cations. We calculated resource selection ratios(RSR) as the ratios between the proportions ofused and available resource units that are in a givencategory (Manly et al. 2002). According to thismethod, RSR = 1 indicates no selection,1 < RSR < ¥ indicates that the category is posi-tively selected, and 0 < RSR < 1 indicates that thecategory is negatively selected, i.e. used less thanavailable. Selection ratios were calculated at thelevel of: (1) the home-range within the study area(second-order habitat selection according to John-son 1980, or design II according to Thomas & Tay-lor 1990) and (2) the selection of sites within thehome-range (third-order habitat selection accord-ing to Johnson 1980 or design III according to Tho-mas & Taylor 1990). In the former case, habitatavailability for all birds was defined as the bound-aries of the 100% MCP calculated using all fixes ofall radio-tracked birds; in the latter case, habitatavailability for each bird was determined within theboundary of 100% MCP calculated on N-fixes.Three Chi-squared tests were used to analyse habi-tat selection (see Manly et al. 2002 for furtherdetails): (1) an overall test of the null hypothesis ofselection in proportion to availability (design II andIII); (2) a test of the null hypothesis that animalswere using resource categories in the same propor-tions (design II); and (3) a test of selectivity at anindividual level (design III).
Home-range estimation and resource selectionanalysis were performed by means of the ADEHABI-
TAT v. 1.7 package (Calenge 2006) of the statisticalsoftware R v. 2.11.1 (R Development Core Team2010). Proportional data (i.e. proportion of activefixes during the day or night) were analysed usinggeneralized linear mixed models (GLMM) witha binomial error distribution and bird identity as arandom intercept (Crawley 2007), fitted usingRLME4 package (Bates & Maechler 2010). Signifi-
Day (n = 9), y = 100(1 − e−0.13x)Night (n = 11), y = 100(1 − e−0.12x)
No. of fixes0 10 20 30 40 50 60 70
020
4060
8010
00
2040
6080
100
Day (n = 9), y = 100(1 − e−0.07x)Night (n = 11), y = 100(1 − e−0.09x)
Are
a (%
of t
he a
sym
ptot
e)
Figure 2. Incremental area analysis. Home-range areas
(mean ± se) were estimated as 95% LoCoH (above) or 95%
MCP (below) and reported as percentage of the asymptotic
area calculated according to the formula y = 100 (1 ) e)bx).
Results are shown for bird ⁄ period (day–night) combinations for
which there were more than 40 fixes.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
710 C. Caccamo et al.

cance was assessed using likelihood-ratio tests(LRT).
Night spotlight counts
Given both the low precision of fixes and the highlevel of habitat fragmentation in the study area, itwas not possible to analyse in detail the birds’ hab-itat choice with radiotracking data. To gain someinsight into this, especially regarding the relation-ship between foraging behaviour and farmlandhabitat, we analysed habitat preferences in theagricultural areas at the riverside by means of noc-turnal counts of foraging birds combined with veg-etation measurements. Data were collected duringtwo years (2007 and 2008) in seven foraging areaslocated near the riverbed (Fig. 1), which were cho-sen by taking into consideration previous dataobtained through evening observations, crepuscularpoint counts of calling birds and radiotracking.Distances from the riverbanks of these areas(mean = 355 m, median = 320 m, range = 40–1030 m) were chosen to be well within the rangesof commuting flights documented for radiotrackedStone-curlews. Each area consisted of a variablenumber of fields (range = 3–10) of different sizes(mean = 2.5 ha, median = 2.1 ha, range = 0.1–12.1 ha), delimited by roads or ditches. Birdcounts were undertaken by two observers duringthe night (between 0:40 h and 5:30 h after localsunset) every 10 days from May to July. We madenine counts per field in 2007 and eight in 2008.Visiting all areas required three consecutive nightsfor each replicate. The order of visits was random-ized. Each night, all fields in the same area werevisited twice. The second visit was performed afterthe first one was completed, leaving at least 1 hbetween the two counts. To reduce pseudoreplica-tion, the maximum number of birds detected inthe two visits was considered for the analysis. Aswalking inside the cultivated fields was notallowed, Stone-curlews were detected using lamps(Lightforce ML170 Striker, diameter 170 mm,30 W) by walking the same transects located on aborder of a given field. Brief illumination usuallydid not immediately scare birds, which often con-tinued their feeding activity until the observer wasquite near. The lack of a tapetum lucidum inthe retina (Martin & Katzir 1994) made the detec-tion of Stone-curlews difficult, especially whenvegetation was high, so birds were also flushed bymaking loud noises and walking a few steps into
each field. With this approach, we are confidentthat all birds that were present in a given fieldwere flushed during counts, especially as they usu-ally produced alarm calls while leaving the area,which increased detectability. Furthermore, birdscould be followed in flight until they landed or leftthe area, thus making double counts during thesame visit unlikely. As evidenced by preliminarysurveys, this census technique was effective onlywell within the range of the spotlight. Preliminarysurveys allowed us to estimate that beyond 150 mfrom each transect the likelihood of missing somebirds was noticeable. For this reason, we consid-ered only detections within this range. The area ofeach field considered in the analysis was reducedaccordingly. Censuses were not undertaken duringperiods of high wind (above 25 km ⁄ h) or heavyprecipitation.
Fields were classified into four categories (fieldtypes): cereals (wheat and barley), forage (alfalfa,permanent pastures, perennial rye-grass and oat),other crops (tomato and fallow land) and piles offarmyard manure (see below). As we were notallowed to walk inside all the cultivated fields, wederived an index of vegetation density by verticallyembedding into the vegetation a 100 · 10 cm che-quered wooden board (cell = 1 · 1 cm), 3 m fromthe transect used during nocturnal counts. Vegeta-tion density was estimated as the maximum heightof 90% obscuration of bands by vegetation, read at3 m distance by the same observer (Sutherland1996, Bibby et al. 2000). Only four replicates perfield were taken at random points, as the consis-tency of measurements within fields was high rela-tive to the extent of differences found among fields(coefficient of intra-class correlation = 0.95; Sokal& Rohlf 1995, van Belle 2008, Wolak et al. 2011).Vegetation density for piles of farmyard manurewas set to 0 cm. Habitat variables were recordedduring the daylight with the same schedule as noc-turnal counts.
Data analysis
Counts were dominated by zeros and single birds,and hence counts were transformed to pres-ence ⁄ absence data, which were analysed usingGLMM with a binomial error distribution. Thevariables considered in the full model were: (1)dependent variable: presence ⁄ absence of birds(1 = at least one bird detected in at least one ofthe two visits to each field in each replicate;
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 711

0 = no birds detected in any of the two visits;logit-transformed); (2) fixed factors: field area inhectares (log-transformed), field distance from theriver in metres (log-transformed), census year (2-level factor: 2007, 2008), month (3-level factor:May, June, July), field type (4-level factor: forage,cereals, other crops, manure), average index of veg-etation density in centimetres; and (3) random fac-tors: field nested within area. After checkingmodel assumptions, the fixed part of the modelwas simplified by means of the Akaike informationcriterion corrected for small sample size (AICc,Burnham & Anderson 2002), considering all themodels between the full model and the modelwhich included only field area, field distance fromthe river and year. Significance of factors retainedin the most parsimonious model was assessed usingLRT. We did not try to simplify the random part ofthe model in order to conserve the hierarchicalstructure of the design and thus we maintained atight linkage between the sampling strategy andthe statistical analysis (van Belle 2008). To betterassess graphically the effect of the two habitat vari-ables (field type and index of vegetation density),we simulated the expected probability of detectingat least one foraging bird ⁄ field under the modelselected using AICc. This was obtained by meansof the logit.mixed model in ZELIG R-package (Bailey
& Alimadhi 2007), using simulations via asymp-totic approximation.
RESULTS
Radiotracking
We recorded between 27 and 142 total fixes fromeach of 17 radiotagged Stone-curlews duringApril–August (Table 1). As expected, the percent-age of active fixes was significantly higher duringthe night (66.7 ± 3.7% se) than during the day(23.5 ± 3.4%; b = 2.09 ± 0.12, v2
1 = 327.41, P <0.001). Birds became active mostly at dusk, andgreatly reduced their activity after dawn. Earlymorning hours still showed some spikes of activity,whereas Stone-curlews were almost completelyinactive during the afternoon (Fig. 3a). This pat-tern of activity closely matched the ranging behav-iour recorded in the two periods. During the day,bird movements were quite short (mean dis-tance = 131 ± 4 m) and c. 80% of D-fixes werewithin 200 m of the capture site. During the night,fewer than 60% of N-fixes were within 200 m ofthe capture site (mean = 418 ± 20 m) and we reg-istered excursions of greater than 1 km (maximumrecorded distance = 3.4 km). This pattern wasrelated to a marked change in habitat use. Almost
Table 1. Summary of data collected from 17 radiotagged birds in 1999–2001.
Individual Year Period Days Total fixes D-fixes N-fixes
1999.747 1999 5 May to 2 July 58 46 26 20
1999.777 11 May to 18 July 68 62 25 37
1999.807 13 May to 30 August 109 61 25 36
1999.837 31 May to 28 June 28 27 11 16
2000.788 2000 1 May to 31 July 91 115 45 70
2000.819 14 May to 10 July 57 70 26 44
2000.853 30 April to 18 July 79 93 39 54
2000.893 27 April to 17 August 112 138 53 85
2000.933 26 April to 17 August 113 135 55 80
2000.951 11 May to 20 June 40 45 19 26
2000.973 21 April to 25 July 95 123 49 74
2001.789 2001 30 April to 27 August 121 142 64 78
2001.819 6 June to 27 August 82 86 42 44
2001.854 8 May to 11 August 95 118 56 62
2001.893 14 May to 9 August 66 88 41 47
2001.933 23 May to 16 August 85 98 43 55
2001.973 2 May to 28 May 26 36 14 22
Min. 26 27 11 16
Median 82 88 41 47
Max. 121 142 64 85
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
712 C. Caccamo et al.
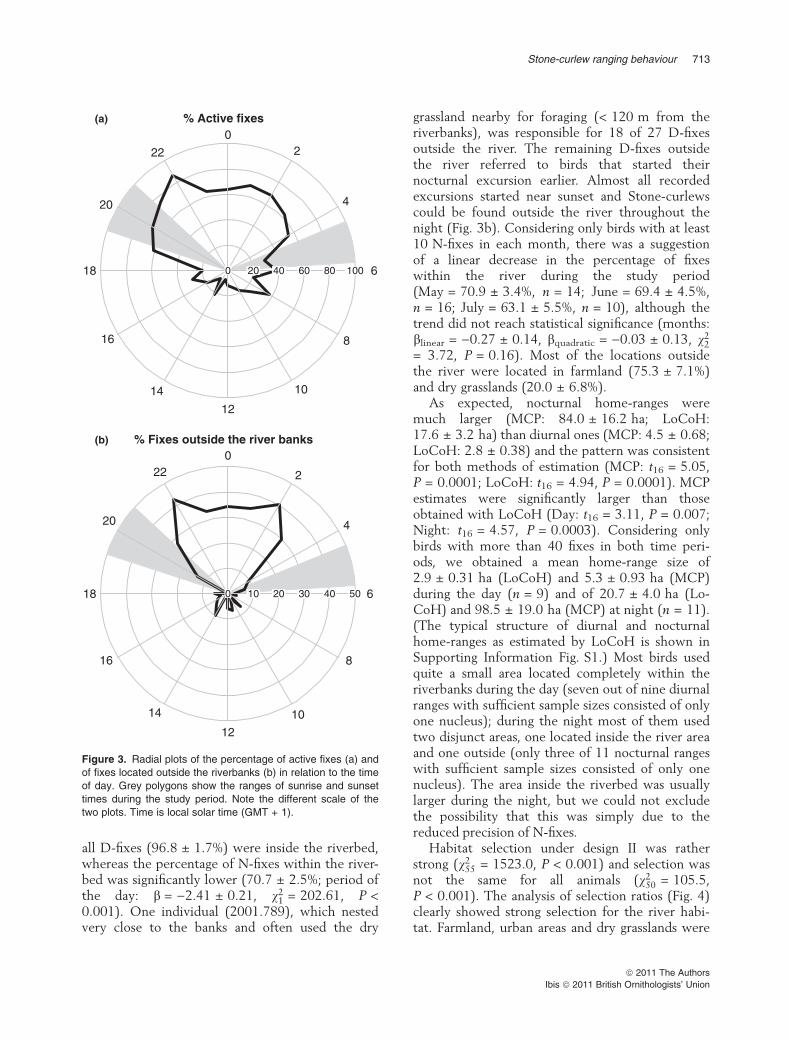
all D-fixes (96.8 ± 1.7%) were inside the riverbed,whereas the percentage of N-fixes within the river-bed was significantly lower (70.7 ± 2.5%; period ofthe day: b = )2.41 ± 0.21, v2
1 = 202.61, P <0.001). One individual (2001.789), which nestedvery close to the banks and often used the dry
grassland nearby for foraging (< 120 m from theriverbanks), was responsible for 18 of 27 D-fixesoutside the river. The remaining D-fixes outsidethe river referred to birds that started theirnocturnal excursion earlier. Almost all recordedexcursions started near sunset and Stone-curlewscould be found outside the river throughout thenight (Fig. 3b). Considering only birds with at least10 N-fixes in each month, there was a suggestionof a linear decrease in the percentage of fixeswithin the river during the study period(May = 70.9 ± 3.4%, n = 14; June = 69.4 ± 4.5%,n = 16; July = 63.1 ± 5.5%, n = 10), although thetrend did not reach statistical significance (months:blinear = )0.27 ± 0.14, bquadratic = )0.03 ± 0.13, v2
2
= 3.72, P = 0.16). Most of the locations outsidethe river were located in farmland (75.3 ± 7.1%)and dry grasslands (20.0 ± 6.8%).
As expected, nocturnal home-ranges weremuch larger (MCP: 84.0 ± 16.2 ha; LoCoH:17.6 ± 3.2 ha) than diurnal ones (MCP: 4.5 ± 0.68;LoCoH: 2.8 ± 0.38) and the pattern was consistentfor both methods of estimation (MCP: t16 = 5.05,P = 0.0001; LoCoH: t16 = 4.94, P = 0.0001). MCPestimates were significantly larger than thoseobtained with LoCoH (Day: t16 = 3.11, P = 0.007;Night: t16 = 4.57, P = 0.0003). Considering onlybirds with more than 40 fixes in both time peri-ods, we obtained a mean home-range size of2.9 ± 0.31 ha (LoCoH) and 5.3 ± 0.93 ha (MCP)during the day (n = 9) and of 20.7 ± 4.0 ha (Lo-CoH) and 98.5 ± 19.0 ha (MCP) at night (n = 11).(The typical structure of diurnal and nocturnalhome-ranges as estimated by LoCoH is shown inSupporting Information Fig. S1.) Most birds usedquite a small area located completely within theriverbanks during the day (seven out of nine diurnalranges with sufficient sample sizes consisted of onlyone nucleus); during the night most of them usedtwo disjunct areas, one located inside the river areaand one outside (only three of 11 nocturnal rangeswith sufficient sample sizes consisted of only onenucleus). The area inside the riverbed was usuallylarger during the night, but we could not excludethe possibility that this was simply due to thereduced precision of N-fixes.
Habitat selection under design II was ratherstrong (v2
55 = 1523.0, P < 0.001) and selection wasnot the same for all animals (v2
50 = 105.5,P < 0.001). The analysis of selection ratios (Fig. 4)clearly showed strong selection for the river habi-tat. Farmland, urban areas and dry grasslands were
% Active fixes0
2
4
6
8
10
12
14
16
18
20
22
0 20 40 60 80 100
(a)
% Fixes outside the river banks0
2
4
6
8
10
12
14
16
18
20
22
0 10 20 30 40 50
(b)
Figure 3. Radial plots of the percentage of active fixes (a) and
of fixes located outside the riverbanks (b) in relation to the time
of day. Grey polygons show the ranges of sunrise and sunset
times during the study period. Note the different scale of the
two plots. Time is local solar time (GMT + 1).
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 713

on average comparably and negatively selected,even although it should be noted that the selectionratios for the last two categories were highly vari-able between animals. The other habitat categorieswere also avoided. The results were almost thesame as regards design III data: habitat selectionwas strong both overall (v2
19 = 654.5, P < 0.001)and at an individual level (v2
1�3 = 25.9–90.6,P < 0.001). The profile of preferences was thesame as design II and thus is not reported here.
Night spotlight counts
Most (70%) non-zero counts referred to singlebirds. Multiple detections (range 2–6) were rathermore frequent on piles of farmyard manure (58%,n = 24) with respect to the other habitat types(forage 13%, n = 39; cereals 25%, n = 4; othercrops 33%, n = 6). Most birds were detected onpiles of manure and on crops characterized by lowvegetation density (Fig. 5a). According to AICc,the best supported model among the consideredset included, apart from field area, field distancefrom the riverbanks and year, index of vegetationdensity and field type (Table 2). Considering
0
2
4
6
8
Glo
bal s
elec
tion
ratio
s (9
5% C
I)
Riv
er
Far
mla
nd
Urb
. are
a
Dry
gra
ss.
Woo
d
Ope
n w
ater
Figure 4. Selection ratios (± 95% CI) according to design II
(Manly et al. 2002) calculated on N-fixes of birds with more
than 40 relocations (n = 11). The horizontal line indicates the
threshold = 1 for positive selection.
0−4 4−10 10−30 > 30
ManureForageCerealsOther crops
Index of vegetation density (cm)
No.
of b
irds
(a)
Exp
ecte
d va
lues
(95
% C
I)
Forage Cereals Other crops Manure
(b)
0 20 40 60 80
0
10
20
30
40
50
0.0
0.2
0.4
0.6
0.8
1.0
0.00
0.05
0.10
0.15
0.20
Index of vegetation density (cm)
Exp
ecte
d va
lues
(95
% C
I)
(c)
Figure 5. (a) Pooled number of birds counted in 2007 and
2008 in relation to the different habitats considered in the analy-
sis, and (b,c) simulated expected values of the probability of
detecting at least one forging bird per field calculated under the
best model selected using AICc. (b) Index of vegetation den-
sity = 0; (c) field type = forage. (b,c) Field area and distance
from the riverbanks were set to their average values (respec-
tively 2.5 ha and 355 m) and the year was set to 2007.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
714 C. Caccamo et al.

Akaike weights, this model had a relatively strongsupport. Indeed, the evidence ratio for this modelvs. the only competitive model among theconsidered set (the second best model) was 4.2.The index of vegetation density and the field typewere included in both models, thus underlying theimportance of both variables. The estimatedparameter in the selected model and the signifi-cance of fixed factors as results of LRT associatedwith variable removal from the model are given inTable 3. All considered variables were significant,except year and field distance from the riverbanks.The effect of the two habitat variables is summa-rized in Figure 5(b,c). It is clear that, when control-ling for vegetation density, piles of farmyardmanure were by far the most preferred sites, andthe other considered categories were much less
used. On the other hand, birds tended to avoidfields with a high level of vegetation density, eventhough their choice seemed relatively flexible.
DISCUSSION
To the best of our knowledge, this paper is onlythe second study of Stone-curlew spatial behav-iour, and the first one of birds nesting in a riverhabitat. The results clearly show the importantrole of this habitat for the population. Almost allof the diurnal fixes and about two-thirds of noctur-nal ones were located within the riverbanks. Thisresult is at least partially expected given that theriver hosts the nest-sites. However, a significantproportion of active fixes (c. 40%) were collectedwithin riverbanks, and, although some fixes couldbe due to other activities, it seems likely that manyconcerned foraging birds, as also confirmed by ournon-systematic diurnal observations of Stone-curl-ews feeding on orthopterans, ants, spiders and evensmall mammals collected from the gravel banks(D. Giunchi, E. Pollonara & C. Caccamo unpubl.data). Indeed, even though most of the foragingactivity was concentrated during the night, asexpected for a chiefly nocturnal species, duringreproduction Stone-curlews also feed during theday, especially in the early morning and in the lateafternoon (Cramp & Simmons 1983, Green et al.2000, E. Pollonara, C. Caccamo & D. Giunchi pers.obs.). These diurnal periods coincide with thepeaks of diurnal activity recorded in this study andalso correspond to the period when almost all birdshad not yet left their breeding sites. This furthersuggests that the riverbed, and specifically a rela-tively small area (usually less than 3 ha), not only
Table 2. Summary of GLMM selection using Akaike’s information criterion for small sample sizes (AICc).
Model k DAICc wi AICc
log (area) + log (distance) + year + vdensity + ftype 10 0.00 0.81
log (area) + log (distance) + year + month + vdensity + ftype 12 2.91 0.19
log (area) + log (distance) + year + ftype 9 10.77 0.00
log (area) + log (distance) + year + month + ftype 11 14.89 0.00
log (area) + log (distance) + year + vdensity 7 30.53 0.00
log (area) + log (distance) + year + month + vdensity 9 32.82 0.00
log (area) + log (distance) + year 6 48.23 0.00
log (area) + log (distance) + year + month 8 52.16 0.00
k, number of parameters; DAICc, difference in AICc between a given model and the model with the lowest AICc; wi AICc, Akaike
weights, which estimate the probability a specified model is the true model, given that one of the models being considered is indeed
the true model; area, field area in hectares; distance, field distance from the riverbanks in metres; vdensity, index of vegetation
density in centimetres (cm); ftype, field type.
Table 3. Estimated parameter (b), with corresponding se and
results of LRT associated with variable removal from the best
model selected using AICc*.
Factor b se v2 df P
(Intercept) )1.72 1.33
Log field area) 0.97 0.30 11.03 1 < 0.001
Log (field distance
from the river)
)0.16 0.24 0.44 1 0.5
Year 3.10 1 0.08
2007 vs. 2008 )0.62 0.37
Index of vegetation
density
)0.05 0.02 12.83 1 < 0.001
Field type 36.71 3 << 0.001
Cereals vs. forage )0.28 0.62
Other crops vs. forage )0.62 0.53
Manure vs. forage 6.10 1.05
*Variance for random factors: area � 0; field nested within
area = 0.45.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 715

provides suitable breeding sites but also part of thefood resources needed for reproduction. However,the movements recorded during the night testifythat these resources do not cover all of the birds’needs. Tagged birds regularly commuted betweenthe riverbed and adjacent habitats, although theextent and frequency of these excursions were var-iable, as indicated by the high variability of noctur-nal home-range sizes. Almost all the fixes collectedoutside the riverbed were classified as active andall birds not immediately flushed by spotlightswere observed feeding, thus confirming that river-side habitats were mostly used for foraging.
In spite of the marked differences in habitat,several aspects of the observed spatial behaviour inand around the riverbed resemble those reportedby Green et al. (2000) for Stone-curlews nesting inheathlands and stony pastures. The asymptoticactive home-range area estimated by Green et al.(2000), using a different method of calculation,was higher (c. 30 ha), but of the same order ofmagnitude as our nocturnal ranges estimated byLoCoH for birds with sufficient sample size(c. 21 ha). Moreover, English birds also used a rela-tively small area around the nest-site and a variablenumber of disjunct areas up to a few kilometresaway, although Green et al. (2000) did not statewhether these foraging areas were chiefly visitedduring the night, as in our case, or throughout theday. The present data confirm the hypothesis byVaughan and Vaughan Jennings (2005) that forag-ing flights of Stone-curlews are probably a wide-spread behavioural trait throughout the range ofthe species during both the breeding and the non-breeding season. This kind of spatial behaviour isnot uncommon among shorebirds: several speciesuse distant foraging areas which may be visitedduring either the day or the night, during bothbreeding (e.g. Whittingham et al. 2000, Hoodless& Hirons 2007, Schekkerman & Beintema 2007)and wintering periods (e.g. Berdeen & Krementz1998, Shepherd & Lank 2004, Duriez et al. 2005a,2005c). However, the almost exclusive nocturnaluse of disjunct foraging areas by our studied Stone-curlews is interesting given that, as stated before,birds regularly forage during the day.
There are probably several reasons for such aclear split between diurnal and nocturnal spatialbehaviour. Gillings and Sutherland (2007) sug-gested that on arable farmlands, shorebird specieswith specialized night vision should have higherintake rates during the night than during the day,
mainly because of the surface activities of largeinvertebrates (especially large anecic earthworms;Paoletti 1999). In addition, as most of the disjunctforaging areas were located in farmlands, some-times close to urban areas, it could be speculatedthat during the day these areas are less profitableowing to human disturbance, which could nega-tively affect birds’ foraging behaviour, as demon-strated for other species of shorebird (Burger &Gochfeld 1991, Goss-Custard & Verboven 1993,Thomas et al. 2003, Yasué 2005). Furthermore,predation by flocks of Hooded Crows Corvus cor-nix on Stone-curlew eggs and chicks recorded inthe study area (Giunchi et al. 2009) forces bothparents actively to defend their clutch, and there-fore one-parent-only parental care during the nightcould be less costly than during the day, as mostnocturnal predators (e.g. foxes) do not search forfood in groups. It should be noted that, as Stone-curlews usually do not leave the eggs or chicks unat-tended (Cramp & Simmons 1983, Vaughan &Vaughan Jennings 2005), this behaviour reducesthe time available to each parent to efficientlyexploit trophic resources, which explains the likelycompensative diurnal foraging activity in the river-bed, which is absent or much reduced outside thebreeding period (E. Pollonara pers. obs.; see alsoVaughan & Vaughan Jennings 2005). The relativelylow basal metabolic rate of Stone-curlews (Duriezet al. 2010) may help them to meet their dailyenergy demands by limiting supplemental diurnalforaging only to the first and last hours of thedaytime. It is interesting to note that a some-what similar trade-off between feeding andpredation has been proposed by Duriez et al.(2005a, 2005b, 2005c) to explain the nocturnal useof disjunct foraging areas by another mainlynocturnal wader, the Woodcock Scolopax rusticola.During the evening, wintering French Woodcocksonly left their relatively safe diurnal roost ⁄ foragingsites located in woods and flew to meadows (whereboth predation risk and food abundance werehigher) if they could not meet their energy require-ments during the day. This pattern of compensatorynocturnal foraging is, however, reversed withrespect to that suggested here for the Stone-curlew(compensatory diurnal foraging), which is probablyrelated both to the different ecology of the two spe-cies, especially regarding habitat selection, and tothe diverse constraints imposed by the markedlydifferent stages of the annual cycle considered bythe two studies (wintering vs. breeding season).
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
716 C. Caccamo et al.

As observed for Woodcocks and other species ofshorebirds (Berdeen & Krementz 1998, Duriezet al. 2005a, Gillings et al. 2005), and as alsoreported by Green et al. (2000) for the Stone-curlew, nocturnal foraging excursions lead to a sig-nificant switch in habitat preference and to anoticeable use of farmland. Indeed, although someindividuals also used dry grasslands and quarries,almost all tagged birds foraged in the agriculturalarea, which is the dominant landscape. Night spot-light counts showed that the preferred foragingsites were piles of farmyard manure. The likeli-hood of finding birds in these spots was relativelyhigh, despite their limited surface area. These pilesare periodically topped-up with fresh manurebefore being spread as an organic fertilizer inAugust–September, at the very end of the Stone-curlew breeding season. The use of heaps of man-ure (chiefly farmyard or poultry manure) beforespreading has also been documented in GreatBritain (Green et al. 2000, Vaughan & VaughanJennings 2005) and the Canary Islands (Martín &Lorenzo 2001, Giannangeli et al. 2004, F. Rodri-guez pers. comm.), and these probably representan important trophic resource for the species infarmland areas. In these habitats, we recorded thehighest frequency of birds foraging in groups(distance between individuals < 10 m), whereas inthe other field types, birds seemed to forage almostalways alone. Provided that the principal ecologicalfactors affecting the tendency for a given species toforage singly or in flocks are food and predators(Alexander 1974, Beauchamp 2005, 2007, Colwell2010), these results seem to suggest a largeravailability of prey in piles of manure. The effect ofpredation on flocking should in fact be marginaland possibly limited to shared vigilance (Roberts1996), given that the different field types probablyshare the same predators (chiefly foxes and feralcats) and that recorded flock sizes were rather small(< six individuals) and thus not very effective indecreasing the individual probability of beingpreyed upon (the dilution effect, Hamilton 1971).Birds using the same heap of manure during a givennight did not necessarily nest in close proximity, asshown by some preliminary colour-ringing data col-lected in 2008 (maximum recorded distancebetween nest-sites of birds trapped on the same pileduring the same night c. 2 km, n = 4; D. Giunchi ,E. Pollonara & C. Caccamo unpubl. data).
Aside from piles of manure, spotlight countsindicated that, as proposed by Green et al. (2000),
the Stone-curlews’ choice of foraging areas ismostly influenced by the quantity and thestructure of vegetation instead of the quality per se(see also Sutherland et al. 2004). Indeed, birds didnot show any significant crop preference that couldnot be attributed to vegetation characteristics. Forthis reason, they used fast-growing forage cropsmore frequently, which were cut at least threetimes throughout the breeding season, rather thanslow-growing cereals (chiefly wheat), which areharvested only once. This preference for short andsparse vegetation, in common with other waderspecies that forage at night (Martin 1990 and refer-ences therein), is probably related to the bird’s for-aging technique. Stone-curlews mainly feedvisually by pecking invertebrates on the soil (Mar-tin & Katzir 1994) so a reduction of vegetationheight and density could increase foraging effi-ciency, especially by means of improved preydetection and forager mobility, as observed forother bird species (Butler & Gillings 2004,Devereux et al. 2004, Duriez et al. 2005a, Wilsonet al. 2005).
The pattern of habitat use in farmlands clearlyshows a close link between Stone-curlew foragingbehaviour and the prevalent farming activity in thestudy area. This area is characterized by a highdensity of quite intensive dairy farms (most ofthem outside the boundary of the natural park)aimed at producing the Parmigiano-Reggianocheese. Meadows and pastures are almost absent,although semi-natural grasslands still survive ina few spots and most of the landscape is domi-nated by species-poor ‘improved’ grasslandsand tillage. However, according to the quiterestrictive regulation of official feeding of dairycows (http://www.parmigianoreggiano.com/consor-tium/rules_regulation_2/feeding_regulation_dairy_cows_1.aspx), cows that produce milk destined forthe production of cheese have to be fed local for-age, at least 50% dry matter of which must beprovided by hay, and any type of silage is forbid-den. As a consequence, crop rotation is widespreadand it is quite easy to find fields with short vegeta-tion throughout the whole breeding season, giventhe high frequency of harvesting of forage grasses,starting in late April. Aside from periodic cuts ofvegetation, farmyard manure is regularly used toincrease the organic content of soils. Previous stud-ies have demonstrated that applications of manureon arable fields have positive effects on bothsurface activity and abundance of soil inverte-
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 717

brates, chiefly earthworms and dung beetles (Curry1976, Tucker 1992, Hutton & Giller 2003, Holeet al. 2005), which are important sources of foodfor Stone-curlews (Amat 1986, Rodriguez & DelCampo 1987, Sanchez Gonzalez 1996, Greenet al. 2000, Giannangeli et al. 2004, Giunchi et al.2009).
In summary, our results indicate that Stone-curlews essentially make use of two differenthabitats in the study area: (1) the gravel river-bed, which provides suitable breeding territoriesand part of the food resources needed for repro-duction, and (2) arable farmland, where the agri-cultural mechanization and the lack of pasturesare probably compensated for by crop rotationand farmyard manure applications which provideconvenient food resources, and which the birdsexploit when human disturbance is lower (atnight). It seems likely that the proximity ofthese two particular situations in the same areais one of the main causes of the high breedingdensity recorded in the area, which is amongstthe highest reported for the species in Europe(distances between simultaneously active nestscould be < 40 m). This suggests that the charac-teristics of the adjacent riverside habitat couldrepresent an important criterion for assessing thesuitability of a given river for the settlement ofthe species.
In the last two decades, a large number of stud-ies have focused on the impact of agriculturalactivities on birds. Agricultural intensification,which negatively affects several critical stages of abird’s life cycle, such as reproduction and foraging,is probably the main cause of the recent and welldocumented negative trends of European farmlandbird species (Wilson et al. 2009). Our study showsthat some degree of intensification of farmingactivities may be tolerated by Stone-curlews, aslong as fairly well-preserved natural areas are alsopresent in the vicinity. This indicates the need fora synergistic management of protected and non-protected areas and further stresses the importanceof habitat heterogeneity for birds and more gener-ally for biodiversity conservation in human-modi-fied landscapes (Benton et al. 2003).
We are grateful to all the people who helped us in thefield: F. Carpita, L. Catellani, E. Chierici, C. Dalle Mura,E. Dimitri, E. Logozzo, S. Masi, F. Piccio and R. Rusti-cali. We would also like to thank the farms Bocchi, Gen-nari, Qualatico and Cà Nova for allowing free access to
their fields and the Taro River Regional Park for support-ing the research. We appreciate the improvements inEnglish usage made by Phil Whitford through the Asso-ciation of Field Ornithologists’ programme of editorialassistance. The comments of F. Rodriguez, D. Chamber-lain and two anonymous reviewers greatly improved anearlier draft of the manuscript.
REFERENCES
Alexander, R.D. 1974. The evolution of social behavior. Annu.
Rev. Ecol. Syst. 5: 325–383.
Amat, J.A. 1986. Information on the diet of the Stone-curlew
Burhinus oedicnemus in Donana, southern Spain. Bird
Study 33: 71–73.
Bailey, D. & Alimadhi, F. 2007. Logit.mixed: mixed effects
logistic regression. In Imai, K., King, G. & Lau, O. (eds)
Zelig: Everyone’s Statistical Software. http://gking.har-
vard.edu/zelig.
Bates, D. & Maechler, M. 2010. Lme4: Linear mixed-effects
models using S4 classes. R package version 0.999375-33.
Beauchamp, G. 2005. Does group foraging promote efficient
exploitation of resources? Oikos 111: 403–407.
Beauchamp, G. 2007. Exploring the role of vision in social for-
aging: what happens to group size, vigilance, spacing,
aggression and habitat use in birds and mammals that for-
age at night? Biol. Rev. 82: 511–525.
van Belle, G. 2008. Statistical Rules of Thumb, 2nd edn.
Hoboken: John Wiley & Sons.
Benton, T.G., Vickery, J.A. & Wilson, J.D. 2003. Farmland
biodiversity: is habitat heterogeneity the key? Trends Ecol.
Evol. 18: 182–188.
Berdeen, J.B. & Krementz, D.G. 1998. The use of fields at
night by wintering American Woodcock. J. Wildl. Manage.
62: 939–947.
Bibby, C.J., Burgess, N.D., Hill, D.A. & Mustoe, S.H. 2000.
Bird Census Techniques. London: Academic Press.
BirdLife International 2004. Birds in Europe. Wageningen:
BirdLife International.
Brichetti, P. & Fracasso, G. 2004. Ornitologia Italiana, vol. 2
Tetraonidae-Scolopacidae. Bologna: Alberto Perdisa Editore.
Burfield, I.J. 2005. The conservation status of steppic birds in
Europe. In Bota, G., Morales, M.B., Manosa, S. & Campro-
don, J. (eds) Conservation of Steppe-land Birds: 119–139.
Barcelona: Lynx Edicions & Centre Tecnologic Forestal de
Catalunya.
Burger, J. & Gochfeld, M. 1991. Human activity influence and
diurnal and nocturnal foraging of Sanderlings (Calidris
alba). Condor 93: 259–265.
Burnham, K. & Anderson, D. 2002. Model Selection and
Multimodel Inference – A Practical Information-Theoretic
Approach. New York: Springer.
Butler, S.J. & Gillings, S. 2004. Quantifying the effects of
habitat structure on prey detectability and accessibility to
farmland birds. Ibis 146: 123–130.
Calenge, C. 2006. The package ‘adehabitat’ for the R soft-
ware: a tool for the analysis of space and habitat use by
animals. Ecol. Model. 197: 516–519.
Calvario, E., Gustin, M., Sarrocco, S., Gallo Orsi, U., Bulga-
rini, F. & Fraticelli, F. 1999. Nuova lista rossa degli uccelli
nidificanti in Italia. Riv. Ital. Ornitol. 69: 3–43.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
718 C. Caccamo et al.

Cody, M.L. (ed.) 1985. Habitat selection in birds. Orlando:
Academic Press.
Colwell, M. 2010. Shorebird Ecology, Conservation, and Man-
agement. Berkeley: University of California Press.
Cramp, S. & Simmons, K. (eds) 1983. The Birds of the Wes-
tern Palearctic, vol. 3. London: Oxford University Press.
Crawley, M.J. 2007. The R Book. Chichester: John Wiley and
Sons.
Curry, J. 1976. Some effects of animal manure on earthworms
in grassland. Pedobiologia 16: 425–438.
Delany, S., Scott, D., Didman, T. & Stroud, D. 2009. An Atlas
of Wader Populations in Africa and Western Eurasia.
Wageningen: Wetlands International.
Devereux, C.L., McKeever, C.U., Benton, T.G. & Whitting-
ham, M.J. 2004. The effect of sward height and drainage
on Common Starlings Sturnus vulgaris and Northern Lap-
wings Vanellus vanellus foraging in grassland habitats. Ibis
146: 115–122.
Duriez, O., Ferrand, Y., Binet, F., Corda, E., Gossmann, F.
& Fritz, H. 2005a. Habitat selection of the Eurasian wood-
cock in winter in relation to earthworms availability. Biol.
Conserv. 122: 479–490.
Duriez, O., Fritz, H., Binet, F., Tremblay, Y. & Ferrand, Y.
2005b. Individual activity rates in wintering Eurasian Wood-
cocks: starvation versus predation risk trade-off? Anim.
Behav. 69: 39–49.
Duriez, O., Fritz, H., Siad, S. & Ferrand, Y. 2005c.
Wintering behaviour and spatial ecology of Eurasian Wood-
cock Scolopax rusticola in western France. Ibis 147: 519–
532.
Duriez, O., Eraud, C. & Bretagnolle, V. 2010. First measure-
ments of metabolic rates in the Stone-curlew, a nocturnal
inland wader. Wader Stud. Group Bull. 117: 119–122.
Getz, W.M., Fortmann-Roe, S., Cross, P.C., Lyons, A.J. &
Ryan, S.J. 2007. LoCoH: nonparametric kernel methods for
constructing home ranges and utilization distributions. PLoS
ONE 2: e207.
Giannangeli, L., de Sanctis, A., Manginelli, R. & Medina,
F.M. 2004. Seasonal variation of the diet of the Stone-
curlew Burhinus oedicnemus distinctus at the island of La
Palma, Canary Islands. Ardea 92: 175–184.
Gillings, S. & Sutherland, W. 2007. Comparative diurnal
and nocturnal diet and foraging in Eurasian Golden Plo-
vers Pluvialis apricaria and Northern Lapwings Vanellus
vanellus wintering on arable farmland. Ardea 95: 243–
257.
Gillings, S., Fuller, R.J. & Sutherland, W.J. 2005. Diurnal
studies do not predict nocturnal habitat choice and site
selection of European Golden Plovers (Pluvialis apricaria)
and Northern Lapwings (Vanellus vanellus). Auk 122:
1249–1260.
Giunchi, D. & Pollonara, E. 2007. On sexing Stone-curlew
Burhinus oedicnemus by cloacal examination. Wader Stud.
Group Bull. 113: 71–72.
Giunchi, D., Pollonara, E. & Baldaccini, N.E. (eds) 2009.
L’occhione (Burhinus oedicnemus): Biologia e conservazi-
one di una specie di interesse comunitario – Indicazioni per
la gestione del territorio e delle aree protette. Collecchio:
Consorzio del Parco Fluviale Regionale del Taro.
Goss-Custard, J.D. & Verboven, N. 1993. Disturbance and
feeding shorebirds on the Exe estuary. Wader Stud. Group
Bull. 68: 59–66.
Green, R. & Griffiths, G. 1994. Use of preferred nesting habi-
tat by Stone-curlews Burhinus oedicnemus in relation to
vegetation structure. J. Zool. 233: 457–471.
Green, R. & Taylor, C. 1995. Changes in Stone-curlew Burhi-
nus oedicnemus distribution and abundance and vegetation
height on chalk grassland at Porton-Down, Wiltshire. Bird
Study 42: 177–181.
Green, R., Tyler, G. & Bowden, C. 2000. Habitat selection,
ranging behaviour and diet of the Stone-curlew (Burhinus
oedicnemus) in Southern England. J. Zool. 250: 161–183.
Hamilton, W.D. 1971. Geometry for the selfish herd. J. Theor.
Biol. 31: 295–311.
Heezen, K.L. & Tester, J.R. 1967. Evaluation of radio-tracking
by triangulation with special reference to deer movements.
J. Wildl. Manage. 31: 124–141.
Hole, D.G., Perkins, A.J., Wilson, J.D., Alexander, I.H.,
Grice, P.V. & Evans, A.D. 2005. Does organic farming
benefit biodiversity? Biol. Conserv. 122: 113–130.
Hoodless, A.N. & Hirons, G.J.M. 2007. Habitat selection and
foraging behaviour of breeding Eurasian Woodcock Scolo-
pax rusticola: a comparison between contrasting land-
scapes. Ibis 149: 234–249.
Huck, M., Davison, J. & Roper, T. 2008. Comparison of two
sampling protocols and four home-range estimators using
radio-tracking data from urban Badgers Meles meles. Wild-
life Biol. 14: 467–477.
Hutton, S.A. & Giller, P.S. 2003. The effects of the intensifica-
tion of agriculture on northern temperate dung beetle com-
munities. J. Appl. Ecol. 40: 994–1007.
Johnson, D.H. 1980. The comparison of usage and availability
measurements for evaluating resource preference. Ecology
61: 65–71.
Kenward, R. 2001. A Manual for Wildlife Radio Tagging.
London: Academic Press.
Manly, B., McDonald, L., Thomas, D., McDonald, T. & Erik-
son, W. 2002. Resource Selection by Animals: Statistical
Design and Analysis for Field Studies. Dordrecht: Kluwer
Academic Publishers.
Mardia, K.V. 1972. Statistics of Directional Data. London: Aca-
demic Press.
Martin, G. 1990. Birds at Night. London: T. & A.D. Poyser.
Martin, G. & Katzir, G. 1994. Visual-fields in the Stone-curlew
Burhinus oedicnemus. Ibis 136: 448–453.
Martın, A. & Lorenzo, J.A. 2001. Aves del Archipielago
Canario. La Laguna: Francisco Lemus Editor.
Marzluff, J.M. & Sallabanks, R. (eds) 1998. Avian conserva-
tion: Research and Management. Washington, DC: Island
Press.
Millspaugh, J.J. & Marzluff, J.M. (eds) 2001. Radio Tracking
and Animal Populations. London: Academic Press.
Mohr, C.O. 1947. Table of equivalent populations of North
American small mammals. Am. Midl. Nat. 37: 223–249.
Paoletti, M.G. 1999. The role of earthworms for assessment of
sustainability and as bioindicators. Agr. Ecosyst. Environ.
74: 137–155.
R Development Core Team 2010. R: A Language and Environ-
ment for Statistical Computing. Vienna: R Foundation for
Statistical Computing. Available at: http://www.R-project.org.
Roberts, G. 1996. Why individual vigilance declines as group
size increases? Anim. Behav. 51: 1077–1086.
Rodriguez, F. & Del Campo, F. 1987. Datos sobre la dieta del
Alcaravan Burhinus oedicnemus distinctus (Bannerman
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
Stone-curlew ranging behaviour 719

1914) en la localidad de Gran Canaria. In Actas del I Cong-
reso Internacional de Aves Esteparias: 175–181. Leon:
Junta de Castilla y Leon.
Sanchez Gonzalez, D.L. 1996. Dieta primaveral del
Alcaravan, Burhinus oedicnemus insularum (Sassi, 1908),
en Alegranza, islas Canarias (Aves, Burhinidae). Vieraea
25: 31–35.
Schekkerman, H. & Beintema, A. 2007. Abundance of inver-
tebrates and foraging success of Black-tailed Godwit Limo-
sa limosa chicks in relation to agricultural grassland
management. Ardea 95: 39–54.
Shepherd, P.C. & Lank, D.B. 2004. Marine and agricultural
habitat preferences of Dunlin wintering in British Columbia.
J. Wildl. Manage. 68: 61–73.
Sokal, R.R. & Rohlf, F.J. 1995. Biometry. New York: W.H.
Freeman and Company.
Sutherland, W. 1996. Ecological Census Techniques: A Hand-
book. Cambridge: Cambridge University Press.
Sutherland, W.J., Newton, I. & Green, R.E. 2004. Bird Ecol-
ogy and Conservation. Oxford: Oxford University Press.
Thomas, D.L. & Taylor, E.J. 1990. Study designs and tests
for comparing resource use and availability. J. Wildl. Man-
age. 54: 322–330.
Thomas, K., Kvitek, R.G. & Bretz, C. 2003. Effects of human
activity on the foraging behavior of Sanderlings Calidris
alba. Biol. Conserv. 109: 67–71.
Tinarelli, R., Alessandria, G., Giovacchini, P., Gola, L., Ien-
tile, R., Meschini, A., Nissardi, S., Parodi, R., Perco, F.,
Taiariol, P.L. & Zucca, C. 2009. Consistenza e distribuzi-
one dell’occhione in Italia: aggiornamento al 2008. In Giun-
chi, D., Pollonara, E. & Baldaccini, N.E. (eds) L’occhione
(Burhinus oedicnemus): Biologia e Conservazione di una
Specie di Interesse Comunitario – Indicazioni per la Gesti-
one del Territorio e delle Aree Protette: 45–50. Collecchio:
Consorzio del Parco Fluviale Regionale del Taro.
Triolo, S., Campobello, D. & Sara, M. 2011. Diurnal habitat
suitability for a Mediterranean steppeland bird, identified by
Ecological Niche Factor Analysis. Wildl. Res. 38: 152–162.
Tucker, G. 1992. Effects of agricultural practices on field use
by invertebrate-feeding birds in winter. J. Appl. Ecol. 29:
779–790.
Tucker, G. & Evans, M. 1997. Habitats for Birds in Europe: A
Conservation Strategy for the Wider Environment. Cam-
bridge: BirdLife International.
Vaughan, R. & Vaughan Jennings, N. 2005. The Stone-
curlew Burhinus oedicnemus. Falmouth: Isabelline Books.
White, G. & Garrott, R. 1990. Analysis of Wildlife Radio-track-
ing Data. San Diego: Academic Press.
Whittingham, M. 1996. The use of radio telemetry to measure
the feeding behavior of breeding European Golden Plovers.
J. Field Ornithol. 67: 463–470.
Whittingham, M., Percival, S. & Brown, A. 2000. Time bud-
gets and foraging of breeding golden plover Pluvialis apri-
caria. J. Appl. Ecol. 37: 632–646.
Wilson, J.D., Whittingham, M.J. & Bradbury, R.B. 2005. The
management of crop structure: a general approach to
reversing the impacts of agricultural intensification on birds?
Ibis 147: 453–463.
Wilson, J.D., Evans, A. & Grice, P. 2009. Bird Conservation
and Agriculture. New York: Cambridge University Press.
van Winkle, W. 1975. Comparison of several probabilistic
home-range models. J. Wildl. Manage. 39: 118–123.
Wolak, M.E., Fairbairn, D.J. & Paulsen, Y.R. 2011. Guide-
lines for estimating repeatability. Methods Ecol. Evol. doi:
10.1111/j.2041-210X.2011.00125.x.
Yasue, M. 2005. The effects of human presence, flock size
and prey density on shorebird foraging rates. J. Ethol. 23:
199–204.
Received 10 February 2011;revision accepted 17 July 2011.Associate Editor: Niall Burton.
SUPPORTING INFORMATION
Additional Supporting Information may be foundin the online version of this article:
Figure S1. Examples of four home-range mapscalculated using 95% LoCoH. Dark poly-gons = diurnal home-ranges; light polygons = noc-turnal home-ranges; filled dots = capture sites; redline = park boundary; sky blue area = riverbed.
Please note: Wiley-Blackwell are not responsiblefor the content or functionality of any supportingmaterials supplied by the authors. Any queries(other than missing material) should be directed tothe corresponding author for the article.
ª 2011 The Authors
Ibis ª 2011 British Ornithologists’ Union
720 C. Caccamo et al.
Copyright © 2022 FDOKUMEN




















