
Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc...
43
Steenstrupia Fjeldså, J., J. Kiure, N. Doggart, L. A. Hansen & A. Perkin. Distribution of higland forest birds accross a potential dispersal barrier in the Eastern Arc Mountains of Tanzania. – Steenstrupia 32 (1): 1–43. Copenhagen, Denmark. February 2010. ISSN 0375-2909. A detailed inventory is provided of the avifauna of montane forests in two highland areas in the Eastern Arc Mountains of Tanzania representing adjacent fault blocks separated by the Great Ruaha River Basin. For the first time detailed survey data is provided for all major montane forests south and north of this potential dispersal barrier. Detailed distribution data and novel interpretations of species boundaries make it possible to tell apart the relative roles of the barrier and of environmental factors affecting the local persistence of populations. Highland forests on opposite sides of the Great Ruaha Basin had virtually identical faunas of resident montane forest birds, and many range-restricted species had dense populations in both highlands. Unlike this, species depending on foothill forest, at least for part of the year, had small or no populations north of the Great Ruaha River. Apparently, the Rubeho area represents sink habitat for many low- or mid-elevation species, and although there are no dispersal barriers in the foothill zone, it is suggested that this zone represents sink habitat which provides little gene flow. Sharp species replacements, although sometimes corresponding to physical barriers, may be secondary phenomena arising as a consequence of population expansion and incompatibility of the involved populations. Physical breaks in the landscape may enhance speciation if they are large enough, but the overwhelming tendency to search for barriers as explanation of speciation may often lead to neglect of other environmental variables that enhance local population persistence. Detailed molecular studies, with nested clade analysis, may greatly improve our understanding of the process of biological diversification within this biodiversity hotspot, and this may guide our decisions about where to focus conservation efforts. Keywords: Aves, Afrotropical, Eastern Arc hotspot, speciation ¹Zoological Museum, Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark. ²P.O. Box 12993, Dar es Salaam, Tanzania. ³Tanzania Forest Conservation Group, P.O. Box 23410, Dar es Salaam, Tanzania. E-mail for Jon Fjeldså: [email protected] Steenstrupia 32 (1): 1–43. Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc Mountains of Tanzania JON FJELDSŹ, JACOB KIURE², NIKE DOGGART³, LOUIS A. HANSEN¹ & ANDREW PERKIN³ INTRODUCTION The Eastern Arc Mountains of Tanzania and Kenya are globally significant because of their high concentration of endemic species of flora and fauna (Burgess et al. 1998, Mittermeier et al. 2000, Eken et al. 2004). Large and small high- lands form a punctuated chain from the Taita hills in south-eastern Kenya to the Iringa highland in south-western Tanzania, where the Eastern Arc Mts are separated by a dry and barren upland plateau called the “Makambako Gap” from the “Southern Highlands” near the Malawi (Nyasa) Rift (Fig. 1). Moreau (1966) lumped all these highlands in a “Tanganyika-Nyasa Mountain Group”, but Lovett (1990) demonstrated a sig- nificant dichotomy, as the Malawi Rift mountains are young and partly volcanic, and influenced by local – and historically unstable – convectional rainfall, whereas those of the Eastern Arc are an- cient crystalline fault-blocks, and under constant influence of humid winds from the Indian Ocean. Thus, the Eastern Arc Mts may have had humid forest continuously since the trans-African mid- Tertiary super-rainforest broke up because of uplift in central Africa, global cooling and a shift to more grass-dominated ecosystems in Africa in the upper Miocene (Vrba et al. 1995, Jacobs et al. 1999, Flenley 2005). They may thus have provided stable forest refuges during the Pleisto-
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc...

1distribution of highland forest birds in tanzania
Steenstrupia Fjeldså, J., J. Kiure, N. Doggart, L. A. Hansen & A. Perkin. Distribution of higland forest birds accross a potential dispersal barrier in the Eastern Arc Mountains of Tanzania. – Steenstrupia 32 (1): 1–43. Copenhagen, Denmark. February 2010. ISSN 0375-2909.
A detailed inventory is provided of the avifauna of montane forests in two highland areas in the Eastern Arc Mountains of Tanzania representing adjacent fault blocks separated by the Great Ruaha River Basin. For the first time detailed survey data is provided for all major montane forests south and north of this potential dispersal barrier. Detailed distribution data and novel interpretations of species boundaries make it possible to tell apart the relative roles of the barrier and of environmental factors affecting the local persistence of populations. Highland forests on opposite sides of the Great Ruaha Basin had virtually identical faunas of resident montane forest birds, and many range-restricted species had dense populations in both highlands. Unlike this, species depending on foothill forest, at least for part of the year, had small or no populations north of the Great Ruaha River. Apparently, the Rubeho area represents sink habitat for many low- or mid-elevation species, and although there are no dispersal barriers in the foothill zone, it is suggested that this zone represents sink habitat which provides little gene flow. Sharp species replacements, although sometimes corresponding to physical barriers, may be secondary phenomena arising as a consequence of population expansion and incompatibility of the involved populations. Physical breaks in the landscape may enhance speciation if they are large enough, but the overwhelming tendency to search for barriers as explanation of speciation may often lead to neglect of other environmental variables that enhance local population persistence. Detailed molecular studies, with nested clade analysis, may greatly improve our understanding of the process of biological diversification within this biodiversity hotspot, and this may guide our decisions about where to focus conservation efforts.
Keywords: Aves, Afrotropical, Eastern Arc hotspot, speciation
¹Zoological Museum, Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark. ²P.O. Box 12993, Dar es Salaam, Tanzania. ³Tanzania Forest Conservation Group, P.O. Box 23410, Dar es Salaam, Tanzania. E-mail for Jon Fjeldså: [email protected]
Steenstrupia 32 (1): 1–43.
Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc Mountains of TanzaniaJON FJELDSŹ, JACOB KIURE², NIKE DOGGART³, LOUIS A. HANSEN¹ & ANDREW PERKIN³
INTRODUCTION
The Eastern Arc Mountains of Tanzania and Kenya are globally significant because of their high concentration of endemic species of flora and fauna (Burgess et al. 1998, Mittermeier et al. 2000, Eken et al. 2004). Large and small high-lands form a punctuated chain from the Taita hills in south-eastern Kenya to the Iringa highland in south-western Tanzania, where the Eastern Arc Mts are separated by a dry and barren upland plateau called the “Makambako Gap” from the “Southern Highlands” near the Malawi (Nyasa) Rift (Fig. 1). Moreau (1966) lumped all these highlands in a “Tanganyika-Nyasa Mountain Group”, but Lovett (1990) demonstrated a sig-
nificant dichotomy, as the Malawi Rift mountains are young and partly volcanic, and influenced by local – and historically unstable – convectional rainfall, whereas those of the Eastern Arc are an-cient crystalline fault-blocks, and under constant influence of humid winds from the Indian Ocean. Thus, the Eastern Arc Mts may have had humid forest continuously since the trans-African mid-Tertiary super-rainforest broke up because of uplift in central Africa, global cooling and a shift to more grass-dominated ecosystems in Africa in the upper Miocene (Vrba et al. 1995, Jacobs et al. 1999, Flenley 2005). They may thus have provided stable forest refuges during the Pleisto-

j. fjeldså et al.2
cene glacial periods (Prell et al. 1980, Marchant et al. 2007). As a result, the Eastern Arc has many highly distinctive endemics (e.g., Davenport et al. 2006), some of them relictual forms whose nearest relatives live far away, for instance in the Guinea-Congolian rainforest, and even on other continents (Burgess et al. 1998).
At the same time it is also an area with strong local differentiation of closely related species, es-pecially of arthropods, plants and herptiles (e.g., Lindqvist & Albert 2001, Huber 2003, Burgess et al. 2006), but also the birds, which will be dis-cussed in this paper, comprise groups of closely related species. The excessive endemism in this area suggests that it may act as a “species pump” providing recruitment to the regional fauna (Jetz et al. 2004), and within this area the Udzungwa Mts have been identified as particularly impor-tant (e.g., as the top priority area for primate conservation in Africa; Jones et al. 2005).
If we want to conserve this potential “spe-cies pump” it is relevant to understand how it works, mechanistically, for instance whether the diversification process is a simple consequence of physical barriers separating populations, or of habitat gaps that arise because of the historical instability of the African climate, or of special lo-cal conditions that allow populations to persist, in spite of the instability of the regional climate. The present study will examine these relationships in a segment of the Eastern Arc Mountains that has hitherto been very incompletely explored (see Isango 2001). We will present the first thorough documentation of how the forest birds are dis-tributed across the highlands flanking a potential barrier in the Eastern Arc, and we will use these data to evaluate alternative interpretations of the biogeographic patterns. The study area includes the north-eastern part of the Udzungwa highland, well known for its extraordinary concentration of endemic species, and the next highland to the north (N in the following), often referred to as the Rubeho Mountains, which has been considered biologically poorer (Burgess et al. 1998) although it has also been less well explored (Doggart et al. 2006).
Complex and species-rich faunas in montane regions (Fig. 2) are generally interpreted in the literature as a result of vicariance caused by physical breaks in the landscape, such as barren
ridges, or deep valleys, isolating populations which once were connected. This seems imme-diately logical in the case of the Eastern Arc Mts, which represent independently uplifted tectonic fault blocks, separated by more or less extensive lowlands (Stuart et al. 1993, Beresford et al. 2004). However, few authors took alternative interpretations into account, for instance that species-replacements across these barriers could be secondary phenomena caused by ecological incompatibility of taxa whose origins are not yet understood (e.g., Louette 1992; García-Moreno & Fjeldså 2000 for Andean birds).
Under the vicariance model we expect co-distributed taxa to have a common history of vicariance events and therefore to share patterns of differentiation and population divergence (Nelson & Platnick 1981). This might even apply to volant groups like birds, considering the reluc-tance of many tropical forest birds to move out of their preferred habitat (e.g., Diamond 1981, Bierregaard et al. 2001, Poulsen 1994).
However, congruent replacement patterns have rarely been documented, and it is therefore relevant to consider whether gene-pools may be separated in other ways than by discrete geo-logical events. Here it is relevant to take into account processes at community level. As the complexity of communities – and the number of competitive interactions – increases, indi-vidual species will be pushed towards extinction thresholds (Ovaskainen & Hanski 2003), with some of them becoming rare and relictual with widely scattered remnant populations, at least at some points in time. This process is independent of geological events, as it is driven by species turnover in local communities, environmental instability and source-sink dynamics. The latter mechanisms assumes that the individual species act as metapopulations – dynamic networks of lo-cal populations of which some are persistent and viable while others, in places with unpredictable ecological conditions, may dwindle away. If their existence depends on spill-over from optimal (“source”) habitats, their evolutionary relevance may be negligable (Pulliam & Danielsen 1991, Holt 1997, Hanski 1999). Range disjunctions are sometimes found in interconnected and seem-ingly uniform landscapes (Diamond 1980), and will profoundly affect the genetic structure of the

3distribution of highland forest birds in tanzania
involved species.Judging from strongly nested distribution
patterns of birds of the Eastern Arc Mts, it can be inferred that once-widespread species have been lost in a fairly predictable sequence outside the main “source” areas (Cordeiro 1998). Thus, the geographical patterns could be linked with intrinsic properties of the different highlands rather than with landscape breaks between them (Jansson & Dynesius 2002). The remaining remnant populations may, under constantly fa-vourable conditions, diverge slowly by genetic drift (Ricklefs & Latham 1992), and, as they are perfectly adapted to these conditions they may at some point start to expand, with population growth (reducing the role of genetic drift) facili-tating adaptation to the more unstable regional climage. As they expand, the populations may meet as different (para-) species, but unlike in the vicariance model, the replacement patterns will not be congruent between lineages. On the other hand we may predict a nestedness of species ranges, and corresponding patterns of genetic diversity within individual species (as revealed by coalescent-based tests of historical population history; e.g. Drummond et al. 2005).
METHODS
Study areasThe two main study areas, the Udzungwa and Rubeho highlands, represent undulating areas at 1100–2500 m elevation, with a mosaic veg-etation of woodlands, grasslands and montane forest. The highlands are separated by a semiarid lowland area through which the Great Ruaha River flows, draining the basin through a narrow erosion canyon (Figs 3 and 4). While baking hot lowland savannas with deciduous Adansonia (baobab) woodlands and Acacia thickets sepa-rates the montane forests of the two highlands, there is only a narrow erosion gap through the humid foothills to the east. This allows us to contrast the well-marked isolation of upper mon-tane forests N and S of the gap with the almost continuous band of eastern foothill habitats (see Figs 3 and 4).
Previously unexplored highland forests near the Great Ruaha Gap were visited mainly in 1997, 1999–2003 and 2006–2007. We focus here on forest habitats above 1500 m (montane and upper montane forest) in the eastern Udzungwa and Rubeho highlands, with outlying smaller mountains (locality names with abbreviations in Fig. 3 being Nyumbanitu [Ny], Ndundulu [Nd], Luhombero [Lu], Kisinga Rugaro [KR], Image [Im], Ukami [Uka] and Mwanihana [Mw] forests and Gologolo Mts [Gg]; Uvidunda highland with Iyungi forest [Iy], Wota Mts with Wota [Wo] and Ligunga [Li] forests, Mangalisa highland [Ma] and Rubeho highland with Msanga [Ms], Ukwiva [Uk] and Mafwemiro [M]; lowland lo-calities are Great Ruaha Basin [GRB], Matundu lowland forest [Mat], Kilosa [Ki], Mpwapwa [Mpw] and Ulaya [Ul]). Because of the lack of previous descriptions for most of the forest tracts we provide fairly detailed habitat descriptions in Appendix 1. The main landscape features of the study area, the cover of evergreen forest and ap-proximate location of study sites appear in Fig. 3, and topographic transects across the Great Ruaha Gap appear in Fig. 4.
The forest cover was calculated from 1:50.000 topographic maps (horizontal projection, count-ing km² or hectare grid-cells as appropriate), after forest boundaries have been revised from satellite images (including Google Earth) and field notes.
The ornithological fieldwork
The new studies that we report here represent more than 30 man-months of ornithological fieldwork in the forests of the study area, and a similar effort has been spent in other highlands in Tanzania, and these studies cover both the rainy and dry seasons. In all study areas, we camped inside the forest, and therefore could record birds throughout the day and often also at night. The principal study method was by quiet walking, watching and listening for birds, supplemented by mist-netting, with 5–10 nets operated near the campsites. During some study periods when JK worked alone (Appendix 1), the effort to mist-net birds limited the amount of exploratory walking. Collected voucher specimens and tissue samples (numbers specified in the text where relevant) are
j. fjeldså et al.4
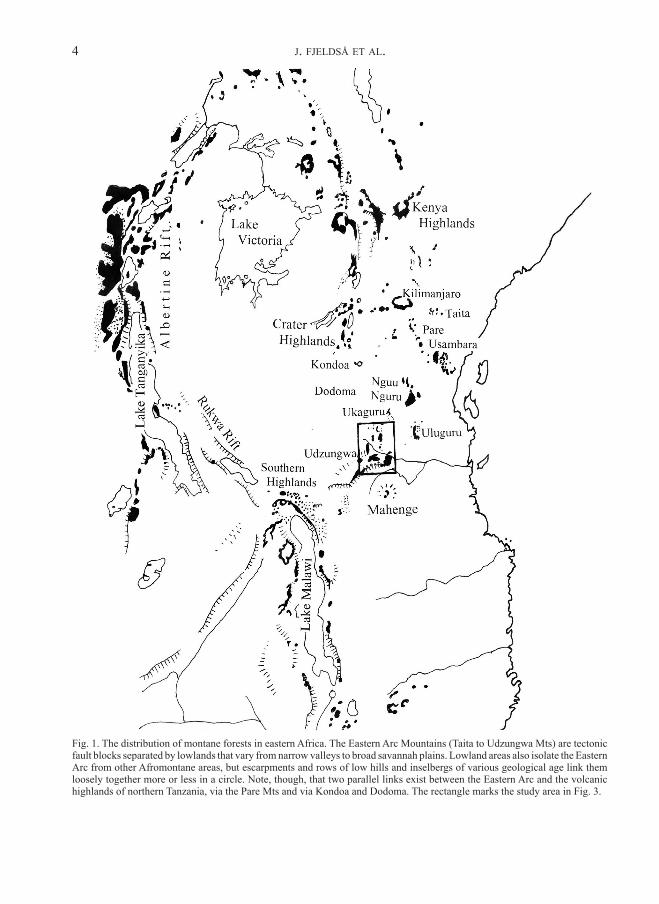
Fig. 1. The distribution of montane forests in eastern Africa. The Eastern Arc Mountains (Taita to Udzungwa Mts) are tectonic fault blocks separated by lowlands that vary from narrow valleys to broad savannah plains. Lowland areas also isolate the Eastern Arc from other Afromontane areas, but escarpments and rows of low hills and inselbergs of various geological age link them loosely together more or less in a circle. Note, though, that two parallel links exist between the Eastern Arc and the volcanic highlands of northern Tanzania, via the Pare Mts and via Kondoa and Dodoma. The rectangle marks the study area in Fig. 3.

5distribution of highland forest birds in tanzania
Fig. 2. The variation in species richness of forest-associated birds in eastern Africa, in a 15’ geographical grid, based on the Copenhagen biodiversity databases, Burgess et al. 1998; darkest tone representing >180 species. See legend of Fig. 1 for further details.

j. fjeldså et al.6
Fig. 3. Detailed map of the highlands flanking the Great Ruaha Basin in central Tanzania, based on topographic maps, interpreta-tions of satellite images and field observations. Black areas mark evergreen forests, which are mainly above 1500 m elevation but extend down to the lowlands in some places (notably in the Mwanihana and Matundu forests). Abbreviations are explained in the main text (p. 5). The map covers an area of 120 x 170 km.

7distribution of highland forest birds in tanzania
deposited at the Zoological Museum, University of Copenhagen (ZMUC) and at the University of Dar es Salaam.
The first author reviewed the bird material from East Africa in the Zoological Museum, University of Copenhagen, National Museums of Kenya (several visits), American Museum in New York (some taxa only), Academy of Natural Sciences of Philadelphia, Field Museum in Chi-cago, Milwaukee Museum, Museum Alexander König in Bonn, Muséum National d’Histoire Na-turelle in Paris, and Naturalis Museum in Leiden. Some material was obtained from The Natural History Museum in Tring as loans. We do not provide an exhaustive historical record of bird distributions, as these data are summarised in various handbooks (e.g., Britton 1980 and most recently Baker & Baker 2002), and we omit data collected in 2006-2007 on lower elevations and partly in other habitats.
The data collected during fieldwork were orga-nized as species lists per locality, with study sites and altitudes specified as appropriate. For certain analyses, the data were pooled per highland. In order to document the variation in community structure between study sites, JF used a “random
walk” census method, which is specially adapted for dense montane forest where there is no estab-lished trail system (Fjeldså 1999; and see legend to Table 1): within ca. 1 km2 in each study area line transects were made randomly, as far as the terrain permits, and all birds seen or heard within 50 m of the transect line were identified and recorded. Line transect data from tropical forest correlate well with results of more precise – but much more time-consuming – spot-mapping (Shahkar Raman 2003) and we therefore assume that this represents a much more time-efficient sampling approach for this kind of terrain. Con-sidering the degree of habitat heterogeneity and the mobility of feeding parties of birds, study plots of c. 1 km2 extension are used. The walking speed varies according to how difficult the terrain is, but is usually ca. 500 m per hour. Because of the many constraints, the sample-size cannot be fully standardised, but for analyses we only used samples with >500 bird identifications. This al-lows making a species richness estimate, drawing a ranked abundance curve for the community, and calculating a Rarity Index, which is the sum of inverse range-sizes (global breeding range recorded in a grid of 1x1 geographical degrees;
Fig. 4. Altitudinal profiles (115 km) along the four stippled lines in Fig. 3. The cross-shading marks areas that are suitable for evergreen forest. The Great Ruaha River, connecting the dry interior of Tanzania with the coastal plains, is 300-500 m.a.s.l., and must have eroded its way through the granite rock continuously since the start of the mountain up-lift in the Miocene. The wide basin in the three upper profiles is dry and hot, and even the narrow canyon further E (in the lower profile) has deciduous forest in the bottom, but towards the western end of the canyon the vegetation becomes evergreen.

j. fjeldså et al.8
see below) for all individual birds, divided by the total number of birds in the sample. This index will reflect the relative dominance in the local avian community of species with very small global distributions.
Our study will primarily focus on birds asso-ciated with high-elevation forest. Most of these are resident or make moderate altitudinal migra-tions (breeding in the highland during the austral summer but leaving the higher ridges during the austral winter, sometimes even reaching the foothill zone; Burgess & Mlingwa 2000, Romdal 2001a). However, we also consider submontane species which are found fairly regularly above 1500 m (see legend to Appendix 2), and in order to judge the importance of landscape features as dispersal barriers we pay particular attention to records of forest birds in non-forest habitats and in small habitat fragments.
Data describing diversification of the regional avifaunaDistributional databases have been developed over many years, using the Worldmap software (Williams 1994) and a grid of 1x1 geographi-cal degrees for all resident bird species of sub-Saharan Africa, and a 15x15 minute grid for forest birds of eastern Africa (Burgess et al. 1998, Brooks et al. 2001, Fjeldså & Tushabe 2005; and see Fig. 2). The first of these databases was used here for an objective assessment of global rarity (see above), by assigning each species a range-size rarity score, which is the inverse number of occupied grid-cells in this database. The second is used (in conjunction with the available evi-dence about systematic relationships of species and subspecies) for the broader geographical analysis and for recording the distribution of geographical replacements along the chain of Eastern Arc Mts.
When reviewing the geographical differentia-tion within the Eastern Arc Mts we will refer to recent molecular studies providing phylogenetic hypotheses for some multi-species groups (Roy et al. 1998; Beresford 2003 and Beresford et al. 2004; Bowie 2003, Bowie et al. 2004a, 2005; Fjeldså et al. 2006, 2007; Fuchs et al. 2006, Nguembock et al. 2006, 2008) and describing historical population structures using large num-
bers of tissue samples and nested-clade analysis, an approach specially designed to tell current processes of gene flow and genetic drift apart from events in the past history of clades (Bowie et al. 2004b, 2006, Bowie & Fjeldså 2005, Fjeldså & Bowie 2008, and unpublished).
RESULTS
Habitat availabilityThe elevational distribution of evergreen forest was found to differ significantly between the highlands south and north of the Great Ruaha Gap (Fig. 5). South of the gap the entire east-facing slope has a high proportion of evergreen trees, but N of the gap there is very little ever-green vegetation as the slopes are dominated by tall grasses and deciduous woodland, mostly “miombo” (Brachystegia). The exact distribu-tion of such semi-evergreen vegetation in the lower montane zone is difficult to define, and observations in 1993 between Kisanga and Ulaya (Ki and Ul in Fig. 3) suggest that most mountain slopes marked as “forest” on the topographic maps are rather dense deciduous “miombo” woodland with evergreen trees and undergrowth bushes admixed only in ravines and below very steep slopes. There are also some patches of groundwater forest at the base of the highland, E of Ukwiva (Fjeldså et al. 1997). More con-tinuously forest-covered elevational gradients are again found in the Ukaguru and Nguru Mts (Romdal 2001b) further N. Further details about habitat quality are provided in Appendix 1.
Variation in avian community parametersWe recorded altogether 308 bird species for these highlands (above 1500 m), including migrant visitors, over-flying raptors and water birds and ubiquitous birds of the open land. Appendix 2 shows the distribution of species per forest (in-cluding records of non-forest birds at the edges and in glades in the forest), and adds a few species of the surrounding open land, when these have restricted or otherwise interesting distributions. The total number of species recorded in forest habitat is 124 for the Udzungwa highland and 114

9distribution of highland forest birds in tanzania
Fig. 5. The distribution of evergreen forest (as per cent) S and N of the Great Ruaha Gap in elevational bands. The southern area is the Udzungwa highland W to 36°15' E (in Kisinga-Rugaro Forest) and the northern area comprises the Mangaliza, Uvidunda and Rubeho highlands. The stippled parts of the curves are marked as forest on the topographic maps but our field observation suggest mainly miombo woodland with local admixture of evergreen trees. For the Matundu forest we regarded half the area as deciduous, as all the small hilltops in this forest tract have this kind of vegetation.

j. fjeldså et al.10
for the Rubeho highland (including Mangalisa). Romdal (2003) demonstrated, for the forested E slope of the Udzungwa highland, a broad species richness peak between 700 and 1500 m elevation (the precise location of the peak being seasonally variable) and a sharp decline above this eleva-tion, which means that our study covers only a subset of the entire forest avifauna.
Table 1 provides some community character-istics for the best-studied forests. Here the num-ber of recorded species is supplemented with an estimated species richness (actually an estimate of the asymptote of data obtained during the “ran-dom walk” surveys, using the Chao 1 formula of Colwell & Coddington 1994). Overall, the total number of species is in good agreement with the estimated value, suggesting a fairly complete record (Table 1). For instance, in the Iyungi forest, four persons recorded 64 bird species (including along forest edges and in gaps inside the forest) over one week, and the estimate based on 23 hours “random walk” surveys by JF was
64.3 species. We undoubtedly missed a few spe-cies inhabiting unvisited ecological zones (e.g., evergreen forest extending ca. 200 m down the southern edge of the Mangalisa plateau, where the unrecorded Terpsiphone viridis and Andropadus virens could be expected). In most cases species richness estimates were made on the basis of one visit per forest, and additional species would probably be found at other seasons. How-ever, most additions during the second or third rendezvous to a forest (Mangalisa, Ukwiva and Mafwemiro in Table 1) were visiting non-forest species. We therefore conclude that the species numbers in Table 1 represent a good record of the forest dependent avifauna.
The highest species numbers were found in the largest forest tracts, without much difference between the Udzungwa (Udzungwa Scarp For-est 78 and Nyumbanitu 81 species) and Rubeho (Ukwiva 85 species) highlands. The Image, Mangalisa and Wota forests, which are located W (inland) of the main rain-capturing ridges, have
Forest Udzungwa Image Mangalisa Iyungi Ukwiva Mafwemiro
Forest area, sq. km 200 89 49 5 100 32
Ca. altitude of study site 1700 2000 2100 1700 1800 2000
Species recorded 75 55 57 64 84 65
Species richness estimated from quantitative surveys
75 54.4 53.5 64.3 72.2 58.5
Phylogenetic relict species 8 3 2 3 4 4
Rarity score in - disturbed forest 0.0263 0.0529 0.0561 0.0345 0.0306 0.0198 - mature forest 0.0397 0.0561 0.0477 0.03671 – 0.01461
- lower altitude2 0.0073 – – – 0.0034 –
1 Although this represents the least disturbed part of this forest it does not really represent mature forest.2 Data from semi-evergreen forest at 1100 m in the Udzungwa Scarp Forest Reserve and in ground-water forest at 700 m
below (E) of Ukwiva forest.
Table 1. Species richness and rarity scores for forests birds for localities where quantitative surveys were made. Species richness estimates were made using the Chao 1 formula of Colwell & Coddington (1994). Rarity scores are the sum of inverse range sizes (see Williams 1994, and using the breeding range, recorded in grid-cells of 1x1 geographical degrees in the Copenhagen distributional databases [Brooks et al. 2001] for all individual birds, divided by the number of individual birds in the sample). Thus, high scores reflect numerical dominance of small-range species, index values around 0.05 corresponding to average range-sizes like that of Cossypha anomala in Fig. 6. Forest area is measured from 1:50.000 maps, after forest boundaries have been adjusted based on field observations).

11distribution of highland forest birds in tanzania
reduced species numbers (57, 57 and ca. 54 spe-cies), and this also applies to the vanishing forest remnants on the top plateau of the Kiboriani Mts N of Mpwapwa (Mpw in Fig. 3).
The Rarity Index (Table 1) reflects the relative dominance of species with very small distribu-tions. Very high scores are mainly caused by high densities of range-restricted species like Batis crypta, Andropadus (tephrolaemus) chlorigula, Sheppardia spp., Artisornis metopias and Nectarinia moreaui, and in some places even extremely local forms like Arcanator orostruthus, Swynnertonia swynnertoni and Nectarinia rufipennis contributed significantly. The high-est scores were in Image (0.0529 and 0.0561) and Mangalisa (0.0561 and 0.0477), which are located adjacent to the Great Ruaha Basin, on opposite sides. However, a small dataset from Kisinga Rugaro forest in the central Iringa highland (based on only 228 identified birds, and therefore not included in Table 1) gave an even higher value of 0.0631, which is interest-ing in view of the palynological documentation of an exceptional climatic stability in this place (Marchant et al. 2007).
In the Udzungwa Scarp Forest Reserve the mature forest was much better for range-re-stricted birds than an adjacent area of strongly disturbed forest (Fjeldså 1999). The difference between mature and disturbed forest was less clear-cut elsewhere (Table 1). However, the high rarity score in the disturbed part of Mangalisa was mainly caused by an exceptional density of Andropadus (tephrolaemus) chlorigula, which is endemic only as a consequence of recent taxonomic splitting of what was once consid-ered a widespread Afromontane biospecies (see below). The high score in the disturbed parts of Mafwemiro was mainly due to very high densi-ties of species with moderately-small ranges (Batis crypta, Laniarius fuelleborni, Cossypha anomala and Andropadus spp.).
Observational evidence of dispersalTrue forest species (see habitat codes in Ap-pendix 2) were mainly observed inside forest. However, lack of observations of certain species in the highest parts of the terrain during the aus-
tral winter, and appearance of highland birds in semi-evergreen foothill forests or even ground-water forests in the lowlands is indirect evidence of local movements (Burgess & Mlingwa 2000, Romdal 2001b and 2003).
The most noteworthy highland residents, never recorded in lowlands, are Xenoperdix udzungwensis and obscurata, Laniarius fuelleborni, Arcanator orostruthus, Turdus olivaceus, Poeoptera kenricki, Sheppardia lowei, Cossypha anomala, Elminia albonotata, Andropadus chlorigula, Phyllastrephus placidus, Sceptomycter winifredae, Apalis thoracica and chapini, Artisornis metopias, Pseudoalcippe abyssinica, Nectarinia moreaui, fuelleborni and “afra“ (see below), Ploceus nicolli, Cryptospiza reichenovii, Serinus melanochrous and Linurgus olivaceus (and Caprimulgus poliocephalus, Lanius marwitzi, Onychognathus tenuirostris [down to 600 m in Kihansi], Dioptrornis fischeri, Cisticola lais, nigriloris and njombe, Apalis alticola and Bradypterus cinnamomeus outside forest habi-tat). There is one lowland record of Sheppardia aurantiithorax, a couple of such records for Modulatrix stictigula and one case in Onychognathus tenuirostris, but since these seem to be rare exceptions we accept also these species as genuine highland residents.
Highland forest birds found fairly regularly at forest edges and even in thickets some distance away from the closed forest are Francolinus squamatus, Stactolaema olivaceus, Laniarius fuelleborni, Cossypha anomala, Dioptrornis fischeri, Bradypterus spp., Apalis chariessa, Pseudoalcippe abyssinicus, Zosterops (senegalensis) stierlingi, Anthreptes neglectus, Nectarinia mediocris, Cryptospiza reichenowii, Serinus whytii and Linurgus olivaceus. During the exceptionally dry and warm austral summer of 2002/3 (when the “short rains” failed almost completely) even other forest birds were seen in woodland outside evergreen Udzungwa highland forest: Coracina caesia, Telephorus nigrifrons, Phyllastrephus placidus and flavostriatus, Apalis chariessa, Nectarinia rufipennis, Anthreptes pallidigaster and Ploceus bicolor. This suggests that even birds which normally appear to be sedentary and reluctant to leave the vegetation cover of the for-est may wander in periods with unusual weather,

j. fjeldså et al.12
maybe as a result of lack of food. Small numbers of juvenile birds, and distinctive growth-bars with lines of weakness (fret marks) on the feath-ers of captured juveniles, suggest that the food supply was insufficient in the 2002/3 season.
Remarks about individual taxaThe distributional record of birds in the studied forests is compiled in Appendix 2 and we com-
ment below on the most interesting findings, mainly of species of very isolated or disjunct distributions. In addition we describe cases where closely related forms replace each other sharply in different parts of the Eastern Arc, espe-cially where recent phylogeographic work with molecular markers suggests hitherto overlooked differentiation (Roy et al. 1998, Fjeldså et al. 2000, 2003 and 2006, Beresford 2003, Bowie 2003, Bowie et al. 2004ab, 2005, 2006, Bowie
Fig. 6. Examples of distribution patterns of forest birds in East Africa using distributional databases in a grid of 15 x 15 geo-graphical minutes implemented in WorldMap software; A the submontane Stactolaema leucotis, B the forest batis group, with the recently described montane Batis crypta and B. (capensis) dimorpha and B. mixta, C montane Arcanator orostruthus, D montane Sheppardia lowei, S. aurantiithorax and S. montana, E montane Cossypha anomala and F montane Nectarinia mediocris, N. usambarae, N. moreaui, N. loveridgei and N. fuelleborni.

13distribution of highland forest birds in tanzania
& Fjeldså 2005). The distribution maps in Fig. 6 are meant to illustrate the great variation in terms of patterns, connectivity and placement of range disjunctions.
PhasianidaGuttera pucherani (Crested Guinea-fowl) is widely but somewhat patchily distributed across Africa, in forest-woodland mosaics in lowlands as well as highlands. Birds in our study area (N = 5 collected) combine distinctive red and blue bare facial skin of subsp. pucherani of East Af-rica and the broad collar of black neck-feathers, which otherwise characterize subsp. barbata of south-eastern Africa and the Malawi Rift region. However, the precise distribution of this interme-diate form is unknown.
Xenoperdix udzungwensis and obscurata (Ud-zungwa and Rubeho Forest-partridges, spp. nov., Dinesen et al. 1994, Bowie & Fjeldså 2005) represent a relictual lineage, as they are the only African taxa in an ancient phasianid clade, which dates back to the Oligocene and is otherwise represented only in the Oriental biogeographic region (Crowe et al. 2006). The first species was discovered in 1991 in the Nyumbanitu and Ndundulu Mts W of the Great Ruaha Gorge, and its range appears to be very restricted within the Udzungwa highland. It is unrecorded further SW and from Image Forest, but could possibly exist in the unexplored Golo-Golo Mts. It was a big surprise when JK in March 2000 found another population in the N part of Mafwemiro Forest, 100 km further N than previously recorded. Mo-lecular data revealed that gene flow between the two forms has probably not taken place for ca. 200.000 years (Bowie & Fjeldså 2005). As the populations also differ in morphological features that may serve as social signals, it has been sug-gested to regard them as different species (Bowie & Fjeldså 2005). Also the Rubeho form is very localized, as scrapes and feathers have only been found in the northern part of the Mafwemiro For-est, despite thorough searching in other forest areas. The northern forest area is very steep, with the nearest village ca. 1000 elevational meters below the forest and therefore faces limited hunt-ing pressure. The Xenoperdix species seem to be associated with mature forest (see also Butynski & Ehart 2003), often with Podocarpus trees and
an open under-storey.Francolinus (Pternistes) squamatus (Scaly
Francolin) is patchily distributed in the forest zones of equatorial Africa. The Eastern Arc forms, subspp. usambarae in the Usambara Mts and uzungwensis (N = 34) in the Udzungwa and Southern Highlands, differ clearly from other populations as black feather-tips form a distinc-tive ornament on an otherwise white face (while other populations have a rather uniform drab-grey face with more or less pronounced scaly pattern). Authors of some recent handbooks, who synonymized all subspecies, have appar-ently overlooked how distinctive the Eastern Arc forms are, and preliminary genetic studies (un-published) suggest that they are not even closely related to other Scaly Francolins. However, a final comparative analysis and taxonomic conclusion must await inclusion of tissues of the Malawi Scaly Francolin (F. squamatus doni). We found Scaly Francolins to be common in many places in the Udzungwa highland and also recorded it in Mangalisa, Rubeho and Kiboriani Mts, which is a range extension. However, possibly due to high hunting pressure (snaring), francolins appear to be absent from many small and easily accessible forests (Appendix 2).
Francolinus (Pternistes) levaillantii (Red-winged Francolin) is patchily distributed in wood- and grasslands of southern Africa, includ-ing the Malawi Rift and Iringa highlands (nomi-nate ssp.), with isolated populations W of the Lake Victoria Basin and in the Kenya highlands to northern Tanzania (kikuyensis). An observa-tion right below the northern edge of Mafwemiro Forest (JK) suggests that it could be more widely present in central Tanzania.
GruiformesSarothura elegans (Buff-spotted Flufftail) is widespread but cryptic and with few published records from the Eastern Arc Mts. Our observa-tions (Appendix 2) suggest that it is present wher-ever there are depressions with muddy areas or grass swamps in and near the highland forests.
FalconiformesAccipiter melanoleucus (Greater Sparrowhawk) is widespread across Africa but sparsely recorded in Tanzania, mainly from the Southern and Ud-

j. fjeldså et al.14
zungwa Highlands, and the mountains in the NE. One was seen in Ukwiva forest and there is also one record from the Nguru Mts. Being shy and quiet, it is probably often overlooked during surveys inside the forest.
PiciformesStactolaema leucotis (White-eared Barbet, map Fig. 6A) is widespread in Mozambique and local in East Africa, mainly in submontane woodlands and semi-evergreen forest. Within our study area, subsp. leucogrammica is poorly documented: Mwanihana and Ukami forests in the Udzungwa area, Mahenge to the S and within Uapaca/Brachystegia woodland of the Man-galisa highland (to 2000 m, exceptionally high for this species). It is also recorded in lowland forest just E of the Uluguru Mts, and possibly in the Nguru Mts (Romdal 2001). Stactolaema olivacea (Green Barbet) has a very local (rel-ictual) distribution in coastal eastern Africa and is more continuous along the Eastern Arc Mts, where Clancey (1989) places all population from coastal Kenya to the Rubeho and Uluguru Mts in the nominate subsp., restricting subsp. howelli to the Udzungwa Mts and subsp. rungweensis to the Southern Highlands and Misuku Hills in N Ma-lawi. Based on recently collected material (N = 48) and molecular data, the montane populations from the Nguru Mts to the Southern Highlands appear to be specifically distinct from those of the Usambara Mts, with considerable genetic and geographical structure, but shared haplotypes across the Great Ruaha Gap (T. Muriel, unpub-lished MSc Thesis).
MusophagiformesTauraco persa (sensu latu, Green Turaco) is widely distributed in forest and woodland savan-nas in Africa, except where replaced by closely related forms. In southern Tanzania and the Eastern Arc N to the Nguu Mts it is generally represented by the form livingstonii (an inde-pendent species in some classifications; Dowsett & Dowsett-Lemaire 1993). However, birds seen in the Rubeho highland were characterised by a long floppy crest and bluish wings and tail, thus corresponding to the schalowi subspecies group (Fjeldså et al. 1997, and field observations in
2002). This latter form is widespread in Zam-besian woodlands E to the Tanganyika-Nyasa region, with an isolated population in the Crater Highlands of N Tanzania and adjacent SW Ken-ya. While only livingstonii has been recorded in the past in the Udzungwa Mts (many sound recordings from 1991 and 1992), the dorsal colouration (green to blue) and crest length seems variable and many individuals tape-recorded and collected in recent years (N = 20) in the Ndundulu and Ukami forests were of the schalowi type. This may suggest a recent population expansion of T. schalowi across the well-wooded hills of northern Tanzania. Typical livingstonii has been collected further W in the Udzungwa highland. No birds in the Kiboriani Mts were seen well enough for confident identification. A thorough analysis of morphological and molecular data is needed to determine the degree of displacement and secondary intergradation between these forms.
StigiformesBubo (poensis) vosseleri (Nduk Eagle Owl) was until recently regarded as endemic to the Usambara Mts (Moreau 1966, Britton 1980). It has now been found to be fairly common in the Uluguru Mts and to also be present elsewhere in the Eastern Arc, including in the Udzungwa Scarp Forest Reserve (Stanley et al. 2002), Nd-undulu Mts (AP) and Luhombero Mts (Butynski & Ehardt 2003). It was heard at both campsites in the Mangalisa forests (1999 & 2000) and in the southern Mafwemiro forest (2002). Local people in Mangalisa knew the species (and had eaten the large young from a nest in a tree that was felled). It seems to prefer tall, undisturbed forest.
CaprimulgiformesCaprimulgus ruwenzorii (Montane Nightjar) is disjunctly distributed in montane heathlands and forest edges of the Albertine Rift Mts, Angola Scarp, with the form (species?) guttifer known from two widely separate areas: the Southern Highlands and Usambara Mts. In recent years it has been collected in several sites in the inter-vening highlands (N = 14) and may possibly be generally distributed on high bracken-covered ridges along the Eastern Arc Mountains.

15distribution of highland forest birds in tanzania
ApodiformesSchoutedenapus myioptilus (Scarce Swift) is lo-cally distributed in the mountains of eastern Af-rica, with published records in the northern (Pare, Usambara) and southern (Udzungwa, Uluguru) Eastern Arc. We saw it over most forests in our study area, and we believe that it is present in all Eastern Arc highlands.
Passeriformes, CorvidaLanius (collaris) marwitzi (Uhehe Fiscal) is a range-restricted (semi) species of open highland areas with scattered trees, including village areas. Mainly known from the Southern and Udzungwa Highlands, and also recorded from Mpwapwa and the Ukaguru Mts (Harris & Franklin 2000). We saw it near Image, Mangalisa, Ukwiva and Mafwemiro forests and Kiboriani Mts. Rubeho birds seem to differ from those recorded in the Udzungwa highland as the white superciliary is supplemented with a more extensively white forehead.
Oriolus chlorocephala (Green-headed Oriole) is endemic to the East African coastal forests, S Malawi and the Eastern Arc, where it has been re-corded in the foothills and lower montane forest of the Usambaras, Nguu, Nguru and Uluguru’s and (commonly) in Mwanihana and Ukami for-ests in the Udzungwa’s. This latter population appears to be isolated, as only the species O. larvatus (African Black-headed Oriole) was seen in the best patch of submontane evergreen forest in the Rubeho foothills.
Campephaga quiscalina (Purple-throated Cuckoo-shrike) is distributed among the high-lands of central Africa to the Crater Highlands of N Tanzania, and very locally in the Eastern Arc (endemic subsp. munzneri): in submontane and lower montane forest in the Udzungwa’s, Mahenge highland S of the Kibashira Swamps, and in the Uluguru Mts, but it is unrecorded in the Rubeho highlands.
Coracina caesia (Grey Cuckoo-shrike) has a wide but patchy Afromontane distribution. It may inhabit all larger highlands in the Eastern Arc, although it is still unrecorded in the Ukagu-ru, Kiboriani, Nguu and Pare Mts and we saw it only in well-matured forest habitat in the largest forest tracts of our study area (Appendix 2).
Dicrurus ludwigii (Square-tailed Drongo) is widespread in forest and thick woodland across Africa, except where replaced by D. atripennis in the Guinea-Congolian rainforest. Collected specimens from the Udzungwa Mts and ground-water forest in the adjacent lowlands (N = 6) are morphologically close to D. atripennis, but until the genetic population structure has been analyzed in detail we prefer to refer all records to D. ludwigii, which is the only forest drongo in East Africa according to the literature. Typically a bird of the lower montane zone, it ascends into montane forest where there is a forest continuum upwards from the submontane zone (Svendsen & Hansen 1995, Romdal 2001a), namely in Ma-henge, Udzungwa, Uluguru, Nguru, Nguu and Usambara Mts. In spite of its noisy and conspicu-ous behaviour it was never recorded N of Iyungi in our study area, but curiously it was recorded in the Kiboriani Mts N of Mpwapwa.
Laniarius aethiopicus (Tropical Boubou) is widespread in savanna thickets and at forest edges across tropical Africa, and forms a species group, according to new molecular data (Nguem-bock et al. 2008). The Eastern Arc Mts may be a contact zone between the widespread form major (with white wing-stripe) and the form (species) sublacteus (with black wings) inhabiting coastal forests of Kenya and northern Tanzania, where it is found inland to Mpwapwa. In the Udzungwa-Rubeho study area, most birds, especially in riparian thickets outside forest) are referable to major, but sublacteus is observed at the edge of highland forest near Mafwemiro and in the Ndun-dulu area, suggesting some geographical overlap. No intermediate individuals were seen here.
Batis crypta (Dark Batis, sp. nov. Fjeldså et al. 2006) is a common bird of montane forests of much of the Eastern Arc, but has previously been considered part of Batis mixta (Forest or Short-tailed Batis) of coastal forests and some montane areas W to the Nguru Mts and volcanic highlands in northern Tanzania (Fig. 6B). The two may not even be sister taxa, as B. crypta is closest to B. (capensis) dimorpha of the Malawi Rift, and B. mixta clustered with B. diops of the Albertine Rift in the molecular phylogeny (Fjeldså et al. 2006). The genetic variation suggests strong genetic divergence between the populations W and E of

j. fjeldså et al.16
the Ukaguru-Nguru gap (crypta vs. mixta), but a complete mix of haplotypes between the Uk-aguru, Rubeho, Udzungwa and Southern High-lands, and similarly there is no morphological differentiation within this latter area.
MuscicapoideaMonticola angolensis (Miombo Rock-thrush) is widespread in the southern woodland sa-vanna zone to the Tanzanian Southern Highlands (Mbeya) with isolated records in Dodoma and in the Rubeho’s near Kilosa. Observations near Iringa, in woodland N of Image, at Mbatwa to the N of the Udzungwa N. P., and in the N part of the Mangalisa plateau as well as in Kiboriani Mts suggest that this species may be regularly present on wooded rocky slopes in the rain-shadow part of our study area.
Turdus olivaceus (Olive Thrush) is widespread in the highlands of southern and eastern Africa, with complex morphological variations. Recent phylogeographic work (Bowie 2003; Bowie et al. 2003, Voelker et al. 2007) demonstrates that two distinctive Eastern Arc forms (helleri in the Taita Hills and roehli in the Usambara and Pare Mts) represent distinctive relictual populations with a basal phylogenetic position compared to the populations inhabiting virtually every other highland of East Africa and Ethiopia (abyssinicus group). Since these populations have been able to maintain their genetic integrity they are probably best regarded as independent species. The form nyikae of the abyssinicus group, is widespread but of varying abundance in the montane forests of our study area (Appendix 2) extending N to Kiboriani and Nguru Mts.
Sheppardia ssp. (akalats) have a complex distribution pattern with some replacements in different highlands and elevational zones. S. sharpei (Sharpe’s Akalat), endemic to the Eastern Arc and Southern Highlands, has been recorded from (800) 1030 to 2160 m, probably reflecting some seasonal elevational movements. Our distributional records (Appendix 2) sug-gest that S. sharpei is most typical of the humid zone near the SE margins of the Udzungwa and Rubeho highlands. Birds in our study area are slightly different in colouration from those of the Nguru (ssp. usambarae) and Uluguru Mts. The
high-altitude “Dryocichloides” clade (Fig. 6D) is subdivided into three reciprocally monophyletic forms (Beresford 2003): S. lowei (Iringa Akalat) in the Southern and Udzungwa Highlands (and Uluguru Mts, one ZMUC specimen from 1962 but no records during recent surveys) is replaced in the Rubeho, Wota, Ukaguru and Kiboriani highlands by S. aurantiithorax (sp. nov.; Beres-ford et al. 2004), and with S. montana (Usam-bara Akalat) isolated in the West Usambara Mts. These forms are generally restricted to montane plateau forests at 1660–2400 m, but occasional records are known from the cold season down to 1400 m, and one aurantiithorax specimen from lowland forest just NE of the Uluguru Mts sug-gests casual straggling.
Cossypha anomala (Olive-flanked Robin-chat, Fig. 6E) is distributed in stunted forest on ridges, near streams and edges in montane forest in the southern Eastern Arc from Udzungwa to the Wota, Kiboriani and Ukaguru Mts, and in the Nguru and Uluguru Mts (ssp., grotei) and isolated in the Mbulu highland in N Tanzania (the melanic form mbuluensis); these forms are replaced by the very different-looking sspp. anomala and macclounii along the Malawi Rift.
Swynnertonia swynnertoni (Swynnerton’s Robin) has a disjunct (relictual?) distribution, along the Zimbabwe/Mozambique border (nomi-nate ssp.), in the East Usambara Mts and in the Udzungwa Mts (rodgersi), mainly in mature humid montane forest (Fjeldså 1999, Dinesen et al. 1993 and 2001). Known only from 1000–1720 m in the Udzungwa’s, and common only in a few places, it may be genuinely absent from the Rubeho highlands.
Myrmecocichla aethiops (Northern Ant Chat) is distributed along Africa’s northern Savannah belt and isolated in the Kenyan highlands and Crater highlands of N Tanzania. An observation on the 17 Nov. 2002 in wooded rocky slopes right above Mpwapwa town, S Kiboriani Mts, suggests that its range may extend S along the rocky ridges that dot central northern Tanzania (see Figs 1 and 2).
Melaenornis fischeri (White-eyed Slaty Fly-catcher) is fairly widespread along the “montane circle” of eastern Africa, with an interesting population structure: birds of the Southern High-

17distribution of highland forest birds in tanzania
lands, Udzungwa, Rubeho, Kiboriani (Appendix 2), Ukaguru and Uluguru Mts (subsp. nyikensis) are morphologically similar to those of the Crater highlands of N Tanzania but distinct from those E of the Rift (Mt Meru, Kilimanjaro and N Pare Mts) and Kenyan highlands (subsp. fischeri). This suggests a faunal connection via the rocky ridges and scattered inselbergs of central north-ern Tanzania.
Poeoptera kenricki (Kenrick’s Starling) is rare and very local in the Kenyan highlands and Eastern Arc (N Pare, Usambaras, Nguu, Nguru, Uluguru, Udzungwa and Iringa highlands to Mdandu Forest, Njombe). It was unrecorded in most of our study sites, but a small breeding group was located in Dec. 2002 at the S end of Mafwemiro forest.
SylvoideaPsalidoprocne albiceps (White-headed Saw-wing) is distributed across Africa’s southern savannah region and central highlands, includ-ing the Udzungwa highland, Crater highlands of N Tanzania and Ukaguru Mts. We found it also in Mangalisa and the Rubeho highlands (with confirmed breeding near the edge of Mafwemiro forest). Being migratory, it is present here only during the rainy season.
Andropadus tephrolaemus (Mountain Green-bul; Arizelocichla following Johansson et al. 2007) has been found to represents a paraphyletic complex (Roy et al. 1998). The form chlorigula represents (along with the morphologically rath-er divergent neumanni of the Uluguru Mts and fusciceps, which replaces it sharply in the South-ern Highland, in an area with no clearcut barrier) represents a distinct phylogenetic branch from the populations of northern Tanzania (nigriceps in the Crater highlands and usambarae in South Pare and Usambara Mts). The form chlorigula is common, with no sign of morphological differ-entiation in all high-altitude forests of our study area, even within the isolated Wota and Ligunga forests. A. olivaceiceps (Stripe-cheeked Green-bul; species rank following Sibley & Monroe 1990), a widespread montane species along the Malawi Rift (subsp. olivaceiceps) and in the East-ern Arc Mts (subsp. striifacies), seems to be rare at high elevations and absent from some of the
high plateau forests in our study area (Appendix 2). Andropadus virens (Little Greenbul; Eurillas according to Johansson et al. 2007) is widespread in evergreen forest and thickets across tropical Africa, where it is mainly found at lower eleva-tions, but ascends to montane forest on slopes with a forest continuum (Appendix 2).
Phyllastrephus flavostriatus (Yellow-streaked Greenbul) is disjunctly distributed in eastern Africa, from sea level to montane forest, and molecular data (S. Lokkugalapatti et al. unpub. data) suggest that different populations represent distinctive lineages that may be best treated as different species. In the Eastern Arc Mts, the distinctive form uzungwensis is generally com-mon throughout the Udzungwa Mts, whereas the form tenuirostris inhabits coastal forests and the humid slopes of the Usambara’s and S Pare Mts as well as extending inland to the Uluguru and Nguru Mts. Between the Nguru and Udzungwa Mts a few individuals (of unknown subspecies) were seen in a groundwater forest in the foothills zone E of the Rubeho Mts.
Cisticola nigriloris (Black-lored Cisticola) is endemic to the highlands of the Rukwa and north-ern Malawi Rifts and adjacent part of the Eastern Arc. Records in bushy re-growth, marshes and forest edges in Image, Mangalisa, Uvidunda and the Ukwiva and Mafwemiro areas represent a range extension to the NE. C. njombe (Churring Cisticola) is endemic to the highlands along the northern Malawi Rift and within the Udzungwa highlands; our records from Image, Msanga, Mafwemiro, Wota and Ligunga represent range extensions. C. lais (Wailing Cisticola) is patchily distributed in southern Africa, the Malawi form semifasciatus extending E to the Udzungwa highland, including tall grass near the northern end of Image forest. We did not record it further N.
Scepomycter winifredae (Winifred’s Warbler) is a relictual Eastern Arc endemic with no near relatives among the cisticolid warblers (Nguem-bock et al. 2006). Associated with high-rainfall areas with tall herbaceous vegetation, mainly in disturbed forest and landslide gaps, it is fairly common in the Uluguru Mts at 1300–2430 m (Svendsen & Hansen 1995). In the Udzungwa highland one duetting pair has been recorded

j. fjeldså et al.18
at 1000 m above Sanje village in the extreme E (Mwanihana, Jensen & Brøgger-Jensen 1992 and pers. comm.), but it was unrecorded by recent visitors to the area (D. Moyer, E. Mulungu and T. Romdal) and information in the literature that it is common here may have no factual basis. Although suitable (but inaccessible) habitat may exist on the steep N face of the Golo-Golo Mts to-wards the Kidatu dam, it seems that the Udzung-wa record may represent a marginal occurrence, or maybe a vanishing population. Scepomycter warblers were found very locally within the Uk-wiva forest at 1800–1900 m (Fjeldså et al. 1998; not found during surveys in 2001, but collected at another site in 2002) and in the highest massif in the NW of the Ukaguru Mts at 1500 and 1880 m (observations by Friedman & Stager 1964 and Evans & Andersen 1992, 1993; specimens collected in year 2000). The Rubeho-Ukaguru form has now been described as an independent species, S. rubehoensis (Bowie, Fjeldså & Kiure 2009).
Apalis thoracica (Bar-throated Apalis) inhab-its montane forests from SE Africa N to northern Tanzania, with several local forms in the northern part of the range being morphologically distinc-tive but genetically closely related (L. Herrmann pers. com.). In the Eastern Arc a “leapfrog” pat-tern is found as the widespread green-backed/yellow-bellied form griseiceps (of Udzungwa to Ukaguru Mts is replaced by the grey-backed/white-bellied form pareensis in the S Pare Mts and an intermediate form (polytopic “murina“) occurs in the Usambara and Nguru Mts and then again along the Malawi Rift.
Apalis chariessa (White-winged Apalis) has a relictual distribution: southern Malawi Rift, Eastern Arc (Udzungwa Mts at 1000–2000 m and foothills of NE Uluguru Mts) and lower Tana River, Kenya (where it is now apparently extinct). In the Udzungwa highland it seems gen-erally to occur in low densities, but on the other hand it seems fairly tolerant to forest disturbance (Fjeldså 1999). It is unrecorded in the highlands N of the Great Ruaha River. Apalis melanocephala (Black-headed Apalis) is somewhat patchily distributed in lowland and highland forests of eastern and south-eastern Africa, with considerable subspecific differentiation. Most of the Eastern Arc Mts are inhabited by the small
and grayish form moschi, but the Udzungwa and Rubeho highlands are inhabited by large birds with black upper-parts, like those of south-western Tanzania along the Malawi Rift. Apalis chapini (Chapin’s Apalis) is endemic to the Ma-lawi Rift Mts and Eastern Arc Mts N to the Nguru Mts. It may inhabit all larger patches of evergreen montane forest in this range, but was unrecorded in Mangalisa, Wota Mts and Msanga forest. The extent of chestnut colouration on forehead and crown varies in the Eastern Arc form chapini, and birds observed in Ukwiva and Mafwemiro forests had the entire crown chestnut. A. alticola (Brown-headed Apalis) is patchily distributed in the highlands of Africa’s southern woodland savannahs, mainly in small thickets and at forest edges, but inside forest in places where A. thoracica is lacking. In the Eastern Arc Mts it is only known from the Udzungwa highland, but an iso-lated population is found in Tanzania’s northern Crater highlands and adjacent SW Kenya.
Bradypterus cinnamomeus (Cinnamon Brack-en-warbler) is disjunctly distributed in highlands of eastern Africa, and may comprise cryptic spe-cies (C. Kahindo pers. com.). In the Eastern Arc Mts it is found at edges and along bushy ridges in the montane forest, from the Southern Highlands and Udzungwa Mts to Mangalisa and Ukwiva and in the Uluguru Mts, and then after a wide disjunction in the Usambara and Pare Mts.
Artisornis metopias (Orthotomus in some classifications, African Tailorbird) is a relict Eastern Arc forest bird, related to Schistolais leucopogon and leontica of the Guinea-Congolian rainforest (Nguembock et al. 2006). In our study area it is common in light-gaps in most forests but unrecorded in Mwanihana and Msanga for-ests. Preliminary genetic studies suggest lack of lineage-sorting across the Great Ruaha River but a marked genetic change between the Nguru and Usambara Mts (R. Bowie in litt.).
Phylloscopus ruficapillus (Yellow-throated Woodland Warbler) is patchily distributed, main-ly in montane forests, in southeastern Africa N to the Eastern Arc (subsp. minulus) and Mahale Mts (ochrogularis, species?) at Lake Tanganyika. In our study area it may be somewhat patchily distributed (Appendix 2).
Illadopsis rufipennis (Pale-breasted Illadop-sis) is a disjunctly distributed Afrotropical spe-

19distribution of highland forest birds in tanzania
cies, with the isolated East African form (distans) very locally distributed in some quite different forest types: certain coastal forests, Usambara Mt foothills (to 1200 m), Nguru foothills (but not Nguu), groundwater forest below the Ulu-guru and Rubeho Mts, but also (with confirmed breeding) at 2000 m in Mafwemiro and Kiboriani Mts (altitudinal record) and in the Udzungwa highland. There is some morphological variation and marked genetic differences between some of these populations (R. Bowie in litt.), which will be defined taxonomically in a separate pub-lication. Highland populations in the Rubeho and Udzungwa Mts seem to represent the same taxon.
PasseroideaThe babbler-like species Modulatrix stictigula (Spotthroat) and Arcanator orostruthus (Dap-plebreast, Fig. 6C) have had a tumultuous taxo-nomic history, but molecular data now suggest that these Eastern Arc near-endemics, together with Kakamega poliothorax (of the Albertine Rift and Cameroon highlands) and the “cape sugarbirds” Promerops, represent basal (rel-ict?) lineages in the Passeroidea (Beresford et al. 2005, Johansson et al. 2008). Modulatrix is present in the majority of Eastern Arc montane forests from N of the Malawi Rift to Nguru and Kanga, and in the Usambaras. The Usambara and Uluguru populations are genetically very diver-gent, but there is only limited genetic structure across the Great Ruaha gap (R. Bowie in litt.). We know of no record outside forest and only very few records below its typical altitudinal range at 1400–2100 m (one specimen from 600 m, near the Uluguru’s, in ZMUC, and once heard at 550 m). There are some interesting distribution gaps: while it was common in the Udzungwa highland and in Mafwemiro forest, few were heard in Ukwiva and only a single alarm note was heard (even with some doubt) in Iyungi. It is unre-corded in Mangalisa and Wota Mts, suggesting restriction to high-rainfall areas. Arcanator is distributed in the Udzungwa highlands (subsp. sanjei, Jensen & Stuart 1982, Dinesen et al. 1993 and 2001, Fjeldså 1999) and is otherwise known only from Mt. Namuli in northern Mozambique (nominate subsp.) and the East Usambara Mts (subsp. amani, which may now be extremely lo-
cal; N. Cordeiro pers. comm.). Within the present study area it is recorded only in the Udzungwa highland, and mainly in the most humid parts at 1300–1800 m, and common only in rather few places.
Macronyx fuellebornii (Fülleborn’s Long-claw) inhabits montane grassland in Angola/Zambia E to the Iringa highland (nominate subsp.) and Ufipa plateau (subsp. ascensi). It was un-recorded in the Rubeho highlands but may be expected to occur here since we paid little atten-tion to open habitats, and since there are speci-men records from the Mbulu Highlands much further to the N (Britton 1980, identity confirmed by N. Cordeiro fide N. Baker in litt.).
Nectarinia rufipennis (Rufous-winged Sun-bird) is a very distinctive form, of isolated systematic position among African sunbirds (Bowie 2003, contrary to the view expressed by Butynski & Ehardt 2003). It is endemic to the NE Udzungwa highland, occurring in mature forest in the lower montane zone, at (600) 1080–1600 (1800) m, S to the middle part of the Udzungwa Scarp Forest Reserve (Jensen & Brøgger-Jensen 1992, Dinesen et al. 1993 and 2001, Fjeldså 1999). It was seen in miombo woodland in the dry rainy season of 2002/3, but is unrecorded in the highlands N of the Great Ruaha River.
Nectarinia mediocris (Eastern Double-col-lared Sunbird), N. moreaui (Moreau’s Sunbird) and N. loveridgei (Loveridge’s Sunbird) divide between them the Kenyan highlands, Crater highlands of N Tanzania, the Eastern Arc Mts and highlands along the Malawi Rift. The range of N. moreaui extends from the eastern part of the Udzungwa highland to the Nguru Mts. Thus, as illustrated in Fig. 6F, it inhabits the area between populations hitherto referred to N. mediocris: ssp. fuelleborni in to the SW and sspp. usambarica and mediocris in the N. Molecular data (Bowie et al. 2004a) now demonstrate that N. mediocris is paraphyletic and should be divided into three species, and that moreaui is sister to loveridgei in the Uluguru Mts. Within moreaui, the Nguru population is somewhat divergent but there is no lineage-sorting across the Great Ruaha gap. However, based on data presently available there is a complete break in gene flow towards fuelleborni within the Udzungwa highland. The earlier suggestion that moreaui could be a hybrid

j. fjeldså et al.20
form (Stuart & van der Willigen 1980, Butynski & Ehardt 2003) can therefore be rejected. The two forms share the highland between them, inhabiting every forest patch within their respec-tive areas, with parapatric contact in the Ukami area, and we must therefore regard them as being incompatible and representing different (para)species.
The Nectarinia afra superspecies (Great-er Double-collared Sunbirds) is distributed in southern Africa with isolated forms in some high-lands N to the Albertine Rift, but with no previous records in the Eastern Arc. We found birds of this group at eastern (drier) forest edges of Udzungwa (Kisinga Rugaro), Image, Wota, Mangalisa and Rubeho Mts. They were seen in evergreen thick-ets in the farmland and at the forest edge and may be particularly common in glades with tall bushy and herbaceous vegetation, where they compete with N. moreaui for Tecomaria capensis flowers. Specimens (N = 11) resemble N. (ludovicensis) whytei, a local endemic of the Nyika plateau in NW Malawi, but differ in some plumage charac-ters. This new form will be formally described elsewhere.
Nectarinia olivacea (Olive Sunbird) is wide-spread in evergreen thickets and forests across Africa, from lowland to highland. We comment on this familiar bird to emphasize the fact that recent molecular studies (samples from across Africa) suggests that ancient populations in the Eastern Arc Mts (two haplotype networks with some gene-flow between them) gave rise to other populations and a rapid population expansion across the continent (Bowie et al. 2004b).
Nectarinia verticalis (Green-headed Sunbird) is widely distributed, mainly in semi-evergreen thickets along the periphery of the Guinea-Congolian rain forest to Zambia, Rukwa and northern Malawi Rifts and the Iringa highland, mainly in lowlands but restricted to highlands in the east. Seen at forest edges and disturbed forest in Kisinga-Rugaro and Image forests, and quite common in disturbed montane forest in the Rubeho highland, which represents a slight range extension and an unusual habitat for the species. Also recorded in montane forest in Kondoa, Mbulu and Marang, suggesting that this popula-tion extends further N along the mountain ridges of central northern Tanzania.
Nectarinia kilimensis (Bronze Sunbird) is fairly widespread in the highlands of East Africa (and Angola Scarp), including the SW part of the Eastern Arc to adjacent eastern Zambia (subsp. arturi). This latter form is morphologically vari-able as some specimens have fairly strong cop-pery or rosy reflections, especially on the rump and lower back, wing-coverts and breast. The single male collected at the edge of Mafwemiro forest was extreme in this respect, approach-ing N. tacazze (Tacazze Sunbird) of the Kenya highlands and Kilimanjaro by appearance, but analysis of mtDNA data placed it with kilimensis (R. Bowie in litt.). More data are needed to work out the historical population structure and rela-tionships between these forms.
Anthreptes pallidigaster (Amani Sunbird) is disjunctly distributed in coastal forests (Arabuko-Sokoke forest in SE Kenya and the East Usam-bara Mts) and apparently isolated in Nyumbanitu and Ndundulu Mts of the Udzungwa highlands (birds with more violet iridescence, observed on rare occasions at 1030–1550 m; Dinesen et al. 1993 and 2001). In the unusual 2002/3 season it was seen in miombo woodland outside the forest in the Ndundulu area (but also inside Ukami and Nyumbanitu forests). This canopy sunbird is still unrecorded in the highlands N of the Great Ruaha River, but could easily have been overlooked.
Anthreptes rubritorques (Banded Green Sun-bird) is uncommon in lower montane forest (300–1600 m), with records in the Usambara Mts, Nguu, Nguru, NE Uluguru’s and eastern Udzungwa highland. However, recent records have been made only from the Usambara and Nguu Mts (Seddon et al. 1999) and the eastern Udzungwa highland (fairly common in Mwani-hana, T. Romdal, pers. com.; rare in Nyumbanitu and Ndundulu). The species tolerates degraded forest (Seddon et al. 1999) but the fragmenta-tion and loss of submontane forest in most parts of the Eastern Arc may nevertheless have led to local extirpation. There are no records between the Udzungwa and Nguru Mts.
Ploceus weavers: Among the forest-inhabiting nuthatch-weavers, Ploceus bicolor (Dark-backed Weaver) is widespread in well-wooded parts of eastern Africa but quite local in the montane regions. In the Eastern Arc (subsp. kersteni) it is fairly common in the Udzungwa Mts but then has

21distribution of highland forest birds in tanzania
a large distribution gap N to the Kiboriani (where recorded at 1750 m), Nguru, Nguu, Usambara and Pare Mts. It is also absent from the Crater highlands of N Tanzania and Kenya. A single pair was found in Iyungi, but apparently the spe-cies is genuinely absent from Image, Mangalisa, Rubeho and Wota Mts. P. nicolli (Tanzanian Weaver) is a rare and local inhabitant in the most humid parts of the Udzungwa and Uluguru Mts (subsp. anderseni) and Usambaras (subsp. nicolli), but unrecorded in the intervening moun-tains (and in the Uluguru’s in recent years). This species requires undisturbed, mossy forest in the lower montane zone, and its distribution seems relictual. An unidentified nuthatch weaver, which differs from nicolli by lacking dark brown/black pigments except on the face, has been observed in Ukwiva (once; Fjeldså et al. 1997) and Mangali-sa (twice in Sep. 2001, JK). This could represent a non-melanic (Harrison 1965) representative of P. nicolli, or an unknown species.
Serinus whytii (Yellow-browed Seedeater) is a bird of bushy vegetation near edges and along bushy ridges in the montane forest from N of the Malawi Rift to the Udzungwa highland. Seen at Madizini in the eastern foothills of the Rubeho highland (Fjeldså et al. 1998) and was common near Nyiumbanito and Mafwemiro forests. S. melanochrous (Kipengere Seedeater) inhabits grassy ridges within the montane forest, from Njombe to the Udzungwa highland, except the most rain-drenched eastern slopes, and it was seen also in Iyungi and Mangalisa forests, and in foothills near Ifakara. Because of its secluded life in the forest understorey we will not exclude that it may be more widespread N of the Great Ruaha gap.
Geographical replacementsNinety-two per cent of all species occurring above 1500 m in our study area (excluding casual visitors of water-birds, raptors etc. for which our data would be inadequate) are recorded on both sides of the Great Ruaha River. Within this area, from the Southern Highlands to the Nguru Mts (excluding the outlying Uluguru Mts) there are five species replacements and ten subspecies replacements, corresponding to four on aver-age, for each of the four breaks between main
mountain blocks. The two species replacements between the Udzungwa and Rubeho highlands, and a possible morphological change in Apalis chapini, is hardly different than what can be expected if replacements were distributed at ran-dom. With such small numbers we did not find it meaningful to statistically test the randomness of replacements.
For a closer analysis we instead sorted the species by elevational zones and considered also the abundance assessments given in Appendix 2, assuming as a rough guiding principle that birds are common (A and C in the Appendix) in source habitats and rare (+ in the Appendix) in sinks (Brown 1984, Pulliam & Danielsen 1991, Romdal 2003). As appears from Table 2, almost all the montane forest birds are recorded as be-ing common, at least locally, to both sides of the Great Ruaha Gap, while many of those in the lower montane zone (1000–1700 m) are uncom-mon in the Rubeho’s, and some of them may be genuinely absent. The same applies to those with wide altitudinal ranges, many of them seasonal migrants which sometimes seek refuge in low-land forests, and species which typically inhabit submontane forests but ascend to the montane forest where there is a fairly continuous forest gradient. The difference in A+C, + and “absence” records in Table 2 is not significant for montane forest birds (χ² 0.221, p>0.50 for the two upper lines in the table) but highly significant for birds that use foothill forest at least in the dry season (χ² 16.9, p<0.001 for the three lower lines in the table).
In order to see the number of replacements across the Great Ruaha Gap in a broader geo-graphical perspective, Table 3 reviews the num-ber of shared species and replacements (different para- or allo-species as well as subspecies, but omitting some clinal and poorly resolved cases) along the entire chain of Eastern Arc Mts in Tanzania. It follows from the difference between figures for shared and vicariant taxa that only a moderate proportion of the represented lineages show marked geographical variation. On the other hand, every break in the mountain chain (excepting the narrow and densely wooded val-ley between the Rubeho and Ukaguru Mts) has at least one taxon replacement, and the Great Ruaha gap does not stand out in this respect. The

j. fjeldså et al.22
most prominent variation in this table relates to the many range disjunctions between the Rubeho and Usambara Mts, which in some cases even extends from the Udzungwa to the Usambara/Pare Mts (350 km), and to different allospecies replacing each other across this broad hiatus.
DISCUSSION
Community parameters and an interpreta-tion of the exceptional abundance of range-restricted species near the Great Ruaha GapThe fact that we recorded every forest-dependent bird species that has previously been found in the area (except Udzungwa populations of Campephaga quiscalina and Scepomycter winifredae) asserts that our record of the resident forest avifauna must be fairly complete. This is also supported by the close fit between observed and estimated species numbers (Table 1). The record of non-forest and non-resident species may be less complete. Overall, the smallest forests, and those on the transition towards the arid inlands, have fewest species. Other differences in species richness could be attributed to the great habitat heterogeneity caused by past or current logging and clearing, and by the presence of azonal habi-tats such as forest swamps and rocky areas. It is therefore more interesting to discuss the variation in numbers of “ancient” species, and of rarity scores (Table 1).
Ancient species, which may represent phy-logenetic and in some cases also biogeographic relicts, are best represented in the eastern parts of the Udzungwa highland (eight species, or seven if we disregard the single historical record of Scepomycter, versus four in the Rubeho highland). This difference is reinforced if we consider other taxonomic groups (Burgess et al. 2006, Rovero et al. 2008), but then much less collecting has been done in the Rubeho highland (Doggart et al. 2006).
The highest rarity values were those for Im-age and Mangalisa forests, which flank the Great Ruaha Basin on opposite sides (but with a small sample suggesting an even higher value in Kis-inga Rugaro forest, Udzungwa). These forests lack some of the Udzungwa specialties, and es-pecially the endemics of the humid submontane zone, but have very high population densities of some truly montane endemics (notably Batis crypta, Cossypha anomala, Andropadus chlorigula, Nectarinia moreaui). The sample site in the Udzungwa Scarp Forest Reserve (outside Fig. 3, to the SW) scores considerably less (Table 1; and see Fjeldså 1999), but this site is W of the assumed centre of endemism in the eastern Udzungwa highland. Based on our general im-pression and the mist-netting record we think that Nyumbanitu and Mwanihana forests would score as high as Image and Mangalisa. Iyungi scored moderately high, even though this forest, just N of the eastern Udzungwa scarp, is tiny and very disturbed. The rarity values for Ukwiva and Mafwemiro are lower but yet very high compared
Table 2. Abundance data for forest birds of the Udzungwa and Rubeho highlands as recorded above 1500 m (Appendix 2). A+C means Abundant or Common, + means present but in small numbers, in many cases just a single observation. Overall, the + and “absent” categories are assumed to represent sinks. Species, which always have low densities, mainly larger birds, are not included.
Udzungwa highland Rubeho highlandAbundance A+C + A+C + absent
Spp. of upper montane forest 9 1 8 2 -Upper and lower montane 15 1 15 1 -Primarily submontane 5 7 - 8 4Lower montane 9 8 4 7 6Wide elevational range 19 6 12 13 -

23distribution of highland forest birds in tanzania
with adjacent lowlands or montane forests out-side the Arc (JF unpublished data).
This high abundance of range-restricted spe-cies in the highlands just adjacent to the Great Ruaha Basin represents a noteworthy deviation from a widely accepted “rule” linking local and global abundance: species that are generally abundant at sites where they occur are also as-sumed to occupy the highest number of sites (e.g., Fuller 1982, Brown 1984 and 1985, Ford 1990, Gaston & Lawton 1990, Lawton 1996) and have the largest geographical ranges (e.g., Ford 1990, Gaston & Blackburn 1996). This “rule”,
implicit in non-deterministic (“neutral”) com-munity models (Hubbell 2001), means that local endemics should generally be rare, but this is not the case in our study area.
High abundance of range-restricted species would be theoretically possible within a non-deterministic framework if we assume low community drift (local species turnover driven by community interactions) in an ecologically stable and semi-isolated environment (Chapter 7 in Hubbell 2001). Johnson (1998) demonstrated a high abundance of range-restricted ancient spe-cies of Australian marsupials, and explained this
Table 3. The number of shared species of highland forest birds between the Eastern Arc highlands, and the number of cases of population differentiation to the right (upper figure representing subspecies as recognized in the current taxonomy, lower figure the well marked allo- or paraspecies, defined from morphology and in some cases from >5% sequence divergence of mtDNA; R. Bowie in litt.).

j. fjeldså et al.24
from past extinctions of the rare range-restricted species. We think that a strong degree of adapta-tion to the local environment represents a more plausible – and simpler – explanation. We would predict this to be the case for species which had a long time to adapt to specific local conditions (as suggested in the Time-Stability Hypothesis of Connell & Orians 1964). A stable environment would also imply co-adaptations in the commu-nity and incompatibility of closely related forms that inhabit adjacent geographical territories (García-Moreno & Fjeldså 2001). Local aggre-gates of endemic species may thus be related to predictable local environments (Fjeldså et al. 1999) in montane regions (Jetz et al. 2004), and under these conditions deterministic processes may override non-deterministic processes in ef-fect. It is worth noting that a recent palynological drill-core from Kisinga Rugaro forest now pro-vides direct evidence of an exceptional climatic stability in this area (Marchant et al. 2008).
This interpretation is also backed up now with the results of molecular data (especially Bowie 2003 and Bowie et al. 2004a and b, 2005, 2006, Bowie & Fjeldså 2005, Fjeldså et al. 2006) suggesting an ancient rooting for many Eastern Arc populations. Genetic mismatch profiles sug-gest that coalescence was followed by steady population growth, unlike bird populations of for example many of the Kenya and Albertine Rift highlands, which are genetically heterogeneous, suggesting considerable changes in the connec-tivity between populations (cit. op.). Accumula-tion of genetic diversity and beneficial mutations can probably start at population sizes of a few hundred individuals, balancing the effects of del-eterious mutations (Franklin & Frankham 1998, Whitlock et al. 2003). Presumably, many Eastern Arc endemics have had a long time to adapt to prevailing local conditions.
The tendency for reduced rarity scores in dis-turbed forest (Table 1) is a commonly observed pattern that may reflect “mass effect” of colonisa-tion by dispersive and widespread species with a broad habitat tolerance, as the increased species turnover may cause a complex competitive situa-tion which will represent “sinks” for the special-ized endemic species (see Brown 1984).
Few species replacements for highland birds
One of the aims of this study was to evaluate the role of the Great Ruaha Valley as a barrier, which could potentially cause speciation. Such a role would mainly be expected to affect resident highland birds, which are not known to venture to low elevations. It is more difficult to imagine a barrier effect for species with wide elevational distributions or those inhabiting patches of ever-green thickets or ground-water forest along the entire eastern foothill zone (Figs 3 and 5).
Our observations do not correspond to these expectations and over all it seems that only the large lowland area separating the northern (Pare, Usambara) and central/southern Eastern Arc Mts represent a significant barrier (Table 3). We found only two cases of species replacements of high-land forest across the Great Ruaha gap, namely Sheppardia lowei / aurantiithorax (Fig. 6D) and Xenoperdix udzungwensis / obscurata, the latter representing an alcient relictual lineage. Some subtle morphological differentiation may exist between local populations S and N of the gap for Apalis chapini, but other highland species seem to be undifferentiated, with little or no evidence of genetic lineage-sorting across the Great Ruaha gap (see accounts for Stactolaema olivacea, Batis crypta, Modulatrix stictigula, Andropadus chlorigula, Artisornis metopias and Nectarinia moreaui). The isolated population of Cossypha anomala (Fig. 6E) in the Volcanic highlands of northern Tanzania and of Phylloscopus umbrovirens in the Uluguru Mts, and the appearance of Tauraco (persa) schalowi in our study area, suggest that some “mountain-hopping” may take place across the interior of northern Tanzania. It is noteworthy that the replacement of Nectarinia (mediocris) fuelleborni and moreaui (Fig. 6F) is located within the Udzungwa highland, in an area with no obvious present-day barrier other than some grass- and woodland separating the remain-ing forest patches, and the same can be said of the replacement of Andropadus chlorigula and fusciceps within the Southern Highlands.
With only two cases of sister taxa (of Xenoperdix and Sheppardia) replacing each other across the Great Ruaha gap, it is difficult to give vi-cariance a significant role in explaining the high

25distribution of highland forest birds in tanzania
number of endemic species in our study area. In mammals, reptiles and amphibians, which have at least 125 endemic and near-endemic species in the Eastern Arc, there are as yet only two cases of vicariance between the Udzungwa and Rubeho highlands (in Rhampholeon dwarf chameleons and Nectophrynoides frogs; Burgess et al. 2006). The Great Ruana River must have separated the two highlands since the start of the tectonic uplift in the Miocene, and this certainly pre-dates the estimated age of divergence of the two avian species pairs (Beresford et al. 2004; Bowie & Fjeldså 2005). The presence of S. aurantiithorax in Wota and Kiboriani Mts, and in a foothill forest near the Uluguru Mts, and of S. lowei in the Uluguru Mts, demonstrates that these birds do sometimes move from one highland to another. Overall, the few cases of sharp species replacements along the Eastern Arc (disregard-ing replacements across the large gap between Nguru and Usaambara Mts) could be secondary phenomena reflecting that these populations are genetically or ecologically incompatible, and each of them therefore unable to settle in the area already occupied by the other. In this light, Tauraco livingstonii/schalowi may provide an interesting case for future studies of secondary contact. The fact that most montane forest birds are distributed across several highlands, and that several congeneric species are broadly sympat-ric, supports the general idea of the Eastern Arc as a “dispersal center”, where speciation is followed by population growth and range expansion (Jetz et al. 2004; Fjeldså & Bowie 2008).
Although our data provide little documenta-tion for non-forest highland birds, they seem to follow a similar pattern with presence on both sides of the Great Ruaha gap. Cisticola lais and the range-restricted Anthus brachyura, Macronyx fuelleborni and Euplectes psammochromicus are unrecorded N of the gap. They may be genuinely absent, as the habitat composition of the high plateaus of the Rubehos suggests that this area may have been more or less completely wooded in the past, unlike the Udzungwa highland, which has large tracts of natural montane grassland and rocky areas. The fact that M. fuellebornii, and the edge-inhabiting Apalis alticola, have been found in the Crater highlands of northern Tanzania, sug-
gest that they may have existed in the Rubeho, Wota and Kiboriani Mts, and they may also have been overlooked in view of how little attention we paid to non-forest habitats.
Disjunct species distributions at lower elevationIn sharp contrast to the situation described for the typical highland birds, those of lower eleva-tions follow another pattern. Many of them be-come rare N of the Great Ruaha Gap, and some are absent here, although some of them pop up again much further N. Such relictual patterns are described above for Arcanator orostruthus (Fig. 6C), Swynnertonia swynnertoni, Anthreptes pallidigaster and Apalis chariessa, with less marked cases for Oriolus chlorocephala, Dicrurus ludwigii, Ploceus bicolor, Anthreptes rubritorques and Ploceus nicolli.
Oriolus chlorocephalus, Dicrurus ludwigii, Andropadus virens, Phyllastrephus flavostriatus and Ploceus bicolor are most abundant in foot-hills and submontane forest. However, they have also been recorded in highland forests, but only on slopes with a forest continuum. Presumably the foothill forest represents prime habitat and the highland forests sink habitat, occupied only where there are opportunities for colonization from below (Romdal 2001a). In this context it is interesting to note that Pogoniulus bilineatus and Dicrurus ludwigii were common, and Ploceus bicolor also present, in the small Iyungi forest (Uvidunda Mts), but they were unrecorded in the Rubeho area. In Iyungi we even found one small group of Procolobus gordonorum, an en-demic primate of the Udzungwa and Southern Highlands. This may represent “spill-over” from Mwanihana Forest just 6 km to the S. Surpris-ingly, Dicrurus ludwigii and Ploceus bicolor were also found in the montane forest of Ki-boriani Mts, N of Mpwapwa, in a district with mostly deciduous forest at lower elevations, but it is possible that suitable habitat exists locally, in ravines with evergreen Ficus trees, and in the adjacent northern foothills of the Ukaguru Mts (Mlali Forest Reserve).
Arcanator, Swynnertonia, Campephaga quiscalina and Ploceus nicolli and the strict Ud-

j. fjeldså et al.26
zungwa endemic Nectarinia rufipennis, are most abundant in mid-altitude forest (Appendix 2 and Romdal 2001a). Also Anthreptes rubritorques, Apalis chariessa and Anthreptes pallidigaster may primarily be sub-montane, and the two latter inhabit lowland forest elsewhere in their range.
Thus, a common feature for the patchily dis-tributed species seems to be that their “source” habitat is at mid-altitude or in the foot-hills, sug-gesting that the montane forest would be a “sink” habitat for them. Because of the small amount of evergreen vegetation in the submontane zone of the Uvidunda and Rubeho Mts these species may be unable to maintain viable populations here. Also a number of more widespread species, which favour mid-elevation forests, or which use lowland forest as “winter refuge”, were recorded only locally and in very small numbers in the Rubeho area.
Anthropogenic or natural causes?In order to interprete the patchy distributions of mid-slope and foot-hill birds it is essential to ask whether the observed pattern of forest distribu-tion (Fig. 5) could be a result of recent habitat loss. The human impact in this area is consider-able and comprises logging and establishment of new cultivated plots inside forest reserves, and large fires lit every year in the woodland zones and grasslands (Appendix 1). We noted a high incidence of fires on the eastern slopes of the Rubeho highland.
However, it is also possible that the more open vegetation here compared with the slopes S of the Great Ruaha Gap may reflect different pat-terns of orographic rainfall. The eastern scarp of the Udzungwa highland rises very steeply above the lowland swamps, facing the monsoon winds almost like a wall, and even outside the rainy season there is an almost constant accumulation of clouds, especially in the Golo-golo Mts. As we follow the Udzungwa scarp to the SW, steep slopes facing E to SE have mainly evergreen for-est (semi-evergreen on the lower slopes) but S-facing slopes typically have deciduous woodland (Fig. 3). Unlike this, large portions of the Uvidun-da and Rubeho Mts appear as more moderately steep hills covered with Brachystegia woodland,
placed in the rain-shadow of moderately high foothill mountains (Pala Ulanga and undulating hills towards Kilosa; Figs 3 and 4), and of the Uluguru Mts (2638 m) nearer to the coast.
Thus, although local rainfall data are lacking, we expect less orographic rain in the Rubeho Mts than in the Udzungwa highlands (Lovett & Pócs 1993), and therefore evergreen forest oc-curs only as small patches in places with suitable inclination and ground water levels, in a matrix of otherwise deciduous woodlands habitat. Dur-ing the arid parts of the Pleistocene the amount of evergreen vegetation may of course have been even more restricted outside the steep east-facing slope of the Udzungwa highlands, maybe restricted to places with local fog formation and around wells in the foothills.
Over all, the transition from woodland to ev-ergreen montane forest, although often accentu-ated by human activity, corresponds to the lower level of clouds and mist-zones in the mountains. Where the landscape is wrapped in the mist, the net capture of water may be significantly increased because of interception rainfall in the forest, and the forest cover may therefore be self-perpetuating, maintaining a constantly humid environment and predictable supply of water further down-slope (see e.g. Bruinzeel 1993, Becker 1999). In addition to this we have noted that local thunderstorms with heavy rainfall often take place inside large clouds that accumulate over forested parts of the highlands, while it does not rain in the adjacent open habitats.
CONCLUSIONS
Although the montane forest birds are hardly ever seen outside their typical habitat, we do not find much evidence suggesting that their distributions are bounded – or interrupted – by the hot and dry Great Ruaha River Basin. Our fieldwork revealed a high abundance of endemic montane forest birds immediately adjacent to the basin, on both sides, and the genetic evidence suggested so far a high level of shared haplotypes across the basin, except in the two documented cases of species replacement. Unexpectedly, it is mainly the birds of lower elevational zones that show range dis-

27distribution of highland forest birds in tanzania
continuities or are present only in small densities N of the Great Ruaha River. The lower montane species wintering in the evergreen thickets of the foothill zone should have no problem with dis-persing along the eastern foothills, but this does not lead to establishment of persistent breeding populations in the Rubeho Area, except for spe-cies that are most tolerant to variation in habitat quality (compare Lens et al. 2002 and Doherty et al. 2003). The loss of species N of the Great Ruaha River is best explained by the scarcity of (semi)-evergreen vegetation in the lower parts of the mountain slopes, which in turn is related to moderate orographic rainfall favouring decidu-ous vegetation (accentuated today by frequent burning of these slopes).
Most highland forest birds appear to be seden-tary and crossings from one highland to another may be relatively rare events. Observations of forest birds in the deciduous woodlands in the exceptionally dry rainy season of 2002–3 would suggest that birds which normally reside inside evergreen forest start wandering when there is little food (LAH, unpublished). Formation of mobile mixed-species feeding parties is mostly seen in the dry season, when there are few insects (T. Lehmberg in litt.). Prigogine (1987) suggest-ed that dispersal could have taken place at times when the climate was more humid and elfin forest (low and stunted, mossy forest) could develop on lower altitudes than today. We do not know yet what is most important, only that dispersal of highland birds and gene flow appears to be suf-ficient to maintain population connectivity in the long term. It is generally assumed, as a guiding principle, that exchange of a single individual per generation is sufficient to maintain genetic homogeneity among populations (Wang 2004).
Some of the most restricted distributions (Xenoperdix spp., Nectarinia moreaui, new sunbird of the Nectarinia afra group) may suggest that such birds find suitable conditions close to the Great Ruaha Basin, on both sides. Also the high rarity scores for Image and Mangalisa (Table 1), and the occurrence of Sheppardia aurantiithorax and Scepomycter rubehoensis in highland forests near the transition towards the dry and warm inland sites, would suggest favourable condi-tions here. The Great Ruaha Basin represents a
local high-pressure cell with a persistently hot and sunny climate. The physical proximity to steep rain-capturing ridges (East slopes of Im-age, Luhombero and Golo-Golo Mts) and high plateaus and mountain domes with persistent blankets of nightly fog may mean that specific vegetation zones can be maintained locally in more or less the same positions, irrespective of the instability of the regional climate.
Some mountain slopes may have lost their forest cover during the coldest and driest parts of the Pleistocene, but because of local mist formation there may still have been patches of evergreen forest in some places in the Udzungwa and Rubeho highlands. In the case of Kisinga Rugaro forest, continuous cover of montane forest has been documented for the last 40.000 years, and this extraordinary stability has now been explained with reference to the circulation model for the Indian Ocean (Marchant et al. 2008). Because of the general stability of the area, and some site-specific factors, local biota may have persisted more or less in situ (Archaux 2004) in spite of the general instability of the Af-rican climate (Hamilton 1981, Nicholson 1994, deMenocal 1995).
The main factor explaining the distribution of endemic species in our study area may thus be constantly favourable conditions in certain places allowing populations to persist long enough for genetic coalescence and emergence of distinctive local lineages. This may require some degree of isolation, but in our area this does not seem to be physical barriers but instead that certain parts of the range function as sinks for some species, because of non-linear dynamics of colonization and extinction (Storch & Šizling 2002) and low tolerance of changing habitat quality or competi-tive situations (Lens et al. 2002). Even for fairly widespread species, part of their range may be sink habitat with small and temporary popula-tions that are not viable in the long term, and this may lead to lack of connectivity and gene flow between persisting source populations.
We will not, with this conclusion, deny that physical barriers play a role in speciation. They will, if they are large enough. However, we find it problematic that many students of speciation do not seem to consider other interpretations than

j. fjeldså et al.28
vicariance. If some landscape structure exists where two paraspe cies meet, then this becomes the cause of speciation, and if no such structure can be seen, then the existence of a past barrier is hypothesized. Indeed other possibilities are possible, also in the framework of allopatric speciation, but high levels of range dynamics may sometimes erase the evidence of how the speciation happened. Detailed data on population genetics may, however, aid the reconstruction of the past. With large numbers of tissue samples available for the Afromontane areas it is now possible to separate the roles of historical isola-tion, population growth and gene flow (Bowie et al. 2006, Fjeldså & Bowie 2008). Particular attention will be needed to tell whether there is geneflow through all parts of a species range, or if some parts represent sink populations and lost genes. If so, range-maps describing distributions with a broad all-inclusive outline will obscure the true geographical population structure.
In the effort to conserve biodiversity we need to consider carefully where we should focus. According to our conclusions, above, efforts to
Bowie, R. C. K., J. Fjeldså & J. Kiure. 2009. Multilocus mo-lecular DNA variation in Winifred's Warbler Scepomycter winifredae suggests cryptic speciation and existence of a threatened species in the Rubeho-Ukaguru Mts. – Ibis 151: 709–719.
Bowie, R. C. K., J. Fjeldså, S. J. Hackett & T. M. Crowe. 2004a. Systematics and biogeography of the double-collared sunbirds (Nectariniidae) of the Eastern Arc Mountains of Tanzania. – The Auk 121: 660–681.
Bowie, R. C. K., J. Fjeldså, S. J. Hackett & T. M. Crowe. 2004b. Molecular evolution in space and through time: mtDNA phylogeography of Olive Sunbirds (Nectarinia olivacea/obscura) throughout continental Africa. – Molecular Phy-– Molecular Phy-Molecular Phy-logenetics and Evolution 33: 56–74.
Bowie, R. C. K., J. Fjeldså, S. J. Hackett, J. M. Bates & T. M. Crowe. 2006. Coalescent models reveal the relative roles of dispersal, vicariance and ancestral polymorphism in shaping phylogeographical structure of an African montane forest robin. – Molecular Phylogenetics and Evolution 38: 171–188.
Bowie, R. C. K., G. Voelker, J. Fjeldså, L. Lens, S. J. Hackett & T. M. Crowe. 2005. Systematics of the olive thrush Turdus olivaceus species complex with reference to the taxonomic status of the endangered Taita Thrush T. helleri. – Journal of Avian Biology 36: 391–404.
Britton, P. L. (ed.). 1980. Birds of East Africa: their Habitat,
maintain the process of diversification should be focused on local environments where popula-tions may persist or even grow, rather than on barriers between such areas.
ACKNOWLEDGEMENTS
The fieldwork for this study was supported by the Danish Science Research Council (Grants No. 21-00-0178 and 21-01-0486), Aage V. Jensen’s Foundation, and for LAH by Queen Margrethe and Prins Henrik’s Foundation and Bøje Benzon’s Foundation). Molecular analysis of a large proportion of the tissue samples col-lected was done by Rauri Bowie (now Museum of Vertebrate Zoology, University of California, Berkeley). Tanzanian Commission for Science and Technology and Wildlife Department are thanked for their compliance with research and export permits. The Curators of the museum col-lections mentioned above are thanked for their courtesy. Neil Burgess and Rauri Bowie are thanked for comments to a manuscript draft.
REFERENCES
Archaux, F. 2004. Breeding upwards when climate is becom-ing warmer: no bird responses in the French Alps. – The Ibis 146: 138–144.
Baker, N. & E. Baker. 2002. Important Bird Areas in Tanzania. Wildlife Conservation Society of Tanzania, Dar es Salaam, 303 pp.
Becker, C. D. 1999. Protecting a Garúa forest in Ecuador: the role of institutions and ecosystem valuation. – Ambio 28: 156–161.
Beresford, P. 2003. Molecular systematics of Alethe, Sheppardia and some other African robins (Muscicapoidea). – The Ostrich 74: 58–73.
Beresford, P., J. Fjeldså, & J. Kiure. 2004. A new species of akalat (Sheppardia) narrowly endemic in the Eastern Arc of Tanzania. – The Auk 121: 23–34.
Bierregaard, R. O., T. E. Lovejoy, C. Gascon & R. Mesquita. 2001. Lessons from Amazonia: the Ecology and Conservation of a Fragmented Forest. Yale University Press, New Haven, 496 pp.
Bowie, R. C. K. 2003. Birds, Molecules and Evolutionary Processes among Africa’s Islands in the Sky. Ph.D. Thesis, University of Cape Town, 1–xvi + 370 pp.
Bowie, R. C. K. & J. Fjeldså. 2005. Genetic and morphological evidence for two species in the Udzungwa forest partridge Xenoperdix udzungwensis. – Journal of East African Natural History Society 94: 191–201.

29distribution of highland forest birds in tanzania
Status and Distribution. East Africa Natural History So-ciety, Nairobi, 271 pp.
Brooks, T., A. Balmford, N. Burgess, J. Fjeldså, L. A. Han-sen, J. Moore, C. Rahbek & P. Williams. 2001. Toward a blueprint for conservation in Africa. – BioScience 51: 613–624.
Brown, J. H. 1984. On the relationship between abundance and distribution of species. – American Naturalist 124: 255–279.
Brown, J. H. 1985. Macroecology. University of Chicago Press, Chicago, 260 pp.
Bruinzeel, L. A. 1993. Hydrology of moist tropical forests and effects of conversion: a state of knowledge review. UNESCO and Free University of Amsterdam, Paris, 224 pp.
Burgess, N. D. & C. O. F. Mlingwa. 2000. Evidence for altitudinal migration of forest birds between montane Eastern Arc and lowland forests in East Africa. – The Ostrich 71: 184–190.
Burgess, N. D., J. Fjeldså & R. Botterweg. 1998. Faunal importance of the Eastern Arc Mountains of Kenya and Tanzania. – Journal of East African Natural History So-ciety 87: 37–58.
Burgess, N. D., T. M. Butynski, N. J. Cordeiro, N. H. Doggart, J. Fjeldså, K. M. Howell, F. B. Kilahama, S. P. Loader, J. C. Lovett, B. Mbilinyi, M. Menegon, D. C. Moyer, E. Nashanda, A. Perkin, F. Rovero, W. T. Stanley & S. N. Stuart. 2006. The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. – Biological Conservation 134: 209–231.
Butynski, T. M. & C. L. Ehardt. 2003. Notes on ten restricted-range birds in the Udzungwa Mountains, Tanzania. – Sco-pus 23: 13–28.
Colwell, R. K. & J. A. Coddington. 1994. Estimating terres-trial biodiversity through extrapolation. – Philosophical Transaction of the Royal Society of London, Series B, 345: 101–118.
Connell, J. H. & E. Orians. 1964. The ecological regulation of species diversity. – American Naturalist 113: 891–833.
Cordeiro, N. 1998. Preliminary analysis of the nestedness patterns of montane forest birds of the Eastern Arc Moun-tains. – Journal of East African Natural History Society 87: 101–117.
Davenport, T. R. B., W. T. Stanley, E. J. Sargis, D. W. De Luca, N. E. Mpunga, S. J. Machaga & L. E. Olson. 2006. A new genus of African Monkey, Rungwecebus: morphol-ogy, ecology, and molecular phylogenetics. – Science 312: 1378–1381.
deMenocal, P. B. 1995. Plio-Pleistocene African climate. – Science 270: 53–59.
Diamond, J. M. 1980. Why are many tropical bird species distributed patchily with respect to available habitat? – Proceedings International Ornithological Congress 17: 968–973.
Diamond, J. M. 1981. Flightlessness and fear of flying in island species. – Nature 193: 507–508.
Dinesen, L., T. Lehmberg, M. C. Rahner & J. Fjeldså. 2001. Conservation priorities for the forests of the Udzungwa Mountains, Tanzania, based on primates, duikers and birds. – Biological Conservation 99: 223–236.
Dinesen, L., T. Lehmberg, J. O. Svendsen & L. A. Hansen.
1993. Range extension and other notes on some restricted-range forest birds from West Kilombero in the Udzungwa Mountains, Tanzania. – Scopus 17: 48–59.
Dinesen, L., T. Lehmberg, J. O. Svendsen, L. A. Hansen & J. Fjeldså. 1994. A new genus and species of perdicine bird (Phasianidae, Perdicini) from Tanzania; a relict form with Indo-Malayan affinities. – The Ibis 136: 2–11.
Doggart, N. K., A. Perkin, J. Kiure, J. Fjeldså, J. Poynton & N. D. Burgess. 2006. Changing places: how the results of new fieldwork in the Rubeho Mountains incluence conservation priorities in the Eastern Arc Mountains of Tanzania. – African Journal of Ecology 44: 134–144.
Doherty, P. F., T. Boulinier & J. D. Nichols. 2003. Local ex-tinction and turnover rates at the edge and interior of spe-cies’ ranges. – Annales Zoologici Fennica 40: 145–153.
Dowsett, R. J. & F. Dowsett-Lemaire. 1993. A contribution to the distribution and taxonomy of Afrotropical and Malagasy birds. Tauraco Press, Liege, Belgium, 389 pp.
Drummond, A. J., A. Rambaut, B. Shapiro & O. G. Pybus. 2005. Bayesian coalescent inference of past population dynamics from molecular sequences. – Molecular Biol-ogy and Evolution 22: 1185–1192.
Eken, G., L. Bennun, T. M. Brooks, W. Darwall, L. D. C. Fishpool, M. Foster, D. Knox, P. Langhammer, P. Matiku, E. Radford, P. Salaman, W. Sechrest, M. L. Smith, S. Spector & A. Tordoff. 2004. Key biodiversity areas as site conservation targets. – BioScience 54: 1110–1118.
Evans, T. D. & G. Q. A. Andersen (eds). 1992. A wildlife survey of the East Usambara and Ukaguru Mountains, Tanzania. ICBP, Cambridge, U.K., 106 pp.
Evans, T. D. & G. Q. A. Andersen. 1993. Results of an or-nithological survey in the Ukaguru and East Usambara Mountains, Tanzania. – Scopus 17: 40–47.
Fjeldså, J. 1999. The impact of human forest disturbance on the endemic avifauna of the Udzungwa Mountains, Tan-zania. – Bird Conservation International 9: 47–62.
Fjeldså, J. & J. Rabøl. 1995. Variation in avian communities between isolated units of the Eastern Arc montane forests, Tanzania. – Gerfaut 85: 3–18.
Fjeldså, J. & H. Tushabe. 2005. Complementarity of species distributions as a tool for prioritizing conservation actions in Africa: testing the efficiency of using coarse-scale dis-tribution data. Pp. 1–24 in Huber, B.A. et al. (Eds) African Biodiversity. Springer.
Fjeldså, J., R. C. K. Bowie & J. Kiure. 2006. The Forest Batis, Batis mixta is two species: description of a new, narrowly distributed Batis species in the Eastern Arc biodiversity hotspot. – Journal of Ornithology 147: 578–590.
Fjeldså, J., K. Howell & M. Andersen. 1997. An ornithologi-cal visit to the Rubeho Mountains. – Scopus 19: 73–82.
Fjeldså, J., M. S. Roy & J. Kiure. 2000. A montane subspe-cies of Sheppardia gunningi (East-coast Akalat) from Tanzania. – Bulletin of British Ornithologists Club 120: 27–33.
Fjeldså, J., D. Ehrlich, D. E. Lambin & E. Prins. 1997. Are biodiversity “hotspots” correlated with current ecocli-matic stability: A pilot study using the NOAA-AVHR remote sensing data? – Biodiversity and Conservation 6: 401–422.
Fjeldså, J., U. Johansson, L. G. S. Lokugalappatti & R. C. K. Bowie. 2007. Diversification of African greenbuls in space

j. fjeldså et al.30
and time: linking ecological and historical processes. – Jour-nal of Ornithology 148 (Suppl. 2): 359–367.
Franklin, I. R. & R. Frankham. 1998. How large must populations be to retain evolutionary potential. – Animal Conservation 1: 60–73.
Friedman, H. & K. E. Stager. 1964. Results of the 1964 Cheney Tanganyikan Expedition, Ornithology. – Con-tributions in Science, Los Angeles Country Museum, 84: 1–50.
Frontier Tanzania. 2001. West Kilombero Scarp Forest Reserve - Management and Summary Report. – 78 pp in Doody, K.Z., Howell, K.M. & Fanning, E. (Eds) Report for the Udzungwa Mountains Management and Biodiversity Conservation Project. MEMA, Iringa, Tanzania.
Fuchs, J., J. Fjeldså & E. Pasquet. 2006. An ancient African ra-diation of corvid birds detected by mitochondrial and nuclear sequence data. – Zoologica Scripta 35: 375–385.
Fuggles-Couchman, N. R. 1939. Notes on some birds of the Eastern Province of Tanganyika Territory. – The Ibis 14: 76–106.
García-Moreno, J. & J. Fjeldså. 2000. Chronology and mode of speciation in the Andean avifauna. – Bonner Zoological Monographs 46: 25–46.
Gaston, K. J. & T. M. Blackburn. 1996. Global-scale macro-ecology: interactions between population size, geograph-ic range size and body size in the Anseriformes. – Journal of Animal Ecology 65: 701–714.
Hamilton, A. C. 1981. The Quaternary history of African forests: its relevance to conservation. – African Journal of Ecology 19: 1–6.
Hanski, I. 1999. Metapopulation Ecology. Oxford University Press, Oxford, 328 pp.
Harrison, C. J. O. 1965. Concealed yellow pigment in the breeding plumage of some weavers. – Bulletin of British Ornithologists Club 85: 44–47.
Holt, R. D. 1997. On the evolutionary stability of sink popula-tions. – Evolutionary Ecology 11: 723–731.
Huber, B. A. 2003. High species diversity in one of the dominant groups of spiders in East African montane for-ests (Araneae: Pholcidae: Buitinga n.gen., Spermophora Hentz). – Zoological Journal of the Linnean Society 137: 555–619.
Hubbell, S. P. 2001. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton U.P., Princeton, 448 pp.
Isango, J. A. 2001. A partially annotated bibliography of biodiversity studies in the Eastern Arc Mountains. TAFORI and MEMA, Iringa, Tanzania, 327 pp.
Jacobs, B. F., J. D. Kingston & L. L. Jacobs. 1999. The origin of grass-dominated ecosystems. – Annals Missouri Bo-tanical Garden 86: 590–643.
Jansson, R. & M. Dynesius. 2002. The fate of clades in a world of recurrent climatic change: Milankovitch oscil-lations and evolution. – Annual Review of Ecology and Systematics 33: 741–777.
Jensen, F. P. & S. Brøgger-Jensen. 1992. The forest avi-1992. The forest avi-fauna of the Uzungwa Mountains, Tanzania. – Scopus 15: 65–83.
Jensen, F. P. & S. N. Stuart. 1982. New subspecies of forest birds from Tanzania. – Bulletin of British Ornithologists’ Club 102: 95–99.
Jetz, W., C. Rahbek & R. K. Colwell. 2004. The coinci-dence of rarity and richness and the potential signature of history in centers of endemism. – Ecology Letters 7: 1180–1191.
Johansson, U. S., J. Fjeldså & R. C. K. Bowie. 2008. Phy-2008. Phy-logenetic relationships within Passerida (Aves: Passeri-formes): a review and a new molecular phylogeny based on three nuclear intron markers. – Molecular Phylogenet-ics and Evolution 48: 858–876.
Johansson, U. S., J. Fjeldså, L. G. S. Lokugalappatti & R. C. K. Bowie. 2007. A molecular phylogeny and proposed taxo-2007. A molecular phylogeny and proposed taxo-nomic revision of African greenbuls (Aves: Passeriformes: Pycnonotidae). – Zoologica Scripta. 36: 417–427.
Johnson, C. N. 1998. Species extinction and the relation-ship between distribution and abundance. – Nature 394: 272–274.
Jones, T., C. L. Ehardt, T. M. Butynski, T. R. B. Davenport, N. E. Mpunga, S. J. Machaga & D. W. De Luca. 2005. The highland mangabey Lophocebus kipunji: a new species of African monkey. – Science 308: 1161–1164.
Lawton, J. H. 1996. Population abundances, geographic ranges and conservation: 1994 Witherby Lecture. – Bird Study 43: 3–19.
Lens, L., S. van Dongen, K. Norris, M. Githiru & R. Mat-thysen. 2002. Avian persistence in fragmented rainforest. – Science 298: 1236–1238.
Lindqvist, C. & V. A. Albert. 2001. A high elevation ancestry for the Usambara Mountains and lowland population of African violets (Saintpaulia, Gesneriaceae). – Systemat-ics and Geography of Plants 71: 37–44.
Louette, M. 1992. Barriers, contact zones and subspeciation in central equatorial Africa. – Bulletin of British Orni-thologists’ Club 112(A): 209–216.
Lovett, J. C. 1990. Classification and status of the moist for-ests of Tanzania. – Mitteilungen Institut von Allgemeine Botanik Hamburg 23a: 287–300.
Lovett, J. C. 2003. Tanzanian forest law. – Pp. 151–180 in Chaytor, B. & Gray, K. (Eds) Environmental Law and Policy in Africa. Kluwer Law International, The Hague, 362 pp.
Lovett, J. C. & T. C. E. Congdon. 1989. Some notes on a visit to Magalisa mountain, Tanzania. – East African Natural History Society Bulletin 19: 25–26.
Lovett, J. C. & T. E. E. Congdon. 1990. Some notes on a visit to Selegu mountain, Tanzania. – East African Natural His-tory Society Bulletin 20: 4–5.
Lovett, J. C. & T. Pócs. 1993. Assesment of the condition of the Catchment Forest Reserves a botanical appraisal. Ministry of Tourism, Natural Resources and Environ-ment, Dar es Salaam, Tanzania, 38 pp.
Lynes, J. 1934. Birds of the Ubena-Uhehe highlands and Iringa Uplands. – Journal für Ornithologie 82 (Sonder-heft): 1–147.
Marchant, R., C. Mumbi, S. Behera & T. Yamagata. 2007. The Indian Ocean dipole - the unsung driver of climatic variability in East Africa. – African Journal of Ecology 45: 4–16.
Mittermeier, R., N. Myers & C. G. Mittermeier (eds). 2000. Hotspots: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. Univ. of Chicago Press, Chicago, 432 pp.

31distribution of highland forest birds in tanzania
Moreau, R. E. 1966. The bird faunas of Africa and its islands. Acad. Press, London, 424 pp.
Nelson, G. & N. I. Platnick. 1981. Systematics and Biogeography: Cladistics and Vicariance. Columbia University Press, New York, 567 pp.
Nguembock, B., J. Fjeldså, A. Couloux & E. Pasquet. 2008. Phylogeny of Laniarius: Molecular data reveal L. liberatus synonymous with L. erlangeri and “plumage coloration” as unreliable morphological characters for defining species and species groups. – Molecular Phylogenetics and Evolu-tion 48: 396–407.
Nguembock, B., J. Fjeldså, A. Tillier & E. Pasquet. 2006. The phylogeny of the Cisticolidae (Aves: Passeriformes) based on nuclear and mitochondrial DNA sequence data, and implications for the evolution of a unique nest-building technique. – Molecular Phylogenetics and Evolution 42: 272–286.
Nicholson, S. E. 1994. Recent rainfall fluctuations in Africa and their relationships to past conditions over the conti-nent. – The Holocene 4: 121–131.
Ovaskainen, O. & I. Hanski. 2003. Extinction threshold in metapopulation dynamics. – Annales Zoologica Fennici 40: 81–97.
Poulsen, B. O. 1994. Movements of single birds and mixed-species flocks between isolated fragments of cloud forest in Ecuador. – Studies on Neotropical Fauna and Environ-ment 29: 149–160.
Prell, W. L., W. H. Hutson, D. F. Williams, A. Be, K. Gei-tzenauer & B. Molfino. 1980. Surface circulation of the Indian Ocean during the Last Glacial Maximum, approximately 18,000 yr BP. – Quaternary Research 14: 309–336.
Prigogine, A. 1987. Disjunctions of montane birds in the Afrotropical region. – Bonner Zoologische Beiträge 38: 195–207.
Pulliam, R. H. & B. J. Danielsen. 1991. Sources, sinks, and habitat selection: a landscape perspective on population dynamics. – American Naturalist 137: 51–66.
Ricklefs, R. E. & R. E. Latham. 1992. Intercontinental correlation of geographical ranges suggests stasis in ecological traits of relict genera of temperate perennial herbs. – American Naturalist 139: 1305–1321.
Romdal, T. S. 2001a. Altitudinal distribution and abundance patterns of bird species in the Eastern Arc Mountains, Tanzania. – Scopus 21: 35–54.
Romdal, T. S. 2001b. An ornithological survey of the Nguru Mountains, Tanzania. – Scopus 22: 49–62.
Romdal, T. S. 2003. Determinants of species distributions along spatial gradients of differing scale: classical similarities and important differences. – Ph.D. Thesis, Copenhagen University, 159 pp.
Rovero, F., M. Menegon, C. Leonard, A. Perkin, N. Doggart, M. Nbilinyi & L. Mlavela. 2008. A previously unsurveyed forest in the Rubeho Mountains of Tanzania reveals new species and range records. – Oryx 42: 16–17.
Rovero, F., G. B. Rathbun, A. Perkin, T. Jones, D. D. Ribble, C. Leonard, R. R. Mwakisoma & N. Doggart. 2008.
A new species of giant sengi or elephant-shrew (genus Rhynchocyon) highlights the exceptional biodiversity of the Udzungwa Mountains of Tanzania. – Journal of Zool-ogy 279: 126–133.
Roy, M. S., P. Arctander & J. Fjeldså. 1998. Speciation and taxonomy of montane greenbuls of the genus Andropadus (Aves: Pycnonotidae). – Steenstrupia 24: 63–78.
Seddon, N., J. M. M. Ekstrom, D. R. Capper, I. S. Isherwood, R. Muna, R. G. Pople, E. Tarimo & J. Timothy. 1999. Notes on the ecology and conservation status of key bird species in Nilo and Nguu North Forest Reserves, Tanza-nia. – Bird Conservation International 9: 9–28.
Shankar Raman, T. R. 2003. Assessment of census techniques for interspecific comparisons of tropical rainforest birds densities: a field evaluation in the Western Ghats, India. – The Ibis 145: 9–21.
Stanley, W. T., D. C. Moyer, E. A. Mulungu, J. Pokines & J. A. Bittner. 2002. Notes on owl pellets found in the Ud-zungwa Scarp Forest Reserve, Tanzania. – Journal of East African Natural History Society 91: 91–94.
Storch, D. & A. L. Šizling. 2002. Patterns in commonness and rarity in central European birds: Reliability of the core-satellite hypothesis within a large scale. – Ecography 25: 405–416.
Stuart, S. N. & T. van der Willigen. 1980. Is Moreau’s Sun-1980. Is Moreau’s Sun-bird Nectarinia moreaui a hybrid species? – Scopus 4: 56–58.
Stuart, S. N., F. P. Jensen, S. Brøgger-Jensen & R. I. Miller. 1993. The zoogeography of the montane forest avifauna of eastern Tanzania. – Pp. 203–28 in Lovett, J.C. & Was-ser, S.K. (Eds), Biogeography and ecology of the rain forests of eastern Africa. Cambridge Univ. Press, Cam-bridge, U.K., 351 pp.
Svendsen, J. O. & L. A. Hansen (eds). 1995. Report on The Uluguru Biodiversity Survey 1993. Royal Society for the Protection of Birds, Sandy, U.K., Part A 87 pp, Part B 112 pp.
Voelker, G., S. Rohwer, R. C. K. Bowie & D. Outlaw. 2007. Molecular systematics of a speciose, cosmopolitan songbird genus: Defining the limits of, and relationships among, the Turdus thrushes. – Molecular Phylogenetics and Evolution 42: 422–434.
Vrba, E. S., G. H. Denton, T. C. Partridge & L. H. Burckle (eds). 1995. Paleoclimate and Evolution, with emphasis on human origins. Yale Univ. Press, New Haven, 558 pp.
Wang, J. 2004. Application of the one-migrant-per-genera-tion rule to conservation and management. – Conserva-tion Biology 18: 332–343.
Whitlock, M. C., C. K. Griswold & A. D. Peters. 2003. Com-pensating for the meltdown: The critical effective size of a population with deleterious and compensatory mutations. – Annales Zoologica Fennici 40: 169–183.
Williams, P. H. 1994. WORLDMAP. Priority areas for bio-diversity. Using version 3. Privately distributed software, London, U.K.

j. fjeldså et al.32
APPENDIX I, STUDY AREAS
The geography of the centralEastern Arc MountainsThe SW part of the Eastern Arc Mts comprises large highlands with extensive tracts of highland forest, altogether ca. 1360 km2 in 56 patches in the Udzungwa highland and 400 km2 in 19 patches in the Rubeho area (including the Mangalisa high-land), and smaller forest patches on some outli-ers on the rain-shadow side. In most highlands a very sharp transition is seen between evergreen montane forest and deciduous woodland, appar-ently corresponding to the usual lower level of clouds or nightly fog. However, some east-facing slopes, especially on the Udzungwa, Uluguru and Nguru Mts, have (had) semi-evergreen forest all the way down to the lowlands, where ground-water forests are found in suitable places. Other east-facing slopes have deciduous woodland or fire-maintained tall grassland (Hyparrenia sp., Themeda triandra in the higher parts).
The Udzungwa highland has extensive high plateaus with fairly dry conditions with a range of open habitats, comprising montane grassland, marshes, heaths and stony and rocky areas, es-pecially in the W towards the dry “Makombako Gap”. Also the Rubeho highland has a consider-able land area, but a more limited spectrum of grassland, swamp and brackens habitats with scattered evergreen thickets (Aguaria, Cussonia, Myrica) suggesting that most parts may once have been forest covered.
The highlands are separated by an area of arid lowland (Great Ruaha Basin, at ca. 500 m) and a deep gorge through which the Great Ruaha River runs from the arid inland to the eastern lowlands at ca. 300 m (Fig. 3). This drainage pattern, which separates the two highlands down to the base, must predate the uplift of the flank-ing mountains. The river gorge has deciduous woodland and the Great Ruaha Basin has flat Acacia savannahs and Adansonia woodlands, and is flanked by enormous wooded hills, mostly miombo (Brachystegia mixed with Combretum, Protea and Pterocarpus), with evergreen aspect on the highest crests near 2000 m. Some huge wooded hills (Mbaga and Ipala, and the steep
but flat-topped Mangalisa highland) are located inside the basin.
The Great Ruaha Basin is flanked to the W by a high forest-covered ridge (Image Forest) extend-ing N from the Udzungwa highland. To the N the basin is bordered by the rather steep and rocky Wota Mts, which have small patches of evergreen forest, and the narrow Nyangolo Range, of kopje-like appearance, which is intersected by a gap through which the Great Ruaha River runs. To the E, the basin is flanked by rolling miombo hills right N of the Ruaha Gorge (Uvidunda Mts, with an evergreen forest patch on the Iyungi peak). The larger highland to the N is the Rubeho Mts (called Usagara Mts in some earlier accounts, although this name may initially have applied to the outliers near Sagara village, now officially called Kiboriani Mts).
To the N, the Rubeho highland is separated by the narrow Mkondoa valley (with road and rail-way connection from Kilosa to Dodoma) from the Ukaguru Mts, with some huge forested hills, ending in steep mountains in the N (Mamwira, 2336 m). Further inland of Mamwira are the dry and rocky Kiboriani Mts, N of Mpwapwa town, with degraded evergreen forest along the top ridge only. The Ukaguru/Kiboriani Mts are, in turn, isolated by 50 km of lowlands from the Nguru Mts (with isolated and very steep outliers Mt Kanga and Nguu Mts).
The climate is seasonal, with a long dry season from June to November. There are no rainfall stations in the study area, but Lovett & Pócs (1993) provide rainfall estimates for the differ-ent forests. High orographic rainfall characterise the eastern scarps, but the plateau forests have a considerable mist precipitation. This may be highest in the steep Golo-golo Mountains, which are most constantly cloud-covered, but also other highlands, such as Mangalisa, are often wrapped in a blanket of fog, which maintains humidity during the dry season.
Forest is being cut steadily, even in forest re-serves where this is illegal, so mature forest exists only in places of difficult access. The forest cover is also limited by frequent burning of the moun-tain slopes, which is often done purposefully, as people prefer open vegetation for easy walking and because of the fear of lions, buffalo and other

33distribution of highland forest birds in tanzania
wildlife, but it also happens incidentally, as the fire spreads due to the burning of bush-land for agriculture, or of trees with honey bees. Even though the Brachystegia trees are fire-resistant, continuous burning means a lack of regeneration and declining tree densities, except in stream ravines, and we assume that even evergreen for-est may gradually change into open woodland in this way.
Fortunately, comparison of old and recent aerial photos (MEMA project, Iringa) shows ex-pansion of low (second-growth) forest in many areas. The existence of extensive areas of low second-growth forest S of Ukwiva, and in the Ukami area, suggest that the forest is gradually growing back in places which are far from current human settlements. In the past, there were prob-ably scattered settlements all over the highland, but large areas were depopulated as a conse-quence of the Wahehe and Maji Maji uprising in 1905–07, and people were also forced (by earlier colonial powers, and in 1965, as a consequence of new Tanzanian legislation) to leave forest areas and settle in villages, which resulted in reduced pressure on some highlands. However, there was a boom of commercial logging in the 1970s and 1980s (Lovett 2003). Furthermore, the deciduous miombo (Brachystegia) woodlands of the Iringa highlands are regenerating very well as a result of a new management approach based on decisions by local communities.
Ornithological explorationA comprehensive literature review over Eastern Arc environments are provided by Isango (2001). Earlier biological exploration in the area covered in this paper was limited to the Udzungwa area (Lynes 1934, Jensen & Brøgger-Jensen 1992, Dinesen et al. 1993, 2001, Fjeldså & Rabøl 1995, Fjeldså 1999, Butynski & Ehardt 2003). There were hardly any ornithological data from the highlands to the N (except Friedman & Stager 1964, Evans & Andersen 1992, 1993, Fjeldså et al. 1997 and informations in Baker & Baker 2002), but all montane forests here were fairly well covered from an ornithological point of view during the years 1997, 1998 and 2000–2004 (see below). Thus the only significant gap is now the
extreme NE corner of the Udzungwa highland (Golo-Golo Mts, Fig. 2), right above the Kidatu dam in the Great Ruaha Gorge, where access is restricted.
The individual forest tracts(1) The Nyumbanitu Mts (55 km2) and Ndun-dulu Mts (240 km2) are located near the NE corner of the Udzungwa highland (7°40'–55'S 36°18–37'E), with the easternmost forested ridge Luhombero (2576 m) overlooking the Great Ruaha Gorge (Dinesen et al. 1994). Forest is continuous at 1200–2400 m, mostly big Albizia, Cassipourea, Cola, Craterispermum, Ficus, Myrianthus and Zanthoxylon, and some gigantic mahogany trees (Entandrophragma excelsum) in the Luhombero area (Butynski & Ehardt 2003). Aphloia, Neoboutonia, Nuxia and Podocarpus take over at 1850–2000 m, and Hagenia and Tecomaria near the mountain tops. The area is quite well described by Lovett & Pócs (1983) and Frontier Tanzania (2001), and ornithological data are reviewed by Jensen & Brøgger-Jensen (1992), Dinesen et al. (1993, 2001), Fjeldså (1999) and Fjeldså & Rabøl (1995). New data were gathered by LAH in 1993-4, 1998 and Nov. 2002 through Mar. 2003, with JK for part of the time, in Nyum-banitu Forest (N and S) and Ukami Forest, and by M. Baker (in lit.). Studies by LAH and JK in 2006-2007, partly on lower elevation and in other habitats (forest edges, swamps and open land) are not covered in this paper. Kisinga-Rugaro forest (7°50’S 35°53–60’E; see Marchant et al. 2006), which covers a large granitic dome in the central Iringa highland, has been visited in short surveys (LAH in 1992, JF 2008).
(2) Ukami forest (7°53–54'S, 36°24–25'E), which is at 1000-1600 m elevation on the southern slope of the Udzungwa highland. The forest grows on steep and rocky terrain, over an area with many parallel mountain ridges. Common forest trees are Allanblackia, Ficus, Parinari, Newtonia and Podocarpus, with some exceptionally tall trees. After permanent human settlements were aban-doned ca. 1955 there is now a strong regeneration of evergreen and deciduous trees on the transition to the adjacent woodlands, which means that this

j. fjeldså et al.34
forest area may become connected with Nyum-banitu, and with the Udzungwa Scarp Forest in the S. A 10-days survey visit was made in 1994 (LAH and J.O. Svendsen, unpublished), and LAH spent 2½ months here primo 2003, one month together with JK. Wildlife is abundant, especially bush-pigs, whose foraging activity creates an open understorey in many places.
(3) Mwanihana Forest (at (7°40–54'S, 36°42–53'E, in the Udzungwa Mountains National Park) covers the high and steep eastern scarp of the Udzungwa highland, right up to the Great Ruaha Gorge (Golo-Golo Mts). This area has continu-ous forests from the lowlands on the eastern scarp and is separated from the Ndundulu and Nyum-banitu forests by a stretch of fire-maintained montane grassland. Ornithological studies have been made since the late 1980s (Jensen & Brøg-ger-Jensen 1992) with recent studies along the full altitudinal gradient by T. Romdal (Romdal 2003). The high Golo-Golo Mts (2279 m) in the N, right on the edge to the Great Ruaha Gorge, is ornithologically unexplored because of steepness and security measures near the Kidatu Dam.
(4) Image Forest Reserve (7°22'15"–33'15"S, 36°08'15"–12'25"E). Brief exploratory visits were made in February 1992 by Lars Dinesen and Thomas Lehmberg, and in 1998 by Elia Mulungu (to the forest patch just N of the Morogoro-Iringa highway). JF, JK, Elia Mulungu and a forester worked in the N part of the forest 4–10 January 2000, entering from Ibumu village.
This forest covers a huge mountain ridge (2454 m, called Selebu or Selegu on some maps, although this name in fact applies to a large rock-face above the village Kilula) projecting N of the Udzungwa highland. The forest (gazetted 8920 ha) is broken into a small southern fragment and a large forest at 1800–2400 m along the high plateau further N. The most striking feature of the mountain range is the huge rock faces and boulders on the W side, but most of the forest is on an undulating plateau. The W slopes are mi-ombo woodland (Brachystegia) with some areas with Protea, Myrica and Erica (ex-Philippia), giving way to semi-evergreen vegetation on the upper slopes. The humid zone may initially have been dominated by Hagenia abyssinica
and Podocarpus latifolius, but was intensively logged in the 1970s, when several logging roads were made. In most parts, only few large for-est trees were left behind. The logging has now stopped, and only a few footpaths are in use, but the regeneration of forest trees proceeds very slowly and there is no regeneration of Hagenia. Thus, most of the area now has low (10–15 m) forest with some emergents, but there is mature HageniaPodocarpus forest in the extreme N. Dominating shrubs and small trees are Acokanthera laevigata, Aphloia theiformis, Cussonia spicata, Maesa lanceolata, Myrica salicifolia, Nuxia congesta, Lasianthus kilimandscharicus, Pavetta kyimbilensis, Rawsonia reticulata and Ziziphos, while larger trees comprise Bersama abyssinica, Croton macrostachyus, Ekebergia capensis, Ixora scheffleri, Podocarpus latifolius, Prunus africana, Rapanea melanophloeos, Sysygium cordatum and Zanthoxylum sp. (Lovett & Congdon 1990). The understorey is mainly tall herbs (labiats and Mimulopsis). In the N there are some large grassy glades and one large marsh. This area is used by elephants and black rhino (the presence of which was confirmed during our survey). On the E slope, the forest changes rapidly into miombo woodland, except for some strips of evergreen riparian scrub.
(5) The Uvidunda Mts with Iyungi (Mt Bismarck, 1900 m) (7°35'S, 36°51'E) are situated right N of the Great Ruaha Gorge. Studies 21–28 Nov. 1997 by JF, D. Moyer, D. Beale, E. Mulungu and bota-nist C. Frimodt-Møller. Fuggles-Couchmanni (1939) collected a small number of bird speci-mens in this highland.
The lower slopes are bush-land or lightly wooded, but the upper slopes have dense human settlements and are almost completely converted for agriculture, except for 5 km2 of evergreen forest. This is very degraded, the glades partly cultivated, so it is only around the steep central granite dome that the forest is undisturbed - but low and gnarled. The evergreen forest had Tabernaemontana, Polyscias and Schefflera spp, Allanblackia sp., Erica (undisturbed low forest on the top), Bridelia sp., Macaranga cf. kilimandscharica, Albizia gummifera, Newtonia sp., Aphloia theiformis, Leucas densiflora, Ocotea usambarensis, Memocylon, Myrianthus holstii,

35distribution of highland forest birds in tanzania
Ensete, Rapanea, Syzygium cf. guineense, Ochna cf. holstii, Phoenix reclinata, Dombeya sp. and Trema orientalis.
(6) Mangalisa (7º7–13’S, 36º23–28’E) is a large, table-like highland in the NE part of the Great Ruaha Basin (Mpwapwa district). JF, JK and Elia Mulungu worked mainly in the N part of the forest 18–24 Dec. 1999 and JK continuing in the S part until new-year. JK, ND and AP worked in the forest 26 Aug.–3 Sep. 2001 (Doggart et al. 2006).
This highland is separated from the Rubeho highland by 3 km of densely populated lowland (with the Kinusi village). The slopes are miombo woodland, mostly of very good quality, and the 92 km2 large top plateau at 2000–2287 m has an interesting vegetation gradient from decidu-ous Brachystegia spiciformis/Uapaca kirkiana woodland in the N to tall Podocarpus-dominated evergreen forest in the S. The middle portion of the plateau is well suited for agriculture (Mangal-isa and Mkwawa villages), and has mostly been turned into fields and fallow land with patches of Protea and Erica woodland and semi-evergreen forest. Evergreen forest, covering 4988 ha in the S, is somewhat logged, some parts having char-acter of secondary forest (Albizia gummifera, only 15 m tall but with closed canopy), thickets (Clutia abyssinica, Hagenia abyssinica, Lippia jaranica and Tecoma nyassae) and vine tangles. Numerous clearings are made for cultivation and large glades with grasses and Protea bushes on shallow stony soil may have been cultivated and grazed in the past. However, the southern half of the forest is mostly tall-growth forest with Podocarpus falcatus and latifolius, Polyscias fulva and Vebris nobilis, with Coffea sp. and tall herbs dominating the under storey. An unusual aspect (considering the elevation) is large numbers of 10-15 m tall palms (Phoenix reclinata). Although the evergreen forest is largely restricted to the top plateau it extends a few hundred meters down the southern scarp.
(7) The Rubeho Mts is an extensive highland E of the Great Ruaha Basin, extending from 6º45’ to 7º30’S. Much of this area is rolling wooded hills of moderate height, especially in the S, and in the NE towards Kidete and Kilosa, and
with a lower foothill mountain isolated to the E (Pala Ulanga, 7º12–13’S, 36º49–50’E). These parts are covered with miombo woodland or grassland with scattered thickets. The highest parts are covered with forest and grassland and bracken, but small thickets (Aguaria, Cussonia, Myrica) here and there in the open land suggest that the forest cover may once have been fairly continuous, without extensive natural grassland and rocky ground as found in the western part of the Udzungwa highland. Studies were conducted in the eastern foothills NW of Pala Ulanga (Sep. 1993, Fjeldså et al. 1997) and in Ukwiva forest 18–23 Sep. 1993 (JF and T. Msangi), 4–16 Aug. 2001 (AP, ND and JK) and Aug–Sep 2002 (JK), in Msanga forest 28 Sep.–8 Oct. 2001 (JK) and in northern Mafwemiro Forest 9–17 May 2000, 18–28 Feb. and 9–28 Mar. 2000 (JK), and in the southern parts 17–23 Aug. 2001 (AP, ND and JK) and 17 Nov–5 Dec 2002 (JF, JK and Jan Bolding Kristensen) (Doggart et al. 2006).
Msanga (Ilole) forest covers the top of the southernmost peak (7º28’S, 36º43’E), separated from the main plateau by a low pass (with the Mikumi-Mololo road). The lower part of the forest is of good quality (Rovero et al. 2008) but the area studied by JK near the summit at 2200 m (reached on foot from the Kasanga mission) has few big trees and is best characterised as second-ary forest of low stature.
Ukwiva Forest (6º58’–7º21’S, 36º34–51’E) covers nearly 100 km², at 1700–2150 m, with Aguaria, Albizea, Bersama, Chrysophylla, Macaranga, Myrianthus, Myrica and Syzygium. Most parts are disturbed by buffalo and elephants, and have quite impenetrable vegetation with brambles, vines (Tinnea), Hibiscus thickets and tall herbs (Fjeldså et al. 1997). There are no signs of recent human activity in this part of the forest, but terraces and pottery found inside the forest indicate that this area was partly cleared in pre-colonial time, maybe by the Wakwiva tribe, who now lives in the foothills to the NE. The southern part is low and stunted second-growth areas like upper Msanga Forest.
The highest parts of Ukwiva are dominated by low Aphloia, Erica and Tecoma shrubs. The eastern pars are rather dry (Agauria salicifolia, Aphloia theiformis, Bridelia micrantha, Catha edulis, Diospyros whyteana, Halleria lucida,

j. fjeldså et al.36
Macaranga kilimandscharica, Maesa lenceolata, Maytenus acuminata, Nuxia congesta, Parinari excelsa, Polyscias fulva), grading towards open woodland (Lovett & Pócs 1993). There are many valley floor wetlands dominated by reeds and sedges.
Mafwemiro forest (6º49–59’S, 36º33–37’E; we use the name given on maps, although local people call it Mafwomero) covers an out-lying ridge W of the main ridge (Kwiva), visited from Mbuga Mission in the S (on the ridge, at 1700 m, SW Kisi) and from Lufusi mission in the NE (at 1700 m, above Lumuma). 3237 ha of montane forest at 1900–2240 m is dominated by Albizia, Bridelia, Maesa, Myrianthus, Olea, Polyscias and Schrebera, with Aphloia, Podocarpus and Ocotea in the highest parts. The forest (especially the northern Chugu hill) is generally undisturbed, but Podocarpus, Ocotea and Syzygium have been logged out in the southern part, which is surronded by villages. A lake, overgrown with reeds, is located right S of the forest, near Mbuga Mission.
(8) Wota Mts in Mpwapwa is a rugged granitic area with no extensive plateaus, isolated NW of the NW end of the Rubeho highland by low Adansonia savannah that connects the lowlands E to Mpwapwa with the Great Ruaha Basin. Vis-ited by JK 14–28 Oct. 2001. Forest cover is very patchy. Wota Forest (6°48'S, 36°22'E, 457–731 m, and some on higher elevation) is 8 km2 of mosaic habitat of good submontane forest and disturbed areas (pit-sawing and fields). Ligunga Forest (6°50'S, 36°17'E), on a small plateau at ca. 2200 m, is situated W Kibakwe.
(9) Kiboriani Mts (6º17’S, 36º29’E) are located N of the Rubeho Mts, near Mpwapwa town, and separated from the northern Ukaguru Mts (Mlali forest) by a narrow valley. Visited by JK 19 Jan.
– 5 Feb. 2004, and again in Nov.–Dec. 2008, with collecting camps at 1750 (Ketua) and 2000 m (Mdote). Located in rainshadow, on the transition towards the dry inland of Dodoma District, this highland is covered by deciduous woodlands, and the upper slopes are mostly barren and rocky, but the top plateau has some evergreen forest, including Albizia, Ficus, Newtonia and Podocarpus. With a dense human population in the highland, the forest was seriously degraded in 2004 and had been converted to agriculture and pasture areas by 2008.
For a better understanding of the biogeographic relationships within of the Eastern Arc Mts, a comment regarding the Uluguru Mts (Fig. 1) is relevant. These high mountains (2635 m) are iso-lated by the Mikumi and Mkata plains 100 km E of the Rubeho Mts. Their avifauna resembles that of the Udzungwa’s except for the lack of seven range-restricted species and the presence of three narrow endemic: Malaconotus alius (Uluguru Bush-shrike), a vanishing species of submontane humid forest; Nectarinia loveridgii (Loveridge’s Sunbird, sister to N. moreaui, see Bowie et al. in press) and Andropadus (fusciceps) neumanni (Uluguru Mountain Greenbul, Roy et al. 1998). When following the mainroad inland (Morogoro to Iringa) one gets the impression that the Ulu-gurus are separated from the other mountains by rather low laying terrain, but in fact the terrain rises gently to the S, and when the landscape is viewed from the top of Iyungi (Uvidunda Mts) it is seen that a row of wooded hilltops (with evergreen submontane forest on Malundwe Hill, 1250 m, 7°24'S, 37°18'E) form a chain between Uvidunda and the south-western outliers of the Uluguru Mts. This provides a potential series of "stepping-stones" for forest species in the most humid climatic periods.

37distribution of highland forest birds in tanzania
APPENDIX 2
Birds of forest and adjacent non-forest habitat (excluding non-forest birds which are widely distributed in eastern Africa, and lowland and submontane species that only exceptionally are seen above 1400 m). For each species a habitat code is given (following Romdal et al., unpub-lished) and the altitudinal range, both these bits of information refer to observations from this part of the species’ ranges and disregards the habitats or altitudinal zones that a species may inhabit in other parts of Africa): F = Forest, E = edges, W
= woodland, O = open land, L = lowlands, below 800 m, S = submontane 800–1500 m, M = mon-tane, above 1500 m, u = ubiquitous, CAPITAL letters equates to dependence or strong associa-tion with a certain habitat or zone, small letters a less strong association. Abundance data for the individual forests are subjective: + meaning that the species was recorded, C that it was common (present in some numbers) and A that it was abundantly present.
Anas sparsa, FSm, 550–2000 + + - - - + - -/- -Xenoperdix udzungwensis, FsM, 1340–2000 + - - - - - - -/- -Xenoperdix obscurata, FsM, 1900–2200 - - - - - - + -/- -Francolinus “squamatus”, fSm, 1000–2000 C + + ? - + + -/- CGuttera pucherani, fLs, 300–2110 C + + - + C + +/- +Pogoniulus leucomystax, fsM, 1140–2240 A C A C C C A C/C +P. bilineatus, fLS, 300–2160 C - - C - + - -/- ?Stactolaema olivacea, flSm, 400–2100 C + + C + + + -/- +S. leucotis, fLs, (400–) 1100–1400 (–2000) + - + - - - - -/- -Indicator variegatus, u, 550–1830 + + + - + + + +/+ +I. minor, u, 1700–2000 + + + - - + + +/- -Campethera abingoni, fLs, 700–2000 + - + - - - - -/- -Dendropicos fuscescens, fLs, 1100–1950 + - + + + + - -/- +D. griseocephalus, FsM ,1340–2160 + + + + + + + +/+ -Apaloderma vittatum, FsM, 1100–2160 C C C C + C C C/C +Phoeniculus purpureus, wfLS, 1100–1720 + + + - - + - -/+ +Ceratogymna brevis, fSu, 300–2410 A C C + + + + +/- +C. buccinator, fLs, 300–1830 C - - - + + + +/- -Tockus alboterminatus, fLS, 300–2160 + + + + + + + + CChrysococcyx klaas, Wfu, 300–2130 + + + C - C + +/- -Cuculus solitarius, wfsu, 1340–2000 C + + ? - + + -/- +
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
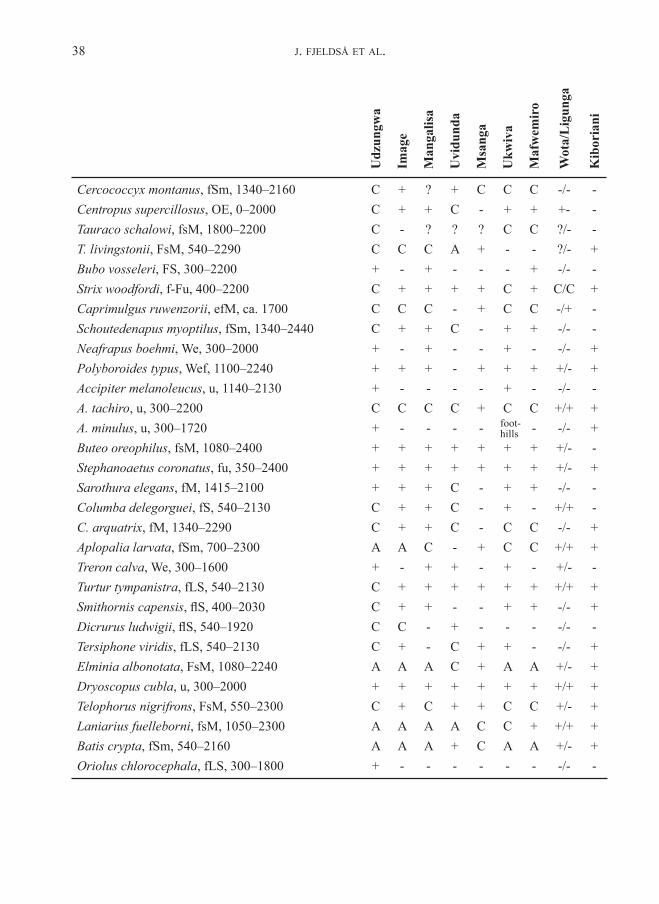
j. fjeldså et al.38
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
Cercococcyx montanus, fSm, 1340–2160 C + ? + C C C -/- -Centropus supercillosus, OE, 0–2000 C + + C - + + +- -Tauraco schalowi, fsM, 1800–2200 C - ? ? ? C C ?/- -T. livingstonii, FsM, 540–2290 C C C A + - - ?/- +Bubo vosseleri, FS, 300–2200 + - + - - - + -/- -Strix woodfordi, f-Fu, 400–2200 C + + + + C + C/C +Caprimulgus ruwenzorii, efM, ca. 1700 C C C - + C C -/+ -Schoutedenapus myoptilus, fSm, 1340–2440 C + + C - + + -/- -Neafrapus boehmi, We, 300–2000 + - + - - + - -/- +Polyboroides typus, Wef, 1100–2240 + + + - + + + +/- +Accipiter melanoleucus, u, 1140–2130 + - - - - + - -/- -A. tachiro, u, 300–2200 C C C C + C C +/+ +A. minulus, u, 300–1720 + - - - - - -/- +Buteo oreophilus, fsM, 1080–2400 + + + + + + + +/- -Stephanoaetus coronatus, fu, 350–2400 + + + + + + + +/- +Sarothura elegans, fM, 1415–2100 + + + C - + + -/- -Columba delegorguei, fS, 540–2130 C + + C - + - +/+ -C. arquatrix, fM, 1340–2290 C + + C - C C -/- +Aplopalia larvata, fSm, 700–2300 A A C - + C C +/+ +Treron calva, We, 300–1600 + - + + - + - +/- -Turtur tympanistra, fLS, 540–2130 C + + + + + + +/+ +Smithornis capensis, flS, 400–2030 C + + - - + + -/- +Dicrurus ludwigii, flS, 540–1920 C C - + - - - -/- -Tersiphone viridis, fLS, 540–2130 C + - C + + - -/- +Elminia albonotata, FsM, 1080–2240 A A A C + A A +/- +Dryoscopus cubla, u, 300–2000 + + + + + + + +/+ +Telophorus nigrifrons, FsM, 550–2300 C + C + + C C +/- +Laniarius fuelleborni, fsM, 1050–2300 A A A A C C + +/+ +Batis crypta, fSm, 540–2160 A A A + C A A +/- +Oriolus chlorocephala, fLS, 300–1800 + - - - - - - -/- -
foot-hills
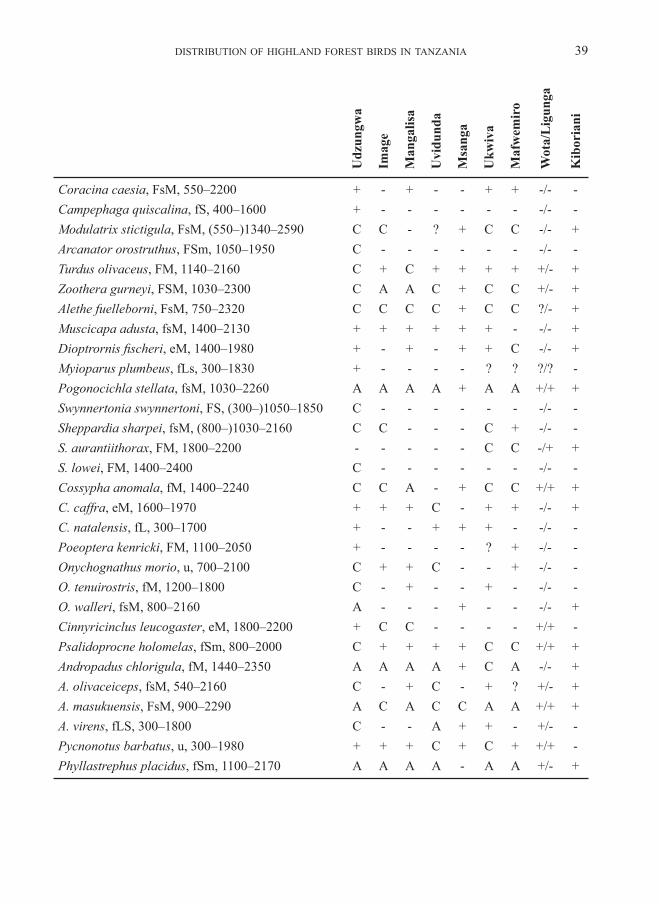
39distribution of highland forest birds in tanzania
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
Coracina caesia, FsM, 550–2200 + - + - - + + -/- -Campephaga quiscalina, fS, 400–1600 + - - - - - - -/- -Modulatrix stictigula, FsM, (550–)1340–2590 C C - ? + C C -/- +Arcanator orostruthus, FSm, 1050–1950 C - - - - - - -/- -Turdus olivaceus, FM, 1140–2160 C + C + + + + +/- +Zoothera gurneyi, FSM, 1030–2300 C A A C + C C +/- +Alethe fuelleborni, FsM, 750–2320 C C C C + C C ?/- +Muscicapa adusta, fsM, 1400–2130 + + + + + + - -/- +Dioptrornis fischeri, eM, 1400–1980 + - + - + + C -/- +Myioparus plumbeus, fLs, 300–1830 + - - - - ? ? ?/? -Pogonocichla stellata, fsM, 1030–2260 A A A A + A A +/+ +Swynnertonia swynnertoni, FS, (300–)1050–1850 C - - - - - - -/- -Sheppardia sharpei, fsM, (800–)1030–2160 C C - - - C + -/- -S. aurantiithorax, FM, 1800–2200 - - - - - C C -/+ +S. lowei, FM, 1400–2400 C - - - - - - -/- -Cossypha anomala, fM, 1400–2240 C C A - + C C +/+ +C. caffra, eM, 1600–1970 + + + C - + + -/- +C. natalensis, fL, 300–1700 + - - + + + - -/- -Poeoptera kenricki, FM, 1100–2050 + - - - - ? + -/- -Onychognathus morio, u, 700–2100 C + + C - - + -/- -O. tenuirostris, fM, 1200–1800 C - + - - + - -/- -O. walleri, fsM, 800–2160 A - - - + - - -/- +Cinnyricinclus leucogaster, eM, 1800–2200 + C C - - - - +/+ -Psalidoprocne holomelas, fSm, 800–2000 C + + + + C C +/+ +Andropadus chlorigula, fM, 1440–2350 A A A A + C A -/- +A. olivaceiceps, fsM, 540–2160 C - + C - + ? +/- +A. masukuensis, FsM, 900–2290 A C A C C A A +/+ +A. virens, fLS, 300–1800 C - - A + + - +/- -Pycnonotus barbatus, u, 300–1980 + + + C + C + +/+ -Phyllastrephus placidus, fSm, 1100–2170 A A A A - A A +/- +

j. fjeldså et al.40
P. flavostriatus, fS, 500–1950 A - - - - + - -/- -P. cerviniventris, fS, 300–1800 + - - + - + - - -Camaroptera brachyura, eu, 300–1830 + + - + + + + +/- -Apalis thoracica, FM, 1340–2320 C C C + + C C +/+ +A. alticola, efM, 1340–2030 + - - - - - - -/- -A. chariessa, FSm, 1000–2000 C - - - - - - -/- -A. melanocephala, FlS, 300–2030 C + - - + - + -/- -A. chapini, FsM, 1100–2200 C C C C + C + -/- -Zosterops senegalensis, fsM, 1200–2290 A + C A - C C C/C +Chloropeta similis, eM, 1700–1950 C + + C + C + -/- -Scepomycter rubehoensis, FsM, 1300–2350 + - - - - C - -/- -Artisornis metopias, FM, 1000–2570 C C C C + C C C/- +Bradypterus mariae, fsM, 700–2200 A A C A + A A C/- +B. cinnamomeus, efM, (1340-)1700–2300 + + + + - + - -/- -Phylloscopus ruficapillus, fsM, 550–2000 C - - + + + + +/+ +Macrosphenus kretchmeri, fLs, 300–1800 + - - - - + - -/- -Illadopsis rufipennis complex, flS, 600–2000 C - - - - + -/- +Pseudalcippe abyssinica, fM, 1180–2290 C A C C + C A +/- -Anthreptes collaris, elS, 300–2030 C - - + + + + +/+ +A. pallidigaster, FlS, 200–1550 + - - - - - - -/- -A. rubritorques, FlS, 250–1550 + - - - - - - -/- -A. neglectus, flS, 300–1800 C - - - - + - -/- -Nectarinia venusta, eS(m?), 1340–2000 + - + + - C C +/+ +N. rufipennis, F, 600–1710 C - - - - - - -/- -N. moreaui, fM, 1350–2240 A A A A C C C -/+ +N. fuelleborni, fM, 1400–2030 C - - - - - - -/- -N. afra group (unnamed), eM, ca. 2000 - + + - - + C -/? -N. olivacea, flSm, 300–2160 A C C A + C A +/+ +N. kilimensis, eoM, 1500–2400 + + - + - + + -/- -N. verticalis, efSm, 1500–2050 + - - - + C C -/- -
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
foot-hills
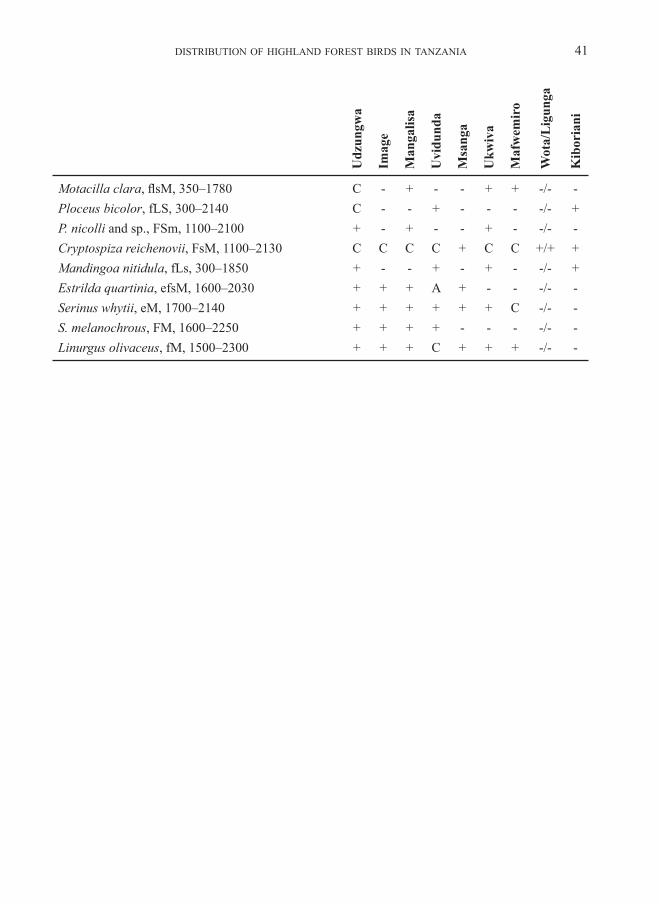
41distribution of highland forest birds in tanzania
Motacilla clara, flsM, 350–1780 C - + - - + + -/- -Ploceus bicolor, fLS, 300–2140 C - - + - - - -/- +P. nicolli and sp., FSm, 1100–2100 + - + - - + - -/- -Cryptospiza reichenovii, FsM, 1100–2130 C C C C + C C +/+ +Mandingoa nitidula, fLs, 300–1850 + - - + - + - -/- +Estrilda quartinia, efsM, 1600–2030 + + + A + - - -/- -Serinus whytii, eM, 1700–2140 + + + + + + C -/- -S. melanochrous, FM, 1600–2250 + + + + - - - -/- -Linurgus olivaceus, fM, 1500–2300 + + + C + + + -/- -
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni

j. fjeldså et al.42
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
APPENDIX 3
Birds of forest and forest edges of the study area (excluding submontane species recorded only exceptionally above 1500 m: Circaetus fasciatus, Apaloderma narina, Accipiter ovampensis, Streptopelia semitorquata, Ceuthmochares aereus, Laniarius aethiopicus/sublacteus, Trochocercus cyanomelas, Platysteira peltata, Muscicapa caerulescens, Nicator gularis, Chlorocichla flaviventris, Apalis flavida, Ploceus spp., Amblyospiza albifrons, Serinus flavicollis and Pyrenestes minor). A short list is appended with species of restricted or otherwise interesting distributions seen outside the highland forests. For each species a habitat code is given (fol-lowing T. Romdal et al., unpublished) and the
altitudinal range, both these bits of information refer to observations from this part of the species’ ranges and disregarding habitats or altitudinal zones that a species may inhabit in other parts of Africa): F = Forest, E = edges, W = woodland, O = open land, L = lowlands, below 800 m, S = submontane 800-1500 m, M = montane, above 1500 m, u = ubiquitous. CAPITAL letters equate to dependence or strong association with a certain habitat or zone, small letters a less strong associa-tion. Abundance data for the individual forests are subjective: + meaning that the species was recorded, C that it was present in some numbers and A that it was abundantly present.
Francolinus afer, Wfu, ?–2200 + + - - - ? ? -/- -Turnix sylvatica, uW, 0–2000 + + - - - - ? -/- -Musophaga porphyreolopha, wfu, ?–1300 + + + - - + - -/- +Circus ranivorus, wo, 300–1700 + - - - - + + -/- -Buteo augur, oM, 1500–2200 + + + + - + + -/- +Merops bullockoides, WE, ?–1400–2100 + + C - - + + ?/? -Lanius marwitzi, oM, above 1500 + + + - - + + -/? +Saxicola promiscua, uSM, 1000–2000 C + + C + C C +/+ -Monticola angolensis, WS, 600–2000 + - + - + - - ?/? +Psalidoprocne albiceps, eM, 1380–2000 C + + - - + C -/- -Cisticola lais, OM, above 1500 + + - - - - - -/- -C. woosmani, wo, 900–1800 + + + + + + ? ?/? -C. nigriloris, eoM, 1100–2750 C + C + + + + -/- -C. njombe, MO,ef, 1830–3000 + + - - + - + +/+ -C. cantans, U, mainly above 1000 - - - + - - - - -C. erythrops, U, 0–2300 - - - + + - - - -Nectarinia amethystina, Wf, ca. 600–1500(–2200) - - - + + ? ? ?/? +

43distribution of highland forest birds in tanzania
Udz
ungw
a
Imag
e
Man
galis
a
Uvi
dund
a
Msa
nga
Ukw
iva
Maf
wem
iro
Wot
a/L
igun
ga
Kib
oria
ni
N. famosa, eoM, 1300–1880 + - + + + - - -/+ -Anthus cinamomeus, oM, ca. 2000 C C C - + + + -/- -A. brachyurus, OM, 1800–2000 + + ? - - - - ?/? -Ploceus bertrandi, oS, 1080–1740 + - - - - - - -/- -P. ocularis, eu, 300–1950 + + + - + + + -/- +Euplectes psammocromius, OM, 1800–300 + ? - - - ? ? -/- +Serinus hypostictus, esM, 1600–2000 C + + + + C C -/- -




















