
Topology of the Shaker Potassium Channel Probed with Hydrophilic Epitope Insertions

Distinction and characterisation of submergence tolerantandsensitive ricecultivars, probedby thefluorescenceOJIPrise kinetics
Ramani Kumar SarkarA,B and Debabrata PandaA
ADivision of Biochemistry, Plant Physiology and Environmental Sciences, Central Rice Research Institute,Cuttack-753 006, India.
BCorresponding author. Email: [email protected]
Abstract. Rice (Oryza sativa L.) plants experience multiple abiotic stresses when they are submerged. In addition to theeffects of submergence on gas exchange, water also creates shading of submerged plants. It is believed that responses tosubmergence are actually responses to low light stress, although during complete submergence in addition to low light otherenvironmental factors like reduce movement of gases affect the plant growth, and therefore, the consequences ofsubmergence are not always alike to shade. We monitored the extent to which shade and submergence change the plantheight, chlorophyll a fluorescence characteristics and CO2 photosynthetic rate in three Indica rice cultivars, namely Sarala,Kalaputia andKhoda, which differed in submergence tolerance. Therewere both similarities and dissimilarities between theconsequence of shade and submergence on rice plants. Under shade conditions, elongation growth was greater insubmergence tolerant cultivars than the sensitive cultivar, whereas elongation growth was greater under submergencein sensitive cultivar. The reduction in chlorophyll content, damage toPSII, anddecrease inCO2photosynthetic ratewasmorenotable under submergence than the shade conditions. Our results show that several JIP-test parameters clearly distinguishbetween submergence tolerant and sensitive cultivars, and responses to submergence among different rice cultivars differdepending on their sensitivity to submergence. Therewere different interactions between cultivar and shade (~low light) andcultivar and submergence.
Additional keywords: chlorophyll fluorescence, photosynthesis, rice, shade, submergence tolerance.
Introduction
Rice is grown in a range of ecologies ranging from irrigated touplands, rainfed lowland, deepwater and tidalwetlands. It is oftenthe only crop that can be grown in flood prone ecosystems, theproductivity of which is greatly affected by the intensity anddurationof rainfall; quality, depth, anddurationof standingwater;and timing and frequency of flooding. Complete submergence byflash flooding is a major production constraint, and at least 16%of the rice lands of the world suffer from flash flooding (Sarkaret al. 2006).
Submergence tolerance is physiologically complex, butgenetically simple (Setter et al. 1997), and is strongly affectedby environmental variables which play a vital role in survival(Jackson and Ram 2003; Sarkar et al. 2006). There are manyenvironmental stresses to which plants are exposed duringcomplete submergence in the field, including reducedmovement of gases to and away from plant surfaces (Colmerand Pedersen 2008a). In addition to its effects on gas exchange,water shades submerged plants logarithmically with increasingdepth (Jackson and Ram 2003), and increasing the depth of waterthe amount of shading promotes submergence injury in plants(Pierik et al. 2005). Further, shaded conditions may be furtherintensified by cloudiness during rainy season; the main rice
cultivation season of South and south-east Asia. Theunderwater light regime is a major controlling factor of CO2
and O2 concentrations, and, hence, greatly affects thephysiological status of submerged plants (Vartapetian 2006;Voesenek et al. 2006; Colmer and Pedersen 2008b).Therefore, the degree of submergence injury depends not onlyupon the species and the timing of floods, but also on lightconditions. The survival rates of rice species were shown to bemuch higher when submerged in diffuse light than in completedark (Panda et al. 2006, 2008), indicating that underwaterphotosynthesis may play an important role in the response ofterrestrial plants to survival under submergence (Vervuren et al.1999; Mommer and Visser 2005; Mommer et al. 2006, 2007).Decreased intensities of light and submergence induce similarsets of traits in plants, such as an increase in leaf length and SLA,and a decline in the underwater compensation point for light(Sarkar et al. 1996; Mommer et al. 2005a, 2005b). Hence, poorplant growth and survival during complete submergence is oftenconsidered a consequence of shading. The resemblance in themorpho-physiological and biochemical responses to low lightand submergence are many, and acclimation to submergencehas been hypothesised to be an acclimation to shade (Vervurenet al. 1999; Boeger and Poulson 2003; Pierik et al. 2005;
CSIRO PUBLISHING
www.publish.csiro.au/journals/fpb Functional Plant Biology, 2009, 36, 222–233
� CSIRO 2009 10.1071/FP08218 1445-4408/09/030222

Voesenek et al. 2006). However, during complete submergencein addition to low light, alteration in theconcentrationsofCO2andO2 occurs, but in the shaded aerated conditions, low light is theonly environmental stimulus which alters the normal functionalcharacteristics of the plant. This indicates that the consequencesof submergence are not always alike to shade.
In the present investigation, fast chlorophyll fluorescence riseand photosynthesis were studied in three rice cultivars (Sarala,high yielding, plant breeders’ variety; Kalaputia and Khoda,local germplasm lines, farmers’ varieties) that differ incomplete submergence tolerance (Panda et al. 2006). Thesephotosensitive rice cultivars are generally grown in the rainyseason under rainfed lowland conditions of eastern India. Theobjectives were to investigate differences between cultivars in:(i) pigment composition, (ii) chloroplast functional and structuralintegrity, (iii) photosynthetic performances, (iv) plant elongationunder submergence and shade, and (v) distinguish betweensubmergence tolerant and sensitive rice cultivars.
Materials and methodsPlant material and growth conditions
Seeds of rice (Oryza sativaL.) cultivars, namely, Sarala (sensitiveto submergence) and Kalaputia and Khoda (tolerant tosubmergence), were sown directly in earthen pots containing2 kgof farmsoil (sandy–clay–loam,pH6.8) and farmyardmanurein a 3 : 1 ratio. Each pot was supplied with 80mg urea, 192mgsingle super phosphate (P2O5) and70mgmurate of potash (K2O).Plants were thinned to five seedlings per pot 10 days aftergermination. Each pot is considered one replicate. Theexperiment was arranged in a randomised complete blockdesign with three replicates for studies of the differentphysiological characteristics.
Experimental set-up
Twenty-day-old seedlings were completely submerged inconcrete tanks (dimension 3� 3� 1.3m) under 120 cm ofwater so that at least 60 cm of water remained above the plantheight. Plants were submerged for 2, 4, 6 or 8 days. This imposedsevere submergence stress, and plants were under completelysubmerged conditions. To study the relevant changes under airadapted conditions, certain pots after 8 days of submergencewerere-exposed to air for 1 day under normal conditions in which riceis cultivated, i.e. with 5–10 cm of stagnant water above soil. Thecharacteristics of thefloodwater in terms of light availabilityweremeasured at 1200 hours (model LI-189, Li-Cor, Lincoln, NE,USA) and water temperature and oxygen concentration weredetermined at 0600 and 1630 hours (model Simplair-F-5, SylandScientific, Heppenheim, Germany). Light intensity at 70 cmwater depth or at the vicinity of canopy level ranged from179 to 194 quantum (mmolm�2 s�1), whereas it was1990–2106 quantum (mmolm�2 s�1) above the water surface.The oxygen concentration at the same water depth was2.6–4.2mgL�1 at 0600 hours and 6.5–10.2mgL�1 at1630 hours. The temperature varied between 28.2 and 33.2�Cthroughout the period of the experiment. To study the effect ofsubmergenceon subsequent re-emergence,we selectedold leavesthat experienced complete submergence stress.
Briefly, the experiment conducted under shade condition wassimilar to submergence.Here, instead of submergence, shadewasapplied. The subtreatments provided under shaded conditionswere exactly the same as under submergence. To achieve a lowirradiance treatment, pots were covere with white muslin cloth sothat the screens produced the same initial degree of light reduction(91%) like column of water so that at the vicinity of canopy level,light intensity remained below200quantum (mmolm�2 s�1). Theaverage intensity of light varied between 172 and 196 quantum(mmolm�2 s�1) at 1200 hours during the entire period ofexperiment. Pots without screens/submergence had a normalirradiance and termed as control. All together, there were twomain treatments, i.e. submergence and shade, and sixsubtreatments, i.e. control, 2, 4, 6, or 8 days of main treatmentand 8 days submergence/shade treatment followed by 1 day air,i.e. return to normal conditions.
Plant height estimation
Plant height was determined after 8 days of treatment in control,submergence and shade treated plant. Absolute elongation of theplant was the height after submergence/shademinus the height ofthe control plants.
CO2 photosynthetic rate measurements
Measurements of CO2 photosynthetic rate weremade on the fullyexpanded leaves of five different plants by using an open systemphotosynthetic gas analyser (model TPS-1, PP Systems,Haverhill, MA, USA) under normal environmental conditions.The second and third leaf from the top was selected andkept inside the chamber under ambient irradiance (PAR821–994mmolm�2 s�1) until a stable reading was achieved.
Fluorescence measurements
After measuring the CO2 photosynthetic rate, the same leaveswere used for the measurement of chlorophyll (Chl) fluorescenceusing a plant efficiency analyser, Handy PEA (HansatechInstruments Ltd, Norfolk, UK) and data were recorded from10ms up to 1 s with a data acquisition of every 10ms for thefirst 300ms, then every 100ms up to 3ms and later every 1ms.Thesignal resolution was 12 bits (0–4000). For each treatment,the Chl a fluorescence transients of 12 individual leaves weremeasured. Leaves were maintained in darkness for 20min beforetaking the data on Chl fluorescence. Themaximal intensity of thelight source, providing an irradiance saturating pulse of3000mmol (photons)m�2 s�1 was used. From the fast O-J-I-Ptransients, several bio-energetic parameters (Table 1) werederived according to the equations of the JIP-test using theprogram BIOLYSER (RM Rodriguez, BioenergeticLaboratory, University of Geneva).
Chlorophyll content
After measuring the CO2 photosynthetic rate and Chlfluorescence characteristics, the same leaves were used for themeasurement of Chl content, which comprised both Chl a andChl b. Samples of 100mg of finely chopped fresh leaves wereplaced in a capped measuring tube containing 25mL of 80%acetone, and placed inside a refrigerator (4�C) for 48 h. Chl
Submergence and shading effects on rice Functional Plant Biology 223
was measured spectrophotometrically following the methodby Porra (2002).
Statistical analysisDifferences between plant height, various Chl fluorescenceparameters and CO2 photosynthetic rates of three rice cultivarswere compared byANOVAusing IRRISTAT (International RiceResearch Institute, Philippines) software’s least significantdifference (l.s.d., *P < 0.05), provided the F-test was significant.
Results
Plant height
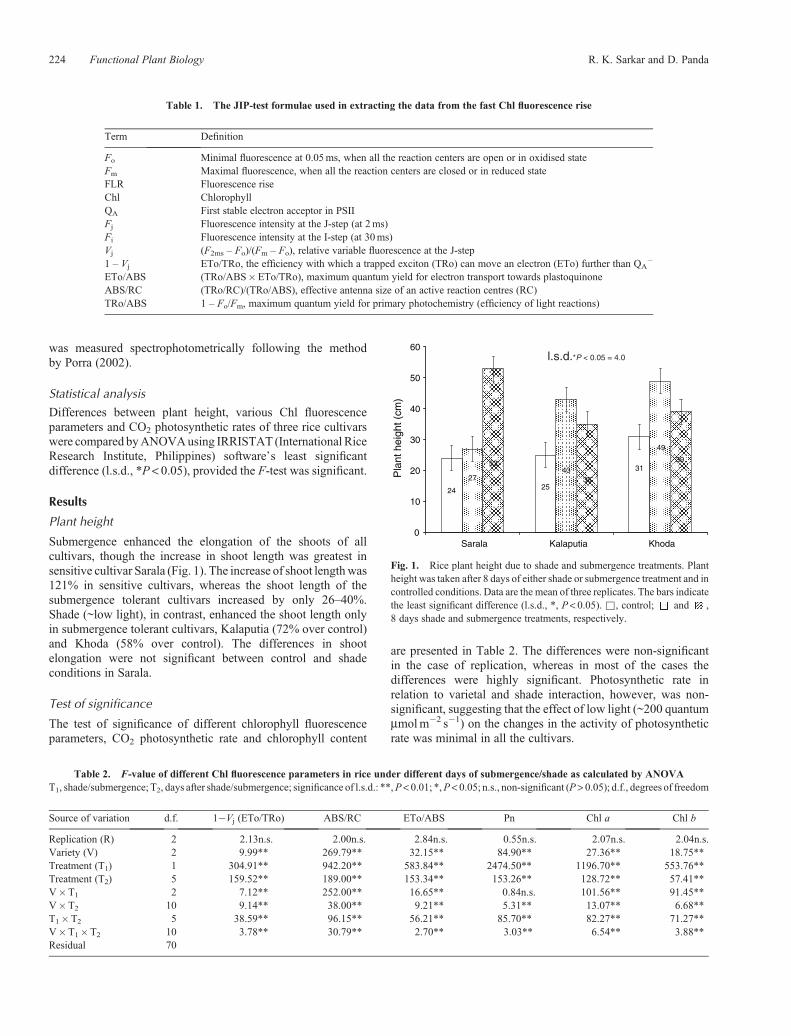
Submergence enhanced the elongation of the shoots of allcultivars, though the increase in shoot length was greatest insensitive cultivar Sarala (Fig. 1). The increase of shoot lengthwas121% in sensitive cultivars, whereas the shoot length of thesubmergence tolerant cultivars increased by only 26–40%.Shade (~low light), in contrast, enhanced the shoot length onlyin submergence tolerant cultivars, Kalaputia (72% over control)and Khoda (58% over control). The differences in shootelongation were not significant between control and shadeconditions in Sarala.
Test of significance
The test of significance of different chlorophyll fluorescenceparameters, CO2 photosynthetic rate and chlorophyll content
are presented in Table 2. The differences were non-significantin the case of replication, whereas in most of the cases thedifferences were highly significant. Photosynthetic rate inrelation to varietal and shade interaction, however, was non-significant, suggesting that the effect of low light (~200 quantummmolm�2 s�1) on the changes in the activity of photosyntheticrate was minimal in all the cultivars.
Table 1. The JIP-test formulae used in extracting the data from the fast Chl fluorescence rise
Term Definition
Fo Minimal fluorescence at 0.05ms, when all the reaction centers are open or in oxidised stateFm Maximal fluorescence, when all the reaction centers are closed or in reduced stateFLR Fluorescence riseChl ChlorophyllQA First stable electron acceptor in PSIIFj Fluorescence intensity at the J-step (at 2ms)Fi Fluorescence intensity at the I-step (at 30ms)Vj (F2ms – Fo)/(Fm – Fo), relative variable fluorescence at the J-step1 – Vj ETo/TRo, the efficiency with which a trapped exciton (TRo) can move an electron (ETo) further than QA
�
ETo/ABS (TRo/ABS�ETo/TRo), maximum quantum yield for electron transport towards plastoquinoneABS/RC (TRo/RC)/(TRo/ABS), effective antenna size of an active reaction centres (RC)TRo/ABS 1 – Fo/Fm, maximum quantum yield for primary photochemistry (efficiency of light reactions)
24 25
3127
43
49
53
35
39
0
10
20
30
40
50
60
Sarala Kalaputia Khoda
Pla
nt h
eigh
t (cm
)
l.s.d.*P < 0.05 = 4.0
Fig. 1. Rice plant height due to shade and submergence treatments. Plantheight was taken after 8 days of either shade or submergence treatment and incontrolled conditions. Data are the mean of three replicates. The bars indicatethe least significant difference (l.s.d., *, P< 0.05). &, control; and ,8 days shade and submergence treatments, respectively.
Table 2. F-value of different Chl fluorescence parameters in rice under different days of submergence/shade as calculated by ANOVAT1, shade/submergence; T2, days after shade/submergence; significance of l.s.d.: **,P< 0.01; *,P< 0.05; n.s., non-significant (P> 0.05); d.f., degrees of freedom
Source of variation d.f. 1�Vj (ETo/TRo) ABS/RC ETo/ABS Pn Chl a Chl b
Replication (R) 2 2.13n.s. 2.00n.s. 2.84n.s. 0.55n.s. 2.07n.s. 2.04n.s.Variety (V) 2 9.99** 269.79** 32.15** 84.90** 27.36** 18.75**Treatment (T1) 1 304.91** 942.20** 583.84** 2474.50** 1196.70** 553.76**Treatment (T2) 5 159.52** 189.00** 153.34** 153.26** 128.72** 57.41**V�T1 2 7.12** 252.00** 16.65** 0.84n.s. 101.56** 91.45**V�T2 10 9.14** 38.00** 9.21** 5.31** 13.07** 6.68**T1�T2 5 38.59** 96.15** 56.21** 85.70** 82.27** 71.27**V�T1�T2 10 3.78** 30.79** 2.70** 3.03** 6.54** 3.88**Residual 70
224 Functional Plant Biology R. K. Sarkar and D. Panda
CO2 photosynthetic rate
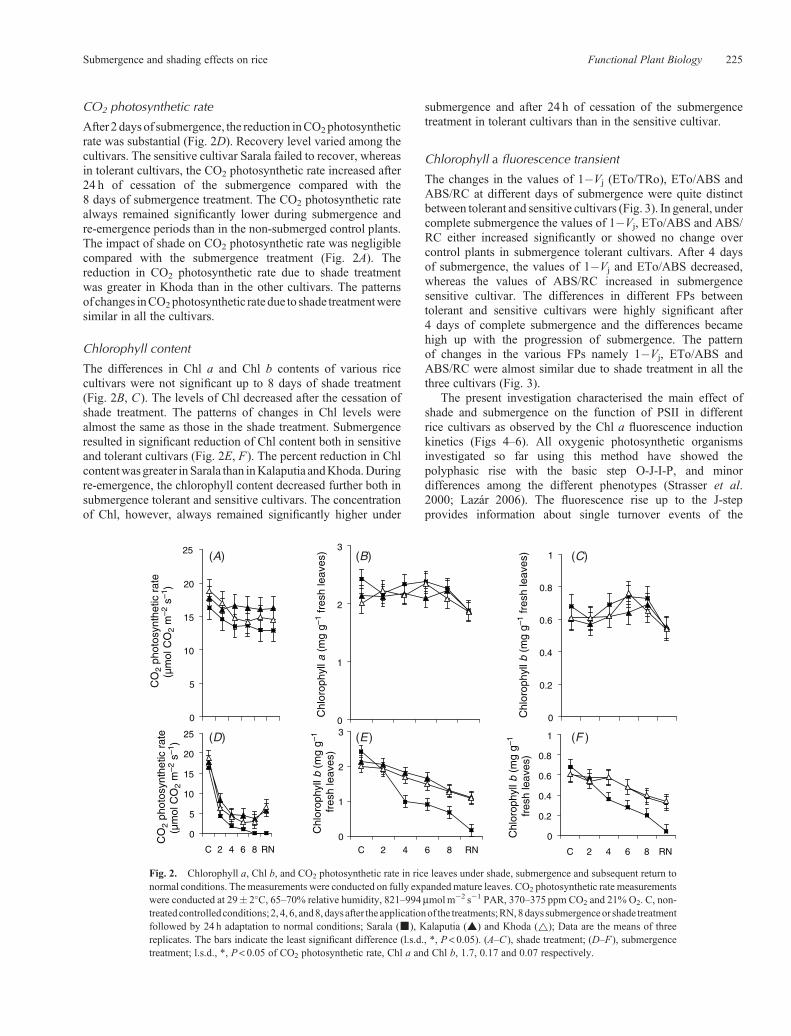
After 2 daysof submergence, the reduction inCO2photosyntheticrate was substantial (Fig. 2D). Recovery level varied among thecultivars. The sensitive cultivar Sarala failed to recover, whereasin tolerant cultivars, the CO2 photosynthetic rate increased after24 h of cessation of the submergence compared with the8 days of submergence treatment. The CO2 photosynthetic ratealways remained significantly lower during submergence andre-emergence periods than in the non-submerged control plants.The impact of shade on CO2 photosynthetic rate was negligiblecompared with the submergence treatment (Fig. 2A). Thereduction in CO2 photosynthetic rate due to shade treatmentwas greater in Khoda than in the other cultivars. The patternsof changes inCO2photosynthetic rate due to shade treatmentweresimilar in all the cultivars.
Chlorophyll content
The differences in Chl a and Chl b contents of various ricecultivars were not significant up to 8 days of shade treatment(Fig. 2B, C). The levels of Chl decreased after the cessation ofshade treatment. The patterns of changes in Chl levels werealmost the same as those in the shade treatment. Submergenceresulted in significant reduction of Chl content both in sensitiveand tolerant cultivars (Fig. 2E, F). The percent reduction in Chlcontentwasgreater inSarala than inKalaputia andKhoda.Duringre-emergence, the chlorophyll content decreased further both insubmergence tolerant and sensitive cultivars. The concentrationof Chl, however, always remained significantly higher under
submergence and after 24 h of cessation of the submergencetreatment in tolerant cultivars than in the sensitive cultivar.
Chlorophyll a fluorescence transient
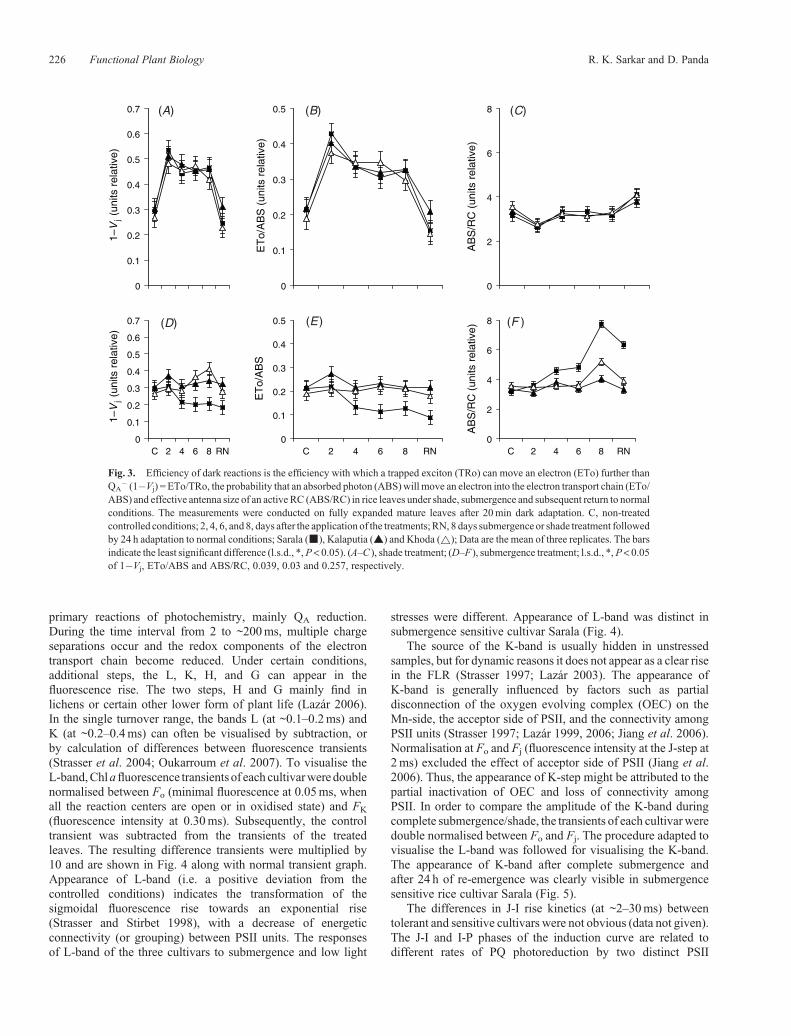
The changes in the values of 1�Vj (ETo/TRo), ETo/ABS andABS/RC at different days of submergence were quite distinctbetween tolerant and sensitive cultivars (Fig. 3). In general, undercomplete submergence the values of 1�Vj, ETo/ABS and ABS/RC either increased significantly or showed no change overcontrol plants in submergence tolerant cultivars. After 4 daysof submergence, the values of 1�Vj and ETo/ABS decreased,whereas the values of ABS/RC increased in submergencesensitive cultivar. The differences in different FPs betweentolerant and sensitive cultivars were highly significant after4 days of complete submergence and the differences becamehigh up with the progression of submergence. The patternof changes in the various FPs namely 1�Vj, ETo/ABS andABS/RC were almost similar due to shade treatment in all thethree cultivars (Fig. 3).
The present investigation characterised the main effect ofshade and submergence on the function of PSII in differentrice cultivars as observed by the Chl a fluorescence inductionkinetics (Figs 4–6). All oxygenic photosynthetic organismsinvestigated so far using this method have showed thepolyphasic rise with the basic step O-J-I-P, and minordifferences among the different phenotypes (Strasser et al.2000; Lazár 2006). The fluorescence rise up to the J-stepprovides information about single turnover events of the
0
5
10
15
20
25
CO
2 ph
otos
ynth
etic
rat
e (µ
mol
CO
2 m
–2 s
–1)
CO
2 ph
otos
ynth
etic
rat
e (µ
mol
CO
2 m
–2 s
–1)
0
5
10
15
20
25
C 2 4 6 8 RN
0
1
2
3
0
1
2
3
C 2 4 6 8 RN
0
0.2
0.4
0.6
0.8
1
Chl
orop
hyll
b (m
g g–1
fres
h le
aves
)
Chl
orop
hyll
a (m
g g–1
fres
h le
aves
)
0
0.2
0.4
0.6
0.8
1
C 2 4 6 8 RN
Chl
orop
hyll
b (m
g g–1
fr
esh
leav
es)
Chl
orop
hyll
b (m
g g–1
fres
h le
aves
)
(A) (B) (C)
(D) (E) (F )
Fig. 2. Chlorophyll a, Chl b, and CO2 photosynthetic rate in rice leaves under shade, submergence and subsequent return tonormal conditions. Themeasurements were conducted on fully expandedmature leaves. CO2 photosynthetic rate measurementswere conducted at 29� 2�C, 65–70% relative humidity, 821–994mmolm�2 s�1 PAR, 370–375 ppm CO2 and 21% O2. C, non-treatedcontrolledconditions;2, 4,6, and8,daysafter the applicationof the treatments;RN,8days submergenceor shade treatmentfollowed by 24 h adaptation to normal conditions; Sarala (&), Kalaputia (~) and Khoda (~); Data are the means of threereplicates. The bars indicate the least significant difference (l.s.d., *, P< 0.05). (A–C), shade treatment; (D–F), submergencetreatment; l.s.d., *, P< 0.05 of CO2 photosynthetic rate, Chl a and Chl b, 1.7, 0.17 and 0.07 respectively.
Submergence and shading effects on rice Functional Plant Biology 225
primary reactions of photochemistry, mainly QA reduction.During the time interval from 2 to ~200ms, multiple chargeseparations occur and the redox components of the electrontransport chain become reduced. Under certain conditions,additional steps, the L, K, H, and G can appear in thefluorescence rise. The two steps, H and G mainly find inlichens or certain other lower form of plant life (Lazár 2006).In the single turnover range, the bands L (at ~0.1–0.2ms) andK (at ~0.2–0.4ms) can often be visualised by subtraction, orby calculation of differences between fluorescence transients(Strasser et al. 2004; Oukarroum et al. 2007). To visualise theL-band,Chlafluorescence transients of each cultivarweredoublenormalised between Fo (minimal fluorescence at 0.05ms, whenall the reaction centers are open or in oxidised state) and FK
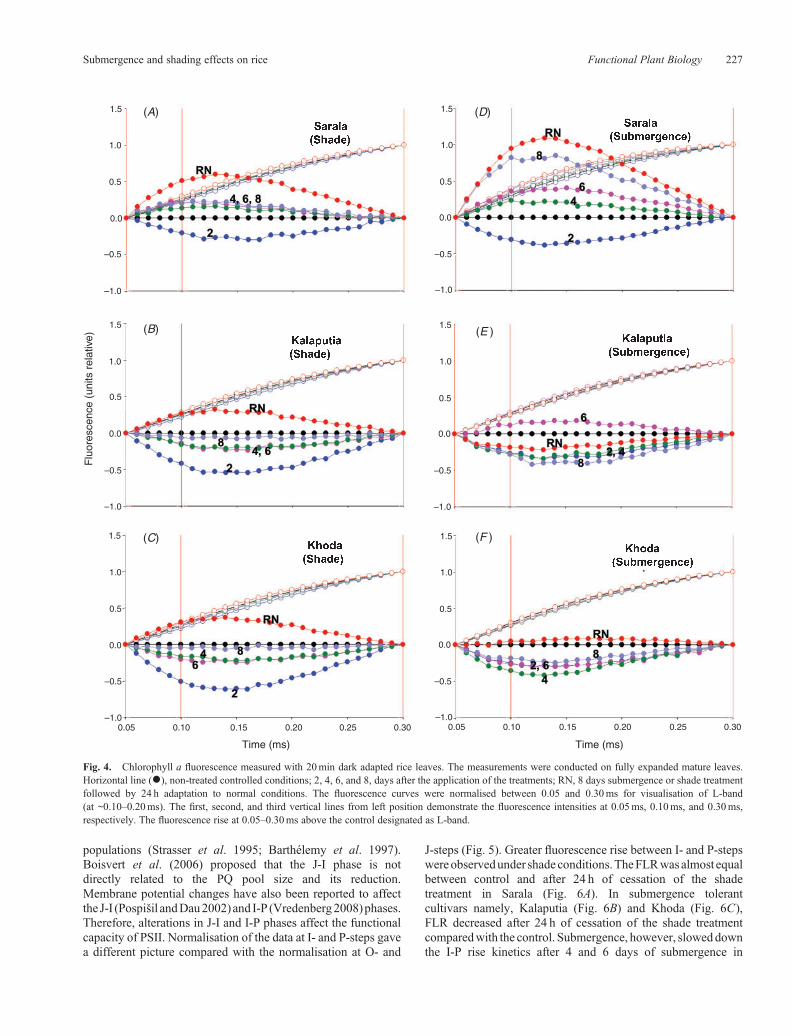
(fluorescence intensity at 0.30ms). Subsequently, the controltransient was subtracted from the transients of the treatedleaves. The resulting difference transients were multiplied by10 and are shown in Fig. 4 along with normal transient graph.Appearance of L-band (i.e. a positive deviation from thecontrolled conditions) indicates the transformation of thesigmoidal fluorescence rise towards an exponential rise(Strasser and Stirbet 1998), with a decrease of energeticconnectivity (or grouping) between PSII units. The responsesof L-band of the three cultivars to submergence and low light
stresses were different. Appearance of L-band was distinct insubmergence sensitive cultivar Sarala (Fig. 4).
The source of the K-band is usually hidden in unstressedsamples, but for dynamic reasons it does not appear as a clear risein the FLR (Strasser 1997; Lazár 2003). The appearance ofK-band is generally influenced by factors such as partialdisconnection of the oxygen evolving complex (OEC) on theMn-side, the acceptor side of PSII, and the connectivity amongPSII units (Strasser 1997; Lazár 1999, 2006; Jiang et al. 2006).Normalisation at Fo and Fj (fluorescence intensity at the J-step at2ms) excluded the effect of acceptor side of PSII (Jiang et al.2006). Thus, the appearance of K-step might be attributed to thepartial inactivation of OEC and loss of connectivity amongPSII. In order to compare the amplitude of the K-band duringcomplete submergence/shade, the transients of each cultivarweredouble normalised between Fo and Fj. The procedure adapted tovisualise the L-band was followed for visualising the K-band.The appearance of K-band after complete submergence andafter 24 h of re-emergence was clearly visible in submergencesensitive rice cultivar Sarala (Fig. 5).
The differences in J-I rise kinetics (at ~2–30ms) betweentolerant and sensitive cultivars were not obvious (data not given).The J-I and I-P phases of the induction curve are related todifferent rates of PQ photoreduction by two distinct PSII
0
0.1
0.2
0.3
0.4
0.5
0.6
0.71–
Vj (
units
rel
ativ
e)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
C 2 4 6 8 RN
1–V
j (un
its r
elat
ive)
0
0.1
0.2
0.3
0.4
0.5
ET
o/A
BS
(un
its r
elat
ive)
0
0.1
0.2
0.3
0.4
0.5
C 2 4 6 8 RN
ET
o/A
BS
0
2
4
6
8
AB
S/R
C (
units
rel
ativ
e)
0
2
4
6
8
C 2 4 6 8 RN
AB
S/R
C (
units
rel
ativ
e)
(A) (B) (C)
(D) (E ) (F )
Fig. 3. Efficiency of dark reactions is the efficiency with which a trapped exciton (TRo) can move an electron (ETo) further thanQA
� (1�Vj) = ETo/TRo, the probability that an absorbed photon (ABS)will move an electron into the electron transport chain (ETo/ABS) and effective antenna size of an active RC (ABS/RC) in rice leaves under shade, submergence and subsequent return to normalconditions. The measurements were conducted on fully expanded mature leaves after 20min dark adaptation. C, non-treatedcontrolled conditions; 2, 4, 6, and 8, days after the application of the treatments;RN, 8 days submergence or shade treatment followedby 24 h adaptation to normal conditions; Sarala (&), Kalaputia (~) and Khoda (~); Data are the mean of three replicates. The barsindicate the least significant difference (l.s.d., *,P< 0.05). (A–C ), shade treatment; (D–F ), submergence treatment; l.s.d., *,P< 0.05of 1�Vj, ETo/ABS and ABS/RC, 0.039, 0.03 and 0.257, respectively.
226 Functional Plant Biology R. K. Sarkar and D. Panda
populations (Strasser et al. 1995; Barthélemy et al. 1997).Boisvert et al. (2006) proposed that the J-I phase is notdirectly related to the PQ pool size and its reduction.Membrane potential changes have also been reported to affectthe J-I (Pospišil andDau2002) and I-P (Vredenberg2008)phases.Therefore, alterations in J-I and I-P phases affect the functionalcapacity of PSII. Normalisation of the data at I- and P-steps gavea different picture compared with the normalisation at O- and
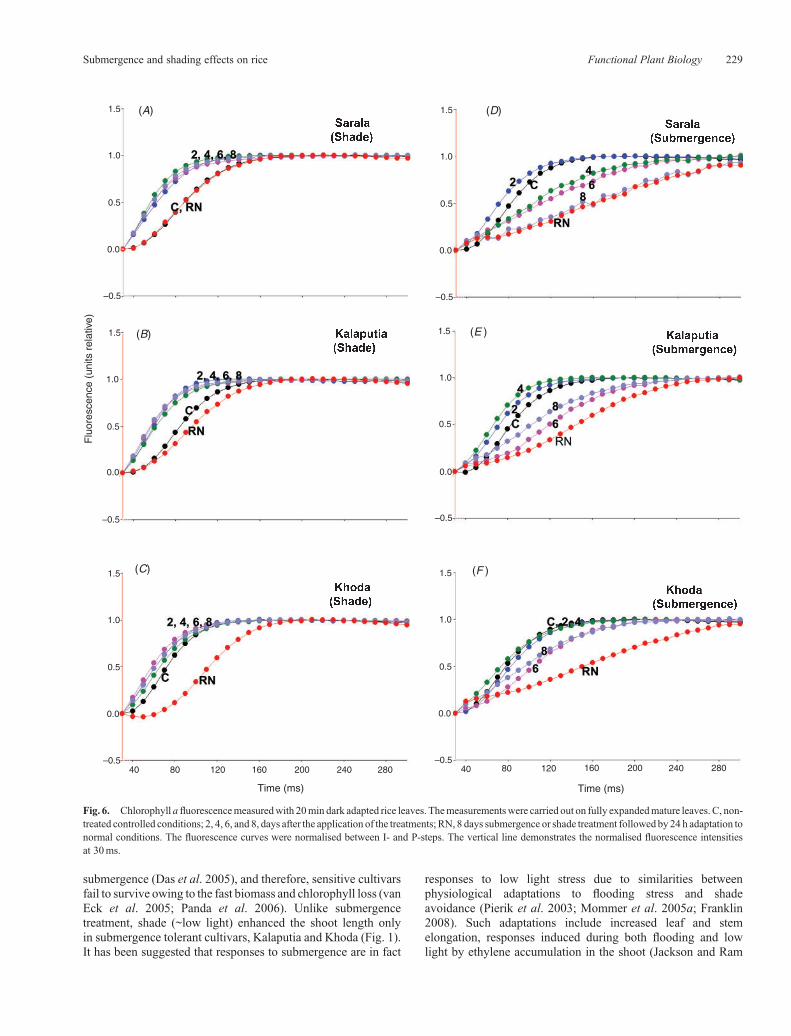
J-steps (Fig. 5). Greater fluorescence rise between I- and P-stepswereobservedunder shade conditions.TheFLRwasalmost equalbetween control and after 24 h of cessation of the shadetreatment in Sarala (Fig. 6A). In submergence tolerantcultivars namely, Kalaputia (Fig. 6B) and Khoda (Fig. 6C),FLR decreased after 24 h of cessation of the shade treatmentcomparedwith the control. Submergence, however, sloweddownthe I-P rise kinetics after 4 and 6 days of submergence in
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
0.05 0.10 0.15 0.20 0.25 0.30 0.05 0.10 0.15 0.20 0.25 0.30
Time (ms) Time (ms)
Flu
ores
cenc
e (u
nits
rel
ativ
e)
(A) (D)
(E )
(F )
(B)
(C)
Fig. 4. Chlorophyll a fluorescence measured with 20min dark adapted rice leaves. The measurements were conducted on fully expanded mature leaves.Horizontal line (*), non-treated controlled conditions; 2, 4, 6, and 8, days after the application of the treatments; RN, 8 days submergence or shade treatmentfollowed by 24 h adaptation to normal conditions. The fluorescence curves were normalised between 0.05 and 0.30ms for visualisation of L-band(at ~0.10–0.20ms). The first, second, and third vertical lines from left position demonstrate the fluorescence intensities at 0.05ms, 0.10ms, and 0.30ms,respectively. The fluorescence rise at 0.05–0.30ms above the control designated as L-band.
Submergence and shading effects on rice Functional Plant Biology 227
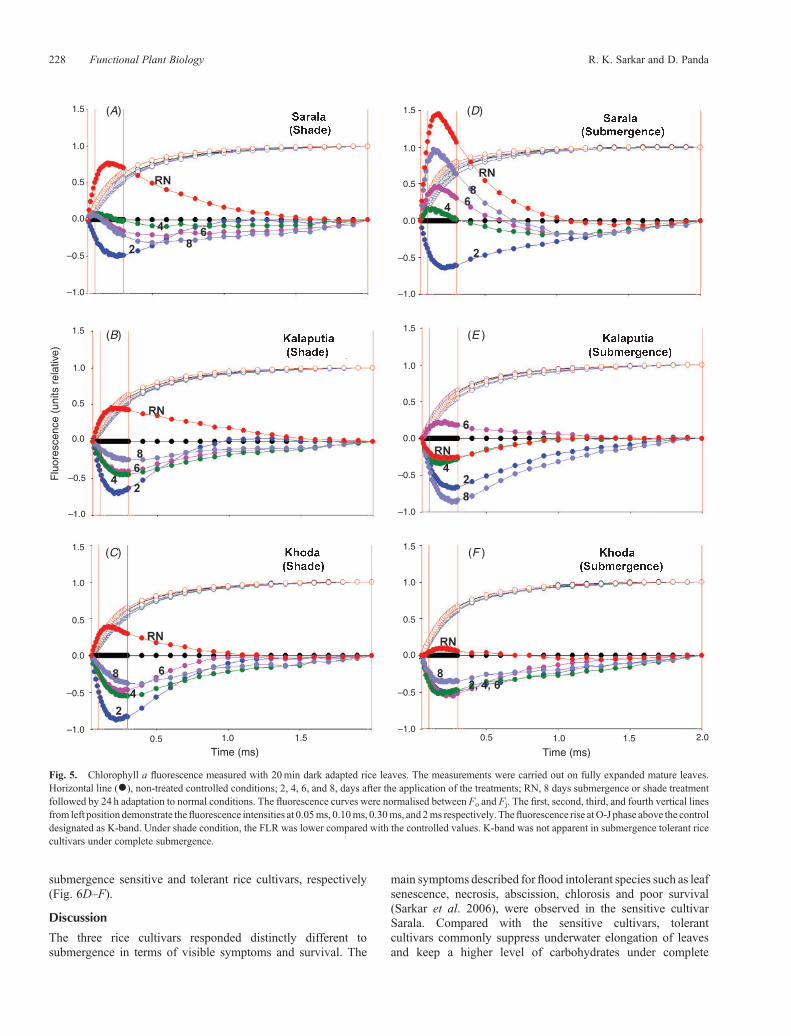
submergence sensitive and tolerant rice cultivars, respectively(Fig. 6D–F).
Discussion
The three rice cultivars responded distinctly different tosubmergence in terms of visible symptoms and survival. The
main symptoms described forflood intolerant species such as leafsenescence, necrosis, abscission, chlorosis and poor survival(Sarkar et al. 2006), were observed in the sensitive cultivarSarala. Compared with the sensitive cultivars, tolerantcultivars commonly suppress underwater elongation of leavesand keep a higher level of carbohydrates under complete
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
1.5
1.0
0.5
0.0
–0.5
–1.0
0.5 1.0 1.5 2.00.5 1.0 1.5
(A) (D)
(E )
(F )
(B)
(C)
Flu
ores
cenc
e (u
nits
rel
ativ
e)
Time (ms) Time (ms)
Fig. 5. Chlorophyll a fluorescence measured with 20min dark adapted rice leaves. The measurements were carried out on fully expanded mature leaves.Horizontal line (*), non-treated controlled conditions; 2, 4, 6, and 8, days after the application of the treatments; RN, 8 days submergence or shade treatmentfollowed by 24 h adaptation to normal conditions. The fluorescence curves were normalised between Fo and Fj. The first, second, third, and fourth vertical linesfrom left position demonstrate thefluorescence intensities at 0.05ms, 0.10ms, 0.30ms, and 2ms respectively. Thefluorescence rise atO-J phase above the controldesignated as K-band. Under shade condition, the FLR was lower compared with the controlled values. K-band was not apparent in submergence tolerant ricecultivars under complete submergence.
228 Functional Plant Biology R. K. Sarkar and D. Panda
submergence (Das et al. 2005), and therefore, sensitive cultivarsfail to survive owing to the fast biomass and chlorophyll loss (vanEck et al. 2005; Panda et al. 2006). Unlike submergencetreatment, shade (~low light) enhanced the shoot length onlyin submergence tolerant cultivars, Kalaputia and Khoda (Fig. 1).It has been suggested that responses to submergence are in fact
responses to low light stress due to similarities betweenphysiological adaptations to flooding stress and shadeavoidance (Pierik et al. 2003; Mommer et al. 2005a; Franklin2008). Such adaptations include increased leaf and stemelongation, responses induced during both flooding and lowlight by ethylene accumulation in the shoot (Jackson and Ram
1.5
1.0
0.5
0.0
–0.5
1.5
1.0
0.5
0.0
–0.5
1.5
1.0
0.5
0.0
–0.5
1.5
1.0
0.5
0.0
–0.5
1.5
1.0
0.5
0.0
–0.5
1.5
1.0
0.5
0.0
–0.5
40 80 120 160 200 240 280 40 80 120 160 200 240 280
(A) (D)
(E )
(F )
(B)
(C)
Flu
ores
cenc
e (u
nits
rel
ativ
e)
Time (ms) Time (ms)
Fig. 6. Chlorophyll afluorescencemeasuredwith 20min dark adapted rice leaves. Themeasurementswere carried out on fully expandedmature leaves. C, non-treated controlled conditions; 2, 4, 6, and 8, days after the application of the treatments; RN, 8 days submergence or shade treatment followed by 24 h adaptation tonormal conditions. The fluorescence curves were normalised between I- and P-steps. The vertical line demonstrates the normalised fluorescence intensitiesat 30ms.
Submergence and shading effects on rice Functional Plant Biology 229

2003; Pierik et al. 2005). Generally, flooding tolerant cultivarsexhibit little or no elongation during complete submergencewhereas the cultivars avoid flooding tended to elongate greaterand greater, avoid complete submergence; suitable for deepwater(>50 cmwater depth) or floating rice ecology (Sarkar et al. 2006).Blocking the endogenous synthesis of ethylene or GAsubstantially improved the survival of sensitive rice cultivarsdue to greater inhibition of elongation growth (Ella et al. 2003;Das et al. 2005). It has been proposed that submergence tolerancein lowland rice is conferred by a specific allele variant of Sub1Athat diminishes ethylene production and GA responsiveness,causing quiescence in growth that correlates with the capacityfor re-growth upon de-submergence (Xu et al. 2006; Bailey-Serres and Voesenek 2008). The greater elongation ability ofsensitive cultivar under complete submergence suggests that thiscultivar might have greater ethylene level or sensitivity andhad resemblance with shade avoidance of other plant speciese.g. Veronica anagallis-aquatica, Rumex palustris (Boeger andPoulson 2003; Mommer et al. 2005a). A different scenario inplant elongation under low light condition compared with thesubmerged condition suggested that probably under shade (~lowlight), signal transduction and perception related to theelongation growth was quite different between submergencetolerant and sensitive cultivars. The ethylene responsive factor(ERF) proteins have important functions in the transcriptionalregulation of a variety of biological processes related to growthand development, as well as various responses to environmentalstimuli including submergence (Nakano et al. 2006; Jackson2008). Probably, the ERF gene family of tolerant cultivarsresponded in a greater extent under shade conditions, and,hence, a greater elongation was noticed in tolerant cultivarscompared with the sensitive cultivar (Fig. 1). In eastern India,there is a prevalence of low light during the rainy season. Thesubmergence sensitive cultivar, Sarala, attained the height of102–108 cm, whereas tolerant cultivars, Kalaputia and Khoda,attained the height of more than 160 cm during the crop growingperiod (data not shown). This shows that Sarala, a high yieldingrice cultivar, was more adaptive to low light conditions in termof lesser extension growth than the submergence tolerantcultivars, Kalaputia and Khoda.
Tolerant cultivars maintained a higher level of both Chl a andbduring submergence and the subsequent period of re-emergencethan the non-tolerant cultivar (Fig. 2). Chl contents decreasedduring submergence and failed to recover following a return tonon-submerged conditions (Sarkar et al. 2001). The patterns ofchanges inChl levels at lower light intensitywere almost the samein three rice cultivars, suggesting that submergence tolerantcultivars need not be tolerant to low light stress. In the presentinvestigation, light intensity at the vicinity of canopy was almostsimilar both under shade and submerged conditions. The greaterchanges in the FPs or greater damage to PSII structural andfunctional capacity in sensitive cultivar Sarala under completesubmergence compared with the shade conditions could berecognised as greater effects of submergence, rather thancombining effects of both shade and submergence. The rapiddrop inCO2photosynthetic rate under submergencewasprobablydue to the structural damage suffered by the photosyntheticapparatus, as evident from different chlorophyll fluorescencecharacteristics. In addition, an irreversible loss in Rubisco
activity (Panda et al. 2008) and plant pigment content mightbe responsible for a poor CO2 photosynthetic rate (Fig. 2).Though there were some aberrations in PSII structural andfunctional activities as evident by chlorophyll fluorescencecharacteristics, the change in CO2 photosynthetic rate was notas severe under shade as under submerged conditions. Thechanges in fluorescence characteristics under low light (~200quantummmolm�2 s�1) conditions were not always in damagingdirection (Figs 3–6). The changes in pigment compositionresultant from the shade treatment were also minimal (Fig. 2).Acclimation of photosynthesis in rice saturates at a relativelylow irradiance (50–200mmolm�2 s�1) (Murchie et al. 2002).Therefore, rice leaves have a considerable flexibility that enablesthem toperformeffectively in a rangeof light environments.Afterthe first day of re-emergence, the CO2 photosynthetic rate startedincreasing in tolerant cultivars, whereas after removal of shadetreatment the changes in CO2 photosynthetic rates were minimalcompared with the 8 days of shade treatment (Fig. 2). Recoveryof photosynthetic activity after complete submergence/shadealso found in other submergence tolerant wetland species,e.g. Phragmites australis (Mauchamp et al. 2001) andEleocharis cellulosa (Macek et al. 2006) or shade tolerant,Posidonia oceanic (Figueroa et al. 2002). Under controlledconditions, the values of different FPs did not differsignificantly between tolerant and sensitive cultivars (Fig. 3).This suggests that responses of both the tolerant and sensitivecultivars were similar to non-stress conditions.
All oxygenic photosynthetic organisms show thecharacteristic changes in O-J-I-P transient (JIP-test) (Strasseret al. 2000, 2004). The JIP-test developed by Strasser andStrasser (1995) is used to translate several biophysicalexpressions that quantify PSII function. Shade caused anincrease in the efficiency of the trapping rate of excitons (TRo)tomove an electron further thanQA
� as evident from the increasein 1 – Vj (= ETo/TRo) values (Fig. 3). The efficiency of themovement of an electron per absorbed photon (ETo/ABS)through the electron transport chain also increased due toshade. The present investigation suggests that under shade[light intensity, ~200 quanta (mmolm�2 s�1)] rice plants utilisethe absorbed energy in an efficient manner compared with thecontrolled plants [light intensity, ~2000 quanta (mmolm�2 s�1)].Under submergence, however, the regulatory mechanism inutilising the absorbed energy was effective only in the tolerantcultivars comparable with controlled plants (Fig. 3). The increasein ABS/RC is due to a decrease in active QA reducing reactioncentres (Strasser and Stirbet 1998). Non-QA reducing reactioncentres exist always in light grown plants acting as heat sink orsilent RCs. This function acts as a heat radiator and protects theplant from high temperature and high light (Strasser et al. 2004).Under submergence though the temperature and light intensitywas not high, yet the damage of the photosynthetic apparatusoccurred. The observation that values in ABS/RC did not changegreatly during submergence in tolerant cultivars compared withthe control sets suggests that tolerant cultivars were better able toregulate the amount of light reaching theRC (Strasser et al. 2004).After 24 h of cessation of the shade treatment, the values of ABS/RC significantly increased in all the three cultivars, suggestingthat all the three cultivars acted in the same mode to protect fromhigh temperature and high light. The values ofABS/RC after 24 h
230 Functional Plant Biology R. K. Sarkar and D. Panda

of cessation of submergence treatment were comparable withcontrol sets in tolerant cultivars.
The progressive reduction of the acceptor side of PSII leads tothree distinct major phases of fluorescence rise from O to P withtwo intermediate steps, J (I1) and I (I2) (Strasser and Govindjee1992; Samson et al. 1999; Boisvert et al. 2006). Most studiesagree that O-J rise is related to PSII primary acceptor (QA)reduction, but several contradictory theories exist for the J-Iand I-P rises (Strasser et al. 1995; Joly et al. 2005; Lazár2006; Vredenberg 2008). Oukarroum et al. (2007) observed arise between Fo (0.05ms) and FK (0.30ms) and at ~0.2–0.4msand termed as L- and K-bands, respectively, and consideredOLKJ as photochemical phase. Changes in respectively theL- and K-bands of the fluorescence transients OJIP conditionswere shown to have predictive value with respect to the vitalityof leaves (Oukarroum et al. 2007). In Sarala (submergencesensitive), the energetic connectivity decreased in response toboth submergence and low light stress, and compared with thesubmergence tolerant cultivars, Kalaputia and Khoda (Fig. 4), itseems that submergence tolerant cultivars maintain the energeticconnectivity between PSII units in greater extent compared withthe submergence insensitive cultivar both under submergenceand low light stresses. An increase in J-step and decrease ofgrouping probability due to submergence as reported earlier(Panda et al. 2006) supported this contention that like hightemperature (Lazár 2006), or drought (Oukarroum et al. 2007),both L- and K-bands appeared due to submergence stress incultivar sensitive to submergence (Figs 4, 5). The results showthat several JIP test parameters, includingL- andK-bands, clearlydifferentiate between submergence tolerant and sensitive ricecultivars. Damage of PSII as visualising through K-band wasnot apparent under low light (~200 quanta mmolm�2 s�1)conditions (Fig. 5). Both L- and K-bands appeared in allthe rice cultivars after cessation of the shade treatment,suggesting that, after returning to normal condition (~highlight) from shade (~low light), damage of PSII occurred in alltypes of rice cultivars.
Our data show that both submergence tolerant and sensitiverice cultivars are equally sensitive to both shade (~low light) andsubmergence (low light with submergence) with respect to J-Ifluorescence rise (data not shown). The FLR at I-P phase,however, increased under shade condition compared with thecontrolled conditions in all rice cultivars (Fig. 6). Strasser et al.(1995) proposed that the release of fluorescence quenchingduring the (J-I) phase is controlled by the PSII donor side(water splitting activity) and the third phase (I-P) correspondsto the release of fluorescence quenching by the oxidised PQpool. Submergence slowed down the I-P rise kinetics after 4and 6 days of submergence in sensitive and tolerant cultivars,respectively. Hence, tolerant cultivars maintained the functionalability of chloroplast for longer period under completesubmergence.
Flooding tolerance vis-à-vis avoidance, appeared to be twoopposite events. The rice species tolerant to completesubmergence showed reduced elongation under completesubmergence, whereas these cultivars showed greaterelongation under low light stress. In literature, acclimation tosubmergence has been hypothesised to be an acclimation to shade(Mommer et al. 2006; Voesenek et al. 2006). This is because
of apparent similarity of simultaneous elongation of leavesand stems due to both low light and submergence stresses.This intention is applicable for those rice cultivars adapted towater-logging conditions (Sarkar et al. 2006). However,reduced elongation under flash-flood conditions (completesubmergence for 1–2 weeks) is vital for survival and plantproductivity, because elongating plants would tend to lodge assoon as the water level recedes. Minimal changes or, in otherwords, protection of vital plant characteristics rather thangreater alteration for acclimation under submergence aftershort-term exposure (e.g. ~1 week) is more beneficial for plantproductivity. This investigation suggests that tolerance tosubmergence is not necessarily a means of tolerance to lowlight stress. Our results also show that a distinction betweensubmergence tolerant and sensitive rice cultivars could be madeby analysing the JIP-test.
References
Bailey-Serres J, Voesenek LACJ (2008) Flooding stress: acclimations andgenetic diversity. Annual Review of Plant Biology 59, 313–339.doi: 10.1146/annurev.arplant.59.032607.092752
BarthélemyX, Popovic R, Franck F (1997) Studies on theO-J-I-P transient ofchlorophyll fluorescence in relation to photosystem II assembly andheterogeneity in plastids of greening barley. Journal of PhotochemistryandPhotobiology.B,Biology39, 213–218.doi: 10.1016/S1011-1344(97)00012-2
Boeger MRT, Poulson ME (2003) Morphological adaptations andphotosynthesis rates of amphibious Veronica anagallis-aquaticaL. (Scrophulariaceae) under different flow regimes. Aquatic Botany 75,123–135. doi: 10.1016/S0304-3770(02)00174-2
Boisvert S, Joly D, Carpentier R (2006) Quantitative analysis of theexperimental O-J-I-P chlorophyll fluorescence induction kinetics:apparent activation energy and origin of each kinetic step. FEBSJournal 273, 4770–4777. doi: 10.1111/j.1742-4658.2006.05475.x
Colmer TD, Pedersen O (2008a) Oxygen dynamics in submerged rice(Oryza sativa). New Phytologist 178, 326–334. doi: 10.1111/j.1469-8137.2007.02364.x
Colmer TD, Pedersen O (2008b) Underwater photosynthesis and respirationin leaves of submerged wetland plants: gas films improve CO2 and O2
exchange. New Phytologist 177, 918–926. doi: 10.1111/j.1469-8137.2007.02318.x
Das KK, Sarkar RK, Ismail AM (2005) Elongation ability and non-structuralcarbohydrate levels in relation to submergence tolerance in rice. PlantScience 168, 131–136. doi: 10.1016/j.plantsci.2004.07.023
Ella ES, Kawano N, Yamauchi Y, Tanaka K, Ismail AM (2003) Blockingethylene perception enhances flooding tolerance in rice seedlings.Functional Plant Biology 30, 813–819. doi: 10.1071/FP03049
Figueroa FL, Jiménez C, Viñegla B, Pérez-Rodríguez E, Aguilera J,Flores-Moya A, Altamirano M, Lebert M, Häder DP (2002) Effects ofsolar UV radiation on photosynthesis of the marine angiospermPosidonia oceanica from southern Spain. In ‘Marine ecologyprogress series. Vol. 230’. (Ed. O Kinne) pp. 59–72. (Oldendorf/Luhe: Germany)
Franklin KA (2008) Shade avoidance. New Phytologist 179, 930–944.doi: 10.1111/j.1469-8137.2008.02507.x
Jackson MB (2008) Ethylene-promoted elongation: an adaptation tosubmergence stress. Annals of Botany 101, 229–248. doi: 10.1093/aob/mcm237
Jackson MB, Ram PC (2003) Physiological and molecular basis ofsusceptibility and tolerance of rice plants to complete submergence.Annals of Botany 91, 227–241. doi: 10.1093/aob/mcf242
Submergence and shading effects on rice Functional Plant Biology 231

Jiang C-D, Shi L, Gao H-Y, Schansker G, Toth SZ, Strasser RJ (2006)Development of photosystems 2 and 1 during leaf growth in grapevineseedlings probed by chlorophyll a fluorescence transient and 820 nmtransmission in vivo. Photosynthetica 44, 454–463. doi: 10.1007/s11099-006-0050-5
Joly D, Bigras C, Harnois J, Govindachary S, Carpentier R (2005) Kineticanalyses of the OJIP chlorophyll fluorescence rise in thylakoidmembranes. Photosynthesis Research 84, 107–112. doi: 10.1007/s11120-004-7763-8
Lazár D (1999) Chlorophyll a fluorescence induction. Biochimica etBiophysica Acta – Bioenergetics 1412, 1–28. doi: 10.1016/S0005-2728(99)00047-X
Lazár D (2003) Chlorophyll a fluorescence rise induced by high lightillumination of dark-adapted plant tissue studied by means of amodel of photosystem II and considering photosystem II heterogeneity.Journal of Theoretical Biology 220, 469–503. doi: 10.1006/jtbi.2003.3140
Lazár D (2006) The polyphasic chlorophyll a fluorescence rise measuredunder high intensity of exciting light. Functional Plant Biology 33, 9–30.doi: 10.1071/FP05095
Macek P, Rejma’nkova’ E, Houdkova’ K (2006) The effect of long-termsubmergence on functional properties of Eleocharis cellulosaTorr. Aquatic Botany 84, 251–258. doi: 10.1016/j.aquabot.2005.11.003
Mauchamp A, Blanch S, Grillas P (2001) Effects of submergence on thegrowth of two Phragmites australis seedlings. Aquatic Botany 69,147–164. doi: 10.1016/S0304-3770(01)00135-8
Mommer L, Visser EJW (2005) Underwater photosynthesis in floodedterrestrial plants: a matter of leaf plasticity. Annals of Botany 96,581–589. doi: 10.1093/aob/mci212
Mommer L, de Kroon H, Pierik R, Bo��gemann GM, Visser EJW (2005a)A functional comparison of acclimation to shade and submergencein two terrestrial plant species. New Phytologist 167, 197–206.doi: 10.1111/j.1469-8137.2005.01404.x
Mommer L, Pons TL, Wolters-Arts M, Venema JH, Visser EJW (2005b)Submergence-induced morphological, anatomical and biochemicalresponses in a terrestrial species affect gas diffusion resistance andphotosynthetic performance. Plant Physiology 139, 497–508.doi: 10.1104/pp.105.064725
Mommer L, Pons TL, Visser EJW (2006) Photosynthetic consequences ofphenotypic plasticity in response to submergence: Rumex palustris as acase study. Journal of Experimental Botany 57, 283–290. doi: 10.1093/jxb/erj015
Mommer L, Wolters-Arts M, Andersen C, Visser EJW, Pedersen O (2007)Submergence-induced leaf acclimation in terrestrial species varying inflooding tolerance. New Phytologist 176, 337–345. doi: 10.1111/j.1469-8137.2007.02166.x
Murchie EH, Hubbart S, Chen Y, Peng S, Horton P (2002) Acclimation ofrice photosynthesis to irradiance under field conditions. Plant Physiology130, 1999–2010. doi: 10.1104/pp.011098
Nakano T, Suzuki K, Fujimura T, Shinshi H (2006) Genome-wide analysis ofthe ERF gene family in Arabidopsis and rice. Plant Physiology 140,411–432. doi: 10.1104/pp.105.073783
Oukarroum A, Madidi SE, Schansker G, Strasser RJ (2007) Probing theresponses of barley cultivars (Hordeum vulgare L.) by chlorophyll afluorescence OLKJIP under drought stress and re-watering.Environmental and Experimental Botany 60, 438–446. doi: 10.1016/j.envexpbot.2007.01.002
Panda D, Rao DN, Sharma SG, Strasser RJ, Sarkar RK (2006) Submergenceeffect on rice genotypes during seedling stage: probing of submergencedriven changes of photosystem 2 by chlorophyll a fluorescence inductionO-J-I-P transients.Photosynthetica 44, 69–75. doi: 10.1007/s11099-005-0200-1
Panda D, Sharma SG, Sarkar RK (2008) Chlorophyll fluorescenceparameters, CO2 photosynthetic rate and regeneration capacity as aresult of complete submergence and subsequent re-emergence in rice(Oryza sativa L.). Aquatic Botany 88, 127–133. doi: 10.1016/j.aquabot.2007.08.012
Pierik R, Visser EJW, de Kroon H, Voesenek LACJ (2003) Ethylene isrequired in tobacco to successfully complete with proximate neighbours.Plant, Cell & Environment 26, 1229–1234. doi: 10.1046/j.1365-3040.2003.01045.x
Pierik R, Millenaar FF, Peeters AJM, Voesenek LACJ (2005) Newperspectives in flooding research: the use of shade avoidance andArabidopsis thaliana. Annals of Botany 96, 533–540. doi: 10.1093/aob/mci208
Porra RJ (2002) The chequered history of the development and use ofsimultaneous equations for accurate determination of chlorophylls aand b. Photosynthesis Research 73, 149–156. doi: 10.1023/A:1020470224740
Pospišil P, Dau H (2002) Valinomycin sensitivity proves that light-inducedthylakoid voltages result inmillisecond phase of chlorophyllfluorescencetransients. Biochimica et Biophysica Acta – Bioenergetics 1554, 94–100.doi: 10.1016/S0005-2728(02)00216-5
SamsonG,PrášilO,YaakoubdB (1999)Photochemical and thermal phasesofchlorophyll a fluorescence. Photosynthetica 37, 163–182. doi: 10.1023/A:1007095619317
Sarkar RK, De RN, Reddy JN, Ramakrishnayya G (1996) Studies on thesubmergence tolerance mechanism in relation to carbohydrate,chlorophyll and specific leaf weight in rice (Oryza sativa L.). Journalof Plant Physiology 149, 623–625.
Sarkar RK, Das S, Ravi I (2001) Changes in certain antioxidative enzymesand growth parameters as a result of complete submergence andsubsequent re-aeration of rice cultivars differing in submergencetolerance. Journal Agronomy & Crop Science 187, 69–74. doi: 10.1046/j.1439-037X.2001.00510.x
Sarkar RK, Reddy JN, Sharma SG, Ismail AM (2006) Physiological basis ofsubmergence tolerance in rice and implications for crop improvement.Current Science 91, 899–906.
Setter TL, Ellis M, Laureles EV, Ella ES, Senadhira D, Mishra SB,Sarkarung S, Datta S (1997) Physiology and genetics of submergencetolerance in rice. Annals of Botany 79, 67–77. doi: 10.1006/anbo.1996.0304
Strasser BJ (1997) Donor side capacity of photosystem II probed bychlorophyll a fluorescence transients. Photosynthesis Research 52,147–155. doi: 10.1023/A:1005896029778
Strasser RJ, Govindjee (1992) The Fo and O-J-I-P fluorescence rise inhigher plants and algae. In ‘Regulation of chloroplast biogenesis’.(Ed. JH Argyroudi-Akoyunogglou) pp. 423–426. (Plenum Press:New York)
Strasser RJ, Stirbet A (1998) Heterogeneity of photosystem II probed by thenumerically simulated chlorophyll a fluorescence rise (O-J-I-P).Mathematics and Computers in Simulation 48, 3–9. doi: 10.1016/S0378-4754(98)00150-5
Strasser BJ, Strasser RJ (1995) Measuring fast fluorescence transients toaddress environmental questions: the JIP test. In ‘Photosynthesis: fromlight to biosphere. Vol. V’. (Ed. P Mathis) pp. 977–980. (KluwerAcademic Publishers: Dordrecht, the Netherlands)
Strasser RJ, Srivastava A, Govindjee (1995) Polyphasic chlorophyll afluorescence transient in plants and cyanobacteria. Photochemistry andPhotobiology 61, 32–42. doi: 10.1111/j.1751-1097.1995.tb09240.x
Strasser RJ, Srivastava A, Tsimilli-Michael M (2000) The fluorescencetransient as a tool to characterize and screen photosynthetic sample.In ‘Probing photosynthesis: mechanism, regulation and adaptation’.(Eds M Yunus, U Pathre, P Mohanty) pp. 445–483. (Taylor & Francis:London)
232 Functional Plant Biology R. K. Sarkar and D. Panda

Strasser RJ, Srivastava A, Tsimilli-Michael M (2004) Analysis of thechlorophyll a fluorescence transient. In ‘Advances in photosynthesisand respiration’. (Eds G Papageorgiou, Govindjee) pp. 321–362.(Kluwer Academic Publishers: Dordrecht, the Netherlands)
van Eck WHJM, Enssen JPM, Rengelink RHJ, Blom CWPM, de Kroon H(2005) Water temperature instead of acclimation stage and oxygenconcentration determines responses to winter floods. Aquatic Botany81, 253–264. doi: 10.1016/j.aquabot.2004.10.006
Vartapetian BB (2006) Plant anaerobic stress as a novel trend in ecological,physiology, biochemistry, andmolecular biology: 2. Further developmentof the problem. Russian Journal of Plant Physiology: a ComprehensiveRussian Journal onModern Phytophysiology 53, 711–738. doi: 10.1134/S102144370606001X
Vervuren PJA, Beurskens SMJH, Blom CWPM (1999) Light acclimation,CO2 response and long-trm capacity of underwater photosynthesis inthree terrestrial plant species. Plant, Cell & Environment 22, 959–968.doi: 10.1046/j.1365-3040.1999.00461.x
Voesenek LACJ, Colmer TD, Pierik R, Millenaar FF, Peeters AJM (2006)How plants cope with complete submergence. New Phytologist 170,213–226. doi: 10.1111/j.1469-8137.2006.01692.x
Vredenberg WJ (2008) Algorithm for analysis of OJDIP fluorescenceinduction curves in terms of photo-and electrochemical events inphotosystems of plant cells: derivation and application. Journalof Photochemistry and Photobiology. B, Biology 91, 58–65.doi: 10.1016/j.jphotobiol.2008.01.005
Xu K, Xia X, Fukao T, Canlas P, Maghirang-Rodriguez R, Heuer S,Ismail AM, Bailey-Serres J, Ronald PC, Mackill DJ (2006) Sub1A isan ethylene response factor-like gene that confers submergencetolerance to rice. Nature 442, 705–708. doi: 10.1038/nature04920
Manuscript received 8 August 2008, accepted 11 December 2008
Submergence and shading effects on rice Functional Plant Biology 233
http://www.publish.csiro.au/journals/fpb
Copyright © 2022 FDOKUMEN




















