
Reinforcing Sustainability Pillars for Robust Growth - Intiland
Upload
independentCategory
view
1download
0
Psychopharmacology (2006) 184: 391–400DOI 10.1007/s00213-005-0183-4
ORIGINAL INVESTIGATION
Matthew I. Palmatier . F. Fay Evans-Martin . Alycia Hoffman .Anthony R. Caggiula . Nadia Chaudhri . Eric C. Donny . Xiu Liu .Sherri Booth . Maysa Gharib . Laure Craven . Alan F. Sved
Dissociating the primary reinforcingand reinforcement-enhancing effects of nicotine usinga rat self-administration paradigm with concurrently availabledrug and environmental reinforcersReceived: 25 April 2005 / Accepted: 29 August 2005 / Published online: 25 October 2005# Springer-Verlag 2005
Abstract Rationale: Nicotine has two effects on rein-forcement in traditional self-administration paradigms. Itserves as a primary reinforcer by increasing the probabilityof behaviors that result in nicotine delivery. However,nicotine also potently enhances behaviors that result in thedelivery of nonpharmacological reinforcers. Objectives: Thepresent study sought to dissociate these two effects ofnicotine on reinforcement. Methods: For one group of rats(2 lever), a nonpharmacological reinforcer [visual stimulus(VS)] was available for pressing one lever. Nicotineinfusions were available for pressing a different lever. Asecond group (NIC + VS) received more traditional self-administration training; both the VS and nicotine weredelivered for pressing a single active lever. Control groupsreceived either nicotine infusions (NIC only) or VSpresentations (VS only) for pressing the active lever.Results: Nicotine alone was a weak reinforcer; the VSalone was slightly more reinforcing than nicotine. Whenthese two reinforcers were combined (NIC + VS), responserates were synergistically increased. For the 2-lever group,responding on the nicotine lever was weak, matching theresponse rates of rats receiving nicotine alone. However,
responding on the VS lever was potently enhanced in thisgroup; equaling the response rates for rats receiving bothreinforcers for making a single response (NIC + VS).Conclusions: These data indicate that the reinforcement-enhancing effects of nicotine are very potent even whenonly moderate quantities of the drug are self-administered.Moreover, they provide the first demonstration that thereinforcement-enhancing and primary reinforcing effects ofnicotine can be dissociated behaviorally.
Keywords Nicotine . Self-administration .Reinforcement . Concurrent schedule . Stimuli .Acetylcholine
Introduction
Smoking tobacco results in rapid delivery of nicotine to thebrain, and the resulting pharmacological action is thoughtto strengthen or “reinforce” subsequent smoking behavior(US Department of Health and Human Services (USDHHS)1988). This view is consistent with a large body of researchwith humans and animals, indicating that nicotine can act asa primary reinforcer, increasing the frequency of variousbehaviors on which nicotine delivery is made contingent.For example, humans, nonhuman primates, and rodentswill learn to perform a response, such as pressing a lever, toreceive intravenous (i.v.) nicotine infusions (e.g., Corrigalland Coen 1994; Goldberg et al. 1981; Harvey et al. 2004).However, in recent years, researchers have come to realizethat non-nicotine stimuli are also important in maintainingcompulsive smoking behavior (e.g., Rose et al. 2000). Forexample, craving or “desire to smoke” is heightened byviewing a lit cigarette in an ashtray or seeing other in-dividuals smoking (Perkins et al. 1994; Niaura et al. 1998).Moreover, neither i.v. nicotine nor the smoking ofdenicotinized cigarettes completely relieves the desire tosmoke in briefly abstinent smokers. Craving is completelyabolished only when subjects smoke denicotinized ciga-
M. I. Palmatier (*) . F. F. Evans-Martin . A. Hoffman .A. R. Caggiula . E. C. Donny . X. Liu . S. Booth .M. Gharib . L. CravenDepartment of Psychology, University of Pittsburgh,3137 Sennott Square, 210 S. Bouquet St.,Pittsburgh, PA 15260, USAe-mail: [email protected].: +1-412-6247345Fax: +1-412-6248558
M. I. Palmatier . N. Chaudhri . A. F. SvedDepartment of Neuroscience, University of Pittsburgh,446 Crawford Hall, 5th and Ruskin,Pittsburgh, PA 15260, USA
Present address:N. Chaudhri .Ernest Gallo Clinic and Research Center,University of California at San Francisco,San Francisco, CA, USA

rettes while receiving an i.v. nicotine infusion (Rose et al.2000), suggesting that both nicotine and nonnicotine stim-uli are critical in tobacco dependence.
Ongoing preclinical research supports the notion thatnonpharmacological (i.e., environmental) stimuli contrib-ute heavily to nicotine addiction (Cohen et al. 2005; Donnyet al. 2003; Le Foll and Goldberg 2005). Recently, we haveargued that nicotine self-administration in rodents reflectstwo dissociable actions of the drug. First, nicotine canfunction as a primary reinforcer of operant behavior; ratswill respond for nicotine in the absence of nonpharmaco-logical stimuli, albeit at very low rates (e.g., Caggiula et al.2002). Second, nicotine also enhances responding for otherreinforcing stimuli, an action that does not require acontingent relationship between drug delivery and operantbehavior. This second effect of nicotine was demonstratedin a series of studies that manipulated the contingenciesbetween operant behavior and the delivery of nicotineinfusions and/or a nonpharmacological visual stimulus(VS; Chaudhri et al. 2005a; Donny et al. 2003). In thisparadigm, the VS served as a primary reinforcer; ratsresponded to deliver the VS in the absence of any otherscheduled outcome. This primary reinforcement by the VSwas potently enhanced by response-contingent or non-contingent (yoked or continuous) nicotine infusions (Donnyet al. 2003). However, noncontingent (yoked) presentationof the VS did not enhance responding for contingentnicotine. One important feature of these studies was thatnoncontingent infusions of nicotine decreased the system-atic pairing of nicotine and the VS; continuous infusion ofnicotine completely eliminated the possibility of NIC–VSpairings. Thus, enhanced responding for the VS could nothave resulted from a pavlovian association between the twostimuli.
The finding that noncontingent nicotine infusionsenhance responding for a nonpharmacological reinforcerexperimentally substantiated the hypothesis that nicotinehas both primary reinforcing and reinforcement-enhancingeffects. However, these findings also raised two importantconcerns about the conditions under which reinforcement-enhancing effects of nicotine may be demonstrated andhow they operate during self-administration. First, in thepreviously described study (Donny et al. 2003), we inferredthat the ability of nicotine to enhance the reinforcing effectsof the VS contributed to robust self-administration understandard training protocols in which administration of bothreinforcers is dependent on the same operant response(NIC + VS). Although this inference was not directlytested, we hypothesized that self-administered nicotinewould enhance responding for other reinforcers. Second,the rats responding for NIC + VS receive significantlymore nicotine than rats self-administering nicotine withoutthe VS; they also determine the amount of nicotine re-ceived by the noncontingent nicotine groups (see Donny etal. 2003). This elevated nicotine intake suggests that thedrug may be more reinforcing when paired with a “self-administered”VS than when given alone and/or that higheramounts of nicotine are required to produce reinforcement-enhancing effects relative to primary reinforcing effects.
None of the three hypotheses described above can betested in a paradigm in which a single response controlsboth reinforcers. Therefore, we used a concurrent rein-forcement paradigm in which nicotine was available formaking one response, and the VS was available for makinganother response. A concurrent paradigm offered at least oneimportant advantage over previously described single-leverprocedures: the ability to isolate the primary reinforcing andreinforcement-enhancing effects of self-administered nico-tine. For example, a single response that delivered nicotineinfusions allowed us to directly measure nicotine’s primaryreinforcing effects compared to a nicotine-only condition.A separate response that controlled the VS allowed us tomeasure its primary reinforcing effects. If the reinforce-ment-enhancing effects of nicotine emerged under aconcurrent reinforcement schedule, they should be evidentas increased responding for the VS relative to a VS-onlycondition. Thus, the primary reinforcing effects of nicotinecan be measured with the nicotine lever, while thereinforcement-enhancing effects can be measured with adifferent operant (VS lever). By unconfounding the re-sponse required to obtain nicotine from the one necessary todeliver the VS, this procedure enables us to more directlyassess the reinforcement-enhancing effects of self-adminis-tered nicotine.
Materials and methods
Subjects Male Sprague–Dawley rats (Harlan Farms, IN)weighing 200–225 g on arrival were housed individuallyin hanging wire mesh cages. The cages were contained ina temperature- and humidity-controlled colony room on areverse 12:12 h light/dark cycle. Rats were allowed un-restricted access to food and water for 3 days after theirarrival. Food access was subsequently restricted as de-scribed in previous studies (Donny et al. 1995).
Apparatus All experimental sessions were conducted in25×31×28 (w×l×h) cm operant conditioning chambers(BRS/LVE Model RTC-020, MD; see Donny et al. 1995for further details). The VS consisted of 1-s illumination ofa white cue light located directly above the “VS” or “active”lever, followed by 1-min deactivation of the house light.During each session, rats were connected to a swivelsystem that delivered intravenous infusions while allowingnearly unrestricted movement in the chamber. See Donnyet al. (1995) for other details about the apparatus.
Lever training Following habituation to the colony room,rats were food deprived for 24 h and then trained to leverpress on both levers for 45 mg food pellets over the courseof three to four daily sessions. During initial sessions,levers were extended (one at a time) into the chamber inrandomized order. After 15 s elapsed or a response wasrecorded, the lever was retracted, and a food pellet wasdelivered. Rats meeting a criterion of 15 responses duringautoshaping sessions were tested for lever preference; theremaining rats were hand-shaped to press both levers
392
before testing. During the test session, both levers wereextended simultaneously; the session ended after 40 totalresponses on either lever. These nonreinforced responseswere used to determine unconditioned lever preferences.
Surgery After lever training, chronic indwelling jugularcatheters were implanted in each rat. Catheters were con-structed as previously described (Donny et al. 1998).Following surgery, catheters were flushed daily by in-fusing 0.1 ml heparinized sterile saline and the antibioticticarcillan plus clavulanate to reduce postsurgical infec-tions. See Donny et al. (1998) for other details about sur-gical procedures.
Drugs Nicotine hydrogen tartrate salt was dissolved in0.9% saline and brought to a pH of 7.0 (±0.2) with a diluteNaOH solution. Throughout the experiment, nicotine wasdelivered at 0.06 mg kg−1 inf−1; dose was calculated fromthe base form of nicotine. This dose was selected becauseprior research demonstrated that it produces the mostreliable self-administration in the nicotine-only condition(Donny et al. 2003). All infusions were delivered in lessthan 1 s at a volume of 0.1 ml kg−1 inf−1.
Self-administration testing (sessions 1–22) During therecovery period, all rats were randomly assigned to one offour groups (2 lever, NIC + VS, NIC only, or VS only)with two constraints. First, left vs right lever preferenceswere counterbalanced as much as allowed by the samples.Second, overall preference ratios for preferred vs non-preferred levers were matched across the groups. Subse-quently, each rat was randomly assigned an “active” and“inactive” lever or an “infusion” and “VS” lever with theconstraints that left vs right was counterbalanced, andpreference ratios were matched across lever assignments.For the 2-lever, VS-only, NIC + VS, and NIC-only groups,the mean (SEM) preference ratio for the infusion/activelever was 0.53 (0.07), 0.48 (0.09), 0.47 (0.07), and 0.53(0.05), respectively.
Nicotine self-administration was carried out in 1-h ses-sions 5 days per week. For each session, rats were placedin the experimental chambers and connected to the drug-delivery swivel system. Illumination of the house lightsignaled the start of the session. For each group, differentcontingencies were delivered as the result of pressing eachlever. For the 2-lever group, pressing the infusion lever
resulted in the delivery of 0.06 mg kg−1 nicotine. Aftereach infusion, nicotine was unavailable for a 1 min time-out period. Pressing the VS lever resulted in 1-s illumi-nation of the cue light followed by 60-s deactivation of thehouse light. Pressing the VS lever had no programmedconsequences during the 61-s VS period. For the VS-onlygroup, training contingencies were identical; however,pressing the infusion lever resulted only in a 0.1 ml kg−1
infusion of 0.9% saline. For the NIC + VS group, pressingthe active lever resulted in a 0.06 mg kg−1 infusion ofnicotine and delivery of the VS. For the NIC-only group,pressing the active lever resulted only in a 0.06 mg kg−1
nicotine infusion. For these latter two groups, pressing theinactive lever had no scheduled consequences (see Table 1).
The first 12 training sessions were conducted on afixed-ratio 1 (FR1) schedule of reinforcement. For oneresponse recorded on a lever, the scheduled reinforcer wasdelivered, and the time-out period began. For sessions 13–22, the schedule was increased to an FR2, meaning thatreinforcers were delivered after two responses wererecorded on a lever. For the 2-lever and VS-only groups,schedules were always the same on both levers and alwaysmatched the schedule requirement for groups with onlyone active lever.
Saline substitution (sessions 23–27) Rats that hadpreviously received nicotine infusions for meeting theFR2 schedule on the infusion or active lever received0.1 ml kg−1 saline infusions instead. During this phase,rats that previously had access to the VS (2-lever, NIC +VS, and VS-only groups) continued to receive the VS forcompleting the FR2 schedule on the VS or active lever.Note that for rats in the VS-only condition, there was nochange in parameters during this phase of the experiment.
Nicotine-primed reinstatement (sessions 28–29) Afterextinction of drug self-administration, noncontingent re-introduction of the training drug often renews the drug-seeking response, an effect termed “reinstatement” (DeWitand Stewart 1981). Here, we were also interested in whethernoncontingent reintroduction of the training drug, nicotine,renews its reinforcement-enhancing effects of respondingfor the VS. Therefore, we delivered noncontingent nicotineinfusions to investigate their effect on nicotine and VSseeking. Under some conditions, nicotine-primed reinstate-ment is variable and unreliable (see Chiamulera et al. 1998;
Table 1 Response–outcome contingencies for each group/lever
Group 2 lever (n=9) VS only (n=9) NIC + VS (n=9) NIC-only (n=11)
Lever INF VS INF VS ACT INACT ACT INACT
Shaping Food Food Food Food Food Food Food FoodSelf-administration NIC VS SAL VS NIC + VS – NIC –Saline substitution SAL VS SAL VS SAL + VS – SAL –Reinstatement test – VS – VS VS – – –
Lever and group assignments were matched according to the response during a preference test that was conducted after food shapingVS Visual stimulus, 1 s cue light-on, 1 min house light-off; NIC nicotine infusion, 0.06 mg kg−1; SAL 0.09% saline infusion; INF infusionlever; ACT active lever; INACT inactive lever
393

Le Sage et al. 2004). In addition, preliminary analysis of the2-lever group suggested that enhancement of respondingfor the VS was positively correlated with the number ofnicotine infusions that rats received across a session. Toinvestigate the role of nicotine’s reinforcement-enhancingeffects in a model of nicotine relapse, nicotine reinstatementtests consisted of a series of eight infusions constructedfrom the pattern of nicotine seeking in the 2-lever and NIC-only groups on sessions 21 and 22. Based on these data,infusions were delivered at minutes 1, 2.5, 5, 10, 15, 20, 28,and 37 of sessions 28 and 29.
A differential reinforcement schedule was included onreinstatement tests to prevent adventitious correlations be-tween nicotine delivery and responding. To do so, scheduledinfusions were delayed if 10 s had not elapsed from therats’ last response on either lever. To ensure that the timingof scheduled infusions did not significantly differ acrosssubjects, the delay was capped at a maximum of 30 s; afterwhich infusions were delivered regardless of the elapsedtime from the previous recorded response. All rats receivedscheduled infusions at least 10 s after their last recordedresponse. The average delay between the schedule andactual receipt of infusions was 0.52 (±0.5) s, there were nodifferences between the various treatment groups, F<1.
Data analyses The primary goal of this study was tocompare the topography of responding when two reinforc-ers were presented for performing a single operant, relativeto presenting each reinforcer for performing separate operants.However, an important aspect of the study was to replicateprevious findings in which nicotine self-administration inthe presence vs absence of nonpharmacological stimuli wascompared. Accordingly, responding was analyzed in twoways. First, “active” lever responding (excluding time-outresponding) was compared according to the reinforcerbeing delivered during each training phase (nicotine leverof the 2-lever group, NIC + VS, and NIC only, or VS leverof the 2-lever group, NIC + VS, and VS only). These anal-yses facilitate comparisons across phases in which nicotinewas response contingent (FR2) or scheduled (nicotine re-instatement). Second, total responding on active vs inactive(NIC + VS and NIC only) or infusion vs VS (2 lever and VSonly) levers were compared. Time-out responding couldnot be excluded from these latter analyses, as no scheduledtime-outs resulted from inactive lever responding. Exclud-ing time-out data might be a more accurate measure of goalor outcome seeking. However, for the NIC-only and 2-levergroups, the time-out from nicotine is unsignaled. Underthese circumstances, time-out responding may reflect nico-tine seeking, depending on what is learned about the timeout (e.g., FI1-min schedule). Therefore, we chose to displaythe data both excluding (Fig. 1) and including (Fig. 2) time-out responding. Data from each reinforcement phase (FR1,FR2, saline substitution, and nicotine reinstatement) wereanalyzed independently with repeated measures analyses ofvariance (ANOVAs) where appropriate. Analyses compar-ing one reinforcement phase to another included phase as awithin-subjects factor. Omnibus ANOVAs are described in
the figure captions, and corrected follow-up comparisons(e.g., Dunnet’s) are described in the text. An alpha level ofp≤0.05 was used to declare statistical significance, multiplep values are reported as “ps.”
Results
Responding for nicotine and/or VS
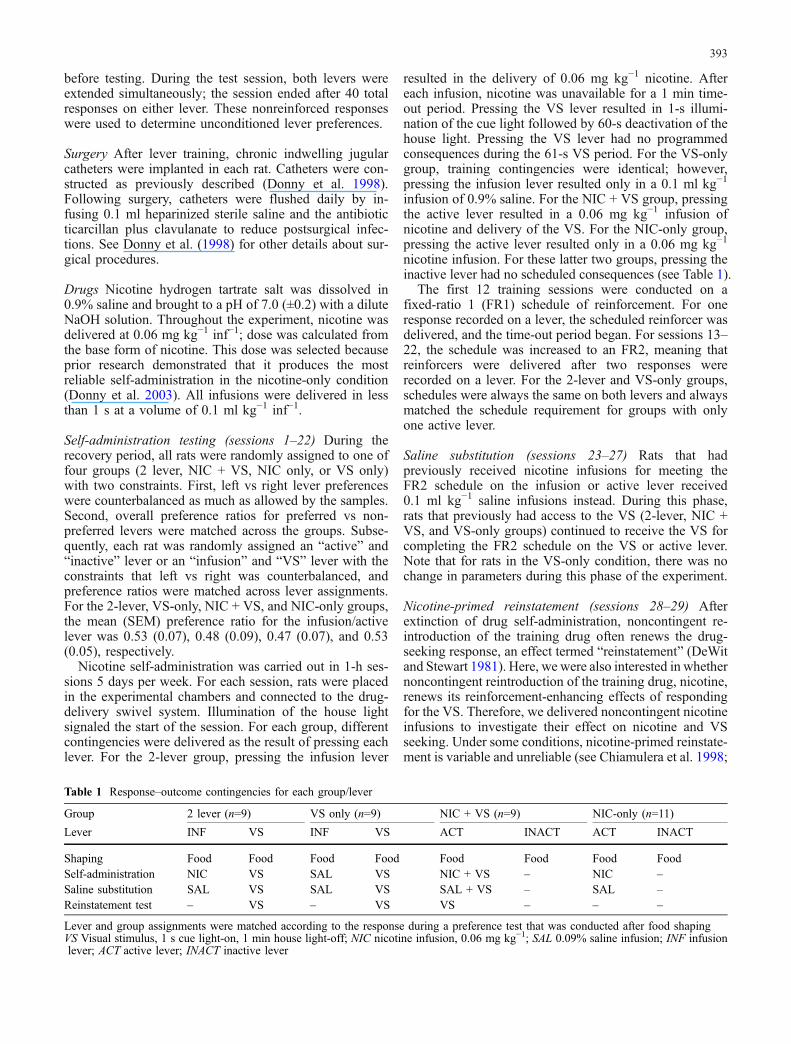
The first main goal of the study was to compare respondingfor two reinforcers when they were delivered for making asingle response (e.g., NIC + VS group), or when each wasconcurrently available (2-lever group) compared to whenthey were delivered alone. Accordingly, average respond-ing for each reinforcer in all groups is illustrated in Fig. 1;time-out responding is not included in the figure orsubsequent analyses.
Self-administration testing During initial testing sessions(left panel), the NIC + VS group responded more on theactive lever than NIC-only controls (ps≤0.034 for sessions1–12). In contrast, responding for nicotine did not differbetween the 2 lever and NIC-only groups (ps>0.18 forsessions 1–12). A similar pattern was observed under anFR2 schedule of reinforcement; NIC + VS rats made moreresponses on the active lever relative to NIC-only controls(ps<0.001 for session 13–22), while nicotine seeking inthe 2-lever group did not differ from NIC-only controls(ps≥0.5). Across FR1 sessions, the NIC + VS groupresponded more for the VS (i.e., active lever) than VS-only controls (ps<0.019 for sessions 3, 4, and 6–8).However, the 2-lever and VS-only groups did not differ inresponding for the VS (ps≥0.10). For FR2 sessions, theNIC + VS group responded more for the VS than VS-onlycontrols (ps<0.05 for sessions 13–22). The 2-lever groupdisplayed more VS seeking than VS-only controls onsessions 17 and 19–22 (ps≤0.03). A subsequent follow-upcomparing NIC + VS and 2-lever rats during FR2 testingrevealed that these groups differed only on session 13(p=0.02).
Saline substitution For the 2-lever group, VS-leverresponding (p=0.006), but not infusion-lever responding(p=0.52), declined on the first session in which salineinfusions replaced nicotine, with no further decreaseacross subsequent sessions (ps≥0.1). Active-lever respond-ing also decreased on the first saline substitution sessionfor the NIC + VS group (p=0.03), with no further decreaseacross subsequent sessions (p=0.42). In contrast, the NIC-only group showed no initial change in responding on theactive lever (p=0.13), but did exhibit a gradual declineacross saline substitution sessions (p=0.008). For the VS-only group, there was no change in the proceduralparameters, and responding did not change from FR2testing to saline substitution testing (p=0.93). Note that asexpected, for the three groups receiving the VS and salineinfusions (NIC + VS, 2 lever, and VS only), responding
394
for the VS was similar during the last two sessions ofsaline substitution (sessions 26–27, ps≥0.32).
Nicotine-primed reinstatement Average (±1 SEM) activelever responses from the nicotine-primed reinstatementtest are illustrated in the far-right panel of Fig. 1. Asillustrated in the figure, noncontingent nicotine infusionsincreased active-lever responding for rats in the NIC + VSgroup (p<0.001) and infusion-lever responding in the 2-lever group (p=0.004). In contrast, this manipulation didnot alter responding in the NIC-only control group(p=0.96). For the 2-lever group, noncontingent nicotineinfusions also reinstated responding on the VS lever(p<0.001).
Overall responding
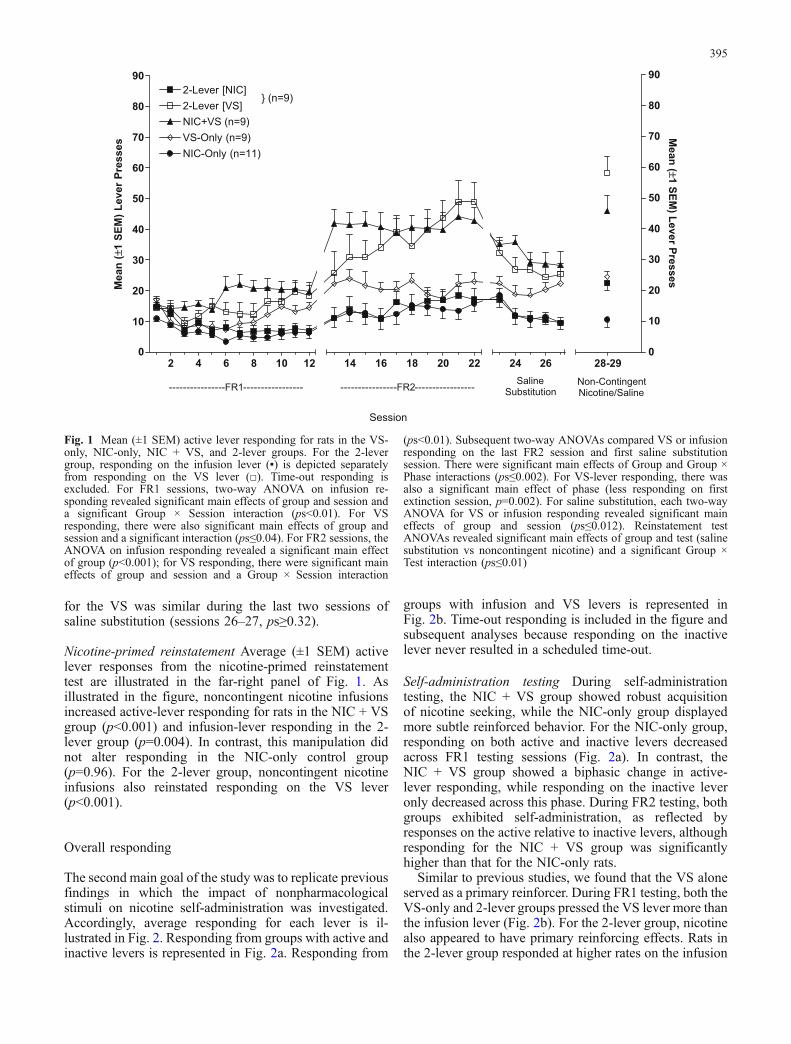
The second main goal of the study was to replicate previousfindings in which the impact of nonpharmacologicalstimuli on nicotine self-administration was investigated.Accordingly, average responding for each lever is il-lustrated in Fig. 2. Responding from groups with active andinactive levers is represented in Fig. 2a. Responding from
groups with infusion and VS levers is represented inFig. 2b. Time-out responding is included in the figure andsubsequent analyses because responding on the inactivelever never resulted in a scheduled time-out.
Self-administration testing During self-administrationtesting, the NIC + VS group showed robust acquisitionof nicotine seeking, while the NIC-only group displayedmore subtle reinforced behavior. For the NIC-only group,responding on both active and inactive levers decreasedacross FR1 testing sessions (Fig. 2a). In contrast, theNIC + VS group showed a biphasic change in active-lever responding, while responding on the inactive leveronly decreased across this phase. During FR2 testing, bothgroups exhibited self-administration, as reflected byresponses on the active relative to inactive levers, althoughresponding for the NIC + VS group was significantlyhigher than that for the NIC-only rats.
Similar to previous studies, we found that the VS aloneserved as a primary reinforcer. During FR1 testing, both theVS-only and 2-lever groups pressed the VS lever more thanthe infusion lever (Fig. 2b). For the 2-lever group, nicotinealso appeared to have primary reinforcing effects. Rats inthe 2-lever group responded at higher rates on the infusion
28-290
10
20
30
40
50
60
70
80
90
Mean
(±1 S
EM
) Lever P
resses
2 4 6 8 10 120
10
20
30
40
50
60
70
80
90
14 16 18 20 22 24 26
2-Lever [NIC]
2-Lever [VS]
NIC+VS (n=9)
VS-Only (n=9)
NIC-Only (n=11)
Mean
(±1
SE
M)
Lever
Pre
sses
Session
----------------FR1----------------- ----------------FR2-----------------Saline
SubstitutionNon-ContingentNicotine/Saline
} (n=9)
Fig. 1 Mean (±1 SEM) active lever responding for rats in the VS-only, NIC-only, NIC + VS, and 2-lever groups. For the 2-levergroup, responding on the infusion lever (▪) is depicted separatelyfrom responding on the VS lever (□). Time-out responding isexcluded. For FR1 sessions, two-way ANOVA on infusion re-sponding revealed significant main effects of group and session anda significant Group × Session interaction (ps<0.01). For VSresponding, there were also significant main effects of group andsession and a significant interaction (ps≤0.04). For FR2 sessions, theANOVA on infusion responding revealed a significant main effectof group (p<0.001); for VS responding, there were significant maineffects of group and session and a Group × Session interaction
(ps<0.01). Subsequent two-way ANOVAs compared VS or infusionresponding on the last FR2 session and first saline substitutionsession. There were significant main effects of Group and Group ×Phase interactions (ps≤0.002). For VS-lever responding, there wasalso a significant main effect of phase (less responding on firstextinction session, p=0.002). For saline substitution, each two-wayANOVA for VS or infusion responding revealed significant maineffects of group and session (ps≤0.012). Reinstatement testANOVAs revealed significant main effects of group and test (salinesubstitution vs noncontingent nicotine) and a significant Group ×Test interaction (ps≤0.01)
395
2-Lever (n=9)
VS-Only (n=9)
A Inactive vs. Active Lever
2 4 6 8 10 120
10
20
30
40
50
60
70
80
90
14 16 18 20 22 24 26
NIC+VS (n=9)
NIC-Only (n=11)M
ean
(±1
SE
M)
Lev
er P
ress
es
Session
-------------- FR1 ---------------- ------------- FR2 --------------Saline
SubstitutionNon-ContingentNicotine/Saline
28-290
10
20
30
40
50
60
70
80
90
Mean
(±1 SE
M) L
ever Presses
B VS Lever vs. Infusion Lever
2 4 6 8 10 120
10
20
30
40
50
60
70
80
90
14 16 18 20 22 24 26
Mea
n (
±1 S
EM
) L
ever
Pre
sses
28-290
10
20
30
40
50
60
70
80
90
Mean
(±1 SE
M) L
ever Presses
Fig. 2 a Mean (±1 SEM) active and inactive lever responding forrats in the NIC-only and NIC + VS groups. b Mean (±1 SEM)infusion and VS-lever responding for rats in the VS-only and 2-levergroups. Filled symbols represent active (a) or infusion levers (b);open symbols represent inactive (a) or VS levers (b). Time-outresponding is included because inactive levers did not result in ascheduled time-out. The three-way ANOVA on active- and inactive-lever responding (a) during FR1 sessions revealed significant maineffects of group, session, and lever (ps<0.01), as well as significantLever × Group and Lever × Session × Group interactions (ps<0.01).For FR2 sessions, the ANOVA revealed significant main effectsof group and lever and a significant Group × Lever interaction(ps<0.01). For saline substitution, there were significant main effectsof group, lever, and session (ps<0.01), as well as significant Group ×
Lever and Lever × Session interactions (ps=0.01). For the three-wayANOVA on reinstatement tests, all main effects and interactions weresignificant (ps≤0.005). For VS- and infusion-lever responding (b)during FR1 testing, the ANOVA revealed significant main effects ofgroup, session, and lever (ps≤0.05), as well as significant Lever ×Session and Group × Lever × Session interactions (ps≤0.03). ForFR2 sessions, there were significant main effects of group, session,and lever (p≤0.02), as well as significant Group × Session, Lever ×Session, and Group × Lever × Session interactions (ps≤0.04). Duringsaline substitution, there were significant main effects of group andlever (p≤0.02). For the three-way ANOVA on reinstatement tests,there were significant main effects of group, phase (sessions 26–27vs sessions 28–29), and lever, as well as a significant Phase × Groupinteraction (ps≤0.01)
396
lever (i.e., for nicotine), relative to the VS-only group (i.e.,for saline). As previously mentioned, the reinforcement-enhancing effects of nicotine emerged during FR2 testing.Rats in the 2-lever group pressed the VS lever more thanVS-only controls.
Saline substitution For NIC + VS and NIC-only groups(Fig. 2a), active lever responding decreased across salinesubstitution sessions, whereas inactive lever pressing didnot change. Because the VS is a mildly reinforcingstimulus (e.g., Donny et al. 2003, also see later discussion),the NIC + VS group maintained a higher rate of re-sponding on the active lever, relative to the NIC-onlycontrols. For the VS-only and 2-lever groups (Fig. 2b),VS-lever responding was higher than responding on theinfusion lever. In addition, responding in the 2-lever groupwas higher than responding in the VS-only group.
Nicotine-primed reinstatement For the NIC + VS group,responding on the active lever, but not the inactive lever,increased on the nicotine-primed reinstatement test. Re-sponding did not change on either lever for the NIC-onlygroup (Fig. 2a). For the 2-lever group, noncontingent nic-otine infusions increased responding on both the infusionand VS levers. Noncontingent saline infusions did not af-fect infusion- or VS-lever responding in the VS-only group(Fig. 2b).
Pattern of nicotine and VS self-administration
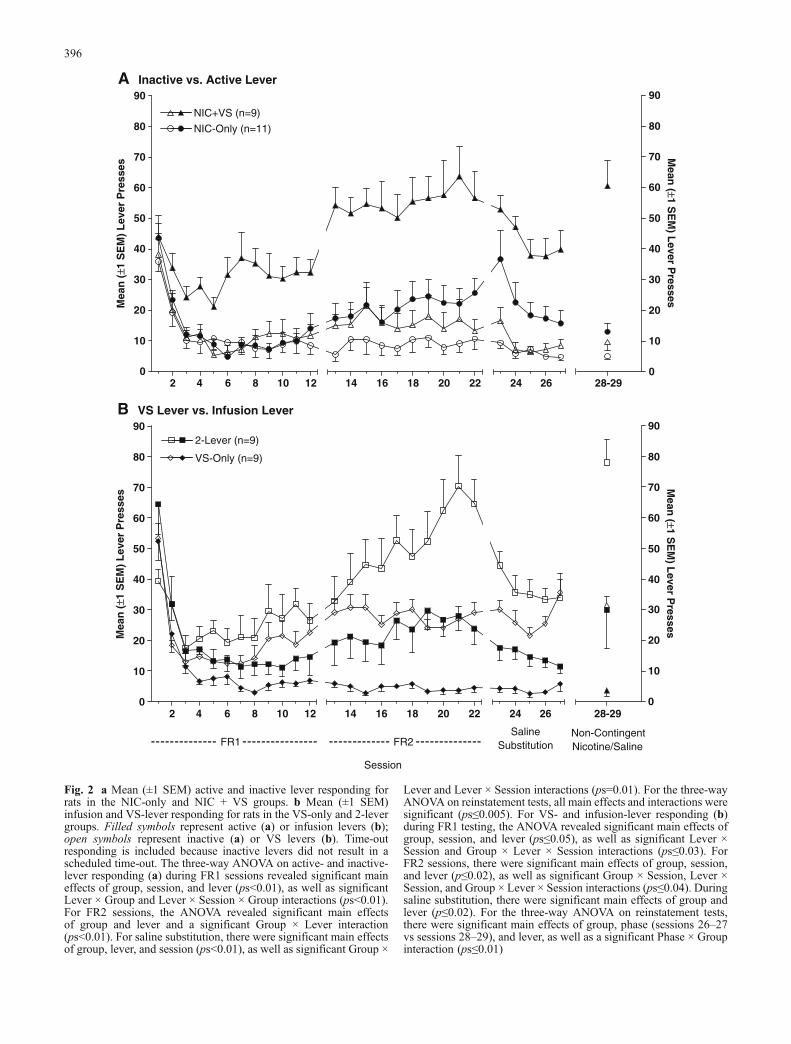
Previous studies (e.g., Lynch and Carroll 1999) havedemonstrated that rats regulate nicotine intake during self-administration sessions. In the present study, the pattern ofnicotine and VS intake across self-administration testingbecomes important because the 2-lever group has theopportunity to regulate the delivery of each reinforcer. The2-lever group displayed a U-shaped pattern of respondingduring FR1 training, presumably due to extinction of food-maintained responding (sessions 1–3) and “emergence” ofnicotine’s reinforcement-enhancing effects (sessions 8–12).To further examine the pattern of nicotine and VS intakeduring self-administration testing, the number of reinforc-ers earned on sessions 7 and 22 were separated into 10-minbins (Fig. 3). Session 22 represents the self-administrationend point, whereas session 7 preceded the emerging rein-forcement-enhancing effects of nicotine with the leastinfluence from prior food training. Figure 3a shows VSpresentations and nicotine infusions for the 2-lever groupon sessions 7 and 22. On session 7, nicotine infusions andVS presentations decreased as the session progressed. Onsession 22, nicotine infusions showed a similar pattern ofdecline across the session; in contrast, VS presentations didnot decline across this session. For comparative purposes,mean (±1 SEM) VS presentations for the 2-lever and NIC +VS groups on sessions 7 and 22 are illustrated in Fig. 3b.Although the pattern of VS deliveries changes drasticallybetween sessions 7 and 22 for the 2-lever group, reinforcer
deliveries for the NIC + VS and VS-only (data not shown)groups did not change over these sessions (ps≥0.1).
For the 2-lever group, the number of pairings betweenthe VS presentations and nicotine infusions was calculatedto determine whether the “reinforcement-enhancing” effectof nicotine could have been the result of adventitiouspairings between the two reinforcers, resulting in VS be-coming a conditioned reinforcer. To investigate this possibil-ity, we sampled 1,075 nicotine infusions from the 2-levergroup over FR1 and FR2 sessions. Each infusion that oc-curred during the 1-min VS or during the 15-s post-VS tracewas scored as “paired,” all other infusions were scored as
A 2-Lever Group
0 10 20 30 40 50 600
1
2
3
4
5
6
7Inf: session 7Inf: session 22
VS: session 22VS: session 7
Time in Session (min)
Re
info
rce
rs E
arn
ed
B 2-Lever and NIC+VS Groups
0 10 20 30 40 50 600
1
2
3
4
5
6
7
Time in Session (min)
VS
Pre
sen
tati
on
s E
arn
ed
2-Lever: session 222-Lever: session 7
NIC+VS: session 7NIC+VS: session 22
Fig. 3 a Mean (±1 SEM) reinforcers earned for rats in the 2-levergroup on sessions 7 and 22. Filled symbols represent infusions; opensymbols represent VS presentations. b Mean (±1 SEM) VSpresentations for rats in the 2-lever and NIC + VS groups. Thethree-way ANOVA revealed significant main effects of reinforcerand interval, and significant Reinforcer × Session and Session ×Interval interactions (ps≤0.03). A follow-up ANOVA included VS-only (data not shown) and 2-lever groups to determine whether thechange in VS delivery resulted from the change in schedule. Therewere significant main effects of group, session, and interval andsignificant Group × Session and Session × Interval interactions(ps≤0.05), indicating that nicotine, rather than schedule, increasedthe number of VS deliveries in the 2-lever group
397

“unpaired.” When pairing is defined in this way, the prob-ability that nicotine and VS would co-occur was 0.05,whereas nicotine infusions and VS deliveries occurred alonewith probabilities of 0.08 and 0.29, respectively. Given thateach stimulus was more likely to occur alone, rather thanpaired, we do not expect Pavlovian associations to have anynoteworthy impact on responding for the VS (Rescorla1967).
Discussion
We have previously argued that nicotine self-administra-tion reflects two dissociable actions of the drug: primaryreinforcement and a reinforcement-enhancing effect(Chaudhri et al. 2005b; Donny et al. 2003). However, tosubstantiate the second (reinforcement-enhancing) effect,we have used experimenter-administered (i.e., noncontin-gent) infusions of nicotine. These procedures raised a fewconcerns with regard to the role of reinforcement-en-hancing effects in nicotine self-administration. First, wehad not directly tested whether self-administered nicotineenhanced responding for nonpharmacological stimuli. Inthe present study, response-contingent nicotine potentlyenhanced responding for a concurrently available non-pharmacological stimulus (2-lever group), confirming thatself-administered nicotine has reinforcement-enhancingeffects. A second concern from previous studies was thatrats receiving both nicotine and the VS for making a singleoperant response (NIC + VS) self-administer much morenicotine than rats taking the drug in the absence ofnonpharmacological cues (NIC only). This finding sug-gests that repeated pairings with the VS may enhancenicotine reinforcement (nicotine seeking). In addition, theoccurrence of reinforcement-enhancing effects are limitedto conditions that induce high levels of drug-intake. In the2-lever group, nicotine infusions were rarely paired withthe VS; each reinforcer was more likely to occur alonerather than in compound. Under these circumstances, thereinforcement-enhancing effect of nicotine was unidirec-tional. Responding for nicotine was maintained at levelsexpected by its primary reinforcing effects (NIC only),while responding for the VS was potently enhancedrelative to rats responding for the VS alone. This findingextends the conditions under which the reinforcement-enhancing effects of nicotine operate by demonstrating thatthe primary reinforcing and reinforcement-enhancingeffects of nicotine occur under similar circumstances.
One finding that deserves further mention was that thereinforcement-enhancing effects of nicotine appeared toemerge more rapidly in the NIC + VS group. The rapidincrease in active-lever responding and subsequent stabi-lization in the NIC + VS group is typical of many drug self-administration studies (e.g., Caggiula et al. 2002; Cohen etal. 2005) and is often characterized as “acquisition” of anassociation between performing the operant response (i.e.,pressing the lever) and the resultant appetitive effectsderived from infusion of the drug. However, results of thepresent study indicate that the acquisition of the response–
drug association is confounded with acquisition of aresponse–VS association and perhaps a VS–nicotine as-sociation (see previous discussion). At least three explana-tions may account for the more rapid development of thereinforcement-enhancing effects of nicotine in the NIC +VS group. First, paired nicotine infusions may have con-ferred conditioned-reinforcing value upon the VS. Thisaccount predicts more rapid “acquisition” for groups with aperfect correlation between nicotine and VS (NIC + VS),relative to groups receiving yoked/continuous infusions (inwhich pairings are much less frequent). However, po-tentiated responding for the VS emerges at similar rateswhether nicotine infusions are contingent (NIC + VS) ornoncontingent (Chaudhri et al. 2005a; Donny et al. 2003).Second, the differences in schedule requirement may havefacilitated receipt of the two reinforcers. NIC + VS groupshad to make one or two responses to get both reinforcers,whereas the 2-lever group would have had to make two orfour responses. However, data from the 2-lever groupsuggest that work requirement had little, if any, impact onthe amount of reinforcers earned. The number of VSdeliveries increased more when the work requirement wasgreater (i.e., FR2), and the number of nicotine infusions didnot change considerably across the schedules. Therefore,the emergence of nicotine’s reinforcement-enhancing effectappeared to depend more on an interaction between thepassage of time and the quantity of nicotine each group wasexposed to. However, further research will be required todetermine the accuracy of this latter hypothesis.
Another interesting finding from the 2-lever group wasthat two separate responses for nicotine and the VS did notinterfere with one another. Introducing alternative responseoptions and reinforcers in drug self-administration is not anovel concept (e.g., Campbell and Carroll 2000). However,in most cases, the alternative reinforcer reduces self-administration of the drug (e.g., Paronis et al. 2002). Thisreduction is probably due to “matching” (Herrnstein 1961)between the response options and the reinforcement derivedfrom each option. In the present study, the concurrentschedule did not seem to result in interference betweennicotine and VS. Nicotine seeking was maintained at levelsthat would be expected for nicotine (NIC-only group), andVS seeking was maintained at or above the levels thatwould be expected for the VS (VS-only group). Parametricfactors such as schedule of reinforcement, session length,and dose are known to influence the effects of alternativereinforcement on drug self-administration (Woolverton1996; Woolverton and Alling 1999; Paronis et al. 2002).Further studies are required to investigate the conditionsunder which nicotine and VS may induce competingbehaviors.
The reinstatement test employed in the present study wasnot the traditional test used to investigate nicotine-primedreinstatement. Rats received eight scheduled noncontingentnicotine infusions over the first 40 min of a 1-h session.Most nicotine-primed reinstatement tests use a single bolusdose delivered at the beginning of the testing interval (e.g.,Le Sage et al. 2004, Shaham et al. 1997; Tessari et al.2004). There were two specific reasons why the present
398

study deviated from more standard nicotine-primed re-instatement techniques. First, we were interested in thereinforcement-enhancing effects of nicotine in the rein-statement model, requiring that we administer a dose knownto produce these effects. Observations from self-adminis-tration testing suggested that eight infusions in this patternshould engage these reinforcement-enhancing effects. Sec-ond, nicotine-induced reinstatement has been reported tobe more reliable in the presence, relative to absence, ofnicotine-paired stimuli (e.g., Le Sage et al. 2004). Indeed, areview of the literature suggests that the best demonstra-tions of nicotine-primed reinstatement include the “time-outcue” either during (Le Sage et al. 2004; Shaham et al. 1997)or just before (Andreoli et al. 2004; Chiamulera et al. 1998;Tessari et al. 2004) the noncontingent nicotine test. Thus,interpretation of responding is somewhat ambiguous because“reinstatement of nicotine seeking” is superimposed on re-sponding that may or may not be motivated by a nicotine-paired reinforcer.
The 2-lever group begins to remove the confoundsbetween stimulus seeking and nicotine seeking in the pres-ent study. Noncontingent nicotine infusions reinstated bothVS-lever and infusion-lever responding to presaline sub-stitution levels. This pattern was not surprising given thatthe reinforcement-enhancing effects of nicotine were morepotent than the primary reinforcing effects of nicotinethroughout self-administration testing. However, it is par-ticularly interesting in light of the stimulus-dependence ofnicotine-primed reinstatement in previous studies. Perhapsthese previous tests (e.g., Chiamulera et al. 1998; Le Sageet al. 2004) were the most reliable because they exploitednicotine’s reinforcement-enhancing properties; that is, nic-otine-primed reinstatement may be less dependent onrelapse to “nicotine seeking” and more dependent on thereturn of a reinforcement-enhancing effect. Future clinicaland preclinical investigation of tobacco relapse shouldinvestigate the relative contribution of nicotine’s primaryreinforcing and reinforcement-enhancing effects.
We have argued previously (e.g., Caggiula et al. 2001;Donny et al. 2003; Chaudhri et al. 2005b) that the weakprimary reinforcing properties of nicotine are a poorcorrelate for the tenacity of the smoking habit. The morepotent reinforcement-enhancing effect of nicotine probablymakes a much stronger contribution to chronic tobacco useand relapse. Recent studies from our laboratory have begunto elucidate the importance of this reinforcement-enhanc-ing effect (Caggiula et al. 2002; Donny et al. 2003;Chaudhri et al. 2005a), and the present study demonstratesthat these two effects of nicotine can be separated be-haviorally. This advance in nicotine self-administration isimportant because it increases the sensitivity with whichthe reinforcement-related effects of nicotine are measured.Accordingly, we may begin to resolve certain ambiguitiesthat exist in nicotine self-administration. For example, whyand how do nicotine’s reinforcement-enhancing effectsemerge over time? Why is nicotine-primed reinstatementso dependent on the presence of nicotine-paired cues (e.g.,
Le Sage et al. 2004)? The present study offers a newtechnique in which we may address these questions moresuccessfully by separating the dual reinforcing effects ofnicotine. This separation, and increased sensitivity, may havea significant impact on our current understanding of tobaccoaddiction.
Acknowledgements The authors would like to thank Dr. NeilLevens for his thoughtful comments on a previous version of thismanuscript. All experiments followed the “Principles of laboratoryanimal care” (NIH #85-23, revised 1985) and were approved by theUniversity of Pittsburgh Institutional Animal Care and Use Com-mittee (assurance #A3187-01). This research was supported by NIHgrants DA-10464, DA-12655, and DA-19278.
References
Andreoli M, Tessari M, Pilla M, Valerio E, Hagan JJ, HeidbrederCA (2004) Selective antagonism at dopamine D3 receptorsprevents nicotine-triggered relapse to nicotine-seeking behav-ior. Neuropsychopharmacology 28:1272–1280
Caggiula AR, Donny EC,White AR, Chaudhri N, Booth S, GharibMA,Hoffman A, Perkins KA, Sved AF (2001) Cue dependency of nic-otine self-administration and smoking. Pharmacol Biochem Behav70:515–530
Caggiula AR, Donny EC, White AR, Chaudhri N, Booth S, GharibMA, Hoffman A, Perkins KA, Sved AF (2002) Environmentalstimuli promote the acquisition of nicotine self-administrationin rats. Psychopharmacology 163:230–237
Campbell UC, Carroll ME (2000) Acquisition of drug self-administration: environmental and pharmacological interven-tions. Exp Clin Psychopharmacol 8:312–325
Chaudhri N, Caggiula AR, Donny EC, Booth S, Gharib MA, CravenLA, Shannon SA, Sved AF, Perkins KA (2005a) Sex differencesin the contribution of nicotine and nonpharmacological stimulito nicotine self-administration in rats. Psychopharmacology 180:258–266
Chaudhri N, Caggiula AR, Donny EC, Palmatier MI, Xiu L, Sved AF(2005b) Complex interactions between nicotine and nonphar-macological stimuli reveal multiple roles for nicotine in re-inforcement. Psychopharmacology (in press)
Chiamulera C, Rezzaghi F, Ardizzon M, Motta C, Marcon C, TessariMA (1998) A novel model to study relapse to nicotine-seekingbehaviour in rats. Arch Pharmacol 358:R578
Cohen C, Perrault G, Griebel G, Soubrié P (2005) Nicotine-associatedcues maintain nicotine-seeking behavior in rats several weeksafter nicotine withdrawal: reversal by the cannabinoid (CB1)receptor antagonist, rimonabant (SR141716). Neuropsychophar-macology 30:145–155
Corrigall WA, Coen KM (1994) Nicotine self-administration andlocomotor activity are not modified by the 5-HT3 antagonistsICS 205-930 and MDL 72222. Pharmacol Biochem Behav 49:67–71
De Wit H, Stewart J (1981) Reinstatement of cocaine-reinforcedresponding in the rat. Psychopharmacology 75:134–143
Donny EC, Caggiula AR, Knopf S, Brown C (1995) Nicotine self-administration in rats. Psychopharmacology 122:390–394
Donny EC, Caggiula AR, Mielke MM, Jacobs KS, Rose C, Sved AF(1998) Acquisition of nicotine self-administration in rats:the effects of dose, feeding schedule, and drug contingency.Psychopharmacology 136:83–90
Donny EC, Chaudhri N, Caggiula AR, Evans-Martin FF, Booth S,Gharib MA, Clements LA, Sved AF (2003) Operant respond-ing for a visual reinforcer in rats is enhanced by noncontingentnicotine: implications for nicotine self-administration and re-inforcement. Psychopharmacology 169:68–76
399

Goldberg SR, Spealman RD, Goldberg DM (1981) Persistent be-havior at high rates maintained by intravenous self-administra-tion of nicotine. Science 214:573–575
Harvey DM, Yasar S, Heishman SJ, Panlilio LV, Henningfield JE,Goldberg SR (2004) Nicotine serves as an effective reinforcerof intravenous drug-taking behavior in human cigarette smokers.Psychopharmacology 175:134–142
Herrnstein RJ (1961) Relative and absolute strength of response as afunction of frequency of reinforcement. J Exp Anal Behav 4:267–272
Le Sage MG, Burroughs D, Dufek M, Keyler DE, Pentel PR (2004)Reinstatement of nicotine self-administration in rats by presen-tation of nicotine-paired stimuli, but not nicotine priming.Pharmacol Biochem Behav 79:507–513
Le Foll B, Goldberg SR (2005) Control of the reinforcing effects ofnicotine by associated environmental stimuli in animals andhumans. Trends Pharmacol Sci 26:287–293
Lynch WJ, Carroll ME (1999) Regulation of intravenously self-administered nicotine in rats. Exp Clin Psychopharmacol 7:198–207
Niaura R, Shadel WG, Abrams DB, Monti PM, Rohsenow DJ,Sirota A (1998) Individual differences in cue reactivity amongsmokers trying to quit: effects of gender and cue type. AddictBehav 23:209–224
Paronis CA, Gasior M, Bergman J (2002) Effects of cocaine underconcurrent fixed ratio schedules of food and IV drug availability:a novel choice procedure in monkeys. Psychopharmacology163:283–291
Perkins KA, Epstein LH, Grobe J, Fonte C (1994) Tobacco ab-stinence, smoking cues, and the reinforcing value of smoking.Pharmacol Biochem Behav 47:107–112
Rescorla RA (1967) Pavlovian conditioning and its proper controlprocedures. Psychol Rev 74:71–80
Rose JE, Behm FM, Westman EC, Johnson M (2000) Dissociatingnicotine and nonnicotine components of cigarette smoking.Pharmacol Biochem Behav 67:71–81
Shaham Y, Adamson LK, Grocki S, Corrigall WA (1997) Reinstate-ment and spontaneous recovery of nicotine seeking in rats.Psychopharmacology 130:396–403
Tessari M, Pilla M, Andreoli M, Hutcheson DM, Heidbreder CA(2004) Antagonism at metabotropic glutamate 5 receptorsinhibits nicotine- and cocaine-taking behaviours and preventsnicotine-triggered relapse to nicotine-seeking. Eur J Pharmacol499:121–133
US Department of Health and Human Services (USDHHS) (1988)Nicotine addiction: a report of the surgeon general. US De-partment of Health and Human Services. Office of the AssistantSecretary for Health, Office on Smoking and Health, Rockville,MD
Woolverton WL (1996) Intravenous self-administration of cocaineunder concurrent VI schedules of reinforcement. Psychophar-macology 127:195–203
Woolverton WL, Alling K (1999) Choice under concurrent VIschedules: comparison of behavior maintained by cocaine orfood. Psychopharmacology 141:47–56
400
Copyright © 2022 FDOKUMEN




















