
Dissociating neural indices of dynamic cognitive control in advance task-set preparation: An ERP...
11
This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial
Transcript of Dissociating neural indices of dynamic cognitive control in advance task-set preparation: An ERP...

This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's p
erso
nal
copyResearch Report
Dissociating neural indices of dynamic cognitive control inadvance task-set preparation: An ERP study of task switching
D.E. Astlea,!, G.M. Jacksonb, R. Swainsona
aSchool of Psychology, University of Nottingham, University Park, Nottingham, NG7 2RD, UKbDivision of Psychiatry, University of Nottingham, UK
A R T I C L E I N F O A B S T R A C T
Article history:Accepted 27 September 2006Available online 7 November 2006
Switching between different tasks is associated with performance deficits, or ‘switch costs’,relative to repeating the same task. Recent evidence suggests that response rather than taskselection processes may be a major cause of switch costs [Schuch, S., Koch, I., 2003. The roleof response selection for inhibition of task sets in task shifting. J Exp Psychol Hum PerceptPerform, 29 (1) 92–105]. Thus, switch costs are not incurred if on the preceding trial a task hasbeen prepared for but no response required (a ‘no-go’ trial). We investigated the relationshipbetween response selection and the subsequent preparation of an alternative task set.While switch costs were absent following ‘no-go’ trials, ERP differences during the precueinginterval showed that response selection has implications for subsequent task preparation aswell as for task performance per se. The results are discussed in relation to the dissociationof intention versus action in behavioural control and the role of inhibition in switchingbetween task sets.
© 2006 Elsevier B.V. All rights reserved.
Keywords:Task switchingERPsTask setCognitive controlExecutive function
1. Introduction
Althoughwe are able to exert a great degree of control over ourbehaviour, it seems clear that the pattern and efficiency of thebehaviour we exhibit is driven not only by our intentions butalso by our recent behaviour. That is, there can be adissociation of intention and action. Such dissociations areparticularly common in patients with lesions of the frontallobe: such patients' behaviour tends to be ‘perseverative’, withrepetition of an action despite its no longer being appropriate(Milner, 1963; Sandson and Albert, 1984); some patients evencomment that they are aware that their behaviour does notmatch their intention (Luria, 1966). A recent example isprovided by the patient JR with a lesion of the supplementaryeye field, described by Husain et al. (2003). Following a changein the rule governing the appropriate eye movement response
to a visual stimulus, JR tended to respond according to theprevious rule before self-correcting the response to reflect thecurrent rule; thus, while he clearly understood the task andwas able to execute the appropriate response, his perfor-mance, at least initially, was still more in line with hisprevious actions than with his current intentions. Suchinstances are not restricted to those with brain damage,however. Duncan et al. (1996) described the phenomenon of‘goal neglect’ among healthy individuals, especially thosewith low general intelligence, whereby instructions which arefully understood are simply not acted upon. In that study,subjects failed to switch attention to an alternative stream ofvisual input in a letter monitoring task, and Duncan et al.pointed out that it is in leaving behind a line of action to switchto another that such failures of intention to control action tendto occur.
B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3
! Corresponding author.E-mail address: [email protected] (D.E. Astle).
0006-8993/$ – see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.brainres.2006.09.092
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res

Autho
r's p
erso
nal
copy
The experimental ‘task-switching’ paradigm has beenmuch used in recent years to examine the nature and limitsof our dynamic control over behaviour. One of the mostintriguing findings from this research is the phenomenon ofthe ‘residual switch cost’ (Rogers and Monsell, 1995). This is acost to performance in terms of speed and/or accuracy whichis incurred on trials where the rule governing respondingdiffers from that used on the previous trial. Crucially, this costcannot be overcome by voluntary preparation undertaken inadvance of the target. The existence of the residual switch costseems to indicate that performance of a task at least once isnecessary to enable subsequent task performance to beefficient, and it appears to demonstrate that even in healthyindividuals of high intelligence, our recent actions have aneffect upon the efficiency of current performance which isinescapable, despite the strength of our current intentions.1
In their work on cognitive control using the task-switchingparadigm,Mayr and Keele (2000) put forward evidence that theresidual switch cost is at least partly due to the presence ofwhat they termed ‘backward inhibition’. By this, they referredto the apparent inhibition of a task set, which was abandonedin order that an alternative task set could be used instead. Theexistence and persisting nature of such inhibition can beinferred from the slowing of response times seen when arecently abandoned task set is switched back to. Mayr andKeele argued that backward inhibition is an executive controlprocess, driven by our intentions to perform a specific taskfrom among a number of competing alternatives. Interest-ingly, although the presence of backward inhibition appearedto depend upon a high-level intention, such as a verbalrepresentation in working memory, apparently it could notbe overcome by an analogous voluntary mechanism. Instead,it survived long preparatory intervals during which theswitched-to task was being prepared. This led Mayr andKeele to highlight the apparent functional dissociationbetween two levels of representation of a task set: a high-level representation which is necessary for eliciting inhi-bition but apparently powerless to remove it, and a lower-level, ‘effective’ representation, which is itself modulated byinhibition.
Despite the existence of such dissociations, most of theresearch into cognitive control has not attempted to distin-guish between the effects of an intention to perform aparticular task and the effects of previous performance. Inall of the examples above, for instance, performance wasmeasured after a switch from an alternative task (or goal, oraction) which was actually carried out, so it is not clearwhether the same behavioural effects would occur when aswitch was required from an alternative course of actionwhich was only intended rather than actually executed.However, a recent study by Schuch and Koch (2003) cleverly
dissociated preparation (intention) from performance in atask-switching paradigm and showed that it is actually theperformance (i.e., selection of a task-specific response inrelation to a target stimulus), not simply the preparation, of analternative task on the previous trial which is responsible forthe existence of switch costs on the current trial. Using acombined task-precueing and ‘go/no-go’ paradigm, theyinduced participants to prepare for a particular task on everytrial, but on 25% of trials, without warning, a target waspresented which required no response (a ‘no-go’ trial). Whilerobust residual switch costs were observed following ‘go’trials, they were absent following ‘no-go’ trials; further, therewas evidence that these residual costs were the result ofovercoming inhibition of the current task, and that thisinhibition had been triggered by performance – but not bypreparation alone – of the alternative task on the previoustrial.
So far we have seen that there can be a dissociationbetween our intentions and actions, in that these can exertdifferential effects upon subsequent performance. But thisonly covers the effect of the dissociation upon observedperformance—what about the potential effects upon prepara-tion processes? Schuch and Koch (2003) found no costs toperformance following ‘no-go’ trials, but it is still an openquestion as to whether preparation for a subsequent taskswitch is affected by prior response selection. This question isdifficult to address using behavioural measures alone as thesenecessarily require performance, but it can be tackled usingthe event-related potential (ERP) technique. In this study, weadopted Schuch and Koch's ‘go/no-go’ task-switching proce-dure to enable us to dissociate task switches away fromalternative tasks which were only prepared and those fromtaskswhichwere also performed, andwe used ERPs to provideindices of neural activity during the preparatory periodseparately from that occurring during task performance.
2. Results
2.1. Behavioural data
2.1.1. PreparationIn order to ascertain whether or not participants were usingthe precue actively to prepare in advance of the target, switchcosts at the short-CTI were compared to those at the long-CTIusing a two-way ANOVA with factors CTI and switching(including only accurate ‘go’ trials preceded by accurate ‘go’trials). This revealed a significant interaction [F(1,16)=4.60,p=0.048]. Switch-trial RTs were reduced from 754 ms in theshort-CTI condition to 683 ms in the long-CTI condition,whereas repeat-trial RTs were reduced from 658 ms to621 ms. There was a reduction in switch costs withpreparation, therefore, from 96 ms in the short-CTI conditionto 62 ms in the long-CTI condition. It may be the case that byblocking CTIs and placing the short-CTI trials at consistentpositions within a block, subjects may have learned topredict their occurrence and become ‘lazy’, not bothering toprepare rapidly on predicted long-CTI trials (Altmann, 2004).Nevertheless, the significant interaction of the switch andCTI effects shows that on average subjects were making use
1 There is evidence that, given appropriate circumstances whichmaximise the motivation to exercise voluntary control, the cost ofswitching may not affect a proportion of individual trials (De Jong,2000); nevertheless, we seem to be limited in the degree to whichwe can exercise such control trial after trial because overall theswitch cost remains, providing evidence of a persisting effect ofthe previous task upon current performance.
95B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
of the CTI to engage in preparatory switching, and so wewould expect to see electrophysiological correlates of suchprocessing in the precue-locked ERPs of the long-CTI trials.Error data were analysed using Wilcoxon matched-pairstests. There was no difference in the error switch costbetween CTI conditions (Z=!0.43, p=0.670) (proportion errorscores, short-CTI: switch trials, 0.16; repeat trials, 0.12; switchcost 0.04; long-CTI: switch trials, 0.14; repeat trials, 0.08;switch cost, 0.06).
2.1.2. Relationship between response sequence and switchcostsLong-CTI trials (from which the ERPs waveforms wereproduced offline) were used to assess the relationshipbetween response sequence and task switching using a two-way ANOVA. This revealed a significant interaction [F(1,16)=20.308, p<0.001] (Fig. 1A), with significant switch costsfollowing a ‘go’ [F(1,16)=33.125, p<0.001] but not following a‘no-go’ [F(1,16)=2.123, p=0.164]. The difference in performancebetween the trials following a ‘no-go’ and those following a‘go’ was apparent both on repeats [F(1,16)=25.618, p<0.001]and switches [F(1,16)=6.508, p=0.021]. There were no signifi-cant switch-related differences for the error data [p>0.05](Fig. 1B), with the difference in repeat trials and switch trialsbetween the two response sequences being nonsignificant[p>0.05]. The pattern of these behavioural data replicates thatof Schuch and Koch (2003, Experiment 1b).
2.2. ERP results
The analysis of the ERP waveforms was split into two separateepochs: one following the precue (‘precue-locked’), and onefollowing the target (‘target-locked’). In each case, switch and
repeat waveforms were compared with each other, separatelyfor each of the response sequences (GGG and GNG).
2.3. Precue-locked
2.3.1. Late parietal positivity (LPP) for switchesWe observed an increased positivity for switch compared withrepeat trials over parietal electrodes. Fig. 2 shows that thiseffect was present for both the GGG and the GNG responsesequences. There was a similar topography for the effect ineach response sequence condition, clustered around Pz(electrode 62), where it reached consecutive significancerelatively late in the epoch (488–646 ms for GGG, and 472–716 ms for GNG).
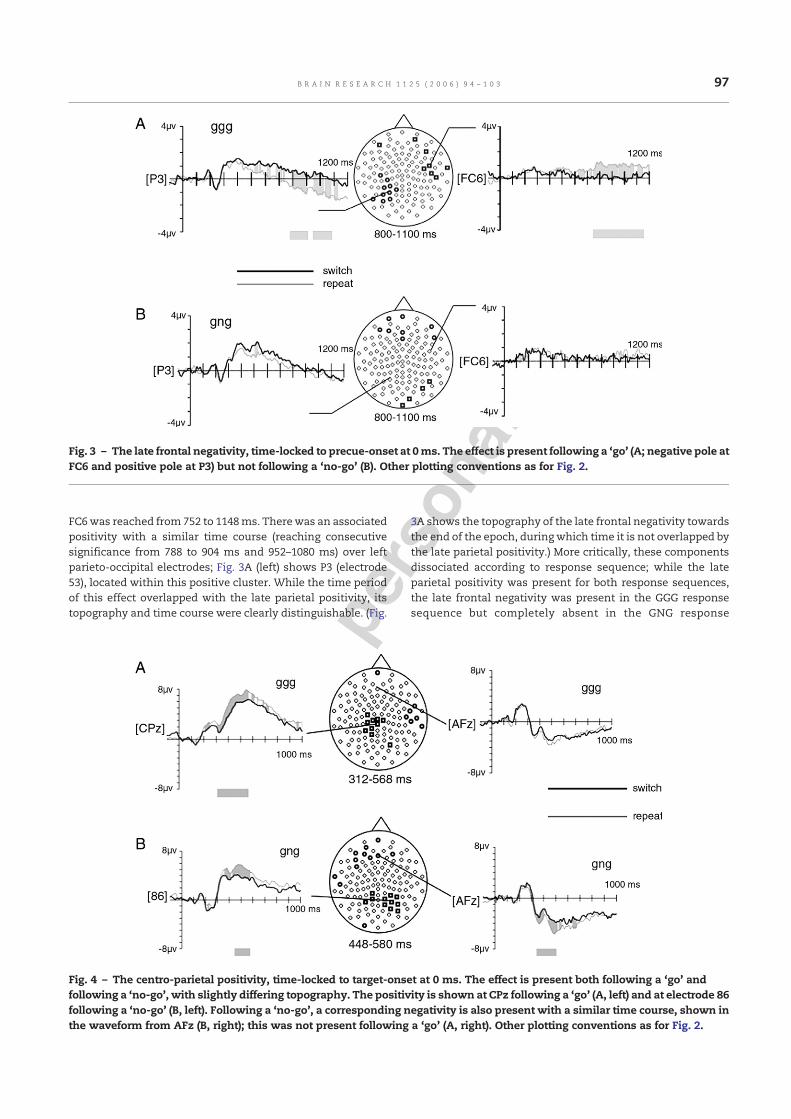
2.3.2. Late frontal negativity (LFN) for switchesA precue-locked late right-frontal negativity for switch versusrepeat trials was seen for the GGG but not the GNG condition.The effect was most prominent over right frontotemporalelectrodes; Fig. 3A (right) shows the effect at FC6 (electrode117). (N.B. Because its time course overlaps with that of thelate parietal positivity, this negativity can also partially beseen in the topographical plot in Fig. 2A.) This effect resemblesa superimposed slow wave somewhat like a contingentnegative variation (CNV) for GGG-switch trials, with a nega-tivity over frontal electrodes towards the end of the epoch (i.e.,towards the time of target onset); consecutive significance at
Fig. 1 – Behavioural data for task switching following a ‘go’(ggg) and following a ‘no-go’ (gng). (A) Mean RT. (B) Meanproportion error.
Fig. 2 – The late parietal positivity, time-locked toprecue-onset at 0 ms. The effect is present both following a‘go’ (A) and following a ‘no-go’ (B). Waveforms are shownfrom electrode Pz, and topographical plots apply to the timeperiod of consecutive significance at Pz. In the topographicalplots, dark circles indicate significantly greater positivity forswitch than repeat trials, and vice versa for dark squares.Arrows indicate the electrode site at which theaccompanying waveforms were recorded.
96 B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3
Autho
r's p
erso
nal
copy
FC6was reached from 752 to 1148ms. Therewas an associatedpositivity with a similar time course (reaching consecutivesignificance from 788 to 904 ms and 952–1080 ms) over leftparieto-occipital electrodes; Fig. 3A (left) shows P3 (electrode53), located within this positive cluster. While the time periodof this effect overlapped with the late parietal positivity, itstopography and time course were clearly distinguishable. (Fig.
3A shows the topography of the late frontal negativity towardsthe end of the epoch, duringwhich time it is not overlapped bythe late parietal positivity.) More critically, these componentsdissociated according to response sequence; while the lateparietal positivity was present for both response sequences,the late frontal negativity was present in the GGG responsesequence but completely absent in the GNG response
Fig. 3 – The late frontal negativity, time-locked to precue-onset at 0ms. The effect is present following a ‘go’ (A; negative pole atFC6 and positive pole at P3) but not following a ‘no-go’ (B). Other plotting conventions as for Fig. 2.
Fig. 4 – The centro-parietal positivity, time-locked to target-onset at 0 ms. The effect is present both following a ‘go’ andfollowing a ‘no-go’, with slightly differing topography. The positivity is shown at CPz following a ‘go’ (A, left) and at electrode 86following a ‘no-go’ (B, left). Following a ‘no-go’, a corresponding negativity is also present with a similar time course, shown inthe waveform from AFz (B, right); this was not present following a ‘go’ (A, right). Other plotting conventions as for Fig. 2.
97B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
condition (Fig. 3B). The pattern of waveforms on switch andrepeat-trial types across the response sequence conditions isinteresting in that it appears (see Fig. 3) that, rather than therebeing an additional frontal negativity for GGG-switch trials,there may in fact be a reduced negativity (or additionalpositivity) for GGG-repeat trials relative to all other trialtypes (and vice versa for the left parieto-occipital positivity).However, additional analyses conducted at electrodes P3 andFC6 showed no significant effect of response sequence oneither repeat trials or switch trials.
2.4. Target-locked
2.4.1. Centro-parietal positivity (C-PP) for repeatsA centro-parietal positivity for repeats was present in bothGGG and GNG sequences. Fig. 4A (left) shows the centro-parietal positivity over CPz (electrode 55) in the GGGcondition (312–568 ms), and Fig. 4B (left) over electrode 86in the GNG condition (448–580 ms); comparison of theirrespective topographies indicates that the GNG centro-positivity was shifted in a right-posterior direction. Fig. 4B(right) also shows an associated negativity over left frontalsites in the GNG condition (344–504 ms at AFz, electrode 16),which was absent in the GGG condition (Fig. 4A, right). (N.B.In the topographical plot for 5A, the lateral positive shift (seebelow and Fig. 5A) is also observable over right hemisphereelectrodes.)
2.4.2. Lateral positive shiftThis component was located over right-temporal sensors,starting relatively early in the epoch, with a gradually increa-
sing switch-versus-repeat difference (with switch more posi-tive than repeat), reaching consecutive significance at 388–496 ms, 572–640 ms, 716–756 ms and 932–996 ms at T8(electrode 109). Fig. 5 demonstrates that this was present inthe GGG condition but absent in the GNG condition.
The lateral positive shift has been reported previously.Swainson et al. (2003) observed something very similar in thetarget-locked epoch of a predictable task-switching design,where the component was visible as a positivity at lateralelectrodes over both hemispheres. Those authors concludedthat while this might appear to be a distinct component in itsown right, it is likely the result of re-baselining on presenta-tion of the target. In the present data set this is also plausible,with electrode T8, where the target-locked lateral positiveshift is clearly seen, also showing a clear precue-lockednegativity for switch trials, part of the precue-locked latefrontal negativity (shown in Fig. 3).
3. Discussion
This study explored the effect of dissociating task perfor-mance from task preparation upon subsequent task switch-ing. Using a precued design, with ‘no-go’ responses requiredon 25% of trials, residual switch costs were found to be absentfollowing a trial on which a task was only prepared and notactually performed (i.e., following a ‘no-go’ response). Of moreimportance, preparatory activity evident in ERPs occurringbefore target onset was shown to be affected by previous taskperformance. While a switch-related late parietal positivity(LPP) was present in the precueing interval following both ‘go’and ‘no-go’ responses, a switch-related late frontal negativity(LFN) in the same interval was only found following a ‘go’response, with repeat and switch not significantly differingfollowing a ‘no-go’. A target-locked centro-parietal positivityfor repeats was present following both ‘go’ and ‘no-go’responses.
We replicated the results of Schuch and Koch (2003),showing that switch costs are absent when a ‘no-go’ responsewas made on the previous trial (i.e., the task was prepared,but not performed). Our data showed the same pattern as inthat original study, with the absence of switch costs followinga ‘no-go’ apparently being due both to an additional cost onrepeat trials and a benefit on switch trials. A number offactors could explain the additional cost on repeat trials, forinstance: elimination of task-set priming (Allport and Wylie,1999); addition of a ‘restart cost’ (Allport and Wylie, 2000;Altmann, 2002); and lack of priming of the ‘go’ response(Schuch and Koch, 2003). We suggest a further alternative,which is that in this type of paradigm, switchingmay occur ontwo levels: the ‘task-set’ level (consonant/vowel versus upper/lower case judgements) and the ‘response-set’ level (‘go’versus ‘no-go’ responding). In order to produce the infrequent‘no-go’ response, subjects probably have to inhibit thetendency to respond immediately upon target onset (the ‘go’response set). When a subsequent ‘go’ response is required,therefore, such inhibition (if it persists) would have to beovercome, potentially producing a cost to performance (seeKoch et al., 2004, for evidence of persisting inhibition ofresponse mode). Whichever of these explanations is true, it
Fig. 5 – The lateral positive shift, time-locked to target-onsetat 0 ms. The effect is present following a ‘go’ (A), but notfollowing a ‘no-go’ (B). Other plotting conventions as forFig. 2.
98 B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
ought to affect both task-switch and task-repeat trials in aGNG sequence. But critically its effect might be counteractedon switch trials by an additional relative benefit—i.e.,absence of the need to overcome task-set inhibition whenno response was selected on the previous trial. Clearly therewas an additional benefit on switch trials in this studybecause there was a significant benefit to performance of apreceding ‘no-go’ on switch trials. In addition, the error dataought to be sensitive to effects of switching ‘task set’ ratherthan those of switching ‘response set’ (in this case, switch-ing from not responding to responding), and these showedeffects only on switch trials. Thus, being in the inappropri-ate ‘task set’ would lead to erroneous responses (responsesbeing driven by the inappropriate task on incongruenttrials), so eliminating the need to switch tasks shouldreduce error rates on switch trials. But inappropriatelyretaining a tendency to inhibit immediate responding (orremaining in a ‘no-go’ response set) ought to lead only todelayed responding rather than increased errors. Whilenonsignificant, this is the pattern (i.e., with a largerdifference on switch trials than repeat) that we observe inthe error data.
3.1. ERP switch effects modulated by prior taskperformance
The important finding in the current study is that the neuralcorrelates associated with preparing to switch versus repeatthe current task on the next trial are different following a ‘no-go’ versus a ‘go’ response. The LFN ERP, seen during thepreparatory interval for switch trials in comparison to repeattrials, was present only for the GGG sequence and not for theGNG sequence. Therefore, it is clear that when a prepared taskis performed (used to produce a task-specific response to atarget stimulus) as opposed to only being prepared, theimplications of this are evident during subsequent prepara-tion of an alternative task set on the next trial (as shown by theLFN effects) as well as at the time of performance (as shown bythe behavioural effects).
Although this issue was not explicitly addressed in thecurrent study, there is reason to speculate that the specificityof the LFN effect to GGG sequences relates to the need toovercome inhibition of the switched-to task. Schuch andKoch (2003) showed that following a ‘no-go’ trial, not onlywere switch costs removed, but also that the specific cost ofswitching back to a task abandoned on the previous trial (i.e.,the cost of performing an ABA relative to a CBA tasksequence) was removed. They suggested that ‘backwardinhibition’ (Mayr and Keele, 2000) of a previous task istriggered at the time of response selection for the new taskand not during the preceding preparation period. Theconsequence of this would be that following a ‘no-go’ trial(on which a new task was only prepared and never actuallyperformed), there would be no cost of switching back to theoriginal task (which had been abandoned on the previous,‘no-go’, trial) because there would be no residual inhibition ofthat task to be overcome. In terms of the dissociation ofpreparatory and performance-based processes upon taskinhibition, then, our behavioural data support the view thatinhibition is neither triggered by preparation (switch costs
being absent following a ‘no-go’ response), nor can it beentirely overcome during the preparation interval (residualswitch costs remaining robustly in the GGG condition despitea substantial precue-target interval). However, the ERPfindings from our study – specifically, the presence of theprecue-locked LFN for GGG but not GNG sequences – suggestthat this residual inhibition may nevertheless be encounteredduring the time when one prepares to return to the previoustask set, and not only at the time of preparing a specificresponse appropriate to that task.
Although we found no significant differences betweenthe two response sequences (GGG versus GNG) on eitherrepeat or switch trials in terms of the LFN waveforms, Fig. 3does suggest that the presence of a task-switch effect onGGG trials may be due to a reduced negativity on GGG-repeat trials at least as much as an increased negativity onGGG-switch trials. How can this be squared with theexplanation offered above that the effect on GGG trialsmay be due to encountering inhibition of a task set on aGGG-switch trial? As discussed above, it may be that thesame need to inhibit a contradictory tendency (or ‘set’)applies to both tasks and types of response. Therefore,performing both repeat and switch trials in the GNGcondition may entail encountering response suppression(but neither task suppression), whereas – if the GNGsequence does indeed prevent the need for persistinginhibition of the switched-to task – only switch trials inthe GGG condition would entail encountering task suppres-sion (but not response suppression). Thus, it is consistentwith our hypothesis (that the LFN reflects processes to dowith encountering persisting inhibition) that a similardegree of negativity should be present in both GNG-trialtypes as in GGG-switch trials, and more than on GGG-repeattrials (when neither persisting response suppression nortask suppression are encountered).
As yet, we cannot be specific as to a potential role of theLFN. For instance, if it is related to the presence of persis-ting ‘backward inhibition’ upon the (recently abandoned)switched-to task, it may index an active attempt to overcomeinhibition, a passive result of encountering inhibition duringpreparation, or the recruitment of resources needed toovercome inhibition once the target is presented. The effectresembles a slow wave (negative over frontal scalp) super-imposed upon the underlying waveforms on switch trials inthe GGG condition. Such slow waves have been associatedwith the allocation of cognitive resources to a particularprocessing module in the cortex (Rosler et al., 1997). One suchslow wave is the contingent negative variation (CNV)—i.e., anegativity which increased towards the time of onset of apredictable stimulus (Walter et al., 1964). An increased frontalCNV has been shown on trials where subjects made aparticular ‘effort’, in anticipation of a target, to perform welland has been interpreted as a reassignment of resources(Falkenstein et al., 2003). The LFN seen here on switch trialsrelative to repeat, following a ‘go’, may similarly representthe recruitment of resources, ready to deal with generating aresponse to a difficult type of target—i.e., one which it isknown will involve a switch in task and the overcoming ofpersisting backward inhibition. We have also seen this effectrecently (Mueller, Swainson and Jackson, submitted) when
99B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
participants switched between tasks which shared responsemeanings (i.e., where responses were bivalent with respect totasks—a left-hand response, for example, being used toindicate either that a letter was a vowel or that a digit wasan odd number) but not between tasks which used separatesets of (univalent) responses. If the LFN does relate toovercoming inhibition of a recently abandoned switched-totask, it may index the need specifically to overcome inhi-bition of the response meanings associated with thattask.
3.2. ERP effects independent of prior task performance
A commonly reported ERP effect associated with task switch-ing is a late positivity over parietal scalp (Barcelo, 2003;Jackson et al., 2001; Karayanidis et al., 2003; Kieffaber andHetrick, 2005; Nicholson et al., 2005; Rushworth et al., 2002;Swainson et al., 2003, 2006) and this effect was replicated inthe present study. Like the LFN effect discussed above, thislate parietal positivity (LPP) occurred prior to the target onsetduring the precue-target interval, but, unlike the LFN, it wasevident following both ‘go’ and ‘no-go’ responses. Thus, itappears to index a process which is independent of switchingbetween performing alternative tasks per se. This componentappears to occur at the time within a trial when ‘endogenous’reconfiguration would be expected to take place—i.e., thataspect of task switching which, it is postulated, can beperformed prior to target-onset, given certain necessaryconditions such as advance information and the expectationof sufficient time in which to use it, but which otherwiseoccurs after target onset (Rogers and Monsell, 1995). Thus, thecomponent has been seen both prior to target onset (Nichol-son et al., 2005; Rushworth et al., 2002) and subsequent totarget onset (Jackson et al., 2001; Swainson et al., 2003, 2006). Infact, its timing seems to be relatively consistent in relation tothe timing of the task cue in these studies. Therefore, we havesuggested that it reflects processing driven by a cue signalingthe need for a task switch (Swainson et al., 2006). (N.B. In thestudy by Karayanidis et al., 2003, tasks switched predictablyand the positivity for task switching was observed to be time-locked to the preceding response, but a task-cueing grid waspresent throughout and the positivity may in fact have beentime-locked to participants’ attending to the grid. The effectmay prove eventually not to be specific to the processing of anexternal cueing stimulus, but none of the studies so far haveshown this to be the case.) The component has a relativelylong latency after precue-onset of around 500–700 ms and inour previous study, where response times were very fast(usually <500 ms), we showed that this component need notprecede the response; therefore, we suggested that it mayindex a nonobligatory switch-related process, such as a shiftfrom controlled to relatively automatic task processing ratherthan an obligatory reconfiguration allowing performance ofthe appropriate task (Swainson et al., 2006). The current dataare consistent with the effect being related to processing of atask cue. Whether this is generated exclusively by parietalregions, or whether this component is composed of theactivity of a number of generators (for instance, includingthe prefrontal cortex; Brass et al., 2005) remains unclear at thisstage. The similarity of the effect between the GGG and the
GNG response sequence conditions indeed implies that theeffect reflects switching at the level of the abstract task set,goal or intended task put in place by the task cue on theprevious trial whether or not that taskwas actually performed,rather than at a level corresponding to the ‘effective task’ andwhich would therefore be dependent upon recent perfor-mance (Mayr and Keele, 2000).
A second component observed regardless of whether therewas a switch in the performed task was the target-lockedcentro-parietal positivity (C-PP) for task repetition. While thiswas observed in both the GGG and GNG response sequenceconditions, there was nevertheless a difference in topography,although to what extent this difference may reflect differentpsychological process occurring following a ‘go’ and followinga ‘no-go’ response is unclear. We have previously observedthis component in the target-locked epoch of a predictabletask-switching paradigm (Swainson et al., 2003, 2006) andhave interpreted it as indexing the current ‘strength’ of task-specific stimulus evaluation, such that when a task isperformed with the knowledge that it will immediately berepeated, it may be actively consolidated, or maintained,thereby allowing improved performance on the subsequenttrial.Wylie et al. (2003) also observed the effect in a predictabletask sequence with three trials in each run (…AAABBBAAA…)where they saw its amplitude increase along with the numberof repetitions. They attributed this increase to a decreasingconflict between the task sets with task repetition, consistentwith a parallel improvement in performance. Similarly,Barcelo (2003) saw that the component increased with taskrepetition and with improved performance on repeat trials.We have previously found the effect to be specific to apredictable task sequence, with no C-PP evident for repeatversus switch trials in random task sequences (Swainson etal., 2006), and suggested that this was because a task set couldpotentially be more rapidly consolidated when subsequentrepetitions were predictable. The data from the present studyand those from Barcelo (2003) indicate, however, that aftermore than one repeat trial, the effect can be observed withoutrepeat trials being predictable (in the current study, all repeattrials occurred after at least two previous trials with the sametask, which was not the case in our previous study). Thisimplies that the same mechanism of task-set consolidationwhich previous data suggests is availablewith predictable taskrepeats still occurs (only more gradually) even withoutpredictability, or at least that the two mechanisms have thesame effect in terms of increasing the C-PP. Unlike theseprevious studies, though, in the present data the componentwas not seen to have direct behavioural consequences. Thatis, in the previous studies an increased C-PP was seen tocoincide with a behavioural repetition benefit, but in the GNGcondition of the current study, despite there being a switch/repeat modulation in the C-PP, there was no behavioural‘repeat benefit’ (or in other words, no switch cost). We suggestthat this may be because the component indexes consolida-tion at a perceptual level, producing speeded stimulusevaluation on repeat trials (Kok, 2001). This would notnecessarily also lead to a speeding of response times,especially if slowing for another reason – such as the need toovercome a ‘no-go’ response tendency – renders speeding of aparallel process impotent to affect response times. It is unclear
100 B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
why the topography of this effect differed between theresponse sequence conditions. It is plausible that the sameconsolidation process occurs both following a ‘go’ and ‘no-go’response yet reaches significance over different electrodes,perhaps due to there being additional processes occurringfollowing a ‘no-go’which affect its observed scalp distributionsuch that its negative pole becomes apparent over frontalsites. Conversely, the difference in topographies could indi-cate that genuinely different psychological processes occur inthe two response sequence conditions, for instance thatfollowing a ‘no-go’ there is a qualitative difference in thetask-consolidation mechanism. As yet either alternative ispossible.
3.3. The relationship between response selection andtask-set preparation
While the LPP seems to be relatively independent fromwhether or not subjects switch between performing (asopposed to only preparing) alternative tasks on successivetrials, this study provides clear evidence that the effects ofprevious action are encountered during preparation for thesubsequent trial (in the form of the LFN). The combination oftask-switching and ‘no-go’ paradigms demonstrates that it ispossible to dissociate neural indices of cognitive control intask switching, between those that relate to changing perfor-mance per se and those involved in changing intendedperformance.
4. Experimental procedures
4.1. Subjects
Eighteen healthy right-handed participants completed acombined EEG and behavioural experiment. Fourteen werefemale, with an average age of 23.63±3.55 (SD) years.Participants provided written informed consent. Eighteensubjects were included in the EEG data set, and 17 in thebehavioural (due to a technical error meaning that somebehavioural data were lost for one subject).
4.2. Behavioural task
Each trial was precued by a fixation cross which changedcolour prior to the onset of the imperative stimulus. Thecolour of the target and fixation cross indicated the task set tobe prepared (the assignment of colour to task being counter-balanced across participants). Each target stimulus wasselected from a set of vowels and consonants in upper andlower case (A,E,I,U,a,e,i,u,G,B,T,D,g,b,t,d). On a ‘go’ trial a singleletter was presented, upon which either of two task sets couldbe implemented, responses being made with a two-buttonserialmouse: discrimination of the letter as either a vowel (leftbutton) or a consonant (right button) or discrimination of theletter as lower (left button) or upper (right button) case. On aquasi-randomly selected 25% of trials, a task-irrelevantstimulus (#) appeared instead of the usual target, to whichno response could be made (‘no-go’ trials). These trials neverappeared in direct succession.
4.3. Experimental design
After practice at performing each task separately and switch-ing between the two (until each participant had achieved 90%accuracy on a ‘switching’ block), the participant performed 17blocks of 60 trials. Each block contained both short (100 ms)and long (1200 ms) CTI (cue-target interval) trials, with theshort-CTI trials being arranged in a short run of 7–9 trials atthe beginning of each block, and again at the midpoint.(These trials were blocked rather than interleaved to avoidintroducing an additional factor of switching between longand short trials.) Participants were not informed of thispattern, only that the time to prepare could be short or longon each trial. Each cue (in fact, a ‘precue’, as all cues werepresented in advance of targets) was on the screen for the fullduration of the CTI, and each target was displayed on thescreen for 160 ms. Following each target there was a responsewindow of 800 ms, which ended upon response execution.This was followed by a subsequent fixed-length responsewindow of either 800 or 1900, depending upon CTI, such thateach trial time lasted 2000 ms+RT regardless of the prepara-tion interval.
ERPs were generated only from long-CTI trials which wereresponded to accurately and which were also preceded by twoother accurate long-CTI trials. The short-CTI trials wereincluded to provide subjects with incentive to engage inpreparation immediately on presentation of the precue and toprovide reaction time information for this to be assessed. As inSchuch and Koch's (2003) study, there were no repeats of ‘no-go’ trials; thus, trials fell at the end of either a go/go/go (GGG)or a go/no-go/go (GNG) ‘response sequence’ triad. But in amodification to their original design, we imposed anotherconstraint that the task pertaining to the third trial in thesetriads also pertained to the first (see Fig. 6). Thus, there waseither a switch back to the task performed two trials ago(switch triads: ABA, where A and B are alternative task sets) orno switch at all within the triad (repeat triads: AAA). Thereason for this was in the cases where trial n!1 required a ‘no-
Fig. 6 – The four specific trial sequences used in the mainanalyses: two repeat and two switch sequences, two with a‘no-go’ on trial n!1 and twowith a ‘go’ on trial n!1. White orblack precues and stimuli indicate one or other of the twotasks ought to be performed on that trial.
101B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
go’ response, neither the ‘switch’ nor the ‘nonswitch’ triadswould entail a switch at the level of performance. The fourtypes of trial included in the analysis were therefore GGG-switch, GGG-repeat, GNG-switch and GNG-repeat. In order toensure that enough of the these specific sequences wereproduced, the trials were generated in a quasi-random fashionusing a software package (Microsoft Excel), giving on average 6of each trial type per block of 60 trials. (N.B. For the analysis ofthe behavioural preparation effect, such a strict criterion oftrial type did not need to be employed. Instead, trials wereclassified as being either switch (BA) or repeat (AA) trials,where both the current and preceding trial required a ‘go’response (GG)).
4.4. EEG recording, processing and ERP formation
EEG was recorded throughout each block, using a 128-channelelectrical geodesic net (Electrical Geodesics, Inc.; Tucker et al.,1994) and digitised at 250 Hz. The recording was performedwith a hardware bandpass filter of 0.01 Hz to 100 Hz. Beforerecording impedance on each of the 128 electrodes wasreduced to <50 k!.
Precue-locked and target-locked epochs were created, witheach epoch starting 100 ms before onset of the respectivestimulus and ending 1200 ms (precue-locked) or 1000 ms(target-locked) afterwards. Segments were rejected if con-taminated by eye-blinks/movements (indicated by EOG activ-ity greater that 70 "v), or an error of response (incorrectresponse or omission of response), or if the trial followed anerror on either of the two previous trials. Trials containingvoltage amplitudes greater than 200 "v or a change greaterthan 100 "v were also removed. Channels that were bad formore than 20% of trials were rejected. With this rationale,approximately on average 20% of trials were removed withinthe precue-locked epoch, and 35% of trials from the target-locked epoch (owing to excessive ocular artefacts). The datafrom each participant included in any ERP waveform had >30waveforms from which to form averages. Based upon thisrationale all 18 participants produced sufficient waveforms tobe included in the precue-locked comparisons, whereas only15 participants produced sufficient to be submitted to thetarget-locked comparisons. In summary, ERPs were formedfrom long-CTI trials only, were free from behavioural errors,and had been preceded by two appropriate (see Fig. 6),accurate and long-CTI trials.
The specific segments were average-referenced to a stan-dard adult 128-electrode montage. Epochs were baseline-corrected for the first 100 ms, before the onset of the stimulus,and epochs lasted for a subsequent 1200ms or 1000ms. Switchand repeat waveforms were explicitly compared (all statisticsconducted using MATLAB v5, scripts developed in-house), bymeans of a T-test for every 4ms sample. Twowaveforms werecounted as significantly different if this significance persistedfor more than 10 samples, and two temporal regions ofconsecutive significance were counted as a continuation ofthe samedifference if the gap between themwas fewer than 10samples. This approach is based upon Rugg et al. (1993) andwas used by Swainson et al. (2003, 2006); Jackson et al. (2001).All ERP comparisons that reached consecutive significance areincluded in the Results section.
Acknowledgment
This study was supported by a BBSRC grant to GMJ.
R E F E R E N C E S
Allport, A., Wylie, G., 1999. Task-switching: positive and negativepriming of task-set. In: Humphreys, G.W., Duncan, J.,Treisman, A.M. (Eds.), Attention, Space and Action: Studiesin Cognitive Neuroscience. Oxford Univ. Press, Oxford, UK,pp. 273–296.
Allport, A., Wylie, G., 2000. ‘Task-switching’, stimulus-responsebindings and negative priming. In: Monsell, S., Driver, J. (Eds.),Attention and Performance XV: Conscious andNonconscious Information Processing. MIT Press,Cambridge, MA, pp. 421–452.
Altmann, E.M., 2002. Functional decay of memory for tasks.Psychol. Res. 66 (4), 287–297.
Altmann, E.M., 2004. The preparation effect in task switching:carryover of SOA. Mem. Cogn. 32 (1), 153–163.
Barcelo, F., 2003. The Madrid card sorting test (MCST): a taskswitching paradigm to study executive attention withevent-related potentials. Brain Res. Brain Res. Protoc. 11 (1),27–37.
Brass, M., Ullsperger, M., Knoesche, T.R., von Cramon, D.Y.,Phillips, N.A., 2005. Who comes first? The role of the prefrontaland parietal cortex in cognitive control. J. Cogn. Neurosci. 17 (9),1367–1375.
De Jong, R., 2000. An intention-activation account of residualswitch costs. In: Monsell, S., Driver, J. (Eds.), Control ofCognitive Processes: Attention and Performance XVIII. MITPress, Cambridge MA, pp. 357–376.
Duncan, J., Emslie, H., Williams, P., Johnson, R., Freer, C., 1996.Intelligence and the frontal lobe: the organization ofgoal-directed behavior. Cogn. Psychol. 30 (3), 257–303.
Falkenstein, M., Hoormann, J., Hohnsbein, J., Kleinsorge, T., 2003.Short-term mobilization of processing resources is revealed inthe event-related potential. Psychophysiology 40 (6), 914–923.
Husain, M., Parton, A., Hodgson, T.L., Mort, D., Rees, G., 2003.Self-control during response conflict by human supplementaryeye field. Nat. Neurosci. 6 (2), 117–118.
Jackson, G.M., Swainson, R., Cunnington, R., Jackson, S.R., 2001.ERP correlates of executive control during repeated lanaguageswitching. Biliguilism: Lang. Cogn. 4 (2), 169–178.
Karayanidis, F., Coltheart, M., Michie, P.T., Murphy, K., 2003.Electrophysiological correlates of anticipatory andpoststimulus components of task switching. Psychophysiology40 (3), 329–348.
Kieffaber, P.D., Hetrick, W.P., 2005. Event-related potentialcorrelates of task switching and switch costs.Psychophysiology 42 (1), 56–71.
Koch, I., Gade, M., Philipp, A.M., 2004. Inhibition of response modein task switching. Exp. Psychol. 51 (1), 52–58.
Kok, A., 2001. On the utility of P3 amplitude as a measure ofprocessing capacity. Psychophysiology 38 (3), 557–577.
Luria, A.R., 1966. Higher Cortical Functions in Man(B. Haigh, Trans), 2nd ed. Basic Books, Inc., New York. revisedand expanded ed.
Mayr, U., Keele, S.W., 2000. Changing internal constraints onaction: the role of backwards inhibition. J. Exp. Psychol. Gen.129, 4–26.
Milner, B., 1963. Effects if different brain lesions on card sorting.Arch. Neurol. 9, 100–110.
Nicholson, R., Karayanidis, F., Poboka, D., Heathcote, A.,Michie, P.T., 2005. Electrophysiological correlates of
102 B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3

Autho
r's p
erso
nal
copy
anticipatory task-switching processes. Psychophysiology42 (5), 540–554.
Rogers, R.D., Monsell, S., 1995. The costs of a predictable switchbetween simple cognitive tasks. J. Exp. Psychol. Gen. 124,207–231.
Rosler, F., Heil, M., Roder, B., 1997. Slow negative brain potentialsas reflections of specific modular resources of cognition. Biol.Psychol. 45 (1–3), 109–141.
Rugg, M.D., Doyle, M.C., Melan, C., 1993. An event-related potentialstudy of the effects of within-and across-modality wordrepetition. Lang. Cogn. Processes 8 (4), 357–377.
Rushworth, M.F., Passingham, R.E., Nobre, A.C., 2002. Componentsof switching intentional set. J. Cogn. Neurosci. 14 (8), 1139–1150.
Sandson, J., Albert, M.L., 1984. Varieties of perseveration.Neuropsychologia 22 (6), 715–732.
Schuch, S., Koch, I., 2003. The role of response selection forinhibition of task sets in task shifting. J. Exp. Psychol. Hum.Percept. Perform. 29 (1), 92–105.
Swainson, R., Cunnington, R., Jackson, G.M., Rorden, C., Peters,A.M., Morris, P.G., Jackson, S.R., 2003. Cognitive controlmechanisms revealed by ERP and fMRI: evidence fromrepeated task-switching. J. Cogn. Neurosci. 15 (6),785–799.
Swainson, R., Jackson, S.R., Jackson, G.M., 2006. Using advanceinformation in dynamic cognitive control: an ERP study oftask-switching. Brain Res. 61–72.
Tucker, D.M., Liotti, M., Potts, G.F., Russell, G.S., Posner, M.I., 1994.Spatiotemporal analysis of brain electrical fields. Hum. BrainMapp. 1, 134–152.
Walter, W.G., Cooper, R., Aldridge, V.J., McCallum, W.C., Winter,A.L., 1964. Contingent negative variation: an electric sign ofsensorimotor association and expectancy in the human brain.Nature 203, 380–384.
Wylie, G.R., Javitt, D.C., Foxe, J.J., 2003. Task switching: ahigh-density electrical mapping study. NeuroImage 20 (4),2322–2342.
103B R A I N R E S E A R C H 1 1 2 5 ( 2 0 0 6 ) 9 4 – 1 0 3




















