
dissertacijaljaminaa.v..pdf - Южно-Уральский ...
241
Федеральное государственное бюджетное образовательное учреждение высшего образования «Самарский государственный медицинский университет» Министерства здравоохранения Российской Федерации На правах рукописи Лямин Артем Викторович НОВЫЕ ПОДХОДЫ К КУЛЬТИВИРОВАНИЮ И ИДЕНТИФИКАЦИИ КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА ACTINOMYCETALES, ВЫДЕЛЕННЫХ ИЗ КЛИНИЧЕСКОГО МАТЕРИАЛА 03.02.03 – Микробиология Диссертация на соискание ученой степени доктора медицинских наук Научный консультант: Жестков Александр Викторович Заслуженный деятель науки РФ, доктор медицинских наук, профессор Самара – 2020
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of dissertacijaljaminaa.v..pdf - Южно-Уральский ...

Федеральное государственное бюджетное образовательное учреждение высшего
образования «Самарский государственный медицинский университет»
Министерства здравоохранения Российской Федерации
На правах рукописи
Лямин Артем Викторович
НОВЫЕ ПОДХОДЫ К КУЛЬТИВИРОВАНИЮ И ИДЕНТИФИКАЦИИ
КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА
ACTINOMYCETALES, ВЫДЕЛЕННЫХ ИЗ КЛИНИЧЕСКОГО МАТЕРИАЛА
03.02.03 – Микробиология
Диссертация на соискание ученой степени
доктора медицинских наук
Научный консультант:
Жестков Александр Викторович
Заслуженный деятель науки РФ,
доктор медицинских наук, профессор
Самара – 2020

2
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ 8
ГЛАВА 1. КИСЛОТОУСТОЙЧИВЫЕ ПРЕДСТАВИТЕЛИ ПОРЯДКА
ACTINOMYCETALES: КУЛЬТИВИРОВАНИЕ, ИДЕНТИФИКАЦИЯ И
КЛИНИЧЕСКОЕ ЗНАЧЕНИЕ (обзор литературы)
22
1.1. Общая характеристика и современная классификация представителей
порядка Actinomycetales
22
1.2. Особенности культивирования отдельных представителей порядка
Actinomycetales
26
1.3. Методы идентификации представителей семейств Mycobacteriaceae,
Nocardiaceae, Gordoniaceae, Tsukamurellaceae, Brevibacteriaceae,
Promicromonosporaceae, Streptomycetaceae
31
1.4. Эпидемиологические особенности и клиническое значение
представителей семейств Mycobacteriaceae, Nocardiaceae, Gordoniaceae,
Tsukamurellaceae, Brevibacteriaceae, Promicromonosporaceae,
Streptomycetaceae
38
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ 46
2.1. Дизайн исследования 46
2.2. Характеристика исследуемых микроорганизмов 48
2.3. Методы выделения и первичной идентификации микроорганизмов при
первичном посеве клинического материала
49
2.3.1. Методика первичного посева материала при обследовании на
туберкулез
49
2.3.2. Методика определения роста контаминирующей микрофлоры 50
2.3.3. Методика определения роста микобактерий на плотных яичных средах 50
2.3.4. Методика определения роста микобактерий на жидкой среде 52
2.3.5. Метод ДНК-гибридизации для первичной идентификации
нетуберкулезных микобактерий
53
2.4. Методика определения видового разнообразия микрофлоры,
выделенной при обследовании на туберкулез
54

3
2.4.1. Методы выделения контаминирующей микрофлоры и
нетуберкулезных микобактерий для последующей идентификации
55
2.4.2. Метод MALDI-ToF масс-спектрометрии для идентификации
микроорганизмов
55
2.5. Молекулярно-генетические методы идентификации микобактерий 58
2.6. Методы оценки культивирования кислотоустойчивых представителей
порядка Actinomycetales на агаризованных средах
58
2.6.1. Метод количественных испытаний агаризованных сред и испытаний
на селективность
58
2.6.2. Метод предварительной идентификации кислотоустойчивых
представителей порядка Actinomycetales при использовании универсальной
хромогенной среды
60
2.6.3. Метод использования универсальной хромогенной среды с
селективной добавкой для культивирования кислотоустойчивых
представителей порядка Actinomycetales
60
2.6.4. Методы оценки возможности выделения кислотоустойчивых
актиномицет из клинического материала
60
2.7. Методы определения состава питательных сред 61
2.8. Методы математической обработки полученных данных 61
2.8.1. Методы анализа белковых профилей, полученных с помощью MALDI-
ToF масс-спектрометрии
61
2.8.2. Методы статистической обработки полученных результатов 62
ГЛАВА 3. СТРУКТУРА И РАСПРОСТРАНЕННОСТЬ
КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА
ACTINOMYCETALES В КОНТАМИНИРУЮЩЕЙ МИКРОФЛОРЕ,
ВЫДЕЛЕННОЙ ИЗ КЛИНИЧЕСКГО МАТЕРИАЛА ПРИ
ОБСЛЕДОВАНИИ НА ТУБЕРКУЛЕЗ
63
3.1. Структура контаминирующей микрофлоры при обследовании на
туберкулез
64

4
3.2. Контаминирующая микрофлора, выделенная при исследовании мочи и
мокроты
68
3.3. Кислотоустойчивые представители порядка Actinomycetales в структуре
контаминирующей микрофлоры
73
3.4. Оценка влияния питательной среды для первичного посева на выделение
представителей порядка Actinomycetales из клинического материала
77
3.5. Резюме 82
ГЛАВА 4. СТРУКТУРА КИСЛОТОУСТОЙЧИВЫХ
ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА ACTINOMYCETALES,
ПРЕДВАРИТЕЛЬНО ИДЕНТИФИЦИРОВАННЫХ КАК
НЕТУБЕРКУЛЕЗНЫЕ МИКОБАКТЕРИИ
86
4.1. Анализ результатов первичной идентификации микрофлоры с
использованием метода ДНК-гибридизации
87
4.2. Структура микроорганизмов, предварительно идентифицированных как
нетуберкулезные микобактерии по результатам MALDI-ToF масс-
спектрометрии
89
4.3. Сравнительный анализ результатов идентификации выделенных
микроорганизмов используемыми методами
94
4.4. Частота выделения и структура кислотоустойчивых актиномицет в
клиническом материале в зависимости от пола и возраста пациентов
98
4.5. Резюме 106
ГЛАВА 5. ОЦЕНКА ВОЗМОЖНОСТИ КУЛЬТИВИРОВАНИЯ
ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА ACTINOMYCETALES НА
АГАРИЗОВАННЫХ СРЕДАХ
111
5.1. Оценка культуральных свойств анализируемых микроорганизмов при
посеве на плотные и жидкие питательные среды для культивирования
микобактерий
111
5.2. Особенности культуральных свойств представителей порядка
Actinomycetales на агаризованных средах
115

5
5.2.1. Оценка возможности культивирования нетуберкулезных
микобактерий на универсальной хромогенной среде и кровяном агаре
115
5.2.2. Влияние температуры культивирования на скорость роста
нетуберкулезных микобактерий на агаризованных средах
121
5.2.3. Оценка возможности использования новых вариантов лабораторной
посуды для культивирования нетуберкулезных микобактерий на
универсальной хромогенной среде
122
5.3. Оценка возможности предварительной идентификации
кислотоустойчивых представителей порядка Actinomycetales при
использовании универсальной хромогенной среды
123
5.4. Оценка использования универсальной хромогенной среды с
селективной добавкой для культивирования кислотоустойчивых
представителей порядка Actinomycetales
131
5.5. Резюме 133
ГЛАВА 6. ОСОБЕННОСТИ ИДЕНТИФИКАЦИИ
КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА
ACTINOMYCETALES МЕТОДОМ MALDI-TOF МАСС-
СПЕКТРОМЕТРИИ
137
6.1. Сравнение методов идентификации представителей семейства
Mycobacteriaceae с использованием методов ДНК-гибридизации, MALDI-
ToF масс-спектрометрии, секвенирования по Сэнгеру
137
6.2. Особенности проведения идентификации кислотоустойчивых
представителей порядка Actinomycetales в зависимости от среды для
культивирования с использованием метода MALDI-ToF масс-
спектрометрии
143
6.2.1. Оценка влияния среды культивирования на результат идентификации
нетуберкулезных микобактерий методом MALDI-ToF масс-спектрометрии
143

6
6.2.2. Оценка влияния среды культивирования на результат идентификации
других микроорганизмов из группы кислотоустойчивых представителей
порядка Actinomycetales методом MALDI-ToF масс-спектрометрии
146
6.3. Сравнение белковых профилей, полученных методом MALDI-ToF масс-
спектрометрии у представителей семейства Mycobacteriaceae, выращенных
на различных питательных средах
147
6.4. Резюме 153
ГЛАВА 7. ФУНДАМЕНТАЛЬНЫЕ И ПРИКЛАДНЫЕ АСПЕКТЫ
ОБОСНОВАНИЯ НОВЫХ ПОДХОДОВ К КУЛЬТИВИРОВАНИЮ И
ИДЕНТИФИКАЦИИ КИСЛОТОУСТОЙЧИВЫХ
ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА ACTINOMYCETALES
156
7.1. Выбор питательной среды для первичного посева как определяющий
фактор выделения и идентификации медленно и быстро растущих
кислотоустойчивых представителей порядка Actinomycetales в клинической
практике
156
7.2. Возможности продленной инкубации для выделения и идентификации
кислотоустойчивых представителей порядка Actinomycetales в рутинной
микробиологической практике
161
7.2.1. Влияние продленной инкубации на агаризованных средах на
возможность выделения кислотоустойчивых представителей порядка
Actinomycetales из клинического материала
161
7.2.2. Особенности идентификации выделенных из клинического материала
культур с использованием метода MALDI-ToF масс-спектрометрии
164
7.3. Протеомный анализ белковых спектров кислотоустойчивых
представителей порядка Actinomycetales как элемент субтипирования
штаммов, выделенных от одного пациента
166
7.4. Резюме 176
ЗАКЛЮЧЕНИЕ 180
ВЫВОДЫ 200

7
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ 202
СПИСОК СОКРАЩЕНИЙ 203
СПИСОК ЛИТЕРАТУРЫ 204

8
ВВЕДЕНИЕ
Актуальность темы исследования и степень ее разработанности
Микробиологическая диагностика инфекционных заболеваний,
обусловленных условно-патогенными микроорганизмами, в последние
десятилетия получила значительное развитие благодаря внедрению новых методов
идентификации и культивирования [32, 65, 91]. Описание новых видов
микроорганизмов, имеющих клиническое значение, и выявление групп риска по
развитию оппортунистических инфекций затронуло практически все направления
классической микробиологии [96]. Необходимо отдельно отметить роль
времяпролетной масс-спектрометрии с матрично-активированной лазерной
десорбцией/ионизацией (MALDI-ToF), благодаря широкому внедрению которой
стала возможной идентификация значительного разнообразия условно-патогенной
микрофлоры, выделенной из клинического материала [183, 270, 288, 291].
Тем не менее, несмотря на значительные успехи в проводимых
исследованиях за последние годы, в клинической микробиологии нерешенными
остаются научные и прикладные вопросы, связанные с выделением,
идентификацией, оценкой клинического значения и эпидемиологическими
аспектами в отношении значительного ряда микроорганизмов. К одной из
проблемных групп относятся кислотоустойчивые представители порядка
Actinomycetales (КУППА), в состав которого входят как облигатно, так и условно-
патогенные микроорганизмы, характеризующиеся разнообразными
биологическими свойствами.
С середины двадцатого века особенно активно идет изучение представителей
семейства Mycobacteriaceae, что обусловлено широким распространением
возбудителя туберкулеза и проблемами его резистентности к антимикробным
химиопрепаратам [16, 196]. Значительные социальные и экономические проблемы
в обществе, связанные с заболеваемостью туберкулезом среди населения, сместили

9
вектор изучения с нетуберкулезных микобактерий (НТМ) и других
микроорганизмов из группы КУППА. Количество публикаций, выполненных на
территории Российской Федерации посвященных M.tuberculosis complex (MTBc)
значительно превышает число работ, в которых приведены данные об
особенностях выделения и идентификации НТМ и других представителей порядка
Actinomycetales. Публикации, в которых приводятся данные о гордониях,
нокардиях и других кислотоустойчивых бактериях в отечественной научной
литературе по медицине практически отсутствуют, или ограничиваются другими
областями науки (ветеринария, сельское хозяйство, биотехнология) [1, 37, 41, 42,
43, 44, 67].
Наиболее активно изучение КУППА стало развиваться с девяностых годов
прошлого века, в связи с эпидемией инфекции, вызванной вирусом
иммунодефицита человека (ВИЧ) и обусловленным ей синдромом приобретенного
иммунодефицита (СПИД) [74, 197]. Рост заболеваемости микобактериозами среди
ВИЧ-инфицированных пациентов стал причиной значительного увеличения
количества работ, посвященных этой проблеме во многих европейских странах
[190, 227, 275, 282].
Параллельно с парадигмой изучения оппортунистических инфекций, как
элемента развития ВИЧ-ассоциированного СПИДа, среди клиницистов различных
специальностей все чаще встает вопрос о росте этиологического значения условно-
патогенных микроорганизмов при иммунодефицитах различной этиологии [10,
222]. Важным доказательством возрастающей роли данной группы
микроорганизмов является четкое обозначение факторов риска по развитию
определенных оппортунистических инфекций у пациентов с первичными
иммунодефицитами (ПИД) и другими врожденными аномалиями;
онкологическими заболеваниями; получающих иммуносупрессивную терапию по
поводу основного заболевания; пациентов после трансплантации органов и тканей;
имеющих хроническую соматическую патологию различного генеза [208].
Увеличение числа пациентов из групп риска значительно расширяет перечень
микроорганизмов, имеющих клиническое значение в той или иной ситуации.

10
На наш взгляд, именно по этой причине в научной литературе все чаще стали
приводится клинические примеры, в которых этиологическим агентом выступают
различные представители порядка Actinomycetales [47]. Условно-патогенные
бактерии из семейств Mycobacteriaceae, Nocardiaceae, Gordoniaceae,
Tsukamurellaceae, Promicromonosporaceae, Brevibacteriaceae, Streptomycetaceae
являются широко распространенными в окружающей среде микроорганизмами,
которые могут входить в состав микробиоты различных органов и систем человека
и животных, имеют ряд особенностей метаболизма, обеспечивающих их
устойчивость к неблагоприятным факторам, которые при определенных
обстоятельствах могут иметь важное патогенетическое значение в случае развития
заболевания с их участием [63, 128, 299, 307, 318].
В дополнение к росту числа пациентов из групп риска немаловажную роль в
диагностике осложнений, связанных с кислотоустойчивыми аэробными
актиномицетами, имеют новые методы идентификации микроорганизмов.
Внедрение в практическую микробиологию MALDI-ToF масс-спектрометрии
создает необходимые условия для идентификации редких и нетривиальных
микроорганизмов, в том числе и представителей порядка Actinomycetales, имеющих
потенциальное клиническое значение [13, 106].
Однако, на наш взгляд, данный факт является причиной определенного
микробиологического парадокса – современные методы идентификации
значительно опередили методы культивирования некоторых микроорганизмов, что
привело к отсутствию единых схем их выделения из клинического материала,
ограниченности данных об их структуре в зависимости от проявлений
патологических процессов, возраста и пола пациентов, отсутствия единого подхода
к практическим аспектам применения новых методов идентификации с учетом
условий культивирования «проблемных» микроорганизмов.
Описанный выше парадокс ставит задачи не только перед микробиологами,
но и перед клиницистами. Именно по причине отсутствия практических
рекомендаций по выделению и идентификации кислотоустойчивых
представителей порядка Actinomycetales в Российской Федерации практически нет

11
статистических данных о распространенности микобактериозов, до конца не ясна
ситуация с критериями верификации клинического диагноза, методами
диагностики и терапии инфекций, вызванных НТМ, нокардиями и другими
актиномицетами. На сегодняшний день ситуация с терапией данной группы
пациентов находится в сфере практической деятельности врачей-пульмонологов и
врачей-фтизиатров с тенденцией выхода из фтизиатрической службы, а
микробиологическая диагностика остается прерогативой лабораторий
противотуберкулезных учреждений в ограниченном микобактериями поле.
Классическая видовая идентификация КУППА в микробиологической
практике долгое время имела значительные ограничения, связанные с
метаболическими, генетическими, культуральными и широким рядом других
особенностей. В современной лабораторной диагностике микобактериозов
используется метод ДНК-гибридизации, предложенный Hain Lifescience GmbH
(Германия) и рекомендованный Всемирной Организацией Здравоохранения в 2008
году как основной метод идентификации НТМ [106]. Важным недостатком этого
метода исследования является ограничение видового разнообразия НТМ и
отсутствие возможности идентификации других КУППА, которые могут быть
причиной патологических состояний, схожих по клинической картине с
туберкулезом и микобактериозами [73].
Таким образом, необходим анализ существующих и разработка новых
подходов к культивированию и идентификации КУППА, выделенных из
клинического материала, что позволит получить данные о распространенности,
структуре, эпидемиологических и клинических особенностях инфекционных
процессов, обусловленных условно-патогенными кислотоустойчивыми аэробными
актиномицетами.

12
Цель исследования
Разработка новых методов культивирования и идентификации
кислотоустойчивых представителей порядка Actinomycetales, выделенных из
клинического материала для улучшения диагностики вызванных ими заболеваний.
Задачи исследования
1. Изучить структуру контаминирующей микрофлоры при исследовании
клинического материала на туберкулез и оценить в ней долю
кислотоустойчивых представителей порядка Actinomycetales.
2. Определить видовой состав кислотоустойчивых представителей порядка
Actinomycetales, первоначально идентифицированных как нетуберкулезные
микобактерии.
3. Провести оценку возможности культивирования кислотоустойчивых
представителей порядка Actinomycetales на агаризованных средах и описать их
свойства в зависимости от условий культивирования.
4. Разработать и внедрить в практическую деятельность микробиологической
лаборатории методы предварительной идентификации кислотоустойчивых
представителей порядка Actinomycetales при культивировании на
универсальных хромогенных средах.
5. Определить особенности проведения идентификации кислотоустойчивых
представителей порядка Actinomycetales в зависимости от среды первичного
посева с использованием метода MALDI-ToF масс-спектрометрии
6. Оценить возможность видовой идентификации методом MALDI-ToF масс-
спектрометрии нетуберкулезных микобактерий, выросших на универсальной
хромогенной среде.
7. Провести протеомный анализ белковых спектров кислотоустойчивых
представителей порядка Actinomycetales для оценки возможного
субтипирования штаммов, выделенных от одного пациента.

13
Методология и методы диссертационного исследования
Исследования по оценке культуральных свойств микроорганизмов,
идентификации с использованием MALDI-ToF масс-спектрометрии, протеомный
анализ полученных белковых спектров проводились на базе кафедры общей и
клинической микробиологии, иммунологии и аллергологии ФГБОУ ВО
«Самарский государственный медицинский университет» Министерства
здравоохранения Российской Федерации (зав. каф. – з.д.н. РФ, д.м.н., проф. А.В.
Жестков). Методологический принцип исследовательской работы заключался в
комплексном подходе по оценке распространенности, структуры, особенностей
культивирования и идентификации, разработке методов предварительной
идентификации и анализе белковых профилей кислотоустойчивых аэробных
актиномицет, выделенных из клинического материала.
Первичный посев клинического материала, отбор культур для включения в
исследование и их идентификация с использованием метода ДНК-гибридизации
проводились на базе бактериологической лаборатории ГБУЗ «Самарский
областной клинический противотуберкулезный диспансер им. Н.В. Постникова»
(зав. – врач-бактериолог высшей категории Т.П. Персиянцева). Проведение
процедур обнаружения ДНК МТВс, их видовое определение методом ПЦР, а также
секвенирования фрагментов ДНК, выделенной из культур, – на базе отдела
молекулярной диагностики и эпидемиологии ФБУН Центральный НИИ
эпидемиологии Роспотребназдора (зав. – к.б.н. Г.А. Шипулин). Определение
состава питательных сред проводилось на базе КДЛ Клиник ФГБОУ ВО
«Самарский государственный медицинский университет» Министерства
здравоохранения Российской Федерации (зав. – д.м.н., доцент О.А. Гусякова).
Материал, включенный в работу собран в период с 2016 по 2019 годы и был
разделен на две части в соответствии с задачами исследования.
В первой части работы исследована микрофлора, полученная из 865 посевов
проб различного клинического материала от 779 пациентов при обследовании на
туберкулез с признаками роста контаминирующей микрофлоры и при условии

14
отсутствия в пробах представителей МТВс. Общее количество штаммов
микроорганизмов, выделенных в первой части исследования составило 1093.
Во второй части работы было проанализировано 316 посевов проб материала
с признаками роста НТМ от 246 пациентов. В исследовании было выделено и
идентифицировано 345 штаммов микроорганизмов.
Все культуры выделенных микроорганизмов были идентифицированы с
использованием метода MALDI-ToF масс-спектрометрии. Штаммы, имеющие
культуральные признаки, характерные для НТМ, были предварительно
идентифицированы с использованием метода ДНК гибридизации.
Всего в исследовании было выделено 402 штамма КУППА. Для всех
представителей кислотоустойчивых актиномицет проведена оценка возможности
культивирования на агаризованных средах (кровяной агар и универсальная
хромогенная среда). Общее количество посевов первичного материала составило
3543. У всех штаммов НТМ был проведен анализ белковых спектров, полученных
при MALDI-ToF масс-спектрометрии с использованием программ flexAnalysis 3.0
и MALDI Biotyper 3.0 Offline Classification (Bruker Daltonik GmbH, Германия).
Дополнительно в исследование было включено 74 образца посевов
клинического материала с признаками роста НТМ, для которых было проведено
сравнение результатов идентификации методами ДНК-гибридизации, MALDI-ToF
масс-спектрометрии и прямого секвенирования по Сэнгеру.
Проведена статистическая обработка полученных результатов с учетом
распространенности отдельных групп микроорганизмов в зависимости от вида
клинического материала, сред для первичного посева, возраста и пола пациентов.
Группировка первичных данных и вычисления проводилась с использованием
пакета программ Microsoft Excel® 2013.
Описание количественных признаков, отличающихся от нормального
распределения представлено в виде медианы. Качественные признаки
представлены в виде долей процентов, абсолютного числа. Оценка структуры
микрофлоры включала в себя расчет частоты встречаемости таксона в отдельных
видах клинического материала и относительного среднего для каждого

15
идентифицированного рода микроорганизмов (доля микроорганизма в
исследуемой совокупности представителей родов, в зависимости от клинического
материала с учетом других факторов). Для оценки силы связи между
анализируемыми факторами, использовали критерий Х2 и степень закономерности
события (р), используемые при анализе сопряженности таблиц. Связь между
признаками статистически расценивали как значимую при уровне значимости р
<0,01 или р <0,05 в зависимости от статистики теста.
Для оценки различий между двумя независимыми выборками по уровню
определенного признака, измеренного количественно применяли критерий Манна-
Уитни. При проверке равенства медиан нескольких выборок использовали
критерий Краскела-Уоллиса.
В работе проводился биоинформационный анализ белковых спектров
выделенных культур с использованием программных пакетов flexAnalysis 3.0 и
MALDI Biotyper 3.0 Offline Classification на базе MALDI-ToF масс-спектрометра
Microflex LT (Bruker Daltonik GmbH, Германия).
Степень достоверности, апробация результатов, личное участие автора
Достоверность полученных результатов достигнута за счет применения в
качестве методологической и теоретической базы фундаментальных трудов в
области микробиологии; соответствия результатов современному уровню методик
проведения исследований.
Основные положения диссертации обсуждены на Национальном конгрессе
по болезням органов дыхания (Москва, 2016; Москва, 2018), Окружной научно-
практической конференции с международным участием «Современные технологии
комплексной медицинской помощи больным туберкулезом и ВИЧ-инфекцией:
реализация, развитие, резервы» (Екатеринбург, 2017), Всероссийской научно-
практической конференции с международным участием «Туберкулез и сочетанные
инфекции: вызовы и перспективы» (Москва, 2017), XIX Международном конгрессе

16
МАКМАХ по антимикробной терапии и клинической микробиологии (Москва,
2017), Российско-Китайском конгрессе по медицинской микробиологии,
эпидемиологии и клинической микологии (XX Кашкинские чтения) (Санкт-
Петербург, 2017), 38-м Ежегодном конгрессе Европейского общества
микобактериологии (Шибенки, 2017), III Российском конгрессе лабораторной
медицины (Москва, 2017), Окружной научно-практической образовательной
конференции «Туберкулез, ВИЧ и сочетанные инфекции: профилактика,
диагностика, лечение» (Екатеринбург, 2018), Международной научно-
практической конференции «Молекулярная диагностика 2018» (Минск, 2018),
Научно-практической конференции по хирургии «Инфекция в хирургии:
международные рекомендации и реальная клиническая практика» (Самара, 2018),
Самарском форуме лабораторной медицины: современные подходы к организации
лабораторной службы, повышению качества и доступности клинических
лабораторных исследований (Самара, 2018), XI Ежегодном Всероссийском
конгрессе по инфекционным болезням с международным участием
«Инфекционные болезни в современном мире: эволюция, текущие и будущие
угрозы» (Москва, 2019), 29-м Европейском конгрессе по клинической
микробиологии и инфекционным заболеваниям (Амстердам, 2019), Российско-
Китайском конгрессе по медицинской микробиологии, эпидемиологии,
клинической микологии и иммунологии (XXII Кашкинские чтения) (Санкт-
Петербург, 2019), 5-м Юбилейном Российском конгрессе лабораторной медицины
(Москва, 2019), Евразийском конгрессе по антимикробной терапии и клинической
микробиологии (Уфа, 2019), Всероссийской научно-практической конференции с
международным участием «Россия на пути ликвидации туберкулеза: реалии и
перспективы» (Москва, 2019), Российской научно-практической конференции с
международным участием «Казанская педиатрическая школа. Булатовские чтения»
«Актуальные вопросы пульмонологии» (Казань, 2020), Форуме специалистов
лабораторной медицины Республики Татарстан «Современные подходы к
организации лабораторной службы, профессиональные стандарты и образование,
перспективные технологии в медицинской практике (Казань, 2020), Научно-

17
практической конференции «Периоперационная антибиотикопрофилактика и
терапия: основные принципы назначения и проблемы» в рамках тридцать восьмой
образовательной недели (Самара, 2020).
Личный вклад автора состоит в непосредственном участии на всех этапах
диссертационного исследования. Основная идея, планирование научной работы,
включая формулировку научной гипотезы, определение методологии и общей
концепции диссертационного исследования, формулировка цели и задач,
разработка дизайна исследования проводились совместно с научным
консультантом з.д.н. РФ, д.м.н., профессором А.В. Жестковым.
Экспериментальные исследования, анализ полученных данных, их интерпретация,
представление результатов работы в научных публикациях и в виде докладов на
конференциях и конгрессах проводились совместно с сотрудниками кафедры
общей и клинической микробиологии, иммунологии и аллергологии ФГБОУ ВО
«Самарский государственный медицинский университет» Министерства
здравоохранения Российской Федерации (зав. каф. – з.д.н. РФ, д.м.н., профессор
А.В. Жестков), бактериологической лаборатории ГБУЗ «Самарский областной
клинический противотуберкулезный диспансер им. Н.В. Постникова» (зав. – врач-
бактериолог высшей категории Т.П. Персиянцева), отдела молекулярной
диагностики и эпидемиологии ФБУН Центральный НИИ эпидемиологии
Роспотребназдора (зав. – к.б.н. Г.А. Шипулин).
Анализ современной отечественной и зарубежной литературы по изучаемой
проблеме, статистическая обработка первичных данных, написание и оформление
рукописи диссертации проведены лично диссертантом.
Положения, выносимые на защиту:
1. В структуре контаминирующей микрофлоры при обследовании на туберкулез
выявлены разнообразные представители порядка Actinomycetales, в том числе
нетуберкулезные микобактерии с атипичными культуральными свойствами.

18
Использование жидкой питательной среды Миддлбрук 7Н9 с селективной
добавкой позволяет более эффективно выделять кислотоустойчивые
актиномицеты.
2. ДНК-гибридизация может быть использована в качестве предварительной
идентификации микробных культур с характерными для нетуберкулезных
микобактерий свойствами. Для окончательной идентификации кислотоустойчивых
представителей порядка Actinomycetales целесообразно использовать
дополнительные методы, в частности MALDI-ToF масс-спектрометрию.
3. Универсальные хромогенные среды могут быть использованы как для
первичного посева клинического материала с целью выделения
кислотоустойчивых аэробных актиномицет, так и для проведения предварительной
идентификации нетуберкулезных микобактерий.
4. Некоторые универсальные хромогенные среды позволяют проводить
идентификацию нетуберкулезных микобактерий с использованием метода
MALDI-ToF масс-спектрометрии также эффективно, как и при использовании
среды Левенштейна-Йенсена.
5. При выборе сред для первичного посева кислотоустойчивых представителей
порядка Actinomycetales необходимо учитывать не только лимитирующие
компоненты роста, но и возможности создания условий для продленной инкубации
с последующей идентификацией с использованием метода MALDI-ToF масс-
спектрометрии.
Научная новизна
Впервые проведена оценка распространенности представителей
кислотоустойчивых актиномицет в структуре контаминирующей микрофлоры,
выделенной из клинического материала при обследовании на туберкулез с
использованием метода MALDI-ToF масс-спектрометрии. Установлено, что при
обследовании на туберкулез возможно выделение вариантов нетуберкулезных

19
микобактерий с атипичными культуральными свойствами и других представителей
порядка Actinomycetales.
Впервые проведено исследование возможности использования
универсальной хромогенной среды как для первичного выделения
нетуберкулезных микобактерий, так и для их предварительной идентификации.
Впервые разработаны варианты лабораторной посуды, замедляющей процесс
дегидратации питательной среды для культивирования медленнорастущих
представителей кислотоустойчивых актиномицет.
Впервые разработаны новые инструменты, позволяющие получать
необходимое количество микробной биомассы для проведения идентификации
культур медленно и быстрорастущих микроорганизмов с использованием метода
MALDI-ToF масс-спектрометрии. Установлено, что универсальные хромогенные
среды могут быть использованы для улучшения результатов идентификации
нетуберкулезных микобактерий при использовании метода MALDI-ToF масс-
спектрометрии.
Впервые установлено значение использования универсальной хромогенной
среды для выявления гетерогенности популяции нетуберкулезных микобактерий,
выделенных от одного пациента.
Теоретическая и практическая значимость
Теоретическая значимость исследования определяется разработкой новых
подходов к культивированию и идентификации кислотоустойчивых бактерий из
порядка Actinomycetales, представленных как сапрофитной, так и условно-
патогенной микрофлорой, которые могут влиять на результаты проведения
микробиологического исследования клинического материала на туберкулез. При
оценке только культуральных свойств микрофлоры, выделенной при первичных
посевах есть вероятность получения как ложноотрицательных результатов в
отношении нетуберкулезных микобактерий с атипичными формами колоний, так и

20
ложноположительных результатов в отношении других кислотоустойчивых
микроорганизмов при идентификации их методом ДНК-гибридизации.
Определено значение использования жидкой питательной среды Миддлбрук 7Н9 с
селективной добавкой для выделения кислотоустойчивых представителей порядка
Actinomycetales, что требует пересмотра алгоритма работы с положительными
культурами в случае определения признаков роста микроорганизмов на ранних
сроках культивирования.
Практическое значение состоит в том, что некоторые универсальные
хромогенные среды являются оптимальными для первичного выделения и
предварительной идентификации значительного количества кислотоустойчивых
актиномицет и могут быть использованы в практической работе
микробиологических лабораторий для улучшения результатов выделения
нетуберкулезных микобактерий и других представителей порядка Actinomycetales
(Патент РФ № 2711954, Патент РФ № 2711957). Пролонгация сроков инкубации
первичных посевов в рутинной микробиологической практике позволяет выделять
представителей порядка Actinomycetales из клинического материала (Патент РФ №
2659155, Патент РФ № 2668406). Для предотвращения дегидратации сред при
пролонгированном инкубировании посевов необходимо использовать
специальную лабораторную посуду, которую можно применять для
культивирования медленно и быстрорастущих аэробных и факультативно-
анаэробных микроорганизмов (Патент РФ № 173302, Патент РФ № 175134, Патент
РФ № 175863).
Для улучшения результатов идентификации микроорганизмов из группы
кислотоустойчивых актиномицет, имеющих особенности культуральных свойств,
необходимо использовать дополнительные инструменты, которые позволяют
получать количество биологической массы достаточное для определения их
видовой принадлежности методом MALDI-ToF масс-спектрометрии (Патент РФ №
187404, Патент РФ № 187421). При выборе сред для работы с микобактериями
необходимо учитывать количество железа и его форму, как лимитирующего
компонента питательной среды, определяющего возможность выделения не только

21
микобактерий туберкулезного комплекса, но и нетуберкулезных микобактерий
(Патент РФ № 2613366).
Внедрение результатов исследования в практику
Результаты исследования внедрены в учебный процесс и используются в
научно-исследовательской деятельности кафедр общей и клинической
микробиологии, иммунологии и аллергологии; инфекционных болезней с
эпидемиологией; фундаментальной и клинической биохимии с лабораторной
диагностикой ФГБОУ ВО «Самарский государственный медицинский
университет» Министерства здравоохранения Российской Федерации.
Результаты диссертационной работы используются в практической
деятельности микробиологического отдела клинико-диагностической лаборатории
КДЛ Клиник ФГБОУ ВО «Самарский государственный медицинский
университет» Министерства здравоохранения Российской Федерации,
бактериологической лаборатории ГБУЗ «Самарский областной клинический
противотуберкулезный диспансер им. Н.В. Постникова», бактериологической
лаборатории ОГБУЗ Белгородской области «Детская областная клиническая
больница», лаборатории бактериологических исследований БУЗ ВО «Воронежский
областной клинический центр профилактики и борьбы со СПИД», лаборатории
клинической бактериологии ГАУЗ «Республиканская клиническая больница
Министерства Здравоохранения Республики Татарстан», городского детского
центра муковисцидоза, расположенного на базе инфекционно-боксированного
отделения №3 СПб ГБУЗ «Детская городская больница Святой Ольги»,
бактериологической лаборатории ГБУЗ «Городская клиническая больница №15
имени О.М. Филатова Департамента здравоохранения города Москвы».

22
ГЛАВА 1. КИСЛОТОУСТОЙЧИВЫЕ ПРЕДСТАВИТЕЛИ ПОРЯДКА
ACTINOMYCETALES: КУЛЬТИВИРОВАНИЕ, ИДЕНТИФИКАЦИЯ И
КЛИНИЧЕСКОЕ ЗНАЧЕНИЕ
(обзор литературы)
1.1. Общая характеристика и современная классификация представителей
порядка Actinomycetales
В соответствии с классификацией микроорганизмов, приведенной в
Определителе бактерий Берджи (девятое издание, редакция 1997 года)
микобактерии и актиномицеты относятся к грамположительным эубактериям,
имеющим клеточную стенку. Микобактерии отнесены к 21 группе и представляют
собой аэробные, неподвижные, споронеобразующие палочковидные бактерии
способные в некоторых случаях к ветвлению, характеризующиеся медленным
ростом от 2 до 40 суток и кислотоустойчивостью. Актиномицеты отнесены к 22-
29-й группам из-за значительного разнообразия представителей данного порядка.
Однако общими для них являются следующие признаки: грамположительные
ветвящиеся нити, способны образовывать гифы в виде мицелия, некоторые
представители имеют жгутики [138]. Приведенные выше свойства часто служат
основополагающими при проведении микроскопических исследований
клинического материала, особенно при подозрении на туберкулез или
микобактериозы, но по литературным данным даже на уровне рода Mycobacterium
возможны различные морфологические варианты бактериальных клеток.
Например, некоторые виды могут быть представлены коккобациллами
(M.lentiflavum, M.interjectum, M.intermedium) [217]. Данный факт может затруднять
интерпретацию результатов микроскопического исследования и приводить к
получению ложноотрицательных результатов, т.к. обнаружение
кислотоустойчивых кокковидных микроорганизмов не является классическим
признаком, характерным для НТМ и МТВс. Основная масса представителей рода

23
Mycobacterium не способны к спорообразованию, однако такая особенность была
выявлена у M.marinum, которые в старых культурах могут образовывать
эндоспоры, при пересеве на свежие среды, дающие вегетативные клетки [303].
В современной литературе таксономическое положение актиномицет и
микобактерий несколько изменилось. Обе условные группы на сегодняшний день
относятся к порядку Actinomycetales, который включает 14 подпорядков и более 45
семейств [236, 269]. Наибольший интерес в медицинской микробиологии в
последнее время проявлен к семействам Actinomycetaceae, Corynebacteriaceae,
Mycobacteriaceae, Nocardiaceae, Gordoniaceae. Представители последних трех
семейств характеризуются кислотоустойчивостью различной степени
выраженности и являются аэробными грамположительными палочками. Роль
представителей других семейств порядка Actinomycetales, обладающих
кислотоустойчивостью в большей или меньшей степени, в частности,
Tsukamurellaceae, все чаще обсуждается в литературных источниках в качестве
потенциально патогенных микроорганизмов, имеющих клиническое значение у
пациентов с иммуносупрессией различной этиологии, а также в качестве
представителей нормобиоты человека [144]. В тоже время представители семейств
Promicromonosporaceae, Brevibacteriaceae и Streptomycetaceae чаще описываются в
качестве представителей сапрофитной микрофлоры [129, 132, 283]. Следует
отметить, что в соответствии с современной классификацией микроорганизмов
семейства Mycobacteriaceae, Nocardiaceae, Gordoniaceae, Tsukamurellaceae
относятся к подпорядку Corynebacterineae; семейства Brevibacteriaceae и
Promicromonosporaceae – к подпорядку Micrococcineae; а семейство
Streptomycetaceae является единственным в подпорядке Streptomycineae [155, 161,
320].
Все КУППА характеризуются признаком «кислотоустойчивость», который,
по литературным данным, обусловлен особенностью строения клеточной стенки,
обеспечивающей их выживание при контакте с концентрациями кислот и щелочей,
оказывающими бактерицидное действие на другие микроорганизмы [57, 109].
Такая особенность связана с высоким содержанием миколовых кислот в клеточной

24
стенке, кроме этого для многих представителей порядка Actinomycetales характерно
преобладание нуклеотидов ГЦ (гуанидин/цитозин) в ДНК. Например, содержание
ГЦ в ДНК микобактерий составляет, по данным зарубежных авторов, 61-71% [155,
265], в отечественной литературе приводятся данные о диапазоне от 62 до 70% [96].
Для других представителей КУППА данный признак находится в следующих
пределах: 68,4-70,8% – для представителей рода Nocardia; 63-69% – для бактерий
рода Gordonia [157, 199]. Данный факт также может использоваться для
предварительной идентификации КУППА.
С точки зрения микробиологической диагностики кислотоустойчивость
является важным критерием, определяющим ход исследования клинического
материала, а также идентификации уже выделенных культур, что отражается в том
числе, и в нормативных документах по диагностике микобактериальных инфекций
[89]. Однако, известно, что не все представители рода Mycobacterium обладают
постоянной кислотоустойчивостью. Несмотря на сложно организованную
клеточную стенку, в состав которой кроме миколовых кислот с количеством
атомов углерода от 70 до 90 входят свободные липиды, воски жирных кислот,
фосфолипиды, микосерозные и фтиеновые кислоты; кислотоустойчивость клетки
может быть утеряна на определенных стадиях роста, особенно у быстрорастущих
НТМ [233]. Таким образом, не выявленная при микроскопии кислотоустойчивость
может приводить к получению ложноотрицательных результатов исследования
чистых культур и клинического материала при использовании окраски по методу
Циль-Нильсена, а также оказывать значительное влияние на проведение
деконтаминации нестерильного клинического материала при подготовке его к
посеву. В связи с этим необходимо внедрять щадящие методы деконтаминации при
пробоподготовке материала, особенно при работе с материалом, в котором
предполагается наличие быстрорастущих НТМ, либо разрабатывать и внедрять для
первичного посева селективные питательные среды, позволяющие осуществлять
посев клинического материала без предварительной деконтаминации [127, 169].
С другой стороны, кроме микобактерий кислотоустойчивость, как было
приведено выше, свойство, характерное для многих представителей порядка

25
Actinomycetales, часть из которых имеет доказанное клиническое значение [156,
166, 276, 317]. Определение таких микроорганизмов в клиническом материале или
чистой культуре требует проведения исследований на МТВс и НТМ, однако в
случае отрицательного результата, дальнейшие алгоритмы работы с таким
материалом или выделенным микроорганизмом не разработаны. Также, как и не
определена роль других микроорганизмов, в частности спорообразующих палочек,
которые могут обладать умеренной или стойкой кислотоустойчивостью, что может
приводить к получению ложноположительных результатов при использовании
стандартных схем исследования материала [107].
В дополнение к нерешенным вопросам дифференциально-диагностической
оценки морфологических и тинкториальных свойств КУППА в научной литературе
последних нескольких лет появляются новые предложения по пересмотру
классификаций отдельных групп микроорганизмов из порядка Actinomycetales
[140]. Особенно активно данный процесс идет в отношении рода Mycobacterium.
Все шире обсуждаются новые подходы к классификации микобактерий,
основанные на результатах генетических исследований. С одной стороны, это
позволяет дополнить классификацию НТМ и выделить несколько крупных и
небольших комплексов, объединяющих генетически наиболее близкие виды. С
другой стороны, такой подход требует серьезного пересмотра оценки результатов
микробиологических исследований на НТМ в частности и КУППА в целом, т.к.
внутри одного комплекса могут быть виды и подвиды с различными свойствами:
чувствительностью к антимикробным препаратам, клиническим значением,
риском развития диссеминированных форм, эпидемиологическими особенностями
[126, 195, 244, 246, 247, 251, 285]. Особенно интересным является вариант новой
классификации микобактерий по кладам, которая предложена в 2018 году группой
микробиологов из Канады. Разделение микобактерий на 5 клад (Abscessus-
Chelonae, Fortuitum-Vaccae, Terrae, Triviale, Tuberculosis-Simiae), основанное на
филогенетических исследованиях показало значительное сродство облигатно-
патогенных, условно-патогенных и непатогенных микобактерий [204]. На
основании молекулярно-генетических исследований были выделены новые роды в

26
семействе Mycobacteriaceae: Mycolicibacter spp., Mycolicibacillus spp.,
Mycobacteroides spp., Mycolicibacterium spp. В связи с активным процессом
изменений в таксономическом положении отдельных групп порядка
Actinomycetales, в работе было принято решение: использовать устоявшуюся в
медицинской микробиологии терминологию без разделения семейства
Mycobacteriaceae на дополнительные роды.
Таким образом, современная классификация кислотоустойчивых
микроорганизмов не ограничена только морфологическими и тинкториальными
свойствами и позволяет рассматривать однородные ранее группы
микроорганизмов в качестве сложноорганизованных консорциумов. Однако
основные диагностические алгоритмы, применяемые в рутинной
микробиологической практике, сохранили архаичные принципы, что оправдано с
экономической точки зрения, но в современных условиях требует разработки
новых алгоритмов диагностического поиска, направленных не на выявление
«абстрактных» групп микроорганизмов, объединенных одним свойством, а на
решение конкретной задачи – верификации этиологической причины развившейся
у пациента патологии.
1.2. Особенности культивирования отдельных представителей порядка
Actinomycetales
Современные возможности культивирования позволяют выделять
значительное количество микроорганизмов с различными потребностями по
набору питательных веществ [2, 21, 33, 88, 321]. Разработка селективных добавок,
специальных и универсальных хромогенных сред значительно расширяет
возможность выделения, а в некоторых случаях идентификации прихотливых
микроорганизмов, а также микроорганизмов, имеющих эпидемиологическое
значение [3, 28, 61, 71, 86]. В последние годы все чаще появляются публикации о
использовании питательных сред для выделения микроорганизмов с

27
определенными механизмами резистентности к антимикробным препаратам [6, 31,
35, 36, 181].
Динамичные изменения по расширению перечня питательных сред
коснулись не только стандартных микробиологических исследований, но и
специфичных направлений клинической микробиологии. Так внедрение жидкой
питательной среды Миддлбрук 7Н9 в работу лабораторий противотуберкулезной
службы позволило сократить время культивирования микобактерий, а в
дополнение к стандартным яичным средам повысило чувствительность
микробиологического метода исследования на туберкулез [103, 106].
Перечисленные преимущества современных питательных сред значительно
расширяют возможности микробиологического метода исследования, но в тоже
время и ставят новые задачи перед практикующими специалистами-
бактериологами. Парадигмы лабораторной диагностики туберкулеза являются
основополагающими с точки зрения выделения определенной облигатно-
патогенной группы микроорганизмов – МТВс, которые были сформированы в
двадцатом веке. Выделение остальных представителей рода Mycobacterium не
является доминирующим с точки зрения диагностического поиска, не говоря о
других КУППА. Формирование данной парадигмы обосновано с точки зрения
приоритетной задачи – выделение широко распространенного на тот момент
микроорганизма. Разработанные и внедренные в повсеместную практику
питательные среды на яичной основе характеризовались значительным по составу
перечнем необходимых для культивирования патогенных микобактерий веществ:
набора определенных минеральных солей и содержащихся в желтке куриного яйца
компонентов [95, 148, 223]. В Российской Федерации к таким средам,
регламентированным на законодательном уровне относятся среды Левенштейна-
Йенсена, Финн-II, Школьниковой [89]. В то же время необходимо учитывать, что
минеральные вещества, аминокислоты и жиры куриного яйца могут обеспечить
необходимые условия для роста и других КУППА, в связи с чем возникают
вопросы о их культуральных особенностях: сроках и условиях появления роста,

28
возможности выделения чистых культур из клинического материала при
использовании таких сред.
Подход, основанный на создании оптимальных условий для выделения одной
группы микроорганизмов, безусловно может использоваться для выделения
сопутствующих групп, но при этом необходимы исследования, подтверждающие
такие предположения. Одним из частных примеров данного факта послужил
алгоритм выделения и предварительной идентификации НТМ на питательных
средах для МТВс, предложенный еще в 1971 году E.Ranyon и J.M.Grange [95].
Согласно этому алгоритму, все НТМ по культуральным свойствам условно делятся
на четыре группы: фотохромогенные образуют пигмент на свету, колонии, как
правило, окрашиваются в различные оттенки желтого цвета; скотохромогенные
образуют пигмент в темноте; нефотохромогенные не образуют пигментов, либо
имеют слабовыраженную окраску колоний. Перечисленные группы НТМ являются
преимущественно медленнорастущими. В отечественной литературе приводятся
данные о преобладающем клиническом значении представителей
медленнорастущих НТМ [15, 58, 110]. В четвертую группу входят быстрорастущие
НТМ, не образующие пигмент на свету. Отличительной особенностью данной
группы являются скорость роста на яичных средах – появление видимого роста в
течение 7-10 суток и отсутствие пигментообразования [32, 230].
Предложенная во второй половине двадцатого века классификация
микобактерий практически не претерпела значительных изменений, несмотря на
появление большого массива данных о новых видах НТМ и их свойствах. Тем не
менее, в зарубежной литературе все чаще приводятся данные о НТМ с указанием,
в первую очередь скорости роста на обычных питательных средах, которые не
используются для выделения МТВс [184, 187]. Временной интервал, который
рассматривают как критерий, разделяющий виды на медленно и быстрорастущие,
при этом сокращен до 7-и суток при посеве культуры из свежеприготовленного
инокулюма на среду Левенштейна-Йенсена и при культивировании в наиболее
оптимальных условиях [237]. Следует отметить, что быстрорастущие НТМ могут
давать видимый рост значительно позже 7-и суток при первичном посеве

29
клинического материала, что может быть связано с рядом факторов: проводимой
антимикробной химиотерапией, методами деконтаминации материала, условиями
его транспортировки до лаборатории и рядом других причин; а медленнорастущие
НТМ могут давать видимый рост значительно быстрее чем за 7 суток [34].
Широкое внедрение жидких питательных сред для посева клинического
материала значительно влияет на скорость роста как медленно-, так и
быстрорастущих НТМ, что безусловно должно учитываться при проведении
исследований. В литературе приводятся данные о значительном сокращении
сроков культивирования представителей МТВс при использовании
автоматических анализаторов для культивирования клинического материала при
исследовании на туберкулез [165, 203]. Однако не опубликованы результаты
комплексных исследований о влиянии условий культивирования при
использовании других сред на рост КУППА, возможности их предварительной
идентификации, которая в некоторых случаях является основополагающей для
выбора антимикробной химиотерапии.
Таким образом, подход к разделению НТМ на медленно и быстрорастущие по
культуральным свойствам оправдан только при строгом соблюдении критериев
оценки скорости появления видимого роста и не может быть использован
однозначно при оценке результатов первичных посевов клинического материала,
так как это может привести к некорректному выбору методов идентификации
выросших культур, а также к получению ложноотрицательных результатов в тех
случаях, когда рост НТМ происходит «атипично» быстро или медленно. Особенно
важно учитывать такую особенность в случае чрезмерно быстрого роста, когда
видимые колонии появляются в течение 24-48 часов, в связи с тем, что образец, в
посеве которого обнаруживаются такие микобактерии чаще рассматривается как
контаминированный посторонней флорой и не берется в работу. Что касается
других КУППА, то для них также характерны различные диапазоны появления
видимого роста на плотных средах, например, для представителей родов Nocardia
и Gordonia при культивировании на 5% кровяном агаре при температуре 37°С
изолированные колонии можно выявить в среднем через шесть суток [258, 304].

30
Параллельно с основным вектором, направленным на выделение МТВс из
клинического материала в качестве доминирующего, и выделение других КУППА
в качестве второстепенного при использовании питательных сред на яичной основе
в научной литературе имеется значительное количество описанных приемов
культивирования МТВс, НТМ и других КУППА на «обычных» питательных
средах: мясо-пептонном агар, кровяном агар, шоколадном агарах, агаре с сердечно-
мозговым экстрактом, агаре Сабуро, средах для выделения Burkholderia cepacia
complex (BCC), а также ряде других элективных сред [146, 147, 167, 170, 245].
Такие подходы описываются не только для первичного выделения
микроорганизмов из группы КУППА, но и для вторичных пассажей культур.
Данный факт является немаловажным для оценки потенциальной возможности
использования уже известных питательных сред для первичного посева, выделения
и, возможно, предварительной идентификации отдельных КУППА.
Дополнительной задачей, которая появляется при анализе состава
искусственных сред и потребностей условно-патогенных микроорганизмов группы
КУППА, является выявление лимитирующих компонентов, которые в избытке или
при их недостатке могут вызывать замедленный рост или гибель микробной
популяции при посеве на такую среду [97, 102, 120, 271, 322]. Частными примерами
влияния компонентов питательных сред, описанными в приведенных выше
ссылках, являются: возможный недостаток ионов железа или избыток нативного
белка, который может значительно подавлять рост сапрофитных микроорганизмов,
естественные условия обитания которых содержит низкие концентрация крупных
белковых соединений [97, 102]. Такие примеры могут являться вариантом
получения искаженных результатов при оценке первичных посевов, когда
предварительная классификация основывается только на скорости роста.
На наш взгляд интересным является возможность оценки использования
универсальных хромогенных питательных сред для первичного выделения и
возможной предварительной идентификации КУППА при работе с клиническим
материалом и культурами из коллекций. Такие среды богаты по своему составу, так
как требуют содержания различного набора питательных веществ, необходимых

31
разнообразным группам бактерий и грибов [114]. В тоже время они не содержат
избытка нативных белков и, как следствие, могут быть использованы для
выделения сапрофитных микроорганизмов, естественной природной средой
которых является окружающая среда. К данной группе можно отнести практически
всех представителей условно-патогенных КУППА. С другой стороны, состав
микроэлементов в таких средах более разнообразен, что делает их интересным
объектом для работы с анализируемой нами группой микроорганизмов.
На отечественном рынке существует несколько производителей различных
универсальных хромогенных сред, которые используются как для выделения
микроорганизмов из отдельного локуса (например, моча), так и дают возможность
выделения патогенов из различных локусов (классические универсальные
хромогенные среды) [72, 76, 101, 108, 179, 309].
Таким образом, на наш взгляд, анализ результатов культивирования
отдельных представителей КУППА не только на классических средах для
выделения МТВс (среды на яичной основе), но и на жидких средах для выделения
МТВс и универсальных хромогенных средах может позволить получить новые
данные, которые могут быть использованы для разработки алгоритмов выделения
и предварительной идентификации НТМ и других КУППА, выделенных из
клинического материала.
1.3. Методы идентификации представителей семейств Mycobacteriaceae,
Nocardiaceae, Gordoniaceae, Tsukamurellaceae, Brevibacteriaceae,
Promicromonosporaceae, Streptomycetaceae
Идентификация КУППА, также, как и других прокариот основана на
фундаментальных принципах строения и метаболизма, тинкториальных,
культуральных, филогенетических и протеомных свойствах микроорганизмов.
Генетическое сродство КУППА, с одной стороны, дает возможность их
идентификации до рода или семейства по общим алгоритмам, с другой стороны

32
значительно затрудняет видовую идентификацию в связи со схожестью целого
ряда признаков [273, 284].
На практике видовая идентификация КУППА имеет значительные
ограничения, что обусловлено, в первую очередь, низким метаболическим
потенциалом, который затрудняет проведение широко распространенного метода
идентификации микроорганизмов, основанном на определении биохимических
свойств [32, 177, 221, 224, 240, 272]. Использование других методов зачастую
ограничено научными целями, например, метод высокоэффективной жидкостной
хроматографии может использоваться для идентификации большого числа видов
КУППА, однако практически не используется из-за сложностей преаналитического
этапа [48, 210]. Вышеперечисленные особенности стали основополагающими с
точки зрения определенной отстраненности от КУППА практикующих врачей-
бактериологов с сохранением интереса к данной группе микроорганизмов у
бактериологов немедицинского направления.
В медицинской микробиологии до сих пор во многих лабораториях
используется принцип групповой идентификации НТМ, основанный на скорости
роста и культуральных свойствах, который, как было описано выше, имеет ряд
серьезных ограничений, особенно с учетом накопления значительного объема
данных о клиническом значении многих видов КУППА наряду с сапрофитами и
комменсалами, не имеющими этиологической роли в патологии у человека [214].
Результатом этого является низкий уровень диагностики микобактериозов в
Российской Федерации, ограниченный возможностью федеральных центров при
работе с редкими видами НТМ, и верификация этиологического участия других
КУППА в качестве причины патологических состояний представленная
единичными случаями в виде «редких» находок [11, 17, 45, 49, 94].
Основная масса видов из КУППА является представителями IV группы
патогенности в соответствии с Российским санитарно-эпидемиологическим
законодательством [100]. Это обусловлено низким патогенным потенциалом
большинства актиномицет, их широким распространением в окружающей среде и
низкой контагиозностью. Несмотря на это, чаще всего работа с КУППА проводится

33
в специализированных лабораториях противотуберкулезной службы, и, как
правило, она ограничена выделением и идентификацией МТВс, а также наиболее
широко распространенных НТМ. Другие виды из группы кислотоустойчивых
аэробных актиномицет остаются вне поля интересов и возможностей таких
лабораторий.
Первоначальная идентификация КУППА проводится с использованием двух
наиболее простых и доступных методов – микроскопического и
микробиологического. Кислотоустойчивость, характерная для большинства
КУППА, позволяет использовать в рутинной практике доступные методы
идентификации этого признака, определение которого у микроорганизмов в
материале значительно сужает диагностический поиск [75]. Некоторые авторы в
научной и учебной литературе приводят информацию о возможности
использования микроскопического метода исследования для предварительной
видовой идентификации НТМ, хотя на практике данный прием оказывается
неэффективным. Различная морфология клеток КУППА: палочки, ветвящиеся
палочки, палочки с утолщениями на концах, кокки, а также определенные
признаки, основанные на взаимном расположении клеток в мазках, описаны для
многих видов НТМ и некоторых других видов КУППА и не являются строго
специфичными [150, 234, 244]. На практике микроскопический метод
исследования, имея высокую специфичность (89-100%), оказался важным, но лишь
ориентировочным методом определения кислотоустойчивых микроорганизмов в
клиническом материале [32, 96]. Особенно с учетом накопленных данных о
кислотоустойчивости как непостоянном, вариабельном признаке [237, 319].
Возможности микробиологического метода частично описаны выше, однако
в рамках оценки культуральных свойств микроорганизмов из группы КУППА он
ограничен в первую очередь работой с НТМ. При работе с условно патогенными
КУППА необходимо учитывать их первоначальный ареал обитания, что влияет на
выбор не только сред для их выделения, но и условий культивирования. Кроме
обязательной для всех КУППА потребности в кислороде для многих из них
оптимальными по температуре являются условия, отличные от культивирования

34
классических возбудителей заболеваний человека. По данным литературы,
температурный оптимум для многих кислотоустойчивых актиномицет значительно
варьируется в пределах от 28 до 45°С [178, 219, 262, 296]. Данный факт ставит еще
один принципиальный вопрос о возможности культивирования НТМ и других
КУППА в условиях лабораторий противотуберкулезной службы, в которых, в
связи с приоритетностью выделения туберкулезных микобактерий, чаще создается
температурный оптимум в 37°С [5, 20, 111].
Значительное варьирование скорости роста НТМ и КУППА не может быть
однозначно ограничено алгоритмом отбраковки посевов по скорости появления
роста при обследовании на туберкулез. Раннее появление роста, по нормативным
документам, расценивается как контаминация посторонней микрофлорой при
условии отсутствия в материале ДНК МТВс, данный образец будет отбракован,
несмотря на возможное наличие в нем клинически значимых видов из группы
КУППА [89].
Если учесть факт возможного выделения НТМ и других КУППА на
агаризованных средах, реальна ли идентификация данной группы
микроорганизмов в рамках микробиологического метода? Без использования
современных методов идентификации ответ будет однозначным – нет. С учетом
отсутствия настороженности у врачей-бактериологов в отношении
кислотоустойчивых аэробных актиномицет, как потенциальных патогенов, данные
микроорганизмы либо не будут идентифицированы по причине их слабой
биохимической активности, либо будут расценены как посторонняя микрофлора,
не имеющая клинического значения. Особенно сложно решить данную проблему в
отношении медленнорастущих НТМ, которые на агаризованных средах дают
дисгонический рост, описываемый как стелющийся, слабый или выпотевающий
[32, 96]. Данный рост при стандартном микробиологическом исследовании может
быть расценен как незначимый, особенно в случае выделения на таком фоне
«классических» возбудителей инфекций.
Биохимическая идентификация как элемент микробиологического метода
исследования при работе с КУППА не показала своей состоятельности. Часто

35
замедленный рост, потребность в определенном наборе питательных веществ и
зависимость от избытка других компонентов ограничивают ее использование в
рутинной микробиологической практике. Отдельно следует отметить отсутствие
наборов коммерческого производства для идентификации КУППА по
биохимической активности. В лабораториях противотуберкулезной службы
биохимическая идентификация ограничена дифференциацией представителей
МТВс и некоторых НТМ [64]. Определение нитратредуктазной, арилсульфатазной,
амидазной активностей (способностей к утилизации ацетата, мочевины,
никотинамида, пиразинамида, аллантаина, сукцинамида), возможности выделять
каталазу, гидролизировать Твин-80, восстанавливать теллурит калия и способность
роста на средах с хлористым натрием позволяет идентифицировать девять
наиболее часто встречающихся видов НТМ [138]. Большинство из этих тестов
ограничены в использовании лабораториями нетуберкулезной службы, что
затрудняет идентификацию даже этих девяти видов в условиях непрофильных
лабораторий. В целом, по данным литературы, биохимическая идентификация
позволяет проводить определение порядка 45-и видов НТМ [24, 48, 69, 96].
Серьезным событием в лабораторной диагностике инфекционных
заболеваний, обусловленных КУППА, особенно микобактериозов стало внедрение
в практику молекулярно-генетических методов исследования. Определенная
степень доступности данных методов привела к тому, что в начале двадцать
первого века Всемирная организация здравоохранения рекомендовала
использовать некоторые из них в качестве приоритетных методов идентификации
МТВс, НТМ и определения механизмов резистентности к антимикробным
препаратам [14, 40, 54, 56, 58, 69].
Отечественные и зарубежные производители, научно-производственные
объединения активно разрабатывают различные варианты молекулярно-
генетической идентификации клинически значимых видов КУППА, однако и в
данном случае речь идет, в первую очередь, о представителях рода Mycobacterium,
в то время как другие микроорганизмы из семейств Nocardiaceae, Gordoniaceae,
Tsukamurellaceae, Brevibacteriaceae, Promicromonosporaceae, Streptomycetaceae по-

36
прежнему остаются вне поля научных и практических интересов. На сегодняшний
день разработаны методы идентификации микобактерий, основанные на
полимеразной цепной реакции (ПЦР) в различных модификациях [19, 70, 82, 85,
137, 158, 163].
Несмотря на значительные научно-практические наработки в данном
направлении наиболее широким распространением в Российской Федерации
пользуется метод идентификации НТМ, основанный на технологии обратной
гибридизации продукта ПЦР с видоспецифичными зондами, иммобилизованными
на ДНК-стрипах. Для идентификации НТМ таким способом разработаны два
набора ступенчатой идентификации коммерческого производства Hain Lifsciense
GmbH (Германия). Данный метод позволяет идентифицировать 28 наиболее часто
встречающихся видов НТМ. Его основными недостатками являются небольшое
число видов (с учетом современных данных о доказанном клиническом значении,
определенном более чем для 50 видов НТМ), отсутствие возможности
идентификации других представителей КУППА, несмотря на возможность
верификации культур с высоким содержанием ГЦ [106].
«Золотым» стандартом идентификации микроорганизмов являются
различные варианты секвенирования генов микроорганизмов. Методы
секвенирования позволяют провести идентификацию патогена как в чистой
культуре, так и в клиническом материале с высокой степенью специфичности и
чувствительности [131, 149, 215, 216]. Однако высокая стоимость одного
определения значительно ограничивает данный метод не только в рутинной
микробиологической практике, но и в научных целях [154, 160, 185, 228, 229, 242].
Использование метода высокоэффективной жидкостной хроматографии
(ВЭЖХ) также значительно ограничено на практике, несмотря на имеющиеся
работы в этом направлении. Идентификация микроорганизмов, основанная на
хроматографии миколовых кислот, является дорогостоящей и требует
значительных трудозатрат на подготовку образца для проведения исследования
[164, 209].

37
Значительным событием в микробиологии в последние десятилетия стало
внедрение в практику научных и практических лабораторий метода, основанного
на MALDI-ToF масс-спектрометрии [29, 30, 50, 51, 52, 90]. Данный метод
значительно расширил видовое разнообразие микроорганизмов, идентификация
которых стала доступной на достаточно высоком уровне для любых лабораторий,
оснащенных масс-спектрометрами. Именно широкое внедрение масс-
спектрометрии можно рассматривать как ключевой триггер, который значительно
повысил практический и научный интерес к КУППА. На сегодняшний день в базах
масс-спектрометров имеются белковые профили основных аэробных актиномицет,
имеющих доказанное или потенциальное клиническое значение. Так, например,
основная библиотека масс-спектрометра Microflex LT (Bruker Daltonik GmbH,
Германия) в версии 2019 года содержит 75 спектров микобактерий, 213 спектров
представителей семейства Nocardiaceae, 22 – Gordoniaceae, 7 – Tsukamurellaceae,
22 – Brevibacteriaceae, 2 – Promicromonosporaceae, 20 – Streptomycetaceae. Такой
перечень микроорганизмов, идентификация которых значительно ограничена при
использовании других методов делает КУППА доступным и перспективным
объектом для изучения в клинической микробиологии.
На ряду с перечисленными преимуществами, метод MALDI-ToF масс-
спектрометрии имеет и достаточный перечень недостатков и сложностей,
преодоление которых является важным с точки зрения оптимизации его
использования при работе со сложными и редкими микроорганизмами. В первую
очередь данный факт был описан в отношении микобактерий. При разработке
дополнительной библиотеки белковых спектров МТВс и НТМ выяснилось, что
питательная среда, на которой был получен рост культуры, влияет на качество
спектров, их интенсивность и возможность использования в рутинной
идентификации. В связи с этим разработчиками было принято решение о
включении в библиотеку спектров микроорганизмов, выросших на жидких и
плотных средах [151, 310].
Дополнительной проблемой, с которой могут столкнуться пользователи
масс-спектрометров, является сложность идентификации некоторых родов

38
КУППА из-за особенности архитектоники колоний (образования сухих колоний со
сложным строением) и строения клеточной стенки, которая значительно снижает
вероятность получения достаточного количества рибосомальных белков, точно
определяющих идентификацию культуры [141, 182, 211, 212, 213, 238, 305]. Данная
проблема частично решается использованием не стандартного протокола
пробоподготовки методом прямого нанесения, а применением расширенного
методами прямого нанесения и экстракции муравьиной кислотой, которые могут
значительно повысить качество проводимой идентификации [99].
Таким образом, масс-спектрометрия является важным элементом в
современной системе видовой идентификации микроорганизмов. Оптимизация
протоколов пробоподготовки и оценка влияния на идентификацию новых приемов
культивирования микроорганизмов могут значительно повысить качество
идентификации как клинически значимых микроорганизмов из группы КУППА,
так и сапрофитной микрофлоры, клиническое значение которой не доказано на
данный момент. В таком случае точная видовая идентификация и разработка
алгоритмов ее применения позволят улучшить диагностику заболеваний,
обусловленных КУППА.
1.4. Эпидемиологические особенности и клиническое значение
представителей семейств Mycobacteriaceae, Nocardiaceae, Gordoniaceae,
Tsukamurellaceae, Brevibacteriaceae, Promicromonosporaceae, Streptomycetaceae.
Естественным резервуаром для большинства аэробных актиномицет
является окружающая среда: в частности, почва, остатки гниющих растений, вода
из различных источников, в том числе из централизованных систем водоснабжения
[239]. КУППА формируют разнообразные микробные сообщества, которые в
окружающей среде могут выполнять важные микроэкологические функции, в
частности по биодеструкции различных загрязняющих веществ [27, 138, 236, 286].
С другой стороны, данная группа микроорганизмов обладает универсальными

39
механизмами выживания в воде и почве в условиях низких концентраций
питательных веществ и может образовывать биопленки, что может являться
важным фактором не только в формировании микробных консорциумов в
окружающей среде, но и в организме случайного хозяина [26, 136, 138]. В связи с
этим основной причиной развития заболеваний, обусловленных КУППА является
не наличие микроорганизма или его попадание в организм человека, а наличие
факторов риска, которые обуславливают развитие заболевания в случае
транзиторной колонизации сапрофитами окружающей среды. В настоящее время
клиницисты выделяют целый ряд факторов риска по развитию микобактериозов,
нокардиозов и других инфекционных осложнений, вызванных КУППА. Развитие
патологических процессов, связанных с НТМ в бронхолегочной системе чаще
всего происходит у пациентов с анатомо-физиологическими аномалиями легких
различной этиологии (бронхоэктазами, эмфиземой); у пациентов с
иммунологическими и генетическими заболеваниями (муковисцидозом,
первичной цилиарной дискинезии, дефицитом альфа-1-антитрипсина, синдромами
Вильямса-Кемпбелла, Мунье-Куна, Шегрена, альвеолярным протеинозом,
иммунодефицитами различной этиологии) [38, 39, 56, 171, 227, 277, 278, 293, 302].
В последние годы все чаще обсуждается вопрос о включении в группу риска по
развитию микобактериозов любых пациентов с хроническими бронхолегочными
заболеваниями и ВИЧ-инфицированных пациентов, а также пациентов с
аномалиями бронхолегочной системы неизвестной этиологии [9, 22, 40, 55, 59,
112]. Для нокардиозов и инфекций, вызванных гордониями и другими КУППА
дополнительно выделяют следующие факторы риска: медикаментозная
иммуносупрессия (после пересадки органов и тканей у пациентов, получающих
терапию системными стероидами, а также терапию по поводу онкологических или
ревматологических заболеваний), хроническая алкогольная интоксикация,
сахарный диабет, ожоги, попадание в организм инородных тел, колонизированных
кислотоустойчивыми микроорганизмами [192, 235, 286, 297, 324].
За исключением НТМ для других КУППА не описаны случаи передачи
микроорганизмов от человека к человеку. Чаще всего заражение или колонизация

40
происходит при контакте с предметами окружающей среды. В то же время в
литературе имеются данные об определенных географических особенностях,
связанных с распространенностью заболеваний, вызванных определенными
видами. Так, например, Nocardia brasiliensis чаще встречается в тропическом и
субтропическом климатах [139]. Наиболее изучены в этом плане
эпидемиологические особенности микобактериозов, для которых определены
региональные особенности распространения даже в пределах одной страны. В
Японии частота выделения Mycobacterium avium выше на севере страны, а
Mycobacterium intracelullare на востоке [176, 295]. Кроме географических и
климатических факторов, влияющих на распространение микобактериозов,
нокардиозов и других форм заболеваний, вызванных аэробными актиномицетами,
нельзя исключать роль культурных и национальных особенностей [145, 301].
Немаловажную роль при эпидемической оценке распространенности
патологии, обусловленной КУППА, играет экономическое развитие региона,
состояние лабораторной службы в частности, и здравоохранения в целом. Частота
распространенности микобактериозов в Северной Америке в период с 1988 по 2010
годы варьировалась от 5,6 в США, до 9,8 случаев в Канаде на 100 тыс. населения.
При этом видовое разнообразие значительно отличалось с преобладанием
медленнорастущих микобактерий в целом [225, 264, 281]. В странах Южной
Америки также преобладают медленнорастущие микобактерии со сменой
доминирующего вида, начиная с 2000-х годов, на Mycobacterium kansasii, с общим
уровнем заболеваемости в пределах 1,0 случая на 100 тыс. населения [263]. В
Европе заболеваемость микобактериозами ниже, чем в Канаде практически в 4
раза, в среднем 1,7 на 100 тыс. населения по данным 2010 года [260]. Для всех
вышеперечисленных стран в последнее десятилетие отмечается неуклонный рост
заболеваемости микобактериозами, особенно с учетом миграционной политики
[172, 173, 174, 175, 218, 226, 243, 279, 287].
Для большого числа регионов мира информация по заболеваемости
микобактериозами и другими формами инфекций, связанных с аэробными
актиномицетами, практически не представлена. В первую очередь это обусловлено

41
отсутствием официальных данных. Такая же ситуация складывается в Российской
Федерации. В последние пять лет в научной литературе стали появляться
публикации об эпидемиологических исследованиях микобактериозов. По данным
Центрального научно-исследовательского института г. Москвы преобладающими
в России являются медленнорастущие НТМ, среди которых были выделены как
клинически значимые виды, так и микобактерии, не имеющие доказанного
значения [63]. По данным Новосибирского научно-исследовательского института
туберкулеза, в период с 2013 по 2015 годы отмечен практически 4-х кратный рост
выделения медленнорастущих микобактерий от пациентов с подозрением на
микобактериоз [4]. Безусловно, такие работы имеют большое значение, но их
недостаточно для оценки реальной картины заболеваемости микобактериозами в
Российской Федерации [9, 11, 12, 63, 92, 104]. Информация об инфекционных
осложнениях, связанных с другими КУППА в отечественной научной литературе,
практически отсутствует.
С точки зрения этиопатогенеза развития микобактериозов, нокардиозов и
других инфекций, связанных с аэробными актиномицетами, инфицирование чаще
всего происходит аэрозольным механизмом [162, 191, 222, 259, 261, 280, 297, 312,
325]. Для большинства КУППА характерно факультативное внутриклеточное
паразитирование в макрофагах, что приводит к формированию клинических
процессов, схожих с туберкулезом [136, 159, 168].
Наиболее распространенными являются следующие формы локализации
инфекционного процесса, вызванного нокардиями и НТМ: поражение легких,
диссеминированные формы, лимфадениты, раневые инфекции. При диссеминации
наиболее тяжелые процессы локализуются в центральной нервной системе
(абсцессы мозга и менингиты). Отдельно необходимо выделить раневые инфекции,
связанные с установкой имплантатов и катетеров, развитие которых возможно не
только на фоне иммуносупрессии, но и у иммунокомпетентных пациентов [142,
188, 266, 286, 323]. Клиническое значение представителей рода Gordonia также
широко описано в этиологии целого спектра заболеваний. Так установлена
причастность G.terrae к развитию холецистита, поражения кожи и подкожной

42
клетчатки, абсцесса века, мастита, абсцесса мозга и менингита [133, 177, 202, 289],
G.bronchialis обнаруживается при рецидивирующих абсцессах молочных желез и
других мягких тканей, при вентрикулите [256, 289], G.polyisoprenivorans – при
эндокардитах [200], G.otitidis – при наружном отите и бронхитах [200, 201], G.sputi
– при эндокардите и медиастените [133], G.araii – при артритах [198]. Кроме того,
все вышеперечисленные микроорганизмы могут вызвать бактериемию, связанную
с использованием центрального венозного катетера [133, 134, 201, 256].
Для некоторых представителей кислотоустойчивых аэробных актиномицет
нет достоверных данных об их участии в патологических процессах у человека
(Brevibacteriaceae, Promicromonosporaceae, Streptomycetaceae) [132, 194, 283]. Для
других данные о клиническом значении представлены в виде единичных
публикаций (Tsukamurellaceae) [144, 267, 294, 316].
Следует отметить, что инфекционные процессы, вызванные КУППА
возможно встречаются намного чаще, однако из-за отсутствия алгоритмов их
выделения и идентификации большинство изолятов вероятнее всего
идентифицируются неправильно. Определение вида для кислотоустойчивых
актиномицет, на наш взгляд имеет важное клиническое значение. Даже на примере
НТМ, по литературным данным, многие близкородственные виды могут иметь
разную степень клинического значения. Определение вида для верификации
диагноза играет ключевое значение и с точки зрения особенностей природной
резистентности многих КУППА к различным антимикробным препаратам, и с
точки зрения накопления данных о новых потенциальных патогенах, клиническое
значение которых ранее не было описано и доказано.
В качестве примера деления микроорганизмов из группы КУППА по
клиническому значению можно привести классификацию быстро и
медленнорастущих НТМ, предложенную Американским обществом
микробиологов, основанную на наблюдении за этиологической причиной случаев
микобактериозов. Все НТМ, таким образом, предложено разделить на три группы:
имеющие доказанное клиническое значение, вызывающие единичные случаи
заболеваний, непатогенные для человека виды [141, 143, 193, 252, 253, 294, 306].

43
Такая условная классификация НТМ имеет важное значение не только для
клиницистов, но и для врачей-бактериологов, особенно в случае выделения
близкородственных видов НТМ, относящихся к разным группам по клиническому
значению. Перечень видов НТМ, распределенных на группы по клиническому
значению, представлен в таблицах 1, 2 и 3.
Таблица 1 – Виды нетуберкулезных микобактерий с доказанным
клиническим значением
Быстрорастущие
микобактерии
Медленнорастущие микобактерии
M.abscessus
M.bacteremicum
M.boenickei
M.canariasense
M.chelonae
M.cosmeticum
M.fortuitum
M.franklinii
M.goodii
M.houstonense
M.immunogenum
M.mageritense
M.mucogenicum
M.neoaurum
M.peregrinum
M.porcinum
M.senegalense
M.smegmatis
M.algericum
M.arosiense
M.arupense
M.asiaticum
M.avium
M.bohemicum
M.branderi
M.celatum
M.chimaera
M.colombiense
M.conspicuum
M.doricum
M.europaeum
M.florentinum
M.genavense
M.haemophilum
M.paraterrae
M.parmense
M.heidelbergense
M.heraklionense
M.interjectum
M.intracellulare
M.bouchedurhonense
M.saskatchewanense
M.heckeshornense
M.kansasii
M.koreense
M.lentiflavum
M.longobardum
M.malmoense
M.mantenii
M.marinum
M.marseillense
M.monteofiorense
M.parakoreense
M.parascrofulaceum
M.pseudoshottsii
M.riyadhense
M.scrofulaceum
M.sherrisii
M.shigaense
M.shimodei
M.shinjukuense
M.shottsii
M.simiae
M.stomatepiae
M.szulgai
M.timonense
M.triplex
M.tusciae
M.ulcerans
M.xenopi
M.yongonense
Среди НТМ, имеющих доказанное клиническое значение, преобладающими
являются медленнорастущие виды.
По данным на 2015 год, для более чем 70-и видов НТМ доказано клиническое
значение и еще для 29-и видов клиническое значение описано в единичных случаях
[193,248,249,252,253,255,306]. Естественно, что тест-системы, основанные на
методе ДНК-гибридизации, которые позволяют идентифицировать 28 видов НТМ,

44
не могут удовлетворить потребность в этиологической расшифровке
микобактериозов. И это частный пример, который затрагивает только
представителей одного рода из группы КУППА, при этом, безусловно, необходимо
анализировать данные о клиническом значении и других видов кислотоустойчивых
аэробных актиномицет.
Таблица 2 – Виды нетуберкулезных микобактерий, для которых
описаны единичные случаи выделения из клинического материала
Быстрорастущие микобактерии Медленнорастущие микобактерии
M.aubagnense
M.brisbanense
M.brumae
M.elefantis
M.iranicum
M.lacticola
M.monacense
M.moriokaense
M.neworleansense
M.novocastrense
M.phocaicum
M.septicum
M.setense
M.wolinsky
M.farcinogenes
M.fragae
M.intermedium
M.kumamotonense
M.kyorinense
M.lacus
M.nebraskense
M.nonchromogenicum
M.palustre
M.paraffinicum
M.paraseoulense
M.senuense
M.seoulense
M.terrae
M.vulneris
Для «редких» видов НТМ, имеющих клиническое значение, можно отметить
практически равномерное распределение среди быстро- и медленнорастущих
видов. Как правило, для таких микроорганизмов описаны единичные случаи
этиологической роли в развитии микобактериозов.
Среди непатогенных для человека видов значительно преобладают
быстрорастущие НТМ, однако следует отметить, что в группу микобактерий с
недоказанным клиническим значением относятся и достаточно часто описываемые
в отечественной литературе виды медленнорастущих НТМ, таких как M.gastri и
M.gordonae.

45
Таблица 3 – Непатогенные для человека виды нетуберкулезных
микобактерий
Быстрорастущие микобактерии Медленнорастущие
микобактерии
M.agri
M.aichiense
M.alvei
M.gadium
M.gilvum
M.aurum
M.chitae
M.chubuense
M.confluentis
M.crocinum
M.diernhoferi
M.duvali
M.fallax
M.flavescens
M.hassiacum
M.hodleri
M.holsaticum
M.fluoranthenivorans
M.frederiksbergense
M.aromaticivorans
M.austroafricanim
M.chlorophenolicum
M.insubricum
M.komossense
M.litorale
M.llatzerense
M.madagascariense
M.psychrotolerans
M.salmoniphilum
M.termoresistibile
M.murale
M.obuense
M.pallens
M.parafortuitum
M.phlei
M.poriferae
M.pyrenivorans
M.rhodesiae
M.rufum
M.rutilum
M.sphagni
M.tokaiense
M.vaccae
M.vanbaalenii
M.botniense
M.cookii
M.engbackii
M.gastri
M.gordonae
M.hiberniae
M.kubicae
M.minnesotense
M.noviomagense
M.paragordonae
M.pulveris
M.triviale
Таким образом, кислотоустойчивые аэробные актиномицеты – неоднородная
группа микроорганизмов, схожих по условиям культивирования,
морфологическим и тинкториальным признакам, характерной особенностью
которой является значительное разнообразие микроорганизмов при оценке их
клинического значения. Широкое распространение КУППА в природе и
увеличение числа пациентов из групп риска по развитию обусловленных ими
заболеваний требует разработки новых подходов для их выделения из
клинического материала, культивирования и идентификации. Без оптимизации
имеющихся на сегодняшний день алгоритмов микробиологической диагностики
невозможно решить вопрос с оценкой эпидемиологических и клинических
особенностей инфекционных осложнений, вызванных кислотоустойчивыми
микроорганизмами, и, как следствие, невозможна их адекватная антимикробная
терапия и профилактика.

46
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для достижения поставленной цели и решения соответствующих задач
исследования проведен анализ результатов 1 255 посевов образцов клинического
материала, полученных от 1 099 пациентов в период с 2016 по 2019 годы. Общее
количество выделенных штаммов микроорганизмов – 1 515. Все посевы,
включенные в исследование, были разделены по культуральным свойствам на две
группы: с признаками роста контаминирующей микрофлоры и с признаками роста
НТМ.
2.1. Дизайн исследования
В первой части исследования проводилась оценка видового разнообразия
контаминирующей микрофлоры, выделенной из клинического материала при
обследовании на туберкулез. Дополнительно проводилась оценка влияния
питательной среды, используемой для первичного посева на выделение КУППА из
клинического материала.
Во второй части исследования анализировалась структура микроорганизмов,
которые по культуральным свойствам были предварительно идентифицированы
как НТМ. Для всех культур была проведена первичная идентификация с
использованием метода ДНК-гибридизации с последующим сравнением с
результатами, полученными при использовании метода MALDI-ToF масс-
спектрометрии. Дополнительно была проанализирована частота выделения
кислотоустойчивых актиномицет и их структура в зависимости от пола и возраста
пациентов. Схема дизайна исследования приведена на рисунке 1.
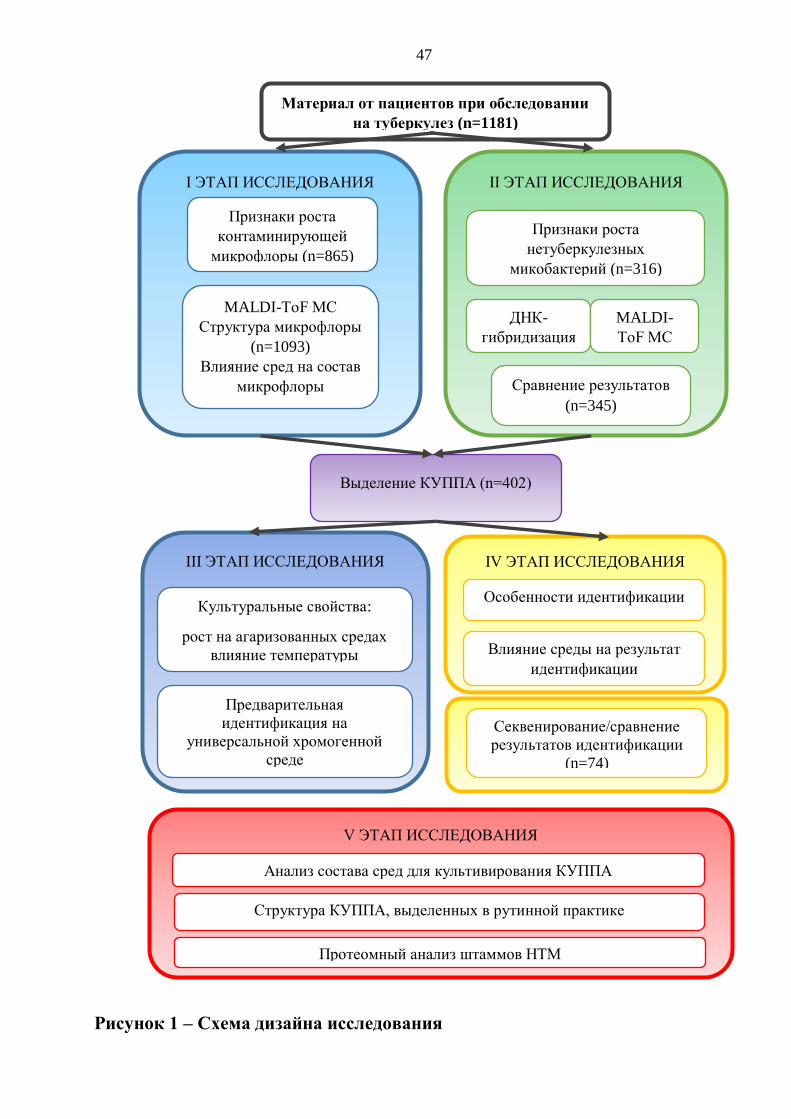
47
Рисунок 1 – Схема дизайна исследования
IV ЭТАП ИССЛЕДОВАНИЯ
Материал от пациентов при обследовании
на туберкулез (n=1181)
V ЭТАП ИССЛЕДОВАНИЯ
Анализ состава сред для культивирования КУППА
Структура КУППА, выделенных в рутинной практике
Протеомный анализ штаммов НТМ
I ЭТАП ИССЛЕДОВАНИЯ
Признаки роста
контаминирующей
микрофлоры (n=865)
MALDI-ToF МС
Структура микрофлоры
(n=1093)
Влияние сред на состав
микрофлоры
Выделение КУППА (n=402)
II ЭТАП ИССЛЕДОВАНИЯ
Признаки роста
нетуберкулезных
микобактерий (n=316)
ДНК-
гибридизация
MALDI-
ToF МС
Сравнение результатов
(n=345)
III ЭТАП ИССЛЕДОВАНИЯ
Культуральные свойства:
рост на агаризованных средах
влияние температуры
Предварительная
идентификация на
универсальной хромогенной
среде
Секвенирование/сравнение
результатов идентификации
(n=74)
Особенности идентификации
Влияние среды на результат
идентификации

48
Третий этап исследования был посвящен оценке возможности
культивирования КУППА, выделенных в первых двух этапах работы, на
агаризованных средах. Анализировались скорость роста микроорганизмов в
зависимости от питательной среды, их культуральные признаки, влияние
температуры. Отдельно была проведена оценка возможности культивирования и
предварительной идентификации КУППА с использованием универсальной
хромогенной среды.
Четвертый этап исследования включал в себя анализ возможности
использования метода MALDI-ToF масс-спектрометрии для идентификации
КУППА, выделенных на универсальной хромогенной среде.
На пятом этапе был проведен анализ состава питательных сред для
выделения КУППА, возможность использования предлагаемого алгоритма работы
с кислотоустойчивыми актиномицетами в рутинной микробиологической
практике, оценка гетерогенности популяции НТМ, выделенной из клинического
материала от одного пациента.
2.2. Характеристика исследуемых микроорганизмов
Из 1 515 штаммов микроорганизмов методом MALDI-ToF масс-
спектрометрии были идентифицированы мицелиальные и дрожжеподобные грибы,
некислотоустойчивые и кислотоустойчивые представители порядка
Actinomycetales, представители других групп грамположительных
микроорганизмов, представители порядка Enterobacterales, неферментирующие
грамотрицательные бактерии. Структура выделенной микрофлоры по группам
представлена в таблице 4.
Дополнительно в работе проанализированы штаммы КУППА, выделенные из
клинического материала в микробиологическом отделе КДЛ Клиник ФГБОУ ВО
СамГМУ Минздрава России в период с 2016 по 2019 годов, в которых было
идентифицировано 90 штаммов представителей порядка Actinomycetales.

49
Таблица 4 – Структура выделенной микрофлоры по группам,
идентифицированных в исследовании
Группа микроорганизмов n %
Микромицеты 295 19,5
Alphaproteobacteria 4 0,3
Betaproteobacteria 10 0,6
Gammaproteobacteria 162 10,6
Bacilli 513 33,9
Clostridia 3 0,2
Actinobacteria 528 34,9
Из всех представителей класса Actinobacteria 90,7% (479 штаммов) оказались
кислотоустойчивыми видами. Среди КУППА доминирующими были бактерии из
рода Mycobacterium, которые были идентифицированы в 75,2% случаях (360
штаммов) от всех кислотоустойчивых актиномицет.
В качестве контрольных культур при оценке ростовых свойств КУППА на
тестируемых агаризованных средах использовались штаммы из коллекции
Американской коллекции типовых культур (АТСС): Escherichia coli ATCC 25922,
Pseudomonas aeruginosa ATCC 27853.
2.3. Методы выделения и первичной идентификации микроорганизмов при
первичном посеве клинического материала
2.3.1. Методика первичного посева материала при обследовании на
туберкулез
Первичный посев клинического материала при обследовании на туберкулез
проводился в соответствии с Приказом № 109 МЗ РФ от 21 марта 2003 года «О
совершенствовании противотуберкулезных мероприятий в Российской Федерации
(с изменениями от 5 июня 2017 года) [89].
Посев диагностического материала проводился на плотные питательные
среды Левенштейн-Йенсена и Финн-II после его обработки 10% раствором натрия

50
фосфорнокислого трехзамещенного (Na3РO4). Пробирки с посевами помещались
горизонтально на «диваны» в термальной комнате, через три дня пробирки с
«диванов» переставлялись в штативы в вертикальное положение с просмотром
посевов. В дальнейшем просмотр всех пробирок, находящихся в термокамере, на
наличие роста микроорганизмов производился один раз в неделю. Рост
микроорганизмов оценивался по следующим критериям: время появления от
момента посева, форма и цвет колоний, наличие или отсутствие изменений цвета
среды и её консистенции.
2.3.2. Методика определения роста контаминирующей микрофлоры
Проба расценивалась как контаминированная посторонней микрофлорой в
случае выявления следующих визуальных признаков: интенсивный рост в течение
первой недели инкубации; изменение консистенции среды (нарушение
целостности скошенной части (трещины) или лизис среды); изменение цвета
среды. Последний признак характерен для некоторых представителей
контаминирующей микрофлоры, которые способны разлагать компоненты среды с
образованием кислых продуктов, приводить к снижению рН и переходу
малахитового зеленого в свободное состояние, что приводит к окрашиванию среды
в зеленый цвет. Рост контаминирующей микрофлоры может характеризоваться
отсутствием видимых колоний на фоне вышеописанных изменений. В случае
появления роста нехарактерного для микобактерий в более поздние сроки, посевы
также рассматривались как контаминированные посторонней микрофлорой.
2.3.3. Методика определения роста микобактерий на плотных яичных средах
Рост МТВс в среднем появляется через 3–4 недели от даты посева.
Исключение составляли случаи, когда материал сильно обсеменен МТВс, в таком

51
случае рост может наблюдаться уже через 7–10 суток от даты посева. Важно
отметить, что рост МТВс, а также полное отсутствие роста микроорганизмов не
приводит к изменению цвета и консистенции среды. На 10-12 неделе возможно
осветление скошенной части среды без признаков роста за счет деградации
молекул малахитового зеленого.
Признаком роста НТМ рассматривали появление на среде колоний различной
формы и окраски, без качественных изменений самой среды, а также появление
колоний на 10-12 неделе. Такие культуры подвергались процедуре идентификации
методом ДНК-гибридизации на тест-системах GenoType Mycobacterium CM и
GenoType Mycobacterium AS (Hain Lifescience GmbH, Германия).
Все культуры, вызывающие сомнения по своей принадлежности к МТВс
после визуальной оценки колоний, направлялись на дополнительные методы
идентификации: иммунохроматографический экспресс-тест SD BIOLINE TB Ag
MPT64 RAPID (Abbott, Франция), посев на чашку с кровяным агаром,
бактериоскопия культуры окраской по методу Циля-Нильсена и аурамин-
родамином, биохимические тесты – посев в пробирки со средой Левенштейн-
Йенсена с добавлением следующих реактивов: нитрата натрия (0,5 %); натрия
салициловокислого (1 мг/мл); паранитробензойной кислоты (500 мкг/мл); хлорида
натрия (5 %); гидразида тиофен-2-карбоксиловой кислоты (1–5 мкг/мл). Инкубация
посевов за исключением чашек с кровяным агаром проводилась в течение трех
недель при 37 °С.
Результаты биохимических тестов оценивались следующим образом: рост
МТВс наблюдался только на средах, содержащих нитрат натрия и гидразид
тиофен-2-карбоксиловой кислоты. Рост на других средах типичен для
представителей НТМ. При подозрении на рост НТМ культура отправлялась на
молекулярно-генетическое исследование GenoType Mycobacterium CM и GenoType
Mycobacterium AS (Hain Lifescience GmbH, Германия).

52
2.3.4. Методика определения роста микобактерий на жидкой среде
Посев материала на жидкую питательную среду проводился в специальные
пробирки BACTEC MGIT 960 (Becton Dickinson, США), содержащие питательную
среду Middlebrook 7H9 и специальную добавку, содержащую смесь антибиотиков:
полимиксина В, амфотерицина В, налидиксовой кислоты, триметоприма,
азлоциллина (PANTA) (Becton Dickinson, США). Обработка материала
производилась реактивом MycoPrep (Becton Dickinson, США), содержащим 4 %
раствор гидроксида натрия, 2,9 % раствор натрия лимоннокислого и растворенный
в них ацетилцистеин. Пробирки после посева помещали в прибор BACTEC 960
(Becton Dickinson, США) для дальнейшей инкубации.
Детекция роста производилась прибором в автоматическом режиме, по
изменению «светимости» пробирок, которая меняется в случае потребления
микроорганизмами кислорода, который находится в связанном виде с
флюорофорами в специальном слое на дне каждой пробирки. Для всех проб,
которые система прибора выдала как «положительные», была проведена
идентификация культур, включающая в себя: иммунохроматографический
экспресс-тест SD BIOLINE TB Ag MPT64 RAPID (Abbott, Франция), посев на
чашку с кровяным агаром, бактериоскопию культуры с окраской по методу Циля-
Нильсена и аурамин-родамином (наличие КУМ в образце).
Результат оценивался как «рост посторонней микрофлоры» в том случае,
если экспресс-тест показывал отрицательный результат, при бактериоскопии не
были обнаружены КУМ (как правило, в таких случаях в препарате определялись
другие микроорганизмы). Рост культуры на кровяном агаре мог отсутствовать.
Критерии интерпретации полученных результатов микробиологического
исследования при подозрении на рост НТМ представлены в таблице 5.

53
Таблица 5 – Интерпретация полученных результатов при подозрении на
рост НТМ, выросших на жидкой питательной среде Р
езульта
т
эксп
рес
с-те
ста
Рез
ульта
т п
осе
ва
на
чаш
ку с
кровян
ым
ага
ром
Рез
ульта
т
бак
тери
оск
оп
ии
Интерпретация результатов исследования
+ - + Рост МТВс
- + - Рост контаминирующей микрофлоры
- +/- + Возможный рост НТМ
+ + +/- Рост МТВс с контаминацией
+ - - Возможный рост МТВс, необходимо повторить
бактериоскопию
При необходимости для полученной культуры проводилось биохимическое
типирование, как и в случае с культурой, выращенной на плотных питательных
средах.
2.3.5. Метод ДНК-гибридизации для первичной идентификации
нетуберкулезных микобактерий
В работе использовался метод ДНК-гибридизации, основанный на
технологии обратной гибридизации продукта ПЦР с видоспецифичными зондами,
иммобилизованными на ДНК-стрипах, предложенный Hain Lifescience GmbH
(Германия).
В работе были использованы два набора для ступенчатой идентификации 28
видов. В первом наборе представлены наиболее часто встречающиеся
микобактерии: M.avium, M.chelonae, M.abscessus, M.fortuitum, M.gordonae,
M.intracellulare, M.scrofulaceum, M.interjectum, M.kansasii, M.malmoense,
M.marinum/M.ulcerans, M.peregrinum, M.xenopi; во втором – менее

54
распространенные виды: M.simiae, M.mucogenicum, M.goodii, M.celatum,
M.smegmatis, M.genavense, M.lentiflavum, M.heckeshornense,
M.szulgai/M.intermedium, M.phlei, M.haemophilum, M.kansasii, M.ulcerans, M.gastri,
M.asiaticum, M.shimoidei. Для идентификации НТМ были использованы культуры
микроорганизмов, выросших на жидких или плотных яичных средах в
соответствии с рекомендацией производителя.
ДНК-гибридизацию проводили на ДНК-амплификаторе GeneAmp 2720
(Thermo Fisher Scientific, США) и термостате электрическом для проведения ПЦР
амплификации ДНК GeneAmp PCR System 9700 (Applied Biosystems Inc., США), с
использованием наборов GenoType Mycobacterium CM и GenoType Mycobacterium
AS (Hain Lifescience GmbH, Германия).
2.4. Методика определения видового разнообразия микрофлоры, выделенной
при обследовании на туберкулез
В связи с отсутствием возможности проведения видовой идентификации на
базе бактериологической лаборатории противотуберкулезного диспансера
отрицательные на наличие представителей МТВс пробы с признаками роста
контаминирующей микрофлоры были переданы в микробиологический отдел КДЛ
Клиник ФГБОУ ВО СамГМУ Минздрава России. Идентификацию всех культур
проводили с использованием метода MALDI-ToF масс-спектрометрии на приборе
Microflex LT (Bruker Daltonik GmbH, Германия).
Для всех проб была проведена идентификация напрямую из пробирок с
плотными и жидкими средами. Если таким способом не удавалось получить
приемлемый результат, проводился пересев микроорганизмов на 5% кровяной агар
с бараньей кровью (HiMedia, Индия) и универсальную хромогенную среду
UriSelect (BioRad, США), агар Сабуро (HiMedia, Индия).

55
2.4.1. Методы выделения контаминирующей микрофлоры и
нетуберкулезных микобактерий для последующей идентификации
Пересев на агаризованные среды проводили одноразовыми петлями (10 мкл)
методом истончающегося штриха. Посевы инкубировали в термостате при
температуре 37 °С в течение трех суток с последующим культивированием в
течение 27-и суток при температуре 28 °С в аэробной среде. Для предотвращения
дегидратирования питательной среды использовали парафильм. Чашки при 28 °С
хранили в пластиковых контейнерах с крышкой.
Для части посевов были апробированы предложенные варианты
лабораторной посуды: чашка для культивирования микроорганизмов с
двухпозиционной резьбой (Патент РФ № 17302), флакон для культивирования
микроорганизмов (Патент РФ № 175134), флакон для культивирования
микроорганизмов со съемной крышкой (Патент РФ № 175863) [77, 78, 79].
2.4.2. Метод MALDI-ToF масс-спектрометрии для идентификации
микроорганизмов
В исследовании применялся метод матрично-активированной лазерной
десорбции/ионизации, основанный на многократном воздействии импульсов
лазера на бактериальные клетки, предварительно покрытые матрицей, с
последующей ионизацией белков. В качестве матрицы было использовалось
вещество – α-циано-4-гидроксикоричная кислота (НССА), (Bruker Daltonik GmbH,
Германия). Полученные в результате воздействия лазерного излучения
ионизированные белки микроорганизма попадали во времяпролетный анализатор
(Time-of-Flight, TOF) и двигались к детектору, который находился в конце
анализатора. Программное обеспечение прибора оценивало время пролета частиц
и преобразовывало эту информацию в спектр молекулярных масс (масс-спектр),
представленный в виде отношения массы и заряда в диапазоне от 2 000 Да до 20
000 Да. Полученный масс-спектр неизвестного микроорганизма сравнивался с

56
референсными спектрами из базы данных и, на основании наличия определенных
белков, их массы, интенсивности и частоты встречаемости происходила
идентификация микроорганизма.
Для идентификации использовалась стандартная библиотека спектров
компании Bruker Daltonik GmbH и дополнительные библиотеки спектров НТМ
(Mycobacteria Library версия 4.0). Последняя версия библиотеки для
идентификации НТМ содержит 880 референсных спектров, позволяющих
идентифицировать 159 видов микобактерий.
В работе были использованы три основных способа пробоподготовки
микроорганизмов согласно рекомендациям производителя, для использования с
методом MALDI-ToF масс-спектрометрии: метод прямого нанесения,
расширенный метод прямого нанесения, метод экстракции муравьиной кислотой.
Метод прямого нанесения выполнялся непосредственным нанесением
одиночной колонии (или ее части) тонким слоем на единичную лунку 96-луночной
мишени, начиная от середины лунки. После нанесения биоматериал покрывался 1
мкл свежеприготовленным раствором матрицы (НССА). Покрытие матрицей
проводилось в течение часа после нанесения материала.
Расширенный метод прямого нанесения заключался в том, что после
нанесения биоматериала тонким слоем непосредственно на точку мишени, начиная
от середины точки, после высыхания материал покрывался 1 мкл 70 % муравьиной
кислоты. После высыхания материал дополнительно покрывался раствором
матрицы (НССА).
Метод экстракции муравьиной кислотой использовался для повышения
качества данных, позволяющих идентифицировать культуры с максимальной
точностью [99]. Метод реализовался последовательностью действий:
1) в 300 мкл деионизированной воды вносили биоматериал в количестве от 1 до 5
колоний;
2) после вортексирования добавляли 900 мкл этанола и проводили повторное
вортексирование;

57
3) после центрифугирования в течение двух минут на максимальной скорости
(13 000 об/мин), пипеткой удаляли супернатант. В случае необходимости
процедуру центрифугирования проводили повторно для удаления остатков
этанола;
4) осадок высушивали при комнатной температуре в течение двух-десяти минут;
5) к высушенному осадку добавляли 70% муравьиной кислоты пропорционально
количеству осадка (от 1 до 80 мкл), материал перемешивали пипетированием;
6) в пробирку добавляли ацетнитрил в количестве равном объему муравьиной
кислоты и аккуратно перемешивали пипеткой;
7) после центрифугирования в течение двух минут на максимальной скорости
(13 000 об/мин) отбирали 1 мкл супернатанта и наносили на точку мишени;
8) после полного высыхания материала на точке, его покрывали 1 мкл раствора
матрицы (HCCA).
Дополнительно проводилось исследование, в котором на лунку мишени
наносили не супернатант, а осадок полученный в пункте 7, с последующим его
высушиванием и покрытием 1 мкл раствора матрицы (НССА).
Для всех культур за исключением грибов и КУППА был использован метод
прямого нанесения. Для части грибов и КУППА дополнительно использовали
расширенный метод прямого нанесения и метод экстракции муравьиной кислотой.
Для анализа гетерогенности популяции использовали метод экстракции
муравьиной кислотой.
Оценка результата идентификации с использованием MALDI-ToF масс-
спектрометра проводилась автоматически с помощью программного обеспечения
MALDI Biotyper RTC по уровню коэффициента совпадения (Score) от 0 до 3, при
этом низкодостоверная идентификация рассматривалась при уровне 0,000-1,699;
высокодостоверная идентификация до рода – 1,700-1,999; высокодостоверная
идентификация до вида – 2,000-2,999; в соответствии с рекомендациями
производителя оборудования.

58
2.5. Молекулярно-генетические методы идентификации микобактерий
В исследовании проводилось обнаружение ДНК МТВс в 74-х пробах с
признаками роста НТМ методом ПЦР c гибридизационно-флуоресцентной
детекцией (АмплиСенс® MTC-FL, ФБУН ЦНИИ Эпидемиологии
Роспотребназдора, Россия). В случае выделения ДНК МТВс проводилась их
внутривидовая идентификация методом ПЦР c гибридизационно-флуоресцентной
детекцией (АмплиСенс® MTC-diff-FL, ФБУН ЦНИИ Эпидемиологии
Роспотребназдора, Россия). Для проведения секвенирования по Сэнгеру
проводилась предварительная ПЦР с праймерами, фланкирующими фрагменты
генов 16S рРНК, hsp65, rpoB, sodА. Секвенирование проводилось с
использованием генетического анализатора Applied Biosystems® 3500, с
последующим анализом данных с помощью интернет-ресурса
RIDOM (http://rdna.ridom.de) и GenBank (http://www.ncbi.nlm.nih.gov/genbank/).
2.6. Методы оценки культивирования кислотоустойчивых представителей
порядка Actinomycetales на агаризованных средах
При оценке роста культур на агаризованных средах учитывались время
появления роста в сутках, культуральные свойства, дифференцирующие свойства
питательной среды; подготовку инокулюма культур, проведение количественных
испытаний и испытаний на селективность проводили в соответствии с методиками,
описанными в клинических рекомендациях «Внутрилабораторный контроль
качества питательных сред для клинических микробиологических исследований»,
2014 год.
2.6.1. Метод количественных испытаний агаризованных сред и испытаний
на селективность
Использовался инокулят микроорганизмов с концентрацией клеток 103
КОЕ/мл. Для оценки селективности и отсутствия подавления роста исследуемых

59
культур использовали инокулюм с концентрацией 104 КОЕ/мл. Оптическую
плотность инокулюма определяли по стандарту МакФарланда с использованием
денситометра Biosan (Латвия). Изначальная концентрация микроорганизмов
соответствовала 0,5 по МакФарланду.
На чашку с агаризованной средой осуществляли посев 0,1 мл смеси с
количеством 100 клеток, с последующим подсчетом колоний на каждой
анализируемой среде. Подготовку инокулюма проводили по схеме, описанной в
таблице 6.
Таблица 6 – Приготовление инокулюма микроорганизмов
Дополнительно рассчитывали критерий продуктивности – количество
клеток, давших видимый рост, выраженный в процентах. Отдельно сравнивали
показатели в трех группах: менее 50%, 50-69%, 70% и более.
№
пробирки
Количество
физ. раствора,
мл
Объем вносимой
суспензии, мл
Количество
микробных
клеток в 1 мл
взвеси
Количество
микробных
клеток в 0,1
мл взвеси
1 4,5
0,5 из исходной
взвеси 0,5 по
МакФарланду
107 1 000 000
2 4,5 0,5 из пробирки
№1 106 100 000
3 4,5 0,5 из пробирки
№2 105 10 000
4 4,5 0,5 из пробирки
№3 104 1000
5 4,5 0,5 из пробирки
№4 103 100
6 4,5 0,5 из пробирки
№5 102 10

60
2.6.2. Метод предварительной идентификации кислотоустойчивых
представителей порядка Actinomycetales при использовании универсальной
хромогенной среды
Исследование проводилось по авторской методике с использованием
универсальной хромогенной среды, на которую был получен Патент РФ № 2711954
[83]. В качестве универсальной хромогенной среды была использована плотная
питательная среда UriSelect (BioRad, США), на которую методом штриха
пересевалась чистая культура микроорганизмов, по свойствам соответствующая
нетуберкулезным микобактериям. В состав универсальной хромогенной среды
UriSelect входят хромогенные субстраты для обнаружения бактериальных
ферментов β-глюкуронидаз, β-галактозидаз и триптофан [114]. Инкубация посевов
проводилась при температуре 28°С до появления роста изолированных колоний с
последующим описанием культуральных свойств выросших микроорганизмов.
2.6.3. Метод использования универсальной хромогенной среды с селективной
добавкой для культивирования кислотоустойчивых представителей порядка
Actinomycetales
Исследование проводилось по авторской методике с использованием
универсальной хромогенной среды с селективной добавкой, на которую был
получен Патент РФ № 2711957 [84]. Первичный посев клинического материала без
предварительной деконтаминации проводился на питательную среду UriSelect
(BioRad, США), содержащую в качестве селективной добавки 200 ME бацитрацина
и 300000 ME полимиксина сульфата из расчета на 1000 мл готовой среды.
2.6.4. Методы оценки возможности выделения кислотоустойчивых
актиномицет из клинического материала
Для оценки возможности выделения и идентификации КУППА были
использованы модифицированные методы, адаптированные для клинического

61
материала, полученного от пациентов с различными нозологическими формами,
описанные в Патентах РФ № 2659155 и № 2668406 [80, 81].
2.7. Методы определения состава питательных сред
Определение состава питательных сред проводили в соответствии с МУК
4.2.2316-08 «Методы контроля бактериологических питательных сред» [60] в
модификации на биохимическом анализаторе Roche Hitachi 911 Chemistry Analyzer,
(Швейцария).
2.8. Методы математической обработки полученных данных
2.8.1. Методы анализа белковых профилей, полученных с помощью MALDI-
ToF масс-спектрометрии
Анализ белковых профилей, полученных методом MALDI-ToF масс-
спектрометрии проводили с использованием пакета программ flexAnalisis 3.0
(Bruker Daltonik GmbH, Германия). При анализе масс-спектров был использован
алгоритм детекции пиков Centroid. Порог сигнала к шуму был установлен на
уровне 2. Порог относительной интенсивности – 0 %, минимальная интенсивность
порога – 600 условных единиц, максимальное количество анализируемых пиков –
300. Для вычисления базовой линии использовали метод TopHat, сглаживание
спектров проводили по алгоритму SavitzkyGolay с массой 5 m/z за один цикл.
Результаты статистического протеомного сравнения масс-спектров были
визуализированы с помощью программного обеспечения MALDI Biotyper 3.0
Offline Classification (Bruker Daltonik GmbH, Германия). Для используемых в
исследовании штаммов были получены изображения спектров в виде псевдогелей
в GelView, а также проведен расчет составного индекса корреляции – Composite
Correlation Index методом BioTyper Composite Correlation Standart Method.
Идентификатор метода 180fcbb1-2349-428a-916b-5a87db00fae9, версия метода 1.1,

62
нижняя граница масс – 3000, верхняя граница масс – 12000, CCI параметры:
интервал 8, LaginDA – 10.
Процедуры субтипирования проводили в соответствии с официальными
рекомендациями производителя MALDI-ToF масс-спектрометра Microflex LT
(Bruker Daltonik GmbH, Германия).
2.8.2. Методы статистической обработки полученных результатов
Статистическую обработку полученных результатов проводили с учетом
распространенности отдельных групп микроорганизмов в зависимости от вида
клинического материала, сред для первичного посева, возраста и пола пациентов,
количества колоний культур микроорганизмов, сроков появления роста.
Группировку первичных данных и вычисления проводили с использованием пакета
программ Microsoft Excel® 2013.
Описание количественных признаков, отличающихся от нормального
распределения, представлено в виде медианы. Качественные признаки
представлены в виде долей процентов, абсолютного числа. Оценка структуры
микрофлоры включала расчет частоты встречаемости таксона в отдельных видах
клинического материала и относительного среднего для каждого
идентифицированного рода микроорганизмов (доля микроорганизма в
исследуемой совокупности представителей родов в зависимости от клинического
материала с учетом других факторов). Для оценки силы связи между
анализируемыми факторами, использовали критерий Х2 и степень закономерности
события (р), используемые при анализе сопряженности таблиц. Связь между
признаками статистически расценивали как значимую при уровне значимости
р<0,01 или р<0,05 в зависимости от статистики теста.
Для оценки различий между двумя независимыми выборками по уровню
определенного признака, измеренного количественно применяли критерий Манна-
Уитни. При проверке равенства медиан нескольких выборок использовали
критерий Краскела-Уоллиса.

63
ГЛАВА 3. СТРУКТУРА И РАСПРОСТРАНЕННОСТЬ
КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА
ACTINOMYCETALES В КОНТАМИНИРУЮЩЕЙ МИКРОФЛОРЕ,
ВЫДЕЛЕННОЙ ИЗ КЛИНИЧЕСКГО МАТЕРИАЛА ПРИ
ОБСЛЕДОВАНИИ НА ТУБЕРКУЛЕЗ
В связи с тем, что исследование на туберкулез требует обязательного
проведения процедур деконтаминации клинического материала, полученного из в
норме нестерильных локусов, выделение кислотоустойчивой микрофлоры в
данной группе микробиологических исследований является наиболее вероятным.
Кроме этого практически все актиномицеты характеризуются замедленным
ростом, что значительно ограничивает возможности стандартных исследований в
условиях классических микробиологических лабораторий. Использование
специальных питательных сред и удлиненный период инкубации в совокупности с
деконтаминацией материала перед посевом создают благоприятные условия для
роста значительного количества видов аэробных актиномицет. Однако в
микробиологических лабораториях противотуберкулезной службы отсутствуют
возможности для их видовой идентификации за исключением представителей
МТВс и некоторых видов НТМ. В связи с этим нами было принято решение
проанализировать распространенность и видовое разнообразие
кислотоустойчивых представителей порядка Actinomycetales в клиническом
материале при обследовании на туберкулез.
На сегодняшний день, контаминирующая микрофлора рассматривается
врачами-бактериологами как балласт, мешающий проведению исследований
клинического материала на микобактерии. Ее выделение при работе с материалом
не подразумевает идентификации полученных культур, и тем более оценки
потенциального клинического значения. Данный факт, на наш взгляд, является
недооцененным, так как вместе с классическими контаминантами,
представленными сапрофитной, гноеродной и гнилостной микрофлорой, в
клиническом материале могут находиться потенциальные патогены, строение

64
клеточной стенки которых способствует выживанию в процессе деконтаминации.
В первую очередь к ним можно отнести разнообразных представителей порядка
Actinomycetales: Brevibacterium spp., Cellulosimicrobium spp., Gordonia spp., НТМ,
Nocardia spp., Streptomyces spp., Tsukamurella spp.
3.1. Структура контаминирующей микрофлоры при обследовании на
туберкулез
Для оценки структуры контаминирующей микрофлоры нами был разработан
алгоритм, по которому исследован видовой состав контаминантов при
обследовании на туберкулез в 865 пробах различного клинического материала,
засеянного на 3 среды: Левенштейна-Йенсена, Финн-II и Миддлбрук 7Н9. Все
пробы были отрицательны в отношении представителей МТВс. Во всех пробах
выявлены признаки роста контаминирующей микрофлоры. Из пробирок с
контаминирующим ростом, которые были включены в исследование, произведен
пересев на плотные агаризованные питательные среды, перечисленные в главе 2.
Общее время инкубации составило 21 сутки. Выбор условий культивирования был
обусловлен замедленным ростом основной массы кислотоустойчивых
микроорганизмов и их умеренной психрофильностью.
Все выросшие культуры идентифицированы с использованием MALDI-ToF
масс-спектрометра Microflex LT (Bruker Daltonik GmbH, Германия) методом
прямого нанесения. В случае получения неприемлемого результата идентификации
использовали метод расширенного прямого нанесения и метод экстракции
муравьиной кислотой в соответствии с рекомендациями производителя
оборудования. При идентификации грибов дополнительно использовали
микроскопический метод. Из 865 проб всего было выделено и идентифицировано
1093 штамма микроорганизмов.
Посевы, включенные в исследование, были представлены различным
клиническим материалом, который подвергался процессу деконтаминации и
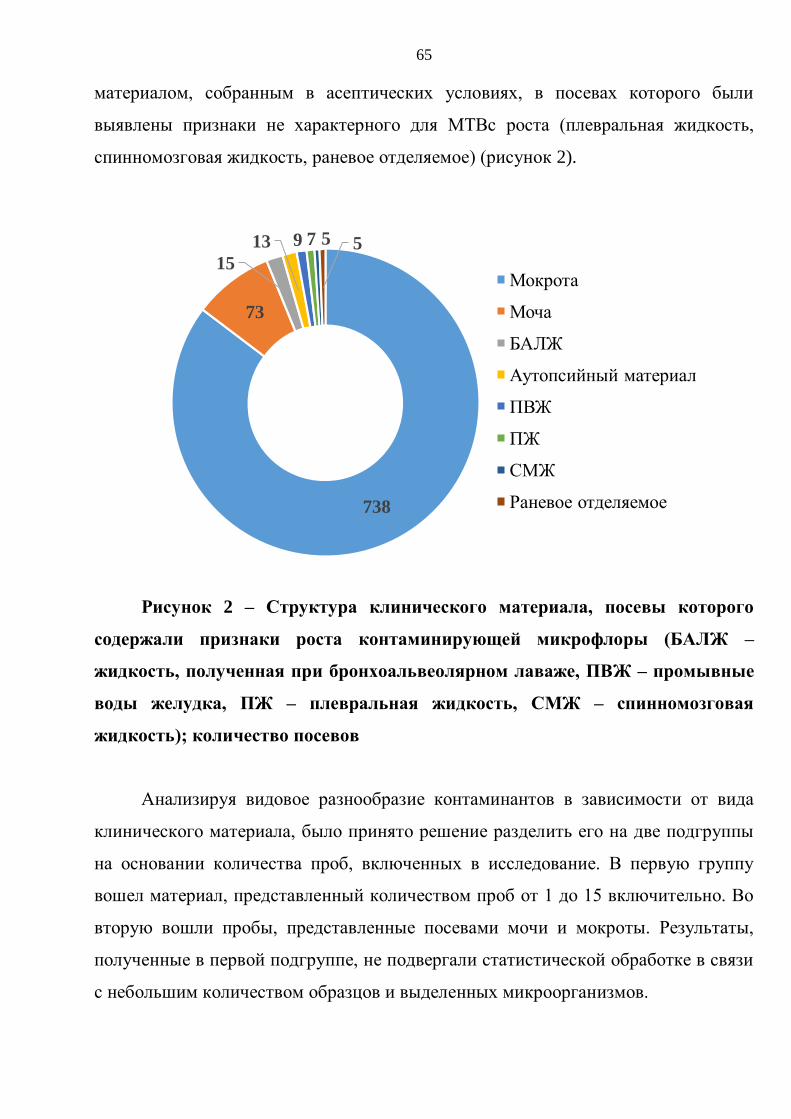
65
материалом, собранным в асептических условиях, в посевах которого были
выявлены признаки не характерного для МТВс роста (плевральная жидкость,
спинномозговая жидкость, раневое отделяемое) (рисунок 2).
Рисунок 2 – Структура клинического материала, посевы которого
содержали признаки роста контаминирующей микрофлоры (БАЛЖ –
жидкость, полученная при бронхоальвеолярном лаваже, ПВЖ – промывные
воды желудка, ПЖ – плевральная жидкость, СМЖ – спинномозговая
жидкость); количество посевов
Анализируя видовое разнообразие контаминантов в зависимости от вида
клинического материала, было принято решение разделить его на две подгруппы
на основании количества проб, включенных в исследование. В первую группу
вошел материал, представленный количеством проб от 1 до 15 включительно. Во
вторую вошли пробы, представленные посевами мочи и мокроты. Результаты,
полученные в первой подгруппе, не подвергали статистической обработке в связи
с небольшим количеством образцов и выделенных микроорганизмов.
738
73
15
13 9 7 5 5
Мокрота
Моча
БАЛЖ
Аутопсийный материал
ПВЖ
ПЖ
СМЖ
Раневое отделяемое

66
Из аутопсийного материала было выделено 17 штаммов микроорганизмов: в
девяти пробах был выделен один вид, в четырех пробах контаминирующая
микрофлора была представлена ассоциацией из двух видов. В аутопсийном
материале не было выделено нитчатых грибов и кислотоустойчивых актиномицет.
В трех пробах был выделен Enterococcus faecalis двух случаях был
идентифицирован Staphylococcus epidermidis. Остальные микроорганизмы были
выделены однократно: Acinetobacter baumanii, Candida tropicalis, Citrobacter
freundii, Citrobacter braakii, Klebsiella pneumoniae, Escherichia coli, Enterobacter
cloacae, Enterococcus faecium, Pseudomonas aeruginosa, Pseudomonas lundensis,
Serratia liquefaciens, Staphylococcus aureus. Таким образом, доминирующей
группой в контаминирующей микрофлоре при обследовании аутопсийного
материала оказались бактерии порядка Enterobacterales, которые были определены
в 46,2% проб.
В структуре контаминирующей микрофлоры, выделенной из промывных вод
желудка, были выделены грибы, которые оказались доминирующими и были
представлены одним штаммом Aspergillus nidulans, двумя штаммами Aspergillus
terreus, по одному штамму грибов рода Candida – C.albicans и C.parapsilosis. Из
бактериальной микрофлоры было выделено два штамма S.epidermidis и один
штамм Nocardia nova. Из одной пробы не удалось получить рост микроорганизмов
в условиях исследования.
Из плевральной жидкости были выделены три штамма энтеробактерий (E.coli
и K.pneumoniae), два штамма грибов из рода Penicillium (один из штаммов был
идентифицирован только до рода, второй штамм был идентифицирован как
P.chrysogenum), по одному штамму Pseudomonas monteilii и M.avium. Следует
отметить, что рост штамма M.avium был получен на обеих плотных средах,
характеризовался отсутствием кислотоустойчивости при окраске по методу Циль-
Нильсена, колонии были слизистыми, ярко-желтыми. Культуральные свойства
были схожи с некоторыми штаммами S.aureus, выделенного на плотных яичных
средах.

67
В пробах с посевами спинномозговой жидкости, несмотря на признаки
контаминации первичных посевов, в трех случаях не удалось получить рост в
условиях исследования. В одном случае был выделен гриб рода Sporothrix, в одном
– P.aeruginosa.
В структуре контаминирующей микрофлоры, выделенной из раневого
отделяемого, был получен рост монокультур E. faecalis, Enterococcus raffinosus,
Staphylococcus haemolyticus, а также одна ассоциация из M.chelonae и Micrococcus
luteus. Из одной пробы рост микрофлоры в условиях исследования получить не
удалось.
Контаминирующая микрофлора, выделенная из жидкости
бронхоальвеолярного лаважа была выделена в 14 образцах из 15, включенных в
исследование. В данном виде материала получено значительное разнообразие
групп микроорганизмов, были выделены мицелиальные грибы,
неферментирующие грамотрицательные бактерии, грамположительные и
грамотрицательные кокки, нетуберкулезные микобактерии. В одном образце была
выделена ассоциация, представленная двумя штаммами бактерий: S.aureus и
Ralstonia pickettii. В остальных случаях микроорганизмы выделены в
монокультурах: A.baumanii, Aspergillus versicolor, E.coli, E.cloacae, E.faecalis,
K.pneumoniae, Moraxella osloensis, Mucor spp., Sporotrix spp. – по одному штамму,
M.avium и P.aeruginosa – по два штамма.
Оба выделенных штамма M.avium обладали атипичными тинкториальными и
культуральными свойствами, схожими со штаммом M.avium, выделенном из
плевральной жидкости. Несмотря на небольшой объем выборки в данной части
исследования из 54 образцов клинического материала в пяти пробах были
выделены представители кислотоустойчивых аэробных актиномицет, что
составило 9,3% от общего количества проб из первой подгруппы клинического
материала. Важной является выявленная в исследовании особенность трех
штаммов M.avium, которые по тинкториальным и культуральным свойствам были
схожи с классическими представителями контаминирующей микрофлоры.
Выделение одного штамма N.nova из промывных вод желудка можно расценивать

68
как случайную находку, тем не менее, имеющую значение как примера выделения
нокардий из клинического материала.
3.2. Контаминирующая микрофлора, выделенная при исследовании мочи и
мокроты
Преаналитический этап исследования мокроты не может исключить
контаминацию собранного материала сапрофитной микрофлорой со слизистых
оболочек верхних дыхательных путей. Схожая ситуация складывается и при
исследовании мочи. Данный факт требует обязательной деконтаминации обоих
видов клинического материала, описание микробного состава которого будет
представлено ниже.
Особенностью мокроты в данном случае является то, что многие
представители кислотоустойчивых аэробных актиномицет могут вызывать
патологические состояния бронхолегочной системы, сопровождающиеся
клинической картиной туберкулезного процесса. В связи с этим анализ структуры
контаминирующей микрофлоры в отношении распространенности в ней
представителей порядка Actinomycetales приобретает элементы диагностического
поиска, особенно если учитывать, что среди данной группы микроорганизмов есть
и клинически значимые, и сапрофитные виды [34].
Моча, как объект микробиологического исследования длительное время
считалась условно стерильной, однако в последние годы появляется все больше
фактов, подтверждающих ее значительную контаминацию различными видами
сапрофитных и условно-патогенных микроорганизмов, в связи с этим важно
оценить в структуре микрофлоре долю кислотоустойчивых актиномицет [62,117].
Структура контаминирующей микрофлоры, выделенной из образцов
мокроты, включенных в исследование была представлена 794 штаммами. В
отличие от образцов из первой подгруппы, в мокроте значительно чаще были
выделены многокомпонентные ассоциации. Так, трехкомпонентные ассоциации
были выделены в 10 случаях (1,4%), а состоящие из двух видов микроорганизмов

69
– в 103 (14,0%). В условиях исследования из 59 образцов (8,0%) с признаками
контаминации, не удалось получить рост микрофлоры. При анализе структуры
контаминирующей микрофлоры в моче было выделено 76 штаммов
микроорганизмов. Без роста оказалось семь образцов, что составило 9,6%, от всех
образцов мочи, включенных в исследование. Двухкомпонентные ассоциации были
определены в десяти случаях (13,7%), многокомпонентных ассоциаций не
выявлено.
Общая информация по частоте выделения и структуре отдельных групп
микроорганизмов представлена в таблицах 7,8 и 9. Микроорганизмы, которые были
выделены однократно приведены в таблицах в условных объединенных группах,
обозначенных «другие».
Таблица 7 – Частота выделения и структура грибов, выделенных из
мокроты и мочи
Группа
микроорганизмов
Вид клинического материала
Уровень p* Мокрота Моча
n %** n %**
Candida spp. 63 28,1 0 0,0 0,011
Aspergillus spp. 79 35,3 2 40,0 0,036
Exophiala spp. 2 0,9 1 20,0 0,132
Magnusiomyces spp. 16 7,1 0 0,0 0,216
Mucor spp. 14 6,3 0 0,0 0,243
Penicillium spp. 30 13,3 2 40,0 0,611
Rhizomucor spp. 2 0,9 0 0,0 0,662
Scopulariopsis spp. 10 4,5 0 0,0 0,325
Trichophyton spp. 2 0,9 0 0,0 0,662
Trichosporon spp. 2 0,9 0 0,0 0,662
Другие 4 1,8 0 0,0 0,535
Итого: 224 100,0 5 100,0 p<0,001
Примечание: *Связь между признаками статистически значима при уровне
p<0,05 при расчете связи между видом клинического материла и микрофлорой,
выделенной из него по отношению к общему количеству штаммов, выделенных
из мокроты и мочи. **Доля от общего числа выделенных микроорганизмов в
подгруппе из соответствующего вида материала.

70
Микробное разнообразие, выделенное из мокроты, значительно преобладало
над микрофлорой, выделенной из мочи. В 19 образцах мокроты были однократно
выделены следующие микроорганизмы: Achromobacter xylosoxidans, Acremonium
strictum, Actinocorallia libanotica, Agromyces mediolanus, Bordetella petrii,
Clostridium interstinale, Cohnella fontinalis, Erwinia persicina, Ewingella americana,
Filifactor vilosus, Fusarium proliferatum, Lactococcus lactis, Morganella morganii,
Ohrobactrum anthropi, Pediococcus pentosaceus, Rhisopus spp., Rothia mucilaginosa,
Sphingobacterium mizutaii. Из мочи в качестве единичного представителя был
выделен Pseudochrobactrum asaccharolyticum.
Таблица 8 – Частота выделения и структура грамположительных
микроорганизмов, выделенных из мокроты и мочи
Группа
микроорганизмов
Вид клинического материала
Уровень p* Мокрота Моча
n %** n %**
Некислотоустойчивые представители порядка Actinomycetales
Corynebacterium spp. 19 76,0 11 84,6 p<0,001
Arthrobacter spp. 2 8,0 0 0,0 0,662
Microbacterium spp. 2 8,0 2 15,4 0,662
Другие 2 8,0 0 0,0 0,662
Итого: 25 100,0 13 100,0 p<0,001
Грамположительные микроорганизмы из других групп
Staphylococcus spp. 232 65,7 20 57,1 0,590
Enterococcus spp. 36 10,2 9 25,7 0,007
Lactobacillus spp. 28 7,9 4 11,4 0,444
Bacillus spp. 28 7,9 1 2,9 0,305
Lysinibacillus spp. 3 0,9 0 0,0 0,592
Paenibacillus spp. 9 2,5 0 0,0 0,351
Streptococcus spp. 7 2,0 0 0,0 0,411
Micrococcus spp. 3 0,9 0 0,0 0,592
Другие 7 2,0 1 2,9 0,706
Итого: 353 100,0 35 100,0 0,797
Примечание: *Связь между признаками статистически значима при
уровне p <0,05 при расчете связи между видом клинического материла и
микрофлорой, выделенной из него по отношению к общему количеству
штаммов, выделенных из мокроты и мочи. **Доля от общего числа выделенных
микроорганизмов в подгруппе из соответствующего вида материала.

71
По частоте выделения в контаминирующей микрофлоре из мокроты,
преобладали различные виды стафилококков и грибов, количество которых
оказалось 232 (29,3%) и 223 (28,1%) штаммов соответственно. Следует отметить,
что грибы в целом, без разделения на виды (p<0,001), а также представители родов
Candida и Aspergillus, встречались статистически достоверно чаще в
контаминирующей микрофлоре, выделенной из мокроты, чем из мочи. В моче было
выделено всего 20 штаммов стафилококков и пять штаммов грибов, что составило
соответственно 26,3% и 6,6%.
Таблица 9 – Частота выделения и структура грамотрицательных
микроорганизмов, выделенных из мокроты и мочи
Группа
микроорганизмов
Вид клинического материала
Уровень p* Мокрота Моча
Абс %** Абс %**
Представители порядка Enterobacterales
Escherichia spp. 12 23,1 9 64,4 p<0,001
Serratia spp. 9 17,3 0 0,0 0,351
Enterobacter spp. 7 13,5 1 7,1 0,706
Proteus spp. 2 3,8 1 7,1 0,132
Klebsiella spp. 19 36,5 3 21,4 0,411
Другие 3 5,8 0 0,0 0,592
Итого: 52 100,0 14 100,0 p<0,001
Неферментирующие грамотрицательные бактерии
Pseudomonas spp. 23 50,0 1 16,7 0,421
Ralstonia spp. 3 6,5 4 66,6 p<0,001
Acinetobacter spp. 7 15,3 1 16,7 0,706
Sphingomonas spp. 3 6,5 0 0,0 0,592
Stenotrophomonas spp. 5 10,9 0 0,0 0,488
Methylobacterium spp. 2 4,3 0 0,0 0,662
Другие 3 6,5 0 0,0 0,592
Итого: 46 100,0 6 100,0 0,463
Примечание: *Связь между признаками статистически значима при уровне
p <0,05 при расчете связи между видом клинического материла и микрофлорой,
выделенной из него по отношению к общему количеству штаммов, выделенных
из мокроты и мочи. **Доля от общего числа выделенных микроорганизмов в
подгруппе из соответствующего вида материала.

72
Отдельно от всех микроорганизмов была рассмотрена распространенность в
контаминирующей микрофлоре, выделенной из мокроты и мочи
кислотоустойчивых представителей порядка Actinomycetales. Данная группа
оказалась третьей по численности группой микроорганизмов, выделенных из
мокроты. Общее количество идентифицированных видов в мокроте составило 94
(11,9%), что в целом сопоставимо с распространенностью кислотоустойчивых
аэробных актиномицет в других видах клинического материала, описанных в
подглаве 3.1. Подробная структура представителей данной группы, выделенных из
мокроты и мочи представлена в таблице 10.
Таблица 10 – Частота выделения и структура кислотоустойчивых
представителей порядка Actinomycetales, выделенных из мокроты и мочи
Группа
микроорганизмов
Вид клинического материала
Уровень р* Мокрота Моча
n %** n %**
Gordonia spp. 5 5,3 0 0,0 0,488
Mycobacterium spp. 32 34,0 0 0,0 0,075
Brevibacterium spp. 5 5,3 2 66,7 0,063
Nocardia spp. 9 9,6 0 0,0 0,351
Cellulosimicrobium spp. 2 2,1 1 33,3 0,132
Rhodococcus spp. 2 2,1 0 0,0 0,662
Streptomyces spp. 36 38,3 0 0,0 0,058
Tsukamurella spp. 3 3,3 0 0,0 0,592
Итого: 94 100,0 3 100,0 0,037
Примечание: * Связь между признаками статистически значима при уровне
p <0,05 при расчете связи между видом клинического материла и микрофлорой,
выделенной из него по отношению к общему количеству штаммов, выделенных
из мокроты и мочи. ** Доля от общего числа выделенных кислотоустойчивых
актиномицет из соответствующего вида материала.
Различия по частоте выделения кислотоустойчивых аэробных актиномицет в
целом, выделенных из мокроты в сравнении с мочой оказались статистически
достоверны (р=0,037). Однако следует отметить, что по отдельным представителям
достоверных различий выявить не удалось. Тем не менее, несмотря на то, что не
были получены статистически значимые различия, отдельные роды порядка

73
Actinomycetales, таких как Gordonia, Mycobacterium, Nocardia, Rhodococcus,
Streptomyces и Tsukamurella были выделены только в составе контаминирующей
микрофлоры из материала, представленного мокротой.
3.3. Кислотоустойчивые представители порядка Actinomycetales в структуре
контаминирующей микрофлоры
Анализируя видовое разнообразие выделенных из контаминирующей
микрофлоры кислотоустойчивых представителей аэробных актиномицет было
идентифицировано 29 видов. Роды Gordonia, Rhodococcus, Tsukamurella были
выявлены только в составе контаминирующей микрофлоры из мокроты и были
представлены одним видом: G.sputi, R.equi, T.paurometabola соответственно. Все
штаммы гордоний и родококков были выделены в монокультуре. Два штамма
T.paurometabola были определены в монокультуре, один – в трехкомпонентной
ассоциации с Paenibacillus rhizosphaerae и Arthrobacter polychromogenes. Данный
факт можно рассматривать в качестве примера истинной контаминации.
Нокардии были выделены в основном из мокроты, за исключением одного
штамма, выделенного из промывных вод желудка. Всего было выделено и
идентифицировано десять штаммов нокардий: по три штамма N.cyriacigeorgica и
N.farcinica, два штамма N.brevicatena, по одному штамму N.carnea и N.nova. Оба
штамма N.brevicatena были выделены в ассоциации с Corynebacterium urealyticum.
Один штамм N.cyriacigeorgica был выделен совместно с Bacillus horneckiae. Все
остальные штаммы нокардий были выделены в монокультуре. Ассоциации с
другими микроорганизмами в данном случае можно рассматривать как критерий
истинной контаминации. Однако выделение представителей рода Nocardia в
других случаях можно рассматривать, как факт, имеющий относительную
клиническую значимость.
В отличие от представителей Gordonia, Rhodococcus, Tsukamurella и
Nocardia, имеющих потенциальное клиническое значение в составе

74
контаминирующей микрофлоры были выделены и сапрофитные виды из
окружающей среды, а также комменсалы кожи и слизистых оболочек. К ним были
отнесены представители родов Brevibacterium и Cellulosimicrobium. В работе было
выделено семь штаммов Brevibacterium: шесть штаммов – B.casei, один из которых
был выделен в ассоциации с S.haemolyticus и один штамм B.borstelensis,
выделенный в монокультуре. Все три штамма C.cellulans были выделены в
монокультуре.
Значительное видовое разнообразие было отмечено для двух родов
кислотоустойчивых аэробных актиномицет: Streptomyces и Mycobacterium.
Стрептомицеты были представлены шестью видами: S.phaeochromogenes (12
штаммов), S.albus (1 штамм), S.avidinii (1 штамм), S.badius (2 штамма), S.chartreusis
(2 штамма), S.griseus (1 штамм), S.hirsutus (1 штамм), S.lavendulae (2 штамма),
S.violaceoruber (10 штаммов). Для четырех штаммов не удалось провести видовую
идентификацию. В двух случаях была выявлена ассоциация, состоящая из двух
видов стрептомицет: S.badius+ S.griseus, S.violaceoruber+ S.phaeochromogenes, в
трех случаях была выявлена ассоциация стрептомицет с нетуберкулезными
микобактериями: две ассоциации S.lavendulae+M.chelonae и одна
S.hirsutus+M.celatum. Один штамм S.chartreusis был выделен в ассоциации с
Bacillus atrophaeus. Таким образом для стрептомицет больше, чем для других видов
кислотоустойчивых актиномицет характерны ассоциации с другими
контаминантами (шесть случаев двухкомпонентных ассоциаций).
Отдельно следует остановится на нетуберкулезных микобактериях, которые
были выделены в качестве представителей контаминирующей микрофлоры. Как
было сказано выше, при описании культуральных и тинкториальных свойств
штаммов M.avium, характеризующихся признаками нетипичными для НТМ,
практически у всех остальных штаммов микобактерий обнаружены схожие с
классической контаминирующей микрофлорой признаки: ускоренный рост на
плотных яичных средах, выраженная пигментация, мукоидный тип колоний,
изменение цвета среды. В случае выделения в составе ассоциаций с другими
микроорганизмами было выявлено разжижение яичных сред. Всего было выделено

75
и идентифицировано 36 штаммов НТМ. Все штаммы были идентифицированы до
вида. Видовое разнообразие и частота встречаемости в составе контаминирующей
микрофлоры представлена на рисунке 3.
Рисунок 3 – Структура нетуберкулезных микобактерий, выделенных при
анализе контаминирующей микрофлоры (абсолютные значения)
По данным научной литературы, проанализированной в Главе 1, из
выделенных НТМ доказанное клиническое значение имеют представители
следующих видов: M.avium, M.fortuitum, M.chelonae, M.senegalense, M.porcinum,
M.abscessus, M.celatum, M.malmoensae, M.marseillense, которые оказались
доминирующими в структуре НТМ – 28 штаммов. M.septicum, представленная
двумя штаммами в качестве этиологического агента при патологии у человека
описана в единичных случаях. Не имеют клинического значения представители
видов M.chitae (1 штамм) и M.gordonae (5 штаммов). Таким образом, в
исследовании НТМ, имеющие потенциальное клиническое значение микобактерии
оказались преобладающей группой и составила 77,8% от всех штаммов НТМ,
8
6
55
3
2
2
11
1 1 1 M.avium
M.fortuitum
M.chelonae
M.gordonae
M.senegalense
M.porcinum
M.septicum
M.abscessus
M.chitae
M.celatum
M.malmoensae
M.marseillense

76
выделенных из контаминирующей микрофлоры. Важным является факт выделения
практически всех штаммов НТМ из органов дыхательной системы, особенно с
учетом того, что НТМ в качестве этиологического агента вызывают поражения
данной локализации.
Часть НТМ была выделена в ассоциациях с другими микроорганизмами. В
трех случаях была выявлена ассоциация с представителями рода Streptomyces: две
ассоциации M.chelonae+S.lavendulae, одна – M.celatum+S.hirsutus. В двух случаях
была выявлена ассоциация трех видов НТМ –
M.fortuitum+M.senegalense+S.epidermidis. Единственный штамм M.chitae был
выявлен в ассоциации со Stenotrophomonas maltophilia. Интересной оказалась
ассоциация M.malmoensae с Aspergillus niger, так как оба микроорганизма могут
вызвать поражения органов дыхания. В трех случаях была выявлена
двухкомпонентная ассоциация M.gordonae с коагулазонегативными
стафилококками (в двух случаях с S.epidermidis, в одном – S.hominis). Один из
штаммов M.chelonae был выделен совместно с M.luteus.
Анализ полученных данных дает четкую картину возможности выделения
КУППА из клинического материала с признаками роста контаминирующей
микрофлоры при обследовании на туберкулез. Изучаемая группа микроорганизмов
наиболее часто встречалась в материале, представленном мокротой. При этом, если
условно рассчитать долю кислотоустойчивых аэробных актиномицет, имеющих
клиническое значение (по литературным данным), то она составит 4,6% (50
штаммов) от всех выделенных в исследовании микроорганизмов, а для всех
кислотоустойчивых актиномицет вне зависимости от их потенциального
клинического значения – 9,3% (102 штамма).

77
3.4. Оценка влияния питательной среды для первичного посева на
выделение представителей порядка Actinomycetales из клинического
материала
Полученные результаты исследования, описанные в первых трех частях
данной главы, показали достаточно широкую распространенность
кислотоустойчивых представителей порядка Actinomycetales в составе
контаминирующей микрофлоры, выделенной из различных видов клинического
материала. Данный факт подтверждает предположение о том, что искусственные
питательные среды для культивирования представителей МТВс являются
подходящими для выделения кислотоустойчивых актиномицет. Однако среды, на
которые проводился первичный посев материала, включенного в исследование,
значительно отличаются по составу. Обе плотные среды представлены яичными
средами, в которых в значительных количествах присутствуют белки и жиры, а
жидкая питательная среда Миддлбрук 7Н9 - полусинтетическая среда более
стандартизированной с одной стороны, а с другой, по результатам многочисленных
исследований характеризующаяся появлением роста микобактерий в более
короткие сроки. В связи с этим было принято решение проанализировать влияние
используемой для первичного посева питательной среды на качественный состав
контаминирующей микрофлоры и выделение кислотоустойчивых представителей
порядка Actinomycetales.
Анализ роста контаминирующей микрофлоры на средах для первичного
посева выявил определенные закономерности: максимальное количество посевов с
признаками контаминирующего роста было выявлено на среде Левенштейна-
Йенсена, меньше всего – на среде Миддлбрук 7Н9. Каждая пятая проба имела
признаки контаминации сразу на двух плотных средах, при этом в случае
выявления роста на жидкой питательной среде Миддлбрук 7Н9 роста на плотных
питательных средах не наблюдалось. В 19,4% проб признаки роста
контаминирующей микрофлоры были выявлены на обеих плотных питательных
средах. Особенности роста контаминирующей микрофлоры представлены на
рисунке 4.

78
Рисунок 4 – Рост контаминирующей флоры на средах для первичного посева
(абсолютные значения); в скобках указаны значения в процентах от общего
числа посевов, включенных в исследование
При пересеве на дополнительные среды в 69 случаях не было выявлено роста
микроорганизмов в условиях исследования. Со среды Левенштейна-Йенсена при
пересеве не удалось получить рост в 17 случаях, со среды Финн-II – 16, со среды
Миддлбрук 7Н9 – 36 случаях. На среде Левенштейна-Йенсена было выделено и
идентифицировано 520 штаммов, на среде Финн-II – 366 штаммов, на среде
Миддлбрук 7Н9 – 207 штаммов различных микроорганизмов. На среде
Левенштейна-Йенсена были выявлены двух и трехкомпонентные ассоциации в 70
случаях (15,3%) и в семи случаях (1,5%) соответственно. На среде Финн-II
многокомпонентные ассоциации встречались реже, также состояли из двух – в 57
случаях (17,7%) и трех микроорганизмов в трех случаях (0,9%). На среде
Миддлбрук 7Н9 ассоциации, состоящие из двух и трех микроорганизмов, были
выделены, в 27 (13,0%) и в двух случаях (1,0%) соответственно.
В структуре контаминирующей микрофлоры, давшей рост на среде
Левенштейна-Йенсена преобладающими, оказались представители следующих
494
(57,1%)
322
(37,2%)214
(24,7%) 168
(19,4%)
0
100
200
300
400
500
600

79
родов: Aspergillus spp., Bacillus spp., Candida spp., Corynebacterium spp.,
Enterococcus spp., Escherichia spp., Klebsiella spp., Lactobacillus spp., Magnusiomyces
spp., Mucor spp., Mycobacterium spp., Staphylococcus spp., Pseudomonas spp.,
Penicillium spp. Представители других групп микроорганизмов были выделены в
единичных случаях. Таким образом, основными микроорганизмами-
контаминантами на среде Левенштейна-Йенсена оказались грибы – представители
нормальной микрофлоры слизистых оболочек – и энтеробактерии.
Нетуберкулезные микобактерии составили 3,5% от общего количества штаммов,
выросших на среде Левенштейна-Йенсена, все штаммы которых по культуральным
свойствам значительно отличались от классического варианта роста.
Структура контаминирующей микрофлоры, выросшей на среде Финн-II, по
составу и преобладающим микроорганизмам со средой Левенштейна-Йенсена. На
ней, как и на среде Левенштейна-Йенсена доминирующими оказались
микромицеты, стафилококки, энтерококки и лактобактерии. Представители рода
Mycobacterium составили 3,8% от всех выросших культур. При этом на среде Финн-
II значительно реже выделялись энтеробактерии и неферментирующие
грамотрицательные микроорганизмы. Рост нетуберкулезных микобактерий,
которые при первичной оценке были отнесены к контаминирующей микрофлоре
был также атипичен.
Максимальные различия в составе контаминирующей микрофлоры в
сравнении с плотными питательными средами оказались со средой Миддлбрук
7Н9. Видовое разнообразие классических контаминантов на данной среде
оказалось меньше, а представители кислотоустойчивых актиномицет были
выделены в значительно большем количестве. Доминирующей микрофлорой
остались представители различных видов стафилококков, а вот второе место в
структуре заняли не грибы, а бактерии рода Streptomyces, которые вместе с
Mycobacterium spp., Nocardia spp. и Brevibacterium spp. были выделены в 26,1% от
всех посевов с контаминирующей микрофлорой из среды Миддлбрук 7Н9.
Дополнительно были выделены представители родов Cellulosimicrobium spp.,
Gordonia spp., Rhodococcus spp., Tsukamurella spp., рост которых не был получен на
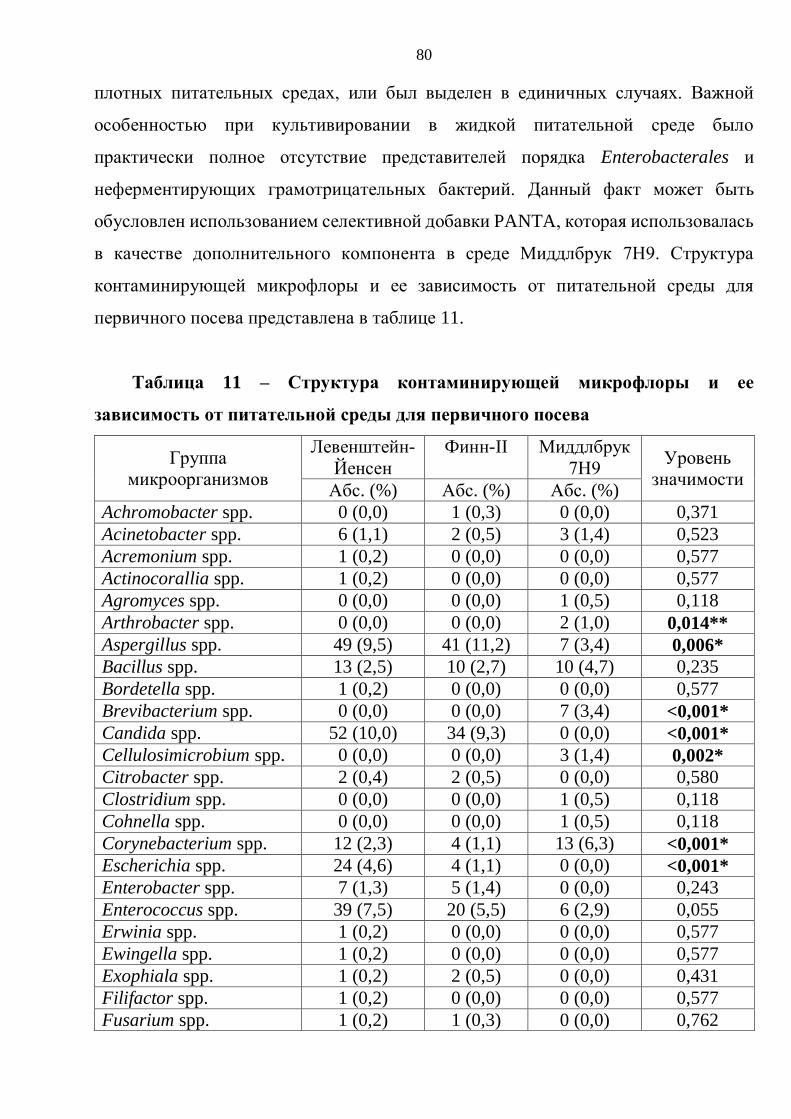
80
плотных питательных средах, или был выделен в единичных случаях. Важной
особенностью при культивировании в жидкой питательной среде было
практически полное отсутствие представителей порядка Enterobacterales и
неферментирующих грамотрицательных бактерий. Данный факт может быть
обусловлен использованием селективной добавки PANTA, которая использовалась
в качестве дополнительного компонента в среде Миддлбрук 7Н9. Структура
контаминирующей микрофлоры и ее зависимость от питательной среды для
первичного посева представлена в таблице 11.
Таблица 11 – Структура контаминирующей микрофлоры и ее
зависимость от питательной среды для первичного посева
Группа
микроорганизмов
Левенштейн-
Йенсен
Финн-II Миддлбрук
7Н9 Уровень
значимости Абс. (%) Абс. (%) Абс. (%)
Achromobacter spp. 0 (0,0) 1 (0,3) 0 (0,0) 0,371
Acinetobacter spp. 6 (1,1) 2 (0,5) 3 (1,4) 0,523
Acremonium spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Actinocorallia spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Agromyces spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Arthrobacter spp. 0 (0,0) 0 (0,0) 2 (1,0) 0,014**
Aspergillus spp. 49 (9,5) 41 (11,2) 7 (3,4) 0,006*
Bacillus spp. 13 (2,5) 10 (2,7) 10 (4,7) 0,235
Bordetella spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Brevibacterium spp. 0 (0,0) 0 (0,0) 7 (3,4) <0,001*
Candida spp. 52 (10,0) 34 (9,3) 0 (0,0) <0,001*
Cellulosimicrobium spp. 0 (0,0) 0 (0,0) 3 (1,4) 0,002*
Citrobacter spp. 2 (0,4) 2 (0,5) 0 (0,0) 0,580
Clostridium spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Cohnella spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Corynebacterium spp. 12 (2,3) 4 (1,1) 13 (6,3) <0,001*
Escherichia spp. 24 (4,6) 4 (1,1) 0 (0,0) <0,001*
Enterobacter spp. 7 (1,3) 5 (1,4) 0 (0,0) 0,243
Enterococcus spp. 39 (7,5) 20 (5,5) 6 (2,9) 0,055
Erwinia spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Ewingella spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Exophiala spp. 1 (0,2) 2 (0,5) 0 (0,0) 0,431
Filifactor spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Fusarium spp. 1 (0,2) 1 (0,3) 0 (0,0) 0,762

81
Группа
микроорганизмов
Левенштейн-
Йенсен
Финн-II Миддлбрук
7Н9 Уровень
значимости Абс. (%) Абс. (%) Абс. (%)
Gordonia spp. 0 (0,0) 0 (0,0) 3 (1,4) 0,002*
Klebsiella spp. 23 (4,4) 18 (4,9) 0 (0,0) 0,007*
Lactobacilus spp. 22 (4,2) 19 (5,2) 4 (1,9) 0,167
Lactococcus spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Lysinibacillus spp. 0 (0,0) 0 (0,0) 2 (1,0) 0,014**
Mycobacterium spp. 18 (3,5) 14 (3,8) 8 (3,9) 0,947
Magnusiomyces spp. 11 (2,1) 11 (3,0) 1 (0,5) 0,130
Microbacterium spp. 0 (0,0) 0 (0,0) 4 (1,9) <0,001*
Micrococcus spp. 1 (0,2) 0 (0,0) 2 (1,0) 0,093
Methylobacter spp. 1 (0,2) 1 (0,3) 0 (0,0) 0,762
Moraxella spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Morganella spp. 1 (0,2) 1 (0,3) 0 (0,0) 0,762
Mucor spp. 10 (1,9) 5 (1,4) 1 (0,5) 0,339
Nocardia spp. 1 (0,2) 1 (0,3) 8 (3,9) <0,001*
Oсhrobactrum spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Paenibacillus spp. 0 (0,0) 1 (0,3) 6 (2,9) <0,001*
Pediococcus spp. 1 (0,2) 1 (0,3) 0 (0,0) 0,762
Penicillium spp. 13 (2,5) 11 (3,0) 12 (5,8) 0,075
Proteus spp. 2 (0,4) 1 (0,3) 1 (0,5) 0,919
Pseudochrobactrum spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Pseudomonas spp. 27 (5,2) 8 (2,2) 1 (0,5) 0,003*
Ralstonia spp. 0 (0,0) 1 (0,3) 7 (3,4) <0,001*
Rhizomucor spp. 2 (0,4) 1 (0,3) 0 (0,0) 0,671
Rhodococcus spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Rhodotorula spp. 1 (0,2) 0 (0,0) 0 (0,0) 0,577
Scopulariopsis spp. 7 (1,3) 3 (0,8) 0 (0,0) 0,222
Serratia spp. 7 (1,3) 5 (1,4) 0 (0,0) 0,243
Sphingobacterium spp. 0 (0,0) 0 (0,0) 1 (0,5) 0,118
Sporothrix spp. 0 (0,0) 0 (0,0) 2 (1,0) 0,014**
Staphylococcus spp. 147 (28,2) 130 (35,5) 50 (24,1) 0,009*
Stenotrophomonas spp. 4 (0,8) 2 (0,5) 1 (0,5) 0,876
Streptococcus spp. 4 (0,8) 3 (0,8) 3 (1,4) 0,667
Streptomyces spp. 0 (0,0) 2 (0,5) 32 (15,4) <0,001*
Trichophyton spp. 1 (0,2) 2 (0,5) 1 (0,5) 0,659
Tsukamurella spp. 1 (0,2) 0 (0,0) 2 (1,0) 0,093
Итого штаммов 520 (100,0) 366 (100,0) 207 (100,0) –
Примечание * Связь между признаками статистически значима при уровне p
<0,01. ** Связь между признаками статистически значима при уровне p <0,05.

82
Анализируя полученные данные, можно сделать заключение о том, что
плотные питательные среды, используемые для первичного посева клинического
материала при обследовании на туберкулез более подвержены росту классической
контаминирующей микрофлоры. Использование жидкой питательной среды
Миддлбрук 7Н9 позволяет расширить перечень микроорганизмов, которые могут
быть выделены из клинического материала при обследовании на МТВс, ставит
вопрос о разработке алгоритмов их идентификации, оценке клинического значения
у пациентов и, в случае подтверждения их роли в этиологии патологического
процесса, вопроса об определении чувствительности к антимикробным препаратам
и критериев интерпретации полученных результатов. Статистически достоверно
влияние выбора среды Миддлбрук 7Н9 на повышение высева из клинического
материала кислотоустойчивых представителей порядка Actinomycetales и снижение
высева некоторых микромицет, Escherichia spp, Klebsiella spp., бацилл и
неферментирующих грамотрицательных бактерий.
3.5. Резюме
Контаминирующая микрофлора при обследовании на туберкулез
клинического материала на туберкулез – разнообразная группа микроорганизмов.
Кислотоустойчивые аэробные актиномицеты представлены в ней различными
представителями родов Mycobacterium spp., Nocardia spp., Gordonia spp.,
Rhodococcus spp., Tsukamurella spp., Brevibacterium spp., Cellulosimicrobium spp.,
Streptomyces spp. Среди них могут быть как клинически незначимые
микроорганизмы – сапрофиты окружающей среды и комменсалы слизистых
оболочек тела человека, так и потенциальные патогены. По данным нашего
исследования каждая десятая культура, выделенная из проб с признаками
контаминации, оказалась представителем кислотоустойчивых аэробных
актиномицет.

83
Безусловно, на данный момент отсутствуют единые алгоритмы работы
лабораторий различных служб, которые могли бы оптимизировать
микробиологическое исследование при обследовании на МТВс и расширить его
возможности в вопросах выделения и идентификации других микроорганизмов,
способных вызывать схожую с туберкулезом клиническую картину. Однако,
увеличение количества иммунокомпрометированных пациентов из групп риска по
развитию заболеваний, вызванных кислотоустойчивыми актиномицетами, с
каждым годом будет требовать от врачей-бактериологов разработки новых
алгоритмов, позволяющих выделять и идентифицировать их из клинического
материала.
Полученные данные о влиянии питательной среды для первичного посева на
структуру контаминирующей микрофлоры могут быть использованы для
оптимизации работы как микробиологических лабораторий противотуберкулезной
службы, так и микробиологических лабораторий, в работе которых не проводятся
исследования на МТВс.
Проведение такого рода исследований необходимо для оценки
распространенности потенциальных патогенов, выделение которых затруднено
при рутинном микробиологическом исследовании, что связано в первую очередь с
короткими сроками инкубации посевов в обычной микробиологической практике.
Отдельно необходимо анализировать и описывать случаи выделения НТМ с
измененными культуральными свойствами, при выделении их из клинического
материала. Важно отметить, что по нашим данным в 3,5-3,9% вне зависимости от
среды первичного посева были выделены представители как медленно, так и
быстрорастущих микобактерий, характерной особенностью которых является
атипичный, не характерный для микобактерий рост, схожий с ростом
контаминирующей гноеродной и гнилостной микрофлоры.

84
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ,
ИЗЛОЖЕННЫМ В ГЛАВЕ 3
Статьи в журналах, рекомендованных ВАК:
1. Лямин, А.В. Микобактериозы: особенности эпидемиологии и лабораторной
диагностики / А.В. Лямин, Д.Д. Исматуллин // Аспирантский вестник Поволжья. –
2016. – № 5-6. – С. 204-208.
2. Лямин, А.В. Лабораторная диагностика микобактериозов / А.В. Лямин, А.В.
Жестков, Д.Д. Исматуллин [и др.] // Вестник современной клинической медицины.
– 2017. – Т. 10. – № 1. – С. 29-35.
3. Родионова, Ю.Д. Оценка влияния условия хранения мокроты на витальные
свойства микобактерий туберкулеза / Ю.Д. Родионова, О.А. Гусякова, А.В. Лямин
// Туберкулез и болезни легких. – 2017. – Т. 95. – № 1. – С. 42-46.
4. Лямин, А.В. Лабораторная диагностика микобактериозов у пациентов с
муковисцидозом (обзор литературы) / А.В. Лямин, Д.Д, Исматуллин, А.В. Жестков
[и др.] // Клиническая лабораторная диагностика. – 2018. – Т. 63. – № 5. – С. 315-
320.
5. Лямин, А.В. Общая характеристика и клиническое значение представителей
родов Nocardia и Gordonia / А.В. Лямин, А.В. Жестков, Т.Р. Никитина [и др.] //
Инфекция и иммунитет. – 2019. – Т. 9. – № 3-4. – С. 429-436.
6. Лямин, А.В. Контаминирующая микрофлора при обследовании на
туберкулез: зависимость от питательных сред для первичного посева / А.В. Лямин
// Астраханский медицинский журнал. – 2019. – Т. 14. – № 4. С. 29-36.
7. Лямин, А.В. Контаминирующая микрофлора при обследовании на
туберкулез: сапрофиты или потенциальные патогены? / А.В. Лямин, Д.Д,
Исматуллин, А.В, Жестков [и др.] // Иммунопатология, аллергология,
инфектология. – 2019. – №3. – С. 63-70.
Публикации в других изданиях:
1. Ковалев, А.М. Структура контаминирующей флоры, выделенной при посеве
диагностического материала на туберкулез / А.М. Ковалев, Т.П. Персиянцева, Н.М.

85
Баранова [и др.] // Лабораторная служба. – 2017. – Т. 6. – № 3. – Тезисы III
Российского конгресса лабораторной медицины. – М. – С. 80.
2. Козлов, А.В. Сравнение методов деконтаминации мокроты при исследовании
микобактерий / А.В. Козлов, А.В. Лямин, А.А. Ерещенко [и др.] // Лабораторная
служба. – 2017. – Т. 6. – № 3. – Тезисы III Российского конгресса лабораторной
медицины. – М. – С. 81.
3. Исматуллин, Д.Д. Новые возбудители заболеваний респираторного тракта у
иммунокопрометированных пациентов (обзор литературы) / Д.Д, Исматуллин,
М.О. Золотов, А.В. Лямин [и др.] // Наука и инновации в медицине. – 2019. – Т. 4.
– № 3. – С. 19-24.

86
ГЛАВА 4. СТРУКТУРА КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ
ПОРЯДКА ACTINOMYCETALES, ПРЕДВАРИТЕЛЬНО
ИДЕНТИФИЦИРОВАННЫХ КАК НЕТУБЕРКУЛЕЗНЫЕ
МИКОБАКТЕРИИ
Вторая часть исследования включала в себя анализ структуры выделенных из
клинического материала при обследовании на туберкулез микроорганизмов,
которые были предварительно идентифицированы как НТМ. Основными
критериями в данном случае были культуральные признаки при оценке роста
микроорганизмов на плотных яичных средах. В случае выявления признаков роста
НТМ культура идентифицировалась с использованием метода ДНК-гибридизации.
Все штаммы микроорганизмов реидентифицировались с использованием метода
MALDI-ToF масс-спектрометрии. При невозможности получения приемлемых
результатов идентификации из первичных посевов повторная идентификация
проводилась после пересева материала с первичных сред на агаризованные среды:
кровяной 5 % агар и универсальную хромогенную среду, – для получения чистых
культур микроорганизмов.
В исследование были включены пробы, собранные в период с 2016 по 2019
годы на базе бактериологической лаборатории ГБУЗ «Самарский областной
клинический противотуберкулезный диспансер им. Н.В. Постникова». Общее
количество образцов материала, включенных в исследование – 316 проб. При
анализе результатов идентификации методом ДНК-гибридизации количество
культур было условно принято за количество проб. При реидентификации
выделенных культур с использованием MALDI-ToF масс-спектрометрии
анализировались все выделенные культуры вне зависимости от принадлежности к
группе НТМ.

87
4.1. Анализ результатов первичной идентификации микрофлоры с
использованием метода ДНК-гибридизации
Во всех посевах на яичных средах не выявлено признаков роста
контаминирующей микрофлоры, не отмечено разжижения сред, визуальных
признаков роста мицелиальных грибов. При оценке культуральных свойств
признаков роста нескольких культур в пробе выявлено не было. При тестировании
иммунохроматографическим методом все пробы в отношении МТВс были
отрицательными. Идентификацию выросших культур проводили без
дополнительных пересевов напрямую из пробирок с первичным ростом.
С помощью ДНК-гибридизации провели видовую идентификацию 188
штаммов, что составило 59,5% от общего числа культур, включенных в
исследование. Из 128 оставшихся культур для 22 штаммов (7,0% от общего числа
культур) была определена принадлежность к роду Mycobacterium, для 67 штаммов
– не было получено приемлемого результата идентификации (21,2% от числа
культур), 39 штаммов (12,3% от числа культур) идентифицированы как
микроорганизмы с высоким содержанием ГЦ. Принадлежность к НТМ без учета
результатов видовой идентификации была определена для 68,7% культур.
Анализ структуры НТМ показал, что преобладающей группой оказались
медленнорастущие виды (M.avium, M.gordonae, M.kansasii), которые составили
67,0% от всех идентифицированных микобактерий. Среди быстрорастущих НТМ
доминирующими видами были M.fortuitum и M.abscessus, которые в совокупности
составили 26,6% от общего количества идентифицированных до вида НТМ.
Остальные микобактерии, идентифицированные до вида, были представлены
единичными штаммами (рисунок 5).
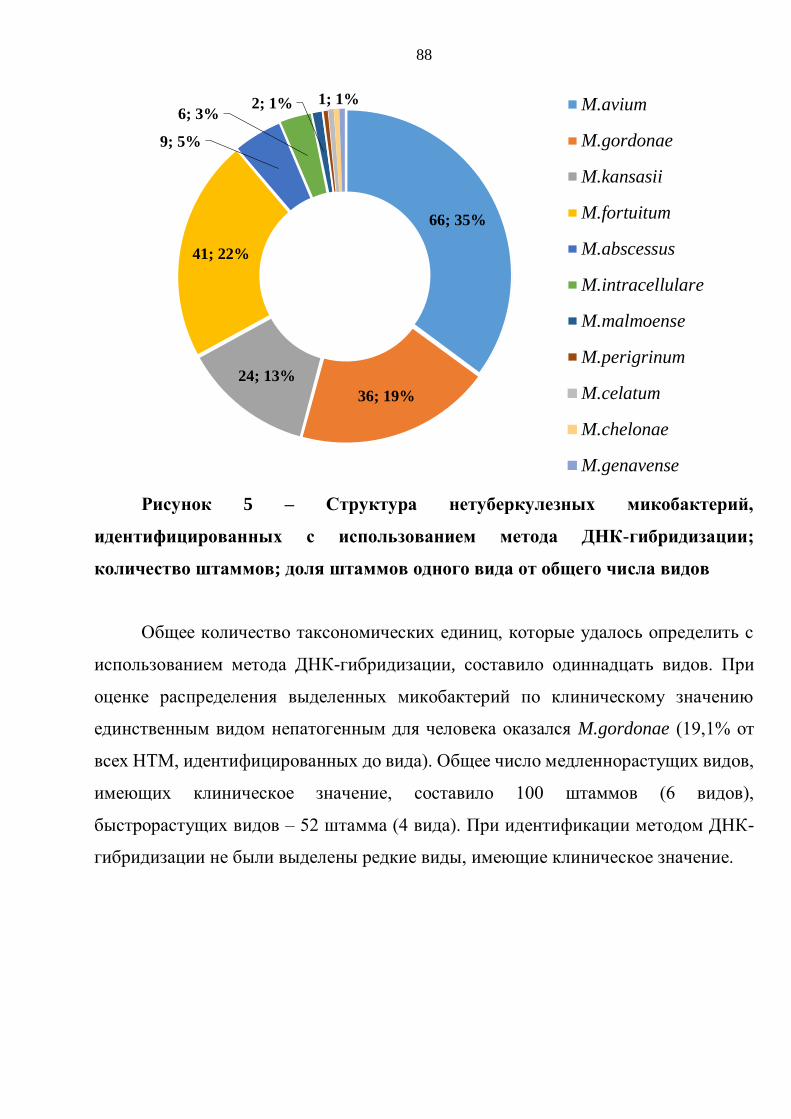
88
Рисунок 5 – Структура нетуберкулезных микобактерий,
идентифицированных с использованием метода ДНК-гибридизации;
количество штаммов; доля штаммов одного вида от общего числа видов
Общее количество таксономических единиц, которые удалось определить с
использованием метода ДНК-гибридизации, составило одиннадцать видов. При
оценке распределения выделенных микобактерий по клиническому значению
единственным видом непатогенным для человека оказался M.gordonae (19,1% от
всех НТМ, идентифицированных до вида). Общее число медленнорастущих видов,
имеющих клиническое значение, составило 100 штаммов (6 видов),
быстрорастущих видов – 52 штамма (4 вида). При идентификации методом ДНК-
гибридизации не были выделены редкие виды, имеющие клиническое значение.
66; 35%
36; 19%
24; 13%
41; 22%
9; 5%
6; 3%2; 1% 1; 1% M.avium
M.gordonae
M.kansasii
M.fortuitum
M.abscessus
M.intracellulare
M.malmoense
M.perigrinum
M.celatum
M.chelonae
M.genavense

89
4.2. Структура микроорганизмов, предварительно идентифицированных как
нетуберкулезные микобактерии по результатам MALDI-ToF масс-
спектрометрии
Результаты, полученные при анализе видового разнообразия
микроорганизмов, идентифицированных с использованием MALDI-ToF масс-
спектрометрии, с одной стороны, закономерны благодаря более широким
возможностям данного метода исследования, с другой стороны, определяют ряд
вопросов: достаточно ли визуальной оценки роста микрофлоры на плотных яичных
средах для предварительного отбора культур в группу нетуберкулезных
микобактерий, насколько ДНК-гибридизация позволяет решать вопросы видовой
идентификации таких культур, насколько широко представлены другие виды из
группы КУППА в образцах клинического материала с признаками роста НТМ?
С использованием MALDI-ToF масс-спектрометрии было
идентифицировано 345 штаммов микроорганизмов. С учетом пересевов на
плотные агаризованные среды было выявлено 28 полимикробных ассоциаций с
преобладанием двухкомпонентных – 26 (92,9%). Две ассоциации состояли из трех
микроорганизмов. При идентификации культур напрямую из пробирок с
первичным посевом для материала, содержащего ассоциации не удалось получить
приемлемые результаты идентификации. Во всех случаях материал был пересеян
на универсальную хромогенную среду и кровяной агар. Одна трехкомпонентная
ассоциация имела следующий состав: M.pseudoshottsii + M.szulgai + S.lavendulae.
Вторая – M.malmoense + Rhodococcus triatomae + C.albicans.
Среди двухкомпонентных преобладали ассоциации, состоящие из двух видов
НТМ: M.abscessus + M.chaelonae (n=1), M.avium + M.malmoense (n=3), M.fortuitum
+ M.farcinogenes (n=1), M.fortuitum + M.peregrinum (n=2), M.septicum +
M.peregrinum (n=1), M.gordonae + M.avium (n=1), M.kansasii + M.avium (n=1),
M.monacense + M.flavescens (n=1), M.malmoense + M.kansasii (n=1), M.gordonae +
M.chimera/intracellulare (n=1). Общее количество ассоциаций, состоящих из
микобактерий было выявлено в 13-и пробах (4,1% от всех проб). Преобладали

90
ассоциации, в состав которых входили M.malmoense и M.avium в различных
сочетаниях.
На втором месте по частоте встречаемости были ассоциации, состоящие из
НТМ и микроорганизма из другой группы: M.avium + K.pneumoniae (n=1), M.avium
+ C.albicans (n=1), M.avium + E.faecalis (n=1), M.szulgai + Staphylococcus warnerii
(n=1), M.gordonae + Streptococcus parasanguinis (n=2), M.gordonae + Aspergillus
sydowii (n=1), M.gordonae + S.epidermidis (n=1), M.gordonae + Kocuria marina (n=1),
M.gordonae + P.chrysogenum (n=1), M.fortuitum + S.phaeochromogenes (n=1),
M.gordonae + Staphylococcus lugdunensis (n=1). Общее количество таких
ассоциаций было выявлено в 12-и пробах (3,8% от общего количества проб). В
отличие от первой группы ассоциаций, в межгрупповых комбинациях чаще
выделялась M.gordonae, которая, по литературным данным, не относится к
патогенным для человека видам.
Выявление ассоциаций НТМ между собой и с другими микроорганизмами
поднимает еще два вопроса: насколько возможно выделение таких ассоциаций без
дополнительного разделения культур с использованием агаризованных сред и
имеют ли комбинации, содержащие в своем составе НТМ, клиническое значение?
Целью работы не являлось решение данных вопросов, но сам факт выделения
комбинаций, содержащих НТМ, на наш взгляд, требует в дальнейшем тщательного
изучения.
В ходе исследования была выделена одна комбинация, не содержащая
микроорганизмов из группы КУППА – Clostridium spp. + S.lugdunensis.
Для одного образца не удалось получить приемлемый результат
идентификации при использовании MALDI-ToF масс-спектрометрии и роста
культуры при пересевах на агаризованные среды. Для четырех штаммов была
определена только родовая принадлежность.
Преобладающей группой микроорганизмов, идентифицированных методом
MALDI-ToF масс-спектрометрии оказались микобактерии, среди которых
определены 283 штамма относящиеся к 27 видам (82,0% от всех
идентифицированных культур). Два штамма были идентифицированы как M.bovis,
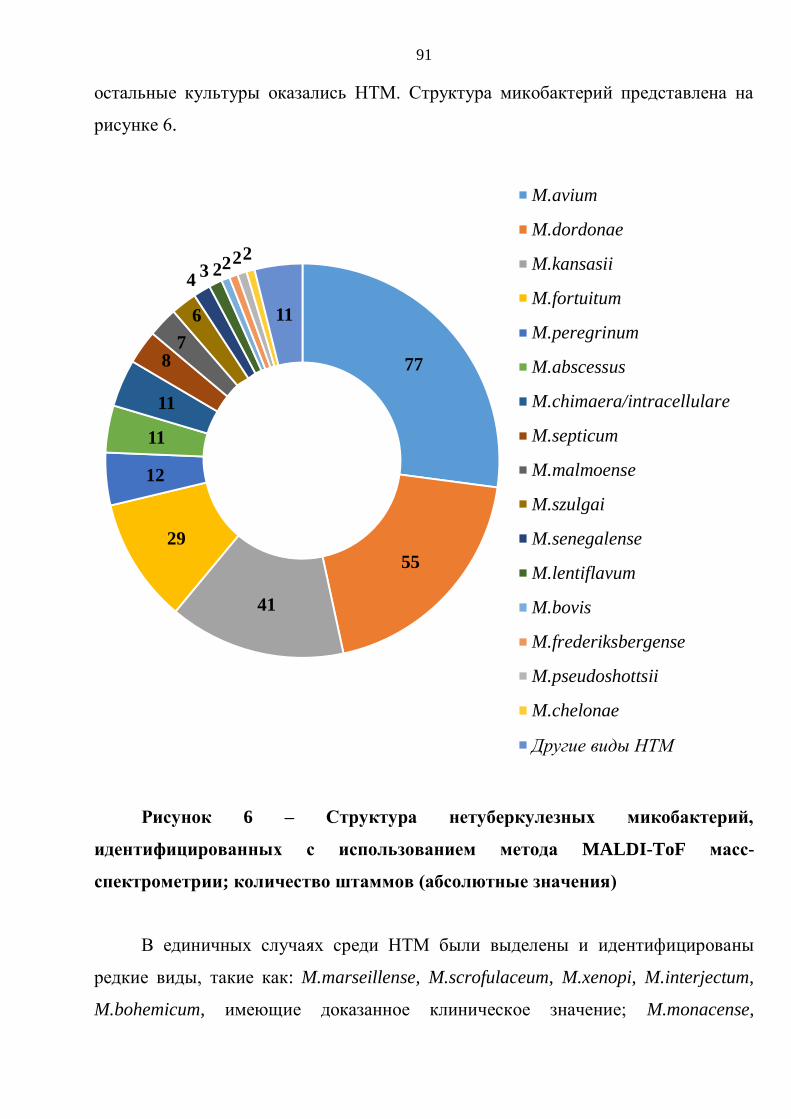
91
остальные культуры оказались НТМ. Структура микобактерий представлена на
рисунке 6.
Рисунок 6 – Структура нетуберкулезных микобактерий,
идентифицированных с использованием метода MALDI-ToF масс-
спектрометрии; количество штаммов (абсолютные значения)
В единичных случаях среди НТМ были выделены и идентифицированы
редкие виды, такие как: M.marseillense, M.scrofulaceum, M.xenopi, M.interjectum,
M.bohemicum, имеющие доказанное клиническое значение; M.monacense,
77
55
41
29
12
11
11
87
6
43 2222
11
M.avium
M.dordonae
M.kansasii
M.fortuitum
M.peregrinum
M.abscessus
M.chimaera/intracellulare
M.septicum
M.malmoense
M.szulgai
M.senegalense
M.lentiflavum
M.bovis
M.frederiksbergense
M.pseudoshottsii
M.chelonae
Другие виды НТМ

92
M.farcinogenes, M.iranicum, M.seoulense – виды НТМ, для которых описаны
единичные случаи выделения из материала от пациентов; M.gastri, M.flavescens,
являющиеся непатогенными для человека видами.
При оценке распределения выделенных микобактерий по клиническому
значению непатогенными для человека оказались 20,9% НТМ. Преобладающими
оказались виды, имеющие доказанное клиническое значение – 74,9% от общего
количества НТМ, редкие виды были идентифицированы в 4,2% случаев.
На втором месте по количеству идентифицированных микроорганизмов
оказались другие представители семейств КУППА: Nocardiaceae, Gordoniaceae,
Tsukamurellaceae, Brevibacteriaceae, Streptomycetaceae. Общее количество
штаммов – 20, что составило 5,8% от общего числа выделенных культур. Структура
кислотоустойчивых актиномицет за исключением микобактерий представлена в
таблице 12.
Таблица 12 – Кислотоустойчивые аэробные актиномицеты,
идентифицированные с использованием метода MALDI-ToF масс-
спектрометрии (n=20)
Выделенный микроорганизм
Количество
штаммов, шт.
Доля от всех
кислотоустойчивых
актиномицет, %
Brevibacillus celere 1 5,0%
Gordonia rubripertincta 4 20,0%
Gordonia sputi 3 15,0%
Nocardia brevicatena 1 5,0%
Nocardia farcinica 3 15,0%
Rhodococcus equi 1 5,0%
Rhodococcus triatomae 1 5,0%
Streptomyces hirsutus 1 5,0%
Streptomyces lavendulae 1 5,0%
Streptomyces phaeochromogenes 1 5,0%
Tsukamurella paurometabola 2 10,0%
Tsukamurella spp. 1 5,0%

93
Следует отметить, что для 75,0% идентифицированных микроорганизмов из
группы КУППА описано клиническое значение.
Остальные 42-е выделенные и идентифицированные культуры относятся к
классической условно-патогенной микрофлоре. Преобладающими группами
микроорганизмов оказались спорообразующие палочки и грибы.
Спорообразующие бактерии преимущественно относились к порядку Bacillales.
Были выделены и идентифицированы следующие виды: три штамма Bacillus cereus
и по одному штамму Bacillus oleronius, Bacillus mojavensis, Paenibacillus lactis,
Paenibacillus glucanolyticus, Paenibacillus larvae. Также был выделен один штамм
Clostridium spp. Всего было идентифицировано 15 штаммов спорообразующих
микроорганизмов, что составило 4,3% от всех выделенных культур.
В исследовании были выделены как дрожжеподобные, так и мицелиальные
виды. Структура микромицет, выделенных в исследовании представлена в таблице
13.
Таблица 13 – Микроскопические грибы, идентифицированные с
использованием метода MALDI-ToF масс-спектрометрии (n=11)
Выделенный микроорганизм Количество
штаммов, шт.
Доля от всех
микромицет, %
Aspergillus sydowii 1 9,1%
Aspergillus spp. 1 9,1%
Penicillium chrysogenum 1 9,1%
Magnusiomyces capitatus 1 9,1%
Rhodotorula mucilaginosa 1 9,1%
Ulocladium spp. 1 9,1%
Trichosporon asahii 1 9,1%
Candida tropicalis 1 9,1%
Candida albicans 3 27,2%
Среди микроскопических грибов также были выделены виды, имеющие
потенциальное клиническое значение, что требует рассмотрения возможности их
выделения и идентификации при работе с материалом от пациентов
противотуберкулезной службы.

94
Остальные микроорганизмы за исключением двух штаммов K.pneumoniae
относятся к грамположительной сапрофитной микрофлоре. Было
идентифицировано по пять штаммов Staphylococcus spp. и Corynebacterium spp.; по
два штамма S.parasanguinis, K.marina, E.faecalis и один штамм A.polychromogenes.
4.3. Сравнительный анализ результатов идентификации выделенных
микроорганизмов используемыми методами
Полученные результаты идентификации выделенных микроорганизмов
используемыми методами показали значительные различия в видовом составе
НТМ. Для определения причин расхождения результатов после идентификации
выделенных микроорганизмов методом ДНК-гибридизации все культуры были
разделены на четыре группы: первично идентифицированные до вида НТМ,
идентифицированные до рода микобактерии, микроорганизмы с высоким
содержанием ГЦ и неидентифицированные культуры.
При сравнении результатов идентификации методами ДНК-гибридизации и
MALDI-ToF масс-спектрометрии для штаммов, идентифицированных до вида
НТМ были получены следующие результаты. Для 138 штаммов,
идентифицированных методом ДНК-гибридизации, получены аналогичные
результаты при реидентификации, что составило 73,4% от количества
идентифицированных НТМ. Для 8 (4,3%) культур выявлено частичное совпадение
результата идентификации, при этом во всех образцах методом MALDI-ToF масс-
спектрометрии было идентифицировано два различных вида НТМ, один из
которых совпадал с результатом идентификации методом ДНК-гибридизации. В
22,3% (42 культуры) случаев получены расхождения в результате идентификации.
Девять штаммов были идентифицированы как микроорганизмы, не относящиеся к
роду Mycobacterium, что составило 21,4% от штаммов, для которых были получены
различные результаты идентификации и 4,8% от общего числа штаммов,
идентифицированных методом ДНК-гибридизации как НТМ. В остальных случаях

95
методом MALDI-ToF масс-спектрометрии был идентифицирован другой вид
микобактерии. Расхождения результатов идентификации по видам НТМ
представлены в таблице 14.
Таблица 14 – Расхождения результатов идентификации НТМ с
использованием анализируемых методов (количество штаммов)
ДНК-гибридизация n MALDI-ToF масс-спектрометрия n
Идентифицированная культура
M.abscessus 1 Tsukamurella spp. 1
M.avium 8
M.chimaera-intracellulare 1
Gordonia rubripertincta 1
M.kansasii 1
M.malmoense 2
M.seoulense 1
Streptomyces hirsutus 1
Gordonia sputi 1
M.celatum 1 M.chimaera-intracellulare 1
M.chimaera-
intracellulare 2
M.fortuitum 1
M.marseillense 1
M.fortuitum 21
Bacillus cereus 1
M.senegalense 4
Paenibacillus glucanolyticus 1
M.peregrinum 7
M.septicum+M.peregrinum 1
M.septicum 6
Bacillus cereus 1
M.genavense 1 M.pseudoshottsii+M.szulgai+Streptomyces
lavendulae 1
M.gordonae 4
M.chimaera-intracellulare 1
Clostridium spp.+Staphylococcus lugdunensis 1
M.szulgai+Staphylococcus warneri 1
Paenibacillus larvae 1
M.kansasii 2 M.bovis 1
M.gastri 1
M.malmoense 1 M.scrofulaceum 1
M.peregrinum 1 M.fortuitum 1
Максимальное количество расхождений выявлено для культур,
первоначально идентифицированных как M.fortuitum (51,2% от штаммов данного

96
вида). Для M.chimaera-intracellulare доля расхождений результатов идентификации
составила 33,3%, для M.avium – 12,1%, для M.gordonae – 11,1%, M.kansasii – 8,3%.
Значительные расхождения результатов идентификации для M.fortuitum
возможно связаны с тем, что 18 штаммов, определенных методом MALDI-ToF
масс-спектрометрии, в качестве представителей других видов входят в M.fortuitum
complex и являются близкородственными. Однако в трех случаях культура,
идентифицированная как M.fortuitum была определена как спорообразующая
палочка из порядка Bacillales.
Следует отдельно отметить факт реидентификации одного штамма
M.kansasii как представителя МТВс – M.bovis.
Полученные результаты расхождений идентификации также могут быть
обусловлены выделенными ассоциациями микроорганизмов.
Анализ идентификации второй группы микроорганизмов, которые были
отнесены к роду Mycobacterium методом ДНК-гибридизации показал следующие
результаты: при использовании MALDI-ToF масс-спектрометрии была определена
видовая принадлежность для всех выделенных микроорганизмов. За исключением
одного штамма, идентифицированного как Brevibacterium celere для всех культур,
была подтверждена принадлежность к микобактериям. Таким образом, из 21-го
штамма были идентифицированы по четыре штамма M.avium и M.kansasii, по три
штамма – M.lentiflavum и M.szulgai, два штамма M.fortuitum, по одному штамму –
M.xenopi, M.gordonae, M.peregrinum, M.interjectum, также был идентифицирован
один штамм НТМ, который не мог быть идентифицирован до вида с
использованием тест-системы Hain Lifescience GmbH (Германия) –
M.frederiksbergense.
В 39 образцах при проведении исследования методом ДНК-гибридизации
был получен результат о принадлежности выделенной культуры к группе
грамположительных микроорганизмов с высоким содержанием ГЦ. Всего было
выделено 42 штамма, в одном образце культуру выделить и идентифицировать не
удалось, в четырех случаях выделены двухкомпонентные ассоциации.

97
В данной группе также были идентифицированы НТМ: M.gordonae – пять
штаммов, M.avium и M.chimaera/intracellulare по три штамма, M.kansasii и
M.abscessus по два штамма. Остальные виды НТМ были идентифицированы по
одному штамму: M.fredericbergense, M.monacense, M.flavescens, M.szulgai,
M.malmoense, M.bohemicum, M.septicum.
Среди КУППА кроме НТМ были выделены: три штамма Nocardia farcinica,
по 1 штамму Gordonia rubripertincta, Gordonia sputi, Nocardia brevicatena,
Tsukamurella paurometabola.
Остальные микроорганизмы были представлены грамположительными
бактериями и грибами: четыре штамма C.albicans, два штамма K.marina, по одному
штамму Magnusiomyces capitatus, P.chrysogenum, A.polychromogenes, E.faecalis,
S.epidermidis, S.lugdunensis, C.amycolatum.
Значительное видовое разнообразие выявлено в группе из 67 образцов, для
которых не удалось провести приемлемую идентификацию. В данной группе
выделено и идентифицировано 68 культур.
Доминирующими в четвертой группе микроорганизмов оказались
микобактерии, которые составили 70,6% от всех выделенных микроорганизмов,
первичная идентификация которых с использованием метода ДНК-гибридизации
не дала результата. Среди НТМ были выделены: четырнадцать штаммов
M.gordonae, по одиннадцать штаммов M.avium и M.kansasii, шесть штаммов
M.fortuitum, по одному штамму M.abscessus, M.chimaera/intracellulare, M.iranicum,
M.pseudoshottsii, M.peregrinum. Также был выделен один представитель МТВс –
M.bovis.
Второй по численности группой первоначально не идентифицированных
микроорганизмов оказались грамположительные палочки – представители родов
Corynebacterium и Bacillus, которые составили 11,8%. Было идентифицировано три
штамма C.amycolatum, по одному штамму C.kroppenstedii, B.cereus, B.mojavensis,
B.oleronius, Paenibacillus lactis.

98
Также в данной группе микроорганизмов кроме НТМ оказались и другие
КУППА: три штамма Gordonia rubipertincta и по одному штамму T.paurometabola,
Rhodococcus equi, S.phaeochromogenes.
Кроме бактерий в группе не идентифицированных микроорганизмов
оказались четыре штамма грибов, два из которых не удалось идентифицировать с
помощью масс-спектрометрии до вида: Rhodotorula mucilaginosa, Aspergillus spp.,
Ulocladium spp., Trichosporon asahii. Единственная микробная ассоциация,
выявленная в данной группе микроорганизмов представлена M.fortuitum и
S.phaeochromogenes.
4.4. Частота выделения и структура кислотоустойчивых актиномицет в
клиническом материале в зависимости от пола и возраста пациентов
Для оценки частоты выделения КУППА из клинического материала в
зависимости от пола и возраста пациентов были использованы результаты
идентификации методом MALDI-ToF масс-спектрометрии.
Наиболее распространенные виды микобактерий были проанализированы
более детально. Для сравнения всех выборок использовались непараметрические
тесты ввиду малого количества данных и отличия распределения возраста от
нормального.
Всего в исследовании был получен клинический материал с признаками
роста НТМ от 246 пациентов. Для оценки частоты выделения из клинического
материала M.abscessus, M.avium, M.fortuitum, M.gordonae и M.kansasii были
построены графики box-plot, представленные на рисунке 7.

99
Рисунок 7 – Частота выделения из клинического материала основных
видов НТМ в зависимости от возраста пациентов
При анализе полученных данных было выявлено, что отдельные виды
микроорганизмов в зависимости от возраста пациента имеют среднее, не
отличающееся статистически от остальных видов. Это подтверждает и тест
Краскела-Уоллиса со статистикой H=4,174 и уровнем значимости равной 0,382.
Хотя анализ сравнения всех микроорганизмов и показал отсутствие
значимого различия в средних, по графику можно увидеть, что попарное сравнение
может уточнить этот результат. Действительно, если провести попарное сравнение
различия в средних с помощью теста Манна-Уитни, то можно заметить, что
средние значения возраста M.abscessus и M.gordonae будут отличаться значимо со
значением статистики U=70,5 и достигаемым уровнем значимости, равной 0,04.
Таким образом, M.gordonae достоверно чаще выделяется среди у пациентов со
средним возрастом 50 лет, а M.abscessus среди пациентов со средним возрастом
41,5 года.
Анализ данных по распространенности микроорганизмов основных групп по
частоте встречаемости с учетом пола и возраста продемонстрировал больше
статистически значимых различий, представленных на рисунке 8.

100
Рисунок 8 – Частота выделения из клинического материала основных
видов НТМ в зависимости от возраста и пола пациентов (m-мужчины, f-
женщины)
Попарные тесты на равенство медиан возраста для мужчин и женщин
выявили значимые различия в средних возрастах для некоторых групп с учетом
вида выделенных микобактерий. Данные представлены в таблицах 15 и 16.
Таблица 15 – Результаты попарных тестов на равенство медиан возраста
для мужчин с учетом вида выделенных микобактерий
1 микроорганизм 2 микроорганизм Значение U Значение р-value
M.abscessus M.avium 97,5 0,076
M.abscessus M.fortuitum 39,0 0,062
M.abscessus M.gordonae 39,0 0,047*
M.abscessus M.kansasii 52,0 0,076
M.avium M.fortuitum 486,5 0,088
M.avium M.gordonae 487,5 0,039*
M.avium M.kansasii 682,0 0,241
M.fortuitum M.gordonae 240,0 0,374
M.fortuitum M.kansasii 256,0 0,212
M.gordonae M.kansasii 282,5 0,274
*различия статистически значимы при значении р < 0,05

101
M.gordonae статистически достоверно чаще выделяются из клинического
материала пациентов мужского пола старших возрастных групп, чем M.abscessus и
M.avium.
Таблица 16 – Результаты попарных тестов на равенство медиан возраста
для женщин с учетом вида выделенных микобактерий
1 микроорганизм 2 микроорганизм Значение U Значение р-value
M.abscessus M.avium 80,0 0,319
M.abscessus M.fortuitum 34,5 0,32
M.abscessus M.gordonae 31,5 0,208
M.abscessus M.kansasii 47,5 0,404
M.avium M.fortuitum 233,5 0,112
M.avium M.gordonae 289,5 0,316
M.avium M.kansasii 255,5 0,029*
M.fortuitum M.gordonae 141,0 0,245
M.fortuitum M.kansasii 192,0 0,5
M.gordonae M.kansasii 151,0 0,077
*различия статистически значимы при значении р < 0,05
M.kansasii статистически достоверно чаще выделяются из клинического
материала у пациентов женского пола более молодого возраста чем M.avium.
В целом, при расчете теста Краскела-Уоллиса для мужчин, по частоте
выделения отдельных видов микобактерий был получен результат Н=6,026, при
уровне достигаемом уровне значимости р=0,197, а для женщин – Н=6,07 при
достигаемом уровне значимости р=0,194. Данные анализа параметров возраста и
пола в целом не показали статистически достоверных различий, несмотря на
полученные различия в парных сравнениях.
В связи с тем, что часть микроорганизмов была выделена в небольших
количествах было принято решение дополнительно укрупнить анализируемые
группы. Отдельно были проанализированы по возрасту и полу частота выделения
из клинического материала быстрорастущих, медленнорастущих НТМ и других
микроорганизмов группы КУППА; частота выделения в материале микобактерий
в целом и других представителей группы КУППА; а также частота выделения в

102
материале микроорганизмов по их потенциальному клиническому значению (с
учетом данных литературы).
Анализ влияния возраста и пола пациентов на частоту выделения быстро и
медленнорастущих НТМ, и других микроорганизмов из группы КУППА
представлен на рисунках 9 и 10.
Рисунок 9 – Частота выделения из клинического материала
быстрорастущих (RG), медленнорастущих (SG) микобактерий и других
КУППА (NonMB) в зависимости от возраста пациентов
Рисунок 10 – Частота выделения из клинического материала
быстрорастущих (RG), медленнорастущих (SG) микобактерий и других
КУППА (NonMB) в зависимости от возраста и пола пациентов (m-мужчины,
f-женщины)

103
Различия по возрасту и полу в укрупненных группах быстро и
медленнорастущих НТМ, и других КУППА оказались статистически не
значимыми. Результаты теста Краскела-Уоллиса для всей выборки: Н=0,293, р-
value=0,864; для мужчин: Н=0,565, р-value=0,754; для женщин: Н=0,820,
р-value=0,664).
Анализ влияния возраста и пола пациентов на частоту выделения НТМ и
других микроорганизмов из группы КУППА в целом представлен на рисунках 11 и
12.
Рисунок 11 – Частота выделения из клинического материала
нетуберкулезных микобактерий в (yes) целом и других КУППА (no) в
зависимости от возраста пациентов
Рисунок 12 – Частота выделения из клинического материала
нетуберкулезных микобактерий в целом (yes) и других КУППА (no) в
зависимости от возраста и пола пациентов (m-мужчины, f-женщины)

104
Тест Манна-Уитни для всей выборки показал следующие результаты:
U=1038,5, p-value=0,248, что означает незначимость различий результатов по
половому признаку пациентов в среднем возрасте, в клиническом материале
которых были выделены микобактерии и другие КУППА. Для мужчин U=468,5, р-
value=0,185; для женщин U=106,5, p-value=0,493, что так же не показывает
значительной разницы полученных результатов.
Анализ влияния возраста и пола пациентов на частоту выделения клинически
значимых и не имеющих клинического КУППА, по литературным данным,
представлен на рисунках 13 и 14.
Рисунок 13 – Частота выделения из клинического материала КУППА в
зависимости от их клинического значения
Рисунок 14 – Частота выделения из клинического материала КУППА в
зависимости от их клинического значения (m-мужчины, f-женщины)

105
Критерий Манна-Уитни для двух выборок дал следующие результаты:
U=1319,5, p-value=0,364, что указывает на незначительные различия по половому
признаку в средних значениях по возрасту. Критерий Манна-Уитни отдельно для
каждого пола дал следующие результаты: для мужчин: U=661,0, p-value=0,422; для
женщин: U=107,0, p-value=0,367. Оба результата означают незначимость различия
в средних. Подробная информация по количеству выделенных НТМ и возрастных
данных пациентов представлены в таблице 17.
Таблица 17 – Результаты описательной статистики для отдельных видов
НТМ и возраста пациентов, в клиническом материале от которых они были
выделены
Вид микобактерии n Mean Std Min 25% 50% 75% Max
M.abscessus 8 41,9 12,8 20,0 34,75 41,5 50,25 60,0
M.avium 64 50,8 15,8 14,0 39,00 46,0 62,50 82,0
M.bohemicum 1 81,0 - 81,0 81,00 81,0 81,00 81,0
M.celatum 1 45,0 - 45,0 45,00 45,0 45,00 45,0
M.chelonae 1 59,0 - 59,0 59,00 59,0 59,00 59,0
M.chimaera/intracelullare 11 52,6 17,0 32,0 39,50 43,0 64,50 80,0
M.flavescens 1 61,0 - 61,0 61,00 61,0 61,00 61,0
M.fortuitum 28 51,5 16,5 22,0 41,00 48,5 65,00 85,0
M.frederiksbergense 2 45,0 14,1 35,0 40,00 45,0 50,00 55,0
M.gastri 1 45,0 - 45,0 45,00 45,0 45,00 45,0
M.gordonae 30 54,8 19,6 14,0 39,50 54,0 68,00 84,0
M.interjectum 1 60,0 - 60,0 60,00 60,0 60,00 60,0
M.iranicum 1 46,0 - 46,0 46,00 46,0 46,00 46,0
M.kansasii 35 49,5 13,8 27,0 38,50 47,0 59,00 82,0
M.lentiflavum 3 44,7 13,6 34,0 37,00 40,0 50,00 60,0
M.malmoense 6 54,0 16,3 39,0 40,50 50,5 63,5 79,0
M.marseillense 1 46,0 - 46,0 46,00 46,0 46,00 46,0
M.monacense 1 61,0 - 61,0 61,00 61,0 61,00 61,0
M.peregrinum 12 49,4 16,5 28,0 32,50 50,5 63,50 69,0
M.pseudoshottsii 2 24,0 28,3 4,0 14,00 24,0 34,00 44,0
M.scrofulaceum 1 59,0 - 59,0 59,00 59,0 59,00 59,0
M.senegalense 4 61,5 16,0 50,0 50,00 56,0 67,50 84,0
M.seoulense 1 42,0 - 42,0 42,00 42,0 42,00 42,0
M.septicum 8 50,8 17,1 32,0 37,50 49,0 61,00 80,0
M.szulgai 4 42,8 26,8 5,0 35,75 49,0 56,00 68,0
M.xenopi 1 51,0 - 51,0 51,00 51,0 51,00 51,0

106
n – количество пациентов; Mean – среднее значение возраста;
Std – стандартное отклонение возраста, для микроорганизмов, которые были
выделены у одного пациента стандартное отклонение не может быть рассчитано;
Min, Max – минимальный и максимальный возраст соответственно;
25%, 75% – 25 квантиль и 75 квантиль возраста соответственно;
50% – медианное значение возраста
Подробная информация по количеству выделенных микроорганизмов, за
исключением НТМ и возрастных данных пациентов представлены в таблице 18.
Таблица 18 – Результаты описательной статистики для других
клинически значимых КУППА и возраста пациентов, в клиническом
материале которых они были выделены
Микроорганизм n Mean Std Min 25% 50% 75% Max
Gordonia spp. 8 51,9 13,9 40,0 41,75 46,0 60,25 79,0
Nocardia spp. 6 52,2 8,2 43,0 45,50 53,0 56,00 64,0
Tsukamurella spp. 3 51,0 13,9 43,0 43,00 43,0 55,0 67,0
n - количество пациентов; Mean – среднее значение возраста;
Std – стандартное отклонение возраста;
Min, Max – минимальный и максимальный возраст соответственно;
25%, 75% - 25 квантиль и 75 квантиль возраста соответственно;
50% - медианное значение возраста
Минимальный возраст пациентов, в клиническом материале от которых были
выделены НТМ – 4 года, максимальный – 82 года. Для других КУППА
минимальный возраст значительно больше – 43 года, максимальный возраст – 79
лет. Средний возраст для пациентов в клиническом материале которых были
выделены представители основных групп НТМ варьирует в пределах 41,9-54,8 лет.
4.5. Резюме
Анализируя полученные результаты можно выявить определенные
закономерности, связанные с условным ограничением использования метода ДНК-
гибридизации в отношении некоторых видов НТМ. Данным методом было

107
идентифицировано порядка 60% штаммов микроорганизмов. Преобладающими
оказались медленно растущие НТМ, среди которых лидирующее положение
занимали представители M.avium complex, а также M.gordonae и M.kansasii. Среди
быстро растущих НТМ преобладали M.fortuitum.
Безусловно, необходимо учитывать, что используемая в исследовании тест-
система исключает идентификацию микобактерий, виды которых в нее не
включены, однако по нашим результатам при ее использовании не удалось
получить видовую идентификацию и для тех видов, которые должны были быть
идентифицированы. Из 22-и образцов, в которых микобактерии были
идентифицированы до рода, в 20-и с использованием метода масс-спектрометрии
были определены виды, которые входят в перечень тест-системы.
Среди культур, для которых методом ДНК-гибридизации не удалось
провести приемлемую идентификацию, оказались широко как распространенные
виды НТМ: M.gordonae, M.avium, M.kansasii, так и редкие виды – M.monacense,
M.farcinogenes, M.iranicum, M.seoulense. Также немаловажным является, то, что
среди данной группы микроорганизмов были выделены другие кислотоустойчивые
аэробные актиномицеты, в том числе имеющие потенциальное клиническое
значение: Gordonia rubipertincta, Tsukamurella paurometabola, Rhodococcus equi.
Полученные результаты могут быть обусловлены несколькими причинами:
значительной вариабельностью внутри и между отдельными видами НТМ, что
может приводить к неточной идентификации при использовании метода ДНК-
гибридизации; определенным ограничением возможности разделения микст-
культур и микробных ассоциаций в условиях микробиологических лабораторий
противотуберкулезной службы; отсутствием возможности выделения
«прихотливых», трудно культивируемых и других микроорганизмов, которые
могли затруднять идентификацию в рамках проведенного исследования.
Наиболее интересными оказались результаты, полученные при
идентификации культур, содержащих высокие концентрации ГЦ. Именно в эту
группу попали виды НТМ, выделенные в единичных случаях: M.fredericbergense,

108
M.szulgai, M.malmoense, M.bohemicum, M.septicum, а также представители родов
Nocardia, Gordonia, Tsukamurella.
Таким образом, использование методов, основанных на ДНК-гибридизации,
с одной стороны, позволяет достаточно точно идентифицировать часть наиболее
часто встречающихся видов НТМ, с другой стороны, имеет значительные
ограничения. Отсутствие результатов по точной идентификации части видов,
которые включены в тест-систему требует проведения дополнительных
исследований и поиска причин полученных результатов идентификации. MALDI-
ToF масс-спектрометрия является важным методом, позволяющим проводить
определение до вида большинства КУППА. Однако необходимы алгоритмы,
позволяющие стандартизировать методы их выделения из клинического материала,
а также рекомендации по интерпретации полученных результатов и оценке
клинического значения выделенных культур.
Анализ распространенности отдельных групп КУППА показал практически
полное отсутствие статистической достоверности по видам и укрупненным
кластерам микроорганизмов в зависимости от таксономического положения,
классификационных признаков и клинического значения, пола и возраста
пациентов, что необходимо учитывать при работе с материалом, в котором
выявлены признаки роста нетуберкулезных микобактерий и требует разработки
алгоритмов точной видовой идентификации, с учетом факта выделения
значительного числа (более 20,0%) микобактерий и других КУППА с не
доказанным клиническим значением.

109
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ,
ИЗЛОЖЕННЫМ В ГЛАВЕ 4
Статьи в журналах, рекомендованных ВАК:
1. Лямин, А.В. Микобактериозы: особенности эпидемиологии и
лабораторной диагностики / А.В. Лямин, Д.Д. Исматуллин // Аспирантский вестник
Поволжья. – 2016. – № 5-6. – С. 204-208.
2. Лямин, А.В. Нетуберкулезные микобактерии: современные возможности
видовой идентификации / А.В. Лямин, А.М Ковалёв, А.В. Жестков [и др.] //
Клиническая микробиология и антимикробная химиотерапия. – 2017. – Т. 19. – №1.
– С. 11-14.
3. Лямин, А.В. Лабораторная диагностика микобактериозов / А.В. Лямин,
А.В. Жестков, Д.Д. Исматуллин [и др.] // Вестник современной клинической
медицины. – 2017. – Т. 10. – № 1. – С. 29-35.
4. Лямин, А.В. Сравнительный анализ методов идентификации
нетуберкулезных микобактерий, выделенных из клинического материала / А.В.
Лямин, Д.Д. Исматуллин, А.В. Жестков [и др.] // Инфекция и иммунитет. – 2017. –
Т. 7. – №3. – С. 285-291.
5. Лямин, А.В. Общая характеристика и клиническое значение
представителей родов Nocardia и Gordonia / А.В. Лямин, А.В. Жестков, Т.Р.
Никитина [и др.] // Инфекция и иммунитет. – 2019. – Т. 9. – № 3-4. – С. 429-436.
6. Лямин, А.В. Редкие виды в структуре кислотоустойчивых представителей
порядка Actinomycetales, выделенных из клинического материала / А.В. Лямин //
Клиническая лабораторная диагностика. – 2020. – Т. 65. – №2. – С. 111-115.
7. Калимулина, К.Р. Представители Mycobacterium abscessus complex у
пациентов с бронхолегочной патологией: распространенность, особенности
культивирования и идентификации / К.Р. Калимулина, Д.Д. Исматуллин, А.В.
Лямин [и др.] // Клиническая лабораторная диагностика. – 2020. – Т. 65. – №5. – С.
316-320.

110
Публикации в других изданиях:
1. Лямин, А.В. Структура нетуберкулезных микобактерий, выделенных из
клинического материала в Самарской области / А.В. Лямин, Исматуллин Д.Д., А.В.
Жестков [и др.] // Клиническая микробиология и антимикробная химиотерапия. –
2017. – Т. 19. – Приложение 1. – Тезисы XIX Международного конгресса МАКМАХ
по антимикробной химиотерапии и клинической микробиологии. – М. – С. 27-28.
2. Исматуллин, Д.Д. Идентификация нетуберкулезных микобактерий,
выделенных из клинического материала методом MALDI-ToF масс-спектрометрии
/ Д.Д. Исматуллин, А.В. Лямин, А.В. Жестков [и др.] // Проблемы медицинской
микологии. – 2017. – Т.19. - №2. – Тезисы докладов Всероссийской научно-
практической конференции по медицинской микробиологии, эпидемиологии и
клинической микологии (XX Кашкинские чтения). – СПб. – С. 71.
3. Kontsevaya, I. Non-tuberculous Mycobacteria isolated from Russian
tuberculosis patients over a one year period / I. Kontsevaya, A. Kovalyov, T. Simak [at
al.] // Abstracts of 38th Annual Congress of the European Society of Mycobacteriology. –
2017. – Sibenik. – P. 79.
4. Kovalyov, A. The identification of non-tuberculous mycobacteria species in
Russian tuberculosis patients using MALDI-ToF mass spectrometry // A. Kovalyov, A.
Lyamin, D. Ismatullin [at al.] // Abstracts of 38th Annual Congress of the European
Society of Mycobacteriology. – 2017. – Sibenik. – P. 88.
5. Исматуллин, Д.Д. Новые возбудители заболеваний респираторного тракта
у иммунокопрометированных пациентов (обзор литературы) / Д.Д, Исматуллин,
М.О. Золотов, А.В. Лямин [и др.] // Наука и инновации в медицине. – 2019. – Т. 4.
– № 3. – С. 19-24.
Учебное пособие:
1. Лямин А.В., Жестков А.В., Исматуллин Д.Д. Лабораторная диагностика
микобактериозов: учебное пособие для студентов, Самара: ООО «Издательско-
полиграфический комплекс «Право», 2019. – 72 с. ISBN 978-5-6041835-8-8

111
ГЛАВА 5. ОЦЕНКА ВОЗМОЖНОСТИ КУЛЬТИВИРОВАНИЯ
ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА ACTINOMYCETALES НА
АГАРИЗОВАННЫХ СРЕДАХ
Принципы, по которым НТМ были разделены на четыре группы до
сегодняшнего дня остаются широко распространенными в практической
микробиологии, несмотря на внедрение новых принципов классификации данной
группы микроорганизмов. Для оценки возможности использования культуральных
свойств изолятов НТМ, выделенных из клинического материала, нами был
проведен ряд исследований с целью оценки культуральных свойств микобактерий
и других КУППА и их сопоставление с видовой принадлежностью. Принцип
разделения НТМ на фотохромогенные, скотохромогенные, нефотохромогенные и
быстрорастущие виды был использован как основополагающий при описании
полученных результатов. Оценку культуральных свойств проводили на плотной
яичной среде Левенштейна-Йенсена и жидкой среде Миддлбрука 7Н9, а также
агаризованных средах: кровяном агаре и универсальной хромогенной среде.
5.1. Оценка культуральных свойств анализируемых микроорганизмов при
посеве на плотные и жидкие питательные среды для культивирования
микобактерий
Оценку культуральных свойств КУППА, представленную в данной главе
проводили у НТМ, а также других кислотоустойчивых актиномицет, описанных в
Главах 3 и 4, идентификация которых была проведена с использованием метода
MALDI-ToF масс-спектрометрии. Общее количество микроорганизмов,
включенных в исследование составило 402 штамма. Структура КУППА по родам
представлена на рисунке 15.

112
Рисунок 15 – Структура КУППА, идентифицированных с
использованием метода MALDI-ToF масс-спектрометрии; количество
штаммов; абсолютные значения и значения в процентах от общего
количества штаммов
На среде Левенштейна-Йенсена при пересеве удалось получить рост всех
штаммов микроорганизмов.
Медленно растущие пигментообразующие культуры были выделены в 85-и
случаях, что составило 21,1% от всех КУППА. В данной группе были
идентифицированы 41 штамм M.kansasii и 2 штамма M.pseudoshottsii –
классических представителей фотохромогенных НТМ. Дополнительно было
отмечено, что 4 штамма M.kansasii имели менее яркий желтый цвет, чем остальные
культуры.
319
79%
39
10%
14
3%
10
2%
8
2%
6
2%
3
1%
3
1% Mycobacterium spp.
Streptomyces spp.
Nocardia spp.
Gordonia spp.
Brevibacterium spp.
Tsukamurella spp.
Rhodococcus spp.
Cellulosimicrobium
spp.

113
В эту же группу вошли 42 штамма (49,4% от всех фотохромогенных НТМ),
идентифицированные как M.avium. Классически M.avium относится к
нефотохромогенным НТМ, которые не образуют пигмент, или колонии которых
имеют слабовыраженную окраску [119]. Однако для всех 42-х штаммов M.avium,
которые были отнесены к фотохромогенным НТМ, была отмечена четкая желтая
пигментация колоний различной степени выраженности.
Скотохромогенные медленнорастущие НТМ были выделены в 74 случаях
(18,4% от всех КУППА). Большинство культур имели ярко-желтый, в некоторых
случаях желто-оранжевый цвет, характерный для M.gordonae. Среди выделенных
культур были идентифицированы 60 штаммов M.gordonae, 6 штаммов M.szulgai, 3
штамма M.lentiflavum и по 1 штамму M.scrofulaceum, M.xenopi, M.bochemicum,
M.interjectum и M.seoulense. У штаммов M.szulgai, M.lentiflavum, M.scrofulaceum,
M.xenopi и M.interjectum цвет колоний был менее насыщенный – желтый или
светло-желтый.
Нефотохромогенные медленнорастущие микроорганизмы были выделены в
68-и случаях, что составило 16,9% от числа всех анализируемых культур. Среди
данной группы было идентифицировано 43 штамма M.avium (63,2% от всех
нефотохромогенных НТМ) с классическим вариантом колоний, 11 штаммов
M.chimaera/intracellulare, 8 штаммов M.malmoense, 2 штамма M.marseillense и по 1
штамму M.celatum и M.gastri. Колонии всех культур имели светло-кремовый или
светло-бежевый цвет. Все штаммы M.malmoense дали нехарактерный для
микобактерий слизистый рост.
Самой разнообразной по культуральным свойствам оказалась группа
быстрорастущих НТМ. Все представители КУППА за исключением
вышеописанных видов НТМ, стрептомицет и представителей рода
Cellulosimicrobium дали два типа колоний – пигментированные и
непигментированные. Часть штаммов стрептомицет и Cellulosimicrobium spp.
вызвали значительное изменение питательной среды, в некоторых случаях до ее
разжижения или дали скудный рост, характерный для мицелиальных грибов –
мелкие колонии белого цвета, покрытые нитями воздушного мицелия. В связи с

114
этим культуральные свойства данных микроорганизмов не учитывались при
оценке внешнего вида колоний у быстрорастущих КУППА.
В итоге анализ культуральных свойств был проведен для 133-х штаммов
быстрорастущих микроорганизмов. Все культуры были разделены на не
образующие пигмент (цвет колоний светло-бежевый, светло-кремовый или светло-
желтый) и пигментообразующие, которые дали окрашенные колонии.
Общее количество культур с окрашенными колониями составило 22 штамма
(15,8% от всех быстрорастущих микроорганизмов). В данной группе были
идентифицированы: 10 штаммов Gordonia spp., 7 штаммов Nocardia spp., 2 штамма
M.frederiksbergense, по 1 штамму M.iranicum, M.monacense, M.flavescens. Гордонии
и нокардии дали оранжевые и желтые колонии соответственно. НТМ в данной
группе образовали колонии желто-оранжевого цвета, характерного для
M.gordonae.
Самой многочисленной группой оказались быстрорастущие виды, которые
не образовывали пигмент на среде Левенштейна-Йенсена (111 штаммов). В данной
группе были идентифицированы 35-ть штаммов M.fortuitum, среди которых 7
культур оказались нетипичной для микобактерий мукоидной формы, по 12
штаммов M.abscessus и M.peregrinum, 10 штаммов M.septicum, 8 штаммов
Brevibacterium spp., по 7 штаммов Nocardia spp., M.senegalense и M.chelonae, 6
штаммов Tsukamurella spp., 3 штамма Rhodococcus spp., 2 штамма M.porcinum и по
1 штамму M.chitae и M.farcinogenes, которая также дала колонии мукоидного типа.
Колонии всех штаммов M.abscessus, M.chelonae и M.peregrinum были бежевого
цвета, в некоторых случаях с признаками врастания в среду.
Культуральные свойства микроорганизмов КУППА, выросших в жидкой
питательной среде Миддлбрука были менее разнообразными. Все культуры дали
осадок. Окрашенный в желто-оранжевый цвет, слизистый осадок был выявлен для
всех штаммов M.gordonae. Светло-желтый среднедисперсный осадок – для
M.kansasii. Желтый однородный осадок был выявлен у 7-и штаммов нокардий,
который дали пигментированные колонии на плотной среде.

115
Для остальных культур был выявлен осадок белого цвета различной
консистенции без определенных закономерностей. Следует отметить, что для
медленнорастущих видов осадок был обильным, для быстрорастущих более
скудным.
5.2. Особенности культуральных свойств представителей порядка
Actinomycetales на агаризованных средах
5.2.1. Оценка возможности культивирования нетуберкулезных микобактерий
на универсальной хромогенной среде и кровяном агаре
В главе 1 были приведены данные о возможности культивирования НТМ на
различных агаризованных средах, как для первичного посева клинического
материала, так и для работы с чистыми культурами НТМ и другими
кислотоустойчивыми аэробными актиномицетами. Для оценки возможности
такого варианта культивирования был проведен ряд экспериментов.
Представители доминирующих групп НТМ были засеяны на 5% кровяной
агар с бараньей кровью и универсальную хромогенную среду UriSelect (BioRad,
США). Был использован инокулюм для посева с ориентировочным содержанием
клеток культуры 1000 КОЕ/1мл (100 КОЕ/0,1 мл). Посевы инкубировались при
температуре 37°С в течение сорока суток с ежедневной оценкой роста и фиксацией
срока появления видимых колоний с их последующим подсчетом и статистической
обработкой полученных результатов.
Следует учесть, что на агаризованных средах не удалось получить рост у 12-
и штаммов M.avium и трех штаммов M.gordonae. Количественная оценка
полученных результатов при культивировании на кровяном агаре и универсальной
хромогенной средой представлены в таблицах 19 и 20.
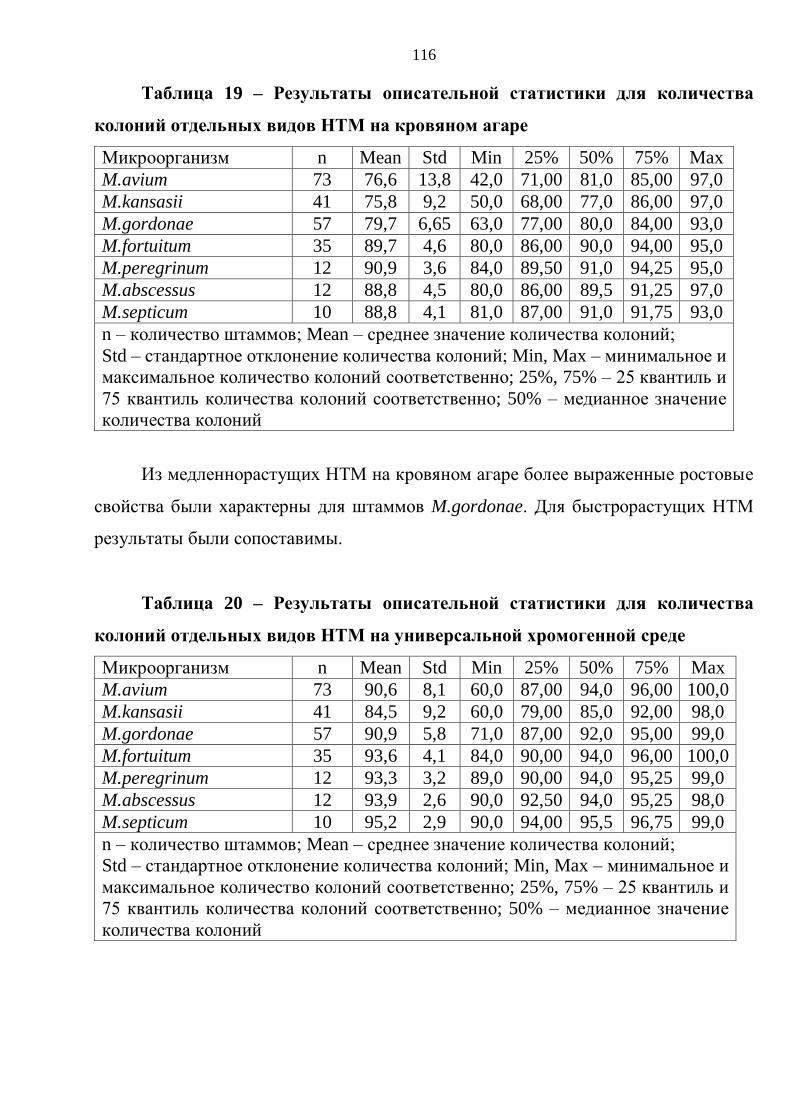
116
Таблица 19 – Результаты описательной статистики для количества
колоний отдельных видов НТМ на кровяном агаре
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 73 76,6 13,8 42,0 71,00 81,0 85,00 97,0
M.kansasii 41 75,8 9,2 50,0 68,00 77,0 86,00 97,0
M.gordonae 57 79,7 6,65 63,0 77,00 80,0 84,00 93,0
M.fortuitum 35 89,7 4,6 80,0 86,00 90,0 94,00 95,0
M.peregrinum 12 90,9 3,6 84,0 89,50 91,0 94,25 95,0
M.abscessus 12 88,8 4,5 80,0 86,00 89,5 91,25 97,0
M.septicum 10 88,8 4,1 81,0 87,00 91,0 91,75 93,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний
Из медленнорастущих НТМ на кровяном агаре более выраженные ростовые
свойства были характерны для штаммов M.gordonae. Для быстрорастущих НТМ
результаты были сопоставимы.
Таблица 20 – Результаты описательной статистики для количества
колоний отдельных видов НТМ на универсальной хромогенной среде
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 73 90,6 8,1 60,0 87,00 94,0 96,00 100,0
M.kansasii 41 84,5 9,2 60,0 79,00 85,0 92,00 98,0
M.gordonae 57 90,9 5,8 71,0 87,00 92,0 95,00 99,0
M.fortuitum 35 93,6 4,1 84,0 90,00 94,0 96,00 100,0
M.peregrinum 12 93,3 3,2 89,0 90,00 94,0 95,25 99,0
M.abscessus 12 93,9 2,6 90,0 92,50 94,0 95,25 98,0
M.septicum 10 95,2 2,9 90,0 94,00 95,5 96,75 99,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний

117
Из медленной растущих НТМ на универсальной хромогенной среде более
выраженные ростовые свойства были характерны для штаммов M.avium и
M.gordonae. Для быстрорастущих НТМ результаты также были сопоставимы.
Для оценки достоверности полученных различий между количеством
колоний на кровяном агаре и универсальной хромогенной среде был рассчитан
критерий Манна-Уитни, результаты которого представлены в таблице 21.
Таблица 21 – Результат сравнительной статистики количества колоний
при культивировании на анализируемых средах
Микроорганизм Критерий Манна-Уитни
Статистика p-value
M.avium 825,0 <0,001
M.kansasii 514,5 0,001
M.gordonae 307,0 <0,001
M.fortuitum 330,0 <0,001
M.peregrinum 49,0 0,090
M.abscessus 21,5 0,002
M.septicum 8,0 <0,001
При p-value <0,05 различия статистически достоверны
Для всех тестируемых видов НТМ, за исключением M.peregrinum количество
колоний на универсальной хромогенной среде достоверно больше, чем на
кровяном агаре.
Дополнительно был рассчитан критерий продуктивности для которого было
определено количество штаммов, которые дали на тестируемых средах более 50%
и более 70% колоний, сравнительная статистика которого приведена в таблице 22.

118
Таблица 22 – Оценка числа культур по критерию продуктивности при
культивировании на анализируемых средах, сравнение медианных значений
Микроорганизм
Продуктивность
на кровяном
агаре, %
Продуктивность
на универсальной
хромогенной
среде, % p-value
50-69 ≥70 50-69 ≥70
1 2 3 4 1&3 2&4
M.avium 56,0 83,0 64,5 94,0 0,09 <0,001
M.kansasii 59,0 83,5 62,0 85,0 0,30 0,109
M.gordonae 65,5 81,0 - 92,0 - 8,288
M.fortuitum - 90,0 - 94,0 - <0,001
M.peregrinum - 91,0 - 94,0 - 0,095
M.abscessus - 89,5 - 94,0 - 0,002
M.septicum - 91,0 - 95,5 - <0,001
При p-value <0,05 различия статистически достоверны
Анализируя полученные результаты можно сделать заключение о том, что
для штаммов M.avium, M.fortuitum, M.abscessus и M.septicum значения по критерию
продуктивности достоверно отличаются при значении ≥70%. Для остальных
культур и продуктивности в пределах 50-69% результаты значимо не отличаются.
Параллельно с анализом возможности культивирования НТМ на
универсальной хромогенной среде было проведено исследование, направленное на
выявление особенностей сроков появления видимого роста на агаризованных
средах. В качестве среды сравнения была использована плотная яичная среда
Левенштейна-Йенсена. Критерием появления роста считали обнаружение первой
колонии на поверхности агаризованной среды или яичной среды при температуре
культивирования 37°С. Для оценки достоверности использовали критерий
Краскела-Уоллиса. Полученные результаты приведены в таблицах 23-26.

119
Таблица 23 – Сроки выявления роста НТМ на кровяном агаре
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 73 19,3 5,6 6,0 17,00 20,0 23,00 28,0
M.kansasii 41 14,8 2,1 10,0 14,00 15,0 16,00 19,0
M.gordonae 57 14,6 3,1 6,0 13,00 15,0 17,00 19,0
M.fortuitum 35 2,1 2,0 1,0 1,00 1,0 2,00 7,0
M.peregrinum 12 1,9 1,3 1,0 1,00 1,5 2,00 5,0
M.abscessus 12 1,8 1,2 1,0 1,00 1,5 2,00 5,0
M.septicum 10 1,7 0,8 1,0 1,00 1,5 2,00 3,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний
Таблица 24 – Сроки выявления роста НТМ на универсальной
хромогенной средой
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 73 15,7 5,7 2,0 12,00 17,0 19,00 29,0
M.kansasii 41 12,3 2,3 8,0 11,00 12,0 14,00 18,0
M.gordonae 57 11,8 2,5 5,0 11,00 12,0 13,00 17,0
M.fortuitum 35 1,6 0,8 1,0 1,00 1,0 2,00 3,0
M.peregrinum 12 1,7 0,5 1,0 1,00 2,0 2,00 2,0
M.abscessus 12 1,6 0,5 1,0 2,00 2,0 2,00 2,0
M.septicum 10 1,3 0,5 1,0 1,00 1,0 1,75 2,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний
Максимальный диапазон появления роста на агаризованных средах был
выявлен для M.avium, который составил 22 суток на кровяном агаре и 27 суток на
универсальной хромогенной среде. Все штаммы быстрорастущих НТМ дали рост
в среднем в течение 2 суток культивирования.
При анализе роста культур на среде Левенштейна-Йенсена учитывался рост
для всех анализируемых штаммов.

120
Таблица 25 – Сроки выявления роста НТМ на среде Левенштейна-
Йенсена
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 85 17,3 7,5 2,0 12,00 17,0 20,00 37,0
M.kansasii 41 14,0 1,7 10,0 13,00 14,0 15,00 17,0
M.gordonae 60 16,1 2,7 10,0 14,75 16,0 18,00 22,0
M.fortuitum 35 2,3 2,1 1,0 1,00 1,0 3,00 7,0
M.peregrinum 12 2,0 1,8 1,0 1,00 1,0 2,25 6,0
M.abscessus 12 1,8 1,4 1,0 1,00 1,0 2,25 5,0
M.septicum 10 1,7 0,9 1,0 1,00 1,0 2,75 3,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний
Таблица 26 – Результат сравнительной статистики сроков появления
роста при культивировании на анализируемых средах
Микроорганизм Критерий Краскела-Уоллиса
Статистика p-value
M.avium 24,4 <0,001
M.kansasii 23,7 <0,001
M.gordonae 55,8 <0,001
M.fortuitum 0,9 0,63
M.peregrinum 0,5 0,76
M.abscessus 0,3 0,86
M.septicum 1,3 0,52
При p-value <0,05 различия статистически достоверны
Анализируя полученные результаты, можно сделать заключение об
отсутствии статистических значимых отличий для быстрорастущих НТМ по
скорости роста при использовании исследуемых агаризованных сред и среды
Левенштейна-Йенсена. Медленнорастущие виды НТМ на среде Левенштейна-
Йенсена и универсальной хромогенной среде давали видимый рост колоний
достоверно быстрее, чем на кровяном агаре.

121
5.2.2. Влияние температуры культивирования на скорость роста
нетуберкулезных микобактерий на агаризованных средах
В данной части эксперимента было проанализировано влияние температуры
28°С на сроки появления видимого роста колоний микроорганизмов при
использовании универсальной хромогенной среды в сравнении со сроками
появления роста при температуре 37°С. Результаты описательной статистики для
сроков роста у анализируемых культур при температуре 28°С приведены в таблице
27, для температуры 37°С были описаны в таблице 24.
Таблица 27 – Сроки выявления роста НТМ на универсальной
хромогенной средой при температуре 28°С
Микроорганизм n Mean Std Min 25% 50% 75% Max
M.avium 73 14,7 5,6 2,0 11,00 16,0 19,00 28,0
M.kansasii 41 12,3 2,4 9,0 11,00 12,0 14,00 18,0
M.gordonae 57 10,8 2,3 5,0 10,00 11,0 12,00 15,0
M.fortuitum 35 1,3 0,6 1,0 1,00 1,0 1,00 3,0
M.peregrinum 12 1,2 0,4 1,0 1,00 1,0 1,00 2,0
M.abscessus 12 1,2 0,4 1,0 1,00 1,0 1,00 2,0
M.septicum 10 1,3 0,5 1,0 1,00 1,0 1,75 2,0
n – количество штаммов; Mean – среднее значение количества колоний;
Std – стандартное отклонение количества колоний; Min, Max – минимальное и
максимальное количество колоний соответственно; 25%, 75% – 25 квантиль и
75 квантиль количества колоний соответственно; 50% – медианное значение
количества колоний
Для оценки статистической значимости влияния температуры
культивирования штаммов НТМ на универсальной хромогенной среде был
проведен расчет p-value для критерия Манна-Уитни, результаты которого
приведены в таблице 28.

122
Таблица 28 – Результат сравнительной статистики сроков появления
роста при культивировании на универсальной хромогенной среде при
температуре 37°С и 28°С
Микроорганизм Критерий Краскела-Уоллиса
Статистика p-value
M.avium 3413,0 0,267
M.kansasii 825,0 0,444
M.gordonae 1374,0 0,012
M.fortuitum 477,0 0,028
M.peregrinum 36,0 0,008
M.abscessus 42,0 0,021
M.septicum 50,0 0,481
При p-value <0,05 различия статистически достоверны
Таким образом, сроки появления роста на универсальной хромогенной среде
статистически значимо меньше практически для всех анализируемых видов
быстрорастущих НТМ и M.gordonae.
5.2.3. Оценка возможности использования новых вариантов лабораторной
посуды для культивирования нетуберкулезных микобактерий на
универсальной хромогенной среде
Полученные результаты исследований демонстрируют возможность
использования универсальной хромогенной среды для культивирования
большинства штаммов НТМ. Однако при пролонгировании времени инкубации
чашек с посевами в термостате была выявлена особенность достаточно быстрой
дегидратации среды, которая затрудняла продолжение исследования. Вариант
использования для посева пробирок со скошенной средой был исключен по
причине значительного ограничения данного метода посева первичного материала
в случае выявления в материале полимикробных ассоциаций.
В связи с вышеперечисленными особенностями нами было принято решение
по разработке новых вариантов лабораторной посуды, использование которой
отвечало бы нескольким параметрам: большая площадь поверхности среды для

123
возможности получения изолированных колоний микроорганизмов, создание
условий для предотвращения преждевременной дегидратации питательной среды
при продленном инкубировании, возможность создания аэробных условий за счет
различных вариантов аэрирования внутренней среды сосуда.
В итоге было разработано и апробировано три варианта лабораторной
посуды: чашка для культивирования микроорганизмов с двухпозиционной резьбой
(Патент РФ № 17302), флакон для культивирования микроорганизмов (Патент РФ
№ 175134), флакон для культивирования микроорганизмов со съемной крышкой
(Патент РФ № 175863).
В случае отсутствия возможности использования новых вариантов
лабораторной посуды преждевременную дегидратацию универсальной
хромогенной среды можно предотвратить, используя для упаковки чашек при
культивировании парафильма или контейнеров с плотно прилегающими
крышками.
5.3. Оценка возможности предварительной идентификации
кислотоустойчивых представителей порядка Actinomycetales при
использовании универсальной хромогенной среды
Особенности культуральных свойств НТМ, описанные в разделах 5.1 и 5.2
показали, что при использовании классических приемов культивирования на
плотных яичных средах возможно получение результатов ошибочного
определения НТМ по культуральным признакам к неверной группе микобактерий,
что, в свою очередь, может негативно отразится как на выборе приемов не только
для идентификации полученных культур, но и для определения чувствительности
микроорганизмов к антибактериальным препаратам. Ускоренный рост
медленнорастущих видов и замедленный рост быстрорастущих видов может быть
описан не только при работе с классическими средами, которые используются в

124
работе лабораторий противотуберкулезной службы, но и при использовании
агаризованных сред.
Для решения данного вопроса были проанализированы культуральные
свойства выросших на универсальной хромогенной среде НТМ и других видов
КУППА. Полученные результаты были сопоставлены с учетом классификации
НТМ по скорости роста и пигментообразованию.
Для всех фотохромогенных медленнорастущих культур НТМ были получены
следующие результаты: на универсальной хромогенной среде был выявлен
дисгонический рост, характеризующийся появлением мелких прозрачных или
слегка пигментированных колоний, размером 1-2 мм, легко снимающихся с
поверхности среды. Сроки появления признаков видимого роста находились в
пределах одной-двух недель, при пересеве на хромогенную среду с последующей
инкубацией при температуре 28°С. Такой рост был получен для 41-го штамма
M.kansasii и двух штаммов M.pseudoshottsii, что составило 100,0% от всех
фотохромогенных медленнорастущих культур НТМ.
Для скотохромогенных медленнорастущих культур НТМ был характерен
рост точечных колоний размером 1-2 мм желтого или оранжевого цвета, который
появлялся на универсальной хромогенной среде в течение одной-двух недель
культивирования при температуре 28°С. Описанные результаты оценки
культуральных свойств были получены для 57-и штаммов M.gordonae (95,0%
культур данного вида), трех штаммов M.lentiflavum и по одному штамму
M.scrofulaceum, M.bochemicum, M.interjectum и M.seoulense. Для всех штаммов
M.szulgai (n=6), M.xenopi (n=1) и трех штаммов M.gordonae рост при описанных
условиях культивирования получить не удалось. Таким образом, предварительная
идентификация с использованием универсальной хромогенной среды была
проведена для 64-х штаммов, что составило 86,5% от всех скотохромогенных
медленнорастущих культур НТМ.
В группу нефотохромогенных медленнорастущих НТМ было принято
решение включить все штаммы M.avium, вне зависимости от образования пигмента
на яичных средах. Большинство штаммов при пересеве на универсальную

125
хромогенную среду дали мелкие желтоватые или кремовые колонии размером 1-2
мм, с появлением видимого роста в течение 7-28 суток при культивировании при
температуре 28°С.
Следует отметить, что желтоватого цвета колонии дали штаммы M.avium,
которые образовали пигмент и при культивировании на яичных средах. Такой же
тип колоний был получен для штаммов M.malmoense и M.marseillense. Штаммы
M.avium с классическим вариантом колоний, штаммы M.chimaera/intracellulare,
M.celatum и M.gastri дали колонии светло-бежевого цвета. В данной группе
микроорганизмов также были выявлены штаммы, которые не дали рост в
проводимых условиях. Из 110-и штаммов не удалось получить видимый рост для
12-и штаммов M.avium с классическим вариантом колоний и 3-х штаммов
M.chimaera/intracellulare, что составило 13,6% от всех нефотохромогенных
медленнорастущих видов НТМ.
Быстрорастущие НТМ и другие бактерии из группы КУППА дали на
универсальной хромогенной среде окрашенные в различные оттенки синего цвета
колонии размером от 2 до 5 мм (рисунок 2), в некоторых случаях формируя
сложные структуры больших размеров (рисунок 3). Рост колоний отмечен в первые
двое-трое суток культивирования, однако характерное окрашивание колонии
приобретали к пятым-седьмым суткам.
Несмотря на то, что и быстрорастущие НТМ, и другие виды из группы
КУППА давали на среде одинаково окрашенные колонии, при анализе
культуральных свойств, была выявлена особенность, характерная только для
нокардий, гордоний и стрептомицет, которые в отличие от НТМ давали вросшие в
среду колонии, а стрептомицеты дополнительно на пятые-седьмые сутки
культивирования дополнительно приобретали беловатый налет на поверхности
колоний. На рисунках 16-25 представлены различные варианты колоний отдельных
видов НТМ, нокардий, гордоний и стрептомицет.

126
Рисунок 16 – Изолированные колонии M.fortuitum на универсальной
хромогенной среде (размер колоний 4-5 мм)
Рисунок 17 – Колонии M.fortuitum в виде консорциумов на
универсальной хромогенной среде (размер консорциумов – 9-12 мм)

127
Рисунок 18 – Колонии M.septicum на универсальной хромогенной среде
(размер колоний 3-4 мм)
Рисунок 19 – Колонии M.senegalense на универсальной хромогенной
среде (размер колоний 2-3 мм)
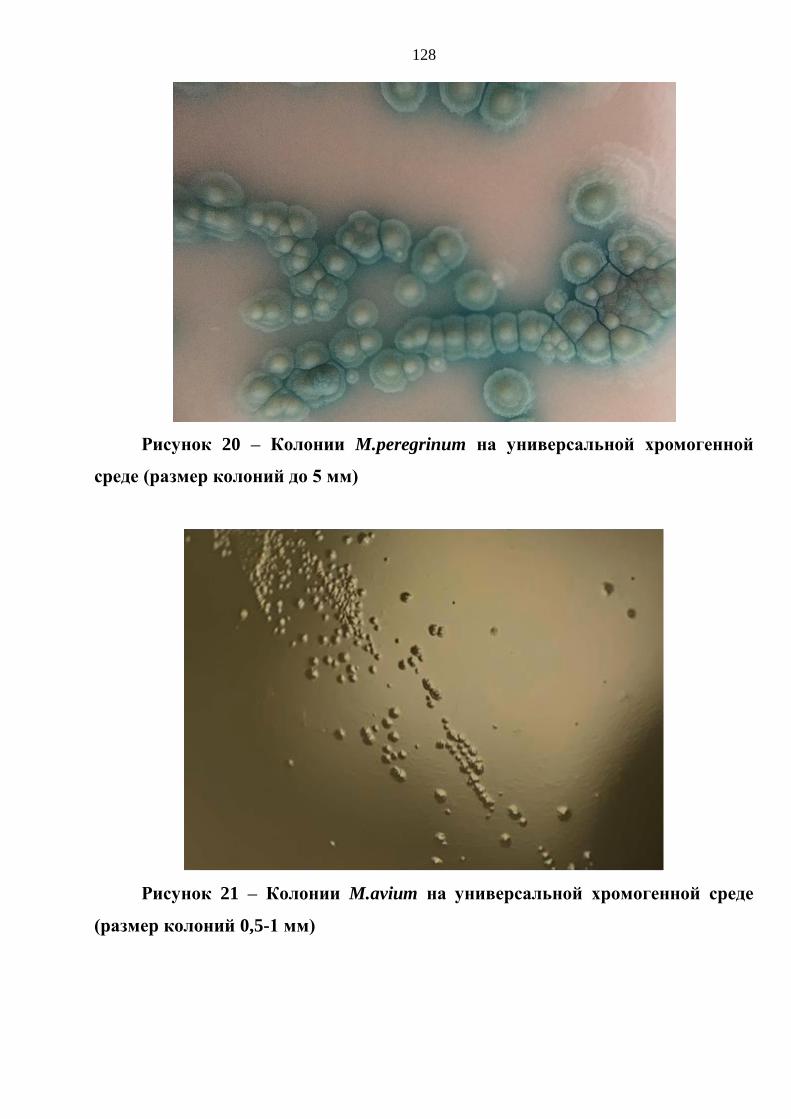
128
Рисунок 20 – Колонии M.peregrinum на универсальной хромогенной
среде (размер колоний до 5 мм)
Рисунок 21 – Колонии M.avium на универсальной хромогенной среде
(размер колоний 0,5-1 мм)
129
Рисунок 22 – Колонии Streptomyces violaceoruber на универсальной
хромогенной среде (размер колоний до 5 мм)
Рисунок 23 – Колонии Nocardia carnea на универсальной хромогенной
среде (размер колоний 1-2 мм)

130
Рисунок 24 – Колонии Gordonia sputi на универсальной хромогенной
среде (размер колоний 2-3 мм)
Рисунок 25 – Колонии Streptomyces phaeochromogenes на универсальной
хромогенной среде (размер колоний 2-3 мм)
Описанные свойства были выявлены у всех представителей быстрорастущих
НТМ, а также для всех видов нокардий, гордоний и стрептомицет. Представители
других родов КУППА на универсальной хромогенной среде давали скудный рост
или совсем его не проявляли. На предложенный способ предварительной

131
идентификации НТМ с использованием универсальной хромогенной среды был
получен Патент РФ № 2711954.
Таким образом, универсальная хромогенная среда, используемая в
эксперименте может быть использована для предварительной идентификации
групп НТМ, а также для дифференциации быстрорастущих НТМ от других
представителей группы КУППА. Однако, универсальные хромогенные среды
являются оптимальными для роста значительного числа микроорганизмов, что
может иметь значение при их использовании для первичного посева материала, так
как даже быстрорастущие микроорганизмы имеют определенную задержку роста
при культивировании на плотных питательных средах. В связи с этим было
принято решение оценки возможности использования с хромогенной средой
селективной добавки, подавляющей рост «классической» микрофлоры,
характерной для нестерильных локусов тела человека.
5.4. Оценка использования универсальной хромогенной среды с селективной
добавкой для культивирования кислотоустойчивых представителей порядка
Actinomycetales
В качестве селективной добавки к универсальной хромогенной среде была
использована смесь из двух антибактериальных препаратов: бацитрацина и
полимиксина сульфата, не оказывающих действия на КУППА, для подавления
сопутствующей грамположительной и грамотрицательной микрофлоры.
Концентрации антибактериальных препаратов были выбраны из расчета
аналогичного для селективной добавки при выделении Burkholderia cepacia
complex, так как имеются работы, в которых описана возможность выведения
некоторых быстрорастущих НТМ на данной среде. Результаты оценки роста
штаммов на среде представлены в таблице 29.

132
Таблица 29 – Оценка интенсивности роста некоторых видов
кислотоустойчивых актиномицет на универсальной хромогенной среде с
селективной добавкой
Микроорганизм
Количество
штаммов
(n)
Доля штаммов с соответствующим
типом роста, n (%)
Обильный Умеренный Скудный
M.avium 73 55 (75,3) 18 (24,7) -
M.gordonae 57 43 (75,4) 14 (24,6) -
M.kansasii 41 31 (75,6) 10 (24,4) -
M.fortuitum 35 35 (100,0) - -
M.peregrinum 12 12 (100,0) - -
M.abscessus 12 12 (100,0) - -
M.septicum 10 10 (100,0) - -
M.chimaera/intracelullare 8 6 (75,0) 2 (25,0) -
M.malmoense 8 8 (100,0) - -
M.senegalense 7 7 (100,0) - -
M.chelonae 7 7 (100,0) - -
M.lentiflavum 3 3 (100,0) - -
M.pseudoshottsii 2 2 (100,0) - -
M.marseillense 2 1 (50,0) 1 (50,0) -
M.porcinum 2 2 (100,0) - -
M.frederiksbergense 2 2 (100,0) - -
M.celatum 1 1 (100,0) - -
M.gastri 1 - 1 (100,0) -
M.scrofulaceum 1 - 1 (100,0) -
M.bohemicum 1 - 1 (100,0) -
M.interjectum 1 - 1 (100,0) -
M.seoulense 1 - 1 (100,0) -
M.iranicum 1 1 (100,0) - -
M.monacense 1 1 (100,0) - -
M.flavescens 1 1 (100,0) - -
M.chitae 1 1 (100,0) - -
M.farcinogenes 1 1 (100,0) - -
Streptomyces spp. 39 27 (69,3) 12 (30,7) -
Nocardia spp. 14 9 (64,3) 5 (35,7) -
Gordonia spp. 10 8 (80,0) 2 (20,0) -
Обильный рост – сплошной рост изолированных колоний с типичными
свойствами
Умеренный рост – значительный рост колоний с типичными свойствами (100
и более колоний)
Скудный рост – единичные колонии с типичными свойствами (до 100
колоний)

133
Тестирование возможного подавления роста с использованием селективной
добавки проводили по наличию роста культуры при инокуляции на поверхность
агара суспензии, содержащей 1000-10000 КОЕ/мл.
В качестве тестируемых культур были выбраны быстрорастущие,
медленнорастущие виды НТМ, а также гордонии, нокардии и стрептомицеты.
Время инкубирования для быстрорастущих видов составило до семи суток, для
медленнорастущих видов до 28-и суток. В исследование были включены штаммы,
которые дали рост на универсальной хромогенной среде по результатам,
описанным в разделе 5.2 и 5.3. В результате 100% штаммов из группы КУППА при
посеве на универсальную хромогенную среду с селективной добавкой дали
обильный или умеренный рост.
Для контрольных штаммов Escherichia coli ATCC 25922, P.aeruginosa ATCC
27853, S.aureus ATCC 25923 рост на универсальной хромогенной среде с
селективной добавкой не был получен.
Предлагаемый способ посева микроорганизмов был апробирован на
клиническом материале. Получен Патент РФ № 2711957 «Способ первичного
посева клинического материала для выделения нетуберкулезных микобактерий».
5.5. Резюме
Анализируя полученные данные можно сделать заключение: классификация
НТМ, основанная на скорости роста и культуральных свойствах микроорганизмов
имеет ряд ограничений, особенно в отношении видов, имеющих схожие по
морфологии и цвету колонии при использовании классических сред для выделения
МТВс. Также были выявлены определенные сходства культуральных свойств
быстрорастущих НТМ и других КУППА при инкубировании посевов на плотных
яичных средах, что может затруднять выбор методов для их дальнейшей
идентификации. Таким образом, при визуальной оценке роста на среде
Левенштейна-Йенсена нельзя достоверно разделить микроорганизмы не только

134
внутри группы НТМ, но и с другими КУППА. Использование среды Левенштейна-
Йенсена позволяет выделять все виды НТМ, которые были включены в
исследование, однако не является оптимальной для культивирования других видов
КУППА. Часть КУППА не дали роста на среде Левенштейна-Йенсена или вызвали
значительное ее изменение, что характерно для классической контаминирующей
микрофлоры.
Ростовые свойства основных видов НТМ на универсальной хромогенной
среде достоверно более выражены чем на кровяном агаре по количественным
признакам, за исключением M.peregrinum. По критерию продуктивности «более
70%» на универсальной хромогенной среде были получены достоверно лучше
результаты для штаммов M.avium, M.fortuitum, M.abscessus, M.septicum.
Сроки появления роста основных видов НТМ на универсальной хромогенной
среде сопоставимы с результатами, полученными на среде Левенштейна-Йенсена
и достоверно отличаются от сроков появления колоний на кровяном агаре в
сторону сокращения. Влияние температуры культивирования статистически не
отличается при культивировании M.avium и M.kansasii и M.septicum, но при
температуре 28°С рост остальных культур был получен достоверно в более
короткие сроки. Тем не менее, использование агаризованных сред с учетом
пролонгирования сроков инкубации требует разработки и внедрения новой
лабораторной посуды для предотвращения преждевременной дегидратации среды.
Использование универсальной хромогенной среды позволяет проводить
предварительную идентификацию НТМ и других КУППА по культуральным
свойствам, а влияние селективной добавки не снижает ростовых свойств КУППА,
что может быть использовано при работе как с клиническим материалом, так и для
выделения чистых культур КУППА из смесей с другими микроорганизмами.

135
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ,
ИЗЛОЖЕННЫМ В ГЛАВЕ 5
Статьи в журналах, рекомендованных ВАК:
1. Лямин, А.В. Лабораторная диагностика микобактериозов / А.В. Лямин, А.В.
Жестков, Д.Д. Исматуллин [и др.] // Вестник современной клинической медицины.
– 2017. – Т. 10. – № 1. – С. 29-35.
2. Лямин, А.В. Лабораторная диагностика микобактериозов у пациентов с
муковисцидозом (обзор литературы) / А.В. Лямин, Д.Д, Исматуллин, А.В. Жестков
[и др.] // Клиническая лабораторная диагностика. – 2018. – Т. 63. – № 5. – С. 315-
320.
3. Лямин, А.В. Общая характеристика и клиническое значение представителей
родов Nocardia и Gordonia / А.В. Лямин, А.В. Жестков, Т.Р. Никитина [и др.] //
Инфекция и иммунитет. – 2019. – Т. 9. – № 3-4. – С. 429-436.
Публикации в других изданиях:
1. Козлов, А.В. Сравнение методов деконтаминации мокроты при исследовании
микобактерий / А.В. Козлов, А.В. Лямин, А.А. Ерещенко [и др.] // Лабораторная
служба. – 2017. – Т. 6. – № 3. – Тезисы III Российского конгресса лабораторной
медицины. – М. – С. 81.
2. Исматуллин, Д.Д. Новые возбудители заболеваний респираторного тракта у
иммунокопрометированных пациентов (обзор литературы) / Д.Д, Исматуллин,
М.О. Золотов, А.В. Лямин [и др.] // Наука и инновации в медицине. – 2019. – Т. 4.
– № 3. – С. 19-24.
Патенты:
1. Патент РФ № 173302 «Чашка для культивирования микроорганизмов с
двухпозиционной резьбой» (Бюл. № 24, 2017 г.). Авторы: Лямин А.В., Тонеев И.Р.,
Козлов А.В. [и др.].
2. Патент РФ № 175134 «Флакон для культивирования микроорганизмов» (Бюл.
№ 33, 2017 г.). Авторы: Лямин А.В., Тонеев И.Р., Козлов А.В. [и др.].

136
3. Патент РФ № 175863 «Флакон для культивирования микроорганизмов со
съемной крышкой» (Бюл. № 36, 2017 г.). Авторы: Лямин А.В., Тонеев И.Р., Козлов
А.В. [и др.].
4. Патент № 2711954 «Способ предварительной идентификации
нетуберкулезных микобактерий с использованием универсальной хромогенной
среды» (Бюл. № 3, 2020 г.) Авторы: Лямин А.В., Исматуллин Д.Д., Кондратенко
О.В. [и др.].
5. Патент № 2711957 «Способ первичного посева клинического материала для
выделения нетуберкулезных микобактерий» (Бюл. № 3, 2020 г.) Авторы: Лямин
А.В., Исматуллин Д.Д., Кондратенко О.В. [и др.].

137
ГЛАВА 6. ОСОБЕННОСТИ ИДЕНТИФИКАЦИИ
КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ ПОРЯДКА
ACTINOMYCETALES МЕТОДОМ MALDI-TOF МАСС-СПЕКТРОМЕТРИИ
6.1. Сравнение методов идентификации представителей семейства
Mycobacteriaceae с использованием методов ДНК-гибридизации, MALDI-ToF
масс-спектрометрии, секвенирования по Сэнгеру
Полученные результаты сравнения использования методов идентификации
НТМ: ДНК-гибридизации и MALDI-ToF масс-спектрометрии, описанные в Главе
4, выявили ряд вопросов в отношении расхождений результатов определения
видовой принадлежности микобактерий, для решения которых было проведено
секвенирование по Сэнгеру дополнительно для 74 культур НТМ, рост которых был
получен из клинического материала на среде Левенштейна-Йенсена от 74
пациентов.
Все образцы не имели признаков роста нескольких культур и предварительно
все образцы были идентифицированы с использованием метода ДНК-
гибридизации. После этого была проведена идентификация методом MALDI-ToF
масс-спектрометрии. Все образцы дополнительно были протестированы на
наличие ДНК МТВс, в случае обнаружения которой определяли вид МТВс. Оба
этапа связанных с обнаружением ДНК МТВс и видового определения проводили
методом ПЦР. На последнем этапе проводили идентификацию методом
секвенирования генов 16S, hsp65, rpob, sod по Сэнгеру.
Всеми анализируемыми методами были идентифицированы представители
рода Mycobacterium, идентифицированные до рода, вида или комплекса.
При исследовании 74-х культур методом ДНК-гибридизации видовая
принадлежность была определена для 63-х штаммов (85,1%). Одна культура была
идентифицирована до комплекса, 12 культур удалось идентифицировать только до
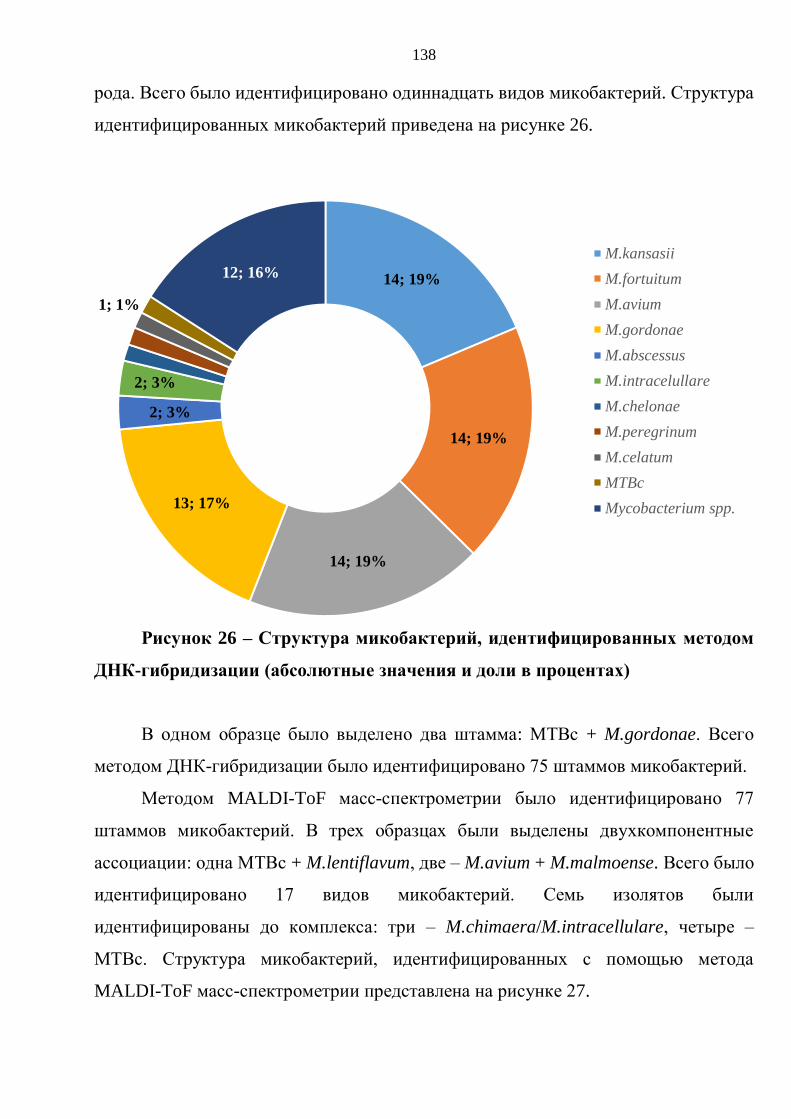
138
рода. Всего было идентифицировано одиннадцать видов микобактерий. Структура
идентифицированных микобактерий приведена на рисунке 26.
Рисунок 26 – Структура микобактерий, идентифицированных методом
ДНК-гибридизации (абсолютные значения и доли в процентах)
В одном образце было выделено два штамма: МТВс + M.gordonae. Всего
методом ДНК-гибридизации было идентифицировано 75 штаммов микобактерий.
Методом MALDI-ToF масс-спектрометрии было идентифицировано 77
штаммов микобактерий. В трех образцах были выделены двухкомпонентные
ассоциации: одна МТВс + M.lentiflavum, две – M.avium + M.malmoense. Всего было
идентифицировано 17 видов микобактерий. Семь изолятов были
идентифицированы до комплекса: три – M.chimaera/M.intracellulare, четыре –
МТВс. Структура микобактерий, идентифицированных с помощью метода
MALDI-ToF масс-спектрометрии представлена на рисунке 27.
14; 19%
14; 19%
14; 19%
13; 17%
2; 3%
2; 3%
1; 1%
12; 16%
M.kansasii
M.fortuitum
M.avium
M.gordonae
M.abscessus
M.intracelullare
M.chelonae
M.peregrinum
M.celatum
MTBc
Mycobacterium spp.

139
Рисунок 27 – Структура микобактерий, идентифицированных методом
MALDI-ToF масс-спектрометрии (абсолютные значения и доли в процентах)
С помощью метода секвенирования по Сэнгеру видовая идентификация была
установлена для 72-х изолятов. Используемый набор генов не позволил провести
видовую дифференциацию между близкородственными видами M.porcinum и
M.peregrinum в двух образцах культур. Всего было идентифицировано 16-ть видов
микобактерий, видовая структура которых представлена на рисунке 28.
16; 22%
14; 19%
9; 12%
10; 13%
3; 4%
3; 4%
3; 4%
2; 3%
1; 1%
4; 5%
M.kansasii
M.avium
M.gordonae
M.fortuitum
M.peregrinum
M.szulgai
M.chimaera/intracellulare
M.abscessus
M.septicum
M.senegalense
M.lentiflavum
M.malmoense
M.chelonae
M.frederiksbergense
MTBc
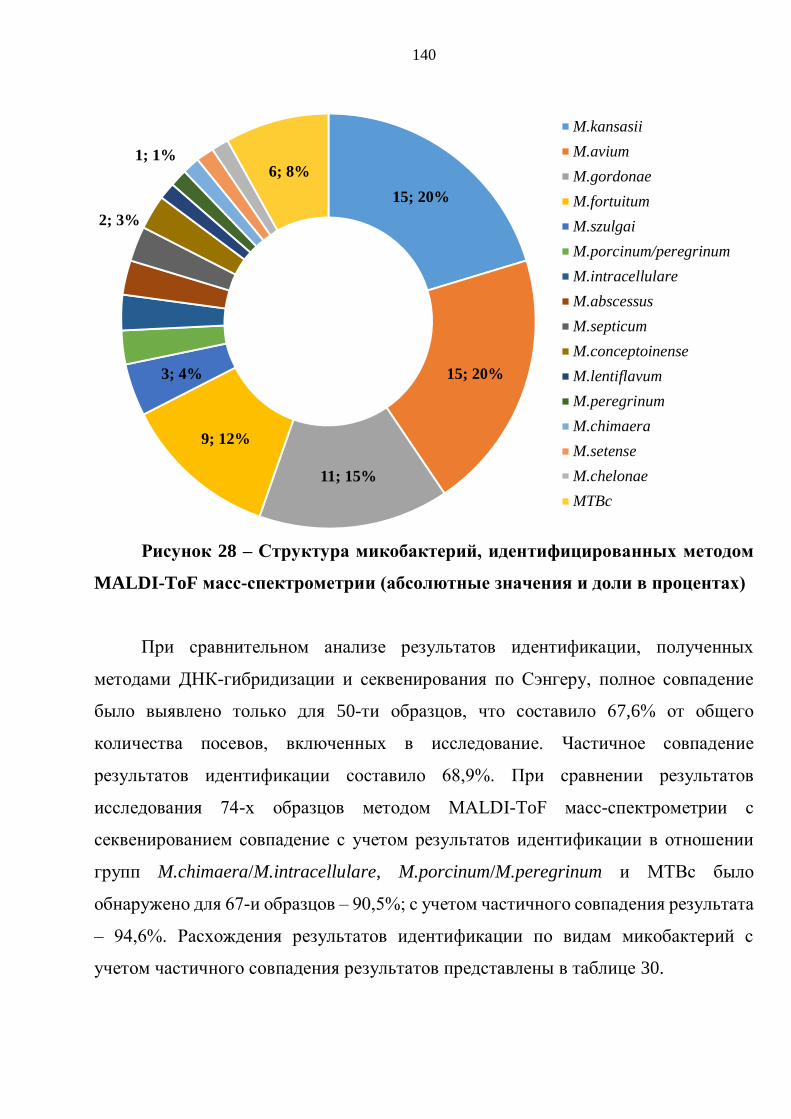
140
Рисунок 28 – Структура микобактерий, идентифицированных методом
MALDI-ToF масс-спектрометрии (абсолютные значения и доли в процентах)
При сравнительном анализе результатов идентификации, полученных
методами ДНК-гибридизации и секвенирования по Сэнгеру, полное совпадение
было выявлено только для 50-ти образцов, что составило 67,6% от общего
количества посевов, включенных в исследование. Частичное совпадение
результатов идентификации составило 68,9%. При сравнении результатов
исследования 74-х образцов методом MALDI-ToF масс-спектрометрии с
секвенированием совпадение с учетом результатов идентификации в отношении
групп M.chimaera/M.intracellulare, M.porcinum/M.peregrinum и МТВс было
обнаружено для 67-и образцов – 90,5%; с учетом частичного совпадения результата
– 94,6%. Расхождения результатов идентификации по видам микобактерий с
учетом частичного совпадения результатов представлены в таблице 30.
15; 20%
15; 20%
11; 15%
9; 12%
3; 4%
2; 3%
1; 1%6; 8%
M.kansasii
M.avium
M.gordonae
M.fortuitum
M.szulgai
M.porcinum/peregrinum
M.intracellulare
M.abscessus
M.septicum
M.conceptoinense
M.lentiflavum
M.peregrinum
M.chimaera
M.setense
M.chelonae
MTBc

141
Таблица 30 – Расхождения результатов идентификации микобактерий с
использованием анализируемых методов
ДНК-гибридизация MALDI-ToF масс-
спектрометрия
Секвенирование по
Сэнгеру
ДНК
МТBc
Mycobacterium spp. M.kansasii M.kansasii -
M.fortuitum M.septicum M.septicum -
M.fortuitum M.septicum M.septicum -
M.peregrinum M.fortuitum M.fortuitum -
M.fortuitum M.peregrinum M.peregrinum -
M.fortuitum M.senegalense M.conceptionense -
M.celatum M.chimaera/intracelullare M.chimaera -
Mycobacterium spp. M.lentiflavum M.lentiflavum -
M.fortuitum M.senegalense M.conceptionense -
M.gordonae M.kansasii M.kansasii -
M.kansasii MTBc M.tuberculosis +
M.fortuitum M.fortuitum M.setense -
Mycobacterium spp. M.fortuitum M.fortuitum -
Mycobacterium spp. M.frederiksbergense M.tuberculosis +
Mycobacterium spp. M.kansasii M.kansasii -
M.intracellulare M.chimaera/intracelullare M.intracellulare -
M.intracellulare M.chimaera/intracelullare M.intracellulare -
M.avium M.avium+M.malmoense M.avium -
Mycobacterium spp. M.szulgai M.szulgai -
Mycobacterium spp. M.szulgai M.szulgai -
Mycobacterium spp. M.szulgai M.szulgai -
M.kansasii M.kansasii M.avium +
Mycobacterium spp. MTBc+M.lentiflavum M.tuberculosis +
Mycobacterium spp. MTBc M.tuberculosis +
Mycobacterium spp. MTBc M.tuberculosis +
MTBc+M.gordonae M.gordonae M.tuberculosis +
M.avium M.avium+M.malmoense M.avium -
Mycobacterium spp. M.peregrinum M.porcinum
/M.peregrinum
-
M.fortuitum M.peregrinum M.porcinum
/M.peregrinum
-
Важно отметить, что максимальные расхождения результатов
идентификации с помощью масс-спектрометрии и секвенирования среди
быстрорастущих НТМ были выявлены для представителей M.fortuitum complex (3

142
несоответствия – 100,0% от числа несовпадений результатов по данной группе
НТМ).
Среди медленнорастущих видов максимальное количество расхождений
между тремя анализируемыми методами было получено в случае выявления
ассоциации микобактерий или обнаружения ДНК представителей МТВс.
Для всех трех ассоциаций, идентифицированных с помощью метода MALDI-
ToF масс-спектрометрии, удалось выделить чистую культуру НТМ на
универсальной хромогенной среде с ее последующей идентификацией со
значением Score более 2,00.
Для оценки чувствительности и специфичности видовой идентификации
НТМ методами ДНК-гибридизации и MALDI-ToF масс-спектрометрии в
сравнении с методом секвенирования, как «золотого» стандарта был проведен
ROC-анализ, результаты которого приведены в таблице 31.
Таблица 31 – Результаты оценки чувствительности и специфичности
анализируемых методов в сравнении с методом секвенирования
Метод Чувствительность, % Специфичность, % AUC p
ДНК-гибридизация 87,5 100,0 0,938 <0,001
MALDI-ToF МС 98,6 100,0 0,993 <0,001
AUC – площадь под кривой, p – уровень достоверности
ДНК-гибридизация, как метод идентификации НТМ показал приемлемые
чувствительность и специфичность, однако для масс-спектрометрии была
определена достоверно более высокая чувствительность с сопоставимой
специфичностью. Таким образом, расхождение результатов видовой
идентификации НТМ, описанных в главе 4, обусловлено более низкой
чувствительностью метода ДНК-гибридизации в сравнении с методом MALDI-ToF
масс-спектрометрией, что необходимо учитывать при анализе полученных
результатов видовой идентификации НТМ сравниваемыми методами.

143
6.2. Особенности проведения идентификации кислотоустойчивых
представителей порядка Actinomycetales в зависимости от среды для
культивирования с использованием метода MALDI-ToF масс-спектрометрии
6.2.1. Оценка влияния среды культивирования на результат идентификации
нетуберкулезных микобактерий методом MALDI-ToF масс-спектрометрии
Известно, что при проведении идентификации НТМ методом MALDI-ToF
масс-спектрометрии была выявлена важная особенность, которая в конечном счете
определила структуру библиотеки спектров для микобактерий. Оказалось, что при
культивировании на различных средах (Левенштейна-Йенсена и Миддлбрука)
микобактерии дают спектры, отличающиеся по количеству и интенсивности пиков
[151,310]. Включение в библиотеку спектров микобактерий, рост которых получен
на плотной и жидкой средах значительно повысил возможности видовой
идентификации НТМ. В связи с этим было принято решение по сравнению
результатов идентификации НТМ, выросших на плотной яичной среде и
универсальной хромогенной среде.
В данной части исследования были проанализированы результаты
идентификации штаммов НТМ, описанные в главе 5, которые дали рост на
универсальной хромогенной среде. Всего были проанализированы результаты
видовой идентификации 292-х штаммов НТМ. Перед проведением процедур
идентификации все штаммы были пересеяны на среды Левенштейна-Йенсена и
универсальную хромогенную среду. Посевы инкубировались при температуре
37°С в первые сутки и 28°С в последующие сутки культивирования до появления
роста. Идентификацию проводили в день появления роста культуры, достаточной
для идентификации методом прямого нанесения. Результаты идентификации
представлены в таблице 32.

144
Таблица 32 – Сравнение результатов идентификации микобактерий в
зависимости от методики нанесения материала и среды для культивирования
Вид
микобактерии n
Результат видовой
идентификации на среде
Левенштейна-Йенсена
Результат видовой
идентификации на среде
универсальной среде
Метод
прямого
нанесения
n (%)
Метод
экстракции
n (%)
Методом
прямого
нанесения
n (%)
Методом
расширенного
нанесения
n (%)
M.avium 73 54 (74,0) 73 (100,0) 69 (94,5) 73 (100,0)
M.gordonae 57 41 (71,9) 57 (100,0) 54 (94,7) 57 (100,0)
M.kansasii 41 32 (78,0) 41 (100,0) 38 (92,7) 41 (100,0)
M.fortuitum 35 30 (85,7) 35 (100,0) 35 (100,0) 35 (100,0)
M.peregrinum 12 8 (66,7) 12 (100,0) 12 (100,0) 12 (100,0)
M.abscessus 12 6 (50,0) 12 (100,0) 12 (100,0) 12 (100,0)
M.septicum 10 7 (70,0) 10 (100,0) 10 (100,0) 10 (100,0)
M.malmoense 8 5 (62,5) 8 (100,0) 8 (100,0) 8 (100,0)
M.chimaera/
intracellulare 8 6 (75,0) 8 (100,0) 6 (75,0) 8 (100,0)
M.senegalense 7 7 (100,0) 7 (100,0) 7 (100,0) 7 (100,0)
M.chelonae 7 6 (85,7) 7 (100,0) 7 (100,0) 7 (100,0)
M.lentiflavum 3 2 (66,7) 3 (100,0) 3 (100,0) 3 (100,0)
M.pseudoshottsii 2 1 (50,0) 2 (100,0) 1 (50,0) 2 (100,0)
M.marseillense 2 2 (100,0) 2 (100,0) 2 (100,0) 2 (100,0)
M.frederiksbergens 2 1 (50,0) 2 (100,0) 2 (100,0) 2 (100,0)
M.porcinum 2 1 (50,0) 2 (100,0) 2 (100,0) 2 (100,0)
M.scrofulaceum 1 1 (100,0) 1 (100,0) 0 (0,0%) 1 (100,0)
M.bochemicum 1 0 (0,0%) 1 (100,0) 0 (0,0%) 1 (100,0)
M.interjectum 1 0 (0,0%) 1 (100,0) 0 (0,0%) 1 (100,0)
M.seoulense 1 1 (100,0) 1 (100,0) 1 (100,0) 1 (100,0)
M.celatum 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
M.gastri 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
M.iranicum 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
M.monacense 1 1 (100,0) 1 (100,0) 1 (100,0) 1 (100,0)
M.flavescens 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
M.chitae 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
M.farcinogenes 1 0 (0,0%) 1 (100,0) 1 (100,0) 1 (100,0)
ИТОГО 292 212 (72,6) 292 (100,0) 276 (94,5) 292 (100,0)
Штаммы, засеянные на среду Левенштейна-Йенсена, идентифицировали с
пробоподготовкой методом прямого нанесения; в случае отсутствия результата

145
видовой или группой идентификации использовали пробоподготовку методом
экстракции муравьиной кислотой. Штаммы, рост которых получен на
универсальной хромогенной среде, исследовали с пробоподготовкой методом
прямого нанесения. Если данным методом не был получен приемлемый результат
видовой идентификации, дополнительно использовали пробоподготовку
расширенным методом прямого нанесения.
При расчете оценки значимости различий результатов идентификации НТМ
на среде Левенштейна-Йенсена анализируемыми методами пробоподготовки
значение критерия Х2=92,698, уровень значимости p <0,001. При расчете оценки
значимости различий результатов идентификации НТМ на универсальной
хромогенной среде с различными методами пробоподготовки значение критерия
Х2=16,451, уровень значимости p <0,001. При расчете оценки значимости различий
результатов идентификации НТМ на среде Левенштейна-Йенсена и универсальной
хромогенной среде с проведением пробоподготовки методом прямого нанесения
значение критерия Х2=51,060, уровень значимости p <0,001. Таким образом, для
всех сравниваемых параметров выявлены статистически значимые результаты
влияния среды на результат идентификации НТМ. Данные по сравнению
пробоподготовки методом прямого нанесения с методом экстракции и
расширенным методом прямого нанесения оказались менее приемлемыми с точки
зрения конечного результата идентификации. Однако результаты идентификации,
с пробоподготовкой методом прямого нанесения в зависимости от среды,
продемонстрировали достоверно более высокий уровень при культивировании
НТМ на универсальной хромогенной среды.
Отрицательные результаты при проведении идентификации методом
прямого нанесения и методом экстракции муравьиной кислотой при работе с
КУППА обусловлены спецификой культуральных свойств данных
микроорганизмов. В случае, если культура была получена в виде вросших колоний
или при дисгоническом росте, использование стандартных инструментов для
нанесения материала на мишень вызывало определенные трудности. Для решения
данного вопроса были разработаны и апробированы новые инструменты для сбора

146
колоний микроорганизмов с поверхности плотных питательных сред (Патент РФ
№ 187404, Патент РФ № 187421).
Использование универсальной хромогенной среды, анализируемой в
исследовании позволяет получить приемлемые результаты идентификации НТМ с
использованием метода прямого нанесения, что значительно упрощает процесс
работы с данной группой микроорганизмов в случае получения роста на
агаризованных средах и расширяет возможность работы с НТМ в лабораториях, не
связанных с противотуберкулезной службой.
6.2.2. Оценка влияния среды культивирования на результат идентификации
других микроорганизмов из группы кислотоустойчивых представителей
порядка Actinomycetales методом MALDI-ToF масс-спектрометрии
Дополнительно было принято решение проанализировать влияние среды
культивирования на результат идентификации других наиболее распространенных
бактерий из группы КУППА: Streptomyces spp., Nocardia spp., Gordonia spp. Общее
количество штаммов 63. Условия культивирования и методы идентификации были
сопоставимы с описанными для НТМ. Результаты идентификации представлены в
таблице 33.
Таблица 33 – Сравнение результатов идентификации микобактерий в
зависимости от методики нанесения материала и среды для культивирования
Вид
микобактерии n
Результат видовой
идентификации на среде
Левенштейна-Йенсена
Результат видовой
идентификации на среде
универсальной среде
Метод
прямого
нанесения
n (%)
Метод
экстракции
n (%)
Методом
прямого
нанесения
n (%)
Методом
расширенного
нанесения
n (%)
Streptomyces spp. 39 6 (15,4,0) 27 (69,2) 30 (76,9) 39 (100,0)
Nocardia spp. 14 9 (64,2) 11 (78,6) 7 (50,0) 14 (100,0)
Gordonia spp. 10 4 (40,0) 8 (80,0) 9 (90,0) 10 (100,0)
ИТОГО 63 19 (30,2) 46 (73,0) 46 (73,0) 63 (100,0)

147
При расчете оценки значимости различий результатов идентификации
стрептомицет, нокардий и гордоний на среде Левенштейна-Йенсена различными
методами значение критерия Х2=23,166, уровень значимости p <0,001. При расчете
оценки значимости различий результатов идентификации стрептомицет, нокардий
и гордоний на универсальной хромогенной среде различными методами значение
критерия Х2=19,651, уровень значимости p <0,001. При расчете оценки значимости
различий результатов идентификации анализируемых микроорганизмов на среде
Левенштейна-Йенсена и универсальной хромогенной среде методом прямого
нанесения значение критерия Х2=23,166, уровень значимости p <0,001. Таким
образом, для всех сравниваемых параметров выявлены статистически значимые
результаты влияния среды на результат идентификации других микроорганизмов
из группы КУППА. Можно сделать заключение о том, что среда Левенштейна-
Йенсена не является оптимальной для идентификации представителей рода
Streptomyces, а метод прямого нанесения при использовании универсальной
хромогенной среды является приемлемым только в отношении видовой
идентификации гордоний.
6.3. Сравнение белковых профилей, полученных методом MALDI-ToF масс-
спектрометрии у представителей семейства Mycobacteriaceae, выращенных
на различных питательных средах
В научной литературе имеются данные, которые показывают значение среды
культивирования НТМ на результат идентификации методом MALDI-ToF масс-
спектрометрии. В связи с этим в библиотеки спектров НТМ для масс-
спектрометров стали включать результаты идентификации культур, выросших на
средах Левенштейна-Йенсена и Миддлбрука [151, 310]. Еще одна важная
особенность при идентификации НТМ данным методом заключается в
необходимости использования специальной подготовки культуры перед
идентификацией.
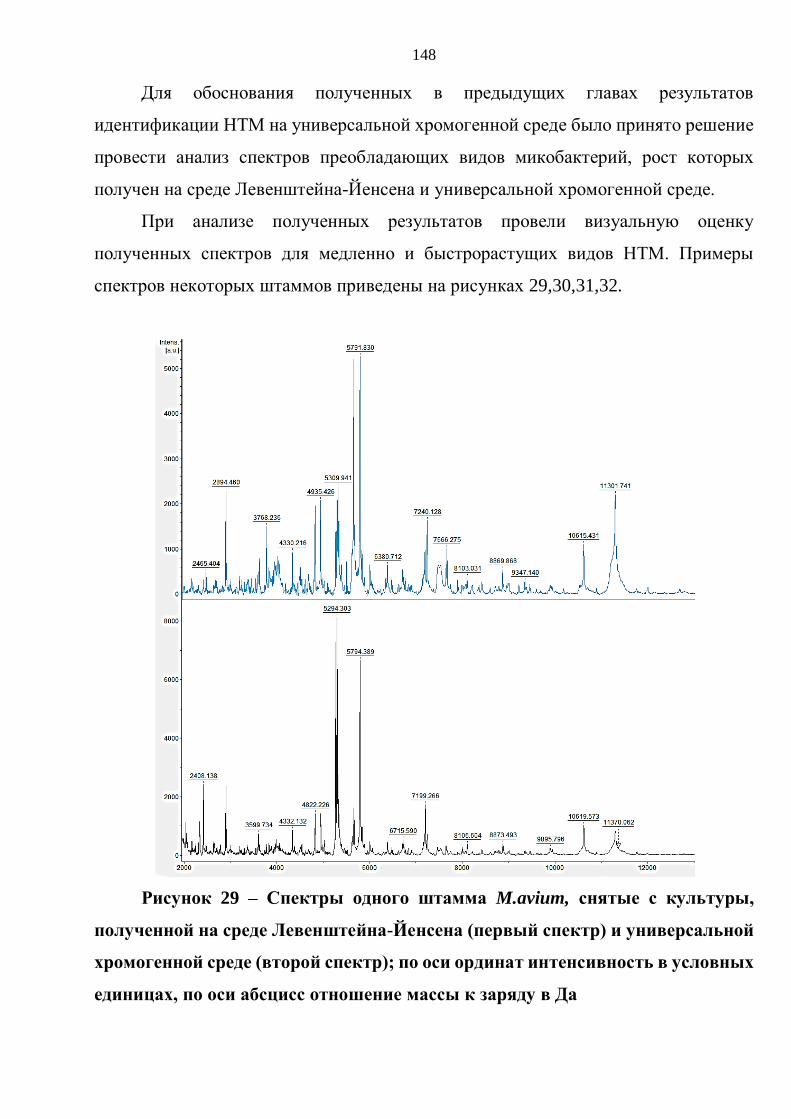
148
Для обоснования полученных в предыдущих главах результатов
идентификации НТМ на универсальной хромогенной среде было принято решение
провести анализ спектров преобладающих видов микобактерий, рост которых
получен на среде Левенштейна-Йенсена и универсальной хромогенной среде.
При анализе полученных результатов провели визуальную оценку
полученных спектров для медленно и быстрорастущих видов НТМ. Примеры
спектров некоторых штаммов приведены на рисунках 29,30,31,32.
Рисунок 29 – Спектры одного штамма M.avium, снятые с культуры,
полученной на среде Левенштейна-Йенсена (первый спектр) и универсальной
хромогенной среде (второй спектр); по оси ординат интенсивность в условных
единицах, по оси абсцисс отношение массы к заряду в Да
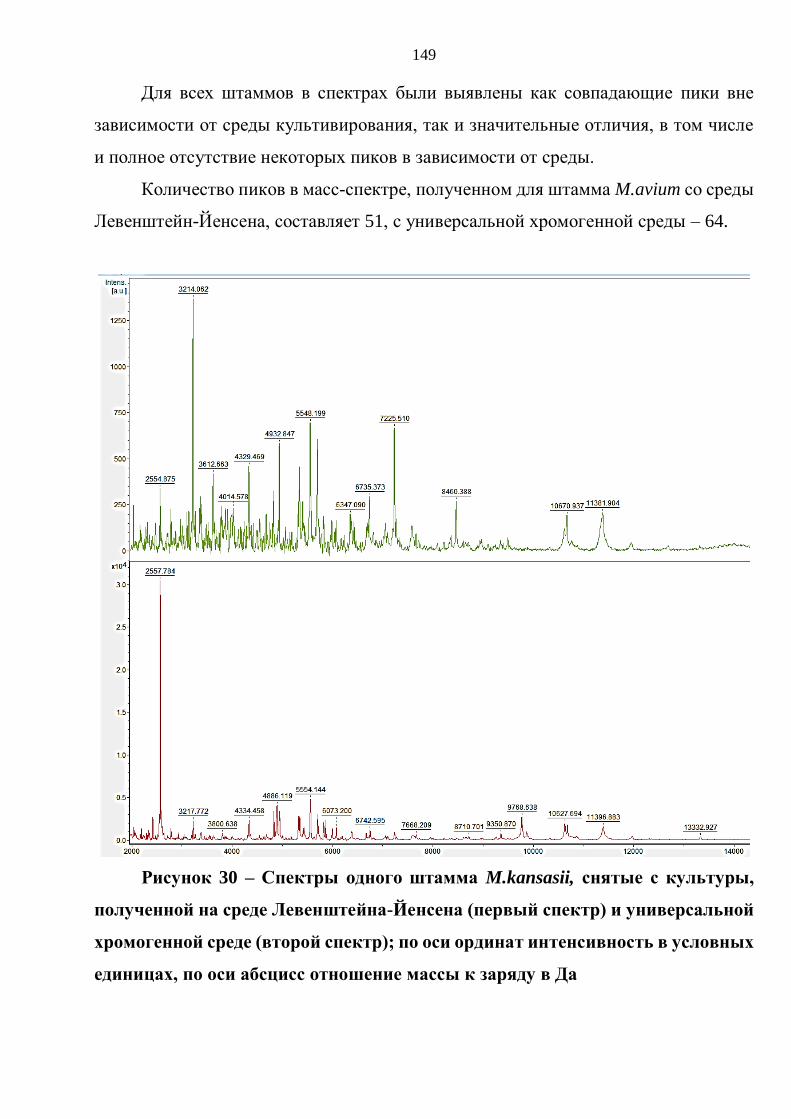
149
Для всех штаммов в спектрах были выявлены как совпадающие пики вне
зависимости от среды культивирования, так и значительные отличия, в том числе
и полное отсутствие некоторых пиков в зависимости от среды.
Количество пиков в масс-спектре, полученном для штамма M.avium со среды
Левенштейн-Йенсена, составляет 51, с универсальной хромогенной среды – 64.
Рисунок 30 – Спектры одного штамма M.kansasii, снятые с культуры,
полученной на среде Левенштейна-Йенсена (первый спектр) и универсальной
хромогенной среде (второй спектр); по оси ординат интенсивность в условных
единицах, по оси абсцисс отношение массы к заряду в Да

150
Количество пиков в масс-спектре, полученном для штамма M.kansasii со
среды Левенштейн-Йенсена, составляет 31, с универсальной хромогенной среды –
64.
Рисунок 31 – Спектры одного штамма M.fortuitum, снятые с культуры,
полученной на среде Левенштейна-Йенсена (первый спектр) и универсальной
хромогенной среде (второй спектр); по оси ординат интенсивность в условных
единицах, по оси абсцисс отношение массы к заряду в Да
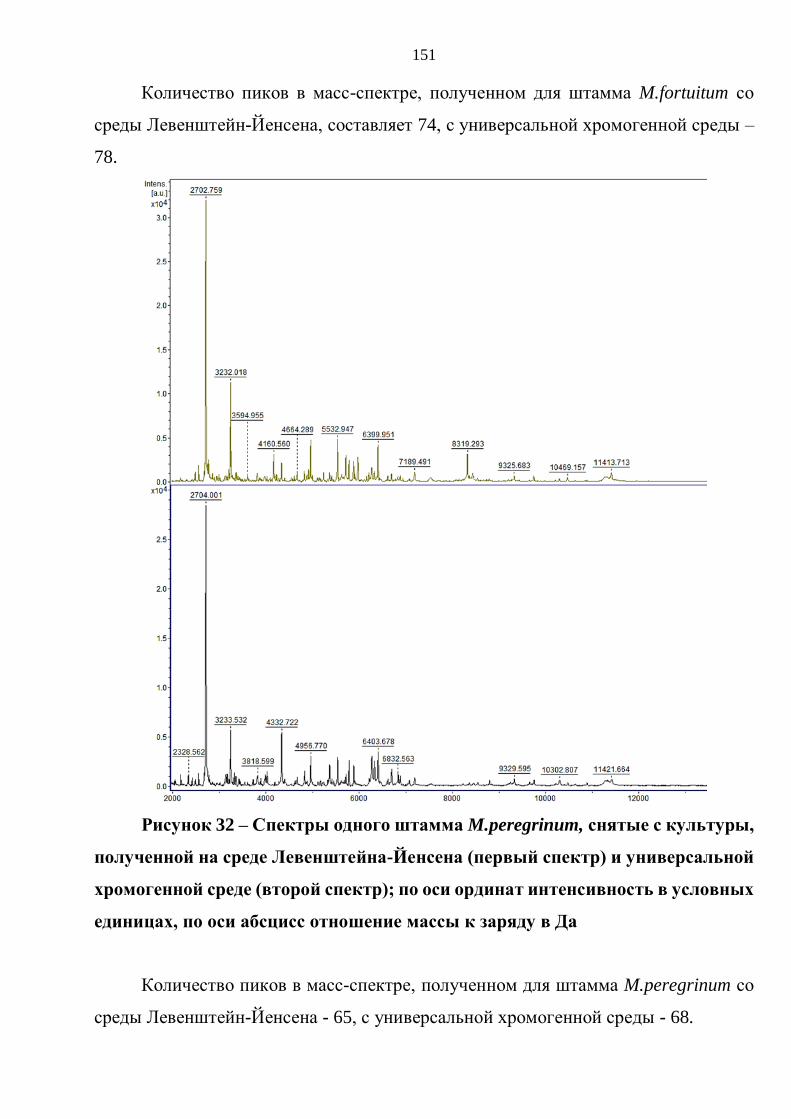
151
Количество пиков в масс-спектре, полученном для штамма M.fortuitum со
среды Левенштейн-Йенсена, составляет 74, с универсальной хромогенной среды –
78.
Рисунок 32 – Спектры одного штамма M.peregrinum, снятые с культуры,
полученной на среде Левенштейна-Йенсена (первый спектр) и универсальной
хромогенной среде (второй спектр); по оси ординат интенсивность в условных
единицах, по оси абсцисс отношение массы к заряду в Да
Количество пиков в масс-спектре, полученном для штамма M.peregrinum со
среды Левенштейн-Йенсена - 65, с универсальной хромогенной среды - 68.
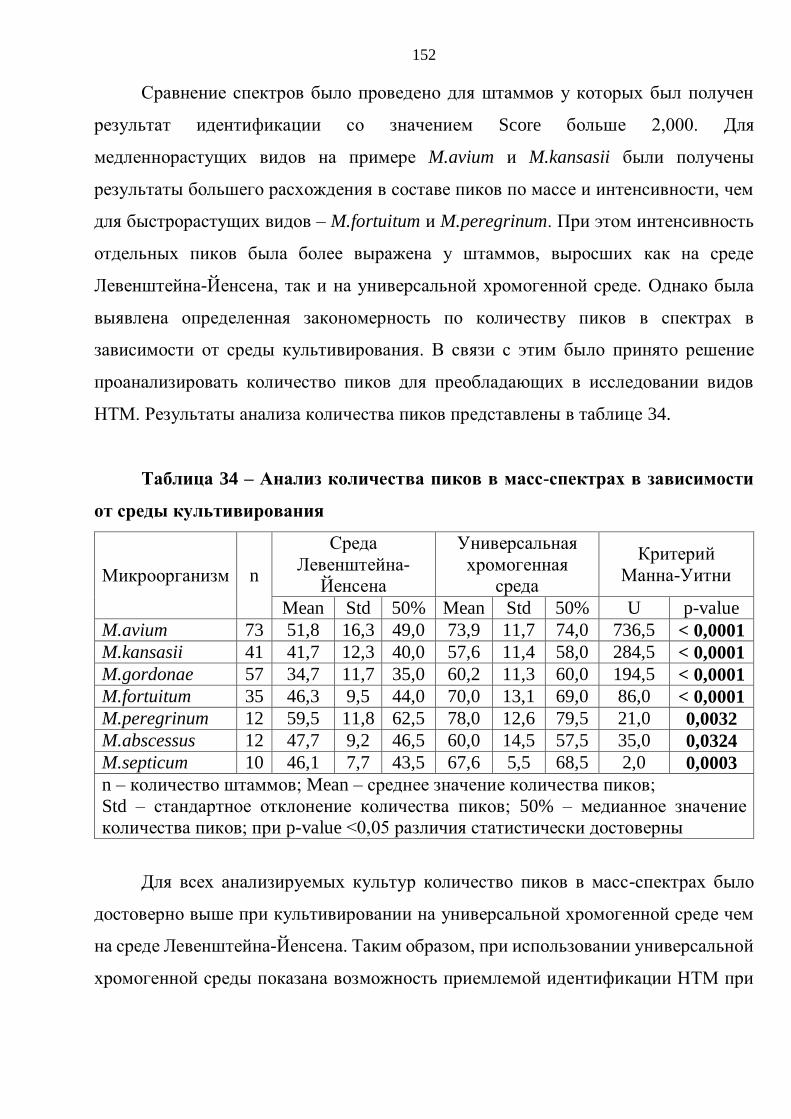
152
Сравнение спектров было проведено для штаммов у которых был получен
результат идентификации со значением Score больше 2,000. Для
медленнорастущих видов на примере M.avium и M.kansasii были получены
результаты большего расхождения в составе пиков по массе и интенсивности, чем
для быстрорастущих видов – M.fortuitum и M.peregrinum. При этом интенсивность
отдельных пиков была более выражена у штаммов, выросших как на среде
Левенштейна-Йенсена, так и на универсальной хромогенной среде. Однако была
выявлена определенная закономерность по количеству пиков в спектрах в
зависимости от среды культивирования. В связи с этим было принято решение
проанализировать количество пиков для преобладающих в исследовании видов
НТМ. Результаты анализа количества пиков представлены в таблице 34.
Таблица 34 – Анализ количества пиков в масс-спектрах в зависимости
от среды культивирования
Микроорганизм n
Среда
Левенштейна-
Йенсена
Универсальная
хромогенная
среда
Критерий
Манна-Уитни
Mean Std 50% Mean Std 50% U p-value
M.avium 73 51,8 16,3 49,0 73,9 11,7 74,0 736,5 < 0,0001
M.kansasii 41 41,7 12,3 40,0 57,6 11,4 58,0 284,5 < 0,0001
M.gordonae 57 34,7 11,7 35,0 60,2 11,3 60,0 194,5 < 0,0001
M.fortuitum 35 46,3 9,5 44,0 70,0 13,1 69,0 86,0 < 0,0001
M.peregrinum 12 59,5 11,8 62,5 78,0 12,6 79,5 21,0 0,0032
M.abscessus 12 47,7 9,2 46,5 60,0 14,5 57,5 35,0 0,0324
M.septicum 10 46,1 7,7 43,5 67,6 5,5 68,5 2,0 0,0003
n – количество штаммов; Mean – среднее значение количества пиков;
Std – стандартное отклонение количества пиков; 50% – медианное значение
количества пиков; при p-value <0,05 различия статистически достоверны
Для всех анализируемых культур количество пиков в масс-спектрах было
достоверно выше при культивировании на универсальной хромогенной среде чем
на среде Левенштейна-Йенсена. Таким образом, при использовании универсальной
хромогенной среды показана возможность приемлемой идентификации НТМ при

153
использовании библиотек спектров, снятых с культур, выросших на средах для
культивирования МТВс.
6.4. Резюме
Идентификация НТМ представляет сложную микробиологическую задачу и
требует разработки, внедрения и валидации новых методов. Использование метода
идентификации НТМ MALDI-ToF масс-спектрометрии демонстрирует более
высокий уровень качества идентификации микроорганизмов, чем применяемый в
большинстве лабораторий противотуберкулезной службы метод ДНК-
гибридизации. Обнаруженные дискордантные результаты идентификации
штаммов с помощью MALDI-ToF спектрометрии в сравнении с секвенированием
по Сэнгеру связаны с рядом причин. Для быстрорастущих НТМ, чаще всего,
расхождение результатов обусловлено принадлежностью выделенных
микроорганизмов к близкородственным видам. Для медленнорастущих НТМ
расхождение результатов, возможно, связано с наличием в образцах микробных
ассоциаций, часть из которых была выявлена при использовании нескольких
методов. Данный факт необходимо учитывать в случае получения противоречивых
результатов при использовании нескольких методов идентификации. Выявленные
ассоциации поднимают важный вопрос о необходимости разработки доступных
методов возможного разделения микстов НТМ с МТВс, между разными видами
НТМ, а также микобактерий с другими микроорганизмами, что, безусловно,
повысит диагностическую ценность микробиологического метода. Однако,
использование в работе с клиническим материалом от пациентов при обследовании
на туберкулез классических сред на яичной основе и среды Миддлбрука
значительно ограничивает такую возможность.
Использование универсальной хромогенной среды для культивирования
НТМ позволяет получить приемлемые результаты идентификации методом
MALDI-ToF масс-спектрометрии при использовании стандартной библиотеки. При
этом результаты видовой идентификации для всех тестируемых штаммов, рост

154
которых был получен на универсальной хромогенной среде, удалось получить
методом расширенного нанесения без необходимости проводить экстракцию
белков муравьиной кислотой. Это позволяет идентифицировать НТМ более
простым и доступным способом.
Идентификация других видов из группы КУППА также оказалась более
эффективной при использовании метода расширенного нанесения. При этом
оказалось, что культуры, рост которых был получен на среде Левенштейна-
Йенсена методом MALDI-ToF масс-спектрометрии, идентифицировались до вида
достоверно менее эффективно, чем при использовании универсальной
хромогенной среды.
Выявленные особенности по количеству пиков в масс-спектрах являются
косвенным признаком более приемлемых условий для культивирования НТМ на
универсальной хромогенной среде, что на наш взгляд объясняет полученные
результаты, при сравнении идентификации методом MALDI-ToF масс-
спектрометрии, выросших культур на среде Левенштейна-Йенсена с хромогенной
средой. Данный результат был получен для всех анализируемых видов НТМ.
Таким образом, использование MALDI-ToF масс-спектрометрии в
совокупности с универсальной хромогенной средой позволяет решать
определенный перечень задач, связанных с получением результатов видовой
идентификации КУППА.

155
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ,
ИЗЛОЖЕННЫМ В ГЛАВЕ 6
Статьи в журналах, рекомендованных ВАК:
1. Лямин, А.В. Нетуберкулезные микобактерии: современные возможности
видовой идентификации / А.В. Лямин, А.М Ковалёв, А.В. Жестков [и др.] //
Клиническая микробиология и антимикробная химиотерапия. – 2017. – Т. 19. – №1.
– С. 11-14.
2. Лямин, А.В. Сравнительный анализ методов идентификации
нетуберкулезных микобактерий, выделенных из клинического материала / А.В.
Лямин, Д.Д. Исматуллин, А.В. Жестков [и др.] // Инфекция и иммунитет. – 2017. –
Т. 7. – №3. – С. 285-291.
Публикации в других изданиях:
1. Лямин, А.В. Сравнение методов в идентификации нетуберкулезных
микобактерий: ДНК-гибридизация, масс-спектрометрия, секвенирование / А.В.
Лямин, Исматуллин Д.Д., А.В. Жестков [и др.] // Сборник трудов Международной
научно-практической конференции «Молекулярная диагностика 2018. – 2018. –
Минск – С. 197-198.
Патенты:
1. Патент РФ № 187404 «Устройство для сбора точечных и мелких колоний
микроорганизмов с поверхности плотных питательных сред» (Бюл. № 7, 2019 г.).
Авторы: Исматуллин Д.Д., Лямин А.В., Тонеев И.Р. [и др.].
2. Патент РФ № 187421 «Устройство для сбора колоний микроорганизмов с
поверхности плотных питательных сред» (Бюл. № 7, 2019 г.). Авторы: Исматуллин
Д.Д., Лямин А.В., Тонеев И.Р. [и др.].
Учебное пособие:
1. Лямин А.В., Жестков А.В., Исматуллин Д.Д. Лабораторная диагностика
микобактериозов: учебное пособие для студентов, Самара: ООО Издательско-
полиграфический комплекс «Право», 2019. – 72 с. ISBN 978-5-6041835-8-8

156
ГЛАВА 7. ФУНДАМЕНТАЛЬНЫЕ И ПРИКЛАДНЫЕ АСПЕКТЫ
ОБОСНОВАНИЯ НОВЫХ ПОДХОДОВ К КУЛЬТИВИРОВАНИЮ И
ИДЕНТИФИКАЦИИ КИСЛОТОУСТОЙЧИВЫХ ПРЕДСТАВИТЕЛЕЙ
ПОРЯДКА ACTINOMYCETALES
7.1. Выбор питательной среды для первичного посева как определяющий
фактор выделения и идентификации медленно и быстро растущих
кислотоустойчивых представителей порядка Actinomycetales в клинической
практике
Известно, что МТВс являются микроорганизмами со сложными
метаболическими процессами, которые требуют значительного количества макро-
и микронутриентов для роста в искусственных условиях, что связано с абсолютным
паразитизмом данной группы бактерий [32, 98]. Исторически сложилось, что НТМ
длительное время культивировались на средах для выделения МТВс, в том числе,
среде Левенштейна-Йенсена и Финн-II [89, 95]. В последние годы все чаще
обсуждается вопрос о возможности выделения НТМ на средах синтетического
состава. Широкое распространение получили различные модификации среды
Миддлбрук, на которых отмечена большая скорость роста как НТМ, так и МТВс
[152, 153, 207, 257, 313].
В нашей работе получены результаты, демонстрирующие то, что
универсальная хромогенная среда является приемлемой для выделения
значительного числа видов НТМ и других КУППА как с точки зрения скорости
роста культуры, так и с точки зрения показателя продуктивности. В связи с этим
возникает вопрос, с чем связаны полученные результаты, если универсальная
хромогенная среда, используемая в работе, значительно отличается по составу от
яичных сред?
Микобактерии, как и другие аэробные актиномицеты в своей
жизнедеятельности ограничены рядом эссенциальных факторов роста, к которым

157
можно отнести, например, железо [25]. Яичные среды содержат высокие
концентрации железа за счет органических форм, в частности, оно входит в состав
овотрансферина [274, 311]. По данным департамента сельского хозяйства США
количество железа в яичной массе составляет 0,0175 мг/г. Такая концентрация
соответствует данным о потребностях основной массы неприхотливых
микроорганизмов в железе от 0,002 мг/мл [25, 290]. Однако при анализе
приведенных данных необходимо учитывать форму железа, в которой оно
представлено в соответствующих веществах. В курином яйце железо связано с
белком, который, предположительно, является одним из основных депо и
способствует снижению доступности железа для микроорганизмов и, таким
образом, становится фактором, лимитирующим их рост и размножение [268, 308].
В то же время, приведенная минимальная концентрация железа рассчитана из
неорганической соли (FeSO4*7H2O), что также может влиять на возможность
использования микобактериями железа в данной форме [105].
Из-за дефицита легкодоступных форм железа в природе микобактерии в
процессе эволюции приобрели способность выхватывать, транспортировать и
конкурировать за данный микроэлемент в ходе своей жизнедеятельности. Речь
идет о способности синтезировать ряд молекул – сидерофоров, функция которых
заключается в связывании и транспортировке молекул железа в бактериальную
клетку [314, 315]. При этом известно, что аффинность сидерофоров микобактерий
к железу выше, чем у трансферрина и других белков [23, 53, 105].
Следует учитывать, что у сапрофитных видов микроорганизмов сидерофоры
направлены на поглощение из окружающей среды железа из малорастворимых и
нерастворимых соединений, а патогенные микроорганизмы чаще используют
сидерофор-опосредованный транспорт железа из металлопротеинов [53, 105]. В
связи с данным фактом возникает вопрос о большей доступности ионов железа в
растворе солей по сравнению с формами железа, связанными трансферринами и
другими аналогичными белками для НТМ и других КУППА. Среди КУППА, за
исключением нескольких видов, основная масса бактерий является сапрофитами,
широко распространенными в окружающей среде. В связи с этим можно

158
предположить, что основные механизмы поглощения железа у них направлены на
нерастворимые неорганические формы. Такой механизм потребления железа
клеткой решает вопрос о его дефиците в микроорганизме, но в тоже время требует
значительных временных и энергетических затрат, что может оказывать влияние
на скорость роста микобактерий на питательных средах.
Различные модификации среды Миддлбрука содержат варианты железа в
виде аммонийного цитрата в концентрации 0,04 г/л [205]. Несмотря на отсутствие
яичной массы, данные среды являются оптимальными для культивирования МТВс
и НТМ. По результатам нашего исследования в бульоне Миддлбрука 7Н9
статистически достоверно лучше растут и другие представители группы КУППА.
Результаты сравнения культивирования НТМ и других КУППА на
универсальной хромогенной среде UriSelect (BioRAD, США) и среде Левенштейна-
Йенсена потребовали анализа состава данных сред для объяснения полученных
закономерностей. Данные биохимического исследования состава сред приведены в
таблице 35.
Таблица 35 – Количественная характеристика компонентов
исследуемых питательных сред
Компонент UriSelect Левенштейна-
Йенсена Финн-II
Миддлбрук
7Н9
Ионизированное
железо, г/л 0,001 0,007 0,006 0,008
Общий белок, г/л 6,6 71,1 73,1 1,1
Альбумин, г/л 0,8 16,3 22,6 1,1
Калий, г/л 0,48 1,13 2,45 0,62
Натрий, г/л 0,75 0,87 1,42 0,69
Хлориды, г/л 0,38 1,13 1,09 0,24
Кальций общий, г/л 0,01 0,11 0,25 0,01
Фосфор, г/л 0,09 0,36 0,57 0,49
Уровень железа, определенный колориметрически, оказался минимальным
для универсальной хромогенной среды в сравнении с классическими средами,
применяемыми для выделения МТВс. Однако в связи с отсутствием данных о

159
форме железа в составе хромогенной среды можно предположить, что ее
доступность для утилизации микобактериями оказалась достаточной для роста
большинства штаммов, включенных в исследование. Этим можно объяснить
отсутствие роста отдельных штаммов M.avium, M.gordonae,
M.chimaera/intracellulare и M.szulgai на универсальной хромогенной среде.
Несмотря на сопоставимые данные по уровню железа в средах на яичной
основе и Миддлбрука, остается открытым вопрос о доступности железа в яичных
средах для роста сапрофитных НТМ и других КУППА, которые участвуют в
патологических процессах у человека, особенно с учетом отсутствия у
микобактерий протеолитических ферментов, повышающих уровень свободного
железа в среде до оптимального для роста и размножения микроорганизмов. С
другой стороны, следует учитывать, что избыток ионов железа может оказывать
ингибирующее действие на рост микроорганизмов, в связи с чем при выявлении
ассоциации двух и более микроорганизмов на яичных средах возможно подавление
роста одного из них, если другой обладает протеолитической активностью [241].
Для оценки возможности использования неорганической формы железа в
качестве ростовой добавки был разработан способ культуральной диагностики
инфицированности туберкулезом, при котором мокрота перед посевом на среду
Левенштейна-Йенсена обрабатывалась 0,04 М оксидом железа в натрий фосфатном
буфере 1 М, рН 6,8 (Патент РФ № 2613366). Данный способ позволил получить
ускоренный рост МТВс на среде Левенштейна-Йенсена, несмотря на изначально
высокие уровни железа в данной среде, что косвенно подтверждает факт различной
степени доступности железа для микроорганизмов в зависимости от формы, в
которой оно добавлено в среду. Неорганическая форма железа, добавленная даже
не в среду, а на этапе пробоподготовки оказала положительное воздействие на
МТВс, что рассматривается нами как элемент адаптации жизнедеятельности
микобактерий при переходе из организма человека в искусственные условия
питательной среды.
На рост сапрофитных микроорганизмов также могут оказывать влияние и
избыточные концентрации определенных компонентов среды. Так в средах на

160
яичной основе было выявлено высокое содержание общего белка, что закономерно
и соответствует составу [95]. При этом в научной литературе обозначена роль
липидов желтка куриного яйца как важного ростового фактора для микобактерий,
но известно, что избыток нативных белков может являться лимитирующим
фактором при выделении сапрофитной флоры [97, 102]. Тем не менее полное
исключение из состава белков также является недопустимым. Полученные нами
результаты роста большинства видов КУППА на универсальной хромогенной
среде и среде Миддлбрука 7Н9, возможно, обусловлены более низкими
концентрациями белка, или наличием белка в более доступной форме для роста
НТМ и других КУППА. Известно, что в состав среды UriSelect входят четыре
пептона, которые в общей совокупности составили концентрацию общего белка
почти в десять раз меньшую, чем в средах на яичной основе, но и большую, чем в
синтетической среде Миддлбрука. Наличие легкодоступных источников
аминокислот в совокупности с микроэлементным составом схожим по основным
компонентам на среду Миддлбрука возможно позволило получить приемлемые
результаты идентификации НТМ методом MALDI-ToF масс-спектрометрии за счет
увеличения количества пиков в спектрах.
Совокупность выявленных фактов о составе универсальной хромогенной
среды UriSelect подтверждает ее использование для выделения КУППА, а
дополнительные преимущества по составу хромогенных субстратов для
обнаружения бактериальных ферментов β-глюкуронидаз, β-галактозидаз и
триптофана позволяют проводить предварительную идентификацию при
первичном посеве и дают потенциал ее широкого использования в
микробиологической практике, оставляя возможность для дальнейшего
совершенствования ее состава с целью выделения большего числа видов НТМ и
других КУППА из клинического материала.

161
7.2. Возможности продленной инкубации для выделения и идентификации
кислотоустойчивых представителей порядка Actinomycetales в рутинной
микробиологической практике
Для оценки возможности использования предложенной методики
продленной инкубации посевов на универсальной хромогенной среде при
температуре 28°С 2018 и 2019 годы, к стандартному микробиологическому
исследованию дополнительно культивировались все чашки с посевами раневого
отделяемого, отделяемого из урогенитального тракта и мокроты от пациентов. В
качестве контрольного был выбран период за 2016 и 2017 годы, когда не
проводилась продленная инкубация посевов. В данное исследование были
включены посевы без признаков роста в стандартных условиях.
7.2.1. Влияние продленной инкубации на агаризованных средах на
возможность выделения кислотоустойчивых представителей порядка
Actinomycetales из клинического материала
За контрольный период было выделено девять случаев выделения
Actinomyces spp., три случая – Mycobacterium spp., два случая Nocardia spp.
Штаммы актиномицет были выделены на кровяном агаре. Всего выделено и
идентифицировано девять штаммов Actinomyces spp. Видовое разнообразие
выделенных актиномицет представлено в таблице 36.

162
Таблица 36 – Структура представителей рода Actinomyces, выделенных
из клинического материала
Вид n Вид клинического
материала
Сопутствующая
микрофлора
Диагноз
A.oris
3 Мокрота
Содержимое абсцесса
Ассоциации с
условно-
патогенной
микрофлорой
Муковисцидоз
ХОБЛ
Паратонзиллярный
абсцесс
A.urogenitalis
2 Моча
Отделяемое из
цервикального канала
Ассоциации с
условно-
патогенной
микрофлорой
Хроническая
почечная
недостаточность
Цервицит
A.neuii 1 Эякулят Монокультура Хронический
простатит
A.europaeus 2 Отделяемое из
брюшной полости
Монокультура Абсцесс
A.turicensis 1 Раневое отделяемое Монокультура Парапроктит
Среди микобактерий было выделено три штамма M.abscessus из мокроты от
пациентов с хронической бронхолегочной патологией. Было выделено два штамма
N.farcinica из крови пациента с деструктивным процессом в правом легком в
отдаленном периоде после трансплантации почки. Все штаммы выделены на
универсальной хромогенной среде. За два года в обычных условиях
культивирования было выделено четырнадцать штаммов представителей порядка
Actinomycetales.
За анализируемый период, с учетом продленной инкубации было выделено
23 штамма Actinomyces spp., рост которых был отмечен также на кровяном агаре.
Видовое разнообразие выделенных актиномицет представлено в таблице 37.
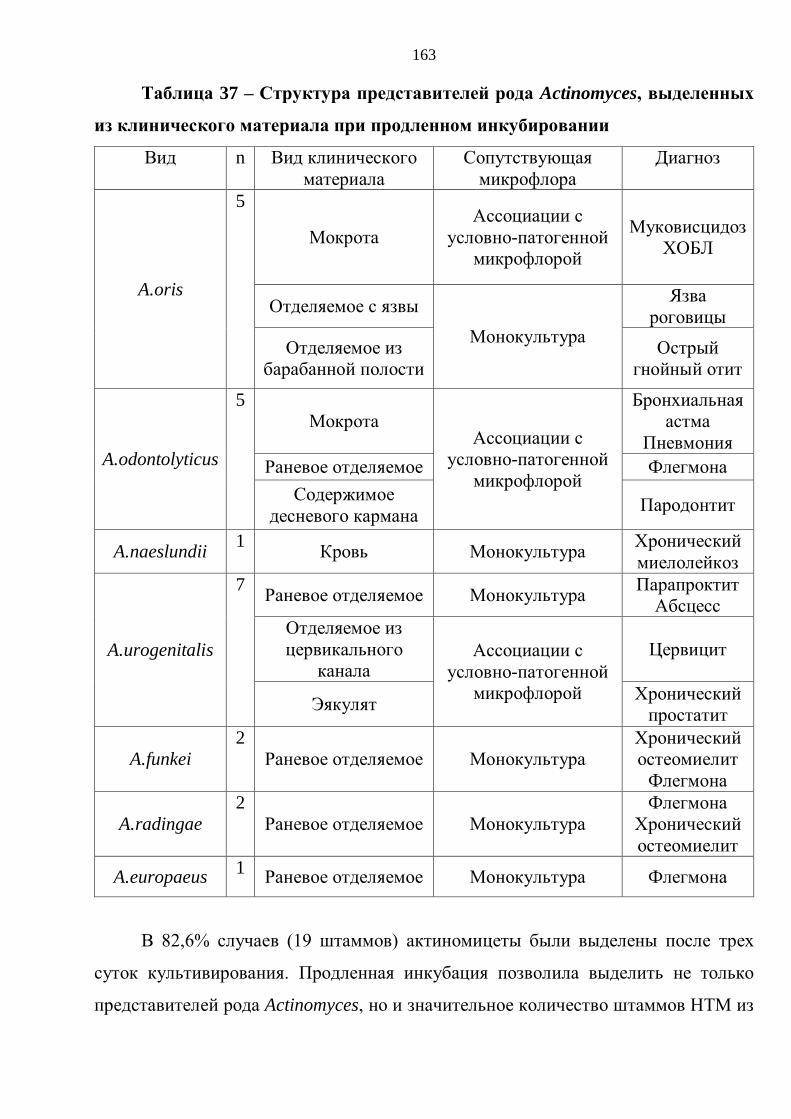
163
Таблица 37 – Структура представителей рода Actinomyces, выделенных
из клинического материала при продленном инкубировании
Вид n Вид клинического
материала
Сопутствующая
микрофлора
Диагноз
A.oris
5
Мокрота
Ассоциации с
условно-патогенной
микрофлорой
Муковисцидоз
ХОБЛ
Отделяемое с язвы
Монокультура
Язва
роговицы
Отделяемое из
барабанной полости
Острый
гнойный отит
A.odontolyticus
5
Мокрота Ассоциации с
условно-патогенной
микрофлорой
Бронхиальная
астма
Пневмония
Раневое отделяемое Флегмона
Содержимое
десневого кармана Пародонтит
A.naeslundii 1
Кровь Монокультура Хронический
миелолейкоз
A.urogenitalis
7 Раневое отделяемое Монокультура
Парапроктит
Абсцесс
Отделяемое из
цервикального
канала Ассоциации с
условно-патогенной
микрофлорой
Цервицит
Эякулят Хронический
простатит
A.funkei
2
Раневое отделяемое Монокультура
Хронический
остеомиелит
Флегмона
A.radingae
2
Раневое отделяемое Монокультура
Флегмона
Хронический
остеомиелит
A.europaeus 1
Раневое отделяемое Монокультура Флегмона
В 82,6% случаев (19 штаммов) актиномицеты были выделены после трех
суток культивирования. Продленная инкубация позволила выделить не только
представителей рода Actinomyces, но и значительное количество штаммов НТМ из

164
различного клинического материала: 42 штамма M.abscessus от пациентов с
муковисцидозом и другой бронхолегочной патологией, один штамм M.avium от
пациента с ХОБЛ; один M.fortuitum из раневого отделяемого от пациента с
абсцессом кожи; один штамм M.monacense от пациента с флегмоной нижней
конечности. Таким образом, всего за два года было выделено 45 штаммов НТМ.
Из других КУППА были выделены по одному штамму: N.cyriacigeorgica в
монокультуре от ВИЧ-инфицированного пациента с пневмонией, N.farcinica и
Rhodococcus globerulus в ассоциации с условно-патогенной микрофлорой от
пациентов с муковисцидозом, Rhodococcus fascians из мокроты от пациента с
хронической бронхолегочной патологией в монокультуре; два штамма
Streptomyces violaceoruber в ассоциации с условно-патогенной микрофлорой из
мокроты пациента с муковисцидозом и один из фрагмента мочеточника при
трансплантации почки; два штамма Streptomyces badius выделены от одной
пациентки с хроническим миелолекозом и деструктивной пневмонией из крови и
бронхоальвеолярного лаважа. Всего выделено восемь штаммов из группы КУППА.
Рост всех штаммов получен на 5-28-е сутки культивирования на универсальной
хромогенной среде, кровяном агаре или селективной среде для выделения
Burkholderia cepacia complex (ВСС).
Для выделения микроорганизмов, описанных в данной части работы, были
разработаны подходы по выбору питательных сред и продлении инкубации
посевов, основные принципы которых приведены в патентах на изобретение РФ
(Патент РФ № 2659155, Патент РФ № 2668406).
7.2.2. Особенности идентификации выделенных из клинического материала
культур с использованием метода MALDI-ToF масс-спектрометрии
Все культуры, описанные в предыдущей главе, были идентифицированы с
использованием метода MALDI-ToF масс-спектрометрии. Штаммы Actinomyces
spp. и Rhodococcus spp. были идентифицированы методом прямого нанесения со

165
среды, на которой были получены колонии. Для других штаммов КУППА не
удалось получить приемлемый результат идентификации до вида во всех случаях.
Результат идентификации зависел от питательной среды, на которой был получен
рост микроорганизмов. Из 48-и штаммов НТМ первичный рост культур был
получен на универсальной хромогенной среде только для десяти (20,8%) штаммов.
Остальные микроорганизмы были выделены на кровяном агаре или селективной
среде для выделения ВСС. Результат идентификации до вида при использовании
метода прямого нанесения удалось получить для десятки штаммов (20,8%)
первично выделенных на универсальной хромогенной среде. Остальные культуры
НТМ удалось идентифицировать до рода (79,2%) с использованием метода прямого
нанесения. С использованием метода расширенного прямого нанесения был
получен результат видовой идентификации для 32-х штаммов НТМ, для шести
штаммов (12,5%) идентификация не была проведена. Применение метода
экстракции муравьиной кислотой не было использовано в данной части
исследования. Для всех штаммов, идентификация которых была ограничена
родовой принадлежностью или не была проведена, апробировали алгоритм
пересева чистой культуры на универсальную хромогенную среду с последующей
инкубацией при температуре 37°С в течение первых суток с последующим
культивированием при температуре 28°С в течение 21-и суток. Все 38 штаммов
НТМ при пересеве на универсальную хромогенную среду дали видимые колонии,
достаточные для проведения идентификации методом прямого нанесения на 2-7-е
сутки культивирования. Все штаммы, рост которых был получен на универсальной
хромогенной среде, идентифицированы методом прямого нанесения до вида.
Для штаммов нокардий (n=4) и стрептомицет (n=4) идентификация до рода
была проведена вне зависимости от среды первичного выделения. Однако
приемлемый результат видовой идентификации был получен только при пересеве
полученных культур на универсальную хромогенную среду или в случае
первоначального роста на ней. Следует учесть, что в отличие от НТМ, нокардии и
стрептомицеты были успешно идентифицированы до вида на 3-4 сутки с момента
появления видимого роста колоний.

166
7.3. Протеомный анализ белковых спектров кислотоустойчивых
представителей порядка Actinomycetales как элемент субтипирования
штаммов, выделенных от одного пациента
MALDI-ToF масс-спектрометрия, кроме видовой идентификации, позволяет
использовать разнообразные подходы для анализа белковых спектров, что, по
нашему мнению, может быть использовано в рутинной микробиологической
практике в лабораториях, оснащенных данным оборудованием. НТМ, также, как и
другие виды КУППА, являются широко распространенными в окружающей среде
микроорганизмами, в связи с чем при проведении диагностики обусловленных ими
заболеваний часто встает вопрос о верификации их клинического значения. Данная
задача частично решается в случае выделения НТМ или других актиномицет из в
норме стерильных локусов, однако для образцов, полученных из дыхательных
путей, мочи или со слизистых оболочек верхних дыхательных путей вопрос
остается не решенным. В соответствии с рекомендациями Американского
торакального общества диагноз микобактериоз устанавливается в случае если
имеется соответствующая клиническая картина, а из мокроты выделяется один вид
НТМ при трехкратном посеве мокроты с интервалом в одну неделю. Такие
рекомендации разработаны по причине высокого риска контаминации материала,
полученного от пациента НТМ из окружающей среды. Следует учитывать, что у
такого подхода к диагностике микобактериозов имеются значительные
расхождения с лабораторной диагностикой туберкулеза. С учетом факта
определенного сходства клинической картины и общих организационных этапов
проведения обследования в системе противотуберкулезной службы нередко НТМ
выделяются при подозрении на туберкулез, а, как следствие, культура НТМ может
быть выделена из нескольких образцов, собранных в один день, что противоречит
критериям постановки диагноза микобактериоз с одной стороны, а с другой,
требует проведения ряда дополнительных тестов, что увеличивает время до
постановки диагноза.
Известно, что микроорганизмы, в норме заселяющие экологические ниши
природных объектов, чаще всего представляют собой сложные гетерогенные

167
популяции, характеризующиеся различными возможностями приспособления к
тем или иным условиям в пределах одного объекта внешней среды. В организме
человека такие процессы возможны при длительно протекающих инфекционных
процессах, примером которых может служить муковисцидоз. Для определения
возможности использования масс-спектрометрии в качестве дополнительного
инструмента в подтверждении диагноза микобактериоз было принято решение
проанализировать белковые профили четырех штаммов НТМ, выделенных от двух
пациентов. Первые два штамма M.fortuitum выделены из двух образцов мокроты,
полученной от одного пациента, с клиническими и рентгенологическими
признаками микобактериоза легких. Другие два штамма M.peregrinum были
выделены из одного образца мокроты, у пациента без клинических признаков
микобактериоза. Рост обоих штаммов M.peregrinum был выявлен при пересеве
культуры, полученной на среде Левенштейна-Йенсена на универсальную
хромогенную среду, культуральные свойства представлены на рисунке 33.
Рисунок 33 – Гетерогенная популяция M.peregrinum на универсальной
хромогенной среде
Один тип колоний характеризовался неправильной формой с приподнятым
окрашенным центром и бесцветной периферией. Второй тип колоний был
представлен классической S-формой, окрашивание которой в светло-голубой цвет
происходило на 3-5-е сутки культивирования. При пересеве данных штаммов на

168
универсальную хромогенную среду выявленные морфотипы колоний сохранили
свои свойства. Дальнейшей диссоциации культуры не было выявлено.
При анализе культуральных свойств для штаммов M.fortuitum, выделенных
из двух разных образцов, не было выявлено признаков гетерогенности при
культивировании на универсальной хромогенной среде. Все колонии были S-
формы, характерной для быстрорастущих НТМ голубого цвета. При продленной
инкубации диссоциации штаммов не выявлено. Культуральные свойства одного из
штаммов представлены на рисунке 34.
Рисунок 34 – Однородная популяция M.fortuitum на универсальной
хромогенной среде
Для всех четырех штаммов была проведена пробоподготовка методом
экстракции с муравьиной кислотой. Анализ полученных спектров проводили с
помощью программ flexAnalysis 3.0 и MALDI Biotyper 3.0 Offline Classification.
Полученные спектры интересующих нас штаммов анализировались
визуально в программе flexAnalysis 3.0 путем выявления отдельных пиков белков
и создания листа выявленных масс белков. Сравнение спектров проводили с
использованием инструментов статистического обсчета и визуализации в
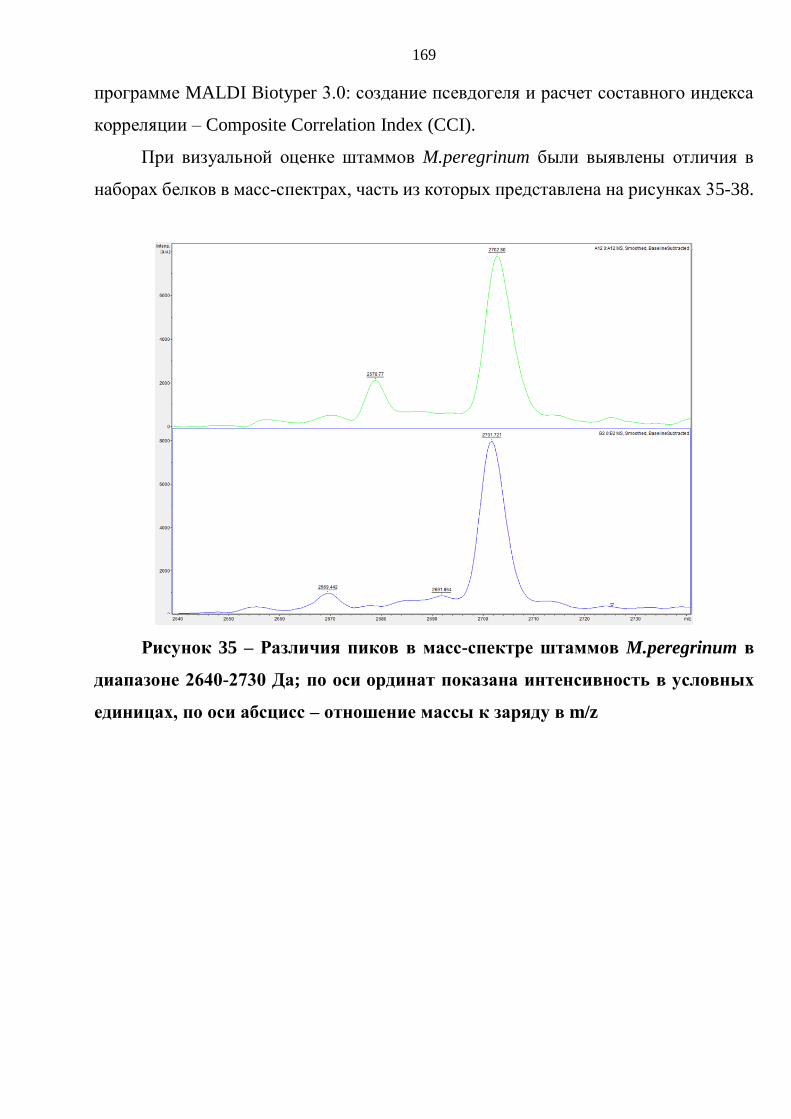
169
программе MALDI Biotyper 3.0: создание псевдогеля и расчет составного индекса
корреляции – Composite Correlation Index (CCI).
При визуальной оценке штаммов M.peregrinum были выявлены отличия в
наборах белков в масс-спектрах, часть из которых представлена на рисунках 35-38.
Рисунок 35 – Различия пиков в масс-спектре штаммов M.peregrinum в
диапазоне 2640-2730 Да; по оси ординат показана интенсивность в условных
единицах, по оси абсцисс – отношение массы к заряду в m/z

170
Рисунок 36 – Различия пиков в масс-спектре штаммов M.peregrinum в
диапазоне 3450-3850 Да; по оси ординат показана интенсивность в условных
единицах, по оси абсцисс – отношение массы к заряду в m/z
Рисунок 37 – Различия пиков в масс-спектре штаммов M.peregrinum в
диапазоне 4100-4500 Да; по оси ординат показана интенсивность в условных
единицах, по оси абсцисс – отношение массы к заряду в m/z

171
Рисунок 38 – Различия пиков в масс-спектре штаммов M.peregrinum в
диапазоне 7050-7450 Да; по оси ординат показана интенсивность в условных
единицах, по оси абсцисс – отношение массы к заряду в m/z
В целом для двух штаммов M.peregrinum были выявлены различия в 37-и
пиках; тридцать пиков были одинаковыми в масс-спектрах обоих штаммов. У
первого штамма дополнительно было определено 26-ть уникальных пиков, у
второго – 11. Таким образом, процент уникальных белков в масс-спектрах для
первого штамма составил 46,4%, для второго – 26,8%.
Для подтверждения полученных результатов были получены изображения
спектров в виде псевдогелей в GelView, результаты которых приведены на рисунке
39.

172
Рисунок 39 – Спектры сравниваемых штаммов M.peregrinum в виде
псевдогелей, по оси ординат отмечены штаммы, по оси абсцисс – отношение
массы к заряду в m/z
Дополнительно был проведен сравнительный протеомный анализ
полученных спектров с использованием составного индекса корреляции для
анализируемых штаммов и двух штаммов M.peregrinum, выделенных из различных
источников в разное время. При анализе результатов можно выявить степень
совпадения штаммов от полного совпадения – темно-красный цвет, до полного
отсутствия совпадения штаммов – темно-синий цвет.
Полученные результаты демонстрируют низкий уровень совпадения двух
штаммов M.peregrinum, выделенных из одного клинического материала, что
подтверждает гетерогенность популяции данного микроорганизма. CCI Score для
первого и второго штаммов составил 0,702; для трех и четвертого – 0,779. При этом
значение CCI Score равное 1 свидетельствует о полной корреляции между
спектрами, а значение, равное 0, – о полном несовпадении спектров. Таким

173
образом, между штаммами, выделенными из одного образца, меньше совпадений
по составу белков, чем между контрольными штаммами. Полученная CCI матрица
приведена на рисунке 40.
Рисунок 40 – CCI матрица в виде «тепловой карты», где 1 и 2 –
анализируемые штаммы M.peregrinum, 3 и 4 – контрольные штаммы
M.peregrinum
При визуальной оценке штаммов M.fortuitum были выявлены единичные
отличия в масс-спектрах: основная часть пиков имела практически полное
совпадение, что представлено в виде частного примера на рисунке 41.

174
Рисунок 41 – Совпадения пиков в масс-спектре штаммов M.fortuitum в
диапазоне 3000-3400 Да; по оси ординат показана интенсивность в условных
единицах, по оси абсцисс – отношение массы к заряду в m/z
В целом для двух штаммов M.fortuitum были выявлены различия в 11-и пиках
при полном совпадении 51-го пика в масс-спектрах. У первого штамма
дополнительно было определено четыре пика, у второго – семь. Таким образом,
несовпадения пиков в масс-листах для первого штамма составили 7,3%, для
второго – 12,1%.
Для подтверждения полученных результатов были получены изображения
спектров в виде псевдогелей в Gel View, результаты которого приведены на
рисунке 42.

175
Рисунок 42 – Спектры сравниваемых штаммов M.fortuitum в виде
псевдогелей, по оси ординат отмечены штаммы, по оси абсцисс – отношение
массы к заряду в m/z
Также, как и для штаммов M.peregrinum, для штаммов M.fortuitum были
получены результаты составного индекса корреляции, которые
продемонстрировали высокий уровень совпадения двух штаммов M.fortuitum,
выделенных из клинического материала от одного пациента, что подтверждает
схожесть штаммов, выделенных в разное время, но из одного источника. CCI Score
для анализируемых штаммов составил 0,8516; для 1 и 2 – 0,6986.
Сравнительный протеомный анализ полученных спектров с использованием
составного индекса корреляции, для анализируемых штаммов, и двух штаммов
M.fortuitum, выделенных из различных источников в разное время, приведен в виде
CCI матрицы на рисунке 43.

176
Рисунок 43 – CCI матрица в виде «тепловой карты», где 1 и 2 –
контрольные штаммы M.fortuitum, 3 и 4 – анализируемые штаммы
M.fortuitum.
Выявленные сходства в спектрах штаммов M.fortuitum, выделенных от
одного пациента в совокупности с отсутствием признаков гетерогенности
популяции по культуральным свойствам можно рассматривать как фактор,
подтверждающий один источник выделения и высокую вероятность участия
данного микроорганизма в патологическом процессе.
7.4. Резюме
Выявленные в ходе работы особенности состава сред для культивирования
НТМ и других КУППА позволили сделать предположение о роли железа как
лимитирующего, а нативных белков в качестве потенциально ингибирующего

177
факторов, которые необходимо учитывать при выделении актиномицет из
клинического материала. Данный факт подтверждается результатами, которые
были получены при оценке влияния выбора питательной среды на выделение
КУППА при первичных посевах, а также при сравнении культуральных свойств
при использовании среды Левенштейна-Йенсена и универсальной хромогенной
среды.
Небольшой объем выборки, описанной в данной части исследования, не
позволяет провести полноценную статистическую обработку полученного
материала. Все примеры, за исключением выделения M.abscessus из дыхательных
путей, представлены единичными случаями. Однако и по данным,
опубликованным в научной литературе на текущий момент, основная информация
ограничена описанием клинических примеров и случаев выделения нокардий,
родококков или редких видов НТМ из материала от пациентов с различной
патологией. Так, некоторые виды актиномицет, выделенных за четыре года
наблюдений, описаны как комменсалы слизистых оболочек полости рта (A.oris,
A.naeslundii), однако в работе один штамм A.naeslundii был выделен из крови у
пациента с онкогематологической патологией [225]. Другие виды описаны в
качестве не только сапрофитной микрофлоры, но и этиологической причины
развития генерализованных форм инфекции, что подтверждается и нашими
наблюдениями: A.urogenitalis, A.odontolyticus, A.neuii [123, 231, 272]. Также были
выделены и идентифицированы виды, для которых единичные случаи
этиологического значения были описаны за последние 5-10-ть лет: A.europaeus,
A.turicensis, A.funkei, A.radingae [122, 130, 298].
R. fascians и R. globerulus не были выделены в качестве микроорганизмов с
доказанным клиническим значением, однако в научной литературе приводятся
данные об их этиологической роли в нескольких примерах [292].
Стрептомицеты являются крайне редкими возбудителями патологии
человека [143]. Однако единичный случай верифицированного клинического
значения штамма S.badius из двух в норме стерильных локусов у пациента с
онкогематологической патологией является частным примером возможного

178
участия данной группы микроорганизмов в качестве этиологических агентов у
пациентов из групп риска по развитию оппортунистических инфекций.
Таким образом, полученные результаты позволяют расширить возможности
выделения редких видов КУППА, которые сводятся к методике продленной
инкубации и идентификации их с использованием универсальной хромогенной
среды. При этом если лаборатория оснащена MALDI-ToF масс-спектрометром, то
выделение и идентификация данной группы микроорганизмов не требует
значительных изменений в работе лаборатории, внедрения сложных методов
идентификации или приобретения дорогостоящих реактивов.
Использование MALDI-ToF масс-спектрометрии в совокупности с
культивированием НТМ на универсальной хромогенной среде позволяет
проводить протеомный анализ выделенных культур, что может быть использовано
как дополнительный критерий определения клинического значения у пациентов в
случае выделения НТМ и других КУППА из в норме не стерильных локусов.

179
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ,
ИЗЛОЖЕННЫМ В ГЛАВЕ 7
Статьи в журналах, рекомендованных ВАК:
1. Лямин, А.В. Железо как эссенциальный фактор роста микобактерий / А.В.
Лямин, А.В. Халиулин, Д.Д. Исматуллин [и др.] // Известия Самарского научного
центра Российской академии наук. – 2016. – Т. 18. – №5(2). – С. 320-327.
2. Козлов, А.В. Микобактериозы у пациентов с муковисцидозом: причина
или следствие микроэкологических изменений в бронхолегочной системе / А.В.
Козлов, А.В. Лямин, О.В. Кондратенко [и др.] // Астраханский медицинский
журнал. – 2020. – №1. – С. 57-65.
Патенты:
1. Патент РФ № 2613366 «Способ культуральной диагностики
инфицированности туберкулезом» (Бюл. №8, 2017 г.). Авторы: Гильмиярова Ф.Н.,
Радомская В.М., Родионова Ю.Д. [и др.].
2. Патент РФ № 2659155 «Способ сбора и первичного посева жидкости
назального лаважа от пациентов с муковисцидозом для микробиологического
исследования» (Бюл. № 19, 2018 г.). Авторы: Кондратенко О.В., Лямин А.В.,
Медведева Е.Д. [и др.].
3. Патент РФ № 2668406 «Способ первичного посева биоматериала,
выделенного из нижних дыхательных путей пациентов с муковисцидозом» (Бюл.
№ 28, 2018 г.). Авторы: Кондратенко О.В., Лямин А.В., Исматуллин Д.Д.
Учебные пособия:
1. Лямин А.В., Жестков А.В., Исматуллин Д.Д. Лабораторная диагностика
микобактериозов: учебное пособие для студентов, Самара: ООО «Издательско-
полиграфический комплекс «Право», 2019. – 72 с. ISBN 978-5-6041835-8-8
2. Руководство по микробиологической диагностике инфекций дыхательных
путей у пациентов с муковисцидозом / С.В. Поликарпова, С.В. Жилина, О.В.
Кондратенко [и др.], М.–Тверь: ООО «Издательство «Триада», 2019. – 128 с. ISBN
978-5-94789-905-4

180
ЗАКЛЮЧЕНИЕ
Рост числа пациентов с иммуносупрессией различной этиологии является
актуальной проблемой современной медицины и общества в целом [7, 8, 113].
Внедрение новых высокотехнологичных методов оказания медицинской помощи
неизбежно влияет на структуру возбудителей оппортунистических инфекций и их
широкое распространение среди пациентов из групп риска, что косвенно
увеличивает вероятность все большего распространения ранее редких
микроорганизмов в популяции в целом [18, 56, 66, 68, 87, 300].
Исторически сложилась парадигма, определяющая подходы к новым
заболеваниям, которую наглядно можно продемонстрировать на примере
инфекций. Процесс изучения любой инфекционной патологии получает
максимальный стимул с момента открытия этиологической причины,
представленной конкретным возбудителем или группой микроорганизмов. Именно
с момента разработки приемлемых в рутинной практике лабораторий методов
культивирования и идентификации патогенного или условно-патогенного
микроорганизма начинается активный процесс диагностики, который закономерно
влечет за собой разработку стандартов терапии и профилактики инфекционного
заболевания.
Таким образом, микробиологическая диагностика становится краеугольным
камнем, определяющим развитие целых направлений в медицине. Примером
такого явления можно назвать выделение отдельной специальности – фтизиатрии,
развитие которой было связано с доказательством этиологической роли MTBc в
качестве возбудителя туберкулеза. Следствием данных процессов стало
формирование отдельного направления в классической микробиологии, целью
которого является выделение МТВс, изучение особенностей их культивирования и
методов идентификации. Если учитывать необходимость создания целого ряда
условий, которые обеспечивают эффективную жизнедеятельность МТВс для

181
успешной диагностики, то это абсолютно оправдано. Но такое разделение чревато
и определенными сложностями в развитии микробиологии как целостной науки.
Результатом этого процесса стал определенный микробиологический парадокс: в
лабораториях противотуберкулезной службы сформировался полноценный
комплекс методов, позволяющих эффективно выделять медленнорастущие,
сложнокультивируемые микроорганизмы, но практически отсутствует
возможность идентификации возбудителей других инфекционных процессов. В
противовес этому в лабораториях, не профильных по туберкулезу, приоритетным
направлением стала работа с быстрорастущей микрофлорой, идентификация
которой, наряду с редкими микроорганизмами вышла на новый уровень с
внедрением в практическую работу метода MALDI-ToF масс-спектрометрии. При
этом для обоих направлений микробиологической диагностики актуальным
является вопрос о сокращении сроков исследований, что неизбежно приводит к
получению ложноотрицательных результатов в отношении выделения целого ряда
медленнорастущих микроорганизмов [93, 115, 116, 125, 232].
В сложившихся условиях возникает закономерный вопрос: какое
направление практической бактериологии должно взять на себя работу с
микроорганизмами, которые по своим свойствам находятся на стыке деятельности
двух этих взаимоисключающих по функциям лабораторий? Кислотоустойчивые
актиномицеты – частный пример таких микроорганизмов. Особенности их
метаболизма, морфологические и культуральные свойства, новые принципы
классификации делают данную группу прокариот интересным объектом научного
поиска, а всевозрастающая роль их в качестве возбудителей заболеваний человека
ставит конкретные практические задачи.
Частично этот вопрос решен в отношении НТМ, исторически данная группа
КУППА была передана для работы в лаборатории противотуберкулезной службы,
в то время как остальные кислотоустойчивые микроорганизмы были отнесены к
контаминантам, априори не имеющим клинического значения [89]. Однако в
последние годы микобактериология как раздел микробиологии, в интересы
которого входит изучение представителей семейства Mycobacteriaceae, сделала

182
значительный научный прорыв, обусловленный открытием новых видов НТМ для
которых описано доказанное и возможное клиническое значение [143, 193, 248,
249, 252, 253, 256, 306]. Этот процесс получил свою максимальную
интенсификацию за последние 10-20 лет. Его следствием стал пересмотр
классификации данных микроорганизмов, выделение новых родов, разработка и
внедрение новых методов идентификации. Параллельно с микобактериями, для
других микроорганизмов из группы КУППА были получены и накоплены
аналогичные сведения [139, 141, 213, 294].
Результатом анализа современных знаний о микобактериях стал пересмотр
практических вопросов терапии пациентов с микобактериозами с передачей их из
сферы профессиональной деятельности врачей-фтизиатров в пульмонологию,
инфекционные болезни, хирургию, в зависимости от формы инфекционного
процесса, который был вызван НТМ. При этом микробиологическая диагностика
осталась в большей степени прерогативой лабораторий противотуберкулезной
службы. В связи с этим возникает ряд вопросов. Насколько широко
распространены микроорганизмы из группы КУППА в структуре
контаминирующей микрофлоры при обследовании на туберкулез? Есть ли риски
неточной идентификации НТМ и других микроорганизмов из группы КУППА в
случае выявления роста, характерного для микобактерий?
При исследовании микробного состава, выделенного из 865-и проб
различного клинического материала с признаками роста контаминирующей
микрофлоры, было выделено 1 093 штамма микроорганизмов, идентификацию
которых проводили методом MALDI-ToF масс-спектрометрии.
В материале, представленном промывными водами желудка, плевральной
жидкостью, спинномозговой жидкостью, раневом отделяемом и аутопсийном
материалом было выделено 54-е штамма микроорганизмов, из которых пять
оказались представителями КУППА, что составило 9,3% от общего числа
микроорганизмов. В трех случаях были выделены НТМ с атипичными признаками
роста на плотных питательных средах: отсутствием кислотоустойчивости при

183
окраске по методу Циль-Нильсена, образованием слизистых ярко-желтых колоний
на вторые-третьи сутки культивирования.
Анализ структуры микрофлоры, выделенной из мокроты и мочи как
доминирующих видов клинического материала показал определенные
статистически достоверные закономерности: преобладание микромицет в целом и
представителей родов Candida и Aspergillus в мокроте, коринебактерий и
энтерококков, энтеробактерий в целом и E.coli, в частности, в моче. Частота
выделения КУППА не имела статистически достоверных различий по отдельным
родам, но данная группа микроорганизмов достоверно чаще выделялась из
мокроты (p=0,037).
В целом, в структуре контаминирующей микрофлоры было выделено 29-ть
видов микроорганизмов из группы КУППА. Представители родов Gordonia,
Rhodococcus, Tsukamurella были выделены только из мокроты. Нокардии –
преимущественно из мокроты, за исключением одного штамма,
идентифицированного в промывных водах желудка. Следует отметить, что в семи
случаях из десяти нокардии выделены в монокультуре, что можно рассматривать в
качестве критерия относительной клинической значимости.
В поставленные задачи нашей работы не входило сопоставление выделенной
микрофлоры с клинической картиной у отдельных пациентов, но по данным
литературы все вышеперечисленные микроорганизмы из группы КУППА имеют
доказанное клиническое значение у иммунокомпрометированных пациентов и
могут вызвать патологические процессы, схожие с туберкулезом [73, 220].
Дополнительно в структуре контаминирующей микрофлоры были выделены
классические сапрофитные виды КУППА: представители родов Brevibacterium и
Cellulosimicrobium.
Максимальное видовое разнообразие среди кислотоустойчивых актиномицет
было отмечено для двух родов: Streptomyces и Mycobacterium. Клиническое
значение стрептомицет является предметом научных дискуссий последних лет. В
отечественной литературе практически отсутствует информация об их
клиническом значении, а также о возможности выделения данной группы

184
микроорганизмов из клинического материала. С другой стороны, в зарубежной
научной литературе в последние годы все чаще встречаются публикации в которых
приводятся данные не только о единичных клинических случаях выделения
стрептомицет из клинического материала, но и крупные обзоры таких случаев
[143].
Всего в исследовании было выделено шесть видов рода Streptomyces,
доминирующими из которых оказались S.phaeochromogenes и S.violaceoruber,
которые были выделены соответственно в 12-и и 10-и случаях из 32-х культур
стрептомицет, идентифицированных в структуре контаминирующей микрофлоры.
Отдельно следует остановиться на НТМ, которые были выделены в качестве
представителей контаминирующей микрофлоры. Для большинства штаммов
микобактерий были описаны культуральные признаки, нетипичные для НТМ.
Практически у всех штаммов обнаружены схожие с классической
контаминирующей микрофлорой свойства: ускоренный рост на плотных яичных
средах, выраженная пигментация, мукоидный тип колоний, изменение цвета
среды. В случае выделения в составе ассоциаций с другими микроорганизмами
было выявлено разжижение яичных сред. Всего было выделено и
идентифицировано 36-ть штаммов НТМ.
Среди НТМ, выделенных в первой части исследования, преобладающей
группой оказались виды НТМ, имеющие доказанное или потенциальное
клиническое значение, которые составили 77,8% от всех штаммов микобактерий,
выделенных из контаминирующей микрофлоры.
Анализ полученных данных дает четкую картину возможности выделения
КУППА из клинического материала с признаками роста контаминирующей
микрофлоры при обследовании на туберкулез. При этом, если условно рассчитать
долю кислотоустойчивых аэробных актиномицет, имеющих клиническое значение
по литературным данным, то она составляет 4,6% (50 штаммов) от всех
выделенных в первой части исследования микроорганизмов, а для всех
кислотоустойчивых актиномицет вне зависимости от их потенциального
клинического значения – 9,3% (102 штамма). При этом выбор среды для

185
первичного посева статистически достоверно влияет на частоту выделения
КУППА при использовании бульона Миддлбрук 7Н9 с селективной добавкой, что
требует разработки и внедрения алгоритмов их идентификации и оценки
клинического значения. Особенно актуальными полученные результаты
становятся с учетом достоверного снижения частоты высева на жидкой
питательной среде истинных контаминантов – некоторых микромицет (для
Aspergillus spp., р=0,006, для Candida spp., р<0,001), Escherichia spp. (р<0,001),
Klebsiella spp. (0,007) и неферментирующих грамотрицательных бактерий
(Pseudomonas spp., р=0,003).
Полученные результаты позволяют сделать заключение о необходимости
проведения видовой идентификации «контаминантов», выделенных, в первую
очередь, на жидкой питательной среде Миддлбрук 7Н9, с целью высева из
клинического материала НТМ с атипичными признаками роста и других
микроорганизмов из группы КУППА, особенно в случае отсутствия
положительного результата на МТВс, а также в случае получения биологического
материала от пациентов из групп риска по развитию заболеваний, вызванных
кислотоустойчивыми аэробными актиномицетами.
Для анализа возможности идентификации НТМ в случае выявления
признаков роста, которые для них характерны, была проведена вторая часть
исследования. В нее были отобраны 316-ть проб посевов клинического материала,
в которых были выявлены признаки роста НТМ. Для первичной идентификации
использовали метод ДНК-гибридизации, с помощью которого было
идентифицировано 188 штаммов НТМ, что составило 59,5% от общего числа
культур, включенных в исследование. Для 22-х штаммов была определена
принадлежность к роду Mycobacterium, 39-ть штаммов были идентифицированы
как микроорганизмы с высоким содержанием ГЦ. Общее количество
таксономических единиц, которые были идентифицированы методом ДНК-
гибридизации, составило 11 видов.
При реидентификации описанных выше культур методом MALDI-ToF масс-
спектрометрии было идентифицировано значительно большее число штаммов

186
микроорганизмов – 346. В связи с тем, что работа по реидентификации
микроорганизмов проводилась в микробиологической лаборатории, не
профильной по работе с МТВс, в случае отсутствия приемлемого результата
идентификации было принято решение о пересеве проб, содержащих
потенциальные микробные ассоциации на агаризованные среды с последующей
идентификацией всех выросших культур. Данный этап невозможно выполнить в
условиях работы лаборатории противотуберкулезной службы. В выявленных
трехкомпонентных ассоциациях были идентифицированы пять видов из группы
КУППА: M.pseudoshottsii + M.szulgai + S.lavendulae. Вторая – M.malmoense +
Rhodococcus triatomae + C.albicans.
Дополнительно были выявлены двухкомпонентные ассоциации, состоящие
из двух видов НТМ, которые составили 4,1% от всех проб. Полученные данные по
межвидовым ассоциациям НТМ являются важным элементом, определяющим
возможные ошибки в этиологической расшифровке при постановке диагноза
микобактериоз. Также было выявлено 12-ть проб (3,8% от общего числа проб), в
которых были выявлены ассоциации, состоящие из НТМ и микроорганизмов из
других групп. Следует отметить, что во всех 27-и пробах, в которых были
выявлены ассоциации, содержащие НТМ, не было визуальных признаков роста
двух или трех культур, что необходимо учитывать при работе с посевами, в
которых выявлены признаки роста НТМ, но не удалось получить приемлемый
результат видовой идентификации с использованием метода ДНК-гибридизации.
Особенно важен анализ полученных результатов с точки зрения клинической роли
таких полимикробных ассоциаций. В задачи нашего исследования не входило
решение данного вопроса, но он, безусловно, требует более пристального изучения
совместно с клиницистами.
Методом MALDI-ToF масс-спектрометрии было идентифицировано 27-м
видов микроорганизмов, при этом микобактерии составили 283-и штамма (82,0%
от всех идентифицированных культур). Кроме НТМ в исследовании было
выделено два штамма M.bovis, которые не были идентифицированы с
использованием метода ДНК-гибридизации. Среди выделенных НТМ были

187
идентифицированы редкие виды, выделенные в единичных случаях, такие как:
M.marseillense, M.scrofulaceum, M.xenopi, M.interjectum, M.bohemicum, имеющие
доказанное клиническое значение; M.monacense, M.farcinogenes, M.iranicum,
M.seoulense – виды НТМ, для которых описаны единичные случаи выделения из
материала от пациентов; M.gastri, M.flavescens – непатогенные для человека виды.
Анализ видовой структуры НТМ выявил, что каждая пятая
идентифицированная культура микобактерий не имеет, по данным литературы,
клинического значения, что, безусловно, играет важную роль с точки зрения
дальнейших действий в отношении пациентов: активный поиск причин развития
патологического процесса; снятие предполагаемого диагноза микобактериоз, а в
некоторых случаях анализ возможной роли выделенной непатогенной
микобактерии в качестве этиологического агента как казуистического, но
возможный вариант, позволяющий получить новые данные о роли редких и ранее
непатогенных видов НТМ в развитии патологии у человека.
Кроме НТМ во второй части исследования были выделены другие
представители кислотоустойчивых актиномицет. В частности, было
идентифицировано двадцать штаммов, принадлежащих к родам Brevibacillus,
Gordonia, Nocardia, Rhodococcus, Streptomyces, Tsukamurella; для которых по
данным литературы у 75,0% описано возможное или доказанное клиническое
значение [68]. Данные культуры могут быть идентифицированы с помощью метода
ДНК-гибридизации, применяемого в лабораториях противотуберкулезной службы
только до уровня микроорганизмов с высоким содержанием ГЦ, но дальнейшие
алгоритмы работы с ними не разработаны, видовой идентификации для них не
проводится, и, как следствие, априори исключается возможность постановки
соответствующего диагноза в случае их клинического значения. Важно понимать,
что в случае отрицательного результата при работе с такими микроорганизмами в
лабораториях противотуберкулезной службы выделение их в обычных
микробиологических лабораториях также является маловероятным, так как они,
как правило дают замедленный рост, выявление которого без дополнительных

188
методов культивирования становится невозможным, особенно в условиях
тенденции к сокращению сроков микробиологических исследований.
Для выявления возможных причин расхождения результатов
идентификации, полученных методом ДНК-гибридизации и MALDI-ToF масс-
спектрометрии, было принято решение о проведении сравнения полученных
результатов. Максимальное количество расхождений было выявлено для культур,
первоначально идентифицированных как M.fortuitum (51,2% от штаммов данного
вида). Для M.chimaera-intracellulare доля расхождений результатов идентификации
составила 33,3%, для M.avium – 12,1%, для M.gordonae – 11,1%, M.kansasii – 8,3%.
Значительные расхождения результатов идентификации для M.fortuitum,
возможно, связаны с тем, что 18-ть штаммов, определенных методом MALDI-ToF
масс-спектрометрии, как представители других видов входят в M.fortuitum complex
и являются близкородственными. Однако в трех случаях культура,
идентифицированная как M.fortuitum, была определена как спорообразующая
палочка из порядка Bacillales. Полученные результаты расхождений
идентификации также могут быть обусловлены выделенными ассоциациями
микроорганизмов.
Отдельно были проанализированы результаты реидентификации
микроорганизмов, которые были идентифицированы с использованием метода
ДНК-гибридизации как представители рода Mycobacterium. Из 21-го штамма были
идентифицированы по четыре штамма M.avium и M.kansasii, по три штамма –
M.lentiflavum и M.szulgai, два штамма M.fortuitum, по одному штамму – M.xenopi,
M.gordonae, M.peregrinum, M.interjectum, которые должны быть
идентифицированы с помощью тест-системы, используемой в исследовании.
Также был идентифицирован один штамм НТМ, который не мог быть
идентифицирован до вида с использованием тест-системы Hain Lifescience GmbH
(Германия) – M.frederiksbergense.
Схожие результаты были выявлены и при сравнении результатов для
культур, идентифицированных как микроорганизмы с высоким содержанием ГЦ, и
для тех штаммов, идентификацию которых не удалось провести методом ДНК-

189
гибридизации. Во всех случаях методом MALDI-ToF масс-спектрометрии были
идентифицированы различные микроорганизмы из группы КУППА: НТМ,
гордонии, нокардии, представители родов Tsukamurella, Rhodococcus. И если
полученные результаты для части КУППА являются логичными, так как тест-
система Hain Lifescience GmbH (Германия) не подразумевает идентификацию
данных микроорганизмов, то результаты, полученные в отношении НТМ, ставят
вопрос о необходимости пересмотра алгоритмов действий врача-бактериолога в
случае получения отрицательных результатов, полученных методом ДНК-
гибридизации. Основной причиной таких расхождений, на наш взгляд, могут быть
разнообразные ассоциации микроорганизмов, из которых только часть была
выявлена в нашей работе; особенно если учесть факт, что нами не был
проанализирован весь возможный пласт прокариот: облигатные анаэробы, другие
виды, требующие особенных условий культивирования. Тем не менее, необходимо
продолжать исследования в данном направлении для более точного обоснования
полученных результатов.
Таким образом, ДНК-гибридизация является важным методом, который
решает целый ряд вопросов с идентификацией части НТМ, выделенных из
клинического материала. Однако необходимо учесть, что проведенный в нашей
работе анализ распространенности отдельных групп КУППА показал практически
полное отсутствие статистической достоверности по видам и укрупненным
кластерам микроорганизмов в зависимости от таксономического положения,
классификационных признаков и клинического значения, пола и возраста
пациентов, что необходимо принимать во внимание при работе с клиническим
материалом, в котором выявлены признаки роста нетуберкулезных микобактерий,
что требует разработки алгоритмов точной видовой идентификации, с учетом
факта выделения значительного числа (более 20,0%) микобактерий и других
КУППА с не доказанным клиническим значением.
По результатам, полученным в первых двух этапах исследования можно
сделать заключение, что КУППА как сложная и разнородная группа
микроорганизмов имеет ряд схожих свойств, которые в определенной мере

190
затрудняют их идентификацию при использовании питательных сред,
предназначенных для работы с МТВс. Культуральные свойства некоторых НТМ
могут значительно различаться в пределах одного вида или комплекса, что
подтверждает выявление диссоциаций штаммов M.avium, которые на среде
Левенштейна-Йенсена дали два типа роста: характерный для классических
вариантов (63,2% от всех нефотохромогенных НТМ), чаще описанный в
отечественной литературе, и в виде пигментированных колоний (49,4% от всех
фотохромогенных НТМ), что описано в зарубежных источниках [32, 46, 119, 124].
Таким образом, даже на уровне одного вида есть определенные сложности
при попытках предварительной идентификации НТМ при использовании
классических вариантов культивирования. Еще более сложной оказалась ситуация
при попытке выявить четкие критерии предварительной дифференциации
быстрорастущих НТМ и некоторых представителей из других групп КУППА, в
частности для 111-и штаммов, не образующих пигмент при визуальной оценке
роста на среде Левенштейна-Йенсена не было выявлено четких различий. В то же
время 22-а штамма быстрорастущих КУППА имели определенные различия по
культуральным свойствам, но при этом пять представителей НТМ дали колонии
желто-оранжевого цвета, более характерного для M.gordonae. В целом, в работе
были проанализированы культуральные свойства 402-х штаммов КУППА,
выделенных при обследовании на туберкулез, более 20% из которых можно
выделить, но невозможно идентифицировать в условиях работы лабораторий
противотуберкулезной службы.
В связи с вышеизложенным, были поставлены задачи провести оценку
возможности культивирования КУППА на агаризованных средах, разработать и
внедрить в практическую деятельность микробиологической лаборатории методы
предварительной идентификации КУППА при культивировании микроорганизмов
на универсальной хромогенной среде. Для этого было проведено исследование,
включающее в себя оценку ростовых свойств КУППА при культивировании на
кровяном агаре и универсальной хромогенной среде. Анализ был проведен для
представителей семи наиболее часто встречающихся видов НТМ, выделенных в

191
предыдущих этапах исследования: M.avium (n=73), M.kansasii (n=41), M.gordonae
(n=57), M.fortuitum (n=35), M.peregrinum (n=12), M.abscessus (n=12), M.septicum
(n=10). В результате сравнительной статистики количества колоний при
культивировании на анализируемых средах при расчете критерия Манна-Уитни
были получены достоверные различия, подтверждающие более подходящие
условия для выделения НТМ на универсальной хромогенной среде, чем на
кровяном агаре.
Для оценки влияния среды на скорость появления роста при температуре
культивирования 37°С были выбраны среда Левенштейна-Йенсена, кровяной агар
и универсальная хромогенная среда. Анализируя полученные результаты можно
сделать заключение об отсутствии статистически значимых отличий для
быстрорастущих НТМ по скорости роста при использовании исследуемых
агаризованных сред и среды Левенштейна-Йенсена. Медленнорастущие виды НТМ
на среде Левенштейна-Йенсена и универсальной хромогенной среде давали
видимый рост колоний достоверно быстрее, чем на кровяном агаре.
В связи с тем, что для многих видов НТМ, по данным литературы описан
температурный оптимум ниже 37°С, было принято решение проанализировать
возможность культивирования НТМ на универсальной хромогенной среде при
температуре 28°С [128, 250, 254]. Полученные данные соответствовали
опубликованным ранее в отношении всех культур за исключением M.avium и
M.kansasii, рост которых статистически достоверно не зависел от температуры
культивирования, р=0,267 и р=0,444 соответственно.
Таким образом, при использовании универсальной хромогенной среды
UriSelect (BioRAD, США) в условиях лаборатории, непрофильной по работе с
МТВс, можно получить сопоставимые результаты с данными при использовании
среды Левенштейна-Йенсена при культивировании наиболее часто встречающихся
видов НТМ. Сроки появления роста при этом статистически достоверно снижаются
при культивировании посевов при температуре 28°С.

192
Анализируя культуральные свойства КУППА, рост которых получен на
универсальной хромогенной среде, выявлены дополнительные особенности, на
основании которых предложена схема предварительной идентификации КУППА.
Для всех фотохромогенных медленнорастущих культур НТМ на
универсальной хромогенной среде был выявлен дисгонический рост в течение
одной-двух недель при температуре 28°С, характеризующийся появлением мелких
прозрачных или слегка пигментированных колоний, размером 1-2 мм, легко
снимающихся с поверхности среды. Для скотохромогенных медленнорастущих
культур НТМ был характерен рост точечных колоний размером 1-2 мм желтого или
оранжевого цвета, который появлялся на универсальной хромогенной среде в
течение одной-двух недель культивирования при температуре 28°С.
Нефотохромогенные НТМ в исследуемых условиях культивирования дали рост в
виде мелких желтоватых или кремовых колоний размером 1-2 мм, с появлением
видимого роста в течение одной-четырех недель. Быстрорастущие НТМ и другие
бактерии из группы КУППА давали на универсальной хромогенной среде
окрашенные в различные оттенки синего цвета колонии размером от 2 до 5 мм с
появлением характерного окрашивания колонии на пятые-седьмые сутки
культивирования. При этом гордонии, нокардии, стрептомицеты
характеризовались образованием в разной степени вросших в среду колоний, что
было нехарактерно для НТМ.
При реализации поставленных задач была выявлена сложность
предотвращения дегидратации среды при пролонгированном культивировании
посевов, которая была решена различными способами, включая авторскими, на
которые были получены патенты РФ на полезные модели.
Дополнительно была проанализирована возможность использования для
работы с КУППА универсальной хромогенной среды, в которую внесена
селективная добавка для подавления роста истинно контаминирующей
микрофлоры. Полученные результаты при оценке интенсивности роста
анализируемых культур были расценены как приемлемые, так как все тестируемые
штаммы дали обильный или умеренный рост.

193
Результаты, полученные на третьем этапе исследования, показали не только
возможность выделения, но и предварительной идентификации КУППА при
использовании универсальной хромогенной среды.
По данным литературы известно, что MALDI-ToF масс-спектрометрия
является высокоточным методом идентификации для многих видов
микроорганизмов, но в то же время имеет ряд особенностей при работе с
некоторыми бактериями и грибами [29, 50, 52]. В частности, это касается
идентификации микобактерий, для которых производителем масс-спектрометра
были разработаны отдельные протоколы подготовки культур для анализа, а также
специальные библиотеки, включающие спектры штаммов, выросших на жидких и
плотных средах для выделения МТВс [151, 310]. Возможность использования
универсальной хромогенной среды для культивирования КУППА определила
следующий вопрос научного поиска: насколько такой вариант выделения
кислотоустойчивых аэробных актиномицет влияет на результат идентификации с
использованием метода MALDI-ToF масс-спектрометрии?
Однако результаты сравнения методов идентификации КУППА,
приведенных в главе 4, поставили дополнительный вопрос, решение которого было
необходимо для проведения оценки возможности использования MALDI-ToF
масс-спектрометрии для работы с культурами, выросшими на универсальной
хромогенной среде. Насколько результаты идентификации НТМ методом ДНК-
гибридизации и MALDI-ToF масс-спектрометрии коррелируют с результатами
секвенирования? Для решения этого вопроса было проведено дополнительное
исследование сравнения идентификации 74-х образцов с признаками роста НТМ,
которые были выделены из клинического материала на среде Левенштейна-
Йенсена. Из данного этапа исследования были исключены все культуры,
идентифицированные хотя бы одним из методов, как не относящиеся к
микобактериям. Все пробы дополнительно тестировались на наличие ДНК МТВс
методом ПЦР.
При сравнительном анализе результатов идентификации, полученных
методами ДНК-гибридизации и секвенирования по Сэнгеру, полное совпадение

194
было выявлено только для 50-и образцов, что составило 67,6% от общего
количества посевов, включенных в исследование. Частичное совпадение
результатов идентификации составило 68,9%. При сравнении результатов
исследования 74-х образцов методом MALDI-ToF масс-спектрометрии с
секвенированием совпадение с учетом результатов идентификации в отношении
групп M.chimaera/M.intracellulare, M.porcinum/M.peregrinum и МТВс было
обнаружено для 67-и образцов – 90,5%; с учетом частичного совпадения результата
– 94,6%.
Максимальные расхождения результатов идентификации с помощью масс-
спектрометрии и секвенирования среди быстрорастущих НТМ были выявлены для
представителей M.fortuitum complex. Среди медленнорастущих видов
максимальное количество расхождений между тремя анализируемыми методами
было получено в случае выявления ассоциации микобактерий или обнаружения
ДНК представителей МТВс. Это отчасти подтверждает результаты, полученные в
главе 4. Если в образце выявляется нечистая культура микроорганизмов, то
возрастает риск получения неточного результата идентификации методом ДНК-
гибридизации. Полученные приемлемые результаты использования метода
MALDI-ToF масс-спектрометрии соответствуют опубликованным работам по
данному направлению [121, 135, 180, 186]. Полученные данные, на наш взгляд,
дают возможность более широкого использования метода MALDI-ToF масс-
спектрометрии в микробиологической практике лабораторий различного
направления.
Для решения вопроса о возможности проведения видовой идентификации
КУППА при культивировании на универсальной хромогенной среде были
проанализированы результаты видовой идентификации 292-х штаммов НТМ и 63-
х штаммов микроорганизмов из других групп КУППА. Для анализа полученных
результатов использовался критерий Х2. На данном этапе исследования
проводилось сравнение влияния различных методов пробоподготовки на
получение результата видовой идентификации в зависимости от среды

195
культивирования. В качестве среды сравнения была выбрана среда Левенштейна-
Йенсена.
Данные по сравнению пробоподготовки методом прямого нанесения с
методом экстракции и расширенным методом прямого нанесения оказались менее
приемлемыми с точки зрения конечного результата идентификации. Однако
полученные результаты идентификации с пробоподготовкой методом прямого
нанесения в зависимости от среды продемонстрировали достоверно более высокий
уровень при культивировании НТМ на универсальной хромогенной среде.
Полученные отрицательные результаты при проведении идентификации
методом прямого нанесения и методом экстракции муравьиной кислотой при
работе с КУППА обусловлены спецификой культуральных свойств данных
микроорганизмов. В случае, если культура была получена в виде вросших колоний
или при дисгоническом росте использование стандартных инструментов для
нанесения материала на мишень масс-спектрометра вызывало определенные
трудности. Для решения данного вопроса были разработаны и апробированы новые
инструменты для сбора колоний микроорганизмов с поверхности плотных
питательных сред.
Для обоснования полученных результатов идентификации НТМ был
проведен анализ белковых профилей культур, выращенных на различных
питательных средах. Для всех штаммов в спектрах были выявлены как
совпадающие пики вне зависимости от среды культивирования, так и значительные
отличия, в том числе и полное отсутствие некоторых пиков в зависимости от среды.
Однако определена закономерность, обусловленная большим количество пиков в
масс-спектрах для культур, выросших на универсальной хромогенной среде: при
расчете критерия Манна-Уитни для всех анализируемых культур количество пиков
в масс-спектрах было достоверно выше при культивировании на универсальной
хромогенной среде, чем на среде Левенштейна-Йенсена.
Использование универсальной хромогенной среды для культивирования
НТМ позволяет получить приемлемые результаты идентификации методом
MALDI-ToF масс-спектрометрии при использовании стандартной библиотеки. При

196
этом результаты видовой идентификации для всех тестируемых штаммов, рост
которых был получен на универсальной хромогенной среде, удалось получить
методом расширенного нанесения без необходимости проводить экстракцию
белков муравьиной кислотой. Это позволяет идентифицировать НТМ более
простым и доступным способом.
Полученные результаты определили следующий аспект научного поиска:
можно ли рассматривать выявленные особенности по количеству пиков в масс-
спектрах как косвенный признак более приемлемых условий для культивирования
НТМ на универсальной хромогенной среде?
КУППА, как группа классических сапрофитных микроорганизмов, имеет ряд
особенностей метаболизма, обусловленный эволюционными механизмами
выживания в окружающей среде в условиях жесткой ресурсной конкуренции.
Подходы, сформированные на использовании сред на яичной основе для
выделения патогенных микобактерий, являются обоснованными и логичными с
точки зрения потребностей МТВс в факторах роста, но, на наш взгляд, не является
обоснованием для экстраполяции этих данных на всю группу КУППА.
Известно, что железо играет важную роль в качестве лимитирующего
фактора, необходимого для большинства строго аэробных микроорганизмов, в
связи с чем они имеют сложные транспортные системы по доставке этого
микроэлемента из окружающей среды в клетку [314, 315]. При этом для патогенных
микобактерий приоритетным является механизм получения железа из белковых
молекул, а для сапрофитных видов – из содержащихся в окружающей среде
малорастворимых и нерастворимых форм [23, 53, 105]. В яичных средах железо
содержится в виде различных комплексов с белками, в частности с
овотрансферином, в то время, как в различных вариантах среды Миддлбрука
железо представлено в виде минеральной соли [205]. При этом рост микобактерий
на средах Миддлбрука по данным литературы, происходит быстрее, чем на яичных
средах [152, 153, 257].
Результаты сравнения культивирования НТМ и других КУППА на
универсальной хромогенной среде UriSelect (BioRAD, США) и среде Левенштейна-

197
Йенсена потребовали анализа состава данных сред для объяснения полученных
закономерностей. Уровень железа, определенный колориметрически, оказался
минимальным для универсальной хромогенной среды в сравнении с классическими
средами, применяемыми для выделения МТВс. Однако в связи с отсутствием
данных о форме железа в составе хромогенной среды, можно предположить, что ее
доступность для утилизации микобактериями оказалась достаточной для роста
большинства штаммов, включенных в исследование. С другой стороны, низкий
уровень железа мог повлиять на отсутствие роста отдельных штаммов M.avium,
M.gordonae, M.chimaera/intracellulare и M.szulgai на универсальной хромогенной
среде.
Параллельно с определением уровня железа в анализируемых средах был
определен уровень белка. Закономерно, что в средах на яичной основе уровень
белка был высоким и составил более 70 г/л, в то время как в среде Миддлбрук 7Н9
и универсальной хромогенной среде количество общего белка было значительно
ниже. В связи с этим нами было сделано предположение о возможном негативном
влиянии избытка белка на условно-патогенные микроорганизмы из группы
КУППА, что подтверждается результатами, описанными в главе 3, в которой
приведены данные о том, что среда Миддлбрука 7Н9 является достоверно более
приемлемой для выделения кислотоустойчивых актиномицет, а также
подтверждается литературными данными [118, 189, 206].
Наличие легкодоступных источников аминокислот в совокупности с
микроэлементным составом, схожим по основным компонентам со средой
Миддлбрука, возможно позволило получить приемлемые результаты
идентификации НТМ методом MALDI-ToF масс-спектрометрии за счет
увеличения количества пиков в масс-спектрах.
Для оценки возможности практического использования результатов,
полученных в нашем исследовании, дополнительно был проведен анализ работы
микробиологического отдела КДЛ Клиник ФГБОУ ВО СамГМУ Минздрава
России после внедрения метода продленного инкубирования на универсальной

198
хромогенной среде UriSelect (BioRAD, США) различных видов клинического
материала с целью выделения КУППА.
За контрольный период с 2016 по 2017 годы, при котором посевы
инкубировались в стандартных условиях было выделено 14-ть штаммов КУППА и
представителей рода Actinomyces, в то время, как за анализируемый период с 2018
по 2019 годы 45-ть штаммов. С учетом сопоставимого общего количества посевов
за анализируемые периоды, мы считаем полученные результаты важным
критерием, подтверждающим возможность выделения КУППА в рутинной
микробиологической практике.
Использование универсальной хромогенной среды в условиях оснащения
лаборатории MALDI-ToF масс-спектрометром открывает дополнительные
возможности, выявленные в нашей работе. Нетуберкулезные микобактерии, также,
как и другие виды КУППА, являются широко распространенными в окружающей
среде микроорганизмами, в связи с чем, при проведении диагностики
обусловленных ими заболеваний часто встает вопрос о верификации их
клинического значения. С учетом того, что микроорганизмы, в норме заселяющие
экологические ниши природных объектов, чаще всего представляют собой
сложные гетерогенные популяции, характеризующиеся различными
возможностями приспособления к тем или иным условиям в пределах одного
объекта внешней среды. В организме человека такие процессы возможны при
длительно протекающих инфекционных процессах, что, на наш взгляд, можно
использовать при анализе протеомных профилей выделенных штаммов.
Приведенные в главе 7 примеры такого анализа демонстрируют возможность
субтипирования штаммов, выделенных на универсальной хромогенной среде в
случае обнаружения в посеве признаков гетерогенности популяции по
культуральным свойствам.
Выявление сходств и отсутствие значимых различий в спектрах штаммов
КУППА, выделенных от одного пациента в разное время в совокупности с
отсутствием признаков гетерогенности популяции по культуральным свойствам
можно рассматривать как фактор, подтверждающий один источник выделения и

199
высокую вероятность участия данного микроорганизма в патологическом
процессе.
Таким образом, разработанные новые подходы к культивированию
кислотоустойчивых представителей порядка Actinomycetales, выделенных из
клинического материала, основанные на использовании универсальной
хромогенной среды позволяют по многим параметрам достоверно повысить
качество выделения данной группы микроорганизмов, проводить их
предварительную идентификацию по культуральным свойствам и точную видовую
идентификацию при условии использования метода MALDI-ToF масс-
спектрометрии. Внедрение полученных результатов в работу лабораторий
различного уровня позволит интенсифицировать процесс их выделения из
клинического материала, и, как следствие, улучшит диагностику заболеваний,
вызванных различными условно-патогенными видами из группы аэробных
кислотоустойчивых актиномицет.
Перспективы дальнейшей разработки темы:
Дальнейшее развитие работы планируется в следующих направлениях:
1. Исследование предложенными в работе методами структуры
кислотоустойчивых актиномицет в сопоставлении с клинической картиной
патологических состояний для верификации их клинического значения и
уточнения лабораторных критериев диагностики.
2. Основываясь на собранной во время исследования коллекции штаммов
кислотоустойчивых актиномицет, проведение комплексного анализа
биологических свойств микроорганизмов, оценки факторов их патогенности и
определения чувствительности к антимикробным препаратам.
3. Провести подробный протеомный анализа штаммов
кислотоустойчивых актиномицет из собранной коллекции и полученных из других
источников для выяснения возможности подвидового типирования и оценки
других свойств отдельных групп микобактерий, нокардий, гордоний и
стрептомицет в зависимости от источника выделения.

200
ВЫВОДЫ
1. Контаминирующая микрофлора при обследовании на туберкулез
содержит в своем составе микроорганизмы из группы кислотоустойчивых
представителей порядка Actinomycetales, в том числе нетуберкулезные
микобактерии с атипичными культуральными свойствами. Доля таких
микроорганизмов составила 9,3% от общего количества выделенных штаммов.
2. В структуре кислотоустойчивых представителей порядка
Actinomycetales, предварительно идентифицированных как нетуберкулезные
микобактерии, дополнительно были реидентифицированы методом MALDI-ToF
масс-спектрометрии различные по составу ассоциации микроорганизмов, штаммы
микобактерий, для которых не был получен приемлемый результат видовой
идентификации методом ДНК-гибридизации и другие представители порядка
Actinomycetales.
3. В исследовании подтверждена возможность культивирования
основных представителей порядка Actinomycetales на агаризованных средах со
статистически достоверным преимуществом универсальной хромогенной среды.
4. Использование универсальной хромогенной среды, содержащей
хромогенные субстраты для обнаружения бактериальных ферментов β-
глюкуронидаз, β-галактозидаз и триптофан позволяет проводить предварительную
идентификацию кислотоустойчивых представителей порядка Actinomycetales по
культуральным свойствам.
5. Идентификация с использованием MALDI-ToF масс-спектрометрии
кислотоустойчивых представителей порядка Actinomycetales, выделенных из
клинического материала на универсальной хромогенной среде, возможна при
менее трудозатратном способе пробоподготовки методом расширенного нанесения
без необходимости экстракции белков муравьиной кислотой.
6. Использование универсальной хромогенной среды для
культивирования нетуберкулезных микобактерий позволяет получить приемлемые

201
результаты идентификации методом MALDI-ToF масс-спектрометрии при
использовании стандартной библиотеки масс-спектров.
7. Выявление сходств и отсутствие значимых различий в спектрах
штаммов микобактерий, выделенных от одного пациента в разное время в
совокупности с отсутствием признаков гетерогенности популяции по
культуральным свойствам можно рассматривать как фактор, подтверждающий
один источник выделения и предполагать высокую вероятность участия данного
микроорганизма в патологическом процессе.

202
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
1. С целью повышения частоты выделения кислотоустойчивых
актиномицет из клинического материала при обследовании на туберкулез
необходимо проведение видовой идентификации всех культур, выросших на среде
Миддлбрука 7Н9 с селективной добавкой.
2. Отрицательный результат идентификации методом ДНК-гибридизации
штаммов с культуральными свойствами, характерными для нетуберкулезных
микобактерий, требует более тщательного анализа полученных изолятов для
выявления микробных ассоциация, в том числе состоящих из микобактерий
различных видов.
3. Использование универсальной хромогенной среды в комбинации с
продленной инкубацией позволяет выделять различные виды кислотоустойчивых
актиномицет (Патент РФ № 17302, Патент РФ № 175134, Патент РФ № 175863,
Патент РФ № 2659155, Патент РФ № 2668406).
4. С помощью универсальной хромогенной среды, содержащей
хромогенные субстраты для обнаружения бактериальных ферментов β-
глюкуронидаз, β-галактозидаз и триптофан возможно проведение предварительной
идентификации кислотоустойчивых актиномицет по культуральным свойствам
(Патент РФ № 2711954, Патент РФ № 2711957).
5. Необходимо рассмотреть возможность межлабораторного
взаимодействия лабораторий туберкулезной службы и бактериологических
лабораторий вне ее для решения вопроса об идентификации культур из группы
кислотоустойчивых актиномицет.
6. Для уточнения лабораторных критериев диагностики микобактериозов
и других инфекций, вызванных кислотоустойчивыми актиномицетами необходимо
разрабатывать и внедрять методы, основанные на субтипировании выделенных
штаммов по их белковым профилям.

203
СПИСОК СОКРАЩЕНИЙ
ВИЧ – вирус иммунодефицита человека
ГЦ – гуанин-цитозин
ДНК – дезоксирибонуклеиновая кислота
КОЕ – колониеобразующая единица
КУППА – кислотоустойчивые представители порядка Actinomycetales
МУК – методические указания
НТМ – нетуберкулезные микобактерии
ПИД – первичный иммунодефицит
ПЦР – полимеразная цепная реакция
ХОБЛ – хроническая обструктивная болезнь легких
ATCC – American Type Culture Collection
ВСС – Burkholderia cepacia complex
HCCA – α-cyano-4-hydroxycinnamic acid
MALDI-ToF – Matrix Assisted Laser Desorption/Ionization Time of Flight
MTBc – Mycobacterium tuberculosis complex
PANTA – Polymyxin B/Amphotericin B/Nalidixic acid/Trimethoprim/
Azlocillin

204
СПИСОК ЛИТЕРАТУРЫ
1. Агаева, А. А. Влияние кислотности и растворимого алюминия на
развитие штаммов рода Nocardia, выделенных из Баиловских нефтезагрязненных
почв / А. А. Агаева // Успехи современного естествознания. – 2010. – № 4. – С. 9–
12.
2. Алгоритм микропроцессорного управления параметрами
культивирования автотрофных микроорганизмов / A. A. Шевцов, A. B. Дранников,
A. B. Пономарев [и др.] // Автоматизация и современные технологии. – 2011. – №
8. – С. 33–35.
3. Алгоритм ускоренного бактериологического исследования с
использованием хромогенных питательных сред / О. В. Полухина, Т. Н. Суборова,
С. А. Егорова [и др.] // Справочник заведующего КДЛ. –2015. – № 1. – С. 3–8.
4. Альховик, О. И. Распространенность нетуберкулезных микобактерий в
Сибири / О. И. Альховик, М. А. Дымова, А. Г. Чередниченко // Проблемы
медицинской микологии. – 2016. – Т. 18, № 2. – С. 37.
5. Аникин, В. А. Проблемы микробиологической диагностики
туберкулеза / В. А. Аникин, В. К. Гришин // Военно-медицинский журнал. – 2012.
– Т. 333, № 3. – С. 33–36.
6. Антибиотикорезистентность биопленочных бактерий / И. В. Чеботарь,
А. Н. Маянский, Е. Д. Кончакова [и др.] // Клиническая микробиология и
антимикробная химиотерапия. – 2012. – Т. 14, № 1. – С. 51–58.
7. Беляков, Н. А. Коморбидные и тяжелые формы ВИЧ-инфекции в
России / Н. А. Беляков // ВИЧ-инфекция и иммуносупрессии. – 2016. – Т. 8, № 3. –
С. 9–25.
8. Беляков, Н. А. Противодействие ВИЧ-инфекции и рост заболеваемости
в России / Н. А. Беляков, В. В. Рассохин, А. С. Бобрешова // ВИЧ-инфекция и
иммуносупрессии. – 2017. – Т. 9, № 2. – С. 82–90.

205
9. Бердников, Р. Б. Летальный случай генерализованного микобактериоза
у больного с терминальной стадией ВИЧ-инфекции / Р. Б. Бердников,
Л. М. Гринберг, А. Ю. Евсеев // Туберкулез и болезни легких. – 2016. – № 94(4). –
С. 57–62.
10. Видовая идентификация микобактерий нетуберкулезного комплекса
методом амплификации и секвенирования генов 16S рРНК / А. А. Майорова, В. Н.
Степаншина, О. В. Коробова [и др.] // Журнал молекулярной генетики,
микробиологии, вирусологии. – 2004. – № 3. – С. 11–20.
11. Возможности и перспективы бактериологической диагностики
микобактериоза / Т. Ф. Оттен, И. В. Мокроусов, О. В. Нарвская [и др.] // Проблемы
туберкулеза. – 2004. – № 5. – С. 32–35.
12. Волков, Д. В. Источники микобактерий и распространение
микобактериозов свиней в Новосибирской области / Д. В. Волков, К. В. Авдеенко,
П. В. Бушмелева // Сибирский вестник сельскохозяйственной науки. – 2009. – № 4.
– С. 45–49.
13. Выделение и идентификация нетуберкулезных микобактерий /
В. И. Литвинов, М. В. Макарова, М. А. Краснова [и др.] // Вестник РАМН. – 2010.
– № 3. – С. 7–11.
14. Выявление нетуберкулезных микобактерий в республике Марий Эл /
Л. В. Петрова, Е. И. Мельникова, Ю. А. Соловьева [и др.] // Туберкулез и болезни
легких. – 2018. – Т. 96, № 2. – С. 41–46.
15. Выявление нетуберкулезных микобактерий, циркулирующих в разных
регионах Сибири, и анализ их лекарственной устойчивости / О. И. Альховик, И. О.
Мешков, Т. И. Петренко [и др.] // Туберкулез и болезни легких. – 2019. – Т. 97, №
10. – С. 5–11.
16. Генетическое маркирование полирезистентных штаммов
Mycobacterium tuberculosis, выделенных на Северо-Западе России / О. В. Нарвская,
И. В. Мокроусов, Т. Ф. Оттен [и др.] // Проблемы туберкулеза. – 1999. – № 3. –
С. 39–41.

206
17. Гунтупова, Л. Д. Микобактериозы во фтизиопульмонологической
практике: обзор литературы и собственный опыт / Л. Д. Гунтупова, С. Е. Борисов,
И. П. Соловьева // Практическая медицина. – 2011. – № 3(51). – С. 39–50.
18. Диагностика микобактериоза у больных ВИЧ-инфекцией / В.
Н. Зимина, М. В. Альварес Фигероа, С. Ю. Дегтярева [и др.] // Инфекционные
болезни. – 2016. – Т. 14, № 4. – С. 63–70.
19. Диагностика сочетания туберкулеза и микобактериоза при ВИЧ-
инфекции / М. В. Альварес Фигероа, Ю. Р. Зюзя, А. В. Прокопенко [и др.] //
Туберкулез и социально значимые заболевания. – 2015. – № 4. – С. 50–57.
20. Диагностика туберкулеза микробиологическими методами в
учреждениях противотуберкулезной службы и общей лечебной сети / Л. И.
Русакова, В. В. Пунга, Л. П. Капков [и др.] // Туберкулез и болезни легких. – 2011.
– Т. 88, № 5. – С. 139–140.
21. Диализное культивирование микроорганизмов как адекватная модель
контроля популяции при исследовании экосистем / А. Ф. Лебедева, Е. Л. Барский,
Я. В. Саванина [и др.] // Вестник Моск. ун-та. Сер. Биология. – 2010. – № 2. – С. 15–
20.
22. Дифференциальная диагностика туберкулеза и микобактериоза у
больных ВИЧ-инфекцией / А. М. Пантелеев, О. В. Никулина, А. С. Христусев [и
др.] // Туберкулез и болезни легких. – 2017. – Т. 95, № 10. – С. 47–54.
23. Добин, В. Л. Обмен железа у микобактерий / В. Л. Добин, В. Г.
Демихов, М. П. Жарикова // Туберкулез и болезни легких. – 2016. – Т. 94, № 7. –
С. 6–10.
24. Елисеев, П. И. Роль молекулярно-генетических методов в повышении
эффективности диагностики туберкулеза с лекарственной устойчивостью
микобактерий и микобактериозов : специальность 14.01.16 «Фтизиатрия» :
автореферат диссертации на соискание ученой степени кандидата медицинских
наук / Елисеев Платон Иванович ; ГБУВ ВПО «Северный государственный
медицинский университет» МЗ РФ. – Санкт-Петербург, 2013. – 23 с.

207
25. Железо как эссенциальный фактор роста микобактерий / А. В. Лямин,
А. В. Халиулин, Д. Д. Исматуллин [и др.] // Известия Самарского научного центра
РАН. – 2016. – Т. 18, № 5(2). – С. 320–327.
26. Зорина, А. С. Биопленкообразование моно- и смешанных культур
штаммов Alcaligenes faecalis 2 и Rhodococcus ruber gt / А. С. Зорина,
Ю. Г. Максимова, В. А. Демаков // Микробиология. – 2019. – Т. 88, № 2. – С. 175–
183.
27. Иммобилизация клеток штамма Rhodococcus wratislaviensis кт112-7 как
способ повышения эффективности биодеструкции модифицированных
полихлорбифенилов / Д. О. Егорова, М. Г. Первова, Т. И. Горбунова [и др.] //
Биотехнология. – 2019. – Т. 35, № 2. – С. 58–70.
28. Исаева, Г. Ш. Отечественная хромогенная среда для выделения
Helicobacter pylori / Г. Ш. Исаева, О. К. Поздеев // Клиническая лабораторная
диагностика. – 2009. – № 8. – С. 35–37.
29. Использование времяпролетной масс-спектрометрии с матрично-
активированной лазерной десорбцией/ионизацией (MALDI-TоF MS) для
идентификации бактериальных и грибковых возбудителей III-IV групп
патогенности / И. В. Чеботарь, С. В. Поликарпова, Ю. А. Бочарова [и др.] //
Лабораторная служба. – 2018. – Т. 7, № 2. – С. 78–86.
30. Использование методов MALDI-ToF масс-спектрометрии и
количественной ПЦР для быстрой диагностики септических состояний /
Т. В. Припутневич, А. Р. Мелкумян, О. В. Бурменская [и др.] // Клиническая
микробиология и антимикробная химиотерапия. – 2014. – Т. 16, № 1. – С. 4–9.
31. Кимайкина, О. В. Результаты эксперимента: использование Chromagar
Staphyloccocus с добавлением цефокситина для раннего выявления
метициллинрезистентности стафилококков, возбудителей
имплантассоциированных инфекций (ИАИ) в травматологии-ортопедии / О. В.
Кимайкина, Л. Г. Григоричева, Е. А. Супрун // Клиническая микробиология и
антимикробная терапия. – 2017. – № 19(51). – С. 20.

208
32. Клиническая лабораторная диагностика: национальное руководство : в
2 томах. Т. 2 / под ред. проф. В. В. Долгова, проф. В. В. Меньшикова. – Москва :
ГЭОТАР-Медиа, 2013. – 805 с.
33. Кокиева, Г. Е. Исследование аппарата для культивирования
микроорганизмов / Г. Е. Кокиева // Научно-технический вестник Поволжья. – 2014.
– № 4. – С. 123–125.
34. Контаминирующая микрофлора при обследовании на туберкулез:
сапрофиты или потенциальные патогены? / А. В. Лямин, Д. Д. Исматуллин, А. В.
Жестков [и др.] // Иммунопатология, аллергология, инфектология. – 2019. – № 3. –
С. 63–70.
35. Коробова, А. Г. Выявление продукции бета-лактамаз расширенного
спектра у энтеробактерий с помощью селективных сред / А. Г. Коробова, Г. А.
Клясова // Медиаль. – 2014. – Т. 13, № 3. – С. 116.
36. Коробова, А. Г. Использование селективной хромогенной среды для
детекции энтеробактерий с продукцией бета-лактамаз / А. Г. Коробова, И. Н.
Фролова, Г. А. Клясова // Клиническая лабораторная диагностика. –2015. – Т. 60,
№ 11. – С. 53–57.
37. Костина, Л. В. Извлечение ионов тяжелых металлов клетками
Rhodococcus ruber ИЭГМ 231 / Л. В. Костина, М. С. Куюкина, И. Б. Ившина //
Вестник Пермского гос. ун-та. Серия Биология. – 2013. – Вып. 2. – С. 42–48.
38. Лабораторная диагностика микобактериозов у пациентов с
муковисцидозом (обзор литературы) / А. В. Лямин, Д. Д. Исматуллин, А. В.
Жестков [и др.] // Клиническая лабораторная диагностика. – 2018. – Т. 63, № 5. – С.
315–320.
39. Ларионова, Е. Е. Инфицирование микобактериями больных с
кистозным фиброзом / Е. Е. Ларионова // Вестник Центрального научно-
исследовательского института туберкулеза. – 2019. – № S1. – С. 191–199.
40. Лаушкина, Ж. А. Случаи диагностики микобактериоза легких у
больного диссеминированным туберкулезом / Ж. А. Лаушкина, О. И. Ольховик
// Туберкулез и болезни легких. – 2018. – № (1). – С. 47–50.

209
41. Лискова, Е. А. Метод деконтаминации патологического материала от
мышей, заражённых rhodococcus equi / Е. А. Лискова, К. Н. Слинина // Евразийский
союз ученых. – 2015. – № 5-5(14). – С. 168–169.
42. Лискова, Е. А. Новый подход к выделению микобактерий,
нокардиоформных актиномицетов и коринебактерий / Е. А. Лискова,
К. Н. Слинина // Вестник Алтайского государственного аграрного университета. –
2016. – № 9(143). – С. 128–131.
43. Лискова, Е. А. Профилактика актиномицетных инфекций у телят
/ Е. А. Лискова, К. Н. Слинина, А. А. Блохин // Вестник АГАУ. – 2017. – № 6(152).
– С. 144–149.
44. Литвиненко, Л. В. Способность актинобактерий родов Dietzia,
Gordonia и Rhodococcus аккумулировать ионы никеля / Л. В. Литвиненко //
Микробиология. – 2019. – Т. 88, № 2. – С. 207–216.
45. Литвинов, В. И. Нетуберкулезные микобактерии / В. И. Литвинов,
М. В. Макарова, М. А. Краснова. – Москва : МНПЦБТ, 2008. – 256 с.
46. Лямин, А. В. Лабораторная диагностика микобактериозов : учебное
пособие для студентов / А. В. Лямин, А. В. Жестков, Д. Д. Исматуллин. – Самара :
Право, 2019. – 72 с.
47. Лямин, А. В. Редкие виды в структуре кислотоустойчивых
представителей порядка Actinomycetales, выделенных из клинического материала /
А. В. Лямин // Клиническая лабораторная диагностики. – 2020. – № 65(2). – С. 111–
115.
48. Майрова, А. А. Индентификация нетуберкулезных микобактерий и
выбор оптимальной комбинации методов для их видовой дифференциации :
специальность 03.00.07 «Микробиология» : автореферат диссертации на соискание
учетной степени кандидата биологических наук / Майорова Ангелина
Александровна ; ФГУЗ «Московский научно-исследовательский институт
эпидемиологии и микробиологии им. Г.Н. Габричевского» Роспотребнадзора РФ. –
Москва, 2007. – 26 с.

210
49. Макарова, М. В. Изучение чувствительности нетуберкулезных
микобактерий, выделенных на плотных и жидких питательных средах к
противотуберкулезным препаратам / М. В. Макарова, Г. Е. Фрейман // Проблемы
туберкулеза. – 2009. – № 9. – С. 49–51.
50. МАLDI-ToF масс-спектрометрический анализ клеточных белков в
идентификации представителей рода Leptospira / Н. В. Бренева, М. В. Афанасьев,
М. Б. Шаракшанов [и др.] // Журнал микробиологии, эпидемиологии и
иммунобиологии. – 2014. – № 4. – С. 36–43.
51. МАLDI-TOF масс-спектрометрия в рутинной работе
микробиологической лаборатории / Н. А. Маянский, А. Н. Калакуцкая, О. В.
Мотузова [и др.] // Вопросы диагностики в педиатрии. – 2011. – № 3(5). – С. 20–25.
52. Масс-спектрометрическая оценка карбапенемазной активности
pseudomonas aeruginosa / Ю. А. Бочарова, И. В. Чеботарь, О. А. Крыжановская [и
др.] // Клиническая лабораторная диагностика. –2018. – Т. 63, № 2. – С. 99–105.
53. Маянский, А. Н. Патогенетическая микробиология : руководство /
А. Н. Маянский. – Н. Новгород : Изд–во Нижегородской государственной
медицинской академии, 2006. – 520 с.
54. Микобактериозы легких: критерии диагностики в современных
условиях / Л. К. Суркова, Е. М. Скрягина, О. М. Залуцкая [и др.] // Смоленский
медицинский альманах. – 2015. – № 3. – С. 145–151.
55. Микобактериозы органов дыхания: эпидемиология,
микробиологические и клинические аспекты диагностики / Л. Д. Гунтупова, С. Е.
Борисов, М. В. Макарова [и др.] // Эпидемиология и инфекционные болезни. –
2012. – № (2). – С. 8–14.
56. Микобактериозы: современное состояние проблемы / В. Н. Зимина,
С. Ю. Дегтярева, Е. Н. Белобородова [и др.] // Клиническая микробиология и
антимикробная химиотерапия. – 2017. – Т. 19, № 4. – С. 276–282.

211
57. Микробиологические методы диагностики туберкулеза: теоретическое
учебное пособие для проведения курсов обучения: «Выявление туберкулеза
методом микроскопии» и «Культуральные методы диагностики туберкулеза» /
Центр. НИИ туберкулеза РАМН [и др.]. – Москва ; Тверь : Триада, 2008. – 39 с. :
цв. ил.
58. Мониторинг видового разнообразия нетуберкулезных микобактерий в
ряде областей РФ с использованием ДНК-стрипов Genotype Mycobacterium CM/AS
(HAINLIFESCIENCE, Германия) / Т. Г. Смирнова, С. Н. Андреевская, Е. Е.
Ларионова [и др.] // Туберкулез и болезни легких. – 2017. – Т. 95, № 5. – С. 54–59.
59. Морфологическая верификация ВИЧ-ассоциированного
микобактериоза, вызванного нетуберкулезными микобактериями Myc. avium
complex / Ю. Р. Зюзя, Ю. Г. Пархоменко, В. Н. Зимина [и др.] // Клиническая и
экспериментальная морфология. – 2015. – № 3(1). – С. 11–21.
60. МУК 4.2.2316-08 «Методы контроля бактериологических питательных
сред». – Текст : электронный // Техэксперт. Консорциум Кодекс. – URL:
http://docs.cntd.ru/document/1200067870 (дата обращения: 23.04.2020).
61. Неизвестные условно-патогенные бактерии представители
микрофлоры человека как возможные возбудители пневмонии / Г. В. Тец,
Д. С. Викина, М. Ф. Вечерковская [и др.] // Атмосфера. Пульмонология и
аллергология. – 2013. – № 1. – С. 32–34.
62. Нестерильность мочи здорового человека - новая парадигма в
медицине / В. И. Коган, Ю. Л. Набока, Х. С. Ибмиев [и др.] // Урология. – 2014. –
№ 5. – С. 48–52.
63. Нетуберкулезные микобактерии у пациентов с заболеваниями органов
дыхания (клинико-лабораторное исследование) / А. Э. Эргешов, Е. И. Шмелев,
М. Н. Ковалевская [и др.] // Пульмонология. – 2016. – Т. 26, № 3. – C. 303–308.
64. Нетуберкулезные микобактерии: современные возможности видовой
идентификации / А. В. Лямин, А. М. Ковалёв, А. В. Жестков [и др.] // Клиническая
микробиология и антимикробная химиотерапия. – 2017. – Т. 19, № 1. – С. 11–14.

212
65. NGS: высокопроизводительное секвенирование / Д. В. Ребриков,
Д. О. Коростин, Е. С. Шубина [и др.]. – Москва : БИНОМ. Лаборатория знаний,
2014. – 235 c.
66. Новые возбудители заболеваний респираторного тракта у
иммунокопрометированных пациентов (обзор литературы) / Д. Д. Исматуллин,
М. О. Золотов, А. В. Лямин [и др.] // Наука и инновации в медицине. – 2019. – Т. 4,
№ 3. – С. 19–24.
67. Норовсурэн, Ж. Актиномицеты редких родов в каштановых почвах и
их антагонистические свойства / Ж. Норовсурэн, Д. Дарам, Б. Цэцэг // Известия
АНМ. – 2003. – Т. 170, № 4. – С. 25–29.
68. Общая характеристика и клиническое значение представителей родов
Nocardia и Gordonia / А. В. Лямин, А. В. Жестков, Т. Р. Никитина [и др.] //
Инфекция и иммунитет. – 2019. – Т. 9, № 3-4. – С. 429–436.
69. Определитель бактерий Берджи : 2 томах. Т. 2 : пер. с англ. / под ред.
Дж. Хоулта, Н. Крига, П. Снита [и др.]. – Москва : Мир, 1997. – 386 с.
70. Оптимизация лабораторной диагностики туберкулеза с
использованием современных бактериологических и молекулярно-биологических
методов / Я. М. Балабанова, Ф. Дробниевский, И. М. Федорин [и др.] // Проблемы
туберкулеза и болезней легких. – 2011. – № 2. – С. 36–42.
71. Опыт применения микробиологических методов исследований при
инфекционных осложнениях тяжелых травм / С. А. Свистунов, А. А. Кузин, Т. Н.
Суборова [и др.] // Инфекция и иммунитет. – 2016. – Т. 6, № 4. – С. 373–378.
72. Отечественная хромогенная среда для дифференциации клебсиелл /
Р. Ю. Юнусова, В. Г. Горелова, Э. Д. Степанова [и др.] // Клиническая
лабораторная диагностика. – 2010. – № 3. – С. 49–51.
73. Оттен, Т. Ф. Микобактериоз / Т. Ф. Оттен, А. В. Васильев. – Санкт–
Петербург, 2005. – 218 с. : ил.
74. Оттен, Т. Ф. Генерализованный микобактериоз у пациента с ВИЧ-
инфекцией на стадии СПИДа / Т. Ф. Оттен, Н. В. Фоменкова, М. Ю. Майская //
Туберкулез и болезни легких. – 2015. – № 8. – С. 57–62.

213
75. Оттен, Т. Ф. Характеристика нетуберкулезных микобактерий -
потенциальных возбудителей заболеваний человека / Т. Ф. Оттен // Проблемы
туберкулеза. – 1994. – № 3. – С. 56–59.
76. Оценка диагностической эффективности алгоритма выделения и
ускоренной идентификации условно-патогенных энтеробактерий с
использованием отечественных хромогенных питательных сред / М. М. Меджидов,
Э. Д. Степанова, Р. Ю. Юнусова [и др.] // Клиническая лабораторная диагностика.
– 2012. – № 5. – С. 51–54.
77. Патент RU 173302 U1 Российская Федерация, МПК C12M 1/22
(2006.01). Чашка для культивирования микроорганизмов с двухпозиционной
резьбой : заявка № 2017108813, 16.03.2017 : опубл. 21.08.2017 / А. В. Лямин, И. Р.
Тонеев, А. В. Козлов [и др.]. – Бюл. № 24. – 7 с.
78. Патент RU 175134 U1 Российская Федерация, МПК C12M 1/22
(2006.01). Флакон для культивирования микроорганизмов : заявка № 2017116263,
10.05.2017 : опубл. 22.11.2017 / А. В. Лямин, И. Р. Тонеев, А. В. Козлов [и др.]. –
Бюл. №33. – 5 с.
79. Патент RU 175863 U1 Российская Федерация, МПК С12М 1/22
(2006.01). Флакон для культивирования микроорганизмов со съемной крышкой :
заявка № 2017116235, 10.05.2017 : опубл. 21.12.2017 / А. В. Лямин, И. Р. Тонеев, А.
В. Козлов [и др.]. – Бюл. №36. – 6 с.
80. Патент RU 2659155 C1 Российская Федерация, МПК G01N 33/487
(2006.01). Способ сбора и первичного посева жидкости назального лаважа от
пациентов с муковисцидозом для микробиологического исследования : заявка №
2017122649, 27.06.2017 : опубл. 28.06.2018 / О. В. Кондратенко, А. В. Лямин, Е. Д.
Медведева, А. В. Ермолаева. – Бюл. № 19. – 6 с.
81. Патент RU 2668406 C1 Российская Федерация, МПК C12Q 1/04
(2006.01), C12N 1/20 (2006.01), C12R 1/01 (2006.01). Способ первичного посева
биоматериала, выделенного из нижних дыхательных путей пациентов с
муковисцидозом : заявка № 2017122657, 27.06.2017 : опубл. 28.09.2018 / О. В.
Кондратенко, А. В. Лямин, Д. Д. Исматуллин. – Бюл. № 28. – 9 с.

214
82. Патент RU 2689800 C1 Российская Федерация, МПК G01N 33/569
(2006.01), C12Q 1/6876 (2018.01). Способ детекции изолятов Mycobacterium
tuberculosis Beijing 94-32-кластера в формате реального времени : заявка №
2017142885, 07.12.2017 : опубл. 29.05.2019 / И. В. Мокроусов, А. А. Вязовая, Е. Н.
Черняева [и др.]. – Бюл. № 16. – 11 с.
83. Патент RU 2711954 C1 Российская Федерация, МПК C12N 1/20
(2006.01), C12R 1/01 (2006.01). Способ предварительной идентификации
нетуберкулезных микобактерий с использованием универсальной хромогенной
среды : заявка № 2019114859, 14.05.2019 : опубл. 23.01.2019 / А. В. Лямин, Д. Д.
Исматуллин, О. В. Кондратенко [и др.]. – Бюл. № 3. – 6 с.
84. Патент RU 2711957 C1 Российская Федерация, МПК C12N 1/20
(2006.01), C12R 1/01 (2006.01). Способ первичного посева клинического материала
для выделения нетуберкулезных микобактерий : заявка № 2019114858, 14.05.2019
: опубл. 23.01.2020 / А. В. Лямин, Д. Д. Исматуллин, О. В. Кондратенко [и др.]. –
Бюл. № 3. – 5 с.
85. Першикова, Н. Л. Использование пцр-тест-систем для диагностики
туберкулеза и микобактериозов / Н. Л. Першикова // Сибирский вестник
сельскохозяйственной науки. – 2009. – № 2(194). – С. 59–62.
86. Питательная среда для выделения Neisseria meningitidis / Л. С.
Черкасова, И. С. Королева, И. М. Грубер [и др.] // Журнал микробиологии,
эпидемиологии и иммунобиологии. – 2008. – № 1. – С. 69–71.
87. Пневмоцистная и цитомегаловирусная пневмония как проявление
синдрома восстановления иммунной системы / С. Ю. Дегтярева, В. Н. Зимина,
Е. Н. Белобородова [и др.] // Трудный пациент. – 2019. – Т. 17, № 11-12. – С. 40–43.
88. Подбор оптимальных параметров культивирования штаммов
молочнокислых бактерий, перспективных в качестве стартерных культур при
разработке закваски прямого внесения / Э. Нагызбеккызы, Н. Б. Молдагулова [и
др.] // Международный журнал прикладных и фундаментальных исследований. –
2019. – № 7. – С. 14–18.

215
89. Приказ № 109 МЗ РФ от 21 марта 2003 года «О совершенствовании
противотуберкулезных мероприятий в Российской Федерации / Министерство
здравоохранения российской федерации. – Текст : электронный // Техэксперт.
Консорциум Кодекс. – URL: http://docs.cntd.ru/document/901868614 (дата
обращения: 23.04.2020).
90. Применение MALDI-TOF масс-спектрометрии для идентификации
микроорганизмов в гемокультурах пациентов с подозрением на сепсис /
Г. Г. Ломинадзе, Е. А. Семенова, О. В. Мотузова [и др.] // Вопросы диагностики в
педиатрии. – 2013. – № 5(2). – С. 28–32.
91. Припутневич, Т. В. Масс-спектрометрия - новое слово в клинической
микробиологии / Т. В. Припутневич, А. Р. Мелкумян // Клиническая лабораторная
диагностика. – 2016. – Т. 61, № 12. – С. 842–848.
92. Пушкарева, Е. Ю. Диагностика микобактериоза легких, вызванного m.
abscessus (случай из практики) / Е. Ю. Пушкарева, О. И. Альховик // Туберкулез и
болезни легких. – 2014. – Т. 91, № 1. – С. 47–50.
93. Разработка тест-системы для количественной детекции ДНК бактерий
Escherichia coli, Streptococcus agalactia, а также представителей рода Klebsiella
методом ПЦР-РВ в экспресс-диагностике перинатальных инфекций / Н. Н.
Володин, Л. А. Дегтярева, А. Н. Шкопоров [и др.] // Инфекционные болезни. –
2010. – № 2. – С. 42–46.
94. Распространенность нетуберкулезных микобактерий, проблемы
диагностики и лечения микобактериозов / Л. К. Суркова, Е. Р. Сагальчик, Т. Д.
Борисенко [и др.] // Известия национальной академии наук Беларуси. Серия
медицинских наук. – 2011. – № 2. – С. 12–18.
95. Руководство по медицинской микробиологии : в 3 томах. Т. 2.
Оппортунистические инфекции: клинико–эпидемиологические аспекты / под ред.
А. С. Лабинской, Н. Н. Костюкова [и др.]. – Москва : Бином, 2014. – 880 с.
96. Руководство по медицинской микробиологии : в 3 томах.
Т. 1. Оппортунистические инфекции: возбудители и этиологическая диагностика /
под ред. А. С. Лабинская, Н. Н. Костюкова. – Москва : Бином, 2013. – 752 с.

216
97. Руководство по медицинской микробиологии. Общая и санитарная
микробиология / под ред. А. С. Лабинской, Е. Г. Волиной. – Москва : БИНОМ,
2008. – 1080 с.
98. Руководство по медицинской микробиологии. Частная медицинская
микробиология и этиологическая диагностика инфекций / под ред.
А. С. Лабинской, Н. Н. Костюковой, С. М. Ивановой. – Москва : БИНОМ, 2010. –
1152 с.
99. Руководство по микробиологической диагностике инфекций
дыхательных путей у пациентов с муковисцидозом / С. В. Поликарпова, С. В.
Жилина, О. В. Кондратенко [и др.]. – Москва ; Тверь : Триада, 2019. – 128 с.
100. Санитарные правила СП 1.3.2322-08 Безопасность работы с
микроорганизмами III-IV групп патогенности (опасности) и возбудителями
паразитарных инфекций : постановление от 28 янв. 2008 г. № 4 / Главный
государственный санитарный врач РФ. – Текст : электронный // Техэксперт.
Консорциум Кодекс. – URL: http://docs.cntd.ru/document/902091086 (дата
обращения: 23.04.2020).
101. Сачивкина, Н. П. Дифференциально-диагностические аспекты
кандидоза свиней / Н. П. Сачивкина, Е. М. Ленченко, А. В. Лисейцев //
Ветеринария. – 2018. – № 11. – С. 26–30.
102. Сбойчаков, В. Б. Санитарная микробиология / В. Б. Сбойчаков. –
Москва : ГЭОТАР-Медиа, 2007. – 192 с.
103. Севастьянова, Э. В. Современные алгоритмы микробиологической
диагностики туберкулеза / Э. В. Севастьянова, Л. Н. Черноусова // Туберкулез и
болезни легких. – 2018. – Т. 96, № 7. – С. 11–17.
104. Случай микобактериоза органов дыхания, вызванный M. kansasii /
Л. Д. Гунтупова, С. Е. Борисов, Т. П. Гришина [и др.] // Инфекционные болезни. –
2016. – № 2(15). – С. 65–72.
105. Современная микробиология. Прокариоты : в 2 томах. Т. 1 : пер. с англ.
/ под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. – Москва : Мир, 2005. – 656 с.

217
106. Сравнительный анализ методов идентификации нетуберкулезных
микобактерий, выделенных из клинического материала / А. В. Лямин, Д. Д.
Исматуллин, А. В. Жестков [и др.] // Инфекция и иммунитет. – 2017. – № 7(3). – С.
285–291.
107. Тец, В. В. Споробиота: свойства и роль в патологии человека /
В. В. Тец, М. Ф. Вечерковская, Г. В. Тец // Лечебное дело. – 2018. – № 4. – С. 90–
96.
108. Фёдорова, А. В. Использование селективной хромогенной среды для
детекции ванкомицинорезистентных энтерококков / А. В. Фёдорова, Г. А. Клясова
// Клиническая микробиология и антимикробная химиотерапия. – 2018. – Т. 20, №
1. – С. 55–61.
109. Фирсов, Н. Н. Микробиология: словарь терминов / Н. Н. Фирсов. –
Москва : Дрофа, 2006. – 52 с.
110. Частота обнаружения разных видов нетуберкулезных микобактерий в
Москве / М. В. Макарова, М. А. Краснова, Г. Е. Фрейман [и др.] // Туберкулез и
болезни легких. – 2009. – № 9. – С. 29–32.
111. Чункаева, Д. Д. К вопросу о микробиологической диагностике
туберкулеза и лекарственной устойчивости. обзор литературы / Д. Д. Чункаева, А.
А. Мансурова // Наука и здравоохранение. – 2017. – № 6. – С. 116–130.
112. Чучалин, А. Г. Респираторная медицина / А. Г. Чучалин. – Москва :
ГЭОТАР-Медиа, 2007. – 800 с.
113. Шабунова, А. А. Обзор мировых и региональных тенденций
заболеваемости ВИЧ-инфекцией и обусловленной ей смертности / А. А. Шабунова,
О. Н. Калачикова, А. В. Короленко. – Текст : электронный // Социальные аспекты
здоровья населения. – 2017. – № 2. – URL:
http://vestnik.mednet.ru/content/view/824/30/lang,ru_RU.CP1251/ (дата обращения:
23.04.2020).
114. Шепелин, А. П. Питательные среды для энтеробактерий /
А. П. Шепелин, И. А. Дятлов. – Оболенск : Династия, 2017. – 230 с.

218
115. Экспрессдиагностика микроорганизмов, поражающих дыхательные
пути больных муковисцидозом / О. Л. Воронина, М. С. Кунда, Е. И. Аксенова [и
др.] // Клиническая лабораторная диагностика. – 2013. – № 11. – С. 53–58.
116. Экспресс-диагностика острого стрептококкового тонзиллита /
А. С. Дарманян, А. Е. Малахова, Е. В. Старовойтова [и др.] // Вопросы диагностики
в педиатрии. – 2012. – № 4(1). – С. 53–56.
117. Этиологическая структура и антибиотикочувствительность
уропатогенов при хронической рецидивирующей инфекции нижних мочевых
путей / Ю. Л. Набока, И. А. Гудима, Х. С. Ибишев [и др.] // Урология. – 2011. – №
6. – С. 12–15.
118. A comparative study for the detection of Mycobacteria by BACTEC MGIT
960, Lowenstein Jensen media and direct AFB smear examination / S. Rishi, P. Sinha, B.
Malhotra [et al.] // Indian J. Med. Microbiol. – 2007. – Oct., Vol. 25(4). – P. 383–386.
119. A definition of the Mycobacterium avium complex for taxonomical and
clinical purposes, a review / J. van Ingen, C. Y. Turenne, E. Tortoli [et al.] // Int. J. Syst.
Evol. Microbiol. – 2018. – Nov., Vol. 68(11). – P. 3666–3677.
120. A novel culture medium for isolation of rapidly-growing mycobacteria from
the sputum of patients with cystic fibrosis / C. L. Preece, A. Perry, B. Gray [et al.] // J.
Cyst. Fibros. – 2016. – Vol. 15. – P. 186–191.
121. Accurate differentiation of Mycobacterium chimaera from Mycobacterium
intracellulare by MALDI-TOF MS analysis / A. B. Pranada, E. Witt, M. Bienia [et al.] //
J. Med. Microbiol. – 2017. – May, Vol. 66(5). – P. 670–677.
122. Actinomyces Species Isolated from Breast Infections / A. U. Bing, S. F. Loh,
T. Morris [et al.] // J. Clin. Microbiol. – 2015. – Oct., Vol. 53(10). – P. 3247–3255.
123. Actinomycosis: etiology, clinical features, diagnosis, treatment, and
management / F. Valour, A. Sénéchal, C. Dupieux [et al.] // Infect. Drug. Resist. – 2014.
– Jul. 5, Vol. 7. – P. 183–197.
124. Adelman, M. H. Management of nontuberculous mycobacterial pulmonary
disease / M. H. Adelman, D. J. Addrizzo-Harris // Curr. Opin. Pulm. Med. – 2018. – May,
Vol. 24(3). – P. 212–219.

219
125. Advances in Rapid Identification and Susceptibility Testing of Bacteria in
the Clinical Microbiology Laboratory: Implications for Patient Care and Antimicrobial
Stewardship Programs / F. P. Maurer, M. Christner, M. Hentschke // Infect. Dis. Rep. –
2017. – Mar. 30, Vol. 9(1). – P. 6839.
126. American Thoracic Society/Centers for Disease Control and
Prevention/Infectious Diseases Society of America: controlling tuberculosis in the United
States / American Thoracic Society [et al.] // Am. J. Respir. Crit. Care Med. – 2005. –
Vol. 172. – P. 1169–1227.
127. An evaluation of methods for the isolation of nontuberculous mycobacteria
from patients with cystic fibrosis, bronchiectasis and patients assessed for lung
transplantation / D. Stephenson, A. Perry, M. R. Appleby [et al.] // BMC Pulm. Med. –
2019. – Jan. 21, Vol. 19(1). – P. 19.
128. An official ATS/IDSA statement: diagnosis, treatment, and prevention of
nontuberculous mycobacterial diseases / D. E. Griffith, T. Aksamit, B. A. Brown-Elliott
[et al.] // Am. J. Respir. Crit. Care Med. – 2007. – Feb. 15, Vol. 175(4). – P. 367–416.
129. Anderson, A. S. The taxonomy of Streptomyces and related genera /
A. S. Anderson, E. M. H. Wellington // Int. J. Syst. Evol. Microbiol. – 2001. – Vol. 51. –
P. 797–814.
130. Antimicrobial susceptibility of clinical isolates of Actinomyces and related
genera reveals an unusual clindamycin resistance among Actinomyces urogenitalis strains
/ C. Barberis, M. Budia, S. Palombarani [et al.] // J. Glob. Antimicrob. Resist. – 2017. –
Mar., Vol. 8. – P. 115–120.
131. Arenskotter, M. Biology of the metabolically diverse genus Gordonia /
M. Arenskotter, D. Broker, A. Steinbuchel // Appl. Environ Microbiol. – 2004. – Vol. 70,
N 3. – P. 195–204.
132. Assignment of Brevibacterium stationis (ZoBell and Upham 1944) Breed
1953 to the genus Corynebacterium, as Corynebacterium stationis comb. nov., and
emended description of the genus Corynebacterium to include isolates that can alkalinize
citrate / K. A. Bernard, D. Wiebe, T. Burdz [et al.] // International Journal of Systematic
and Evolutionary Microbiology. – 2010. – Vol. 60 (Pt 4, number 4). – P. 874–879.

220
133. Bacteremia and endocarditis caused by a Gordonia species in a patient with
a central venous catheter / O. Lesens, Y. Hansmann, P. Riegel [et al.] // Emerg. Infect.
Dis. – 2000. – Vol. 6, N 4. – P. 382–385.
134. Bacteremia caused by Gordonia bronchialis in a patient with sequestrated
lung / L. H. Sng, T. H. Koh, S. R. Toney [et al.] // J. Clin. Microbiol. – 2004. – Vol. 42,
N 6. – P. 2870–2871.
135. Balada-Llasat, J. M. Identification of mycobacteria from solid and liquid
media by matrix-assisted laser desorption ionization-time of flight mass spectrometry in
the clinical laboratory / J. M. Balada-Llasat, K. Kamboj, P. Pancholi // J. Clin. Microbiol.
– 2013. – Sep., Vol. 51(9). – P. 2875–2879.
136. Beaman, B. L. Interaction of Nocardia asteroides at different phases of
growth with in vitro-maintained macrophages obtained from lungs of normal and
immunized rabbits / B. L. Beaman // Infect. Immun. – 1979. – Vol. 26. – P. 355–361.
137. Bensi, E. P. Incidence of tuberculous and non-tuberculous mycobacteria,
differentiated by multiplex PCR, in clinical specimens of a large general hospital /
E. P. Bensi, P. C. Panunto, M. C. Ramos // Clinics (Sao Paulo). –2013. – Vol. 68(2). – P.
179–184.
138. Bergey's manual of determinative bacteriology / D. H. Bergey, J. G. Holt. –
9th ed. – Baltimore : Williams & Wilkins, 1994.
139. Brown, J. M. Nocardia, Rhodococcus, Gordonia, Actinomadura,
Streptomyces, and other aerobic actinomycetes / J. M. Brown, M. M. McNeil // Manual
of clinical microbiology / ed. P. R. Murray, E. J. Baron, M. A. Pfaller [et al.]. – 8th ed. –
Washington : American Society for Microbiology, 2003. – P. 502–531.
140. Brown-Elliott, B. A. Clinical and taxonomic status of pathogenic
nonpigmented or late-pigmenting rapidly growing mycobacteria / B. A. Brown-Elliott,
R. J. Jr. Wallace // Clin. Microbiol. Rev. – 2002. – Vol. 15. – P. 716–746.
141. Bruker biotyper matrix-assisted laser desorption ionization-time of flight
mass spectrometry system for identification of Nocardia, Rhodococcus, Kocuria,
Gordonia, Tsukamurella, and Listeria species / P. R. Hsueh, T. F. Lee, S. H. Du [et al.]
// J. Clin. Microbiol. – 2014. – Jul., Vol. 52(7). – P. 2371–2379.

221
142. Central venous catheter-associated Nocardia bacteremia in cancer patients /
F. Akhrass, R. Hachem, J. A. Mohamed [et al.] // Emerg. Infect. Dis. – 2011. – Vol. 17,
N 9. – P. 1651–1658.
143. Chater, K. F. Recent advances in understanding Streptomyces / K. F. Chater.
– Text : electronic // F1000 Res. – 2016. – Nov. 30, Vol. 5. – 2795. – URL:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5133688/ (date of access: 09.03.2020).
144. Chen, C. H. Tsukamurella tyrosinosolvens bacteremia with coinfection of
Mycobacterium bovis pneumonia: case report and literature review / C. H. Chen, C. T.
Lee, T. C. Chang // Springer Plus. – 2016. – Vol. 5(1). – P. 2033.
145. Chou, M. P. A spatial epidemiological analysis of nontuberculous
mycobacterial infections in Queensland, Australia / M. P. Chou, A. C. Clements, R. M.
Thomson // BMC Infect. Dis. – 2014. – May 21, Vol. 14. – P. 279.
146. Clinical and microbiological profile of non-tuberculous mycobacterial
endophthalmitis-experience in a tertiary eye care centre in Southern India / R. M. Paulose,
J. Joseph, R. Narayanan [et al.] // J. Ophthalmic. Inflamm. Infect. – 2016. – Dec., Vol.
6(1). – P. 27.
147. Coban, A. Y. Evaluation of blood agar medium for the growth of
mycobacteria / A. Y. Coban, A. Akgüneş, B. Durupınar // Mikrobiyol. Bul. – 2011. –
Oct., Vol. 45(4). – P. 617–622.
148. Comparison of culture methods for isolation of nontuberculous
mycobacteria from surface waters / N. Radomski, E. Cambau, L. Moulin [et al.] // Appl.
Environ Microbiol. – 2010. – Jun., Vol. 76(11). – P. 3514–3520.
149. Comparison of in-house and commercial 16S rRNA sequencing with high-
performance liquid chromatography and genotype AS and CM for identification of
nontuberculous mycobacteria / P. Daley, A. Petrich, K. May [et al.] // Diagn Microbiol.
Infect. Dis. – 2008. – Vol. 61. – P. 284–293.
150. Comparison of LED and conventional fluorescence microscopy for detection
of acid fast bacilli in a lowincidence setting / J. Minion, M. Pai, A. Ramsay [et al.]. – Text
: electronic // PLoSONE. – 2011. – Vol. 6. – e22495. – URL:
https://www.ncbi.nlm.nih.gov/pubmed/21811622 (date of access: 09.03.2020).

222
151. Comparison of MALDI-TOF MS with HPLC and nucleic acid sequencing
for the identification of Mycobacterium species in cultures using solid medium and broth
/ B. W. Buchan, K. M. Riebe, M. Timke [et al.] // Am. J. Clin. Pathol. – 2014. –
Vol. 141(1). – P. 25–34.
152. Comparison of the performance of TK system with LJ and MGIT methods
in the diagnosis of tuberculosis / B. Feyzioglu, M. Dogan, O.O. Sanli [et al.] // Int. J. Clin.
Exp. Med. – 2014. – Apr. 15, Vol. 7(4). – P. 1084–1088.
153. Comparison of the RGM medium and the mycobacterial growth indicator
tube automated system for isolation of non-tuberculous mycobacteria from sputum
samples of cystic fibrosis patients in Belgium / A. Scohy, S. Gohy, V. Mathys [et al.] //
J. Clin. Tuberc. Other Mycobact. Dis. – 2018. – Aug. 1, Vol. 13. – P. 1–4.
154. Comprehensive subspecies identification of 175 nontuberculous
mycobacteria species based on 7547 genomic profiles / Y. Matsumoto, T. Kinjo, D.
Motooka [et al.] // Emerg. Microbes Infect. – 2019. – Vol. 8(1). – P. 1043–1053.
155. Conville, P. S. Nocardia and other aerobic actinomycetes / P. S. Conville,
F. G. Witebsky // Microbiology and Microbial Infections, Bacteriology / ed.
S. P. Boriello, P. R. Murray, G. Funke. – 10th ed. – London : United Kingdom, 2005. –
Vol. 2. – P. 1137–1180.
156. Conville, P. S. Nocardia, Rhodococcus, Gordonia, Actinomadura,
Streptomyces, and other aerobic actinomycetes / P. S. Conville, F. G. Witebsky // Manual
of Clinical Microbiology / ed. P. R. Murray, E. J. Baron, J. H. Jorgensen. – 9th ed. –
Washington : DC, 2007. – Vol. 1. – P. 515–542.
157. Cummins, C. S. Chemical composition and antigenic structure of cell walls
of Corynebacterium, Mycobacterium, Nocardia, Actinomyces and Arthrobacter /
C. S. Cummins // J. Gen. Microbiol. – 1962. – Vol. 28. – P. 35–50.
158. Current methods in the molecular typing of Mycobacterium tuberculosis and
other mycobacteria / T. Jagielski, J. van Ingen, N. Rastogi [et al.]. – Text : electronic //
Biomed. Res. Int. – 2014. – 645802. – URL:
https://www.ncbi.nlm.nih.gov/pubmed/24527454 (date of access: 09.03.2020).

223
159. Davis-Scibienski, C. Interaction of Nocardia asteroides with rabbit alveolar
macrophages: association of virulence, viability, ultrastructural damage and phagosome-
lysosome fusion / C. Davis-Scibienski, B. L. Beaman // Infect. Immun. – 1980. – Vol. 28.
– P. 610–661.
160. De Zwaan, R. Utility of rpoB gene sequencing for identification of
nontuberculous mycobacteria in the Netherlands / R. de Zwaan, J. van Ingen, D. van
Soolingen // J. Clin. Microbiol. – 2014. – Jul., Vol. 52(7). – P. 2544–2551.
161. Description of Bogoriellaceaefam. nov., Dermacoccaceae fam. nov.,
Rarobacteraceae fam. nov. and Sanguibacteraceae fam. nov. and emendation of some
families of the suborder Micrococcineae / E. Stackebrandt, P. Schumann // Int. J. Syst.
Evol. Microbiol. – 1997. – Vol. 50(Part 3). – P. 1279–1285.
162. Detection and characterization of nocardia from patients diagnosed as
tuberculosis in Egypt / Z. H. Helal, M. I. Khan, M. S. Ashour [et al.] // Int. J. Biomed.
Sci. – 2008. – Sep., Vol. 4(3). – P. 179–184.
163. Detection and differentiation of Mycobacterium tuberculosis and
nontuberculous mycobacterial isolates by real-time PCR / N. K. Shrestha, M. J. Tuohy,
G. S. Hall [et al.] // J. Clin. Microbiol. – 2003. – Nov., Vol. 41(11). – P. 5121–5126.
164. Determination of antibiotic resistance and high-performance liquid
chromatography profiles for Mycobacterium species / T. Toka Özer, E. Yula, M. Doğan
[et al.]. – Text : electronic // J. Clin. Lab. Anal. – 2018. – Sep., Vol. 32(7). – e22459. –
URL: https://onlinelibrary.wiley.com/doi/ full/10.1002/jcla.22459 (date of access:
09.03.2020).
165. Differences in time to detection and recovery of Mycobacterium spp.
between the MGIT 960 and the BacT/ALERT MB automated culture systems /
N. Parrish, K. Dionne, A. Sweeney [et al.] // Diagn. Microbiol. Infect. Dis. – 2009. – Vol.
63. – P. 342–345.
166. Distribution of Nocardia species in clinical samples and their routine rapid
identification in the laboratory / G. Wauters, W. Avesani, J. Charlier [et al.] // J. Clin.
Microbiol. – 2005. – Vol. 43. – P. 2624–2628.

224
167. Drancourt, M. Cost-effectiveness of blood agar for isolation of mycobacteria
/ M. Drancourt, D. Raoult. – Text : electronic // PLoS Negl. Trop. Dis. – 2007. – Nov. 28,
Vol. 1(2). – e83. – URL: https://www.ncbi.nlm.nih.gov/ pubmed/18060087 (date of
access: 09.03.2020).
168. Effect of human neutrophils and monocytes on Nocardia asteroides: failure
of killing despite occurrence of the oxidative metabolic burst / G. A. Filice, B. L. Beaman,
J. A. Krick [et al.] // J. Infect. Dis. – 1980. – Vol. 142. – P. 432–438.
169. Effects of decontamination, DNA extraction, and amplification procedures
on the molecular diagnosis of Mycobacterium ulcerans disease (Buruli ulcer) /
D. Affolabi, N. Sanoussi, M. Odoun [et al.] // J. Clin. Microbiol. – 2012. – Vol. 50. – P.
1195–1198.
170. Elborn, J. S. Identification and management of unusual pathogens in cystic
fibrosis / J. S. Elborn // J. R. Soc. Med. – 2008. – Jul., Vol. 101, (Suppl 1). – P. S2–S5.
171. Epidemiology of and risk factors for extrapulmonary nontuberculous
mycobacterial infections in Northeast Thailand / I. Kham-Ngam, P. Chetchotisakd, P.
Ananta [et al.]. – Text : electronic // PeerJ. – 2018. – Aug. 16, Vol. 6. – e5479. – URL:
https://www.ncbi.nlm.nih.gov/pubmed/30128214 (date of access: 09.03.2020).
172. Epidemiology of Nontuberculous Mycobacteria in French Polynesia /
M. Phelippeau, D. A. Osman, D. Musso // J. Clin. Microbiol. – 2015. – Dec., Vol. 53(12).
– P. 3798–3804.
173. Epidemiology of Nontuberculous Mycobacteria Infection in Children and
Young People With Cystic Fibrosis: Analysis of UK Cystic Fibrosis Registry / A. I.
Gardner, E. McClenaghan, G. Saint [et al.] // Clin. Infect. Dis. – 2019. – Feb. 15,
Vol. 68(5). – P. 731–737.
174. Epidemiology of nontuberculous mycobacterial infections in the U.S.
Veterans Health Administration / M. M. Jones, K. L. Winthrop, S. D. Nelson [et al.]. –
Text : electronic // PLoS One. – 2018. – Jun. 13, Vol. 13(6). – e0197976. – URL:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5999224/ (date of access: 09.03.2020).

225
175. Epidemiology of Nontuberculous Mycobacterial Lung Disease and
Tuberculosis, Hawaii, USA / J. Adjemian, T. B. Frankland, Y. G. Daida [et al.] // Emerg.
Infect. Dis. – 2017. – Mar., Vol. 23(3). – P. 439–447.
176. Epidemiology of Pulmonary Nontuberculous Mycobacterial Disease, Japan
/ H. Namkoong, A. Kurashima, K. Morimoto [et al.] // Emerg. Infect. Dis. – 2016. – Jun.,
Vol. 22(6). – P. 1116–1117.
177. Etiological misidentification by routine biochemical tests of bacteremia
caused by Gordonia terrae infection in the course of an episode of acute cholecystitis /
E. Gil-Sande, M. Brun-Otero, F. Campo-Cerecedo [et al.] // J. Clin. Microbiol. – 2006. –
Jul., Vol. 44(7). – P. 2645–2647.
178. Evaluating the clinical significance of nontuberculous mycobacteria isolated
from respiratory samples in Iran: an often-overlooked disease / Z. Mortazavi, A.
Bahrmand, F. Sakhaee // Infect. Drug Resist. – 2019. – Jul. 3, N 12. – P. 1917–1927.
179. Evaluation of a new chromogenic medium, Uriselect 4, for the isolation and
identification of urinary tract pathogens / J. D. Perry, L. A. Butterworth, A. Nicholson [et
al.] // J. Clin. Pathol. – 2003. – Jul., Vol. 56(7). – P. 528–531.
180. Evaluation of a Novel MALDI Biotyper Algorithm to Distinguish
Mycobacterium intracellulare From Mycobacterium chimaera / L. E. Epperson, M.
Timke, N. A. Hasan [et al.] // Front Microbiol. – 2018. – Dec. 18, Vol. 9. – P. 3140.
181. Evaluation of isolation procedures and chromogenic agar media for the
detection of MRSA in nasal swabs from pigs and veal calves / H. Graveland, E. van
Duijkeren, A. van Nes [et al.] // Veterinary Microbiology. – 2009. –Vol. 139, N 1/2. – P.
121–125.
182. Evaluation of matrix-assisted laser desorption ionization-time of flight mass
spectrometry for identification of Nocardia species / A. Verroken, M. Janssens, C. Berhin
[et al.] // J. Clin. Microbiol. – 2010. – Nov., Vol. 48(11). – P. 4015–4021.
183. Evaluation of matrixassisted laser desorption ionization-time-of-flight mass
spectrometry in comparison to 16S rRNA gene sequencing for species identification of
nonfermenting bacteria / A. Mellmann, J. Cloud, T. Maier [et al.] // J. Clin. Microbiol. –
2008. – Vol. 46. – P. 1946–1954.

226
184. Evaluation of RGM Medium for Isolation of Nontuberculous Mycobacteria
from Respiratory Samples from Patients with Cystic Fibrosis in the United States / R.
Plongla, C. L. Preece, J. D. Perry [et al.] // J. Clin. Microbiol. – 2017. – May, Vol. 55(5).
– P. 1469–1477.
185. Evaluation of the MicroSeq system for identification of mycobacteria by 16S
ribosomal DNA sequencing and its integration into a routine clinical mycobacteriology
laboratory / L. Hall, K. A. Doerr, S. L. Wohlfiel [et al.] // J. Clin. Microbiol. – 2003. –
Apr., Vol. 41(4). – P. 1447–1453.
186. Evaluation of the performance of MALDI-TOF MS and DNA sequence
analysis in the identification of mycobacteria species / I. Akyar, C. Çavuşoğlu, M. Ayaş
[et al.] // Turk J. Med. Sci. – 2018. – Dec. 12, Vol. 48(6). – P. 1351–1357.
187. Evaluation of Various Culture Media for Detection of Rapidly Growing
Mycobacteria from Patients with Cystic Fibrosis / Clair L. Preece, Thomas A.
Wichelhaus, Audrey Perry [et al.] // Journal of Clinical Microbiology. – 2016. – Jun., Vol.
54(7). – P. 1797–1803.
188. Extensive cerebral nocardiosis cured with antibiotic therapy alone /
N. Kirmani, C. U. Tuazon, J. A. Ocuin [et al.] // Case report. J. Neurosurg. –1978. – Vol.
49, N 6. – P. 924–928.
189. Factors Associated with Missed Detection of Mycobacterium tuberculosis
by Automated BACTEC MGIT 960 System / Y. Pang, B. Su, H. Zheng [et al.]. – Text :
electronic // Biomed. Res. Int. – 2016. – 5972021. – URL:
https://www.ncbi.nlm.nih.gov/pubmed/28078294 (date of access: 09.03.2020).
190. Falkinham, J. O. Epidemiology of infection by nontuberculous mycobacteria
/ J. O. Falkinham // Clin. Microbiol. Rev. – 1996. – Vol. 9. – P. 178–215.
191. Falkinham, J. O. Mycobacterial aerosols and respiratory disease /
J. O. Falkinham // Emerg. Infect. Dis. – 2003. – Jul., Vol. 9(7). – P. 763–767.
192. First identification of Gordonia sputi in a post-traumatic endophthalmitis
patient - a case report and literatures review / W. Fang, J. Li, H. S. Cui [et al.] // BMC
Ophthalmol. – 2017. – Vol. 17, N 1. – P. 190.

227
193. First isolation of Mycobacterium kyorinense from clinical specimens in
Brazil / C. E. Dias Campos, P. C. de Souza Caldas, H. Ohnishi [et al.] // J. Clin.
Microbiol. – 2012. – Vol. 50. – P. 2477–2478.
194. Gao, B. Phylogenetic Framework and Molecular Signatures for the Main
Clades of the Phylum Actinobacteria / B. Gao, R. S. Gupta // Microbiology and
Molecular Biology Reviews. – 2012. – Vol. 76(1). – P. 66–112.
195. Genetic diversity and population structure of Mycobacterium marinum: new
insights into host and environmental specificities / V. Broutin, A. L. Banuls, A. Aubry [et
al.] // J. Clin. Microbiol. – 2012. – Vol. 50. – P. 3627–3634.
196. Global incidence of multidrug-resistant tuberculosis / M. Zignol,
M. S. Hosseini, A. Wright [et al.] // Infect. Dis. – 2006. – Vol. 194. – P. 479–485.
197. Global tuberculosis report 2012 / World Health Organization. – Switzerland,
Geneva, 2012. – 98 p. – URL: http://www.who/int/tb/publications /global_report/en/
(date of access: 09.03.2020). – Text : electronic.
198. Gordonia araii infection associated with an orthopedic device and review of
the literature on medical device-associated Gordonia infections / D. P. Jannat-Khah, E. S.
Halsey, B. A. Lasker [et al.] // J. Clin. Microbiol. – 2009. – Vol. 47, N. 2. – P. 499–502.
199. Gordonia bronchialis Bacteremia and pleural Infection: case report and
review of the literature / J. A. Johnson, A. B. Onderdonk, L. A. Cosimi [et al.] // J. Clin.
Microbiol. – 2011. – Vol. 49, N. 4. – P. 1662–1666.
200. Gordonia otitidis sp. nov., isolated from a patient with external otitis /
S. Iida, H. Taniguchi, A. Kageyama [et al.] // Int. J. Syst. Evol. Microbiol. – 2005. –Vol.
55, N 5. – P. 1871–1876.
201. Gordonia species: emerging pathogens in pediatric patients that are
identified by 16S ribosomal RNA gene sequencing / A. J. Blaschke, J. Bender,
C. L. Byington [et al.] // Clin. Infect. Dis. – 2007. – Vol. 45. – P. 483–486.
202. Gordonia terrae: a difficult-to diagnose emerging pathogen? / V. Blanc,
M. Dalle, A. Markarian [et al.] // J. Clin. Microbiol. – 2007. – Vol. 45, N 3. – P. 1076–
1077.

228
203. Growth detection failures by the non-radiometric Bactec MGIT 960
mycobacterial culture system / J. A. Pena, M. J. Ferraro, C. G. Hoffmann [et al.] // J. Clin.
Microbiol. – 2012. – Vol. 50. – P. 2092–2095.
204. Gupta, R. S. Phylogenomics and Comparative Genomic Studies Robustly
Support Division of the Genus Mycobacterium into an Emended Genus Mycobacterium
and Four Novel Genera / R. S. Gupta, B. Lo, J. Son // Front Microbiol. – 2018. – Feb. 13,
Vol. 9. – P. 67.
205. Handbook of media for clinical microbiology / by J. W. Snyder, R. M. Atlas.
– 2-nd ed. – CRC Press, 2006. – 544 p.
206. Hillemann, D. Use of the BACTEC Mycobacteria Growth Indicator Tube
960 automated system for recovery of Mycobacteria from 9,558 extrapulmonary
specimens, including urine samples / D. Hillemann, E. Richter, S. Rüsch-Gerdes // J. Clin.
Microbiol. – 2006. – Nov., Vol. 44(11). – P. 4014–4017.
207. How we determined the most reliable solid medium for studying treatment
of tuberculosis / C. M. Heilig, P. J. Feng, M. L. Joloba [et al.] // Tuberculosis (Edinb). –
2014. – May, Vol. 94(3). – P. 317–322.
208. Huitt, G. A. Nontuberculous mycobacteria / G. A. Huitt, C. L. Daley // Clin.
Chest. Med. – 2015. – Vol. 36(1). – P. 11–12.
209. Identification and Differentiation of Clinically Relevant Mycobacterium
Species Directly from Acid-Fast Bacillus-Positive Culture Broth / Haijing Li, Vedat
Turhan, Laxmi Chokhani [et al.] // Journal of Clinical Microbiology. – 2009. – Nov.,
Vol. 47(12). – P. 3814–3820.
210. Identification of Mycobacteria Species by HPLC and Species Distribution
during Five Years at Ulsan University Hospital / J. Jeong, S. R. Kim, C. L. Chang [et al.]
// Korean J. Lab. Med. – 2008. – Feb., Vol. 28(1). – P. 24–33.
211. Identification of mycobacterium spp. and nocardia spp. from solid and liquid
cultures by matrix-assisted laser desorption ionization-time of flight mass spectrometry
(MALDI-TOF MS) / V. Girard, S. Mailler, M. Welker [et al.] // Diagn. Microbiol. Infect.
Dis. – 2016. – Nov., Vol. 86(3). – P. 277–283.

229
212. Identification of Nocardia Species by Matrix-Assisted Laser Desorption
Ionization-Time of Flight Mass Spectrometry / P. D. Khot, B. A. Bird, R. J. Durrant [et
al.] // J. Clin. Microbiol. – 2015. – Oct., Vol. 53(10). – P. 3366–3369.
213. Identification of Nocardia species using matrix-assisted laser
desorption/ionization-time-of-flight mass spectrometry / S. Segawa, M. Nishimura, K.
Sogawa [et al.] // Clin. Proteomics. – 2015. – Mar. 7, Vol. 12(1). – P. 6.
214. Identification of nontuberculous mycobacteria by partial gene sequencing
and public databases / Ines Joao, Paula Cristovao, Liliana Antunes [et al.] // International
Journal of Mycobacteriology. – 2014. – Vol. 3, Issue 2. – P. 144–151.
215. Identification of non-tuberculous mycobacteria: utility of the GenoType
Mycobacterium CM/AS assay compared with HPLC and 16S rRNA gene sequencing /
A. S. Lee, P. Jelfs, V. Sintchenko // Journal of medical microbiology. – 2009. – Vol. 58,
Issue 7. – P. 900–904.
216. Identification of pathogenic Nocardia species by reverse line blot
hybridization targeting the 16S rRNA and 16S-23S rRNA gene spacer regions / M. Xiao,
F. R. Kong, T. C. Sorrell [et al.] // J. Clin. Microbiol. – 2010. – Vol. 48. – P. 503–511.
217. Impact of genotypic studies in mycobacterial taxonomy: the new
mycobacteria of the 1990s / E. Tortoli // Clin. Microbiol. Rev. – 2003. – Vol. 16. –
P. 319–354.
218. Increasing Prevalence of Nontuberculous Mycobacteria in Respiratory
Specimens from US-Affiliated Pacific Island Jurisdictions1 / C. Lin, C. Russell, B. Soll
[et al.] // Emerg. Infect. Dis. – 2018. – Mar., Vol. 24(3). – P. 485–491.
219. Insights from the Genome Sequence of Mycobacterium paragordonae, a
Potential Novel Live Vaccine for Preventing Mycobacterial Infections: The Putative Role
of Type VII Secretion Systems for an Intracellular Lifestyle Within Free-Living
Environmental Predators / B. J. Kim, G. Y. Cha, B. R. Kim [et al.] // Front Microbiol. –
2019. – Jul. 3, Vol. 10. – P. 1524.
220. Intrathoracic Mycobacterium avium complex infection in immunocompetent
children: case report and review / J. E. Fergie, T. W. Milligan, B. M. Henderson [et al.] //
J. Infect. Dis. – 1997. – Vol. 24. – P. 250–253.

230
221. Isolation and phenotypic identification of non-tuberculous mycobacteria
existing in Isfahan different water samples / N. E. Bahram, S. Ensieh, M. Shrareh [et al.].
– Text : electronic // Adv. Biomed. Res. – 2012. – Vol. 1, N 18. –
URL: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3507016/ (date of access:
09.03.2020).
222. Isolation of nontuberculous mycobacteria (NTM) from household water and
shower aerosols in patients with pulmonary disease caused by NTM / R. Thomson, C.
Tolson, R. Carter [et al.] // J. Clin. Microbiol. – 2013. – Sep., Vol. 51(9). – P. 3006–3011.
223. Isolation of Nontuberculous Mycobacteria from the Environment of Ghanian
Communities Where Buruli Ulcer Is Endemic / S. Y. Aboagye, E. Danso, K. A. Ampah
[et al.] // Appl. Environ Microbiol. – 2016. – Jun. 30, Vol. 82(14). – P. 4320–4329.
224. Isolation, biochemical and molecular identification of Nocardia species
among TB suspects in northeastern, Tanzania; a forgotten or neglected threat? /
A. S. Hoza, S. G. S. Mfinanga, I. Moser [et al.] // BMC Infect. Dis. – 2017. – Jun. 8,
Vol. 17(1). – P. 407.
225. Jeffery-Smith, A. Is the Presence of Actinomyces spp. in Blood Culture
Always Significant? / A. Jeffery-Smith, C. Nic-Fhogartaigh, M. Millar // J. Clin.
Microbiol. – 2016. – Apr., Vol. 54(4). – P. 1137–1139.
226. Jeon, D. Infection Source and Epidemiology of Nontuberculous
Mycobacterial Lung Disease / D. Jeon // Tuberc. Respir. Dis. (Seoul). – 2019. – Apr.,
Vol. 82(2). – P. 94–101.
227. Kendall, B. A. Update on the epidemiologyof pulmonary nontuberculous
mycobacterial infections / B. A. Kendall, K. L. Winthrop // Semin. Respir. Crit. Care
Med. – 2013. – Vol. 34. – P. 87–94.
228. Kim, J. U. Multiplex real-time PCR assay and melting curve analysis for
identifying Mycobacterium tuberculosis complex and nontuberculous mycobacteria / J.
U. Kim, C. H. Cha, H. K. An // J. Clin. Microbiol. – 2012. – Feb., Vol. 50(2). – P. 483–
487.

231
229. Kim, S. H. Identification of nontuberculous mycobacteria using multilocous
sequence analysis of 16S rRNA, hsp65, and rpoB / S. H. Kim, J. H. Shin. – Text :
electronic // J. Clin. Lab. Anal. – 2018. – Jan., Vol. 32(1). – e22184. – URL:
http://www.scicombinator.com/articles/3097641 (date of access: 09.03.2020).
230. Koh, W. J. Nontuberculous mycobacterial pulmonary diseases in
immunocompetent patients / W. J. Koh, O. J. Kwon, K. S. Lee // Korean J. Radiol. –
2002. – Jul.-Sep., Vol. 3(3). – P. 145–157.
231. Könönen, E. Actinomyces and related organisms in human infections /
E. Könönen, W. G. Wade // Clin. Microbiol. Rev. – 2015. – Apr., Vol. 28(2). – P. 419–
442.
232. Kozel, T. R. Point-of-Care Testing for Infectious Diseases: Past, Present, and
Future / T. R. Kozel, A. R. Burnham-Marusich // J. Clin. Microbiol. – 2017. – Aug.,
Vol. 55(8). – P. 2313–2320.
233. Kremer, L. A waxy tale, by Mycobacterium tuberculosis / L. Kremer,
G. S. Besra // Tuberculosis and the TubercleBacillus / S. T. Cole, K. D. Eisenach,
D. N. McMur ray [et al.]. – Washington : ASM Press, 2005. – P. 287–305.
234. LED fluorescence microscopy for thediagnosis of pulmonary tuberculosis: a
multi-country cross-sectional evaluation / L. E. Cuevas, A. Al-Sonboli, L. Lawson [et
al.]. – Text : electronic // PLoS Med. – 2011. – Jul., Vol. 8(7). – e1001057. – URL:
https://www.ncbi.nlm.nih.gov/pubmed/21765809 (date of access: 09.03.2020).
235. Long, P. F. A retrospective study of Nocardia infections associated with the
acquired immune deficiency syndrome (AIDS) / P. F. Long // Infection. – 1994. Vol. 2.
– P. 362–364.
236. LPSN – List of Prokaryotic names with Standing in Nomenclature (LPSN)
provides comprehensive information on the nomenclature of prokaryotes and much more.
– Website section «Order Actinomycetales». – URL: https://lpsn.dsmz.de/order/
actinomycetales (date of access: 09.03.2020).
237. Manual of clinical microbiology / ed. J. H. Jorgensen, M. A. Pfaller,
K. C. Carroll. – 11th ed. – Washington : ASM Press, 2015. – Vol. 1. – 2730 p. – URL:
https://b-ok.cc/book/2606391/f27334 (date of access: 09.03.2020). – Text : electronic.

232
238. Matrix-assisted laser desorption ionization-time of flight mass spectrometry
(MALDI-TOF) score algorithm for identification of Gordonia species / M. Ercibengoa
Arana, M. Alonso, P. Idigoras [et al.] // AMB Express. – 2018. – Jul. 23, Vol. 8(1). – P.
121.
239. McNeil, M. M. The medically important aerobic actinomycetes:
epidemiology and microbiology / M. M. McNeil, J. M. Brown // Clin. Microbiol. Rev. –
1994. – Vol. 7, N 3. – P. 357–417.
240. Methods of phenotypic identification of non-tuberculous mycobacteria /
G. S. Bhalla, M. S. Sarao, D. Kalra [et al.]. – Text : electronic // Pract. Lab. Med. – 2018.
– Jul. 18, Vol. 12. – e00107. – URL : https://www.ncbi.nlm.nih.gov/pmc/articles
/PMC6074605/ (date of access: 09.03.2020).
241. Microcalorimetric investigation of the toxic action of ammonium ferric (III)
sulfate on the metabolic activity of pure microbes / F. Wang, J. Yao, L. Tian [et al.] //
Environmental toxicology and pharmacology. – 2008. – Vol. 25. – P. 351–357.
242. Molecular Identification of Nontuberculous Mycobacteria in Humans in
Zimbabwe Using 16S Ribosequencing / N. Chin'ombe, B. Muzividzi, E. Munemo [et al.]
// Open Microbiol. J. – 2016. – May 26, Vol. 10. – P. 113–123.
243. Multicenter study of prevalence of nontuberculous mycobacteria in patients
with cystic fibrosis in France / A. L. Roux, E. Catherinot, F. Ripoll [et al.] // J. Clin.
Microbiol. – 2009. – Dec., Vol. 47(12). – P. 4124–4128.
244. Multicentre evaluation of Ziehl-Neelsen and light-emitting diode
fluorescence microscopy in China / H. Xia, Y. Y. Song, B. Zhao [et al.] // Int. J. Tuberc.
Lung. Dis. – 2013. – Vol. 17. – P. 107–112.
245. Mycobacterium and aerobic actinomycete culture: are two medium types and
extended incubation times necessary? / P. J. Simner, K. A. Doerr, L. K. Steinmetz [et al.]
// J. Clin. Microbiol. – 2016. – Vol. 54, N 4. – P. 1089–1093.
246. Mycobacterium arosiense sp. nov., a slowly growing, scotochromogenic
species causing osteomyelitis in an immunocompromised child / D. Bang, T. Herlin, M.
Stegger [et al.] // Int. J. Syst. Evol. Microbiol. – 2008. – Vol. 58. – P. 2398–2402.

233
247. Mycobacterium avium and Mycobacterium abscessus complex target distinct
cystic fibrosis patient subpopulations / E. Catherinot, A. L. Roux, M. A. Vibet [et al.] //
J. Cyst. Fibros. – 2013. – Vol. 12. – P. 74–80.
248. Mycobacterium europaeum sp. nov., a scotochromogenic species related to
the Mycobacterium simiae complex / E. Tortoli, E. Böttger, A. Fabio [et al.] // Int. J. Syst.
Evol. Microbiol. – 2011. – Vol. 61. – P. 1606–1611.
249. Mycobacterium fragaesp. nov., anon-chromogenicspecies isolated from
human respiratory specimens / J. P. Ramos, C. E. Campos, P. C. Caldas [et al.] // Int. J.
Syst. Evol. Microbiol. – 2013. – Vol. 63. – P. 2583–2587.
250. Mycobacterium haemophilum and lymphadenitis in children /
L. E. Bruijnesteijn van Coppenraet, E. J. Kuijper, J. A. Lindeboom [et al.] // Emerg.
Infect. Dis. – 2005. – Jan., Vol. 11(1). – P. 62–68.
251. Mycobacterium kansasii pericarditis in a kidney transplant recipient: a case
report and comprehensive review of the literature / J. H. Cho, C. H. Yu, M. K. Jin [et al.]
// Transpl. Infect. Dis. – 2012. – Vol. 14. – P. E50–E55.
252. Mycobacterium paraseoulense sp. nov., a slowly growing,
scotochromogenic species related genetically to Mycobacterium seoulense / H. K. Lee,
S. A. Lee, I. K. Lee [et al.] // Int. J. Syst. Evol. Microbiol. – 2010. – Vol. 60. – P. 439–
443.
253. Mycobacterium riyadhense pulmonary infection, France and Britain /
S. Godreuil, H. Marchandin, A. L. Michon [et al.] // Emerg. Infect. Dis. – 2012. – Vol.
18. – P. 176–178.
254. Mycobacterium xenopi and related organisms isolated from stream waters in
Finland and description of Mycobacterium botniense sp. nov / P. Torkko,
S. Suomalainen, E. Iivanainen [et al.] // Int. J. Syst. Evol. Microbiol. – 2000. – Jan.,
Vol. 50 Pt. 1. – P. 283–289.
255. Mycobacterium yongonense sp. nov., a slow-growing non-chromogenic
species closely related to Mycobacterium intracellulare / B. J. Kim, R. K. Math, C. O.
Jeon [et al.] // Int. J. Syst. Evol. Microbiol. – 2013. – Vol. 63. – P. 192–199.

234
256. Native valve endocarditis due to Gordonia polyisoprenivorans: case report
and review of literature of bloodstream infections caused by Gordonia species / P. Verma,
J. M. Brown, V. H. Nunez [et al.] // J. Clin. Microbiol. – 2006. – Vol. 44, N 5. – P. 1905–
1908.
257. Naveen, G. Comparison of the Lowenstein-Jensen Medium, the
Middlebrook 7H10 Medium and MB/BacT for the Isolation of Mycobacterium
tuberculosis (MTB) from Clinical Specimens / G. Naveen, B. V. Peerapur // J. Clin.
Diagn. Res. – 2012. – Dec., Vol. 6(10). – P. 1704–1709.
258. Nocardia aurea sp. nov., a novel actinobacterium isolated from a karstic
subterranean environment / B. Z. Fang, M. X. Han, L. Y. Zhang [et al.] // Int. J. Syst.
Evol. Microbiol. – 2018. – Vol. 69, N 1. – P. 159–164.
259. Nocardiosis in immune disorder disease / M. F. Bafghi, S. S. Eshraghi,
P. Heidarieh [et al.] // Malays J. Med. Sci. – 2014. – Jan., Vol. 21(1). – P. 75–76.
260. Nontuberculous mycobacteria in non-HIV patients: epidemiology, treatment
and response / M. T. Henry, L. Inamdar, D. O'Riordain [et al.] // European Respiratory
Journal. – 2004. – Vol. 23. – P. 741–746.
261. Nontuberculous mycobacteria in respiratory tract infections, eastern Asia /
S. Simons, J. van Ingen, P. R. Hsueh [et al.] // Emerg. Infect. Dis. – 2011. – Mar.,
Vol. 17(3). – P. 343–349.
262. Nontuberculous mycobacteria isolated from specimens of pulmonary
tuberculosis suspects, Northern Tunisia: 2002-2016 / R. Gharbi, B. Mhenni, S. Ben Fraj
[et al.] // BMC Infect. Dis. – 2019. – Sep. 18, Vol. 19(1). – P. 819.
263. Non-tuberculous mycobacterial lung disease prevalence at four integrated
health care delivery systems / D. R. Prevots, P. A. Shaw, D. Strickland [et al.] // Am. J.
Respir. Crit. Care Med. – 2010. – Vol. 182. – Р. 970–976.
264. Non-tuberculous mycobacterial pulmonary infections / J. D. Chalmers,
T. Aksamit, A. C. C. Carvalho [et al.] // Pulmonology. – 2018. – Vol. 24, Issue 2. – P.
120–131.

235
265. Novel mycolic acid-containing bacteria in the family Segniliparaceae fam.
nov., including the genus Segniliparus gen. nov., with descriptions of Segniliparus
rotundus sp. nov. and Segniliparus rugosus sp. nov / R. W. Butler, M. M. Floyd, J. M.
Brown [et al.] // Int. J. Syst. Evol. Microbiol. – 2005. – Vol. 55. – P. 1615–1624.
266. Optimizing diagnosis and management of Nocardia keratitis, scleritis, and
endophtalmatis: 11-year microbial and clinical overview / F. C. DeCroos, P. Garg, A. K.
Reddy [et al.] // Ophthalmology. – 2011. – Vol. 118. – P. 1193–1200.
267. Outbreak of Tsukamurella species bloodstream infection among patients at
an oncology clinic, West Virginia, 2011-2012 / I. See, D. B. Nguyen, S. Chatterjee [et
al.] // Infect. Control Hosp Epidemiol. – 2014. – Mar., Vol. 35(3). – P. 300–306.
268. Ovotransferrin Plays a Major Role in the Strong Bactericidal Effect of Egg
White against the Bacillus cereus Group / F. Baron, S. Jan, F. Gonnet [et al.] // Journal of
food protection. – 2014. – Vol. 77. – P. 955–962.
269. Parte, A. C. LPSN--list of prokaryotic names with standing in nomenclature
/ A. C. Parte // Nucleic Acids Res. – 2014. – Jan. 42(Database issue). – P. D613–D616.
270. Performance of matrix-assisted laser desorption ionization-time of flight
mass spectrometry for identification of bacterial strains routinely isolated in a clinical
microbiology laboratory / A. Bizzini, C. Durussel, J. Bille [et al.] // J. Clin. Microbiol. –
2010. – Vol. 48. – P. 1549–1554.
271. Performance of RGM Medium for Isolation of Nontuberculous
Mycobacteria from Respiratory Specimens from Non-Cystic Fibrosis Patients /
S. Rotcheewaphan, O. E. Odusanya, C. M. Henderson [et al.]. – Text : electronic // J.
Clin. Microbiol. – 2019. – Jan. 30, Vol. 57(2). – e01519-18. – URL:
https://jcm.asm.org/content/57/2/e01519-18 (date of access: 09.03.2020).
272. Phenotypic identification of Actinomyces and related species isolated from
human sources / N. Sarkonen, E. Könönen, P. Summanen [et al.] // J. Clin. Microbiol. –
2001. – Nov., Vol. 39(11). – P. 3955–3961.
273. Phylogeny of mycolic acid-containing actinomycetes / J. Chun, S. O. Kang,
Y. C. Hah [et al.] // J. Ind. Microbiol. – 1996. – Vol. 17. – P. 205–213.

236
274. Physiological roles of ovotransferrin / Francesco Giansanti, Loris Leboffe,
Giuseppina Pitari [et al.] // Biochimica et Biophysica Acta (BBA) - General Subjects. –
2012. – Vol. 1820, Issue 3. – P. 218–225.
275. Polyclonal Mycobacterium avium complex infections in patients with
nodular bronchiectasis / R. J. Wallace, Y. Jr. Zhang, B. A. Brown [et al.] // Am. J. Respir.
Crit. Care Med. – 1998. – Vol. 158. – P. 1235–1244.
276. Prescottia equi gen. nov., comb. nov.: a new home for an old pathogen /
A. L. Jones, I. C. Sutcliffe, M. Goodfellow // Antonie Van Leeuwenhoek. – 2013. –
Vol. 103. – P. 655–671.
277. Prevalence and risk factors of pulmonary nontuberculous mycobacterial
infections in the Zhejiang Province of China / J. Xu, P. Li, S. Zheng [et al.]. – Text :
electronic // Epidemiol. Infect. – 2019. – Sep. 11, Vol. 147. – e269. – URL:
https://www.pubfacts.com/detail/31506134/ (date of access: 09.03.2020).
278. Prevalence of nontuberculous mycobacterial lung disease in U.S. Medicare
beneficiaries / J. Adjemian, K. N. Olivier, A. E. Seitz [et al.] // American Journal of
Respiratory and Critical Care Medicine. – 2012. – Vol. 185. – P. 881–886.
279. Prevalence of Nontuberculous Mycobacterial Pulmonary Disease, Germany,
2009-2014 / F. C. Ringshausen, D. Wagner, A. de Roux [et al.] // Emerg. Infect. Dis. –
2016. – Jun., Vol. 22(6). – P. 1102–1105.
280. Prevention of aerosol isolation of nontuberculous mycobacterium from the
patient's bathroom / K. Morimoto, A. Aono, Y. Murase [et al.]. – Text : electronic // ERJ
Open Res. – 2018. – Jul. 3, Vol. 4(3). – 00150-2017. – URL:
https://openres.ersjournals.com/content/4/3/00150-2017 (date of access: 09.03.2020).
281. Prevots, D. R. Epidemiology of human pulmonary infection with
nontuberculous mycobacteria: a review / D. R. Prevots, T. K. Marras // Clin. Chest. Med.
– 2015. – Mar., Vol. 36(1). – P. 13–34.
282. Prognostic value of American Thoracic Society criteria for non-tuberculous
mycobacterial disease: a retrospective analysis of 120 cases with four years of follow-up
/ H. Kotilainen, V. Valtonen, P. Tukiainen [et al.] // Scand. J. Infect. Dis. – 2013. –
Vol. 45. – P. 194–202.

237
283. Promicromonospora kroppenstedtii sp. nov., isolated from sandy soil /
P. Alonso-Vega, R. I. Santamaría, E. Martínez-Molina [et al.] // Int. J. Syst.
Evol. Microbiol. – 2008. – Vol. 58. – P. 1476–1481.
284. Proposal for a new hierarchic classification system, Actinobacteria classis
nov / E. Stackebrandt, F. A. Rainey, N. L. WardRainey // Int. J. Syst. Bacteriol. – 1997.
– Vol. 47. – P. 479–491.
285. Pseudo-outbreak of Mycobacterium gordonae associated with water from
refrigerated fountains / V. Lalande, F. Barbut, A. Vernerot [et al.] // J. Hosp. Infect. –
2001. – Vol. 48. – P. 76–79.
286. Pulmonary nocardiosis revisited: a case series / D. Aggarwal, K. Garg,
J. Chander [et al.] // Lung India. – 2015. – Vol. 32, N 2. – P. 165–168.
287. Pulmonary nontuberculous mycobacterial disease Ontario, Canada, 1998-
2010 / T. K. Marras, D. Mendelson, A. Marchand-Austin [et al.] // Emerg. Infect. Dis. –
2013. – Nov., Vol. 19(11). – P. 1889–1891.
288. Rapid identification and typing of listeria species by matrix-assisted laser
desorption ionization-time of flight mass spectrometry / S. B. Barbuddhe, T. Maier, G.
Schwarz [et al.] // Appl. Environ Microbiol. – 2008. – Vol. 74. – P. 5402–5407.
289. Recurrent breast abscess caused by Gordonia bronchialis in an
immunocompetent patient / A. M. Werno, T. P. Anderson, S. T. Chambers [et al.] // J.
Clin. Microbiol. – 2005. – Vol. 43, N 6. – P. 3009–3010.
290. Réhault-Godbert, S. The Golden Egg: Nutritional Value, Bioactivities, and
Emerging Benefits for Human Health / S. Réhault-Godbert, N. Guyot, Y. Nys //
Nutrients. – 2019. – Mar 22, Vol. 11(3). – P. 684.
291. Revisited distribution of nonfermenting Gramnegative bacilli clinical
isolates / E. Carbonnelle, J. L. Beretti, S. Cottyn [et al.] // Eur. J. Clin. Microbiol. Infect.
Dis. – 2011. – Dec., Vol. 30(12). – P. 1579–1586.
292. Rhodococcus fascians infection after haematopoietic cell transplantation: not
just a plant pathogen? / M. C. Austin, T. S. Hallstrand, D. R. Hoogestraat [et al.]. – Text
: electronic // JMM Case Rep. – 2016. – Mar. 3, Vol. 3(2). – e005025. – URL:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5330220/ (date of access: 09.03.2020).

238
293. Risk for Mycobacterial Disease among Patients with Rheumatoid Arthritis,
Taiwan, 2001-2011 / T. L. Liao, C. H. Lin, G. H. Shen [et al.] // Emerg. Infect. Dis. –
2015. – Aug., Vol. 21(8). – P. 1387–1395.
294. Safaei, S. Role of Tsukamurella species in human infections: first literature
review / S. Safaei, M. Fatahi-Bafghi, O. Pouresmaeil // New Microbes New Infect. –
2017. – Oct. 10, Vol. 22. – P. 6–12.
295. Sakatani, M. Treatment of non-tuberculous pulmonary mycobacteriosis /
M. Sakatani, Y. Nakajima // Kekkaku: [Tuberculosis]. – 2006. – Vol. 81, N 1. – Р. 35–
50.
296. Saleeb, P. Pulmonary nontuberculous mycobacterial disease: new insights
into risk factors for susceptibility, epidemiology, and approaches to management in
immunocompetent and immunocompromised patients / P. Saleeb, K. N. Olivier // Curr.
Infect. Dis. Rep. – 2010. – May, Vol. 12(3). – P. 198–203.
297. Saubolle, M. A. Nocardiosis review of clinical and laboratory experience /
M. A. Saubolle, D. Sussland // J. Clin. Microbiol. – 2003. – Vol. 41, N 10. – P. 4497–
4501.
298. Seven isolates of Actinomyces turicensis from patients with surgical
infections of the anogenital area in a Czech hospital / E. Chudácková, L. Geigerová,
J. Hrabák [et al.] // J. Clin. Microbiol. – 2010. – Jul., Vol. 48(7). – P. 2660–2661.
299. Shaer, A. Tsukamurella peritonitis associated with continuous ambulatory
peritoneal dialysis / A. Shaer, C. Gadegbeku // Clin. Nephrol. – 2001. – Vol. 56. – P. 241–
246.
300. Shourian, M. Resistance and Tolerance to Cryptococcal Infection: An
Intricate Balance That Controls the Development of Disease / M. Shourian, S. T. Qureshi
// Front Immunol. – 2019. – Jan. 29, Vol. 10. – P. 66.
301. Spatial clusters of nontuberculous mycobacterial lung disease in the United
States / J. Adjemian, K. N. Olivier, A. E. Seitz [et al.] // Am. J. Respir. Crit. Care Med. –
2012. – Sep. 15, Vol. 186(6). – P. 553–558.

239
302. Species-Specific Risk Factors, Treatment Decisions, and Clinical Outcomes
for Laboratory Isolates of Less Common Nontuberculous Mycobacteria in Washington
State / E. S. Ford, D. J. Horne, J. A. Shah [et al.] // Ann. Am. Thorac. Soc. – 2017. – Jul.,
Vol. 14(7). – P. 1129–1138.
303. Sporulation in mycobacteria / J. Ghosh, P. Larsson, B. Singh [et al.] // Proc.
Natl. Acad. Sci U S A. – 2009. – Vol. 106. – P. 10781–10786.
304. Staneck, J. L. Simplified approach to identification of aerobic actinomycetes
by thin-layer chromatography / J. L. Staneck, G. D. Roberts // Appl. Microbiol. – 1974.
– Vol. 28, N 2. – P. 226–231.
305. Sternal wound infection caused by Gordonia bronchialis: identification by
MALDI-TOF MS / J. Rodriguez-Lozano, E. Pérez-Llantada, J. Agüero [et al.]. – Text :
electronic // JMM Case Rep. – 2016. – Oct. 31, Vol. 3(5). – e005067. – URL:
https://www.pubfacts.com/detail/28348789/Sternal-wound-infection-caused-by-
identification-by-MALDI-TOF-MS (date of access: 09.03.2020).
306. Survey of 150 strains belonging to the Mycobacterium terrae complex and
description of Mycobacterium engbaekiisp. nov., Mycobacterium heraklionense sp. nov.
and Mycobacterium longobardum sp. nov / E. Tortoli, Z. Gitti, H. P. Klenk [et al.] // Int.
J. Syst. Evol. Microbiol. – 2013. – Vol. 63. – P. 401–411.
307. Ten year review of pulmonary nocardiosis: a series of 55 cases / K. Zia,
T. Nafees, M. Faizan [et al.] // Cureus. – 2019. – Vol. 11(5). – P. 4759.
308. The anti-bacterial iron-restriction defence mechanisms of egg white; the
potential role of three lipocalin-like proteins in resistance against Salmonella /
L. A. Julien, F. Baron, S. Bonnassie [et al.] // Biometals. – 2019. – Vol. 32. – P. 453–467.
309. The emergence of non-albicans candidemia and evaluation of HiChrome
Candida differential agar and VITEK2 YST® platform for differentiation of Candida
bloodstream isolates in teaching hospital Kandy, Sri Lanka / M. Kothalawala, J. A. A. S.
Jayaweera, S. Arunan [et al.] // BMC Microbiol. –2019. – Jun. 21, Vol. 19(1). – P. 136.
310. The influence of culture conditions on the identification of Mycobacterium
species by MALDI-TOF MS profiling / T. Balážová, J. Makovcová, O. Sedo [et al.] //
FEMS Microbiol. Lett. – 2014. – Apr. 17, Vol. 353(1). – P. 77–84.

240
311. The Nutraceutical Properties of Ovotransferrin and Its Potential Utilization
as a Functional Food / F. Giansanti, L. Leboffe, F. Angelucci [et al.] // Nutrients. – 2015.
– Nov. 4, Vol. 7(11). – P. 9105–9115.
312. The Rise of Non-Tuberculosis Mycobacterial Lung Disease / C. N.
Ratnatunga, V. P. Lutzky, A. Kupz [et al.]. – Text : electronic // Front Immunol. – 2020.
– Mar. 3. – URL: https://www.frontiersin.org/articles/10.3389/fimmu. 2020.00303/full
(date of access: 09.03.2020).
313. The Role of MGIT 960 Culture Medium in Resolving the Diagnostic
Dilemma for Genital Tuberculosis Patients Presenting with Infertility / N. Jindal,
S. Gainder, L. K. Dhaliwal [et al.] // J. Obstet. Gynaecol. India. – 2018. – Apr., Vol. 68(2).
– P. 123–128.
314. The salicylate–derived mycobactin siderophores of Mycobacterium
tuberculosis are essential for growth in macrophages / J. J. De Voss, K. Rutter, B. G.
Schroeder [et al.] // Proc. Natl. AcadSci U S A. – 2000. – Feb. 1, Vol. 97(3). – P. 1252–
1257.
315. The struggle for iron a metal at the host–pathogen interface / M. Nairz,
A. Schroll, T. Sonnweber [et al.] // Cellular Microbiology. – 2010. – Vol. 12, N 12. – P.
1691–1702.
316. Tsukamurella catheter-related bloodstream infection in a pediatric patient
with pulmonary hypertension / K. A. Wendorf, C. M. Espinosa, W. D. Lebar [et al.]. –
Text : electronic // Infect. Dis. Rep. – 2010. – Mar. 17, Vol. 2(1). – e5. – URL:
https://www.pagepress.org/journals/index.php/idr/article/view/1649 (date of access:
09.03.2020).
317. Uesaka, I. On the morphology of Nocardia, especially on its acid-fastness /
I. Uesaka, N. M. McClung // Jpn. J. Tuberc. – 1961. – Vol. 8. – P. 116–117.
318. Vazquez-Boland, J. A. Rhodococcus equi: the many facets of a pathogenic
actinomycete / J. A. Vazquez-Boland // Vet. Microbiol. – 2013. – Vol. 167(1/2). – P. 9–
33.

241
319. Vilchèze, C. Acid-fast positive and acid-fast negative Mycobacterium
tuberculosis: The koch paradox / C. Vilchèze, L. Kremer // Tuberculosis and the Tubercle
Bacillus / Jr. W. Jacobs, H. McShane, V. Mizrahi [et al.]. – Washington : ASM Press,
2017. – P. 519–532.
320. Wayne, L. G. Mycobacteria / L. G. Wayne, G. P. Kubica // Bergey’s
Manual of Systemic Bacteriology / ed. P. H. A. Sneath. – Baltimore : William’s &
Wilkins, 1986. – Vol. 2. – P. 1435–1457.
321. Wheeler, P. R. General metabolism and biochemical pathways of tubercle
bacilli / P. R. Wheeler, J. S. Blanchard // Tuberculosis and the Tubercle Bacillus / ed.
S. T. Cole, K. D. Eisenach, D. N. McMurray. – Washington : ASM Press, 2005. – P. 309–
339.
322. Williams, M. D. Effect of Cetylpyridinium Chloride (CPC) on Colony
Formation of Common Nontuberculous Mycobacteria / M. D. Williams, J. O. Falkinham
// Pathogens. – 2018. – Oct. 5, Vol. 7(4). – P. 79.
323. Willis, M. S. Nocardia meningitis / M. S. Willis, D. Cavuoti, R. Gander //
Lab. Med. – 2001. – Vol. 32, N 11. – P. 695–699.
324. Wilson, J. W. Nocardiosis: updates and clinical overview / J. W. Wilson //
Mayo. Clin. Proc. – 2012. – Vol. 87, N 4. – P. 403–407.
325. Zink, M. C. Experimental infection of piglets by aerosols of Rhodococcus
equi / M. C. Zink, J. A. Yager // Can. J. Vet. Res. – 1987. – Jul., Vol. 51(3). – P. 290–
296.




















