
Diminished Production of Human Immunodeficiency Virus Type 1 in Astrocytes Results from Inefficient...
11
1999, 73(1):352. J. Virol. A. McPhee and Damian F. J. Purcell L. Anderson, Anthony Cunningham, Deborah Adrian, Dale Paul R. Gorry, Jane L. Howard, Melissa J. Churchill, Jenny despite Efficient Expression of Tat and Rev mRNAs nef , and env , gag Translation of Astrocytes Results from Inefficient Immunodeficiency Virus Type 1 in Diminished Production of Human http://jvi.asm.org/content/73/1/352 Updated information and services can be found at: These include: REFERENCES http://jvi.asm.org/content/73/1/352#ref-list-1 at: This article cites 51 articles, 21 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on January 11, 2014 by guest http://jvi.asm.org/ Downloaded from on January 11, 2014 by guest http://jvi.asm.org/ Downloaded from
Transcript of Diminished Production of Human Immunodeficiency Virus Type 1 in Astrocytes Results from Inefficient...

1999, 73(1):352. J. Virol.
A. McPhee and Damian F. J. PurcellL. Anderson, Anthony Cunningham, Deborah Adrian, Dale Paul R. Gorry, Jane L. Howard, Melissa J. Churchill, Jenny despite Efficient Expression of Tat and Rev
mRNAsnef, and env, gagTranslation ofAstrocytes Results from InefficientImmunodeficiency Virus Type 1 in Diminished Production of Human
http://jvi.asm.org/content/73/1/352Updated information and services can be found at:
These include:
REFERENCEShttp://jvi.asm.org/content/73/1/352#ref-list-1at:
This article cites 51 articles, 21 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
on January 11, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

JOURNAL OF VIROLOGY,0022-538X/99/$04.0010
Jan. 1999, p. 352–361 Vol. 73, No. 1
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Diminished Production of Human Immunodeficiency VirusType 1 in Astrocytes Results from Inefficient Translation
of gag, env, and nef mRNAs despite EfficientExpression of Tat and Rev
PAUL R. GORRY,1,2 JANE L. HOWARD,1 MELISSA J. CHURCHILL,3,5 JENNY L. ANDERSON,1,3
ANTHONY CUNNINGHAM,4,5 DEBORAH ADRIAN,4,5 DALE A. MCPHEE,1,5
AND DAMIAN F. J. PURCELL1*
AIDS Cellular1 and Molecular3Biology Units, Macfarlane Burnet Centre for Medical Research, Fairfield 3078, andDepartment of Medical Laboratory Science, RMIT University, Melbourne 3001,2 Victoria, Centre for Virus Research,
Westmead Institutes of Health Research, Westmead Hospital, University of Sydney, Westmead 2145,New South Wales,4 and National Centre in HIV Virology Research, Fairfield 3078, Victoria,5Australia
Received 18 May 1998/Accepted 14 October 1998
Astrocytes infected with human immunodeficiency virus type 1 (HIV-1) produce only minimal quantities ofvirus. The molecular events that limit acute-phase HIV-1 infection of astrocytes were examined after inducingacute-phase replication by transfection with the pNL4-3 proviral plasmid. The levels of HIV-1 mRNA weresimilarly high in both astrocytes and HeLa cells, but astrocytes produced approximately 50-fold less super-natant p24 than HeLa cells. We found that diminished HIV-1 production in astrocytes resulted from inefficienttranslation of gag, env, and nef mRNAs that were efficiently transported to the cytoplasm. Tat- or Rev-dependent reporter constructs showed no defect in Tat or Rev function in astrocytes compared with HeLa cells.HIV-1 mRNAs were correctly spliced, but only Rev and Tat proteins were efficiently translated from their nativemRNAs. Pulse-chase labelling and immunoblot experiments revealed no defect in protein processing, but levelsof Gag, Env, or Nef protein expressed were dramatically reduced in astrocytes compared to HeLa cells. Theseresults demonstrate that inefficient translation of HIV-1 structural proteins underlies the restricted infectionof astrocytes. The efficient expression of functional Tat and Rev by astrocytes may contribute to HIV-1neuropathogenesis.
Human immunodeficiency virus type 1 (HIV-1) infection ofthe central nervous system occurs early in the course of HIV-1disease. HIV-1 productively infects brain macrophages andmicroglial cells (3, 7, 18, 24, 43). A subpopulation of astrocytesis also consistently infected (3, 41, 43), but astrocyte infectionyields little progeny virus (6, 14, 21, 25, 36, 49, 50). However,nonproductively infected astrocytes may be activated to pro-duce virus that can be transferred to other susceptible cells (6,19, 50). Thus, infected astrocytes might serve as a reservoir ofHIV-1 in the brain and could be a sanctuary site for HIV-1 thatmight thwart virus eradication by current therapeutic regi-mens. Astrocytes are terminally differentiated in adult braintissue and are crucial for neuronal function and survival.Therefore, elimination of HIV-1-infected astrocytes by thera-peutic or immunological means would be detrimental. A pref-erable approach would permanently lock HIV-1 into a dor-mant state in astrocytes. However, to achieve this, the precisemechanisms that initiate and maintain the latent infection ofastrocytes must be understood. Our present study examinedthe initial events during acute-phase viral replication that driveHIV-1 infection of astrocytes toward a dormant state.
Most previous studies on HIV-1 replication in astrocyteshave focused on the transcriptional control of stably integratedHIV-1 in long-term- or latently infected cells. During this dor-
mant phase that follows the initial infection, restricted expres-sion of virus results from low-level basal long terminal repeat(LTR) activity which can be modestly induced by cytokines orother chemical stimuli (13, 32, 40, 42). In contrast, duringacute-phase virus replication in astrocytes, low-level virus pro-duction appears to be controlled posttranscriptionally, sincehigh levels of HIV-1 mRNA are synthesized after transfectionwith a proviral plasmid (19, 49). These observations suggestthat there may be two phases of entry into the dormant state,one operating initially to suppress virion production despitehigh-level RNA synthesis and another that eventually sup-presses RNA transcription. Transcriptional repression in long-term-infected cells is not unique to astrocytes (10, 16, 17), butthe action of a novel central nervous system-derived moleculethat promotes TAR-independent transcription in the presenceof Tat indicates that unique transcriptional regulatory mecha-nisms may exist in astrocytes (28, 45–47, 53).
Earlier studies using a persistently HIV-1-infected astrocytecell line, TH4-7-5, demonstrated that inefficient HIV-1 Revfunction prevented the nucleocytoplasmic export and subse-quent expression of gag and env RNAs bearing the Rev-re-sponsive element (RRE) (31). Here, we examined acute-phaseHIV-1 replication in astrocytes following transfection of aninfectious molecular clone to examine whether an inactiveRev-RRE regulatory axis leads to the restricted expression ofvirus during the initial infection. We showed that astrocytesacutely infected by HIV-1 produce high levels of viral mRNA,which is correctly spliced and efficiently translocated to thecytoplasm. Efficient RNA expression in astrocytes is associatedwith normal expression and function of Rev and Tat proteins.
* Corresponding author. Mailing address: Macfarlane Burnet Cen-tre for Medical Research, P.O. Box 254, Fairfield, Victoria 3078, Aus-tralia. Phone: 61-3-9282-2256. Fax: 61-3-9282-2100. E-mail: [email protected].
352
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

However, the major HIV-1 structural and Nef proteins areexpressed at low levels in astrocytes during the acute infectionphase, despite high levels of available mRNA and the efficienttranslation of a coexpressed, non-HIV-1 reporter. We showedthat diminished expression of these proteins in astrocytes re-sults from an HIV-1-specific restriction in the translation ofgag, env, and nef mRNAs to proteins. This severely reduces thesynthesis of progeny virions and initiates the nonproductive orlatent infection. The efficient translation of Tat and Rev pro-teins by HIV-1-infected astrocytes may contribute to AIDSneuropathogenesis.
MATERIALS AND METHODS
Cell culture. The astrocytoma cell line U251MG (5) was obtained from J.Kort, Department of Medicine, Albany Medical College, Albany, N.Y., and wasmaintained in Dulbecco’s modified Eagle medium (DMEM) supplemented with10% (vol/vol) fetal calf serum, penicillin (100 U/ml), streptomycin (100 mg/ml),and glutamine (25 mg/ml) (DMEM-10). Primary fetal astrocytes (PFA), obtainedfrom fetal abortuses and judged to be .99% pure by positivity to the astrocyte-specific marker glial fibrillary acidic protein (GFAP), were cultured inDMEM-10 for up to 10 weeks postisolation. HeLa cells (39) were also culturedin DMEM-10.
Plasmids and cell transfection. The HIV-1 proviral plasmids pNL4-3 (1) andpNL4-3Tat2, which has two premature stop codons introduced into the tat openreading frame (23), were obtained from M. Martin (Laboratory of MolecularMicrobiology, National Institute of Allergy and Infectious Diseases, NationalInstitutes of Health, Bethesda, Md.). pNL4-3Rev2 contains an ACG pointmutation at the initiating Met codon of the rev gene and was prepared viasite-directed mutagenesis by using the pALTER system (Promega, Madison,Wis.). The Rev reporter plasmid, pDM128, expresses the chloramphenicolacetyltransferase (CAT) gene from an intron bearing the RRE in the presence ofHIV-1 Rev protein (22). pEGFP-N1 (Clontech, Palo Alto, Calif.) producesgreen fluorescent protein (GFP) under the control of a cytomegalovirus pro-moter. pLTR-EGFP, in which the AseI-to-HindIII cytomegalovirus promoterfragment of pEGFP-N1 is replaced with the XhoI-to-HindIII 39-LTR fragmentfrom pNL4-3, expresses high levels of GFP in the presence of HIV-1 Tat.pRSV-Rev produces Rev protein under the direction of a Rous sarcoma viruspromoter (22). In pCMV-hGH, the GFP fragment of pEGFP-N1 was replacedwith the human growth hormone (hGH) gene. Unless indicated otherwise, cellswere cotransfected with 20 mg of proviral plasmid plus 5 mg of GFP-expressingplasmid and 2 mg of pCMV-hGH plasmid by the calcium phosphate method aspreviously described (19).
Antibodies. BB10 (a gift from E. Dax, National HIV Reference Laboratory,Fairfield, Victoria, Australia) is pooled human HIV-1-immune serum that rec-ognizes the major HIV-1 structural proteins p24, p55, and gp120. Sheep anti-Nef15–27 (provided by A. Greenway) is an affinity-purified polyclonal antibodyraised against a peptide corresponding to amino acids 15 to 27 of HIV-1NL Nef(20).
hGH and HIV-1 p24 assays. HIV-1 p24 antigen production was measured byenzyme immunoassay in accordance with the manufacturer’s instructions (Or-ganon Teknika, Durham, N.C.). hGH production was measured from 100 ml ofculture supernatant by radioimmunoassay (Nichols Institute Diagnostics, SanJuan Capistrano, Calif.).
Analysis of HIV-1 mRNA. Total cell RNA was extracted by using TRIzolreagent in accordance with the manufacturer’s protocol (BRL Life Technologies,Gaithersburg, Md.). Cytoplasmic RNA extracts were prepared from 106 trans-fected cells that were washed three times in phosphate-buffered saline (PBS) andthen hypotonically swollen in 3 ml of a solution consisting of 10 mM KCl, 1.5 mMMgCl2 and 10 mM HEPES (pH 7.9) for 10 min on ice. Cells were lysed in 200ml of ice-cold cytoplasmic extraction buffer (150 mM NaCl; 10 mM Tris-HCl, pH8.0; 2 mM MgCl2; 0.5% [vol/vol] Nonidet P-40 [NP-40]; 10 mM ribonucleosidecomplexes), and nondisruption of cell nuclei was confirmed by light microscopy.Cell lysates were vortexed for 10 s and centrifuged at 1,700 3 g for 5 min to pelletintact nuclei. The supernatant was collected and centrifuged again to remove anyresidual nuclei. One hundred eighty microliters of supernatant was added to 1 mlof TRIzol reagent, the mixture was gently inverted, and RNA was purified asabove. Northern blot detection of the three major HIV-1 mRNA classes intotal-cell and cytoplasmic RNA extracts was performed with a 39 HIV-1 LTRregion PCR product labelled with [a-32P]dCTP as previously described (19).
First-strand cDNA was synthesized from total RNA extracts with avian my-eloblastosis virus reverse transcriptase, using a cDNA cycle kit and an oligo(dT)primer (Invitrogen, Carlsbad, Calif.). PCR for spliced HIV-1 mRNA species wasperformed by a method described previously (34), with the following modifica-tions: for PCR analysis of cDNA from the 4.0-kb mRNA class, the oligonucle-otide primers Odp.045 (59-CTGAGCCTGGGAGCTCTCTG-39, positions 477to 499 of HIV-1NL) and Odp.084 (59-TCATTGCCACTGTCTTCTGCTCT-39,positions 6202 to 6225 of HIV-1NL) were used. Amplification products wereradiolabelled by performing a single round of PCR, replacing dCTP with 10 mCi
of [a-32P]dCTP, and then analyzed by electrophoresis on a 6% polyacrylamide-urea gel and autoradiographed. Individual HIV-1 mRNA species were namedaccording to the nomenclature of Purcell and Martin (34).
CAT assays. Cells were trypsinized 72 h after cotransfection, washed twice inPBS, resuspended in 120 ml of 250 mM Tris-HCl (pH 7.8) with 0.5% (vol/vol)NP-40, and then subjected to three rapid freeze-rapid thaw cycles to induce lysis.The tubes were vortexed for 10 s between each freeze-thaw cycle. Lysates werecentrifuged at 15,300 3 g for 5 min at 4°C, supernatants were heat inactivated at65°C for 10 min and then centrifuged at 15,300 3 g for 5 min at 4°C, and theresultant supernatants were harvested for CAT assay. To control for transfectionefficiency, the amount of cell lysate assayed was adjusted according to hGHproduction by dilution with 1 M Tris-HCl (pH 7.8), and then 25 ml of thissupernatant was added to 25 ml of 1 M Tris-HCl (pH 7.8) and reacted with 10 mlof acetyl coenzyme A (Boehringer, Mannheim, Germany; 3.5 mg/ml) and 5 ml ofD-threo[dichloracetyl-1,2-14C]chloramphenicol (NEN, Boston, Mass.; 57 mCi/mmol) at 37°C for 6 to 24 h, depending on the experiment. Incubation times werevaried to measure CAT activity in the linear range (,30% conversion). Reactionmixtures were extracted with ethyl acetate and analyzed by thin-layer chroma-tography. Acetylation by CAT protein was assessed by autoradiography andquantified with a model FLA-2000 phosphorimager (Fuji, Tokyo, Japan).
Assessment of GFP fluorescence. Cells transfected with pEGFP-N1 or pLTR-EGFP were fixed in 1% (vol/vol) formaldehyde (ultrapure electron microscopygrade; Polysciences, Warrington, Pa.) and analyzed for the percentage and meanfluorescence of GFP-expressing cells on a FACSCalibur flow cytometer (BectonDickinson, San Jose, Calif.) at 48 or 72 h posttransfection. To account for boththe number and intensity of fluorescent cells in experiments using the pLTR-EGFP reporter, the amount of fluorescence was determined from the fluores-cence-activated cell sorter (FACS) profiles by multiplying the percentage ofpositive cells by the mean channel fluorescence.
Immunoblotting. At 72 h after transfection, cells (106) were washed twice inPBS and resuspended in 200 ml of ice-cold lysis buffer (0.5% [vol/vol] NP-40;0.5% [wt/vol] sodium deoxycholate; 50 mM NaCl; 25 mM Tris-HCl, pH 8.0; 10mM EDTA; 5 mM benzamidine HCl; 10 mM phenylmethylsulfonyl fluoride) for10 min, and then cellular debris was removed by centrifugation at 15,300 3 g for10 min. Lysate concentrations were standardized for transfection efficiencyagainst hGH production, and 20 ml of lysate was added to 5 ml of 53 sodiumdodecyl sulfate (SDS) gel loading buffer (10% [wt/vol] SDS; 500 mM dithiothre-itol; 300 mM Tris-HCl, pH 6.8; 0.001% [wt/vol] bromophenol blue). Sampleswere boiled for 3 min and then electrophoresed on SDS–13% polyacrylamidegels, and proteins were transferred to Hybond-C nitrocellulose membranes (Am-ersham, Buckinghamshire, England). Nonspecific binding of membranes wasblocked with Tris-buffered saline containing 3% (wt/vol) casein and 0.3% (vol/vol) Tween 20 for 16 h. After being washed four times with Tris-buffered salinecontaining 0.3% (vol/vol) Tween 20, the membranes were probed with eitherpolyclonal HIV-1 antiserum BB10 (1:5,000) or sheep anti-Nef15–27 (1:500). Afterbeing subjected to four more washing steps as described above, the membranewas incubated with a horseradish peroxidase-conjugated antibody to human orsheep immunoglobulin G (1:5,000) for 1 h and then washed seven times asdescribed above, and immunoreactive proteins were detected by enhancedchemiluminescence (ECL; Amersham).
Pulse-chase labelling and immunoprecipitation. Cells were transfected with20 mg of pNL4-3 and 5 mg pEGFP-N1 and cultured until 24 h before the peak ofp24 production (48 h for HeLa cell transfectants or 72 h for U251MG transfec-tants). Transfected (or mock-transfected) cells (6 3 106) were washed twice inPBS and starved in 6 ml of DMEM lacking methionine and cysteine (ICN, CostaMesa, Calif.) but containing 2% dialyzed fetal calf serum for 30 min at 37°C.Cells were pulse-labelled for 1 h by adding 500 mCi of 35S-labelled methionine-cysteine (NEN) after suspension in 600 ml of starvation medium. Labelled cellswere washed twice in PBS, resuspended in 6 ml of DMEM-10 medium, anddivided into six tubes (106 cells each). Cells were incubated at 37°C and harvestedat 0, 1, 2, 3, 4, and 6 h post-pulse-labelling. Preparation of cell and virion lysatesand immunoprecipitation of HIV-1 structural proteins with pooled HIV-1-im-mune serum were performed as previously described (51). Immunoprecipitation forRev protein was performed with a sheep antiserum to glutathione S-transferase–Rev(ICN) from lysates of 6 3 106 transfected HeLa or U251MG cells that were radio-labelled with 500 mCi of 35S-labelled methionine-cysteine for 16 h.
RESULTS
Inefficient expression of p24 antigen in PFA and U251MGastrocytoma cells acutely replicating HIV-1. In vitro infectionof astrocytes with cell-free HIV-1 is highly inefficient at initi-ating measurable virus replication. To ensure sufficient syn-chronous expression of HIV-1, we cotransfected either PFA orthe U251MG astrocytoma cell line with the pNL4-3 HIV-1proviral plasmid, as well as with pEGFP-N1 to monitor thetransfection efficiency. When compared to control HeLa celltransfections, the primary astrocytes and U251MG astrocy-toma cells synthesized 22- and 30-fold less p24 antigen, respec-
VOL. 73, 1999 RESTRICTED TRANSLATION OF HIV-1 mRNAs IN ASTROCYTES 353
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
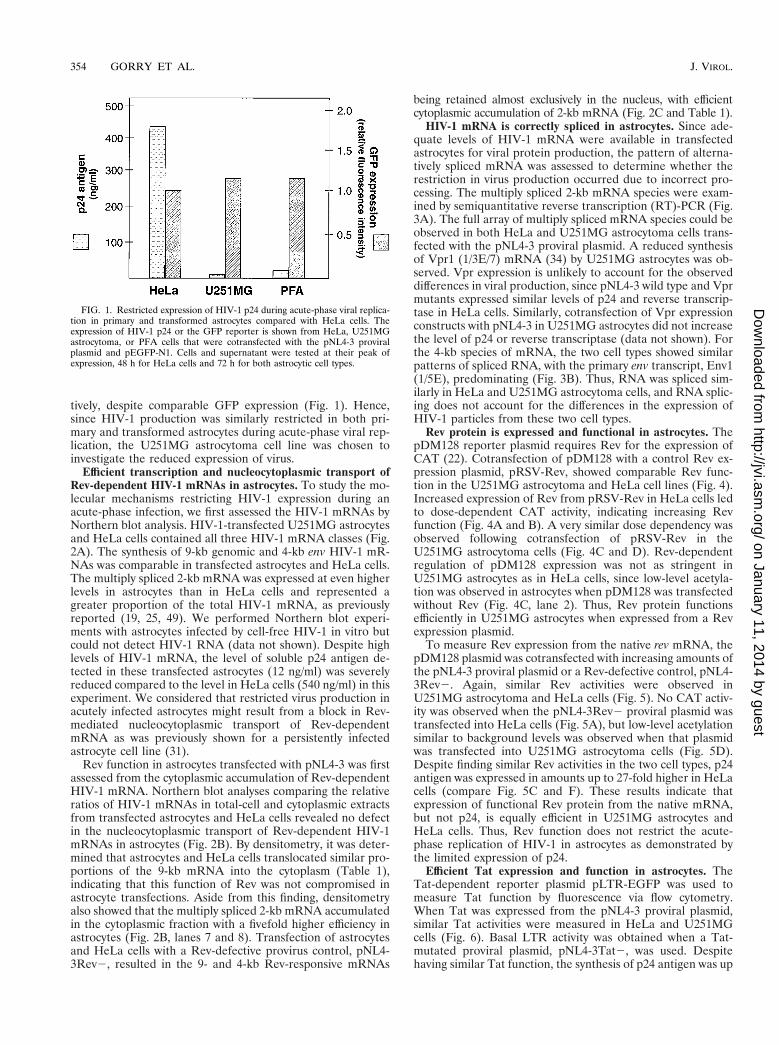
tively, despite comparable GFP expression (Fig. 1). Hence,since HIV-1 production was similarly restricted in both pri-mary and transformed astrocytes during acute-phase viral rep-lication, the U251MG astrocytoma cell line was chosen toinvestigate the reduced expression of virus.
Efficient transcription and nucleocytoplasmic transport ofRev-dependent HIV-1 mRNAs in astrocytes. To study the mo-lecular mechanisms restricting HIV-1 expression during anacute-phase infection, we first assessed the HIV-1 mRNAs byNorthern blot analysis. HIV-1-transfected U251MG astrocytesand HeLa cells contained all three HIV-1 mRNA classes (Fig.2A). The synthesis of 9-kb genomic and 4-kb env HIV-1 mR-NAs was comparable in transfected astrocytes and HeLa cells.The multiply spliced 2-kb mRNA was expressed at even higherlevels in astrocytes than in HeLa cells and represented agreater proportion of the total HIV-1 mRNA, as previouslyreported (19, 25, 49). We performed Northern blot experi-ments with astrocytes infected by cell-free HIV-1 in vitro butcould not detect HIV-1 RNA (data not shown). Despite highlevels of HIV-1 mRNA, the level of soluble p24 antigen de-tected in these transfected astrocytes (12 ng/ml) was severelyreduced compared to the level in HeLa cells (540 ng/ml) in thisexperiment. We considered that restricted virus production inacutely infected astrocytes might result from a block in Rev-mediated nucleocytoplasmic transport of Rev-dependentmRNA as was previously shown for a persistently infectedastrocyte cell line (31).
Rev function in astrocytes transfected with pNL4-3 was firstassessed from the cytoplasmic accumulation of Rev-dependentHIV-1 mRNA. Northern blot analyses comparing the relativeratios of HIV-1 mRNAs in total-cell and cytoplasmic extractsfrom transfected astrocytes and HeLa cells revealed no defectin the nucleocytoplasmic transport of Rev-dependent HIV-1mRNAs in astrocytes (Fig. 2B). By densitometry, it was deter-mined that astrocytes and HeLa cells translocated similar pro-portions of the 9-kb mRNA into the cytoplasm (Table 1),indicating that this function of Rev was not compromised inastrocyte transfections. Aside from this finding, densitometryalso showed that the multiply spliced 2-kb mRNA accumulatedin the cytoplasmic fraction with a fivefold higher efficiency inastrocytes (Fig. 2B, lanes 7 and 8). Transfection of astrocytesand HeLa cells with a Rev-defective provirus control, pNL4-3Rev2, resulted in the 9- and 4-kb Rev-responsive mRNAs
being retained almost exclusively in the nucleus, with efficientcytoplasmic accumulation of 2-kb mRNA (Fig. 2C and Table 1).
HIV-1 mRNA is correctly spliced in astrocytes. Since ade-quate levels of HIV-1 mRNA were available in transfectedastrocytes for viral protein production, the pattern of alterna-tively spliced mRNA was assessed to determine whether therestriction in virus production occurred due to incorrect pro-cessing. The multiply spliced 2-kb mRNA species were exam-ined by semiquantitative reverse transcription (RT)-PCR (Fig.3A). The full array of multiply spliced mRNA species could beobserved in both HeLa and U251MG astrocytoma cells trans-fected with the pNL4-3 proviral plasmid. A reduced synthesisof Vpr1 (1/3E/7) mRNA (34) by U251MG astrocytes was ob-served. Vpr expression is unlikely to account for the observeddifferences in viral production, since pNL4-3 wild type and Vprmutants expressed similar levels of p24 and reverse transcrip-tase in HeLa cells. Similarly, cotransfection of Vpr expressionconstructs with pNL4-3 in U251MG astrocytes did not increasethe level of p24 or reverse transcriptase (data not shown). Forthe 4-kb species of mRNA, the two cell types showed similarpatterns of spliced RNA, with the primary env transcript, Env1(1/5E), predominating (Fig. 3B). Thus, RNA was spliced sim-ilarly in HeLa and U251MG astrocytoma cells, and RNA splic-ing does not account for the differences in the expression ofHIV-1 particles from these two cell types.
Rev protein is expressed and functional in astrocytes. ThepDM128 reporter plasmid requires Rev for the expression ofCAT (22). Cotransfection of pDM128 with a control Rev ex-pression plasmid, pRSV-Rev, showed comparable Rev func-tion in the U251MG astrocytoma and HeLa cell lines (Fig. 4).Increased expression of Rev from pRSV-Rev in HeLa cells ledto dose-dependent CAT activity, indicating increasing Revfunction (Fig. 4A and B). A very similar dose dependency wasobserved following cotransfection of pRSV-Rev in theU251MG astrocytoma cells (Fig. 4C and D). Rev-dependentregulation of pDM128 expression was not as stringent inU251MG astrocytes as in HeLa cells, since low-level acetyla-tion was observed in astrocytes when pDM128 was transfectedwithout Rev (Fig. 4C, lane 2). Thus, Rev protein functionsefficiently in U251MG astrocytes when expressed from a Revexpression plasmid.
To measure Rev expression from the native rev mRNA, thepDM128 plasmid was cotransfected with increasing amounts ofthe pNL4-3 proviral plasmid or a Rev-defective control, pNL4-3Rev2. Again, similar Rev activities were observed inU251MG astrocytoma and HeLa cells (Fig. 5). No CAT activ-ity was observed when the pNL4-3Rev2 proviral plasmid wastransfected into HeLa cells (Fig. 5A), but low-level acetylationsimilar to background levels was observed when that plasmidwas transfected into U251MG astrocytoma cells (Fig. 5D).Despite finding similar Rev activities in the two cell types, p24antigen was expressed in amounts up to 27-fold higher in HeLacells (compare Fig. 5C and F). These results indicate thatexpression of functional Rev protein from the native mRNA,but not p24, is equally efficient in U251MG astrocytes andHeLa cells. Thus, Rev function does not restrict the acute-phase replication of HIV-1 in astrocytes as demonstrated bythe limited expression of p24.
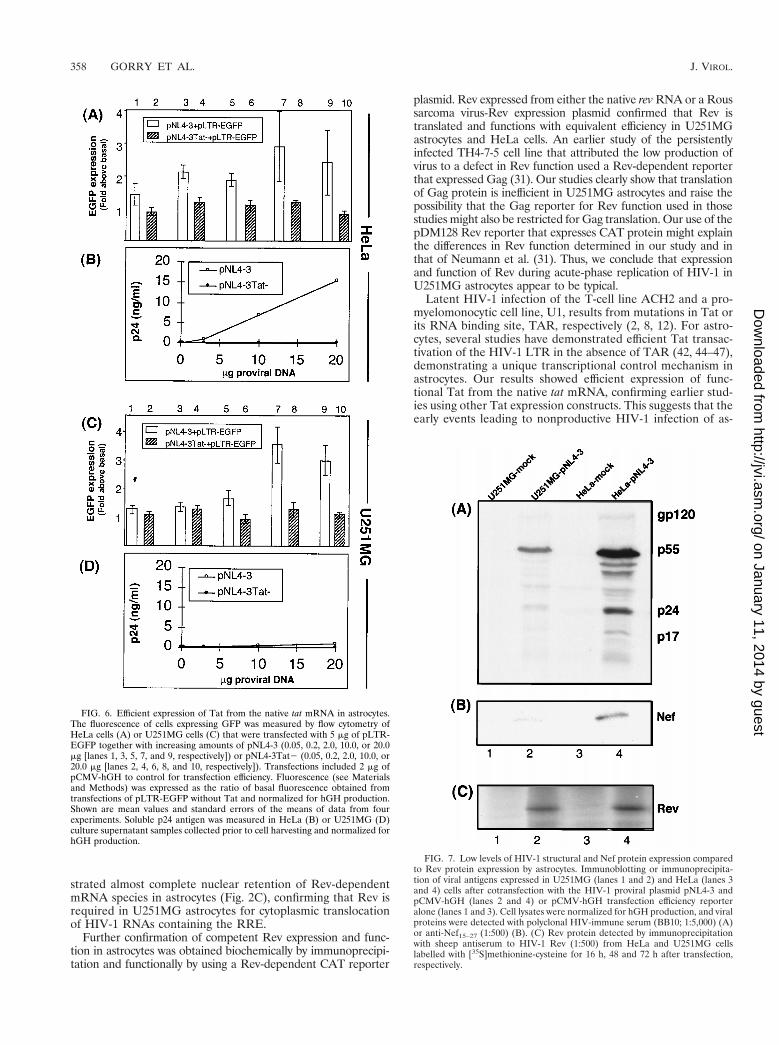
Efficient Tat expression and function in astrocytes. TheTat-dependent reporter plasmid pLTR-EGFP was used tomeasure Tat function by fluorescence via flow cytometry.When Tat was expressed from the pNL4-3 proviral plasmid,similar Tat activities were measured in HeLa and U251MGcells (Fig. 6). Basal LTR activity was obtained when a Tat-mutated proviral plasmid, pNL4-3Tat2, was used. Despitehaving similar Tat function, the synthesis of p24 antigen was up
FIG. 1. Restricted expression of HIV-1 p24 during acute-phase viral replica-tion in primary and transformed astrocytes compared with HeLa cells. Theexpression of HIV-1 p24 or the GFP reporter is shown from HeLa, U251MGastrocytoma, or PFA cells that were cotransfected with the pNL4-3 proviralplasmid and pEGFP-N1. Cells and supernatant were tested at their peak ofexpression, 48 h for HeLa cells and 72 h for both astrocytic cell types.
354 GORRY ET AL. J. VIROL.
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

to 23-fold higher in HeLa cell supernatants that producedequivalent levels of hGH (compare Fig. 6B and D). This showsthat restricted virus production in U251MG cells was not dueto inefficient Tat protein function. These results indicate that
Tat protein is efficiently synthesized from its native mRNA inU251MG astrocytes and is fully functional for transactivatingtranscription from the HIV-1 LTR.
The restricted acute-phase infection of astrocytes resultsfrom a specific downmodulation in the translation of HIV-1structural and Nef proteins. The preceding experimentsshowed that astrocytes express and process HIV-1 RNA in thesame way as HeLa cells. We therefore measured the produc-tion of HIV-1 proteins after transfection with pNL4-3, usingimmunoblotting or pulse-chase labelling and immunoprecipi-tation. Immunoblotting revealed very low-level synthesis ofHIV-1 proteins in pNL4-3-transfected astrocytes compared toHeLa cells (Fig. 7A). Similarly, there were fivefold lower levelsof Nef in astrocytes than in HeLa cells (Fig. 7B), despite theexistence of fivefold higher levels of nef mRNA (Fig. 2A and3A). The ratio of intracellular p24 (measured by enzyme-
FIG. 2. Efficient transcription and nucleocytoplasmic transport of HIV-1 mRNA in astrocytes. Northern blotting, using a 39 HIV-1 LTR region probe designed todetect all HIV-1 mRNAs, was performed with total cellular RNA from HeLa (lanes 1 and 2) and U251MG cells (lanes 3 and 4) transfected with pNL4-3 (lanes 2 and4) or mock transfected (lanes 1 and 3) (A) or with cytoplasmic (C) or total (T) RNA extracted from HeLa (lanes 1 to 4) and U251MG (lanes 5 to 8) cells transfectedwith pNL4-3 (lanes 3, 4, 7, and 8) or mock transfected (lanes 1, 2, 5, and 6) (B) or with cytoplasmic or total RNA extracted from HeLa (lanes 1 to 4) and U251MG(lanes 5 to 8) cells transfected with pNL4-3 Rev2 (lanes 3, 4, 7, and 8) or mock transfected (lanes 1, 2, 5, and 6) (C).
TABLE 1. Nuclear export of 9-kb Rev-dependent HIV-1 mRNA
Cell line Plasmid transfected Cytoplasmic mRNA(% of total)a
HeLa pNL4-3 26U251MG pNL4-3 24HeLa pNL4-3Rev2 0.8U251MG pNL4-3Rev2 0.5
a The percent accumulation of 9-kb HIV-1 mRNA detected in the cytoplasmicfraction relative to that in the total-cell RNA of astrocytes and HeLa cellstransfected with pNL4-3 and pNL4-3Rev2 (Fig. 2B and C) was determined fromdensitometric analysis of Northern blot pNL4-3Rev2 assays.
VOL. 73, 1999 RESTRICTED TRANSLATION OF HIV-1 mRNAs IN ASTROCYTES 355
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

linked immunosorbent assay of cell lysates) to hGH (measuredfrom transfected-cell supernatant) in astrocytes (0.073 ng/ng ofhGH) was 15-fold lower than that in HeLa cells (1.1 ng/ng ofhGH). In contrast, Rev was expressed with similar efficienciesby HeLa and U251MG astrocytes, since equivalent amounts ofRev were immunoprecipitated from the respective cell lysatesafter normalization for trichloroacetic acid (TCA)-precipitablecounts (Fig. 7C, compare lanes 2 and 4).
To determine whether the reduced levels of HIV-1 struc-tural proteins expressed by astrocytes resulted from abnormalprotein processing or from a specific block in translation ofthese mRNAs, pulse-chase labelling and immunoprecipitationwere performed with equal numbers of U251MG and HeLacells previously cotransfected with pEGFP-N1 and pNL4-3plasmids. Approximately 10% more U251MG cells were trans-fected than HeLa cells, but the two cell types expressed GFPwith equivalent mean intensities (Fig. 8). Despite a higheroverall level of expression of GFP by astrocytes, they synthe-sized 25-fold less pulse-labelled HIV-1 protein than did HeLacells after normalization for TCA-precipitable counts in therespective cell lysates (compare Fig. 9A and C). This indicatesthat astrocytes have a specific block in expression of HIV-1structural proteins. However, HIV-1 Gag and Env proteinprocessing was identical in the two cell types, because theprocessing of Env precursor gp160 to gp120-SU and gp41-TMand that of Gag Pr55 to p24-CA and p17-MA were comparableat all chase time points in lysates from both cell types (Fig. 9Aand C) and their progeny virions (Fig. 9B and D).
Thus, the restricted virus production from astrocytes acutelyreplicating HIV-1 results from inefficient translation of struc-tural protein from high levels of correctly processed HIV-1mRNA.
DISCUSSION
We examined the mechanisms restricting acute-phaseHIV-1 production in astrocytes by using a transfection systemto optimize expression during early viral replication. Despitenormal transcription, splicing, and transport of HIV-1 mRNA,immunoblot and radioimmunoprecipitation experimentsshowed that the translation of a subgroup of HIV-1 mRNAs is
FIG. 3. Astrocytes correctly splice HIV-1 mRNAs. RT-PCR analysis of mul-tiply spliced 2.0-kb (A) and singly spliced 4.0-kb (B) HIV-1 mRNAs from total-cell RNA extracted from HeLa (lane 1) or U251MG (lane 2) cells transfectedwith pNL4-3. cDNAs were amplified by PCR, labelled, and analyzed on a 6%acrylamide-urea gel. Nonspecific amplification was not observed in reactionslacking reverse transcriptase (data not shown).
FIG. 4. Rev-dependent expression of a CAT reporter in astrocytes. Acetyla-tion of chloramphenicol was measured in extracts of HeLa (A and B) orU251MG (C and D) cells that were transfected with 5 mg of the Rev-dependentCAT reporter pDM128 alone (lane 2) or together with increasing amounts ofRev from the pRSV-Rev expression plasmid (0.05, 0.2, 2.0, or 5.0 mg [lanes 3, 4,5, and 6, respectively]). Cells were cotransfected with 2 mg of pCMV-hGH tocontrol for transfection efficiency. Mock transfections received pCMV-hGHalone (lane 1). CAT assays were performed on cell lysates that were normalizedfor hGH production. Acetylation was quantitated by phosphorimaging, withbasal acetylation defined as CAT activity by pDM128 without Rev (lane 2), andmeans and standard errors of the means of the CAT activity for data from threeexperiments were determined after subtracting basal acetylation (B and D).
356 GORRY ET AL. J. VIROL.
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

highly inefficient during acute-phase viral replication in theU251MG astrocyte cell line. The cytoplasmic gag, env, and nefRNAs were poor substrates for protein expression, producing25-fold less protein in astrocytes than in HeLa cells. Pulse-chase labelling showed that reduced accumulation of HIV-1structural proteins during acute-phase viral replication was dueto their inefficient synthesis in the astrocyte cell line and wasnot due to any abnormal processing or targeted degradation ofthese proteins.
All HIV-1 mRNA classes were expressed at high levels inU251MG cells (Fig. 2A) and were correctly spliced (Fig. 3) butyielded 50-fold lower p24 levels than HeLa cells expressing
comparable levels of HIV-1 mRNA. Previous studies of apersistently HIV-1-infected astrocyte cell line, TH4-7-5, dem-onstrated that in that model, restricted infection resulted froman astrocyte-specific block in HIV-1 Rev function, causinginefficient nucleocytoplasmic transport of unspliced and singlyspliced HIV-1 mRNA species (31). In our acute-phase repli-cation model, this mechanism did not account for the reducedexpression of virus because the Rev-dependent mRNA classeswere readily detected in cytoplasmic RNA extracts of astro-cytes (Fig. 2B), showing that Rev-dependent mRNAs wereefficiently translocated to the cytoplasm. Control transfectionswith a Rev-deficient proviral plasmid, pNL4-3Rev2, demon-
FIG. 5. Efficient expression of Rev from the native rev mRNA in astrocytes. Acetylation of chloramphenicol was measured in extracts of HeLa (A and B) orU251MG (D and E) cells that were transfected with 5 mg of pDM128 without HIV-1 provirus (lane 2) or together with increasing amounts of pNL4-3 (lanes 3, 5, 7,9, and 11) or pNL4-3Rev2 (lanes 4, 6, 8, 10, and 12). Cells were transfected with 2 mg of pCMV-hGH to control for transfection efficiency. Mock transfections receivedpCMV-hGH alone (lane 1). CAT assays were performed on cell lysates that were normalized for hGH production. Acetylation was quantitated by phosphorimaging,with basal acetylation defined as CAT activity by pDM128 without Rev (lane 2), and means and standard errors of the means of the CAT activity for data from threeexperiments were determined after subtracting basal acetylation (B and E). Soluble p24 antigen was measured in culture supernatant samples collected prior to cellharvesting and normalized for hGH production (C and F).
VOL. 73, 1999 RESTRICTED TRANSLATION OF HIV-1 mRNAs IN ASTROCYTES 357
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from
strated almost complete nuclear retention of Rev-dependentmRNA species in astrocytes (Fig. 2C), confirming that Rev isrequired in U251MG astrocytes for cytoplasmic translocationof HIV-1 RNAs containing the RRE.
Further confirmation of competent Rev expression and func-tion in astrocytes was obtained biochemically by immunoprecipi-tation and functionally by using a Rev-dependent CAT reporter
plasmid. Rev expressed from either the native rev RNA or a Roussarcoma virus-Rev expression plasmid confirmed that Rev istranslated and functions with equivalent efficiency in U251MGastrocytes and HeLa cells. An earlier study of the persistentlyinfected TH4-7-5 cell line that attributed the low production ofvirus to a defect in Rev function used a Rev-dependent reporterthat expressed Gag (31). Our studies clearly show that translationof Gag protein is inefficient in U251MG astrocytes and raise thepossibility that the Gag reporter for Rev function used in thosestudies might also be restricted for Gag translation. Our use of thepDM128 Rev reporter that expresses CAT protein might explainthe differences in Rev function determined in our study and inthat of Neumann et al. (31). Thus, we conclude that expressionand function of Rev during acute-phase replication of HIV-1 inU251MG astrocytes appear to be typical.
Latent HIV-1 infection of the T-cell line ACH2 and a pro-myelomonocytic cell line, U1, results from mutations in Tat orits RNA binding site, TAR, respectively (2, 8, 12). For astro-cytes, several studies have demonstrated efficient Tat transac-tivation of the HIV-1 LTR in the absence of TAR (42, 44–47),demonstrating a unique transcriptional control mechanism inastrocytes. Our results showed efficient expression of func-tional Tat from the native tat mRNA, confirming earlier stud-ies using other Tat expression constructs. This suggests that theearly events leading to nonproductive HIV-1 infection of as-
FIG. 6. Efficient expression of Tat from the native tat mRNA in astrocytes.The fluorescence of cells expressing GFP was measured by flow cytometry ofHeLa cells (A) or U251MG cells (C) that were transfected with 5 mg of pLTR-EGFP together with increasing amounts of pNL4-3 (0.05, 0.2, 2.0, 10.0, or 20.0mg [lanes 1, 3, 5, 7, and 9, respectively]) or pNL4-3Tat2 (0.05, 0.2, 2.0, 10.0, or20.0 mg [lanes 2, 4, 6, 8, and 10, respectively]). Transfections included 2 mg ofpCMV-hGH to control for transfection efficiency. Fluorescence (see Materialsand Methods) was expressed as the ratio of basal fluorescence obtained fromtransfections of pLTR-EGFP without Tat and normalized for hGH production.Shown are mean values and standard errors of the means of data from fourexperiments. Soluble p24 antigen was measured in HeLa (B) or U251MG (D)culture supernatant samples collected prior to cell harvesting and normalized forhGH production.
FIG. 7. Low levels of HIV-1 structural and Nef protein expression comparedto Rev protein expression by astrocytes. Immunoblotting or immunoprecipita-tion of viral antigens expressed in U251MG (lanes 1 and 2) and HeLa (lanes 3and 4) cells after cotransfection with the HIV-1 proviral plasmid pNL4-3 andpCMV-hGH (lanes 2 and 4) or pCMV-hGH transfection efficiency reporteralone (lanes 1 and 3). Cell lysates were normalized for hGH production, and viralproteins were detected with polyclonal HIV-immune serum (BB10; 1:5,000) (A)or anti-Nef15–27 (1:500) (B). (C) Rev protein detected by immunoprecipitationwith sheep antiserum to HIV-1 Rev (1:500) from HeLa and U251MG cellslabelled with [35S]methionine-cysteine for 16 h, 48 and 72 h after transfection,respectively.
358 GORRY ET AL. J. VIROL.
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

trocytes involve suppressed translation of HIV-1 structuralproteins, as opposed to the transcriptional repression that un-derpins the latent infection of T cells and monocytes/macro-phages. The suppressed translation of HIV-1 structural pro-teins in the face of efficient HIV-1 mRNA transcription mayexplain the discrepancies in measurements of the numbers ofHIV-1-infected astrocytes in vivo. Measurements of HIV-1structural proteins may significantly underrepresent the distri-bution and transcriptional activity of HIV-1-infected astrocytesin vivo compared to RNA or DNA detection (3, 7, 18, 41, 43).Furthermore, efficient expression of the neurotoxic Tat proteinby astrocytes in vivo may contribute to neuropathogenesis inAIDS patients (30, 37).
Our studies showed that Nef protein is expressed at fivefoldlower levels in U251MG cells than in HeLa cells, despite thefivefold higher level of nef mRNA synthesis in the astrocytes.This reduced Nef synthesis contrasts with the high levels mea-sured in the TH4-7-5 cell line that was selected for Nef expres-sion (6) and may indicate that changes in the native nef mRNAalleviate the inefficient translation. The high-level expressionof Nef in postmortem astrocytes from pediatric and adultAIDS patients (35, 38, 48) might demonstrate the existence ofin vivo mechanisms that overcome the inefficient translation ofNef, possibly modulated by local cytokines. However, the sup-pressed translation of nef mRNA observed in our study is inagreement with other reports showing low-level Nef synthesisthat could be relieved by coculture with monocytes/macro-phages (15) or tumor necrosis factor alpha stimulation (49).
The mechanism underlying the inefficient translation of the
FIG. 8. Efficient translation of a non-HIV-1 fluorescence reporter by astro-cytes. Flow cytometry profiles showing the transfection-translation efficiency ofGFP (light shading) by U251MG astrocytes (A and C) and HeLa cells (B and D)transfected with 5 mg of pEGFP-N1 alone (A and B) or together with 20 mg ofthe HIV-1 proviral plasmid pNL4-3 (C and D). FACS analysis was performed atthe peak of expression posttransfection (72 h for U251MG and 48 h for HeLacells). Mock transfections (dark shading) contained either no DNA (A and B) orpNL4-3 alone (C and D). The region R1 shows the percentage and the meanchannel fluorescence (MCF) of cells expressing GFP above the mock-transfec-tion values.
FIG. 9. Inefficient translation but correct processing of HIV-1 structural proteins by astrocytes. HeLa (A) and U251MG (C) cells were pulse-labelled with[35S]methionine for 1 h at 48 h (HeLa) or 72 h (U251MG) after cotransfection with pEGFP-N1 alone (mock) or together with 20 mg of HIV-1 proviral plasmid pNL4-3(pulse). Labelled cells were then incubated at 37°C for 1, 2, 3, 4, and 6 h in complete medium, and HIV-1 proteins were detected by immunoprecipitation from celllysate volumes (standardized by total TCA-precipitable 35S-labelled proteins) with pooled HIV-1 immune serum (BB10). Virions pelleted from the supernatant of HeLa(B) or U251MG (D) transfections were lysed and subjected to immunoprecipitation for analysis of HIV-1 structural proteins.
VOL. 73, 1999 RESTRICTED TRANSLATION OF HIV-1 mRNAs IN ASTROCYTES 359
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

gag, env, and nef mRNAs is unclear. The strength of the Kozaksignal surrounding the initiating AUG codon does not corre-late with the translation efficiency of these mRNAs, becauseHIV-1NL tat, gag, env, and nef mRNAs all have strong Kozaksignals whereas rev mRNA has a weak Kozak signal (26, 27).The translational control mechanism accounting for our resultsmust accommodate (i) the selective repression of HIV-1 struc-tural protein synthesis over that of the other reporter proteinsused here (hGH and GFP), (ii) suppression mediated by 59HIV-1 noncoding RNA proximal to TAR, and (iii) a relief oftranslational suppression for tat and rev mRNAs that is medi-ated by a short stretch of unique 59 noncoding RNA.
Several reports have shown that the translation of HIV-1proteins may be downmodulated by the alpha-interferon-in-ducible double-stranded RNA-activated protein kinase R(PKR) (4, 33). It is possible that astrocytes exhibit high-levelconstitutive expression of activated PKR that might inhibitengagement of initiator tRNAMet at the 7-methyl-Gppp capstructure at the 59 end of the RNA (reviewed by Clemens [9]).However, PKR-mediated translational control has not beenreported to discriminate between mRNAs translated by typicalribosome scanning. The existence of polypyrimidine tractsproximal to the initiating AUG of tat and rev mRNAs (34)raises the possibility that the translation of these mRNAs es-capes a cap-dependent translational suppression imposed onthe other HIV-1 RNAs by direct interactions of the 40S ribo-somal subunit with polypyrimidine tract binding proteins, in amanner resembling the activity of the internal ribosome entrysites of some viruses, e.g., picornavirus (11).
The efficient expression of Tat protein in astrocytes raisesthe possibility that this viral protein contributes to the trans-lation control observed in these cells. Recently, the secondcoding exon of HIV-1 Tat was shown to directly interact witha protein of the human translation machinery, translation elon-gation factor 1 delta (52). This interaction caused downmodu-lated translation of CAT protein in vitro and of CD4 protein intransfected HeLa cells. Interestingly, unlike CD4 cells, thetranslation of HIV-1 structural proteins was not diminished inHeLA cells cotransfected with CD4 and HIV-1 proviral plas-mids (52). This discriminatory modulation of translation inHeLa cells mediated by the second coding exon of Tat raisesthe possibility that efficient expression of HIV-1 Tat in astro-cytes contributes to the observed downmodulated translationof Gag, Env, and Nef. Another role for Tat might result fromthe reported direct interaction of the first coding exon of Tat(Tat1–72) with PKR causing an inhibition of autophosphoryla-tion and activation of PKR (29). In addition, the second codingexon of Tat (Tat73–86) is itself a substrate for phosphorylationby PKR (29). An accumulation of Tat in astrocytes might resultin interactions with PKR that preferentially restrict translationof HIV-1 structural proteins. Further experiments to examinethe translational control mechanisms of astrocytes that causethe diminished expression of HIV-1 are under way. What isclear from our present results is that the translational controlmechanism of astrocytes discriminates between two subgroupsof HIV-1 mRNAs.
With regard to identifying the relevant differences betweenthe HIV-1 mRNAs, it is of interest that the pDM128 reporter,which has the CAT gene inserted at the translation initiationsite for Env, efficiently expresses CAT protein in astrocytes,whereas Env is inefficiently expressed from the native envmRNA. The significant differences in the 59 untranslated re-gions (UTRs) of the pDM128 and the native env mRNAs (22)provide insight into the RNA elements that might increase thetranslation efficiency of the CAT mRNA as well as rev and tatmRNA. The 59 end of the pDM128 mRNA has up to 428 bases
of the 59 UTR from simian virus 40 large T antigen joined tothe bulge of the HIV-1 TAR region. The HIV-1 sequenceextends to the end of the U5 region but excludes 102 basesupstream of the major 59 splice site and includes noncodingmRNA 39 of the seventh codon of Tat. In pDM128, the startsite for Rev has been mutated and the CAT gene has beeninserted at the Env initiation codon. Thus, pDM128 includesup to 125 extra bases of the 59 UTRs from rev and tat mRNAsthat are not in the predominant env mRNA. Thus, uniqueRNA elements from the rev and tat mRNAs, such as thepolypyrimidine tracts of the splice acceptor sites for rev andenv, are included into the 59 UTR of pDM128. These RNAelements are spliced from the native env and nef mRNAs andmight confer a more-efficient translation to some mRNAs inastrocytes. However, translation modulation by sequences inthe Env coding frame cannot be excluded.
In the adult brain, astrocytes are unrenewable and crucialfor neurological function. The elimination of astrocytes la-tently infected by HIV-1 may be neurologically detrimental.However, this dormant infection provides a sanctuary forHIV-1 that may thwart viral eradication and serve as a sourcefor renewed systemic infection. Inducing permanent viral dor-mancy may be a strategy to control HIV-1. Our results showthat the dormant infection of astrocytes originates from a blockin the translation of gag, env, and nef mRNAs. Translation ofthe rev and tat mRNAs was unaffected. Understanding themechanisms modulating the translation of HIV-1 structuralproteins may identify strategies to sustain the suppression ofHIV-1 in viral sanctuaries such as brain astrocytes.
ACKNOWLEDGMENTS
This work was supported by the National Health and Medical Re-search Council (D.F.J.P., J.L.H., and J.L.A.; Reg Key 970558), theNational Centre for HIV Virology Research (D.A.M., M.J.C., A.C.,and D.A.), RMIT University (P.R.G.), and the Research Fund of theMacfarlane Burnet Centre for Medical Research (P.R.G., J.L.H.,M.J.C., J.L.A., D.A.M., and D.F.J.P.).
We thank J. Mills, A. Jaworowski, and D. Gabuzda for review of themanuscript, G. Paukovics and A. Meikle (Flow Cytometry Unit, Mac-farlane Burnet Centre for Medical Research) for FACS analysis, andB. Rumble (Department of Medical Laboratory Science, RMIT Uni-versity) for advice and support in this project.
REFERENCES1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and
M. A. Martin. 1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with aninfectious molecular clone. J. Virol. 59:284–291.
2. Adams, M., L. Sharmeen, J. Kimpton, J. M. Romeo, J. V. Garcia, B. M.Peterlin, M. Groudine, and M. Emerman. 1994. Cellular latency in humanimmunodeficiency virus-infected individuals with high CD4 levels can bedetected by the presence of promoter-proximal transcripts. Proc. Natl. Acad.Sci. USA 91:3862–3866.
3. Bagasra, O., E. Lavi, L. Bobroski, K. Khalili, J. P. Pestaner, R. Tawadros,and R. J. Pomerantz. 1996. Cellular reservoirs of HIV-1 in the centralnervous system of infected individuals: identification by the combination ofin situ polymerase chain reaction and immunohistochemistry. AIDS 10:573–585.
4. Benkirane, M., C. Neuveut, R. F. Chun, S. M. Smith, C. E. Samuel, A.Gatignol, and K.-T. Jeang. 1997. Oncogenic potential of TAR RNA bindingprotein TRBP and its regulatory interaction with RNA-dependent proteinkinase PKR. EMBO J. 16:611–624.
5. Bigner, D. D., S. H. Bigner, J. Ponten, B. Westermark, M. S. Mahaley, E.Ruoslahti, H. Herschman, L. F. Eng, and C. J. Wilkstrand. 1981. Hetero-geneity of genotypic and phenotypic characteristics of fifteen permanent celllines derived from human gliomas. J. Neuropathol. Exp. Neurol. 40:201–229.
6. Brack-Werner, R., A. Kleinschmidt, A. Ludvigsen, W. Mellert, M. Neumann,R. Herrmann, M. C. L. Khim, A. Burny, N. Muller-Lantzsch, D. Stavrou,and V. Erfle. 1992. Infection of human brain cells by HIV-1: restricted virusproduction in chronically infected human glial cell lines. AIDS 6:273–285.
7. Brew, B. J., M. Rosenblum, K. Cronin, and R. W. Price. 1995. AIDS de-mentia complex and HIV-1 brain infection: clinical-virological correlations.Ann. Neurol. 38:563–570.
360 GORRY ET AL. J. VIROL.
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from

8. Cannon, P., S.-H. Kim, C. Ulich, and S. Kim. 1994. Analysis of Tat functionin human immunodeficiency virus type 1-infected low-level-expression celllines U1 and ACH-2. J. Virol. 68:1993–1997.
9. Clemens, M. J. 1996. Protein kinases that phosphorylate eIF2 and eIF2B,and their role in eukaryotic cell translational control, p. 139–172. In J. W. B.Hershey, M. B. Mathews, and N. Sonenberg (ed.), Translational control.Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
10. Clouse, K. A., D. Powell, I. Washington, G. Poli, K. Strebel, W. Farrar, P.Barstad, J. Kovacs, A. S. Fauci, and T. M. Folks. 1989. Monokine regulationof human immunodeficiency virus-1 expression in a chronically infected Tcell clone. J. Immunol. 142:431–438.
11. Ehrenfeld, E. 1996. Initiation of translation by picornavirus RNAs, p. 549–573. In J. W. B. Hershey, M. B. Mathews, and N. Sonenberg (ed.), Trans-lational control. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y.
12. Emiliani, S., C. Van Lint, W. Fischle, P. Paris, Jr., M. Ott, J. Brady, and E.Verdin. 1996. A point mutation in the HIV-1 Tat responsive element isassociated with postintegration latency. Proc. Natl. Acad. Sci. USA 93:6377–6381.
13. Ensoli, F., H. Wang, V. Fiorelli, S. L. Zeichner, M. R. De Cristofaro, G. Luzi,and C. J. Thiele. 1997. HIV-1 infection and the developing nervous system:lineage-specific regulation of viral gene expression and replication in distinctneuronal precursors. J. Neurovirol. 3:290–298.
14. Erfle, V., P. Stoeckbauer, A. Kleinschmidt, B. Kohleisen, W. Mellert, D.Stavrou, and R. Brack-Werner. 1991. Target cells for HIV in the centralnervous system: macrophages or glial cells? Res. Virol. 142:139–144.
15. Fiala, M., R. H. Rhodes, P. Shapshak, I. Nagano, O. Martinez-Maza, A.Diagne, G. Baldwin, and M. Graves. 1996. Regulation of HIV-1 infection inastrocytes: expression of Nef, TNF-a and IL-6 is enhanced in coculture ofastrocytes with macrophages. J. Neurovirol. 2:158–166.
16. Folks, T., D. M. Powell, M. M. Lightfoote, S. Benn, M. A. Martin, and A. S.Fauci. 1986. Induction of HTLV-III/LAV from a nonvirus-producing T-cellline: implications for latency. Science 231:600–602.
17. Folks, T. M., J. Justement, A. Kinter, C. A. Dinarello, and A. S. Fauci. 1987.Cytokine-induced expression of HIV-1 in a chronically infected promono-cyte cell line. Science 238:800–802.
18. Glass, J. D., H. Fedor, S. L. Wesselingh, and J. C. McArthur. 1995. Immu-nocytochemical quantitation of human immunodeficiency virus in the brain:correlations with dementia. Ann. Neurol. 38:755–762.
19. Gorry, P., D. Purcell, J. Howard, and D. McPhee. 1998. Restricted HIV-1infection of human astrocytes: potential role of nef in the regulation of virusreplication. J. Neurovirol. 4:377–386.
20. Greenway, A. L., D. A. McPhee, E. Grgacic, D. Hewish, A. Lucantoni, I.Macreadie, and A. Azad. 1994. Nef 27, but not Nef 25 isoform of humanimmunodeficiency virus-type 1 pNL4.3 down-regulates surface CD4 andIL-2R expression in peripheral blood mononuclear cells and transformed Tcells. Virology 198:245–256.
21. Hatch, W. C., E. Pousada, L. Losev, W. K. Rashbaum, and W. D. Lyman.1994. Neural cell targets of human immunodeficiency virus type 1 in humanfetal organotypic cultures. AIDS Res. Hum. Retroviruses 10:1597–1607.
22. Hope, T. J., X. Huang, D. McDonald, and T. G. Parslow. 1990. Steroid-receptor fusion of the human immunodeficiency virus type 1 Rev transacti-vator: mapping cryptic functions of the arginine-rich motif. Proc. Natl. Acad.Sci. USA 87:7787–7791.
23. Huang, L., A. Joshi, R. Willey, J. Ornstein, and K. T. Jeang. 1994. Humanimmunodeficiency viruses regulated by alternative trans-activators: geneticevidence for a novel non-transcriptional function of Tat in virion infectivity.EMBO J. 13:2886–2896.
24. Johnson, R. T., J. D. Glass, J. C. McArthur, and B. W. Chesebro. 1996.Quantitation of human immunodeficiency virus in brains of demented andnondemented patients with acquired immunodeficiency syndrome. Ann.Neurol. 39:392–395.
25. Kleinschmidt, A., M. Neumann, C. Moller, V. Erfle, and R. Brack-Werner.1994. Restricted expression of HIV-1 in human astrocytes: molecular basisfor viral persistence in the CNS. Res. Virol. 145:147–153.
26. Kozak, M. 1986. Point mutations define a sequence flanking the AUGinitiator codon that modulates translation by eukaryotic ribosomes. Cell44:283–293.
27. Kozak, M. 1989. The scanning model for translation: an update. J. Cell Biol.108:229–241.
28. Kurth, J., J. M. Buzy, L. Lindstrom, and J. E. Clements. 1996. In vivotranscriptional regulation of the human immunodeficiency virus in the cen-tral nervous system in transgenic mice. J. Virol. 70:7686–7694.
29. McMillan, N. A., R. F. Chun, D. P. Siderovski, J. Galabru, W. M. Toone,C. E. Samuel, T. W. Mak, A. G. Hovanessian, K. T. Jeang, and B. R.Williams. 1995. HIV-1 Tat directly interacts with the interferon-induced,double-stranded RNA-dependent kinase, PKR. Virology 213:413–424.
30. Nath, A., K. Psooy, C. Martin, B. Knudsen, D. S. K. Magnuson, N. Haugheyand J. D. Geiger. 1996. Identification of a human immunodeficiency virus type1 Tat epitope that is neuroexcitatory and neurotoxic. J. Virol. 70:1475–1480.
31. Neumann, M., B. K. Felber, A. Kleinschmidt, B. Froese, V. Erfle, G. N.Pavlakis, and R. Brack-Werner. 1995. Restriction of human immunodefi-
ciency virus type 1 production in a human astrocytoma cell line is associatedwith a cellular block in Rev function. J. Virol. 69:2159–2167.
32. Niikura, M., G. Dornadula, H. Zhang, M. Mukhtar, D. Lingxun, K. Khalili,O. Bagasra, and R. Pomerantz. 1996. Mechanisms of transcriptional trans-activation and restriction of human immunodeficiency virus type 1 replica-tion in an astrocytic glial cell. Oncogene 13:313–322.
33. Park, H., M. V. Davies, J. O. Langland, H.-W. Chang, Y.-S. Nam, J. Tarta-glia, E. Paoletti, B. L. Jacobs, R. J. Kaufman, and S. Venkatesan. 1994. TARRNA-binding protein is an inhibitor of the interferon induced protein ki-nase, PKR. Proc. Natl. Acad. Sci. USA 91:4713–4717.
34. Purcell, D. F. J., and M. A. Martin. 1993. Alternative splicing of humanimmunodeficiency virus type 1 mRNA modulates viral protein expression,replication, and infectivity. J. Virol. 67:6365–6378.
35. Ranki, A., M. Nyberg, V. Ovod, M. Haltia, I. Elovaara, R. Raininko, H.Haapaasalo, and K. Krohn. 1995. Abundant expression of HIV Nef and Revproteins in brain astrocytes in vivo is associated with dementia. AIDS9:1001–1008.
36. Rytik, P. G., V. F. Eremin, Z. B. Kvacheva, N. N. Poleschuk, S. A. Popov,H. C. Schroder, M. Bachmann, B. E. Weiler, and W. E. G. Muller. 1991.Susceptibility of primary human glial fibrillary acidic protein-positive braincells to human immunodeficiency virus infection in vitro: anti-HIV activity ofmemantine. AIDS Res. Hum. Retroviruses 7:89–95.
37. Sabatier, J.-M., E. Vives, K. Mabrouk, A. Benjouad, H. Rochat, A. Duval, B.Hue, and E. Bahraoui. 1991. Evidence for neurotoxic activity of tat fromhuman immunodeficiency virus type 1. J. Virol. 65:961–967.
38. Saito, Y., L. R. Sharer, L. G. Epstein, J. Michaels, M. Mintz, M. Louder, K.Golding, T. A. Cvetkocitch, and B. M. Blumberg. 1994. Overexpression of nefas a marker for restricted HIV-1 infection of astrocytes in postmortempediatric central nervous tissue. Neurology 44:474–481.
39. Scherer, W. F., J. T. Syverton, and G. O. Gey. 1953. Viral multiplication ina stable strain of human malignant epithelial cells (strain HeLa) derivedfrom an epidermoid carcinoma of the cervix. J. Exp. Med. 97:695–710.
40. Shahabuddin, M., B. Volsky, H. Kim, K. Sakai, and D. J. Volsky. 1992.Regulated expression of human immunodeficiency virus type 1 in humanglial cells: induction of dormant virus. Pathobiology 60:195–205.
41. Sharer, L. R., Y. Saito, A. De Cunha, D. Padiwath, H. A. Gelbard, L. G.Epstein, and B. M. Blumberg. 1996. In situ amplification and detection ofHIV-1 DNA in fixed pediatric AIDS brain tissue. Hum. Pathol. 27:614–617.
42. Swingler, S., A. Easton, and A. Morris. 1992. Cytokine augmentation ofHIV-1 LTR-driven gene expression in neural cells. AIDS Res. Hum. Ret-roviruses 8:487–493.
43. Takahashi, K., S. L. Wesselingh, D. E. Griffin, J. C. McArthur, R. T. John-son, and J. D. Glass. 1996. Localization of HIV-1 in human brain usingpolymerase chain reaction/in situ hybridization and immunocytochemistry.Ann. Neurol. 39:705–711.
44. Taylor, J. P., M. Kundu, and K. Khalili. 1993. TAR-independent activationof HIV-1 requires the activation domain but not the RNA-binding domain ofTat. Virology 195:780–785.
45. Taylor, J. P., R. Pomerantz, O. Bagasra, M. Chowdhury, J. Rappaport, K.Khalili, and S. Amini. 1992. TAR-independent transactivation by Tat in cellsderived from the CNS: a novel mechanism of HIV-1 gene regulation. EMBOJ. 11:3395–3403.
46. Taylor, J. P., R. J. Pomerantz, J. W. Oakes, K. Khalili, and S. Amini. 1995.A CNS-enriched factor that binds to NF-kB and is required for interactionwith HIV-1 Tat. Oncogene 10:395–400.
47. Taylor, J. P., R. J. Pomerantz, G. V. Raj, F. Kashanchi, J. N. Brady, S.Amini, and K. Khalili. 1994. Central nervous system-derived cells express akB-binding activity that enhances human immunodeficiency virus type 1transcription in vitro and facilitates TAR-independent transactivation byTat. J. Virol. 68:3971–3981.
48. Tornatore, C., R. Chandra, J. R. Berger, and E. O. Major. 1994. HIV-1infection of subcortical astrocytes in the pediatric central nervous system.Neurology 44:481–487.
49. Tornatore, C., K. Meyers, W. Atwood, K. Conant, and E. Major. 1994.Temporal patterns of human immunodeficiency virus type 1 transcripts inhuman fetal astrocytes. J. Virol. 68:93–102.
50. Tornatore, C., A. Nath, K. Amemiya, and E. O. Major. 1991. Persistenthuman immunodeficiency virus type 1 infection in human fetal glial cellsreactivated by T-cell factor(s) or by the cytokines tumor necrosis factor alphaand interleukin-1 beta. J. Virol. 65:6094–6100.
51. Willey, R. L., T. Klimkait, D. M. Frucht, J. S. Bonifacino, and M. A. Martin.1991. Mutations within the human immunodeficiency virus type 1 gp160envelope glycoprotein alter its intracellular transport and processing. Virol-ogy 184:319–329.
52. Xiao, H., C. Neuveut, M. Benkirane, and K. T. Jeang. 1998. Interaction ofthe second coding exon of Tat with human EF-1d delineates a mechanism forHIV-1-mediated shut-off of host mRNA translation. Biochem. Biophys. Res.Comm. 244:384–389.
53. Yang, L., G. F. Morris, J. M. Lockyer, M. Lu, Z. Wang, and C. B. Morris.1997. Distinct transcriptional pathways of TAR-dependent and TAR-inde-pendent human immunodeficiency virus type-1 transactivation by Tat. Vi-rology 235:48–64.
VOL. 73, 1999 RESTRICTED TRANSLATION OF HIV-1 mRNAs IN ASTROCYTES 361
on January 11, 2014 by guesthttp://jvi.asm
.org/D
ownloaded from




















