
Dietary variation in spectacled flying foxes ( Pteropus conspicillatus ) of the Australian Wet...
12
Introduction Large pteropodids (Megachiroptera: Pteropodidae), also known as Old World fruit bats or flying foxes (Pteropus spp.), rely on a variety of plant resources and have varied for- aging and feeding strategies (Courts 1998; Augee and Ford 1999; Dumont and O’Neal 2004). Australian flying foxes exhibit these varied feeding and foraging strategies with dietary flexibility or by moving in response to changes in resource availability (Nelson 1965; Parry-Jones and Augee 1991; Eby 1998; Markus and Hall 2004; McDonald-Madden et al. 2005). Pteropus conspicillatus has long been consid- ered the only flying fox on the Australian continent to be a rainforest-fruit specialist (Richards 1990b; Birt et al. 1997; Garnett et al. 1999). This conclusion is supported by its asso- ciation with rainforest habitat (Richards 1990a), observa- tions of the species foraging on the fruits of rainforest trees and the morphology of their tongue and teeth, which sug- gests a frugivorous diet (Birt et al. 1997). However, specific details of the diet and habitat use of P. conspicillatus are scarce, and the presence of many camps outside of rainforest (Richards 1990b; Garnett et al. 1999) along with theoretical predictions that megachiropteran bats cannot meet their nutritional requirements on a pure-fruit diet (Law 1992; Courts 1998) suggest that P. conspicillatus may not be restricted entirely to rainforest habitats and fruit. Phytophagous bats have been shown to be ecologically important for a variety of ecosystem functions, such as angiosperm pollination, seed dispersal (Williams and Adam 1994; Shilton et al. 1999; Banack and Grant 2002; Singaravelan and Marimuthu 2004; Nyhagen et al. 2005) and possibly nutrient recycling (Feeley 2005). The black flying fox Pteropus alecto plays a significant role in main- taining rainforest fragments in the Northern Territory (Woinarski et al. 1992; Palmer et al. 2000), and it is possible that P. conspicillatus plays a similar role in the Wet Tropics region of Australia (Westcott et al. 2001). Determining the role that P. conspicillatus plays in maintaining ecosystem function requires knowledge of the dietary habits of the species and how such habits may vary among camps. Previous dietary analysis of P. conspicillatus has concen- trated on the identification of seeds in faeces, often as part of frugivory and seed-dispersal studies. However, seed identifi- cation from bat faeces suffers some significant shortcomings (Thomas 1988; Shilton 1999). First, it commonly excludes large seeded fruits, because these animals are able to ingest only small seeds (Richards 1990b). Second, a focus on fruit means that other plant parts go undetected, particularly flowers that require pollen analysis. A fast gut-passage time in pteropodids also means that faeces collected at the day roost is likely to consist primarily of the last feed of the night, even though a gut retention time of >12 h has been shown (Shilton et al. 1999). The lack of studies that included other faecal components useful for diet determination, such as fruit pulp and pollen, means that these dietary indicators and the habi- Australian Journal of Zoology, 2006, 54, 417–428 10.1071/ZO06092 0004-959X/06/060417 © CSIRO 2006 Jennifer G. Parsons A,C , Andi Cairns A , Christopher N. Johnson A , Simon K. A. Robson A , Louise A. Shilton B and David A. Westcott B A School of Marine andTropical Biology, James Cook University, Qld 4811, Australia. B Tropical Forest Research Centre, CSIRO Sustainable Ecosystems, PO Box 780, Atherton, Qld 4883, Australia. C Corresponding author. Email: [email protected] Abstract. The diet of Pteropus conspicillatus, a large flying fox, was examined by collecting faeces in traps beneath daytime roost trees in four geographically distinct camps in the Wet Tropics bioregion of North-eastern Queensland, Australia. Faecal analyses revealed that P. conspicillatus utilise a broad variety of plant resources from a variety of habitats. Seed and pulp from figs (Ficus spp., Moraceae) and pollen from the family Myrtaceae were most frequently represented in the faeces from a range of both wet sclerophyll and rainforest habitats. The dietary composition of P. conspicillatus at individual camps could not be predicted by the habitats located within a typical foraging distance of each camp (20 km), and although consistent dietary changes were seen across all camps over time, each camp had a unique dietary signature indicative of feeding on a distinct subset of available vegetation. The unique diet of each camp and the variety of dietary items consumed suggest that camps may need to be managed on an individual camp-specific basis, and that P. conspicillatus are utilising a broader range of resources than would be expected if the species was a strict ‘rainforest-fruit specialist’. Dietary variation in spectacled flying foxes (Pteropus conspicillatus) of the Australian Wet Tropics www.publish.csiro.au/journals/ajz CSIRO PUBLISHING
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Dietary variation in spectacled flying foxes ( Pteropus conspicillatus ) of the Australian Wet...

IntroductionLarge pteropodids (Megachiroptera: Pteropodidae), alsoknown as Old World fruit bats or flying foxes (Pteropusspp.), rely on a variety of plant resources and have varied for-aging and feeding strategies (Courts 1998; Augee and Ford1999; Dumont and O’Neal 2004). Australian flying foxesexhibit these varied feeding and foraging strategies withdietary flexibility or by moving in response to changes inresource availability (Nelson 1965; Parry-Jones and Augee1991; Eby 1998; Markus and Hall 2004; McDonald-Maddenet al. 2005). Pteropus conspicillatus has long been consid-ered the only flying fox on the Australian continent to be arainforest-fruit specialist (Richards 1990b; Birt et al. 1997;Garnett et al. 1999). This conclusion is supported by its asso-ciation with rainforest habitat (Richards 1990a), observa-tions of the species foraging on the fruits of rainforest treesand the morphology of their tongue and teeth, which sug-gests a frugivorous diet (Birt et al. 1997). However, specificdetails of the diet and habitat use of P. conspicillatus arescarce, and the presence of many camps outside of rainforest(Richards 1990b; Garnett et al. 1999) along with theoreticalpredictions that megachiropteran bats cannot meet theirnutritional requirements on a pure-fruit diet (Law 1992;Courts 1998) suggest that P. conspicillatus may not berestricted entirely to rainforest habitats and fruit.
Phytophagous bats have been shown to be ecologicallyimportant for a variety of ecosystem functions, such as
angiosperm pollination, seed dispersal (Williams and Adam1994; Shilton et al. 1999; Banack and Grant 2002;Singaravelan and Marimuthu 2004; Nyhagen et al. 2005)and possibly nutrient recycling (Feeley 2005). The blackflying fox Pteropus alecto plays a significant role in main-taining rainforest fragments in the Northern Territory(Woinarski et al. 1992; Palmer et al. 2000), and it is possiblethat P. conspicillatus plays a similar role in the Wet Tropicsregion of Australia (Westcott et al. 2001). Determining therole that P. conspicillatus plays in maintaining ecosystemfunction requires knowledge of the dietary habits of thespecies and how such habits may vary among camps.
Previous dietary analysis of P. conspicillatus has concen-trated on the identification of seeds in faeces, often as part offrugivory and seed-dispersal studies. However, seed identifi-cation from bat faeces suffers some significant shortcomings(Thomas 1988; Shilton 1999). First, it commonly excludeslarge seeded fruits, because these animals are able to ingestonly small seeds (Richards 1990b). Second, a focus on fruitmeans that other plant parts go undetected, particularlyflowers that require pollen analysis. A fast gut-passage time inpteropodids also means that faeces collected at the day roostis likely to consist primarily of the last feed of the night, eventhough a gut retention time of >12 h has been shown (Shiltonet al. 1999). The lack of studies that included other faecalcomponents useful for diet determination, such as fruit pulpand pollen, means that these dietary indicators and the habi-
Australian Journal of Zoology, 2006, 54, 417–428
10.1071/ZO06092 0004-959X/06/060417© CSIRO 2006
Jennifer G. ParsonsA,C, Andi CairnsA, Christopher N. JohnsonA, Simon K. A. RobsonA, Louise A. ShiltonB and David A. WestcottB
ASchool of Marine andTropical Biology, James Cook University, Qld 4811, Australia.BTropical Forest Research Centre, CSIRO Sustainable Ecosystems, PO Box 780, Atherton, Qld 4883, Australia.
CCorresponding author. Email: [email protected]
Abstract. The diet of Pteropus conspicillatus, a large flying fox, was examined by collecting faeces in trapsbeneath daytime roost trees in four geographically distinct camps in the Wet Tropics bioregion of North-easternQueensland, Australia. Faecal analyses revealed that P. conspicillatus utilise a broad variety of plant resources froma variety of habitats. Seed and pulp from figs (Ficus spp., Moraceae) and pollen from the family Myrtaceae weremost frequently represented in the faeces from a range of both wet sclerophyll and rainforest habitats. The dietarycomposition of P. conspicillatus at individual camps could not be predicted by the habitats located within a typicalforaging distance of each camp (20 km), and although consistent dietary changes were seen across all camps overtime, each camp had a unique dietary signature indicative of feeding on a distinct subset of available vegetation. Theunique diet of each camp and the variety of dietary items consumed suggest that camps may need to be managedon an individual camp-specific basis, and that P. conspicillatus are utilising a broader range of resources than wouldbe expected if the species was a strict ‘rainforest-fruit specialist’.
Dietary variation in spectacled flying foxes (Pteropus conspicillatus)of the Australian Wet Tropics
www.publish.csiro.au/journals/ajz
CSIRO PUBLISHING

J. Parsons et al.418 Australian Journal of Zoology
tats in which they are located are potentially undetected.Foraging in ‘fruit-poor, pollen-rich’ habitats such as wet anddry sclerophyll forests is unlikely to be detected by faecalstudies that concentrate on seeds only. This may explain whyflower feeding by P. conspicillatus in non-rainforest habitatshas only been infrequently recorded through directbehavioural observations (Richards 1990b; Eggert 1994).
In the present study, we described the fruit pulp, seed andpollen content of faecal samples collected beneath fourcamps of P. conspicillatus located in the Wet Tropics bio-region of Australia in order to clarify the dietary habits ofthis species, the level of dietary variation among camps andthe extent to which P. conspicillatus is a rainforest specialist.
Materials and MethodsStudy sites
P. conspicillatus faecal samples were collected in traps set below roosttrees at four camp sites (Fig. 1): three on the Atherton Tablelands(Whiteing Road, Tolga Scrub and Gadaloff Road) and one in the adja-cent lowlands (at Gordonvale). The climate of the area is tropical withtwo distinct seasons, the wet season occurring from December to Apriland the dry season from May to November. The highly variable rainfall,climate, soil types and topography of this region have resulted in theformation of a variety of vegetational communities (Adam 1992). TheAtherton Tablelands consists of a complex mosaic of habitat types withlarge areas cleared for agriculture. This has created small rainforestfragments of varying plant composition that occur throughout the land-scape. Large areas of wet sclerophyll forest occur to the west of theTablelands. Upland P. conspicillatus camps have been known to migrateto lowland areas during winter months (Garnett et al. 1999), a timeperiod that was not covered during the present study. The camp size esti-mates provided are from L.A. Shilton, P. Latch, A. McKeown andD. A. Westcott (unpublished data).
The Whiteing Road camp (145°36.3!E, 17°33.36!S, at 760 m a.s.l.)is in a fragmented highland rainforest west of Milla Milla, surroundedby pastoral land. The vegetation of the camp is Complex Notophyll VineForest Type 5a (Tracey 1982) and has many epiphytes and woody lianes.Sclerophyllous habitat is absent within 10 km of the camp (Fig. 1). Thisis a seasonally occupied maternal camp used by both P. conspicillatusand P. scapulatus (little red flying foxes). During this study, numbers ofP. conspicillatus fluctuated between 12000 and 20000 individuals.
The Tolga Scrub camp (145°28.8!E, 17°13.86!S, at 770 m a.s.l.) islocated in a small and isolated fragment of Complex Notophyll VineForest Type 5b (Tracey 1982), outside the township of Atherton. Thistype of forest was once widespread on the Atherton Tablelands butowing to heavy land clearance is now restricted to small patches offorest near Atherton (Tracey 1982). Approximately 88% of the landwithin a 10-km radius of the camp has been cleared of native vegetation(Stanton and Stanton 2005), whereas a relatively small area of rain-forest and substantial areas of sclerophyll forest are located within a20-km radius (Fig. 1). The forest canopy height is uneven and manycanopy species are semi-deciduous. There is a dense shrub layer, dom-inated by Hodgkinsonia frutescens and a high abundance of Calamuscaryotoides that, together with a paucity of epiphytes, is typical of drierrainforest communities (Tracey 1982). Tolga Scrub is occupied byP. conspicillatus throughout the year (a permanent maternal roost), andnumbers of P. conspicillatus fluctuated from 10000 to 25000 during theperiod of the study. Large numbers (>100000) of P. scapulatus werealso present at this site during the study period.
The Gadaloff Road camp (145°42.6!E, 17°20.82!S, at 720 m a.s.l.)is in a steep valley of revegetated dry rainforest with Acacia celsa as a
successional dominant, and is adjacent to large areas of farmland aswell as wet rainforest and sclerophyll forest (Fig. 1). P. conspicillatusoccupy this camp seasonally, and numbers during this study fluctuatedbetween 70 and 5000 animals. The camp was comprised predominantlyof young male or non-breeding female P. conspicillatus (L. A. Shilton,P. Latch, A. McKeown and D. A. Westcott, unpublished data), unlikeany of the other camps sampled. No other flying fox species used theGadaloff Road camp site during the present study.
The Gordonvale camp (145°46.74!E, 17°4.86!S, at 40 m a.s.l.) is inlow to medium mixed shrubland and woodland with Melaleuca quin-quinerva as the dominant species. It is surrounded by agricultural land(mainly sugarcane plantations) and suburban housing. Around 74% ofthe natural vegetation within a 10-km radius of the camp has beencleared (Stanton and Stanton 2005). Flying foxes at this camp had readyaccess to continuous rainforest and sclerophyll forest (Fig. 1). This wasthe largest camp sampled, with over 40000 P. conspicillatus presentduring the present study. This is a permanent maternal camp. No otherflying fox species were sighted at the Gordonvale camp during thepresent study.
The field work for the present study was undertaken during the wetseason from November 2004 to March 2005. This period covered thebreeding season of P. conspicillatus and included the peak fruiting of rain-forest and commercially grown trees in the area (Westcott et al. 2001).
km
Fig. 1. Map of P. conspicillatus camp study sites, displayingvegetation types with radii of 5, 10 and 20 km surrounding each camp.Base data reproduced with permission of Wet Tropics ManagementAuthority.

Australian Journal of Zoology 419
Experimental procedures
P. conspicillatus diet was documented by collecting faeces at each of thefour camps. Collection traps were 1 m2, and consisted of a fine syn-thetic woven material (~1 mm diameter mesh) on a PVC piping frame.Each trap was suspended beneath roost trees in P. conspicillatus campsby attaching fishing line to vegetation to limit seed predation byrodents. Flying foxes were not handled and faecal traps were addedduring the day. There was no obvious disturbance to flying foxes duringdaytime recovery of faecal material. Faecal traps were placed randomlyin areas that were accessible, provided that they were under roostingtrees in which only P. conspicillatus were present. P. conspicillatus areeasily distinguished from other Pteropus spp. in the area owing to theirdistinctive markings of pale fur surrounding the eyes and on thecollar/mantle. P. conspicillatus are also notably larger than the othercommon flying fox species in the area, P. scapulatus. Roosting trees areusually only occupied by one species (Richards 1990a); however, smallnumbers of P. conspicillatus have been observed roosting within thesame trees as P. scapulatus (L. Shilton, personal observation). Trapswere left in position for the duration of the study but were cleaned andrepaired before each sampling period. No mixed species roosting treeswere observed over traps at any time during sampling.
To determine the optimum number of 1-m2 traps required to repre-sent diet diversity at each camp, a pilot study was conducted over fivedays at Whiteing Road and Tolga Scrub camps. This involved placing20 traps at each camp and collecting trap contents each day. The meannumber of dietary species (represented by seeds) collected per trap wasplotted against the number of traps analysed. This showed that, onaverage, 84.21% of plant species were documented using only 10 traps.Consequently, 10 traps were used per site for the main study.
Each of the four sites were visited on every second day for a periodof 10 days (five trapping days per site) per month, to ensure (1) thatfaecal material was collected when still fresh, allowing for fruit pulpidentification on the basis of colour and texture, and (2) that samplesfrom different camps were collected concurrently. Faecal material wascollected from each of the four sites for a total of 1600 trapping nights.Tolga Scrub and Whiteing Road were sampled in November andDecember 2004 and in January and March 2005. Gordonvale andGadaloff Road were sampled in December 2004 and January 2005.
Faecal content identification
To eliminate bias associated with faecal trapping, all seeds, fruit pulpand pollen were identified to plant species level wherever possible. Allother material was identified or recorded as leaf fragments, bark, flowerstamens, lichen, other flower parts, hair or insect exoskeleton.
Faeces were collected, prepared and analysed individually. Sampleswere placed in marked click-seal plastic bags and refrigerated as soonas possible. Climatic conditions were recorded at each site on each sam-pling day. We also recorded the number of P. conspicillatus roosting inthe canopy above each trap and the presence of any other Pteropus spp.at the site.
Samples were diluted with water and strained through a fine mesh(1 mm2), and the retained solid matter was placed in a Petri dish, driedand identified under a Leica MZ6 stereo-dissecting microscope(ranging from 6.5 to 40" magnification). Seeds were identified usingseed reference material at CSIRO’s Tropical Forest Research Centre(CSIRO TFRC) and in comparison with seeds already identified duringthe present study. When #5 seeds of a particular type were collected infaecal material, these seeds were kept as reference material and not ger-minated. Seeds surplus to the first five were planted in soil to germinatein greenhouses in Atherton and Townsville. Seeds were grown to thetenth leaf stage wherever possible, and identified using botanical keys(Hyland et al. 2002) and in consultation with botanists. Fruit pulp wasidentified where possible from the colour, texture and odour in com-parison with reference fruits.
Pollen grains were extracted from a smaller sub-sample of faecalmaterial using the potassium hydroxide-acetolysis method (Faegri andIverson 1975). These were then suspended in silicone oil and mountedon a microscope slide. Pollen samples were examined at "400 magnifi-cation using a Leitz Dialux microscope. Plant taxa represented bypollen in each sample were identified with the assistance of referencematerial collected during the course of the present study, electronicpollen reference collections (maintained by the University of Arizona,USA, University of Newcastle, NSW, and the Australian NationalUniversity, ACT) and published sources (Pike 1955; Sedgley et al.1993). Presence/absence of a plant species was recorded.
Statistical analysis
To quantify patterns in the data, the number of species represented bypollen or seeds in each faecal sample was first reduced using principalcomponents analysis (PCA). To meet the normality assumptions ofPCA, it was necessary to remove species that were only rarely found infaeces (Quinn and Keough 2002). Abundances were ranked, and thosespecies that comprised either less than 1% of total faecal samples, orwere present in less than 50% of these eight camp " month categories,were excluded from the analyses. Individual frequencies (the number offaecal samples) were converted into the percentage of the total numberof faecal samples within each camp " month group. PCA analysis usedthe extracted covariance matrix to give greater weighting to the relativeabundance of each species in the diet. A clustering function (FAST-CLUS) was then used to determine if the camp " month groups couldbe clustered into distinct groups based on their PCA factor scores. Thisfunction used nearest centroid sorting to place camp " month groupsinto a predetermined number of groups based on the overall similarityof their PCA scores. Finally, a canonical discriminant analysis(CANDISC) was used to determine if the groups defined by the clus-tering function were statistically discrete.
To determine if there was a relationship between the diet of P. con-spicillatus and available habitat, the correlation between the availablehabitat and diet was examined for two vegetation types (rainforest andsclerophyll). Spearman’s rank correlation was used for this analysis,and the proportion of habitat types was determined using Stanton veg-etation mapping (Stanton and Stanton 2005). Radii of 5, 10 and 20 kmfrom each camp were chosen for habitat comparison, because P. con-spicillatus generally forage within 20 km of a camp (L. A. Shilton,P. Latch, A. McKeown and D. A. Westcott, unpublished data).
Frequency analyses used either chi-square analysis or Monte CarloRxC contingency analysis (Engels 1988) with 10000 replicates. FAST-CLUS and CANDISC were performed with SAS version 8.2. All otherstatistical analyses were performed with SPSS version 11. In all cases,the formal analysis of temporal and/or spatial patterns in resource useused the balanced data set in which diet information had been obtainedfor all four camps during the two sample months of December andJanuary.
Results
Dietary analysis
A total of 2071 faecal samples were analysed for the pres-ence of seed or any other recognisable organic matter. Therewere significant differences in the frequency of occurrenceof seed, hair, bark, and insects sampled in P. conspicillatusfaeces among the four sites ($2 = 349.09, d.f. = 12,P < 0.001).
During the present study, a total of 43 plant taxa wereidentified in the diet of P. conspicillatus using both seed/pulpand pollen analysis (Table 1). In total, the known diet of
Dietary variation in Pteropus conspicillatus

J. Parsons et al.420 Australian Journal of Zoology
P. conspicillatus includes the fruit of 49 native species,19 exotic species and floral resources from 25 native species(Richards 1990b; Eggert 1994). The present study added sixplant taxa from which fruits are eaten and 18 plant taxa fromwhich floral resources are consumed by P. conspicillatus,increasing the published dietary inventory of P. conspi-cillatus by 25.8%.
Spatial variation
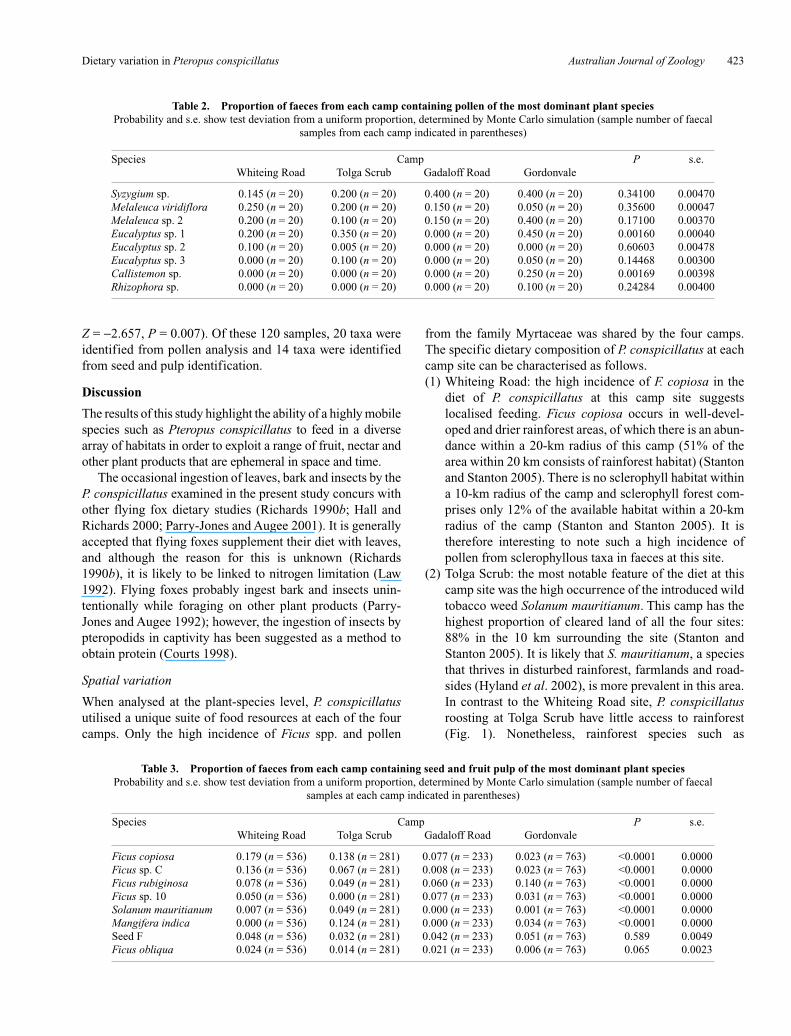
P. conspicillatus from each of the four camps utilised a dis-tinct suite of habitats and plant species over the course of thepresent study; this included fruit and flower resources from43 plant taxa (Table 1), of which 22 taxa were not sharedamong camps. To characterise the diet profile of each camp,we used the five most frequently recorded plant species,which accounted for 77 to 88% of dietary resources presentin faecal samples (Figs 2 and 3). The only taxa that showedsignificant probability values (against an expected distri-bution) calculated for pollen were Callistemon sp. and
Eucalyptus sp. 1 (Monte Carlo, P = 0.016, s.e. = 0.0039 andP = 0.0016, s.e. = 0.0004 respectively) (Table 2). The pro-portion of total faecal samples containing particular plantspecies varied significantly among camps for almost all ofthe most frequently occurring seed and fruit pulp species(Table 2). For example, Ficus copiosa varied from propor-tions of 0.179 at Whiteing Road to proportions of 0.023 atGordonvale (P < 0.0001, s.e. = 0.000) (Table 3). The onlyplant species that did not vary based on seed/fruit pulp pres-ence in faeces was an undescribed species, Seed F(P = 0.589, s.e. = 0.0049) and Ficus obliqua (P = 0.065,s.e. = 0.0023) (Table 4). Fruit and pulp remnants were mostlyrepresented by 13 spp. from the Ficus genus (Table 1). Figdetritus was present in 757 faecal samples (s.d. = 52.31,n = 2071) and comprised 59.19% of all the fruits recorded.Pollen from the family Myrtaceae was the most frequentlyrepresented (Table 1) and was identified in 96 faecal samples(s.d. = 4.91, n = 120), representing 79.9% of plant taxa iden-tified from pollen in samples.
Table 1. A list of plant species present in P. conspicillatus faecal materials fromfour camps (November 2004 to March 2005)
Also listed is the percentage of each plant species represented in faecal material and ifit is represented by pollen (P), seed (S) or other (O)
Family Genus/species P (%) S (%) O (%)
Anacardiaceae Mangifera indica 4.7Arecaceae Calamus sp. 0.07Combretaceae Terminalia sp. 0.6Dodonaea 1 unknown species 0.6Elaeocarpaceae Elaeocarpus sp. 1.21 0.15Euphorbiaceae Mallotus sp. 0.6Lecythidiaceae Baringtonia sp. 0.6Moraceae F. copiosa 13.36
F. obliqua 2.11F. pleurocarpa 0.39F. racemosa 2.03F. rubiginosa 21.428 unidentified Ficus sp. 19.85
Myrtaceae Calllistemon sp. 4.87Corymbia sp. 1.2Eucalyptus tereticornis 1.823 unidentified Eucalyptus sp. 24.39Melaleuca viridiflora 12.8Melaleuca sp. 15.24Psidium. guajava 0.78Syzygium sp. 20.73
Passifloraceae Passiflora sp. 1.56Proteaceae Banksia sp. 3.04
Grevillea sp. 2.43Rhizophoraceae Rhizophora sp. 1.21Rosaceae Rubus sp. 0.39Rubiaceae Timonius timon 0.54
1 unidentified sp. 1.82Rutaceae 2 unidentified sp. 1.21Sapindaceae Litchi chinensis 0.31
1 unidentified sp. 0.6Solanaceae Solanum mauritianum 15.4
S. torvum 1.64

Australian Journal of Zoology 421
We did not find a clear relationship between local habitat(within a 20-km radius) and P. conspicillatus diet. Forexample, there was no correlation between the percentage ofrainforest or sclerophyll plant taxa represented in the diet andthe percentage of rainforest or sclerophyll around the camps.This was tested for all camps with rainforest available within5 km (Spearman’s rS = 0.688, n = 4, P = 0.332), 10 km(rS = 0.533, n = 4, P = 0.447) and 20 km (rS = 0.443, n = 4,P = 0.557), and with sclerophyll within 5 km (rS = 0.727,n = 4, P = 0.273), 10 km (rS = 0.685, n = 4, P = 0.315) and20 km (rS = 0.552, n = 4, P = 0.448).
Temporal variation
For plant species identified from seed and fruit pulp in faecalsamples, the amount of variation explained by the first fiveprinciple components and the factor loadings of each are pre-
sented in Table 4. When combined, the cumulative total vari-ation explained by the first four components extracted fromthe PCA was 36.87, 64.23, 80.3 and 90.8% respectively. Themajor loadings on each principal component appeared toreflect the presence of Ficus spp. in the diet. For example, thefirst principal component was most heavily weighted by thepercentage of Ficus copiosa (factor loading = %15.32),F. rubiginosa (factor loading = 10.92) and Ficus sp. 10(factor loading = 9.07). Similarly, the second principal com-ponent was most heavily weighted by the percentage ofF. rubiginosa, Ficus sp. C and F. copiosa in the diet (factorloading scores = 13.11, 10.26 and %5.65 respectively).
The first two principal components suggest that the camp" month groups fall within three categories, with groupstending to clump together on the basis of the months inwhich they were sampled (Fig. 4). For example, all four sites
Dietary variation in Pteropus conspicillatus
Ficus copiosa Ficus sp. C Seed T Ficus rubiginosa
Ficus sp. 100
20
40
60
80
100
Num
ber o
f fae
cal s
ampl
es
Whiteing Rd
n = 685 faecal samples
Solanum mauritianum
Ficus rubiginosa
Ficus sp. C Ficus copiosa Mangifera indica
0
50
100
150
200 Tolga Scrub
n = 390 faecal samples
Ficus rubiginosa
Seed F Mangifera indica
Ficus sp. 10 Ficus sp. C0
20
40
60
80
100
120 Gordonvale
n = 763 faecal samples
Ficus copiosa Ficus sp. 10 Ficus rubiginosa
Seed F Ficus obliqua0
5
10
15
20 Gadaloff Rd
n = 233 faecal samples
(a) (b)
(c) (d)
Fig. 2. The most frequently occurring seed and fruit pulp species represented in faecal samples at four P. conspicillatus camps over the 4 monthsof the study.

J. Parsons et al.422 Australian Journal of Zoology
that were sampled during December clustered together, asdid three of the four sites sampled in January (WhiteingRoad, Tolga Scrub and Gadaloff Road). The single remainingcamp " month group (Gordonvale in January) appeared to beseparate from each of the preceding two groups.
The FASTCLUS function confirmed that the clustersindicated in Fig. 4 represent the most appropriate threegroupings for the eight camp " month points. The canonicaldiscriminant analysis indicated that these three clusters dif-fered significantly from each other (Wilks’ & = 0.009, F4,8. =19.02, P < 0.001). When the same analysis was run forpollen, no discrete groups were found.
It is worth noting that the clustering patterns found in thisanalysis were not sensitive to the exclusion principals used.Repeating this analysis with the inclusion of all data, irre-
spective of its rarity, still produced principal componentsheavily weighted by figs. It also produced the same forma-tion of three clusters supported by the clustering function,and the detection of significant differences among the threeclusters based on their PCA scores.
Comparison of number of taxa identified from seedand pollen
Owing to the time-consuming processing used to extractpollen from faecal samples, 120 samples were randomlyselected to represent the four camps and the time periodsampled. Of these samples, 69.6% contained pollen. Whencomparing seed with pollen in the sub-sample, significantlymore plant taxa were found using pollen analysis than usingseed or fruit pulp analyses (Mann–Whitney U-test, n = 24,
–P.
Syzygium sp.
Melaleuca viridiflora
Melaleuca sp. 2
Eucalyptus sp. 1
Eucalytpus sp. 2
0
2
4
6
8
10
12 Whiteing Rd
n = 40 faecal samples
Eucalyptus sp. 1
Melaleuca viridiflora
Syzygium sp.
Eucalytpus sp. 3
Melaleuca sp. 2
0
2
4
6
8
10Tolga Scrub
n = 40 faecal samples
Eucalyptus sp. 1
Melaleuca sp. 2
Syzygium sp.
Callistemon Rhizophora
0
2
4
6
8
10 Gordonvale
n = 20 faecal samples
Syzygium sp. Melaleuca sp. 2 Melaleuca viridiflora0
2
4
6
8Gadaloff Rd
n = 20 faecal samples
Num
ber o
f fae
cal s
ampl
es
(a) (b)
(c) (d)
Fig. 3. The most frequently occurring pollen species represented in faecal samples at four P. conspicillatus camps over the 4 months of the study.
Australian Journal of Zoology 423
Z = %2.657, P = 0.007). Of these 120 samples, 20 taxa wereidentified from pollen analysis and 14 taxa were identifiedfrom seed and pulp identification.
Discussion
The results of this study highlight the ability of a highly mobilespecies such as Pteropus conspicillatus to feed in a diversearray of habitats in order to exploit a range of fruit, nectar andother plant products that are ephemeral in space and time.
The occasional ingestion of leaves, bark and insects by theP. conspicillatus examined in the present study concurs withother flying fox dietary studies (Richards 1990b; Hall andRichards 2000; Parry-Jones and Augee 2001). It is generallyaccepted that flying foxes supplement their diet with leaves,and although the reason for this is unknown (Richards1990b), it is likely to be linked to nitrogen limitation (Law1992). Flying foxes probably ingest bark and insects unin-tentionally while foraging on other plant products (Parry-Jones and Augee 1992); however, the ingestion of insects bypteropodids in captivity has been suggested as a method toobtain protein (Courts 1998).
Spatial variation
When analysed at the plant-species level, P. conspicillatusutilised a unique suite of food resources at each of the fourcamps. Only the high incidence of Ficus spp. and pollen
from the family Myrtaceae was shared by the four camps.The specific dietary composition of P. conspicillatus at eachcamp site can be characterised as follows.(1) Whiteing Road: the high incidence of F. copiosa in the
diet of P. conspicillatus at this camp site suggestslocalised feeding. Ficus copiosa occurs in well-devel-oped and drier rainforest areas, of which there is an abun-dance within a 20-km radius of this camp (51% of thearea within 20 km consists of rainforest habitat) (Stantonand Stanton 2005). There is no sclerophyll habitat withina 10-km radius of the camp and sclerophyll forest com-prises only 12% of the available habitat within a 20-kmradius of the camp (Stanton and Stanton 2005). It istherefore interesting to note such a high incidence ofpollen from sclerophyllous taxa in faeces at this site.
(2) Tolga Scrub: the most notable feature of the diet at thiscamp site was the high occurrence of the introduced wildtobacco weed Solanum mauritianum. This camp has thehighest proportion of cleared land of all the four sites:88% in the 10 km surrounding the site (Stanton andStanton 2005). It is likely that S. mauritianum, a speciesthat thrives in disturbed rainforest, farmlands and road-sides (Hyland et al. 2002), is more prevalent in this area.In contrast to the Whiteing Road site, P. conspicillatusroosting at Tolga Scrub have little access to rainforest(Fig. 1). Nonetheless, rainforest species such as
Dietary variation in Pteropus conspicillatus
Table 2. Proportion of faeces from each camp containing pollen of the most dominant plant speciesProbability and s.e. show test deviation from a uniform proportion, determined by Monte Carlo simulation (sample number of faecal
samples from each camp indicated in parentheses)
Species Camp P s.e.Whiteing Road Tolga Scrub Gadaloff Road Gordonvale
Syzygium sp. 0.145 (n = 20) 0.200 (n = 20) 0.400 (n = 20) 0.400 (n = 20) 0.34100 0.00470Melaleuca viridiflora 0.250 (n = 20) 0.200 (n = 20) 0.150 (n = 20) 0.050 (n = 20) 0.35600 0.00047Melaleuca sp. 2 0.200 (n = 20) 0.100 (n = 20) 0.150 (n = 20) 0.400 (n = 20) 0.17100 0.00370Eucalyptus sp. 1 0.200 (n = 20) 0.350 (n = 20) 0.000 (n = 20) 0.450 (n = 20) 0.00160 0.00040Eucalyptus sp. 2 0.100 (n = 20) 0.005 (n = 20) 0.000 (n = 20) 0.000 (n = 20) 0.60603 0.00478Eucalyptus sp. 3 0.000 (n = 20) 0.100 (n = 20) 0.000 (n = 20) 0.050 (n = 20) 0.14468 0.00300Callistemon sp. 0.000 (n = 20) 0.000 (n = 20) 0.000 (n = 20) 0.250 (n = 20) 0.00169 0.00398Rhizophora sp. 0.000 (n = 20) 0.000 (n = 20) 0.000 (n = 20) 0.100 (n = 20) 0.24284 0.00400
Table 3. Proportion of faeces from each camp containing seed and fruit pulp of the most dominant plant speciesProbability and s.e. show test deviation from a uniform proportion, determined by Monte Carlo simulation (sample number of faecal
samples at each camp indicated in parentheses)
Species Camp P s.e.Whiteing Road Tolga Scrub Gadaloff Road Gordonvale
Ficus copiosa 0.179 (n = 536) 0.138 (n = 281) 0.077 (n = 233) 0.023 (n = 763) <0.0001 0.0000Ficus sp. C 0.136 (n = 536) 0.067 (n = 281) 0.008 (n = 233) 0.023 (n = 763) <0.0001 0.0000Ficus rubiginosa 0.078 (n = 536) 0.049 (n = 281) 0.060 (n = 233) 0.140 (n = 763) <0.0001 0.0000Ficus sp. 10 0.050 (n = 536) 0.000 (n = 281) 0.077 (n = 233) 0.031 (n = 763) <0.0001 0.0000Solanum mauritianum 0.007 (n = 536) 0.049 (n = 281) 0.000 (n = 233) 0.001 (n = 763) <0.0001 0.0000Mangifera indica 0.000 (n = 536) 0.124 (n = 281) 0.000 (n = 233) 0.034 (n = 763) <0.0001 0.0000Seed F 0.048 (n = 536) 0.032 (n = 281) 0.042 (n = 233) 0.051 (n = 763) 0.589 0.0049Ficus obliqua 0.024 (n = 536) 0.014 (n = 281) 0.021 (n = 233) 0.006 (n = 763) 0.065 0.0023

J. Parsons et al.424 Australian Journal of Zoology
F. copiosa and F. racemosa were represented in their dietin high proportions during the study. This finding alsosuggests that P. conspicillatus traveled over a large dis-tance (>20 km) from Tolga Scrub to access these foodresources.
(3) Gadaloff Road: the fruit diet of P. conspicillatus at thiscamp consisted almost entirely of the fruit of Ficus spp.;additionally, this camp yielded the least number ofspecies represented by pollen, and Eucalyptus pollen wasnot recorded in faecal samples. The incidence ofSyzygium spp. in samples was high. P. conspicillatusroosting at this camp have access to both rainforest andsclerophyll forest within 10 km (Stanton and Stanton2005). The diet of P. conspicillatus at this site suggestsregular foraging in rainforest habitat, with several rain-forest species frequently occurring in the diet. In con-trast, the habitat surrounding the camp site represents avariety of different habitat types, including sclerophyll.
(4) Gordonvale: As Gordonvale was the only lowland campsite in the present study, we expected resource use byP. conspicillatus to reflect a greater use of lowland vege-tation types. Although some plant species rarely recordedin the diet were unique to the faecal material collected atthis site, the samples indicated a similar reliance on thefruits of Ficus spp. and myrtaceous floral resources. Therewas also a high incidence of Mangifera indica (mango)pulp in faeces. This is not surprising, considering that P.conspicillatus in this area have access to commercialorchards within a 20-km radius (Stanton and Stanton2005). This camp is surrounded by a greater variety ofhabitat types and was the only camp from which P. con-spicillatus showed evidence of foraging in mangroves(Rhizophora sp.). This may explain why it was the onlycamp in the PCA to deviate from temporal clustering.
The reasons why the P. conspicillatus roosting at thesefour camp sites used such different plant resources areunknown. One of the most interesting results from thepresent study is that camp location and proximity to foodplants did not account for differences in diet. Although theenergetic benefits of roosting close to foraging areas arelikely to be high, this advantage is apparently over-ridden bythe temporal distribution of resources across the landscape aswell as within different habitat types. Perhaps the competi-tion for accessible food resources surrounding camps is high,which forces individual P. conspicillatus to forage furtheraway from the camp.
Different fruiting and flowering phenologies clearly havean effect on how P. conspicillatus uses the landscape but whenseveral resources are available at the same time at varying dis-tances from camps; (i.e. peak rainforest fruiting and peakEucalyptus spp. flowering occur at the same time inDecember), why are these flying foxes traveling such largedistances to use these resources? We suggest that some addi-tional benefits are gained from using resources further a field.If megachiropterans are nitrogen-limited when feeding onfruits (Law 1992; Courts 1998) and are able to supplementthis deficiency by feeding on floral resources, this couldexplain why P. conspicillatus are traveling so far to reachfloral resources. This suggests that a combined diet of fruitsand flowers is necessary to sustain P. conspicillatus, evenduring periods of high fruit availability in the wet season. Totest this possibility, a much more detailed study of P. con-spicillatus diet in conjunction with phenological studies offruiting and flowering is needed. Alternatively, P. con-spicillatus could be displaying a preference for ephemerallyabundant food resources. At the time of the present study,Eucalyptus spp. were in mass flower and presented an easilylocatable, densely distributed and super-abundant foodresource. Rainforest fruits, though at their peak of availability,are likely to have been more dispersed and less easily located(M. Bradford, personal communication).
Dietary variation
Old world fruit bats are known to have a reliance on Ficusspp. (Shanahan et al. 2001), with the diet of localised popu-lations of some species consisting almost entirely of figs(Shilton 1999). This trend is also apparent in the diet ofP. conspicillatus. The bias towards only small-seeded speciesoccurring in faeces results from the small lumen diameter ofthe oesophagus (Richards 1990b), which could account thistrend. However, the consistent high abundance of Ficus spp.seeds in faecal samples from each camp and throughout theperiod of study suggests that Ficus spp. are an importantcomponent of the diet of P. conspicillatus.
Our results show a high incidence of pollen from thefamily Myrtaceae, a family that is known to be an importantcomponent of the diet of many other AustralianMegachiroptera (Parry-Jones and Augee 1992; Law 2001).
Table 4. The effect of each plant species determined by thepresence of seed or pulp on the first four component scores
of the principal component analysisAlso listed is the % variance and cumulative % variance explained
by each component
Species Component1 2 3 4
Ficus rubiginosa 10.92 –13.11 –4.57 2.14Ficus copiosa –15.32 –5.65 4.5 2.64Ficus sp. C 2.4 10.26 –2.75 7.51Seed F –4.87 –3.77 –0.56 –2.43Ficus sp. 10 9.07 2.35 9.86 –3.55Mangifera indica –2.83 4.56 –7.29 –7.15Ficus obliqua 2.6 2.75 2.28 –0.44Ficus racemosa –2.28 0.11 –0.44 0.17Solanum mauritianum –1.17 2.36 –2.44 –0.34Ficus sp. 9 1.71 0.65 1.62 0.37Solanum torvum –0.22 –0.52 –0.21 1.07% variance explained 36.87 27.36 16.08 10.49Cumulative % variance 36.87 64.23 80.3 90.8

Australian Journal of Zoology 425
It is likely that flying foxes are not only ingesting pollen butalso drinking the nectar from flowers on which they arefeeding. This finding demonstrates the range of habitattypes being used by P. conspicillatus. A large portion of theplants from the family Myrtaceae found in P. conspicillatusfaeces occur in sclerophyll or non-rainforest habitats (e.g.Eucalyptus spp. and Melaleuca spp.). Although flower-feeding by P. conspicillatus has been previously recorded(Richards 1990b; Eggert 1994), the prevalence of floralresources in the diet of P. conspicillatus has not previouslybeen recognised. It is apparent that individuals in theWhiteing Road camp are traveling large distances (>10 km)to access sclerophyll trees at times when there are rainforestresources available within a much shorter distance; this isshown by the presence of rainforest fruits in faeces coinci-dent with Myrtaceous pollen. Studies of movement patternsby P. conspicillatus in the same population also suggestlarge foraging commutes by individual animals(L. A. Shilton, P. Latch, A. McKeown and D. A. Westcott,unpublished data).
The paucity of commercially grown fruits in the diet ofP. conspicillatus suggests that these fruits formed only aminor component of their diet during the period of thisstudy. However, because larger seeded fruits were generallyexcluded by our sampling methods (even though pulp infaeces was recorded and identified wherever possible), it islikely that many large-seeded commercial crops (e.g.mango, lychee, stonefruit) would be under-represented.Nevertheless, previous studies that used similar samplingmethods recorded a much larger range of commerciallyavailable fruits in the diet of P. conspicillatus (Richards1990b; Eggert 1994; Tidemann et al. 1997). In part, thismay have been a result of a bias attributed to the ease ofpopulation estimation in orchards compared with remoteareas of natural habitat (Richards 1990b). A feeding prefer-ence and nutritional benefit of native forest foods over com-mercially grown resources has been demonstrated for otherPteropus species (Banack 1998; Nelson et al. 2000). Thiscould lead to the conclusion that although these exotic fruitsdo form part of the diet of P. conspicillatus, these resources
Dietary variation in Pteropus conspicillatus
–1.50 –1.00 –0.50 0.00 0.50 1.00 1.50
1.00
0.00
–1.00
–2.00
REGR factor score 1
Gadaloff Road -December
Gadaloff Road -January
Gordonvale -December
Gordonvale -January
Tolga Scrub -December
Tolga Scrub -January
Whiteing Road -December
Whiteing Road -January
RE
GR
fact
or s
core
2
Camp-Month
Fig. 4. Scatterplots of two principal component analysis factor scores explaining 64% of thevariation in the diet of P. conspicillatus. The effect of principle factors 1 and 2 on each camp " monthpoint is shown. This forms three distinct groups.

J. Parsons et al.426 Australian Journal of Zoology
will be used sparingly when native resources are availableand abundant.
Temporal variation
It is likely that P. conspicillatus would vary its diet inresponse to the differing flower and fruiting phenologies ofangiosperms. It has been suggested that flying foxes aggre-gate during times of peak flowering (Parry-Jones and Augee2001). The time period covered by this study incorporatedthe peak of rainforest and commercial fruiting, in addition tothe flowering peak of many myrtaceous species on theAtherton Tablelands (M. Bradford, personal communication;L. A. Shilton, P. Latch, A. McKeown and D. A. Westcott,unpublished data). A temporal trend was detected for fruitusing PCA. Our results suggest that, with the exception ofGordonvale, different P. conspicillatus camps display asimilar pattern of variation in their fruit diet over time.Gordonvale was the only lowland camp site examined and,because it is also semi-urban, it provides access to a widervariety of habitat types (Fig. 1). There was no consistentpattern of change for pollen taxa. Although the seasonalpulses of flowering are well known (Waser et al. 1996), it ispossible that they were not detected in this study owing to itsshort duration. The identification of pollen to species level isoften not possible and, as a result, fine-scale flowering pulsesof different myrtaceous species within the same genus mayhave also been missed. By extending the time period coveredby this study and increasing the sample sizes for pollenanalysis, a comprehensive assessment of the effect of theseflowering and fruiting phenologies on P. conspicillatus couldbe undertaken in the future.
Is P. conspicillatus a rainforest fruit specialist?
Describing a species as being either a generalist or specialistcan be problematic, owing to the relative nature of this termand the need for a clear understanding of the dietary habitsof the species with which it is being compared. For example,species with relatively low and high dietary overlap are oftenreferred to as generalists and specialists respectively (Eby1998).
Pteropodids are often considered dietary generalists whencompared with other bats (Fleming 1993), but the degree ofgeneralisation or specialisation appears to vary within thegroup as a whole. Within Australian flying fox species,Pteropus alecto has been called a dietary generalist becauseit feeds on a combination of nectar, pollen and rainforestfruits (Richards 1995; Palmer et al. 2000; Markus and Hall2004). Pteropus poliocephalus has been termed both a gen-eralist (Eby 1998) and a more specialised nectivore (Markusand Hall 2004), a term more commonly associated withP. scapulatus (Nelson 1965; Birt et al. 1997; Markus andHall 2004). Other flying foxes such as P. samoensis andP. tonganus have been described as ‘sequential specialists’(Banack 1998) because they are considered to specialise sea-
sonally on one or a few plant species among a group ofpotential food types at any one time (Marshall 1985).Previous studies of P. conspicillatus noted that rainforestfruits are a primary component of their diet, and concludedthat this species is a rainforest specialist (Richards 1990b;Eggert 1994). In this study we extended these results anddemonstrated that although P. conspicillatus can roost inrainforest and consume a variety of rainforest taxa, they alsoconsume a variety of non-rainforest plants from other habi-tats such as wet sclerophyll. Although it might be difficult tosay where P. conspicillatus falls on the spectrum of dietaryspecialist v. generalist, these studies together suggest thatP. conspicillatus may be more similar in foraging habits toother Australian flying foxes than has been recognised todate (Richards 1990b; Parry-Jones and Augee 1992; Birtet al. 1997; Eby 1998; Markus and Hall 2004). Therefore, theconservation management of the species could present manyof the same issues as for other mainland flying foxes.
The continued survival of Australian flying foxes intoday’s transformed landscape has been attributed to theirability to interchange between dietary resources (Nelson1965; Birt et al. 1997). The results presented here indicatethat the dietary resources utilised by P. conspicillatus aremore diverse than previously thought, suggesting that thisspecies may be better able to respond to future landscapechanges owing to increased urbanisation and global warmingthan previously thought. However the presence of campswith unique dietary signatures and the use of a variety of dif-ferent habitats highlights the need for management decisionsto take a broad-scale approach to understanding the ecologyof this species. Studies of single camps in single habitattypes would not be sufficient.
Acknowledgments
This study was supported by the Rainforest CooperativeResearch Centre and James Cook University. Botanicaladvice was provided by Dr Dale Dixon (NT Herbarium),Matt Bradford and Andrew Ford (CSIRO). Assistance wasprovided by Dr Jon Luly and The School of Earth andEnvironmental Sciences at James Cook University. Adviceand support was also provided from J. Maclean and Tolga BatResearch and Rescue, L. Ezzy, D. Bower and J. Rowley.
ReferencesAdam, P. (1992). ‘Australian Rainforests.’ (Oxford University Press:
Oxford.)Augee, M. L., and Ford, D. (1999). Radio-tracking studies of grey-
headed flying foxes, Pteropus poliocephalus, from the Gordoncolony, Sydney. Proceedings of the Linnean Society of New SouthWales 121, 61–70.
Banack, S. A. (1998). Diet selection and resource use by flying foxes(Genus Pteropus). Ecology 79, 1949–1967. doi:10.2307/176701
Banack, S. A., and Grant, G. S. (2002). Spatial and temporal movementpatterns of the flying fox, Pteropus tonganus, in American Samoa.The Journal of Wildlife Management 66, 1154–1163.

Australian Journal of Zoology 427
Birt, P., Hall, L. S., and Smith, G. C. (1997). Ecomorphology of thetongues of Australian Megachiroptera (Chiroptera: Pteropodidae).Australian Journal of Zoology 45, 369–384. doi:10.1071/ZO97005
Courts, S. E. (1998). Dietary strategies of Old World Fruit Bats(Megachiroptera, Pteropodidae): how do they obtain sufficientprotein? Mammal Review 28, 185–194. doi:10.1046/j.1365-2907.1998.00033.x
Dumont, E. R., and O’Neal, R. (2004). Food hardness and feedingbehavior in old world fruit bats (Pteropodidae). Journal ofMammalogy 85, 8–14. doi:10.1644/BOS-107
Eby, P. (1998). An analysis of diet specialisation in frugivorousPteropus poliocephalus (Megachiroptera) in Australian sub-tropicalrainforest. Australian Journal of Ecology 23, 443–456. doi:10.1111/j.1442-9993.1998.tb00752.x
Eggert, C. (1994). Is tick paralysis in the spectacled flying fox,Pteropus conspicillatus, related to a change in the foragingbehaviour of P. conspicillatus? Honours thesis, Southern CrossUniversity, Lismore.
Engels, W. (1988). Monte Carlo RxC Contingency Table Test. Availableat http://engels.genetics.wisc.edu/ [Verified 6 December 2006].
Faegri, K., and Iverson, J. (1975). ‘Textbook of Pollen Analysis.’(Hafner Press: New York.)
Feeley, K. (2005). The role of clumped defecation in the spatial distri-bution of soil nutrients and the availability of nutrients for plantuptake. Journal of Tropical Ecology 21, 99–102. doi:10.1017/S0266467404001701
Fleming, T. H. (1993). Plant-visiting bats. American Scientist 81,460–467.
Garnett, S., Whybird, O., and Spencer, H. (1999). The conservationstatus of the spectacled flying fox Pteropus conspicillatus inAustralia. Australian Zoologist 31, 38–54.
Hall, L., and Richards, G. (2000). ‘Flying Foxes, Fruit and BlossomBats of Australia.’ (UNSW Press: Sydney.)
Hyland, B. P. M., Whiffin, T., Christophel, D. C., Gray, B., andElick, R. W. (2002). Key to Australian Tropical Rainforest Plants:Trees, Shrubs and Vines. (CD-ROM) (CSIRO Publishing:Melbourne.)
Law, B. S. (1992). Physiological factors affecting pollen use byQueensland blossom bats (Syconycteris australis). FunctionalEcology 6, 257–264. doi:10.2307/2389515
Law, B. S. (2001). The diet of the common blossom bat (Syconycterisaustralis) in upland tropical rainforest and the importance of ripar-ian areas. Wildlife Research 28, 619–626. doi:10.1071/WR00058
Markus, N., and Hall, L. (2004). Foraging behaviour of the black flying-fox (Pteropus alecto) in the urban landscape of Brisbane,Queensland. Wildlife Research 31, 345–355. doi:10.1071/WR01117
Marshall, A. G. (1985). Old World phytophagous bats(Megachiroptera) and their food plants: a survey. ZoologicalJournal of the Linnean Society 83, 351–369.
McDonald-Madden, E., Schreiber, G. S. G., Forsyth, D. M., Choquenot,D., and Clancy, T. F. (2005). Factors affecting grey-headed flyingfox (Pteropus poliocephalus: Pteropodidae) foraging in theMelbourne meropolitan area, Australia. Austral Ecology 30,600–608. doi:10.1111/j.1442-9993.2005.01492.x
Nelson, J. E. (1965). Movements of Australian flying foxes(Pteropodidae: Megachiroptera). Australian Journal of Zoology 13,53–73. doi:10.1071/ZO9650053
Nelson, J. E., Miller, M. A., Heske, E. J., and Fahey, G. C., Jr (2000).Nutritional consequences of a change in diet from native to agricul-tural fruits for the Samoan fruit bat. Ecography 23, 393–401.doi:10.1034/j.1600-0587.2000.230402.x
Nyhagen, D. F., Turnbull, S. D., Olesen, J. M., and Jones, C. G. (2005).An investigation into the role of the Mauritian flying fox, Pteropus
niger, in forest regeneration. Biological Conservation 122,491–497. doi:10.1016/j.biocon.2004.08.012
Palmer, C., Price, O., and Bach, C. (2000). Foraging ecology of theblack flying fox (Pteropus alecto) in the seasonal tropics of theNorthern Territory. Australian Wildlife Research 27, 169–178.doi:10.1071/WR97126
Parry-Jones, K., and Augee, M. L. (1991). Food selection by grey-headed flying foxes (Pteropus poliocephalus) occupying a summercolony site near Gosford, New South Wales. Wildlife Research 18,111–124. doi:10.1071/WR9910111
Parry-Jones, K. A., and Augee, M. L. (1992). Movements of Grey-headed flying foxes (Pteropus poliocephalus) to and from a colonysite on the central coast of New South Wales. Wildlife Research 19,331–340. doi:10.1071/WR9920331
Parry-Jones, K. A., and Augee, M. L. (2001). Factors affecting the occu-pation of a colony site in Sydney, New South Wales by the grey-headed flying fox Pteropus poliocephalus (Pteropodidae). AustralEcology 26, 47–55. doi:10.1046/j.1442-9993.2001.01072.x
Pike, K. M. (1955). Pollen morphology of Myrtaceae from the south-west Pacific area. Australian Journal of Botany 4, 13–53.doi:10.1071/BT9560013
Quinn, G. P., and Keough, M. J. (2002). ‘Experimental Design and DataAnalysis for Biologists.’ (Cambridge University Press: Cambridge.)
Richards, G. C. (1990a). The spectacled flying fox, Pteropus conspicil-latus (Chiroptera: Pteropodidae), in North Queensland 1. Roostsites and distribution patterns. Australian Mammalogy 13, 17–24.
Richards, G. C. (1990b). The spectacled flying fox, Pteropus conspicil-latus (Chiroptera:Pteropodidae), in North Queensland. 2. Diet, seeddispersal and feeding ecology. Australian Mammalogy 13, 25–31.
Richards, G. C. (1995). Review of ecological interactions of fruit batsin Australian ecosystems. In ‘Ecology, Evolution and Behaviour ofBats’. (Eds P. A. Racey and S. M. Swift.) pp. 79–96. (OxfordScientific Publications: London.)
Sedgley, M., Sierp, M., Wallwork, M. A., Fuss, A. M., and Thiele, K.(1993). Pollen presenter and pollen morphology of Banksia L.f.(Proteaceae). Australian Journal of Botany 41, 439–464.doi:10.1071/BT9930439
Shanahan, M., So, S., Compton, S. G., and Corlett, R. (2001). Fig-eating by vertebrate frugivores: a global review. Biological Reviews76, 529–572.
Shilton, L. A. (1999). Seed dispersal by fruit bats on the KrakatauIslands, Indonesia. PhD thesis, University of Leeds.
Shilton, L. A., Altringham, J. D., Compton, S. G., and Whittaker, R. J.(1999). Old world fruit bats can be long-distance seed dispersersthrough extended retention of viable seeds in the gut. Proceedingsof the Royal Society of London. Series B. Biological Sciences 266,219–223. doi:10.1098/rspb.1999.0625
Singaravelan, N., and Marimuthu, G. (2004). Nectar feeding and pollencarrying from Cieba pentandra by Pteropodid bats. Journal ofMammalogy 85, 1–7. doi:10.1644/1545-1542(2004)085<0001:NFAPCF>2.0.CO;2
Stanton, J. P., and Stanton, D. J. (2005). The Vegetation of the WetTropics of Queensland Bioregion [digital data], version 1.2. (WetTropics Management Authority: Cairns.)
Thomas, D. W. (1988). Analysis of the diets of plant-visiting bats. In‘Ecological and Behavioural Methods for the Study of Bats’. (Ed.T. H. Kunz.) pp. 211–219. (Smithsonian Institution Press:Washington, DC.)
Tidemann, C. R., Kelson, S. L., and Jamieson, G. I. (1997). Flying-foxdamage to orchard fruit in Australia: incidence, extent and eco-nomic impact. Australian Biologist 10, 177–184.
Tracey, J. G. (1982). ‘The Vegetation of the Humid Tropical Region ofNorth Queensland.’ (CSIRO: Melbourne.)
Dietary variation in Pteropus conspicillatus

J. Parsons et al.428 Australian Journal of Zoology
Waser, N. M., Chittka, L., Price, M. V., Williams, N. M., and Ollerton, J.(1996). Generalization in pollination systems and why it matters.Ecology 77, 1043–1060. doi:10.2307/2265575
Westcott, D. A., Dennis, A. J., McKeown, A., Bradford, M., andMargules, C. A. (2001). ‘The Spectacled Flying Fox, Pteropus con-spicillatus, in the Context of the World Heritage Values of the WetTropics World Heritage Area.’ (CSIRO: Atherton.)
Williams, G., and Adam, P. (1994). A review of rainforest pollinationand plant-pollinator interactions with particular reference toAustralian subtropical rainforests. Australian Zoologist 29,177–212.
Woinarski, J. C. Z., Whitehead, P. J., Bowman, D. M. J. S., and Russell-Smith, J. (1992). Conservation of mobile species in a variableenvironment: the problem of reserve design in the NorthernTerritory, Australia. Global Ecology and Biogeography Letters 2,1–10. doi:10.2307/2997325
Manuscript received 3 November 2006, accepted 10 November 2006
http://www.publish.csiro.au/journals/ajz




















