
Developmental and evolutionary basis for drought tolerance of the Anopheles gambiae embryo
9
Genomes & Developmental Control Developmental and evolutionary basis for drought tolerance of the Anopheles gambiae embryo Yury Goltsev a, 1 , Gustavo L. Rezende a,b, 1 , Karen Vranizan a , Greg Lanzaro c , Denise Valle b, ⁎, Michael Levine a, ⁎ a Department of Mol. Cell Biology, Division of Genetics Genomics and Development, Center for Integrative Genomics, University of California, Berkeley, CA 94720, USA b Laboratório de Fisiologia e Controle de Artrópodes Vetores, Instituto Oswaldo Cruz, Fiocruz, Rio de Janeiro, Brazil c Department of Veterinary Medicine, University of California, Davis, CA 94720, USA abstract article info Article history: Received for publication 18 November 2008 Revised 19 February 2009 Accepted 23 February 2009 Available online 17 March 2009 Keywords: Mosquito Anopheles gambiae Embryo Extraembryonic membranes Serosa Serosal cuticle Evolution Microarray Gene regulatory network Drought tolerance During the evolution of the Diptera there is a dramatic modification of the embryonic ectoderm, whereby mosquitoes contain separate amnion and serosa lineages while higher flies such as Drosophila melanogaster contain a single amnioserosa. Whole-genome transcriptome assays were performed with isolated serosa from Anopheles gambiae embryos. These assays identified a large number of genes implicated in the production of the larval cuticle. In D. melanogaster, these genes are activated just once during embryogenesis, during late stages where they are used for the production of the larval cuticle. Evidence is presented that the serosal cells secrete a dedicated serosal cuticle, which protects A. gambiae embryos from desiccation. Detailed temporal microarray assays of mosquito gene expression profiles revealed that the cuticular genes display biphasic expression during A. gambiae embryogenesis, first in the serosa of early embryos and then again during late stages as seen in D. melanogaster. We discuss how evolutionary modifications in the well- defined dorsal–ventral patterning network led to the wholesale deployment of the cuticle biosynthesis pathway in early embryos of A. gambiae. © 2009 Elsevier Inc. All rights reserved. Introduction The serosa and amnion are extraembryonic membranes (EM) produced during early embryogenesis in most insects (Panfilio, 2008). The serosa is a protective membrane that can enclose the entire embryo. In most cases the amnion and serosa are destroyed before the completion of embryogenesis and do not contribute to a definitive larval organ (Schmidt-Ott, 2005). The developmental origins of the amnion and serosa are used as a phylogenetic trait (Cobben, 1968), and there is considerable informa- tion about the evolutionary origins of these tissues in primitive insects and non-insect hexapods (Machida, 2006). Most taxa, including both hemimetabolous and holometabolous insects, manifest distinct amnion and serosa tissues (Dorn, 1976; Panfilio, 2008). However, higher Dipteran lineages radiating within the Cyclorrhapha taxon, specifically the Schizophora (Rafiqi et al., 2008) as represented by the best-studied insect, Drosophila melanogaster , contain a single extra- embryonic membrane, the amnioserosa, in place of separate amnion and serosa membranes (Demerec, 1950; Panfilio, 2008; Schmidt-Ott, 2000). Here, we investigate the evolutionary and physiological basis for this reduction in complexity. In most insects the amnion and serosa are tightly linked to blastokinesis — a process of spatial reorientation of the embryo during development (Cobben, 1968; Panfilio, 2008). These tissues have also been adapted for a variety of functions in different insects. For example, the amnioserosa of D. melanogaster is required for germband elongation, retraction (Lamka and Lipshitz, 1999), and dorsal closure (Scuderi and Letsou, 2005). In some species the serosa is involved in water retention, and is primarily responsible for the elaboration of an extracellular cuticle, the serosal cuticle (Hinton, 1981), which is intimately associated with the inner side of the maternally provided chorion. Recently, we have shown that the secretion of the serosal cuticle coincides with the acquisition of drought tolerance in the mosquito Aedes aegypti, the main dengue vector (Rezende et al., 2008). The multilayer structure of the serosal cuticle varies dramatically between species (Hinton, 1981), but is generally composed of two types: an external (so-called yellow cuticle in crickets) hydrophobic epicuticle that was hypothesized to be enriched in lipids (Beament, 1946a,b, 1949; Beckel, 1958; Harwood and Horsfall, 1959; McFarlane, 1960) and an internal multilayer endocuticle containing chitin (so- called white cuticle) (Campbell, 1929; Jahn, 1935; Slifer, 1937b). The Developmental Biology 330 (2009) 462–470 ⁎ Corresponding authors. E-mail addresses: dvalle@ioc.fiocruz.br (D. Valle), [email protected] (M. Levine). 1 Authors contributed equally. 0012-1606/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2009.02.038 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Developmental and evolutionary basis for drought tolerance of the Anopheles gambiae embryo

Developmental Biology 330 (2009) 462–470
Contents lists available at ScienceDirect
Developmental Biology
j ourna l homepage: www.e lsev ie r.com/deve lopmenta lb io logy
Genomes & Developmental Control
Developmental and evolutionary basis for drought tolerance of theAnopheles gambiae embryo
Yury Goltsev a,1, Gustavo L. Rezende a,b,1, Karen Vranizan a, Greg Lanzaro c, Denise Valle b,⁎, Michael Levine a,⁎a Department of Mol. Cell Biology, Division of Genetics Genomics and Development, Center for Integrative Genomics, University of California, Berkeley, CA 94720, USAb Laboratório de Fisiologia e Controle de Artrópodes Vetores, Instituto Oswaldo Cruz, Fiocruz, Rio de Janeiro, Brazilc Department of Veterinary Medicine, University of California, Davis, CA 94720, USA
⁎ Corresponding authors.E-mail addresses: [email protected] (D. Valle), m
(M. Levine).1 Authors contributed equally.
0012-1606/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.ydbio.2009.02.038
a b s t r a c t
a r t i c l e i n f o© 2009 Elsevier Inc. All rights reserved.
Article history:Received for publication 18 November 2008Revised 19 February 2009Accepted 23 February 2009Available online 17 March 2009
Keywords:MosquitoAnopheles gambiaeEmbryoExtraembryonic membranesSerosaSerosal cuticleEvolutionMicroarrayGene regulatory networkDrought tolerance
During the evolution of the Diptera there is a dramatic modification of the embryonic ectoderm, wherebymosquitoes contain separate amnion and serosa lineages while higher flies such as Drosophila melanogastercontain a single amnioserosa. Whole-genome transcriptome assays were performed with isolated serosafrom Anopheles gambiae embryos. These assays identified a large number of genes implicated in theproduction of the larval cuticle. In D. melanogaster, these genes are activated just once during embryogenesis,during late stages where they are used for the production of the larval cuticle. Evidence is presented that theserosal cells secrete a dedicated serosal cuticle, which protects A. gambiae embryos from desiccation.Detailed temporal microarray assays of mosquito gene expression profiles revealed that the cuticular genesdisplay biphasic expression during A. gambiae embryogenesis, first in the serosa of early embryos and thenagain during late stages as seen in D. melanogaster. We discuss how evolutionary modifications in the well-defined dorsal–ventral patterning network led to the wholesale deployment of the cuticle biosynthesispathway in early embryos of A. gambiae.
Introduction
The serosa and amnion are extraembryonic membranes (EM)produced during early embryogenesis in most insects (Panfilio, 2008).The serosa is a protective membrane that can enclose the entireembryo. In most cases the amnion and serosa are destroyed before thecompletion of embryogenesis and do not contribute to a definitivelarval organ (Schmidt-Ott, 2005).
The developmental origins of the amnion and serosa are used as aphylogenetic trait (Cobben, 1968), and there is considerable informa-tion about the evolutionary origins of these tissues in primitive insectsand non-insect hexapods (Machida, 2006). Most taxa, including bothhemimetabolous and holometabolous insects, manifest distinctamnion and serosa tissues (Dorn, 1976; Panfilio, 2008). However,higher Dipteran lineages radiating within the Cyclorrhapha taxon,specifically the Schizophora (Rafiqi et al., 2008) as represented by thebest-studied insect, Drosophila melanogaster, contain a single extra-embryonic membrane, the amnioserosa, in place of separate amnion
l rights reserved.
and serosa membranes (Demerec, 1950; Panfilio, 2008; Schmidt-Ott,2000). Here, we investigate the evolutionary and physiological basisfor this reduction in complexity.
In most insects the amnion and serosa are tightly linked toblastokinesis— a process of spatial reorientation of the embryo duringdevelopment (Cobben, 1968; Panfilio, 2008). These tissues have alsobeen adapted for avariety of functions in different insects. For example,the amnioserosa of D. melanogaster is required for germbandelongation, retraction (Lamka and Lipshitz, 1999), and dorsal closure(Scuderi and Letsou, 2005). In some species the serosa is involved inwater retention, and is primarily responsible for the elaboration of anextracellular cuticle, the serosal cuticle (Hinton, 1981), which isintimately associated with the inner side of the maternally providedchorion. Recently, we have shown that the secretion of the serosalcuticle coincides with the acquisition of drought tolerance in themosquitoAedes aegypti, themain dengue vector (Rezende et al., 2008).The multilayer structure of the serosal cuticle varies dramaticallybetween species (Hinton, 1981), but is generally composed of twotypes: an external (so-called yellow cuticle in crickets) hydrophobicepicuticle that was hypothesized to be enriched in lipids (Beament,1946a,b, 1949; Beckel, 1958; Harwood and Horsfall, 1959; McFarlane,1960) and an internal multilayer endocuticle containing chitin (so-called white cuticle) (Campbell, 1929; Jahn, 1935; Slifer, 1937b). The
463Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
outer layer contains tyrosinase and manifests dopamine-basedsclerotization (Furneaux and McFarlane, 1965a,b; McFarlane, 1960).
We previously examined the genetic changes underlying thetransition from two EMs in the lower Diptera as represented byAnopheles gambiae to one in the higher flies as in D. melanogaster(Goltsev et al., 2004, 2007). The expansion of the dorsal ectoderm inA. gambiae (as compared with D. melanogaster) embryos is due, atleast in part, to changes in the regulation of Sog expression, while theseparation of dorsal cell lineages depends on the localized expressionof specialized repressors. In this study we investigate the generegulatory networks governing the function of the serosa in A.gambiae. Transcriptome profiling of the A. gambiae serosa, along withmicroarray analysis of staged embryos, suggests that many compo-nents of the larval cuticle gene pathway display biphasic expressionduring A. gambiae embryogenesis. In contrast, there is just a singlephase of expression in D. melanogaster, during late stages ofembryogenesis. The expression of the cuticle genes in the A. gambiaeserosa is coincident with the timing of embryonic resistance todesiccation. We propose that the failure to deploy the cuticle genebattery in the D. melanogaster amnioserosa is due to subtle changes inthe dorsal–ventral patterning network.
Results
Serosal cuticle formation coincides with desiccation resistance
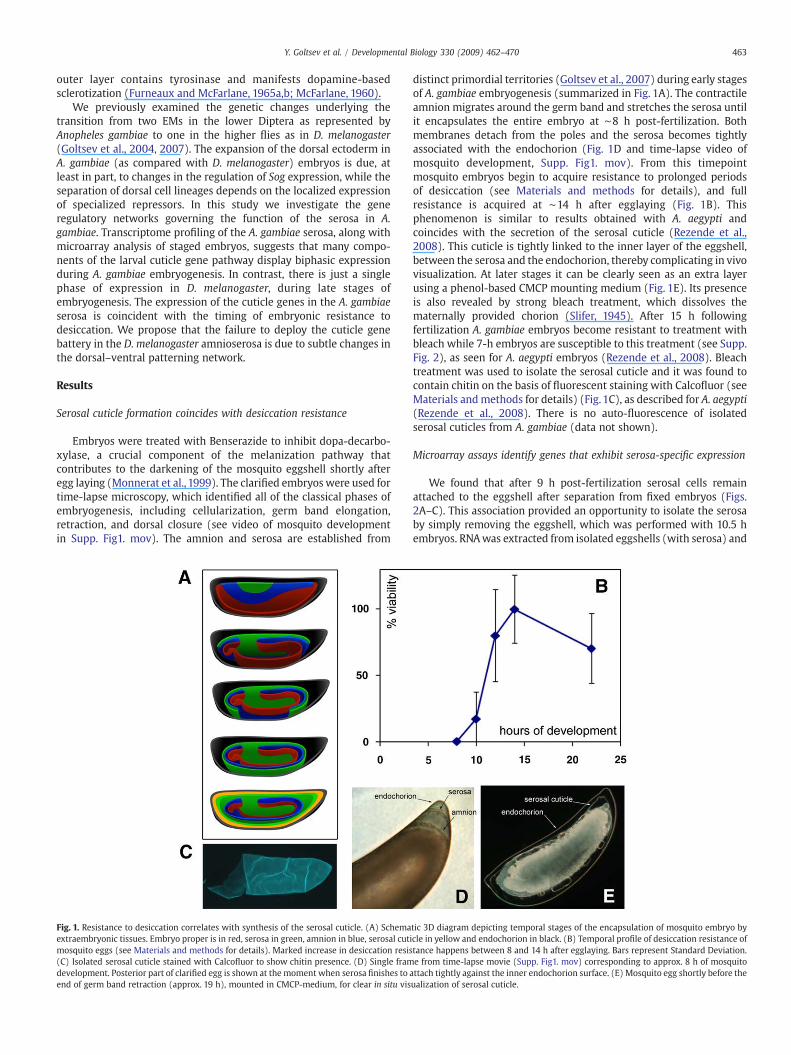
Embryos were treated with Benserazide to inhibit dopa-decarbo-xylase, a crucial component of the melanization pathway thatcontributes to the darkening of the mosquito eggshell shortly afteregg laying (Monnerat et al., 1999). The clarified embryos were used fortime-lapse microscopy, which identified all of the classical phases ofembryogenesis, including cellularization, germ band elongation,retraction, and dorsal closure (see video of mosquito developmentin Supp. Fig1. mov). The amnion and serosa are established from
Fig. 1. Resistance to desiccation correlates with synthesis of the serosal cuticle. (A) Schemaextraembryonic tissues. Embryo proper is in red, serosa in green, amnion in blue, serosal cutimosquito eggs (see Materials and methods for details). Marked increase in desiccation resis(C) Isolated serosal cuticle stained with Calcofluor to show chitin presence. (D) Single framdevelopment. Posterior part of clarified egg is shown at the moment when serosa finishes to aend of germ band retraction (approx. 19 h), mounted in CMCP-medium, for clear in situ vis
distinct primordial territories (Goltsev et al., 2007) during early stagesof A. gambiae embryogenesis (summarized in Fig. 1A). The contractileamnion migrates around the germ band and stretches the serosa untilit encapsulates the entire embryo at ∼8 h post-fertilization. Bothmembranes detach from the poles and the serosa becomes tightlyassociated with the endochorion (Fig. 1D and time-lapse video ofmosquito development, Supp. Fig1. mov). From this timepointmosquito embryos begin to acquire resistance to prolonged periodsof desiccation (see Materials and methods for details), and fullresistance is acquired at ∼14 h after egglaying (Fig. 1B). Thisphenomenon is similar to results obtained with A. aegypti andcoincides with the secretion of the serosal cuticle (Rezende et al.,2008). This cuticle is tightly linked to the inner layer of the eggshell,between the serosa and the endochorion, thereby complicating in vivovisualization. At later stages it can be clearly seen as an extra layerusing a phenol-based CMCP mounting medium (Fig. 1E). Its presenceis also revealed by strong bleach treatment, which dissolves thematernally provided chorion (Slifer, 1945). After 15 h followingfertilization A. gambiae embryos become resistant to treatment withbleach while 7-h embryos are susceptible to this treatment (see Supp.Fig. 2), as seen for A. aegypti embryos (Rezende et al., 2008). Bleachtreatment was used to isolate the serosal cuticle and it was found tocontain chitin on the basis of fluorescent staining with Calcofluor (seeMaterials andmethods for details) (Fig. 1C), as described for A. aegypti(Rezende et al., 2008). There is no auto-fluorescence of isolatedserosal cuticles from A. gambiae (data not shown).
Microarray assays identify genes that exhibit serosa-specific expression
We found that after 9 h post-fertilization serosal cells remainattached to the eggshell after separation from fixed embryos (Figs.2A–C). This association provided an opportunity to isolate the serosaby simply removing the eggshell, which was performed with 10.5 hembryos. RNAwas extracted from isolated eggshells (with serosa) and
tic 3D diagram depicting temporal stages of the encapsulation of mosquito embryo bycle in yellow and endochorion in black. (B) Temporal profile of desiccation resistance oftance happens between 8 and 14 h after egglaying. Bars represent Standard Deviation.e from time-lapse movie (Supp. Fig1. mov) corresponding to approx. 8 h of mosquitottach tightly against the inner endochorion surface. (E) Mosquito egg shortly before theualization of serosal cuticle.

464 Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
dechorionated embryos. The efficacy of the eggshell extractionmethod was tested by quantitative PCR assays: Chitin Synthase 1transcripts, used as an internal control of this procedure (Rezendeet al., 2008), were readily detected in the eggshell RNA extract, butonly background levels were seen in dechorionated embryos (Fig. 2D).
Microarray assays identified 538 probe sets representing 359 genesthat are significantly upregulated in the eggshell fraction as comparedwith dechorionated embryos (Fig. 2E, Supp. Table 2).While these genesrepresent a variety of processes (see GO-terms enrichment analysis inSupp. Table 1 and manually selected groups in Supp. Table 3), manycluster within biosynthetic pathways controlling chitin synthesis anddeposition, tyrosinemetabolism and catecholamine synthesis, and longchain fatty acid (presumably — wax) synthesis. All of these pathwayshave been implicated in the elaboration of the larval cuticle in D.melanogaster (Payre, 2004;Wigglesworth,1948). Accordingly it appearsthat theyare alsoused for the production of the chitinizedwhite layer, aswell as the tyrosine-mediated cross-linking and synthesis of the yellowwaxy layer of the serosal cuticle (Slifer, 1937a) (see in Discussion).
Representative genes from each class were used for in situhybridization assays in A. gambiae embryos (Fig. 3). All of the genesthat were tested are activated either before or during the deposition ofthe serosal cuticle and display selective expression in the serosa. Someof the genes are expressed in the presumptive serosa prior to itsspreading around the embryo (Figs. 3A, G, M), while others areexpressed during (Fig. 3K) or after (Fig. 3F) spreading.
Not all of the genes expected to participate in insect cuticlesynthesis were detected by the microarray assays. The mosquitoorthologs of Ddc (dopa-decarboxylase), pale (tyrosine hydroxylase),and kkv (chitin synthase 1) (AGAP009091, AGAP006023, andAGAP001748 respectively) fell below the cutoff. Nonetheless in situ
Fig. 2. Isolation of serosa cells attached to eggshells for microarray analysis. (A) Fixed 10.5(B) Same field as in A but after embryo extraction from shell. White embryo is seen in the boreveal monolayer of serosal cells attached to the inner eggshell surface after embryo extractiofixed embryos with shells and extracted embryos. Chitin Synthase 1 is augmented in shellplotted against log2 (Serosa Enrichment Fold) for all mosquito genes. Color coding is used towhich was subjected to GO-terms overrepresentation analysis (Supp.Table1) to reveal pathwstatistically significant genes upregulated in serosa.
hybridization indicates that all three genes are indeed expressed in theserosa, although the genes are activated at somewhat divergent stages.Chitin synthase 1 is expressed in themature serosa and can be detectedby in situ hybridization only in isolated eggshells (Fig. 3F). Apparentlythe failure to identify chitin synthase 1 is due to a flaw in the design ofthe specific probe sets on the A. gambiaemicroarray (data not shown).The failure to identify dopa-decarboxylase and tyrosine hydroxylasemay be due to their early onset of expression (Figs. 3G, M), prior to thestage when the serosa can be isolated by removal of eggshells.
In addition to the aforementioned biosynthetic pathways we alsoobserved upregulation of several potential estrogen dehydrogenases —members of a family of enzymes (Gilbert, 2004) involved in thematuration of steroid hormones (Supp. Table 3, part2). This observationis consistent with studies suggesting production of ecdysone-likesteroids by extraembryonic tissues (Kozlova and Thummel, 2003).Genes involved in programmed cell death (Supp. Table 3, part1) are alsoexpressed in the serosa, probably foreshadowing its destruction ∼28 hafter fertilization (as inferred from time-lapse movies). A number ofserine peptidases and serine protease inhibitors are also upregulated inthe serosa (Supp. Table 3, part2). These genes have been implicated ininnate immunity by inducing an enzymatic cascade leading to nuclearimport of rel− family transcription factors (Reichhart, 2005). Finally, alarge number of enzymes potentially involved in detoxification such ascytochromes, GSTs and active radical scavenging (superoxide dismu-tases) are also expressed in the serosa (Supp. Table 3, part2).
Serosal genes show biphasic profiles during mosquito development
The identification of cuticle gene transcripts in the serosa raises thepossibility that these genes might display biphasic expression
h embryo inside eggshell on the surface of sticky tape covered slide before extraction.ttom right of the image. (C) An eggshell prepared similarly to B was stained with DAPI ton. (D) Presence of An. Chitin Synthase 1 examined by qPCR on RNA from separate shells,preparation. (E) Volcano plot. The negative log10-transformed moderated p-values arehighlight the gene matching probesets with log2 fold change above 1 (two-fold change)ays augmented in serosa. Red line indicates moderated p-value threshold used to select

Fig. 3. Embryonic expression of serosal genes. Colorimetric (A, B, D, E, G, H, J, K, M, N) in situ hybridizations were performedwith 6.5 h pre-gastrulation (A, D, G, J, M) and 7.5 h (B, E, H,K, N) mosquito embryos. Fluorescent (C, F, I, L, O) in situ hybridizations were performed with eggshells mounted on slides and anti-sense dig-labeled RNA probes for the followinggenes were used (A–C) AGAP011812 (ELOVL, elongase) presumably involved inwax synthesis (Jakobsson et al., 2006). (D–F) AGAP001748 (Chitin Synthase 1, ortholog of kkv) criticaleffector of cuticular chitin production (Moussian et al., 2005). (G–I) AGAP006023 (tyrosine hydroxylase, pale) enzyme involved in dopamine-based cuticle pigmentation/Sclerotization (Neckameyer and Quinn, 1989). (J–L) AGAP011936 (Serpentine) chitin-binding protein involved in regulating chitin deposition (Luschnig et al., 2006). (M–O)AGAP009091 (dopa-decarboxylase, Ddc) enzyme involved in dopamine-based cuticle pigmentation/Sclerotization (Wright, 1996).
465Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
patterns during A. gambiae embryogenesis. Microarray assays wereperformed on staged A. gambiae embryos, from fertilization to 46 h ofdevelopment (which is close to hatching at ∼50 h post-fertilization).RNA was extracted from staged embryos roughly every 3 h afterfertilization, and then hybridized to the A. gambiae transcriptomemicroarray. Putative serosal genes were selected from the datasetbased on the previous analysis of isolated serosa (see Supp. Table 2).We found that the temporal profiles of mosquito serosal genesgenerally manifest two peaks — early expression coincides with thesynthesis of the serosal cuticle, while late expression probably reflectsformation of the larval cuticle during terminal phases of embryo-genesis (Figs. 4B–D). The biphasic mode of cuticle gene expressionwas validated by qPCR assays on selected genes (Fig. 4E). Orthologuesof A. gambiae serosal genes were identified in previously publishedD. melanogaster microarray assays. Approximately 200 gene pairswere compared in A. gambiae (Fig. 4A top) and D. melanogaster (Fig.4A bottom). Strikingly, most of the genes implicated in the formationof the A. gambiae serosal cuticle are expressed only during advancedperiods of D. melanogaster embryogenesis, while most of the genesdisplay biphasic expression in A. gambiae.
Thus, larval cuticle genes are deployed in the early A. gambiaeembryo to form the serosal cuticle.D.melanogaster lacks a serosa, has noassociated serosal cuticle, and the genes responsible for the formationofthe larval cuticle are activated only during late embryogenesis.
Discussion
Cuticle gene battery
Sclerotization, wax and chitin deposition are used at differentstages of the insect life cycle for the synthesis of serosal (Campbell,
1929; Jahn, 1935; McFarlane, 1960; Slifer, 1937b), larval (Moussianet al., 2006), pupal and adult cuticles (Wigglesworth, 1948).Sclerotization and chitin are also used for the production of theperitrophic matrix (Shao et al., 2001), tracheal lumen (Araujo et al.,2005), and wound healing (Mace et al., 2005). It is not knownwhether the same gene batteries are used for all of these processes, orif specialized versions of the cuticle genes are used for differentpurposes. The tight association of the A. gambiae serosa with theeggshell provided an opportunity to isolate a homogenous tissue atthe time when the serosal cuticle is synthesized. A large number ofgenes encoding components of the larval cuticle were found to exhibitan early wave of expression in the developing serosa. These resultsstrongly suggest that the same gene battery is used for the synthesis ofthe serosal and larval cuticles.
The absence of the serosa is a phylogenetic hallmark of higherDiptera (Rafiqi et al., 2008; Schmidt-Ott, 2000). Basal insects such ascrickets (Slifer, 1937b) and beetles (van der Zee et al., 2005) containserosal tissues similar to those seen in mosquitoes. Thus, during theevolution of the Diptera there is a transition in the expression of thecuticle gene battery from a biphasic mode (serosa and larvalepidermis) to the singular mode (larval only) characteristic of theD. melanogaster embryo. In principle, a limited number of mutations inthe DV gene network are sufficient to account for the loss of the serosaand serosal cuticle, as we discuss below.
Evolution of serosa transcription networks
What is the basis for the selective activation of the cuticle genes inthe A. gambiae serosa and the loss of this activation inD. melanogaster?The microarray assays using isolated serosa identified a number ofregulatory genes (Supp. Table 2) that may be lost in the amnioserosa

466 Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470

467Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
of D. melanogaster. Included in this list are: AGAP005311 — Anophelesortholog of Drosophila homeobrain, AGAP010623 — Xbox bindingfactor, and AGAP005137— a lim homeobox gene. Transcription factorsgenerally belong to relatively low abundance molecules and aretherefore underrepresented in a population of upregulated mRNAs.To compensate for this we used a lower cut-off value whencomparing serosa vs. embryo transcripts for putative regulatorygenes (0.65 rather than 1.0) and identified additional genesincluding AGAP011625 — mosquito AP-2, AGAP009515 — mosquitoDorsal, and AGAP007327 — a major transcriptional effector ofecdysone signaling Eip74 (Fletcher and Thummel, 1995). In situhybridization assays confirmed that the lim homeobox gene, ho-meobrain and Dorsal are indeed expressed in the developing serosaof the early A. gambiae embryo (Fig. 5). Dorsal is also expressed inthe T. castaneum serosa (da Fonseca et al., 2008). None of thesegenes are expressed in the amnioserosa of the D. melanogasterembryo (BDGP fly in situ database — data not shown). Previousstudies demonstrated that ecdysone signaling (Chavez et al., 2000) iscritically involved in the regulation of cuticle synthesis both at larvaland late stages of Drosophila development. Moreover, cuticle genessuch as Ddc and pale (Mace et al., 2005) are induced duringwounding of the D. melanogaster epidermis. The characterization ofminimal cis-regulatory sequences identified grainyhead (grh) as akey activator of Ddc (Mace et al., 2005). Grh is known to be a criticalregulator of proper integument function in D. melanogster, C. elegansand mice (Jane et al., 2005). We found that grh exhibits selectiveexpression in the serosa anlagen of A. gambiae embryos (Fig. 5C), butit is also expressed in the dorsal ectoderm of early D. melanogasterembryos (Huang et al., 1995), so it is not sufficient to account for thedeployment of the cuticle gene battery in A. gambiae. Interestingly,the analysis of the D. melanogaster Ddc enhancer raised thepossibility that additional regulatory factors such as rel− familygenes (exemplified by dorsal and its orthologues in insects) mightparticipate in its activation (Mace et al., 2005).
Many insects (Dorn and Romer,1976; Imboden and Lanzrein,1982;Lagueux et al., 1979; Scalia and Morgan, 1982) including D. melano-gaster (Kozlova and Thummel, 2003) manifest a mid-embryonic peakof ecdysone production, yet Eip74EF is not expressed in the Drosophilaamnioserosa (BDGP in situ database, data not shown). Perhaps Eip74EFis needed to activate at least some of the serosal cuticle genes inresponse to ecdysone. It is possible that a number of mosquito-specificserosal transcription factors such as hunchback, tramtrack, ems(Goltsev et al., 2007), dorsal, AP-2, and the lim homeobox geneAGAP005137 facilitate serosa-specific Eip74EF induction by EcR(ecdysone receptor). dorsal might act synergistically with grh toactivate genes such as Ddc and pale that are independent of ecdysonesignaling. Thus, the expression of just a few serosa-specific transcrip-tion factors, possibly under the control of the homeobox gene zen(Goltsev et al., 2007), might be sufficient to account for thedeployment of the cuticle gene battery in the A. gambiae embryo.
Role of serosa in drought resistance
One of the major factors determining the occurrence of malaria insub-Saharan Africa is the capacity of A. gambiae to actively maintainmalaria transmission in the driest areas and months of the year. This
Fig. 4. Transition from biphasic (in A. gambiae) to singular in (D. melanogaster) expressionenriched in serosa (top), compared to the developmental expression profiles of D. melanogastet al., 2007). Each row is a gene, and each column is a time point (indicated in grey along threpression (green) for each gene. Black means no change of expression compared to averamatched to Drosophila orthologues are shown) are sorted according to enrichment in serosaprofiles of selected genes (picked based onmanual analysis of annotations and public databasTyrosine based crosslinking and (D) Wax synthesis. (E) Expression profiles for selected mosqthe profile of AGAP001748 was obtained by qPCR (see Materials and methods for details). Reof the Mean. Drosophila profiles on B, C and D represent expression values from data colle(Serpentine), and (Hooper et al., 2007) for the rest. The corresponding mosquito profiles redata from this study.
drought tolerance is a major adaptation contributing to the apparentcontinuing expansion in the range of A. gambiae (Beier et al., 1990;Onyabe et al., 2003; Toure et al., 1994). So far two major strategies forsurvival during the dry seasons have been suggested: reproduction inlocalized moist niches and aridity tolerance of adults (Gray andBradley, 2004, 2005), larvae and eggs (Beier et al., 1990; Minakawaet al., 2001; Shililu et al., 2004). There are several factors that mightinfluence embryonic drought resistance, including: the structure ofthe eggshell, the capacity of the early embryo to retain water, and theability of older embryos to undergo diapause.
The present study provides insights into the mechanisms ofembryonic drought resistance. It has been proposed that crosslinkingand melanization of the mosquito eggshell are the major mechanismsfor protecting the embryo against desiccation (Li and Li, 2006).However, we believe that it is unlikely that these reactions contributesignificantly to drought resistance since the early embryo is highlysensitive to desiccation (Fig. 1B). Resistance is not acquired until 14 hafter fertilization, during the time when the serosal cuticle is formed.We also note that there is an early spike of aquaporin gene expression,which might contribute to the ability of the early embryo to retainwater (Y.G. unpublished observations). We propose that the serosalcuticle produced by the A. gambiae serosa is primarily responsible forthe desiccation resistance of the early embryo. Since the same genebattery is used for the serosal and larval cuticle it is possible thatmutations affecting embryonic drought tolerance will also impair theresistance of adultmosquitoes. The identity of serosal genes uncoveredin our study provides a foundation for population control through theproduction of transgenic mosquitoes with compromised droughttolerance.
Materials and methods
Mosquito stocks and egg collection
A. gambiae population was reared at 27 °C, 75% humidity, with a12-h light/dark cycle. Adults were maintained on a 10% sucrosesolution and females were blood-fed on anesthetized hamsters. Forsynchronized embryo collection the females were placed in the darkat 27 °C for 1 h inside 15 cm Petri dish lined up with circles of wetWhatman paper. The developmental time was counted starting fromthe moment Whatman paper was moisturized with water. Mosquitoembryo fertilization happens at the moment of egglaying. Due toconstraints in experimental setup, after 2 h following the start ofcollection the eggs were shifted from 27 °C and constantly kept at25 °C. As our unpublished observations show 2 h incubation at 27 °Caugments egglaying effectiveness yet does not change the develop-mental dynamics of time course as sampled at 25 °C.
Time lapse microscopy of mosquito embryogenesis
Eggs were collected into 500 μMBenserazide (Sigma, St. Louis, MO,Catalog # B-7283) solution in water and glued with double-sidedsticky tape to the bottom of Petri dish filled with collection solution.The images were recorded with Nikon microscope fitted withsubmersible 25×objective. Due to the limit in the size of the viewfield the images of anterior most part of the embryo were not
of serosal/cuticular genes. (A) Developmental expression profiles of A. gambiae geneser orthologues of mosquito serosa genes (bottom). Fly data was extracted from (Hoopere diagrams). The intensities of the colors indicate the magnitudes of induction (red) orge expression level across the time course. Mosquito genes (only those that could be, Drosophila orthologues are sorted according to mosquito sorting. Individual expressiones seemore details in Supp. Table 3) involved in (B) Chitin synthesis and deposition, (C)uito genes generated from microarray data were independently checked by qPCR. In B,d and green lines are AgCHS1a and AgCHS1b, respectively. Bars represent Standard Errorcted by (Arbeitman et al., 2002) for CG2666 (kkv), (Manak et al., 2006) for CG32209present z-transformed (value-average(acrossprofile))/stdev(acrossprofile) microarray

Fig. 5. Expression of serosal transcription factors in pre-gastrulating embryo.Colorimetric in situ hybridizations were performed with 6.5 h pre-gastrulationmosquito embryos. Antisense dig-labeled RNA probes for the following genes wereused (A) AGAP009515 (dorsal, dl), (B) AGAP005311 (homeobrain, hbn), (C) AGAP005564(grainyhead, grh), (D) AGAP005137 (lim homeobox, CG32105).
468 Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
captured. Yet the photographed abdominal and posterior partsprovided sufficient details of the main embryonic movements aswell as the dynamics of extraembryonic tissues.
Isolation of serosa cuticles by bleach treatment and Calcofluor staining
Serosal cuticle (SC) was isolated from fully developed embryos.Complete digestion of exo and endochorion was attained with 50%household bleach (2.6% active sodiumhypochlorite) during 25–30mininside a Falcon cell strainer (70 μm Nylon, Becton Dickinson). Theremaining SCs, with larvae inside, were then thoroughly washed with25mMpH 6.2 sodium phosphate buffer. This last procedure breaks theSC, often at the line of dehiscence (Judson and Hokama, 1965). IsolatedSCs were incubated with 1 mg/ml Calcofluor (Sigma, St. Louis, MO,also named Fluorescent Brightener 28, Catalog # F-3543) during10 min in the dark, thoroughly washed with 25 mM pH 6.2 sodiumphosphate buffer and analyzed under a microscope with fluorescence,dark field or DIC. Calcofluor specificity and presence of chitin inserosal cuticle is further discussed elsewhere (Rezende et al., 2008).
Analysis of mosquito egg viability in arid conditions
Synchronized developing eggs obtained as indicated above weretransferred fromwet to dry conditions at different embryonic ages: 8,10, 12, 14 and 22 h after egglaying (HAE). In each case, groups of 100eggs were taken from moisture Whatman No. 1 filter paper andtransferred onto dry polycarbonate filters (2.5 cm diameter, 8 μmpore, Poretics Corporation). These filters were then blotted on dryWhatman No. 1 filter paper to remove all water and eggs were leftdrying by air-exposure for 20 h (e.g. 8 HAE embryos were left on dryfrom 8 until 28 HAE,10 HAE embryos were left on dry from 10 until 30HAE and so on). Following this 20 h period on dry the eggs wereplaced into a Petri dish filled with water until the end of embryogen-esis (∼50 HAE). From 50 HAE on, larvae hatching was counted every30 min, until no more hatching occurred. Viability was scored aspercentage of total hatched larvae related with the total amount ofeggs, normalized by parallel control samples kept on water through-out thewhole embryogenesis. Experiments were performed at least intriplicates inside an incubator at 25 °C with 45–60% relative humidity.
Real time quantitative PCR (qPCR)
The cDNA was synthesized with Superscript III (Invitrogen) withthe oligo-d(T)15 primer according to manufacturer instructions. Afinal concentration of 4 ng RNA/μL was employed at the final volumeof the reverse transcriptase reaction. Final concentrations above 8 ngRNA/μL inhibits the proportional synthesis of the cDNA pool (data notshown). The qPCR reactions were carried out in ABI Prism 7300(Applied Biosystems) with the QuantiTest SYBR Green Kit (QIAGEN).The following primers were used.
AGAP011812 (ELOVL, elongase):ag.elov.F2 (5′ GCTTCATTGGCGTCAAGTACTTT 3′),ag.elov.R2 (5′ TCGCGGCCAGCATGTAGT 3′),AGAP001748 (Chitin Synthase, AgCHS1). Mosquito CHS1 possessestwo splice variants (Rezende et al., 2008) and specific primerswere designed for each of the variants:chs1ExA:ag.chsExA.F1 (5′ AAGTGGCCGTTGGGTGTTAA 3′),ag.chsExA.R1 (5′ CGGTTCCAGCTGCAGATACTC 3′),chs1ExB:ag.chsExB.F2 (5′ AGACAAGCTGCACATCATTTGG 3′),ag.chsExA.R2 (5′ TCGGTTCCAGCTGCAGATACT 3′),AGAP011936 (Serpentine):ag.serp.F2 (5′ ACGTACTTCGTGTCGCACAAGT 3′),ag.serp.R2 (5′ CTCATCATCGTTGTGGGTGATAG 3′),AGAP004500 (Knickkopf):ag.knk.F4 (5′ GCGATCCGGCAGTGTTG 3′),ag.knk.R4 (5′ GCTATCGACGATGTGTATCTTCCA 3′).
The primers 5gambexpRP1 (5′ GCTATGATAAACTCGCTCCCAA 3′)and 3aeaquaRP1b (5′ TCATCAGCACCTCCAGCTC 3′) were employed toamplify the constitutive gene rp49 (AGAP002122), used as an internalnormalizer for qPCR (Gentile et al., 2005).
Identification of mosquito serosa genes and statistical analysis
10.5 h mosquito embryos were collected and fixed as describedpreviously (Goltsev et al., 2004). Following embryo dissectionprocedure, the empty eggshells remaining on double-sticky tape(Scotch3M) were washed off the tape by Xylenes and equilibrated inethanol. Total RNA was prepared from fixed eggs, dissected embryosor purified shells using RecoverALL kit (Ambion). Biotin-labeledprobes were synthesized using two-step amplification kit (Affyme-trix) and hybridized to Plasmodium–Anopheles expression arrays(Affymetrix). Three biological replicates were obtained for everyeach of the purified shells (serosa) or extracted embryo groups at10.5 h. Log base 2 expression values all probesets on the array werecomputed using RMAExpress (Bolstad et al., 2003). Log2Fold valuesfor each probeset were computed by subtracting the averageexpression value in the whole embryo group from the averageexpression value in the serosa group. A p-value testing each Log2Foldvalue against zero was obtained using a moderated two-sample t-test(Smyth, 2005). A false discovery rate (FDR) was obtained by adjustingthese p-values by the Benjamini–Hochberg method for correction formultiple hypothesis testing (Benjamini and Hochberg, 1995). Custombuilt table reassigning the probesets according to most recent Ano-pheles genome annotation was used for matching of Plasmodium–
Anopheles microarray probes to mosquito genes (Supp. Table 2).Probesets matching genes with statistically significant differentialexpression in serosa were chosen using cut-off values of Log2Foldgreater than 1 (two-fold change) plus p-valueb0.1. The volcano plot inFig. 2E highlights the 359 probesets satisfying the criteria. GenMapp2software (Salomonis et al., 2007) was used to identify GO-terms

469Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
enriched in the population of serosal genes. GoElite (http://www.genmapp.org/go_elite/go_elite.html) software was used to removeredundant nested GO-terms fromMAPPFinder results. The microarraydata can be downloaded from http://www.ncbi.nlm.nih.gov/geo/,accession number GSE14851.
Mosquito developmental microarray timecourse
Total RNA from unfixed staged eggs at sequential (2 h, 4 h, 6 h,7 h, 8 h, 10 h, 13 h, 16 h, 19 h, 22 h, 25 h, 28.5 h, 31 h, 34 h, 37 h,40 h, 43 h, and 46 h) time points was purified initially by Trizol(Invitrogen) and then further by RNAeasy (Qiagen) kit. Biotin-labeled probes were synthesized using regular labeling kit (Affyme-trix) and hybridized to Plasmodium–Anopheles expression arrays(Affymetrix). All time points were sampled with three independentbiological replicates. RMA-express software was used for processingof raw expression data and estimation of log base 2 expressionvalues (Supp. Table 4). Normalized expression levels depicted inheatmap and time profiles in Fig. 4 were obtained for each probesetby subtracting the mean and dividing by the standard error of thelog base 2 expression values of the 18 time points. The microarraydata can be downloaded from http://www.ncbi.nlm.nih.gov/geo/,accession number GSE15001.
Cross species microarray analysis
Drosophila Anopheles ortholog pairs were downloaded usingBIOMART interface of ENSEMBL website. Drosophila developmentaldatacourse from (Hooper et al., 2007) was used to create heatmap ofDrosophila orthologues of Anopheles serosa genes in Fig. 4A, CG2666(kkv) profile in Fig. 4B was extracted from (Arbeitman et al., 2002),profile for CG32209 (Serpentine) was extracted from (Manak et al.,2006) using TiMAT software.
Whole-mount In Situ hybridization
Mosquito embryoswere collected andfixed as describedpreviously(Goltsev et al., 2004). The hybridization dig-labeled anti-sense RNAprobes against specific A. gambiae genes were generated by RT-PCRamplification from embryonic RNA and reverse transcription. A 26 bptail encoding the T7 RNA polymerase promoter (TAATACGACTCACTA-TAGGGAGA) was included on the 5′ side of the reverse primer. Thefollowing primer pairs were used to amplify each of the indicated A.gambiae genes. (The T7 promoter sequence is denoted by the symbol,[T7].):
AGAP005564 (grainyhead):GACGTTGGCTTCAAATACTATCTGGAA,aag[T7]AACACTAGTATCACGCGGATCTTCGAA;AGAP011812 (ELOVL, elongase): TCGTGCTAGTGTATGAAGGAATT-GAGG, aag[T7]CGTGTGGAAGAACACAATGATGAACT,AGAP006023 (tyrosine hydroxylase, pale): CACATTGACTACACCGA-GACGGAAA, aag[T7]CCCAAACTCAACCGTGAACCAGTATA,AGAP011936 (Serpentine): AAGTTTAAGAACAAGGAGCGCAAGGT,aag[T7]GTAATGTTGGCGTACTTCTCGATGATG,AGAP009091 (dopa-decarboxylase, Ddc): AAGCTGCGCGGCCTGA-AGGCGGACGAA, aag[T7]ACCGGCGACCGAGCGGGATCTGCCAGT,AGAP001748 (Chitin Synthase, AgCHS1): TTCATCGTCTGGCCGCTG-CTCGAGAAT, aag[T7]AAGAAGCACGGATCATCGTTGCGAATA,AGAP005311 (homeobrain): GTACAGGTTTGGTTCCAGAATCGTC, aag[T7]TGCTACTGTTACTGCTGCTAAGAATGC,AGAP009515 (dorsal): CGAGAAGGTGGTAAAGGAGGACATTAA, aag[T7]AAGCACTTGCATTGTGGTAGGTATAAA,AGAP005137 (lim homeobox, CG32105): AAACATCTCGTCTACCAT-CTGCACTG, aag[T7]AGCTTGATTGATTTGTCCTCCTTGTC
Acknowledgments
This study was funded by an NIH grant (GM46638) and the MooreFoundation to M.L. G.L.R was a fellowship of Conselho Nacional deDesenvolvimento Científico e Tecnológico (CNPq, Brazil).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ydbio.2009.02.038.
References
Araujo, S.J., Aslam, H., Tear, G., Casanova, J., 2005. mummy/cystic encodes an enzymerequired for chitin and glycan synthesis, involved in trachea, embryonic cuticle andCNS development — analysis of its role in Drosophila tracheal morphogenesis. Dev.Biol. 288, 179–193.
Arbeitman, M.N., Furlong, E.E.M., Imam, F., Johnson, E., Null, B.H., Baker, B.S., Krasnow,M.A., Scott, M.P., Davis, R.W., White, K.P., 2002. Gene expression during the life cycleof Drosophila melanogaster. Science 297, 2270–2275.
Beament, J.W.L., 1946a. The formation and structure of the chorion of the egg in anhemipteran, Rhodnius prolixus. Q. J. Microsc. Sci. 87, 392–439.
Beament, J.W.L., 1946b. The waterproofing process in eggs of Rhodnius-prolixus stahl.Proc. R. Soc. Lond. B Biol. Sci. 133, 407–418.
Beament, J.W.L., 1949. The penetration of insect egg-shells. 2. The Properties andpermeability of sub-chorial membranes during development of Rhodnius-prolixus,Stal. Bull. Entomol. Res. 39, 467–488.
Beckel, W.E., 1958. Investigation of permeability, diapause, and hatching in the eggs ofthe mosquito Aedes hexodontus Dyar. Can. J. Zool. 36, 541–553.
Beier, J.C., Copeland, R., Oyaro, C., Masinya, A., Odago, W.O., Oduor, S., Koech, D.K.,Roberts, C.R., 1990. Anopheles gambiae complex egg-stage survival in dry soil fromlarval development sites in western Kenya. J. Am. Mosq. Control Assoc. 6, 105–109.
Benjamini, Y., Hochberg, Y., 1995. Controlling the false discovery rate — a practical andpowerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 57, 289–300.
Bolstad, B.M., Irizarry, R.A., Astrand,M., Speed, T.P., 2003. A comparison of normalizationmethods for high density oligonucleotide array data based on variance and bias.Bioinformatics 19, 185–193.
Campbell, F.L., 1929. The detection and estimation of insect chitin; and the irrelation ofthe cuticula of the american cockroach, Periplaneta americana L. Ann. Entomol. Soc.Am. 22, 401.
Chavez, V.M., Marques, G., Delbecque, J.P., Kobayashi, K., Hollingsworth, M., Burr, J.,Natzle, J.E., O'Connor, M.B., 2000. The Drosophila disembodied gene controls lateembryonic morphogenesis and codes for a cytochrome P450 enzyme that regulatesembryonic ecdysone levels. Development 127, 4115–4126.
Cobben, R.H., 1968. Evolutionary trends in Heteroptera, Part 1, Eggs, Architecture of theShell, Gross Embryology and Eclosion. Center for Agricultural Publishing andDocumentation, Wageningen.
da Fonseca, R.N., von Levetzow, C., Kaischeuer, P., Basal, A., van der Zee, M., Roth, S.,2008. Self-regulatory circuits in dorsoventral axis formation of the short-germbeetle Tribolium castaneum. Dev. Cell 14, 605–615.
Demerec, M., 1950. Biology of Drosophila. New York - John Wiley & Sons, Inc.; London -Chapman & Hall, Limited.
Dorn, A., 1976. Ultrastructure of embryonic envelopes and integument of Oncopeltus-fasciatus Dallas (Insecta, Heteroptera). 1. Chorion, amnion, serosa, integument.Zoomorphologie 85, 111–131.
Dorn, A., Romer, F., 1976. Structure and function of prothoracic glands and enocytes inembryos and last larval instars of Oncopeltus-fasciatus Dallas (Insecta, Heteroptera).Cell Tissue Res. 171, 331–350.
Fletcher, J.C., Thummel, C.S., 1995. The Drosophila E74 gene is required for the properstage-specific and tissue-specific transcription of ecdysone-regulated genes at theonset of metamorphosis. Development 121, 1411–1421.
Furneaux, P.J.S., McFarlane, J.E., 1965a. Identification, estimation, and localization ofcatecholamines in eggs of the house cricket, Acheta domesticus (L.). J. Insect Physiol.11, 591–600.
Furneaux, P.J.S., McFarlane, J.E.,1965b. A possible relationship between the occurrence ofcatecholamines and water absorption in insect eggs. J. Insectphysiol. 11, 631–635.
Gentile, C., Lima, J.B.P., Peixoto, A.A., 2005. Isolation of a fragment homologous to therp49 constitutive gene of Drosophila in the Neotropical malaria vector Anophelesaquasalis (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz 100, 545–547.
Gilbert, L.I., 2004. Halloween genes encode P450 enzymes that mediate steroidhormone biosynthesis in Drosophila melanogaster. Mol. Cell. Endocrinol. 215, 1–10.
Goltsev, Y., Fuse, N., Frasch, M., Zinzen, R.P., Lanzaro, G., Levine, M., 2007. Evolution ofthe dorsal–ventral patterning network in the mosquito, Anopheles gambiae.Development 134, 2415–2424.
Goltsev, Y.,Hsiong,W., Lanzaro,G., Levine,M., 2004.Different combinationsof gap repressorsfor common stripes in Anopheles and Drosophila embryos. Dev. Biol. 275, 435–446.
Gray, E.M., Bradley, T.J., 2004. Water balance in two tropical mosquitoes, Anophelesgambiae and An. arabiensis. Integr. Comp. Biol. 44, 561.
Gray, E.M., Bradley, T.J., 2005. Physiology of desiccation resistance in Anopheles gambiaeand Anopheles arabiensis. Am. J. Trop. Med. Hyg. 73, 553–559.
Harwood, R.F., Horsfall, W.R., 1959. Development, structure and function for coveringsof eggs of floodwatermosquitoes. III. Functions of coverings. Ann. Entomol. Soc. Am.52, 113–116.

470 Y. Goltsev et al. / Developmental Biology 330 (2009) 462–470
Hinton, H.E., 1981. Biology of Insect Eggs. Pergamon Press.Hooper, S.D., Boue, S., Krause, R., Jensen, L.J., Mason, C.E., Ghanim, M., White, K.P.,
Furlong, E.E.M., Bork, P., 2007. Identification of tightly regulated groups of genesduring Drosophila melanogaster embryogenesis. Mol. Syst. Biol. 3, 72.
Huang, J.D., Dubnicoff, T., Liaw, G.J., Bai, Y.J., Valentine, S.A., Shirokawa, J.M., Lengyel, J.A.,Courey, A.J., 1995. Binding sites for transcription factor NTF-1/Elf-1 contribute tothe ventral repression of decapentaplegic. Genes Dev. 9, 3177–3189.
Imboden, H., Lanzrein, B., 1982. Investigations on ecdysteroids and juvenile hormonesand on morphological aspects during early embryogenesis in the ovoviviparouscockroach Nauphoeta-cinerea. J. Insect Physiol. 28, 37–46.
Jahn, T.L., 1935. Nature and permeability of the grasshopper egg membranes. II.Chemical composition of membranes. Proc. Soc. Exp. Biol. Med. xxxiii.
Jakobsson, A., Westerberg, R., Jacobsson, A., 2006. Fatty acid elongases in mammals:their regulation and roles in metabolism. Prog. Lipid Res. 45, 237–249.
Jane, S.M., Ting, S.B., Cunningham, J.M., 2005. Epidermal impermeable barriers inmouseand fly. Curr. Opin. Genet. Dev. 15, 447–453.
Judson, C.L., Hokama, Y., 1965. Formation of the line of dehiscence in Aedine mosquitoeggs. J. Insect Physiol. 11, 337–345.
Kozlova, T., Thummel, C.S., 2003. Essential roles for ecdysone signaling duringDrosophilamid-embryonic development. Science 301, 1911–1914.
Lagueux, M., Hetru, C., Goltzene, F., Kappler, C., Hoffmann, J.A., 1979. Ecdysone titer andmetabolism in relation to cuticulogenesis in embryos of Locusta-migratoria. J. InsectPhysiol. 25, 709–723.
Lamka, M.L., Lipshitz, H.D., 1999. Role of the amnioserosa in germ band retraction of theDrosophila melanogaster embryo. Dev. Biol. 214, 102–112.
Li, J.S., Li, J., 2006. Major chorion proteins and their crosslinking during chorionhardening in Aedes aegypti mosquitoes. Insect Biochem. Mol. Biol. 36, 954–964.
Luschnig, S., Batz, T., Armbruster, K., Krasnow, M.A., 2006. serpentine and vermiformencode matrix proteins with chitin binding and deacetylation domains that limittracheal tube length in Drosophila. Curr. Biol. 16, 186–194.
Mace, K.A., Pearson, J.C., McGinnis, W., 2005. An epidermal barrier wound repairpathway in Drosophila is mediated by grainy head. Science 308, 381–385.
Machida, R., 2006. Evidence from embryology for reconstructing the relationships ofhexapod basal clades. Arthropod Syst. Phylogeny 64, 95–104.
Manak, J.R., Dike, S., Sementchenko, V., Kapranov, P., Biemar, F., Long, J., Cheng, J., Bell, I.,Ghosh, S., Piccolboni, A., Gingeras, T.R., 2006. Biological function of unannotatedtranscription during the early development of Drosophila melanogaster. Nat. Genet.38, 1151–1158.
McFarlane, J., 1960. Structure and function of the egg shell as related towater absorptionby the eggs of Acheta domesticus (L). Can. J. Zool. 38, 231–241.
Minakawa, N., Githure, J.I., Beier, J.C., Yan, G., 2001. Anopheline mosquito survivalstrategies during the dry period in western Kenya. J. Med. Entomol. 38, 388–392.
Monnerat, A.T., Soares, M.J., Lima, J.B.P., Rosa-Freitas, M.G., Valle, D., 1999. Anophelesalbitarsis eggs: ultrastructural analysis of chorion layers after permeabilization.J. Insect Physiol. 45, 915–922.
Moussian, B., Schwarz, H., Bartoszewski, S., Nusslein-Volhard, C., 2005. Involvementof chitinin exoskeleton morphogenesis in Drosophila melanogaster. J. Morphol. 264, 117–130.
Moussian, B., Seifarth, C., Muller, U., Berger, J., Schwarz, H., 2006. Cuticle differentiationduring Drosophila embryogenesis. Arthropod Struct. Develop. 35, 137–152.
Neckameyer, W.S., Quinn, W.G., 1989. Isolation and characterization of the gene forDrosophila tyrosine-hydroxylase. Neuron 2, 1167–1175.
Onyabe, D.Y., Vajime, C.G., Nock, I.H., Ndams, I.S., Akpa, A.U., Alaribe, A.A., Conn, J.E.,2003. The distribution of M and S molecular forms of Anopheles gambiae in Nigeria.Trans. R. Soc. Trop. Med. Hyg. 97, 605–608.
Panfilio, K.A., 2008. Extraembryonic development in insects and the acrobatics ofblastokinesis. Dev. Biol. 313, 471–491.
Payre, F., 2004. Genetic control of epidermis differentiation in Drosophila. Int. J. Dev.Biol. 48, 207–215.
Rafiqi, A.M., Lemke, S., Ferguson, S., Stauber, M., Schmidt-Ott, U., 2008. Evolutionaryorigin of the amnioserosa in cyclorrhaphan flies correlates with spatial andtemporal expression changes of zen. Proc. Natl. Acad. Sci. U. S. A. 105, 234–239.
Reichhart, J.M., 2005. Tip of another iceberg: Drosophila serpins. Trends Cell Biol. 15,659–665.
Rezende, G.L., Martins, A.J., Gentile, C., Farnesi, L.C., Pelajo-Machado, M., Peixoto, A.A.,Valle, D., 2008. Embryonic desiccation resistance in Aedes aegypti: presumptive roleof the chitinized serosal cuticle. BMC Dev. Biol. 8, 82.
Salomonis, N., Hanspers, K., Zambon, A.C., Vranizan, K., Lawlor, S.C., Dahlquist, K.D.,Doniger, S.W., Stuart, J., Conklin, B.R., Pico, A.R., 2007. GenMAPP 2: new features andresources for pathway analysis. Bmc Bioinformatics 8, 217.
Scalia, S., Morgan, E.D.,1982. A reinvestigation on the ecdysteroids during embryogenesisin the desert locust, Schistocerca-gregaria. J. Insect Physiol. 28, 647–654.
Schmidt-Ott, U., 2000. The amnioserosa is an apomorphic character of cyclorrhaphanflies. Dev. Genes Evol. 210, 373–376.
Schmidt-Ott, U., 2005. Insect serosa: a head line in comparative developmentalgenetics. Curr. Biol. 15, R245–R247.
Scuderi, A., Letsou, A., 2005. Amnioserosa is required for dorsal closure in Drosophila.Dev. Dyn. 232, 791–800.
Shao, L., Devenport, M., Jacobs-Lorena, M., 2001. The peritrophic matrix of hemato-phagous insects. Arch. Insect Biochem. Physiol. 47, 119–125.
Shililu, J.I., Grueber, W.B., Mbogo, C.M., Githure, J.I., Riddiford, L.M., Beier, J.C., 2004.Development and survival of Anopheles gambiae eggs in drying soil: influence of therate of drying, egg age, and soil type. J. Am. Mosq. Control Assoc. 20, 243–247.
Slifer, E.H., 1937a. The origin and fate of the membranes surrounding the grasshopperegg, together with some experiments on the source of the hatching enzyme. Q. J.Microsc. Sci. 79, U412–U493.
Slifer, E.H., 1937b. The origin and fate of the membranes surrounding the grasshopperegg; together with some experiments on the source of the hatching enzyme. Quart.Journ. Micr. Sci. 79, 497–508.
Slifer, E.H., 1945. Removing the shell from living grasshopper eggs. Science 102, 282.Smyth, G.K., 2005. Limma: linear models for microarray data. In: Gentleman, R., Carey,
V., Dudoit, S., Irizarry, R., Huber, W. (Eds.), Bioinformatics and ComputationalBiology Solutions using R and Bioconductor. Springer, New York, pp. 397–420.
Toure, Y.T., Petrarca, V., Traore, S.F., Coulibaly, A., Maiga, H.M., Sankare, O., Sow, M.,Dideco, M.A., Coluzzi, M., 1994. Ecological genetic-studies in the chromosomalform mopti of Anopheles–gambiae S-Str in Mali, West-Africa. Genetica 94,213–223.
van der Zee, M., Berns, N., Roth, S., 2005. Distinct functions of the Tribolium zerknulltgenes in serosa specification and dorsal closure. Curr. Biol. 15, 624–636.
Wigglesworth, V.B., 1948. The insect cuticle as a living system. Trans. Faraday Soc. 44,172–177.
Wright, T.R.F., 1996. Phenotypic analysis of the dopa decarboxylase gene clustermutants in Drosophila melanogaster. J. Heredity 87, 175–190.




















