
Determinants of abundance and effects of blood-sucking flying insects in the nest of a hole-nesting...
8
Oecologia (2008) 156:305–312 DOI 10.1007/s00442-008-1001-6 123 POPULATION ECOLOGY - ORIGINAL PAPER Determinants of abundance and eVects of blood-sucking Xying insects in the nest of a hole-nesting bird Gustavo Tomás · Santiago Merino · Josué Martínez-de la Puente · Juan Moreno · Judith Morales · Elisa Lobato Received: 26 April 2007 / Accepted: 30 January 2008 / Published online: 4 March 2008 © Springer-Verlag 2008 Abstract Compared to non-Xying nest-dwelling ectopar- asites, the biology of most species of Xying ectoparasites and its potential impact on avian hosts is poorly known and rarely, if ever, reported. In this study we explore for the Wrst time the factors that may aVect biting midge (Diptera: Cera- topogonidae) and black Xy (Diptera: Simuliidae) abun- dances in the nest cavity of a bird, the hole-nesting blue tit Cyanistes caeruleus, and report their eVects on adults and nestlings during reproduction. The abundance of biting midges was positively associated with nest mass, parental provisioning eVort and abundance of blowXies and black Xies, while negatively associated with nestling condition. Furthermore, a medication treatment to reduce blood par- asitaemias in adult birds revealed that biting midges were more abundant in nests of females whose blood parasitae- mias were experimentally reduced. This Wnding would be in accordance with these insect vectors attacking preferen- tially uninfected or less infected hosts to increase their own survival. The abundance of black Xies in the population was lower than that of biting midges and increased in nests with later hatching dates. No signiWcant eVect of black Xy abun- dance on adult or nestling condition was detected. Blood- sucking Xying insects may impose speciWc, particular selec- tion pressures on their hosts and more research is needed to better understand these host–parasite associations. Keywords Biting midges · Black Xies · Blood parasite– vector–vertebrate host relationship · Cyanistes caeruleus · Nest mass Introduction While colonial avian species are more often attacked by wingless ectoparasites such as ticks, mites and bugs, soli- tary nesters may be more infested by Xying parasites like Xies (Poulin 1991; Loye and Carroll 1995). However, com- pared to non-Xying nest-dwelling ectoparasites, the biology of most species of Xying ectoparasites and its potential impact on avian hosts is poorly known and rarely, if ever, reported (Loye and Carroll 1995). Here, we will refer as Xying insects to blood-feeding Xies (Diptera) belonging to the group Culicimorpha, namely members of families Ceratopogonidae (biting mid- ges) and Simuliidae (black Xies). Biting midges are among the most abundant, and lesser known, haematophagous insects, occurring on almost all large land masses through- out the world (Mellor et al. 2000). Biting midges are short- lived small Xies measuring 1–3 mm in size; most of them are crepuscular or nocturnal and currently many species are considered to form species complexes with diYcult reliable identiWcation (Mellor et al. 2000). Black Xies are also wide- spread throughout the world, being found on all continents Communicated by Markku Orell. G. Tomás · S. Merino · J. Martínez-de la Puente · J. Moreno · J. Morales · E. Lobato Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), J. Gutiérrez Abascal 2, 28006 Madrid, Spain G. Tomás (&) Centre for Terrestrial Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Postbus 40, 6666 ZG Heteren, The Netherlands e-mail: [email protected]; [email protected] Present Address: J. Morales Departamento de Ecología Evolutiva, Instituto de Ecología, Universidad Nacional Autónoma de México, AP 70-275, 04510 México D.F., México
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Determinants of abundance and effects of blood-sucking flying insects in the nest of a hole-nesting...

Oecologia (2008) 156:305–312
DOI 10.1007/s00442-008-1001-6POPULATION ECOLOGY - ORIGINAL PAPER
Determinants of abundance and eVects of blood-sucking Xying insects in the nest of a hole-nesting bird
Gustavo Tomás · Santiago Merino · Josué Martínez-de la Puente · Juan Moreno · Judith Morales · Elisa Lobato
Received: 26 April 2007 / Accepted: 30 January 2008 / Published online: 4 March 2008© Springer-Verlag 2008
Abstract Compared to non-Xying nest-dwelling ectopar-asites, the biology of most species of Xying ectoparasitesand its potential impact on avian hosts is poorly known andrarely, if ever, reported. In this study we explore for the Wrsttime the factors that may aVect biting midge (Diptera: Cera-topogonidae) and black Xy (Diptera: Simuliidae) abun-dances in the nest cavity of a bird, the hole-nesting blue titCyanistes caeruleus, and report their eVects on adults andnestlings during reproduction. The abundance of bitingmidges was positively associated with nest mass, parentalprovisioning eVort and abundance of blowXies and blackXies, while negatively associated with nestling condition.Furthermore, a medication treatment to reduce blood par-asitaemias in adult birds revealed that biting midges weremore abundant in nests of females whose blood parasitae-mias were experimentally reduced. This Wnding would bein accordance with these insect vectors attacking preferen-tially uninfected or less infected hosts to increase their own
survival. The abundance of black Xies in the population waslower than that of biting midges and increased in nests withlater hatching dates. No signiWcant eVect of black Xy abun-dance on adult or nestling condition was detected. Blood-sucking Xying insects may impose speciWc, particular selec-tion pressures on their hosts and more research is needed tobetter understand these host–parasite associations.
Keywords Biting midges · Black Xies · Blood parasite–vector–vertebrate host relationship · Cyanistes caeruleus · Nest mass
Introduction
While colonial avian species are more often attacked bywingless ectoparasites such as ticks, mites and bugs, soli-tary nesters may be more infested by Xying parasites likeXies (Poulin 1991; Loye and Carroll 1995). However, com-pared to non-Xying nest-dwelling ectoparasites, the biologyof most species of Xying ectoparasites and its potentialimpact on avian hosts is poorly known and rarely, if ever,reported (Loye and Carroll 1995).
Here, we will refer as Xying insects to blood-feedingXies (Diptera) belonging to the group Culicimorpha,namely members of families Ceratopogonidae (biting mid-ges) and Simuliidae (black Xies). Biting midges are amongthe most abundant, and lesser known, haematophagousinsects, occurring on almost all large land masses through-out the world (Mellor et al. 2000). Biting midges are short-lived small Xies measuring 1–3 mm in size; most of themare crepuscular or nocturnal and currently many species areconsidered to form species complexes with diYcult reliableidentiWcation (Mellor et al. 2000). Black Xies are also wide-spread throughout the world, being found on all continents
Communicated by Markku Orell.
G. Tomás · S. Merino · J. Martínez-de la Puente · J. Moreno · J. Morales · E. LobatoDepartamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), J. Gutiérrez Abascal 2, 28006 Madrid, Spain
G. Tomás (&)Centre for Terrestrial Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Postbus 40, 6666 ZG Heteren, The Netherlandse-mail: [email protected]; [email protected]
Present Address:J. MoralesDepartamento de Ecología Evolutiva, Instituto de Ecología, Universidad Nacional Autónoma de México, AP 70-275, 04510 México D.F., México
123

306 Oecologia (2008) 156:305–312
except Antarctica, and playing very important ecologicalroles in certain ecosystems (Malmqvist et al. 2004a). BlackXies are slightly larger Xies (2–6 mm) with a strongly con-vex thorax giving a humpbacked appearance; most of themare diurnal and may occur in large numbers in some areas(Malmqvist et al. 2004a). Both groups share a similar lifecycle, including egg, several larval stages requiring wateror moisture, pupa, and Xying adult. In both groups, malesand females feed on a sugar source (e.g. plant sap), whilethe females, but not the males, of many species are alsoblood-feeders, requiring blood for egg development (Leh-ane 2005). Variable proportions of species in both familiesare ornithophilic, depending on the area and the speciescomposition of the community (Adler et al. 1999; Melloret al. 2000; Malmqvist et al. 2004b).
Although the few published studies reporting the eVectsof blood-sucking Xying insects on avian hosts have under-taken quite rough qualitative estimates of their abundances(Tomás et al. 2008), their impact does not appear to be neg-ligible. Black Xies have been shown to be a nuisance foradult (Bukacijski and Bukacijska 2000; Rohner et al.2000; Ojanen et al. 2002; Ratti et al. 2006) and nestlingbirds (Smith et al. 1998; Bukacijski and Bukacijska 2000),causing dermatitis and skin injuries (Hunter et al. 1997;Smith et al. 1998; Bukacijski and Bukacijska 2000). Theymay provoke blood loss and anaemia (Hunter et al. 1997)and are vectors of Wlarial nematodes (Anderson 1956), try-panosomes (Votypka et al. 2002), and haemosporidianssuch as diVerent Leucocytozoon (Khan and Fallis 1971;Herman et al. 1975) and Haemoproteus species (Fallis andBennett 1961). Overall, black Xies can cause dramaticreductions in reproductive success and even mortality inbirds (Hunter et al. 1997; Smith et al. 1998; Bukacijski andBukacijska 2000). Even less known is the impact of bitingmidges on birds, though it has been shown that they canalso be vectors of Wlarial nematodes (Weinmann et al.1979; Mellor et al. 2000), several Haemoproteus species(Fallis and Wood 1957; Valkiínas et al. 2002; Garvin andGreiner 2003; Valkiínas and Iezhova 2004a; Mullens et al.2006) and viruses (Naugle et al. 2004; Lehane 2005). Simi-lar detrimental eVects associated with blood feeding byblack Xies may be expected for biting midges. Overall, thedirect and indirect eVects of this ectoparasite guild on theiravian hosts are largely unknown (Mellor et al. 2000; Roh-ner et al. 2000; Malmqvist et al. 2004a), as well as thecauses determining their abundances in avian nests (Atkin-son and Van Riper 1991; Smith et al. 1998; Leppert et al.2004).
In this study we explore for the Wrst time the factors thatmay aVect Xying insect abundances in the nest of a bird, thehole-nesting blue tit Cyanistes caeruleus, and report theireVects on adults and nestlings during reproduction. We spe-ciWcally look for associations with host breeding phenology,
female condition and health state, nestling quality andabundance of other ectoparasites. We focus on femalesbecause they spend prolonged periods in the nest duringincubation and brooding. In addition, we also explore theeVect of a medication treatment to reduce adult blood par-asitaemias on Xying insect abundances. This treatment hasdetectable eVects on blood parasitaemias in females but notin males (see Martínez-de la Puente et al. 2007). The abun-dance of Xying insects may be lower in nests of medicatedfemales as shown for nest-dwelling ectoparasites (Tomáset al. 2007a). Alternatively, the abundance of Xying insectsmay be higher in nests of medicated females if Xyinginsects preferentially attack hosts uninfected or lessinfected by blood parasites, which increases insect survival(Desser and Yang 1973; Valkiínas and Iezhova 2004b).
Materials and methods
The study was carried out during the 2004 breeding seasonin a Pyrenean Oak Quercus pyrenaica deciduous forestlocated in Valsaín (Segovia, central Spain, 40°53�N,4°01�W, 1,200 m a.s.l.). The study site lies in the uppersupraMediterranean bioclimatic belt, with a mean annualtemperature range of 8–12°C and 750–1,000 mm of annualrainfall (Izco 1984). Around the study area, there are somepermanent streams and numerous dispersed seasonal smallstreams and ponds. A population of blue tits Cyanistescaeruleus breeding in wooden nest boxes hanging frombranches at about 5 m above the ground has been studiedsince 1991 in this area (e.g. Tomás et al. 2006). Every year,nest boxes are periodically inspected and the dates of clutchinitiation, clutch sizes, hatching dates and Xedging successare determined as described in Tomás et al. (2007a).
An antimalarial treatment was applied to adult birdstrapped at nest boxes at the nestling age of 3 days (hatchingdate = age 0) (Martínez-de la Puente et al. 2007). In brief,birds attending nests of similar clutch size (§ one egg) andlaying date (§ one day) were randomly assigned to one oftwo treatments (medicated and control). Medicated birdswere injected subcutaneously in the abdominal region with0.1 mg of the antimalarial primaquine (Sigma, St Louis,Mo.) diluted in 0.1 ml of saline solution and control birdswere injected with the same volume of saline solution. Thistreatment generated pairs with both individuals medicated,pairs with both controls and pairs with one medicated andone control bird. For determination of blood parasite loads,thin blood smears were made from a drop of blood obtainedfrom the brachial vein of adults with the aid of a needle anda capillary tube. Blood smears were immediately air-driedand later Wxed with ethanol (96%) and stained with Giemsa(1/10 v/v) for 45 min. Half of the symmetrical smear wasscanned at £200 magniWcation in search of large blood
123

Oecologia (2008) 156:305–312 307
parasites such as Trypanosoma or Leucocytozoon, whereassmall intra-erythrocytic parasites such as Haemoproteuswere detected on the other half of the smear using £1,000magniWcation (Merino and Potti 1995a; Merino et al.1997). Intensity of infection (abundance) by Haemoproteusparasites was estimated as the number of infected cells per2,000 erythrocytes, a reliable method to estimate intensitiesof infection by these parasites as shown by Godfrey et al.(1987). We used presence/absence indexes for Trypano-soma and Leucocytozoon parasites due to their low intensi-ties of infection. Other blood parasites were present in lowprevalences and therefore not included in analyses. Adultbirds were banded individually with numbered aluminiumrings if unringed. To measure nestling-provisioning rate,both males and females were equipped with uniquely iden-tiWable transponders attached to two colour rings on theirtarsus. Provisioning rate was recorded on day 12 of nestlingage with a datalogger (Trovan, EID Iberica, Madrid) con-nected to an antenna at the nest box entrance for a mini-mum of 90 min (mean 164 § SE 14 min). Some nests forwhich one of the adults was not recorded at the nest werediscarded from the analyses. Male and female provisioningrates were added as a measure of total parental provisioningeVort. Results express provisioning rates relative to 1-hperiods. Nest mass, as an estimate of the amount of nestmaterial (see Tomás et al. 2006), was recorded to the near-est 0.1 g when nestlings were 3 days old. We have shown ina previous study that nest mass is related to female healthstatus (Tomás et al. 2006).
Adult birds were trapped in their nest boxes again whentheir nestlings were 13 days old. A second thin bloodsmear was prepared for determination of blood parasiteloads as in the Wrst capture. At this capture, female masswas recorded with an electronic balance to the nearest0.1 g and tarsus length was measured with a digital calliperto the nearest 0.01 mm. Tarsus length of all nestlings wasmeasured and nestling mass was recorded at 13 days ofage as for females. We used mass/tarsus length, as a mea-sure of female and nestling body condition. We used thesum of nestling masses at 13 days of age (brood mass) as asurrogate of the amount of heat and CO2 that the broodproduce and that may serve as a cue for nest detection byXying insects. Replacing this variable by the number ofnestlings did not change the results. After the nestlings hadXedged, nests were removed and nest material was care-fully dismantled in order to count pupae of the ectopara-sitic blowXy Protocalliphora azurea (Merino and Potti1995b).
To estimate Xying insect abundance we placed a piece ofplastic tape (10 £ 4 cm; 40 cm2) horizontally supported onwire netting attached with two drawing pins inside andclose to the roof of all blue tit nest boxes when nestlingswere 12 days old. The lower surface of the plastic tape was
covered with 0.5 ml of commercially available body gel–oil(Johnson’s baby chamomilla; Johnson & Johnson, Dussel-dorf). The gel–oil comprises paraVinum liquidum, hexyllaurate, ethylene/propylene/styrene copolymer, cyclope-ntasiloxane, butylene/ethylene/styrene copolymer, Cham-omilla recutita, bisabolol and fragrance (FPT1353). Thissticky trap allows the capture of Xying insects when they goto land on the inner part of the roof and encounter the lowerside of the tape. We did not record whether the trappedinsects were engorged or not. However, in another study,6.3% of the biting midges trapped in nest boxes wereengorged, suggesting that they are ornithophilic and feed onblue tits (Tomás et al. 2008, see Mullens et al. 2006). Fur-thermore, Xying insects do not just rest inside the nestboxes because no insects were trapped in empty nest boxesand in nest boxes just after nestlings Xedged (Tomás et al.2008). The lack of Xying insects in empty nest boxes alsoindicates that the gel–oil does not attract Xying insects fromoutside the nest box (Tomás et al. 2008). The wire nettingsupported the plastic tape in the correct position and pre-vented birds from contacting it or reaching the trappedinsects (Tomás et al. 2008). The next day (day 13 of nest-ling age) the device was removed from nest boxes. Plastictapes were stored frozen until examination under a binocu-lar lens for Xying insect quantiWcation.
Statistical analyses
The eVect of the medication treatment was analysed bymeans of analyses of covariance (ANCOVAs) with Wnalintensity of infection by Haemoproteus majoris as depen-dent variable and initial intensity as covariate. Final preva-lences of Trypanosoma avium and Leucocytozoon majoriswere also compared between medicated and control birds.Maximum sample sizes available were employed for theseanalyses (data on initial and Wnal haematozoa infection inrelation to treatment was available for 79 females and 75males).
We conducted general regression models (GRM), apply-ing backward stepwise solution to identify signiWcant vari-ables associated with Xying insect abundances. Bitingmidge or black Xy abundance was included as the depen-dent variable; female medication treatment, and femaleprevalences of T. avium and L. majoris at 13 days of nest-ling age as categorical predictors; and hatching date, nestmass, female body condition, female intensity of infectionby H. majoris at 13 days of nestling age, nestling body con-dition, brood mass, parental provisioning rates per hour,and abundances of blowXies and either biting midges orblack Xies, as continuous predictors. Fifty-six nests wereincluded in the GRM, those for which data for all variableswere available. The results did not change when applying for-ward stepwise solutions or when including male medication
123
308 Oecologia (2008) 156:305–312
treatment. No interaction was included in the models.Residuals of the models were tested for normality. Becausecollinearity between independent variables might confoundthe analyses, we checked for redundancy by investigatingtolerance levels for the variables, which proved adequatelyhigh in all cases (more than 0.67; Quinn and Keough 2002).We also looked at variance inXation factors for individualpredictors, which were always lower than 1.5, a value farbelow those accepted as giving unbiased results (Chaterjeeet al. 2000). Thus, collinearity should not have seriouslybiased our regression models.
To explore further the possibility that Xying insectsmay preferentially attack hosts with fewer blood parasites(see Results), Xying insect abundances were analysed inrelation to initial blood parasitaemias of female blue tits.Again, sample sizes for these analyses were larger (n = 88nests) than for the GRM because a reduced number ofvariables involved in the analyses led to fewer missingvalues.
To explore the potential eVects of Xying insects and toconWrm further the relationships of biting midge abundancewith nestling condition and parental provisioning rate (seeResults), we carried our further analyses with either bitingmidge abundance or black Xy abundance as continuous pre-dictors and either nestling condition (n = 88 nests), parentalprovisioning rate (n = 67 nests), Xedging success (n = 96nests), female condition (n = 89 nests) and female Haemo-proteus intensity, Trypanosoma prevalence and Leucocyto-zoon prevalence (n = 76 nests) at 13 days of nestling age asthe dependent variable. All analyses were done with STAT-ISTICA, version 7 (Statsoft, Tulsa, Okla.). Parasite abun-dances were logarithmically transformed and Xedgingsuccess (Xedglings/hatchlings) was arcsin-square roottransformed to ensure normality. Means § SE are shownexcept otherwise stated.
Results
The medication treatment reduced the intensity of infectionby H. majoris in females (ANCOVA: treatment,F1,76 = 5.26, P = 0.025; initial infection, F1,76 = 403.01,P < 0.0001) but not in males (ANCOVA: treatment,F1,72 = 0.30, P = 0.583; initial infection, F1,72 = 439.11,P < 0.0001). Final prevalences of T. avium and L. majorisdid not diVer between medicated and control birds, in malesand in females (Pearson �2, all P > 0.40).
Of the 56 nests that were included in the GRM, bitingmidges were present in 55 of these nests (98.2%) with anaverage of 41.9 § 10.6 biting midges per nest (range 0–528). Black Xies were present in 26 nests (46.4%) with anaverage of 0.9 § 0.2 black Xies per nest (range 0–6).
Determinants of abundance of Xying insects
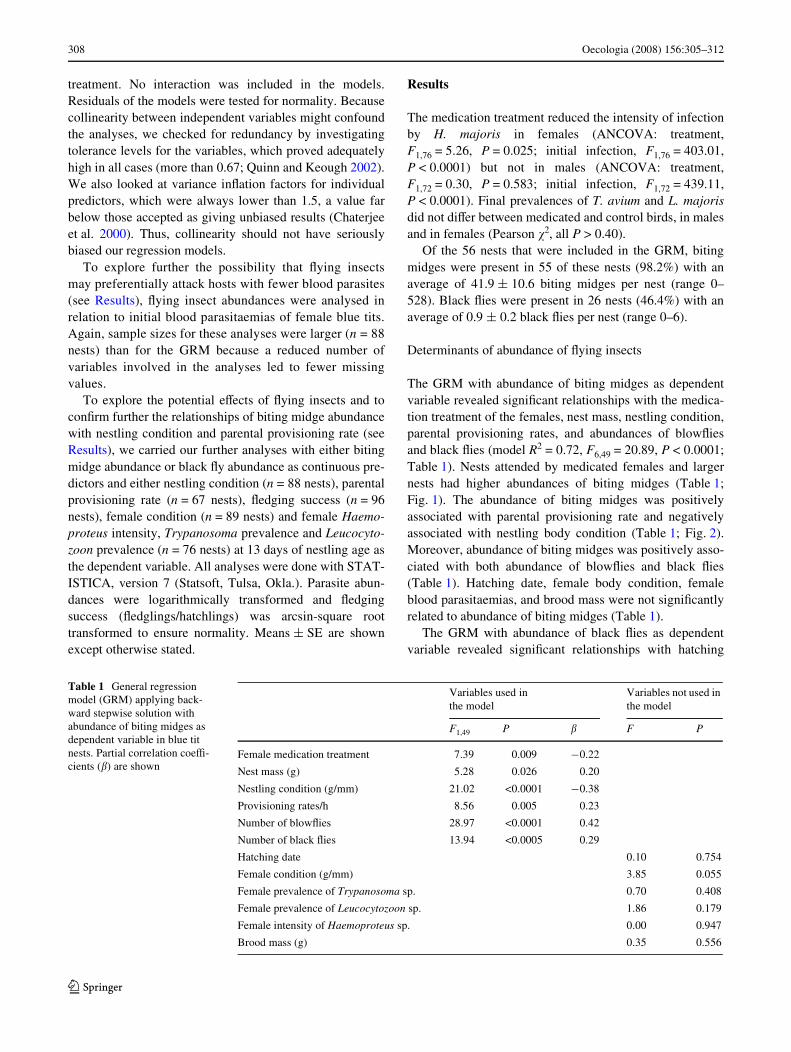
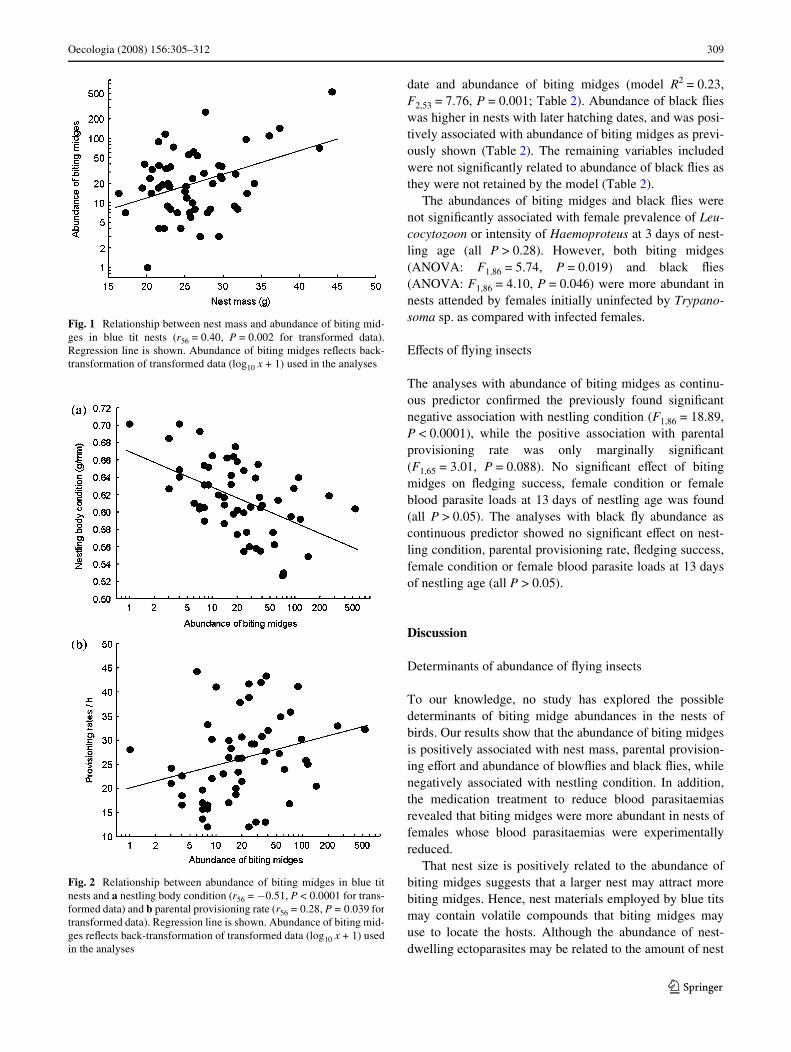
The GRM with abundance of biting midges as dependentvariable revealed signiWcant relationships with the medica-tion treatment of the females, nest mass, nestling condition,parental provisioning rates, and abundances of blowXiesand black Xies (model R2 = 0.72, F6,49 = 20.89, P < 0.0001;Table 1). Nests attended by medicated females and largernests had higher abundances of biting midges (Table 1;Fig. 1). The abundance of biting midges was positivelyassociated with parental provisioning rate and negativelyassociated with nestling body condition (Table 1; Fig. 2).Moreover, abundance of biting midges was positively asso-ciated with both abundance of blowXies and black Xies(Table 1). Hatching date, female body condition, femaleblood parasitaemias, and brood mass were not signiWcantlyrelated to abundance of biting midges (Table 1).
The GRM with abundance of black Xies as dependentvariable revealed signiWcant relationships with hatching
Table 1 General regression model (GRM) applying back-ward stepwise solution with abundance of biting midges as dependent variable in blue tit nests. Partial correlation coeY-cients (�) are shown
Variables used in the model
Variables not used in the model
F1,49 P � F P
Female medication treatment 7.39 0.009 ¡0.22
Nest mass (g) 5.28 0.026 0.20
Nestling condition (g/mm) 21.02 <0.0001 ¡0.38
Provisioning rates/h 8.56 0.005 0.23
Number of blowXies 28.97 <0.0001 0.42
Number of black Xies 13.94 <0.0005 0.29
Hatching date 0.10 0.754
Female condition (g/mm) 3.85 0.055
Female prevalence of Trypanosoma sp. 0.70 0.408
Female prevalence of Leucocytozoon sp. 1.86 0.179
Female intensity of Haemoproteus sp. 0.00 0.947
Brood mass (g) 0.35 0.556
123
Oecologia (2008) 156:305–312 309
date and abundance of biting midges (model R2 = 0.23,F2,53 = 7.76, P = 0.001; Table 2). Abundance of black Xieswas higher in nests with later hatching dates, and was posi-tively associated with abundance of biting midges as previ-ously shown (Table 2). The remaining variables includedwere not signiWcantly related to abundance of black Xies asthey were not retained by the model (Table 2).
The abundances of biting midges and black Xies werenot signiWcantly associated with female prevalence of Leu-cocytozoon or intensity of Haemoproteus at 3 days of nest-ling age (all P > 0.28). However, both biting midges(ANOVA: F1,86 = 5.74, P = 0.019) and black Xies(ANOVA: F1,86 = 4.10, P = 0.046) were more abundant innests attended by females initially uninfected by Trypano-soma sp. as compared with infected females.
EVects of Xying insects
The analyses with abundance of biting midges as continu-ous predictor conWrmed the previously found signiWcantnegative association with nestling condition (F1,86 = 18.89,P < 0.0001), while the positive association with parentalprovisioning rate was only marginally signiWcant(F1,65 = 3.01, P = 0.088). No signiWcant eVect of bitingmidges on Xedging success, female condition or femaleblood parasite loads at 13 days of nestling age was found(all P > 0.05). The analyses with black Xy abundance ascontinuous predictor showed no signiWcant eVect on nest-ling condition, parental provisioning rate, Xedging success,female condition or female blood parasite loads at 13 daysof nestling age (all P > 0.05).
Discussion
Determinants of abundance of Xying insects
To our knowledge, no study has explored the possibledeterminants of biting midge abundances in the nests ofbirds. Our results show that the abundance of biting midgesis positively associated with nest mass, parental provision-ing eVort and abundance of blowXies and black Xies, whilenegatively associated with nestling condition. In addition,the medication treatment to reduce blood parasitaemiasrevealed that biting midges were more abundant in nests offemales whose blood parasitaemias were experimentallyreduced.
That nest size is positively related to the abundance ofbiting midges suggests that a larger nest may attract morebiting midges. Hence, nest materials employed by blue titsmay contain volatile compounds that biting midges mayuse to locate the hosts. Although the abundance of nest-dwelling ectoparasites may be related to the amount of nest
Fig. 1 Relationship between nest mass and abundance of biting mid-ges in blue tit nests (r56 = 0.40, P = 0.002 for transformed data).Regression line is shown. Abundance of biting midges reXects back-transformation of transformed data (log10 x + 1) used in the analyses
Fig. 2 Relationship between abundance of biting midges in blue titnests and a nestling body condition (r56 = ¡0.51, P < 0.0001 for trans-formed data) and b parental provisioning rate (r56 = 0.28, P = 0.039 fortransformed data). Regression line is shown. Abundance of biting mid-ges reXects back-transformation of transformed data (log10 x + 1) usedin the analyses
123

310 Oecologia (2008) 156:305–312
material due to density-dependent mechanisms (Heeb et al.1996, 2000; see also Wasylik 1971), to our knowledge suchan association between nest size and abundance of Xyinginsect vectors (that do not dwell in the nest) has never beenreported. Both black Xy and blowXy abundance were posi-tively associated with that of biting midges suggesting thatthese insect vectors in which the Xying adults search forhosts use similar cues for host detection. However, blackXy abundance was correlated only with biting midge abun-dance but not with nest mass.
Our results also show that nests attended by femaleswhose blood parasitaemia was experimentally reduced holdhigher abundances of biting midges than control birds. Thiscontrasts with the previous observation that experimentalreductions of female blood parasitaemias are associated withreduced levels of nest-dwelling ectoparasitism in the nest(Tomás et al. 2007a). Flying insects may preferentiallyattack uninfected hosts or hosts less infected by blood para-sites, given that survival of biting midges and black Xies isinversely related to the intensity of parasitaemia in the birdsthey have fed on (Desser and Yang 1973; Valkiínas andIezhova 2004b), as reported also for other insect vectors(Lambrechts et al. 2006). This possibility is partially sup-ported by the fact that abundances of both biting midges andblack Xies were higher in nests attended by females unin-fected by trypanosomes as compared with infected femalesat the beginning of the nestling period, when females arestill brooding their young and spend much time at the nest.
Alternatively, through being released from the drainimposed by blood parasites, females in the medicated groupmay have increased their provisioning visits to the nest(Merino et al. 2000; Tomás et al. 2007a), and hence mayhave made their nests more easy to locate by Xying insects.Blood-sucking Xying insects can locate their hosts guidedby visual cues and follow moving objects (SutcliVe 1986;
Lehane 2005; Yuval 2006). Hence, biting midges may takeadvantage of the activity of the parents at the nest as a cueto locate hosts, locating more easily those nests where par-ent birds are provisioning the nestlings at a higher rate.
To our knowledge, no study has explored the possibledeterminants of black Xy abundances in the nests of birds.Our results show that black Xies are more abundant later inthe season in the blue tit population studied (see also Oja-nen et al. 2002; Malmqvist et al. 2004a), implying thatblack Xies may impose a selective force for earlier breedingin temperate areas.
EVects of Xying insects
To our knowledge, no study has explored the eVects of bit-ing midges on breeding birds. When exploring the potentialeVects of biting midges on reproductive parameters, wefound a signiWcant negative relationship of biting midgeabundances with nestling condition and a marginally sig-niWcant positive relationship with provisioning rates. Ourresults may suggest that parent blue tits increase their pro-visioning eVort as the abundance of biting midgesincreases, presumably in an attempt to counteract its delete-rious eVects on the oVspring. However, this was not thecase, since nestling condition was negatively related to theabundance of biting midges. As for other ectoparasites,Xying insects may increase the energy requirements of nes-tlings, inducing them to beg more (Tripet and Richner1997) and thus receive more food from the adults (Tripetand Richner 1997; Hurtrez-Boussès et al. 1998). Femalebody condition was not signiWcantly impaired by Xyinginsects, probably because adults increase their eVort tobuVer the detrimental eVects of ectoparasitism on nestlings,but only to a certain level to avoid severe costs to them-selves (Merino et al. 1998; Tomás et al. 2007b).
Table 2 GRM applying back-ward stepwise solution with abundance of black Xies as dependent variable in blue tit nests. � are shown
Variables used in the model
Variables not used in the model
F1,53 P � F P
Hatching date 7.29 0.009 0.33
Number of biting midges 8.86 0.004 0.36
Female medication treatment 0.08 0.775
Nest mass (g) 0.00 0.967
Female condition (g/mm) 0.40 0.530
Female prevalence of Trypanosoma sp. 0.38 0.542
Female prevalence of Leucocytozoon sp. 0.04 0.838
Female intensity of Haemoproteus sp. 1.39 0.243
Provisioning rates/h 0.08 0.785
Nestling condition (g/mm) 2.09 0.154
Brood mass (g) 0.03 0.867
Number of blowXies 2.17 0.147
123

Oecologia (2008) 156:305–312 311
Blood parasitaemias of female blue tits at 13 days ofnestling age were not associated with Xying insect abun-dances, in spite of both biting midges (Fallis and Wood1957; Valkiínas et al. 2002; Garvin and Greiner 2003; Val-kiínas and Iezhova 2004a; Mullens et al. 2006) and blackXies (Fallis and Bennett 1961; Khan and Fallis 1971; Her-man et al. 1975; Votypka et al. 2002) being potential vec-tors of several blood parasites in birds. It has been shownthat female blood parasitaemia is indeed associated withabundance of nest-dwelling ectoparasites (Tomás et al.2007b), which may be already present in the nest duringincubation and brooding (when females may be moreexposed to ectoparasites) because some parts of their popu-lations overwinter in nest boxes (Burtt et al. 1991; Harperet al. 1992). However, data indicate that abundances ofXying insects in nests during incubation are low (Tomáset al. 2008), with the onset of infestation frequently occur-ring after the nestlings hatch (Smith et al. 1998), althoughthere could be strong environmental inXuences on Xyinginsect abundances and on the seasonality of their emergingpatterns (Smith et al. 1998; Mellor et al. 2000; Malmqvistet al. 2004a). It could be interesting to explore nestling par-asitaemias, because Xying insects may attack nestlingsmore frequently than adults due to their conWnement to thenest. However, prepatency periods of blood parasites, i.e.the period between initial infection and release of gameto-cytes into the blood, may have made the detection of newinfections diYcult by the time we sampled birds (Tomáset al. 2007b). In this respect, only Trypanosoma sp. andLeucocytozoon sp., both having short prepatency periods,have been detected in 15-day-old nestlings of this blue titpopulation (Fargallo and Merino 2004), whereas new infec-tions by Haemoproteus sp. are more diYcult to detect(Merino and Potti 1995a; Lobato et al. 2005).
Very few studies have explored the eVects of black Xieson birds (Hunter et al. 1997; Smith et al. 1998; Bukacijskiand Bukacijska 2000; Rohner et al. 2000). Black Xies arefar less abundant than biting midges in the blue tit popula-tion studied and no signiWcant eVect on Xedging successand adult or nestling body condition was detected, althoughthe variance explained by the model was quite low (23%)compared to variance explained for biting midges (72%).Black Xies may be more detrimental to their bird hostsin situations of low food availability or inclement weather(Hunter et al. 1997; Loye and Carroll 1998; Smith et al.1998).
In conclusion, our results show that Xying insects notonly can have severe eVects during mass outbreaks asshown for colonial (Bukacijski and Bukacijska 2000) andsolitary breeding host species (Hunter et al. 1997; Smithet al. 1998), but that also in smaller numbers, they canhave detrimental eVects for solitary breeding birds. Blood-sucking Xying insects exhibit ecological and evolutionary
diVerences with other ectoparasites like nest-dwelling ecto-parasites and thus the selection pressures imposed on theevolution of avian life histories may well be diVerent (Loyeand Carroll 1995). Further research on these host–parasiteassemblages is urged, because their impact has been largelyneglected until now.
Acknowledgements We thank two anonymous reviewers for valu-able comments on the manuscript. We thank Javier Donés (Director ofMontes de Valsaín) for permission to work in the study area. The Juntade Castilla y León authorized the ringing and handling of birds. Thisstudy was funded by projects BOS2003-05724 and CGL2006-14129-C02-01 from the Ministerio de Educación y Ciencia (to S.M.) andCGL2004-00787 (to J. Moreno). J. M.-P. is supported by a grant fromEl Ventorrillo Weld station. J. Morales was supported by a FPI and apostdoctoral grant from the Ministerio de Ciencia y Tecnología. E. L.was supported by a FPU grant from MEC. G. T. was supported atdiVerent stages by a FPI grant from the Comunidad de Madrid, an I3Ppostdoctoral contract from CSIC and a postdoctoral grant from MEC.This study is a contribution to the research developed at El VentorrilloWeld station and complies with the current laws of the country in whichit was performed.
References
Adler PH, Malmqvist B, Zhang Y (1999) Black Xies (Diptera: Simulii-dae) of northern Sweden, with notes on their taxonomy, chromo-somes, and bionomics. Entomol Scand 29:361–382
Anderson RC (1956) The life cycle and seasonal transmissions of Or-nithoWlaria falliesensis Anderson: a parasite of domestic and wildducks. Can J Zool 34:485–525
Atkinson CT, Van Riper III C (1991) Pathogenicity and epizootiologyof avian haematozoa: Plasmodium, Leucocytozoon, and Haemo-proteus. In: Loye JE, Zuk M (eds) Bird–parasite interactions.Ecology, evolution and behaviour. Oxford University Press, Ox-ford, pp 19–48
Bukacijski D, Bukacijska B (2000) The impact of mass outbreaks ofblack Xies (Simuliidae) on the parental behaviour and breedingoutput of colonial common gulls (Larus canus). Ann Zool Fenn37:43–49
Burtt EH Jr, Chow W, Babbitt GA (1991) Ocurrence and demographyof mites of tree swallow, house wren, and eastern bluebird nests.In: Loye JE, Zuk M (eds) Bird–parasite interactions. Ecology,evolution and behaviour. Oxford University Press, Oxford, pp104–122
Chaterjee S, Hadi AS, Price B (2000) Regression analysis by example.Wiley, New York
Desser SS, Yang YJ (1973) Sporogony of Leucocytozoon spp. in mam-malophilic simuliids. Can J Zool 51:793–793
Fallis AM, Bennett GF (1961) Sporogony of Leucocytozoon and Hae-moproteus in simuliids and ceratopogonids and a revised classiW-cation of the Haemosporidiida. Can J Zool 39:215–228
Fallis AM, Wood DM (1957) Biting midges (Diptera: Ceratopogoni-dae) as intermediate hosts for Haemoproteus of ducks. Can J Zool35:425–435
Fargallo JA, Merino S (2004) Clutch size and haemoparasite speciesrichness in adult and nestling blue tits. Ecoscience 11:168–174
Garvin MC, Greiner EC (2003) Ecology of Culicoides (Diptera: Cera-topogonidae) in southcentral Florida and experimental Culicoidesvectors of the avian hematozoan Haemoproteus danilewskyiKruse. J Wildl Dis 39:170–178
Godfrey RD Jr, Fedynich AM, Pence DB (1987) QuantiWcation of he-matozoa in blood smears. J Wildl Dis 23:558–565
123

312 Oecologia (2008) 156:305–312
Harper GH, Marchant A, Boddington DG (1992) The ecology of thehen Xea Ceratophyllus gallinae and the moorhen Xea Dasypsyllusgallinulae in nestboxes. J Anim Ecol 61:317–327
Heeb P, Werner I, Richner H, Kölliker M (1996) Horizontal transmis-sion and reproductive rates of hen Xeas in great tit nests. J AnimEcol 65:474–484
Heeb P, Kölliker M, Richner H (2000) Bird–ectoparasite interactions,nest humidity, and ectoparasite community structure. Ecology81:958–968
Herman CM, Barrow JH Jr, Tarshis IB (1975) Leucocytozoonosis inCanada geese at the Seney National Wildlife Refuge. J Wildl Dis11:404–411
Hunter DB, Rohner C, Currie DC (1997) Mortality in Xedging greathorned owls from blackXy hematophaga and Leucocytozoonosis.J Wildl Dis 33:486–491
Hurtrez-Boussès S, Blondel J, Perret P, Fabreguettes J, Renaud FR(1998) Chick parasitism by blowXies aVects feeding rates in aMediterranean population of blue tits. Ecol Lett 1:17–20
Izco J (1984) Madrid Verde. Instituto de Estudios Agrarios, Pesquerosy Alimentarios. Ministerio de Agricultura, Pesca y Alimentación.Comunidad de Madrid, Madrid
Khan RA, Fallis AM (1971) Speciation, transmission, and schizogonyof Leucocytozoon in corvid birds. Can J Zool 49:1363–1367
Lambrechts L, Chavatte J-M, Snounou G, Koella JC (2006) Environ-mental inXuence on the genetic basis of mosquito resistance tomalaria parasites. Proc R Soc Lond B 273:1501–1506
Lehane M (2005) The biology of blood-sucking in insects, 2nd edn.Cambridge University Press, Cambridge
Leppert LL, Layman S, Bragin EA, Katzner T (2004) Survey for he-moparasites in imperial eagles (Aquila heliaca), steppe eagles(Aquila nipalensis), and white-tailed sea eagles (Haliaeetus albi-cilla) from Kazakhstan. J Wildl Dis 40:316–319
Lobato E, Moreno J, Merino S, Sanz JJ, Arriero E (2005) Haematolog-ical variables are good predictors of recruitment in nestling piedXycatchers Ficedula hypoleuca. Ecoscience 12:27–34
Loye JE, Carroll SP (1995) Birds, bugs and blood: avian parasitism andconservation. Trends Ecol Evol 10:232–235
Loye JE, Carroll SP (1998) Ectoparasite behaviour and its eVect onavian nest site selection. Ann Entomol Soc Am 91:159–163
Malmqvist B, Adler PH, Kuusela K, Merritt RW, Wotton RS (2004a)BlackXies in the boreal biome: key organisms in both terrestrialand aquatic environments. Ecoscience 11:187–200
Malmqvist B, Strasevicius D, Hellgren O, Adler PH, Bensch S (2004b)Vertebrate host speciWcity of wild-caught blackXies revealed bymitochondrial DNA in blood. Proc R Soc Lond B (Suppl)271:S152–S155
Martínez-de la Puente J, Merino S, Tomás G, Moreno J, Morales J,Lobato E, García-Fraile S (2007) Can the host immune systempromote multiple invasions of erythrocytes in vivo? DiVerentialeVects of medication treatment and host sex in a wild malaria-likemodel. Parasitology 134:651–655
Mellor PS, Boorman J, Baylis M (2000) Culicoides biting midges:their role as arbovirus vectors. Annu Rev Entomol 45:307–340
Merino S, Potti J (1995a) High prevalence of hematozoa in nestlings ofa passerine species, the pied Xycatcher (Ficedula hypoleuca). Auk112:1041–1043
Merino S, Potti J (1995b) Mites and blowXies decrease growth and sur-vival in nestling pied Xycatchers. Oikos 73:95–103
Merino S, Potti J, Fargallo JA (1997) Blood parasites of some passer-ine birds from central Spain. J Wildl Dis 33:638–641
Merino S, Moreno J, Potti J, De León A, Rodríguez R (1998) Nest ec-toparasites and maternal eVort in pied Xycatchers. Biol Cons Fau-na 102:200–205
Merino S, Moreno J, Sanz JJ, Arriero E (2000) Are avian blood para-sites pathogenic in the wild? A medication experiment in blue tits(Parus caeruleus). Proc R Soc Lond B 267:2507–2510
Mullens BA, Cardona CJ, McClellan L, Szijj CE, Owen JP (2006)Culicoides bottimeri as a vector of Haemoproteus lophortyx toquail in California, USA. Vet Parasitol 140:35–43
Naugle DE, Aldridge CL, Walker BL, Cornish TE, Moynahan BJ,Holloran MJ, Brown K, Johnson GD, Schmidtmann ET, MayerRT, Kato CY, Matchett MR, Christiansen TJ, Cook WE, Creek-more T, Falise RD, Rinkes ET, Boyce MS (2004) West Nile virus:pending crisis for greater sage-grouse. Ecol Lett 7:704–713
Ojanen U, Rätti O, Adler PH, Kuusela K, Malmqvist B, Helle P(2002) Blood feeding by black Xies (Diptera: Simuliidae) onthe black grouse (Tetrao tetrix) in Finland. Entomol Fenn13:153–158
Poulin R (1991) Group-living and infestation by ectoparasites in passe-rines. Condor 93:418–423
Quinn GP, Keough MJ (2002) Experimental design and data analysisfor biologists. Cambridge University Press, Cambridge
Rätti O, Ojanen U, Helle P (2006) Increasing group size dilutes blackXy attack rate in black grouse. Ornis Fenn 83:86–90
Rohner C, Krebs CJ, Hunter DB, Currie DC (2000) Roost site selectionof great horned owls in relation to black Xy activity: an anti-para-site behaviour? Condor 102:950–955
Smith RN, Cain SL, Anderson SH, Dunk JR, Williams ES (1998)BlackXy-induced mortality of nestling red-tailed hawks. Auk115:368–375
SutcliVe JF (1986) Black Xy host location: a review. Can J Zool64:1041–1053
Tomás G, Merino S, Moreno J, Sanz JJ, Morales J, García-Fraile S(2006) Nest weight and female health in the blue tit (Cyanistescaeruleus). Auk 123:1013–1021
Tomás G, Merino S, Moreno J, Morales J, Martínez-de la Puente J(2007a) Impact of blood parasites on immunoglobulin level andparental eVort: a medication Weld experiment on a wild passerine.Funct Ecol 21:125–133
Tomás G, Merino S, Moreno J, Morales J (2007b) Consequences ofnest reuse for parasite burden and female health and condition inblue tits, Cyanistes caeruleus. Anim Behav 73:805–814
Tomás G, Merino S, Martínez-de la Puente J, Moreno J, Morales J,Lobato E (2008) A simple trapping method to estimate abun-dances of blood-sucking Xying insects in avian nests. Anim Be-hav 75:723–729
Tripet F, Richner H (1997) Host responses to ectoparasites: food com-pensation by parent blue tits. Oikos 78:557–561
Valkiínas G, Iezhova TA (2004a) The transmission of Haemoproteusbelopolskyi (Haemosporida: Haemoproteidae) of blackcap byCulicoides impunctatus (Diptera: Ceratopogonidae). J Parasitol90:196–198
Valkiínas G, Iezhova TA (2004b) Detrimental eVects of Haemo-proteus infections on the survival of biting midge Culicoidesimpunctatus (Diptera: Ceratopogonidae). J Parasitol 90:194–196
Valkiínas G, Liutkevicius G, Iezhova TA (2002) Complete develop-ment of three species of Haemoproteus (Haemosporida, Haemo-proteidae) in the biting midge Culicoides impunctatus (Diptera,Ceratopogonidae). J Parasitol 88:864–868
Votýpka J, Oborník M, Volf P, Svobodová M, Luken J (2002) Trypan-osoma avium of raptors (Falconiformes): phylogeny and identiW-cation of vectors. Parasitology 125:253–263
Wasylik A (1971) Nest types and the abundance of mites. Ekol Polska19:689–699
Weinmann CJ, Murphy K, Anderson JR, DeMartini JC, LonghurstWM, Connolly G (1979) Seasonal prevalence, pathology, andtransmission of the quail heartworm, SplendidoWlaria californi-ensis (Wehr and Herman, 1956), in northern California. Can JZool 57:1871–1877
Yuval B (2006) Mating systems of blood-feeding Xies. Annu RevEntomol 51:413–440
123




















