
Kv3.1b and Kv3.3 channel subunit expression in murine spinal dorsal horn GABAergic interneurones
Upload
independentCategory
view
0download
0
Brain Research, 275 (1983) 1-11 1 Elsevier
Research Reports
Depression of Activities of Dorsal Horn Convergent Neurones by Propriospinal Mechanisms Triggered by Noxious Inputs; Comparison with Diffuse Noxious
Inhibitory Controls (DNIC)
SAMUEL W. CADDEN*, LUIS VILLANUEVA, DJAMEL CHITOUR* * and DANIEL LE BARS* * *
Unit# de Recherches de Neurophysiologie Pharmacologique, de I'INSERM (U. 161), 2 rue d'Al#sia, 75014 Paris (France)
(Accepted February 1st, 1983)
Key words: dorsal horn neurons - - propriospinal neurons - - nociception - - inhibition
The ability of heterotopic noxious stimuli to inhibit the activity of dorsal horn convergent neurones was investigated in both intact anesthetized, and spinal unanesthetized rats.
Forty-four convergent neurones in lumbar dorsal horn were recognized by their ability to respond to both noxious and non-noxious natural stimuli and by their characteristic responses corresponding to A- and C-fibre activity following electrical stimulation of their cutaneous excitatory receptive fields on the ipsilateral hindpaw. The application of a sustained pinch to the excitatory receptive field resulted in an initial phasic activation of the neurone, which adapted to a stable tonic level of activity (mean 31.8 + 2.2 spikes/s). The levels of activity produced in this fashion were not appreciably different between the two types of preparation.
In the intact anesthetized rat, the tonic activity produced by the sustained pinch could be strongly depressed by noxious conditioning stimuli applied to various parts of the body for all 10 neurones studied: heating the tail or pinching the contralateral hindpaw, the tail or a forepaw during 30 s each resulted in comparable inhibitions which had mean values in the order of 80% and which were always marked by post-effects lasting for upwards of 30 s. These inhibitory effects have been called Diffuse Noxious Inhibitory Controls (DNIC).
In the spinal unanaesthetized rat, the tonic activity was depressed to some extent by the same conditioning stimuli, for only 16/34 neurones studied. By comparison with the intact animals these inhibitions were weak, adapted to base-line levels within 30 s and were more marked for conditioning stimuli applied to structures proximal (tail, contralateral hindpaw) to the excitatory receptive field than for stimuli applied more distally (forepaws).
The differences between the inhibitions found in the intact and spinal preparations were subsequently confirmed in a series of exper- iments in which single convergent neurones were studied before and after the pharmacological blocking of the cervical spinal cord in anaesthetized rats.
The results in the spinal preparations provide evidence for the existence of some propriospinal modulatory processes, triggered by the onset of noxious stimulation and acting on convergent neurones. These processes appear to be different from those mediating DNIC, which have been shown to involve supraspinal structures, to concern all convergent neurones, to be very potent and associated with long-lasting post-effects whether the conditioning noxious stimuli are applied to parts of the body proximal or distal to the excita- tory receptive field.
INTRODUCTION
Prev ious e x p e r i m e n t s in this l abo ra to ry have
shown that nox ious s t imula t ion o f w idesp read areas
of the body can depress the act ivi ty of s o m e neu rones
in the spinal dorsa l ho rn o f the rat 12 (see also refs. 2,
3, 9-11) . This p h e n o m e n o n which has been cal led
Diffuse Nox ious Inh ib i to ry Con t ro l s ( D N I C ) , is
charac te r ized by its s t rong efficacy, its long-last ing
post -effects and its abil i ty to affect a lmos t all conver -
gent neu rones (i .e. those n e u r o n e s which can be ex-
ci ted by bo th noxious and non-nox ious st imuli) , but
* Present address: Department of Physiology, New Medical and Dental Building, The University, Leeds LS2 9NQ, U.K. ** Present address: Laboratoire de Physiologic, Institut des Scienes M6dicales, Alger, Alg6rie.
*** To whom correspondence should be addressed.
0006-8993/83/$03.00 (~ 1983 Elsevier Science Publishers B.V.

not other types of neurone in the dorsal horn 12.13. We have previously reported that DNIC are not found in
anaesthetized rats following cervical section of the spinal cord 13. However, subsequent studies by Ger- hart et al. 7 in the monkey and by Fitzgerald5 in the rat
have revealed the ability of remote noxious stimuli to
depress to some extent the activity of some dorsal horn neurones even following spinal section. We have therefore re-investigated this possibility using unanaesthetized (decerebrate) spinal rats.
For simplicity, we have restricted the study to in-
vestigate the activity of convergent neurones and in particular, the activity produced by the application of a sustained pinch to the excitatory receptive field of
the neurone. The relatively low levels of activity pro- duced in this fashion are, in principle, easier to inhibit than phasic activity such as that resulting from elec- trical or noxious thermal stimulation; furthermore,
the tonic activity provides a steady good baseline against which the extent and time course of any inhi- bitions can be monitored. It may be noted that activ- ity produced in a similar fashion was investigated in the other studies mentioned above 5,7.12.13. The re-
striction of the study to convergent neurones was made since previous experience in this laboratory had shown that these were the only neurones for which a stable level of tonic activity could be pro- duced by pinching the receptive field and also since DNIC apply almost exclusively to these neurones.
In order to make a direct comparison between any effects found in spinal preparation, and the type of inhibition already described for the intact animal, we also studied a small number of convergent neurones in anaesthetized intact rats, using a similar, strict pro- tocol. In addition, results are presented from a sepa- rate series of experiments in which the relevant prop- erties of single convergent neurones were studied be- fore and after the blocking of activity in the cervical
spinal cord of anaesthetized rats.
MAI"ERIALS AND METHODS
The experiments were performed on 25 male Spra- gue-Dawley rats weighing 200-300 g. in general, the preparation of the animals was as previously de- scribed 12 and therefore, in the present paper only a
brief account will be given. In the initial stages of the experiments, the animals
were deeply anaesthetized with 2% halothane in a mixture of nitrous oxide and oxygen (0.66:(1.33), and
following cannulation of the trachea and a jugular vein, the lumbar spinal cord was exposed with the ap- propriate vertebrae being mounted into a rigid
frame. Thereafter, in 14 experiments, the spinal cord was sectioned at the level of the rostral border of the second cervical vertebra, an ischaemic decerebration was performed and the anaesthetic regime was dis- continued; in the remaining experiments, the spinal
cord was left intact, no decerebration was performed and anaesthesia was maintained throughout the ex- periments with the level of halothane reduced to 0.5%. In neither group of animals were recordings
made sooner than 1 h after the cessation, or diminu- tion of the level, of anaesthesia.
The animals were paralyzed by gallamine triethio- dide and were artificially ventilated, the rate and vol- ume being adjusted to maintain a normal acid-base equilibrium 6. The electrocardiogram was monitored
throughout the experiments and the central tempera- ture was maintained at 37 _+ 0.5 °C by means of a thermostatic blanket system. Special attention was paid to the state of the peripheral circulation by con- sidering the colour of the hindpaw extremities and their ability to regain normal colour after the applica- tion of pressure.
Extracellular unitary recordings were made using glass micropipette electrodes filled with a mixture of 5% NaC1 and pontamine sky blue dye (resistance 8-20 Mr2). The cord was stabilized during the record-
ing sessions by a covering of Ringer agar gel. Convergent neurones were identified by their re-
sponses to stimuli applied to their excitatory recep- tive fields on the ipsilateral hindpaw. All the conver- gent neurones studied: (a) responded to innocuous and noxious mechanical stimuli and to noxious ra- diant heat applied to their receptive fields; and (b) gave responses with latencies correspoding to A- and C-fibre inputs following transcutaneous electrical stimulation of their receptive fields 16.
The experimental protocol consisted of producing a tonic discharge from an identified convergent neu- rone and then testing the effects of noxious condi- tioning stimuli applied to various parts of the body. The discharge was produced by applying a small ar- tery clip to the center of the excitatory receptive field of the neurone: the resulting response (see Fig. 1)

consisted of an initial phasic component which gener- ally adapted within 1-2 min, and was followed by a tonic discharge which remained relatively constant throughout the period of stimulation. In order to al- low comparison between results obtained in spinal and intact animals, the conditioning stimuli were ap- plied according to a strict protocol as represented schematically in Fig. 1; no conditioning stimuli were applied during the first 3 min following the applica- tion of the artery clip to allow for the adaptation of the phasic component of the evoked discharge. Thereafter, conditioning stimuli were applied for pe- riods of 30 s every 2.5 min in the spinal animals and every 4 min in the intact animals; the larger interval was employed in the intact animals because of the long-lasting post-effects which had previously been observed following noxious conditioning stimuli in this type of preparation (see Introduction). The con- ditioning stimuli consisted of: (a) noxious heat ap- plied to the distal 2/3 of the tail by means of immer- sion in hot water (52 °C); (b) noxious calibrated pinch applied to the contralateral hindpaw; (c) nox- ious calibrated pinch applied to the tail; and (d) nox- ious calibrated pinch applied to either forepaw. The noxious pinch stimuli were applied in a standard fash- ion using a pair of non-toothed forceps. In some ex- periments, after completing these standard tests, non-noxious conditioning stimuli (brushing, rubbing movements) were als0. applied both close to the excit- atory receptive fields and to the same areas which had been tested with the noxious conditioning stimu- li.
In an additional 5 experiments in which the prop- erties of single convergent neurones were studied be- fore and after blocking the cervical spinal cord, the preparation and protocol were generally similar to those described above for the experiments on intact anaesthetized rats except that: (1) the cervical spinal cord (C1-2) was exposed at an early stage of the ex- periments for subsequent injection of xylocaine (25 gl, 5%) and surgical section; and (2) the only condi- tioning stimuli employed were, immersion of the tail in hot water (52 °C) and noxious pinch of the contra- lateral hindpaw.
In all the experiments the evoked discharges were recorded using conventional techniques. The inte- grated spike-discharges were subsequently analyzed and any effects of the conditioning procedures were
calculated as the percentage changes in the activity
compared with a 30 s control period immediately pre-
ceding the conditioning stimulus. The location of neurones was marked by electro-
phoretic deposition of pontamine sky blue from the microelectrodes for subsequent histological recon- struction of electrode tracts.
RESULTS
A total of 49 convergent neurones (15 in intact, and 34 in spinal animals) were studied; the results for the 5 neurones in anaesthetized rats, which were studied before and after blocking activity in the cervi- cal spinal cord, are presented separately at the end of this section.
The locations of the cells were found to lie in the deeper layers of the dorsal horn (laminae IV-VI); all the neurones responded[ ' to both innocuous and nox- ious stimulation of their cutaneous receptive fields, and gave responses corresponding to A- and C-fibre inputs following transcutaneous electrical stimula- tion. The application of a small artery clip to the cuta- neous receptive field always produced an initial pha- sic component followed by a tonic discharge from the neurones (see Figs. 1, 2 and 4) with the mean level of activity being 31.8 + 2.2 spikes/s (as determined dur- ing the 30 s period immediately preceding the first conditioning stimulus). In these patterns of re- sponses, ther~ were no obvious differences between the neurones studied in the intact, and those studied in the spinal animals. Indeed, the mean levels of ac- tivity produced by application of the artery clip were very similar for the two groups of animals: 33.5 + 3.2 spikes/s for cells in intact animals: 31.3 + 2.6 spikes/s for cells in spinal animals.
Some of the conditioning stimuli were found to af- fect this kind of activity in both types of preparation. Non-noxious conditioning stimuli were only effective when applied in the vicinity of, and generally adja- cent to, the excitatory receptive field; again, no ob- vious differences were seen between the two groups of animals. By contrast, potent inhibitory effects were obtained by the application of noxious stimuli to widespread parts of the body and these effects dif- fered markedly between the two groups of animals.

Hz !00 -
poriplWel ,11----~ excitatory '
an c b d
60-
0 - 6 10
'1 16 2O N N I 16
Fig. 1. Example of inhibitory effects induced by various noxious stimuli in the intact anaesthetized rat. The application of sustained pinch (open arrowhead, broken line) to the peripheral excitatory field of the convergent neurone resulted in a phasic response fol- lowed by a tonic discharge (bin width; 2 s). Various noxious conditioning stimuli (filled arrowheads, solid lines; a: immersion of the tail in a 52 °C water bath; b: pinch of the contralateral hindpaw; c: pinch of the tail; d: pinch of a forepaw) induced strong and long-lasting inhibitory effects.
Effects of noxious conditioning stimuli in intact ani- mals
The experiments in the anaesthetized intact ani- mals were per formed to provide a control group for comparison with the results obtained in the unanaes- thetized spinal preparations. The results obtained us- ing the present protocol were essentially similar to those which have previously been reportedl0,]2. As shown for an individual example (Fig. 1), noxious stimuli applied to widespread areas of the body in- duced strong and long-lasting inhibitory effects: the
evoked discharge of this neurone was reduced to 20.7%, 5.3%, 30.3% and 14.7% of its control values during the immersion of the tail in hot water (52 °C) and during the application of pinches to the contralat- eral hindpaw, the tail and the contralateral forepaw, respectively. Note that, on each occasion, the inhibi- tory effects outlasted the period of conditioning; these 'post-effects ' were particularly marked (lasting several minutes) following the application of noxious heat to the tail (Fig. 1 'a ' ) and of noxious pinch to the
contralateral forepaw (Fig. 1 'd ') .

Consistent with the above example, the activity of
all 10 neurones under study was strongly depressed
by the application of each of the conditioning stimuli.
For individual neurones the inhibitions produced
during the application of the different conditioning
stimuli were often of different magnitudes; however,
the pooled results summarized in Fig. 3 (left) show
that, overall, these inhibitions were of the same or-
der. It is also clear from Fig. 3 (left) that the inhibi-
tions required a certain time to reach their maximal
effect (within 10 s) which then remained fairly con-
stant until the end of the conditioning period and which were consistently followed by post-effects of
long duration.
HZ
A '° ] |
0 5 I|
B" t 0 0 6 10
c ,.°] 0
u 5 10
°'t 0 u 5 m m !o
Fig. 2. Examples of tonic discharges from 4 different convergent neurones (A, B, C, D) recorded in spinal unanaesthetized rats, against which various conditioning stimuli were tested (symbols and abbreviations as in Fig. 1). Depressions of activity were obtained with all 4 conditioning stimuli for neurone A, with 3 for neurone B, and with only I for neurone C. Note in every case the rapid recov- ery of the neuronal activity. Neurone D was not affected by any of those conditioning stimuli.

Effects of noxious conditioning stimuli in spinal ani- mals
In contrast with the inhibitions which were found
for all convergent neurones in the intact animals, the
activities of only 16 of 34 such neurones studied in un-
anaesthetized spinal preparations, were depressed
I anaesthetized intact rats
IO~
by one or more of the noxious condit ioning stimuli.
Of these 16 neurones, only 4 were affected by all 4
conditioning stimuli, 6 by 3 of the condit ioning stimu-
li, 3 by 2 of the conditioning stimuli and the remaining
3 by only one conditioning stimulus. It may be noted
that no significant differences were observed be-
J unanaesthetized [ spinal rats I
pinch centralateral hindpaw 1
Ioo
o 0 1 2 3 m l f l 0 I
pinch tail
52"C--tail 0 1 2
!oo
0
pinch forepaw
lo(
1 2
0 (
Fig. 3. Histograms summarizing data obtldl~l in intact (left) al~Slli~d(x!llht) rats. TI~ pooled data shown were obtained after nor- malizing individual results with respect to the mean levels of activity recorded during the 30 s immediately preceding each conditioning stimulus (bin width: 10 s). Individual results were used only when an inhibitory effect was found for the particular conditioning stimu- lus (numbers shown as proportions of total numbers of neurones in bottom right hand comer of each histogram). Note that for the in- tact animals all the neurones were inhibited by all the conditioning stimuli whereas this was not the case for the spinal animals (see Text). The hatched areas indicate the periods of application of the various conditioning stimuli. Bars indicate standard errors of the mean.
tween the mean levels of activity of the neurones
which were inhibited by all of the conditioning stimuli (31.8 + 4.8 spikes/s), f rom those which where inhib- ited by some of the conditioning stimuli (30.1 + 4.4 spikes/s) or even from those unaffected by the condi- tioning stimuli (32.0 + 4.0 spikes/s). Examples of re- sults obtained from 4 neurones in spinal preparat ions
are shown in Fig. 2. Fig. 2A, B and C illustrate the ac- tivities of cells which were depressed respectively by 4, 3 and 1 of the conditioning stimuli. It can be seen from these examples that the inhibitions, when pre-
sent, were relatively weak and generally of shorter duration than the 30 s conditioning period. The pooled results for cells exhibiting inhibitions by each of the conditioning stimuli are illustrated in Fig. 3
(right) in which it can be seen that these inhibitions were most marked immediately following the appli-
cation of the conditioning stimulus. Of the 4 conditioning stimuli, noxious pinch of the
contralateral hindpaw most frequently produced in- hibitions (14 cells). Noxious stimulation of the tail af-
fected a comparable number of cells (noxious pinch, 11 cells; noxious heat, 13 cells) whereas pinching a
forepaw only affected 5 cells (ipsilateral forepaw, 2 cells; contralateral forepaw, 3 cells). This order of ef-
ficacy was reflected also by the extent of the inhibi- tions observed in those cells affected by more than
one conditioning stimulus. As an example, for the 4
neurones whose activities were depressed by all the conditioning stimuli, noxious pinch of the contralat-
Hz
'°° 1 50-
O- 5 10 15
100 -
50-
0-
-D
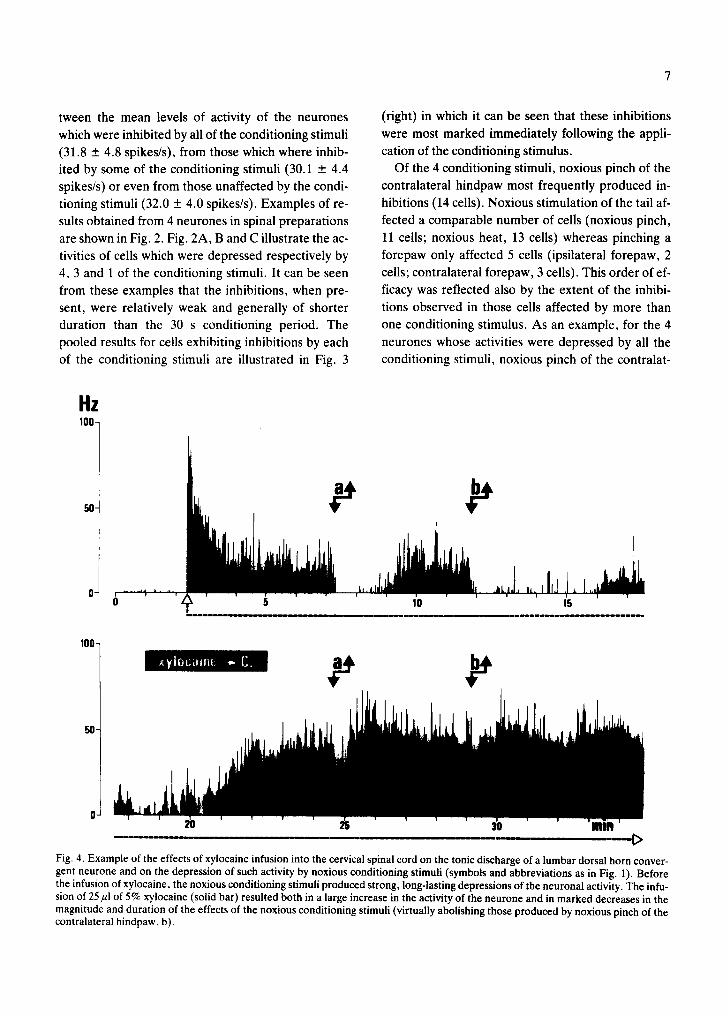
Fig. 4. Example of the effects of xylocaine infusion into the cervical spinal cord on the tonic discharge of a lumbar dorsal horn conver- gent neurone and on the depression of such activity by noxious conditioning stimuli (symbols and abbreviations as in Fig. 1). Before the infusion of xylocaine, the noxious conditioning stimuli produced strong, long-lasting depressions of the neuronal activity. The infu- sion of 25/zl of 5% xylocaine (solid bar) resulted both in a large increase in the activity of the neurone and in marked decreases in the magnitude and duration of the effects of the noxious conditioning stimuli (virtually abolishing those produced by noxious pinch of the contralateral hindpaw, b).

eral hindpaw reduced the activity during the condi-
tioning period to 53.2 + 8.9% of its control value whereas noxious pinch of the forepaw only reduced the activity to 68.0 + 5.0%.
Another trend in the extent of the inhibitory ef- fects was the direct relationship between the number of conditioning stimuli which affected a cell and the
degree of the inhibitions produced: the activity of neurones affected by all 4 conditioning stimuli was reduced to a mean level of 63.0 + 6.3% during the
conditioning periods while the corresponding figures for neurones affected by 3, 2 and f conditioning stim- ulus were 67.0 + 5.9%, 72.8 + 3.0% and 84.5 +
2.3%, respectively, during the application of the ef- fective conditioning stimuli.
The marked differences between the inhibitions
observed in the intact and spinal animals can be seen by comparing Fig. 1 with Fig. 2 and left part with right part of Fig. 3. It must be noted that Fig. 3 (right) shows the pooled results only for neurones which were inhibited by each particular conditioning stimu- lus and thus effectively reduced the contrast between these results and those presented in Fig. 3 (left) which includes all the results obtained from intact an-
imals. It is clear that the following differences existed be-
tween the two types of inhibitory effects: (a) the max-
imal effects of the conditioning stimuli were obtained generally after several seconds in the case of intact animals whereas these were achieved almost imme- diately following the application of the conditioning stimuli in the spinal animals; (b) both the maximal and mean inhibitory effects were generally much greater in the intact, than in the spinal animals; (c) the inhibitory effects in the intact animals were gen- erally sustained throughout the conditioning period whereas in the spinal animals the effects began to adapt, normally within the first 10 s, and with few ex- ceptions had disappeared before the end of the con- ditioning period and (d) the inhibitions in the intact animals always outlasted the period of conditioning, whereas such post-effects were never observed in the spinal animals.
Comparison of the effects of noxious conditioning stimuli before and after cervical spinal block
Subsequent to the experiments described above, we investigated, in the anaesthetized rat, the prop-
erties of single convergent neurones before and after blocking activity in the cervical spinal cord by the in-
fusion of the local anaesthetic, xylocaine. Typical re- sults from one such experiment are presented in Fig.
4 in which it can be seen that following the infusion of 25/~1 of 5% xylocaine at the C2 level, the inhibitory effects produced by the immersion of the tail in hot
water were greatly reduced, and those produced by noxious pinch of the contralateral hindpaw virtually abolished.
In all 5 such preparations, the conditioning stimuli initially produced powerful inhibitions but following the infusion of xylocaine at C2, clear inhibitory ef- fects remained only for one cell during the applica- tion of hot water to the tail and for 4 cells during nox-
ious pinch of the contralateral hindpaw; furthermore the remaining inhibitions were greatly reduced in both magnitude and duration. These results are therefore in general agreement with those which we have presented above and, indeed, subsequent sec- tion of the cervical cord had no further effect on these inhibitions.
However, it may be noted from Fig. 4 that the infu- sion of xylocaine also resulted in a large rise in the ac- tivity of the neurone, a finding which was obtained in all 5 preparations and which is at variance with our previous observation that the application of the ar- tery clip produced similar levels of activity in spinal and intact animals. The most likely explanation for this apparent discrepancy is that it resulted from a difference between the immediate and longer term effects of blocking activity in the cervical spinal cord (see also ref. 8).
DISCUSSION
The results presented above provide evidence that the activity of dorsal horn convergent neurones can be depressed by noxious stimulation of widespread areas of the body, not only in the intact animal but al.so in spinal preparations. However, there were no- ticeable differences between the effects observed in these types of preparations in terms of potency, pat- tern and time-course.
The data we have obtained in intact anaesthetized rats were similar to those we previously reported!0.~2: the inhibitory effects were consistently found for all convergent neurones regardless of the site of the con-

ditioning stimulus; these effects were powerful, often
completely silencing the neuronal activity, and were always marked by long-lasting post-effects.
By comparison, the inhibitions in spinal unanaes- thetized rats were seen for less than half of the con- vergent neurones studied and were more often pro- duced by stimulation of parts of the body proximal to the excitatory receptive field (contralateral hindpaw, tail) than by stimulation of more distal structures (forepaws). These effects were less powerful than in the intact animal, especially during stimulation of dis- tal structures, generally adapted rapidly and were never followed by post-effects. In addition, it was found that, while inhibitions could repeatedly be pro- duced for a given cell, the strength of these inhibi- tions exhibited a noticeable variability.
These findings in the spinal preparation are in keeping with the recent reports of Fitzgerald5 in the rat and of Gerhart et al. 7 in the monkey. Our results are in particular agreement with those of Fitzgerald 5 who, using essentially the same preparation, also found that approximately half the convergent neu- rones were inhibited by noxious, but not by non-nox- ious stimulation of the contralateral hindpaw or tail; furthermore, the time course and extent of the inhibi- tions described by Fitzgerald were similar to those found in the present study and by Gerhart et al. 7 in the spinal anaesthetized monkey. Gerhart et al. also reported that the inhibitions resulting from the nox- ious stimulation of the forepaw were clearly weaker than those evoked by stimuli applied more proximal to the excitatory receptive field of the neurone, which is again consistent with our results. However, it may be noted that Gerhart et al. obtained inhibi- tions for all 6 cells which they studied in spinal prepa- rations; in addition they reported similar patterns of inhibition in both spinal and intact monkeys in that these inhibitions were always of relatively short dura- tion, adapted and generally had no post-effects (re- sembling those which we observed in the spinal rat). The contrast between our results in the intact rat and those of Gerhart et al. in the monkey may be due to species and/or methodological (e.g. anaesthetic re- gimes) differences.
In a previous study13 we found no evidence for in- hibitions resulting from noxious stimulation of wide- spread areas of the body in the spinal anaesthetized rat. The findings of the present experiments using the
spinal unanaesthetized preparation, together with
those subsequently obtained in spinal anaesthetized animals suggest that the sample tested in the previous experiments may have been too small; in those ear- lier experiments, the effects of distant noxious stimu- li were tested against the activity evoked by a sus- tained pinch for only 5 convergent neurones and it should be noted that in the present experiments 18/39 convergent neurones were unaffected by such condi- tioning stimuli. In the previous study we also investi- gated the effects of the conditioning procedures against activities produced by electrical stimulation (18 cells), without finding any obvious inhibitions. It is probable that the synchronized volleys resulting from such stimuli resulted in a greater degree of syn- aptic safety and hence the responses were less ame- nable to any inhibitory effects. This is consistent with an additional finding in some of our present experi- ments that, for cells in spinal animals for which we had already found inhibitions of the activity evoked by tonic pinch, there was little or no evidence of inhi- bitions of the responses evoked by an electrical stim- ulus sufficient to excite both A- and C-fibres. It would appear from the results illustrated by Fitzge- rald 5, that she also found that the responses evoked by electrical stimuli were more difficult to modulate than those evoked by sustained pinch. This insuscep- tibility of the responses evoked by electrical stimuli, to modification by remote noxious stimuli, is a fur- ther clear difference between the properties of con- vergent neurones in the intact and spinal prepara- tions.
With regard to the mechanisms responsible for the inhibitions reported here, we have discussed else- where 2,3,1°-13 some of those responsible in the intact
animal. In the spinal animal, it is necessary to pro- pose some form of propriospinal mechanism (see ref. 7); although our present data are not helpful in fur- ther defining such a system, there is both anatomi- cal 1,15,18-22 and electrophysiological20 evidence that
the neural pathways exist, at least in other species, whereby such mechanisms might be mediated. Due to the fact that the inhibitions which we observed in the spinal animal, were poorly reproducible at a quantitative level, it has not been feasible to date, to investigate pharmacologically the neural mech- anisms which are involved. Nor on the basis of pre- sent evidence are we able to postulate whether the fi-

10
nal mechanisms in these inhibitions are pre- or post-
synaptic. In view of the marked differences which we
have described above, between the inhibitions ob-
served in intact animals and those observed in spinal
animals, it is most likely that the mechanisms in-
volved are different. It is, however, possible that the propriospinal system which we have proposed above,
and the system responsible for the inhibitions in the
intact animal, may share some common neuronal el-
ements.
It may be stressed that, from a methodological
point of v iew, it is highly unlikely that the results of
previous experiments in which we investigated the phenomenon of DNIC2, 3.9-13 were influenced by the
inhibitions which we have now observed in the spinal
animal. The experiments on D N I C have generally
employed electrical stimulation as a means of excit-
ing convergent neurones and, under these circum-
stances, as previously reported 13 and confirmed
above, it is very difficult to demonstrate inhibitory ef-
fects in spinal animals.
We have discussed in some detail elsewhere, the possible functional roles of D N I C in sensori-discrimi-
native processes, including a possible role in the pro-
duction of analgesia by certain counter-irritation pro- cedures 13. Similar roles may now be proposed for the
inhibitory mechanisms whose effects we have dem-
onstrated in the spinal animal. It is also possible that
these types of inhibitions might be involved in some
reflex mechanisms, and indeed, the brief duration of
the effects seen in the spinal animals, together with
the relatively short pathways by which they are pre-
sumably mediated, tend to suggest such a role. If this
were the case, then the present results would be in
good agreement with reports that, in the spinal cat,
the flexor reflex of the hindlimb can be inhibited by
contralateral nerve stimulation 4 and that polysynap-
tic but not monosynaptic reflexes in forelimb muscles
can be inhibited by stimulation of the hindlimbslT;
furthermore in this latter example, it was reported
that the inhibition could be produced by natural stim-
uli, only when these were of an intensity sufficient to
produce pain in man (again cf. present study). The
possibility that the inhibitions which we have report-
ed for intact and spinal rats, may be involved both in
sensori-discriminative and in reflex mechanisms,
gains some support from the results of a recent study in man in which it was found that the pain and a noci-
ceptive flexor reflex thresholds were similarly raised
during noxious conditioning stimulation of hetero-
topic parts of the body TM.
ACKNOWLEDGEMENTS
We wish to thank Mr. H. de Pommery for the pho-
tography, Miss M. Hoch for secretarial help and Dr.
Ji M. Besson for advice in the preparation of this manuscript. S.W.C. was supported by a Royal Socie-
ty European Science Exchange Programm fellowship
in conjunction with the Centre National de la Re-
cherche Scientifique (CNRS). L.V., on leave from
the Department of Pharmacology of the University of Chili, was supported by a scholarship from the
French government. This work was supported by l 'Institut National de la Sant6 et de la Recherche
M6dicale ( INSERM), PRC no. 120034.
REFERENCES
1 Burton, H. and Loewy, A. D., Descending projections from the marginal cell layer and other regions of the mon- key spinal cord, Brain Research, 116 (1976) 485-491.
2 Chitour, D., Dickenson, A. H. and Le Bars, D., Pharma- cological evidence for the involvement of serotonergic mechanisms in Diffuse Noxious Inhibitory Controls (DNIC), Brain Research, 236 (1982) 329-337.
3 Dickenson, A. H., Rivot, J. P., Chaouch, A., Besson, J. M. and Le Bars, D., Diffuse Noxious Inhibitory Controls (DNIC) in the rat with or without pCPA pretreatment, Brain Research, 216 (1981) 313-321.
4 Eccles, J. C. and Sherrington, C., Studies on the flexor re- flex. VI - - Inhibition, Proc. Roy Soc., B, 109 (1931) p. 91-113.
5 Fitzgerald, M., The contralateral input to the dorsal horn of
the spinal cord in the decerebrate spinal rat, Brain Re- search, 236 (1982) 275-287.
6 Freminet, A., Bursaux, E. et Poyart, C., Mesure de la vi- tesse de renouvellement du lactate chez le rat par perfusion de 14 CU (L) Lactate +, Pflfigers Arch. ges. Physiol., 334 (1972) 293-302.
7 Gerhart, K. D., Yezierski, R. P., Giesler, Jr., G. J. and Willis, W. D., Inhibitory receptive fields of primate spino- thalamic tract cells, J. Neurophysiol., 46 (1981) 1309-1325.
8 Le Bars, D. and Chitour, D., Do convergent neurones in the spinal dorsal horn discriminate nociceptive from non- nociceptive information?. Pain, in press.
9 Le Bars, D., Chitour, D. and Clot, A. M., The encoding of thermal stimuli by Diffuse Noxious Inhibitory Controls (DNIC), Brain Research, 230 (1981) 394--399.
10 Le Bars, D., Chitour, D., Kraus, E., Clot, A. M., Dicken- son, A. H. and Besson, J. M., The effect of systemic mor-

phine upon Diffuse Noxious Inhibitory Controls (DNIC) in the rat: evidence for a lifting of certain descending inhibito- ry controls of dorsal horn convergent neurones, Brain Re- search, 215 (1981) 257-274.
11 Le Bars, D., Chitour, D., Kraus, E., Dickenson, A. H. and Besson, J. M., Effect of naloxone upon Diffuse Noxious In- hibitory Controls (DNIC) in the rat, Brain Research, 204 (1981) 387--402.
12 Le Bars, D., Dickenson, A. H. and Besson, J. M., Diffuse Noxious Inhibitory Controls (DNIC). I - - Effects on dorsal horn convergent neurpnes in the rat, Pain, 6 (1979) 283--304.
13 Le Bars, D., Dickenson, A. H. and Besson, J. M., Diffuse Noxious Inhibitory Controls (DNIC). II - - Lack of effect on non-convergent neurones, supraspinal involvement and theoretical implications, Pain, 6 (1979) 305-327.
14 Le Bars, D., Roby, A. and Wilier, J. C., Effects of hetero- topic thermal stimuli on a nociceptive flexion reflex and the related pain sensation in man, J. Physiol. (Lond.), 336 (1983) 31P-32P.
15 Matsushita, M., Ikeda, M. and Hosoya, Y., The location of spinal neurons with long descending axons (long descend- ing propriospinal tract neurons) in the rat: a study with the horseradish peroxidase technique, J. comp. Neurol., 184 (1979) 63--80.
16 Menetrey, D., Giesler, Jr., G. J. and Besson, J. M., An analysis of response properties of spinal cord dorsal horn
11
neurones to nonnoxious and noxious stimuli in the spinal rat, Exp. Brain Res., 27 (1977) 15-33.
17 Miller, S., Reitsma, D. J. and Van der Meche, F. G. A., Functional organisation of long ascending propriospinal pathways linking lumbosacral and cervical segments in the cat, Brain Research, 62 (1973) 169-188.
18 Molenaar, I. and Kuypers, H. G. J. M., Cells of origin of propriospinal fibers and of fibres ascending to supraspinal levels. An HRP study in the cat and rhesus monkey, Brain Research, 152 (1978) 429:-450.
19 Ramon y Cajal, S., Histologie du Syst~rne Nerveux de l'Homme et des Vertdbrds, Vol. l., Inst. Cajal, Madrid, 1909, reprinted 1972.
20 Skinner, R. D., Coulter, J. D., Adams, R. J. and Remmel, R. S., Cells of origin of long descending propriospinal fibers connecting the spinal enlargements in cat and monkey de- termined by horseradish peroxidase and electrophysiological techniques, J. comp. Neurol., 188 (1979) 443-454.
21 Szentagothai, J., Propriospinal pathways and their syn- apses. In J. C. Eccles and J. P. Schad~ (Eds.), Organization of the Spinal Cord, Progress in Brain Research, Vol. 11, Elsevier, Amsterdam, 1964, pp. 155-177.
22 Yezierski, R. P., Culberson, J. L. and Brown, P. B., Cells of origin of propriospinal connections to cat lumbosacral gray as determined with horseradish peroxidase, Exp. Neu- rol., 69 (1980) 493-512.
Copyright © 2022 FDOKUMEN




















