
Delauny et al 2009(2)
10
Genetic Tracing of Subpopulation Neurons in the Prethalamus of Mice (Mus musculus) DELPHINE DELAUNAY, 1,2 KATHARINA HEYDON, 1,2 ANDRES MIGUEZ, 1,2 MARKUS SCHWAB, 3 KLAUS-ARMIN NAVE, 3 JEAN LEON THOMAS, 1,2 NATHALIE SPASSKY, 1,2 SALVADOR MARTINEZ, 4 AND BERNARD ZALC 1,2 * 1 Inserm, U711, Ho ˆpital de la Salpe ˆtrie `re, 75013 Paris, France 2 Universite ´ Pierre et Marie Curie, Faculte ´ de me ´decine, IFR70, 75013 Paris, France 3 Max-Planck-Institute of Experimental Medicine, D-37075 Goettingen, Germany 4 Experimental Embryology Laboratory, Instituto de Neurociencias Alicante, Campus de San Juan, San Juan de Alicante, 03550, Spain ABSTRACT Genetic labeling based on the Cre/lox reporter system has allowed the creation of fate maps for progenitor cells and their offspring. In the diencephalon, pools of progenitors express the plp transcripts in the zona limitans intrathal- amica (ZLI), the basal plate of the diencephalon (bpD), and the posterior part of the hypothalamus. We used plp-Cre transgenics crossed with either Rosa26-lox-lacZ (R26R) or actin-lox gfp (Z/EG) reporter mice to investigate the progeny of plp-expressing ventricular cells in the diencephalon. We describe the subpopulations of prethalamic neurons de- rived from plp-activated progenitors, their possible migra- tory routes as development proceeds, and their final posi- tional identity. Neurons derived from plp-expressing progenitors issued from the ZLI contribute to GABAergic cells in the zona incerta, the subgeniculate nucleus, the ventral lateral geniculate, and the intergeniculate leaflet. Plp progenitors in the bpD and posterior hypothalamus appear to generate glutamatergic neurons in the subtha- lamic nucleus and GABAergic neurons in the mammillary and retromammillary tegmentum derivatives. In all these nuclei the contribution of plp progenitors is only partial, illustrating the heterogeneity of origin of neurons in pretha- lamic and caudal hypothalamic nuclei. J. Comp. Neurol. 512: 74 – 83, 2009. © 2008 Wiley-Liss, Inc. Indexing terms: diencephalon; hypothalamus; intergeniculate leaflet; subgeniculate nucleus; subthalamic nucleus; ventral lateral geniculate nucleus; zona incerta; zona limitans intrathalamica In the adult central nervous system (CNS), the proteolipid protein (PLP) is the major protein of myelin and oligodendro- cytes (Folch and Lees, 1951). During embryonic development, expression of the corresponding transcript is detectable as early as embryonic day (E)9.5, i.e., at least 10 –15 days before any myelin is deposited around axons. In the developing em- bryo, plp mRNA has been detected in discrete ventricular foci distributed along the neural tube (Timsit et al., 1992, 1995; Spassky et al., 1998) and the plp transcript has been reported as a marker of a subpopulation of oligodendrocyte precursor cells (at E12.5–E14.5), that do not depend on PDGFR signal- ing for their survival and proliferation (Spassky et al., 2001a,b). In the diencephalon, we have recently shown that plp is ex- pressed first (at E9.5) by neuroepithelial cells and later (at E13.5) by radial glial cells (Delaunay et al., 2008). Using in vitro and in vivo complementary approaches, we have provided evidence that the plp progenitors of the diencephalon have a restricted neuronal fate of differentiation at E9.5, while they exhibit a restricted glial cell fate at E13.5. Therefore, we have proposed a two-step model for the specification of neural progenitors in the diencephalon, with neuronal progenitors (neuroblasts) segregating at E9.5 and glial progenitors (glio- blasts) appearing at E12.5–E13.5. Grant sponsor: DD was a fellow of the Ministe ` re de l’Enseignement Supe ´ rieur et de la Recherche and European Leukodystrophy Association; Grant sponsor: INSERM; Grant sponsor: Association de Recherche sur la Scle ´ rose En Plaques (to J.L.T.); Grant sponsor: National Multiple Sclerosis Society; Grant number: TR-3762-A-1 (to B.Z.); Grant sponsor: DFG (CMPB) and BMBF (to K.A.N.); Grant sponsor: EEC; Grant number: LSHG-CT-2004- 512003; Grant sponsor: EUREXPRESS; Grant numbers: ACOMP06/076, BFU2005-23722-E/BFI, MEC BFU2005-09085; Grant sponsor: ELA Fonda- tion de Recherche, France (to S.M.). The first two authors contributed equally to this work. *Correspondence to: B. Zalc, INSERM U711; UPMC, Biologie des Inter- actions Neurones/Glie, Hopital de la Salpetriere, 47, Bd de l’Hopital, 75651 Paris Cedex 13, France. E-mail: [email protected] Received 16 February 2007; Revised 28 April 2008; Accepted 27 Sep- tember 2008 DOI 10.1002/cne.21904 Published online in Wiley InterScience (www.interscience.wiley.com). The Journal of Comparative Neurology 512:74 – 83 (2009) © 2008 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Delauny et al 2009(2)

Genetic Tracing of Subpopulation Neurons in thePrethalamus of Mice (Mus musculus)
DELPHINE DELAUNAY,1,2 KATHARINA HEYDON,1,2 ANDRES MIGUEZ,1,2 MARKUS SCHWAB,3
KLAUS-ARMIN NAVE,3 JEAN LEON THOMAS,1,2 NATHALIE SPASSKY,1,2 SALVADOR MARTINEZ,4
AND BERNARD ZALC1,2*1Inserm, U711, Hopital de la Salpetriere, 75013 Paris, France2Universite Pierre et Marie Curie, Faculte de medecine, IFR70, 75013 Paris, France3Max-Planck-Institute of Experimental Medicine, D-37075 Goettingen, Germany4Experimental Embryology Laboratory, Instituto de Neurociencias Alicante, Campus de San Juan, San Juan de Alicante, 03550,Spain
ABSTRACTGenetic labeling based on the Cre/lox reporter system hasallowed the creation of fate maps for progenitor cells andtheir offspring. In the diencephalon, pools of progenitorsexpress the plp transcripts in the zona limitans intrathal-amica (ZLI), the basal plate of the diencephalon (bpD), andthe posterior part of the hypothalamus. We used plp-Cretransgenics crossed with either Rosa26-lox-lacZ (R26R) oractin-lox gfp (Z/EG) reporter mice to investigate the progenyof plp-expressing ventricular cells in the diencephalon. Wedescribe the subpopulations of prethalamic neurons de-rived from plp-activated progenitors, their possible migra-tory routes as development proceeds, and their final posi-
tional identity. Neurons derived from plp-expressingprogenitors issued from the ZLI contribute to GABAergiccells in the zona incerta, the subgeniculate nucleus, theventral lateral geniculate, and the intergeniculate leaflet.Plp� progenitors in the bpD and posterior hypothalamusappear to generate glutamatergic neurons in the subtha-lamic nucleus and GABAergic neurons in the mammillaryand retromammillary tegmentum derivatives. In all thesenuclei the contribution of plp� progenitors is only partial,illustrating the heterogeneity of origin of neurons in pretha-lamic and caudal hypothalamic nuclei. J. Comp. Neurol. 512:74–83, 2009. © 2008 Wiley-Liss, Inc.
Indexing terms: diencephalon; hypothalamus; intergeniculate leaflet; subgeniculate nucleus;subthalamic nucleus; ventral lateral geniculate nucleus; zona incerta; zonalimitans intrathalamica
In the adult central nervous system (CNS), the proteolipidprotein (PLP) is the major protein of myelin and oligodendro-cytes (Folch and Lees, 1951). During embryonic development,expression of the corresponding transcript is detectable asearly as embryonic day (E)9.5, i.e., at least 10–15 days beforeany myelin is deposited around axons. In the developing em-bryo, plp mRNA has been detected in discrete ventricular focidistributed along the neural tube (Timsit et al., 1992, 1995;Spassky et al., 1998) and the plp transcript has been reportedas a marker of a subpopulation of oligodendrocyte precursorcells (at E12.5–E14.5), that do not depend on PDGFR� signal-ing for their survival and proliferation (Spassky et al., 2001a,b).In the diencephalon, we have recently shown that plp is ex-pressed first (at E9.5) by neuroepithelial cells and later (atE13.5) by radial glial cells (Delaunay et al., 2008). Using in vitroand in vivo complementary approaches, we have providedevidence that the plp� progenitors of the diencephalon have arestricted neuronal fate of differentiation at E9.5, while theyexhibit a restricted glial cell fate at E13.5. Therefore, we haveproposed a two-step model for the specification of neural
progenitors in the diencephalon, with neuronal progenitors(neuroblasts) segregating at E9.5 and glial progenitors (glio-blasts) appearing at E12.5–E13.5.
Grant sponsor: DD was a fellow of the Ministere de l’EnseignementSuperieur et de la Recherche and European Leukodystrophy Association;Grant sponsor: INSERM; Grant sponsor: Association de Recherche sur laSclerose En Plaques (to J.L.T.); Grant sponsor: National Multiple SclerosisSociety; Grant number: TR-3762-A-1 (to B.Z.); Grant sponsor: DFG (CMPB)and BMBF (to K.A.N.); Grant sponsor: EEC; Grant number: LSHG-CT-2004-512003; Grant sponsor: EUREXPRESS; Grant numbers: ACOMP06/076,BFU2005-23722-E/BFI, MEC BFU2005-09085; Grant sponsor: ELA Fonda-tion de Recherche, France (to S.M.).
The first two authors contributed equally to this work.*Correspondence to: B. Zalc, INSERM U711; UPMC, Biologie des Inter-
actions Neurones/Glie, Hopital de la Salpetriere, 47, Bd de l’Hopital, 75651Paris Cedex 13, France. E-mail: [email protected]
Received 16 February 2007; Revised 28 April 2008; Accepted 27 Sep-tember 2008
DOI 10.1002/cne.21904Published online in Wiley InterScience (www.interscience.wiley.com).
The Journal of Comparative Neurology 512:74–83 (2009)
© 2008 Wiley-Liss, Inc.

The aim of the present article is to provide a detailed fatemap for the neuronal plp-activated progenitors of the dien-cephalon, which are localized in the zona limitans intrathalamica(ZLI), the basal plate (bpD), and in the posterior hypothalamus,i.e., the mammillary and retromammillary tegmentum, at E9.5.Using CRE-recombinase-based genetic labeling we show thatthe progeny of diencephalic and hypothalamic plp-activatedprogenitors can be divided into two groups, which are in theirmajority both anterior to the ZLI. The neurons derived from theZLI segregate into the zona incerta (ZI), the subgeniculate nu-cleus, the ventral lateral geniculate, and the intergeniculate leaf-let. The involvement of the latter, since it is a thalamic structure,suggests that after a radial migration through the ZLI, some cellsmove caudally. Rostrally and ventrally, the basal plate cells of theretromammillary and mammillary tegmentum generate neuronsforming the subthalamic nucleus and contributing to the neuro-nal populations in the caudal part of the posterior hypothalamusand mammillary nuclei.
MATERIALS AND METHODSAnimals
Animal handling was carried out according to ethical regu-lations and guidelines from the Guide for the Care and Use ofLaboratory Animals (National Research Council 1996) and theEuropean Communities Council Directive (86/609/ECC). Theplp-cre mice (Delaunay et al., 2008) were crossed with eitherthe R26R (Soriano, 1999) or the Z/EG reporter mice (Novak etal., 2000). The plp construct contained 3.74 kb of 5� flankingplp DNA, all of exon 1, and the 8.15 kb long intron 1 of themurine plp gene. This construct has been repeatedly ob-served to faithfully drive a plp-like expression of reportergenes (Spassky et al., 1998; Leone et al., 2003; Delaunay et al.,2008). A cDNA encoding Cre recombinase (from pMC-Cre; Guet al., 1994) was then fused into plp exon 2. The plp-shble-lacZ animals were generated in DBA2 � C57Bl6/N F1 usingthe same plp-expression cassette and maintained homozy-gous in the C57Bl6/N strain (Spassky et al., 1998, 2001a).Heterozygous plp-shble-lacZ mice were derived from thesehomozygous lines by crossing with OF1 animals (Iffa Credo,France). The midpoint of the dark interval during which matingoccurred was designated day 0, and the embryos were con-sidered to be E0.5 on the morning after fertilization.
Detection of �-galactosidase enzymatic activityEmbryos were dissected in phosphate-buffered saline
(PBS, pH 7.4), fixed in 4% paraformaldehyde (PFA) for 2 min-utes, washed twice in PBS, and stained at room temperature(RT) overnight. The staining solution contained 2 mM of5-bromo-3-indolyl-�-D-galactoside (Bluo-gal; Life Technolo-gies, Gaithersburg, MD), 20 mM potassium ferrocyanide, 20mM potassium ferricyanide, and 2 mM MgCl2 in PBS.
ImmunohistochemistryTissues were either fixed by immersion (for embryos) or
perfused (for postnatal/adult brains) with 4% PFA, then post-fixed overnight at 4°C. Embryonic fixed tissues were washedin PBS, cryoprotected in PBS/15% sucrose for 12 hours at4°C, embedded in sucrose-gelatin (15%–7.5% in PBS), andfrozen in melting isopentane (�40°C to �50°C). Postnatal andadult fixed tissues were directly embedded in OCT and frozenin dry ice. All tissues were cut (16–20 �m) on a cryostat
(Leica). Alternatively, the embryos were embedded in low-melting-point agarose and cut on a vibratome (Leica) in 50–60�m thick sections. Sections were preblocked in PBS contain-ing 10% FCS / 0.2% TritonX-100 and incubated with primaryantibody overnight at 4°C. After washing, sections were incu-bated with secondary antibody for 1–2 hours at RT in the samesolution. For immunoperoxidase staining, endogenous perox-idase was inhibited by immersion of the sections in 1.5% H2O2
in PBS solution for 10 minutes at RT. After blocking, sectionswere incubated with the first antibody O/N at 4°C, then withthe biotin-conjugated secondary antibody for 1 hour at RT.Immunolabeled sections were treated with the Vectastain-Elite-ABC reagent (AbCys, Paris, France). After two washes in0.1 M Tris-HCl, pH 7.6, the peroxidase activity was revealedusing 3,3�-diaminobenzidine tetrahydrochloride (Dakopatts,Glostrup, Denmark) as a chromogen at a concentration of 1mg/mL in 0.1 M Tris-HCl, pH 7.6. The reaction was stopped bychanging the pH by addition of distilled water. For betteranatomical identification a cresyl violet counterstain was per-formed and the sections were dehydrated in graded ethanol,transferred to xylene, and mounted with Eukitt (Prolabo, Paris,France). Tissue sections were examined and photographedunder a Leica DMR microscope equipped with a photographiccamera DC500. Photographs were stored in Adobe Photo-shop (San Jose, CA) program under TIFF format (the originalformat of the camera software). The figures were composedwith Adobe Photoshop software (v. 7.0), using the Free Handprogram for labeling and drawings. In some cases, whenrequired, image brightness and contrast were adjusted toobtain homogeneous color tonalities and picture back-grounds. All antibodies used in this study are listed in Table 1.
Antibody characterizationCB. The calbindin D28k antibody reacts specifically to a
single band at 28 kDa on Western blot of rat brain extract.Immunostaining of mouse embryo brain sections produces aspecific staining pattern similar to that previously published(Haverkamp and Wassle, 2000).
CR. This antibody reacts specifically with a 29-kDa bandon Western blots of rat brain and nerve tissue extract. Thispurified antibody immunostained the expected cellular pat-tern of interneurons in the adult mouse neocortex (Liodis etal., 2007).
Cre. This monoclonal antibody recognized the epitopelocated between amino acids 77 and 343 of the Cre recombi-nase. No staining was detected on brain sections of wildtypemice.
GABA. The monoclonal anti-GABA antibody stained aband of 57 kDs on Western blot of mouse brain, and has beenevaluated for specificity by dot blot immunoassay by themanufacturer. No crossreaction was observed with BSA, L-�-aminobutyric acid, L-glutamic acid, L-aspartic acid, glycine,D-aminovaleric acid, L-threonine, L-glutamine, taurine, pu-trescine, L-alanine, and carnosine. A weak crossreaction wasobserved with �-alanine and amino-caproic acid.
�-Gal. According to the manufacturer, assay by immuno-electrophoresis of this antibody resulted in a single precipitinarc against antigoat serum as well as purified and partiallypurified �-galactosidase (�-gal) from E. coli. Crossreactivityagainst �-gal from other tissues and species may occur buthas not been specifically determined.
The Journal of Comparative Neurology
75SUBPOPULATIONS OF PRETHALAMIC NEURONS

GFP (rabbit). According to the manufacturer, this anti-GFP antibody is suited for detection of native GFP, and mostGFP fusion proteins by Western blot and immunochemistryanalysis using transgenic mice expressing the GFP geneproduct. No staining was detected on brain sections fromwildtype mice.
GFP (rat). According to the manufacturer, this antibodyreacts with GFP (Western blot, immunohistochemistry, andenzyme-linked immunosorbent assay [ELISA]) and crossre-acts with BlueFluorescent Protein (BFP).
Glu. The antibody has been calibrated against a spec-trum of antigens to assure hapten selectivity and properaffinity. According to the manufacturer, there exists nomeasurable crossreactivity (<1:1,000) against glutamate inpeptides or proteins. No measurable glutaraldehyde-fixedtissue crossreactivity (<1:1,000) against L-alanine,�-aminobutyrate, 1-amino-4-guanidobutane, D/L-arginine,D/L-aspartate, L-citrulline, L-cysteine, D/L-glutamine, glu-tathione, glycine, L-lysine, L-ornithine, L-serine, taurine,L-threonine, L-tryptophan, L-tyrosine. Modest crossreac-tivity (1:20) against D-glutamate. Significant crossreactivity(1:8) against free NAAG in competition assays. However, itis of note that NAAG is not retained in glutaraldehyde-fixedtissue.
NeuN. The monoclonal antibody to vertebrate neuron-specific nuclear protein, called NeuN (or Neuronal Nuclei),reacts with most neuronal cell types throughout the ner-vous system of mice including cerebellum, cerebral cortex,hippocampus, thalamus, spinal cord, and neurons in theperipheral nervous system, including dorsal root ganglia,sympathetic chain ganglia, and enteric ganglia. Develop-mentally, immunoreactivity is first observed shortly afterneurons have become postmitotic; no staining has beenobserved in proliferative zones. The immunohistochemicalstaining is primarily localized in the nucleus of the neurons,with lighter staining in the cytoplasm. The few cell types notreactive with MAB377 include Purkinje, mitral, and photo-receptor cells.
PV. According to the manufacturer, the PV-28 antibodyrecognizes a 28-kDa band on Western blot of the brain extractof various mammals. The distribution of the stained cellscoincides exactly with those of neurons expressing parvalbu-min mRNA. The antibody PV-28 does not stain the brain andmuscle of parvalbumin-knockout mice.
TH. This antibody reacts specifically with a 60-kDa bandon Western blot of rat caudate lysate and of PC-12 cellsstimulated by okadaic acid.
Cell countsTo count the number of neurons in each nucleus, coronal
sections of two P15 plp-cre;R26R animals were immuno-stained for �-gal and neuN, then examined under a Leicaconfocal microscope. Each nucleus containing �-gal� cellswas identified and the number of double-labeled neurons wascounted on a series of rostral to caudal photographed sec-tions and in both hemispheres. Quantification was performedwith the software program Image J. The number of cells socounted was corrected post-hoc by using the Abercrombieformula according to Guillery (2002).
RESULTSValidation of plp-cre mouse
To establish a fate map of the plp-activated progenitorsgenerated during embryonic development, we have taken ad-vantage of a genetic labeling procedure based on the CRErecombinase strategy. We have generated a plp-cre mouseusing the regulatory sequences of the plp gene to drive theexpression of the CRE recombinase (see Materials and Meth-ods). This mouse has been crossed to transgenic mice ex-pressing lox-reporters (either �-gal or GFP) under the controlof an ubiquitous reporter, either Rosa26 (R26R; Soriano, 1999)or actin (Z/EG; Novak et al., 2000). In the resulting doubletransgenic (plp-cre;R26R or plp-cre;Z/EG) all the cells containthe loxP-reporter sequence, but only the cells expressing plpalso express the CRE-recombinase. In the plp� cells, the
TABLE 1. Antibody Reagents Used in This Study
Target Antigen Species Dilution Source, catalog number
Primary antibodies1
Calbindin D28k Purified rat calbindin D28k Mouse 1:5000 Swant C9638Calretinin Recombinant rat calretinin Rabbit 1:1000 Chemicon AB5054Cre-recombinase Recombinant Cre recombinase fusion protein Mouse 1:100 Euromedex CRE-2D8-ASGABA Purified GABA conjugated to BSA Mouse 1:100 Sigma A0310Galactosidase-� �-galactosidase from E. coli Goat 1:700 Biotrend 4600-1409GFP GFP isolated directly from Aequorea victoria Rabbit 1:500 Invitrogen A11122GFP His-GFP (full length) fusion protein Rat 1:1000 Nacalai Tesque 0440426Glutamate Glutamate-glutaraldehyde conjugate Rabbit 1:100 Chemicon AB5018NeuN Immunopurified nuclei from mouse brain Mouse 1:100 Chemicon MAB377Parvalbumin Rat muscle parvalbumin Rabbit 1:1000 Swant PV28Tyrosine hydroxylase SDS-denatured tyrosine hydroxylase purified
from rat pheochromocytomaRat 1:500 PelFreez Biologicals Rogers
40101-1
Antibody Species Dilution Source, catalog number
Secondary antibodiesAntisheep/goat total IgG biotinylated Donkey 1/200 Amersham France #RPN 1025Antirabbit total IgG biotinylated Goat 1/200 Sigma France #B7389Antirat total IgG biotinylated Goat 1/200 Sigma France #B7139Antimouse IgG2b Alexa 594 nm conjugated Goat 1/2000 Molecular Probes/Interchim France #A21145Antirabbit IgG(H�L) Alexa 488 or 594 nm conjugated Goat 1/2000 Molecular Probes/Interchim France #A11008 & A11012Antigoat IgG(H�L) Alexa 488 or 594 nm conjugated Donkey 1/2000 Molecular Probes/Interchim France #A110055 & A110058Antirat IgG(H�L) Alexa 594 nm conjugated Goat 1/2000 Molecular Probes/Interchim France #A11007Antimouse IgG(H�L) Alexa 488 or 594 nm conjugated Goat 1/2000 Molecular Probes/Interchim France #A11029 & A11005Streptavidin Alexa 488 or 594 nm conjugated Chicken
egg1/1000 Molecular Probes/Interchim France #S32354 & S32356
1Details on the specificity controls are described in Materials and Methods.
The Journal of Comparative Neurology
76 D. DELAUNAY ET AL.
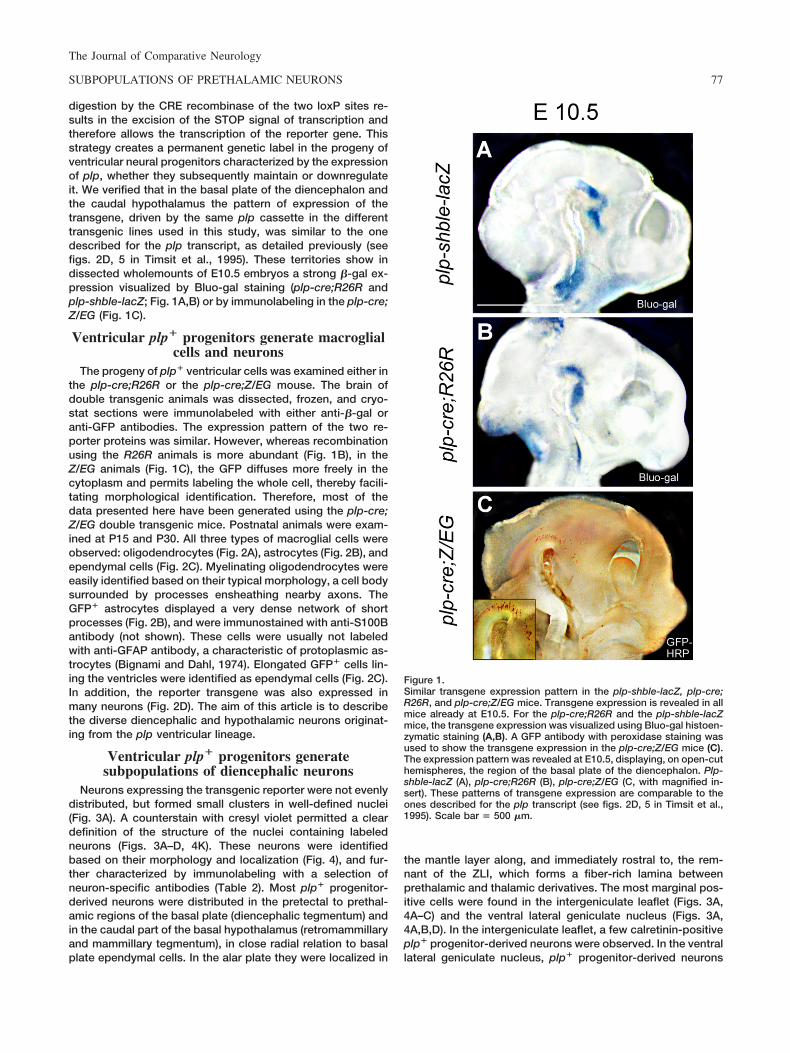
digestion by the CRE recombinase of the two loxP sites re-sults in the excision of the STOP signal of transcription andtherefore allows the transcription of the reporter gene. Thisstrategy creates a permanent genetic label in the progeny ofventricular neural progenitors characterized by the expressionof plp, whether they subsequently maintain or downregulateit. We verified that in the basal plate of the diencephalon andthe caudal hypothalamus the pattern of expression of thetransgene, driven by the same plp cassette in the differenttransgenic lines used in this study, was similar to the onedescribed for the plp transcript, as detailed previously (seefigs. 2D, 5 in Timsit et al., 1995). These territories show indissected wholemounts of E10.5 embryos a strong �-gal ex-pression visualized by Bluo-gal staining (plp-cre;R26R andplp-shble-lacZ; Fig. 1A,B) or by immunolabeling in the plp-cre;Z/EG (Fig. 1C).
Ventricular plp� progenitors generate macroglialcells and neurons
The progeny of plp� ventricular cells was examined either inthe plp-cre;R26R or the plp-cre;Z/EG mouse. The brain ofdouble transgenic animals was dissected, frozen, and cryo-stat sections were immunolabeled with either anti-�-gal oranti-GFP antibodies. The expression pattern of the two re-porter proteins was similar. However, whereas recombinationusing the R26R animals is more abundant (Fig. 1B), in theZ/EG animals (Fig. 1C), the GFP diffuses more freely in thecytoplasm and permits labeling the whole cell, thereby facili-tating morphological identification. Therefore, most of thedata presented here have been generated using the plp-cre;Z/EG double transgenic mice. Postnatal animals were exam-ined at P15 and P30. All three types of macroglial cells wereobserved: oligodendrocytes (Fig. 2A), astrocytes (Fig. 2B), andependymal cells (Fig. 2C). Myelinating oligodendrocytes wereeasily identified based on their typical morphology, a cell bodysurrounded by processes ensheathing nearby axons. TheGFP� astrocytes displayed a very dense network of shortprocesses (Fig. 2B), and were immunostained with anti-S100Bantibody (not shown). These cells were usually not labeledwith anti-GFAP antibody, a characteristic of protoplasmic as-trocytes (Bignami and Dahl, 1974). Elongated GFP� cells lin-ing the ventricles were identified as ependymal cells (Fig. 2C).In addition, the reporter transgene was also expressed inmany neurons (Fig. 2D). The aim of this article is to describethe diverse diencephalic and hypothalamic neurons originat-ing from the plp ventricular lineage.
Ventricular plp� progenitors generatesubpopulations of diencephalic neurons
Neurons expressing the transgenic reporter were not evenlydistributed, but formed small clusters in well-defined nuclei(Fig. 3A). A counterstain with cresyl violet permitted a cleardefinition of the structure of the nuclei containing labeledneurons (Figs. 3A–D, 4K). These neurons were identifiedbased on their morphology and localization (Fig. 4), and fur-ther characterized by immunolabeling with a selection ofneuron-specific antibodies (Table 2). Most plp� progenitor-derived neurons were distributed in the pretectal to prethal-amic regions of the basal plate (diencephalic tegmentum) andin the caudal part of the basal hypothalamus (retromammillaryand mammillary tegmentum), in close radial relation to basalplate ependymal cells. In the alar plate they were localized in
the mantle layer along, and immediately rostral to, the rem-nant of the ZLI, which forms a fiber-rich lamina betweenprethalamic and thalamic derivatives. The most marginal pos-itive cells were found in the intergeniculate leaflet (Figs. 3A,4A–C) and the ventral lateral geniculate nucleus (Figs. 3A,4A,B,D). In the intergeniculate leaflet, a few calretinin-positiveplp� progenitor-derived neurons were observed. In the ventrallateral geniculate nucleus, plp� progenitor-derived neurons
Figure 1.Similar transgene expression pattern in the plp-shble-lacZ, plp-cre;R26R, and plp-cre;Z/EG mice. Transgene expression is revealed in allmice already at E10.5. For the plp-cre;R26R and the plp-shble-lacZmice, the transgene expression was visualized using Bluo-gal histoen-zymatic staining (A,B). A GFP antibody with peroxidase staining wasused to show the transgene expression in the plp-cre;Z/EG mice (C).The expression pattern was revealed at E10.5, displaying, on open-cuthemispheres, the region of the basal plate of the diencephalon. Plp-shble-lacZ (A), plp-cre;R26R (B), plp-cre;Z/EG (C, with magnified in-sert). These patterns of transgene expression are comparable to theones described for the plp transcript (see figs. 2D, 5 in Timsit et al.,1995). Scale bar � 500 �m.
The Journal of Comparative Neurology
77SUBPOPULATIONS OF PRETHALAMIC NEURONS

were GABA-immunoreactive and calbindin-positive, but werenot labeled by antibodies against tyrosine hydroxylase, cal-retinin, or parvalbumin (Table 2). Medial and rostral to theventral lateral geniculate nucleus, the subgeniculate nucleus(Fig. 4A,E,F) and the ventral and medial region of the ventralthalamus, the zona incerta (ZI) also showed reporter-positivecells (Fig. 4A,G,H). In the subgeniculate nucleus, transgene-expressing neurons were calretinin- and parvalbumin-positive. In the ZI they were also calretinin-positive, but rarelyparvalbumin-positive (Table 2). In most of these nuclei plp�-derived neurons represented on average 10% of neurons,except for the subgeniculate nucleus, where they accountedfor only 5% (Table 2).
With regard to the hypothalamic basal plate derivatives,placed rostral to the subgeniculate nucleus and the ZI, plp�
progenitor-derived neurons were present in the subthalamicnucleus, representing around 10% of neurons (Fig. 4A,I,J;
Table 2). These plp� progenitor-derived neurons were gluta-matergic, calretinin-positive, and negative for the other testedmarkers (TH, calbindin, parvalbumin). More rostrally, the lastgroup of basal plate neurons derived from plp� ventricularprogenitors were numerous dispersed polygonal neurons inthe posterior hypothalamus (Fig. 4A,K,L), where they repre-sented 56% of neurons (Table 2). These neurons wereGABAergic and expressed calretinin (Table 2). In addition,plp� ventricular progenitors generated scattered tegmentalreticular neurons in the prerubral and retromammillary teg-mental areas.
Detection of plp progenitor-derived neuronsduring diencephalic development
To investigate the timing of genesis of plp� progenitor-derived neurons during diencephalic development, we alsoused the plp-cre;Z/EG double transgenic. At E9.5–E10.5, the
Figure 2.Plp� progenitors give rise to all types of neural cells in the adult diencephalon. Staining with an anti-GFP Ab in a plp-cre;Z/EG adult mouse (P30)revealed progeny of plp� neuroepithelial cells. In the diencephalon, plp progenitors-derived cells were macroglia: oligodendrocytes (A),astrocytes (B), ependymal cells (C), and neurons (D). Scale bar � 35 �m. [Color figure can be viewed in the online issue, which is available atwww.interscience.wiley.com.]
Figure 3.Plp� progenitor-derived neurons are distributed in a subset of diencephalic nuclei. A: Coronal section across the diencephalon of a P15plp-cre;Z/EG mouse, immunostained with anti-GFP and counterstained with cresyl violet for better identification of the nuclear structurescontaining labeled cells. B: More detailed identification of the alar region showing the dorsal and ventral laterate geniculate nuclei and theintergeniculate leaflet. Note the presence of glial plp-positive cells populating these nuclear structures. C: Labeled neurons in the subthalamicnucleus, same section as (A). D: Partial overview of a coronal diencephalic section mouse showing the dorsal region of the posteriorhypothalamus (PH). Areas and nuclear structures containing labeled cells are identified. CZI, caudal zona incerta; dLG, dorsal lateral geniculatenucleus; IGL, intergeniculate leaflet; LH, lateral hypothalamic area; ped, peduncle; PH, posterior hypothalamic area; RZI, rostral zona incerta;subG, subgeniculate nucleus; sth, subthalamic nucleus; vLG, ventral lateral geniculate nucleus; VPL, ventro posterolateral thalamic nucleusVPM, ventro posteromedial thalamic nucleus ZI, zona incerta; FF, field of Forel (1877). Scale bars � 300 �m in A,D; 150 �m in B; 80 �m in C.
The Journal of Comparative Neurology
78 D. DELAUNAY ET AL.

pattern of expression of GFP-reporter expressing cells wasrestricted to the ventricular layer of the latero-basal plate andwas similar to that described for the plp-shble-lacZ (Spasskyet al., 1998) embryos examined at similar developmentalstages (Delaunay et al., 2008) (Fig. 1C).
At E12.5, a large number of GFP-positive cells were stilldetected in the ventricular layer of the third ventricle (Fig.
5A–D). In the prethalamus, at the anterior limit of the dien-cephalon with the hypothalamus (Fig. 5B), GFP� cells weremost frequently bipolar and seemed to be in the process ofmigration. The central part of the diencephalon showed twogroups of GFP-positive cells (Fig. 5C,D). A dorsal subpopu-lation was situated in the alar plate, at the base of the ZLI,between the thalamus and the prethalamus (Fig. 5C). A
Figure 4.Localization and morphological identification of plp� progenitor-derived neurons in prethalamic nuclei. A: Overview of a coronal diencephalicsection of an adult plp-cre;Z/EG mouse showing the global distribution of plp� progenitor-derived neurons. Neurons were revealed byexpression of GFP and recognized based on their morphology. B: The intergeniculate leaflet and the ventral lateral geniculate nucleus containplp� progenitor-derived neurons. C: Higher magnification of the intergeniculate leaflet, which presented few GFP-labeled neurons. D: Highermagnification of GFP-positive neurons in the ventral lateral geniculate nucleus. E: Transgene expressing neurons in the subgeniculate nucleus.F: Higher magnification of (E). G: Typical plp� progenitor-derived neurons detected in the zona incerta. H: Higher magnification of (G). I: Thesubthalamic nucleus presents long bipolar GFP� neurons. J: Higher magnification of (I). K: Caudal part of posterior hypothalamus presentinga group of GFP� neurons, with cresyl violet counterstain. L: Higher magnification of hypothalamic neurons with long processes and large cellbodies, detected in the caudal hypothalamus (low magnification, insert). DM, dorsomedial hypothalamic nucleus; IGL, intergeniculate leaflet; Hy,hypothalamus; PH, posterior hypothalamic area; sth, subthalamic nucleus; subG, subgeniculate nucleus; vLG, ventral lateral geniculate nucleus;ZI, zona incerta;. Scale bars � 300 �m in A; 150 �m in B,E,G,I,K; 37 �m in C,D,F,H,J,L. [Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
The Journal of Comparative Neurology
79SUBPOPULATIONS OF PRETHALAMIC NEURONS

subpopulation, lying rostrally to the ZLI, corresponded tothe basal plate of the prethalamus (bpD in Fig. 5D). Clustersof GFP� cells formed a longitudinal continuum that coveredthe entire basal plate of the diencephalon, down to thepretectal tegmentum (Fig. 5D), always excluding the epi-chordal floor plate. Some of these ventricular GFP-positivecells extended long processes toward the pial surface,similar to radial glial cells. At E15.5, GFP-expressing cells inthe ventricular layer were less numerous, but adopted atypical radial glia morphology (Fig. 4E–G), with the cellbodies lying close to the ventricular lumen, forming withtheir neighbors a cell-cluster anchored to the third ventri-cle, and exhibiting a long process extending across themantle layer toward the pial surface, where they displayedcharacteristic endfeet structures (Fig. 5H,I). At the mostrostral sections, such GFP� cells with a radial glia morphol-ogy formed two distinct boundaries: the ZLI, located trans-versally in the alar plate between thalamus and prethalamus, andthe limit between the diencephalon and the secondary prosen-cephalon, the diencephalic–hypothalamic boundary (DHB),found at the limit between the diencephalon (prosomere 3) andthe secondary prosencephalon (Fig. 5E). Numerous immatureGFP� cells, either uni- or bipolar, were lined up along the radialglial processes of the ZLI. At the pial end of the ZLI, GFP� cellscontributed to the ventral lateral geniculate nucleus in the pre-thalamus (p3), and, caudally, in the intergeniculate leaflet insidethe thalamus (p2) (Fig. 5E,F). Ventral to the ventral lateral genic-ulate nucleus GFP� cells were detected more medially in thezona incerta (Fig. 5I). More ventrally, GFP� cells in the basalplate, already detected at E12.5, had a more mature appearance(Fig. 5J).
At birth, the ZLI and DHB were still visible as two thinboundaries formed by plp progenitor-derived radial glial cells(Fig. 5K–N). The GFP� cells located along the radial glial cellprocesses were, however, much less numerous than at em-bryonic stages (E12.5–15.5). In contrast, the GFP� neuronswere accumulated either close to the marginal end of the ZLI(in the intergeniculate leaflet and ventral lateral geniculate) orpopulated rostrally the subthalamic nucleus close to the DHB(Fig. 5K). Part of the GFP� cells formed a stream between theDHB and ZLI, suggesting that they had left the radial gliatracts after an initial radial migration (Fig. 5K,L). Medially,GFP� cells were still observed among the radial glia cellbodies of the ventricular lumen, and in the medial ZI, close tothe basal plate (Fig. 5M,N).
DISCUSSIONUsing a genetic labeling based on the CRE recombinase
strategy we have characterized the neurons generated fromplp� ventricular cells in the diencephalon and caudal hypo-thalamus. It is worth noting that in a study dissecting theregulatory sequences controlling plp transcription, discreteregions located 5� of exon 1 and in intron 1 of the plp genehave been reported to be associated with neuronal expres-sion (Tuason et al., 2008). Diencephalic plp� ventricularcells are neuroepithelial cells localized in the ZLI, and moreventrally in the basal plate, extending from the pretectal toprethalamic tegmental regions, and the posterior hypothal-amus and the retromammillary and mammillary regions,according to the revised version of the prosomeric model(Puelles and Rubenstein, 2003). In Figure 6 we propose a
Figure 5.Time-course localization of plp� progenitor-derived neurons during diencephalic development. A–D: GFP expression in a E12.5 plp-cre;Z/EGembryo. Coronal sections are presented from the rostral part of the diencephalon to the most caudal part. A: Diagram showing a sagittal viewof the diencephalon at E12.5 with the different planes of section. B: GFP expression in the prethalamus, in the diencephalic–hypothalamicboundary, and the hypothalamus. C: Section showing the ZLI and the basal plate of the diencephalon. D: Ventricular and mantle layer labeledcells in the basal plate of the caudal diencephalon (pretectum), with long radial processes reaching the pial surface. Note that labeled cells areexcluded from the median floor plate. E–J: GFP expression in the plp-cre;Z/EG mouse at E15.5. E,F: GFP� neurons are seen in the ventral lateralgeniculate nucleus and the intergeniculate leaflet. These nuclei seemed to be formed by the ventricular cells originating in the ZLI, which extendlong processes toward them. Note that ventral to the ZLI, the diencephalic–hypothalamic boundary (DHB) is visible (E). G: High magnificationof (F): the ZLI shows immature bipolar cells migrating along the GFP� radial processes. H: Formation of the zona incerta, which is crossed bya multitude of radial glial fibers reaching from the ventricle to the pial surface (p). I: High magnification of the ventral lateral geniculate nucleusalready formed at this developmental stage. J: GFP� cells in the basal plate of the diencephalon, many having already left the ventricular zone.K–N: At birth, the ZLI and the diencephalic-hypothalamic boundary are still visible and some cells seem to bridge these two limits (L). The ZIis well-formed and GFP-positive cells are still present in the basal plate of the diencephalon (M), some of them displaying a typical radial gliamorphology (N). bpD, basal plate of diencephalon; DHB, diencephalic-hypothalamic boundary; dLG, dorsal lateral geniculate; IGL, intergenicu-late leaflet; Hy, hypothalamus; PH, posterior hypothalamic area; pth, pre-thalamus; sth, subthalamic nucleus; th, thalamus; vLG, ventral lateralgeniculate nucleus; ZLI, zona limitans intrathalamica; ZI, zona incerta. Scale bars � 400 �m in C,D,K; 260 �m in B,E,F,L; 200 �m in H,J,M; 133�m in I; 100 �m in G; 80 �m in N. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
TABLE 2. Neuronal Markers Expressed by plp� Progenitor-Derived Neurons
Territory Plp-activated neurons (%SEM) Calretinin Calbindin Parvalbumin GABA TH Glutamate
IGL 10.4 3.1 � � � � � �vLG 15.4 4.2 � � � � � �subG 5.0 2.8 � � � � � �ZI 10.4 1.8 � � � � � �sth 10.1 1.5 � � � � � �PH 56.3 9.6 � � � � � �
The neuronal phenotype of the plp� progenitor-derived neurons was assessed on P15 plpCRE;R26R mouse coronal sections double-stained with both anti-�-galactosidase andspecific neuronal markers: either calretinin, calbindin, or parvalbumin. The neurotransmitters expressed by plp� progenitor-derived neurons were examined using Abs directedagainst GABA, tyrosine hydroxylase, and glutamate. To quantify the plp� progenitor-derived neurons within each nucleus, coronal sections were immunostained with bothanti-�-galactosidase and neuN Abs. IGL, intergeniculate leaflet; PH, posterior hypothalamic area; subG, subgeniculate nucleus; sth, subthalamic nucleus, TH, tyrosine hydroxylase;vLG, ventral lateral geniculate nucleus; ZI, zona incerta.
The Journal of Comparative Neurology
80 D. DELAUNAY ET AL.

tentative model of the migration pathways followed by plp-derived diencephalic neurons. The neurons arising from theZLI were found in the ZI, the subgeniculate nucleus, theventral lateral geniculate, and the intergeniculate leaflet.The presence of plp-activated neurons in the IGL suggests
a caudal tangential movement into the neighboring subpialthalamus after radial migration through the ZLI radial glia.Plp-activated neurons originating from the basal plate, theDHB, and the retromammillary area seem to contribute tothe formation of parts of alar prethalamus (e.g., ZI), as well
Figure 5
The Journal of Comparative Neurology
81SUBPOPULATIONS OF PRETHALAMIC NEURONS

as of the subthalamic nucleus and posterior hypothalamus(Fig. 6).
The majority of neurons derived from plp� progenitors co-incide with GABAergic populations. The ZI is a prethalamicregion principally composed of GABAergic neurons involvedin the integration of somatosensory signals (Lin et al., 1990;Kimet al., 1992; Kolmac and Mitrofanis, 1998). Neurons in theZI project to the thalamus, the cortex, and the superior col-liculus. Recently, it has been proposed that scattered neuronsfrom the ZI, characterized by their expression of hypocretin,project to the intergeniculate leaflet and are therefore a com-ponent controlling the modulation of the circadian rhythm(Vidal et al., 2005). The projection of the ZI neurons to the IGLis of interest with regard to the present study, since this is theonly circuitry of the diencephalon connecting plp� progenitor-derived neurons. The ventral lateral geniculate, subgeniculate,and intergeniculate nuclei are three closely related structuresalso containing GABAergic neurons. These nuclei have affer-ent and efferent connections in the visual, auditory, and cir-cadian systems (Harrington, 1997; Kas et al., 1999).
Our data show that only a subpopulation of neurons withineach nucleus derives from plp-activated progenitors. Thisfinding is in agreement with previous studies. Based on theexpression of different neurotransmitters and calcium-bindingproteins, Kolmac and Mitrofannis (1998) have shown hetero-geneity of neurons in the ZI. This heterogeneity has beenfurther illustrated and confirmed in the prethalamus by usingnumerous transcription factors such as LIM-homeodomaingenes (Isl1, Lhx1, Lhx2, Lhx5, and Lhx9) to mark subsets ofnuclei (Nakagawa and O’Leary, 2001).
A causal relationship between plp expression and sonichedgehog (Shh) has been reported in the telencephalon (Neryet al., 2001; Olivier et al., 2001). Our observations suggest thatin the diencephalon the plp expression is also regulated byShh, which may confer extensive migratory properties to theneural cells born in this domain, such as in other Shh� terri-tories (Lavdas et al., 1999; Olivier et al., 2001; Marin andRubenstein, 2003). Postmitotic neurons derived from plp�
progenitors display morphological features in accordancewith such migratory behavior. We observed bipolar GFP-positive cells with a leading process directed toward the pialsurface, which are closely related to the GFP-positive radialglia fibers of the ZLI (Fig. 5H). The former cells may very likelybe neurons derived from the ZLI epithelium (from progenitorseither similar to or different from the radial glia cells) whichmigrate along radial glia fibers, to then segregate either cau-dalward into the superficial region of the thalamus, where theypopulate the IGL, or rostralward, entering the vLG nucleus(Figs. 5E, 6). Neurons labeled in the subthalamic nucleus,which are glutamatergic and calretinin-positive, provided sim-ilar observations, showing a ventrodorsal migration. The sub-thalamic nucleus, also known as the corpus of Luys, is themain regulator of motor functions related to the basal ganglia.Subthalamic neurons originate in the basal plate of the caudalhypothalamus (Muccielli et al., 2004; Martin et al., 2004).These authors discuss migration of prospective subthalamicneurons out of the retromammillary area. The basal plate ofthe caudal hypothalamus contains abundant GFP� progeni-tors, but in a salt-and-pepper distribution, as has also beenshown to be the case for neural stem cells in the wall of thelateral ventricle (Merkle et al., 2007). This could account forthe fact that only a fraction of the neurons in the subthalamicnucleus and other hypothalamic nuclei are GFP-positive,while the bulk of these neurons apparently derive from plp-negative progenitors from the same area. The subthalamicnucleus has gained special attention, since a major break-through in the treatment of advanced Parkinson’s disease hasbeen the discovery that high-frequency deep brain stimulationby electrodes, implanted in the subthalamic nucleus, allevi-ates akinesia/dyskinesia and rigidity, the cardinal symptomsof patients suffering from this illness (Benabid et al., 1994).
Finally, hypothalamic neurons were also generated fromplp� progenitors in the basal plate. All these neurons appearas reticular neurons, with a large cell body and a heteroge-neous distribution. By permanently marking a subpopulationof neural progenitors and their progeny we have been able tostudy in detail the neuronal component of the derived cellpopulations and we illustrate the heterogeneous final locationof these neurons, which contribute to tegmental diencephalic,intergeniculate, prethalamic, and caudal basal hypothalamicforebrain structures.
Figure 6.Three-dimensional diagram showing a lateral view of the mouse di-encephalon. Along the ventricular surface, plp expression domains inthe basal plate and the ZLI are illustrated in red. Derivates from thebasal plate in the mantle layer as well as the alar plate nuclei arerepresented in gray tones. GFP-positive cells, originating from plp-activated progenitors are mapped with red dots. Red arrows illustratethe putative migratory streams followed by these cells to reach theirrespective nuclei. IGL, intergeniculate leaflet; MM, mammillary area;ped, telencephalic peduncle; PH, posterior hypothalamic area; pTh,prethalamus; RT, reticular nucleus; RM, retromammillary area; subG,subgeniculate nucleus; sth, subthalamic nucleus; Th, thalamus; vLG,ventral lateral geniculate nucleus; ZI, zona incerta.
The Journal of Comparative Neurology
82 D. DELAUNAY ET AL.

LITERATURE CITEDBenabid AL, Pollak P, Gross C, Hoffmann D, Benazzouz A, Gao DM,
Laurent A, Gentil M, Perret J. 1994. Acute and long-term effects ofsubthalamic nucleus stimulation in Parkinson’s disease. StereotactFunct Neurosurg 62:76 – 84.
Bignami A, Dahl D. 1974. Astrocyte-specific protein and neuroglial differ-entiation. An immunofluorescence study with antibodies to the glialfibrillary acidic protein. J Comp Neurol 153:27–38.
Delaunay D, Heydon K, Cumano A, Schwab M, Thomas JL, Nave K-A,Suter U, Zalc B, Spassky N. 2008. Early neuronal and glial fate restric-tion of embryonic neural stem cells. J Neurosci 28:2551–2562.
Folch J, Lees M. 1951. Proteolipides, a new type of tissue lipoproteins;their isolation from brain. J Biol Chem 191:807– 817.
Forel A. 1877. Untersuchungen ueber die Hauben-region und ihre ober-enVerknuepfungen im Gehirne des Menschen und einiger Saeugeth-iere, mit Beitragen zu den Methoden der Gehirnuntersuchung. ArchPsychiat Nervenkrankh 7:393– 495.
Guillery RW, 2002. On counting and counting errors. J Comp Neurol447:1–7
Harrington ME. 1997. The ventral lateral geniculate nucleus and the inter-geniculate leaflet: interrelated structures in the visual and circadiansystems. Neurosci Biobehav Rev 21:705–727.
Haverkamp S, Wassle H. 2000. Immunocytochemical analysis of themouse retina. J Comp Neurol 424:1–23.
Kim U, Gregory E, Hall WC. 1992. Pathway from the zona incerta to thesuperior colliculus in the rat. J Comp Neurol 321:555–575.
Kolmac CI, Power BD, Mitrofanis J. 1998. Patterns of connections be-tween zona incerta and brainstem in rats. J Comp Neurol 396:544 –555.
Lavdas AA, Grigoriou M, Pachnis V, Parnavelas JG. 1999. The medialganglionic eminence gives rise to a population of early neurons in thedeveloping cerebral cortex. J Neurosci 19:7881–7888.
Leone DP, Genoud S, Atanasoski S, Grausenburger R, Berger P, MetzgerD, Macklin WB, Chambon P, Suter U. 2003. Tamoxifen-inducible glia-specific Cre mice for somatic mutagenesis in oligodendrocytes andSchwann cells. Mol Cell Neurosci 22:430 – 440.
Lin CS, Nicolelis MA, Schneider JS, Chapin JK. 1990. A major directGABAergic pathway from zona incerta to neocortex. Science 248:1553–1556.
Liodis P, Denaxa M, Grigoriou M, Akufo-Addo C, Yanagawa Y, Pachnis V.2007. Lhx6 activity is required for the normal migration and specifica-tion of cortical interneuron subtypes. J Neurosci 27:3078 –3089.
Marin O, Rubenstein JL. 2003. Cell migration in the forebrain. Annu RevNeurosci 26:441– 483.
Martin DM, Skidmore JM, Philips ST, Vieira C, Gage PJ, Condie BG,Raphael Y, Martinez S, Camper SA. 2004. PITX2 is required for normal
development of neurons in the mouse subthalamic nucleus and mid-brain. Dev Biol 267:93–108.
Merkle FT, Mirzadeh Z, Alvarez-Buylla A. 2007. Mosaic organization ofneural stem cells in the adult brain. Science 317:381–384.
Muccielli ML, Martinez S, Pattyn A, Goridis C, Brunet JF. 1996. Otlx2, anOtx-related homeobox gene expressed in the pituitary gland and in arestricted pattern in the forebrain. Mol Cell Neurosci 8:258 –271.
Nakagawa Y, O’Leary DD. 2001. Combinatorial expression patterns ofLIM-homeodomain and other regulatory genes parcellate developingthalamus. J Neurosci 21:2711–2725.
Nery S, Wichterle H, Fishell G. 2001. Sonic hedgehog contributes tooligodendrocyte specification in the mammalian forebrain. Develop-ment 128:527–540.
Novak A, Guo C, Yang W, Nagy A, Lobe CG. 2000. Z/EG, a double reportermouse line that expresses enhanced green fluorescent protein uponCre-mediated excision. Genesis 28:147–155.
Soriano P. 1999. Generalized lacZ expression with the ROSA26 Cre re-porter strain. Nat Genet 21:70 –71.
Spassky N, Goujet-Zalc C, Parmantier E, Olivier C, Martinez S, Ivanova A,Ikenaka K, Macklin W, Cerruti I, Zalc B, Thomas JL. 1998. Multiplerestricted origin of oligodendrocytes. J Neurosci 18:8331– 8343.
Spassky N, Heydon K, Mangatal A, Jankovski A, Olivier C, Queraud-Lesaux F, Goujet-Zalc C, Thomas JL, Zalc B. 2001a. Sonic hedgehog-dependent emergence of oligodendrocytes in the telencephalon: evi-dence for a source of oligodendrocytes in the olfactory bulb that isindependent of PDGFRalpha signaling. Development 128:4993–5004.
Spassky N, Olivier C, Cobos I, LeBras B, Goujet-Zalc C, Martinez S, ZalcB, Thomas JL. 2001b. The early steps of oligodendrogenesis: insightsfrom the study of the plp lineage in the brain of chicks and rodents. DevNeurosci 23:318 –26.
Timsit SG, Bally-Cuif L, Colman DR, Zalc B. 1992. DM-20 mRNA isexpressed during the embryonic development of the nervous system ofthe mouse. J Neurochem 58:1172–1175.
Timsit S, Martinez S, Allinquant B, Peyron F, Puelles L, Zalc B. 1995.Oligodendrocytes originate in a restricted zone of the embryonic ven-tral neural tube defined by DM-20 mRNA expression. J Neurosci15:1012–1024.
Tuason M, Rastikerdar A, Goujet C, Zalc B, Dib S, Friedman A, Peterson A.2008. Separate PLP/DM20 enhancers serve different lineages andstages of development. J Neurosci 28:6895– 6903.
Vidal L, Blanchard J, Morin LP. 2005. Hypothalamic and zona incertaneurons expressing hypocretin, but not melanin concentrating hor-mone, project to the hamster intergeniculate leaflet. Neuroscience134:1081–1090.
Xu Q, Tam M, Anderson SA. 2008. Fate mapping Nkx2.1-lineage cells inthe mouse telencephalon. J Comp Neurol 506:16 –29.
The Journal of Comparative Neurology
83SUBPOPULATIONS OF PRETHALAMIC NEURONS




















