
Defining sustainability of plantation forests through identification of site quality indicators...
13
Defining sustainability of plantation forests through identification of site quality indicators influencing productivity—A national view for New Zealand Michael S. Watt a, * , Graham Coker a , Peter W. Clinton a , Murray R. Davis a , Roger Parfitt c , Robyn Simcock d , Loretta Garrett b , Tim Payn b , Brian Richardson b , Andrew Dunningham b a Centre for Sustainable Forest Management, Forest Research, P.O. Box 29237, Fendalton, Christchurch 4800, New Zealand b Centre for Sustainable Forest Management, Forest Research, Private Bag 3020, Rotorua, New Zealand c Landcare Research, Private Bag 11052, Palmerston North, New Zealand d Landcare Research, Private Bag 92170, Auckland, New Zealand Received 1 April 2005; received in revised form 19 May 2005; accepted 20 May 2005 Abstract New Zealand is committed to developing sustainable forest management practices as evidenced through Government involvement in international forestry agreements such as the Montreal Process, and the forestry sector’s adoption of forest certification mechanisms. In support of this commitment, it has been identified that there is little quantitative evidence of the interactions of plantation forestry on site quality and long-term site productivity. To address this issue, a nationwide study of site quality was initiated at 35 key sites covering the range of edaphic and environmental conditions representing the productivity envelope for New Zealand plantation forests. At each location, within the productivity envelope, eight short-term site quality plots were planted at a very high stand density (40,000 stems ha 1 ) to rapidly identify key soil indicators of growth which may be useful for determining site sustainability. In addition, a permanent sample plot was established by planting seedlings at conventional stem densities (500–1100 stems ha 1 ). At each site, a factorial design was applied with the following three factors: species (Pinus radiata and Cupressus lusitanica), fertiliser (no fertiliser and nutrients supplied in excess of crop demands) and disturbance (low and high disturbances). After two years of increment, initial analyses are presented which partition treatment and site effects on increment and identify key soil properties that influence increment of the two species. Volume increment over the two-year period was most strongly influenced by site, ranging 50-fold and 15-fold across sites for C. lusitanica and P. radiata, respectively. For the treatments, species accounted for most of the variance in increment, with mean volume increment across all sites of P. radiata significantly exceeding that of C. lusitanica by 56%. Fertilisation also significantly influenced volume increment inducing mean gains of 30%. Disturbance had a significant, but comparatively weak effect, reducing mean volume increment by 9%. After correction had been made for environment (temperature and rainfall), soil properties that were most strongly related to volume increment for both species included CN ratio, total soil nitrogen, total soil www.elsevier.com/locate/foreco Forest Ecology and Management 216 (2005) 51–63 * Corresponding author. Tel.: +64 3 364 2949; fax: +64 3 364 2812. E-mail address: [email protected] (M.S. Watt). 0378-1127/$ – see front matter # 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.foreco.2005.05.064
Transcript of Defining sustainability of plantation forests through identification of site quality indicators...

Defining sustainability of plantation forests through
identification of site quality indicators influencing
productivity—A national view for New Zealand
Michael S. Watt a,*, Graham Coker a, Peter W. Clinton a, Murray R. Davis a,Roger Parfitt c, Robyn Simcock d, Loretta Garrett b, Tim Payn b,
Brian Richardson b, Andrew Dunningham b
a Centre for Sustainable Forest Management, Forest Research, P.O. Box 29237, Fendalton, Christchurch 4800, New Zealandb Centre for Sustainable Forest Management, Forest Research, Private Bag 3020, Rotorua, New Zealand
c Landcare Research, Private Bag 11052, Palmerston North, New Zealandd Landcare Research, Private Bag 92170, Auckland, New Zealand
Received 1 April 2005; received in revised form 19 May 2005; accepted 20 May 2005
Abstract
New Zealand is committed to developing sustainable forest management practices as evidenced through Government
involvement in international forestry agreements such as the Montreal Process, and the forestry sector’s adoption of forest
certification mechanisms. In support of this commitment, it has been identified that there is little quantitative evidence of the
interactions of plantation forestry on site quality and long-term site productivity. To address this issue, a nationwide study of site
quality was initiated at 35 key sites covering the range of edaphic and environmental conditions representing the productivity
envelope for New Zealand plantation forests. At each location, within the productivity envelope, eight short-term site quality
plots were planted at a very high stand density (40,000 stems ha�1) to rapidly identify key soil indicators of growth which may
be useful for determining site sustainability. In addition, a permanent sample plot was established by planting seedlings at
conventional stem densities (500–1100 stems ha�1). At each site, a factorial design was applied with the following three factors:
species (Pinus radiata and Cupressus lusitanica), fertiliser (no fertiliser and nutrients supplied in excess of crop demands) and
disturbance (low and high disturbances). After two years of increment, initial analyses are presented which partition treatment
and site effects on increment and identify key soil properties that influence increment of the two species.
Volume increment over the two-year period was most strongly influenced by site, ranging 50-fold and 15-fold across sites for
C. lusitanica and P. radiata, respectively. For the treatments, species accounted for most of the variance in increment, with mean
volume increment across all sites of P. radiata significantly exceeding that of C. lusitanica by 56%. Fertilisation also
significantly influenced volume increment inducing mean gains of 30%. Disturbance had a significant, but comparatively weak
effect, reducing mean volume increment by 9%. After correction had been made for environment (temperature and rainfall), soil
properties that were most strongly related to volume increment for both species included CN ratio, total soil nitrogen, total soil
www.elsevier.com/locate/foreco
Forest Ecology and Management 216 (2005) 51–63
* Corresponding author. Tel.: +64 3 364 2949; fax: +64 3 364 2812.
E-mail address: [email protected] (M.S. Watt).
0378-1127/$ – see front matter # 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2005.05.064

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6352
1. Introduction
New Zealand is a member of the 12 country
Working Group on Criteria and Indicators for the
Conservation and Sustainable Management of Tem-
perate and Boreal Forests. ‘‘The Montreal Process’’
(Anon., 1995) and the New Zealand Government have
agreed to report on progress towards sustainable forest
management as measured by indicators grouped
within seven criteria. Criterion 2, ‘Maintenance of
Productive Capacity of Forest Ecosystems’, and
criterion 4, ‘Conservation and Maintenance of Soil
and Water Resources’, address the important issue of
whether plantations can be grown for an indefinite
number of rotations without adversely affecting soil
quality or the site’s capacity for net primary
production. Although the Montreal Process and
certification schemes identify potential indicators that
can be used to describe site quality, foresters need
tools, guidelines or management systems to enable
them to turn these high-level, subjective sustainability
goals into indicators that can be measured quantita-
tively (Richardson et al., 1999).
Despite the importance of sustainability for the
long-term viability of the forestry sector, there is
little direct evidence to indicate how successive
rotations, characteristic of plantation forestry, influ-
ence site productivity. As well-designed long-term
field trials for investigating sustainability are rare,
evidence demonstrating changes in productivity are
uncommon, even at a regional level (Dyck and Cole,
1990). Within New Zealand, there are considerable
data that may be used to examine temporal changes
in productivity of the most commonly planted
plantation species Pinus radiata, as annual growth
has been measured in permanent sample plots in
some plantations for over 60 years (Woollons, 2000).
However, these data are of questionable value in
assessing temporal changes in soil quality because
they are invariably confounded by changes over time
in management practices, tree genotype and climate
(Morris and Miller, 1994; Richardson et al., 1999;
Woollons, 2000). Mechanistic models are also of
limited use in assessing changes in site productivity
as our knowledge of many of the key ecosystem
processes is incomplete (Yarie, 1990).
The importance of well-designed long-term field
experiments for detecting changes in site productivity
over time has been emphasised by many researchers
(Adlard et al., 1984; Dyck and Cole, 1990; Richardson
et al., 1999). Ideally, these trials should be installed
across a wide environmental range and run over a
number of successive rotations. Within the trials, it is
vital to control management and genotype factors and
monitor climatic parameters, so that any changes in
productivity can be correctly attributed to alterations
in site quality (Morris and Miller, 1994).
Although long-term field trials are important for
determining if site productivity changes over time,
these trials do little to elucidate the mechanisms
causing alterations to site quality (Adlard et al., 1984;
Richardson et al., 1999). Understanding these
mechanisms is essential as it provides the information
necessary for managers to predict and prevent or
ameliorate declining site quality. Measurements of
soil properties are useful in this regard as indicators of
changes in site quality and provide considerable
information on factors which result in changes in site
quality and productivity over time. As forest
productivity is an important indicator of sustainability,
these soil properties must be related to measures of
performance such as net primary production (Richard-
son et al., 1999). Use of this information and the
development of relationships between various man-
agement practices and soil properties would enable
site-specific recommendations to be made on manage-
ment practices necessary to maintain the productive
capacity of the site (Richardson et al., 1999).
One approach that could be used to rapidly
determine the key soil properties influencing site
phosphorus, organic phosphorus and depth of the A horizon. When soil properties were included in combination, the best
predictive models of volume increment formulated for both species included rainfall, temperature, the product of total soil
nitrogen and total soil phosphorus and depth of the A horizon.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Sustainability; Forest productivity; Soil quality; Indicators; Pinus radiata; Cupressus lusitanica; New Zealand

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 53
productivity is experimentation in miniature. This
method, which uses small scale, highly stocked plots
to rapidly induce site productivity limitations and
compress the length of the rotation, has considerable
merit (Amateis et al., 2003a). Previous research shows
that trees grown at these high stand densities over short
periods mimic growth patterns of trees growing
over longer periods at conventional stand densities
(Amateis et al., 2003a,b; Sharma et al., 2003). As the
experimental cycle is shortened, this approach is more
efficient and cost effective than the same experiment
conducted at an operational scale. These greater
efficiencies allow relatively large numbers of plots to
be installed, which permits increased coverage across
a wide range of environments. Another key advantage
of this approach is that the small plot area used is
subject to less edaphic variation, which makes it easier
to characterise soil properties within the plot (Amateis
et al., 2003a).
Although these small plots are relatively short-
lived, the use of high stand densities should ensure that
there is significant demand for resources from the site
relatively quickly, thus enabling the key soil properties
influencing productivity to be identified. Experimen-
tation in miniature has been successfully used to link
productivity with changes in key soil properties
induced by different management practices at a single
site (Kelting et al., 1999). In this study, small highly
stocked plots (spacing 0.3 m � 0.3 m; stand density of
111,111 stems ha�1) of Pinus taeda grown over a one-
year period were used to validate a soil quality index
(SQI) model developed from literature based suffi-
ciency curves for five key growth determining
attributes of forest soils. Using predetermined
sufficiency curves developed from juvenile and
mature stands (see Kelting et al., 1999 for references),
the SQI accounted for 60% of the variation in tree
volume, with three of the five attributes (water table
depth, soil aeration depth and net N mineralisation)
exhibiting significant relationships with volume.
These results suggest that experimentation in minia-
ture may provide a useful means of rapidly identifying
key soil properties influencing tree growth and site
sustainability.
Following this rationale, a nationwide study of site
quality using mini plantations was initiated at 35 key
sites covering the range of edaphic and environmental
conditions for New Zealand plantation forests. An
important component of this study is the establishment
of new permanent sample plots at each study site to
determine the long-term effects of forest and forest
operations on soil quality and site productivity and
validate the short-term experimentation.
This paper describes the plot location, layout and
measurements taken and documents the range of
climatic and edaphic properties occurring across the
trial series. Using data collected from two years of
increment in the productivity envelope plots, addi-
tional objectives were to: (i) investigate how treat-
ments (species, fertilisation and disturbance) and site
influence productivity; (ii) identify key soil indicators
of productivity; (iii) examine whether these soil
indicators vary between species.
2. Materials and methods
2.1. Location of site quality plots
Sites were selected to represent the range in soil
properties on which plantation forests are currently
grown in New Zealand (Fig. 1). Soils were grouped
according to the New Zealand soil classification
(Hewitt, 1998), which recognises 15 soil orders.
Plantation forests within New Zealand occur on 9 of
these 15 soil orders, namely the Allophanic, Brown,
Oxidic, Pallic, Podzol, Pumice, Raw, Recent and Ultic
soil orders. The trial series was established on all of
these soil orders apart from Oxidic, which represents a
small proportion of the area (0.7%) on which the forest
estate is planted (R. Simcock, personal communica-
tion). The number of sites established on each soil
order was weighted to be representative of the
corresponding plantation area on which the soil order
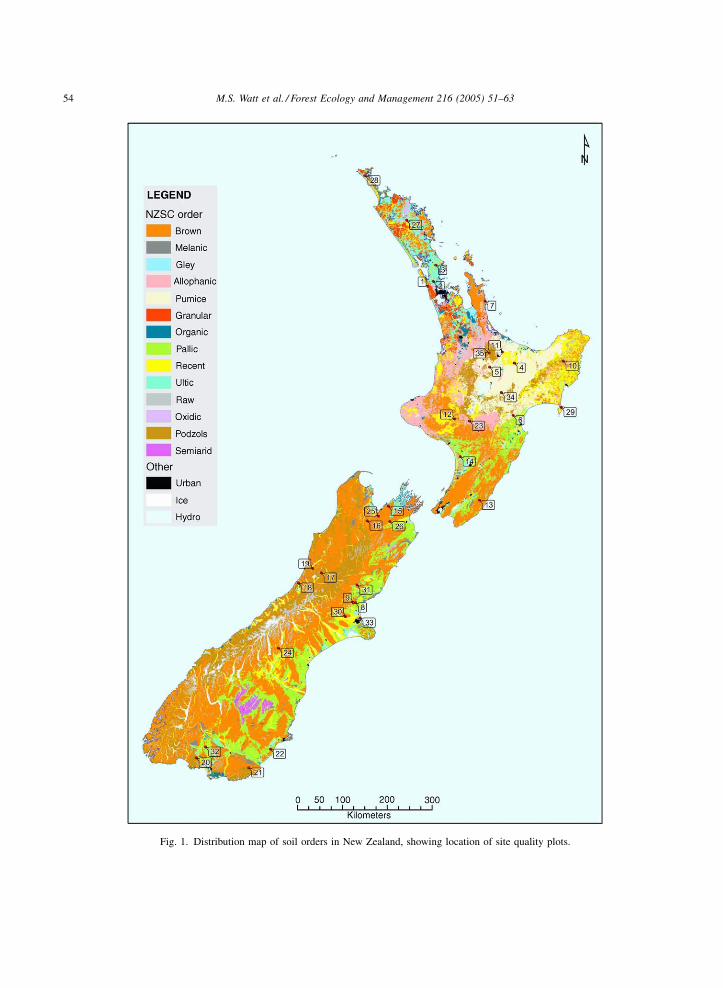
is found (Fig. 1).
Sites were further screened using climatic surfaces
to ensure that selected areas represented the con-
siderable range in meteorological conditions found
throughout New Zealand forest plantations. Long-
term climate data obtained from thin-plate spline
surfaces (Hutchinson and Gessler, 1994) fitted to
meteorological station data (Leathwick and Stephens,
1998) were used to determine climatic conditions at
each selected site. When compared to long-term
average values for all plantation forests (Anon., 1983),
the 35 selected sites almost completely encompass the
M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6354
Fig. 1. Distribution map of soil orders in New Zealand, showing location of site quality plots.
M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 55
Table 1
Comparison of the range in mean annual temperature, solar radiation, relative humidity (recorded at 09:00 NZST) and total annual rainfall, at the
selected 35 sites, with that found throughout New Zealand plantation forests
Selected 35 sites Plantation forests
Minimum Mean Maximum Minimum Maximum
Temperature (8C) 8.6 11.5 (0.3) 15.5 8.0 15.6
Solar radiation (MJ m�2) 12.0 14.3 (0.16) 15.4 12.0 15.4
Relative humiditya (%) 69 79 (0.8) 87 66 87
Rainfall (mm) 609 1560 (121) 3718 609 3718
The mean � standard error for the selected 35 sites is shown.a Recorded at 09:00 NZST.
range in total annual rainfall and mean annual
temperature, relative humidity and solar radiation
(Table 1).
2.2. Experimental design
Site quality plots were located at each of the 35
selected locations. At each location, a series of eight
plots was installed using a factorial design with the
following three factors: species (P. radiata and
Cupressus lusitanica), fertiliser (no fertiliser and
nutrients supplied in excess of crop demands) and
disturbance (low and high disturbances). P. radiata
was chosen as it is the most common plantation
species within New Zealand, while C. lusitanica was
selected as it is a commonly planted species on more
fertile farm sites. Quantities of fertiliser were applied
to treated plots to ensure that there were no nutrient
limitations. Disturbed plots were located on areas
compacted by previous harvesting operations. Each
plot was small in size (3 m � 3 m) and contained
nine measurement trees spaced at 0.5 m � 0.5 m
(40,000 stems ha�1) surrounded by a two-row buffer.
Regular applications of herbicide were made to
ensure weed-free conditions. All sites were planted
with seed stock of the same origin (growth and
form factor of 19 (Vincent and Dunstan, 1989) for
P. radiata and 99/275 for C. lusitanica) sourced from
the Forest Research nursery in Rotorua. At each of
the 35 selected locations, permanent 40 m � 40 m
sample plots were installed adjacent to the site
quality plots at conventional stand densities (500–
1100 stems ha�1) using P. radiata seedlings with a
growth and form factor of 19. The site quality plots
were installed over a three-year period from 2000 to
2002.
2.3. Measurements
Elevation, longitude, latitude, aspect and slope
were recorded at each location. Within the site quality
plots tree ground-line diameter, height and crown
width are measured annually. Measurements of
photosynthetically active photon flux density, air
temperature and relative humidity are taken from
sensors installed on a 3 m tower located adjacent to the
experimental plots. A tipping bucket rain gauge
positioned on top of the tower records above-canopy
rainfall.
A comprehensive set of soil chemical and physical
measurements were taken from the site quality plots
prior to planting and fertilisation. Soil chemistry
samples were taken from 0 to 100 mm depth at five
points in each of the four undisturbed plots to give 20
cores per bag. This process was repeated for the four
disturbed plots at each location. Sampling points were
at least 1 m apart near the centre of the plots. These
samples were bulked by disturbance class, then
analysed for soil moisture, pH in water, total carbon
(C), total soil nitrogen (N), total soil phosphorus (P),
organic P, Bray P, Olsen P, P retention, exchangeable
bases and CEC, following the methods described by
Blakemore et al. (1987). Results are expressed on an
oven dry basis (105 8C). Soil physical properties weremeasured on samples of mineral soil taken from the
edges of disturbed and undisturbed plots. Air capacity
(at 10 kPa), macroporosity (at 5 kPa), bulk density,
particle density, cone penetration resistance (at
10 kPa) and total available water were determined
from 5 to 200 mm depth, following the procedures
described by Gradwell (1972).
A soil pit was excavated in undisturbed soil
adjacent to each site quality plot to supplement soil

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6356
property measurements taken from the upper mineral
horizons. From this pit, all previously described
physical and chemical soil properties were determined
by horizon to depths of up to 1 m. Particle size was
measured by dispersing the field-moist soil in water
with an ultrasonic probe and separating the <2, 2–63
and >63 mm fractions by sedimentation. The depth of
all horizons, profile drainage class (Hewitt, 1993),
rooting depth and depth to mottling were also
recorded. Each soil was described pedologically and
classified to subgroup level following the New
Zealand Soil Classification (Hewitt, 1993). As organic
horizons were not always present and were usually
less than 30 mm thick, data from these layers have not
been included.
2.4. Data analyses
Analyses presented in this paper use data collected
from the complete series of 35 site quality plots over the
first two years of the experiment. Data for four plots
were excluded from all analyses due to the excessive
mortality within these plots. All analyses were under-
taken in SAS (SAS Institute, 1996). Variables were
tested for normality and homogeneity of variance and
transformations made as necessary to meet the under-
lying statistical assumptions of the models used.
A two-way analysis of variance (ANOVA) was
used to test for the main effects of disturbance and site
on soil physical and chemical properties within the top
100 mm of the mineral horizon. The main and
interactive influence of site and treatment (fertiliser,
species and disturbance) on height, diameter and
volume increment over the first two years (defined as
final values less initial values), was tested using a
generalised linear model (GLM). Initial size was
included as a covariate in the model when found to be
significantly related to increment.
Key soil properties influencing volume increment
were determined for each species using data from
unfertilised plots for which soil property measure-
ments were available. Stem volumewas selected as the
dependant variable as it is considered to be a better
measure of productivity than either height or diameter
(Jackson and Gifford, 1974). Stem volume (V) was
determined from tree height (h) and diameter
measured at ground level (D) for both P. radiata
and C. lusitanica using the following equation
previously found to be applicable to stands covering
a range of ages and stockings (Beets, unpublished
data):
V ¼�p
�D
2
�2
h
�0:25 (1)
To remove the effect of climate, base models of
volume increment were separately constructed for each
species using temperature and rainfall as independent
variables. Univariate relationships between corrected
volume increment and soil properties were then
examined by singly adding soil properties to the two
base models. Significance, functional form and strength
of these univariate relationships were compared
between species.
A multiple regression model for each species was
also constructed using a combination of edaphic
properties. Using appropriate functional forms and
any necessary transformations, variables were sequen-
tially introduced into each base model. Variables were
only retained if inclusion significantly improved the
model and parameter values for the included variables
were significant (P < 0.05). In constructing the two
final models, emphasis was placed on developing
simple equations with very little apparent bias.
3. Results
3.1. Impact of site and disturbance on soil
properties
The selected sites included extremes in soil texture
ranging from single-grained scoria and sandy soils to
clay loam soils. Variation in soil texture was most
pronounced for the sand (5–96%) and silt fractions (3–
79%). All physical and chemical properties sampled
significantly (P < 0.001) varied across sites (Table 2).
Disturbance had a significant impact on all soil
physical properties apart from total available water
(Table 2). The closely related variables, air capacity
and macroporosity, were most strongly affected by
disturbance (partial r2 = 0.16 and 0.12, respectively).
Of the soil physical properties examined, macro-
porosity exhibited the greatest range with values in the
undisturbed treatment exceeding those in the disturbed
treatment by 47% (21 versus 14 v/v). For chemical
M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 57
Table 2
Characteristics of soil physical and chemical properties in the top 100 mm of the mineral soil for disturbed and undisturbed plots sampled at 31
sites before planting
Undisturbed plots Disturbed plots Analysis of variance
Mean Range Mean Range Site Dist.
Physical properties
Bulk density (g cm�3) 1.0 0.5–1.4 1.1 0.5–1.6 0.90*** 0.04***
Particle density (g cm�3) 2.5 2.2–3.0 2.5 2.2–3.0 0.99*** 0.005*
Penetration resistance (MPa) 0.8 0.3–2.1 1.1 0.2–2.1 0.81*** 0.07***
Total porosity (%, v/v) 62 45–78 58 46–79 0.85*** 0.07***
Macroporosity (%) 21 4–49 14 4–32 0.69*** 0.16***
Air capacity (%) 24 4–45 18 5–38 0.78*** 0.12***
Total available water (%, v/v) 19 9–32 19 4–35 0.91*** ns
Chemical properties
Carbon (%) 6.0 0.7–21.6 5.2 1.0–28.9 0.86*** 0.02*
Total N (%) 0.29 0.02–0.67 0.25 0.03–0.79 0.89*** 0.02*
CN ratio 22 11–46 22 11–38 0.97*** ns
pH 5.2 4.1–6.0 5.2 4.2–6.2 0.94*** ns
CEC (cmol g�1) 22 2–72 21 3–76 0.97*** ns
Exchangeable Na (cmol g�1) 0.19 0.02–0.77 0.16 0.01–0.56 0.85*** ns
Exchangeable K (cmol g�1) 0.47 0.01–1.15 0.43 0.03–0.85 0.88*** ns
Exchangeable Mg (cmol g�1) 1.9 0.1–12.0 1.7 0.3–9.8 0.97*** ns
Exchangeable Ca (cmol g�1) 4.4 0.2–18.7 4.1 0.4–18.7 0.94*** ns
Sum bases (cmol g�1) 7.0 0.3–23.6 6.4 0.8–22.2 0.95*** ns
Base saturation (%) 33 7–95 34 5–89 0.96*** ns
P retention (%) 42 3–96 40 0–89 0.97*** ns
Olsen P (mg g�1) 8.3 2.3–29.0 7.6 0.9–24.2 0.88*** ns
Bray P (mg g�1) 20 3–82 20 3–106 0.94*** ns
Inorganic P (mg g�1) 150 15–326 140 18–457 0.93*** ns
Organic P (mg g�1) 314 21–675 281 28–595 0.93*** ns
Total P (mg g�1) 464 37–937 420 62–838 0.93*** 0.01*
The influence of disturbance (Dist.) and site on each soil property is shown by partial r2-values followed by significance levels. ns: non-
significant at P = 0.05.* Significant at P = 0.05.
*** Significant at P = 0.001.
properties, disturbance significantly reduced concen-
trations of total soil N, P and C (Table 2).
3.2. Site and treatment influences on tree
increment
Increment over the two-year period was most
strongly influenced by site (Table 3). Site significantly
(P < 0.001) influenced increment in all characteristics
accounting for 61, 76 and 49% of the total variance for
tree diameter, height and volume increment, respec-
tively. Stem volume increment was a more sensitive
indicator of site quality than either tree height or
diameter increment, exhibiting 50-fold and 15-fold
variations in volume increment, respectively, for
C. lusitanica and P. radiata (Fig. 2).
Treatment influences on tree characteristics
became more pronounced as the experiment pro-
gressed, especially for stem volume (Fig. 3). The
maximum treatment differences for stem volume
(averaged across sites) increased from 33% (4.8–
6.4 m3 ha�1) at the end of year 1 to two-fold (7.5–
17.4 m3 ha�1) at the end of year 2. Treatment effects,
particularly those associated with species, were least
marked for tree height increment (Table 3).
For all characteristics examined increment of
P. radiata significantly (P < 0.001) exceeded that of
C. lusitanica (Tables 3 and 4). Fertilisation also
significantly influenced all tree characteristics (P <0.001), inducing gains of 30, 8 and 13% for stem
volume, height and diameter increment, respectively
(Tables 3 and 4). Disturbance significantly influenced
M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6358
Table 3
Main and interactive effects of initial size, site and treatments
(species, disturbance and fertilisation) on tree diameter, height
and volume increment over two years
Dependant variable
Diameter Height Volume
Initial size ns ns 0.25*
Site 0.61*** 0.76*** 0.49***
Species (Sp.) 0.10*** 0.02*** 0.05***
Fertilisation (Fert.) 0.04*** 0.01*** 0.03***
Disturbance (Dist.) ns 0.01** 0.01*
Site � Sp. 0.07*** 0.06*** 0.05***
Site � Fert. ns 0.03*** 0.03*
Site � Dist. 0.06*** 0.04*** 0.04***
Sp. � Fert. ns ns ns
Sp. � Dist. ns ns ns
Dist. � Fert. ns ns ns
Sp. � Dist. � Fert. ns ns ns
The partial r2 of each significant variable is shown with the
significance level.* Significant at P = 0.05.** Significant at P = 0.01.*** Significant at P = 0.001.
Fig. 2. Boxplots showing site variation in growth increment over
two years for: (a) tree diameter, (b) tree height and (c) tree volume
for Cupressus lusitanica and Pinus radiata. The boundaries of the
boxplot closest to zero indicates the 25th percentile, the centre lines
within the boxes indicate the median and the boundaries of the boxes
farthest from zero indicate the 75th percentile. Error bars are the 5th
and 95th percentiles, and filled symbols represent data outlying the
5th and 95th percentiles.
tree height (P < 0.01) and volume increment
(P < 0.05) but not diameter increment. Reductions
in increment induced by disturbance were consider-
ably lower than gains obtained through fertilisation for
all tree characteristics apart from height. In contrast to
fertilisation, tree height was more strongly influenced
by disturbance than tree diameter (Table 3).
The interactions were significant for almost all
combinations of site by treatment, with the single
exception being the fertiliser by site interaction for tree
diameter (Table 3). Site interactions were strongest for
species, intermediate for disturbance and weakest for
fertiliser. In contrast, none of the interactions examined
between the species, fertiliser and disturbance treat-
ments were significant (P > 0.05) suggesting that
disturbance and fertilisation have independent influ-
ences on increment and both species respond similarly
to fertiliser and disturbance. This was confirmed by
examination of treatment means which show a
consistent ranking in disturbance and fertiliser treat-
ments (UF > DF > UN > DN) for both species
(Table 4).
3.3. Identification of key soil properties
For both species, temperature and rainfall were
significantly related to volume increment. Inclusion of
both variables in the model as second-order poly-
nomials accounted for 46 and 33% of total variation in
volume increment for P. radiata and C. lusitanica,
respectively.
After correction had been made for climate, the soil
properties most strongly related to increment for both
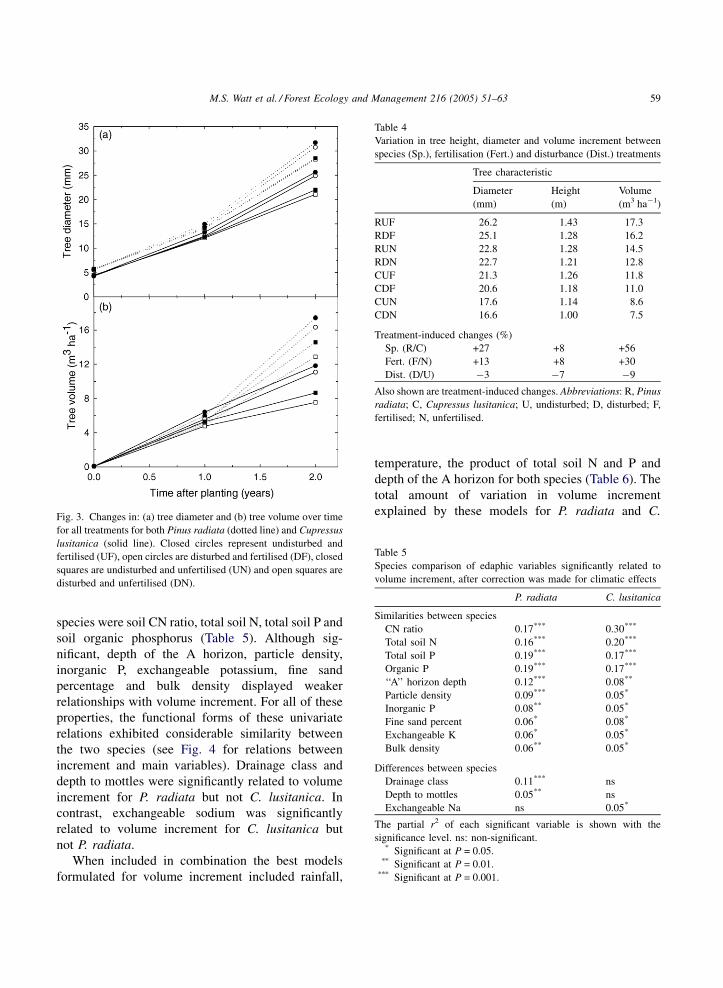
M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 59
Fig. 3. Changes in: (a) tree diameter and (b) tree volume over time
for all treatments for both Pinus radiata (dotted line) and Cupressus
lusitanica (solid line). Closed circles represent undisturbed and
fertilised (UF), open circles are disturbed and fertilised (DF), closed
squares are undisturbed and unfertilised (UN) and open squares are
disturbed and unfertilised (DN).
Table 4
Variation in tree height, diameter and volume increment between
species (Sp.), fertilisation (Fert.) and disturbance (Dist.) treatments
Tree characteristic
Diameter
(mm)
Height
(m)
Volume
(m3 ha�1)
RUF 26.2 1.43 17.3
RDF 25.1 1.28 16.2
RUN 22.8 1.28 14.5
RDN 22.7 1.21 12.8
CUF 21.3 1.26 11.8
CDF 20.6 1.18 11.0
CUN 17.6 1.14 8.6
CDN 16.6 1.00 7.5
Treatment-induced changes (%)
Sp. (R/C) +27 +8 +56
Fert. (F/N) +13 +8 +30
Dist. (D/U) �3 �7 �9
Also shown are treatment-induced changes. Abbreviations: R, Pinus
radiata; C, Cupressus lusitanica; U, undisturbed; D, disturbed; F,
fertilised; N, unfertilised.
Table 5
Species comparison of edaphic variables significantly related to
volume increment, after correction was made for climatic effects
P. radiata C. lusitanica
Similarities between species
CN ratio 0.17*** 0.30***
Total soil N 0.16*** 0.20***
Total soil P 0.19*** 0.17***
Organic P 0.19*** 0.17***
‘‘A’’ horizon depth 0.12*** 0.08**
Particle density 0.09*** 0.05*
Inorganic P 0.08** 0.05*
Fine sand percent 0.06* 0.08*
Exchangeable K 0.06* 0.05*
Bulk density 0.06** 0.05*
Differences between species
Drainage class 0.11*** ns
Depth to mottles 0.05** ns
Exchangeable Na ns 0.05*
The partial r2 of each significant variable is shown with the
significance level. ns: non-significant.* Significant at P = 0.05.** Significant at P = 0.01.*** Significant at P = 0.001.
species were soil CN ratio, total soil N, total soil P and
soil organic phosphorus (Table 5). Although sig-
nificant, depth of the A horizon, particle density,
inorganic P, exchangeable potassium, fine sand
percentage and bulk density displayed weaker
relationships with volume increment. For all of these
properties, the functional forms of these univariate
relations exhibited considerable similarity between
the two species (see Fig. 4 for relations between
increment and main variables). Drainage class and
depth to mottles were significantly related to volume
increment for P. radiata but not C. lusitanica. In
contrast, exchangeable sodium was significantly
related to volume increment for C. lusitanica but
not P. radiata.
When included in combination the best models
formulated for volume increment included rainfall,
temperature, the product of total soil N and P and
depth of the A horizon for both species (Table 6). The
total amount of variation in volume increment
explained by these models for P. radiata and C.

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6360
Fig. 4. Response curves of volume increment for Pinus radiata (thick line) andCupressus lusitanica (thin line) plotted against: (a) total soil P, (b)
total soil N, (c) CN ratio and (d) A horizon depth. All other variables in the model were held at mean values when each response curve was
generated.
lusitanica was 72 and 57%, respectively. Both models
exhibited little apparent bias when residuals were
plotted against either predicted values or independent
variables.
Table 6
Summary of statistics for the final predictive models of volume
increment for Pinus radiata and Cupressus lusitanica
P. radiata C. lusitanica
Rainfall 0.31*** 0.23***
Temperature 0.15*** 0.08*
N � P 0.20*** 0.18***
‘‘A’’ horizon depth 0.06*** 0.06**
Total r2 0.72 0.57
The partial r2 of each significant variable is shown with the
significance level of the variable tested using the F-test. Also shown
is the total r2 for the complete model.* Significant at P = 0.05.** Significant at P = 0.01.*** Significant at P = 0.001.
4. Discussion
Indicators of productivity found in this study have
been regularly cited as determinants of site quality in
both forestry and agricultural settings (see Schoen-
holtz et al., 2000 for review). Total soil N and P
concentrations are the most commonly cited elemental
indicators of productivity. Although not as widely
employed, soil CN ratio has been used to model
growth (Page, 1976) and provides an index of the rate
of nitrogen mineralisation. Soil bulk density is
commonly used as an important determinant of
productivity as it can affect root growth and a host
of soil properties and processes which influence water
and oxygen supply (Schoenholtz et al., 2000). Depth
to mottling and total porosity have been previously
used as variables for determining sufficiency of
aeration within site quality models (Gale et al., 1991).

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 61
For P. radiata, the variables selected for the final
model in this study (Table 6) are very similar to those
found to be the dominant influences on increment in
mature stands. In a nationwide study covering a
similar range of sites to this study, Jackson and Gifford
(1974) found 66% of periodic volume increment was
attributable to mean annual rainfall, seasonal dis-
tribution of rainfall, temperature departures from
optimum, soil depth, total soil N and available P.
Although A horizon depth was not measured in the
Jackson and Gifford study, more recent analysis
(Hunter and Gibson, 1984; Woollons et al., 2002) has
found average depth of the A horizon to be a
significant determinant of site index (mean height of
the 100 largest trees per hectare in terms of diameter,
at age 20 years) for P. radiata. The close correspon-
dence in soil properties related to productivity found
between our study and mature P. radiata stands
supports findings by Kelting et al. (1999) and suggests
that experimentation in miniature may provide a
useful and rapid means of assessing sustainability.
Research strongly suggests that trees grown at very
high stand densities follow a similar developmental
pattern as operational stands. Studies using P. taeda
show that the pattern of diameter and height growth for
trees grown at operational stand densities over 16
years closely corresponds to the pattern of develop-
ment over 4 years when the distance between trees is
reduced to 1/16th of the operational stand distance
(Amateis et al., 2003a,b; Sharma et al., 2003).
Recently obtained data from our oldest site quality
plots (Watt, unpublished data) confirm this similarity
in growth pattern and also show that the average rate of
volume increment (mean annual increment) in site
quality plots over the first four years closely matches
that of stands growing at final operational stockings
(300 stems ha�1) over a 30-year period. These results
suggest that young trees grown at very high stockings
may impose the same annual demand for resources on
a site as older trees grown at operational stockings.
An important consideration in modelling site
quality is to determine how selected soil properties
differ between species. In this study, both species
exhibited very similar growth responses to the most
important soil chemical properties, and the variables
included in the final model of volume increment were
identical between species. Despite these similarities,
P. radiata exhibited greater sensitivity to the drainage
class of the soil and depth to mottles, which suggests
that this species has a greater requirement for a well
aerated soil than C. lusitanica. Subtle differences in
responses to soil properties between species were also
apparent for total soil N, P and the CN ratio. Total soil
N and CN ratio were more important indicators of
productivity for C. lusitanica than total and organic P,
whereas the opposite held for P. radiata. This may
reflect their contrasting mycorrhizal status, with P.
radiata associating with ectomycorrhizal species,
whereas C. lusitanica associates with endomycor-
rhizal species. As well as facilitating the acquisition of
nutrient ions, especially of N and P, recent studies have
shown ectomycorrhizae may mobilise N from organic
polymers as well, enabling the host species to obtain N
directly from plant and microbial detritus sources
(Read and Perez-Moreno, 2003). Thus, N is likely to
be less limiting for P. radiata than C. lusitanica.
Soil disturbance from harvesting induced significant
and substantial differences in soil physical properties.
Results indicate that most of the variation between
disturbed and undisturbed treatmentswas attributable to
macroporosity, air capacity, cone penetration resistance,
total porosity and to a lesser extent bulk density, a
finding which is consistent with previous research
(McMahon et al., 1999; Merino and Edeso, 1999;
Merino et al., 1998). In the present study, concentrations
of total soil C, N and P were reduced by disturbance.
Other researchers have also found that disturbance
significantly reduces soil concentrations of N (Merino
and Edeso, 1999) and P (Mroz et al., 1985; Tuttle et al.,
1985). Reductions in concentrations of these elements
may have been caused through removal of the humus
layer and topsoil in disturbed plots during harvesting,
reducing the quantity of available nutrients.
Despite the clear disturbance-induced alterations to
soil physical properties, increment reductions attri-
butable to disturbance were relatively small, and did
not appear to be strongly related to the most influenced
properties, macroporosity or cone penetration resis-
tance. This may be due to the low number of sites in
which macroporosity was below the 10% threshold
thought to be limiting to root growth (Greacen and
Sands, 1980). Similarly, cone penetration resistance
values did not exceed values (>2.5 MPa) cited by
Greacen and Sands (1980) as being limiting to root
growth. In contrast, bulk density was more strongly
related to increment for both species between sites

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–6362
than air-filled porosity, macroporosity or cone
penetration resistance. High bulk densities have been
shown to cause low porosity and low water infiltration
(Rab, 1996) and the values found at some of these
sites were sufficiently high (>1.3 g cm�3) to cause
impedance to root elongation and therefore a
reduction in plant growth (Froehlich, 1979; Gale
et al., 1991). Disturbance-induced differences for
increment were also attributable to total soil N, a result
which is consistent with the strong influence of this
variable on increment across sites.
The strong and significant influence of fertilisation
on volume increment found in this study is consistent
with previous research across a broad range of species
(see Fox, 2000 for review). Gains obtained through
fertilisation were three-fold higher than reductions in
volume increment induced by disturbance. In contrast,
the site interaction was weaker for fertilisation than
disturbance, indicating that fertiliser effects were
relatively consistent across sites. The relative impor-
tance of fertilisation compared to disturbance is
reinforced by the modelling which shows significant
soil indicators to be predominantly chemical proper-
ties. These results highlight the importance of fertiliser
as a silvicultural treatment which can be used to
improve site quality.
Although preliminary, results from this study
highlight soil properties likely to be most important
for monitoring sustainability of site quality within
New Zealand plantations. The soil properties included
in the final model are consistent with key drivers of
productivity within mature stands. If these relation-
ships are confirmed by final measurements in the site
quality plots, the key soil properties will be monitored
within the long-term permanent sample plots. This
monitoring should provide a basis for site-specific
recommendations on management practices necessary
to maintain the productive capacity of New Zealand’s
plantation forests.
Acknowledgements
We are indebted to the numerous forest companies
and private owners for providing sites for the trial
series, and Hugh Wilde, Trevor Webb, Amy Taylor,
Wim Rijkse, Craig Ross and Malcolm Mcleod who
assisted in selection, identification and classification
of soils. The help of technicians at the Landcare
Research Environmental Chemistry Laboratory in
analysing samples is gratefully acknowledged. Dr. E.
Mason and Mr. M. Kimberley provided advice on the
statistical analyses. We are also grateful to helpful
comments provided by the two anonymous referees.
This project was funded by the New Zealand
Foundation for Research Science and Technology
under Contract No. C04X0304, ‘Protecting and
Enhancing the Environment through Forestry’.
References
Adlard, P.G., Johnson, J.A., Evans, J., 1984. A strategy for
detecting productivity change in tropical plantations. In: Grey,
D.C., Schonau, A.P.G., Schutz, C.J. (Eds.), Proceedings,
IUFRO Symposium on Site and Productivity of Fast Growing
Plantations. Pretoria and Pietermaritzburg, South Africa, 30
April–11 May 0-621-08513-8, pp. 857–869.
Amateis, R.L., Sharma, M., Burkhart, H.E., 2003a. Scaling growth
relationships from seedling plots using similarity analysis. For.
Sci. 49, 188–195.
Amateis, R.L., Sharma,M., Burkhart, H.E., 2003b. Using miniature-
scale plantations as experimental tools for assessing sustain-
ability issues. Can. J. For. Res. 33, 450–454.
Anon., 1983. Summaries of Climatological Observations to 1980.
New Zealand Meteorological Service Misc. Pub. 177, Well-
ington, New Zealand.
Anon., 1995. Criteria and indicators for the conservation and
sustainable management of temperate and boreal forests—the
Montreal Process. Canadian Forest Service, Natural Resources
Canada, Hull, Que., Canada, 27 pp.
Blakemore, L.C., Searle, P.L., Daly, B.K., 1987. Methods for
Chemical Analysis of Soils. Department of Scientific and
Industrial Research, New Zealand Soil Bureau Scientific Report
No. 80.
Dyck, W.J., Cole, D.W., 1990. Requirements for site productivity
research. In: Dyck,W.J., Mees, C.A. (Eds.), Impacts of Intensive
Harvesting on Forest Site Productivity. Proceedings of the IEA/
BE A3 Workshop, South Island, New Zealand, March 1989.
IEA/BE T6/A6 Report No. 2. Forest Research Institute, Rotorua,
New Zealand, FRI Bulletin No. 159, pp. 159–170.
Froehlich, H.A., 1979. Soil compaction from logging equipment:
effects on growth of young ponderosa pine. J. Soil Water Cons.
48, 136–139.
Fox, T.R., 2000. Sustained productivity in intensively managed
forest plantations. For. Ecol. Manage. 138, 187–202.
Gale, M.R., Grigal, D.F., Harding, R.B., 1991. Soil productivity
index: predictions of site quality for white spruce plantations.
Soil Sci. Soc. Am. J. 55, 1708–1710.
Gradwell, M.W., 1972. Methods for Physical Analysis of Soils. New
Zealand Soil Bureau Scientific Report No. 10C. Wellington,
DSIR.

M.S. Watt et al. / Forest Ecology and Management 216 (2005) 51–63 63
Greacen, E.L., Sands, R., 1980. Compaction of forest soils, a review.
Aust. J. Soil Res. 18, 163–189.
Hewitt, A.E., 1993. Methods and Rationale of the New Zealand
Soil Classification. Landcare Research Science Series No. 2.
Manaaki-Whenua-Landcare Research New Zealand Ltd.,
Lincoln, New Zealand, 71 pp.
Hewitt, A.E., 1998. New Zealand Soil Classification. Landcare
Research Science Series No.1, second ed. Manaaki-Whenua-
Landcare Research New Zealand Ltd., Lincoln, New Zealand.
Hunter, I.R., Gibson, A.R., 1984. Predicting Pinus radiata site index
from environmental variables. N. Z. J. For. Sci. 14, 53–64.
Hutchinson, M.F., Gessler, P.E., 1994. Splines—more than just a
smooth interpolator. Geoderma 62, 45–67.
Jackson, D.S., Gifford, H.H., 1974. Environmental variables influ-
encing the increment of radiata pine. (1) Periodic volume
increment. N. Z. J. For. Sci. 4, 3–26.
Kelting, D.L., Burger, J.A., Patterson, S.C., Aust, W.M., Miwa, M.,
Trettin, C.C., 1999. Soil quality assessment in domesticated
forests—a southern pine example. For. Ecol. Manage. 122, 167–
185.
Leathwick, J.R., Stephens, R.T.T., 1998. Climate Surfaces for New
Zealand. Landcare Res. Contract Report LC9798/126. Landcare
Research, Lincoln, New Zealand, 19 pp.
McMahon, S., Simcock, R., Dando, J., Ross, C., 1999. A fresh look
at operational soil compaction. N. Z. J. For. 44, 33–37.
Merino, A., Edeso, J.M., Gonzalez, M.J., Marauri, P., 1998. Soil
properties in a hilly area following different harvesting manage-
ment practices. For. Ecol. Manage. 103, 235–246.
Merino, A., Edeso, J.M., 1999. Soil fertility rehabilitation in young
Pinus radiata D.Don. plantations from northern Spain after
intensive site preparation. For. Ecol. Manage 116, 83–91.
Morris, L.A., Miller, R.E., 1994. Evidence for long-term produc-
tivity change as provided by field trials. In: Dyck, W.J., Cole,
D.W., Comerford, N.B. (Eds.), Impacts of Forest Harvesting on
Long-Term Site Productivity. Chapman and Hall, London, pp.
41–80.
Mroz, G.D., Jurgensen, M.F., Frederick, D.J., 1985. Soil nutrient
changes following whole tree harvesting on three northern
hardwood sites. Soil Sci. Soc. Am. J. 57, 246–255.
Page, G., 1976. Quantitative evaluation of site potential for spruce
and fir in Newfoundland. For. Sci. 22, 131–143.
Rab, M.A., 1996. Soil physical and hydrological properties
following logging and slash burning in the Eucalyptus regnans
forest of southeastern Australia. For. Ecol. Manage. 84, 159–
176.
Read, D.J., Perez-Moreno, J., 2003. Mycorrhizas and nutrient
cycling in ecosystems—a journey towards relevance? New
Phytol. 157, 475–492.
Richardson, B., Skinner, M.F., West, G., 1999. The role of forest
productivity in defining the sustainability of plantation forests in
New Zealand. For. Ecol. Manage. 122, 125–137.
SAS Institute, 1996. SAS/STAT Software: Changes and Enhance-
ments through Release 6.11. SAS Institute Inc., Cary, NC.
Schoenholtz, S.H., Van Miegroet, H., Burger, J.A., 2000. A review
of chemical and physical properties as indicators of forest soil
quality: challenges and opportunities. For. Ecol. Manage. 138,
335–356.
Sharma, M., Amateis, R.L., Burkhart, H.E., 2003. Forest stand
dynamics and similarity theory. Ecol. Model. 167, 165–180.
Tuttle, C.L., Golden, M.S., Meldahl, R.S., 1985. Surface soil
removal and herbicide treatment: effects on soil properties
and loblolly early growth. Soil Sci. Soc. Am. J. 49, 1558–
1562.
Vincent, T.G., Dunstan, J.S., 1989. Register of Commercial Seedlots
Issued by the New Zealand Forest Service. Ministry of Forestry,
Rotorua. Forest Research Institute Bulletin No. 144. Forest
Research Institute, Rotorua, New Zealand, 155 pp.
Woollons, R.C., 2000. Comparison of growth of Pinus radiata over
two rotations in the central North Island of New Zealand. Int.
For. Rev. 2 (2), 84–89.
Woollons, R.C., Skinner, M.F., Richardson, M.F., Rijske, W.C.,
2002. Utility of ‘‘A’’ horizon soil characteristics to separate
pedological groupings, and their influence with climatic and
topographic variables on Pinus radiata height growth. N. Z. J.
For. Sci. 32 (2), 195–207.
Yarie, J., 1990. Role of computer models in predicting the con-
sequences of management on forest productivity. In: Dyck, W.J.,
Mees, C.A. (Eds.), Impacts of Intensive Harvesting on Forest
Site Productivity. Proceedings of the IEA/BE A3 Workshop,
South Island, New Zealand, March 1989. IEA/BE T6/A6 Report
No. 2. Forest Research Institute, Rotorua, New Zealand, FRI
Bulletin No. 159, pp. 3–18.




















