
Decontamination of ochratoxin A by yeasts: possible approaches and factors leading to toxin removal...
13
MINI-REVIEW Decontamination of ochratoxin A by yeasts: possible approaches and factors leading to toxin removal in wine Leonardo Petruzzi & Milena Sinigaglia & Maria Rosaria Corbo & Daniela Campaniello & Barbara Speranza & Antonio Bevilacqua Received: 5 March 2014 /Revised: 30 April 2014 /Accepted: 1 May 2014 # Springer-Verlag Berlin Heidelberg 2014 Abstract Biological decontamination of mycotoxins using microorganisms is one of the well-known strategies for the management of mycotoxins in foods and feeds. Yeasts are an efficient biosorbant, used in winemaking to reduce the con- centration of harmful substances from the must which affect alcoholic fermentation (medium-chain fatty acids) or which affect wine quality in a negative way (ethyl phenols and sulphur products). In recent years, several studies have dem- onstrated the ability of yeasts to remove ochratoxin A (OTA) by live cells, cell walls and cell wall extracts, yeast lees. In spite of the physical and chemical methods applied to re- move the toxin, the biological removal is considered a prom- ising solution, since it is possible to attain the decontamina- tion without using harmful chemicals and without losses in nutrient value or palatability of decontaminated food. In addition, adsorption is recognized as economically viable, technically feasible and socially acceptable. This paper in- tends to review the current achievements of OTA removal mediated by yeasts, the recent updates in the selection of strains acting at the same time as starters and as biological tools to remove OTA and the factors affecting the removal process. Keywords Yeasts . Ochratoxin A . Removal . Starter cultures . Adsorbing tools Introduction The contamination of food and feed by mycotoxins [toxic metabolites of fungi] has been recently characterized by the World Health Organization (WHO) as one of the most signif- icant sources of food-borne illnesses (Anly and Bayram 2009); in fact, according to the Food and Agricultural Organization (FAO) of the United Nations, up to 25 % of the world’ s agricultural commodities have been estimated to be significantly contaminated with mycotoxins (Jard et al. 2011). On the other hand, at least 100 countries have regula- tions regarding levels of mycotoxins in the food and feed industry (van Egmond et al. 2007). Currently, approximately 400 secondary metabolites with toxigenic potential produced by more than 100 moulds have been reported (Jard et al. 2011), and this number is expected to increase, probably due to the rising number of extreme weath- er events (Paterson and Lima 2011). Aspergillus, Penicillium and Fusarium fungal genera were considered the main sources of contamination (Reddy et al. 2010). Some mycotoxins with a strong impact on human health are aflatoxins (AFs), ochratoxin A (OTA), trichothecenes (deoxynivalenol (DON) and T-2 toxin), zearalenone (ZEN) and fumonisins (FMNs) (Afsah-Hejri et al. 2013 ). Unfortunately, these toxic compounds are generally thermo- stable and can remain present in crops even after all signs of the fungus itself have been removed (Inoue et al. 2013). In recent years, OTA has received a special focus due to its toxic effects and high incidence in a wide range of foods, including cereal products, coffee, spices, beer, grape and its derivates and products of animal origin (Covarelli et al. 2012). Ochratoxin A is a nephrotoxin with immunosuppressive, teratogenic and carcinogenic properties, as stated in 1993 by the International Agency for Research on Cancer (IARC). In humans, OTA is frequently reported as the possible causative agent of an endemic kidney disease observed in the Balkans L. Petruzzi : M. Sinigaglia : M. R. Corbo : D. Campaniello : B. Speranza : A. Bevilacqua (*) Department of the Science of Agriculture, Food and Environment, University of Foggia, Via Napoli 25, 71122 Foggia, Italy e-mail: [email protected] A. Bevilacqua e-mail: [email protected] Appl Microbiol Biotechnol DOI 10.1007/s00253-014-5814-4
Transcript of Decontamination of ochratoxin A by yeasts: possible approaches and factors leading to toxin removal...

MINI-REVIEW
Decontamination of ochratoxin A by yeasts: possible approachesand factors leading to toxin removal in wine
Leonardo Petruzzi & Milena Sinigaglia &
Maria Rosaria Corbo & Daniela Campaniello &
Barbara Speranza & Antonio Bevilacqua
Received: 5 March 2014 /Revised: 30 April 2014 /Accepted: 1 May 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Biological decontamination of mycotoxins usingmicroorganisms is one of the well-known strategies for themanagement of mycotoxins in foods and feeds. Yeasts are anefficient biosorbant, used in winemaking to reduce the con-centration of harmful substances from the must which affectalcoholic fermentation (medium-chain fatty acids) or whichaffect wine quality in a negative way (ethyl phenols andsulphur products). In recent years, several studies have dem-onstrated the ability of yeasts to remove ochratoxin A (OTA)by live cells, cell walls and cell wall extracts, yeast lees. Inspite of the physical and chemical methods applied to re-move the toxin, the biological removal is considered a prom-ising solution, since it is possible to attain the decontamina-tion without using harmful chemicals and without losses innutrient value or palatability of decontaminated food. Inaddition, adsorption is recognized as economically viable,technically feasible and socially acceptable. This paper in-tends to review the current achievements of OTA removalmediated by yeasts, the recent updates in the selection ofstrains acting at the same time as starters and as biologicaltools to remove OTA and the factors affecting the removalprocess.
Keywords Yeasts . OchratoxinA . Removal . Startercultures . Adsorbing tools
Introduction
The contamination of food and feed by mycotoxins [toxicmetabolites of fungi] has been recently characterized by theWorld Health Organization (WHO) as one of the most signif-icant sources of food-borne illnesses (Anly and Bayram2009); in fact, according to the Food and AgriculturalOrganization (FAO) of the United Nations, up to 25 % ofthe world’s agricultural commodities have been estimated tobe significantly contaminated with mycotoxins (Jard et al.2011). On the other hand, at least 100 countries have regula-tions regarding levels of mycotoxins in the food and feedindustry (van Egmond et al. 2007).
Currently, approximately 400 secondary metabolites withtoxigenic potential produced by more than 100 moulds havebeen reported (Jard et al. 2011), and this number is expected toincrease, probably due to the rising number of extreme weath-er events (Paterson and Lima 2011). Aspergillus, PenicilliumandFusarium fungal genera were considered the main sourcesof contamination (Reddy et al. 2010).
Some mycotoxins with a strong impact on human healthare aflatoxins (AFs), ochratoxin A (OTA), trichothecenes(deoxynivalenol (DON) and T-2 toxin), zearalenone (ZEN)and fumonisins (FMNs) (Afsah-Hejri et al. 2013).Unfortunately, these toxic compounds are generally thermo-stable and can remain present in crops even after all signs ofthe fungus itself have been removed (Inoue et al. 2013).
In recent years, OTA has received a special focus due to itstoxic effects and high incidence in a wide range of foods,including cereal products, coffee, spices, beer, grape and itsderivates and products of animal origin (Covarelli et al. 2012).
Ochratoxin A is a nephrotoxin with immunosuppressive,teratogenic and carcinogenic properties, as stated in 1993 bythe International Agency for Research on Cancer (IARC). Inhumans, OTA is frequently reported as the possible causativeagent of an endemic kidney disease observed in the Balkans
L. Petruzzi :M. Sinigaglia :M. R. Corbo :D. Campaniello :B. Speranza :A. Bevilacqua (*)Department of the Science of Agriculture, Food and Environment,University of Foggia, Via Napoli 25, 71122 Foggia, Italye-mail: [email protected]
A. Bevilacquae-mail: [email protected]
Appl Microbiol BiotechnolDOI 10.1007/s00253-014-5814-4

(Balkan endemic nephropathy and related urinary tract tu-mors) (Quintela et al. 2013).
Ochratoxin A is continuously recovered from the blood ofhealthy humans, and this datum confirms the continuousexposure to the toxin (Pozo-Bayón et al. 2012); a reason forthis high incidence is probably the importance of wine in thediet (ca. 13 % of the total dietary intake) (ScientificCooperation on Questions relating to Food-SCOOP, task3.2.7; European Commission 2002).
Some authors reviewed in the past the physical, chemicaland biological strategies to remove and/or decrease the con-tent of OTA (Shetty and Jespersen 2006; Amézqueta et al.2009; Abrunhosa et al. 2010; Quintela et al. 2013); however,there is no comprehensive report on yeasts as biological toolsto achieve the bioremediation of the toxin. Therefore, the aimof this paper was to specially focus on the role of yeasts asbiological and bioadsorbent tools to remove the toxin in wine,as well as on the factors acting on this complex phenomenon.
Basic facts on OTA contamination in grape product chain
The presence of OTA in grape juice and wine was reported forthe first time by Zimmerli and Dick (1995, 1996), and sincethen, the toxin was recovered in grapes, musts and wines inseveral Mediterranean countries such as France (Ospital et al.1998), Portugal (Ratola et al. 2004), Greece (Soufleros et al.2003), Italy (Brera et al. 2008) and Spain (Bellí et al. 2004).
Although Penicillium verrucosum and Aspergillusochraceus are considered to be the main OTA-producingspecies, there is strong evidence for the role of Aspergilluscarbonarius in OTA contamination in grape product chain(Quintela et al. 2013). A. carbonarius is a saprophyte in thetop layer of soil beneath vines and grows in berries injuredby biotic agents, pests and diseases, and by abiotic factors.The grape berry moth (Lobesia botrana) is a major cause ofcolonization by black Aspergillus species (section Nigri),due to its active role in transporting spores into injuredberries (Cubaiu 2008). The temperature, the moisture, theaeration, the period of infection and the interaction betweendifferent fungi play a significant role in mycotoxin diffusion(Esti et al. 2012).
The level of OTA relies upon the types of wines and vineproducts, regions and vintages (Solfrizzo et al. 2010). In fact,red wines have generally higher levels than white wines, dueto the increased time of contact between berry skins and grapejuice during the mashing stage (Cubaiu 2008). In addition, thecontent of the toxin is higher in sweet and special wines, ifcompared to dry ones due to the different oenological prac-tices (Covarelli et al. 2012).
A survey performed by the European Commission on1,470 samples revealed that the average value of toxin is0.36 μg l−1, although the wines from Southern Europe suffer
a higher contamination (15.6 μg l−1). High levels of OTAwerealso reported on dried vine fruits (e.g. sultanas and raisins), theoverall mean level being 3.10 μg kg−1 as a result of a surveyon 593 samples (European Commission 2002).
Based on the available scientific toxicological and expo-sure data, Regulation (EC) No. 123/2005 (EuropeanCommission 2005) set the limit of OTA to 2 μg l−1 for wine,grape juice, grape nectar and grape must intended for directhuman consumption; however, many trade agreements requirelower limits (e.g. 0.5 μg l−1) (Solfrizzo et al. 2010). On theother hand, the maximum level of OTA in dried vine fruitswas set to 10 μg kg−1 (European Commission 2005).
Measures for preventing OTA appearanceand possibilities of its removal
Both preventive and corrective approaches were proposed toreduce the incidence of OTA contamination; the preventionstrategies include the use of biocontrol agents and fungicidestowards A. carbonarius and insecticides against L. botrana.However, these approaches cannot completely prevent theproblem, and severe contamination of wine can occur espe-cially for some susceptible grape varieties in certain high-riskregions or vintages with climatic conditions promoting theinfection by A. carbonarius (Solfrizzo et al. 2010).
Physical decontamination of OTA involves first the remov-al of mouldy grapes or bunches before entering in thewinemaking process (Leong et al. 2006). According toQuintela et al. (2013), this method may be able to reduceOTA incidence up to 98 %, but it might not be economicallyfeasible for the wine industry.
Wine filtration through a 0.45-μmmembrane reduced OTAlevel by 80 %; on the other hand, a filtration through a 10-μmmembrane does not reduce significantly the level of the toxin(Quintela et al. 2013).
Another approach to remove OTA from contaminatedwine involves the use of inorganic adsorbent such as alumi-nosilicates, zeolites, bentonites, clays and activated carbon. Alimit for their use is that they could decrease the nutritivevalue and organoleptic properties and increase the cost offood production (Piotrowska et al. 2013). In addition, somefining agents commonly used in winemaking process couldcause adverse reactions in susceptible wine consumers(Quintela et al. 2013).
Oak wood fragments can be used to reduce the levels ofOTA in wine. The effectiveness of this treatment relies uponthe wood format (chips or powder), quantity, time of contactand wine composition (Quintela et al. 2013). The rules for theuse of this kind of approach were reported in CommissionRegulation (EC) No. 1507/2006 (European Commission2006); however, the use of oak wood fragments is forbiddenin some European regions (Quintela et al. 2013). Finally,
Appl Microbiol Biotechnol

chitosan, chitin, chitin glucan or chitin glucan hydrolysatefrom fungal origin may also be useful ancillaries for reducingOTA levels (Bornet and Teissedre 2008), but in some cases,they were responsible for a decrease of total polyphenols andtotal anthocyans (Quintela et al. 2013).
Bioremediation of OTA: scientific backgroundfor the application of yeasts as adsorbing tools
The ability to degrade OTA was observed for some bacteria(Streptococcus, Bifidobacterium and Bacillus) and fungi(Aspergillus, Alternaria, Botrytis and Penicillum genera). Inaddition, microbial-derived enzymes with carboxypeptidaseA activity (CPA) can also degrade OTA (Amézqueta et al.2009). However, there are still some concerns about the tox-icity of the products of enzymatic degradation and the unde-sired effects of non-native microorganisms on wine quality(Cubaiu 2008).
Another approach for the bioremediation is the adsorptionof OTA on cell surface/cell surface components (Amézquetaet al. 2009). Adsorption effects were reported forLactobacillus rhamnosus (Turbic et al. 2002), Lactobacillusspp. (i.e. L. acidophilus CH-5, L. rhamnosus GG, L.plantarum BS, L. brevis and L. sanfranciscensis)(Piotrowska and Zakowska 2005) and some wine lactic acidbacteria (LAB) (i.e. Oenococcus oeni RM11, L. plantarumCECT 748T and L. brevisRM273) (Del Prete et al. 2007). Thepeptidoglycan and polysaccharides are probably involved inthe toxin binding (Del Prete et al. 2007).
On the other hand, conidia of black Aspergillus spp. (i.e. A.niger, A. carbonarius and A. japonicus) showed adsorbingproperties toward OTA in grape juices and musts (Bejaouiet al. 2005).
Yeasts could retain different wine compounds; thus, theiruse as adsorbing tools to eliminate harmful and/or negativecompounds (medium-chain fatty acids, ethyl phenols andsulphur products) could be of interest (Jiménez-Moreno andAncín-Azpilicueta 2009).
Some researchers reported the idea that wine yeastscould efficiently remove OTA (Amézqueta et al. 2009);this idea was supported by five key factors: (1) it was notpossible to find wine products from the degradation ofOTA (Bejaoui et al. 2004; Cecchini et al. 2006), (2) acidor heat treatment retained the ability to bind the toxin(Bejaoui et al. 2004; Núñez et al. 2008), (3) the recoveryof OTA in yeast lees (Caridi et al. 2006; Cecchini et al.2006), (4) yeasts without cell wall (protoplast) were notable to adsorb the toxin (Piotrowska 2012) and (5) thefate of radiolabeled OTA during the fermentation of grapejuice (Lataste et al. 2004).
Generally, a surface phenomenon is the result of complexinteractions (van der Walls, resonance and electrostatic forces,
and hydrogen bonding) amongst the adsorbent, adsorbateand solvent. The molecular size and the physicochemicalproperties of OTA as well as the physical structure of theadsorbent, including the total charge and its distribution,the size of the pores and the surface area, play a signifi-cant role in OTA-binding by adsorbent materials (Huwiget al. 2001).
Why is there an interaction between OTA and the yeast cellwall? The answer relies upon the chemical traits of both ofthem. Ochratoxin A is a complex organic compound,consisting of chlorine-containing dehydroisocoumarin linkedthrough the 7-carboxyl group to 1-β-phenylalanine; phenoland carboxyl are the main functional groups involved in somedifferent adsorption mechanisms. First, OTA is consideredwith zearalenone as the less polar mycotoxin and could thenbe bound on hydrophobic surfaces through the phenol groupand via interactions of two-π-electron orbital. Moreover, thepKa of the carboxyl group of phenylalanine moiety is 4.4;thus, the toxin is partially dissociated at the pH of wine andcarries a positive charge on the amine function (NH3
+) (Leonget al. 2006).
Yeast biomass may be regarded as a good source ofadsorbent material, due to the presence in the cell wall ofsome specific macromolecules, such as mannoproteins andβ-D-glucans (Ringot et al. 2007). For example, the cellwall of Saccharomyces consists of two layers: an innerlayer made of β-1,3-glucan and chitin, which representsabout 50–60 % of the cell wall dry weight, and an outerlayer, which consists of β-1,6-glucan and heavily glyco-sylated mannoproteins. The mannoproteins are glycopro-teins having carbohydrate fractions made of around 98 %mannose and 2 % glucose; they are covalently linked to theinner cell wall layer, either directly to the β-1,3-glucanmatrix or indirectly via a β-1,6-glucan branch (Gonzalez-Ramos and Gonzalez 2006). This structure is highly dy-namic and strain-dependent, since about 1,200 genes drivethe synthesis of these cell wall components. Culture con-ditions, including pH, temperature, oxygenation rate, kindof medium and concentration or nature of the carbonsource, strongly modulate the quantity and structural prop-erties of β-D-glucans, mannans and chitin in cell walls;moreover, the cell cycle stage also interacts with the cellwall composition. For example, budding induces strongchanges in the distribution of the structural componentsof the cell wall such as chitin (Jouany et al. 2005).
Despite the strong evidence of OTA adsorption to cellwalls (Bejaoui et al. 2004; Garcia-Moruno et al. 2005;Leong et al. 2006; Ringot et al. 2007; Núñez et al. 2008;Bizaj et al. 2009; Piotrowska 2012), Angioni et al. (2007)found that some yeasts could reduce OTA levels in wine,although they did not adsorb OTA; therefore, they suggestedthe existence of a possible pathway for the degradation of thetoxin, different from the mechanism involving L-β-
Appl Microbiol Biotechnol

phenylalanine and ochratoxin α; however, they could not findany product of hydrolysis.
Application of yeasts or yeast-derived productsas adsorbing tools to remove OTA from syntheticand natural grape juices, wines or model wine systems
Some studies focused on the reduction of OTA in syn-thetic and natural grape juices, wines or model winesystems by yeasts or inactive dry yeast (IDY) preparations(i.e. inactive yeast, yeast autolysates, yeast extracts andyeast hulls or walls) (Pozo-Bayón et al. 2009); Table 1reports a short overview of the available references. Theexperiments were carried out under laboratory conditions,and viability was not a prerequisite to achieve the removalof the toxin. Indeed, yeasts were intended simply asadsorbing tools.
The first comprehensive report on the use of yeasts andyeast-derived products intended as winemaking tools for theremoval of OTAwas conducted by Bejaoui et al. (2004). Theyreported that heat- and acid-treated cells were able to bindsignificantly higher levels of OTA than the viable ones in asynthetic grape juice medium. Viable yeasts bound up to 35%of OTA, depending on yeast concentration, whilst heat- andacid-treated cells decreased the toxin by 90.80 and 73 %,respectively. In addition, a comparative experiment betweenheat-treated cells and commercial yeast walls, at two differentconcentrations (0.2 and 6.7 g l−1) in a contaminated grapejuice, showed the highest efficiency by heat-treated cells ableto completely remove the toxin in 5 min. This trend could bedue to the fact that heating might cause changes in the surfaceproperties of cells, like the denaturation of proteins or theformation of Maillard reaction products. On the other hand,acidic conditions could affect polysaccharides by releas-ing monomers, which are further fragmented into alde-hydes after the breaking of glycosidic bonds. Thesereleased products could harbour higher adsorption sitesthan viable cells with an enhancement of OTA removal(Piotrowska et al. 2013).
Several winemaking practices involve a prolonged contactbetween yeasts or IDY preparations and wine (i.e. ageing onlees and the production of sparkling wines), thus suggestingthat yeast cells could play a significant role in OTA removalalso at the end of the fermentation process (Núñez et al. 2006).During this period, some genes encoding proteins involved incell wall biosynthesis are strongly up-regulated (fks1, gsc2,ssd1 and mpt5) (Pradelles et al. 2008); in addition, variableamounts of mannoproteins and glucans can be released intowines due to yeast autolysis interacting with wine phenoliccompounds, decreasing their astringency and/or acting asprotective colloids, and enhancing the color stability of redwines (Del Barrio-Galán et al. 2012). Mannoproteins can also
interact with OTA, as recently demonstrated by Núñez et al.(2008).
The most recent winemaking technologies are focusedon the use of vinification on lees (used successfully withwhite wines) for red wines (Garcia-Moruno et al. 2005);Garcia-Moruno et al. (2005) reported that OTA removalwas different using white and red lees wine. After 7 days ofcontact (20 g l−1 of lees), the white lees wine reduced OTAby 70.90 % whilst the red lees wine decreased the toxin by51.50 %. After 80 days. OTA reduction was ca. 73.90 %with the white lees and 63.10 % with the red ones. Thisdifference was attributed to competition on the bindingsites between OTA and polyphenols, and this hypothesiswas supported by Meca et al. (2010), who reported thatphenols, proteins, organic acid and colloidal particlesshowed the same ability of OTA to react with differentcompounds of the cell wall.
Another trend is the use of commercial yeast derivativepreparations, as an alternative to lees; these products couldreduce the time required to obtain wines with physico-chemical and sensory characteristics similar to those agedon lees (Del Barrio-Galán et al. 2012). Piotrowska et al.(2013) added a thermally inactivated biomass ofSaccharomyces cerevisiae yeast to wines from white grapeand blackcurrant juices and decreased the content of OTAby ca. 60 %. The heat treatment (85 °C for 10 min) wasalso proposed by Núñez et al. (2008) to increase the re-moval yield of whole yeast cells and yeast cell walls in amodel system up to 95 %. According to Piotrowska et al.(2013), the use of inactivated yeasts as OTA-adsorbingtools might be highly advantageous since such biomassdoes not change organoleptic features of end products; onthe other hand, Núñez et al. (2008) reported that heat- andacid-treated cells could negatively affect the quality ofwine. Nevertheless, the existence of potential unexpectedeffects related to odorant compounds initially present in theIDY preparations (Pozo-Bayón et al. 2009) should be care-fully evaluated.
Since the natural autolysis is a long-lasting process, it isusual to use model systems to obtain results in shorterperiods of time (Martínez-Rodríguez et al. 2001); Petruzziet al. (2014a) tested two commercial (BM45 and RC212)and three wild S. cerevisiae strains (W13, W47 and Y28)and a commercial cell wall preparation to remove OTA. AspH, temperature and ethanol are the major factors affectingautolysis in a model system, different pHs (3.0 and 3.5)temperatures (25 and 30 °C) and ethanol concentrations (5,10 and 15 %) were used. They reported that yeasts removedthe toxin by 3.69–81.87 %; S. cerevisiae Y28 showed thehigher removing percentages (24.60–81.87 %), whereas thestrain RC212 removed up to 25 % of the toxin. On the otherhand, yeast cell walls reduced OTA by 2.47–50.00 %. Thedecrease of the toxin was higher at 30 °C and pH 3.0 and
Appl Microbiol Biotechnol

with 15 % of ethanol. The viable count of yeasts was belowthe detection limit after 3 days in all the samples, thussuggesting that the bioremediation was not strictly relatedto cell viability.
Two wild strains (S. cerevisiae W47 and Y28) werepreviously studied for their ability to remove OTA in alaboratory medium (Petruzzi et al. 2013); the strainsdecreased the toxin by 36–42 % in a medium adjusted
Table 1 Review of wine yeasts and yeast-derived products used as adsorbing tools to remove ochratoxin A (OTA) from synthetic and natural grapejuices, wines or model wine systems
Yeast or yeast-derived product
State of cells Type of assay Experimental conditions Adsorbentamount(g l−1)
OTA level(μg l−1)
OTA removal(%)
References
Yeast cell walls Not treated Red wine Incubation at 25 °C 0.5–1.0 1.9 12–30 Silva et al. (2003)
Saccharomycescerevisiae
Viable Synthetic grape juice Incubation for 2 h, 30 °C 6.7–27.4 2 17.00–35.00 Bejaoui et al. (2004)
Heat-treated Synthetic grape juice Incubation for 2 h, 30 °C 6.7–27.4 2 75.00–90.80
Acid-treated Synthetic grape juice Incubation for 2 h, 30 °C 6.7 2 73
Heat-treated Red juice Incubation for 2 h, 30 °C 0.2–6.7 10 100
Yeast wallsadditive
Not treated Red juice Incubation for 2 h, 30 °C 0.2–6.7 10 100
Saccharomycescerevisiae
IY Red wine Stirring for the first 90 minand then stored for7 days at ca. 20 °Cwithout further shaking
1–4 3.04–3.30 25–60.30 Garcia-Moruno et al.(2005)
Saccharomycescerevisiae
Lees from whitemust fermentation
Red wine Stirring for the first 90 minand then stored for 7 or80 days at ca. 20 °Cwithout further shaking
20 1.65–7.09 70.90–73.90
Lees from red mustfermentation
Red wine Stirring for the first 90 minand then stored for 7 or80 days at ca. 20 °Cwithout further shaking
20 2.49–4.12 51.50–63.10
Yeast hulls Not treated Red wine Incubation for 2 days, atca. 22 °C
2.00–5.00 5 28.00–43.00 Leong et al. (2006)
NI IY Red wine Stirring for 90 min andthen stored for 30 or90 min at ca. 20 °Cwithout further shaking
1–4 2.55–2.78 20.00–46.50 Savino et al. (2006)
IY Red wine Stirring for the first 90 minand then stored for 7 or80 days at ca. 20 °Cwithout further shaking
4 2.84 62.00–81.00
Saccharomycescerevisiae
Rehydrated IY Model wine Incubation for 214 h 1 10 0.8 Núñez et al. (2008)
Rehydrated IYandheat-treated
Model wine Incubation for 214 h 1 10 95.4
Yeast cell walls Not treated Model wine Incubation for 214 h 1 10 18.8
Heat-treated Model wine Incubation for 214 h 1 10 95.0
Candida spp.,Kloeckera spp.,Rhodotorulaglutinis,Cryptococcuslaurentii
Biomass fromculture broth
White wine Incubation for 4 h, 25 °C NI 10 4.75–21.40 Var et al. (2009)
Biomass thermallyinactivated
White wine Incubation for 4 h, 25 °C NI 10 8.08–30.45
Saccharomycescerevisiae
Biomass fromculture broth
Model wine Incubation for 9 days at 25or 30 °C, with ethanolcontent of 5, 10 or15 %, at pH 3.0 or 3.5
10 2 3.69–81.87 Petruzzi et al. (2014a)
Yeast cell walls Not treated Model wine Incubation for 9 days at 25or 30 °C, with ethanolcontent of 5, 10 or15 %, at pH 3.0 or 3.5
10 2 2.47–50.00
Saccharomycescerevisiae
Biomass thermallyinactivated
White wine Incubation for 24 h, 30 °C 0.005 1,000 64.40 Piotrowska et al. (2013)Red wine Incubation for 24 h, 30 °C 0.005 1,000 62.40
IY inactive yeast, NI not indicated
Appl Microbiol Biotechnol

to pH 3.5, containing 10 % of ethanol and incubated at37 °C.
Based on available data, we could suppose that somevariables play a major role in OTA removal efficiency of yeastderivatives, i.e.:
& The enzymatic and chemical protocol for their production,as the enzymatic and thermal procedure currentlyemployed for their production could have a strong influ-ence on their physical and compositional characteristics(Núñez et al. 2006);
& The amount (Silva et al. 2003; Bejaoui et al. 2004), al-though most of the researchers performed the experimentsusing amounts of yeast cell wall preparations of up to0.40 g/l, i.e. the legal limit set by Commission Regulation(EC) No. 1410/2003 (European Commission 2003);
& The physiological state of yeast cells before the produc-tion of autolysates, as the content of mannoproteins andglucans is usually higher in the lysates from viable cellsthan from rehydrated cells (Guilloux-Benatier andChassagne 2003).
Application of yeasts to remove OTA throughout alcoholicfermentation and factors affecting its removal
Some basic requirements for an ideal method to remove OTAare the low cost, the simplicity of use and the absence ofeffects on the quality of wine (Amézqueta et al. 2009).Unfortunately, inactivated cells or IDY products could nega-tively affect the quality of wine (Núñez et al. 2008; Pozo-Bayón et al. 2009), whilst the use of viable cells does not showthis drawback; moreover, active yeasts have a long history ofsafe use as starter cultures (Abrunhosa et al. 2010).
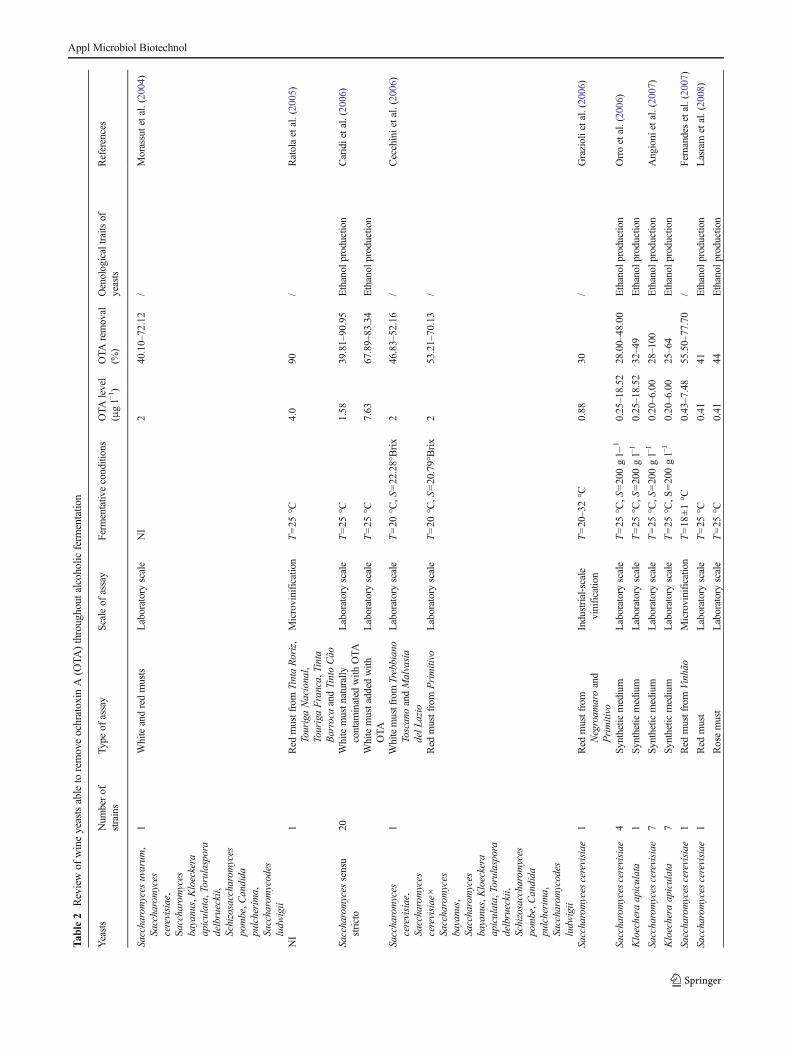
Over the last years, some authors identified yeasts fromSaccharomyces and non-Saccharomyces genera able to de-crease OTA content by 0.60–100 % throughout fermentation(Table 2). Some key factors able to influence this phenomenonare the conditions for the assay (laboratory or industrial scale,in vitro or in vivo conditions), kind of the strain, cell dimen-sion, flocculence, cell sedimentation kinetics and toxin con-centration (Caridi et al. 2006; Caridi 2007; Cecchini et al.2006; Angioni et al. 2007; Ponsone et al. 2009; Petruzzi et al.2014b, d).
Kind of fermentation, scale and OTA content
The level of OTA in wine is significantly lower than in grapes,due to toxin removal throughout vinification, and especiallyduring alcoholic fermentation, as a result of yeast activity and/or the interaction between OTA and grape constituents (Paster2008). In red vinification, most of OTA is removed in the
solid–liquid separation stages, when the wine or the juice isseparated from the skins (Bizaj et al. 2009). In white vinifica-tion, grapes are pressed before fermentation and OTA mightbind more effectively to the grape proteins and solids, whichare removed during clarification. In addition, other steps (e.g.additional clarification steps) allow higher OTA-binding rates(Paster 2008). We could suppose also an interaction betweenOTA and the natural microbiota of grape must, probablycontributing to the modification of the removal yield of inoc-ulated starter strains.
Differences in OTA removal may be related to the scale ofassay, as the management of the fermentation could be diffi-cult in macro-scale experiments; on the other hand, changes inOTA content during small-scale experiments might be simplymanaged.
Under laboratory conditions (from 10 ml to 2 l), OTAwasremoved by 32.6–90.95 %, depending on the strain and themedium (Caridi et al. 2006; Meca et al. 2010; Cecchini et al.2006; Esti et al. 2012).
OTA removal was also found in higher volumes. Fernandeset al. (2007) reported that the toxin was reduced by 55.5–77.7 % in microvinification trials consisting of 20 kg ofgrapes; however, the mass balance performed in eachvinification trial revealed that this decrease was duepredominantly by the adsorption onto suspended solids inmusts and wines. Ponsone et al. (2009) showed that a signif-icant reduction of OTA (i.e. 77–100 %) after fermentation in100-l tanks was caused by an extensive adsorption onto thesolid parts of the grapes and yeast lees. Only one study wascarried out at industrial scale (800-hl tanks) by Grazioli et al.(2006); they reported that maceration increased OTA contentby 45 %, whilst alcoholic fermentation caused its reduction(30 %).
Concerning the effect of the initial amount of OTA,Piotrowska et al. (2013) studied OTA removal by two strainsof S. cerevisiae: Malaga LOCK 0173, able to remove OTA(1,000μg l−1) by 82.80, 10.70 and 35.40%, respectively, fromwhite juice, blackcurrant juice and synthetic medium, and thestrain Syrena LOCK 0201 respectively reducing toxin con-centration by 85.10, 65.20 and 21.00 %. Csutorás et al. (2013)reported that a commercial S. cerevisiae type “Fermol PremierCru” removed OTA (initial content 4,000 μg l−1) by 90, 85and 73 % from red, rose and white must, respectively.
Chemico-physical factors acting on OTA removal
Although many papers focused on the role of some physico-chemical factors on the course of wine fermentation (Belyet al. 2003, 2008; Hernández-Orte et al. 2006; Torija et al.2003), few data are available on their influence on the removalof OTA by yeasts. This issue is of great concern consideringthat some oenological parameters can modify the surfaceproperties of cell wall (Vasserot et al. 1997); thus, they could
Appl Microbiol Biotechnol
Tab
le2
Reviewof
wineyeastsableto
removeochratoxin
A(O
TA)throughout
alcoholic
ferm
entatio
n
Yeasts
Num
berof
strains
Type
ofassay
Scaleof
assay
Ferm
entativeconditions
OTA
level
(μgl−1 )
OTA
removal
(%)
Oenologicaltraitsof
yeasts
References
Saccharomyces
uvarum
,Saccharomyces
cerevisiae,
Saccharomyces
bayanus,Kloeckera
apiculata,Torulaspora
delbrueckii,
Schizosaccharomyces
pombe,C
andida
pulcherima,
Saccharomycodes
ludw
igii
1Whiteandredmusts
Laboratoryscale
NI
240.10–72.12
/Morassutetal.(2004)
NI
1Red
mustfrom
TintaRoriz,
TourigaNacional,
TourigaFranca,Tinta
Barroca
andTintoCão
Microvinification
T=25
°C4.0
90/
Ratolaetal.(2005)
Saccharomyces
sensu
stricto
20Whitemustn
aturally
contam
inated
with
OTA
Laboratoryscale
T=25
°C1.58
39.81–90.95
Ethanol
productio
nCaridietal.(2006)
Whitemustadded
with
OTA
Laboratoryscale
T=25
°C7.63
67.89–83.34
Ethanol
productio
n
Saccharomyces
cerevisiae,
Saccharomyces
cerevisiae×
Saccharomyces
bayanus,
Saccharomyces
bayanus,Kloeckera
apiculata,Torulaspora
delbrueckii,
Schizosaccharomyces
pombe,C
andida
pulcherima,
Saccharomycodes
ludw
igii
1WhitemustfromTrebbiano
ToscanoandMalvasia
delL
azio
Laboratoryscale
T=20
°C,S
=22.28°Brix
246.83–52.16
/Cecchinietal.(2006)
Red
mustfrom
Primitivo
Laboratoryscale
T=20
°C,S
=20.79°Brix
253.21–70.13
/
Saccharomyces
cerevisiae
1Red
mustfrom
Negroam
aroand
Primitivo
Industrial-scale
vinificatio
nT=20–32°C
0.88
30/
Graziolietal.(2006)
Saccharomyces
cerevisiae
4Sy
nthetic
medium
Laboratoryscale
T=25
°C,S
=200gl−
10.25–18.52
28.00–48.00
Ethanol
productio
nOrroetal.(2006)
Kloechera
apiculata
1Sy
nthetic
medium
Laboratoryscale
T=25
°C,S
=200gl−1
0.25–18.52
32–49
Ethanol
productio
n
Saccharomyces
cerevisiae
7Sy
nthetic
medium
Laboratoryscale
T=25
°C,S
=200gl−1
0.20–6.00
28–100
Ethanol
productio
nAngioni
etal.(2007)
Kloechera
apiculata
7Sy
nthetic
medium
Laboratoryscale
T=25
°C,S
=200gl−1
0.20–6.00
25–64
Ethanol
productio
n
Saccharomyces
cerevisiae
1Red
mustfrom
Vinhão
Microvinification
T=18
±1°C
0.43–7.48
55.50–77.70
/Fernandesetal.(2007)
Saccharomyces
cerevisiae
1Red
must
Laboratoryscale
T=25
°C0.41
41Ethanol
productio
nLasram
etal.(2008)
Rosemust
Laboratoryscale
T=25
°C0.41
44Ethanol
productio
n
Appl Microbiol Biotechnol

Tab
le2
(contin
ued)
Yeasts
Num
berof
strains
Type
ofassay
Scaleof
assay
Ferm
entativeconditions
OTA
level
(μgl−1 )
OTA
removal
(%)
Oenologicaltraitsof
yeasts
References
Saccharomyces
cerevisiae
1Sy
nthetic
medium
Laboratoryscale
T=20
°C,S
=180gl−1
4.95
2.60
Sugarconsum
ptionand
volatileacidity
productio
n
Bizajetal.(2009)
Saccharomyces
cerevisiae
1Sy
nthetic
medium
Laboratoryscale
T=20
°C,S
=180gl−1
3.48
2.00
Sugarconsum
ptionand
volatileacidity
productio
nSaccharomyces
cerevisiae
3Whitemustfrom
Moscato
ofSaracena
Laboratoryscale
T=16–18°C
,S=38°B
rix
10>40
/Blaiotta
etal.(2009)
Saccharomyces
cerevisiae
2Red
mustfrom
Tempranilo
Microvinification
T=24–28°C
,S=
24.70°Brix
683–86
Sugarconsum
ption,
ethanoland
acids
productio
n
Ponsoneetal.(2009)
Red
mustfrom
Bonarda
Microvinification
T=24–28°C
,S=
21.50°Brix
0.3
77–100
Sugarconsum
ption,
ethanoland
acids
productio
nSaccharomyces
cerevisiae
16Whitemustfrom
Moscato
Laboratoryscale
T=25
°C,S
=22°B
rix
1032.60–50.40
/Mecaetal.(2010)
Saccharomyces
cerevisiae
48Whitemustfrom
Zibibbo
Laboratoryscale
T=25
°C4.10
7.80-81.95
/Caridietal.(2012)
Saccharomyces
cerevisiae
1Whitemustfrom
Sauvignonblanc
Laboratoryscale
T=20
°C,S
=20.5°B
rix
1562
/Estietal.(2012)
Saccharomyces
cerevisiae
1 1Red
mustfromCesanesedi
Affile
Laboratoryscale
T=20
°C,S
=22°B
rix
1579.00
/
Whitemustfrom
Sauvignonblanc
Laboratoryscale
T=20
°C,S
=20.5°B
rix
1567.00
/
Saccharomyces
cerevisiae
Rosemust
Microvinification
T=12
°C10–4,000
83.00–86.00
/Csutorásetal.(2013)
Whitemust
Microvinification
T=12
°C10–4,000
73.00–76.00
/
Red
must
Microvinification
T=12
°C10–4,000
88.00–90.00
/
Saccharomyces
cerevisiae
1Sy
nthetic
medium
Laboratoryscale
T=30
°C,S
=20
gl−1
1,000
35.40
/Piotrowskaetal.(2013)
Whitejuice
Laboratoryscale
T=30
°C,S
=12°B
rix
1,000
82.80
/
Blackcurrantjuice
Laboratoryscale
T=30
°C,S
=12°B
rix
1,000
10.70
/
Saccharomyces
cerevisiae
1Sy
nthetic
medium
Laboratoryscale
T=30
°C¸S=20
gl−1
1,000
21.00
/
Whitejuice
Laboratoryscale
T=30
°C,S
=12°B
rix
1,000
85.10
Ethanol,glyceroland
acidsproductio
nBlackcurrantjuice
Laboratoryscale
T=30
°C,S
=12°B
rix
1,000
65.20
Ethanol,glyceroland
acidsproductio
nSaccharomyces
cerevisiae
2Sy
nthetic
medium
Laboratoryscale
T=25
or30
°C,S
=200or
250gl−1
25.41–49.58
Sugarconsum
ption,
ethanol,glycerol
and
volatileacidity
productio
n
Bevilacqua
etal.(2014)
Saccharomyces
cerevisiae
1Sy
nthetic
medium
Laboratoryscale
T=25
or30
°C¸S=200or
250gl−1,N
=250or
310mgl−1
26.00–25.66
Sugarconsum
ption,
ethanoland
glycerol
productio
n;phenotypicaltraits
Petruzzietal.(2014b)
Saccharomyces
cerevisiae
2Sy
nthetic
medium
Laboratoryscale
T=25
or30
°C,S
=200or
250gl−1,N
=250or
310mgl−1
22.82–27.53
Sugarconsum
ption,
ethanoland
glycerol
Petruzzietal.(2014c)
Appl Microbiol Biotechnol

affect the binding capacity of yeasts. Petruzzi et al. (2014b)studied the kinetics of OTA removal by a potential starterstrain (i.e. S. cerevisiae W13) as affected by the most impor-tant environmental parameter (e.g. temperature) (Pizarro et al.2008), as well as by two major nutrients acting on the fate ofmust fermentation (sugar and nitrogen) (Heinisch and Rodicio2009). At this scope, laboratory-scale fermentations werecarried out at two temperatures (25 and 30 °C) and sugarlevels (200 and 250 g l−1), with or without supplementationof medium with diammonium phosphate (DAP) (300 mg l−1).The yeast was able to reduce OTA up to 57.21 %, with apositive effect of temperature and sugar. These resultsconfirmed the data by Patharajan et al. (2011), who studiedsix different temperatures (10, 15, 20, 25, 30 and 35 °C) tooptimize OTA removal in a liquid medium by three yeasts(Metschnikowia pulcherrima MACH1, Pichia guilliermondiiM8, Rhodococcus erythropolis AR14) and found the highestremoval at 30 °C. Petruzzi et al. (2014b) suggested that themajor ability to remove OTA at 30 °C could be associated tothe release of cell wall polysaccharides. The effects of sugarconcentration could be related to the level of mannoproteins,as these compounds are strictly dependent on sugar (Mecaet al. 2010). Finally, Petruzzi et al. (2014b) found that theremoval was highest after 3 days of fermentation; then, thetoxin was partially released in the medium and the perfor-mances of the strain decreased (i.e. 6.0–25.66 % reduction). Arelease of toxins after their removal was also recovered forother compounds; Peltonen et al. (2001) reported the releaseof aflatoxin B1 (AFB1) by lactobacilli (i.e. L. amylovorus andL. rhamnosus). They suggested that this phenomenon couldbe associated to the weak bond in lactobacilli/AFB1 com-plexes. Since hydrogen bonding and ionic or hydrophobicinteraction seem to be related to OTA adsorption mechanismby S. cerevisiae (Cecchini et al. 2006), Petruzzi et al. (2014b)considered this statement as a possible explanation for OTArelease by yeasts.
In another study, Petruzzi et al. (2014c) tested two wildS. cerevisiae strains (W28 and W46) under the same experi-mental conditions reported for the strain W13. As a result, thestrains W28 and W46 were able to decrease the toxin by ca.70 %, with the highest removing effect observed after 3 daysat 30 °C in the presence of 250 g l−1 of sugars and with DAP;after 10 days, the toxin was partially released into the mediumand the performances of the strains decreased (i.e. 2.82–27.53 % reduction). As reported above, sugar and temperatureplayed a major role, with a significant effect of DAP supple-mentation, too. As regards nitrogen, we could suppose that thestrains exhibited a significant ability to remove OTA onlywhen the conditions were closer to the optimal ones (i.e.greater YAN content in the must), probably due to a higherrelease of cell wall polysaccharides (Giovani et al. 2010). Thedifferent trends of the three strains (i.e. W13, W28 and W46)toward DAP addition could be due to the strong strainT
able2
(contin
ued)
Yeasts
Num
berof
strains
Type
ofassay
Scaleof
assay
Ferm
entativeconditions
OTA
level
(μgl−1 )
OTA
removal
(%)
Oenologicaltraitsof
yeasts
References
productio
n;phenotypicaltraits
Saccharomyces
cerevisiae
4Sy
nthetic
medium
Laboratoryscale
T=25
or30
°C,S
=200or
250gl−1
20.60–42.80
Sugarconsum
ption,
ethanol,glycerol
and
volatileacidity
productio
n;phenotypicaltraits
Petruzzietal.(2014d)
NInotindicated,/
none
oenologicaltraitassessed¸Ttemperature,S
sugarcontent,Nnitrogen
contentasyeastassim
ilablenitrogen
(YAN)
Phenotypicaltraitsincludethefollo
wing:resistance
tosingleandcombinedstressconditions,enzymaticactiv
ities,hydrogensulphide
productio
n,interactionwith
phenoliccompounds
andbiogenicam
ines
form
ation;
1°B
rixmeans
1weightp
ercentageof
reducing
sugars
Appl Microbiol Biotechnol

dependence on the response to changes in environmentalconditions (Giovani et al. 2010).
OTA removal as a new trait to select functional yeast startercultures
Recently, a new approach for the study of OTA-removingyeasts was proposed by Petruzzi et al. (2014b, c, d), i.e. yeaststrains intended as functional starter cultures, as they showboth the benefits of traditional starters and a health- orproduct-focused function. In this sense, a drawback in theliterature is that few data are available on the oenological traitsof OTA-removing yeasts (see Table 2), whilst this aspect is ofgreat concern for the possible selection of functional strainsacting at the same time as starters and as biological tools toremove the toxin.
Thus, the removal of OTA could be an interesting andattractive trait to complement the classical oenological char-acterization based on technological and qualitative key traits,such as the tolerance and high ethanol production, exhaus-tion of sugars, growth at high sugar concentration, goodglycerol production, growth at high temperatures, low hy-drogen sulphide and volatile acidity production, resistance tosulphur dioxide and good enzymatic profile (Nikolaou et al.2006). This idea is supported by the fact that the removal oftoxin by yeasts is genetically controlled and is a polygenicinheritable trait of wine yeasts (Caridi et al. 2012). However,the toxin could be released back into the medium after itsadsorption (Bevilacqua et al. 2014; Petruzzi et al. 2013,2014b, c), probably due to the nature of the adsorptionmechanism involved (hydrogen bonding, ionic or hydropho-bic interaction) (Cecchini et al. 2006). Strain dependencecould play a significant role also on the stability of thecomplex cell/toxin, as well as on the yeast cell wall compo-sition in response to a variety of fermentative/growth condi-tions (Bevilacqua et al. 2014).
Therefore, Petruzzi et al. (2014d) used a polyphasic ap-proach, consisting of the genotypic identification and theevaluation of the phenotypic traits and fermentative perfor-mances in a model system (temperature, 25 and 30 °C; sugarlevel, 200 and 250 g l−1), to select wine starters of S. cerevisiaefrom 30 autochtonous isolates from Uva di Troia cv., a redwine grape variety of the Apulian region (Southern Italy). Theability to remove OTAwas used by the authors as a desirablefunctional trait to improve the safety of wine. As a result, 11biotypes were identified and a strain, representative of eachbiotype, was further studied. Four strains (Y20, W21, W40and W41) were able to reduce OTA by 0.60–42.80 %; inaddition, the strains W21 and W40 were promising in termsof ethanol, glycerol and volatile acidity production, as well asfor their enzymatic and stress resistance characteristics. In thissense, interesting traits were also possessed by S. cerevisiaestrains W13 (Petruzzi et al. 2014b), W28 and W46 (Petruzzi
et al. 2014c), e.g. high tolerance to single and combined stressconditions; β-D-glucosidase, pectolytic and xylanase activi-ties; low-to-medium level of hydrogen sulphide production;low-to-medium parietal interaction with phenolic compounds;and non-decarboxylase activity, potentially related to the pres-ence of biogenic amines in the wine.
Thus, the selected strains could be considered as promisingfunctional starter cultures, acting at the same time as biolog-ical tools to remove OTA throughout the fermentation.
Conclusion and future trends
To our knowledge, this represents the first review regardingexclusively the biological removal of OTAmediated by yeastsor yeast-derived products in winemaking process. Based onthe available reports, toxin removal was dependent on theyeast strain or on the use of yeast-derived products. In addi-tion, several physico-chemical factors have been found toaffect OTA removal ability throughout alcoholic fermentation(i.e. temperature and sugar and nitrogen concentration) orduring the ageing of wine with yeasts or yeast cell walls (i.e.ethanol, temperature and pH). However, the toxin could bereleased back into the medium after its adsorption and this traitis strain-dependent.
Thus, ochratoxin A removal by yeasts is a promising andfriendly solution to attain the decontamination without usingharmful chemicals and without losses in nutrient value orpalatability of decontaminated food; however, the availableresults could be labeled as the first step to design practicalcommercial technologies at a larger scale. In fact, furtherintensive screening of yeasts may lead to the detection ofefficient and applicable cultures acting at the same time asstarters and as biological tools to remove OTA, with lowOTA-release ability. On the other hand, further investigationsare required, since the use of yeast-derived products or heat-and acid-treated cells could negatively affect the quality ofwine.
As a future perspective, the use of molecular biologytechniques could help design strategies for the genetic im-provement of OTA-removing yeasts, whilst the application ofpredictive models to estimate the effects of toxin concentra-tion, environmental and nutritional variables on OTA removalcould be a very useful tool for wine industry.
An open question on the link yeast/bioremediation is asfollows: Are yeasts considered as adsorbing tools or po-tential starter strains? Based on the available reports, wecan conclude that OTA levels can be reduced up to accept-able limits (2 μg l−1) in both cases; however, well-characterized starter cultures (mainly S. cerevisiae) withhigh adsorbing abilities can represent a significant andpromising frontiers goal.
Appl Microbiol Biotechnol

Acknowledgments The authors wish to thank Miss. SilvanaRendinella for her kind co-operation.
References
Abrunhosa L, Paterson RRM, Venâncio A (2010) Biodegradation of och-ratoxin A for food and feed decontamination. Toxins 2:1078–1099
Afsah-Hejri L, Jinap S, Hajeb P, Radu S, Shakibazadeh S (2013) Areview on mycotoxins in food and feed: Malaysia case study.Compr Rev Food Sci F 12:629–651
Amézqueta S, González-Peñas E, Murillo-Arbizu M, López de Cerain A(2009) Ochratoxin A decontamination: a review. Food Control 20:326–333
Angioni A, Caboni P, Garau A, Farris A, Orro D, Budroni M, Cabras P(2007) In vitro interaction between ochratoxin A and differentstrains of Saccharomyces cerevisiae and Kloeckera apiculata. JAgric Food Chem 55:2043–2048
Anly E, Bayram M (2009) Ochratoxin A in wines. Food Rev Int 25:214–232
Bejaoui H, Mathieu F, Taillandier P, Lebrihi A (2004) Ochratoxin Aremoval in synthetic and natural grape juice by selected oenologicalSaccharomyces cerevisiae strains. J Appl Microbiol 97:1038–1044
Bejaoui H, Mathieu F, Taillandier P, Lebrihi A (2005) Conidia of blackaspergilli as new biological adsorbents for ochratoxin A in grapejuices and musts. J Agric Food Chem 53:8224–8229
Bellí N, Marín S, Duaigües A, Ramos AJ, Sanchis V (2004) OchratoxinA in wines, musts and grapes juices from Spain. J Sci Food Agr 84:591–594
Bely M, Rinaldi A, Dubourdieu D (2003) Influence of assimilable nitro-gen on volatile acidity production by Saccharomyces cerevisiaeduring high sugar fermentation. J Biosci Bioeng 96:507–512
Bely M, Stoeckle P, Masneuf-Pomarède I, Dubourdieu D (2008) Impactof mixed Torulaspora delbrueckii-Saccharomyces cerevisiae cultureon high-sugar fermentation. Int J Food Microbiol 122:312–320
Bevilacqua A, Petruzzi L, Corbo MR, Baiano A, Garofalo C, Sinigaglia M(2014) Ochratoxin A released back into the medium bySaccharomyces cerevisiae as a function of the strain, washingmediumand fermentative conditions. J Sci Food Agr. doi:10.1002/jsfa.6683
Bizaj E, Mavri J, Čuš F, Raspor A (2009) Removal of ochratoxin A inSaccharomyces cerevisiae liquid cultures. S Afr J Enol Vitic 30:151–155
Blaiotta G, Meca G, Ritieni A (2009) OTA adsorption kinetic bySaccharomyces cerevisiae strains during fermentation of must ob-tained from dry grapes “Moscato of Saracena”, a southern Italypassito-wine (Calabria Region). In: Book of abstract of 2nd interna-tional symposium “Wine Microbiology and Safety: from the vine-yard to the bottle (Microsafetywine),” Martina Franca (TA), Italy,19–20 November 2009
Bornet A, Teissedre PL (2008) Chitosan, chitin-glucan and chitin effectson minerals (iron, lead, cadmium) and organic (ochratoxin A) con-taminants in wines. Eur Food Res Technol 226:681–689
Brera C, Debegnach F, Minardi V, Prantera E, Pannunzi E, Faleo S, deSantis B, Miraglia M (2008) Ochratoxin A contamination in Italianwine samples and evaluation of the exposure in the Italian popula-tion. J Agric Food Chem 56:10611–10618
Caridi A (2007) New perspectives in safety and quality enhancement ofwine through selection of yeasts based on the parietal adsorptionactivity. Int J Food Microbiol 120:167–172
Caridi A, Galvano F, Tafuri A, Ritieni A (2006) Ochratoxin A removalduring winemaking. Enzym Microb Technol 40:122–126
Caridi A, Sidari R, Pulvirulenti A, Meca G, Ritieni A (2012) OchratoxinA adsorption phenotype: an inheritable yeast trait. J Gen ApplMicrobiol 58:225–233
Cecchini F, Morassut M, Garcia-Moruno E, Di Stefano R (2006)Influence of yeast strain on ochratoxin A content during fermenta-tion of white and red must. Food Microbiol 23:411–417
Covarelli L, Beccari G, Marini A, Tosi L (2012) A review on theoccurrence and control of ochratoxigenic fungal species and ochra-toxin A in dehydrated grapes, non-fortified dessert wines and driedvine fruit in the Mediterranean area. Food Control 26:347–356
Csutorás CS, Rácz L, Rácz K, Fűtő P, Forgó P, Kiss A (2013)Monitoringof ochratoxin A during the fermentation of different wines byapplying high toxin concentrations. Microchem J 107:182–184
Cubaiu L (2008) Prevention and biocontrol of ochratoxin A in wine. PhDDissertation, Università degli Studi di Sassari, Sassari, Italy
Del Barrio-Galán R, Pérez-Magariño S, Ortega-Heras M (2012) Effect ofthe aging on lees and other alternative techniques on the lowmolecular weight phenols of Tempranillo red wine aged in oakbarrels. Anal Chim Acta 732:53–63
Del Prete V, Rodriguez H, Carrascosa AV, de las Rivas B, Garcia-MorunoE, Muñoz R (2007) In vitro removal of ochratoxin A by wine lacticacid bacteria. J Food Prot 70:2155–2160
Esti M, Benucci I, Liburdi K, Acciaro G (2012)Monitoring of ochratoxinA fate during alcoholic fermentation of wine-must. FoodControl 27:53–56
European Commission (2002) Reports on tasks for scientific cooperation.Report of experts participating in Task 3.2.7. Assessment of dietaryintake of ochratoxin A by the population of EU member states.Directorate-General Health and Consumer Protection, January 2002
European Commission (2003) Commission Regulation (EC) No.1410/2003 of 7 August 2003 amending Regulation (EC) No.1622/2000 laying down certain detailed rules for implementingCouncil Regulation (EC) No. 1493/1999 on the common organisa-tion of the market in wine and establishing a Community code ofoenological practices and processes. Off J Eur Union L201:9–11
European Commission (2006) Commission Regulation (EC) No.1507/2006 of 11 October 2006 amending regulations (EC) No.1622/2000, (EC) No. 884/2001 and (EC) No. 753/2002 concerningcertain detailed rules implementing regulation (EC) No. 1493/1999on the common organisation of the market in wine, as regards theuse of pieces of oak wood in winemaking and the designation andpresentation of wine so treated. Off J Eur Union L280:9–11
European Commission (2005) Commission Regulation (EC) No. 123/2005 of 26 January 2005 amending Regulation (EC) No. 466/2001as regards ochratoxin A. Off J Eur Union L25:3–5
Fernandes A, Ratola N, Cerdeira A, Alves A, Venâncio A (2007)Changes in ochratoxin A concentration during winemaking. Am JEnol Viticult 58:92–96
Garcia-Moruno E, Sanlorenzo C, Beccaccino B, Di Stefano R (2005)Treatment with yeast to reduce the concentration of ochratoxin A inred wine. Am J Enol Vitic 56:73–76
Giovani G, Canuti V, Rosi I (2010) Effect of yeast strain and fermentationconditions on the release of cell wall polysaccharides. Int J FoodMicrobiol 137:303–307
Gonzalez-Ramos D, Gonzalez R (2006) Genetic determinants of therelease of mannoproteins of enological interest by Saccharomycescerevisiae. J Agric Food Chem 54:9411–9416
Grazioli B, Fumi MD, Silva A (2006) The role of processing on ochra-toxin A content in Italian must and wine: a study on naturallycontaminated grapes. Int J Food Microbiol 111:S93–S96
Guilloux-Benatier M, Chassagne D (2003) Comparison of componentsreleased by fermented or active dried yeasts after aging on lees in amodel wine. J Agric Food Chem 51:746–751
Heinisch JJ, Rodicio R (2009) Physical and chemical stress factors inyeast. In: König H, Unden G, Fröhlich J (eds) Biology of microor-ganisms on grapes, inmust and in wine. Springer, Berlin, pp 275–291
Hernández-Orte P, Ibarz MJ, Cacho J, Ferreira V (2006) Impact ofammonium additions on volatile acidity, ethanol, and aromaticcompound production by different Saccharomyces cerevisiae strains
Appl Microbiol Biotechnol

during fermentation in controlled synthetic media. Aust J GrapeWine Res 12:150–160
Huwig A, Freimund S, Kappeli O, Dutler H (2001) Mycotoxin detoxi-cation of animal feed by different adsorbents. Toxicol Lett 122:179–188
Inoue T, Nagatomi Y, Uyama A,Mochizuki N (2013) Fate of mycotoxinsduring beer brewing and fermentation. Biosci Biotechnol Biochem77:1410–1415
Jard G, Liboz T, Mathieu F, Guyonvarc’h A, Lebrihi A (2011) Reviewof mycotoxin reduction in food and feed: from prevention in thefield to detoxification by adsorption or transformation. FoodAddit Contam Part A Chem Anal Control Expo Risk Assess28:1590–1609
Jiménez-Moreno N, Ancín-Azpilicueta C (2009) Sorption of volatilephenols by yeast cell walls. J Wine Res 1:11–18
Jouany JP, Yiannikouris A, Bertin G (2005) The chemical bonds betweenmycotoxins and cell wall components of Saccharomyces cerevisiaehave been identified. Archiva Zootechnica 8:26–50
Lasram S,Mani A, ZaiedC, Chebil S, Abid S, BachaH,Mliki A, GhorbelA (2008) Evolution of ochratoxin A content during red and rosevinification. J Sci Food Agric 88:1696–1703
Lataste C, Guérin L, Solanet D, Berger JL, Cottereau P, Molot B (2004)Évolution de la contamination en ochratoxine A du vignobleFrançais au vin. Prog Agric Vitic 121:57–64
Leong SL, Hocking AD, Varelis P, Giannikopoulos G, Scott ES (2006)Fate of ochratoxin A during vinification of Semillon and Shirazgrapes. J Agric Food Chem 54:6460–6464
Martínez-Rodríguez AJ, Polo MC, Carrascosa AV (2001) Structural andultrastructural changes in yeast cells during autolysis in a modelwine system and in sparkling wines. Int J Food Microbiol 71:45–51
MecaG, Blaiotta G, Ritieni A (2010) Reduction of ochratoxinA during thefermentation of Italian red wine Moscato. Food Control 21:579–583
Morassut M, Cecchini F, Garcia-Moruno E, Savino M, Cedroni A (2004)Ocratossina A: decontaminazione biologica dei mosti nel corso dellafermentazione alcolica. 29–30 novembre. Rapporti ISTISAN 05/421°Congresso nazionale “Le micotossine nella filiera agro-alimentare” (pp. 142–145). Roma: Istituto Superiore di Sanità
Nikolaou E, Soufleros EH, Bouloumpasi E, Tzanetakis N (2006)Selection of indigenous Saccharomyces cerevisiae strains accordingto their oenological characteristics and vinification results. FoodMicrobiol 23:205–211
Núñez YP, Carrascosa AV, Gonzalez R, Polo MC, Martínez-RodríguezAJ (2006) Isolation and characterization of a thermally extractedyeast cell wall fraction potentially useful for improving the foamingproperties of sparkling wines. J Agric Food Chem 54:7898–7903
Núñez YP, Pueyo E, Carrascosa AV, Martínez-Rodríguez AJ (2008)Effects of aging and heat treatment on whole yeast cells and yeastcell walls and on adsorption of ochratoxin A in a wine modelsystem. J Food Prot 71:1496–1499
Orro D, Budroni M, Angioni A, Cubaiu L, Cabras P, Farris GA (2006)Interazione tra lieviti vinari e ocratossina A in fase fermentativa. VQ4:40–47
Ospital M, Cazabeil JM, Betbeder AM, Tricard C, Creppy E, Medina B(1998) Ochratoxin A in wines. Rev Fr Oenol 169:16–18
Paster N (2008) Means to prevent contamination with patulin in apple-derived produce and with ochratoxin A in wines. In: Barkai-GolanR, Paster N (eds) Mycotoxins in fruits and vegetables. Elsevier, SanDiego, pp 351–386
Paterson RRM, Lima N (2011) Further mycotoxin effects from climatechange. Food Res Int 44:2555–2566
Patharajan S, Reddy KRN, Karthikeyan V, Spadaro D, Lore A, GullinoML, Garibaldi A (2011) Potential of yeast antagonists on in vitrobiodegradation of ochratoxin A. Food Control 22:290–296
Peltonen K, El-Nezami H, Haskard C, Ahokas J, Salminen S (2001)Aflatoxin B1 binding by dairy strains of lactic acid bacteria andbifidobacteria. J Dairy Sci 84:2152–2156
Petruzzi L, Sinigaglia M, Corbo MR, Beneduce L, Bevilacqua A (2013)Ochratoxin A removal by Saccharomyces cerevisiae strains: effectsof wine-related physicochemical factors. J Sci Food Agric 93:2110–2115
Petruzzi L, CorboMR, Sinigaglia M, Bevilacqua A (2014a) Yeast cells asadsorbing tools to remove ochratoxin A in a model wine. Int J FoodSci Technol 49:936–940
Petruzzi L, Bevilacqua A, Baiano A, Beneduce L, Corbo MR,Sinigaglia M (2014b) Study of Saccharomyces cerevisiae W13as a functional starter for the removal of ochratoxin A. FoodControl 35:373–377
Petruzzi L, Bevilacqua A, Baiano A, Beneduce L, Corbo MR, SinigagliaM (2014c) In vitro removal of ochratoxin A by two strains ofSaccharomyces cerevisiae and their performances under fermenta-tive and stressing conditions. J Appl Microbiol 116:60–70
Petruzzi L, Bevilacqua A, Corbo MR, Garofalo C, Baiano A, SinigagliaM (2014d) Selection of autochthonous Saccharomyces cerevisiaestrains as wine starters using a polyphasic approach and ochratoxinA removal. J Food Prot. In press
Piotrowska M (2012) Adsorption of ochratoxin A by Saccharomycescerevisiae living and non-living cells. Acta Aliment Hung 41:1–7
Piotrowska M, Zakowska Z (2005) The elimination of ochratoxin A bylactic acid bacteria strains. Pol J Microbiol 54:279–286
PiotrowskaM, Nowak A, Czyzowska A (2013) Removal of ochratoxin Aby wine Saccharomyces cerevisiae strains. Eur Food Res Technol236:441–447
Pizarro FJ, Jewett MC, Nielsen J, Agosin E (2008) Growth temperatureexerts differential physiological and transcriptional responses inlaboratory and wine strains of Saccharomyces cerevisiae. ApplEnviron Microbiol 74:6358–6368
Ponsone ML, Chiotta ML, Combina M, Dalcero AM, Chulze SN (2009)Fate of ochratoxin A content in Argentinean red wine during a pilotscale vinification. Rev Argent Microbiol 41:245–250
Pozo-BayónMA, Andújar-Ortiz I, Moreno-ArribasMV (2009) Scientificevidences beyond the application of inactive dry yeast preparationsin winemaking. Food Res Int 42:754–761
Pozo-Bayón MA, Monagas M, Bartolomé B, Moreno-Arribas VM(2012) Wine features related to safety and consumer health: anintegrated perspective. Crit Rev Food Sci Nutr 52:31–54
Pradelles R, Alexandre H, Ortiz-Julien A, Chassagne D (2008) Effects ofyeast cell-wall characteristics on 4-ethylphenol sorption capacity inmodel wine. J Agric Food Chem 56:11854–11861
Quintela S, VillaránMC, deArmentia IL, Elejalde E (2013) Ochratoxin Aremoval in wine: a review. Food Control 30:439–445
Ratola N, Martins L, Alves A (2004) Ochratoxin A in wines-assessingglobal uncertainty associated with the results. Anal Chim Acta 513:319–324
Ratola N, Abade E, Simões T, Venâncio A (2005) Evolution of ochra-toxin A content from must to wine in port wine microvinification.Anal Bioanal Chem 382:405–411
Reddy KRN, Salleh B, Saad B, Abbas HK, Abel CA, Shier WT (2010)An overview of mycotoxin contamination in foods and its implica-tions for human health. Toxin Rev 29:3–26
Ringot D, Lerzy B, Chaplain K, Bonhoure J, Auclair E, Larondelle Y(2007) In vitro biosorption of ochratoxin A on the yeast industry by-products: comparison of isotherm models. Bioresour Technol 98:1812–1821
Savino M, Cecchini F, Morassut M, Di Stefano R, Garcia-Moruno E(2006) Trattamenti utili per abbattere il contenuto di ocratossina Anei vini. L’Enologo Dicembre:97–102
Shetty HP, Jespersen L (2006) Saccharomyces cerevisiae and lactic acidbacteria as potential mycotoxin decontaminating agents. TrendsFood Sci Technol 17:48–55
Silva A, Galli R, Grazioli B, Fumi MD (2003) Metodi di riduzione diresidui di ocratossina A nei vini. Industrie delle Bevande Ottobre32(187):467–472
Appl Microbiol Biotechnol

SolfrizzoM, Avantaggiato G, Panzarini G, Visconti A (2010) Removal ofochratoxin A from contaminated red wines by repassage over grapepomaces. J Agric Food Chem 58:317–323
Soufleros EH, Tricard C, Boloumpasi EC (2003) Occurence of ochratox-in A in Greek wines. J Sci Food Agric 83:173–179
Torija MJ, Rozès N, Poblet M, Guillamón JM, Mas A (2003) Effects offermentation temperature on the strain population of Saccharomycescerevisiae. Int J Food Microbiol 80:47–53
Turbic A, Ahokas JT, Haskard CA (2002) Selective in vitro binding ofdietary mutagens, individually or in combination, by lactic acidbacteria. Food Addit Contam 19:144–152
van Egmond HP, Schothorst RC, Jonker MA (2007) Regulations relatingto mycotoxins in food: perspectives in a global and Europeancontext. Anal Bioanal Chem 389:147–157
Var I, Erginkaya Z, Kabak B (2009) Reduction of ochratoxin Alevels in white wine by yeast treatments. J Inst Brew 115:30–34
Vasserot Y, Caillet S, Maujean A (1997) Study of anthocyanin adsorptionby yeast lees. Effect of some physicochemical parameters. Am JEnol Vitic 48:433–437
Zimmerli B, Dick R (1995) Determination of ochratoxin A at the ppt levelin human blood, serum, milk and some foodstuffs by HPLC withenhanced fluorescence detection and immunoaffinity column clean-up: methodology and Swiss data. J Chromatogr B Biomed Appl666:85–99
Zimmerli B, Dick R (1996) Ochratoxin A in table wine and grapejuice: occurrence and risk assessment. Food Addit Contam13:655–668
Appl Microbiol Biotechnol




















