
Everyday Listening to auditory displays: Lessons from Acoustic Ecology
Upload
alfredadlerCategory
view
0download
0
doi:10.1016/j.jmb.2008.07.084 J. Mol. Biol. (2008) 382, 1113–1120
Available online at www.sciencedirect.com
COMMUNICATION
Crystal Structure of the Ig1 Domain of the Neural CellAdhesion Molecule NCAM2 Displays Domain Swapping
Kim K. Rasmussen1,2†, Nikolaj Kulahin1,2,3,4⁎†, Ole Kristensen2,Jens-Christian N. Poulsen1,2,3,5, Bent W. Sigurskjold1,2,3,6,Jette S. Kastrup2, Vladimir Berezin1, Elisabeth Bock1,Peter S. Walmod1 and Michael Gajhede2
*Corresponding author. Protein LaboUniversity of Copenhagen, Blegdam† K.K.R. and N.K., contributed eqAbbreviations used: CAM, cell ad
0022-2836/$ - see front matter © 2008 E
The crystal structure of the first immunoglobulin (Ig1) domain of neural celladhesion molecule 2 (NCAM2/OCAM/RNCAM) is presented at a reso-lution of 2.7 Å. NCAM2 is a member of the immunoglobulin superfamily ofcell adhesion molecules (IgCAMs). In the structure, two Ig domains interactby domain swapping, as the two N-terminal β-strands are interchanged. β-Strand swapping at the terminal domain is the accepted mechanism ofhomophilic interactions amongst the cadherins, another class of CAMs, butit has not been observed within the IgCAM superfamily. Gel-filtrationchromatography demonstrated the ability of NCAM2 Ig1 to form dimers insolution. Taken together, these observations suggest that β-strand swappingcould have a role in the molecular mechanism of homophilic binding forNCAM2.
© 2008 Elsevier Ltd. All rights reserved.
1Protein Laboratory,Department of Neuroscience andPharmacology, Faculty of HealthSciences, University ofCopenhagen, Blegdamsvej 3,DK-2200 Copenhagen,Denmark2Biostructural Research,Department of MedicinalChemistry, Faculty ofPharmaceutical Sciences,University of Copenhagen,Universitetsparken 2,DK-2100 Copenhagen,Denmark3ENKAM Pharmaceuticals A/S,Fruebjergvej 3, DK-2100Copenhagen, Denmark4Receptor Systems BiologyLaboratory, Hagedorn ResearchInstitute, Niels Steensens Vej 6,DK-2820 Gentofte, Denmark5Department of Chemistry,Faculty of Science, University ofCopenhagen, Universitetsparken5, DK-2100 Copenhagen,Denmark6Department of Biology,Faculty of Science, University ofCopenhagen, Universitetsparken13, DK-2100 Copenhagen,Denmark
ratory, Department of Neuroscience and Pharmacology, Faculty of Health Sciences,svej 3, DK-2200 Copenhagen, Denmark. E-mail address: [email protected] to this work.hesion molecule; NCAM, neural cell adhesion molecule.
lsevier Ltd. All rights reserved.

1114 NCAM2 Ig1 Crystal Displays Domain Swapping
Received 29 April 2008;received in revised form29 July 2008;accepted 29 July 2008Available online6 August 2008
Edited by I. Wilson
Keywords: crystal structure; differential scanning calorimetry; domainswapping; gel filtration; neural cell adhesion molecule 2Neural cell adhesion molecules (CAMs) have mul-tiple functions and their adhesive properties are ofpivotal importance for the self-assembly of thebillions of interconnections between cells in themammalian brain. Many families of neural CAMshave been identified, including the cadherins, theimmunoglobulin (Ig) superfamily of CAMs, theintegrins and the neurexins and neuroligins.1 Celladhesion molecules are generally multi-domainproteins with the extracellular part composed of Ig-like domains in tandem arrangement. The molecularbasis for their functions and binding specificities isstarting to emerge.It is known that cadherins interact via so-called
domain swapping between two N-terminal cad-herin domains.2,3 The cadherin domain is a memberof the class of Ig-like domains, and the 3-D structureof the cadherin domain resembles that of the Igdomains found in IgCAMs. The swapping is morespecifically a β-strand exchange, where the two N-terminal β-strands are interchanged between thetwo interacting cadherin molecules forming a so-called strand-dimer. The cadherin-mediated celladhesion process can thus be described as an equili-brium between three states: a state with conven-tional packing of β1-strand in the Ig-like sandwich(closed monomers); a state with the β1-strand of theIg1 domain swapped out but not engaged indimerization (open monomers); and a state withthe strand-dimer formed.3
The molecular basis for Ca2+-independent homo-philic CAM interaction has been investigated withinthe superfamily of IgCAMs. For example, thecrystal structures of the N-terminal Ig domainsIg1-Ig24 and of Ig1-Ig2-Ig35 of neural cell adhesionmolecule (NCAM) have been determined. Thesestructures reveal interactions between Ig1 and Ig2that are interpreted as interactions between NCAMmolecules on the same cell (cis-interactions) and amore complex scheme has been suggested on thebasis of the Ig1-Ig2-Ig3 structure, which alsoaccounts for interactions between opposing cells(trans-interactions), involving, for example, interac-tions between the Ig2 and Ig3 domains. Dimerformation has been reported for ICAM-1, but in thiscase the side-to-side interactions observed betweentwo Ig4 domains result from partial unfolding ofthe domains.6
Neural cell adhesion molecule 2 (NCAM2) wasoriginally described as a membrane-associated125 kDa protein.7–9 NCAM2 exists in transmem-
brane and GPI-anchored isoforms resulting fromalternative splicing of the transcript from a singlegene. NCAM2 transcripts are found in many regionsof the brain, and in several other tissues.10 In mouse,NCAM2 is reported to be expressed almost exclu-sively in neuronal tissue and predominantly in theolfactory sensory system.11,12 Furthermore, theexpression level of the transmembrane isoform ofNCAM2 inmouse brain is generally higher than thatof GPI-anchored NCAM2. The expression ofNCAM2 has been suggested to influence certaintypes of neurological diseases and to modulate neu-rite outgrowth, axonal guidance, synapse formationin the olfactory system and formation of dendriticbundles in the brain.10,13–15 Studies have indicatedthat the expression of NCAM2 has a role in thedevelopment of certain carcinomas.16–18
NCAM2 is a paralogue of NCAM, and the extra-cellular part of NCAM2 consists, as in NCAM, offive N-terminal Ig domains (Ig1–Ig5) followed bytwo fibronectin type III domains (FnIII1 and FnIII 2).NCAM2 contains eight or nine potential sites forN-linked glycosylation, none of which is situated inthe Ig1 domain. It has been demonstrated that fibro-blasts transfected with NCAM2 aggregate morestrongly than the parental non-transfected cells,indicating that NCAM2 mediates homophilic trans-interactions.12 Data from in vitro binding assayssupport this notion, but the domains involved in theinteractions are not identified.12 It has been demon-strated that NCAM2 does not participate in hetero-philic interactions with NCAM.12
We report here the crystal structure of the Ig1domain of NCAM2 (Table 1) and discuss the impli-cations for homophilic cell–cell interactions.
The NCAM2 Ig1 domain forms adomain-swapped dimer
The crystallized Ig1 domain of human NCAM2adopts the topology of an intermediate (I)-setdomain.19 In crystals, the domain forms a dimer,and a swapping of the N-terminal β-strand (strandA) between two molecules (molecules A and B)has occurred, resulting in a dimer with twomolecules related by a crystallographic twofoldaxis (Fig. 1a). Residues 2–6 (Gln2, Val3, Thr4, Ile5and Ser6) from molecule A are part of the β-sheetof molecule B (Fig. 1b). The following residues,Leu7-Ser8, form a hinge region, and at Lys9 the

Table 1. Data collection and refinement statistics
X-ray sourceI911-5 MAX-Lab,Lund, Sweden
A. Data collectionWavelength (Å) 0.907Space group I4122Unit-cell parameters
a (Å) 47.28b (Å) 47.28c (Å) 196.72
Resolution (Å) 15.2–2.7(2.85–2.7)
No. observations 34,231No. unique observations 3356Redundancy 10.2 (10.4)Completeness (%) 99.6 (99.2)I/σ(I) 17.3 (7.1)Rsym (%) 3.6 (9.6)
B. RefinementRwork (%) 22.7Rfree (%) 26.8rmsd from ideal
Bond lengths (Å) 0.007Bond angles (°) 1.059
Mean B-factor (Å2) 48.9Ramachandran plot regionsa
Most favored (%) 90.4Allowed (%) 8.4Generally allowed (%) 1.2Disallowed (%) 0.0
Screening for initial crystallization hits was done with an Oryx 8System (Douglas Instruments Ltd) with the commercial JCSG+,PACT (Qiagen) and Index (Hampton Research) crystallizationscreens. Crystallization was achieved in hanging-drop experi-ments by mixing 1 μl of protein (3 mg/ml) and 1 μl of reservoirsolution (0.1 M citric acid, pH 3.5, 20% (w/v) PEG 3350). X-raydata were collected (I911-5, MAX-Lab, Lund, Sweden) at 100 Kfrom a cryo-protected crystal (reservoir solution containing 20%(v/v) glycerol). The crystals diffracted anisotropically, and data to2.7 Å were indexed, integrated and scaled using the programsMOSFLM and SCALA.23 The structure was determined by themolecular replacement method using the program PHASER24
with NCAM Ig1 as a search model (PDB code 1EPF). Modelbuilding was performed using the program ARP/wARP25 andmanual correction using the program COOT.26 The programPHENIX was used for refinement27 and the quality of the finalmodel was estimated by PROCHECK analysis.28 Values inparentheses are for the highest resolution shell.Rsym=∑hkl(∑i[∑Ihkl,i–bIhklN|])/∑hkl,ibIhklN, where Ihkl, i is the inten-sity of an individual measurement of the reflection with Millerindices h, k, and l, and bIhklN is the mean intensity of that reflection.Rwork=∑hkl(||Fo, hkl|–|Fc,hkl||)/|Fo,hkl|,where|Fo,hkl|and|Fc, hkl|are the observed and calculated structure factor amplitudes.Rfree is equivalent to Rwork, but is calculated with 5% of reflectionsomitted from the refinement process.
a According to PROCHECK definitions.28
1115NCAM2 Ig1 Crystal Displays Domain Swapping
usual intramolecular β-sheet interactions resumewithin molecule A.The dimer interface was analyzed using the PISA
server.20 A buried interface area of 2820 Å2 wasreported. There are 24 hydrogen bonds in the inter-face, related pairwise by the twofold symmetry, butthese hydrogen bonds seem not to be the majorcontributors to the dimer interaction energy, sincesimilar interactions can be formed between closedmonomers and solvent. The sequence of the first
eight residues of NCAM2 (excluding the signalpeptide preceding the Ig1 domain) is LQVTISLS,which has a characteristic pattern of hydrophobicresidues at every odd-numbered position. Theseresidues contribute to the core of the β-sandwich ofthe Ig domain, where Leu1, Val3, Ile5 and Leu7 ofmolecule A become part of the hydrophobic core ofmolecule B (Fig. 1b). These hydrophobic residues arethe main contributors to the interaction energy witha total contribution of –6.7 kcal/mol relative to theisolated open monomers. However, except for Leu7,these residues do not seem to contribute to theenergy difference between the closed state and thestrand-dimer state, as similar interactions areexpected in the closed state, based on comparisonto the known structure of the NCAM Ig1 domain(Fig. 2).4,5 Val3 and Ile5 are fully conserved in bothNCAM2 and NCAM, while Leu7 is replaced byanother hydrophobic residue, a proline, in NCAM.Instead, the main contributions to the differencebetween the closed state and the strand-dimer stateenergymost likely come from the enhanced burial ofresidues Leu7 and Val10 in a hydrophobic clustergenerated by symmetry, in addition to shielding ofthe Phe19 side-chains by interactions with Cβ ofSer8, which is also located in the strand-exchangehinge region (Fig. 1a).In summary, the crystal structure suggests that the
domain-swapped dimer is also stable in solution.
NCAM2 Ig1-Ig1 dimers are present in solution
Gel-filtration chromatography was used to inves-tigate oligomerization in solution. Chromato-graphy of concentrated recombinant Ig1 resultedin three elution peaks corresponding to higholigomer aggregates, dimers and monomers, respec-tively (see Fig. 3a, pink line). To investigate thedimer/monomer equilibrium, the fraction corres-ponding to monomeric Ig1 was collected, concen-trated and subjected again to gel-filtration chroma-tography. The sample eluted as a single peak,suggesting slow transition to equilibrium betweenthe monomeric and dimeric forms (Fig. 3a, blackline). To test this suggestion, monomeric Ig1 wasincubated for two days before the chromatographywas performed. Under these conditions, the sampleeluded as two peaks (corresponding to dimer andmonomer, respectively), suggesting that a fractionof the monomeric form had adopted the dimericform (Fig. 3a, green line). A competition experimentwas performed to further investigate the monomer–dimer equilibrium (Fig. 3a, red line), in whichdimeric Ig1 was incubated in the presence of a ten-fold molar excess of a monomeric peptide corres-ponding to the N-terminal part of NCAM2. Still,no monomeric Ig1 was observed after gel-filtrationchromatography, indicating that dimeric Ig1 is verystable once formed.In summary, these experiments showed that
dimerization of recombinant NCAM2 Ig1 occurs insolution, and that a monomer–dimer equilibrium is
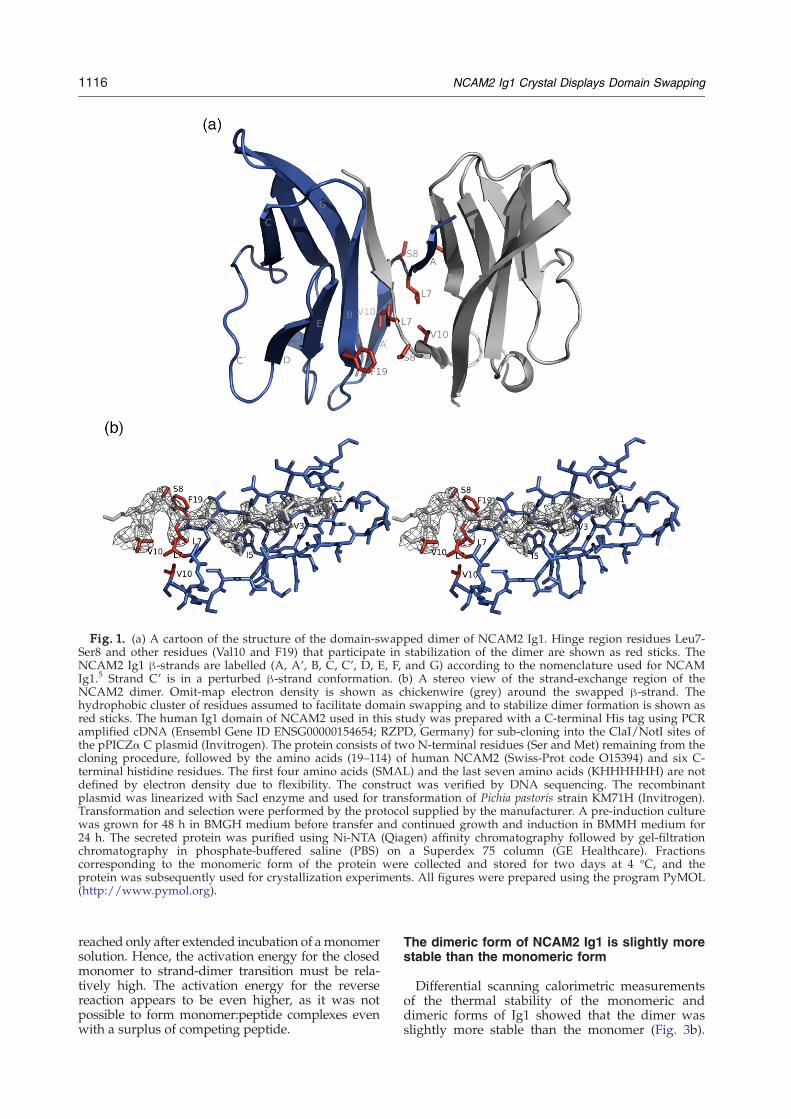
Fig. 1. (a) A cartoon of the structure of the domain-swapped dimer of NCAM2 Ig1. Hinge region residues Leu7-Ser8 and other residues (Val10 and F19) that participate in stabilization of the dimer are shown as red sticks. TheNCAM2 Ig1 β-strands are labelled (A, A', B, C, C', D, E, F, and G) according to the nomenclature used for NCAMIg1.5 Strand C' is in a perturbed β-strand conformation. (b) A stereo view of the strand-exchange region of theNCAM2 dimer. Omit-map electron density is shown as chickenwire (grey) around the swapped β-strand. Thehydrophobic cluster of residues assumed to facilitate domain swapping and to stabilize dimer formation is shown asred sticks. The human Ig1 domain of NCAM2 used in this study was prepared with a C-terminal His tag using PCRamplified cDNA (Ensembl Gene ID ENSG00000154654; RZPD, Germany) for sub-cloning into the ClaI/NotI sites ofthe pPICZα C plasmid (Invitrogen). The protein consists of two N-terminal residues (Ser and Met) remaining from thecloning procedure, followed by the amino acids (19–114) of human NCAM2 (Swiss-Prot code O15394) and six C-terminal histidine residues. The first four amino acids (SMAL) and the last seven amino acids (KHHHHHH) are notdefined by electron density due to flexibility. The construct was verified by DNA sequencing. The recombinantplasmid was linearized with SacI enzyme and used for transformation of Pichia pastoris strain KM71H (Invitrogen).Transformation and selection were performed by the protocol supplied by the manufacturer. A pre-induction culturewas grown for 48 h in BMGH medium before transfer and continued growth and induction in BMMH medium for24 h. The secreted protein was purified using Ni-NTA (Qiagen) affinity chromatography followed by gel-filtrationchromatography in phosphate-buffered saline (PBS) on a Superdex 75 column (GE Healthcare). Fractionscorresponding to the monomeric form of the protein were collected and stored for two days at 4 °C, and theprotein was subsequently used for crystallization experiments. All figures were prepared using the program PyMOL(http://www.pymol.org).
1116 NCAM2 Ig1 Crystal Displays Domain Swapping
reached only after extended incubation of amonomersolution. Hence, the activation energy for the closedmonomer to strand-dimer transition must be rela-tively high. The activation energy for the reversereaction appears to be even higher, as it was notpossible to form monomer:peptide complexes evenwith a surplus of competing peptide.
The dimeric form of NCAM2 Ig1 is slightly morestable than the monomeric form
Differential scanning calorimetric measurementsof the thermal stability of the monomeric anddimeric forms of Ig1 showed that the dimer wasslightly more stable than the monomer (Fig. 3b).
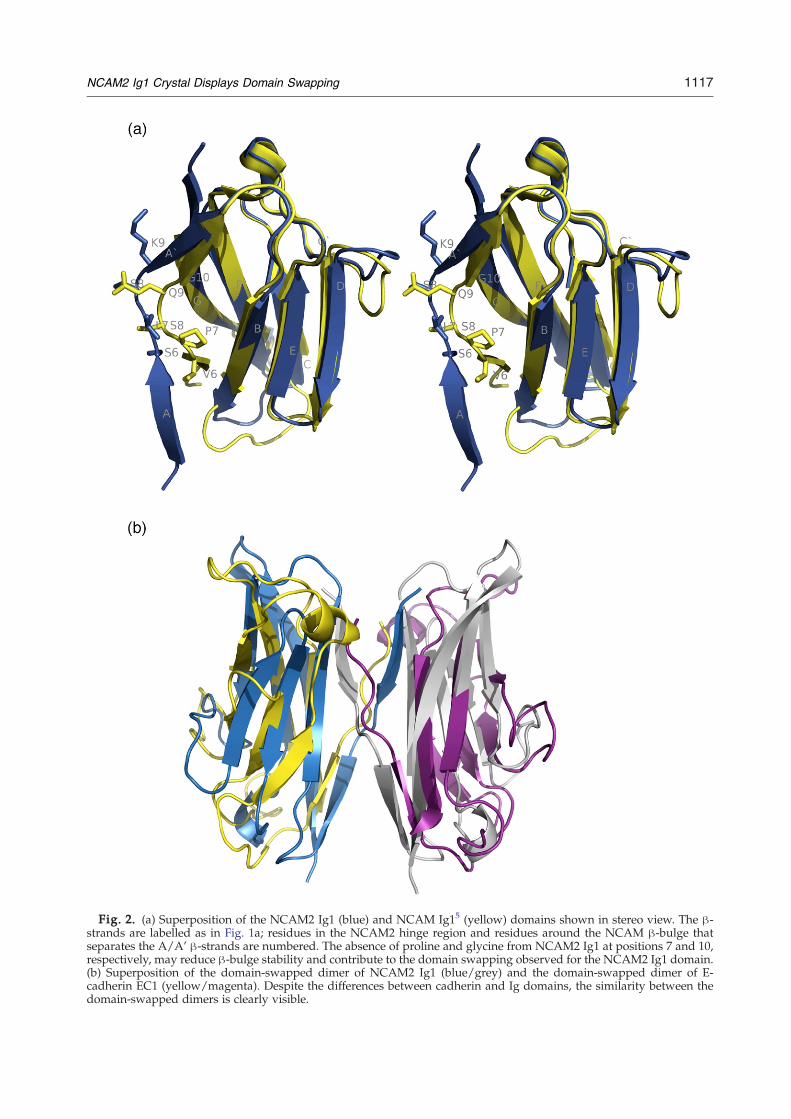
Fig. 2. (a) Superposition of the NCAM2 Ig1 (blue) and NCAM Ig15 (yellow) domains shown in stereo view. The β-strands are labelled as in Fig. 1a; residues in the NCAM2 hinge region and residues around the NCAM β-bulge thatseparates the A/A' β-strands are numbered. The absence of proline and glycine from NCAM2 Ig1 at positions 7 and 10,respectively, may reduce β-bulge stability and contribute to the domain swapping observed for the NCAM2 Ig1 domain.(b) Superposition of the domain-swapped dimer of NCAM2 Ig1 (blue/grey) and the domain-swapped dimer of E-cadherin EC1 (yellow/magenta). Despite the differences between cadherin and Ig domains, the similarity between thedomain-swapped dimers is clearly visible.
1117NCAM2 Ig1 Crystal Displays Domain Swapping
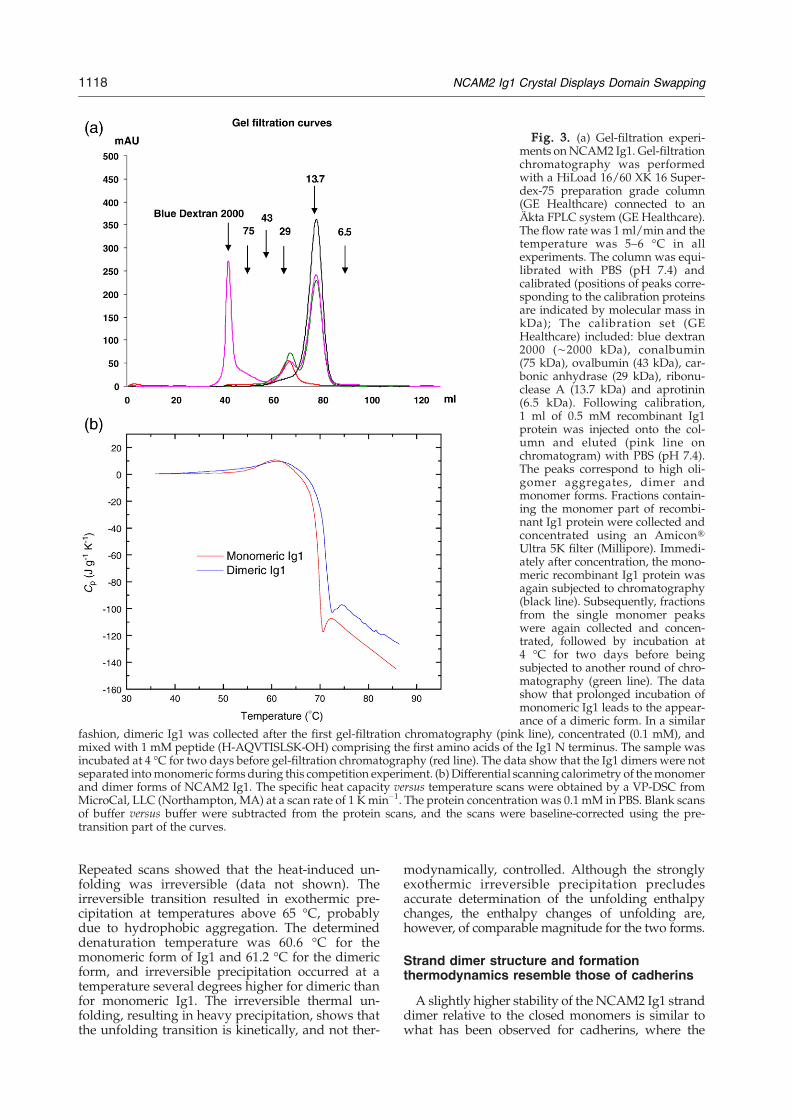
Fig. 3. (a) Gel-filtration experi-ments onNCAM2 Ig1. Gel-filtrationchromatography was performedwith a HiLoad 16/60 XK 16 Super-dex-75 preparation grade column(GE Healthcare) connected to anÄkta FPLC system (GE Healthcare).The flow rate was 1 ml/min and thetemperature was 5–6 °C in allexperiments. The column was equi-librated with PBS (pH 7.4) andcalibrated (positions of peaks corre-sponding to the calibration proteinsare indicated by molecular mass inkDa); The calibration set (GEHealthcare) included: blue dextran2000 (∼2000 kDa), conalbumin(75 kDa), ovalbumin (43 kDa), car-bonic anhydrase (29 kDa), ribonu-clease A (13.7 kDa) and aprotinin(6.5 kDa). Following calibration,1 ml of 0.5 mM recombinant Ig1protein was injected onto the col-umn and eluted (pink line onchromatogram) with PBS (pH 7.4).The peaks correspond to high oli-gomer aggregates, dimer andmonomer forms. Fractions contain-ing the monomer part of recombi-nant Ig1 protein were collected andconcentrated using an Amicon®Ultra 5K filter (Millipore). Immedi-ately after concentration, the mono-meric recombinant Ig1 protein wasagain subjected to chromatography(black line). Subsequently, fractionsfrom the single monomer peakswere again collected and concen-trated, followed by incubation at4 °C for two days before beingsubjected to another round of chro-matography (green line). The datashow that prolonged incubation ofmonomeric Ig1 leads to the appear-ance of a dimeric form. In a similar
fashion, dimeric Ig1 was collected after the first gel-filtration chromatography (pink line), concentrated (0.1 mM), andmixed with 1 mM peptide (H-AQVTISLSK-OH) comprising the first amino acids of the Ig1 N terminus. The sample wasincubated at 4 °C for two days before gel-filtration chromatography (red line). The data show that the Ig1 dimers were notseparated intomonomeric forms during this competition experiment. (b) Differential scanning calorimetry of themonomerand dimer forms of NCAM2 Ig1. The specific heat capacity versus temperature scans were obtained by a VP-DSC fromMicroCal, LLC (Northampton, MA) at a scan rate of 1 Kmin−1. The protein concentration was 0.1 mM in PBS. Blank scansof buffer versus buffer were subtracted from the protein scans, and the scans were baseline-corrected using the pre-transition part of the curves.
1118 NCAM2 Ig1 Crystal Displays Domain Swapping
Repeated scans showed that the heat-induced un-folding was irreversible (data not shown). Theirreversible transition resulted in exothermic pre-cipitation at temperatures above 65 °C, probablydue to hydrophobic aggregation. The determineddenaturation temperature was 60.6 °C for themonomeric form of Ig1 and 61.2 °C for the dimericform, and irreversible precipitation occurred at atemperature several degrees higher for dimeric thanfor monomeric Ig1. The irreversible thermal un-folding, resulting in heavy precipitation, shows thatthe unfolding transition is kinetically, and not ther-
modynamically, controlled. Although the stronglyexothermic irreversible precipitation precludesaccurate determination of the unfolding enthalpychanges, the enthalpy changes of unfolding are,however, of comparable magnitude for the two forms.
Strand dimer structure and formationthermodynamics resemble those of cadherins
A slightly higher stability of the NCAM2 Ig1 stranddimer relative to the closed monomers is similar towhat has been observed for cadherins, where the

1119NCAM2 Ig1 Crystal Displays Domain Swapping
energy difference between the closed monomers andthe strand dimer is relatively low at ∼4 kcal/mollower for the strand dimer when compared to theclosed monomer.3 The existence of a binding site forthe swapped strand in the monomer that is close toidentical with that in the dimer partner decreases thefree energy difference. This had been pointed out as ageneral feature of domain swapping in work withthe diphtheria toxin.21 The energy of the open E-cadherin Ig1 domain (EC1) is ∼7 kcal/mol higherthan the energy of the closed form, 3 which results inlow-affinity binding with a high energy barrier forthe formation of the strand dimer from the mono-mers and an even higher barrier for the oppositereaction. This is considered the basis of the specificityof cell–cell adhesion by cadherins. 3 The results of thegel-filtration experiments on NCAM2 Ig1 are also inagreement with the cadherin strand dimer formationthermodynamics, since the monomer fraction willform dimers only after days, and the dimers appearvery stable in the peptide competition experiments.This further supports the suggestion that the dimerobserved in solution is a strand dimer.The structure of the NCAM2 Ig1 domain-swapped
dimer has been compared to that of EC1 (Fig. 2b).22
Despite the differences between the Ig1 and EC1domains, the organisation of the dimers is identical.The β1 (strand A) of NCAM2 obtains a better incor-poration into the β-sheet of the neighbouring do-main than the β1 strand of EC1, since the hydrogenbonding pattern and ϕ/φ angles are closer to idealfor NCAM2 Ig1 than for those seen in EC1.
Comparison of Ig1 domains of NCAM2 andNCAM
In contrast to NCAM2, domain swapping hasnot been reported in NCAM. The Ig1 domains ofNCAM2 and NCAM share 49% of sequence iden-tity,4,5 and the structures display a root-mean-square deviation (rmsd) of 0.77 Å when super-imposing 583 out of the total of 651 atoms in thetwo domains (Fig. 2a). The major conformationaldifferences are observed in the β1 strand regions.The Ig1 domain of NCAM does not display domainswapping and its short β1 strand (residues 3–6) ispart of a classical β-sandwich structure. NCAM Ig1residues 7–8 form a β-bulge and at residue 9 a β-strand like conformation is resumed. It is note-worthy that 80% of the NCAM Pro7 residue isburied within the Ig1-Ig2 interface; an interface thathas been reported to be responsible for homophilicNCAM cis-interactions.4,5 The fact that domainswapping has not been observed in NCAM Ig1might be related to sequence differences at posi-tions in the domain-swapping NCAM2 hinge re-gion. At the four important positions (Leu7, Ser8,Val10 and Phe19), which are major contributors toNCAM2 dimerization (Fig. 1a and b), NCAMdiffers at two positions: Leu7 is proline and Val10is glycine. If NCAM and NCAM2 are assumed tofunction differently, these substitutions might in-
fluence the conformational flexibility and stabilizethe β-bulge of NCAM in this region, preventing aNCAM2-like hinge region transition as required fordimer formation.
The role of a strand dimer in homophilic cis ortrans cell adhesion interactions
Taken together, these findings suggest that do-main swapping could be involved in the dynamicevent of NCAM2 homophilic binding, as a thermo-dynamically favoured state. Whether NCAM2 Ig1domain swapping is involved in cis- or trans-interactions remains to be resolved. A mechanismthat involves kinetically controlled initial interac-tions like those reported for the NCAM system,5
followed by the formation of thermodynamicallyfavoured strand dimers, is also a possibility. Ela-boration on the mechanism of NCAM2 cell–celladhesion and attempts to model the adhesioncomplex assembly, as done in the NCAM system,will be done in the context of multi-domain struc-tural analysis of NCAM2.
Protein Data Bank accession code
The structure has been deposited in the ProteinData Bank with accession code2VAJ.
Acknowledgements
The beamline scientists at MAXLAB, Lund, Swe-den are gratefully acknowledged for their technicalsupport. This work was supported by grants fromDANSYNC (Danish Center for Synchrotron-BasedResearch), Carlsbergfondet, Beckett-Fonden andAugustinus Fonden.
References
1. Shapiro, L., Love, J. & Colman, D. R. (2007). Adhesionmolecules in the nervous system: structural insightsinto function and diversity. Annu. Rev. Neurosci. 30,451–474.
2. Boggon, T. J., Murray, J., Chappuis-Flament, S., Wong,E., Gumbiner, B. M. & Shapiro, L. (2002). C-cadherinectodomain structure and implications for cell adhe-sion mechanisms. Science, 296, 1308–1313.
3. Chen, C. P., Posy, S., Ben-Shaul, A., Shapiro, L. &Honig, B. H. (2005). Specificity of cell-cell adhesion byclassical cadherins: Critical role for low-affinitydimerization through beta-strand swapping. Proc.Natl Acad. Sci. USA, 102, 8531–8536.
4. Kasper, C., Rasmussen, H., Kastrup, J. S., Ikemizu, S.,Jones, E. Y., Berezin, V. et al. (2000). Structural basis ofcell-cell adhesion by NCAM. Nature Struct. Biol. 7,389–393.
5. Soroka, V., Kolkova, K., Kastrup, J. S., Diederichs, K.,Breed, J., Kiselyov, V. V. et al. (2003). Structure and

1120 NCAM2 Ig1 Crystal Displays Domain Swapping
interactions of NCAM Ig1-2-3 suggest a novel zippermechanism for homophilic adhesion. Structure, 11,1291–1301.
6. Chen, X., Kim, T. D., Carman, C. V., Mi, L. Z., Song, G.& Springer, T. A. (2007). Structural plasticity in Igsuperfamily domain 4 of ICAM-1 mediates cell sur-face dimerization. Proc. Natl Acad. Sci. USA, 104,15358–15363.
7. Schwob, J. E. & Gottlieb, D. I. (1986). The primaryolfactory projection has two chemically distinct zones.J. Neurosci. 6, 3393–3404.
8. Schwob, J. E. & Gottlieb, D. I. (1988). Purification andcharacterization of an antigen that is spatially segre-gated in the primary olfactory projection. J. Neurosci. 8,3470–3480.
9. Kulahin, N. & Walmod, P. S. (2008). The neural celladhesion molecule NCAM2/OCAM/RNCAM, aclose relative to NCAM. Neurochem. Res. 2008 Mar27. [Epub ahead of print].
10. Paoloni-Giacobino, A., Chen, H. & Antonarakis, S. E.(1997). Cloning of a novel human neural cell adhesionmolecule gene (NCAM2) that maps to chromosomeregion 21q21 and is potentially involved in Downsyndrome. Genomics, 43, 43–51.
11. Alenius, M. & Bohm, S. (1997). Identification of anovel neural cell adhesion molecule-related gene witha potential role in selective axonal projection. J. Biol.Chem. 272, 26083–26086.
12. Yoshihara, Y., Kawasaki, M., Tamada, A., Fujita, H.,Hayashi, H., Kagamiyama, H. & Mori, K. (1997).OCAM: A new member of the neural cell adhesionmolecule family related to zone-to-zone projection ofolfactory and vomeronasal axons. J. Neurosci. 17,5830–5842.
13. Bailey, A., Luthert, P., Dean, A., Harding, B., Janota, I.,Montgomery, M. et al. (1998). A clinicopathologicalstudy of autism. Brain, 121, 889–905.
14. Ichinohe, N., Yoshihara, Y., Hashikawa, T. & Rock-land, K. S. (2003). Developmental study of dendriticbundles in layer 1 of the rat granular retrosplenialcortex with special reference to a cell adhesionmolecule, OCAM. Eur. J. Neurosci. 18, 1764–1774.
15. Hamlin, J. A., Fang, H. & Schwob, J. E. (2004).Differential expression of the mammalian homologueof fasciclin II during olfactory development in vivoand in vitro. J. Comp. Neurol. 474, 438–452.
16. Wreesmann, V. B., Shi, W., Thaler, H. T., Poluri, A.,Kraus, D. H., Pfister, D. et al. (2004). Identification ofnovel prognosticators of outcome in squamous cellcarcinoma of the head and neck. J. Clin. Oncol. 22,3965–3972.
17. Xu, L. L., Su, Y. P., Labiche, R., Segawa, T.,Shanmugam, N., McLeod, D. G. et al. (2001). Quan-titative expression profile of androgen-regulatedgenes in prostate cancer cells and identification ofprostate-specific genes. Int. J. Cancer, 92, 322–328.
18. Wittig, I. & Groner, B. (2005). Signal transducer andactivator of transcription 5 (STAT5), a crucial regulatorof immune and cancer cells. Curr. Drug Targets ImmuneEndocr. Metabol. Disord. 5, 449–463.
19. Chothia, C. & Jones, E. Y. (1997). The molecularstructure of cell adhesion molecules. Annu. Rev.Biochem. 66, 823–862.
20. Krissinel, E. & Henrick, K. (2007). Inference ofmacromolecular assemblies from crystalline state. J.Mol. Biol. 372, 774–797.
21. Liu, Y. & Eisenberg, D. (2002). 3Ddomain swapping: asdomains continue to swap. Protein Sci. 11, 1285–1299.
22. Shapiro, L., Fannon, A. M., Kwong, P. D., Thompson,A., Lehmann, M. S., Grubel, G. et al. (1995). Structuralbasis of cell-cell adhesion by cadherins. Nature, 374,327–337.
23. Collaborative Computational Project, Number 4.(1994). The CCP4 suite: programs for protein crystal-lography. Acta Crystallogr. D, 50, 760–763.
24. Storoni, L. C., McCoy, A. J. & Read, R. J. (2004).Likelihood-enhanced fast rotation functions. ActaCrystallogr. D, 60, 432–438.
25. Perrakis, A., Morris, R. & Lamzin, V. S. (1999). Auto-mated protein model building combined with itera-tive structure refinement.Nature Struct. Biol. 6, 458–463.
26. Emsley, P. & Cowtan, K. (2004). Coot: model-buildingtools for molecular graphics. Acta Crystallogr. D, 60,2126–2132.
27. Adams, P. D., Grosse-Kunstleve, R. W., Hung, L. W.,Ioerger, T. R., McCoy, A. J., Moriarty, N. W. et al.(2002). PHENIX: building new software for auto-mated crystallographic structure determination. ActaCrystallogr. D, 58, 1948–1954.
28. Laskowski, R. A., Moss, D. S. & Thornton, J. M. (1993).Main-chain bond lengths and bond angles in proteinstructures. J. Mol. Biol. 231, 1049–1067.
Copyright © 2022 FDOKUMEN




















