
Correlation among Thermosensitive Period, Estradiol Response, and Gonad Differentiation in the Sea...
13
Correlation among Thermosensitive Period, Estradiol Response, and Gonad Differentiation in the Sea Turtle Lepidochelys olivacea Horacio Merchant-Larios,* Salvador Ruiz-Ramirez,² Norma Moreno-Mendoza,* and Alejandro Marmolejo-Valencia* *Department of Cell Biology, Instituto de Investigaciones Biome ´dicas, UNAM, Apartado Postal 70228, Me ´xico, D.F., Me ´xico 04510; and ²Centro de Ecologı ´a Costera, S. P. Melaque, Universidad de Guadalajara, Guadalajara, Me ´xico Accepted May 28, 1997 Reptile embryos with temperature sex determination have a thermosensitive period (TSP). The finding that exogenous estradiol (E2) overcomes the effect of male- promoting temperature led to the idea that temperature may regulate estrogen concentration in the gonad during TSP. Since interspecific variations in TSP and in the effect of exogenous E2 exist, we undertook a study in the olive ridley Lepidochelys olivacea. Four parameters were correlated: the TSP (time dimension), the thermosensi- tive stages (rate of development), gonad development (histological aspect), and the estradiol response. Two kinds of experiments were performed: (1) Eggs were shifted once, at different stages of development, from a male-promoting temperature to a female-temperature (or vice versa) for the remainder of development. (2) Eggs at male-promoting temperature were treated once with 6 or 12 mg of estradiol (E2) at various times of incubation. Sex ratio was established around hatching in each experimental series. We found that the temporal dimen- sion of the TSP was around 7 days (Days 20–27 of incubation) at a male-promoting or a female-promoting temperature. The rate of development of the whole embryo and gonadal growth was faster at female- promoting temperature than at male-promoting tempera- ture. Formation of the genital ridge began at stage 21–22 and histological differentiation of the gonads occurred around stage 26–27. Although these stages coincided with the TSP, at male-promoting temperature the thermo- sensitive stages occurred earlier (from stages 20–21 to stages 23–24) than at female-promoting temperature (from stages 23–24 to stages 26–27). Thus, at male promoting-temperature, sex was determined in embryos with incipient or undifferentiated gonads. In contrast, E2 treatment continued to feminize the gonads of embryos at a male-promoting temperature beyond the TSP up to stage 25–26, but the E2-induced ovaries were signifi- cantly smaller than temperature-induced ovaries. It is suggested that the doses of E2 used were higher than the concentration of endogenous E2 required for normal sex determination. The lack of correlation between sex determination and gonad differentiation suggests that irreversible molecular processes underlying sex determi- nation occur earlier at male- than at female-promoting temperature. Results suggest that the male sex may be the default state and that the female condition must be imposed upon it. r 1997 Academic Press Sexual differentiation of the gonads is dependent on incubation temperature in five sea turtle species: Eret- mochelys imbricata (Dalrymple et al., 1985), Lepidochelys olivacea (Morreale et al., 1982) Caretta caretta (Yntema and Mrosovsky, 1980, 1982), Chelonia mydas (Miller and Limpus, 1980, Morreale et al., 1982), and Dermochelys coriacea (Rimblot et al., 1985). Pivotal temperatures (temperature during incubation at constant tempera- ture which gives 50% individuals of each sexual General and Comparative Endocrinology 107, 373–385 (1997) Article No. GC976946 373 0016-6480/97 $25.00 Copyright r 1997 by Academic Press All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Correlation among Thermosensitive Period, Estradiol Response, and Gonad Differentiation in the Sea...

Correlation among Thermosensitive Period, EstradiolResponse, and Gonad Differentiation in the Sea TurtleLepidochelys olivacea
Horacio Merchant-Larios,* Salvador Ruiz-Ramirez,† Norma Moreno-Mendoza,*and Alejandro Marmolejo-Valencia**Department of Cell Biology, Instituto de Investigaciones Biomedicas, UNAM, Apartado Postal 70228, Mexico, D.F.,Mexico 04510; and †Centro de Ecologıa Costera, S. P. Melaque, Universidad de Guadalajara, Guadalajara, Mexico
Accepted May 28, 1997
Reptile embryos with temperature sex determinationhave a thermosensitive period (TSP). The finding thatexogenous estradiol (E2) overcomes the effect of male-promoting temperature led to the idea that temperaturemay regulate estrogen concentration in the gonad duringTSP. Since interspecific variations in TSP and in theeffect of exogenous E2 exist, we undertook a study in theolive ridley Lepidochelys olivacea. Four parameters werecorrelated: the TSP (time dimension), the thermosensi-tive stages (rate of development), gonad development(histological aspect), and the estradiol response. Twokinds of experiments were performed: (1) Eggs wereshifted once, at different stages of development, from amale-promoting temperature to a female-temperature (orvice versa) for the remainder of development. (2) Eggs atmale-promoting temperature were treated once with 6 or12 mg of estradiol (E2) at various times of incubation.Sex ratio was established around hatching in eachexperimental series. We found that the temporal dimen-sion of the TSP was around 7 days (Days 20–27 ofincubation) at a male-promoting or a female-promotingtemperature. The rate of development of the wholeembryo and gonadal growth was faster at female-promoting temperature than at male-promoting tempera-ture. Formation of the genital ridge began at stage 21–22and histological differentiation of the gonads occurredaround stage 26–27. Although these stages coincidedwith the TSP, at male-promoting temperature the thermo-
sensitive stages occurred earlier (from stages 20–21 tostages 23–24) than at female-promoting temperature(from stages 23–24 to stages 26–27). Thus, at malepromoting-temperature, sex was determined in embryoswith incipient or undifferentiated gonads. In contrast, E2treatment continued to feminize the gonads of embryosat a male-promoting temperature beyond the TSP up tostage 25–26, but the E2-induced ovaries were signifi-cantly smaller than temperature-induced ovaries. It issuggested that the doses of E2 used were higher than theconcentration of endogenous E2 required for normal sexdetermination. The lack of correlation between sexdetermination and gonad differentiation suggests thatirreversible molecular processes underlying sex determi-nation occur earlier at male- than at female-promotingtemperature. Results suggest that the male sex may bethe default state and that the female condition must beimposed upon it. r 1997 Academic Press
Sexual differentiation of the gonads is dependent onincubation temperature in five sea turtle species: Eret-mochelys imbricata (Dalrymple et al., 1985), Lepidochelysolivacea (Morreale et al., 1982) Caretta caretta (YntemaandMrosovsky, 1980, 1982), Chelonia mydas (Miller andLimpus, 1980, Morreale et al., 1982), and Dermochelyscoriacea (Rimblot et al., 1985). Pivotal temperatures(temperature during incubation at constant tempera-ture which gives 50% individuals of each sexual
General and Comparative Endocrinology 107, 373–385 (1997)Article No. GC976946
373
0016-6480/97 $25.00Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

phenotype; Mrosovsky and Pieau, 1991) are known forthe last three species and have been placed around 29°.Temperatures around 27° or 32° are male- or female-promoting, respectively. In C. caretta, it was found thatthe thermosensitive period (TSP) was at stages 21–26(Yntema and Mrosovsky, 1982; Harry and Williams,1991).In L. olivacea, a descriptive study of the morphology,
ultrastructure and some histochemical aspects on theestablishment of the genital ridge and gonads incu-bated at male- or female-promoting temperatures wasmade but did not attempt to establish the TSP (Mer-chant-Larios et al., 1989). In D. coriacea, it has beenfound that higher aromatase activity at female- than atmale-promoting temperature may correspond to TSP(Desvages et al., 1993).Administration of exogenous estradiol to embryos
incubated at masculinizing temperatures overcame theeffect of temperature and resulted in all-femalehatchlings in several reptiles with temperature sexdetermination (TSD) (Bull et al., 1988; Crews et al., 1991;Dorizzi et al., 1991; Wibbels et al., 1991, 1993; Pieau etal., 1994). Moreover, treatment of eggs with aromataseinhibitors incubated at feminizing temperature dis-rupted ovary differentiation or induced testis differen-tiation at temperatures that otherwise yielded bothsexes (Lance and Bogart, 1992; Rhen and Lang, 1994;Crews and Bergeron, 1994; Dorizzi et al., 1994, 1996;Wibbels and Crews, 1994). The sensitive period forestrogen effects on gonadal development apparentlycoincided with the TSP which, in turn, coincided withthe establishment of an undifferentiated gonad and theonset of its histological sex differentiation (Gutzke andChymiy, 1988; Raynaud and Pieau, 1985; Dorizzi et al.,1991; Wibbels et al., 1991, 1993). However, interspecificvariations have been found and estrogen effects on seaturtles are not available.The TSP implies at least three related but different
aspects: the temporal dimension, the stage of develop-ment, and the histological differentiation of the gonad.Since the rate of embryonic development is dramati-cally affected by temperature (Yntema andMrosovsky,1980; Miller, 1985), and temperature shifts affect differ-ent stages of development depending on the back-ground temperature (Yntema, 1979; Pieau and Dorizzi,1981), the TSP (temporal dimension) and the thermo-sensitive stages may be different. Consequently, if the
developmental rate of the gonad follows the develop-mental rate of the embryo, the TSPmay affect differentstages of its histological differentiation at male- orfemale-promoting temperatures. Therefore, a compre-hensive study correlating the temporal dimension, thestage of development, and the histological differentia-tion of the gonad in different species will be the basis tounderstand the physiological basis of temperature-dependent sex determination.In the current work, we undertook a study correlat-
ing the TSP, the thermosensitive stages, and the histo-logical differentiation of the gonad in the sea turtle L.olivacea. The operational criterion for the TSP wasbased on the response of the embryos incubated atconstant temperature to one shift from male-promot-ing temperature to female-promoting temperature orvice versa. Moreover, the effect of exogenous estradioland feminizing temperature were compared. Our find-ings lay the ground work for our studies concernedwith the steroidogenic pattern in developing embryosof L. olivacea.
MATERIALS AND METHODS
Animals. L. olivacea eggs were obtained from ‘‘LaEscobilla’’ beach (96° 278169W, 15° 408369N), Oaxaca,Mexico, on the night they were laid and transported toMexico City within 12 hr by ground transportation.This study used eggs from 12 clutches obtained be-tween 1991 and 1995. After arrival, eggs were placed incovered plastic trays filledwithmoistened vermiculite.Containers of eggs from each clutch were placed inincubators at either 276 0.5° (male-promoting tempera-ture) or 32 6 0.1° (female-promoting temperature) fordifferent experiments.Experiments on critical periods. Nine experiments
using a total of 10 clutches (100–110 eggs/clutch) wereperformed. Containers with 10 eggs each were trans-ferred at different days of incubation from male-promoting temperature to female-promoting tempera-ture or the converse, from female-promotingtemperature to male-promoting temperature, and keptthere until hatching. In each experimental series, around80 eggs were used and four to eight exchanges weremade at different times. Since most exchanges wererepeated one or more times in different experiments,
374 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.
eggs from several clutches were used for each ex-change.Controls: at least 20 eggs were left without transfer
in every experimental series to certify the stability ofincubation conditions for promoting 100% male orfemale hatchlings. One experimental series was dis-carded (95 eggs) where three females were found in theincubator set at male-promoting temperature. One ortwo embryos from each temperature were harvested atevery exchange, their gonadswere processed for histol-ogy, and the stage of development was establishedaccording to Miller’s criteria (1985).Estradiol (E2) treatment. Eggs from five clutches
were used for these experiments. Groups of 10–30 eggsincubated at 27 6 0.5° were treated once with 6 or 12µg of E2 diluted in 3 µl of oil and replaced into themale-promoting temperature. Simultaneously, 5 eggswere treated only with oil. Vehicles with or without E2were administered through the eggshell by droppingthe solution in a plastic ring sealed with paraffin andplacing it on the most vascularized region near theembryo. All groups were kept at male-promotingtemperature until hatching.Histology. Sex was determined by the microscopic
structure of the gonad. Samples were fixed withKarnovsky’s solution and embedded in Epon, as previ-ously described (Merchant-Larios et al., 1989). To com-pare the feminizing effects of E2 and temperature, amorphometric study was undertaken: cross sectionsfrom the medial region of the ovaries of hatchlingswere analyzed. Images obtained with a camera lucidawere outlined on a paper and the area of the cortex andmedulla were measured with a planimeter. Gonadsfrom 26 E2-treated embryos and 26 embryos incubated
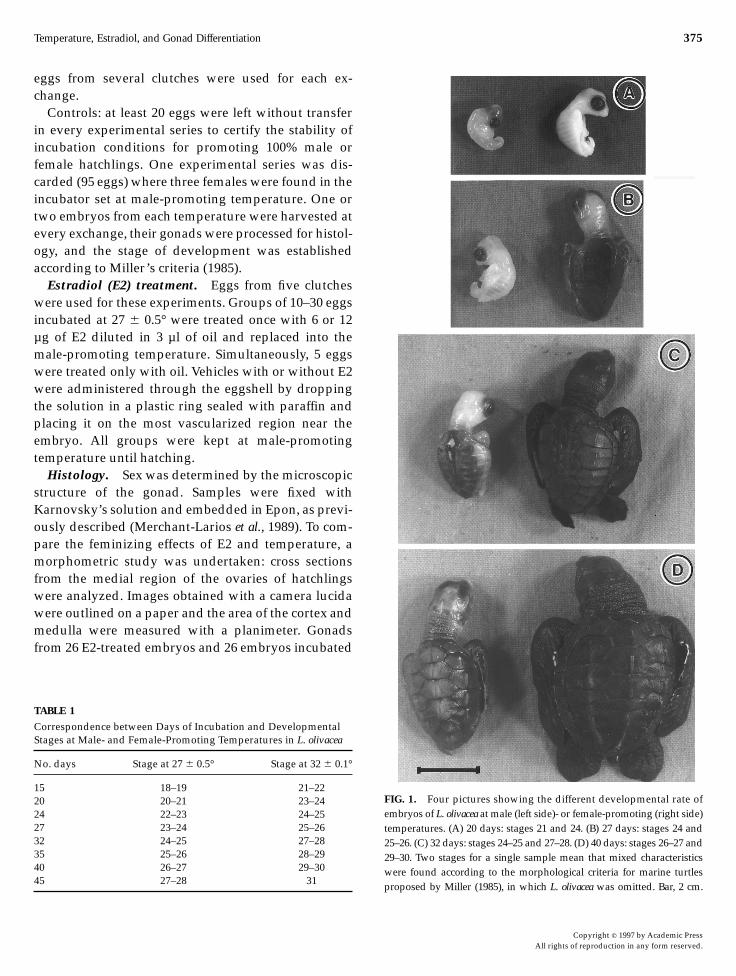
FIG. 1. Four pictures showing the different developmental rate ofembryos of L. olivacea atmale (left side)- or female-promoting (right side)temperatures. (A) 20 days: stages 21 and 24. (B) 27 days: stages 24 and25–26. (C) 32 days: stages 24–25 and 27–28. (D) 40 days: stages 26–27 and29–30. Two stages for a single sample mean that mixed characteristicswere found according to the morphological criteria for marine turtlesproposed by Miller (1985), in which L. olivacea was omitted. Bar, 2 cm.
TABLE 1
Correspondence between Days of Incubation and DevelopmentalStages at Male- and Female-Promoting Temperatures in L. olivacea
No. days Stage at 27 6 0.5° Stage at 32 6 0.1°
15 18–19 21–2220 20–21 23–2424 22–23 24–2527 23–24 25–2632 24–25 27–2835 25–26 28–2940 26–27 29–3045 27–28 31
Temperature, Estradiol, and Gonad Differentiation 375
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

at female-promoting temperature were analyzed.Means and standard deviations were calculated foreach experimental condition. Student’s t test wasapplied using ,0.05.
RESULTS
Embryos incubated at 32° developed faster thanembryos incubated at 27° (Table 1 and Fig. 1). After 20days of incubation, the embryos incubated at highertemperature (female-promoting) reached stages 23–24,whereas at lower temperature (male-promoting) theyreached stages 20–21, around two or three stagesbehind (Fig. 1A). After 27 days, the embryos incubatedat female-promoting temperature were at stages 25–26,whereas those at male-promoting temperature hadreached stages 23–24 (Fig. 1B). After 32 days at female-promoting temperature the embryos reached stages27–28, whereas at male-promoting temperatures theembryos were at stages 24–25 (Fig. 1C). Embryosincubated at female- or male-promoting temperaturesfor 40 days were at stages 29–30 or 26–27, respectively.Embryos developing at feminizing temperature hatchedaround Day 45, while those developed at masculiniz-ing temperature were still at stages 27–28. The latterhatched after around 63–65 days of incubation.In correlation with the faster development of whole
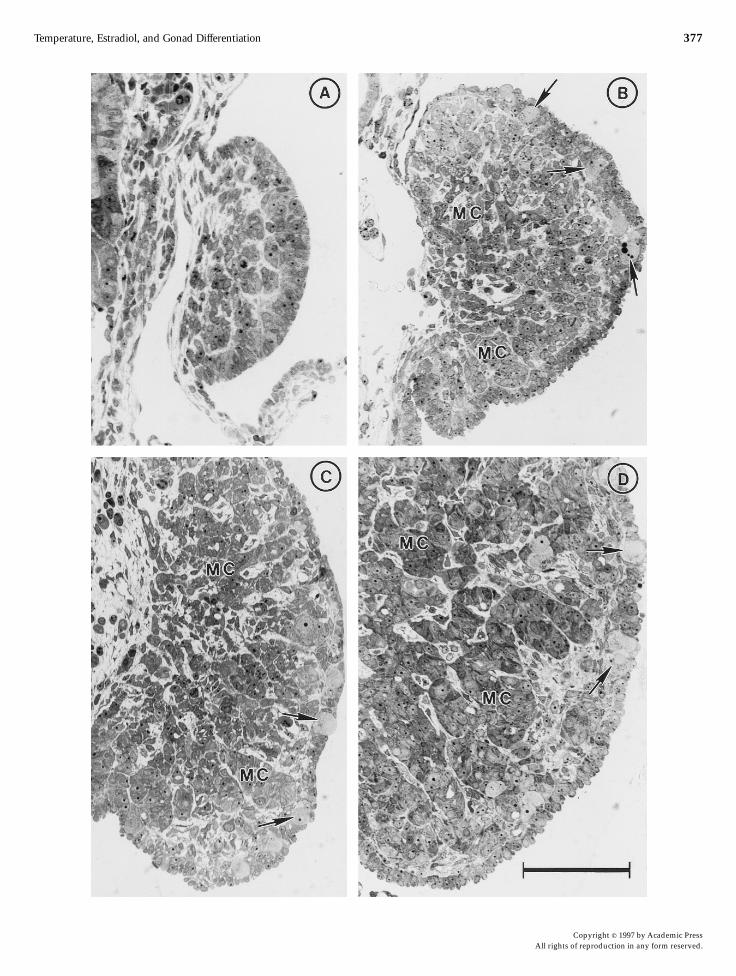
embryos at female-promoting temperature than atmale-promoting temperature, the gonads grew fasterat female-promoting temperature (Fig. 2). The histologi-cal aspect of the genital ridges and gonads at differentstageswas studied in cross sections taken from samplesat 20, 27, 32, and 40 days from different regions.Figures 2 and 3 show the histology of gonads fromembryos at incubation. The histological changes forthe establishment of an undifferentiated gonad weresimilar at both male- and female-promoting tempera-tures: at stages 21–23, proliferating cells became con-densed as clusters near the coelomic epithelium of the
genital ridge (Fig. 2A). Then, around stages 24–26,medullary sex cords were formed and remained con-tinuous with the surface epithelium in some regions(Figs. 2B–2D, 3A, and 3B). Histological sex differentia-tion of the gonads was evident at or after stages 26–27.Gonads from embryos incubated at male-promotingtemperature showed a thin surface epithelium andprominent medullary cords at stage 27 (Fig. 3C). Ingonads at female-promoting temperature, the surfaceepithelium became thick and contained most of thegerm cells, although at stage 27 the medullary cordswere still evident (Fig. 3B). Medullary cords appearedfragmented in female gonads at stage 29 (Fig. 3D).
Temperature Exchanges and Sex DeterminationPeriods
Table 2 summarizes the results of response to oneshift of temperature at different developmental stages.All embryos initially incubated at male-promotingtemperature up to stages 17–19 were feminized whentransferred to 32°C. Between stages 20 and 23 adecreasing number of embryos incubated at male-promoting temperature responded to the female-promoting temperature: 73.6, 57.5, and 47.3%, respec-tively, responded at stages 20, 21, and 22, whereas only7.2% were feminized when transferred at stage 23.From stage 24 onward all hatchlings from embryosinitially incubated at 27° and then shifted to 32°developed as males.In contrast, the response of embryos initially incu-
bated at female-promoting temperature covered moreadvanced stages than embryos incubated at male-promoting temperature: all hatchlings from embryosinitially incubated at female-promoting temperatureand transferred to male-promoting temperature atstage 23, or earlier, developed intomales.When shiftedat stages 24, 25, or 26, the percentage of embryos whichresponded to the male-promoting temperature was94.5, 62.3, and 3.1%, respectively. Finally, embryos
FIG. 2. Histology of the gonads of embryos incubated at male (Aand C)- or female (B and D)-promoting temperature.A–B and C–D show crosssections of gonads at 20 and 27 days of incubation, respectively. This period of around 7 days corresponds to the TSP. Although at thefemale-promoting temperature the gonads are larger (stages 24 and 26) than at the male-promoting temperature (stages 21 and 24), they appearmorphologically undifferentiated. Prominent medullary cords (MC) and several germ cells (arrows) in the coelomic epithelium are observed ingonads from embryos incubated at both temperatures. Bar, 100 µm.
376 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.
Temperature, Estradiol, and Gonad Differentiation 377
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

378 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

incubated at 32° up to stage 27 developed as femalesafter shift to male-promoting temperature.The different response to one shift of temperature
depending on temperature of incubation (male- orfemale-promoting temperatures) led us to investigatethe correlation between stages of development and
days of incubation. As shown in Fig. 4, although thethermosensitive stages appeared different, the thermo-sensitive periods (temporal dimension) were similar.During the first 20 days of incubation, all embryosresponded to the shifted temperature regardless of thestage of development reached during this period.Thereafter, during the next 7 days, a decreasing num-
FIG. 3. Aand B are gonads fixed at 32 days at male- or female-promoting temperatures, respectively. They were from embryos at stages 25 (A)and 27 (B) corresponding to the SDP. At female-promoting temperature (B) the gonad has initiated medullary cord fragmentation (stars); a thickcortex with abundant germ cells (arrow) indicates the start of ovarian differentiation. At male-promoting temperature (A), the gonad still has anundifferentiated aspect with medullary cords (MC) and several germ cells (arrows) in the coelomic epithelium. Pictures C and D show gonadsfixed after 40 days of incubation at stages 27 and 29 at male- or female-promoting temperature, respectively. The testis (C) shows a thin cortexand conspicuous medullary cords (MC) surrounded by stromal tissue (S). Picture D shows an ovary with a thick cortex (CX) and remnants ofmedullary cords (stars). Bar, 100 µm.
TABLE 2
Critical Periods (C.P.) for Sex Determination of L. olivacea
StageaNo.eggsb
No.hatchlings
No.males
No.females
%Responsec C.P.
Eggs shifted from 27 6 0.5° to 32 6 0.1°17 20 (2) 11 0 11 100 UP18 20 (2) 17 0 17 100 UP19 20 (2) 11 0 11 100 UP20 30 (3) 19 5 14 73.6 TSP21 50 (5) 33 14 19 57.5 TSP22 80 (4) 76 40 36 47.3 TSP23 100 (5) 97 90 7 7.2 TSP24 40 (4) 34 34 0 0 SDP25 10 (1) 8 8 0 0 SDP26 20 (2) 17 17 0 0 SDP27 10 (1) 9 9 0 0 SDP
StageNo.
EggsbNo.
HatchlingsNo.Males
No.Females
%Responsea C.P.
Eggs shifted from 32 6 0.1° to 27 6 0.5°17 20 (2) 19 19 0 100 UP18 20 (2) 11 11 0 100 UP19 20 (2) 11 11 0 100 UP20 40 (4) 18 18 0 100 UP21 20 (2) 9 9 0 100 UP22 30 (3) 18 18 0 100 UP23 20 (2) 18 18 0 100 UP24 100 (5) 74 70 4 94.5 TSP25 100 (5) 85 53 32 62.3 TSP26 40 (4) 32 1 31 3.1 TSP27 30 (3) 22 0 22 0 SDP
a According with Miller’s criteria (1985). UP, undeterminedperiod. TSP, temperature-sensitive period. SDP, sex-determinedperiod.
b Number of clutches.c Gonadal sex was determined by histology a few days after
hatching.
FIG. 4. Correlation among sex determination, days of incubation,and stage of development at female- or male-promoting tempera-tures. During the first 20 days of incubation, all embryos respondedto the shifted temperature and it was named the undeterminedperiod (UP). Thereafter, at around 7 days, a decreasing number ofembryos responded to the shifted temperature; this is the thermosen-sitive period (TSP). Afterward, none of the embryos responded tothe shifted temperature and this was considered the sex-determinedperiod (SDP). While at the female-promoting temperature(32 6 0.1°C) the TSP was from stages 23–24 to stages 26–27, at themale-promoting temperature (27 6 0.5°C) the TSP was from stages20–21 to stages 23–24. Moreover, the SDP was initiated at stages27–28 or 24–25 at female- or male-promoting temperature, respec-tively.
Temperature, Estradiol, and Gonad Differentiation 379
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

ber of embryos responded to the shifted temperatureat different stages of development: embryos at male-promoting temperature during these 7 days were atstages 20–23, whereas embryos at female-promotingtemperaturewere at stages 24–26. Finally, after approxi-mately 27 days of incubation, all embryos from bothmale- and female-promoting temperatures were unre-sponsive to the shifted temperature and maintainedthe sex expected with the initial temperature.Assuming that the percentage of embryos respon-
sive to the shifted temperature corresponded to em-bryos where sex was not determined by the initialtemperature, the embryonic development was dividedinto three periods: undetermined (UP), thermosensi-tive (TSP), and sex-determined (SDP) periods (Table 2and Fig. 4). During TSP, the embryos incubated atmale- or female-promoting temperatures were at stages20–23 or 24–26, respectively. At these stages the gonadswere histologically undifferentiated (Fig. 2). The onsetof SDP was around stage 24 for embryos incubated atmale-promoting temperature and around stage 27 forembryos incubated at female-promoting temperature.Although at the female-promoting temperature thehistological differentiation of the female gonad wasdetectable, at male-promoting temperature, the go-nads were still undifferentiated (Figs. 3A and 3B).
Effect of Estradiol (E2) at Different DevelopmentalStages
Embryos incubated at male-promoting temperatureup to stages 24–25 were feminized by E2 treatment.Although some embryos may still respond to exog-enous E2 at stage 26, they did not respond after thisstage (Table 3). The histological study of E2-treatedgonads revealed a reduction or complete disappear-ance of the medullary cords and a thick cortex, whichsuggest ovarian differentiation (Fig. 5). The morpho-metric analysis showed, however, that ovaries fromE2-treated hatchlings were significantly smaller thanovaries from hatchlings incubated at female-promot-ing temperature (Fig. 6).
DISCUSSION
There are some discrepancies between the resultsobtained in the present work and those published
previously (Merchant-Larios et al., 1989; Merchant-Larios and Villalpando, 1990). There is no correspon-dence between the developmental stages since inprevious studies we used the developmental stagesproposed by Cratz (1982) for L. olivacea, which aredifferent from Miller’s stages (Miller, 1985) used in thepresent study. Our conclusion that the female gonaddifferentiates earlier than themale gonadwas based onthe presence of germ cells on the surface epithelium inmale gonads up to hatching, whereas medullary cordsare fragmented in female gonads before hatching.However, in the current study, which uses far moresamples from embryos incubated under more stableconditions, it was found that histological differentia-tion occurs at stages 26–27 at both male- and female-promoting temperatures. The interesting finding inour recent study was that sex determination at male-promoting temperature occurs at stage 24, before thehistological differentiation of the gonad.It is known that in reptiles with TSD, temperature
changes affect sex determination only during a certainperiod of incubation. Mrosovsky and Pieau (1991)defined the thermosensitive period (or thermosensi-tive stages) as ‘‘that interval when variation of tempera-ture (any kind of temperature variation) has beenshown to have some significant effect (any kind ofeffect) on sex ratio.’’ Apart from the interspecificvariations that may exist, the sensitive periods alsovary with the experimental procedures. Nevertheless,there is agreement that reptiles with TSD have tempera-ture-sensitive periods placed around the middle one-
TABLE 3
Effect of Estradiol (6 µg/egg) Applied to Embryos at DifferentStages of Development in L. olivacea Incubated at 27 6 0.5°
Stage No. eggsa No. males No. females No. ovotestis
20 20 (2) 1 14 221 10 (1) 0 4 022 10 (1) 0 8 223 20 (2) 0 17 023–24 10 (1) 0 10 024 30 (2) 0 20 225 10 (1) 0 9 025–26 10 (1) 3 5 026 20 (2) 14 0 027 10 (1) 6 0 028 10 (1) 5 0 0
a Number of clutches.
380 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

third to one-half of embryonic development (Yntema,1979; Bull and Vogt, 1981; Pieau and Dorizzi, 1981;Yntema and Mrosovsky, 1982; Ferguson and Joanen,1983; Bull, 1987; Webb et al., 1987; Wibbels et al., 1991).Therefore, clearly, there are two periods not sensitiveto temperature: at the beginning and at the end ofembryonic development.During the first 20 days of incubation, all L. olivacea
embryos responded to one shift of temperature (male-to female-promoting temperatures or vice versa). Thus,during the first days of incubation, sex was not yetdetermined and all embryos responded to one shift oftemperature when they were left at the new tempera-ture until hatching. Double shifts of temperature dur-ing this period when all embryos were undetermineddid not modify the sex promoted by the backgroundtemperature. Thereafter, sex began to be determined insome embryos and gradually fewer embryos of theexperimental group responded to the shifted tempera-ture. Finally, during the last days of incubation, allembryos were determined. That is, they were insensi-tive to the shifted temperature.The present results suggest that the physiological
basis for the lack of sensitivity to temperature shiftsduring the first and the last days of embryonic develop-ment are different. In double shift experiments, ‘‘insen-sitivity’’ during the first days of development refers tothe period when the embryos were sexually undeter-mined. In contrast, at late developmental stages, ‘‘insen-sitivity’’ to temperature is due to the fact that theembryos were sexually determined.
We divided the development of L. olivacea embryosinto three periods: UP, TSP, and SDP. It must beemphasized that these periods refer to groups and notto individual embryos. Thus, the gradual sex determi-nation of a series of embryos during TSP suggests thatthe embryos became determined at different stages,probably due to genotypic variations. If sex determina-tion is defined as ‘‘the establishment of an irreversiblemolecular process which leads to male or femaledifferentiation,’’ it can be postulated that TSP refers tothe time required to set this process in a series ofdeveloping embryos of a species. Developmental‘‘time,’’ however, has two implications: the temporaldimension and the stage of development. Since thelatter are expressed in terms of morphology, underly-ing molecular events may follow different rates.Most studies on thermosensitive periods are ex-
pressed in terms of thermosensitive stages of develop-ment (Pieau an Dorizzi, 1981; Yntema and Mrosovsky,1982; Bull and Vogt, 1981; Wibbels et al., 1991; Harryand Williams, 1991). In the present study, we foundthat the thermosensitive stages in L. olivacea, arebetween stages 21–22 and stages 26–27. These stagesare similar to the thermosensitive stages reported inCaretta caretta, another sea turtle (Yntema and Mros-ovsky, 1982; Harry and Williams, 1991). Our presentresults, however, suggest that in L. olivacea, the thermo-sensitive stages are different at male- and female-promoting temperatures. Embryos incubated at male-
FIG. 5. Small ovary from a hatchling treated in ovo with 12 µg ofestradiol on Day 20 of incubation at male-promoting temperature. Athick cortex with several germ cells (GC) and remanents of medul-lary cords (MC) can be seen. Bar, 100 µm.
FIG. 6. Morphometric study of gonads from hatchlings incubatedat male-promoting temperature 16 or 12 µg of estradiol (E2) andfrom hatchlings incubated at female-promoting temperature.
Temperature, Estradiol, and Gonad Differentiation 381
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

promoting temperature start sex determination at stages21–22, whereas embryos incubated at female-promot-ing temperature begin to be determined around stage24. Moreover, embryos initially incubated at male-promoting temperature did not respond to female-promoting temperature around stage 24, whereas em-bryos initially incubated at female-promotingtemperature did not respond later, around stage 27.Results suggest that sexual fate at the male-
promoting temperature became irreversible in all em-bryos two or three stages earlier than at the female-promoting temperature. Although less dramatic (oneor two stages earlier), similar findings have beenreported in two freshwater turtles, Emys orbicularis andGraptemys ouchitensis (Pieau and Dorizzi, 1981; Bulland Vogt, 1981). However, in Eublepharis maculariusand Trachemys scripta it was reported that the sexdetermination at female-promoting temperature be-comes irreversible prior to that at male-promotingtemperature (Bull, 1987; Wibbels et al., 1991). Althoughdifferences in the chronology of male and female sexdetermination may represent interspecific variation,they could also be related to the sex-determiningpotencies of the specific temperatures used in eachstudy (Wibbels et al., 1991).According with the current experimental proce-
dures, in L. olivacea, TSPwas placed aroundDays 20–27 ofincubation.During this period (temporal dimension), indi-vidual embryosmay reach the threshold of an irreversiblemolecular process necessary for sex determination. Ex-periments of Lang and Andrews (1994) in A. mississip-piensis involving shifts of 33° to 31° and vice versafound that both the male-promoting temperature andthe female-promoting temperature were equally effec-tive over the same length of time (around 20 days).Interestingly, in A. mississippiensis, sex is determined
one to two stages earlier at female-promoting tempera-ture than at male-promoting temperature. Although intheAmerican alligator (A.mississippiensis) low tempera-ture is female-promoting, whereas in the olive ridley(L. olivacea) it is male-promoting, in both species, sexdetermination occurs earlier in low-growing embryosat lower temperature than in fast-developing embryosat higher temperature. An interaction between geno-type and environmental temperature may explain thedifferent outcomes: asynchronous development of
whole embryos at male- or female-promoting tempera-tures may trigger signals for either male or femaledetermination depending on the species genotype(Mittwoch, 1992).Of particular interest, present results showed that
some embryos of L. olivacea start sex determinationaround Day 20 and all became determined around Day27 of incubation. Differences in the rate of embryonicdevelopment at male- or female-promoting tempera-tures were also reflected in differences between sexdetermination and the histological differentiation ofthe gonads at both temperatures. Whereas at female-promoting temperature, sex determination occurredwhen ovaries had already begun their histologicaldifferentiation (around stage 26), at male-promotingtemperature, sex determination occurred early, whenthe gonads were still undifferentiated (around stage24). This at first appears counterintuitive, as the growthand metabolic rate is clearly greater at the warmertemperature. A possible explanation is that the malesex is the default state and that the female conditionmust be imposed upon it. The present results showedthat stages 24–25 mark the end or the onset of TSD atmale- or female-promoting temperature. Therefore, upto this stage an irreversible male-determining mecha-nism may be set in 100% of embryos at male-promotingtemperature, whereas in embryos incubated at female-promoting temperature, the female-determiningmech-anism may still be switched off.The lack of direct correlation between sex determina-
tion and histological differentiation of the gonad wasalso reported in A. mississippiensis. In this species, thethermosensitive period (shifts from 34° to 30° and viceversa, Ferguson and Joanen, 1983) occurs during go-nadal ridge stages and ends well in advance of thesexual differentiation of the gonads (Deeming andFerguson, 1988; Joss, 1989).Experiments on several reptiles with TSD have
shown that estrogens can overcome the effect ofmale-promoting temperatures and produce femalehatchlings. Furthermore, the use of compounds thatdisturb steroidogenesis, including aromatase inhibi-tors, has led to the idea that estrogen and temperatureact via a common physiological pathway (see reviews:Raynaud and Pieau, 1985; Crews et al., 1994; Dorizzi etal., 1996). Current results demonstrate that in L. oliva-
382 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

cea, as in other species, exogenous estradiol also‘‘feminized’’ the gonads of hatchlings incubated atmale-promoting temperature. Thus, exogenous estra-diol inhibited the development of medullary testicularcords, whereas the gonadal cortex was stimulated.Experiments in Emys orbicularis have shown that aro-matase activity is important for maintenance of differ-entiated ovaries after TSP (Dorizzi et al., 1996).Although in various species the sensitive stages to
exogenous estradiol correspond to the thermosensitiveperiod (Gutzke and Chymiy, 1988; Wibbels et al.,1991b; Dorizzi et al., 1991), we found that, in L. olivacea,the period of estradiol sensitivity was longer than thethermosensitive period from male- to female-promot-ing temperature. This may be explained by a doseeffect of the exogenous estradiol: at female-promotingtemperature, estradiol synthesis may be regulated at acritically lower concentration required for normal dif-ferentiation of the gonad than the concentrationsemployed in the present experiments. In fact, it wasfound that in the estradiol-treated embryos, the ‘‘femi-nized’’ gonads were considerably smaller than ovariesinduced by temperature. Abnormally small ovariesfrom estradiol-treated embryos, however, seemed torecover after hatching. It was found that in hatchlingsfrom estradiol-treated eggs, morphologically normalovaries developed sixmonths after hatching (Merchant-Larios et al., unpublished observations).It has been postulated that estrogen synthesis and,
therefore, aromatase activity may play a key role in sexdetermination of thermosensitive species (Pieau et al.,1994) but the actual mechanism remains to be eluci-dated. Species variations and different experimentalprocedures have complicated the finding of a commonmechanism if it exists at all. In Dermochelys coriacea,another sea turtle, aromatase activity was found to behigher at 30.5° (female-promoting temperature) than at27° (male-promoting temperature). When eggs wereshifted from 27 to 35° at different embryonic stages andexposed to 35° for 6 days, an increase in aromataseactivity was seen only between stages 23 and 27(Desvages et al., 1993). We found that in L. olivacea,embryos incubated at female-promoting temperatureare determined between stages 23–24 and 26–27. It willbe interesting to study if in this species gonadalaromatase activity is induced at female-promotingtemperature.
Collectively, interactions between temperature andgenetic factors may exist for sex determination in allTSD species. If enhanced aromatase activity is theinitial signal for ovary differentiation, at least twopossibilities can be postulated: (i) the enzyme aro-matase directly responds to temperature, and theestrogen produced exerts a positive feedback controlon the aromatase gene (Pieau, 1996) and (ii) a thermo-sensitive process (gene?) located upstream of thearomatase gene controls the female developmentalpathway, including the enhanced expression of thearomatase gene.The present findings that at male-promoting low
temperature sex determination occurs before that atfemale-promoting high temperature suggest that en-dogenous estrogens produced at female-promotinghigh temperature may prevent the expression of amale-determining gene in L. olivacea. Lack or lowconcentration of estrogen may allow accumulation of amale-determining gene product. On the other hand, inA. mississippiensis, exogenous E2 feminizes embryos atmale-promoting high temperature (Lang and An-drews, 1994), suggesting that, in this species, endog-enous estrogens are produced at female-promotinglow temperature. Thus, if feminizing estrogen is pro-duced at different temperatures (and/or developingrates), genetic predisposition may underlie differentmechanisms controlling aromatase activity.Moreover, since sex determination and morphologi-
cal differentiation of the gonad appear as dissociatedevents, female-promoting high temperature may en-hance estrogen production, which may prevent testisdifferentiation and lead the undifferentiated gonad toform an ovary.
ACKNOWLEDGMENTS
The Secretarıa de Pesca and Instituto Mexicano de Pesca gavepermission to collect eggs in La Escobilla, Oaxaca, a Mexican faunalpreserve.We thankMr. Jose G. Baltazar for technical assistance in thelaboratory and Biol. J. Vasconcelos, Director of the Centro Mexicanode la Tortuga, Mazunte, Oax., and Biol. Cuahutemoc Villanueva forassistance in the field work. Mrs. Isabel Perez-Montfort helped withthe English. This study was partially supported by the MexicanCONACYT (4037-N) and PAPIID-UNAM:IN209694.
Temperature, Estradiol, and Gonad Differentiation 383
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

REFERENCES
Bull, J. J. (1987). Temperature-dependent sex determination inreptiles: validity of sex diagnosis in hatchling lizards. Can. J. Zool.65, 1421–1424.
Bull, J. J., Gutzque, W. H. N., and Crews, D. (1988). Sex reversal byestradiol in three reptilian orders. Gen. Comp. Endocrinol. 70,425–428.
Bull, J. J., and Vogt, R. C. (1981). Temperature-sensitive periods of sexdetermination in emydid turtle. J. Exp. Zool. 218, 435–440.
Cratz, F., (1982). Embryological stages of the marine turtle Lepido-chelys olivacea (Escholtz). Rev. Biol. Trop. 30, 113–120.
Crews, D., Bull, J. J., andWibbels, T. (1991). Estrogen and sex reversalin turtles: A dose-dependent phenomenon. Gen. Comp. Endocrinol.81, 357–364.
Crews, D., and Bergeron, J. M. (1994). Role of reductase andaromatase in sex determination in the red-eared slider (Trachemysscripta), a turtle with temperature-dependent sex determination. J.Endocrinol. 143, 279–289.
Crews, D., Bergeron, J. M., Bull, J. J., Flores, D., Tousignant, A.,Skiper, J. K., and Wibbels, T. (1994). Temperature-dependent sexdetermination in reptiles: proximate mechanisms, ultimate out-comes, and practical applications.Dev. Gent. 15, 297–312.
Dalrymple, G. H., Hampp, J. C., and Wellins, D. J. (1985). Male-biased sex ratio in a cold nest of a hawksbill sea turtle Eretmochelysimbricata. J. Herpetol. 19, 1219–1229.
Desvages, G., Girondot, M., and Pieau, C. (1993). Sensitive stages forthe effects of temperature on gonadal aromatase activity inembryos of the marine turtle Dermochelys coriacea. Gen. Comp.Endocrinol. 92, 54–61.
Deeming, D. C., and Ferguson, M. W. J. (1988). Enviromentalregulation of sex determination in reptiles. Philos. Trans. R. Soc.London Ser. B 322, 19–39.
Dorizzi, M., Mignot, T. M., Guichard, A., Desvages, G., and Pieau, C.(1991). Involvement of oestrogens in sexual differentiation ofgonads as a function of temperature in turtles. Differentiation. 47,9–17.
Dorizzi, M., Richard-Mercier, N., Desvages, G., Girondot, M., andPieau, C. (1994). Masculinization of gonads by aromatase inhibi-tors in a turtle with temperature-dependent sex determination.Differentiation. 58, 1–8.
Dorizzi, M., Richard-Mercier, N., and Pieau, C. (1996). The ovaryretains male potential after the thermosensitive period for sexdetermination in the turtle Emys orbicularis. Differentiation 60,193–201.
Ferguson, M. W. J., and Joanen, T. (1983). Temperature dependentsex determination in Alligator mississippiensis. J. Zool. 200, 143–177.
Gutzke, W. H. N., and Chymiy, D. B. (1988). Sensitive periods duringembryogeny for hormonally induced sex determination in turtles.Gen. Comp. Endocrinol. 71, 265–267.
Harry, J. L., and Williams, K. L. (1991). Differential growth of maleand female urogenital systems of Caretta carreta, within thesex-determining period. J. Exp. Zool. 258: 204–211.
Joss, J. M. P. (1989). Gonadal development and differentiation inAlligator mississippiensis at male and female producing incubationtemperatures. J. Zool. 218, 679–687.
Lance, V. A., and Bogart, M. H. (1992). Disruption of ovariandevelopment in alligator embryos trated with an aromataseinhibitor. Gen Comp. Endocrinol. 86, 59–71.
Lang, J. F., and Andrews, H. V. (1994). Temperature-dependent sexdetermination in crocodilians. J. Exp. Zool. 270, 28–44.
Merchant-Larios, H., Villalpando-Fierro, I., and Centeno-Urruiza, B.(1989). Gonadal morphogenesis under controlled temperature insea turtle Lepidochelys olivacea. Herpetol. Monogr. 3, 43–61.
Merchant-Larios, H., and Villalpando-Fierro, I. (1990). Effect oftemperature on gonadal sex differentiation in the sea turtleLepidochelys olivacea: An organ culture study. J. Exp. Zool. 254,327–331.
Miller, J. D., and Limpus, C. J. (1980). Incubation period and sexualdifferentiation in the green turtle, Chelonia mydas L. In ‘‘ZoologicalBoard of Victoria’’ (C. B. Banks and A. A. Martin, Eds), pp. 66–73,Proc. Melbourne Herp. Symp., Victoria.
Miller, J. D. (1985). Embryology of marine turtles. In ‘‘Biology ofReptilia’’ (C. Gans, F. Billet, and P. F. A. Maderson, Eds.), Vol. 14,pp. 270–328. Wiley, New York.
Mittwoch, U. (1992). Sex determination and sex reversal: Genotype,phenotype, dogma and semantics.Hum. Genet. 89, 467–479.
Mrosovsky, N., and Pieau, C. (1991). Transitional range of tempera-ture, pivotal temperatures and thermosensitive stages for sexdetermination in reptiles. Amphibia-Reptilia 12, 169–179.
Morreale, S. J., Ruiz, G. J., Spotila, J. R., and Standora, E. A. (1982).Temperature-dependent sex determination: Current practicesthreaten conservation of sea-turtles. Science 216, 1245–1247.
Pieau, C., and Dorizzi, M. (1981). Determination of temperaturesensitive stages for sexual differentiation of the gonads in embryosof the turtle, Emys orbicularis. J. Morphol. 170, 373–382.
Pieau, C., Girondot, M., Richard-Mercier, N., Desvajes, G., Dorizzi,M., and Zaborski, P. (1994). Temperature sensitivity of sexualdifferentiation of gonads in the European Pord turtle: Hormonalinvolvement. J. Exp. Zool. 270, 86–94.
Pieau, C. (1996). Temperature variation and sex determination inreptiles. BioEssays 18, 19–26.
Raynaud, A., and Pieau, C. (1985). Embryonic development of thegenital system. In ‘‘Biology of the Reptilia’’ (S. Gans and F. Billeu,Eds.), pp. 149–299. Wiley, New York.
Rhen, T., and Lang, J.W. (1994). Temperature-dependent sex determi-nation in the snapping turtle: Manipulation of the embryonic sexsteroid environment. Gen. Comp. Endocrinol. 96, 243–254.
Rimblot, F., Fretey, J., Mrosovsky, J., Lescure, J., and Pieau, C. (1985).Sexual differentiation as a function of incubation temperature ofeggs in the sea turtleDermochelys coriacea (Vandelli, 1761).Amphibia-Reptilia 6, 83–92.
Webb, G. J. W., Beal, A. M., Manolis, S. C., and Dempsey, K. E. (1987).The effects of incubation temperature on sex determination andembryonic development rate in Crocodylus johnstoni and C. poro-sus. In: Wildlife Management: Crocodiles and Alligators (G. J. W.Webb, S. C. Manolis, and P. J. Whitehead, Eds.), pp. 507–531.Surrey Beatty and Sons, Winnellie.
384 Merchant-Larios et al.
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

Wibbels, T., Bull, J. J., and Crews, D. (1991a). Chronology andmorphology of temperature-dependent sex determination. J. Exp.Zool. 260, 371–381.
Wibbels, T., Bull, J. J., and Crews, D. (1991b). Synergism of tempera-ture and estradiol: a common pathway in sex determination. J.Exp. Zool. 260, 130–134.
Wibbels, T., Gideon, P., Bull, J. J., and Crews, D. (1993). Estrogen andtemperature-induced medullary cord regression during gonadaldifferentiation in a turtle.Differentiation 53, 149–154.
Wibbels, T., and Crews, D. (1994). Putative aromatase inhibitorinduces male sex determination in a female unisexual lizard and
in a turtle with temperature-dependent sex determination. J.Endocrinol. 141, 295–299.
Yntema, C. L. (1979). Temperature levels and periods of sex determi-nation during incubation of eggs of Chelydra serpentina. J. Morphol.159, 17–28.
Yntema, C. L., and Mrosovsky, N. (1980). Sexual differentiation inhatchling loggerheads (Caretta caretta) incubated at different con-trolled temperatures.Herpetologica 36, 33–36.
Yntema, C. L., andMrosovsky, N. (1982). Critical periods and pivotaltemperatures for sexual differentiation in logger-head sea-turtles.Can. J. Zool. 60, 1012–1016.
Temperature, Estradiol, and Gonad Differentiation 385
Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.




















