
This PDF is the Sample PDF taken from our Comprehensive Study ...
Upload
independentCategory
view
0download
0
1
1
COPPER IS TAKEN UP EFFICIENTLY FROM ALBUMIN AND ALPHA-2-
MACROGLOBULIN BY CULTURED HUMAN CELLS BY MORE THAN ONE
MECHANISM
Mizue Moriya1, Yi-Hsuan Ho
1, Anne Grana
1, Linh Nguyen
1, Arrissa Alvarez
1, Rita Jamil
1, M. Leigh Ackland
2,
Agnes Michalczyk2, Pia Hamer
1, Danny Ramos
1, Stephen Kim
1, Julian F.B. Mercer
2, and Maria C. Linder
1
1Department of Chemistry and Biochemistry and Institute for Molecular Biology and Nutrition, California State
University, Fullerton, CA 92834-6866, U.S.A., and 2Centre for Cellular and Molecular Biology, Deakin
University, Burwood, Victoria 3125, Australia
Running head: Cu uptake from plasma proteins by hepatic and mammary cells
Contact information (To whom correspondence should be addressed):
Maria C. Linder
Department of Chemistry and Biochemistry
Fullerton, CA 92834-6866
Tel. 714-278-3621
FAX 714-278-5316
Email: [email protected]
Articles in PresS. Am J Physiol Cell Physiol (June 25, 2008). doi:10.1152/ajpcell.00029.2008
Copyright © 2008 by the American Physiological Society.

2
2
ABSTRACT
Ionic copper entering the blood plasma binds tightly to albumin and the macroglobulin, transcuprein.
It then goes primarily to the liver and kidney, except in lactation where a large portion goes directly to the
mammary gland. Little is known about how this copper is taken up from these plasma proteins. To examine
this, the kinetics of uptake from purified human albumin and 2-macroglobulin, and effects of inhibitors, were
measured using human hepatic (HepG2) and mammary epithelial (PMC42) cell lines. At physiological
concentrations (3-6 M) both cell types took up copper from these proteins independently and at rates similar
to each other and to those for Cu-di-histidine (His) or Cu-nitrilotriacetate (NTA). Uptakes from 2-
macroglobulin indicated a single saturable system in each cell type, but with different kinetics, and 65-80%
inhibition by Ag(I) in the HepG2 but not the PMC42 cells. Uptake kinetics for Cu-albumin were more complex
and also differed with cell type (as was the case for Cu-histidine and NTA); and there was little or no
inhibition by Ag(I). High Fe(II) concentrations (100-500 M) inhibited copper uptake from albumin 20-30% in
both cell types, that from 2-macroglobulin 0-30% and with no inhibition of the latter by Mn(II) or Zn(II). We
conclude that the proteins mainly responsible for the plasma exchangeable copper pool deliver the metal to
mammalian cells efficiently and by several different mechanisms. 2-Macroglobulin delivers it primarily to
CTR1 in hepatic but not mammary epithelial cells; and additional as yet unidentified copper transporters or
systems for uptake from these proteins remain to be identified.
Key words: Transcuprein, uptake kinetics, iron competition, silver competition, HepG2 cells, PMC42 cells

3
3
INTRODUCTION
In normal humans and animals, most of the copper entering the blood in ionic form (such as after
absorption from the intestine) goes directly to the liver and kidney, before re-appearing in the blood plasma in
newly-formed ceruloplasmin (35-39,58). This has been shown repeatedly in studies of rodents and other
mammals, using radioactive and heavy isotopes of copper. However, as first shown in rats with trace
amounts of radioactive copper (16) and now also corroborated in mice (Kenny Chu, Eric Sauble, Maria C.
Linder, unpublished results), this is not the case in lactating animals. Here, a large proportion of the copper
entering the blood is diverted from the liver and kidney to the mammary gland, where it rapidly incorporates
into the milk and milk ceruloplasmin. In preparation for studying what controls these different patterns of
copper distribution among organs, we have used cultured hepatic and mammary epithelial cell models to
examine how copper is taken up from the blood plasma proteins to which it is firmly bound, and to obtain
clues that will help us identify the receptors and/or transporters involved of which expression and/or activity
are regulated by lactational hormones.
Little is actually known about how copper is delivered to and enters mammalian cells, including those
of the liver and mammary epithelium. This is despite numerous studies with various cell lines in culture. In
normal (non-lactating) rats and humans, we know that ionic copper entering the blood becomes part of an
“exchangeable copper pool” in the plasma (35-39,58). This pool does not include ceruloplasmin, as
ceruloplasmin copper is buried in the structure and not dialyzable at neutral pH. Rather, it is composed
primarily or exclusively of copper bound to high affinity sites on albumin and a macroglobulin we named
transcuprein (4,35-39,44,68). In the case of the rat, the macroglobulin is 1-inhibitor-3 (21,44); in the case of
the human, it is 2-macroglobulin (44), which shares a very similar, histidine and cysteine-rich region. There
is no solid evidence that any of the exchangeable copper is bound to amino acids, except perhaps as a
ternary albumin-copper-his complex (31,35,39). Indeed, as calculated by Rae et al (56), there are virtually
no “free” copper ions in biological systems. Copper on transcuprein and albumin is rapidly exchanged
between them, and in vitro is released only very slowly to various buffers, including those containing high
concentrations of histidine (37,38). This implies that for cell uptake, the copper is being handed from one (or
both) of these plasma proteins directly to transporters in the cell membrane, and is not first released to amino
acids (or present as the free ion) before binding to a transporter or transport system.

4
4
It has become evident that this phenomenon of “handing” copper from protein to protein is what
occurs within cells as well, where a series of copper chaperones (proteins that transport incoming copper to
specific intracellular sites/proteins) have been identified (23,25,27,52-54). Thus, present evidence strongly
implies that protein-bound copper in the blood plasma, attached to albumin and transcuprein, is the direct
source of copper entering cells of the liver and kidney after intestinal absorption or after it otherwise enters
the blood in ionic form. Our previous studies with rats indicate that this is also the case with mammary
epithelial cells during lactation (16). Several questions thus arise. The first is whether
transcuprein/macroglobulin and/or albumin directly deliver copper to hepatic and mammary epithelial cells,
and whether other plasma factors are required. If both proteins deliver copper, then the question is whether
they interact with the same transport system in the cell membrane. The ultimate questions are of course the
identity of the transporter or transport mechanisms involved, and details about the mechanism(s) of entry.
As concerns albumin, the results of several studies suggested that it may actually impede rather than
facilitate copper uptake. The groups of Ettinger and McArdle showed with cultured rat and mouse
hepatocytes and fibroblasts that the presence of albumin markedly lowered initial rates of copper uptake
supplied as Cu(II) with or without histidine (15,48,49,64). In addition, we showed that copper uptake by the
liver was not dependent upon the presence of albumin (63). Thus in analbuminemic rats, uptake of i.p.
injected tracer copper was (if anything) accelerated (63). Moreover, receptors for albumin have not been
detected on hepatic cells. Thus, the function of copper binding to albumin in the blood plasma may not be
for it to deliver the metal to cells. Rather, it may have a protective role in the plasma, allowing safe
sequestration of excess copper ions entering the blood. [Because of its abundance in the plasma, albumin is
capable of binding a great deal of copper and will do so when excessive amounts are administered (12).]
Transcuprein is a macroglobulin (44) and so an obvious possibility is that copper bound to this
protein enters cells via receptor-mediated endocytosis, using the macroglobulin receptor (8,59).
Macroglobulins are well known for their ability to act as “traps” for proteases, clearing them from the plasma.
Hepatocytes have macroglobulin receptors, and these cells are thought to be the main removal route for
macroglobulin complexes (59). Thus, for example, almost all 125
I-labeled PSA- 2-macroglobulin injected i.v.
into rats went directly to the liver (9), with small amounts also going to the kidney and spleen, the same initial
distribution pattern that occurs with radioactive copper tracer (68). However, other cells also have these

5
5
receptors, including fibroblasts (8), macrophages (30), retinal pigment epithelial cells (24), and those of the
mammary epithelium (6).
Alternatively, or in addition, transcuprein and/or albumin may deliver copper to specific transporters
on the cell membrane. The obvious possibilities are copper transporter 1 (CTR1) and divalent metal
transporter 1 (DMT1, also known as Nramp 2 and DCT1). CTR1 is ubiquitous (52,74) but expressed more
highly in liver and kidney than in most other tissues (28). However, CTR1 may not be the only transporter
involved (33). DMT1 (also expressed in most tissues) does appear to play a role in uptake of ionic copper
across the intestinal brush border (3,41). Then there is the hepatocyte (and fibroblast) copper transporter
studied by Ettinger and McArdle and their collaborators (17,35,48), using CuCl2 or the di-histidine complex as
a source, which does not appear to be identical to CTR1 or DMT1 (see later). In addition, there is CTR2,
initially reported to be associated only with internal vesicles and organelles (57,61) but now apparently also
in the plasma membrane of some cells (5).
In this report we focused on determining whether 2-macroglobulin (transcuprein) and albumin
deliver copper directly to hepatic and mammary epithelial cells, and initial characterization of the uptake
mechanisms involved. We studied uptake of copper from the purified human proteins in comparison with Cu-
di-histidine, using cultured human hepatic and mammary epithelial cell models (HepG2 and PMC42). The
results reported here indicate that both plasma proteins independently deliver copper efficiently to both cell
types, but that they do so by mechanisms that differ not only with the protein involved but also among the cell
types.

6
6
METHODS AND MATERIALS
Cell culturing and uptake studies. HepG2 cells were obtained from ATCC (Manassas, VA) and
cultured in MEM with non-essential amino acid (0.1mM), sodium pyruvate (1mM), and 10% fetal bovine
serum at 37o in 5% CO2. PMC42 cells, a human adenocarcinoma cell line derived from the pleural effusion
of a breast carcinoma patient (69), were cultured in RPMI1640 with non-essential amino acids (0.1mM),
sodium pyruvate (1mM), and 10% fetal bovine serum. Measurements of rates of copper (and other metal
ion) uptake were carried out in 6-well plates (HepG2 cells), or (for PMC42 cells) 12-well, bicameral Transwell
plates (Corning, Corning, NY). In the latter case, 7.5 x 104 cells were plated on 1:5 diluted Matrigel (BD
Biosciences, San Jose, CA) in each well and grown for 14 days in monolayers, to reach a resistance of 300
Ohms. Uptake studies with 64
Cu-labeled proteins, Cu(II)-dihistidine or Cu(II)-NTA (1:5) were carried out for
30 min in Hepes buffer (50 mM Hepes, pH 7.4, containing 5 mM glucose, 10 mM KCl, 1 mM MgSO4, 130 mM
NaCl), as previously described for Caco2 cell monolayers (41,66), except that with the PMC42 cells, the
metal ions were delivered from the basal chamber (“blood” side). [The copper content of the growth medium
was in the range of 0.6 M, that in the Hepes buffer less than 0.1 M).] Uptake rates were linear for at least
60 min. Uptake over 30 min was used to calculate initial rates and uptake kinetics. Uptake was total
radioactivity in washed cells (HepG2 cells), plus (in the case of the PMC42 cells) radioactivity in the apical
medium. (Cells were washed with Hepes buffer containing 100 histidine.) Overall transfer across the
monolayer was calculated from the radioactivity in the apical medium. ). Kinetic data were analyzed with
Prism 5 software (Graphpad Software, Inc.). 64
Cu (specific activity 20-300 mCi/ g) was obtained from MIR
Radiological Sciences at Washington University Medical School (St. Louis, MO). In one set of experiments,
Caco2 cell monolayers were also used, as previously described, to follow uptake of 59
Fe(II) (1 M) in 1 mM
ascorbate and the same Hepes buffer (72). Other metal ions were added in the form of Ag(I)NO3, ZnCl2, and
Mn(II)-histidine (1:10).
Purification of 2-macroglobulin from human plasma. Purification followed the protocol kindly
provided by Salvatore Pizzo and his laboratory (Duke University, Durham, North Carolina). For this, 300 ml
batches of human plasma, obtained from subjects being treated for iron overload due to hemochromatosis
(through the kindness of Dr. Richard Ajioka; University of Utah Medical Center, Salt Lake City, UT), and
under our university IRB (HSR# 05-004), were fractionated with PEG 8000, the 4-16% saturation fraction

7
7
being subjected to Zn(II)-charged immobilized metal affinity chromatography (IMAC). For the IMAC, sample
was applied in 100 mM Na phosphate buffer, pH 6.5, containing 800 mM NaCl. The column was washed
with 20 mM Na phosphate, pH 6.0, containing 150 mM NaCl; and the 2-macroglobulin was eluted with 10
mM Na acetate, pH 5.0, containing 150 mM NaCl. In some cases, the samples were further purified by size
exclusion chromatography on Sephacryl S300, equilibrated with (low copper) 20 mM K phosphate, pH 7.0.
The resulting protein was pure, as indicated in Figure 1A (left side) which shows an overloaded native PAGE
gel (4.5% acrylamide) stained with Coomassie Blue. Treatment with 200 mM methylamine to convert the
open to the closed form of the protein (Figure 1A) indicates that the open form had been purified, since it
migrated more slowly in native PAGE. SDS-PAGE indicated subunits of 180 kDa (data not shown).
Endogenous copper was removed by 2 day dialysis in 100 M histidine in 20mM Na phosphate buffer pH
7.0, and further dialysis into the Hepes cell incubation buffer. The resulting protein had <0.04 Cu atoms per
subunit.
Purification of human albumin. Purification from human plasma was by a combination of
pseudoaffinity chromatography with Cibacron Blue (Affi-Gel Blue, BioRad, Richmond, CA), using 20mM Na
phosphate buffer, pH 7 to apply the plasma and wash the gel column, and 1M NaCl in the same buffer to
elute the albumin; followed by size exclusion chromatography of concentrated extracts on Sephadex G150,
or removal of residual contaminants with 50% saturation ammonium sulfate. Attached copper was removed
by dialysis against histidine, as described for 2-macroglobulin. The resulting albumin was pure (Figure 1A,
right) and had <0.01 Cu atom per molecule.
Copper loading of 2-macroglobulin and albumin. The stoichiometry of high affinity copper binding to
2-macroglobulin was assessed by mixing 1.25 nmol of the copper-free tetrameric protein with various
amounts of 64
Cu-labeled Cu(II)-nitrilotriacetate (Cu-NTA,1:2 molar ratio) in the presence of 5 nmol albumin
and in 20 mM K phosphate, pH 7.0, and after 1h of incubation, separating the resulting 0.5 ml mixture on 25
ml (1 x 25 cm) Sephadex G150 columns. Columns were standardized with a mixture of ferritin (480 kDa),
IgG (158 kDa), transferrin (80 kDa) and ovalbumin (45 kDa). The nmol of Cu bound to the macroglobulin
were calculated from the proportion of radioactivity in that peak. For cell uptake studies, purified 2-
macroglobulin and albumin (stripped of endogenous copper) were loaded with 64
Cu-labeled Cu-NTA at
specific molar Cu-protein ratios. In some studies, the two proteins were labeled with tracer 64
Cu in situ, by

8
8
addition of pg quantities of tracer (in 0.1 mM HCl and 25 M NTA) to whole human plasma, and separating
the proteins on Sephadex G150 equilibrated with the Hepes buffer used for copper uptake studies.
Protein determinations. These were performed using the Bradford reagent and instructions from
BioRad, with bovine serum albumin as the standard.
Statistics. Calculations were performed via ANOVA, p values <0.05 for differences between means
being considered significant.

9
9
RESULTS
Uptake of copper from albumin and 2-macroglobulin by HepG2 and PMC42 cells. To study the
relative abilities of the two plasma proteins to deliver copper to hepatic and mammary cells, we used these
human cell lines as models and incubated them with the purified human proteins loaded with known amounts
of 64
Cu-labeled Cu(II). To occupy the single high affinity Cu binding site at the N-terminus (10,31), albumin
was loaded with 1 Cu atom per molecule, using Cu(II)-NTA (1:2 molar ratio). Specific binding was verified by
EPR (J. Goto, M Moriya, M. Linder, unpublished data). For 2-macroglobulin we made a preliminary
assessment of the copper binding stoichiometry by incubating 1.25 nmol of the pure, copper-stripped protein
(or 5 nmol of the 180 kDa subunit) with increasing amounts of 64
Cu-labeled Cu(II)-NTA (see Methods) in the
presence of 5 nmol of albumin, then separating the components in size exclusion chromatography (SEC).
We loaded the macroglobulin with copper in the presence of albumin so the latter would prevent non-specific
binding. Radioactivity in the two peaks obtained by SEC reflected that proportions of copper bound to each
protein. Figure 1B-D shows examples obtained with three different amounts of added copper.
Immunoblotting and column standardizations verified that the first peak contained only Cu- 2-macroglobulin
(Mr 770k), and the second one only Cu-albumin (Mr 70k).
The first thing evident is that, when very little copper was added (1.25 nmol), a larger proportion
bound to the macroglobulin than albumin, confirming what we have known for some time, namely that
transcuprein ( 2-macroglobulin in humans, 1-inhibitor-3 in rodents) has an even higher affinity for copper
than albumin. When larger amounts of copper were added, increasing proportions bound to albumin (Figure
1C and D). The amount bound to 2-macroglobulin appeared to plateau at 0.5 Cu atoms per subunit, or 2
Cu atoms per molecule of 4 subunits. This is in the range of what might be expected from in vivo data that
show about 100 ng Cu associated with macroglobulin per ml of human and rat plasma (35,36) and
concentrations of about 1 mg/ml 2-macroglobulin (59). This ratio of Cu:protein was used in all subsequent
uptake experiments.
The question was now whether Cu, attached to either albumin and 2-macroglobulin would be taken
up by cells at rapid rates. To study this, we used copper-protein concentrations (1-2 M) at the low end of
the physiological range, of which about 6 M copper is bound mainly to these proteins. For comparison, we
also examined uptake of 1 M Cu from the di-histidine complex. Figure 2A shows that both cell types took

10
10
up copper from both proteins rapidly. At 1 M Cu, rates of uptake from albumin (hatched bars) and 2-
macroglobulin (dark bars) were in the same range. This was true for both cell types (left vs. right), although
in both cases, rates for Cu-albumin were significantly lower (p<0.001). In the hepatic cells (left side), rates of
uptake from copper histidine were in the same range as those for Cu on 2-macroglobulin. In the mammary
epithelial cell model (PMC42 cell monolayers) (right side), Cu-di-histidine was significantly more effective
than either of the copper proteins at this concentration (p<0.001). It should be noted that PMC42 cells were
in the form of a monolayer with tight junctions, growing on filters in bicameral chambers, and the 64
Cu-loaded
proteins (and Cu-histidine) were applied on the basolateral (“blood”) side. These results showed clearly, for
the first time, that both albumin and the macroglobulin were independently capable of transferring copper
directly to cells. Uptake of copper as the free ion was not occurring (after dissociation of the copper from the
proteins), except perhaps in the process of binding to a transporter, transport system or reductase on the cell
surface. No low molecular weight 64
Cu was detected in the conditioned medium (example in Figure 1E)
fractionated on SEC columns as for Figure 1B-D; and dialysis of the 64
Cu-loaded proteins in PBS resulted in
<5% release over 24h (as previously described; 35). Preliminary EPR analysis indicated specific copper
binding (J. Goto, M. Moriya, M.C. Linder, unpublished results).
To assess whether uptake of copper from albumin would also be robust if only a small proportion
had copper bound to it (as is the case in vivo), we also examined uptake from tracer 64
Cu-NTA-treated whole
plasma fractions, separated by size exclusion chromatography as in Figure 1B-D. With whole plasma, the
tracer bound to the albumin versus macroglobulin fractions in a ratio of 14:1, and the ratio of apo to holo Cu-
albumin was about 250:1. Nevertheless, as shown in Figure 2B, there was good uptake of copper. More
was taken up from albumin than from the macroglobulin because the peak albumin fraction had about 10x
more 64
Cu than that of the macroglobulin, and there was about 150 x more albumin than macroglobulin
offered to the cells on a molar basis (~46 M albumin, ~ 0.3 M macroglobulin.)
Kinetics of copper uptake from albumin, 2-macroglobulin, and histidine. The next step was to
monitor the effects of different concentrations of the Cu-loaded proteins on initial rates of uptake, and to
compare them to the results for Cu-di-histidine, for both cell types. Figure 3 shows the results for HepG2
cells. Covering the range of copper concentrations (as Cu-protein complexes) from 0.1 to 20 M, there
appeared to be one saturable uptake mechanism for Cu delivered on 2-macroglobulin (Figure 3A).

11
11
Software analysis indicated a Km of 3 M, and a Vmax of 22 pmol /min/mg cell protein. Thus, the uptake
mechanism for copper delivered by this protein was sensitive and efficient. For Cu-albumin, there appeared
to be two separate uptake mechanisms in the physiological range from 0.1 to 6 M (Figure 3B and C), one
higher affinity, lower capacity system (Km 0.36 M; Vmax 2.0 pmol/min/mg cell protein) (Figure 3B), the
other a lower affinity, higher capacity one (Figure 3C) (Km 2.4 M, Vmax 18 pmol/min/mg cell protein)
resembling that for Cu- 2-macroglobulin. The data for Cu-di-histidine (Figure 3D) suggested there might be
three different uptake systems, with Kms of about 1, 4 and 10 M, the latter two with the highest uptake
capacities. Kinetic analysis of the data indicated one low capacity (Vmax 2.9) high affinity (Km 0.6 M)
system and ambiguous kinetics above 3 M Cu. Except at the lowest copper concentrations, uptake of Cu
from histidine was higher than that from the two proteins, being about twice as high at 4 M, and 3-4 times
higher at 10 M. Thus, apart from the plasma proteins being able to deliver copper at about the same rate
as Cu-histidine in the physiological range of copper concentrations of the exchangeable copper pool (3-6 M
or below 10 M), the data indicated that there was more than one mechanism of uptake in the case of ionic
copper or that delivered on albumin, in contrast to a single mechanism in the case of Cu- 2-macroglobulin.
Figure 4 shows the data for uptake by the mammary epithelial cells (PMC42). Uptake of copper from
2-macroglobulin (Figure 4A) was as robust as for the hepatic cells, but had different kinetics, with a lower
Km (22 versus 3 M) but higher capacity (Vmax 100 versus 22 pmol/min/mg). In contrast to what occurred
with the HepG2 cells, the kinetics of Cu uptake from albumin did not show a saturable transport system in
the 0-4 M range (Figure 4B). Rather, uptake rates increased almost linearly with Cu-albumin
concentrations up to 25 M. This suggested that Cu-albumin might be diffusing paracellularly through the
monolayer. However, we have shown that when grown in bicameral chambers under the conditions used,
the monolayers have tight junctions and a resistance of 300 Ohms (which was the case here as well), and
thus contain no gaps through which ions could diffuse. In addition, application of 14
C-mannitol or phenol red
to the basal chamber did not result in its leakage into the apical chamber under the same conditions. The
overall kinetics of the uptake systems for Cu-di-histidine (Figure 4C) in PMC42 cells had some similarity to
those for the HepG2 cells (Figure 3D); but kinetic analysis indicated a lower affinity (“Km” 21 M) saturable
system with twice the capacity. In the PMC42 cells, uptake of copper from histidine was again about twice
that for 2-macroglobulin, and 4-fold higher than from albumin. Uptake of copper from the NTA complex was

12
12
also examined in the PMC42 cell monolayers (Figure 4D), showing even higher rates of uptake (Vmax 140
pmol/min/mg) and a Km of 11 M in the physiological range (bottom), and increasing rates at higher
concentrations (top). Taken together, the results of these kinetic studies in two very different kinds of cells
indicated that there were one or more uptake mechanisms with different kinetics for Cu delivered either
tightly bound to albumin or 2-macroglobulin, or kept soluble in the medium by chelation with histidine or
NTA.
The mammary epithelial cell monolayers (PMC42) allowed us not only to measure rates of copper
uptake but also the overall transfer (of the entering copper radioisotope) from the basal to the apical medium.
Transfer from cells to the apical medium would represent copper secreted across the apical membrane in
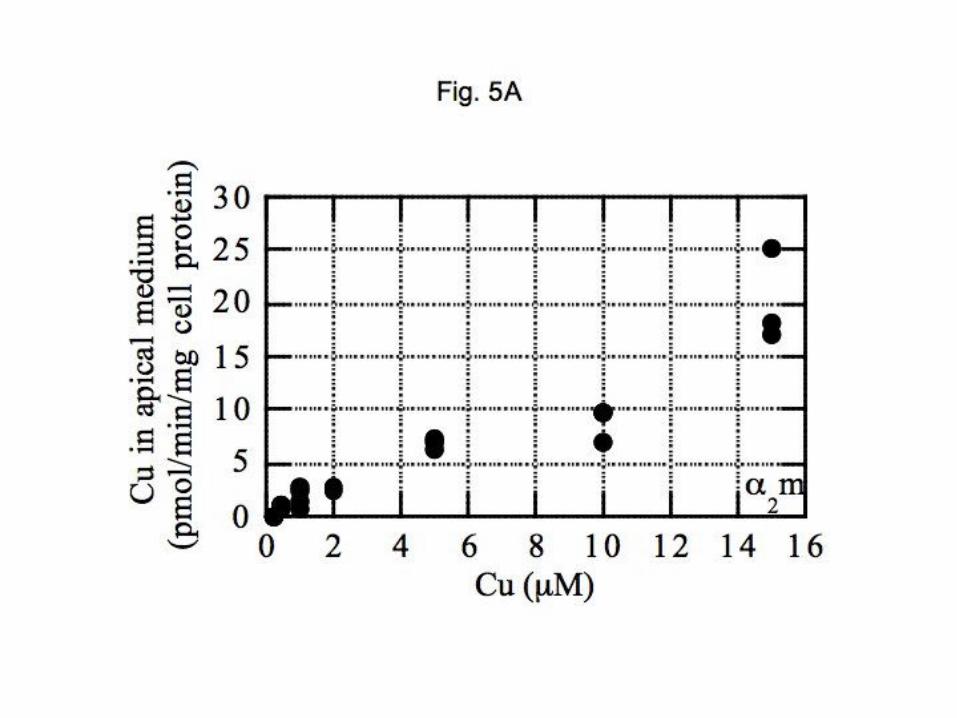
these polarized monolayers, akin to what occurs when milk is secreted. The data presented in Figure 5
show rates of appearance of copper in the apical medium with different Cu-protein (or chelate)
concentrations administered to the basolateral side of the monolayer. Calculations of copper release rates
(pmol/min/mg cell protein) were made on the basis of 64
Cu specific activity in the basal medium. Rates
generally increased in linear fashion in relation to basal copper concentrations, and rates were similar for Cu
on both plasma proteins and Cu-histidine, in the physiological range (<10 M). For Cu-NTA, rates were
higher. Cu-albumin was the least efficient, and there was a distinct lag in apical secretion until the Cu-
albumin concentration exceeded 3 M, implying potential differences in the intracellular routes taken by
copper entering from the two plasma proteins.
Differentiation of copper uptake mechanisms by responses to the presence of high concentrations of
other metal ions. The uptake systems or transporters that could be involved in uptake of copper may at least
in part also absorb other transition metal ions. Large concentrations of such potentially competing ions
would thus be expected to inhibit uptake of copper, depending upon which transporter is involved. For
example, the hepatic copper uptake system characterized by Ettinger et al (17) was markedly inhibited by
Mn(II) and Cd(II) but not by Ni(II), and only marginally by Zn(II), at molar ratios of 5:1 (inhibitor:Cu) and using
10 M Cu(II)-di-histidine. DMT1 is also used by ions of iron, copper and manganese (3,19,22,41,60),
although its importance for Fe(II) uptake has been emphasized. CTR1 on the other hand, is thought to be
more specific, but is inhibited by (the rare) silver ion [Ag(I)] (32,33,75). If the transporter(s) involved in uptake
of copper from albumin and transcuprein (and histidine or NTA) are identical, one would expect that various,

13
13
potentially competing metal ions would identically influence copper uptake from these proteins and chelators.
We thus examined the effects of ions of iron and silver on copper uptake from these factors, in both cell
types. Cells were incubated with Cu-proteins or Cu-chelators at two concentrations (1-2 and 8-10 M Cu) in
the presence and absence of high concentrations (50-200 M) of the potentially competing metal ions.
The results for our studies with HepG2 cells are given in Figure 6. High (50 M) concentrations of
iron (as Fe(II)-NTA) had little or no statistically significant inhibitory effects on uptake of copper delivered by
either 2-macroglobulin, albumin, or histidine (Figure 6A; top); and this was the case at both the higher and
lower concentrations of the Cu-complexes tested (see legend for indications). In contrast, uptake of 10 M
copper from NTA was significantly (~40%) impaired. Much higher concentrations of Fe(II)-NTA (200 M) had
a more severe inhibitory effect on 10 M copper uptake from NTA (60% inhibition), and the same was the
case for 10 M copper delivered on histidine (Figure 6A, bottom). These high concentrations of Fe(II) also
inhibited copper delivery from the plasma proteins, but to a lesser degree. Thus, the uptake mechanisms
active at high concentrations of copper chelate were only affected by very large amounts of ionic iron; and
there was much less or no effect on the uptake mechanisms for copper delivered on albumin and 2-
macroglobulin (perhaps because proteins might scavenge some of the iron under these very unphysiological
conditions and thus mitigate harm to the uptake systems).
The effects of Ag(I) were much more significant with regard to uptake of copper from the plasma
proteins by the hepatic cells (Figure 6B). Here, there was a clear distinction between delivery of copper on
albumin versus 2-macroglobulin: Uptake from the latter was markedly inhibited, but there was only a slight
inhibitory effect on uptake from albumin. This was true at both concentrations of the silver ions (Figure 6B,
top and bottom); and in the case of 8 M Cu-albumin, 50 M Ag(I) actually stimulated copper uptake. The
presence of high concentrations of ascorbate during uptake of copper from 2-macroglobulin did not alter
uptake rates (Figure 6C), implying no (additional) need for copper reduction; but the presence of this
reductant somewhat enhanced the inhibitory effects of Ag(I) (compare grey and black bars). These results
clearly indicate that albumin and 2-macroglobulin are mainly delivering copper to different cellular uptake
systems in the HepG2 cells, and suggest that the macroglobulin is interacting with CTR1 [inhibited by Ag(I)].
Silver ions did inhibit 10 M (but not 1 M) copper uptake by HepG2 cells from histidine (50% with 50 M

14
14
Ag(I); 70% with 100 M Ag(I)], but there was no significant inhibition of uptake from Cu-NTA (Figure 6B),
suggesting some involvement of CTR1 with copper uptake from histidine but not NTA.
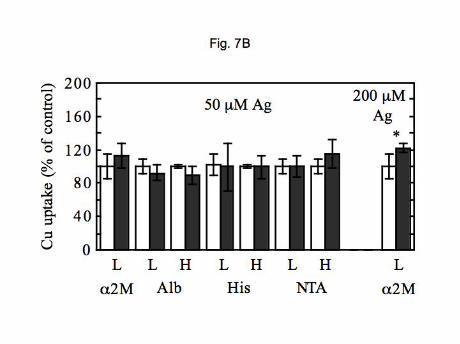
The same parameters were investigated with PMC42 cell monolayers (Figure 7). As in the hepatic
cells, iron had little or no effect on uptake of copper from the two plasma proteins (Figure 7A), except in the
case of 1 M Cu-albumin where 20-30% inhibition was observed. Uptake from histidine was also relatively
unaffected. In the case of silver ions (Figure 7B), there also were no statistically significant inhibitory effects
on uptake of copper from albumin, histidine, or NTA. In contrast to the HepG2 cells, Ag(I) also had no effect
on uptake of 10 M Cu delivered as the di-histidine complex. More importantly, uptake of copper from 2-
macroglobulin was not inhibited. This was a highly reproducible result and confirms kinetic indications that
the two cell types are taking up copper from 2-macroglobulin by different mechanisms.
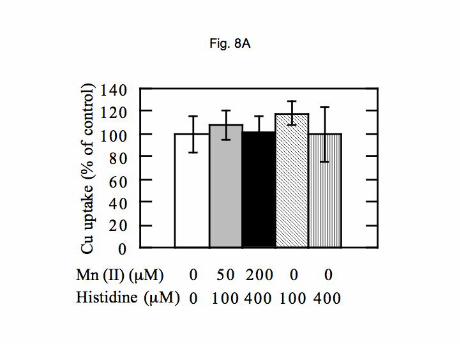
Effects of Mn(II) and Zn(II) on copper uptake from 2-macroglobulin by HepG2 cells. Since the
hepatocyte transporter characterized some years ago by Murray Ettinger and his group (17,35) showed that
uptake from Cu-di-histidine by hepatocytes was inhibitable by manganese and zinc ions, we examined
whether this was the case for copper delivered to HepG2 cells on 2-macroglobulin. As shown in Figure 8,
an excess of neither Mn(II) nor Zn(II) inhibited uptake. Adding Mn(II) at concentrations of 50 and 200 M
(Figure 8A; grey and black bars), as the 1:5 histidine complex, made no difference to uptake of 1 M Cu from
2-macroglobulin. Neither did 100 and 400 M concentrations of histidine alone (hatched and striped bars).
Zinc (200 M (Figure 8B) also had no effect.
No effects of Ag(I) on Fe(II) uptake by Caco2 cell monolayers. To eliminate the possibility that the
inhibitory effects of Ag(I) in HepG2 cells were due to inhibition of DMT1, we investigated whether uptake of
Fe(II) across the brush border of Caco2 cell monolayers (an excellent model for the intestinal mucosa;
3,41,43,73) would be affected by this metal ion. It is well established that in this cell model as well as the
intestinal mucosa, in vivo, DMT1 is the primary (and perhaps only) transporter for uptake of Fe(II) from the
intestinal lumen. As shown in Figure 9, no inhibition was observed at ratios of Ag:Fe of 25 and 50:1. This
indicates that the transport function of DMT1 is not inhibited by silver ions. Therefore, HepG2 cell Cu uptake
from 2-macroglobulin (inhibited by silver) cannot be ascribed to DMT1.

15
15
DISCUSSION
The studies described here were initiated as a first step to understand not only how copper is taken
up from the blood plasma by mammalian cells but how this is regulated in the physiological condition of
lactation, during which considerable copper is transferred from the mother to the newborn through the milk.
In this condition, a substantial proportion of copper associated with the plasma exchangeable copper pool
(bound by albumin and macroglobulin) enters the mammary gland and milk, and less is taken up by the liver
(16). This suggests that specific transporters or transport systems for copper are upregulated in mammary
epithelial cells, and down-regulated in hepatic cells in the suckling period. These transporters might include
CTR1, DMT1, the uptake system described by Ettinger et al (17), and even CTR2, as well as receptor-
mediated endocytosis via the macroglobulin receptor.
The cell line we used to model the mammary epithelial cell (PMC 42) was originally established from
the pleural effusion of a breast carcinoma patient (69). Based on morphological criteria and responsiveness
to lactational hormones, the cells have retained many of the characteristics of mammary epithelium. In
culture, the PMC42 cells differentiate into several cell types, some showing characteristics of secretory cells
(swollen endoplasmic reticulum and secretory vesicles) and others characteristics of myoepithelial cells
(contractile fibers). As with primary cultures of breast epithelium (13,14), PMC42 cells grown as monolayers
as organoids on filters coated with extracellular matrix can be stimulated towards milk production by
sequential treatment with estradiol and progesterone, then insulin, hydrocortisone and prolactin (1,70,71).
These treatments not only induce secretion of lipid (characteristic of milk) but synthesis of the milk-specific
protein, -casein. The cell line also expresses a range of luminal-specific markers (cytokeratin 18, and cell
adhesion molecule E-cadherin) (2), and has the morphology of mammary epithelial cells, with apical microvilli
and nuclei in the cell basal region. Like human breast epithelial cells, they express CTR1 and both ATP7A
and ATP7B (50; A Michalczyk, L Ackland, unpublished results). The hepatic cell model used (HepG2) also
retains many of the characteristics of normal hepatocytes and expresses CTR1, DMT1, and the
macroglobulin receptor (M Moriya, MC Linder, unpublished results; 55).
Since exchangeable copper in blood plasma is tightly bound to proteins (yet exchangeable), and the
same is the case within cells, we would expect that protein-protein interactions are required for transfer of the
metal ion from blood carriers to the transport systems on the cell surface. Binding and cell uptake might then
be driven by mass action, or alternatively coupled with enzymatic reduction, as occurs in the case of Fe(III)

16
16
seeking to enter the enterocyte from the intestinal lumen. In the latter, a cell surface reductase
(dCytb/Cybird1) produces Fe(II) which can then cross the apical membrane through DMT1 (51).
Analogously, a copper reductase (NADH oxidase; 62) on the hepatocyte or mammary epithelial cell surface
might reduce Cu(II) attached to albumin or macroglobulin, to make it available for entry via CTR1.
The results reported here clearly show that both plasma proteins that tightly bind ionic copper in the
blood plasma efficiently and independently deliver the trace element to the two cell types investigated.
Moreover, they do so as efficiently as complexes of ionic copper with amino acids. Human blood plasma
(like that of the rat) contains about 1200 ng Cu/ml, 65-70% of which is on ceruloplasmin and/or other
components of that size (12,35,36). About 350 ng Cu/ml (6 M) are with the exchangeable copper pool,
comprised principally (or exclusively) of albumin and 2-macroglobulin (transcuprein). Estimates from size
exclusion chromatographic profiles indicate that about 60% of that is associated with the very abundant
albumin, and 40% with 2-macroglobulin (44). Thus, the concentration of plasma copper in the
exchangeable pool is 5-6 M, with albumin-Cu at 3-3.6 M, and 2-macroglobulin-Cu at about 2-2.4 M.
Alpha2-macroglobulin itself is present at a concentration of about 1 mg/ml, or 1.3 M; albumin is about 70%
of plasma protein, and thus about 710 M. From these calculations, the molar ratio of Cu: 2-macroglobulin
would be about 2:1, and that for albumin about 0.005. Our findings that about 2 copper atoms bind one
tetrameric 2-macroglobulin molecule fits closely with the estimates just described and indicates that the high
affinity copper binding sites on this protein are more or less saturated, in vivo. The high affinity copper
binding sites on albumin, on the other hand, are far from saturated in vivo (which has been clear for a long
time). These data add strength to the finding that 2-macroglobulin has an even higher affinity for copper
than albumin, as also indicated by the preferential binding of copper to the macroglobulin in the presence of
albumin seen in our loading studies.
The studies we carried out with pure Cu-albumin and Cu- 2-macroglobulin were done with
concentrations right in the physiological range. In HepG2 cells, maximum rates of uptake in the physiological
range of 3-6 M Cu were 12-15 pmol/min/mg cell protein with Cu- 2-macroglobulin, 10-15 pmol/min/mg with
Cu-albumin, and 7-20 pmol/min/mg with Cu-dihistidine. In the mammary epithelial-type cells (PMC42),
maximum rates of uptake in the same physiological range were 10-20 pmol/min/mg cell protein with Cu- 2-
macroglobulin, 5-13 pmol/min/mg for Cu-albumin, and 20-40 pmol/min/mg with Cu-di-histidine (25-60 with

17
17
Cu-NTA). Where saturable, the Kms for the uptake systems for copper from these proteins were in or
somewhat above the physiological range. With Cu- 2-macroglobulin, the kinetics indicated a single
saturable uptake system for both cell types, but with different kinetics. The system in HepG2 cells was of
much higher affinity, with half the capacity of that in the PMC42 cells. In addition, the former but not the
latter was inhibited by Ag(I), suggesting involvement of CTR1. Thus, different uptake systems for Cu- 2-
macroglobulin are involved. For Cu-albumin, different uptake mechanisms may also be operating in the two
cell types, kinetic analysis indicating saturable systems in the physiological range (to 10 uM), and a much
lower affinity higher capacity one (though with similar uptake rates) in the PMC42 cells. Uptake kinetics for
Cu-di-histidine and Cu-NTA were more complex and ambiguous, consistent with multiple carriers and
diffusion, especially at higher (unphysiological) concentrations. [Earlier studies implicated diffusion in uptake
of ionic copper by rat intestinal mucosa, at high copper concentrations (10-90 M; 11).] There was little or no
inhibition by Ag(I) or Fe(II), except at very high concentrations of the latter.
The main points and significance of these findings is that both plasma proteins are capable of
donating copper to more than one cell type independently, and that uptake occurs by different mechanisms
both within and between cell types. Uptake of copper from these proteins is just as efficient as when
delivered as ionic copper complexes (including Cu-di-histidine). As previously reviewed (36,39), there is no
solid evidence that copper-histidine actually exists as a complex in the blood plasma, although it may form a
trimeric complex with Cu-albumin (31,35). Size exclusion fractionations of blood plasma have never
indicated the presence of radioactive or actual copper in components of the size of amino acids. In our
recent HPLC-ICP-MS studies, the lowest molecular weight component detected in mouse, human and rat
plasma was about 11 kDa (12). Binding constants of albumin for copper at the high affinity site have ranged
from 1012
to 1017
M, and 1022
M for the albumin-Cu-histidine complex, compared to a constant of 1017
M for
histidine (31,46). Moreover, albumin (at 700 M) is present in much higher concentrations than histidine (30-
100 M); and the constant for 2-macroglobulin is higher than that for albumin; all of which indicates that
copper would preferentially bind to these two proteins in the plasma rather than histidine, the amino acid with
the highest copper affinity. This is of course also the case within cells, where copper is tightly bound to, and
distributed to other proteins and compartments, by its protein chaperones (52,53).

18
18
As concerns the potential involvement of CTR1 in uptake of copper from 2-macroglobulin by HepG2
(but not PMC42) cells, copper transport through CTR1 is known to be inhibited by Ag(I) (32,33,75), and this
metal ion markedly inhibited copper uptake from the macroglobulin in the hepatic cells, where CTR1 is
known to be highly expressed. However, we now know that at least one other copper transporter (CTR2) is
also inhibited by Ag(I) (5). It seems less likely that CTR2 is involved, because it is not abundantly expressed
in hepatic cells (5) and is mainly confined to the membranes of internal vesicles and organelles (57,61),
although it has been detected in the plasma membrane of transfected COS-7 cells (5). Still other
transporters, not yet identified, might also be inhibited by Ag(I). The studies by Ettinger et al (17)
characterizing a hepatic copper transporter in primary rat and mouse hepatocytes did not examine the
effects of Ag(I). However, that transport system (which used Cu-di-histidine) was markedly inhibited by
Mn(II) and Zn(II), and we found no such inhibition with copper delivered on 2-macroglobulin, tending to
confirm the concept that CTR1 is involved. [Further definitive studies are in progress to answer these
questions.] It should be noted that CTR1 is thought only to transport Cu(I), which could mean that the
copper bound to 2-macroglobulin is in that state. The alternative, namely that Cu(II) on transcuprein
requires a cell surface reductase (47,62) for release and binding or its copper to CTR1 seems unlikely, as
large concentrations of ascorbate capable of reducing Cu(II) had no effect on uptake rates. Our data also
suggest that uptake from higher concentrations of Cu-di-histidine also partly involves CTR1 in HepG2 (but
not PMC42) cells.
It also seems unlikely that in our studies, Cu- 2-macroglobulin was delivering copper (even partly)
through receptor-mediated endocytosis via the macroglobulin receptor, which is well known to be abundant
on hepatocytes (18,67) and has also been identified on mammary epithelial cells (5). The macroglobulin
receptor takes up the compact form of this protein, which is produced when the “bait region” inside the
subunits is cleaved (as by proteases; 59), resulting in a marked conformational change that exposes regions
to interact with the receptor and remove it from the plasma (18,20,67). We purified and used the “open” form
of the protein, which is the most abundant in the blood plasma. In addition, although both cell types are
thought to express the macroglobulin receptor, our data indicated that the macroglobulin was delivering
copper to different uptake systems in the two cell types, one inhibited and the other not inhibited by Ag(I).
It also seems unlikely that DMT1 is involved, since we observed no inhibition by excess iron (at a
ratio of 25:1) on copper uptake from 2-macroglobulin, and the same was true for excess manganese, which

19
19
is also thought to use DMT1 for uptake (19,22,60). This is consistent with extensive further evidence from
our laboratory, utilizing not just cell culture but also the Belgrade rat, which produces an inactive form of
DMT1 (M. Moriya, Y.H. Ho, E. Sauble, M.C. Linder, unpublished), showing that copper uptake is unaltered.
As concerns the uptake system(s) to which albumin was delivering copper, there was no inhibition by
silver or ferrous iron at ratios of 50:1, and only 20% inhibition at 100:1, indicating that neither CTR1, CTR2 or
DMT1 are likely to be involved. Although uptake of copper from albumin by hepatocytes and fibroblasts has
previously been demonstrated (17,48,49,65), our studies are in the first to examine Cu-albumin uptake
kinetics. Our HepG2 cell data indicated two saturable systems in the physiological range that were not seen
in the PMC42 cells, where uptake rates were almost linear with Cu-albumin concentrations, indicating a
difference in response of the two cell types to this form of copper.
Previous studies had shown that the addition of given concentrations of albumin to solutions of CuCl2
or Cu-di-histidine decreased uptake of that copper by rat hepatocytes, fibroblasts and even trophoblasts
(7,45,64). For example, albumin added in a 1:1 molar ratio to Cu(II) inhibited uptake by rat hepatocytes 50%
and about 8-fold in the case of fibroblasts (65). Histidine added to CuCl2 also decreased uptake rates (64).
A single more detailed published study where uptake of copper in the presence of a range of albumin
concentrations was studied showed that the protein had an increasingly inhibitory effect (7). Uptake of 2 M
67Cu-dihistidine by rat hepatocytes fell dramatically with the addition of increasing amounts of albumin, falling
from about 90 pmol/min/mg cell protein to about 8 when albumin concentrations reached 0.1 mg/ml or more
(>90% lower rates). This might suggest that with actual plasma albumin concentrations (which are orders of
magnitude higher- about 45 mg/ml), copper uptake rates might dwindle to nothing. However, we found that
when 64
Cu tracer (pg amounts) on albumin (obtained by addition to whole human plasma) was used for
uptake studies, rapid uptake still occurred (Figure 2B). In this case, only about 0.4% of the total was bound
to the metal. The idea of albumin inhibiting copper uptake in vivo thus seems untenable.
The rates of copper uptake and the Kms we obtained with our two cell types were of the same order
of magnitude but lower than those reported by others for primary cultures of primary hepatocytes. For
example, Waldrop et al (65) reported rates of 40 pmol/min/mg for rat hepatocytes with 5 M Cu(II)-albumin
compared to our rates of about 12 for the HepG2 cells. Bingham and McArdle (7) obtained rates of 80
pmol/min/mg with rat hepatocytes and 2 M Cu-di-histidine, ten-fold higher than our rates with HepG2 cells,
but only 4-fold higher than what we obtained with the mammary epithelial cell line. As concerns the kinetics

20
20
of uptake from Cu-di-histidine in HepG2 cells, our evidence for high affinity (Km 0.6 M) low capacity
saturable system, and non saturable kinetics at higher concentrations, differs from that of Ettinger et al (17)
for primary hepatocytes, where the Km was in the range of 11 M. All of these differences are most likely
due to differences between primary and immortalized cells.
The strong implication of our findings that CTR1 plays little or no role in uptake of copper from the
blood plasma proteins to which it is bound, especially in the mammary epithelial cell model, may at first seem
counterintuitive, as this transporter is ubiquitously expressed in mammalian cells and clearly does play a role
in copper uptake elsewhere. Kuo et al (28) demonstrated strong expression in the epithelium of ducts and
alveoli of mouse mammary gland in lactation; Kelleher and Lonnerdal (26) showed expression in the
lactating gland of rats. However, the data from both these groups indicated that normally, most or all of the
CTR1 was confined to cytoplasmic organelles, in part colocalizing with transferrin receptor containing
endosomes (26), and thus largely unavailable for mediating uptake at the cell surface. Treating the cells with
prolactin, or having pups suckle the rat dams (inducing prolactin release) greatly and transiently enhanced
the proportion of CTR1 on the plasma membrane (26), which correlated with a 50% increase in the rate of
copper uptake (from 67
CuCl2). Moreover, prolactin induced CTR1 to migrate both to the basolateral and
apical portions of the HC11 cells, suggesting that it plays a role not just in uptake of copper from the blood
but also in re-uptake from the developing milk (unless CTR1 might work in both directions – which is not
inconceivable). Since our studies with the PMC42 cells were done in the absence of prolactin, when little or
no CTR1 might be on the basal cell surface, it is not surprising that Ag(I) did not inhibit copper uptake.
(Studies on the effects of prolactin are in progress.)
In all, our findings indicate that although CTR1 does play a role in uptake of copper from the blood by
hepatocytes, this is only when delivered by 2-macroglobulin and does not happen in the mammary cells.
More importantly and as detailed earlier, our studies indicate that the remaining known uptake systems that
could be involved in uptake of copper from albumin and 2-macroglobulin (DMT1, macroglobulin receptor,
CTR2, the Ettinger system) do not play a major role. Uptake from Cu-albumin also differs between the two
cell types, particularly at lower copper concentrations, judging from differences in kinetics. In the PMC42
cells, the observed differential kinetics of copper release from the mammary epithelial cell monolayers at the
apical surface, in response to Cu-albumin and Cu-macroglobulin in the basal medium, additionally implies
either that copper delivered by these proteins is entering by mechanisms that lead to different intracellular

21
21
pools and routes across the cell and into the apical fluid, and/or that the two Cu-proteins trigger different
intracellular responses of copper transporters/transport mechanisms that lead to apical copper release.

22
22
ACKNOWLEDGEMENTS
We are grateful for the help of Dr. Salvatore Pizzo at Duke University for the protocol to purify human
2-macroglobulin, and to Dr. Richard Ajioka at University of Utah for the human plasma used for that purpose
and for purifying human albumin.
GRANTS
This work was supported by U.S. Public Health Service Grants RO1 HD46949 (M.C. Linder, PI; M.L.
Ackland and J.F.P. Mercer, co-PIs) and IR24CA86307 (MIR, Washington University, St. Louis) for
subsidizing of 64
Cu production.

23
23
REFERENCES
1. Ackland ML, Michalczyk A, Whitehead RH. PMC42, a novel model for the differentiated breast. Exp
Cell Res 263: 14-22, 2001.
2. Ackland ML, Newgreen DF, Price J, Fridman M, Waltham MC, Arvanitis A, Minichiello J, Price JT,
Thompson A. Epidermal growth factor-induced epithelio-mesenchymal transition in human breast
carcinoma cells. Lab Invest 83: 435-448, 2003.
3. Arredondo M, Munoz P, Mura CV, Nunez MT. DMT1, a physiologically relevant apical Cu+1 transporter
of intestinal cells. Am J Physiol 284: C1525-1530, 2003.
4. Barrow L, Tanner MS. Copper distribution among serum proteins in paediatric liver disorders and
malignancies. Eur J Clin Invest 18: 555-560, 1988.
5. Bertinato J, Swist E, Plouffe LJ, Brooks SPJ, L’Abbe MR. Ctr2 is partially localized to the plasma
membrane and stimulates copper uptake in COS-7 cells. Biochem J, Oct 19, 2007
6. Beshgetoor D, Lonnerdal B. Identification of alpha2-macroglobulin receptor in human mammary
epithelial cells. J Nutr 129: 152-157, 1999.
7. Bingham MJ, McArdle HJ. A comparison of copper uptake by liver plasma membranes vesicles and
uptake by isolated cultured rat hepatocytes. Hepatol 20: 1024-1031, 1994. 8
8. Birkenmeier G, Heidrich K, Glaeser C, Handschug K, Fabricius EM, Frank R, Reissig D. Different
expression of the alpha-2-macroglobulin receptor/low-density lipoprotein receptor-related protein in human
keratinocytes and fibroblasts. Arch Dermatol Res 290:561-568, 1998.
9. Birkenmeier G, Struck F, Gebhardt R. Clearance mechanism of prostate specific antigen and its
complexes with 2-macroglobulin and 1-antichymotrypsin. J Urol 162: 897-901, 1999.
10. Breslow E. Comparison of cupric ion-binding sites in myoglobin derivatives and serum albumin. J Biol
Chem 239: 3252-3259, 1964.
11. Bronner F, Jost J.H. Saturable and nonsaturable copper and calcium transport in mouse duodenum.
Am J Physiol 249: G108-G112, 1985.
12. Cabrera A, Alonzo E, Chu YL, Linder MC, Sato D, Mason AZ. Copper binding components of blood
plasma and organs, and their responses to influx of large doses of 65
Cu, in the mouse. Biometals, in press.

24
24
13. Darcy KM, Shoemaker SF, Lee PP, Vaughn MM, Black JD, Ip MM. Prolactin and epidermal growth
factor regulation of the proliferation, morphogenesis, and functional differentiation of normal rat mammary
epithelial cells in three dimensional primary culture. J Cell Physiol 163: 346-364, 1995
14. Darcy KM, Shoemaker SF, Lee PP, Ganis BA, Ip MM. Hydrocortisone and progesterone regulation of
the proliferation, morphogenesis, and functional differentiation of normal rat mammary epithelial cells in three
dimensional primary culture. J Cell Physiol 163: 365-379, 1995.
15. Darwish HM, Cheney JC, Schmitt C, Ettinger MJ. Mobilization of copper (II) from plasma components
and mechanisms of hepatic copper transport. Am J Physiol (Gastrointest Liver Physiol) 9: G72-G79, 1984.
16. Donley SA, Ilagan BJ, Rim H, Linder MC. Copper transport to mammary gland and milk during
lactation in rats. Am J Physiol 284: G739-G747, 2001.
17. Ettinger MJ, Darwish HM, Schmitt RC. Mechanisms of copper transport from plasma to hepatocytes.
Fed Proc 45: 2800-2804, 1986.
18. Falkenberg C, Allhorn M, Thogersen IB, Valnickova Z, Pizzo SV, Salvesen G, Akerstrom B,
Enghild JJ. 1Microglobulin destroys the proteinase inhibitory activity of 1inhibitor3 by complex formation.
J Biol Chem 270: 4478-4483, 1995.
19. Garrick MD, Kuo HC, Vargas F, Singleton S, Zhao L, Smith JJ, Paradhar P, Roth JA, Garrick LM.
Comparison of mammalian cell lines expressing distinct isoforms of divalent metal transporter 1 in a
tetracycline-regulated fashion. Biochem J 398: 539-546, 2006.
20. Gliemann J, Sottrup-Jensen L. Rat plasma 1inhibitor3 binds to receptors for 2macroglobulin. FEBS
Lett 221: 55-60, 1987.
21. Goforth J, Vivas E, Liu NM, Askary HS, Lo LSL, Linder MC. Correspondence between rat
transcuprein and human alpha-2-macoglobulin in copper binding. FASEB J 15: Abstract 237.6, 2001.
22. Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, Nussberger S, Gollan
JL, Hediger MA (1997) Cloning and characterization of a mammalian proton-coupled metal-ion transporter.
Nature 388, 482-488.
23. Hamsa I, Faisst A, Prohaska J, Chen J, Gruss P, Gitlin JD. The metallochaperone Atox1 plays a
critical role in perinatal copper homeostasis. Proc Natl Acad Sci USA 98: 6848-6852, 2001.

25
25
24. Hollborn M, Birkenmaier G, Saalbach A, Iandiev I, Reichenbach A, Wiedemann P, Koben L.
Expression of LRP1 in retinal pigment epithelial cells and its regulation by growth factors. Invest Ophthalmol
Vis Sci 45: 2033-2038, 2004.
25. Huffman DL, O’Halloran TV. Function, structure, and mechanism of intracellular copper trafficking
proteins. Annu Rev Biochem 70: 677-701, 2001.
26. Kelleher SI, Lonnerdal B. Mammary gland copper transport is stimulated by prolactin through
alterations in CTR1 and ATP7A localization. Am J Physiol 291: R1181-R1191, 2006.
27. Klomp LW, Lin SJ, Yuan DS, Klausner RD, Culotta VC, Gitlin JD. Identification and functional
expression of HAH1: a novel human gene involved in copper homeostasis. J Biol Chem 272: 9221-9226,
1997.
28. Kuo Y-M, Gybina AA, Pyatskowit JW, Gitschier J, Prohaska JR. Copper transport protein (Ctr1)
levels in mice are tissue specific and dependent on copper status. J Nutr 136: 21-26, 2006.
29. Kuo Y-M, Zhou B, Cosco D, Gitschier J. The copper transporter CTR1 provides an essential function
in mammalian embryonic development. Proc Natl Acad Sci USA 98: 6836-6841, 2001.
30. Kurdowska A. Alden SM, Noble JM, Stevens MD, Carr FK. Involvement of alpha-2-macroglobulin
receptor in clearance of interleukin 8-alpha-2-macroglobulin complexes by human alveolar macrophages.
Cytokine 12: 1046-1053, 2000.
31. Lau S, Sarkar B. Ternary coordination complex between human serum albumin, copper(II) and L-
histidine. J Biol Chem 246: 5938-5943, 1971.
32. Lee J, Pena MMO, Nose Y, Thiele, DJ. Biochemical characterization of the human copper transporter
Ctr1. J Biol Chem 277: 4380-4387, 2002.
33. Lee J, Petris MJ, Thiele DJ. Characterization of mouse embryonic cells deficient in the Ctr1 high affinity
copper transporter: Identification of a CTR1-independent copper transport system. J Biol Chem 277: 40253-
40259, 2002.
34. Lee J, Prohaska JR, Thiele DJ. Essential role for mammalian copper transporter Ctr1 in copper
homeostasis and embryonic development. Proc Natl Acad Sci USA 98, 6842-6847, 2001.
35. Linder MC. Biochemistry of Copper. New York: Plenum, 1991.
36. Linder MC. Biochemistry and molecular biology of copper in mammals. In: Handbook of Copper
Pharmacology and Toxicology, edited by Massaro EJ. Totowa, NJ: Humana, 2002, pp. 2-32.

26
26
37. Linder MC, Cerveza P, Cotton S, Wooten L, Shulze R., Lomeli N. Copper transport. Am. J. Clin. Nutr.
67: 965S-971S, 1998.
38. Linder MC, Donley S, Dominguez D, Wooten L, Mehrbod F, Cerveza P, Shulze R, Cotton S,
Sridhar A, Lomeli N, Hallock M. Copper transport and ceruloplasmin during lactation and pregnancy. In:
Metals and Genetics, edited by Sarkar B. New York: Kluwer/Plenum, 1999, pp. 117-1129.
39. Linder MC, Hazegh-Azam M. The biochemistry and molecular biology of copper. Am. J. Clin. Nutr.
Supplement, 63: 797S-811S, 1996.
40. Linder MC, Lomeli NA, Donley S, Mehrbod F, Cerveza P, Cotton S, Wooten L. Copper transport in
mammals. In: Copper Transport and its Disorders, edited by Leone A and Mercer JFB. New York:
Kluwer/Plenum, New York, 1999, pp. 1-16.
41. Linder MC, Moriya M, Whon A, Kassa A, Gilley C. Recent studies on vesicular transport of Fe and
interactions with other metal ions in intestinal absorption, as studied in polarized Caco2 cell monolayers.
Biol. Res. 39: 143-156, 2005.
42. Linder MC, Wooten L, Cerveza P, Cotton S, Shulze R, Lomeli N. Copper transport. Am J Clin Nutr
(Suppl.) 67: 965S-971S, 1998.
43. Linder MC, Zerounian NR, Moriya M, Malpe R. Iron and copper homeostasis and intestinal absorption
using the Caco2 cell model. Biometals 16: 145-160, 2003.
44. Liu NM, Lo LSL, Askary SH, Goforth J, Vivas E, Tsai MT, Linder MC. Transcuprein is a
macroglobulin regulated by copper and iron. J Nutr Biochem. 18: 597-608, 2007.
45. Mas A, Sarkar B. Uptake of 67
Cu by isolated human trophoblast cells. Biochim Biophys Acta 1135:
123-128, 1992.
46. Masuoka J, Hegenauer J, Van Dyke BR, Saltman P. Intrinsic stoichiometric equilibrium constants for
the binding of zinc(II) and copper(II) to the high affinity site of serum albumin. J Biol Chem 268: 21533-
21537, 1993.
47. McArdle HJ, Bingham MJ, Summer K, Ong TJ. Cu metabolism in the liver. Adv Exp Med Biol 448:
29-37, 1999.
48. McArdle HJ, Guthrie JR, Ackland ML, Danks DM. Albumin has no role in the uptake of copper by
human fibroblasts. J. Inorg. Biochem. 31: 123-131, 1987.

27
27
49. McArdle HJ, Gross SM, Danks DM. Uptake of copper by mouse hepatocytes. J Cell Physiol 136: 373-
378, 1988.
50. Michalczyk A, BNastow E, Greenough M, Camakaris J, Freestone D, Taylor P, LOinder M, Mercer
J, Ackland ML. ATP7B expression in human breast epithelial cells is mediated by lactational hormones. J
Histochem Cytochem Jan 7 (EPub), 2008.
51. Oakhill JS, Marritt SJ, Gareta EG, Cammack R, McKie AT. Functional characteristization of human
duodenal cytochrome b (Cybrd1): Redox properties in relation to iron and ascorbate metabolism. Biochim
Biophys Acta Dec 23 (EPub), 2007.
52. Pena MMO, Lee J, Thiele DJ. A delicate balance: Homeostatic control of copper uptake and
distribution. J. Nutr. 129: 1251-1260, 1999.
53. Portnoy ME, Schmidt PJ, Rogers RS, Culotta VC. Metal transporters that contribute copper to
metallochaperones in Saccharomyces cerevisiae. Mol Genet Genomics 265: 873-882, 2001.
54. Pufahl RA, Singer CP, Peariso KL, Lin SJ, Schmidt PJ, Fahmi CJ, Culotta VC, Penner-Hahn JE,
O’Halloran TV. Metal ion chaperone function of the soluble Cu(I) receptor Atx1. Science 278: 853-856,
1997.
55. Quinn KA, Grimsley PG, Dai YP, Tapner M, Chesterman CN, Owensby DA. Soluble low density
lipoprotein receptor-related protein (LRP) circulates in hman plasma. J Biol Chem 272: 23946-23951, 1997.
56. Rae TD, Schmidt PJ, Pufahl RA, Culotta VC, O’Halloran TV. Undetectable intracellular free copper:
the requirement of a copper chaperone for superoxide dismutase. Science 284: 805-808, 1999.
57. Rees EM, Thiele DJ. Identification of a vacuole-associated metalloreductase and its role in Ctr2-
mediated intracellular copper mobilization. J Biol Chem 282: 21629-21638, 2007.
58. Scott KC, Turnlund JR. Compartmental model of copper metabolism in adult men. J Nutr Biochem 5:
342-350, 1994.
59. Sottrup-Jensen L. Macroglobulins: Structure, shape, and mechanism of proteinase complex
formation. J Biol Chem 264: 11539-11542, 1989.
60. Thompson K, Molina RM, Donaghey T, Schwob JE, Brain JD, Wessling-Resnick M. Olfactory
uptake of manganese requires DMT1 and in enhanced by anemia. FASEB J 21: 223-230, 2007.

28
28
61. van den Berghe PV, Folmer DE, Malingre HE, can Beurden E, Klomp AE, van de Sluis B, Merkx M,
Klomp LW. Human copper transporter 2 is localized in late endosomes and lysosomes and facilitates
cellular copper uptake. Biochem J 407: 49-59, 2007.
62. Van den Berghe GJ, McArdle HJ. A plasma membrane NADH oxidase is involved in copper uptake by
plasma mambrane vesicles isolated from rat liver. Biochim Biophys Acta 1195: 276-280, 1994.
63. Vargas EJ, Shoho AR, Linder MC. Copper transport in the Nagase analbuminemic rat. Am. J. Physiol.
267: G259-G269, 1994.
64. Waldrop GL, Ettinger MJ. Effects of albumin and histidine on kinetics of copper transport by
fibroblasts. Am J Physiol (Gastrointest Liver Physiol) 22: G212-G218, 1990.
65. Waldrop GL, Fakhruddin AP, Hadi M, Lonergan PA, Ettinger MJ. Effect of albumin on net copper
accumulation by fibroblasts and hepatocytes. Am J Physiol (Gastrointest Liver Physiol) 22: G219-G225,
1990.
67. Ward MD, Ajioka R, Kaplan J. Cohort movement of different ligands and receptors in the intracellular
endocytic pathway of alveolar macrophages. J Biol Chem 264: 8164-8170, 1989.
68. Weiss KC, Linder MC. Copper transport in rats involving a new plasma protein. Am J Physiol 249: E77-
E88, 1985.
69. Whitehead RH, Bertoncello I, Webber LM, Pederson JS. A new human breast carcinoma cell line
(PMC42) with stem cell characteristics. I. Morphological characterization. J Natl Cancer Inst 70: 649-661,
1983a.
70. Whitehead RH, Monaghan P, Webber LM, Bertoncello I, Vitali AA. A new human breast carcinoma
cell line (PMC42) with stem cell characteristics. II. Characterization of cells growing as organoids. J Natl
Cancer Inst 71: 1193-1203, 1983b.
71. Whitehead RH, Quirk SJ, Vitali AA, Funder JW, Sutherland RL, Murphy LC. A new human breast
carcinoma cell line (PMC42) with stem cell characteristics. III. Hormone receptor status and responsiveness.
J Natl Cancer Inst 73: 643-648, 1984.
72. Zerounian NA, Linder MC. Effects of copper and ceruloplasmin on iron transport in the Caco2 cell
intestinal model. J. Nutr. Biochem. 13, 138-148, 2002.
73. Zerounian NA, Redekosky C, Malpe M, Linder MC. Regulation of copper absorption by copper
availability in the Caco2 cell intestinal model. Am. J. Physiol. (GI) 284, G739-G747, 2003.

29
29
74. Zhou B, Gitschier J. hCTR1: a human gene for copper uptake identified by complementation in yeast.
Proc Natl Acad Sci USA 94, 7481-7486, 1997.
75. Zhou H, Thiele, DJ. Identification of a novel high affinity copper transport complex in the fission yeast
Schizosaccharomyces pombe. J Biol Chem 276: 20529-20535, 2001.

30
30
FIGURE LEGENDS
Fig. 1. Purity and copper loading of 2-macroglobulin in the presence of albumin and its retention in
conditioned medium. (A) Native PAGE of purified 2-macroglobulin (left) and SDS-PAGE of purified albumin
(right). In the case of the macroglobulin, samples shown were (+) and were not (-) pretreated with
methylamine, to convert the open to the closed form of the protein. SDS-PAGE gave subunits of 180 kDa
(data not shown). (B-D) Sephadex G 150 chromatographic separation of 64
Cu-labeled 2-macroglobulin
( 2M), albumin (Alb) and Cu-NTA, after preincubating 5 nmol of pure 2-macroglobulin subunit, 5 nmol pure
albumin, and 1.25 (B), 2.5 (C) and 7.5 (D) nmol 64
Cu-labeled Cu(II) (as the NTA complex) in 0.50 ml (see
Methods). The nmol copper bound to 2-macroglobulin were calculated from the proportion of radioactivity
associated with that protein peak. For (A), (B), and (C), the proportions were 63, 54 and 26%, respectively.
(E) Elution of radioactive copper on 2-macroglobulin following use in uptake studies with cultured (PMC42)
cells. Conditioned medium, from cultures incubated with 2 M 64
Cu-labeled copper bound to pure human 2-
macroglobulin, was separated by Sephadex G150 chromatography, as in B-D, except that a slightly larger (1
x 28 cm) column was used. Radioactivity was still bound to 2-macroglobulin (also verified by
immunoblotting of the 64
Cu peak fractions).
Fig. 2. Copper is taken up efficiently by HepG2 and PMC42 cells from human 2-macroglobulin or albumin,
and from the Cu-di-histidine complex. A. Data show initial rates of uptake (pmol/min/mg cell protein) from
pure 2-macroglobulin (grey bar), albumin (hatched bar), and Cu-di-histidine (white bar) by HepG2 cells (left),
and PMC 42 cell monolayers (right), after administration of copper (1 M) bound to these proteins. Values
are Means + SD, for N = 12. All differences, except between uptake from macroglobulin and histidine in the
PMC42 cells, were statistically significant (p<0.001). B. Initial rates of uptake of 64
Cu by HepG2 cells from
albumin and macroglobulin fractions of whole human plasma. For this, tracer 64
Cu-NTA was added to whole
plasma, and the macroglobulin (a2M) and albumin fractions separated on Sephadex G150 prior to incubation
of equal volumes of the peak fractions with the cells for 30 min. Cell radioactivity (cpm/mg cell protein)
values are Means + SD, for N = 4. Radioactivity could not be translated into nmol copper, as actual copper
concentrations and thus 64
Cu specific activity were not determined.
Fig. 3. Kinetics of copper uptake by HepG2 cells from 2-macroglobulin, albumin, and histidine. (A) Initial
rates of copper uptake (pmol/min/mg cell protein) from 2-macroglobulin ( 2m) at different Cu- 2-

31
31
macroglobulin concentrations, from 0.1 to 20 M Cu. Kinetic analysis indicated a Vmax of 22 pmol/min/mg,
and a “Km” of 3 M. (B) Initial rates of copper uptake from albumin (Alb) at concentrations from 0.1 to 0.4
M. Kinetic analysis indicated a Vmax of 2.0 pmol/min/mg and a “Km” of 0.36 M. (C) Cu-albumin uptake
data for 0.4-10 M gave a Vmax of 18 pmol/min/mg and a “Km” of 2.3 M. (D) Initial rates of copper uptake
from Cu(II)-di-histidine at concentrations from 0.1-20 M (top) and 0-3.5 M (bottom). The latter gave a
Vmax of 2.9 pmol/min/mg and a “Km” of 0.6 M. Data for higher concentrations did not yield standard kinetic
values.
Fig. 4. Kinetics of copper uptake by PMC42 cells from 2-macroglobulin, albumin, and histidine and NTA.
(A) Initial rates of copper uptake (pmol/min/mg cell protein) from 2-macroglobulin ( 2m) at concentrations
from 0.1 to 15 M Cu. Kinetic analysis indicated a Vmax of 100 pmol/min/mg, and a “Km” of 22 M. (B)
Initial rates of copper uptake from albumin (Alb) at concentrations from 0.1 to 25 M gave values for Vmax
and “Km” of 194 pmol/miin/mg and 131 M, respectively. (C) Initial rates of copper uptake from Cu(II)-
dihistidine (His) concentrations of 0.1-10 M, gave Vmax and “Km” values of 206 pmol/min/mg and 21 M,
respectively. (D) Initial rates of copper uptake from Cu(II)-NTA (NTA) concentrations of 0.1-20 M (top), and
0-8 M (bottom). Only the latter gave kinetic values for Vmax (140 pmol/min/mg) and “Km” (11 M).
Fig. 5. Transfer of copper from mammary epithelial cells (PMC42) to the apical medium, after delivery of
64Cu-labeled copper on 2-macroglobulin (A) ( 2m), albumin (B) (Alb), histidine (C) (His), or NTA (D) to the
basolateral surface of the monolayer. Data are 64
Cu absorbed from the basal chamber that was released to
the apical medium after administration of various concentrations of Cu attached to the proteins or chelators,
using the specific radioactivity of the original 64
Cu for calculations of release rates in pmol/min/mg cell
protein.
Fig. 6. Effects of iron and silver ions on copper uptake from plasma proteins and chelators by HepG2 cells.
Uptake (30 min) of copper attached to 2-macroglobulin ( 2M), albumin (Alb), histidine (His) or NTA, at lower
(L) or higher (H) concentrations, was measured in the absence (open bars) and presence (closed/gray bars)
of (A) 50 or 200 M Fe(II)-NTA (1:5), or (B) 50 or 100 - 200 M Ag(I). Uptake from 2M was only at the
lower concentration of 2 M Cu, 1 M protein. L and H copper concentrations otherwise were 1 and 8 M for
Cu-albumin; 1 and 10 M for Cu-dihistidine and Cu-NTA. (C) Uptake of copper (2 M) from 2-
macroglobulin (1 M), in the absence (white and hatched bars) and presence (grey and black bars) of 50 M

32
32
Ag(I), measured with (black and hatched bars) or without (white and grey bars) added 1mM ascorbate in the
medium. Values, presented as percent of control (no competing ions) (open bar) within a given experiment
(combining data from several experiments), are given as Means + SD. For A and B, N= 6-12 for Cu- 2-
macroglobulin; 5-15 for Cu-albumin; 9-27 for Cu-dihistidine; and 6-14 for Cu-NTA. For (C), N=6. Starred
values indicate statistically significant differences from the controls (*p>0.01 or **p< 0.001). Actual (control)
copper uptake rates (in pmol/min/mg cell protein) for (A-C) were 3.4-5.7 for 2M; 2.2-3.3 and 11-15 for 1 and
8 M Cu-albumin; 3.7-4.2 and 38-54 for 1 and 10 M Cu-dihistidine; and 7.0-7.5 and 16-35 for 1 and 10 M
Cu-NTA, respectively.
Fig. 7. Effects of iron and silver ions on copper uptake from plasma proteins and chelators by PMC42 cell
monolayers. Uptake (30 min) of copper attached to 2-macroglobulin ( 2M), albumin (Alb), histidine (His) or
NTA, at lower (L) or higher (H) concentrations, was measured in the absence (open bars) and presence
(closed/gray bars) of (A) 50-500 M Fe(II)-NTA (1:5), as indicated; or (B) 50 or 200 M Ag(I). Uptake from
2M was only at the lower (L) concentration of 2 M Cu, 1 M protein. Otherwise, L and H copper
concentrations were 1 and 8 M for Cu-albumin; 1 and 10 M for Cu-dihistidine and Cu-NTA. Values,
presented as percent of control (no competing ions) (open bar) within a given experiment (combining data
from several experiments), are given as Means + SD, with N= 6-12 for Cu- 2-macroglobulin; 4-9 for Cu-
albumin; 4-12 for Cu-dihistidine; and 6-12 for Cu-NTA. Starred values indicate statistically significant
differences from the controls, *p< 0.01-**p< 0.001. Actual (control) copper uptake rates (in pmol/min/mg cell
protein) for (A-B) were 3.6-4.3 for 2M; 1.9 and 10 for 1 and 8 M Cu-albumin; 3.7 and 60 for 1 and 10 M
Cu-dihistidine; and 6.0-7.2 and 54 for 1 and 10 M Cu-NTA, respectively.
Fig. 8. No effects of manganese and zinc ions on uptake of copper from 2-macroglobulin by HepG2 cells.
(A) Effects of 50 and 200 M Mn(II)-histidine (1:2) on uptake of 2 M Cu on 1 M 2-macroglobulin,
presented as percent of control from several experiments (Mean + SD). Controls – no competing ions (white
bars, N=12); 50 M Mn(II) (grey bars, N= 3); 200 M Mn(II) (black bars, N = 4); 100 M histidine alone
(hatched bar, N= 6) and 400 M histidine alone (lined bar, N= 4). Actual Cu uptake rates for controls
averaged 5.5 + 1.9 pmol/min/mg cell protein. (B) Effect of 200 M Zn(II) (black bar, N=6) on uptake of
copper from 2 M Cu on 1 M 2-macroglobulin, compared to control (white bar, N=6). Actual Cu uptake
rates for controls averaged 3.4 + 0.1 pmol/min/mg cell protein.

33
33
Fig. 9. No effects of Ag(I) on uptake of Fe(II) by Caco2 cell monolayers. Monolayers with tight junctions
grown on filters in 12-well Transwells, were administered 1 M 59
Fe(II) (in 1 mM ascorbate) in the apical fluid,
and uptake into cells and basal medium were recorded over 30 min, in the absence and presence of two
concentrations of Ag(I), as indicated. The results are given as percent of control (Mean + SD, N=4-8),
combining data from two separate experiments.
























Copyright © 2022 FDOKUMEN




















