
Listeria monocytogenes-Infected Human Dendritic Cells: Uptake and Host Cell Response
Upload
independentCategory
view
0download
0
1
Construction and characterization of an attenuated Listeria 1
monocytogenes cancer immunotherapeutic for clinical use. 2
3
4
Attenuated Listeria monocytogenes as immunotherapeutic 5
6
7
Anu Wallecha*1, Paulo Cesar Maciag
1, Sandra Rivera
1, Yvonne Paterson
2, Vafa Shahabi
1 8
9
10
1 Research and Development, Advaxis Inc., North Brunswick, NJ-08902 11
2 Department of Microbiology, University of Pennsylvania, Philadelphia, PA-19104
12
13
14
* Corresponding author 15
Anu Wallecha, Ph. D 16
Senior Scientist, Advaxis Inc. 17
675 US Highway One, Suite 120 18
Technology Center of New Jersey, North Brunswick, NJ-08902 19
Tel No-91- 732-545-1590 extn-3202 20
Fax- 91-732-545-1591 21
23
ACCEPTED
Copyright © 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Clin. Vaccine Immunol. doi:10.1128/CVI.00274-08 CVI Accepts, published online ahead of print on 19 November 2008

2
Abstract 1
Listeria monocytogenes (Lm) has been exploited previously as a vaccine vector for the 2
delivery of heterologous proteins such as tumor specific antigens for active cancer 3
immunotherapy. However for effective use of live vector in clinic, safety is a major concern. In 4
the present study, we describe an irreversibly attenuated and highly immunogenic Lm platform, 5
Lm dal(-) dat(-) actA(-) strain that expresses the human prostate specific antigen (PSA) using an 6
antibiotic resistance marker free plasmid (LmddA142). Despite limited in vivo survival 7
LmddA142 was able to elicit efficient immune responses required for tumor clearance. Our 8
results showed that immunization in mice with LmddA142 caused the regression of the tumors 9
established by prostate adenocarcinoma cell line expressing PSA. An evaluation of immunologic 10
potency indicated that LmddA142 elicits a high frequency of PSA specific immune responses. 11
Interestingly, immunization with LmddA142 induced significant infiltration of PSA specific T 12
cells in the intratumoral milieu. Collectively, our data suggests that LmddA142 is safe and potent 13
vector for clinical use and this platform may be further exploited as potential candidate to 14
express other single or multiple antigens for cancer immunotherapy. 15
16
17
18
19
20
21
22
ACCEPTED

3
Introduction 1
Biological and immunological characteristics of Listeria monocytogenes (Lm) make this 2
gram-positive bacterium an ideal vaccine vector. Lm triggers potent cellular immune responses 3
in an infected host due to its ability to survive in both phagocytic and cytosolic compartments. 4
Several groups have shown recombinant Lm to be an effective agent for immunotherapy against 5
infection and cancer (2, 16, 19-21, 24, 27, 28). Currently, there are two methods to genetically 6
modify Lm for expressing heterologous antigens in vivo. These include the insertion of a 7
heterologous gene in the bacterial chromosome either by homologous recombination, (7, 16) or 8
phage specific insertion (13) and the transformation of Lm with a plasmid carrying a foreign 9
antigen (7, 26). The plasmid-based strategy has the advantage of multi-copy expression but relies 10
on complementation for the maintenance of the plasmid in vivo. To address this, two 11
mechanisms have been described previously for Lm. One is based on the complementation of a 12
prfA deficient Lm strain (XFL7) with a copy of episomal prfA (a major gene transcription 13
activator for several virulence genes in Lm) (7). This complementation ensures the retention of 14
the plasmid in vivo but requires the presence of antibiotic resistance genes for in vitro selection. 15
The second approach uses complementation with alanine racemase (dal), an enzyme involved in 16
the synthesis of the cell wall component D-alanine (35). 17
In Lm, the D-alanine metabolism is regulated by two genes, dal and dat (34). The 18
complementation of Lm dal dat (Lmdd) with either one of these genes is sufficient for restoring 19
the synthesis of D-alanine both in vivo and in vitro (34, 35). Based on this property, Verch et al 20
designed the shuttle vector, pTV3 that is devoid of antibiotic resistance markers but harbored a 21
copy of the Lm-dal gene. This plasmid could complement the growth of both Escherichia coli 22
ala drx (MB2159) and Lmdd mutants in vivo and in vitro (35). The strain Lmdd contains 82% 23
ACCEPTED

4
deletion of the dal gene and the remaining 18% corresponds to 44 bp at the N-terminus and 158 1
bp of the C-terminus (33). The 18% homology between the episomal Lm-dal and Lmdd 2
chromosome is low for reverse recombination to create a revertant. However, due to regulatory 3
concerns for using this plasmid in humans, it was safer to replace Lm-dal gene in pTV3 with the 4
non-homologous Bs-dal gene. Previously, Zhao et al have shown that (37) Bacillus subtilis dal 5
(Bs-dal) gene can complement the growth of the Lmdd strain in vivo and in vitro. 6
Lmdd strains containing the pTV3-based plasmids are often attenuated merely due to the 7
metabolic burden caused by the expression of a foreign antigen. Therefore, there is always a 8
probability that these strains might regain their original virulence, if the antigen expression is 9
reduced or lost for any reason. This prompted us to construct an attenuated strain that exhibits a 10
reduction in virulence due to an irreversible deletion of a major Lm virulence gene i.e. actA. 11
ActA is a major virulence factor of Lm that is involved in actin polymerization and is necessary 12
for cytoplasmic movement and cell-cell spread of the organism (12). Here, we tested this new 13
generation of antibiotic free, Bs-dal plasmid antigen expression system by complementing the 14
attenuated Lmdd∆actA (LmddA) strain using the tumor antigen, human prostate specific antigen 15
(PSA). We tested the resulting LmddA142 (LmddA expressing PSA) strain in a previously 16
described mouse model for prostate cancer for its ability to eradicate established tumors and to 17
induce cell mediate immunity to PSA (27). 18
PSA is a kallikrein serine protease (KLK3) secreted by prostatic epithelial cells (36) and 19
used as a serum marker for Prostate Cancer (PCa) (32). The potential of PSA as an antigen in 20
immunotherapy is due to its over-expression in malignant cells of prostate, in contrast to its low 21
expression by normal prostate epithelial cells and other organs such as small intestine and testis 22
(3). There is a great deal of information regarding the potential of this antigen for the treatment 23
ACCEPTED

5
of PCa (1, 10, 15, 17, 25). Previously, we showed that a recombinant Lm vaccine expressing 1
PSA (Lm-LLO-PSA) can cause the regression of solid tumors expressing PSA in a murine model 2
for prostate cancer. (27). The application of Lm-LLO-PSA is not ideal for a clinical setting due to 3
its virulence and the presence of two chloramphenicol resistance genes (CAT) for in vitro 4
selection in gram-negative (E. coli) and gram-positive (Lm) bacteria. Here we show that the Bs-5
dal based antibiotic free plasmid in an attenuated LmddA backbone is a more efficient delivery 6
vector when compared to our previously described Lm-LLO-PSA vaccine. The expression of the 7
heterologous antigens from LmddA based on the antibiotic free shuttle vectors is potentially more 8
compatible with the international regulatory requirements and could be developed further for 9
clinical use. 10
11
Materials and Methods: 12
Peptides, oligos, antibodies and FACS reagents: Peptides were synthesized by EZbiolabs 13
(Westfield, IN) and oligos by Invitrogen (Carlsbad, CA). The reagents for flow cytometry were 14
obtained from Becton Dickinson (San Diego, CA). The antibodies used for staining in flow 15
cytometry were: CD3e-PerCP-Cy5.5 (clone 142-2C11); CD4-PE (clone RM4-5); CD8-FITC 16
(clone 53-6.7); CD25-APC (clone PC61); CD62L-APC (clone MEL-14); IFN-γ-PE (clone 17
XMG1.2); Anti-Caspase-3-PE (clone C92-605); Anti-FoxP3 (Miltenyi Biotec, Auburn, CA). 18
Culture media and supplements were obtained from Gibco (Carlsbad, CA) or Sigma (St. Louise, 19
MO). ELISpot antibodies were obtained from Mabtech (Cincinnati, OH). PSA tetramers were 20
prepared by Emory University (Atlanta, GA). All other reagents, unless indicated, were from 21
Sigma. 22
ACCEPTED

6
Mice, cell lines and media. C57BL/6 mice and C57BL/6 Ifng tm1tms
(GKO-/-
) were purchased from 1
Jackson laboratories (Bar Harbor, ME) and were maintained at Cook campus animal facility at 2
Rutgers University, New Brunswick, NJ. Experiments with mice were performed after written 3
approval with the Institutional Animal Care and Use committee at Rutgers University. The 4
construction of the prostate adenocarcinoma cell line expressing human PSA (TPSA23) was 5
described previously (27). TPSA23 cells were grown and maintained in Dulbecco’s modified 6
Eagle’s medium (DMEM) supplemented with 4 mM glutamine and adjusted to contain 1.5 g/L 7
of sodium bicarbonate, 4.5 g/L glucose, 5µg/ml insulin, 10 nM dehydroisoandrosterone, 5% fetal 8
bovine serum (FBS), 5% Nuserum IV (BD biosciences, CA) in the presence of 5µg/ml 9
Blasticidin (Invitrogen). MC57G fibrosarcoma cells were maintained in Eagle’s minimum 10
essential medium (EMEM) with 2 mM glutamine, 1.5 g/L sodium bicarbonate, 0.1 mM non-11
essential amino acids, 1.0 mM sodium pyruvate and 10% FBS. EL4 lymphoma cells were 12
maintained in DMEM with 4 mM glutamine adjusted to contain 1.5 g/L sodium bicarbonate, 4.5 13
g/L glucose and 10% FBS. J774A.1, a murine macrophage like cell line was maintained in RPMI 14
1640 medium with 4 mM glutamine, 1.5 g/L sodium bicarbonate, 4.5 g/L glucose and 10% FBS. 15
Complete RPMI (C-RPMI) contained RPMI 1640 complemented with 2 mM glutamine, 0.1 mM 16
non-essential amino acids, 1.0 mM sodium pyruvate and 10% FBS, Penn/Strep (1%), Hepes 17
buffer (1 mM). For immunological assays such as intracellular cytokine staining and cytotoxic T 18
cell assays, 2-mercaptoethanol was added to C-RPMI medium to a final concentration of 55µM. 19
20
Construction of the LmddA strain. This strain was constructed using the method of 21
homologous recombination as described previously in Mata et al (16). The construction of Lmdd 22
strain has been described previously (34). The Lmdd strain is based on the Lm genetic 23
ACCEPTED

7
background 10403S, which contains a streptomycin resistance gene integrated in the 1
chromosome. To delete actA from Lmdd, the chromosomal region corresponding to the upstream 2
(657 bp) and downstream (625 bp) of actA was amplified and joined by SOEing PCR. The 3
primers used for the amplification of DNA region upstream of actA was UActA-F1-4
ccGGATCCgcgccaaatcattggttgattg and UActA-R1- 5
cgtcgtatggttccctgggtcggggttaatcgtaatgcaattggc and downstream region were DActA-F2- 6
acgattaaccccgacccagggaaccatacgacgttaattcttgc and DActAR2- 7
ccGGATCCgctaggcctaatttataaaacgc. The resulting 1282 bp DNA segment was subsequently 8
cloned in the temperature sensitive shuttle plasmid, pKSV7 at its BamHI restriction site 9
(indicated in the primers). Lmdd strain was transformed with the plasmid ∆actA/pKSV7. 10
Repeated passaging of this strain in BHI/D-alanine (100 µg/ml) medium at two different 11
temperatures, 30°C and 42°C resulted in the desired recombination events that created the 12
LmddA strain. The deletion of the actA region was verified by PCR using primers: forward-13
tgggatggccaagaaattc and reverse-ctaccatgtcttccgttgcttg that anneal externally to the recombinant 14
gene. 15
16
Construction of the pAdv142 plasmid. Several modifications were introduced into the original 17
pTV3 plasmid (35) to create the next generation of E.coli/Lm antibiotic free shuttle plasmid. To 18
delete prfA, pTV3 was digested with the restriction enzymes (RE) XmaI and EheI (New England 19
Biolabs). The plasmid pTV3 was first linearized with XmaI and its ends were filled with Klenow 20
polymerase. The pTV3/XmaI linearized and end-filled fragment was further digested with EheI 21
restriction enzyme, resulting in the release of 1571 bp DNA fragment containing prfA. The 22
remaining 6000 bp of the plasmid backbone was blunt-ligated resulting in the pTV3∆prfA 23
ACCEPTED

8
plasmid. The cassette for p60 promoter-Listeria dal at the NheI/PacI restriction sites was 1
replaced by p60 promoter-Bacillus subtilis dal resulting in the plasmid, pTV3Bs-dal∆prfA. The 2
Bs-dal gene (protein ID: NP 388345.1) was amplified from the chromosomal DNA of the 3
Bacillus subtilis 168 (ATCC) using the forward primer, dal F-4
gagaggagttttccatgagcacaaaacctttttacagagatacgtgggcgg which contained a 5’ overhang to join with 5
the p60 promoter by SOEing PCR and the reverse primer dal R- 6
cGCTAGCttaattgcttatatttacctgcaataaaggatttc which contained a NheI site (underlined). The klk3 7
was amplified from our previously described plasmid, pAdv34 (27) using oligos; F- 8
gtgCTCGAGattgtgggaggctgggagtg and R- gttCCCGGGttaggggttggccacgatgg. The RE sites in 9
the oligos are underlined. The human PSA gene, klk3 was cloned in the pTV3Bs-dal∆prfA at its 10
XhoI/XmaI restriction sites resulting in the plasmid, pAdv142. 11
12
Construction of the strains, Lmdd142 and LmddA142. The Lmdd and LmddA were 13
transformed with pAdv142 by electroporation resulting in the strains Lmdd142 and LmddA142. 14
The expression and secretion of LLO-PSA fusion protein was confirmed in the culture 15
supernatants of both these strains using anti-PSA and anti-LLO antibodies, according to the 16
previously described protocols (27). Lmdd142 and LmddA142 were passaged twice in mice as 17
previously reported (23). Murine macrophage like cell line, J774A.1 was used to examine the 18
ability of LmddA142 to grow intracellularly as described previously (27). 19
20
In vitro and in vivo stability studies. Plasmid maintenance in vitro was determined by serial 21
passages under selective and non-selective conditions. Bacteria were cultured in 10 ml of 22
BHI/Strp and subcultured daily at 1/10,000 dilution into fresh medium in the presence or absence 23
ACCEPTED

9
of D-alanine (100 µg/ml). Bacterial titers were determined daily on BHI plates +/- 100 µg/ml D-1
alanine. Streptomycin (100 µg/ml) was added to the medium as a control to select for Lm and to 2
reduce the potential growth of contaminants. 3
Plasmid maintenance in vivo was determined by intravenous injection of 5 x 107 CFU of 4
LmddA142. Viable bacterial loads were determined in the spleens homogenized in PBS on day 1, 5
2 and 3 (2 mice/day). CFU was determined at each time point by plating on BHI/Strp plates in 6
the presence or absence of 100 µg/ml D-alanine. 7
8
In vivo virulence and clearance studies with Lmdd142 and LmddA142. The C57BL/6 (WT) 9
male mice were immunized i.p. with different doses of Lmdd142 (107
and 108 CFU) and 10
LmddA142 (108
and 109 CFU) and were monitored for the signs of sickness for a period of 10 11
days. The in vivo clearance of the strains was examined by immunizing mice (10/group) with 2 x 12
106 CFU of Lmdd142 (WT) or 1 x 10
8 CFU of LmddA142 (WT and GKO
-/-). Viable CFU in the 13
homogenized spleens and livers was determined from two mice in each group on day 1, 2, 3, 7 14
and 10 after plating cell suspension on BHI/Strp medium. 15
16
Tumor regression study. This was performed using the murine adenocarcinoma prostate tumor 17
model expressing PSA, TPSA23 (27). Three groups of male, C57BL/6 mice (8/group) were 18
implanted with 2 x 106
TPSA23 cells on day 0 and were immunized with 107 CFU of Lm-LLO-19
PSA (27), 108 CFU LmddA142 on day 6, 13 and day 20 or untreated (naïve). Tumor growth was 20
monitored once a week using electronic calipers for a period of 8 weeks. The mice were 21
sacrificed when the tumor size was found to be greater than 15 mm. 22
ACCEPTED

10
Immunogenicity studies in mice. Male C57BL/6 mice (2/group) were immunized twice with 1
LmddA142 (108 CFU), Lm-LLO-PSA as positive control (10
7 CFU) at one-week interval or left 2
untreated (naïve). The immune responses elicited for the PSA H-2Db peptide epitope 3
(65
HCIRNKSVIL74
) (22) were determined on day 6 after the boost using ELISpot, PSA tetramer 4
staining and intracellular cytokine staining for IFN-γ, as described previously (27). The data 5
were analyzed for tetramer staining and IFN-γ staining using CellQuest Pro software. 6
7
Cytotoxic T cell assay. Splenocytes from immunized and naïve mice were stimulated in vitro 8
for 5 days in C-RPMI containing 20 U/ml of IL-2 (Sigma) in the presence of mitomycin C 9
treated MC57G cells infected with PSA/vaccinia at an effector: stimulator ratio of 20:1 as 10
described previously (27). The cytotoxicity of effector T cells was examined using a FACS 11
assay, which is based on the staining for cleaved caspase-3 and thus, measures the number of 12
apoptotic target cells (9). The target cells EL4 were labeled with 0.6 µM DDAO-SE (Molecular 13
Probes, Invitrogen) for 20 min at 37°C. Labeled cells were washed once with C-RPMI, 14
resuspended at a concentration of 2 x 106/ml and pulsed with 1 µg/ml of PSA peptide for 1 hour. 15
Labeled EL4 cells pulsed with PSA were then washed once in C-RPMI and were finally 16
resuspended at a concentration of 1 x 106/ml for use in the CTL assay. Effector cells were mixed 17
with the targets at effector/target (E:T) ratio of 10:1, 3:1, 1:1, 0.3:1 and 0.1:1 and incubated for 3 18
h at 37°C, 5% CO2. After 3 h, these cells were fixed with paraformaldehyde for 20 min, washed 19
twice in FACS buffer (PBS, 2% FBS), permeabilized with perm-wash (BD) and labeled with 20
anti-caspase-3-PE antibody (BD) for 1 h. The induction of apoptosis in the target cells was 21
measured by determining the number of caspase+/DDAO-SE
+ cells at each effector/target (E:T) 22
ratio. Specific lysis was considered to be directly related to the percentage of caspase+ cells and 23
ACCEPTED

11
described as (% caspase+
DDAO-SE+ EL4 PSA peptide pulsed cells) – (% caspase
+DDAO-SE
+ 1
EL4 cells). 2
3
Analysis of tumor infiltrating lymphocytes (TILs). Male C57BL/6 mice (3/group) were 4
implanted with TPSA23 tumors on day 0 immunized with 108 CFU of LmddA142 and 10
8 CFU 5
of irrelevant Lm (Lm-LLO-E7) on day 7 and day 14. On day 20, tumors embedded in Matrigel 6
were excised from the mice. The PSA specific TILs and regulatory T cells were determined in 7
the tumors embedded in Matrigel and spleens using the protocol described previously (27). 8
9
Statistical Analyses. Non-parametric Kruskal Wallis test was applied to compare the tumor 10
sizes among different treatment groups. Tumor sizes were compared on day 40 for statistical 11
analysis, because this was the latest time-point with the highest number of mice in each group. 12
The Kaplan-Meier test was applied to compare the survival of mice in different groups. A p-13
value of less than 0.05 was considered statistically significant in these analyzes. 14
15
Results: 16
Construction of actA deleted Lm strain (LmddA) that expresses tLLO-PSA fusion protein 17
by means of an antibiotic resistance free plasmid. To construct an attenuated LmddA strain, an 18
in frame deletion of actA was generated in Lmdd background to avoid any polar effects on the 19
expression of the downstream genes. The resulting strain, LmddA contains the first 19 amino 20
acids at the N-terminus and 28 amino acid residues at the C-terminus with a deletion of 591 21
amino acids of the chromosomal ActA. The deletion of actA was verified by PCR using primers 22
that anneal externally to this region of the chromosome on the genomic DNA isolated from 23
ACCEPTED

12
Lmdd or LmddA (data not shown). The deletion of actA was further confirmed by DNA 1
sequencing. 2
To increase the stability and safety of the previously constructed pTV3 (35), we 3
introduced two modifications in this plasmid. First, an unnecessary copy of prfA gene in this 4
plasmid was deleted and further, the Lm-dal gene was replaced with Bs-dal gene. The new 5
plasmid, pAdv142 (Fig. 1A) expressed Bs-dal under the control of Lm p60 promoter. The 6
plasmid, pAdv142 was able to complement the growth of MB2159 (33) as well as Lmdd or 7
LmddA in the absence of exogenous D-alanine. The antigen expression cassette in pAdv142 8
consists of hly promoter from Lm and the tLLO-PSA fusion protein. 9
Both Lmdd and LmddA were transformed with pAdv142 resulting in Lmdd142 and 10
LmddA142. The expression and secretion of the fusion protein tLLO-PSA was confirmed in 11
these strains by Western blots using both anti-PSA and anti-LLO antibodies (Fig. 1B). These 12
strains were passaged twice in vivo in C57BL/6 mice to select for stable vaccine strains (23). 13
Lmdd142 and LmddA142 retained the expression and secretion of tLLO-PSA fusion protein after 14
two passages (data not shown) suggesting that both these constructs were stable in vivo. 15
16
ActA deletion results in 2 logs attenuation of Lmdd vaccine strain. We determined the 17
maximum safe dose (MSD) of Lmdd142 and LmddA142 in C57BL/6 mice after administering 18
different doses of each construct. Our results indicated that a dose of 106
CFU for Lmdd142 and 19
108
CFU for LmddA142 was one-tenth minimum dose observed to have adverse effects in mice 20
and thus, was used as safe dose for further studies. These virulence studies showed that 21
LmddA142 was at least 2 logs more attenuated than Lmdd142. Additionally, we observed that 22
GKO-/-
mice, which are very susceptible to wild type Lm (8) were also resistant to 108
CFU of 23
ACCEPTED

13
LmddA142 suggesting that high levels of IFN-γ released during Lm innate immune responses 1
control infection possibly by preventing cell to cell spread. 2
We further examined the in vivo clearance of both of these strains after intraperitoneal 3
administration of the MSD in C57BL/6 mice and GKO-/-
mice. We did not detect any live 4
LmddA142 in the livers and spleens of either of these strains on day 3 post-injection. This 5
suggests that LmddA142 is completely cleared after 48-72 h post injection (Fig 2A). In contrast, 6
Lmdd142, containing the chromosomal actA, could survive in vivo for up to 7 days reaching a 7
peak growth in the liver and spleen on day 3 post-infection (Fig. 2B). Our data demonstrate that 8
LmddA142 is a safer vaccine construct than its unattenuated counterpart. Therefore, we chose 9
LmddA142 to evaluate its stability and therapeutic efficacy. 10
11
LmddA142 is stable both in vitro and in vivo and can grow intracellularly. We examined the 12
in vitro stability of the plasmid pAdv142 by passaging LmddA142 in the presence or absence of 13
selective pressure for seven days. Total CFUs were determined each day after plating on 14
selective and non-selective media. It was expected that a loss of plasmid would result in higher 15
CFU counts after plating on non-selective medium (BHI+D-alanine). As depicted in Fig. 3A, 16
there was no significant difference between the number of CFUs in selective and non-selective 17
medium and this shows that LmddA142 retains the plasmid pAdv142 for at least 60 generations. 18
The in vivo stability of pAdv142 in LmddA142 was tested after immunizing C57BL/6 19
mice once with this strain intravenously and examining the number of viable bacteria at different 20
time points such as 24 h, 48 h and 72 h by plating on both selective and non-selective media. We 21
observed no significant differences (p= 0.1566, using paired student t-test) in the CFUs after 24 h 22
by plating under either condition suggesting that pAdv142/LmddA142 was stable in vivo (Fig. 23
ACCEPTED

14
3B). However, this does not exclude the possibility of plasmid loss. No colonies were recovered 1
after 48 h and 72 h post injection, which was due to the rapid in vivo clearance of LmddA142. 2
Furthermore, to determine if the deletion of actA caused any deleterious effect on the 3
ability of LmddA142 to infect macrophages and its intracellular growth, we performed a cell 4
infection assay. We observed that both Lmdd142 and LmddA142 were able to infect and grow in 5
this cell line. The more virulent strain Lmdd142 displayed a similar growth rate to that of the 6
wild type Lm 10403S. However fewer colonies were recovered with LmddA142 at different time 7
points of growth (Fig. 3C). The differences in the uptake of Lmdd142 and LmdA142 were 3-4 8
fold at 0 h but at time 8 h were 37 fold. Thus, there is a greater difference at 8 h when compared 9
to 0 h suggesting that the loss of cell to cell spread in LmddA142 contributes to the reduction in 10
its intracellular growth. 11
12
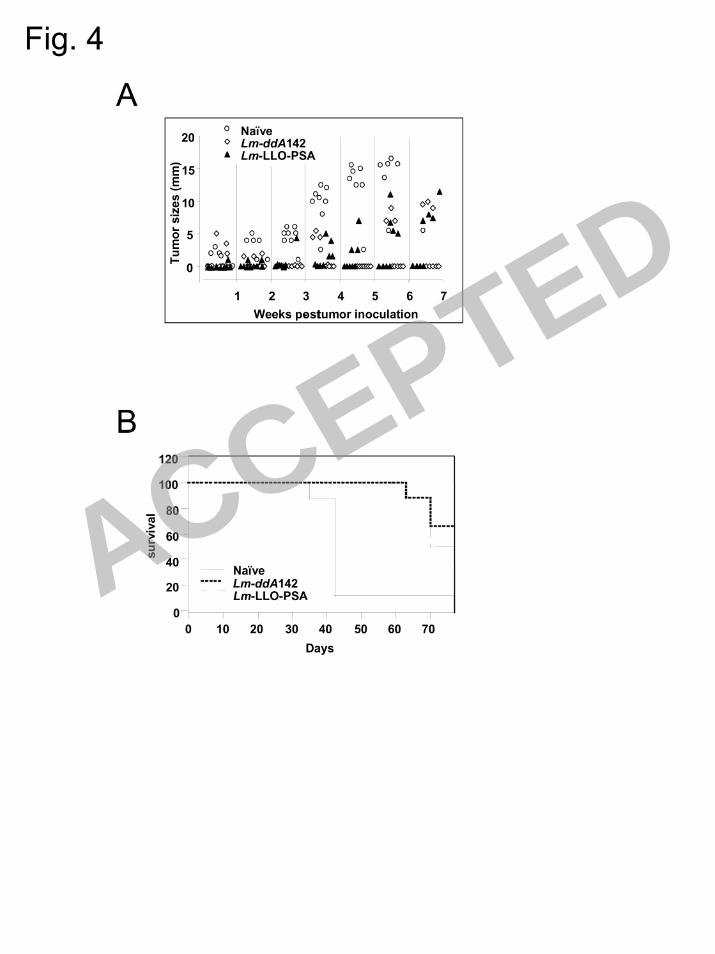
LmddA142 mediates the regression of established tumors expressing human PSA. After 13
initial characterization, we further extended the study to evaluate the therapeutic efficacy of 14
LmddA142 using a murine prostate adenocarcinoma cell line engineered to express human PSA, 15
i.e. TPSA23 (27). Naïve mice developed tumors gradually and all were sacrificed before day 45 16
(Fig. 4A). Immunization of mice with Lm-LLO-PSA, which served as a positive control for the 17
study resulted in the complete regression of three out of eight tumors. In contrast, five out of 8 18
mice immunized with LmddA142 became tumor free and remained in this state until the 19
experiment was terminated, day 70 (Fig. 4A). The statistical differences between each group 20
were examined on day 40 using non-parametric Kruskal-Wallis test. The results indicate that 21
immunization with the strain, LmddA142 makes a significant impact on the TPSA23 tumor 22
ACCEPTED

15
growth (p=0.001). Thus, LmddA142 caused complete tumor regression in 60% of the 1
experimental animals. 2
Additionally, the average survival of mice in each group was determined using Kaplan-3
Meier analysis (Fig. 4B). The mean survival of mice immunized with Lm-LLO-PSA (70+ 2 days) 4
and LmddA142 (73 + 2 days) was two-fold higher than the naïve mice (42 + 2 days) and the 5
overall comparison with log rank test showed that p = 0.01. These results suggest that 6
vaccination with LmddA142 significantly impacts on the survival of mice. 7
8
Immunization with LmddA142 elicits PSA specific cellular immune responses in mouse 9
spleens. We have shown previously that the ability of recombinant Lm based vaccines to regress 10
the growth of established solid tumors is associated with the generation of antigen specific T cell 11
responses. We examined PSA specific immune responses elicited by LmddA142 in C57BL/6 12
mice using PSA specific tetramer staining, intracellular cytokine staining (IFN-γ) and ELISpot 13
assays. Staining with the PSA specific tetramer showed that after two immunizations with 14
LmddA142, 23% of activated CD8+ CD62L
low T cells were PSA specific in the splenocytes (Fig. 15
5A). This was a 2-fold high PSA tetramer specific T cell when compared to the mice that 16
received Lm-LLO-PSA. 17
The results for the intracellular staining for IFN-γ were consistent with those obtained 18
from the tetramer staining. There was a two fold increase in IFN-γ+
CD8+
CD62Llow
cells (5 %) 19
in LmddA142 immunized splenocytes relative to the mice immunized with Lm-LLO-PSA (2.2 %) 20
(Fig. 5B). Using ELISpot assay we observed that there was a 10 fold increase in the number of 21
IFN-γ secreting cells in splenocytes from LmddA142 immunized mice in response to in vitro 22
pulsing with PSA H-2Db
peptide when compared to no peptide (data not shown). 23
ACCEPTED

16
The functional activity of the T cells generated against PSA was determined using an in 1
vitro CTL assay that detects the cleavage of caspase 3 in target cells as a function of cell killing 2
(9). At an effector to target ratio of 10:1, 70 % of cleaved caspase 3+ EL4/H-2D
b PSA peptide 3
pulsed cells were detected when incubated with in vitro stimulated effector T cells from either 4
LmddA142 or Lm-LLO-PSA (Fig. 5C). This response was reduced in proportion to the decrease 5
in effector : target ratio. Taken together, these assays show that LmddA142 is highly 6
immunogenic in mice as implied by the detection of high frequency of PSA specific CD8+ T cell 7
responses. 8
9
Immunization with LmddA142 results in infiltration of tumors by PSA specific 10
lymphocytes. Further we investigated the ability of PSA specific CD8+ lymphocytes generated 11
by vaccination with LmddA142 to infiltrate into the tumors. We observed that a very low number 12
of PSA-specific TILs (2 %) were present in the tumors harvested from both naïve and irrelevant 13
Lm immunized mice. However, there was about 8-fold increase in the percentage of PSA-14
specific TILs (16 %), in the tumors of the mice immunized with LmddA142 when compared to 15
the naïve mice (Fig. 6A). 16
In addition, we determined the presence of CD4+/CD25
+/Foxp3
+ Tregs in the tumors and 17
spleens of untreated or Lm immunized mice. Interestingly, we observed that immunization with 18
Lm (LmddA142 or irrelevant Lm) resulted in a 2-3 fold decrease in the number of Tregs in the 19
tumors but not in the spleens. However, LmddA142 showed a relatively stronger impact in 20
decreasing the frequency of Tregs in tumors when compared to the irrelevant Lm immunized 21
group (Fig. 6B). 22
23
ACCEPTED

17
Discussion 1
In the present study, we describe the construction of an irreversibly attenuated Lm 2
delivery vector LmddA that harbors an antibiotic resistance free plasmid to express a tumor 3
specific antigen. The auxotropic mutant Lmdd was constructed with an aim to create an 4
attenuated Lm-based platform for clinical use (34). However, Lmdd elicited potent immune 5
responses only if administered in the presence of D-alanine, which may not be appropriate for an 6
immunotherapeutic (34). Alternatively, trans complementation of Lmdd with dal not only 7
restores the synthesis of D-alanine but also in vivo virulence. Previously, two mechanisms have 8
been proposed that reduce Lmdd virulence after dal complementation. These mechanisms 9
involve the use of either a plasmid that expresses Bs-dal gene under the control of tightly
10
regulated inducible promoter (14) or a Bs-dal containing suicidal plasmid, which is subsequently 11
sensitive to dilution and degradation in vivo (37). Complementation of Lmdd using both these 12
methods causes the cessation of bacterial growth in vivo after two or three generations, reducing 13
their applicability as potential vaccine platforms. We believe that one of the reasonable 14
approaches to resolve these issues is to attenuate Lmdd preferably by an irreversible deletion of 15
one of its virulence factors. Therefore, we hypothesized that an irreversible in frame deletion of 16
actA would increase the safety of Lmdd backbone as an immunotherapeutic vector. 17
In addition to the modification of the Lmdd vaccine backbone, we also generated an 18
improved plasmid (pAdv142) that is retained by the complementation of Bs-dal. The expression 19
of Bs-dal in pAdv142 was under the control of Lm promoter p60. The p60 promoter was chosen 20
due to its constitutive mode of expression in Lm and its ability to function in E. coli. The 21
complementation of Bs-dal in E. coli ala drx makes this plasmid more amenable for further 22
genetic manipulations and cloning purposes. We expressed the tumor antigen PSA in LmddA 23
ACCEPTED

18
background to demonstrate its applicability as antigen delivery system. Initially, we transformed 1
Lmdd and LmddA with pAdv142 resulting in Lmdd142 and LmddA142 and confirmed that the 2
recombinants Lmdd142 and LmddA142 were expressing and secreting the fusion protein LLO-3
PSA using both anti-LLO and anti-PSA antibodies. Further, we established that Lmdd142 and 4
LmddA142 could infect and grow intracellularly by using cell infection assay. The properties 5
such as expression and secretion of fusion protein and intracellular growth are necessary for 6
successful antigen delivery and presentation when using Lm as an immunotherapeutic vector (7). 7
To verify the attenuation of LmddA142, we performed an in vivo virulence and clearance 8
study in wild type and IFN-γ -/-
mice (GKO-/-
). As expected, LmddA142 was at least two logs 9
more attenuated than Lmdd142 (5). Previous reports demonstrate that GKO-/-
mice are extremely 10
sensitive to infection with wild type Lm by three orders of magnitude when compared to wild 11
type mice (8). A dose of 108 CFU of LmddA142 was easily tolerated by GKO
-/- mice, supporting 12
that IFN-γ is not required for the clearance of this strain. LmddA142 was completely cleared 13
from the spleen and liver of the wild type and GKO-/-
mice in 48- 72 h post-injection, suggesting 14
that early innate or developing immune responses may not be vital for the in vivo clearance of 15
LmddA142. On the contrary, immunization with a low dose (2 x 106 CFU) of virulent Lmdd142 16
resulted in an increased persistence of bacteria in the spleen and liver of C57BL/6 mice for up to 17
168 h, due to its ability to spread from cell to cell. These observations suggest that LmddA may 18
serve as safer platform for antigen delivery as compared to Lmdd. 19
We extended our study to compare the therapeutic efficacy of LmddA142 with our 20
previously published Lm counterpart, Lm-LLO-PSA (27). After immunization with LmddA142, 21
60% of tumors were completely eradicated until day 70. On the other hand, 38% of the mice 22
were tumor free in the group immunized with Lm-LLO-PSA, suggesting that LmddA142 was 23
ACCEPTED

19
more efficacious than Lm-LLO-PSA. One of the reasons for this therapeutic effect may be 1
attributed to the administration of 10 fold high dose of LmddA142. This is supported by the 2
observation that antitumor efficacy of LmddA142 and Lm-LLO-PSA was comparable after the 3
administration of similar doses of each immunotherapeutic vector (data not shown). Another 4
live bacterial vaccine, Salmonella enterica serovar Typhimurium aroA strain expressing PSA has 5
been shown to confer protection against PSA expressing tumors (4). However, the therapeutic 6
efficacy for this construct is not determined. It is well established that the efficacy of active 7
immunotherapy using Lm as vaccine vector is largely dependent on the ability of this bacterium 8
to generate potent innate and adaptive immune responses (2, 18, 30). Our data on immunological 9
assessment showed that LmddA142 elicited high levels of PSA specific T cell responses. 10
However, the efficacy of a vaccine is not only dependent on the magnitude of T cells in the 11
periphery but also creating conditions for them to infiltrate into the tumors. Both LmddA142 and 12
Lm-LLO-PSA (27) cause the infiltration of PSA specific T cells in the tumor microenvironment 13
which are required for TPSA23 tumor regression. Another possible mechanism for tumor 14
regression may involve the down modulation of intra-tumoral regulatory T cells (11, 27). 15
Interestingly, immunization with LmddA142 was shown to decrease the population of regulatory 16
T cells in the tumors. 17
Noteworthy, PSA is a foreign antigen in mice and thus, no tolerance is expected towards 18
this antigen. Previously we have verified the ability of Lm-based vaccines to limit autochthonous 19
tumor growth and break immunologic tolerance in HPV16 E6/E7 and Her2/neu transgenic mice 20
using Lm-LLO-E7 and Lm-Her-2/neu vaccines (29, 31). Thus, it is likely that LmddA142 may be 21
able to break tolerance. Moreover other investigators have shown that tolerance against PSA in 22
humans can be overcome by repeated immunizations (6, 10). As an alternative, male 23
ACCEPTED

20
cynomologus monkeys have shown to express PSA protein, which is 89% homologous to human 1
PSA making this species an attractive candidate for future evaluation of LmddA142 2
immunogenicity as well as side effects that might arise upon vaccination against self antigen 3
(15). 4
Overall, our data show that the LmddA is superior to Lmdd as a vaccine vector due to its 5
attenuation and increased safety as evident from mouse studies. The challenge in designing an 6
ideal Lm vaccine for human use has been to achieve a level of attenuation such that the 7
bacterium is less virulent yet retaining its ability to present antigen to the immune system. Here, 8
we confirmed that the limited in vivo survival of LmddA does not compromise its therapeutic 9
potency. Our aim is to move this prototype vaccine described here towards clinical trial in near 10
future. The broader goal of this study is to extend the application of this vector to design 11
potential constructs expressing other single or multiple heterologous antigens. 12
13
Acknowledgements 14
We thank to MHC Tetramer Core Facility at Emory University vaccine Center for providing us 15
with PSA tetramer. The authors wish to thank Dr. Fred Frankel, Department of Microbiology, 16
University of Pennsylvania (UPENN), for providing the Lmdd strain. We wish to acknowledge 17
Dr. John Rothman, Dr. Kyla-Driscoll Carroll and Dr. Gurpreet Singh Kapoor (UPENN) for 18
careful reading of the manuscript and constructive comments. 19
20
References 21
1. Arlen, P. M., J. L. Gulley, C. Parker, L. Skarupa, M. Pazdur, D. Panicali, P. 22
Beetham, K. Y. Tsang, D. W. Grosenbach, J. Feldman, S. M. Steinberg, E. Jones, C. 23
ACCEPTED

21
Chen, J. Marte, J. Schlom, and W. Dahut. 2006. A randomized phase II study of 1
concurrent docetaxel plus vaccine versus vaccine alone in metastatic androgen-2
independent prostate cancer. Clin Cancer Res 12:1260-1269. 3
2. Bruhn, K. W., N. Craft, and J. F. Miller. 2007. Listeria as a vaccine vector. Microbes 4
and infection / Institut Pasteur 9:1226-1235. 5
3. Cunha, A. C., B. Weigle, A. Kiessling, M. Bachmann, and E. P. Rieber. 2006. Tissue-6
specificity of prostate specific antigens: comparative analysis of transcript levels in 7
prostate and non-prostatic tissues. Cancer letters 236:229-238. 8
4. Fensterle, J., B. Bergmann, C. L. Yone, C. Hotz, S. R. Meyer, S. Spreng, W. Goebel, 9
U. R. Rapp, and I. Gentschev. 2008. Cancer immunotherapy based on recombinant 10
Salmonella enterica serovar Typhimurium aroA strains secreting prostate-specific antigen 11
and cholera toxin subunit B. Cancer Gene Ther 15:85-93. 12
5. Goossens, P. L., and G. Milon. 1992. Induction of protective CD8+ T lymphocytes by 13
an attenuated Listeria monocytogenes actA mutant. Int Immunol 4:1413-1418. 14
6. Gulley, J., A. P. Chen, W. Dahut, P. M. Arlen, A. Bastian, S. M. Steinberg, K. 15
Tsang, D. Panicali, D. Poole, J. Schlom, and J. Michael Hamilton. 2002. Phase I study 16
of a vaccine using recombinant vaccinia virus expressing PSA (rV-PSA) in patients with 17
metastatic androgen-independent prostate cancer. Prostate 53:109-117. 18
7. Gunn, G. R., A. Zubair, C. Peters, Z. K. Pan, T. C. Wu, and Y. Paterson. 2001. Two 19
Listeria monocytogenes vaccine vectors that express different molecular forms of human 20
papilloma virus-16 (HPV-16) E7 induce qualitatively different T cell immunity that 21
correlates with their ability to induce regression of established tumors immortalized by 22
HPV-16. J Immunol 167:6471-6479. 23
ACCEPTED

22
8. Harty, J. T., and M. J. Bevan. 1995. Specific immunity to Listeria monocytogenes in 1
the absence of IFN gamma. Immunity 3:109-117. 2
9. He, L., J. Hakimi, D. Salha, I. Miron, P. Dunn, and L. Radvanyi. 2005. A sensitive 3
flow cytometry-based cytotoxic T-lymphocyte assay through detection of cleaved 4
caspase 3 in target cells. Journal of immunological methods 304:43-59. 5
10. Heiser, A., D. Coleman, J. Dannull, D. Yancey, M. A. Maurice, C. D. Lallas, P. 6
Dahm, D. Niedzwiecki, E. Gilboa, and J. Vieweg. 2002. Autologous dendritic cells 7
transfected with prostate-specific antigen RNA stimulate CTL responses against 8
metastatic prostate tumors. The Journal of clinical investigation 109:409-417. 9
11. Hussain, S. F., and Y. Paterson. 2004. CD4+CD25+ regulatory T cells that secrete 10
TGFbeta and IL-10 are preferentially induced by a vaccine vector. J Immunother 27:339-11
346. 12
12. Kocks, C., E. Gouin, M. Tabouret, P. Berche, H. Ohayon, and P. Cossart. 1992. L. 13
monocytogenes-induced actin assembly requires the actA gene product, a surface protein. 14
Cell 68:521-531. 15
13. Lauer, P., M. Y. Chow, M. J. Loessner, D. A. Portnoy, and R. Calendar. 2002. 16
Construction, characterization, and use of two Listeria monocytogenes site-specific phage 17
integration vectors. Journal of bacteriology 184:4177-4186. 18
14. Li, Z., X. Zhao, C. Zhou, B. Gu, and F. R. Frankel. 2006. A truncated Bacillus subtilis 19
dal gene with a 3' ssrA gene tag regulates the growth and virulence of racemase-deficient 20
Listeria monocytogenes. Microbiology (Reading, England) 152:3091-3102. 21
15. Marshall, D. J., L. R. San Mateo, K. A. Rudnick, S. G. McCarthy, M. C. Harris, C. 22
McCauley, A. Schantz, D. Geng, P. Cawood, and L. A. Snyder. 2005. Induction of 23
ACCEPTED

23
Th1-type immunity and tumor protection with a prostate-specific antigen DNA vaccine. 1
Cancer Immunol Immunother 54:1082-1094. 2
16. Mata, M., Z. J. Yao, A. Zubair, K. Syres, and Y. Paterson. 2001. Evaluation of a 3
recombinant Listeria monocytogenes expressing an HIV protein that protects mice 4
against viral challenge. Vaccine 19:1435-1445. 5
17. Noguchi, M., K. Kobayashi, N. Suetsugu, K. Tomiyasu, S. Suekane, A. Yamada, K. 6
Itoh, and S. Noda. 2003. Induction of cellular and humoral immune responses to tumor 7
cells and peptides in HLA-A24 positive hormone-refractory prostate cancer patients by 8
peptide vaccination. The Prostate 57:80-92. 9
18. Pamer, E. G. 2004. Immune responses to Listeria monocytogenes. Nature reviews 10
4:812-823. 11
19. Pan, Z. K., G. Ikonomidis, A. Lazenby, D. Pardoll, and Y. Paterson. 1995. A 12
recombinant Listeria monocytogenes vaccine expressing a model tumour antigen protects 13
mice against lethal tumour cell challenge and causes regression of established tumours. 14
Nature medicine 1:471-477. 15
20. Pan, Z. K., G. Ikonomidis, D. Pardoll, and Y. Paterson. 1995. Regression of 16
established tumors in mice mediated by the oral administration of a recombinant Listeria 17
monocytogenes vaccine. Cancer research 55:4776-4779. 18
21. Pan, Z. K., L. M. Weiskirch, and Y. Paterson. 1999. Regression of established B16F10 19
melanoma with a recombinant Listeria monocytogenes vaccine. Cancer research 20
59:5264-5269. 21
ACCEPTED

24
22. Pavlenko, M., C. Leder, A. K. Roos, V. Levitsky, and P. Pisa. 2005. Identification of 1
an immunodominant H-2D(b)-restricted CTL epitope of human PSA. The Prostate 64:50-2
59. 3
23. Peters, C., and Y. Paterson. 2003. Enhancing the immunogenicity of bioengineered 4
Listeria monocytogenes by passaging through live animal hosts. Vaccine 21:1187-1194. 5
24. Prins, R. M., K. W. Bruhn, N. Craft, J. W. Lin, C. H. Kim, S. K. Odesa, J. F. Miller, 6
and L. M. Liau. 2006. Central nervous system tumor immunity generated by a 7
recombinant listeria monocytogenes vaccine targeting tyrosinase related protein-2 and 8
real-time imaging of intracranial tumor burden. Neurosurgery 58:169-178; discussion 9
169-178. 10
25. Roos, A. K., M. Pavlenko, J. Charo, L. Egevad, and P. Pisa. 2005. Induction of PSA-11
specific CTLs and anti-tumor immunity by a genetic prostate cancer vaccine. The 12
Prostate 62:217-223. 13
26. Sewell, D. A., V. Shahabi, G. R. Gunn, 3rd, Z. K. Pan, M. E. Dominiecki, and Y. 14
Paterson. 2004. Recombinant Listeria vaccines containing PEST sequences are potent 15
immune adjuvants for the tumor-associated antigen human papillomavirus-16 E7. Cancer 16
research 64:8821-8825. 17
27. Shahabi, V., M. Reyes-Reyes, A. Wallecha, S. Rivera, Y. Paterson, and P. Maciag. 18
2008. Development of a Listeria monocytogenes based vaccine against prostate cancer. 19
Cancer Immunol Immunother 57:1301-1313. 20
28. Singh, R., M. E. Dominiecki, E. M. Jaffee, and Y. Paterson. 2005. Fusion to 21
Listeriolysin O and delivery by Listeria monocytogenes enhances the immunogenicity of 22
ACCEPTED

25
HER-2/neu and reveals subdominant epitopes in the FVB/N mouse. J Immunol 1
175:3663-3673. 2
29. Singh, R., and Y. Paterson. 2007. In the FVB/N HER-2/neu transgenic mouse both 3
peripheral and central tolerance limit the immune response targeting HER-2/neu induced 4
by Listeria monocytogenes-based vaccines. Cancer Immunol Immunother 56:927-938. 5
30. Singh, R., and Y. Paterson. 2006. Listeria monocytogenes as a vector for tumor-6
associated antigens for cancer immunotherapy. Expert review of vaccines 5:541-552. 7
31. Souders, N. C., D. A. Sewell, Z. K. Pan, S. F. Hussain, A. Rodriguez, A. Wallecha, 8
and Y. Paterson. 2007. Listeria-based vaccines can overcome tolerance by expanding 9
low avidity CD8+ T cells capable of eradicating a solid tumor in a transgenic mouse 10
model of cancer. Cancer Immun 7:2. 11
32. Stamey, T. A., N. Yang, A. R. Hay, J. E. McNeal, F. S. Freiha, and E. Redwine. 12
1987. Prostate-specific antigen as a serum marker for adenocarcinoma of the prostate. 13
The New England journal of medicine 317:909-916. 14
33. Strych, U., R. L. Penland, M. Jimenez, K. L. Krause, and M. J. Benedik. 2001. 15
Characterization of the alanine racemases from two mycobacteria. FEMS microbiology 16
letters 196:93-98. 17
34. Thompson, R. J., H. G. Bouwer, D. A. Portnoy, and F. R. Frankel. 1998. 18
Pathogenicity and immunogenicity of a Listeria monocytogenes strain that requires D-19
alanine for growth. Infection and immunity 66:3552-3561. 20
35. Verch, T., Z. K. Pan, and Y. Paterson. 2004. Listeria monocytogenes-based antibiotic 21
resistance gene-free antigen delivery system applicable to other bacterial vectors and 22
DNA vaccines. Infection and immunity 72:6418-6425. 23
ACCEPTED

26
36. Watt, K. W., P. J. Lee, T. M'Timkulu, W. P. Chan, and R. Loor. 1986. Human 1
prostate-specific antigen: structural and functional similarity with serine proteases. 2
Proceedings of the National Academy of Sciences of the United States of America 3
83:3166-3170. 4
37. Zhao, X., Z. Li, B. Gu, and F. R. Frankel. 2005. Pathogenicity and immunogenicity of 5
a vaccine strain of Listeria monocytogenes that relies on a suicide plasmid to supply an 6
essential gene product. Infection and immunity 73:5789-5798. 7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
ACCEPTED

27
Figure legends 1
FIG. 1: (A) Schematic map of the plasmid pAdv142. The antigen expression cassette contains 2
hly promoter followed by its signal sequence and 417 amino acids of the N-terminus of 3
hemolytic protein Listeriolysin O (tLLO) fused to human prostate specific antigen, PSA. The 4
PSA protein lacks its secretory signal sequence. (B) Expression and secretion of tLLO-PSA 5
fusion protein by the strains, Lmdd142 and LmddA142. The protein extracts were prepared by 6
precipitating culture supernatants with 10% TCA. The Western blot represents the expression of 7
tLLO-PSA in the total proteins secreted by Lmdd142 and LmddA142. The blot was stained with 8
1:1000 dilution of both anti-LLO and anti-PSA primary antibody. 9
10
FIG. 2: (A) In vivo clearance of the strains, LmddA142 in WT and GKO-/-
mice and (B) Lmdd142 11
in WT mice. Mice were immunized intraperitoneally with 2 x 106 CFU of Lmdd142 and 10
8 12
CFU of LmddA142. The bacterial load in the organs, liver and spleen were determined after day 13
1, 2, 3, 7 and 10 of immunization. Two mice from each group were sacrificed at different days 14
and bacterial CFU was determined by plating several dilutions of the homogenized spleens and 15
liver on BHI plates containing 100 µg/ml of streptomycin. The plates were incubated at 37°C for 16
24-48 h for bacterial growth. Columns, mean of CFU from each mouse; bars, SD. 17
18
FIG. 3: (A) In vitro stability of the plasmid pAdv142 in the strain LmddA142 after 60 19
generations. The in vitro stability was determined by sub-culturing the strain LmddA142 in 20
selective (BHI) and non-selective medium (BHI plus 100µg/ml D-alanine) for 8 days at 1: 21
10,000 dilution at 37°C and 200 rpm. The average CFU + SD was determined for each day after 22
plating on selective and non-selective plates in triplicate. Streptomycin (100 µg/ml) was added to 23
ACCEPTED

28
the culture medium to restrict the growth of any potential contaminant. (B) In vivo stability was 1
examined by immunizing mice with 5 x 107 CFU LmddA142 intravenously in the tail vein. The 2
CFUs were determined in the homogenized spleens after 24, 48 and 72 h. Viable CFUs were 3
determined after plating on both selective and non-selective medium. No colonies were 4
recovered at the time points, 48 and 72 h. Columns, mean of CFU from each mouse; bars, SD. 5
(C) Intracellular growth of the strains, Lmdd142 and LmddA142 in murine macrophage cells, 6
J774A.1. The CFUs were determined by taking duplicate samples at the indicated time points 7
followed by titration on BHI/plates. The data shown is representative of the average CFU at each 8
time point. The experiment was repeated three times and data is representative of one study. 9
10
FIG. 4: (A) Tumor regression study using TPSA23 as transplantable tumor model. Three groups 11
of eight mice were implanted with 2 x 106 tumor cells on day 0 and two groups were immunized 12
with 107 CFU Lm-LLO-PSA (positive control) and 10
8 CFU LmddA142 on day 6, 13 and 20. 13
Naïve mice did not receive any treatment. Tumors were monitored weekly and mice were 14
sacrificed if the average tumor diameter was 14-16 mm. Each symbol in the graph represents the 15
tumors size of an individual mouse. The experiment was repeated twice and similar results were 16
obtained. (B) The percentage survival of the naïve mice and immunized mice at different days of 17
the experiment. 18
19
FIG. 5: PSA specific immune responses were examined by tetramer staining, intracellular 20
cytokine staining for IFN -γ and ELISpot. Mice were immunized twice with 107 CFU Lm-LLO-21
PSA and 108
CFU LmddA142 at one-week interval. For immune assays, spleens were harvested 22
on day 6 after the boost. Spleens from three mice/group were pooled for this experiment. (A) 23
ACCEPTED

29
PSA specific T cells in the spleen of naïve, Lm-LLO-PSA and LmddA142 immunized mice using 1
PSA specific tetramer staining. Cells were stained with mouse anti-CD8 (FITC), anti-CD3 2
(Percp-Cy5.5), anti-CD62L (APC) and PSA tetramer-PE and analyzed by FACS Calibur. (B) 3
Intracellular cytokine staining to detect the percentage of IFN-γ secreting CD8+ CD62L
low cells 4
in the naïve and immunized mice after stimulation with 1 µM of PSA specific, H-2Db peptide for 5
5 h. (C) PSA specific cytotoxic responses in the mice immunized with Lm-LLO-PSA and 6
LmddA142. Mice were immunized twice with 107 CFU Lm-LLO-PSA and 10
8 CFU LmddA142 at 7
a one-week interval and spleens were harvested on day 6 after the boost. The pooled splenocytes 8
from each group were stimulated in vitro for 5 days after incubating with PSA expressing 9
stimulators. The target cells EL4 were pulsed with PSA H-2Db peptide, HCIRNKSVIL and 10
specific lysis was determined after incubating targets with different ratios of effector cells for 3h. 11
Experiment was repeated three times and a representative data is shown. 12
13
FIG. 6: Effect of vaccination of different Listeria constructs on the presence of (A) PSA specific 14
TIL’s and (B) Tregs in the tumors and spleens. Mice were implanted with TPSA23 tumors 15
embedded in matrigel and were immunized with irrelevant Listeria, Lm-LLO-E7 and LmddA142 16
on day 7 and day 14. On day 20, tumors and spleens were harvested from naïve and immunized 17
mice. The tumors were pooled in each group to obtain adequate number of cells for the staining 18
of tumor infiltrating lymphocytes and regulatory T cells. The cells were stained with surface 19
antibodies, CD8, CD3, anti- CD62L and PSA tetramer. To determine the Treg population in both 20
tumor and spleen, the cells were stained for CD3, CD4, CD25 and Foxp3 antibodies. Experiment 21
was repeated three times and representative data are shown. 22
23
ACCEPTED

ACCEPTED

ACCEPTED

ACCEPTED
ACCEPTED

ACCEPTED

ACCEPTED
Copyright © 2022 FDOKUMEN




















