
Constitutively activated phosphatidylinositol -3 kinase (PI -3K) is involved in the defect of...
31
doi:10.1182/blood-2002-02-0539 Prepublished online July 12, 2002; Peschel and Thomas Decker Ingo Ringshausen, Folker Schneller, Christian Bogner, Susanne Hipp, Justus Duyster, Christian δ in the defect of apoptosis in B-CLL: association with Proteinkinase C Constitutively activated phosphatidylinositol-3 kinase (PI-3K) is involved http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: articles must include digital object identifier (DOIs) and date of initial publication. priority; they are indexed by PubMed from initial publication. Citations to Advance online prior to final publication). Advance online articles are citable and establish publication yet appeared in the paper journal (edited, typeset versions may be posted when available Advance online articles have been peer reviewed and accepted for publication but have not Copyright 2011 by The American Society of Hematology; all rights reserved. of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society For personal use only. on April 27, 2014. by guest bloodjournal.hematologylibrary.org From For personal use only. on April 27, 2014. by guest bloodjournal.hematologylibrary.org From
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Constitutively activated phosphatidylinositol -3 kinase (PI -3K) is involved in the defect of...

doi:10.1182/blood-2002-02-0539Prepublished online July 12, 2002;
Peschel and Thomas DeckerIngo Ringshausen, Folker Schneller, Christian Bogner, Susanne Hipp, Justus Duyster, Christian
δin the defect of apoptosis in B-CLL: association with Proteinkinase CConstitutively activated phosphatidylinositol-3 kinase (PI-3K) is involved
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
articles must include digital object identifier (DOIs) and date of initial publication. priority; they are indexed by PubMed from initial publication. Citations to Advance online prior to final publication). Advance online articles are citable and establish publicationyet appeared in the paper journal (edited, typeset versions may be posted when available Advance online articles have been peer reviewed and accepted for publication but have not
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

1
Constitutively activated phosphatidylinositol-3 kinase (PI-3K) is involved in the
defect of apoptosis in B-CLL: association with Proteinkinase Cδδδδ
Short titel: Constitutively activated PI-3 kinase in B-CLL
Ingo Ringshausen*, Folker Schneller*, Christian Bogner*, Susanne Hipp*, Justus
Duyster*, Christian Peschel* and Thomas Decker*
From the * IIIrd Department of Medicine
Technical University of Munich, Munich, Germany
Total word count: 4861 abstract word count: 250
Correspondence to Ingo Ringshausen, M.D.
IIIrd Department of Medicine
Technical university of Munich
Ismaninger Strasse 15
81675 Munich
Tel.: (-49)-89-4140-4110 Fax.: (-49)-89-4140-4879
e-mail: [email protected]
This work was supported by a research grant from the Technical university of Munich
(KKF 15-00)
Copyright 2002 American Society of Hematology
Blood First Edition Paper, prepublished online July 12, 2002; DOI 10.1182/blood-2002-02-0539For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

2
ABSTRACT
In the present study we analyzed the role of phophatidylinositol-3 kinase (PI-3K) in B-
CLL cells. PI-3 kinase is activated by many stimuli and is linked to several different
signaling pathways. We demonstrated that inhibition of PI-3 kinase by a specific
inhibitor LY294002 induced apoptosis in B-CLL cells in vitro. This effect was specific
for the inhibition of PI-3 kinase as inhibition of other signaling pathways like
extracellular signaling-regulated kinase (ERK), p38 or p70S6 kinase did not affect
spontaneous apoptosis. Furthermore PI-3 kinase was constitutively activated in
freshly isolated B-CLL cells. Corresponding to enhanced apoptosis, LY294002
downregulated expression of the antiapoptotic proteins XIAP and Mcl-1. Next we
investigated which factors downstream of PI-3 kinase were activated in B-CLL cells.
We demonstrated that Proteinkinase B/Akt is expressed in all tested CLL samples
but no activation of Akt was detected. In contrast, we observed a constitutive
activation of PKCδ in freshly isolated B-CLL cells. PKCδ is linked to PI-3 kinase and
phosphorylated at threonine 505 in response to PI-3 kinase activation. We further
demonstrated that tyrosinephosphorylation and activity of PKCδ were depending on
PI-3 kinase activity in B-CLL cells. Inhibition of PKCδ by the specific inhibitor Rottlerin
strikingly enhanced apoptosis. In contrast, peripheral blood B-cells of healthy donors
were resistant to inhibition of PI-3K or PKCδ. We conclude that activated PI-3 kinase
might be important in the pathogenesis of B-CLL and survival signals might be
mediated via PKCδ. Therefore inhibition of PI-3 kinase or PKCδ may be an innovative
approach to treat B-CLL.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

3
INTRODUCTION
Chronic lymphocytic leukemia (CLL) is the most common type of leukemia in the
western world. It is characterized by the accumulation of monoclonal CD 5+ mature
B-cells, with a high percentage of cells arrested in G0/G1 cell cycle phase (1). A
profound defect in programmed cell death as well as in cell cycle progression is a
major step in the pathogenesis of B-CLL (2). Despite the development of new
chemotherapeutic drugs like purine nucleotides the disease remains incurable and
progression occurs in every patient after an unpredictable clinical course. A variety of
chromosomal aberrations have been described such as 13q14 and 11q22 deletions
and trisomy 12 (3), (4), but no general molecular defect has been found and little is
known about the molecular pathogenesis of the disease. Apoptosis resistance has
been associated with high levels of the antiapoptotic protein bcl-2 in many B-CLL
samples (5). In addition, overexpression of the antiapoptotic protein Mcl-1 in B-CLL
has been correlated with failure to achieve complete remission through
chemotherapy (6). However, it is still unknown which factors are responsible for the
disproportion of pro- and antiapoptotic proteins in B- CLL.
There is evidence for the existence of constitutively activated signaling pathways in
CLL, as Frank et al. identified constitutive serine phosphorylation of STAT1 and
STAT3 (7). Furthermore, B-CLL cells contain high levels of NF-κB activity (8), but
involved upstream activators still need to be determined. The biological meaning of
constitutively activated signaling pathways in B-CLL is largely unknown.
Phosphatidylinositol-3 kinase (PI-3K) is an enzyme whose inositol lipid products are
key mediators of several distinct intracellular pathways. PI-3 kinase consists of a 110-
kD catalytic subunit and a tightly associated regulatory subunit p85 (9). PI-3 kinase is
involved in several signaling transduction pathways in B-cells like CD40 signaling
(10), BCR-signaling (11) and signaling of a variety of cytokines. It has been shown
that PI-3 kinase activates the serine/threonine kinase Akt/Proteinkinase B (PKB) (12).
Akt binds to the products of PI-3 kinase, phosphatidylinositol 3,4-bisphosphate and
phosphatidylinositol 3,4,5-trisphosphate. Then Akt becomes activated by
phosphorylation at threonine 308 and serine 473 through the action of the 3-
phosphoinositide-dependent kinase PDK1 (13). One major activity of Akt is to
mediate cell survival in a broad spectrum of cells. Akt overexpression has been found
in cancer cells of pancreatic, ovarian or breast origin (14), (15). Antiapoptotic signals
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

4
mediated by Akt include phosphorylation of the bcl-2 counterpart bad, inhibition of
caspase-9 (16), activation of NF-κB (17) and p70S6-kinase (18). PI-3 kinase
signaling is also linked to novel and atypical proteinkinase C (PKC) isoforms (19).
PKC is a family of isoenzymes classified by their dependence on cofactors and may
be regulated by several independent mechanisms. We investigated PKCδ expression
and activation in B-CLL cells, because this enzyme is expressed in lymphocytes,
activated in response to B-cell receptor ligation (20) and associates with PI-3 kinase
in response to cytokine stimulation of hematopoetic cells (21). PKCδ is involved in
regulation of the cell cycle and the programmed cell death: Overexpression of PKCδcauses cell cycle arrest in a variety of cell types whereas cell cycle transition is not
affected by overexpression of other PKC isoforms (22), (23). PKCδ inhibits the
expression of cycline D1 and cycline E accompanied with an up-regulation of p27 in
vascular smooth muscle cells (24). PKCδ-deficient mice show autonomous
hyperproliferation of B cells, suggesting that PKCδ negatively regulates B cells
proliferation (25). Other findings have shown that PKCδ also might play a role in
apoptosis. In cells undergoing apoptosis, PKCδ is cleaved by caspase 3 and this
cleaving frangment is sufficient to induce apoptosis itself (26). Because of its
ambivalent effects on cell cycle progression and apoptosis PKCδ might also be
important in CLL.
In the present work we have shown for the first time that PI-3 kinase is constitutively
activated in B-CLL cells and that inhibition of PI-3 kinase induces apoptosis in a way
that is independent of the downstream kinase Akt. We further demonstrate that PKCδis activated in freshly isolated B-CLL cells and that inhibition of this enzyme strikingly
induces apoptosis in contrast to normal peripheral blood B-cells. Our findings suggest
that the PI-3 kinase/ PKCδ pathway plays an important role in the defect of
programmed cell death in B-CLL cells.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

5
MATERIAL and METHODS
Cell samples:
After informed consent, peripheral blood was obtained from patients with a diagnosis
of B-CLL according to clinical and immunophenotypic criteria. Patients were either
untreated or had not received chemotherapy/ steroids for a period of at least three
month prior to investigation. At the time of analysis all patients were clinically stable,
free from infectious complications and undergoing routine clinical outpatient review.
Control samples were obtained from peripheral blood of healty donors.
Reagents and antibodies:
Monoclonal antibodies specific for bcl-2, XIAP, PKCδ were purchased from BD
Transduction laboratories (Heidelberg, Germany); MAb against Akt, phospho(473)-
Akt, phosphothreonine(505)-PKCδ were obtained from New England Biolabs
(Schwalbach, Germany). Specific MAb against caspase-3 and bax were from
Pharmingen (San Diego, USA), against β-actin from Sigma; Deisenhofen, Germany.
MAb specific for Mcl-1 was purchased from Santa Cruz biotechnology (Santa Cruz,
USA) and for anti-phosphotyrosine, clone 4G10 from upstate biotechnology (Lake
placid, USA). The proteinkinase inhibitors LY294002, wortmannin, Rottlerin, SB
203580, PD 98059 and Rapamycin were obtained from Calbiochem (Schwalbach,
Germany) as well as the caspase inhibitor zvad.fmk and caspase-3 inhibitor z-devd-
fmk.
Separation procedures:
Peripheral blood mononuclear cells (PBMNC) were isolated from heparinized blood
samples by centrifugation over a Ficoll-Hypaque layer (Biochrom, Berlin, Germany)
of 1.077 g/ml density. For separation of CLL B-cells, PBMNC were incubated with
anti-CD2 and anti-CD14 magnetic beads (Dynabeads M450, Dynal, Oslo, Norway)
according to the manufracturer´s instructions. Such prepared B cells from CLL
patients were >98% pure as assessed by direct immunofluorescence using Coulter
Epics XL (Coulter, Hamburg, Germany). For separation of peripheral blood B-cells of
healthy donors PBMNC were incubated with anti-CD19 magnetic beads and isolated
B-cells were released from CD19 beads by using CD19 DETACHaBEAD (Dynal
biotech, Oslo, Norway ) according to manufracturer´s instructions.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

6
Culture conditions:
Purified normal and leukemic B cells were cultured in RPMI 1640 medium (Biochrom)
supplemented with 10% fetal calf serum (Biochrom), penicillin/streptomycin 50 IU/ml,
Na-pyruvate 1mM, L-glutamine 2mM, L-asparagine 20µg/ml, 2-mercaptoethanol
0.05mM, HEPES 10mM and MEM non-essential amino-acids 0.7x (Biochrom) at
37°C and 5% CO2 in a fully humidified atmosphere in 6 well plates at 1*106 cells/ml.
Analysis of apoptosis:
All flow cytometry analyses were made on a Coulter Epics XL cytoflourometer.
Analysis of annexin V binding to phosphatidylserine (PS) on the cell surface: Cells to
be examined for annexin V expression were washed with PBS and resuspended in
500µl binding buffer (Annexin V-FITC Kit, Immunotech), containing 1µl of Annexin V-
FITC stock and 5µl of 20µg/ml PI to determinate the PS exposure on the outer
plasma membrane. After incubation for 10 minutes at room temperature in a light
protected area, the specimens were quantified by flow cytometry, acquiring 5.000
events.
Analysis of mitochondrial membrane potential: Incorporation of the cationic lipophilic
dye DiOC6 into the mitochondria is proportional to the mitochondrial transmembrane
potential ∆ψ. Cells were incubated for 30 minutes with 20nM DiOC6(3) at 37°C, then
washed once in PBS and subsequently resuspended in 500µl binding buffer (Annexin
V-FITC kit, Immunotech), containing 5µl of 20µg/ml PI. Cells were analyzed via flow
cytometry, aquiring 5.000 events.
PI-3 kinase assay:
PI-3 kinase activitiy in CLL and in Ba/F3 cells was measured as described in (27)
with minor modifications. 200µg of protein from total cell lysate were
immunoprecipitated with anti-p85 PI-3 kinase antibody in the lysis buffer for 2 hours
at 4°C. Immunprecipitates were collected by adding protein A-sepharose beads for 1
hour at 4°C with gently agitation. Then beads were washed twice in lysis buffer,
twice in buffer containing 0.1mol/L Tris/ HCl (ph7.4) and 0.5 mol/L LiCl, and finally in
0.15 mol/L NaCl and 10 mmol/L Tris/ HCl (pH7.4) and 5 mmol/L EDTA. Kinase
reaction was performed for 15 minutes at 37°C in the presence of 0.4 mg/mL
phosphatidylinositol (Sigma), 40 µmol/L ATP, 30 mmol/L MgCl2, and 0.37 MBq
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

7
(10µCi) of [γ-32P] ATP. The reaction was stopped by adding 20µL of 6 N HCl and the
lipids were first extracted with 160µL chloroform/ methanol (1:1; vol/vol). The lipids in
the organic phase were separated by thin-layer chromatography (TLC) using a silica
gel 60 (Merck, Darmstadt, Germany) with chloroform/ methanol / H20 / NH4OH
(43:38:7:5; vol/vol) and visualized by autoradiography.
Immunoblotting:
To investigate cell proteins via Western Blot 1 to 10 x107 cells were cultured in
medium alone or together with LY294002 (10µM) or Rottlerin (5µM). After 12 and 24
hours cells were collected, washed twice in PBS, and were lysed in lysis buffer
(10µM Tris / HCl (ph7,4), 5 mM EDTA, 130 mM NaCl, 1% Triton, 1mM
phenylmethylsulfonyl fluoride, 1mM Na3VO4 and 10mg/ml of each phenantroline,
aprotinin, leupeptin and pepstatin) for 20 minutes at 4°C. Lysates were spun at
12.000 rpm for 20 minutes and supernatants were collected. Protein concentration
was assessed by the Bio-Rad assay method (Bio-Rad Laboratories, Richmond,
Canada). Total extracts (40 to 100µg / lane) were subjected to SDS-Page and
blotting was performed on PVDF membranes (Immobilon-P, Millipore GmbH,
Germany). Blots were developed using a Super Signal chemoluminescent
substrates from Piere Chemical Company (KMF GmbH, Germany).
Akt Kinase assay:
Nonradioactive Akt kinase assay kit was purchased from New England Biolabs
(Schwalbach, Germany). After immunoprecipitation of Akt, kinase reaction was
performed according to the manufracturer´s instructiones using GSK-3 fusion protein
as an exogenous substrate. The kinase reaction was analyzed by immunoblotting,
using a phospho-GSK-3 antibody (Ser21/9).
PKCδ kinase assay:
Cell lysates were immunoprecipitated with an anti-PKC δ antibody (BD Transduction
Laboratories, Heidelberg, Germany) and immunoprecipitates were washed three
times with phosphorylation lysis buffer and three times with kinase buffer (25mM Tris-
HCl [pH 7.4], 5 mM MgCl2, 0.5 mM EGTD, 1mM DTT, 20µg of phosphatidylserine, 20
µM ATP) and were resuspended in 25 µl of kinase buffer containing 5 µg Histone H1
(upstate biotechnology, Lake placid) as an exogenous substrate, to which 20µCi [γ-
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

8
32P] ATP was added. The reaction was incubated for 20 minutes at room
temperature and was terminated by addition of SDS-sample buffer. Proteins were
analyzed by SDS-PAGE and phosphorylated form of Histone H1 was detected by
autoradiography.
Statistical analysis:
Data from individual experiments are presented as mean +/- SEM. Statistical
significances were determined using the Wilcoxon signed rank test and the Mann-
Whitney test as appropriate. A p-value < 0.05 was considered to be statistically
significant.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

9
RESULTS
Inhibition of PI-3 kinase in B-CLL cells induces apoptosis in a dose dependent
manner
We investigated the effect of inhibtion of PI-3 kinase in B-CLL cells by its specific
inhibitor LY294002 (28) in vitro. Cells were incubated with or without the inhibitor and
apoptosis was measured by annexin/PI-staining after 24 hours (Figure 1A). Inhibition
of PI-3 kinase resulted in an increased amount of apoptotic cells compared to
spontaneous apoptosis, which ranged from 3 to 67%. An increased number of
apoptotic cells could be detected in 24 out of 24 tested samples [(34+/-3.5% versus
58+/-3.6%; p<0.0001); Figure 1B]. We also considered that the concentration of
cultivated CLL cells is critical for ex vivo survival of cells, depending on cell- cell
interactions (29). Therefore we tested the impact of LY294002 on increasing
concentrations [1 to 6*106 cells/ml] of cultivated CLL cells. Apoptosis responsiveness
to LY294002 was not affected by the concentration of cells ex vivo (data not shown).
A dose-response analysis revealed an increase of apoptotic cells beginning at a
concentration of 1µM LY 294002 with a maximum at 100µM (Figure 1C). Because
nonspecific and toxic effects cannot be ruled out at 100µM, we chose a working
concentration of 10µM, which is known to be specific for inhibiton of PI-3 kinase. We
next performed DiOC6(3) staining of the cells in the absence or in the presence of
10µM LY294002. Inhibition of PI-3 kinase strikingly led to a loss of the mitochondrial
membrane potential (∆ψ). One representative experiment is shown in figure 1D,
demonstrating an increase of DiOC6(3) negative, apoptotic cells from 10 to 50% with
LY294002 after 24 hours. We confirmed the proapoptotic effect of PI-3 kinase
inhibition by TUNEL-assay (data not shown). Next we examined the effect of
LY294002 on peripheral blood B-cells of healthy donors. No increase in the number
of apoptotic cells was detected after 24 hours of incubation with 10µM LY294002
(Figure 1E). To exclude the possibility that the difference in apoptosis
responsiveness to LY294002 between normal B cells and B-CLL cells was related to
different purification processes, we purified CLL cells either by positive or by negative
selection. CD19-positive selection of B-CLL cells did not affect the suceptibility of
cells towards apoptosis induction by LY294002 (data not shown).
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom
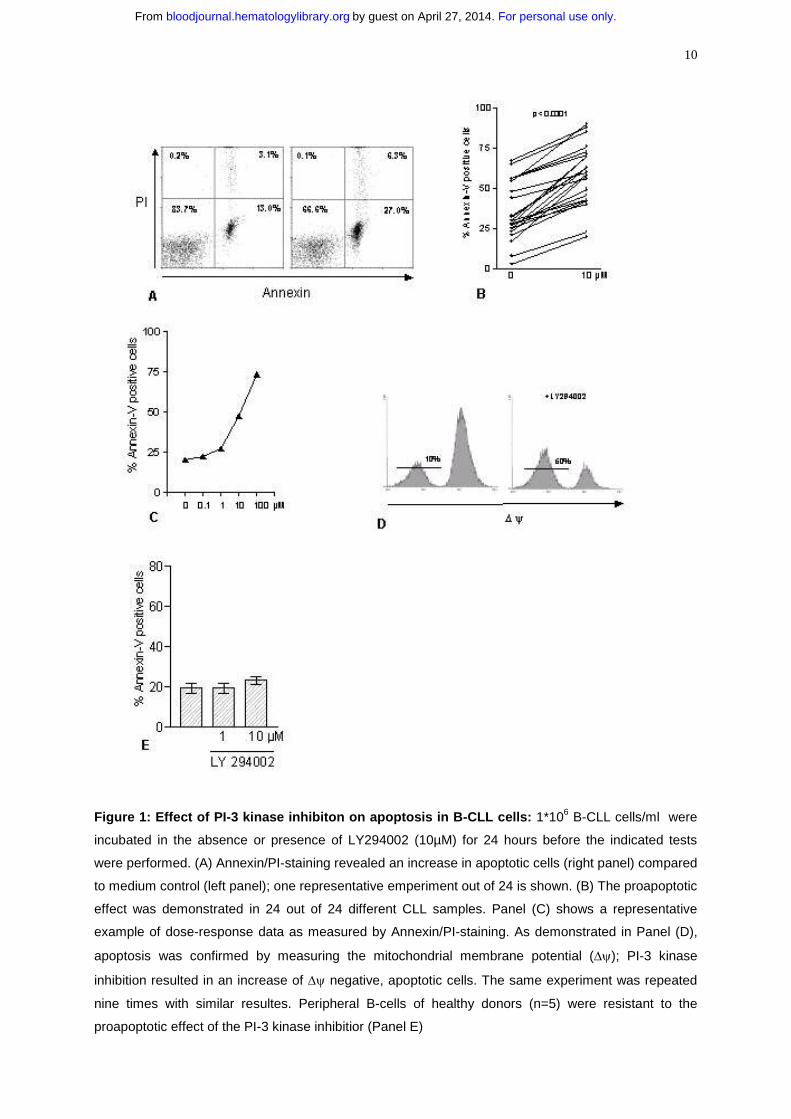
10
Figure 1: Effect of PI-3 kinase inhibiton on apoptosis in B-CLL cells: 1*106 B-CLL cells/ml were
incubated in the absence or presence of LY294002 (10µM) for 24 hours before the indicated tests
were performed. (A) Annexin/PI-staining revealed an increase in apoptotic cells (right panel) compared
to medium control (left panel); one representative emperiment out of 24 is shown. (B) The proapoptotic
effect was demonstrated in 24 out of 24 different CLL samples. Panel (C) shows a representative
example of dose-response data as measured by Annexin/PI-staining. As demonstrated in Panel (D),
apoptosis was confirmed by measuring the mitochondrial membrane potential (∆ψ); PI-3 kinase
inhibition resulted in an increase of ∆ψ negative, apoptotic cells. The same experiment was repeated
nine times with similar resultes. Peripheral B-cells of healthy donors (n=5) were resistant to the
proapoptotic effect of the PI-3 kinase inhibitior (Panel E)
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom
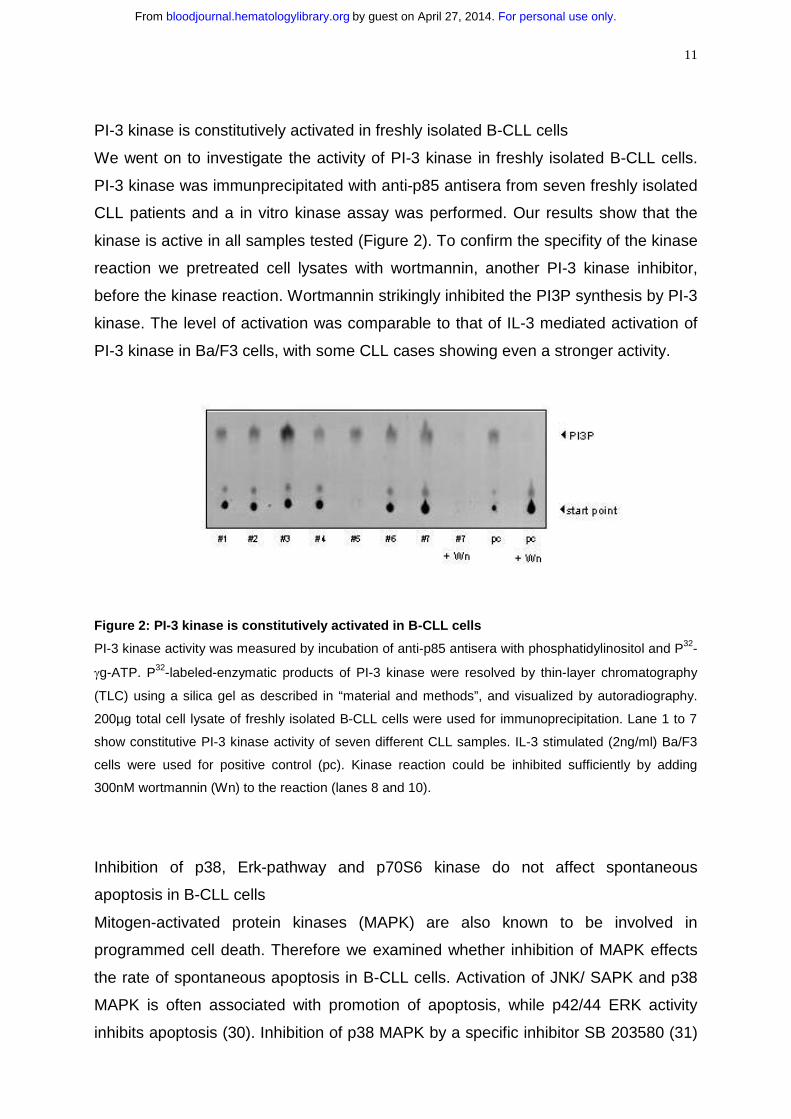
11
PI-3 kinase is constitutively activated in freshly isolated B-CLL cells
We went on to investigate the activity of PI-3 kinase in freshly isolated B-CLL cells.
PI-3 kinase was immunprecipitated with anti-p85 antisera from seven freshly isolated
CLL patients and a in vitro kinase assay was performed. Our results show that the
kinase is active in all samples tested (Figure 2). To confirm the specifity of the kinase
reaction we pretreated cell lysates with wortmannin, another PI-3 kinase inhibitor,
before the kinase reaction. Wortmannin strikingly inhibited the PI3P synthesis by PI-3
kinase. The level of activation was comparable to that of IL-3 mediated activation of
PI-3 kinase in Ba/F3 cells, with some CLL cases showing even a stronger activity.
Figure 2: PI-3 kinase is constitutively activated in B-CLL cells
PI-3 kinase activity was measured by incubation of anti-p85 antisera with phosphatidylinositol and P32-
γg-ATP. P32-labeled-enzymatic products of PI-3 kinase were resolved by thin-layer chromatography
(TLC) using a silica gel as described in “material and methods”, and visualized by autoradiography.
200µg total cell lysate of freshly isolated B-CLL cells were used for immunoprecipitation. Lane 1 to 7
show constitutive PI-3 kinase activity of seven different CLL samples. IL-3 stimulated (2ng/ml) Ba/F3
cells were used for positive control (pc). Kinase reaction could be inhibited sufficiently by adding
300nM wortmannin (Wn) to the reaction (lanes 8 and 10).
Inhibition of p38, Erk-pathway and p70S6 kinase do not affect spontaneous
apoptosis in B-CLL cells
Mitogen-activated protein kinases (MAPK) are also known to be involved in
programmed cell death. Therefore we examined whether inhibition of MAPK effects
the rate of spontaneous apoptosis in B-CLL cells. Activation of JNK/ SAPK and p38
MAPK is often associated with promotion of apoptosis, while p42/44 ERK activity
inhibits apoptosis (30). Inhibition of p38 MAPK by a specific inhibitor SB 203580 (31)
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

12
did not decrease the number of apoptotic cells even at a concentration of 5µM
(Figure 3A) as measured by Annexin/PI staining. To answer the question if the ERK
pathway is involved in apoptosis in B-CLL cells in vitro we also incubated cells with
increasing concentrations of PD98059, a specific inhibitor of the MEK/ERK pathway.
PD98059 acts by blocking activation of ERK by the upstream MAPK kinases MEK-1
and MEK-2 (32). Even at the very high concentration of 500µM, PD98059 did not
increase spontaneous apoptosis (Figure 3B). P70S6 kinase lies downstream of PI-3
kinase and Akt (33). Therefore, we investigated whether inhibition of p70S6 kinase
could mimic the proapoptotic effect of LY294002. The macrolide rapamycin blocks
the activity of mTOR (mammilian target of rapamycin) which is an upstream activator
of p70S6 kinase. As shown in figure 3C rapamycin did not affect the spontaneous
apoptosis of B-CLL cells in vitro. Therefore, neither MAP kinases nor p70S6 kinase
seems to be involved in the defect of apoptosis in B-CLL cells.
Figure 3: P38- and Erk-MAPK as well as p70S6kinase are not involved in spontaneous
apoptosis of B-CLL cells: 1*106 cells/ml were incubated with increasing amounts of SB203580, a
specifc inhibitor of p38-MAPK (A) ; PD98059, which inhibits Erk-MAPK pathway (B) or Rapamycin,
which blocks p70S6kinase (C). Apoptosis was measured by Annexin/PI-staining after incubation for
24 hours. One representative result out of 10 individual experiments is shown.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

13
LY 294002 induced apoptosis can be antagonized by zvad.fmk, a pan-caspase-
inhibitor and by the caspase-3 inhibitor z-devd-fmk.
To confirm the involvement of caspases in LY294002-induced apoptosis we
investigated the impact of caspase inhibitors on B-CLL cells. Zvad.fmk is a broad
spectrum caspase inhibitor targeting caspases I, III,IV and VII (34). As shown in
figure 4 preincubation of B-CLL cells with 100µM zvad.fmk completely antagonized
LY294002 induced apoptosis. Moreover, the level of apoptotic cells was below the
spontaneous rate of apoptosis even in the presence of 10µM LY294002. A protective
effect of zvad.fmk on spontaneous apoptosis was published by Bellosillo et al.
investigating the role of caspases in apoptosis of B-CLL cells (35). We next
investigated the role of caspase-3 by using the caspase-3 inhibtor z-devd-fmk.
Inhibition of caspase-3 also protected cells from LY294002 induced apoptosis.
Figure 4: Antagonisation of the proapoptotic effect of LY294002 by the pan-caspase-inhibitor
zvad.fmk and the caspase-3 inhibitor z-devd-fmk: B-CLL cells were incubated in medium alone or
with 10µM LY294002. Cells were preincubated either with the broad spectrum caspase inhibitor
zvad.fmk (100µM) or the caspase-3 inhibitor z-devd-fmk (200µM) 30 minutes prior to adding
LY294002 to the culture medium. Apoptosis was detected by Annexin/PI- staing after 24 hours. One
representative experiment out of three is shown.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

14
Inhibition of PI-3 kinase results in down-regulation of antiapoptotic proteins
High levels of the antiapoptotic proteins bcl-2 and Mcl-1 are commonly found in
circulating B-CLL cells and might play an important role in the pathophysiology of the
disease. Therefore, we studied the expression of bcl-2 and its counterpart bax, and
Mcl-1 during in vitro culture with or without LY294002. As shown in figure 5, freshly
isolated B-CLL cells expressed high levels of bcl-2, but we did not detect any change
in the level of this protein during the time course of 24 hours with or without
LY294002. In contrast to bcl-2, incubation of cells with LY294002 induced a
proteolytic degradation of its counterpart bax. The proapoptotic bax cleavage product
became detectable within 12 hours. Mcl-1 is also a member of the bcl-2 family and
high levels have been correlated with failure to achieve complete remission to
chemotherapy in CLL. Inhibition of PI-3 kinase resulted in a complete loss of the
Mcl-1 protein compared to resting B-CLL cells, but a spontaneous decrease also was
observed when cells were incubated in medium alone. Finally, XIAP is a member of
the inhibitor of apoptosis (IAP), and because of its ability to directly inhibit some
members of the caspase family (36) it may be important in CLL. Freshly isolated B-
CLL cells expressed high levels of XIAP and no spontaneous decrease in cells
cultured for 24 hours in medium was observed. In contrast, incubating cells with
LY294002 resulted in a decreased XIAP expression. As caspases are known to
mediate important steps in the apoptotic pathway we next investigated procaspase-3
activity. A procaspase-3 cleavage fragment was detected in resting B-CLL cells as
well as in cells incubated with LY294002, but inhibiton of PI-3 kinase resulted in a
stronger cleavage of procaspase 3 (Figure 5).
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

15
Figure 5: Modulation of pro- and antiapoptotic proteins in response to PI-3 kinase inhibition:
Representative immunoblot data show regulation of pro- and antiapoptotic proteins. B-CLL cells were
cultured with or without 10µM LY294002. Cells were harvested after 12 and 24 hours and whole cell
lysates were prepared. A protein content of 40µg was subjected to SDS-PAGE/ immunoblot analysis
by the use of specific antibodies for Bax, Bcl-2, XIAP, Mcl-1 and caspase3.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

16
Proteinkinase B/ Akt is not constitutively activated in B-CLL cells
Proteinkinase B/Akt is activated by PI-3 kinase via the 3-phosphoinosititde-
dependent kinase PDK1. Because of its important role to mediate cell survival, we
investigated whether Akt is constitutively activated in freshly isolated B-CLL cells.
Maximal activation of Akt requires phosphorylation of the residues threonine 308 and
serine 473 (37). As shown in figure 6A, Akt is expressed in all samples tested but no
phosphorylation of Akt at serine 473 was detected. Furthermore we did not detect
any phosphorylation of threonine 308 (data not shown). Next we perfomed an Akt-in
vitro kinase assay to confirm that Akt is not active in B-CLL cells. Again we failed to
detect any activity of Akt (Figure 6B). To rule out the possibility of an in vitro
activation of Akt we incubated cells for 24 hours with or without LY294002 and
repeated western blot. Akt expression remained stable during the time course of 24
hours but no phosphorylation occured in vitro (Figure 6C).
Figure 6: Proteinkinase B/Akt is expressed, but not constitutively phosphorylated in B-CLL:
B-CLL cells were lysed immediately after separation from peripheral blood. (A) Whole protein lysates
(100µg) from seven different patients were analyzed by immunoblotting with the antibodies as
indicated. NIH-3T3 cells treated with platelet derived growth factor (PDGF) severed as a positive
control (pc), without PDGF as a negtative control (nc). (B) Akt-in vitro kinase assay was performed
after immunoprecipitation of Akt from seven different patients; GSK-3 fusion protein served as
exogenous substrate for Akt. Kinase reaction was analyzed by immunobloting with mAb specific for
phospho-GSK-3 (Ser21/9). (C) To rule out in vitro phosphorylation of Akt, B-CLL cells were incubated
with or without 10µM LY294002 and lysed after 12 and 24 hours.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

17
Proteinkinase Cδ is constitutively activated in freshly isolated B-CLL cells:
Dependence on PI-3 kinase activity
PI-3 kinase signaling is also linked to novel and atypical proteinkinase C (PKC)
isoforms. One novel isoform is PKCδ, which is involved in the regulation of cell cycle
and apoptosis. PKCδ is associated with PI-3 kinase following cytokine stimulation
and is activated by PDK1 at threonine 505. Therefore we examined whether PKCδ is
constitutively activated in B-CLL cells. Cell lysates of freshly isolated B-CLL cells
were immunoprecipitated with an antibody against PKCδ, and in vitro kinase assay
was carried out on immunoprecipitates using Histone H1 as an exogenous substrate.
We detected a constitutive activation of PKCδ on eight different samples; kinase
activity was inhibited by the addition of Rottlerin (5µM) to the kinase reaction (Figure
7A, lane 9). Next we examined whether PKCδ was also constitutively phosphorylated
on threonine 505. Figure 7B shows constitutive phosphorylation of PKCδ at threonine
505. The level of phosphorylation was different in individual samples. To investigate
the dependence of tyrosine-phosphorylation of PKCδ on PI-3 kinase activity we
incubated cells with LY294002 over a time period of 12 hours and measured
tyrosine-phosphorylation after PKCδ immunoprecipitation. As shown in figure 7C
inhibition of PI-3 kinase led to a reduction of PKCδ tyrosine-phosphorylation,
becoming detectable after one hour. Accordingly, PKCδ kinase activity declined after
PI-3 kinase inhibition by LY294002 as shown in Figure 7D. These data demonstrate
that Rottlerin inhibits PKCδ kinase activity and that inhibition of PI-3 kinase negatively
affects tyrosine-phosphorylation and activity of PKCδ in B-CLL cells.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

18
Figure 7: PKCδδδδ is constitutively activated in B-CLL cells: (A) 300µg total protein content of eight
different freshly isolated B-CLL cells were immunprecipitated with an antibody against PKCδ and in
vitro kinase assay was performed -in the absence (lane 1-8) or presence (lane 9 ) of Rottlerin- using
Histone H1 as an exogenous substrate. Proteins were analyzed by SDS-PAGE and a phosphorylated
form of Histone H1 was detected by autoradiography. (B) 100µg total protein content of seven different
B-CLL cells were analyzed by immunoblot with an anti-phosphothreonine(505)-antibody specific for
PKCδ. Samples from figure 7A were different from those shown in figure 7B. To investigate the
influence of PI-3 kinase activity on PKCδ cells were incubated with 10µM LY294002 and lysed after
the indicated time period. Then PKCδ was immunoprecipitated with a specific antibody: (C) Tyrosine-
phosphorylation was detected by using an antiphosphotyrosine (4G10)-antibody. (D) PKCδ kinase
activity was measured by a in vitro kinase assay as described above.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

19
Inhibition of PKCδ by Rottlerin induces apoptosis in B-CLL cells and not in peripheral
B-cells of normal controls
We next analyzed the influence of inhibition of PKCδ on apoptosis in B-CLL cells.
Cells were incubated with or without the PKCδ inhibitor Rottlerin (38) for 24 hours
prior to measuring apoptosis. Interestingly, Rottlerin induced apoptosis in B-CLL cells
(Figure 8A) and most times the effect was even stronger compared to PI-3 kinase
inhibition (data not shown). To support the importance of PKCδ and the specifity of its
inhibitor Rottlerin we repeated the experiments with the protein kinase C inhibitor Gö
6976 , which is known to specificly inhibit the Ca2+- dependent PKC isoforms -α,
and -β (39) ; Gö 6976 did not show any impact on the percentage of apoptotic B-CLL
cells in vitro (data not shown). Next we investigated the impact of PKCδ inhibiton on
peripheral B-cells of healthy donors. In contrast to B-CLL cells, Rottlerin did not
induce apoptosis in normal B-cells. Furthermore we saw a trend towards an
antiapoptotic effect on non-malignant cells, but no statistical significance was
reached [(p=0.125); Figure 8B].
Figure 8: Influence of PKCδδδδ inhibtion on viability of B-CLL cells: 1*106 cells/ml were incubated
with or without Rottlerin (5µM), which specificly inhibits PKCδ. After 24 hours apoptosis was measured
by Annexin/PI-staining. Panel (A) shows the result of 12 different experiments. The proapototic effect
was compared to peripheral B-cells of healthy donors (n=5); (B).
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

20
DISCUSSION
Constitutively activated signaling pathways are a common finding in hematological
malignancies (40), (41), (42). They might also play an important role in the
disturbance of apoptosis in B-CLL. Important features are the high level of NF-κB
activity in unstimulated CLL cells in comparison to nonmalignant human B cells (8) as
well as constitutive phosphorylation of STAT1 on serine 727.
In the present work we have shown for the first time that PI-3 kinase is constitutively
activated in CLL cells and that specific inhibition of this kinase increases apoptosis in
B-CLL cells. PI-3 kinase is a ubiquitously expressed protein kinase that is involved in
the regulation of normal and neoplastic cell growth. Many factors which are important
for the development and survival of normal B-cells use the PI-3 kinase pathway,
including CD40 and BCR-ligation (11). Mice that lack the p85 subunit of PI-3 kinase
exhibit profound defects of B-cell development, as well as a diminished proliferation
response and survival (43). Some of the cytokines that protect B-CLL cells from
apoptosis, such as IL-4, transduce their effects through PI-3 kinase in normal B-cells
(44). Recently it has been published that survival effects of IL-4 and the phorbol ester
TPA on B-CLL cells are mediated via the PI-3 kinase pathway (45). Because of its
importance in many critical signaling pathways it is not surprising that the PI-3 kinase
pathway is involved in the development of solid tumors (46) and hematological
malignancies such as multiple myeloma (47). Several different possibilities have to
be taken into consideration to explain the constitutive activation of PI-3 Kinase in B-
CLL cells: (I) PI-3 kinase activity is related to an intrinsic defect of B-CLL cells leading
to a permanent activation of downstream anti-apoptotic factors or (II) PI-3 kinase is
activated by an extrinsic, humoral factor mediating survival signals to the cell. (III) PI-
3 kinase regulation is disturbed in CLL cells resulting in a long lasting activation after
receptor-ligation. The resistance of normal B-cells towards apoptosis induced by PI-3
kinase inhibition demonstrates that kinase activation is critical for B-CLL cells but not
for normal resting peripheral B-cells. This difference is supported by the results of
Aagaard-Tillery et al. in human B-cells, as they failed to detect any kinase activity in
unstimulated peripheral B-cells; PI-3 kinase activity only became detectable after
cross-linking of membrane bound Ig or CD40-ligation (11).
Despite the crucial role of PI-3 kinase with respect to proliferation and survival of B-
cells, little is known about the events following PI-3 kinase activity. The serine/
threonine kinase Akt, or proteinkinase B, is the best characterized downstream
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

21
kinase of PI-3 kinase. The importance of Akt in CLL cells with respect to protection of
apoptosis has recently been investigated. Autologous plasma has been shown to
protect B-CLL cells from spontaneous or cytotoxic induced apoptosis; this relies on a
PI-3 kinase/ Akt dependent pathway (48). In contrast to other reports, Bernal et al.
have demonstrated that engagement of surface IgM on B-CLL cells also improves
cell survival; this occurs via activation of PI-3 kinase and phosphorylation of Akt (49).
In this recently published work no toxic effect or change in viability was observed
when CLL cells were preincubated with LY294002 alone, even at a higher
concentration of 75µM. This appears to be in conflict with our present work; one
explanation for this discrepancy can be the time of preincubation with the PI-3 kinase
inhibitor prior to measuring apoptosis. In that particular experiment the time of
incubation was not described. In our experiments we failed to detect any proapoptotic
effect of LY294002 before 8 hours of incubation (data not shown). Another
explanation might be the employed method of measuring apoptosis; PI-staining might
be too insensitive to reveal slight differences in the number of apoptotic cells. Again,
as mentioned above, we confirmed the proapoptotic effect of PI-3 kinase inhibiton by
several different methods. Although PI-3 Kinase/ Akt is involved in survival signals in
CLL cells mediated by factors which are known to use the PI-3K / Akt pathway, our
results have shown that Akt is not constitutively activated due to the constitutive PI-3
kinase activity.
In the past observations have been made showing that PI 3,4-P2 and PI 3,4,5-P3 can
also activate novel and atypical Proteinkinase C isoforms in vitro (19). This occurs in
context with a marked increase in the autophosphorylation of PKC δ, - ε, and -η. With
respect to PKC δ it has been demonstrated that the PKB kinase PDK1 is responsible
for phosphorylation of threonine 505 in the activation loop of the enzyme (50). It has
to be considered that this phosphorylation, unlike that of corresponding threonines in
other PKC isoformes, is not essential for the formation of functional PKCδ as shown
by site-directed mutagenesis (51), but PKCδ activity is increased approximately
twofold by PDK1 phosphorylation. However, phosphorylation of threonine 505 may
be necessary for other purposes, such as protein-protein interactions. The
significance of tyrosine-phosphorylation of PKCδ also needs to be further determined.
According to in vitro studies, it seems to depend on the substrate whether PKC
activity is elevated or reduced by tyrosine-phosphorylation (52). In the present work
we have shown that inhibition of PI-3 kinase also reduces the level of
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

22
phosphotyrosine of PKCδ in CLL cells, according to a decrease of enzyme activity.
Despite the uncertainty about the tyrosine- and threonine phosphorylation of PKCδwe have shown a biological relevance of PKCδ exclusively in CLL while inhibiton of
the kinase does not affect apoptosis in peripheral B-cells of healthy donors. One of
the most important tasks in understanding the role of PKCδ in the pathogenesis of
CLL is the search for physiological substrates and associated signaling pathways.
PKCδ is linked to serine-phosphorylation of STAT proteins in several different cell
types (53) as well as to NF-κB activation in human neutrophiles (54). Because both
pathways are constitutively activated in B-CLL cells, further in vitro studies are
needed and experiments are in progress in our laboratory.
An increasing amount of publications have demonstrated the importance of protein
kinase C in CLL. PKC activation in CLL cells has been implicated with the
suppression of either spontaneous or drug induced apoptosis. Activation of PKC by
phorbol ester like TPA is able to prevent dexamethason- or chemotherapy induced
apoptosis (35), (55). Bryostatin-1 is a member of the macrocyclic lactones which
structurally mimic the PKC-activating second messenger diacylglycerol. Incubation of
CLL cells with Bryostatin-1 up-regulates the protein levels of Mcl-1 and XIAP in
accordance to an enhanced apoptosis resistance (56). On the other hand it has been
shown that nonselective inhibition of PKC by UCN-01 induces apoptosis in B-CLL
cells (57). However, it has not been shown which isoform(s) of PKC is important in
the regulation of apoptosis. We conclude from our data that PKCδ might have a
predominant role in PKC mediated prevention of apoptosis, because we failed to
detect any impact on apoptosis when B-CLL cells were incubated with inhibitors
known to be specific for inhibition of other isoforms (data not shown).
The knowledge of disturbed signaling pathways can open new opportunities in the
treatment of a disease, as it has been impressively shown for chronic myeloid
leukemia (58). Despite the lack of a unique pathognomonic feature in CLL, different
cellular alterations could end up in the same signaling pathway, causing cell cycle
arrest or prevention of apoptosis. Here we have shown that PI-3 kinase as well as
PKCδ are involved in apoptosis in CLL. To our knowledge, inhibition of PI-3 kinase in
the treatment of a malignant disease has never been investigated in a clinical trial.
Because PI-3 kinase is ubiquitiously expressed and involved in so many
physiological events this does not seem to be a feasable approach. In contrast,
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

23
PKC-inhibitors already have been tested in clinical trials (59) and have shown
antiproliferative activity against various tumors in vitro (60). Bryostatin, which acts as
a PKC inhibitor when cells are exposed for a long-term period, has shown
antileukemic activity in a phase II trial in patients with low-grade Non-Hodgkin
lymphoma and CLL (61). Targeting signaling pathways seems to be a breakthrough
in the treatment of an incurable disease. Inhibition of PKCδ in particular may be a
promising novel approach for the treatment of chronic lymphocytic leukemia.
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

24
REFERENCES
1 O´Bien S., del Giglio A., Keating M.: Advances in the biology and treatment of B-cell
chronic lymphocytic leukemia. Blood 1995; 85: 306 – 318
2 Meinhardt G., Wendtner CM., Hallek M.: Molecular pathogenesis of chronic
lymphocytic leukemia: Factors and signaling pathways regulating cell growth and
survival. J Mol Med 1999; 77 : 282 – 293
3 Dohner H., Stilgenbauer S., Benner A. et al.: Genomic aberrations and survival in
chronic lymphocytic leukemia. N Engl J Med 2000; 343: 1910 - 1916
4 Reed JC.: Molecular biology of chronic lymphocytic leukemia. Semin Oncol 1998;
25 : 11 – 18
5 Hanada M., Delia D., Aiello A., Stadtmauer E., Reed JC.: Bcl-2 gene
hypomethylation and high-level expression in B-cell chronic lymphocytic leukemia.
Blood 1993; 82 : 1820 – 1828
6 Kitada S., Andersen J., Akar S. et al. : Expression of apoptosis-regulating proteins
in chronic lymphocytic leukemia: correlations with in vitro and in vivo
chemoresponses. Blood 1998;91: 3379-3389
7 Frank DA., Mahajan S., Ritz J.: B lymphocytes from patients with chronic
lymphocytic leukemia contain signal transducer and activator of transcription (STAT)1
and STAT3 constitutively phosphorylated on serine residues. J Clin Invest 1997; 100:
3140-3148
8 Furman RR., Asgary Z., Mascarenhas JO., Liou HC., Schattner EJ.: Modulation of
NF-κB activity and apoptosis in chronic lymphocytic leukemia. J Immunol. 2000; 164:
2200 – 2206
9 Domin J., Waterfield MD.: Using structure to define the function of phosphoinositide
3-kinase family members. FEBS Lett. 1997; 410: 91 – 95
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

25
10 Andjelic S., Hsia C., Suzuki H., Kadowaki T., Koyasu S., Liou HC.:
Phosphatidylinositol 3-kinase and NF-kappa B/Rel are at the divergence of CD40
mediated proliferation and survival pathways. J Immunol. 2000; 165 : 3860 - 3867
11 Aagaard-Tillery KM., Jelinek DF.: Phosphatidylinositol 3-kinase activation in normal
human B lymphocytes. J Immunol. 1996 ; 156 : 4543 - 4554
12 Alessi DR., Andejelkovic M., Caudwell B. et al.: Mechanism of activation of protein
kinaseB by insulin and IGF-1. EMBO J 1996; 15 : 6541 – 6551
13 Aoki M., Schetter C., Himly M., Batista O., Chang HW., Vogt PK.: The catalytic
subunit of phosphoinositide 3-kinase: requirements for oncogenicity. J Biol Chem.
2000; 275: 6267-6275
14 Cheng JQ., Godwin AK., Bellacosa A. et al.: AKT2, a putative oncogene encoding
a member of a subfamily of protein-serine/threonine kinase, is amplified in human
ovarian carcinomas. Proc Natl Acad Sci USA. 1992; 89: 9267 – 9271
15 Sun M., Wang G., Paciga JE. et al. : Akt1/ PKBα kinase is frequently elevated in
human cancers and its constitutive activation is required for oncogenic transformation
in NIH3T3 cells. Am J Pathol. 2001; 159: 431 – 437
16 Kandel ES., Hay N. : The regulation and activities of the multifunctional serine /
threonine kinase Akt/PKB. Exp Cell Res. 1999; 253: 210 - 229
17 Romashkova JA., Makarov SS.: NF-kB is a target of Akt in anti-apoptotic PDGF
signaling. Nature 1999; 401: 86-90
18 Aoki M., Blazek E., Vogt K.: A role of the kinase mTOR in cellular transformation
induced by the oncoproteins P3K and Akt. Proc Natl Acad Sci USA. 2001; 98 : 136-
141
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

26
19 Toker A., Meyer M., Reddy KK. et al. : Activation of protein kinase C family
members by the novel polyphosphoinositides PtdIns-3,4-P2 and PtdIns-3,4,5-P3. J
Biol Chem. 1994; 269 : 32358 – 32367
20 Popoff IJ., Deans JP.: Activation and tyrosine phosphorylation of proteinkinase Cδin response to B cell antigen receptor stimulation. Mol Immunol. 1999, 36: 1005 –
1016
21 Ettinger SL., Lauener RW., Duronio V.: Proteinkinase Cδ specifically associates
with phosphathidyinositol 3-kinase following cytokine stimulation. J Biol Chem. 1996;
271 : 14514 - 14518
22 Ashton AW., Watanabe G., Albanese C., Harrington EO., Ware JA., Pestell RG.:
Protein kinase Cδ inhibition of S-phase transition in capillary endothelial cell involves
the cyclin-dependent kinase inhibitor p27. J Biol Chem. 1999; 274 : 20805-20811
23 Watanabe T., Ono Y., Taniyama Y., Hazama K. et al.: Cell division arrest induced
by phorbol ester in CHO cells overexpressing protein kinase C-δ subspecies. Proc
Natl Acad Sci USA. 1992; 89 : 10159 – 10163
24 Fukumoto S., Nishizawa Y., Hosoi M. et al.: Protein Kinase Cδ inhibits the
proliferation of vascular smooth muscle cells by supressing G1 cycline expression. J
Biol Chem. 1997; 272 : 13816 – 13822
25 Miyamoto A., Nakayama K., Imaki H. et al..: Increased proliferation of B cells and
auto-immunity in mice lacking protein kinase Cδ. Nature 2002; 416: 865 – 869
26 Emoto Y., Manome G., Meinhard G. et al.: Proteolytic activation of protein kinase C
delta by an ICE-like protease in apoptotic cells. EMBO J. 1995; 14 : 6148-6156
27 Skorski T., Kanakaraj P., Nieborowska-Skorska M. et al.: Phosphatidylinositol-3
kinase activity is regulated by BRC/ABL and is required for growth of Philadelphia
chromosome-positive cells. Blood 1995; 86; 726 – 736
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

27
28 Vlahos CJ., Matter WF., Hui KY., Brown RF.: A specific inhibitor of
phosphatidylinositol-3 kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one
(LY294002). J Biol Chem. 1994; 269 : 5241 – 5248
29 Pettitt AR., Moran EC., Cawley JC.: Homotypic interactions protect chronic
lymphocytic leukemia cells from spontaneous death in vitro. Leuk Res. 2001; 25:
1003 – 1012
30 Xia Z., Dickens M., Raingeaud J., Davis RJ., Greenberg ME. : Opposing effects of
ERK and JNK-p38 MAP kinases on apoptosis. Sience 1995; 270 : 1326 – 1331
31 Lali FV., Hunt AE., Turner SJ., Foxwell B.: The pyridinyl imidazole inhibitor
SB203580 blocks phosphoinositide-dependent protein kinase activity, protein kinase
B phosphorylation and retinoblastoma hyperphosphorylation in interleukin-2
stimulated T-cells independently of p38 MAP kinase. J Biol Chem. 2000; 275 : 7395
- 7402
32 Dudley DT., Pang L., Decker SJ., Bridges AJ., Saltiel AR.: A synthetic inhibitor of
the mitogen-activated protein kinase cascade. Proc Natl Acad Sci USA. 1995; 92 :
7686-7689
33 Pullen N., Thomas G.: The molecular phosphorylation and activation of
p70S6kinase. FEBS Lett. 1997; 410 : 78 - 82
34 Pronk GJ., Ramer K., Amiri P., Williams LT.: Requirement of an ICE-like protease
for induction of apoptosis and ceramide generation by REAPER. Science 1996; 271:
808 - 810
35 Bellosillo B., Dalmau M., Colomer D., Gil J.: Involement of CED-3/ ICE proteases
in the apoptosis of B-chronic lymphocytic leukemia cells. Blood 1997; 89 : 3378-3384
36 Deveraux QL., Takahashi R., Salvesen GS., Reed JC. : X-linked IAP is a direct
inhibitor of cell death proteases. Nature 1997; 388 : 300 – 304
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

28
37 Gold MR., Scheid MP., Santos L. et al.: The B cell antigen receptor activates the
Akt (protein kinase B)/ glycogen synthase kinase-3 signaling pathway via
phospatidylinositol 3-kinase. J Immunol. 1999; 163: 1894 - 1905
38 Gschwendt M., Muller HJ., Kielbassa K. et al.: Rottlerin, a novel protein kinase
inhibitor. Biochem Biophys Res Commun. 1994; 199 : 93 – 98
39 Gschwendt M., Dieterich S., Rennecke J., Kittstein W., Mueller HJ., Johannes FJ.:
Inhibition of protein kinase Cµ by various inhibitors. Differentiation from protein
kinase c isoenzymes. FEBS Lett. 1996; 392 : 77 – 80
40 Benekli M., Xia Z., Donohue KA. et al.: Constitutive activation of signal transducer
and activator of transcription 3 protein in acute myeloid leukemia blasts is associated
with short disease-free survival. Blood 2002; 99; 252 – 257
41 Tse KF., Mukherjee G., Small D.: Constitutive activation of FLT3 stimulates
multiple intracellular signal transducers and results in transformation. Leukemia 2000;
14: 1766 – 1776
42 Milella M., Kornblau SM., Estrov Z. et al.: Therapeutic targeting of MEK/ MAPK
signal transduction modulate in acute myeloid leukemia. J Clin Invest. 2001; 108 :
851 - 859
43 Fruman DA., Snapper SB., Yballe CM. et al.: Impaired B cell development and
proliferation in absence of phosphoinositide 3 kinase p85α. Sience 1999; 283: 393 -
397
44 Imani F., Rager KJ., Catipovic B., Marsh DG.: Interleukin-4 (IL-4) induces
phosphatidylinositol 3 kinase (p85) dephosphorylation. Implications for the role of
SHP-1 in the IL-4-induced signals in human B cells. J Biol Chem. 1997; 272: 7927 –
7931
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

29
45 Barragan M., Bellosillo B., Campas C., Colomer D., Pons G., Gil J.: Involvement of
protein kinase C and phosphatidylinositol 3-kinase pathways in the survival of B-cell
chronic lymphocytic leukemia. Blood 2002; 99: 2969 – 2976
46 Cantley LC., Neel BG.: New insights into tumor supression: PTEN supresses
tumor formation by restaining the phophoinositol-3 kinase/ AKT pathway. Proc Natl
Acad Sci USA. 1999; 96: 4240 – 4245
47 Tu Y., Gardner A., Lichtenstein A. : The phosphatidylinositol-3 kinase/Akt pathway
in multiple myeloma plasma cells: Role in cytokine-dependent survival and
proliferation responses. Cancer Res. 2000; 60: 6763 - 6770
48 Wickremasinghe RG., Ganeshaguru K., Jones DT. et al.: Autologous plasma
activates Akt/ proteinkinase B and enhances basal survival and resistance to DNA
damage-induced apoptosis in B-chronic lymphocytic leukemia cells. Br J Haematol.
2001; 114: 608 - 615
49 Bernal A., Pastore RD., Asgary Z. et al.: Survival of leukemic B cells promoted by
engagement of antigen receptor. Blood 2001; 98: 3050 – 3057
50 Good JA., Ziegler WH., Parekh DB., Alessi DR., Cohen P., Parker PJ.:
Proteinkinase C isotypes controlled by phosphoinsitide 3 kinase through the protein
kinase PDK1. Sience 1998; 281: 2042 – 2045
51 Stempka L., Girod A., Muller HJ. et al.: Phosphorylation of protein kinase Cδ(PKCδ) at threonine 505 is not a prerequisite for enzymatic activity. Expression of rat
PKCdelta and an alanine 505 mutant in bacteria in a functional form. J Biol Chem.
1997; 272 : 6805 – 6811
52 Gschwendt M., Kielbassa K., Kittstein W., Marks F.; Tyrosine phosphorylation and
stimualtion of proteinkinase Cδ from porcine spleen by src in vitro. Dependence on
the activated state of proteinkinase Cδ. FEBS Lett. 1994; 347 : 85 – 89
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom

30
53 Jain N., Zhang T., Kee WH., Li W., Cao X.: Protein Kinase Cδ associates with
phosphorylates Stat3 in an interleukin-6 dependent manner. J Biol Chem 1999; 274 :
24392 – 24400
54 Vancurova I., Miskolci V., Davidson D.: NF-κB activation in tumor necrosis factor α-
stimulated neutrophils is mediated by proteinkinase Cδ. Correlation to nuclear Ikappa
Balpha. J Biol Chem. 2001; 276 : 19746 – 19752
55 Forbes IJ., Zalewski PD., Giannakis C., Cowled PA.: Induction of apoptosis in CLL
and its prevention by phorbol ester. Exp Cell Res. 1992; 198 : 367 - 372
56 Kitada S., Zapata JM., Andreeff M., Reed JC.: Bryostatin and CD40-ligant enhance
apoptosis resistance and induce expression of cell survival genes in B-CLL. Br J
Haematol. 1999; 106 : 995 – 1004
57 Kitada S., Zapata JM., Andreeff M., Reed JC.: Protein kinase inhibitors flavopiridol
and 7-hydroxy-staurosporine down-regulate antiapoptosis proteins in B-CLL. Blood
2000; 96 : 393 - 397
58 Goldman JM., Druker BJ.: Chronic myeloid leukemia: Current treament options.
Blood 2001; 98 : 2039 – 2042
59 Propper DJ., McDonald AC., Man A. et al.: Phase I and pharmacokinetic study of
PKC412, an inhibitor of proteinkinase C. J Clin Oncol. 2001; 19 : 1485 – 1492
60 Meyer T., Regenass U., Fabbro D. et al.: A derivate of staurosporine (CGP 41 251)
shows selectivity for proteinkinase C inhibition and in vitro anti-proliferative as well as
in vivo anti-tumor activity. Int J Cancer 1989; 43 : 851 – 859
61 Varterasian ML., Mohammad RM., Shurafa S. et al.: Phase II trial of bryostatin 1 in
patients with relapsed low-grade non-Hodgkin´s lymphoma and chronic lymphocytic
leukemia. Clin Cancer Res. 2000; 6 : 825 - 828
For personal use only.on April 27, 2014. by guest bloodjournal.hematologylibrary.orgFrom




















