
COMPARATIVE FEEDING ECOLOGY OF SPINNER DOLPHINS (STENELLA LONGIROSTRIS) AND FRASER'S DOLPHINS...
19
MARINE MAMMAL SCIENCE, 19(1):1-19 (January 2003) 0 2003 by the Society for Marine Mammalogy COMPARATIVE FEEDING ECOLOGY OF SPINNER DOLPHINS (STENELLA LONGIROSTRIS) AND FRASER’S DOLPHINS (LAGENODELPHIS HOSEI) IN THE SULU SEA M. LOUELLA L. DOLAR’ Scripps Institution of Oceanography, University of California, San Diego, La Jolla, California 92093-0204, U.S.A E-mail: [email protected] WILLIAM A. WALKER National Marine Mammal Laboratory, National Marine Fisheries Service, NOAA, 7600 Sand Point Bay, N.E., Seattle, Washington 981 15, U.S.A. GERALD L. KOOYMAN Scripps Institution of Oceanography, University of California, San Diego, La Jolla, California 92093-0204, U.S.A. WILLIAM F. PERRIN Southwest Fisheries Science Center, National Marine Fisheries Service, NOAA, 8604 La Jolla Shores Drive, La Jolla, California 92037, U.S.A. ABSTRACT Fraser’s and spinner dolphins are known to feed on mesopelagic prey, but their diets and feeding ecologies have not been compared in areas where they are observed together. In this study we examined the stomach contents of both species caught incidentally in a driftnet fishery for tuna in the eastern Sulu Sea. Importance of prey items was determined using the percent occurrence, percent number, and volumetric methods. Mesopelagic fishes, particularly myctophids (mainly Ceratoscopelus warmingi, Diaphus spp. and Myctophum asperum), were the most important component in the diet of spinner dolphins, whereas in Fraser’s dolphins, mesopelagic cephalopods (Abraliopsis, Onychoteuthis, Histioteuthis, and Chiroteuthis), and crustaceans (Notostornos elegans, Acanthephyra quadrispinosa, and Acanthephyra carinata) appeared to be equally important as the myctophid ’ Current address: 6363 Lakewood Sr., San Diego, California 92122 1
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of COMPARATIVE FEEDING ECOLOGY OF SPINNER DOLPHINS (STENELLA LONGIROSTRIS) AND FRASER'S DOLPHINS...

MARINE MAMMAL SCIENCE, 19(1):1-19 (January 2003) 0 2003 by the Society for Marine Mammalogy
COMPARATIVE FEEDING ECOLOGY OF SPINNER DOLPHINS (STENELLA LONGIROSTRIS)
AND FRASER’S DOLPHINS (LAGENODELPHIS HOSEI ) IN THE SULU SEA
M. LOUELLA L. DOLAR’ Scripps Institution of Oceanography, University of California, San Diego,
La Jolla, California 92093-0204, U.S.A E-mail: [email protected]
WILLIAM A. WALKER National Marine Mammal Laboratory,
National Marine Fisheries Service, NOAA, 7600 Sand Point Bay, N.E.,
Seattle, Washington 981 15, U.S.A.
GERALD L. KOOYMAN Scripps Institution of Oceanography, University of California, San Diego,
La Jolla, California 92093-0204, U.S.A.
WILLIAM F. PERRIN Southwest Fisheries Science Center,
National Marine Fisheries Service, NOAA, 8604 La Jolla Shores Drive,
La Jolla, California 92037, U.S.A.
ABSTRACT
Fraser’s and spinner dolphins are known to feed on mesopelagic prey, but their diets and feeding ecologies have not been compared in areas where they are observed together. In this study we examined the stomach contents of both species caught incidentally in a driftnet fishery for tuna in the eastern Sulu Sea. Importance of prey items was determined using the percent occurrence, percent number, and volumetric methods. Mesopelagic fishes, particularly myctophids (mainly Ceratoscopelus warmingi, Diaphus spp. and Myctophum asperum), were the most important component in the diet of spinner dolphins, whereas in Fraser’s dolphins, mesopelagic cephalopods (Abraliopsis, Onychoteuthis, Histioteuthis, and Chiroteuthis), and crustaceans (Notostornos elegans, Acanthephyra quadrispinosa, and Acanthephyra carinata) appeared to be equally important as the myctophid
’ Current address: 6363 Lakewood Sr., San Diego, California 92122
1

2 MARINE MAMMAL SCIENCE. VOL. 19. NO. 1. 2003
species. Fraser’s dolphins appeared to feed preferentially on larger prey and had more diversified prey (Shannon’s diversity index = 1.2) than spinner dolphins (diversity index = 0.9). Vertical distributions of the prey items summarized from published literature indicate that spinner dolphins forage in the upper 200 m and probably occasionally to as deep as 400 m, whereas Fraser’s dolphins have a wider vertical foraging range, from near the surface to probably as deep as 600 m.
Key words: Philippines, diet, myctophids, cetaceans, spinner dolphin, Stenella longirostris, Fraser’s dolphin, Lugenodelphis hosei, squids.
The spinner dolphin (Stenella lungimtris) and Fraser’s dolphin (Lagenodelph hosei) both are abundant in the central Philippines. Spinner dolphins occur in both shallow and deep waters, whereas Fraser’s dolphins prefer deep waters (Dolar 1999). In areas where their distributions overlap, e.g., in the eastern Sulu Sea, the two species are occasionally found together forming large schools. The by-catch of these two species in the driftnet fishery accounts for most of the dol- phins caught (Dolar 1994).
Current knowledge on the diets of Fraser’s dolphin comes from the stomach contents of animals stranded on beaches in Japan and France (Tobayama et al. 1973, van Bree et al. 1986), taken for scientific research in Japan and South Afri- ca (Miyazaki and Wada 1978, Sekiguchi et al. 1992), and killed incidentally in fisheries in the Atlantic and eastern tropical Pacific (Caldwell et al. 1976, Robi- son and Craddock 1983). Robison and Craddock (1983) provided the most de- tailed description of the fish prey identified from the otoliths. They suggested that Fraser’s dolphins were deep divers, feeding selectively on relatively large mesopelagic fish at depths between 250 m and 500 m. Watkins et al. (1994), on the other hand, reported surface feeding by a school of Fraser’s dolphins in the Caribbean. Similar behavior was reported for a pod of Fraser’s dolphins off the eastern coast of South Africa (Ross 1984).
Spinner dolphins feed primarily on mesopelagic fish and squid (Cadenat and Doutre 1959, Fitch and Brownell 1968, Perrin et al. 1973, Norris and Doh1 1980, Wiirsig et al. 1994) as these prey migrate to or near the surface at night (Fitch and Brownell 1968, Perrin et al. 1973). A dwarf form in Southeast Asia (not the Philippines), S. longirostris roseiuentris, feeds on shallow benthic and coral reef organisms (Perrin et al. 1989, Perrin et al. 1999).
Thus, both spinner and Fraser’s dolphins appear to feed mainly on mesopelagic prey. Robison and Craddock (1983) postulated that the two species have different feeding strategies in the eastern tropical Pacific. Although both dolphins occur there, their centers of distribution appear to differ (Wade and Gerroderte 1993). In addition, these two species have not been previously found together. We sought to determine if these species exploit the same prey when sharing the same area or exhibit evidence of resource partitioning. We also compare their diet with that of other populations in the eastern tropical Pacific, Hawaii, and South Africa.
METHODS
Collection and Processing of SampleJ
Stomachs were collected from 45 spinner and 37 Fraser’s dolphins incidentally caught in a tuna driftnet fishery operating in the eastern Sulu Sea, Philippines.

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY 3
Both ends of each stomach were tied off before removal from the dolphin. After removal they were injected with and stored in 70% ethanol for a period of one month. The volume of the unsorted contents of the forestomach only was deter- mined to the nearest ml using the water displacement method. The lining of the stomach was thoroughly rinsed through a series of small-sized sieves to recover otoliths. Otoliths were gathered and stored dry in plastic vials. The remaining stomach contents were placed in fresh 70% ethanol and shipped to Scripps Insti- tution of Oceanography (SIO).
In the laboratory, the stomach contents of each dolphin were sorted into the major categories of fish, cephalopods, and crustaceans. The volume of each care- gory was determined using the displacement method. Fish were often semi- digested so that it was not possible to measure their total lengths. Specimens in relatively good condition were preserved in 70% alcohol and identified after the sagittal otoliths were extracted to serve as part of the reference collection. Oto- liths were also extracted from the skulls of semidigested fish, washed and stored dry in plastic vials. Fish and crustacean remains and squid beaks were sorted and stored in 70% ethanol. Otoliths were identified to the lowest possible taxon using the reference collection of one of us (WAW) and of the US. National Mu- seum of Natural History (NMNH) at the Smithsonian Institution. Fish were also collected from the study site from catches by commercial deep-water ring nets. These specimens were used as references to identify the otoliths found in the dol- phin stomachs. In addition, otoliths from identified fish at the N M N H collected from the Philippines (Sulu Sea and other areas) and the tropical Pacific were ex- tracted and used for reference; only fish stored in alcohol with otoliths still intact were used. Published identification guides were also consulted (Fitch and Brow- nell 1968, Von Weiler 1968, Fitch and Gotshall 1972, Harkonen 1986, Gon and Heemsrra 1990, Williams and McEldowney 1990, Gago 1993, Smale et ul. 1995). Some otoliths were in good condition but could not be identified, where- as others were too eroded to be identified. The total number of otoliths was divided by two to estimate the number of fish eaten. The maximum length of minimally eroded otoliths of important fish species was measured to the nearest 0.01 mm using an eyepiece micrometer. Otolith length was used to estimate fish length using available regression equations relating otolith length to fish length (Appendix 1). Fish dentary bones were identified, sorted into right and left, matched and enumerated. The number of viperfishes (Chuzlliodus sloani) eaten was determined this way, since the otoliths of this species were extremely small and some might have been lost. Debrot and Barros (1992) estimated the size of Chuzlliodzls sloani ingested by a Mesoplodon earnopaem using a ratio of 1.8 of mandi- ble to total length. Fish classification followed Nelson (1994).
Beaks were extracted from the cephalopods and the upper and lower mandibles were separated and counted. The highest count plus the number of intact beaks were used to estimate the total number of cephalopods in the stomach. Cephalo- pods were identified from the voucher collection of WAW and by using pub- lished identification guides (Wolff 1982, Clarke 1986a). The lower rostra1 length (LRL) of the beaks of the commonly occurring species Abruliopsis sp. cf. A. linea- ta was measured to the nearest 0.01 m m and was used in estimating mantle length. Classification was based on Sweeney and Roper (1998). Crustaceans were identified using published identification guides (Burukovskii 1982, Takeda and Okutani 1983, Chace 1986) and verified by Mary Wicksten of Texas A&M Uni- versity. Since crustaceans do not have hard indigestible structures like the oto-

4 MARINE MAMMAL SCIENCE, VOL. 19. NO. 1. 2003
liths in fish and beaks in cephalopods, it was not possible to enumerate them ac- curately. Postorbital carapace lengths of relatively intact animals were measured to the nearest 0.1 mm.
Data AnalysiJ
Stomach fullness was estimated as the ratio of the volume of the contents to the volume of the most distended stomach within a particular dolphin size class. There were three size classes established for spinner dolphins (1 30-160 cm, 161-190 cm, 191-220 cm) and four for Fraser’s dolphins (120-160 cm, 161- 200 cm, 201-240 cm, 241-290 cm). In this analysis, only stomachs of dolphins with body length measurements were used (38 spinner and 28 Fraser’s dolphins). Five fullness categories were used: category 1 (empty with ratio = O), category 2 (trace to quarter-full with ratio = 0.1-0.25), category 3 (ratio = 0.26-50), cat- egory 4 (ratio = 0.51-0.75), and category 5 (ratio = 0.76-1.0). The number of stomachs with semidigested prey (i.e., relatively intact prey) was noted.
Three methods were used to quantify the importance of prey: the volumetric, percent occurrence, and number methods. Each of these three methods has weak- nesses, but when used together may give a reasonable estimate (Hyslop 1980). The volumetric method was used to estimate the proportion of the three major food categories, i.e., fish, cephalopods, and crustaceans. This was used for intact, semidigesred, as well as all distinguishable parts of prey including loose otoliths and beaks. To test for differenceshnilarities in mean sizes of prey eaten by the two dolphin species, two-tailed t-tests were used on normally distributed data and Mann-Whitney test on non-normally distributed data. The standard error (SE) is given as a measure of variability.
Shannon’s index of diversity ( H ) and Simpson’s determined for the fish and cephalopod species 1997):
and
where ni = number for each prey species and N =
index of dominance ( D ) were using the equations (Odum
total number of prey
RESLJLTS
The general condition of stomach fullness in the two species was similar. Fifty percent and 61% of the stomachs examined showed trace amounts to quarter-full (category 2) in spinner and Fraser’s dolphins, respectively; 21% and 14% were up to half-full (category 3), 11% and 4% were up to three-quarters full (category 4), 5% and 7% were full (category 5); and 13% and 14% were empty (category 1).
Recovered prey items belonged to three general types: teleosts (bony fishes), cephalopods, and crustaceans. Fish was the dominant prey, more so in spinner dol- phins (87% of the total prey volume) than in Fraser’s dolphins (55%) (Table l). Fish occurred in all spinner dolphin stomachs and in 97% of Fraser’s dolphin

DOLAR ET AL : DOLPHIN FEEDING ECOLOGY 5
Table I . Proportions (%) of general food categories by volume and occurrence found in stomachs of 45 spinner and 37 Fraser’s dolphin.
Spinner Fraser’s
Prev Volume Occurrence Volume Occurrence
Fish 86.6 100 54.6 91.3 Cephalopod 6.4 89.8 30.0 89.2 Crustaceans 7.0 27.0 15.4 67.0
stomachs. Cephalopod remains comprised approximately 30% of the total prey volume in Fraser’s dolphins and were found in approximately 90% of all stom- achs; crustaceans comprised only 15% in volume and were found in approxi- mately 67% of all stomachs examined. In spinner dolphins these two prey categories comprised only approximately 7% each of the total prey volume, although cephalopods occurred in almost 90% of the stomachs examined. Crusta- ceans occurred in only 27% of spinner dolphin stomachs examined.
Fishes
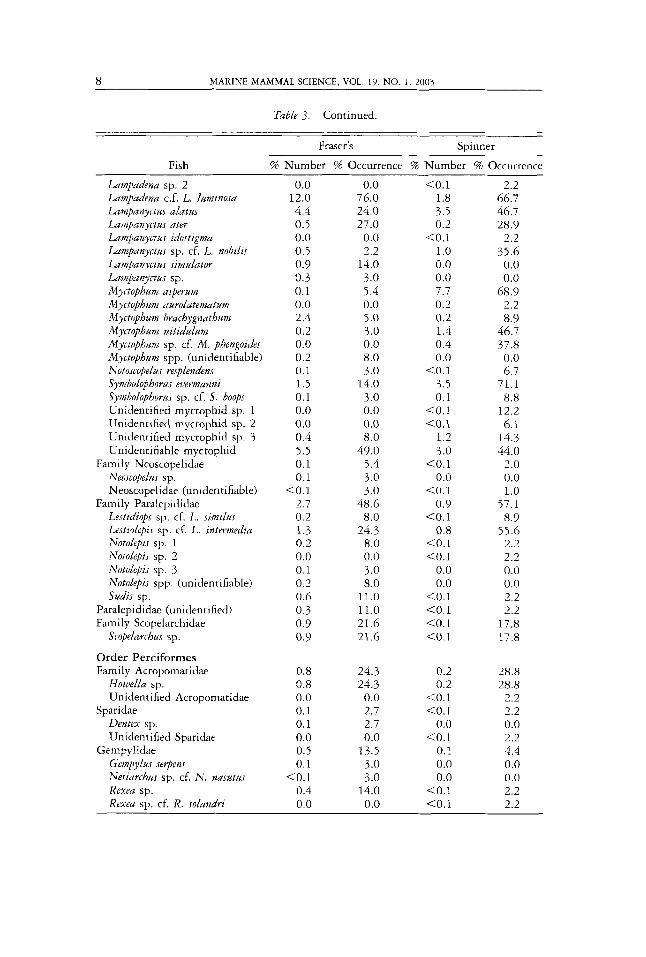
A total of 26,809 otoliths was found in the 45 spinner dolphin stomachs and 2,936 in the 37 Fraser’s dolphin stomachs, representing a total of 92 species. At least 75 species belonging to 16 families were observed in spinner dolphins and 59 species belonging to 20 families in the Fraser’s dolphins (Table 2). Forty-six percent of fish prey species were found in both species of dolphins. The list of fish families and species and the corresponding percentages using the number and occurrence methods for the two dolphin species are shown in Table 3. One to two percent of whole otoliths were not identified to the family level, and 7%-8% were extremely eroded and could not be identified.
Myctophids (Order Myctophiformes) or lanternfishes were the most numerous and the most frequently occurring group, more so in spinner (84% in number and 94% in occurrence) than in Fraser’s dolphins (65% in number and 89% in occurrence) (Table 3). Thirty-two myctophid species were found in spinner dol- phins and only 26 in Fraser’s dolphins; 49% were common to both. Ceratoscopelus wamingi accounted for about 28% in spinner and 14% in Fraser’s dolphins by number. This species was also the most frequently occurring species in the spin- ner dolphin (found in 91% of the stomachs) and the third most frequent species occurring in Fraser’s dolphins.
The other important myctophid species in terms of number and percent occur- rence in spinner dolphin stomachs were Diapbus parri, Myctopbum asperum, Dia- pbus Iutkeni, Symbolophorus euermanni, Lampanyctus alatus, Diogenicbthyes atlanticus, and Lampadena sp. Cf. L. lzlminosa (Table 3).
Although six of the seven species mentioned above were also found in Fraser’s dolphins, the order of importance differed. Lampadena Cf: L. luminosa ranked sec- ond to Ceratoscopelus warmingi in terms of number, but ranked first in frequency of occurrence. Didpbus parri ranked third. Other important prey species in descending order of importance based on number were: Lampanyctus alatus, Bolinicbtbys pyrsobolus, Diaphus lutkeni, Diapbus fragilis, Bolinicbthys sp.2, Myctopbum

6 MARINE MAMMAL SCIENCE, VOL. 19, NO. 1 , 2001
Table 2. Summary of number of fish, cephalopod, and crustacean families and species found in stomachs of spinner and Fraser’s dolphins.
Spinner Fraser’s Total Fish
Number of families 16 20 2 1 Number of species 75 59 92 Species overlap 42 (46%)
Number of families 10 12 12 Number of species 17 21 24 Species overlap 14 (58%)
Number of families 3 2 3
Species overlap 2 (40%)
Cephalopods
Crustaceans
Number of species 3 4 5
Total number of stomachs 45 37 Number of stomachs with food 40 33 Number of otoliths 26,809 2,936 29,745 Number of beaks 588 428 1,016
brachygnathum, Electrona risso, and Symbolophorus evermunni (Table 3). Aside from the rnyctophids, other fish groups appeared to be relatively important in number in Fraser’s dolphins, but not in spinner dolphins. These were the Stomiidae (Order Salmoniformes), Nomeidae (Order Perciformes), and Macrouridae (Order Gadiformes). The size ranges and mean standard lengths of important fish species found in the stomachs of spinner and Fraser’s dolphins are shown in Table 4. The sizes of Lampadena sp. cf. L. luminoJa in the two dolphin species were signifi- cantly different (two-tailed t-test, P < 0.001). The mean for Fraser’s dolphin was larger than for the spinner dolphin. However, those of C. warmingi, S. evermun- ni and C. puuciradiatus were not significantly different (two-tailed t-test, P for S. evermunni = 0.891, C. warmingi = 0.723 and C. paucirudiatus = 0.203).
The mean number of fish species found in the stomachs was significantly lower in Fraser’s dolphin (10 2 1.36) than in the spinner dolphin (15 2 1.25) (Kol- mogorov-Smirnov test, P < 0.001), as well as the mean number of fish per stomach (302 2 47.20 for spinner dolphins, but only 42.30 2 11.9 for Fraser’s dolphins). Number of fish in each stomach ranged from 0 to 416 in Fraser’s dol- phin and from l to l ,63 l in the spinner dolphin.
Cephulopods
A total of 1,016 cephalopod beaks were found in the stomachs of spinner and Fraser’s dolphins, and from these 12 families (all of order Sepiolida) were identi- fied (Table 2). All 12 families were found in Fraser’s dolphin stomachs, but only 10 were found in spinner dolphin stomachs. Of the 12 families, the Enoploreu- thidae (specifically Abruliopsis sp. cf. A. lineata) accounted for the highest number in both species (85% in spinner dolphin and 329% in Fraser’s dolphin) (Table 5). Abraliopsisis sp. cf. A. lineuta alone comprised 79% (in terms of number) of all

DOLAK ET AL.: DOLPHIN FEEDING ECOLOGY 7
Table 3. Fish families and species and their frequency occurrence and percent number in stomachs of Fraser’s and spinner dolphins.
Fraser’s Spinner
Fish % Number % Occurrence % Number % Occurrence
Order Anguilliformes Family Congridae
Family Ophichthyidae Conger sp.
Unidentified Ophichthyidae
Order Beloniformes Family Scomberoscocidae
Colalabis saira
Order Beryciformes Family Diretmidae
Diretmoides parini Family Melamphidae
Melamphae.r sp. Scapelobeyx sp. Scopelogadus sp.
Order Gadiformes Family Bregmacerotidae
Breginaceros sp. Family Macrouridae
Caelorinchus sp. Coryphaenoides sp. Hymenocephalus sp. macrourid (unidentifiable)
Order Myctophiformes Family Myctophidae
Bentbasema fibdatum Bolinichthys pyrsobolus Bolinichthys sp. 1 Bolinichthys sp. 2 Ceratoscopelus warmingi Diaphus fragilis Diaphus garmani Diaphus Iutkeni Diaphus parri Diaphus rafnesquei Diaphus sagamiensis Diaphus suborbitalis Diaphus sp. 1 Diaphus sp. 2 Diaphzls sp. 3 Diaphus spp. (unidentifiable) -. .-___- . ~~ .~
0.0 0.0 0.1 0.1
0.4 0.4
0.9 0.9 0.9 0.2 0.2 0.5
0.9 0.9 2.3 1.4 0.4 0.0 0.1
65.0 0.1 3.7 0.0 2.5
14.0 2.8 0.0 3.5 5.1 0.0 0.0 1.4 0.1 0.1 0.1 0.4
0.0 0.0 3.0 3.0
5.4 5.4
24.0 24.0 35.1
8.0 5 .O
11.0
14.0 14.0 37.8 32.4
5 .O 0.0 3.0
89.2 3 .0 3 .O 0.0
27.0 43.0 45.9 0.0
41 .O 30.0
0.0 0.0
16.0 3 .O 3 .0 3 .O 8.0
c o . 1 c o . 1 c o . 1 c o . 1
0.0 0.0
1.8 1.8 0.2
<o. 1 0.1 0.1
1.2 1.2 0.4 0.2
<0.1 <0.1
0.0
83.6 0.0 0.0
c o . 1 0.1
27.8 1.2 1.5 4.0
19.8 <0.1
0.1 1.7 0.0 0.1 0.0 0.0 ^ ^ .. .
6.7 6.7 2.2 2.2
0.0 0.0
55.6 55.6 12.2 4.4 2.2 8.9
49.0 49.0 28.9 28.9 6.7 2.2 0.0
93.9 0.0 0.0
1 1 . 1 4.4
91.1 53.3 40.0 68.9 97.8 4.4
44.4 44.4
0.0 6.7 0.0 0.0
/-.-
8 MARINE MAMMAL SCIENCE. VOL. 19. NO. 1. 20(11
Table 3. Continued.
Fraser's Spinner
Fish % Number % Occurrence % Number % Occurrence
Lampadena sp. 2 Lawqadena c.f. L. Zuminosa Larizpanyctus alatus Lampanyctus ater Lampanyctus idostigma Lampanyetus sp. cf. L. nobilis Lampanyetus simulator Lampanyetus sp. Myctophum asperum Myctophum aurolatematum Myctophum hrachygnathurn Myi-tophum nitidulum Myctophum sp. cf. M. phengoides Myctophum spp. (unidentifiable) Notoscopelu.r resplendem Symholophorus euemanni Symholophorus sp. cf. S. hoops Unidentified myctophid sp. 1 Unidentified myctophid sp. 2 Unidentified myctophid sp. 3 Unidentifiable myctophid
Neoscopelus sp. Neoscopelidae (unidencifiable)
Lestidiops sp. cf. I>. sirnilus Latiolepis sp. cf. L. intermedia Notolepzs sp. 1 Notolepis sp. 2 Notolepis sp. 3 Notolepis spp. (unidentifiable) Sudis sp.
Family Neoscopelidae
Family Paralepididae
Paralepididae (unidentified) Family Scopelarchidae
Scopelarchus sp.
Order Perciformes Family Acropomatidae
Uowella sp. Unidenrified Acropornatidae
Dentex sp. Unidentified Spari dae
Gempylidae Gempy1u.r .rerpen.r Nesiarchus sp. cf. N. nasutu.r Rexea sp. Rexea SD. cf. R. solandri
Sparidae
0.0 0.0 12.0 76.0 4.4 24.0 0.5 27.0 0.0 0.0 0.5 2.2 0.9 14.0 0.3 3.0 0.1 5.4 0.0 0.0 2.4 5 .0 0.2 3.0 0.0 0.0 0.2 8.0 0.1 3.0 1 . 5 14.0 0.1 3.0 0.0 0.0 0.0 0.0 0.4 8.0 5 .5 49.0 0.1 5.4 0.1 3.0
<0.1 3.0 2.7 48.6 0.2 8.0 1.3 24.3 0.2 8.0 0.0 0.0 0.1 3.0 0.2 8.0 0.6 11.0 0.3 11.0 0.9 21.6 0.9 21.6
0.8 24.3 0.8 24.3 0.0 0.0 0.1 2.7 0.1 2.7 0.0 0.0 0.5 13.5 0.1 3.0
<0.1 3.0 0.4 14.0 0.0 0.0
(0.1 1.8 3.5 0.2
C0.1 1 .0 0.0 0.0 7.7 0.2 0.2 1.4 0.4 0.0
<0.1 3.5 0.1
<0. 1 <0.1
1.2 3 .0
<0.1 0.0
<0.1 0.9
K0.1 0.8
<0.1 <0.1
0.0 0.0
<0.1 C0.1 <0.1 C0.1
0.2 0.2
<0. 1 <0. 1
0.0 <0.1
0.1 0.0 0.0
C0.1 <0.1
2.2 56.7 46.7 28.9
2.2 35.6 0.0 0.0
68.9 2.2 8.9
46.7 37.8 0.0 6.7
71.1 8.8
12.2 6.1
14.3 44.0
2.0 0.0 1.0
57.1 8.9
55.6 2.2 2.2 0.0 0.0 2.2 2.2
17.8 17.8
28.8 28.8
2.2 2.2 0.0 2.2 4.4 0.0 0.0 2.2 2.2

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY 9
Table 3. Continued.
Fraser’s Spinner
Fish % Number % Occurrence 9% Number % Occurrence Family Trichiuridae
Apanophus carbo Family Nomeidae
Cubireps pauciradiatus Cubireps squamiceps Cubiceps sp.
Order Salmoniformes Family Argentinidae
Nansenia sp. Family Bathylagidae
Bathylagus sp. Family Gonostomatidae
Unidentified Gonostomatidae Family Sternoptychidae
Polyipnus indicus Polyipnus lateratus
Family Stomiidae Chauliodus sloani
Unidentified teleosts
Unidentifiable teleosts Total
0.6 0.6 5.4 3.5 0.7 1.2
1.8 1.8 0.2 0.2 0.1 0.1 0.4 0.1 0.3
7.4 7.4 1.4 2 SPP. 7.2
100.0
21.6 21.6 45.9 32.0 11.0 11.0
37.8 37.8 8.0 8.0 5 .0 5 .0 5.4 3.0 3 .O
42.0 42.0 43.2
16.0
0.1 0.1 1 .0 0.4 0.6 0.0
0.0 0.0 0.1 0.1 0.0 0.0 0.0 0.0 0.0
<0. 1 <0. 1
1.7
8.4 100.0
15 spp.
15.6 15.6 59.2 26.7 37.8 0.0
0.0 0.0
13.3 13.3 0.0 0.0 0.0 0.0 0.0
4.4 4.4
53.1
60.0
cephalopods in spinner dolphins and 31% in Fraser’s dolphins (Table 5 ) . Other important cephalopod families in Fraser’s dolphin’s diet were Cranchiidae, Ony- choteuthidae, and Chiroteuthidae. Overall, Fraser’s dolphins took more cephalo- pod species than spinner dolphins (21 us. 17 spp.), with the mean number of species in each stomach equal to 3.5 2 0.26 for Fraser’s and 2.3 ? 0.25 for spinner dolphins, respectively. The percentage of overlap in cephalopod species taken by spinner and Fraser’s dolphins was 58%. The mean number of cephalo- pods per stomach was 9.1 2 1.37 for spinner dolphin and 8.0 2 1.31 for Fras- er’s dolphin.
The mean mantle lengths of the most abundant cephalopod prey, Abraliopsis sp. cf: A. lineatu were significantly different in the two dolphin species (two- tailed t-test, P < 0,001). The mean size in Fraser’s dolphin was greater than in the spinner dolphin.
Crustuceans
Owing to the fact that a large proportion of crustaceans found in the dolphin stomachs were in an advanced stage of digestion, quantification by species was not attempted except for a rough evaluation of occurrence. Four species belonging to two families were found in Fraser’s dolphins: Oplophoridae: Notostornos elegans, Acanthephyra quadrispinosa, Acanthephyra curinata; Sergestidae: unidentified sp.

Tabl
e 4.
M
ean
size
s an
d si
ze ra
nges
of
som
e pr
ey f
ound
in
stom
achs
of
Fras
er's
and
spin
ner
dolp
hins
. n =
= nu
mbe
r of
pre
y m
easu
red.
Pro
babi
lity
valu
es a
lso
show
n fo
r co
mpa
riso
n of
mea
n si
zes
of p
rey
(*tw
o-ta
iled
t-te
st;
**M
ann-
Whi
tney
tes
t).
Prey
Fr
aser
's Sp
inne
r
n n
Ran
ge
P-va
lue
Fam
ily
Spec
ies
Mea
n -C
SE
Ran
ge
Mea
n t S
E
Fish
M
ycto
phid
ae
Nom
eida
e
Cep
halo
pod
Lam
pade
na lu
min
osa
Cera
tosc
opelu
s w
arm
ingi
Sy
mbo
lopb
orus
evm
anni
M
ycto
pbum
niti
dulu
m
Cubi
ceps
pauc
iradi
atus
Eno
plot
euth
ic-e
Ab
ralio
psis
cf: A
. lin
eata
Cru
stac
ean
Opl
opho
rida
e No
tosto
mus
eleg
ans
Acdn
thep
hyra
car
inat
a Ac
anth
ephy
ra q
uadr
ispin
osa
Pena
eida
e H
emip
enae
us s
p.
Serg
estid
ae
Uni
dent
ifie
d se
rges
tid
245
171 43
3 55
30 5 5 19 1
102
+ 0.
87
59.3
+ 0
.36
64.8
+ 0.
53
45 +
4.4
113
+ 1.
70
50.9
+ 1.
39
37.4
+-
4.32
37
.6 2
3.4
9 14
.7 i 0
.56
Non
e 16
81-1
46
42-6
9 5 8
-7 6
37
-5 1
85
-142
37-6
7
23
45
10-1
9 30
-50
241
82.4
i-
0.87
42
0 59
.1 i
- 0.
26
243
64.8
+ 0.
28
166
57.2
+ 0.
45
9 11
8.9
4- 3.
30
112
38.2
i- 1
.03
Non
e N
one
15
14.2
t 0
.40
18
18.9
-C 0
.28
7 15
.3 2
0.3
3
49-1
14
45-7
8 53
-79
41-7
3 11
0-13
4
17-6
3
12-1
8
14-1
6 15
-20
<0.0
01*
0.72
* 0.
89*
0.20
" -
<0.
001*
-
-
>0.
05**
-
-

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY I 1
Table 5. Relative proportions (to total number of cephalopods) and frequency occurrence of cephalopod species in stomachs of Fraser's and spinner dolphins.
~ ~
Fraser's Spinner
Species % Number % Occurrence % Number % Occurrence
Family Lycoteuthidae Lycoteuthis sp.
Family Enoploteuthidae Abraliopsis sp. cf. A . lineata Enoploteuthis chunii Enoplateuthis sp.
Family Octopoteuthidae Ocropoteuthis sp.
Family Onychoteuthidae Onycboteuthis banksi
Family Histioteuthidae Histioteutbis miranda
Family Ctenopteridae Ctenopteryx sicula
Family Brachioteuthidae Brachioteuthis sp.
Family Ommastrephidae Nototodarus c. f. philippinensis Ommastrephidae unid sp.
Thysanoteutbis rhombus
Chiroteuthis c.f. imperator Chiroteuthis sp. Grimalditeutbis Chiroteuthidae unid sp.
Idzotuethis sp. Mastigoteuthis sp. cf. M. hjorti
Family Cranchiidae Cranchia scabra Gals'tcuthiJ sp. Helicocranchia sp. Leachia dislocata Taonius sp. Cranchiidae unid sp.
Family Thysanoteuthidae
Family Chiroteuthidae
Family Mastigoteuthidae
0.8 0.8
32.1 30.9
0.0 1.2 5.7 5.7
12.6 12.6 9.4 9.4 0.8 0.8 0.4 0.4 6.5 6.1 0.4 2.4 2.4
11.4 1.2 8.5 0.0 1.6 4.9 3.7 1.2
13.0 0.4 3.3 0.0 5.3 3.7 0.4
100.0
2.7 0.0 2.7 0.0
45.9 85.0 40.5 79.2
0.0 0.3 5.4 5.5
32.4 1.2 32.4 1.2 35.1 1.4 35.1 1.4 27 .0 1.2 27.0 1.2
5.4 0.0 5.4 0.0 2.7 1.2 2.7 1.2
29.7 2.3 29.7 2.3 2.7 0.0 5.4 0.0 5.4 0.0
43.2 0.9 5.4 0.0
35.1 0.6 0.0 0.3 2.7 0.0
21.6 0.9 21.6 0.9
8. I 0.0 40.5 6.1
2.7 0.3 18.9 1.4 0.0 2.6
16.2 0.6 10.8 0.6 2.7 0.6
100.0
0.0 0.0
80.0 71.2 2.0
22.4 4.1 4.1
13.5 13.5 6.7 6.7 0.0 0.0 8.2 8.2 6.1 6.1 0.0 0.0 0.0 6.6 0.0 4.4 2.2 0.0 6.1 6.1 0.0
24.5 2.0
10.2 10.2 2.0 2.0 4.1
Species belonging to three families were found in spinner dolphin stomachs: Oplophoridae: Acantbepbyra quadrispinnsa and Penaeidae: Hemipenaeus sp., and an unidentified species belonging to the Sergestidae. The carapace mean length and range (without the rostrum since most of the rostra were damaged) of the rela- tively intact specimens are shown in Table 4. Mean carapace lengths of A. qgad- rzspinosa are not significantly different in the two dolphin species (Mann- Whitney test, U(o.os) critical = 200; larger calculated U(o,05) = 163).

12 MARINE MAMMAL SCIENCE, VOL. 19, NO. 1, 2003
DISCUSSION
Time of Feeding
The exact time of dolphin capture was not recorded and can be only inferred from the soak time of the driftnets. This varied with the phase of the moon. Generally, nets were set for three to four hours, starting as early as sunset during dark nights and as late as 0100 when approaching full moon. This means that the dolphins were caught anytime between 1800 and 0500. Because of this wide range of time rhat the dolphins may have been captured, only gross generaliza- tions can be made based on the volume and the digestive state of the stomach contents: (1) there was feeding in the afternoon and at night as indicated by a large proportion of semi-digested prey present in the stomach contents, and (2) the foraging period appeared similar in both species.
Similarity and Diversity of Prey Items
Both Fraser’s and spinner dolphins eat mainly fish, especially myctophids, as shown by all three indices. However, the importance of fish in the diet of spin- ner dolphins appears greater than for Fraser’s dolphins as shown by the relative proportion of fish to other prey (87% fish us. 13% cephalopods and crustaceans, by volume). In Fraser’s dolphins, crustaceans and cephalopods appear as equally important; together, contributing about 45% of total prey volume. The analysis of species diversity and dominance of prey further supports this. Even exclusive of the crustaceans, many of which were poorly preserved, the diversity index of the prey species in Fraser’s dolphins was higher than that of the spinner dolphins (Shannon Index of diversity: Fraser’s dolphins = 1.16; spinner dolphins = 0.91), and conversely, the Simpson’s index of dominance was higher in spinner dolphins (0.16) than in Fraser’s dolphins (0.05). This means that although spinner dol- phins overall fed on a wide variety of prey (at least 95 spp. of fish, cephalopods and crustaceans) they tended to feed more on some prey than on others.
A large overlap was observed in the prey species of the two dolphins: i t was highest in cephalopods (58%), next in fishes (46%) and least in crustaceans (40%). The most numerous species in each general food category was the same for both spinner and Fraser’s dolphins (e.g., fish, C. wamzingi; cephalopod, A. line- ata; crustaceans, A. quadrispinosd); however, the rankings of the rest of the prey species were different. This suggests that both species probably feed on the most abundant food available in the area. C. warmingi and Abraliopsis are among the most abundant mesopelagic fish and squid species found in many areas (Clarke 1973, Young 1978, Karnella 1987, Badcock and Arahjo 1988, Gartner 1993, Miya e t a/. 1995, Young et al. 1996 ); and two of the important cephalopod fam- ilies, Cranchiidae and Onychoteuthidae are among the most important families in the diet of cetaceans all over the world (Clarke 1996). Numerous samples of the shrimp A. quadrispinosa were collected in the Philippines (including the Sulu Sea) during the Albatross Philippine Expedition (Chace 1986).
Depth Distribution of Prey ltems
Depth distributions and sizes of the major prey suggest that these two species of dolphins may have different feeding strategies. Spinner dolphins appear to feed primarily on mesopelagic fishes, squids, and crustaceans that migrate to the

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY 13
upper 200 m at night (Fig. 1-3). Many species of myctophids undertake vertical migrations of several hundred meters from a daytime depth range of about 700- 3,000 m to a nighttime depth of about 200 m to near the surface (Clarke 1973, Wisner 1974, McGinnis 1982, Masuda et al. 1984, Karnella 1987, Miya et a/. 1995). Similarly, the most abundant squids found in the diet, the enoploteu- thids, also exhibit extensive vertical migration, from a daytime range of 400- 700 m to a night time range of 100-150 m (Young 1978). Fishermen in the area who collect squids with dipnets have reported seeing spinner dolphins feed- ing near the surface at night. Spinner dolphins may also forage in as deep as 400 m as indicated by the presence of some deep-water non-migrating species like the squids Nototodarus and Mastigoteuthis. Fitch and Brownell (1968) sug- gested that spinner dolphins feed at depths greater than 250 m. However, the very low occurrence of these deep water species and the overall low importance to the diet suggest that they probably spend much less time feeding at these depths than in the upper 200 m (Fig. 2). Moreover, the absence of the deep water caridean shrimp, N. elegans and non-migrating benthic caridean shrimps A . carinata (Chace 1986) supports this suggestion.
Fraser’s dolphin appears to exploit both the upper 200 m, where numerous mesopelagic species migrate at night, and greater depths (to probably at least 600 m) where non-migrating mesopelagics or even mesobenthic species dwell. It also appears to preferentially feed on larger prey items. Groups like C. warming2 (also found abundant in spinner dolphins) and other mesopelagic fishes exhibit- ing die1 vertical migration, vertically migrating squids like Abraliopsis, Onychoteu- this, Chiroteathis, and the caridean shrimp A. quadrispinosa all suggest feeding in the upper 200 m.
The suggestion that Fraser’s dolphin may also forage frequently in deep waters, probably deeper than the spinner dolphin, is supported firstly by the sig- nificantly larger size of one of its major prey, the myctophid L. luminosa (Table 4). The large, non-migrating individuals of this species live between 600 and 1,000 m and their preferred depth is 2 7 0 0 m (Robison and Craddock 1983). Secondly, deep-dwelling and sometimes benthic macrourids were also found fre- quently and in relatively high abundance in the stomachs of Fraser’s dolphins (Fig. 1). Thirdly, there was a frequent occurrence and relatively high abundance of the deep-water squids Nototodams and Mastigoteuthidae (Fig. 2). Moreover, adult Galiteuthis (as indicated by blackened wings on the beak) were found in relatively high abundance in the stomachs. Adults of this group live at 600 to more than 1,000 m. Lastly, the occurrence in nearly all of the stomachs contain- ing crustaceans of the benthic deep dwelling and non-migrating caridean shrimp, A . carinata, and the mesopelagic N. elegans further supports the deep water feed- ing habit (Fig. 3). Acanthephyra carinata is a benthic species known only from the Philippines and Indonesia and was collected by the Albatross Expedition from many sites in the Philippines, including the Sulu Sea, from depths between 3 15 m and 1,4 12 m (Chace 1986). Notostornos elegans is mesopelagic and has been collected from less than 1,000 m. The sizes of N. elegans collected from Fraser’s dolphins (23-45 mm PCL, mean = 37.4 mm -+ 4.32) however were signifi- cantly greater, with most individuals almost twice as big as those collected by the expedition (which was 17-20 mm PCL, Chace 1986).
Examination of the myoglobin content in the muscles of the spinner and Fras- er’s dolphins indicates that these two dolphins are good divers, with Fraser’s dolphin being better than spinner dolphin (Dolar e t al. 1998). Myoglobin

14 MARINE MAMMAL SCIENCE. VOL. I 9, NO. 1.2003
Pzgure 1. Depth distributions of some of fish prey species of Fraser’s (F) and spinner (S) dolphins. Importance expressed as 96 of total number of fish. Gray bar = day; black bar = night, (? = limit on this end not known). When there is only one bar it includes both day and night distributions. Information on depth distributions summarized from Clarke (1973), Karnella, (1987), McGinnis (1982), Hulley and Lutjehams (1989), Miya et al. (1995), Gartner (1993), Frost and McCrone (1979), Fitch and Brownell (1968), Cohen et al. (1990).
contents of primary swimming muscles were 7.1 g/100 g muscle for Fraser’s dol- phin and 5.5 g/100 g muscle for spinner dolphin. Thus, i t appears that Fraser’s dolphin has greater diving abilities than the spinner dolphin, enabling it to ex- ploit prey-rich greater depths. The distribution of spinner and Fraser’s dolphins in the study area also support the hypothesis that Fraser’s dolphin is a deeper diver than the spinner dolphin and is probably tied to deep waters more than the spinner dolphin. Fraser’s dolphin distribution is directly correlated with bot- tom depth, whereas that of the spinner dolphin is not (Dolar 1999).
Comparison with Other Popzllations
The total number of species taken by the spinner dolphins found in this study (95 spp.) is several-fold higher than has been reported in other areas such as the eastern tropical Pacific, where approximately 15-30 species were identified (Fitch and Brownell 1968, Perrin et al. 1973), and in waters near Hawaii where approxi- mately 21 species were reported eaten by spinner dolphins (Wiirsig et ul. 1994). The difference may be explained by the generally high species diversity characteris- tic of the Philippines and of the Indo-Malayan region in general (Fowler 1934, Heaney et ul. 1998). Despite this difference, the results of this study agree with the previous studies in terms of the most prevalent food item that spinner dolphins rake, i.e. myctophids, and the general raxa of other mesopelagic fishes and squids. The majority of the fish prey genera taken by spinner dolphins in the eastern tropi- cal Pacific were also found in the Sulu Sea population. However, the abundant gen- erakpecies differed (Fitch and Brownell 1968, Perrin et ul. 1973). The epipelagic fish groups found in the stomachs of spinner dolphins off Hawaii were absent from

DOLAR B1' AL DOLPHIN FEEDING ECOLOGY 15
Figure 2. Gross summary of depth distributions of various squid taxa found in spin- ner and Fraser's dolphins. Importance expressed as 5% of total number of all cephalopods. Gray bar = day; black bar = night; broken line black = young squids only; ? = limit of distribution not known; NI = no information; F = Fraser's dolphin; S = spinner dol- phin. Information obtained from: Young (1 978), Fitch and Brownell (1 968), Perrin et al. (1973), Robison and Craddock (1983), Voss (1963), Takeda and Okutani (1983), Voss and Voss (1971), Chun (1975), Clarke (1986a, h).
the diet of the Sulu Sea population. However, the most important cephalopod prey, the Enoploteuthidae, was also the major cephalopod group that spinner dolphins of Hawaii feed on (Clarke 1996). Crustaceans have not been reported present in the stomachs of spinner dolphins from the eastern tropical Pacific (Fitch and Brownell 1968, Perrin et al. 1973) bur have been reported for dolphins in Hawaiian waters (Wursig et al. 1994). Reef-living and benthic organisms found in stomachs of spin- ner dolphins caught in gill nets off northern Australia were not found in dolphin stomachs of the Sulu Sea (Hembree 1986 cited in Perrin et al. 1989).
Thirteen of the 21 fish families and two of the three crustacean families preyed on by Fraser's dolphins in the eastern tropical Pacific were also found in the stom- achs of Fraser's dolphins in the Sulu Sea. There was also a similarity in the most abundant fish prey items; e.g., L. liminosa and Chazlliodzls sloani. Cephalopods were mentioned taken by Fraser's dolphins in the eastern tropical Pacific (Robison and Craddock 1983), but identification was not given. Fraser's dolphins off South Afri- ca appear to feed mainly on squids, with 96% (by mass) of their diet composed of Chiroteuthidae (53.8%), Histioteuthidae (3 1.9%) and Octopoteuthidae (10.3%) (Sekiguchi et al., 1992). These three families were also found important in the diet of Fraser's dolphins of the Sulu Sea. Fish comprised only 4% of the diet and no crustaceans were found in the South African population (Sekiguchi et al. 1992).
ACKNOWLEDGMEN'I'S
This project would not have been possible wlthout the help of many people. We thank George Acogido, Colin Wood, Esther Himoya, Joe Pres Gaudiano, Erwin Dolumbal, Heidi

16 MARINE MAMMAL SCIENCE, VOL. 19, NO. 1, 2003
Notostomos elegans
Acanthephyra carinata
Acanthephyra quadrispinosa
Sergestidae
Hemipenaeus SP.
m w I . c o b ~ ~ o 8 8 8 8 8 8 8 0 0 0
!- 3
P.
Depth (rn)
Figure 3. Summary of depth distributions of crustacean prey species of Fraser’s and spinner dolphins. Information obtained from Chace (1986) and Takeda and Okutani (1983). N = night, D = day.
Bellamide, Isidro Bellamide, Jun Bellamide, and Magenta Villegas for helping in the dis- section of animals and processing of the dolphins’ stomach contents; Moonyeen Alava for providing the student volunteers; Cindy Klepadlo and H. J. Walker of the Scripps Institu- tion of Oceanography Vertebrate Collection for fish identification, Mary Wicksten of Texas A&M University for crustacean identification and verification, Leslie Knapp and Jeff Wil- liams of the National Museum of Natural History of the Smithsonian Institution for giving us access to the fish otolith collection of the Smithsonian Institution; and Barb Taylor, Ri- chard Rosenblatt, Robert Shadwick, Paul Dayton, David Woodruff and Richard Neal for valuable comments. Funding for the project was provided by the US. National Science Foundation (NSF) and the International Foundation for Science (IFS, Sweden). Travel in the first year of data collection was supported by the Whale and Dolphin Conservation Society (WDCS).
Appendix I . Regression equations showing relationship between otolith length ( I ) and fish standard length ( x ) , and squid lower rostra1 length ( Y ) and mantle length (ml ) . For species with no regression equations available, length derived from ratio between otolith length and fish standard length.
Species Regression equation Source
Fish Myctophum nitidulum I = 4.860 + 1 9 . 1 4 2 ~ Unpublished data
(J. Caretta, SWFSC) in Robertson and Chivers (1997)
(W. Walker, NWFSC) Symbolophorus evemanni I = 1.982 + 1 4 . 4 5 2 ~ Unpublished data
Cephalopods Abraliopsis sp. ml = 9.80 -t 1 9 . 2 8 ~ Wolff (1 982)
Fish length: otolith length Lampadena luminosa 13.1: 1.0 Nafpaktitis and Paxton, (1968) Ceratoscopelus warmingi 20:O.l Hecht (1987) Cubiceps pauciradidtus 21:l . l Butler (1 979)

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY 17
LITERATURE CITED
BADCOCK, J., AND T. M. H. ARAUJO. 1988. On the significance of variation in a warm water cosmopolitan species, nominally Cerutoscopelus wurmingi (Pisces, Myctophidae). Bulletin of Marine Science 42:1643.
BUTLER, J. L. 1979. The nomeid genus Cubiceps (Pisces) with a description of a new spe- cies, Bulletin of Marine Science 29:226-241.
BURUKOVSKII, R. N. 1982. Key to shrimps and lobsters. Oxonian Press Pvt. Ltd., New Delhi.
CADENAT, J., AND M. DOUTRE. 1959. Notes sur les Delphinidej Ouest-Africains. 5 , Sur un Prodelphinus B long bec capturC au large des c6tes du Senegal, Prodelphinus longirostris (Gray 1928). Bulletin de L‘Institut Fransais d’Afrique Noire, Ser. A,, 21:787-792, pl. 1-6.
CALDWELL, D. K., M. C. CALDWELL AND R. V. WALKER. 1976. First records for Fraser’s dolphin (Lagenodelphzs hosei) in the Atlantic and melon-headed whale (Peponocephulu electru) in the western Atlantic. Cetology 25:1-4.
CHACE, F. A. JR. 1986. The caridean shrimps (Crustacea: Decapoda) of the Albatross Phil- ippine expedition, 1907-1910, Part 4: Families Oplophoridae and Nemarocarcini- dae. Smithsonian Contributions to Zoology 434. 82 pp.
CHUN, C. 1975. The Cephalopoda. Part I: Oegopsida, Part 11: Myopsida. Scientific results of the German deepsea expedition on board the steamship “Valvidia” 1889-1899. Octopoda. Volume 18. Keter Publishing House Jerusalem Ltd.
CLARKE, M. R. 1 9 8 6 ~ . A handbook for the identification of cephalopod beaks. Clarendon Press, Oxford.
CLARKE, M. R. 19866. Cephalopods in the diet of odontocetes. Pages 281-321 in M. M. Bryden and Harrison, eds. Research on dolphins. Clarendon Press, Oxford.
CLARKE, M. R. 1996. Cephalopods as prey. 111. Cetaceans. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 351:1053-1065.
CLARKE, T. A. 1973. Some aspects of the ecology of lanternfishes (Myctophidae) in the Pacific Ocean near Hawaii. Fishery Bulletin, U.S. 7 1:401-433.
COHEN, D. M., T. INADA, T. IWAMOTO AND N. SCIALABBA. 1990. FA0 species catalogue: Gadiform fishes of the world (order Gadiformes). Volume 10. FAO, Rome.
DEBROT, A. O., AND N. B. BARROS. 1992. Notes on a Gervais’ beaked whale, Mesoplodon ewopueus, and a dwarf sperm whale, Kogiu sirnus, stranded in Curacao, Netherlands Antilles. Marine Mammal Science 8:172-178.
DOLAR, M. L. L. 1994. Incidental takes of small cetaceans in fisheries in Palawan, central Visayas and northern Mindanao in the Philippines. Reports of the International Whaling Commission (Special Issue 15):35 5-363.
DOLAR, M. L. L. 1999. Abundance, distribution and feeding ecology of small cetaceans in the eastern Sulu Sea and Taiion Strait, Philippines. Ph.D. dissertation, University of California, San Diego, CA. 241 pp.
DOLAR, M. L. L., P. SUAREZ, P. J. PONGANIS AND G. L. KOOYMAN. 1998. Myoglobin in pelagic cetaceans. Journal of Experimental Biology 202:227-236.
FITCH, J. E., AND R. L. BROWNELL. 1968. Fish otoliths in cetacean stomachs and their im- portance in interpreting feeding habits. Journal of the Fisheries Research Board of Canada 2 5 :2 5 6 1-2 574.
FITCH, J. E., AND D. W. GOTSHALL. 1972. First record of the black scabbardfish, Aphuno- phus curbo, from the Pacific Ocean with some notes on other Californian trichiurid fishes. Bulletin of the Southern California Academy of Sciences 71:ll-18.
FOWLER, W. W. 1934. Descriptions of new fishes obtained from 1907 to 1910 chiefly in the Philippine Islands and adjacent seas. Proceedings of the Academy of Sciences of Philadelphia 85:282-322.
FROST, B. W., AND L. E. MCCRONE. 1979. Vertical distribution, die1 vertical migration and abundance of some mesopelagic fishes in the eastern subarctic Pacific Ocean in summer. Fishery Bulletin, U.S. 76:75 1-770.

18 MARINE MAMMAL SCIENCE, VOL 19, NO 1, 2001
GAGO, F. J. 1993. Morphology of the saccular otoliths of six species of lanternfishes of the genus Synholupburuj (Pisces: Myctophidae). Bulletin of Marine Science 52:949-960.
GARTNIX, J. V. JR. 1993. Patterns of reproduction in the dominant lanternfish species (Pisces: Myctophidae) of the eastern Gulf of Mexico, with a review of reproduction among tropical subtropical Myctophidae. Bulletin of Marine Science 52:72 1-750.
GON, O., AND P. C. HEEMSTRA, ELIS. 1990. Fishes of the Southern Ocean. CTP Book Printers, Cape Town.
HARKONEN, T. 1986. Otoliths of the bonyfishes of the Northeast Atlantic. Danbiu ApS, Hellerup, Denmark.
HEANEY, L. R., D. S. BALE'rE, M. L. L. L)OI.AR, A. C. ALCALA, T. L. DANS, P. C. GON%ALES, N. R. INGLE, M. V. LEPITEN, W. L. K. OLIVER, P. S. ONG, E. A. RICKART, B. R. TABARANZA J R . A N D R. C. B. UTZURRUM. 1998. A Synopsis of the mammalian fauna of the Philippine Islands. Fieldiana: Zoology. Field Museum of Natural His- tory, Chicago. No. 88. 61 pp.
HECHT, T. 1987. A guide to the otoliths of southern ocean fishes. South African Journal of Antarctic Research 17:l-88.
HUI.I.I;Y, P. A,, AND J. R. E. LI~I,JEHAMS. 1989. Lantetnfishes of the southern Benguela re- gion. Part 3. Pseudoceanic-oceanic interface. Annals of the South African Museum 9 8 : 4 0 9 4 3 5 ,
HYSLOP, E. J. 19x0. Stomach content analysis-a review of methods and their application. Journal of Fishery Biology 17:411-429.
KAKNIXLA, C. 1987. Biology of midwater fishes of Bermuda ocriln acre. Smithsonian Con- tributions to Zoology 452:5 1-1 68.
MASUDA, H. , K. AMAOKA, C. T. AIIAGA, UYENO, AND T. YoskirNo, EDS. 1984. The fishes of Japanese Archipelago. Tokai University Press, Tokyo.
MCGINNIS, R. F. 1982. Biogeography of lanternfishes (Myctophidae) south of 30"s. Pages 1-1 10 in D. L. Pawson, ed. Antarctic Research Series No. 35. American Geophysical Union, Washington, DC.
MIYA, M., M. YAMAGUCIII AND M. OKIYAMA. 1995. Midwater fishes off the Pacific coast of Boso Peninsula, Central Japan: Species composition, abundance, biomass, and zoo- geograp h ic affinities, Japanese Journal of Ichthyology 4 2 : 2 3 7-248.
MIYAZAKI, N., AND S. WADA. 1978. F'raser's dolphin, Lagenod&bis hoJei, in the western North Pacific. Scientific Reports of the Whales Research Institute, Tokyo 30:23 1- 244.
NAPPAK'rlTIS, R . G., AND J. R. PAXI'ON. 1968. Review of the lanternfish genus Lumpadmu with description of the new species. Los Angeles County Museum of Natural His- tory Contributions in Science 138: 1-29.
NELSON, J. S. 1994. Fishes of the world. 3rd edition. John Wiley & Sons, Inc., New York, NY.
NOHRIS, K. S., A N D T. P. DOHL. 1980. Behavior of the Hawaiian spinner dolphin, Stenellu lnngtrristris. Fishery Bulletin, U.S. 77:821-849.
ODIJM, E. 1997. Ecology: A bridge between science and society. Sinauer Associates, Inc Sunderland, MA.
PI~IIRIN, W. F,, R . WAKNEK, C. H . Fiscus AND D. B. HOLX 1973. Stomach contents of porpoise, S t r w h spp., and yellowfin tuna, Tbunnus ulbucurcs, in mixed-species aggre- gations, Fishery Bulletin, U.S. 71:1077-1092.
PERKIN, W. F., N. MIYAZAKI AND T. KASUYA. 1989. A dwarf form of the spinner dolphin (Stenellu longirostris) from Thailand. Marine Mammal Science 5:213-227.
PEl<RlN, w. F., M. L. L. DoLAR A N D D. ROUINEAU. 1999. Spinner dolphins (Stenella lungir- u~tris) of the western Pacific and Southeast Asia: Pelagic and shallow water forms. Marine Mammal Science 15:1029-1053.
ROBERTSON, K. M., AND S. CHIVERS. 1997. Prey occurrence in pantropical spotted dol- phins, StenelEu atfenuutu, From the eastern tropical Pacific. Fishery Bulletin, 1J.S. 9 5 : 3 34-3 4 8.

DOLAR ET AL.: DOLPHIN FEEDING ECOLOGY 19
ROBISON, B., AND J. E. CRADDOCK. 1983. Mesopelagic fishes eaten by Fraser’s dolphin, Lagenodelphis hosei. Fishery Bulletin, US. 81:283-289.
Ross, G. J. B. 1984. The smaller cetaceans of the south east coast of southern Africa. An- nals of the Cape Province Museum (Natural History) 15:173-410.
SEKIGUCHI, K., N. T. W. KLAGES AND P. B. BEST. 1992. Comparative analysis of the diets of smaller odontocete cetaceans along the coast of Southern Africa. South African Journal of Marine Science 12343-861.
SMALE, M. J. S., G. WATSON AND T. HECHT. 1995. Otolith atlas of southern African ma- rine fishes. Ichthyological Monographs of the J.L.B. Smith Institute of Ichthyology, Grahamstown. 253 pp.
SWEENEY, M. J., AND C. F. E. ROPER. 1998. Classification, type localities and type reposi- tories of recent Cephalopoda. Pages 561-595 in M. Voss, M. Veccione, R. B. Toll and M. J. Sweeney, eds. Systematics and biogeography of cephalopods. Smithsonian Contributions to Zoology, Smithsonian Institution Press, Washington, DC.
TAKEDA, M., AND T. OKUTANI. 1983. Crustaceans and mollusks trawled off Suriname and French Guiana. Department of Zoology, National Science Museum, Tokyo.
TOBAYAMA, T., M. NISHIWAKI AND H. C. YANG. 1973. Records of the Fraser’s dolphin (Lagenodelphis hosei) in the western North Pacific. Scientific Reports of the Whales Research Institute, Tokyo 2 5 2 5 1-263.
VAN BREE, P. J. H., A. COLLET, E. DESPORTES, E. HUSSENOT AND J. A. RAGA. 1986. Le dauphin de Fraser, Lagenadelphis hosei (Cetacea, Odontoceti), espke nouvelle pour la faune d’Europe. Mammalia 50:57-86.
VON WEILER, W. 1968. Die Otolithen der bathypelagischen Familie Melamphaidae und ihre systematische Bedeutung (Pisces). Senckenbergiana Biologie 49:221-230.
VOSS, G . L. 1963. Cephalopods of the Philippine Islands. Bulletin of the US. National Museum 234. 180 pp.
Voss, G. L., AND G. W. Voss. 1971. Cephalopods of Hong Kong. Government Press, Hong Kong.
WADE, P., AND T. GERRODETTE. 1993. Estimates of cetacean abundance and distribution in the eastern tropical Pacific. Report of the International Whaling Commission 43:477-493.
WATKINS, W. A,, M. A. DAHER, K. FRISTRIJP AND N. DI SCIARA. 1994. Fishing and acous- tic behavior of Fraser’s dolphin (tagenudelphis hosei) near Dominica, southeast Carib- bean. Caribbean Journal of Science 30:76-82.
WILLIAMS, R., AND A. MCELDOWNEY. 1990. A guide to the fish otoliths from waters off the Australian Antarctic Territory, Heard and Macquarie islands. Anare Research Notes 75. Antarctic Division, Australia. 173 pp.
WISNER, R. L. 1974. The taxonomy and distribution of lanternfishes (family Myctophi- dae) of the eastern Pacific Ocean. Naval Ocean Research and Development Activity Report 3. NORDA, Stennis Field, MS. 229 pp.
WOLFF, G. A. 1982. Beak key for eight eastern tropical Pacific cephalopod species with relationships between their beak dimensions and size. Fishery Bulletin, U S . 80: 3 5 7-370.
WURSIG, B., R. WELU AND K. S. NORRIS. 1994. Food and feeding. Pages 216-231 in K. S. Norris, B. Wursig, R. S. Wells and M. Wursig, eds. The Hawaiian spinner dolphin. University of California Press, Berkeley, CA.
YOUNG, J. W., T. D. LAMB AND R. W. BRADFORD. 1996. Distribution and community structure of midwater fishes in relation to the subtropical convergence off eastern Tasmania, Australia. Marine Biology 126:571-584.
YOUNG, R. E. 1978. Vertical distribution and photosensitive vesicles of pelagic cephalo- pods from Hawaiian waters. Fishety Bulletin, U S . 76:583-615.
Received: 22 December 2000 Accepted: 2 June 2002




















