
Co-existence of Paragonimus harinasutai and Paragonimus bangkokensis metacercariae in fresh water...
6
Co-existence of Paragonimus harinasutai and Paragonimus bangkokensis metacercariae in fresh water crab hosts in central Viet Nam with special emphasis on their close phylogenetic relationship Pham Ngoc Doanh a, b , Hoang Van Hien a , Nariaki Nonaka b , Yoichiro Horii b, ⁎, Yukifumi Nawa c a Institute of Ecology and Biological Resources, Viet Nam Academy of Science and Technology, Hanoi, Viet Nam b Laboratory of Veterinary Parasitic Diseases, Faculty of Agriculture, University of Miyazaki, Miyazaki, Japan c Research Affairs Division, Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand abstract article info Article history: Received 18 October 2011 Received in revised form 30 January 2012 Accepted 30 January 2012 Available online xxxx Keywords: Paragonimus metacercariae P. harinasutai/P. bangkokensis Central Viet Nam During our epidemiological surveys for Paragonimus species in central Viet Nam, we found four morpholog- ically different Paragonimus metacercariae in mountainous crabs. They were identified as metacercariae of Paragonimus westermani, P. bangkokensis, P. proliferus, and P. harinasutai in the order of their prevalence in crab hosts. This is the first discovery of P. harinasutai in Viet Nam, co-inhabiting with P. bangkokensis and other species. Metacercariae of P. harinasutai were given orally to a cat to obtain adult worms. Then, ITS2 and CO1 sequences of metacercariae and adults of P. harinasutai, and metacercariae of P. bangkokensis collect- ed from the same place were determined for analyses of phylogenetic relationships to other P. harinasutai and P. bangkokensis populations as well as related species. The results of molecular analyses showed that P. harinasutai from Quang Binh province of central Viet Nam was almost completely identical with those from Vientiane, Lao PDR; P. bangkokensis from Quang Binh, Viet Nam was also almost completely identical with those from Lao PDR and from Quang Ninh province, Viet Nam. Except for one P. harinasutai isolate from China, all populations of P. harinasutai and P. bangkokensis from Thailand, Lao and Viet Nam make a sin- gle clade in both ITS2 and CO1 trees. In ITS2 sequences, AT deletion and ATC insertion were observed in some isolates of both species, indicating recent gene flow between P. harinasutai and P. bangkokensis. Moreover, be- cause of their extremely high genetic similarities and their co-inhabitation in the same crab hosts found in Thailand, Lao PDR and Viet Nam, they should be considered as the sister species at the early stage of diver- gence. In addition, P. microrchis previously described from Yunnan, China should be placed as the synonym of P. harinasutai, because of their morphological and molecular similarities. © 2012 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Human paragonimiasis is endemic in northern Viet Nam, where Paragonimus heterotremus has been identified as the causative species [1,2]. New patients with P. heterotremus are still encountered in en- demic areas [3]. During epidemiological surveys on Paragonimus metacercariae in freshwater crab hosts in northern Viet Nam, we found, in addition to P. heterotremus, three Paragonimus species in- cluding one new species, P. vietnamensis [4,5], and two new records of P. proliferus [6] and P. bangkokensis [7]. Subsequently we expanded our survey down to central Viet Nam and discovered a new endemic area with very high infection rate of P. westermani metacercariae in crab hosts in Quang Tri province [8]. These series of discoveries suggested the necessity of more extensive survey on the biodiversity of genus Paragonimus in this and neighboring areas. Recently we conducted surveys on Paragonimus in Quang Tri and two adjacent provinces, Quang Binh and Thua Thien Hue, in central Viet Nam. In those study areas, we found P. westermani as the domi- nant species, consistent with our previous data [8]. In addition, meta- cercariae of P. proliferus, P. bangkokensis and P. harinasutai were also found in crab hosts in this area. This is the first report of P. harinasutai in Viet Nam. Since P. harinasutai and P. bangkokensis are phylogenet- ically close with each other [7,9] and often co-inhabiting in the same crab hosts [10–12], their mutual phylogenetic relationship and their relations to other Paragonimus species are discussed. 2. Materials and methods 2.1. Collection of Paragonimus metacercariae Metacercariae of Paragonimus spp. were isolated from individual crabs collected from three provinces, Quang Binh, Quang Tri and Parasitology International xxx (2012) xxx–xxx ⁎ Corresponding author at: Laboratory of Veterinary Parasitic Diseases, Faculty of Ag- riculture, University of Miyazaki, Gakuen-kibanadai-Nishi, Miyazaki 889-2192, Japan. Tel./fax: +81 985 58 7276. E-mail addresses: [email protected] (P.N. Doanh), [email protected] (Y. Horii), [email protected] (Y. Nawa). PARINT-00971; No of Pages 6 1383-5769/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.parint.2012.01.011 Contents lists available at SciVerse ScienceDirect Parasitology International journal homepage: www.elsevier.com/locate/parint Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus harinasutai and Paragonimus bangkokensis metacercariae in fresh wa- ter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016/j.parint.2012.01.011
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Co-existence of Paragonimus harinasutai and Paragonimus bangkokensis metacercariae in fresh water...

Parasitology International xxx (2012) xxx–xxx
PARINT-00971; No of Pages 6
Contents lists available at SciVerse ScienceDirect
Parasitology International
j ourna l homepage: www.e lsev ie r .com/ locate /par in t
Co-existence of Paragonimus harinasutai and Paragonimus bangkokensismetacercariae in fresh water crab hosts in central Viet Nam with special emphasis ontheir close phylogenetic relationship
Pham Ngoc Doanh a,b, Hoang Van Hien a, Nariaki Nonaka b, Yoichiro Horii b,⁎, Yukifumi Nawa c
a Institute of Ecology and Biological Resources, Viet Nam Academy of Science and Technology, Hanoi, Viet Namb Laboratory of Veterinary Parasitic Diseases, Faculty of Agriculture, University of Miyazaki, Miyazaki, Japanc Research Affairs Division, Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand
⁎ Corresponding author at: Laboratory of Veterinary Priculture, University of Miyazaki, Gakuen-kibanadai-NisTel./fax: +81 985 58 7276.
E-mail addresses: [email protected] (P.N. Doanh)(Y. Horii), [email protected] (Y. Nawa).
1383-5769/$ – see front matter © 2012 Elsevier Irelanddoi:10.1016/j.parint.2012.01.011
Please cite this article as: Doanh PN, et al, Cter crab hosts in central Viet Nam w..., Para
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 October 2011Received in revised form 30 January 2012Accepted 30 January 2012Available online xxxx
Keywords:Paragonimus metacercariaeP. harinasutai/P. bangkokensisCentral Viet Nam
During our epidemiological surveys for Paragonimus species in central Viet Nam, we found four morpholog-ically different Paragonimus metacercariae in mountainous crabs. They were identified as metacercariae ofParagonimus westermani, P. bangkokensis, P. proliferus, and P. harinasutai in the order of their prevalence incrab hosts. This is the first discovery of P. harinasutai in Viet Nam, co-inhabiting with P. bangkokensis andother species. Metacercariae of P. harinasutai were given orally to a cat to obtain adult worms. Then, ITS2and CO1 sequences of metacercariae and adults of P. harinasutai, and metacercariae of P. bangkokensis collect-ed from the same place were determined for analyses of phylogenetic relationships to other P. harinasutaiand P. bangkokensis populations as well as related species. The results of molecular analyses showed that P.harinasutai from Quang Binh province of central Viet Nam was almost completely identical with thosefrom Vientiane, Lao PDR; P. bangkokensis from Quang Binh, Viet Nam was also almost completely identicalwith those from Lao PDR and from Quang Ninh province, Viet Nam. Except for one P. harinasutai isolatefrom China, all populations of P. harinasutai and P. bangkokensis from Thailand, Lao and Viet Nam make a sin-gle clade in both ITS2 and CO1 trees. In ITS2 sequences, AT deletion and ATC insertion were observed in someisolates of both species, indicating recent gene flow between P. harinasutai and P. bangkokensis. Moreover, be-cause of their extremely high genetic similarities and their co-inhabitation in the same crab hosts found inThailand, Lao PDR and Viet Nam, they should be considered as the sister species at the early stage of diver-gence. In addition, P. microrchis previously described from Yunnan, China should be placed as the synonymof P. harinasutai, because of their morphological and molecular similarities.
© 2012 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Human paragonimiasis is endemic in northern Viet Nam, whereParagonimus heterotremus has been identified as the causative species[1,2]. New patients with P. heterotremus are still encountered in en-demic areas [3]. During epidemiological surveys on Paragonimusmetacercariae in freshwater crab hosts in northern Viet Nam, wefound, in addition to P. heterotremus, three Paragonimus species in-cluding one new species, P. vietnamensis [4,5], and two new recordsof P. proliferus [6] and P. bangkokensis [7]. Subsequently we expandedour survey down to central Viet Nam and discovered a new endemicarea with very high infection rate of P. westermani metacercariae incrab hosts in Quang Tri province [8]. These series of discoveries
arasitic Diseases, Faculty of Ag-hi, Miyazaki 889-2192, Japan.
Ltd. All rights reserved.
o-existence of Paragonimus hsitol Int (2012), doi:10.1016
suggested the necessity of more extensive survey on the biodiversityof genus Paragonimus in this and neighboring areas.
Recently we conducted surveys on Paragonimus in Quang Tri andtwo adjacent provinces, Quang Binh and Thua Thien Hue, in centralViet Nam. In those study areas, we found P. westermani as the domi-nant species, consistent with our previous data [8]. In addition, meta-cercariae of P. proliferus, P. bangkokensis and P. harinasutai were alsofound in crab hosts in this area. This is the first report of P. harinasutaiin Viet Nam. Since P. harinasutai and P. bangkokensis are phylogenet-ically close with each other [7,9] and often co-inhabiting in the samecrab hosts [10–12], their mutual phylogenetic relationship and theirrelations to other Paragonimus species are discussed.
2. Materials and methods
2.1. Collection of Paragonimus metacercariae
Metacercariae of Paragonimus spp. were isolated from individualcrabs collected from three provinces, Quang Binh, Quang Tri and
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011

2 P.N. Doanh et al. / Parasitology International xxx (2012) xxx–xxx
Thua Thien Hue, of the Central Viet Nam (Fig. 1). The procedure ofcrab examination followed the method described previously [4].Freshly isolated metacercariae were used for morphological observa-tion. For molecular study, metacercariae were separately fixed in 70%ethanol and stored at −20 °C until use.
2.2. Experimental infection
Eight Paragonimus metacercariae, which were morphologicallyidentified as P. harinasutai metacercariae collected for the first timein Viet Nam, were orally given to a cat. The fecal samples of the catwere examined for detection of Paragonimus eggs from 1 monthafter infection, and the cat was sacrificed when Paragonimus eggswere detected in the stool sample. All adult worms were extensivelywashed in phosphate-buffered saline. A small part of their posteriorportion were cut and fixed in 70% ethanol for molecular studies. Formorphological examination, adult worms were flattened betweenslide glasses and fixed in 70% ethanol. They were then stained withcarmine, cleared in xylene, mounted individually onto a slide glasswith Canada balsam, and covered with a cover slip for permanentpreparation.
2.3. Molecular phylogenetic analyses
Four metacercariae that were morphologically identified as P. har-inasutai from Quang Binh province and four adults derived from thesame batch of metacercariae were used for molecular analyses. In ad-dition, three metacercariae of P. bangkokensis from Quang Binh prov-ince were also used because P. harinasutai and P. bangkokensis aregenetically very close to each other [7,9]. For molecular analyses, weamplified the nuclear ribosomal second internal transcribed spacerregion (ITS2) using 3S and A28 primers [13] and partial mitochondri-al cytochrome oxidase subunit 1 gene (CO1) using a primer pair ofCO1-1-forward and CO1-2-reverse [14] with the PCR procedure
Fig. 1. A map showing the study sites in Viet Nam and geographical distribution of P. h
Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus hter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016
described in our previous report [4]. The PCR products were purifiedusing Qiaquick PCR purification Kit (Qiagen Inc.), then primed usingBig-Dye terminator cycle sequencing kit v3.1 (ABI) and both strandswere directly sequenced by Genetic Analyzer 3130 (Applied Biosys-tems). Ten of each ITS2 and CO1 sequences determined in this studywere deposited in the GenBank/EMBL/DDBJ nucleotide sequence da-tabase with accession No. AB673406–AB673425. In addition, all ITS2and CO1 sequences of P. bangkokensis and P. harinasutai, and some se-quences of several other Paragonimus species were obtained fromDNA database for analyses. However, some completely identical se-quences of the same species from the same locality were treated assingle operational taxonomic unit. Two sequence data sets, ITS2 andCO1, were set up for reconstructing the phylogenetic trees. Theywere aligned using Clustal-W [15] with default options. Evolutionaryanalyses were conducted in MEGA5 [16]. The best model of nucleo-tide substitution for each sequence data set was tested and MaximumLikelihood trees were subsequently reconstructed using Kimura 2-parameter model with gamma rate heterogeneity (+G) for ITS2tree and Hasegawa–Kishino–Yano model with invariable sites (+ I)for CO1 tree. The statistical confidence of branching patterns wasevaluated by the bootstrap test.
3. Results
3.1. Epidemiology and morphology of P. harinasutai metacercariae
We examined two mountainous crab species collected in thestudy areas; Potamiscus tannanti in Quang Binh province and Vietopo-tamon aluoiensis, which was tentatively identified as Potamiscus sp. inour previous study [8], in Quang Tri and Thua Thien Hue provinces.Metacercariae of Paragonimus spp. were found in crabs in 10 out of14 surveyed communes, but was negative in crabs collected in 4 com-munes in Thua Thien Hue province (Table 1). We found four morpho-logically distinct Paragonimus metacercariae. The most dominant
arinasutai and P. bangkokensis populations showing indels in the ITS2 sequences.
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011

3P.N. Doanh et al. / Parasitology International xxx (2012) xxx–xxx
species was P. westermani that was found in 10 out of 14 surveyedcommunes with varying infection rates of 2.0–86.0% and account for28.1–99.6% of the total metacercariae collected in each site. The infec-tion rate of P. bangkokensis varied from 2.0% to 31.0% and this speciesoccupied 0.4–40.3% of the total metacercariae. Other two species, P.proliferus and P. harinasutai, were found only in two communes ofQuang Binh province with low infection rates. The infection rates ofP. proliferus in the crabs of two communes were 3.0% and 4.0%, andthose of P. harinasutai were 4.0% in both locations (Table 1).
In morphological studies, metacercariae of P. westermani and P.bangkokensis were both medium in size, about 400 μm in diameter,but were distinguishable from each other easily by the structureand thickness of cyst walls and the shape of the worm with or with-out the space in the cyst [7,8]. Paragonimus proliferus metacercariaeappeared as extremely large (2.5×0.7 mm in average) excystedform [6,17]. Metacercariae of P. harinasutai (Fig. 2a) were large andround in shape measuring 615–800×610–800 μm (mean 718±47×696±49 based on 18 metacercariae). The artificially excystedmetacercariae (2 samples, Fig. 2b) were 1200–1300×560–600 μmin size. Ventral sucker (196–200 μm in diameter) is slightly biggerthan oral sucker (120–150×150–170 μm) in size. There is no wrinklein the excretory bladder. These morphological characters of encystedand excysted metacercariae are completely identical to those of theoriginal description of P. harinasutai [11].
3.2. Experimental infection and morphological description of P.harinasutai adult worms
A cat was infected orally with 8 encysted metacercariae of P. har-inasutai. Paragonimus eggs were detected in feces of the cat at120 days post infection and 6 adult worms (with the sample code:AD7–AD12) were obtained from the lungs of the cat.
Bodies of adult worms are elongate measuring 8.6–12.3×3.2–4.1 mm (Fig. 3). Oral sucker is 410–450×533–718 μm (avg.431×678), and ventral sucker is 677–861×677–820 μm (avg.738×738) situated at about one-third from the anterior end of thebody. Vitelline glands widely distributed on the both sides of body.Testes are branched and 820–845×820–1025 μm in size. Ovary is
Table 1Prevalence of Paragonimus metacercariae in crab hosts collected from central Viet Nam.
Province District Communes Crab hosts No. of examined Parag
Quang Binh Bo Trach Thuong Trach P. tannanti 100 P. weP. ba
Phuoc Trach P. tannanti 100 P. weP. proP. ha
Minh Hoa Thuong Hoa P. tannanti 100 P. weP. baP. proP. ha
Trung Hoa P. tannanti 100 P. weP. ba
Quang Tri Dakrong Dakrong V. aluoiensis 100 P. weP. ba
Ba Nang V. aluoiensis 100 P. weP. ba
Huong Hoa Tan Long V. aluoiensis 100 P. weP. ba
Huong Phung V. aluoiensis 50 P. weP. ba
Thua Thien Hue Phong Dien Phong Xuan V.aluoiensis 100 P. weP. ba
Phong My V.aluoiensis P. weP. ba
Phu Loc Loc Dien V. aluoiensis 100Loc Hoa V. aluoiensis 100
A Luoi Hong Van V. aluoiensis 100Hong Thai V. aluoiensis 100
Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus hter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016
923–968×946–1025 μm in size and finely branched. Eggs are oval,measuring 73–79×41–45 μm (avg. 75×43). Among six adultworms, five of them (AD8-12) have mainly single arrangement of cu-ticular spines, few of them split into two (Fig. 4). This is compatiblewith the morphological characteristics of P. harinasutai [11]. Unex-pectedly, one adult worm (AD7) was covered with grouped cuticularspines that are a characteristic feature of P. bangkokensis. This unusualsample having incongruent features of metacercariae and adult stageis likely to be a hybrid specimen and is currently being analyzed toconfirm hybridization of two Paragonimus species.
3.3. Molecular analyses
A total of 11 samples of Paragonimus from Quang Binh province,including 4 metacercariae of P. harinasutai (MC7-9 and MC49), 4adults derived from P. harinasutai metacercariae (AD7–AD10), and 3metacercariae of P. bangkokensis (MC1-MC3), were used for molecu-lar phylogenetic analyses. By direct sequencing, we successfullyobtained ten ITS2 and eleven CO1 sequences from those Paragonimussamples, except for the ITS2 sequence of AD7 sample, which has twopeaks, suggesting possible hybridization. AD7 is still to be analyzed toconfirm its status. The CO1 sequences of P. harinasutai and P. bangko-kensis populations of central Viet Nam were identical in length(399 bp). In contrast, ITS2 sequences of P. harinasutai (465 bp) were2 bp shorter than that of P. bangkokensis (467 bp) due to AT deletion(Table 2). ITS2 and CO1 sequences of P. harinasutai population fromQuang Binh province were almost completely identical (99.7% and99.3% similarities in ITS2 and CO1, respectively) with those of P. har-inasutai population from Lao PDR. These data are higher than similar-ities (98.6% and 93.9%, respectively) between P. harinasutai and P.bangkokensis populations from the same place in Quang Binh prov-ince, Viet Nam.
Molecular phylogenetic tree analyses revealed that all P. harinasu-tai and P. bangkokensis populations from Thailand, Lao PDR and VietNam, and P. microrchis from China showed very high similarities(99.0% in ITS2 and 94.2% in CO1 sequences) to each other and weregrouped in a single clade with bootstrap values of 93% in ITS2 treeand 81% in CO1 tree (Fig. 5a, b). One P. harinasutai isolate from
onimus metacercariae No. of infected (%) No. of metacercaria found (% of total)
stermani 33 (33.0) 497 (98.4)ngkokensis 4 (4.0) 8 (1.6)stermani 10 (2.0) 34 (53.1)liferus 3 (3.0) 17 (26.6)rinasutai 4 (4.0) 13 (20.3)stermani 8 (8.0) 34 (28.1)ngkokensis 5 (5.0) 31 (25.6)liferus 4 (4.0) 40 (33.1)rinasutai 4 (4.0) 16 (13.2)stermani 2 (2.0) 15 (75.0)ngkokensis 2 (2.0) 5 (25.0)stermani 58 (58.0) 277 (98.9)ngkokensis 2 (2.0) 3 (1.1)stermani 68 (68.0) 342 (88.6)ngkokensis 31 (31.0) 44 (11.4)stermani 22 (22.0) 89 (59.7)ngkokensis 16 (16.0) 60 (40.3)stermani 43 (86.0) 2091 (99.6)ngkokensis 4 (8.0) 8 (0.4)stermani 13 (13.0) 72 (98.9)ngkokensis 2 (2.0) 6 (7.7)stermani 7 (7.0) 20 (80.0)ngkokensis 3 (3.0) 5 (20.0)
0000
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011

Fig. 2. Metacercariae of Paragonimus harinasutai collected from crab (Potamiscus tannanti) caught in Quang Binh province. a) Encysted form b) Excysted form.
4 P.N. Doanh et al. / Parasitology International xxx (2012) xxx–xxx
China did not fall into this big clade, but was grouped with P. paishui-hoensis in ITS2 and P. ohirai in CO1 tree.
In the ITS2 tree (Fig. 5a), this large P. harinasutai/P. bangkokensis/P. microrchis clade was divided into three groups according to nucle-otide indels. The first group which have both AT deletion at 241–242site and ATC insertion at 271–273 site includes P. harinasutai fromViet Nam and Lao PDR and P. microrchis from China; the second grouphaving ATC insertion at 271–273 sites includes P. bangkokensis fromLao PDR and from Quang Binh and Quang Ninh provinces of VietNam; and the third group which do not have either AT deletion or
Fig. 3. An adult of P. harinasutai collected from the experimentally infected cat 120 dayspost infection.
Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus hter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016
ATC insertion includes P. bangkokensis from Lai Chau, Viet Nam, andP. bangkokensis and P. harinasutai from Thailand (Table 2; Fig. 5a). Nu-cleotide differences between these groups are only 1.1–1.5%. In addi-tion, ITS2 sequences of P. harinasutai from China and P. paishuihoensisfrom Thailand, which also have ATC insertion, make a distinct groupclose to this P. harinasutai/P. bangkokensis/P. microrchis clade.
In CO1 tree (Fig. 5b), P. harinasutai from Viet Nam and Lao PDRand P. microrchis from China formed a clade as was seen in the ITS2tree, although there is slight variance between them. Paragonimusbangkokensis from Quang Ninh and Quang Binh provinces, Viet Nam,having completely identical ITS2 sequences, were also grouped witheach other in the CO1 tree. However, P. bangkokensis from Lao PDRwere separated from P. bangkokensis from Quang Binh and QuangNinh provinces of Viet Nam in CO1, although their ITS2 sequenceswere completely identical with each other (Fig. 5b). Paragonimus bang-kokensis populations from Lai Chau, Viet Nam and P. bangkokensis andP. harinasutai from Thailand, which were clustered in the same cladein ITS2 tree (Fig. 5a), were also divided into different clade in CO1 tree(Fig. 5b).
Fig. 4. Arrangement of body spines of P. harinasutai.
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011
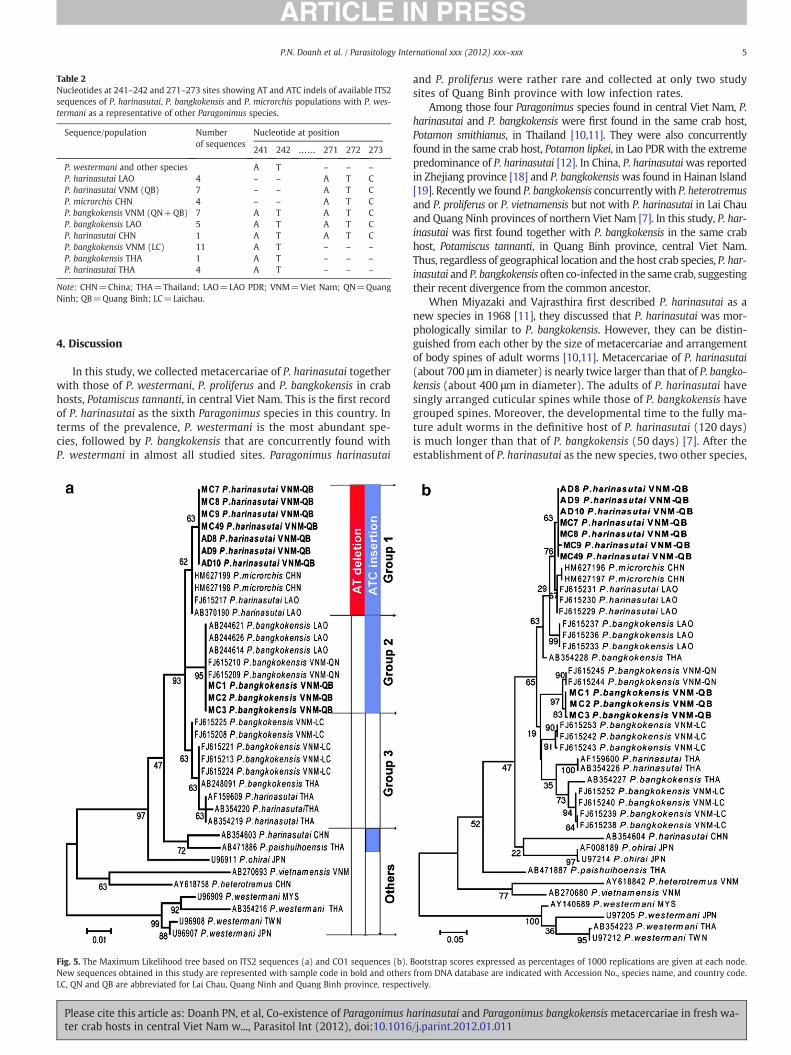
Table 2Nucleotides at 241–242 and 271–273 sites showing AT and ATC indels of available ITS2sequences of P. harinasutai, P. bangkokensis and P. microrchis populations with P. wes-termani as a representative of other Paragonimus species.
Sequence/population Numberof sequences
Nucleotide at position
241 242 …… 271 272 273
P. westermani and other species A T – – –
P. harinasutai LAO 4 – – A T CP. harinasutai VNM (QB) 7 – – A T CP. microrchis CHN 4 – – A T CP. bangkokensis VNM (QN+QB) 7 A T A T CP. bangkokensis LAO 5 A T A T CP. harinasutai CHN 1 A T A T CP. bangkokensis VNM (LC) 11 A T – – –
P. bangkokensis THA 1 A T – – –
P. harinasutai THA 4 A T – – –
Note: CHN=China; THA=Thailand; LAO=LAO PDR; VNM=Viet Nam; QN=QuangNinh; QB=Quang Binh; LC=Laichau.
5P.N. Doanh et al. / Parasitology International xxx (2012) xxx–xxx
4. Discussion
In this study, we collected metacercariae of P. harinasutai togetherwith those of P. westermani, P. proliferus and P. bangkokensis in crabhosts, Potamiscus tannanti, in central Viet Nam. This is the first recordof P. harinasutai as the sixth Paragonimus species in this country. Interms of the prevalence, P. westermani is the most abundant spe-cies, followed by P. bangkokensis that are concurrently found withP. westermani in almost all studied sites. Paragonimus harinasutai
Fig. 5. The Maximum Likelihood tree based on ITS2 sequences (a) and CO1 sequences (b).New sequences obtained in this study are represented with sample code in bold and othersLC, QN and QB are abbreviated for Lai Chau, Quang Ninh and Quang Binh province, respect
Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus hter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016
and P. proliferus were rather rare and collected at only two studysites of Quang Binh province with low infection rates.
Among those four Paragonimus species found in central Viet Nam, P.harinasutai and P. bangkokensis were first found in the same crab host,Potamon smithianus, in Thailand [10,11]. They were also concurrentlyfound in the same crab host, Potamon lipkei, in Lao PDRwith the extremepredominance of P. harinasutai [12]. In China, P. harinasutaiwas reportedin Zhejiang province [18] and P. bangkokensiswas found in Hainan Island[19]. Recentlywe found P. bangkokensis concurrentlywith P. heterotremusand P. proliferus or P. vietnamensis but not with P. harinasutai in Lai Chauand Quang Ninh provinces of northern Viet Nam [7]. In this study, P. har-inasutai was first found together with P. bangkokensis in the same crabhost, Potamiscus tannanti, in Quang Binh province, central Viet Nam.Thus, regardless of geographical location and the host crab species, P. har-inasutai and P. bangkokensis often co-infected in the same crab, suggestingtheir recent divergence from the common ancestor.
When Miyazaki and Vajrasthira first described P. harinasutai as anew species in 1968 [11], they discussed that P. harinasutai was mor-phologically similar to P. bangkokensis. However, they can be distin-guished from each other by the size of metacercariae and arrangementof body spines of adult worms [10,11]. Metacercariae of P. harinasutai(about 700 μm in diameter) is nearly twice larger than that of P. bangko-kensis (about 400 μm in diameter). The adults of P. harinasutai havesingly arranged cuticular spines while those of P. bangkokensis havegrouped spines. Moreover, the developmental time to the fully ma-ture adult worms in the definitive host of P. harinasutai (120 days)is much longer than that of P. bangkokensis (50 days) [7]. After theestablishment of P. harinasutai as the new species, two other species,
Bootstrap scores expressed as percentages of 1000 replications are given at each node.from DNA database are indicated with Accession No., species name, and country code.ively.
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011

6 P.N. Doanh et al. / Parasitology International xxx (2012) xxx–xxx
P. microrchis [20] and P. vietnamensis [5], were reported as morpho-logically similar to P. harinasutai. However, metacercariae of P. hari-nasutai and P. vietnamensis can be distinguished from each other bythe prominent wrinkles of the intestine and the excretory bladderof P. vietnamensis, which could be easily seen in the excysted form[5]. In the adults, the location of ventral sucker of P. harinasutai isabout 1/3 of the anterior part of the bodywhile that of P. vietnamensisis near the middle [5]. The results of molecular analyses clearly showthat P. vietnamensis is far distant from P. harinasutai (Fig. 5a and b). Incontrast, ITS2 and CO1 of P. microrchis from China were almostcompletely identical (100% in ITS2 and 99.7% in CO1 sequences)and clustered with P. harinasutai from Lao PDR (Fig. 5a, b). Moreover,P. microrchis metacercariae were 713 x 664 μm in average size andadult worms have single arranged body spines [20]. These morpho-logical characteristics are identical to P. harinasutai [11]. Therefore,P. microrchis should be placed as a synonym of P. harinasutai.
In this study, the results of molecular analyses revealed that, exceptfor P. harinasutai from China, all P. harinasutai and P. bangkokensis popu-lations from Thailand, Lao PDR and Viet Nam and also P. microrchis fromChina are highly similar to each other to form a species complex basedon both ITS2 and CO1 trees. In comparison with other Paragonimus spe-cies, two types of indels (AT deletion at 241–242 and ATC insertion at271–273) were independently observed in ITS2 sequences amongP. harinasutai/P. bangkokensis/P. microrchis complex. These indels arenot species-specific, but have some tendencies to associate with partic-ular geographical populations (Table 2, Fig. 1). The distribution of theidentical indels in the limited populations of two species indicates recentgene flow between P. harinasutai and P. bangkokensis and recent diver-gence of these two species [21,22]. Among other Paragonimus species,ATC insertion was only found in one ITS2 sequence of P. paishuihoensisThai isolate which was clustered with P. harinasutai isolate from China,indicating their close relationship as was reported previously [23,24].
About 50 species of Paragonimus, most of whichwere found in nature,were identified based on morphological characteristics of metacercariaeand adultworms [19]. The shape, size, cystwall structure and the positionof metacercaria in the cyst have been used for characterization of meta-cercariae. For adult worms, the shape of body and organs, position andratio of suckers, the size and the degree of branching of ovary and testesas well as arrangement of cuticular spines have been used [19]. However,recentmolecular studies revealed significant incongruence betweenmor-phological and molecular identification on Paragonimus species [19,25].Blair et al. [25] proposed that some species were considered to be syno-nyms or subspecies, orwere grouped in a species complex, such as P. wes-termani or P. skrjabini species complex [19,25]. On the other hand, ourprevious [7] and present study indicated that in spite of high molecularsimilarities and co-habitation in the same crabs in the same localities,P. harinasutai or P. bangkokensis still maintain their species identities inevery location. At the present time we tentatively consider them as sisterspecies at early stage of divergence. Since Paragonimus species are, likemost of other trematodes, hermaphroditic, their actualmode of reproduc-tion such as the frequency of heterogamy as well as the possible co-sharing of thedefinitivehostmammals by two species shouldbe exploredin detail to elucidate how can they maintain their species identity.
In conclusion, in the present study, P. harinasutaiwasfirst found as thesixth Paragonimus species in Viet Nam. In central Viet Nam, P. harinasutaiis co-inhabitingwith P. bangkokensis and other two species, P. westermaniand P. proliferus. The combination ofmorphological, biological andmolec-ular data in this study indicated that P. harinasutai and P. bangkokensis aresister species at early stage of divergence. Our series ofwork on P. bangko-kensis and P. harinasutai highlighted the difficulties in determining thespecies boundaries of Paragonimus spp.
Please cite this article as: Doanh PN, et al, Co-existence of Paragonimus hter crab hosts in central Viet Nam w..., Parasitol Int (2012), doi:10.1016
Acknowledgements
One of the authors, Pham Ngoc Doanh, received Postdoctoral Fel-lowship No.P10111 from the Japanese Society for Promotion of Science(JSPS). This study was also supported by Viet NamNational Foundationfor Science and Technology Development (No. 106.12.53.09).
References
[1] De NV, Murrell KD, Cong LD, Cam PD, Chau LV, Toan ND, et al. The food-born trem-atode zoonoses of Viet Nam. The Southeast Asian Journal of Tropical Medicine andPublic Health 2003;34(Suppl. 1):12–34.
[2] Le TH, De NV, Blair D, McManus DP, Kino H, Agatsuma T. Paragonimus heterotremusChen et Hsia, 1964 in Viet Nam: a molecular identification and relationships of iso-lates from different hosts and geographical origins. Acta Tropica 2006;98:25–33.
[3] DoanhPN, ThachDTC,DungDT, ShinoharaA,Horii Y,NawaY.Humanparagonimiasis inViet Nam: epidemiological survey and identification of the responsible species by DNAsequencing of eggs in patients’ sputum. Parasitology International 2011;60:534–7.
[4] Doanh PN, Shinohara A, Horii Y, Habe S, Nawa Y, The DT, et al. Morphological andmolecular identification of two Paragonimus spp., of which metacercariae concur-rently found in a land crab, Potamiscus tannanti, collected in Yenbai Province, VietNam. Parasitology Research 2007;100:1075–82.
[5] Doanh PN, Shinohara A, Horii Y, Habe S, Nawa Y, Le NT. Description of a new lungfluke species, Paragonimus vietnamensis sp. nov. (Trematoda: Paragonimidae),found in Central Viet Nam. Parasitology Research 2007;101:1495–501.
[6] Doanh PN, Shinohara A, Horii Y, Habe S, Nawa Y, Le NT. Discovery of Paragonimusproliferus in Central Viet Nam and their molecular phylogenetic status amonggenus Paragonimus. Parasitology Research 2008;102:677–83.
[7] Doanh PN, Shinohara A, Horii Y, Yahiro S, Habe S, Vannavong N, et al. Morpholog-ical differences and molecular similarities between Paragonimus bangkokensis andP. harinasutai. Parasitology Research 2009;105:429–39.
[8] Doanh PN, Shinohara A, Horii Y, Habe S, Nawa Y. Discovery of Paragonimus wester-mani in Viet Nam and its molecular phylogenetic status in P. westermani complex.Parasitology Research 2009;104:1149–55.
[9] Nawa Y, Doanh PN. Paragonimus bangkokensis and P. harinasutai: reappraisal ofphylogenetic status. J Trop Med Parasitol 2009;32:87–96.
[10] Miyazaki I, Vajrasthira S. On a new lung fluke, Paragonimus bangkokensis sp. nov.in Thailand (Trematoda: Troglotrematidae). Japan J Med Sci Biol 1967;20:243–9.
[11] Miyazaki I, Vajrasthira S. On a new lung fluke found in Thailand, Paragonimus har-inasutai sp. nov. (Trematoda, Troglotrematidae). Annals of Tropical Medicine andParasitology 1968;62:81–7.
[12] Odermatt P, Habe S, Maichanh S, Tran DS, Duong V, ZhangW, et al. Paragonimiasisand its intermediate hosts in a transmission focus in Laos People's Democratic Re-public. Acta Tropica 2007;103:108–15.
[13] Bowles J, Blair D, McManus DP. A molecular phylogeny of the human schisto-somes. Molecular Phylogenetics and Evolution 1995;4:103–9.
[14] Thaenkham U, Waikagul J. Molecular phylogenetic relationship of Paragonimuspseudoheterotremus. The Southeast Asian Journal of Tropical Medicine and PublicHealth 2008;39:217–21.
[15] Thompson JD, Higgins DG, Gibson TJ. CLUSTAL-W: improving the sensitivity of progres-sive multiple sequence alignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Research 1994;22:4673–80.
[16] Tamura K, PetersonD, PetersonN, StecherG,NeiM, Kumar S.MEGA5:molecular evo-lutionary genetics analysis using maximum likelihood, evolutionary distance, andmaximum parsimony methods. Molecular Biology and Evolution 2011;28:2731–9.
[17] Zhou BJ. Studies on Paragonimus proliferus. Chin J Parasitol Parasitic Dis 2004;22:109–12.
[18] Qian BZ, Waikagul J, Dekumyoy P, Yoonuan T, Komalamisra C. Occurrence of Para-gonimus harinasutai in Zhejiang, China. J Trop Med Parasitol 2005;28:22–5.
[19] Blair D, Xu ZB, Agatsuma T. Paragonimiasis and the genus Paragonimus. Advancesin Parasitology 1999;42:113–222.
[20] Hsia TK, Chou PC, Chang HC. A primary report on a new species Paragonimusmicrorchis sp. nov. Data Collection of Kunming Medical College 1978;4:1–9.
[21] Nei M, Kumar S. Molecular evolution and phylogenetics. Oxford; 2000.[22] Britten RJ, Rowen L, Williams J, Cameron RA. Majority of divergence between
closely related DNA samples is due to indels. Proceedings of the National Acade-my of Sciences 2003;100:4661–5.
[23] Cui AL, Chang ZS, ChenMG, Blair D, Chen SH, Zhang YN, et al. Taxonomic status in DNAsequences of five species of genus Paragonimus. Chin J Parasitol Parasitic Dis 2003;21:27–30.
[24] Rangsiruji A, Binchai S, Qian BZ, Morishimia Y, Sugiyama H. A new form of Para-gonimus metacercariae isolated from freshwater crabs: possible discovery of P.paishuihoensis in Thailand. Proc JITMM 2008:137.
[25] Blair D, Agatsuma T, Wang WL. Paragonimiasis. In: Murrell KD, Fried B, editors.World class parasites, vol. 11, food-borne parasitic zoonoses. New York: Springer;2007. p. 117–50.
arinasutai and Paragonimus bangkokensismetacercariae in fresh wa-/j.parint.2012.01.011




















