
Köhler integrators embedded into illumination optics add functionality
Upload
independentCategory
view
2download
0
Gene 426 (2008) 23–31
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r.com/ locate /gene
Cloning, genomic organization and functionality of 5-HT7 receptor splice variantsfrom mouse brain
Evelien Gellynck a,1, Koen Laenen a,1, Kjetil Wessel Andressen b, Béatrice Lintermans a, Kim De Martelaere a,Anne Matthys a, Finn Olav Levy b, Guy Haegeman a, Peter Vanhoenacker a,2, Kathleen Van Craenenbroeck a,⁎,2
a Laboratory for Eukaryotic Gene Expression and Signal Transduction (LEGEST), Ghent University, K.L. Ledeganckstraat 35, 9000 Gent, Belgiumb Department of Pharmacology and Center for Heart Failure Research, Faculty of Medicine, University of Oslo, N-0316 Oslo, Norway
Abbreviations: 5-HT, 5-hydroxytryptamine; 5-CTANOVA, analysis of variance; BSA, bovine serum albumito RNA; CHO, Chinese hamster ovary; DTT, dithiotrereceptor; HEK293, human embryonic kidney; HRP, hphosphate buffered saline; PCR, polymerase chain reactioPCR, reverse transcription PCR; SDS, sodium dodecpolyacrylamide gel electrophoresis; TBS, Tris-buffered s⁎ Corresponding author. Tel.: +32 9/264 51 35; fax: +3
E-mail address: [email protected] Share first authorship.2 Share senior authorship.
0378-1119/$ – see front matter © 2008 Elsevier B.V. Alldoi:10.1016/j.gene.2008.08.011
a b s t r a c t
a r t i c l e i n f oArticle history:
The serotonin (5-HT) 5-HT7 Received 3 July 2008Received in revised form 13 August 2008Accepted 13 August 2008Available online 27 August 2008Keywords:Serotonin receptorG-protein-coupled receptorAlternative splicingCarboxyl terminus
receptors are expressed in both the central nervous system and in peripheraltissues. Receptor distribution studies and pharmacological studies have established that 5-HT7 receptors playan important role in the control of circadian rhythms and thermoregulation. Selective 5-HT7 receptor ligandshave potential therapeutic applications for the treatment of pain and migraine, schizophrenia, anxiety,cognitive disturbances and inflammation.
We have cloned two novel C-terminal splice variants of the 5-HT7 receptor from mouse brain. These twonew splice variants have almost identical sequences as the rat 5-HT7(b) and 5-HT7(c) splice variants and soweregiven the same name. Ligand binding assays ([3H]5-CT), membrane localization and functional studies intransiently transfected cells indicated that all three splice variants are well expressed on the membrane andno major differences in their respective pharmacology and their ability to activate adenylyl cyclase wereobserved. This is in analogy with previous reports comparing either the rat or the human variants.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Serotonin (5-HT, 5-hydroxytryptamine) is one of themost commonneurotransmitters in the brain. Its biological effects aremediated by atleast 14 different serotonin receptors, of which 13 belong to thesuperfamily of membrane-spanning G-protein-coupled receptors(GPCRs).
5-HT7 receptors are expressed in both the central nervous systemand in peripheral tissues. In the central nervous system, they are foundin the thalamus, the hypothalamus (including the suprachiasmaticnucleus), the hippocampus (all CA regions), cerebral cortex (both onpyramidal and GABAergic neurons), amygdala and dorsal raphenucleus (Bonaventure et al., 2004; Thomas and Hagan, 2004). Most5-HT7 receptors are localized on both the soma and axon terminals ofGABAergic neurons. The use of specific agonists and antagonists and a
, 5-carboxamidotryptamine;n; cDNA, DNA complementaryitol; GPCR, G-protein-coupledorse radish peroxidase; PBS,n; PEI, polyethyleneimine; RT-yl sulphate; SDS-PAGE, SDSaline; TBS-T, TBS Tween-20.2 9/264 53 04.e (K. Van Craenenbroeck).
rights reserved.
knock-out mouse helped to better define the roles of the 5-HT7receptors, e.g. the control of circadian rhythms and thermoregulation(Lovenberg et al., 1993; Tsou et al., 1994; Guscott et al., 2003; Hedlundet al., 2003). Involvement of 5-HT7 receptors in learning and memoryis suggested by the observed decrease in synaptic plasticity in CA1 andcontextual fear conditioning in 5-HT7 receptor −/− mice (Thomas andHagan, 2004). Importantly, 5-HT7 receptor agonists can be useful inthe treatment of dysfunctional memory in age-related decline andAlzheimer's disease (Perez-Garcia and Meneses, 2005). Functionalstudies have also implicated 5-HT7 receptors in depression and theactivation of rapid-eye-movement (REM) sleep via the modulation ofsuprachiasmatic nucleus neurons (Thomas and Hagan, 2004). Finally,other observations have led to suggestions that selective 5-HT7receptor ligands may have potential therapeutic applications fortreatment of pain, migraine, schizophrenia, anxiety, cognitive dis-turbances and inflammation (Thomas and Hagan, 2004).
In the periphery, 5-HT7 receptors are found primarily in smoothmuscle cells of blood vessels, but also in the gastrointestinal tract,where they have been proposed to be involved in peristalsis, and in ratlumbar dorsal root and sympathetic ganglia. Lower levels have beendetected in kidney, liver, pancreas and spleen, based on mRNAexpression (Bard et al., 1993). The importance of the 5-HT7 receptor insmooth muscle relaxation might indicate involvement in diseasessuch as irritable bowel syndrome or angina (Coupar et al., 2007).
In 1993, the 5-HT7 receptor gene was discovered independently inseveral different laboratories and has to date been cloned frommouse
24 E. Gellynck et al. / Gene 426 (2008) 23–31
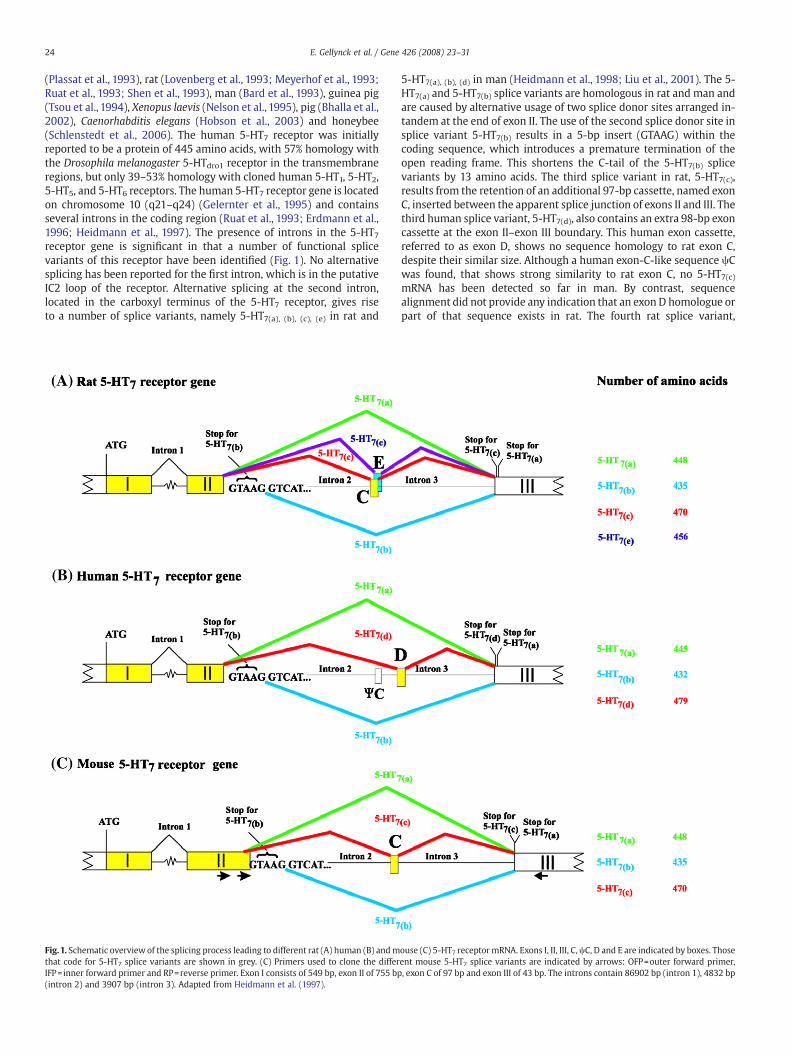
(Plassat et al., 1993), rat (Lovenberg et al., 1993; Meyerhof et al., 1993;Ruat et al., 1993; Shen et al., 1993), man (Bard et al., 1993), guinea pig(Tsou et al., 1994), Xenopus laevis (Nelson et al., 1995), pig (Bhalla et al.,2002), Caenorhabditis elegans (Hobson et al., 2003) and honeybee(Schlenstedt et al., 2006). The human 5-HT7 receptor was initiallyreported to be a protein of 445 amino acids, with 57% homology withthe Drosophila melanogaster 5-HTdro1 receptor in the transmembraneregions, but only 39–53% homology with cloned human 5-HT1, 5-HT2,5-HT5, and 5-HT6 receptors. The human 5-HT7 receptor gene is locatedon chromosome 10 (q21–q24) (Gelernter et al., 1995) and containsseveral introns in the coding region (Ruat et al., 1993; Erdmann et al.,1996; Heidmann et al., 1997). The presence of introns in the 5-HT7receptor gene is significant in that a number of functional splicevariants of this receptor have been identified (Fig. 1). No alternativesplicing has been reported for the first intron, which is in the putativeIC2 loop of the receptor. Alternative splicing at the second intron,located in the carboxyl terminus of the 5-HT7 receptor, gives riseto a number of splice variants, namely 5-HT7(a), (b), (c), (e) in rat and
Fig.1. Schematic overview of the splicing process leading to different rat (A) human (B) andmthat code for 5-HT7 splice variants are shown in grey. (C) Primers used to clone the differIFP= inner forward primer and RP=reverse primer. Exon I consists of 549 bp, exon II of 755 bp(intron 2) and 3907 bp (intron 3). Adapted from Heidmann et al. (1997).
5-HT7(a), (b), (d) in man (Heidmann et al., 1998; Liu et al., 2001). The 5-HT7(a) and 5-HT7(b) splice variants are homologous in rat and man andare caused by alternative usage of two splice donor sites arranged in-tandem at the end of exon II. The use of the second splice donor site insplice variant 5-HT7(b) results in a 5-bp insert (GTAAG) within thecoding sequence, which introduces a premature termination of theopen reading frame. This shortens the C-tail of the 5-HT7(b) splicevariants by 13 amino acids. The third splice variant in rat, 5-HT7(c),results from the retention of an additional 97-bp cassette, named exonC, inserted between the apparent splice junction of exons II and III. Thethird human splice variant, 5-HT7(d), also contains an extra 98-bp exoncassette at the exon II–exon III boundary. This human exon cassette,referred to as exon D, shows no sequence homology to rat exon C,despite their similar size. Although a human exon-C-like sequence ψCwas found, that shows strong similarity to rat exon C, no 5-HT7(c)mRNA has been detected so far in man. By contrast, sequencealignment did not provide any indication that an exon D homologue orpart of that sequence exists in rat. The fourth rat splice variant,
ouse (C) 5-HT7 receptormRNA. Exons I, II, III, C,ψC, D and E are indicated by boxes. Thoseent mouse 5-HT7 splice variants are indicated by arrows: OFP=outer forward primer,, exon C of 97 bp and exon III of 43 bp. The introns contain 86902 bp (intron 1), 4832 bp

Fig. 2. Amplification of different mouse 5-HT7 receptor variants by PCR. (A) PCR withouter forward primer (OFP) and reverse primer (RP), as indicated in Fig. 1C, wasperformed on mouse brain cDNA. (B) Semi-nested PCR was performed with innerforward primer (IFP) and reverse primer (RP) on the PCR product obtained in Fig. 2A(annealing temperature 58 °C). PCR products were separated on a 6% polyacrylamidegel.
25E. Gellynck et al. / Gene 426 (2008) 23–31
5-HT7(e), was discovered after pinpointing a potential 54-bp exoncassette, exon E, which falls just behind exon C and might produce therecently discovered 5-HT7(e) receptor splice variant (Liu et al., 2001). Inguinea pig and pig the 5-HT7 receptor represents a species homologueof the human 5-HT7(a) variant.
Investigation of the distribution of the different splice variants ledto the observation that all variants were present in most tissuesexamined, although relative expression levels differ considerably. Inman and rat, the 5-HT7(a) and 5-HT7(b) splice variants are the mostabundant splice variants, whilst the rat 5-HT7(c) and human 5-HT7(d)splice variants representb4% of the total brain 5-HT7 mRNA(Heidmann et al., 1997; Heidmann et al., 1998; Vanhoenacker et al.,2000; Guthrie et al., 2005).
To date, these splice variants show no major differences in theirrespective pharmacology and signal transduction properties orfunctional coupling to Gs (Heidmann et al., 1998; Krobert et al.,2001). However, there are indications that constitutive adenylylcyclase activity is higher for the human 5-HT7(b) splice variant in aHEK293 stable cell line (Krobert et al., 2002). Furthermore, variation inthe length of their C-termini and in the number of consensus sites forphosphorylation by protein kinases PKA and PKC (Heidmann et al.,1997) raises the possibility that the splice variants can displaydifferences in their desensitisation or trafficking properties. Accord-ingly, the human 5-HT7(d) receptor, in contrast to the 5-HT7(a) and 5-HT7(c) splice variants, has been reported to be constitutivelyinternalized in stable HEK293 T cell lines (Guthrie et al., 2005).Furthermore, the human 5-HT7(b) and the rat 5-HT7(b) and 5-HT7(c)splice variants have been reported to contain recognition sites forbinding of PDZ domain-containing proteins (Hamblin et al., 1998),suggesting that these splice variants may couple to alternative sig-naling pathways.
Here, we report the alternative splicing of the mouse 5-HT7receptor gene, providing at least three splice variants which showdifferences in their C-terminus. The new splice variants are called m5-HT7(b) (accession number FM178515) and m5-HT7(c) (accessionnumber FM178516). Our data do not show a major difference inmembrane expression, pharmacology and functionality between thethree m5-HT7 receptor splice variants.
2. Experimental procedures
2.1. Identification of a mouse 5-HT7 receptor genomic clone
The known cDNA sequence of the mouse 5-HT7(a) receptor(GI:6680328; recently replaced by GI:113865997) was used to searchthe NCBI-database (http://www.ncbi.nlm.nih.gov/BLAST/). We identi-fied a genomic contig (GI:38085349), which contains the completecoding sequence of the mouse 5-HT7 receptor.
2.2. RT-PCR
The cDNAs, corresponding with the different splice variants ofthe mouse 5-HT7 receptor, were detected using semi-nested PCR.PCR amplifications were carried out in a total volume of 30 µl,containing 10 pmol of mouse outer forward primer (TTATCTGTGG-CACCTCCTGT), 10 pmol of mouse reverse primer (GTTCCGCTCTGG-ATCATGTA), 200 µM dNTP, 2.5 U Taq DNA polymerase, 0.5 µgmouse brain cDNA (Marathon-ready mouse cDNA from Clontech,Mountain View, CA) and 2.5 mM MgCl2. The mixture was amplifiedfor 40 cycles, each consisting of a denaturation step (94 °C, 30 s),an annealing step (52 °C–63 °C, 1 min) and an extension step(72 °C, 2 min). PCR products were analyzed by electrophoresis on a6% polyacrylamide gel.
Semi-nested PCR was performed with a mouse inner forwardprimer (GGAATATCAACCGGAAGCTC), and the same mouse reverseprimer under the same conditions as the first PCR.
2.3. Cloning of splice variants
The PCR products, obtained after semi-nested PCR, were isolatedfrom the polyacrylamide gel and cloned into a TOPO TA vector(Invitrogen, Carlsbad, CA). Several clones with different inserts weresequenced and this resulted in the identification of mouse 5-HT7(a),5-HT7(b) and 5-HT7(c) C-termini.
Full-length mouse 5-HT7(a) cDNA was cloned into pcDNA3.1plasmid by inserting the full length receptor cDNA from pAc5-HT7(Obosi et al., 1997), using the restriction enzyme EcoRI. In a next step,mouse 5-HT7(b) and 5-HT7(c) were obtained by exchanging the C-terminus of the 5-HT7(a) pcDNA3.1 vector by the (b) or (c) C-terminalfragments, cloned in the TOPO vectors, using SphI/EcoRI and SphI/PvuII digests, respectively.
2.4. Cell culture and transient transfection
HEK293 and CHO cells were maintained in Dulbecco's modifiedEagle's medium (Invitrogen), supplemented with 100 U/ml penicillin,100 µg/ml streptomycin, and 10% foetal calf serum, at 37 °C, in ahumidified atmosphere with 5% CO2. 4×105 CHO cells weretransiently transfected with JetPEI as described before (Laenen et al.,2007) in 6-well plates with 1 µg mouse 5-HT7 receptor expressionplasmid. 48 h after transfection, the cells were rinsed with ice-coldPBS (phosphate buffered saline) and harvested. HEK293 cells weretransiently transfected in 100 mm dishes with 8 µg DNA using 40 µlLipofectAMINE2000™ (Invitrogen) according to the manufacturer'sinstructions. After transfection, these cells were cultured in Ultra-CULTURE (Cambrex Bio Science Walkersville, Inc., Walkersville, MD),supplemented with L-glutamine (2 mM), penicillin (100 U/ml), andstreptomycin (100 µg/ml) for 48 h.

26 E. Gellynck et al. / Gene 426 (2008) 23–31
2.5. Western immunoblotting
Cells were lysed as previously described (Rondou et al., 2008)in RIPA buffer (50 mM Tris/HCl (pH 7,5), 150 mM NaCl, 1 mMEDTA, 1 mM EGTA, 1% NP40, 0.1% SDS, 0.5% deoxycholic acid),supplemented with freshly added protease inhibitors. For analysisof the lysates, a 5× protein sample buffer (2% SDS, 10% glycerol,0.02% bromophenol blue and 2. 5 mM DTT) was mixed with thelysate. The samples were denatured for 10 min at 95 °C andsubjected to SDS-PAGE. Proteins were subsequently transferredonto a nitrocellulose membrane which was rinsed in TBS (Tris-buffered saline; 10 mM Tris/HCl (pH 8.0), and 150 mM NaCl) andincubated in TBS-T (TBS with 0.1% Tween) containing 5% dried milkfor 1 h at room temperature. Next, the membranes were incubatedovernight at 4 °C with rabbit anti-5-HT7 (Oncogene ResearchProducts, Cambridge, MA) followed by an HRP-conjugated second-
Fig. 3. cDNA sequence alignment of the mouse 5-HT7 receptor variants m5-
ary antibody for 60 min at room temperature. All antibodies werediluted in TBS-T containing 1% dried milk. The membranes werewashed in TBS-T and antigens were visualized by the Luminolchemiluminescence method using a Digital Science Image Station(Kodak, Rochester, NY).
2.6. Immunofluorescence microscopy
CHO cells, grown on glass coverslips were transiently transfected.48 h after transfection, the cells werewashed twicewith PBS and fixedwith PBS, containing 4% paraformaldehyde for 20 min. Aspecificbinding of antibody was reduced by incubating the fixed cells inblocking buffer (5% bovine serum albumin (BSA) and 1% newborn calfserum in PBS) for 2 h at room temperature. Next, cells were incubatedovernight at 4 °C with rabbit anti-5-HT7 (1/500), followed byincubation with fluorescently labeled secondary antibody, anti-rabbit
HT7(a), m5-HT7(b) and m5-HT7(c), emphasizing their different C-termini.
27E. Gellynck et al. / Gene 426 (2008) 23–31
Alexa 488 (1/500; Molecular Probes/Invitrogen) for 1 h at roomtemperature. All antibody dilutions were made in blocking buffer.Finally, cells were incubated with DAPI (4′,6-diamidino-2-phenylin-dole-2HCl) (1/1000 in PBS) for 5 min, mounted with VectaShield(Vector Laboratories, Burlingane, CA) and microscopy was performedusing a Zeiss Axiophot microscope (Oberkirchen, Germany).
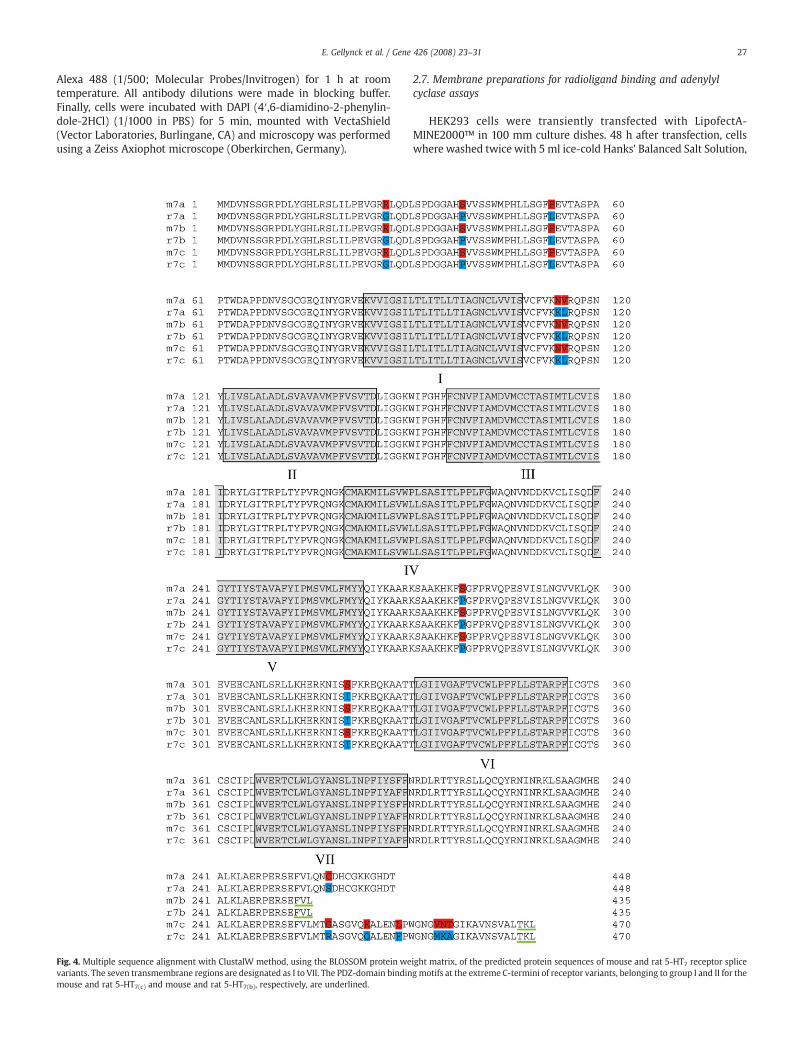
Fig. 4. Multiple sequence alignment with ClustalW method, using the BLOSSOM protein wevariants. The seven transmembrane regions are designated as I to VII. The PDZ-domain bindinmouse and rat 5-HT7(c) and mouse and rat 5-HT7(b), respectively, are underlined.
2.7. Membrane preparations for radioligand binding and adenylylcyclase assays
HEK293 cells were transiently transfected with LipofectA-MINE2000™ in 100 mm culture dishes. 48 h after transfection, cellswhere washed twice with 5 ml ice-cold Hanks' Balanced Salt Solution,
ight matrix, of the predicted protein sequences of mouse and rat 5-HT7 receptor spliceg motifs at the extreme C-termini of receptor variants, belonging to group I and II for the
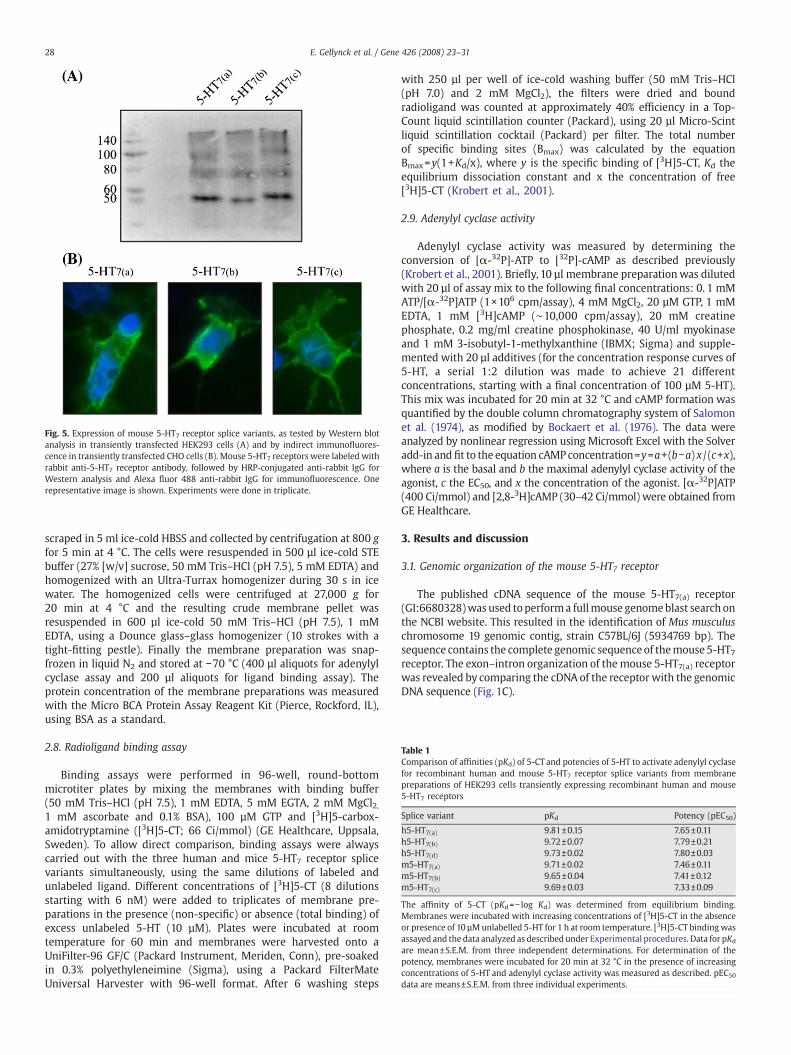
Fig. 5. Expression of mouse 5-HT7 receptor splice variants, as tested by Western blotanalysis in transiently transfected HEK293 cells (A) and by indirect immunofluores-cence in transiently transfected CHO cells (B). Mouse 5-HT7 receptors were labeled withrabbit anti-5-HT7 receptor antibody, followed by HRP-conjugated anti-rabbit IgG forWestern analysis and Alexa fluor 488 anti-rabbit IgG for immunofluorescence. Onerepresentative image is shown. Experiments were done in triplicate.
Table 1Comparison of affinities (pKd) of 5-CT and potencies of 5-HT to activate adenylyl cyclasefor recombinant human and mouse 5-HT7 receptor splice variants from membranepreparations of HEK293 cells transiently expressing recombinant human and mouse5-HT7 receptors
Splice variant pKd Potency (pEC50)
h5-HT7(a) 9.81±0.15 7.65±0.11h5-HT7(b) 9.72±0.07 7.79±0.21h5-HT7(d) 9.73±0.02 7.80±0.03m5-HT7(a) 9.71±0.02 7.46±0.11m5-HT7(b) 9.65±0.04 7.41±0.12m5-HT7(c) 9.69±0.03 7.33±0.09
The affinity of 5-CT (pKd=− log Kd) was determined from equilibrium binding.Membranes were incubated with increasing concentrations of [3H]5-CT in the absenceor presence of 10 µMunlabelled 5-HT for 1 h at room temperature. [3H]5-CT bindingwasassayed and the data analyzed as described under Experimental procedures. Data for pKd
are mean±S.E.M. from three independent determinations. For determination of thepotency, membranes were incubated for 20 min at 32 °C in the presence of increasingconcentrations of 5-HT and adenylyl cyclase activity was measured as described. pEC50
data are means±S.E.M. from three individual experiments.
28 E. Gellynck et al. / Gene 426 (2008) 23–31
scraped in 5 ml ice-cold HBSS and collected by centrifugation at 800 gfor 5 min at 4 °C. The cells were resuspended in 500 µl ice-cold STEbuffer (27% [w/v] sucrose, 50 mM Tris–HCl (pH 7.5), 5 mM EDTA) andhomogenized with an Ultra-Turrax homogenizer during 30 s in icewater. The homogenized cells were centrifuged at 27,000 g for20 min at 4 °C and the resulting crude membrane pellet wasresuspended in 600 µl ice-cold 50 mM Tris–HCl (pH 7.5), 1 mMEDTA, using a Dounce glass–glass homogenizer (10 strokes with atight-fitting pestle). Finally the membrane preparation was snap-frozen in liquid N2 and stored at −70 °C (400 µl aliquots for adenylylcyclase assay and 200 µl aliquots for ligand binding assay). Theprotein concentration of the membrane preparations was measuredwith the Micro BCA Protein Assay Reagent Kit (Pierce, Rockford, IL),using BSA as a standard.
2.8. Radioligand binding assay
Binding assays were performed in 96-well, round-bottommicrotiter plates by mixing the membranes with binding buffer(50 mM Tris–HCl (pH 7.5), 1 mM EDTA, 5 mM EGTA, 2 mM MgCl2,1 mM ascorbate and 0.1% BSA), 100 µM GTP and [3H]5-carbox-amidotryptamine ([3H]5-CT; 66 Ci/mmol) (GE Healthcare, Uppsala,Sweden). To allow direct comparison, binding assays were alwayscarried out with the three human and mice 5-HT7 receptor splicevariants simultaneously, using the same dilutions of labeled andunlabeled ligand. Different concentrations of [3H]5-CT (8 dilutionsstarting with 6 nM) were added to triplicates of membrane pre-parations in the presence (non-specific) or absence (total binding) ofexcess unlabeled 5-HT (10 µM). Plates were incubated at roomtemperature for 60 min and membranes were harvested onto aUniFilter-96 GF/C (Packard Instrument, Meriden, Conn), pre-soakedin 0.3% polyethyleneimine (Sigma), using a Packard FilterMateUniversal Harvester with 96-well format. After 6 washing steps
with 250 µl per well of ice-cold washing buffer (50 mM Tris–HCl(pH 7.0) and 2 mM MgCl2), the filters were dried and boundradioligand was counted at approximately 40% efficiency in a Top-Count liquid scintillation counter (Packard), using 20 µl Micro-Scintliquid scintillation cocktail (Packard) per filter. The total numberof specific binding sites (Bmax) was calculated by the equationBmax=y(1+Kd/x), where y is the specific binding of [3H]5-CT, Kd theequilibrium dissociation constant and x the concentration of free[3H]5-CT (Krobert et al., 2001).
2.9. Adenylyl cyclase activity
Adenylyl cyclase activity was measured by determining theconversion of [α-32P]-ATP to [32P]-cAMP as described previously(Krobert et al., 2001). Briefly, 10 µl membrane preparationwas dilutedwith 20 µl of assay mix to the following final concentrations: 0. 1 mMATP/[α-32P]ATP (1×106 cpm/assay), 4 mM MgCl2, 20 µM GTP, 1 mMEDTA, 1 mM [3H]cAMP (∼10,000 cpm/assay), 20 mM creatinephosphate, 0.2 mg/ml creatine phosphokinase, 40 U/ml myokinaseand 1 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma) and supple-mented with 20 µl additives (for the concentration response curves of5-HT, a serial 1:2 dilution was made to achieve 21 differentconcentrations, starting with a final concentration of 100 µM 5-HT).This mix was incubated for 20 min at 32 °C and cAMP formation wasquantified by the double column chromatography system of Salomonet al. (1974), as modified by Bockaert et al. (1976). The data wereanalyzed by nonlinear regression using Microsoft Excel with the Solveradd-in andfit to the equation cAMPconcentration=y=a+(b−a) x / (c+x),where a is the basal and b the maximal adenylyl cyclase activity of theagonist, c the EC50, and x the concentration of the agonist. [α-32P]ATP(400 Ci/mmol) and [2,8-3H]cAMP (30–42 Ci/mmol) were obtained fromGE Healthcare.
3. Results and discussion
3.1. Genomic organization of the mouse 5-HT7 receptor
The published cDNA sequence of the mouse 5-HT7(a) receptor(GI:6680328)was used toperforma fullmouse genomeblast search onthe NCBI website. This resulted in the identification of Mus musculuschromosome 19 genomic contig, strain C57BL/6J (5934769 bp). Thesequence contains the complete genomic sequenceof themouse 5-HT7receptor. The exon–intron organization of the mouse 5-HT7(a) receptorwas revealed by comparing the cDNA of the receptor with the genomicDNA sequence (Fig. 1C).
29E. Gellynck et al. / Gene 426 (2008) 23–31
3.2. Cloning and sequence analysis of mouse full-length 5-HT7receptor cDNAs
The mouse genomic sequence shows a comparable organization ofintrons and exons as the human 5-HT7 receptor gene. Therefore, weinvestigated the presence of different 5-HT7 receptor splice variantsin the mouse brain. Assuming species comparability, the differentmouse splice variants would also represent splice variants containingthe same final exon and therefore it would be possible to co-amplifyall variants within one single (semi) nested PCR reaction. With the
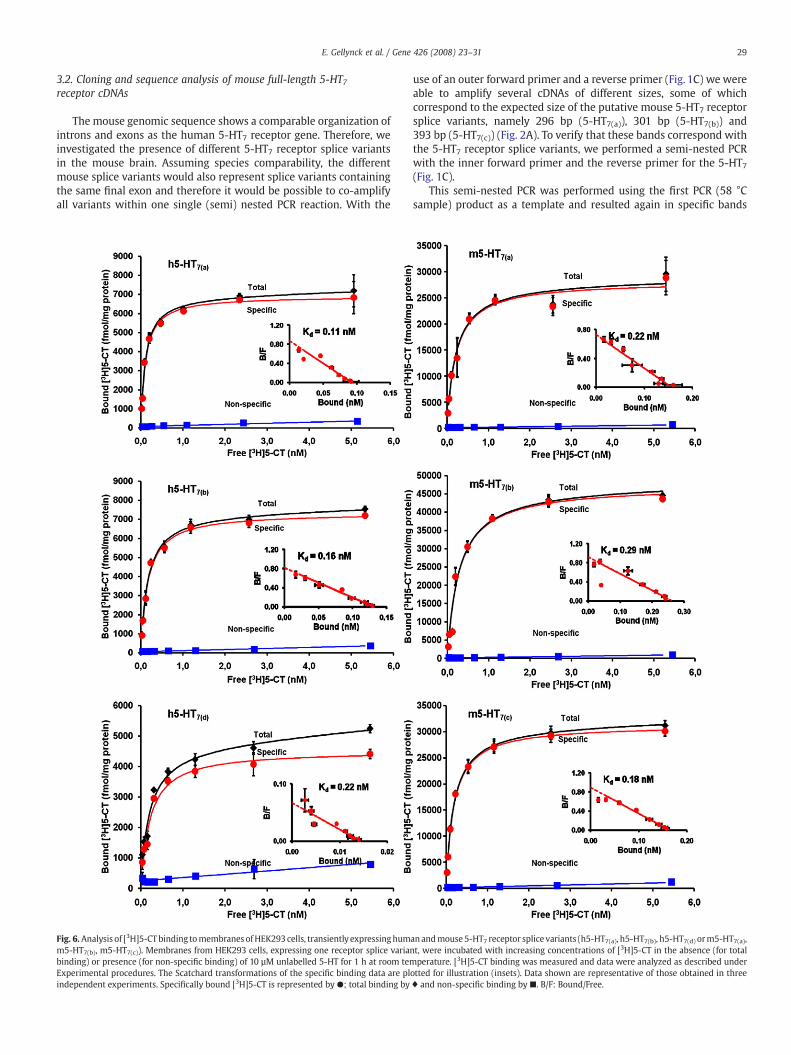
Fig. 6. Analysis of [3H]5-CT binding tomembranes ofHEK293 cells, transiently expressinghumm5-HT7(b), m5-HT7(c)). Membranes from HEK293 cells, expressing one receptor splice varianbinding) or presence (for non-specific binding) of 10 µM unlabelled 5-HT for 1 h at room temExperimental procedures. The Scatchard transformations of the specific binding data are plindependent experiments. Specifically bound [3H]5-CT is represented by ●; total binding by
use of an outer forward primer and a reverse primer (Fig. 1C) we wereable to amplify several cDNAs of different sizes, some of whichcorrespond to the expected size of the putative mouse 5-HT7 receptorsplice variants, namely 296 bp (5-HT7(a)), 301 bp (5-HT7(b)) and393 bp (5-HT7(c)) (Fig. 2A). To verify that these bands correspond withthe 5-HT7 receptor splice variants, we performed a semi-nested PCRwith the inner forward primer and the reverse primer for the 5-HT7(Fig. 1C).
This semi-nested PCR was performed using the first PCR (58 °Csample) product as a template and resulted again in specific bands
an andmouse5-HT7 receptor splice variants (h5-HT7(a), h5-HT7(b), h5-HT7(d) orm5-HT7(a),t, were incubated with increasing concentrations of [3H]5-CT in the absence (for totalperature. [3H]5-CT binding was measured and data were analyzed as described under
otted for illustration (insets). Data shown are representative of those obtained in three♦ and non-specific binding by ■. B/F: Bound/Free.
30 E. Gellynck et al. / Gene 426 (2008) 23–31
with expected lengths for all three 5-HT7 splice variants: 140 bp (5-HT7(a)), 145 bp (5-HT7(b)) and 237 bp (5-HT7(c)) (Fig. 2B). Under thesePCR conditions it was difficult to separate the 5-HT7(a) and 5-HT7(b)splice variants from each other. This is most probably due to the factthat both splice variants only differ by 5 bp in length and that the 5-HT7(a) receptor splice variant most likely predominates and thus willbe more strongly amplified during the semi-nested PCR reaction.
The obtained PCR fragments were isolated from the gel,subcloned into the TOPO TA vector and after sequence verification,recloned into the pcDNA3 vector, giving rise to new variants of themouse 5-HT7 receptor. The full-length mouse 5-HT7 receptor cDNAconsists of 1347 bp for 5-HT7(a), 1308 bp for 5-HT7(b) and 1413 bp for5-HT7(c) (Fig. 3). Cloning and sequencing of the higher bands presentafter the PCR reactions (Fig. 2A: 500 bp and Fig. 2B: 350 bp) indicatedthat these represented non-specific PCR products.
Translation of the cDNA resulted in three open reading frames ofdifferent length, namely 448 amino acids for m5-HT7(a), 435 aminoacids for m5-HT7(b) and 470 amino acids for m5-HT7(c). The alignmentof the deduced amino acid sequences of mouse and rat 5-HT7 splicevariants shows very high homology between the two species (Fig. 4)and therefore, themouse variantswere also designatedm5-HT7(a),m5-HT7(b) and m5-HT7(c). m5-HT7(a) and m5-HT7(b) splice variants bothhave 98% homology with the r5-HT7(a) and r5-HT7(b) splice variants,respectively, and m5-HT7(c) shows 97% homology with r5-HT7(c).
3.3. Expression of the mouse 5-HT7 receptor splice variants in transientlytransfected HEK293 and CHO cells
To investigate whether the cloned cDNAs encode functional 5-HT7receptors, the pcDNA3.1 expression vectors for m5-HT7(a), m5-HT7(b)and m5-HT7(c) were transiently transfected into HEK293 T cells. AfterWestern blot analysis several bands were detected with a 5-HT7receptor specific antibody (Fig. 5A). Analysis of the coding sequence ofthem5-HT7 receptorwithBioEdit software http://www.mbio.ncsu.edu/BioEdit/BioEdit.html, predicted a molecular weight of 49.8 kDa,48.4 kDa and 51.9 kDa for m5-HT7(a), m5-HT7(b) and m5-HT7(c), res-pectively. This is in agreement with the bands observed in Fig. 5A. Thebands at a higher molecular weight most probably represent receptorwith posttranslational modifications, such as ubiquitination or glyco-sylation, or they could represent oligomeric forms of the receptor.
To study the subcellular localization of the mouse 5-HT7 receptor,CHO cells were transiently transfected with the expression plasmidsfor m5-HT7(a), m5-HT7(b) or m5-HT7(c) and receptors were visualizedusing indirect immunofluorescence (Fig. 5B). These results show astronger immunofluorescence on the cell surface as compared to thecytoplasmatic regions, indicating that the different m5-HT7 splice
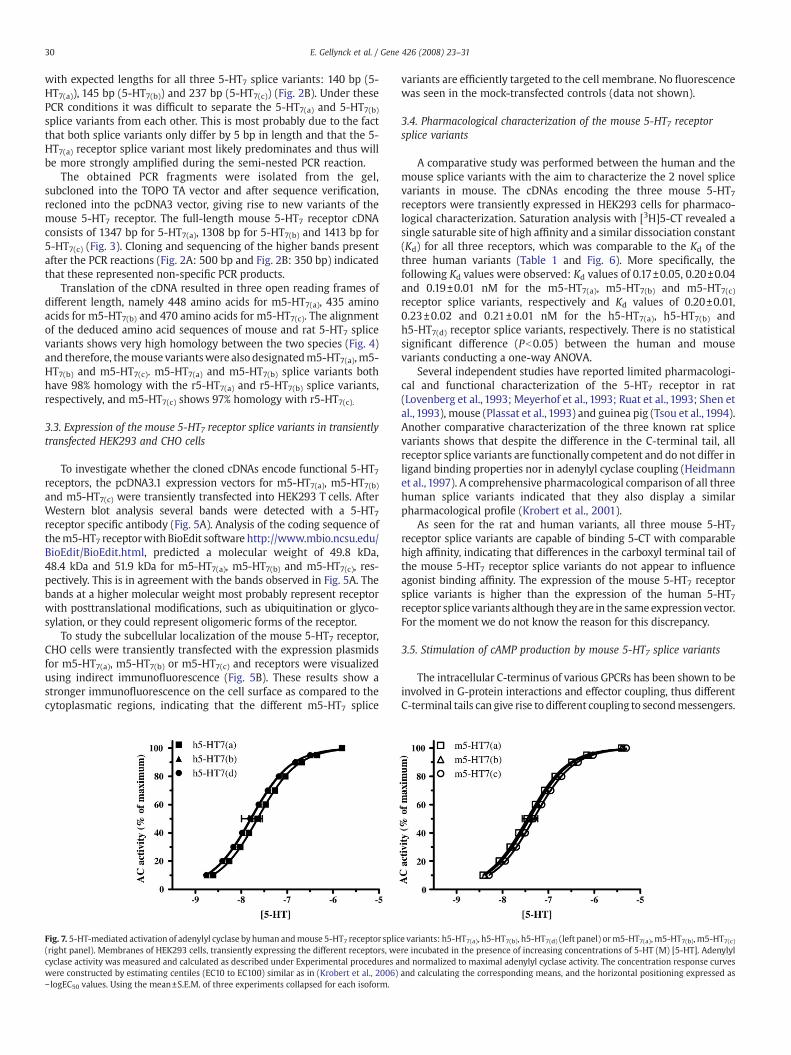
Fig. 7. 5-HT-mediated activation of adenylyl cyclase by human andmouse 5-HT7 receptor splic(right panel). Membranes of HEK293 cells, transiently expressing the different receptors, wecyclase activity was measured and calculated as described under Experimental procedures awere constructed by estimating centiles (EC10 to EC100) similar as in (Krobert et al., 2006)− logEC50 values. Using the mean±S.E.M. of three experiments collapsed for each isoform.
variants are efficiently targeted to the cell membrane. No fluorescencewas seen in the mock-transfected controls (data not shown).
3.4. Pharmacological characterization of the mouse 5-HT7 receptorsplice variants
A comparative study was performed between the human and themouse splice variants with the aim to characterize the 2 novel splicevariants in mouse. The cDNAs encoding the three mouse 5-HT7receptors were transiently expressed in HEK293 cells for pharmaco-logical characterization. Saturation analysis with [3H]5-CT revealed asingle saturable site of high affinity and a similar dissociation constant(Kd) for all three receptors, which was comparable to the Kd of thethree human variants (Table 1 and Fig. 6). More specifically, thefollowing Kd values were observed: Kd values of 0.17±0.05, 0.20±0.04and 0.19±0.01 nM for the m5-HT7(a), m5-HT7(b) and m5-HT7(c)receptor splice variants, respectively and Kd values of 0.20±0.01,0.23±0.02 and 0.21±0.01 nM for the h5-HT7(a), h5-HT7(b) andh5-HT7(d) receptor splice variants, respectively. There is no statisticalsignificant difference (Pb0.05) between the human and mousevariants conducting a one-way ANOVA.
Several independent studies have reported limited pharmacologi-cal and functional characterization of the 5-HT7 receptor in rat(Lovenberg et al., 1993; Meyerhof et al., 1993; Ruat et al., 1993; Shen etal., 1993), mouse (Plassat et al., 1993) and guinea pig (Tsou et al., 1994).Another comparative characterization of the three known rat splicevariants shows that despite the difference in the C-terminal tail, allreceptor splice variants are functionally competent and do not differ inligand binding properties nor in adenylyl cyclase coupling (Heidmannet al., 1997). A comprehensive pharmacological comparison of all threehuman splice variants indicated that they also display a similarpharmacological profile (Krobert et al., 2001).
As seen for the rat and human variants, all three mouse 5-HT7receptor splice variants are capable of binding 5-CT with comparablehigh affinity, indicating that differences in the carboxyl terminal tail ofthe mouse 5-HT7 receptor splice variants do not appear to influenceagonist binding affinity. The expression of the mouse 5-HT7 receptorsplice variants is higher than the expression of the human 5-HT7receptor splice variants although theyare in the sameexpressionvector.For the moment we do not know the reason for this discrepancy.
3.5. Stimulation of cAMP production by mouse 5-HT7 splice variants
The intracellular C-terminus of various GPCRs has been shown to beinvolved in G-protein interactions and effector coupling, thus differentC-terminal tails can give rise to different coupling to secondmessengers.
e variants: h5-HT7(a), h5-HT7(b), h5-HT7(d) (left panel) orm5-HT7(a), m5-HT7(b),m5-HT7(c)re incubated in the presence of increasing concentrations of 5-HT (M) [5-HT]. Adenylylnd normalized to maximal adenylyl cyclase activity. The concentration response curvesand calculating the corresponding means, and the horizontal positioning expressed as

31E. Gellynck et al. / Gene 426 (2008) 23–31
In all species studied, the5-HT7 receptor splice variants positively coupleto adenylyl cyclasewith identical pharmacology (Heidmann et al.,1998;Krobert et al., 2001). By analyzing the conversion of [α-32P]ATP to [32P]cAMP in membranes, we assessed the coupling of the individual mouse5-HT7 splice variants, in comparison with the human 5-HT7 splicevariants, to changes in intracellular adenylyl cyclase activity.
As shown in Table 1 and Fig. 7, the potencies (pEC50) of 5-HT tostimulate adenylyl cyclase activity in transiently transfected HEK293cells are comparable for the different mouse 5-HT7 receptors, rangingfrom 7.33 to 7.80, showing no significant difference conducting a one-way ANOVA (Pb0.05).
Thus, it appears that the differences in the C-terminal ends of themouse 5-HT7 receptors do not translate into an observable differencein functional properties of these receptor splice variants. This is inanalogywith data obtained for both the rat and human 5-HT7 receptorvariants (Heidmann et al., 1998; Krobert et al., 2001).
Acknowledgments
This work was financially supported by Janssen Pharmaceutica N.V.,Beerse, the IWT (Instituut voor de aanmoediging van innovatie doorwetenschap en technologie in Vlaanderen) project 990173 and FWO(Fonds voorwetenschappelijk onderzoekVlaanderen; AMandKVChavea pre- and postdoctoral FWO scholarship, respectively), The NorwegianResearch Council and The Novo Nordisk Foundation. pAc5-HT7 was akind gift from Dr. J. Bermudez (Oxford Brookes University, UK).
References
Bard, J.A., Zgombick, J., Adham, N., Vaysse, P., Branchek, T.A., Weinshank, R.L., 1993.Cloning of a novel human serotonin receptor (5-HT7) positively linked to adenylatecyclase. J. Biol. Chem. 268, 23422–23426.
Bhalla, P., Saxena, P.R., Sharma, H.S., 2002. Molecular cloning and tissue distribution ofmRNA encoding porcine 5-HT7 receptor and its comparison with the structure ofother species. Mol. Cell. Biochem. 238, 81–88.
Bockaert, J., Hunzicker-Dunn, M., Birnbaumer, L., 1976. Hormone-stimulated desensi-tization of hormone-dependent adenylyl cyclase. Dual action of luteninizinghormone on pig graafian follicle membranes. J. Biol. Chem. 251, 2653–2663.
Bonaventure, P., Nepomuceno, D., Hein, L., Sutcliffe, J.G., Lovenberg, T., Hedlund, P.B.,2004. Radioligand binding analysis of knockout mice reveals 5-hydroxytryptamine(7) receptor distribution and uncovers 8-hydroxy-2-(di-n-propylamino)tetralininteraction with alpha(2) adrenergic receptors. Neuroscience 124, 901–911.
Coupar, I.M., Desmond, P.V., Irving, H.R., 2007. Human 5-HT4 and 5-HT7 receptor splicevariants: are they important? Curr. Neuropharmacol. 5, 224–231.
Erdmann, J., et al., 1996. The human serotonin 7 (5-HT7) receptor gene: genomicorganization and systematic mutation screening in schizophrenia and bipolaraffective disorder. Mol. Psychiatry 1, 392–397.
Gelernter, J., Rao, P.A., Pauls, D.L., Hamblin, M.W., Sibley, D.R., Kidd, K.K., 1995.Assignment of the 5HT7 receptor gene (HTR7) to chromosome 10q and exclusion ofgenetic linkage with Tourette syndrome. Genomics 26, 207–209.
Guscott, M.R., et al., 2003. The hypothermic effect of 5-CT in mice is mediated throughthe 5-HT7 receptor. Neuropharmacology 44, 1031–1037.
Guthrie, C.R., Murray, A.T., Franklin, A.A., Hamblin, M.W., 2005. Differential agonist-mediated internalization of the human 5-hydroxytryptamine 7 receptor isoforms.J. Pharmacol. Exp. Ther. 313, 1003–1010.
Hamblin, M.W., Guthrie, C.R., Kohen, R., Heidmann, D.E., 1998. Gs protein-coupledserotonin receptors: receptor isoforms and functional differences. Ann. N. Y. Acad.Sci. 861, 31–37.
Hedlund, P.B., Danielson, P.E., Thomas, E.A., Slanina, K., Carson, M.J., Sutcliffe, J.G., 2003.No hypothermic response to serotonin in 5-HT7 receptor knockout mice. Proc. Natl.Acad. Sci. U. S. A. 100, 1375–1380.
Heidmann, D.E., Metcalf, M.A., Kohen, R., Hamblin, M.W., 1997. Four 5-hydroxytrypta-mine7 (5-HT7) receptor isoforms in human and rat produced by alternativesplicing: species differences due to altered intron–exon organization. J. Neurochem.68, 1372–1381.
Heidmann, D.E., Szot, P., Kohen, R., Hamblin, M.W., 1998. Function and distribution ofthree rat 5-hydroxytryptamine7 (5-HT7) receptor isoforms produced by alternativesplicing. Neuropharmacology 37, 1621–1632.
Hobson, R.J., Geng, J., Gray, A.D., Komuniecki, R.W., 2003. Ser-7b, a constitutively activeGalphas coupled 5-HT7–like receptor expressed in the Caenorhabditis elegans M4pharyngeal motor neuron. J. Neurochem. 87, 22–29.
Krobert, K.A., Levy, F.O., 2002. The human 5-HT7 receptor splice variants: constitutiveactivity and inverse agonist effects. British J. Pharmacol. 135, 1563–1571.
Krobert, K.A., Bach, T., Syversveen, T., Kvingedal, A.M., Levy, F.O., 2001. The clonedhuman 5-HT7 receptor splice variants: a comparative characterization of theirpharmacology, function and distribution. Naunyn Schmiedebergs Arch. Pharmacol.363, 620–632.
Krobert, K.A., Andressen, K.W., Levy, F.O., 2006. Heterologous desensitization is evokedby both agonist and antagonist stimulation of the human 5-HT(7) serotoninreceptor. Eur. J. Pharmacol. 17, 1–10.
Laenen, K., Haegeman, G., Vanhoenacker, P., 2007. Structure of the human 5-HT7receptor gene and characterization of its promoter region. Gene 391, 252–263.
Liu, H., Irving, H.R., Coupar, I.M., 2001. Expression patterns of 5-HT7 receptor isoforms inthe rat digestive tract. Life Sci. 69, 2467–2475.
Lovenberg, T.W., et al., 1993. A novel adenylyl cyclase-activating serotonin receptor (5-HT7) implicated in the regulation of mammalian circadian rhythms. Neuron 11,449–458.
Meyerhof, W., Obermuller, F., Fehr, S., Richter, D., 1993. A novel rat serotonin receptor:primary structure, pharmacology, and expression pattern in distinct brain regions.DNA Cell. Biol. 12, 401–409.
Nelson, C.S., Cone, R.D., Robbins, L.S., Allen, C.N., Adelman, J.P., 1995. Cloning andexpression of a 5HT7 receptor from Xenopus laevis. Recept. Channels 3, 61–70.
Obosi, L.A., Hen, R., Beadle, D.J., Bermudez, I., King, L.A., 1997. Mutational analysis of themouse 5-HT7 receptor: importance of the third intracellular loop for receptor-G-protein interaction. FEBS. Lett. 28, 321–324.
Perez-Garcia, G.S., Meneses, A., 2005. Effects of the potential 5-HT7 receptor agonist AS19 in an autoshaping learning task. Behav. Brain. Res. 163, 136–140.
Plassat, J.L., Amlaiky, N., Hen, R., 1993. Molecular cloning of a mammalian serotoninreceptor that activates adenylate cyclase. Mol. Pharmacol. 44, 229–236.
Rondou, P., Haegeman, G., Vanhoenacker, P., Van Craenenbroeck, K., 2008. BTB proteinKLHL12 targets the dopamine D4 receptor for ubiquitination by a Cul3-based E3ligase. J. Biol. Chem. 283, 11083–11096.
Ruat, M., et al., 1993. Molecular cloning, characterization, and localization of a high-affinity serotonin receptor (5-HT7) activating cAMP formation. Proc. Natl. Acad. Sci.U. S. A. 90, 8547–8551.
Salomon, Y., Londos, C., Rodbell, M., 1974. A highly sensitive adenylate cyclase assay.Anal. Biochem. 58, 541–548.
Schlenstedt, J., Balfanz, S., Baumann, A., Blenau, W., 2006. Am5-HT7: molecular andpharmacological characterization of the first serotonin receptor of the honeybee(Apis mellifera). J. Neurochem. 98, 1985–1998.
Shen, Y., Monsma Jr., F.J., Metcalf, M.A., Jose, P.A., Hamblin, M.W., Sibley, D.R., 1993.Molecular cloning and expression of a 5-hydroxytryptamine7 serotonin receptorsubtype. J. Biol. Chem. 268, 18200–18204.
Thomas, D.R., Hagan, J.J., 2004. 5-HT7 receptors. Curr. Drug Targets CNS Neurol. Disord.3, 81–90.
Tsou, A.P., et al., 1994. Cloning and expression of a 5-hydroxytryptamine7 receptorpositively coupled to adenylyl cyclase. J. Neurochem. 63, 456–464.
Vanhoenacker, P., Haegeman, G., Leysen, J.E., 2000. 5-HT7 receptors: current knowledgeand future prospects. Trends Pharmacol. Sci. 21, 70–77.
Copyright © 2022 FDOKUMEN




















