
Visual target distance, but not visual cursor path length produces shifts in motor behavior
Upload
independentCategory
view
0download
0
http://ini.sagepub.com/Innate Immunity
http://ini.sagepub.com/content/early/2013/10/22/1753425913507096The online version of this article can be found at:
DOI: 10.1177/1753425913507096
published online 22 October 2013Innate ImmunityChang, Cheng-Yeu Wu, Jan Martel, David M Ojcius, Kowit-Yu Chong and John D Young
Tsung-Teng Huang, Hsin-Chih Lai, Young-Bin Chen, Lih-Geeng Chen, Yi-Hui Wu, Yun-Fei Ko, Chia-Chen Lu, Chih-Junginflammasomes in macrophages
-Resveratrol produces anti-inflammatory effects by inhibiting canonical and non-canonicalcis
Published by:
http://www.sagepublications.com
On behalf of:
International Endotoxin & Innate Immunity Society
can be found at:Innate ImmunityAdditional services and information for
http://ini.sagepub.com/cgi/alertsEmail Alerts:
http://ini.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Oct 22, 2013OnlineFirst Version of Record >>
at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from at Chang Gung University on October 24, 2013ini.sagepub.comDownloaded from

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
Original Article
cis-Resveratrol producesanti-inflammatory effects by inhibitingcanonical and non-canonicalinflammasomes in macrophages
Tsung-Teng Huang1,2,3,4, Hsin-Chih Lai1,2,4, Young-Bin Chen5,Lih-Geeng Chen6, Yi-Hui Wu7, Yun-Fei Ko8, Chia-Chen Lu9,Chih-Jung Chang10, Cheng-Yeu Wu1,3,4, Jan Martel1,3,David M Ojcius1,11, Kowit-Yu Chong2 and John D Young1,3,8,12
Abstract
Resveratrol, a natural phenolic compound found in red grapes and wine, exists as cis and trans isomers. Recent studies
have shown that trans-resveratrol possesses anti-inflammatory, anti-oxidant, anti-carcinogenic, anti-tumor and immuno-
modulatory properties. However, it remains unclear whether cis-resveratrol may exhibit similar activities. The objective
of the present study was to examine the effects of cis- and trans-resveratrol on the production of pro-inflammatory
cytokines and mediators in human macrophages. We examined the possibility that cis- and trans-resveratrol may affect
cytokine secretion by modulating inflammasomes, intracellular multi-protein complexes, the assembly of which leads to
caspase-1 activation and secretion of active IL-1b by macrophages. Our results show that pre-treatment of macrophages
with cis-resveratrol not only reduces pro-IL-1b production and IL-1b secretion, but also suppresses ATP-induced tran-
scription and activation of caspase-1 and caspase-4. Notably, cis-resveratrol inhibits the expression of the purinergic
receptor, P2X7R, and the endoplasmic reticulum stress marker, Glc-regulated protein 78, but also reduces reactive
oxygen species production. Moreover, cis-resveratrol attenuates cyclooxygenase-2 expression and prostaglandin
E2 production. cis-Resveratrol also decreases the phosphorylation of p38 MAPK and expression of the c-Jun protein.
These results indicate that cis-resveratrol produces anti-inflammatory effects by inhibiting both the canonical and
non-canonical inflammasomes, and associated pathways in human macrophages.
Keywords
Resveratrol, inflammasome, interleukin-1b, caspase, reactive oxygen species
Date received: 18 April 2013; revised: 29 July 2013; 30 July 2013; accepted: 28 August 2013
1Center for Molecular and Clinical Immunology, Chang Gung University,
Taoyuan, Taiwan, Republic of China2Department of Medical Biotechnology and Laboratory Sciences, College
of Medicine, Chang Gung University, Taoyuan, Taiwan, Republic of China3Laboratory of Nanomaterials, Chang Gung University, Taoyuan, Taiwan,
Republic of China4Research Center of Bacterial Pathogenesis, Chang Gung University,
Taoyuan, Taiwan, Republic of China5Institute of Biotechnology, College of Bio-Resources and Agriculture,
National Taiwan University, Taipei, Taiwan, Republic of China6Department of Microbiology, Immunology and Biopharmaceuticals,
College of Life Sciences, National Chiayi University, Chiayi, Taiwan,
Republic of China7Cancer Research Center, National Cheng Kung University Hospital,
Tainan, Taiwan, Republic of China
8Biochemical Engineering Research Center, Ming Chi University of
Technology, Taipei, Taiwan, Republic of China9Department of Respiratory Therapy, Fu Jen Catholic University, Taipei,
Taiwan, Republic of China10Department of Microbiology and Immunology, College of Medicine,
Chang Gung University, Taoyuan, Taiwan, Republic of China11Molecular Cell Biology, Health Sciences Research Institute, University
of California Merced, Merced, CA, USA12Laboratory of Cellular Physiology and Immunology, Rockefeller
University, New York, NY, USA
Corresponding author:
Hsin-Chih Lai, Center for Molecular and Clinical Immunology,
Department of Medical Biotechnology and Laboratory Sciences, College
of Medicine, Chang Gung University, No. 259, Wen-Hwa 1st Road,
Kweishan, Taoyuan, 333, Taiwan, Republic of China.
Email: [email protected]
Innate Immunity
0(0) 1–16
! The Author(s) 2013
Reprints and permissions:
sagepub.co.uk/journalsPermissions.nav
DOI: 10.1177/1753425913507096
ini.sagepub.com

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
Introduction
Resveratrol (3,4’,5-trihydroxystilbene) represents a nat-ural phenolic phytoalexin found in a wide variety ofplant species, such as red grapes, peanuts and mulber-ries.1,2 It exists as two isomers, cis- and trans-resveratrol. The cis isomer is thought to be producednaturally during grape fermentation as a result of iso-merization of the trans isomer by yeast isomerases; inaddition, cis-resveratrol can be obtained by exposure ofthe trans isomer to sunlight.3,4 Several studies con-ducted within the last few years have shown thattrans-resveratrol exhibits a wide range of biologicalactivities, including anti-inflammatory, anti-oxidant,anti-carcinogenesis, anti-aging, anti-platelet aggrega-tion, immunomodulatory and chemopreventioneffects.5–9 However, only a few studies have been per-formed to examine the biological effects of the cisisomer. cis-Resveratrol has been shown to possessanti-cancer, antimicrobial and anti-platelet aggregationactivities.10–12 Studies are currently under way to deter-mine the properties of cis-resveratrol.
Macrophages are major components of the innateimmune system. The activation of macrophages hasbeen shown to play a pivotal role during the initiationand development of inflammatory responses by produ-cing numerous pro-inflammatory cytokines, such asIL-1b, IL-6 and TNF-a, and other pro-inflammatorymediators, such as NO and prostaglandins.13–15 IL-1band IL-18, which are members of the IL-1 cytokinesuperfamily, promote a variety of innate immune pro-cesses associated with infection, inflammation andautoimmunity.16,17 IL-1b participates in the generationof systemic and local immune responses against variouspathogens, and has been implicated in the pathogenesisof inflammatory diseases, such as gout, asthma, inflam-matory bowel disease, rheumatoid arthritis and athero-sclerosis.18–20 Similarly, IL-18 plays a critical role in theexecution of antimicrobial and antiviral immunity, andthis cytokine has also been associated with severeinflammatory disorders, such as rheumatoid arthritis,Crohn’s disease, psoriasis, lupus, sarcoidosis and mul-tiple sclerosis.21,22 During inflammation or local infec-tion with pathogens, prostaglandin E2 (PGE2)represents the main prostaglandin derived from arachi-donic acid via the action of cyclooxygenase-2(COX-2).23 Recent studies have shown that COX-2and its downstream product PGE2 contribute to thedevelopment of many chronic inflammatory diseases,such as cardiovascular disease, rheumatoid arthritisand inflammatory bowel disease.24,25
The pro-inflammatory cytokines IL-1b and IL-18are synthesized as inactive precursors (i.e. pro-IL-1band pro-IL-18) and accumulate within the cytosol ofmonocytes and macrophages exposed to or ‘primed’with pathogen-associated molecular patterns like the
bacterial endotoxin, LPS.26 However, LPS by itself isusually insufficient to trigger IL-1b and IL-18 secretionfrom macrophages unless danger-associated molecularpatterns provide the second signal responsible for theactivation of inflammasome complexes and the prote-ase caspase-1, processing of pro-IL-1b and pro-IL-18,and secretion of the mature cytokines from thecells.27–29 In addition, a recent study demonstratedthat inflammasome activation also modulates the pro-duction of pro-inflammatory lipid mediators like pros-taglandins in macrophages,30 indicating that thecanonical inflammasome pathway is linked to prosta-glandin production in these cells.
Extracellular ATP acts as a danger signal releasedfrom injured cells during tissue damage and inflamma-tion; it initiates inflammation and further amplifies andsustains cell-mediated immunity through P2 receptor-mediated purinergic signaling.31 Binding of ATP to theP2X7 receptor (P2X7R) leads to inflammasome activa-tion and secretion of IL-1b and IL-18 by primedmacrophages.32
Activation of caspase-1 requires the assembly andactivation of a cytosolic multi-protein complexesknown as the inflammasome, which is typicallyformed by three components consisting of a nucleotidebinding and oligomerization domain-like, leucine-richrepeat-containing receptor (NLR), the adaptor proteinapoptosis-associated speck-like protein containing acaspase recruitment domain (ASC) and pro-caspase-1.33 Upon activation, oligomerized NLRsinteract with ASC, which, in turn, recruits and activatescaspase-1, and leads to cleavage and activation of pro-IL-1b and pro-IL-18.29 The NLRP1 (NALP1) andNLRP3 (NALP3/cryopyrin) inflammasomes are twoof the best-characterized canonical inflammasomesdescribed so far. A large number of stimuli have beenshown to trigger activation of the NLRP3 inflamma-some, including ATP, monosodium urate crystals, chol-esterol crystals, UVB irradiation, pathogen-derivednucleic acids, silica, asbestos and amyloid-b.34–41 Inaddition, treatment with LPS or muramyl dipeptide(along with ATP) has been reported to induce theNLRP1 inflammasome.42,43
More recently, non-canonical inflammasomes con-taining murine caspase-11 have also been reported.44,45
Caspase-11 does not exist in humans, but the functionof this enzyme is functionally equivalent to humancaspase-4 and caspase-5, which also modulate inflam-masome activity as revealed in previous studies.46,47
The main objective of the present study was to inves-tigate whether cis- and trans-resveratrol inhibit theproduction of pro-inflammatory cytokines (IL-1b andIL-18) and the pro-inflammatory mediator (PGE2) inLPS-primed, ATP-stimulated human macrophages.We also aimed to determine whether cis- and trans-resveratrol modulate inflammasome activation in
2 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
macrophages. We found that cis-resveratrol suppressesIL-1b secretion and PGE2 production by inhibiting thenon-canonical inflammasome pathway and COX-2.These findings suggest that cis-resveratrol possessesanti-inflammatory activities that may prove beneficialfor the treatment of inflammatory diseases.
Materials and methods
Chemicals and reagents
ATP, LPS and phorbol 12-myristate 13-acetate (PMA)were purchased from Sigma-Aldrich (St. Louis, MO,USA). cis-Resveratrol and trans-resveratrol were pur-chased from Cayman Chemical Company (Ann Arbor,MI, USA), and a stock solution of 200mM was pre-pared in absolute ethanol and kept at –20�C away fromlight. Cell culture media (RPMI 1640), FBS, penicillinand streptomycin were purchased from LifeTechnologies (Grand Island, NY, USA). For Westernblot analysis, anti-IL-1b, anti-NF-kB p65, anti-IkB-a,anti-p38, anti-p-p38, anti-ERK1/2, anti-p-ERK1/2,anti-JNK and anti-p-JNK were obtained from CellSignaling Technology (Danvers, MA, USA); anti-caspase-4 was from MBL (Nagoya, Japan); anti-ASC,anti-COX-2, anti-c-Fos, anti-c-Jun, anti-Glc-regulatedprotein 78 (GRP78), anti-pro-IL-1b, anti-IL-18 andanti-P2X7R were from Santa Cruz Biotechnology(Santa Cruz, CA, USA); anti-NLRP3 and anti-P2X4Rwere from Sigma-Aldrich; anti-caspase-1 was fromMillipore (Billerica, MA, USA); anti-NLRP1 wasfrom Enzo Life Sciences (Farmingdale, NY, USA);anti-lamin B1 was from Abcam (Cambridge, MA,USA); and anti-b-actin was from Novus Biologicals(Littleton, CO, USA). HRP-conjugated anti-rabbitand anti-mouse secondary Abs were obtained fromSanta Cruz Biotechnology.
Cell culture and treatments
Human acute monocytic leukemia THP-1 cells [TIB-202, American Type Culture Collection (ATCC),Manassas, VA, USA] were cultured in RPMI 1640medium supplemented with 10% (v/v) heat-inactivatedFBS, 100 units/ml of penicillin, and 100 mg/ml ofstreptomycin. Cell lines were cultured at 37�C in ahumidified incubator containing 5% CO2. The THP-1cells were plated in six-well plates at 2� 106 cells/well.Cellular differentiation of the suspended cells to adher-ent macrophages was induced by overnight culture incomplete medium supplemented with 500 ng/ml ofPMA, followed by culture in fresh complete mediumfor an additional 2 d. THP-1 macrophages were pre-treated for 20 h with the indicated concentrations ofresveratrol or with ethanol as a control, followed bytreatment with LPS (0.5 mg/ml) for 3 h and ATP
(5mM) for 1 h. Cell culture supernatants were har-vested at 14,000 g for 5min at 4�C, and the super-natants were collected and stored at –80�C forcytokine assay. In addition, cell lysates were re-sus-pended in lysis buffer for RNA extraction andWestern blot analysis.
Cell viability assay
Cell viability was determined using a commercial MTT-based in vitro toxicology assay (Sigma-Aldrich), whichdetects viable cells colorimetrically based on the pro-duction of purple formazan by viable cells. THP-1 cellswere seeded at a density of 1� 105 cells/well for 24 h.For macrophage differentiation, cells were treated andincubated with PMA as described above. Cell culturemedia were replaced by complete media containing theindicated concentrations of resveratrol and then incu-bated with cells for 24 h. After incubation, 10 ml ofMTT (5mg/ml) were added to each well and theplates were incubated at 37�C for 4 h. The content ofeach well was eluted and the precipitate was dissolvedwith 100 ml of the MTT solubilization solution.Absorbance was read at 570 nm using a VersaMaxmicroplate reader (Molecular Devices, Sunnyvale,CA, USA). Cell viability (%) was calculated as theratio of surviving cells in the resveratrol-treated groupdivided by that of the control group. All treated sam-ples and controls were tested in triplicate.
Measurement of human IL-1�, IL-18, caspase-1and PGE2 by ELISA
THP-1 macrophages were seeded at a densityof 2� 106 cells/well in six-well plates and were pre-incubated with the indicated concentrations of resvera-trol in 1ml of complete medium for 20 h, followed bytreatment with LPS (0.5 mg/ml) for 3 h and with ATP(5mM) for 1 h. Cell culture supernatants were collectedand centrifuged at 10,000 g for 5min at 4�C to removeinsoluble material. Levels of secreted IL-1b, activatedcaspase-1 and PGE2 in cell culture supernatants weremeasured using commercial ELISA kits (R&DSystems, Minneapolis, MN, USA) based on the manu-facturer’s instructions; IL-18 in cell culture super-natants was measured by using a human IL-18ELISA kit (MBL).
Reactive oxygen species production
Total reactive oxygen species (ROS) production inTHP-1 macrophages was measured using the TotalROS/Superoxide detection kit (Enzo Life Sciences,Farmingdale, NY, USA). Briefly, cells were firstseeded at a density of 1� 105 cells/well in 96-wellplates for 24 h. For macrophage differentiation, cells
Huang et al. 3

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
were treated with PMA as described above. Cell culturemedia were replaced by complete media containingresveratrol at the indicated concentrations and thenincubated for 20 h, followed by treatment with LPS(0.5 mg/ml) for 3 h and with ATP (5mM) for 1 h. Inaddition, cells were treated with the ROS inducer, pyo-cyanin (200mM), at 37�C for 30min as a positive con-trol. After treatment, the cells were washed with 200mlof 1�washing buffer and loaded with 100 ml of ROS/superoxide detection reagents prior to incubation at37�C for 1 h. Absorbance was read at 520 nm using amicroplate reader after excitation at 488 nm. Theincrease in relative fluorescence intensity was used todetermine intracellular ROS production.
Protein extraction and Western blot analysis
Total cellular proteins were isolated from cell lines inRIPA lysis buffer (50mMTris-HCl, pH 7.4,150mMNaCl, 0.25% deoxycholic acid, 1% NonidetP-40, 1mM EDTA) (Millipore) containing a proteaseinhibitor cocktail (Roche, Mannheim, Germany). Cellculture supernatants were harvested as described above.Cytosolic and nuclear proteins were extracted usingcytoplasmic and nuclear protein extraction kits(Thermo Fisher Scientific, Rockford, IL, USA).Protein concentration was determined using the Bio-Rad Bradford assay (Bio-Rad, Hercules, CA, USA).Equal amounts of protein were separated by 8–12%SDS-PAGE. After gel electrophoresis, the proteinswere transferred to polyvinylidene difluoride mem-branes (Millipore) and subsequently stained withPonceau S (Sigma-Aldrich) to confirm protein transfer.The membranes were blocked with 5% non-fat drymilk or BSA in TBS-T (0.1% Tween-20 in 1�TBS,pH 7.4) for 1 h and probed overnight with the indicatedprimary Abs. Protein signal was revealed by enhancedchemiluminescence (Millipore) following treatmentwith HRP-conjugated secondary Ab. Quantificationof protein signal obtained by Western blot analysis
was performed with the LAS-3000 chemiluminescencedetection device (Fujifilm, Valhalla, NY, USA). Toadjust for protein loading, the signal of each proteinwas normalized to b-actin.
Quantitative real-time RT-PCR
Total RNA was extracted from THP-1 cells using atotal RNA mini kit (Geneaid, Taipei, Taiwan) accord-ing to the manufacturer’s instructions. Two micro-grams of RNA were reverse-transcribed in a finalreaction volume of 20 ml containing an oligo dTprimer, dNTP and the SuperScript III reverse tran-scriptase (Invitrogen, Carlsbad, CA, USA). ThecDNA for ASC, caspase-1, caspase-4, GRP78, IL-1b,IL-18, NLRP1, NLRP3, P2X4R, P2X7R and b-actinwere amplified by PCR with the primers listed inTable 1. PCR products were electrophoresed using a2% agarose gel and visualized after staining with eth-idium bromide (Sigma-Aldrich). Quantitative real-timePCR was performed using a LightCycler technology(Roche) with FastStart DNA MasterPLUS SYBRGreen I detection (Roche). Each LightCycler capillarywas loaded with a mixture containing template cDNA,250 nM sense and antisense primers, 4 ml of 5� SYBRGreen Master Mix. In all assays, cDNA was amplifiedusing a standard program (10min denaturing step; 55amplification cycles of 10 s at 95�C, 10 s at 55�C and10 s at 72�C). Real-time PCR was performed using thesame primers as described above. For normalization,the mRNA level of the b-actin gene in each RNA prep-aration was determined. Relative gene expression wasdetermined by using the ��Ct method, whereCt¼ threshold cycle. Each PCR experiment was per-formed in triplicate on two separate occasions.
Statistical analysis
For each experiment, data were obtained in triplicateand presented as means� SD. Mean comparisons
Table 1. Sequences for the quantitative real-time RT-PCR primers.
Gene Forward primer (50-30) Reverse primer (50-30)
ASC ATCCAGGCCCCTCCTCAGT GTTTGTGACCCTCCGCGATAAG
Caspase-1 GAATGTCAAGCTTTGCTCCCTAGA AAGACGTGTGCGGCTTGACT
Caspase-4 GGTCATCATTGTCCAGGC CCATTGTGCTGTCTCTCC
GRP78 GCCTGTATTTCTAGACCTGCC TTCATCTTGCCAGCCAGTTG
IL-1b AAAAGCTTGGTGATGTCTGG TTTCAACACGCAGGACAGG
IL-18 GCTGAACCAGTAGAAGACAATTG ATCTGATTCCAGGTTTTCATCATCT
NLRP1 ACCTGATCCCAAGTGACTGC TCTTCTCCAGGGCTTCGATA
NLRP3 CTTCTCTGATGAGGCCCAAG GCAGCAAACTGGAAAGGAAG
P2X4R GGATGTGGCGGATTATGTGATAC AGTGGTCGCATCTGGAATCTC
P2X7R TGTGCCTACAGGTGCTACGCC GCCCTTCACTCTTCGGAAACTC
b-Actin GAGACCTTCAACACCCCAGCC GGATCTTCATGAGGTAGTCAG
4 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
between cis-resveratrol or trans-resveratrol-treated cellsand untreated control cells were analyzed usingStudent’s t-test. P-Values< 0.05 were considered statis-tically significant.
Results
cis- and trans-Resveratrol produce no toxic effecton human macrophages
To observe whether cis- and trans-resveratrol showcytotoxic effects on human THP-1 macrophages, wetreated the cells with either cis- or trans-resveratrol atvarious concentrations for 24 h, followed by monitor-ing cell viability using the MTT assay. The resveratrolisomers showed no toxic effect on THP-1 macrophages(Figure 1).
cis-Resveratrol attenuates the production of IL-1�at the transcriptional and translational levelsin activated macrophages
To evaluate the effects of cis- and trans-resveratrol onthe production of pro-inflammatory cytokines bymacrophages, we treated THP-1 cells with either cis-or trans-resveratrol (1, 10 or 100 mM for 20 h), thenwith LPS (0.5 mg/ml for 3 h) to induce cytokine expres-sion and, finally, with ATP (5mM for 1 h) to induce
secretion of IL-1b and IL-18. ELISA and Westernblot analyses showed that cis-resveratrol pre-treatmentsignificantly inhibited IL-1b secretion in the cell cul-ture supernatants (Figure 2a, e) and pro-IL-1b pro-duction in whole cell lysates (Figure 2e, f). Incontrast, pre-treatment with trans-resveratrolincreased IL-1b secretion in a dose-dependentmanner (Figure 2a). Neither cis-resveratrol nor trans-resveratrol inhibited IL-18 production in macro-phages; actually, cis-resveratrol induced IL-18 secre-tion under these conditions (Figure 2b, e). Based onthese results, we focused our attention in this study onthe inhibitory effects of cis-resveratrol on IL-1b secre-tion, as this anti-inflammatory activity may havepotential applications in treating inflammatory dis-eases in humans.
We examined whether the immunomodulatoryeffects of cis-resveratrol on IL-1b and IL-18 productionmay be due to changes in gene expression of these cyto-kines. Quantitative real-time RT-PCR analysis showedthat pre-treatment of macrophages with 100 mM cis-resveratrol significantly reduced IL-1b mRNA expres-sion (Figure 2c); in contrast, cis-resveratrol used at thesame concentration increased IL-18 mRNA expression(Figure 2d). These findings indicate that cis-resveratrolsuppresses IL-1b production by altering the transcrip-tion and translation of this cytokine in activatedmacrophages.
Figure 1. cis-Resveratrol shows no toxicity on human THP-1 macrophages. THP-1 macrophages were treated with various con-
centrations (1, 10 and 100 mM) of either cis-resveratrol or trans-resveratrol for 24 h, and cell viability was measured by using the MTT
assay as described in the ‘Materials and methods’. Data are presented as means� SD of three experiments preformed in duplicate.
Huang et al. 5

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
Figure 2. Effects of cis-resveratrol on the secretion of IL-1b and IL-18 in human THP-1 macrophages. Cells were pre-treated with
various concentrations (1, 10 and 100 mM) of cis-resveratrol or 10 mM of trans-resveratrol for 20 h, followed by treatment with LPS
(0.5 mg/ml) for 3 h and with ATP (5 mM) for 1 h. Secretion of (a) IL-1b and (b) IL-18 in cell culture supernatants was monitored by
ELISA. mRNA expression levels of (c) IL-1b and (d) IL-18 were quantified using real-time RT-PCR. b-Actin gene expression was used
for normalization. The results were expressed as fold change of untreated cells. (e) The presence of IL-1b and IL-18 in cell lysates and
cell culture supernatants were analyzed by Western blot analysis. b-Actin was used as an internal control. (f) Bands from (e) were
analyzed by scanning densitometry and normalized to b-actin. Data were presented as means� SD of three experiments performed in
duplicate. #P< 0.01 versus untreated cells. *P< 0.01 versus resveratrol-untreated (ethanol-treated) control cells. yP< 0.01 versus
cis-resveratrol (100 mM) treated cells.
6 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:54pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
cis-Resveratrol reduces expression and activation ofcaspase-1 and caspase-4 in macrophages
The cytokine IL-1b is generated as a cytosolic precursorthat requires cleavage by the protease caspase-1 inorder to generate the biologically active cytokine.27
To determine whether caspase-1 gene expression andprotein activation may be affected by cis-resveratrol,we pre-incubated THP-1 macrophages with this com-pound for 20 h, prior to LPS and ATP treatmentsas mentioned above. As shown in Figure 3a, cis-resveratrol reduced caspase-1 mRNA expressionunder these conditions. cis-Resveratrol used at100 mM significantly inhibited both caspase-1 secretionin cell culture supernatants and activated caspase-1 (20ku) in whole cell lysates (Figure 3b–d).
Recent studies have shown that pro-IL-1b is also thesubstrate of non-canonical inflammasomes containingcaspase-4, and that this proteolytic enzyme is able togenerate the biologically active form of IL-1b.46 Weexamined the effect of cis-resveratrol on caspase-4gene expression in activated macrophages. As shownin Figure 3a, pre-treatment with cis-resveratrol resultedin a significant decrease of caspase-4 gene expression inmacrophages stimulated with LPS and ATP. cis-Resveratrol-treated cells also showed a significantreduction of active caspase-4 protein in cell lysates(Figure 3c, d; 30 ku). Taken together, these results sug-gest that the reduction in IL-1b secretion induced bycis-resveratrol in macrophages is associated with down-regulation of both caspase-1 and caspase-4 gene expres-sion and protein activation.
Figure 3. cis-Resveratrol inhibits caspase-1 and caspase-4 gene expression and activation in macrophages. Cells were pre-treated
with the indicated concentrations of cis-resveratrol or trans-resveratrol for 20 h, followed by treatment with LPS (0.5 mg/ml) for 3 h
and with ATP (5 mM) for 1 h. (a) mRNA expression levels of caspase-1 and caspase-4 were quantified using real-time RT-PCR. b-Actin
gene expression was used for normalization. The results were expressed as fold change of untreated cells. (b) Secretion of caspase-1
subunit p20 into the supernatants of THP-1 macrophages was assessed by ELISA. (c) Equal amounts of cell lysates were probed to
detect pro-caspase-1 p45 and caspase-1 subunit p20 or pro-caspase-4 p43 and caspase-4 subunit p30. b-Actin was used as an internal
control. (d) Bands from (c) were analyzed by scanning densitometry and normalized to b-actin. Data were presented as means� SD of
three experiments preformed in duplicate. #P< 0.01 versus untreated cells. *P< 0.01 versus resveratrol-untreated control (ethanol)
cells.
Huang et al. 7

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
cis-Resveratrol does not inhibit the expressionof NLRP1, NLRP3, or ASC
Canonical inflammasomes are formed by three compo-nents that include an NLR family protein (e.g. NLRP1or NLRP3), the adaptor protein ASC and pro-caspase-1.48 To determine whether cis-resveratrol affects theNLRP1 and NLRP3 inflammasomes, we measuredthe mRNA and protein expression levels of NLRP1,NLRP3 and ASC using quantitative real-time RT-PCR and Western blot analyses. As shown in Figure4a, c and d, the mRNA and protein levels of ASC,which decreased following treatment with LPS andATP, appeared to slightly increase in cells pre-treatedwith cis-resveratrol at 100 mM compared to 1 and10 mM. Moreover, pre-treatment of macrophages withcis-resveratrol at 100 mM increased the mRNA and pro-tein levels of NLRP1 and NLRP3 compared with cells
treated only with LPS and ATP (Figure 4b, c, d).We conclude that the inhibitory effect of cis-resveratrolon IL-1b secretion is not due to down-regulation ofNLRP1, NLRP3 or ASC expression.
cis-Resveratrol reduces P2X7 receptor expression
Previous studies have shown that activation of the pur-inergic receptor P2X7R by extracellular ATP activatesthe inflammasome, which leads to processing andrelease of both caspase-1 and mature IL-1b into theculture medium of activated macrophages.49,50 Todetermine whether cis-resveratrol may influence expres-sion of P2X receptors, which represent ATP-gatedchannels, we pre-treated THP-1 macrophages withcis-resveratrol, and then with LPS and ATP, prior tomeasuring mRNA and protein expression of P2X7Rand P2X4R (receptor for lower concentrations of
Figure 4. Effects of cis-resveratrol on the expression of ASC, NLRP1 and NLRP3 in activated macrophages. Cells were pre-treated
with the indicated concentrations of cis-resveratrol or trans-resveratrol for 20 h, followed by treatment with LPS (0.5 mg/ml) for 3 h
and with ATP (5 mM) for 1 h. mRNA expression levels of (a) ASC, as well as (b) NLRP1 and NLRP3, were quantified using real-time RT-
PCR. b-Actin gene expression was used for normalization. The results were expressed as fold change of untreated cells. (c) Cell
lysates were analyzed by Western blot analysis using specific anti-ASC, anti-NLRP1 and anti-NLRP3 Abs. (d) Bands from (c) were
analyzed by scanning densitometry and normalized to b-actin. Data were presented as means� SD of three experiments preformed in
duplicate. #P< 0.01 versus untreated cells.
8 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
ATP than P2X7R) using quantitative real-time PCRand Western blot analyses. As shown in Figure 5aand c, the up-regulation of P2X7R expression inducedby ATP (and LPS) was significantly suppressedby100 mM cis-resveratrol. In contrast, the mRNA andprotein levels of P2X4R increased following treatmentwith cis-resveratrol (Figure 5a, b). These results suggestthat the inhibitory effect of cis-resveratrol on IL-1bsecretion may be due, at least in part, to reducedP2X7R expression.
cis-Resveratrol inhibits endoplasmic reticulum stressand ROS production
Previous studies have reported that activation ofP2X7R by ATP increases the generation of ROS thatare associated with activation of caspase-1 and
processing of IL-1b.51,52 We examined whether theROS production induced by ATP was also affected bycis-resveratrol. We used a commercially available detec-tion kit to measure intracellular ROS productionin macrophages pre-treated with cis-resveratrol,prior to LPS priming and ATP activation. ATPtreatment in LPS-primed macrophages induced ahigh amount of ROS compared with untreatedcells (Figure 5d; pyocyanin was used as a positive con-trol for ROS formation). Notably, ATP-inducedROS production was suppressed in a concentration-dependent manner by cis-resveratrol pre-treatment(Figure 5d).
Previous studies have shown that ROS can directlyinduce endoplasmic reticulum (ER) stress.53–55 In add-ition, caspase-4, a member of the caspase-1 subfamily,has been shown to localize to the ER membrane and to
Figure 5. cis-Resveratrol suppresses P2X7R-mediated ROS production in macrophages. Cells were pre-treated with the indicated
concentrations of cis-resveratrol or trans-resveratrol for 20 h, followed by treatment with LPS (0.5 mg/ml) for 3 h and with ATP (5 mM)
for 1 h. (a) mRNA expression levels of P2X4R and P2X7R were quantified using real-time RT-PCR. b-Actin gene expression was used
for normalization. The results were expressed as fold change of untreated cells. (b, c) Cell lysates were analyzed by Western blot
analysis used specific anti-P2X4R and anti-P2X7R Abs. The relative protein levels were quantified by scanning densitometry and
normalized to b-actin. (d) ROS production was measured with the total ROS detection kit. Pyocyanin (200 mM), a ROS inducer, was
used as a positive control for ROS formation. Data were presented as means� SD of three experiments preformed in duplicate.#P< 0.01 versus untreated cells. *P< 0.01 versus resveratrol-untreated control (ethanol) cells.
Huang et al. 9

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
be cleaved and activated in response to ER stress.56
The prototypical ER stress-responsive gene, GRP78,encodes a molecular chaperone and has been used asa ER stress marker.57 To assess whether ATP-inducedROS production is involved in ER stress, we evaluatedthe mRNA and protein levels of GRP78 using quanti-tative real-time RT-PCR and Western blot analyses. Asshown in Figure 6a, GRP78 mRNA expression wasdown-regulated by pre-treatment with cis-resveratrolcompared with cells treated only with LPS and ATP.Further experiments showed that cis-resveratrol pro-duced a similar effect on GRP78 protein levels(Figure 6b). Taken together, these results suggest thatcis-resveratrol suppresses caspase-4 activation in acti-vated macrophages, at least in part, by reducing ROSproduction and GRP78 expression.
cis-Resveratrol inhibits COX-2 protein expressionand PGE2 production
To investigate further the anti-inflammatory propertiesof cis-resveratrol, we examined the protein level ofanother important pro-inflammatory enzyme, COX-2,by Western blot analysis. Treatment of THP-1 macro-phages with LPS and ATP resulted in a pronouncedup-regulation of COX-2 protein level (Figure 6b).Pre-treatment of the cells with cis-resveratrol(100mM) inhibited COX-2 protein level comparedwith cells treated only with LPS and ATP (Figure 6c).
PGE2 represents a major active product of themetabolism of arachidonic acid that is produced bythe COX pathway in macrophages.58 To determinewhether the inhibitory effect of cis-resveratrol on
Figure 6. cis-Resveratrol suppresses GRP78 and COX-2 expression, and PGE2 production in activated macrophages. Cells were pre-
treated with the indicated concentrations of cis-resveratrol or trans-resveratrol for 20 h, followed by treatment with LPS (0.5 mg/ml)
for 3 h and with ATP (5 mM) for 1 h. (a) mRNA expression levels of GRP78 were quantified using real-time RT-PCR. b-Actin gene
expression was used for normalization. The results were expressed as fold change of untreated cells. (b and c) Cell lysates were
analyzed by Western blot analysis used specific anti-GRP78 and anti-COX-2 Abs. The relative protein levels were quantified by
scanning densitometry and normalized to b-actin. (d) The concentrations of PGE2 in the cell culture supernatants of THP-1
macrophages were assessed by ELISA. Data were presented as means� SD of three experiments preformed in duplicate. #P< 0.01
versus untreated cells. *P< 0.01 versus resveratrol-untreated control (ethanol) cells.
10 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
COX-2 protein level affected the production of PGE2,we performed ELISA to detect PGE2 in cell culturesupernatants. As shown in Figure 6d, stimulation ofmacrophages with LPS and ATP resulted in a markedincrease in PGE2 production compared with theuntreated control. However, cis-resveratrol decreasedthe production of PGE2 (Figure 6c; 100 mM). Theseresults suggest that cis-resveratrol may produce anti-inflammatory effects by inhibiting the COX pathway.
cis-Resveratrol suppresses p38 MAPKphosphorylation and c-Jun protein expression
NF-kB is an important transcription factor that regu-lates gene expression of pro-inflammatory cytokinesand mediators in activated macrophages. NF-kBactivation occurs via phosphorylation and subsequentdegradation of IkB-a, which results in translocation ofNF-kB to the nucleus, causing activation of NF-kB-dependent genes.59 Therefore, we determined whethercis-resveratrol inhibits pro-IL-1b and COX-2 expres-sion by blocking NF-kB activation in THP-1 macro-phages. The cytosolic level of IkB-a and nuclear levelof NF-kB p65 subunit were analyzed by Western blotanalysis. The results showed that pre-treatment withcis-resveratrol at 100 mM led to translocation of NF-kB p65 subunit from the cytosol to the nucleus, sug-gesting that the NF-kB pathway may not be involved indown-regulation of pro-IL-1b and COX-2 expressionby cis-resveratrol (Figure 7a, b).
MAPKs represent a group of proline-directed serine/threonine kinases that comprise three major enzymes:p38 MAPK, c-Jun N-terminal kinases (JNK)/stress-activated protein kinases and extracellular signal-regulated kinases 1 and 2 (ERK1/2). MAPKs arethought to be involved in the production of pro-inflammatory cytokines and mediators by activatedmacrophages in response to various stimuli, includingLPS.60,61 For these reasons, we examined the effect ofcis-resveratrol on MAPKs in activated macrophages.Western blot analysis was carried out using Absraised against the phosphorylated and total forms ofp38 MAPK, ERK1/2 and JNK. We observed that cis-resveratrol at 100 mM significantly suppressed the phos-phorylation of p38 MAPK, but did not suppress thephosphorylation of ERK1/2 or JNK (Figure 7c, d).In addition, cis-resveratrol did not significantly affectthe total protein levels of p38 MAPK, ERK1/2 andJNK in activated macrophages (Figure 7c).
Previous work has shown that pro-inflammatorycytokines and mediators are controlled not only byNF-kB but also by activator protein-1 (AP-1).60 AP-1is a dimeric transcription factor composed of membersof the c-Jun and c-Fos proto-oncogene families.62 Asthe activation of AP-1 can be regulated predominatelyby the MAPK signal transduction pathway,63 we fur-ther examined the effect of cis-resveratrol on the
expression of AP-1 component proteins (c-Jun andc-Fos) in activated macrophages. After pre-incubationwith cis-resveratrol (100mM), the induction of c-Jun byLPS and ATP was significantly reduced (Figure 7e);however, the level of c-Fos protein was not suppressedby cis-resveratrol (Figure 7f). Based on these results, itappears that cis-resveratrol may impair pro-IL-1b andCOX-2 expression by inhibiting p38 MAPK phosphor-ylation and c-Jun protein production.
Discussion
The present study demonstrates that cis-resveratrolinhibits IL-1b secretion in activated macrophages bysuppressing various pro-inflammatory signaling mol-ecules and pathways. To our knowledge, our report isthe first to show a difference between cis-resveratroland trans-resveratrol in activated macrophages. Afteridentifying that cis-resveratrol inhibits IL-1b secretion,we focused our attention on the modulatory effects ofthis compound on the inflammasomes and pro-inflammatory mediators.
LPS stimulates the secretion of a large number ofinflammatory molecules by macrophages, includingIL-1b, IL-6, TNF-a and COX-2.60,64 Our study showsthat LPS and ATP significantly induces the expressionand production of pro-inflammatory cytokines (IL-1band IL-18), and enhances expression of COX-2 andproduction of PGE2 in human THP-1 macrophages.By binding to TLR4, LPS activates several intracellularsignaling pathways that include the IkB kinase–NF-kBpathway and MAPK pathways. In turn, these signalingpathways activate a variety of transcription factors thatinclude NF-kB and AP-1.60 Our study shows that LPSand ATP induces NF-kB activation, phosphorylationof p38 MAPK, ERK1/2 and JNK, and AP-1 compo-nent proteins (c-Jun and c-Fos) expression in activatedmacrophages.
In addition to significantly reducing p38 MAPKphosphorylation and c-Jun protein expression, pre-treatment of macrophages with cis-resveratrol alsoinhibited the expression and production of pro-IL-1b,caspase-1, caspase-4, COX-2 and GRP78 followingactivation with LPS and ATP. A previous study hasdemonstrated that human caspase-1 mRNA is inducedthrough p38 MAPK signaling in cultured human kera-tinocytes.65 p38 MAPK has also been shown to up-regulate the expression of the ER chaperone, GRP78,and to activate ER stress.66 Moreover, p38 MAPK is akey regulator of inflammatory cytokines and medi-ators—an observation which makes this protein anattractive drug target for treatment.67 Interestingly,our study shows, for the first time, that cis-resveratrolinhibits the phosphorylation of p38 MAPK. Our find-ings suggest that the inhibitory effects of cis-resveratrolon p38 MAPK phosphorylation and c-Jun protein
Huang et al. 11

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
Figure 7. cis-Resveratrol inhibits p38 MAPK phosphorylation and c-Jun expression in macrophages. Cells were pre-treated with the
indicated concentrations of cis-resveratrol or trans-resveratrol for 20 h, followed by treatment with LPS (0.5 mg/ml) for 3 h and with
ATP (5 mM) for 1 h. (a, b) After the isolation of cytosolic and nuclear fractions, cytosolic IkB-a and nuclear NF-kB p65 were analyzed
by Western blot analysis. b-Actin and lamin B1 were used as internal controls. The relative protein levels were quantified by scanning
densitometry. (c) Whole cell lysates were analyzed by Western blotting using various Abs against p38 MAPK, ERK1/2, JNK, phospho-
p38 MAPK, phospho-ERK1/2 and phospho-JNK. (d) Bands from (c) were analyzed by scanning densitometry and normalized to total
p38 MAPK, ERK1/2 and JNK. (E and F) AP-1 component proteins (c-Jun and c-Fos) were analyzed by Western blot analysis. The
relative protein levels were quantified by scanning densitometry and normalized to b-actin. Data were presented as means� SD of
three experiments preformed in duplicate. #P< 0.01 versus untreated cells. *P< 0.01 versus resveratrol-untreated control (ethanol)
cells.
12 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
expression may involve the reduction of pro-IL-1b,caspase-1, caspase-4, COX-2 and GRP78 expression.
Macrophages were first shown to secrete IL-18 fol-lowing activation with stimuli such as LPS.68 The NF-kB pathway has been shown to be involved in theproduction of IL-18 induced by LPS in THP-1 cells.69
Cellular signaling leading to IL-18 production alsoinvolves MyD88- and IL-1 receptor associated kinaseTRAF6-dependent pathways, which, in turn, induceNF-kB.22 In addition, LPS-induced NLRP3 mRNAexpression is reduced by NF-kB inhibitors, indicatingthat NF-kB may play an important role in regulatingNLRP3 gene expression.70 In the present study,pre-treatment with cis-resveratrol increased NF-kBactivation and JNK phosphorylation, which may beresponsible for the increased expression and productionof IL-18 in activated macrophages. A recent study hasshown that blockade of either ERK1/2 or JNK, but notp38 MAPK, inhibits LPS-induced expression ofNLRP3 protein in murine macrophages.71 Consistentwith this finding, we found that inhibition of p38MAPK phosphorylation did not affect the expressionof NLRP3, but significantly inhibited IL-1b mRNAand pro-IL-1b protein expression in cis-resveratrolpre-treated macrophages.
It is generally recognized that COX-2 is barelydetectable under normal physiological conditions, butis induced mainly at sites of inflammation in responseto inflammatory stimuli, including pro-inflammatorymediators and mitogenic stimuli, such as cytokines(IL-1 cytokine family), endotoxins (LPS), growth fac-tors, oncogenes and phorbol esters.72 Moreover, COX-2 and its products, particularly PGE2, are known to beassociated with various tumors and chronic inflamma-tory diseases.24,25,73–75 Therefore, COX-2 may rep-resent a promising molecular target for thechemoprevention of cancer and inflammatory diseases.Our results demonstrate that cis-resveratrol at 100 mMmarkedly reduces PGE2 production, as well as COX-2protein synthesis in activated macrophages, allowing usto conclude that cis-resveratrol has significant modula-tory effects on the production of PGE2 by macro-phages. These findings also suggest that reducedPGE2 levels may be due to a reduction of COX-2 bycis-resveratrol pre-treatment. Our results confirm thework of Leiro et al. who also demonstrated that cis-resveratrol blocks ROS production by inflammatorymurine macrophages and inhibits PGE2 productionand COX-2 expression.76 In addition, our findings arein agreement with previous results showing that canon-ical inflammasome activation modulates the productionof prostaglandins in macrophages.30
ATP-induced P2X7R activation promotes the rapidproduction of large amounts of ROS, which, in turn,stimulates activation of the NLRP3 inflammasome.51
P2X7R stimulation in primary human and murinemacrophages is also required for the release of PGE2,
and PGE2 release is independent of inflammasomeactivation.77 In this study, we demonstrate that pre-treatment of LPS-primed macrophages with cis-resveratrol significantly inhibits ATP-induced P2X7Rexpression. Moreover, our results show that ATP-induced ROS production is suppressed by cis-resveratrol. In agreement with these findings, previousstudies have shown that cis-resveratrol displays typicalantioxidant activity, and that this effect may be due toinhibition of NADH/NADPH oxidase enzyme activ-ity.76,78 Our data, which were obtained in humanmacrophages in vitro, also support previous observa-tions suggesting that a decline in PGE2 productionmay be due, at least in part, to inhibitory effects onP2X7R expression.
Caspase-4 expression was shown to be required forcaspase-1 activation and maturation of pro-IL-1b inkeratinocytes and activated THP-1 macrophages, sug-gesting that caspase-4 may act upstream of caspase-1.46
ROS accumulation is known to induce ER stress, whichfurther leads to proteolytic cleavage of caspase-4.56,79
Previous studies have also reported that oxidative chal-lenge can lead to ER stress with up-regulation of theprotective chaperone protein GRP78, which has beenproposed to play a crucial role in preventing oxidativedamage to the ER and promoting cell survival.57,80 Pre-treatment of the cells with cis-resveratrol resulted in asignificant reduction of caspase-4 expression and activa-tion compared with resveratrol-untreated control cells.We also demonstrated that cis-resveratrol inhibits theexpression of the ER stress marker GRP78 and cas-pase-1 activation, indicating that cis-resveratrol mayact upstream of the non-canonical inflammasome andresult in down-regulation of GRP78 expression and cas-pase-4 and caspase-1 activation, thereby leading toreduced IL-1b secretion. However, cis-resveratrol pre-treatment did not impair IL-18 secretion in activatedmacrophages. In this respect, a recent study has shownthat processing and secretion of IL-18 does not requireROS production.81 Based on these observations, wespeculate that the inhibitory effect of cis-resveratrol onROS production may down-regulate caspase-4 and cas-pase-1 activation and lead to reduced IL-1b secretion.
Previous studies have reported that trans-resveratrolis quickly absorbed and rapidly metabolized in vivo.82
Preliminary studies in humans have shown that whiletrans-resveratrol is quickly absorbed following oralconsumption, its bioavailability in the human body isrelatively low.83 Additional studies are needed to exam-ine the absorption, bioavailability and potential bene-ficial effects of cis-resveratrol in vivo.
In conclusion, this study reveals that cis-resveratrolinhibits IL-1b secretion in activated THP-1 macro-phages. The anti-inflammatory effect of cis-resveratrolis associated with reduced P2X7R and GRP78 expres-sion, which leads to suppression of ROS productionand reduced caspase-1 and caspase-4 activation
Huang et al. 13
XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
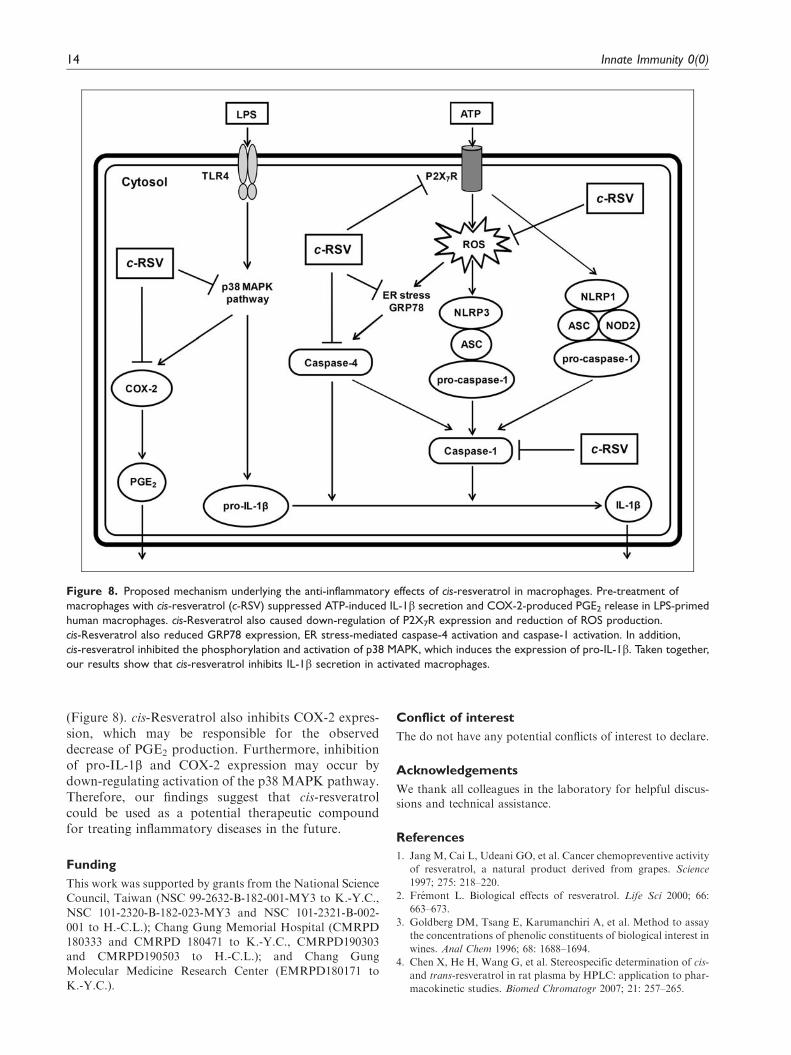
(Figure 8). cis-Resveratrol also inhibits COX-2 expres-sion, which may be responsible for the observeddecrease of PGE2 production. Furthermore, inhibitionof pro-IL-1b and COX-2 expression may occur bydown-regulating activation of the p38 MAPK pathway.Therefore, our findings suggest that cis-resveratrolcould be used as a potential therapeutic compoundfor treating inflammatory diseases in the future.
Funding
This work was supported by grants from the National ScienceCouncil, Taiwan (NSC 99-2632-B-182-001-MY3 to K.-Y.C.,
NSC 101-2320-B-182-023-MY3 and NSC 101-2321-B-002-001 to H.-C.L.); Chang Gung Memorial Hospital (CMRPD180333 and CMRPD 180471 to K.-Y.C., CMRPD190303and CMRPD190503 to H.-C.L.); and Chang Gung
Molecular Medicine Research Center (EMRPD180171 toK.-Y.C.).
Conflict of interest
The do not have any potential conflicts of interest to declare.
Acknowledgements
We thank all colleagues in the laboratory for helpful discus-sions and technical assistance.
References
1. Jang M, Cai L, Udeani GO, et al. Cancer chemopreventive activity
of resveratrol, a natural product derived from grapes. Science
1997; 275: 218–220.
2. Fremont L. Biological effects of resveratrol. Life Sci 2000; 66:
663–673.
3. Goldberg DM, Tsang E, Karumanchiri A, et al. Method to assay
the concentrations of phenolic constituents of biological interest in
wines. Anal Chem 1996; 68: 1688–1694.
4. Chen X, He H, Wang G, et al. Stereospecific determination of cis-
and trans-resveratrol in rat plasma by HPLC: application to phar-
macokinetic studies. Biomed Chromatogr 2007; 21: 257–265.
Figure 8. Proposed mechanism underlying the anti-inflammatory effects of cis-resveratrol in macrophages. Pre-treatment of
macrophages with cis-resveratrol (c-RSV) suppressed ATP-induced IL-1b secretion and COX-2-produced PGE2 release in LPS-primed
human macrophages. cis-Resveratrol also caused down-regulation of P2X7R expression and reduction of ROS production.
cis-Resveratrol also reduced GRP78 expression, ER stress-mediated caspase-4 activation and caspase-1 activation. In addition,
cis-resveratrol inhibited the phosphorylation and activation of p38 MAPK, which induces the expression of pro-IL-1b. Taken together,
our results show that cis-resveratrol inhibits IL-1b secretion in activated macrophages.
14 Innate Immunity 0(0)

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
5. de la Lastra CA and Villegas I. Resveratrol as an anti-
inflammatory and anti-aging agent: mechanisms and clinical
implications. Mol Nutr Food Res 2005; 49: 405–430.
6. de la Lastra CA and Villegas I. Resveratrol as an antioxidant and
pro-oxidant agent: mechanisms and clinical implications.
Biochem Soc Trans 2007; 35: 1156–1160.
7. Shakibaei M, Harikumar KB and Aggarwal BB. Resveratrol
addiction: to die or not to die. Mol Nutr Food Res 2009; 53:
115–128.
8. Shen MY, Hsiao G, Liu CL, et al. Inhibitory mechanisms of
resveratrol in platelet activation: pivotal roles of p38 MAPK
and NO/cyclic GMP. Br J Haematol 2007; 139: 475–485.
9. Baur JA and Sinclair DA. Therapeutic potential of resveratrol:
the in vivo evidence. Nat Rev Drug Discov 2006; 5: 493–506.
10. Bertelli AA, Giovannini L, Bernini W, et al. Antiplatelet activity
of cis-resveratrol. Drugs Exp Clin Res 1996; 22: 61–63.
11. Pettit GR, Grealish MP, Jung MK, et al. Antineoplastic agents.
465. Structural modification of resveratrol: sodium resverastatin
phosphate. J Med Chem 2002; 45: 2534–2542.
12. Kukric ZZ and Topalic-Trivunovic LN. Antibacterial activity of
cis- and trans-resveratrol isolated from Polygonum cuspidatum
rhizome. Acta Period Technol 2006; 37: 131–136.
13. Dinarello CA. Interleukin-1beta, interleukin-18, and the interleu-
kin-1 beta converting enzyme. Ann N Y Acad Sci 1998; 856: 1–11.
14. Gregory JL, Morand EF, McKeown SJ, et al. Macrophage
migration inhibitory factor induces macrophage recruitment via
CC chemokine ligand 2. J Immunol 2006; 177: 8072–8079.
15. Sims JE and Smith DE. The IL-1 family: regulators of immunity.
Nat Rev Immunol 2010; 10: 89–102.
16. Dinarello CA. Immunological and inflammatory functions of the
interleukin-1 family. Annu Rev Immunol 2009; 27: 519–550.
17. Davis BK, Wen H and Ting JP. The inflammasome NLRs in
immunity, inflammation, and associated diseases. Annu Rev
Immunol 2011; 29: 707–735.
18. Dinarello CA and Wolff SM. The role of interleukin-1 in disease.
N Engl J Med 1993; 328: 106–113.
19. Dinarello CA. Biologic basis of interleukin-1 in disease. Blood
1996; 87: 2095–2147.
20. Church LD, Cook GP and McDermott M. Primer: inflamma-
somes and interleukin 1 beta in inflammatory disorders. Nat Clin
Pract Rheumatol 2008; 4: 34–42.
21. Nakanishi K, Yoshimoto T, Tsutsui H, et al. Interleukin-18 regu-
lates both Th1 and Th2 responses. Annu Rev Immunol 2001; 19:
423–474.
22. Arend WP, Palmer G and Gabay C. IL-1, IL-18, and IL-33
families of cytokines. Immunol Rev 2008; 223: 20–38.
23. Lee SH, Soyoola E, Chanmugam P, et al. Selective expression of
mitogen-inducible cyclooxygenase in macrophages stimulated
with lipopolysaccharide. J Biol Chem 1992; 267: 25934–25938.
24. Rocca B and FitzGerald GA. Cyclooxygenases and prostaglan-
dins: shaping up the immune response. Int Immunopharmacol
2002; 2: 603–630.
25. Turini ME and DuBois RN. Cyclooxygenase-2: a therapeutic
target. Annu Rev Med 2002; 53: 35–57.
26. Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know
about danger. J Leukoc Biol 2007; 81: 1–5.
27. Martinon F, Burns K and Tschopp J. The inflammasome:
a molecular platform triggering activation of inflammatory
caspases and processing of proIL-beta. Mol Cell 2002; 10:
417–426.
28. Petrilli V, Dostert C, Muruve DA, et al. The inflammasome: a
danger sensing complex triggering innate immunity. Curr Opin
Immunol 2007; 19: 615–622.
29. Franchi L, Eigenbrod T, Munoz-Planillo R, et al. The inflamma-
some: a caspase-1-activation platform that regulates immune
responses and disease pathogenesis. Nat Immunol 2009; 10:
241–247.
30. von Moltke J, Trinidad NJ, Moayeri M, et al. Rapid induction of
inflammatory lipid mediators by the inflammasome in vivo.
Nature 2012; 490: 107–111.
31. Bours MJ, Swennen EL, Di Virgilio F, et al. Adenosine 50-
triphosphate and adenosine as endogenous signaling molecules
in immunity and inflammation. Pharmacol Ther 2006; 112:
358–404.
32. Di Virgilio F. Liaisons dangereuses: P2X7 and the inflamma-
some. Trends Pharmacol Sci 2007; 28: 465–472.
33. Yu HB and Finlay BB. The caspase-1 inflammasome: a pilot of
innate immune response. Cell Host Microbe 2008; 4: 198–208.
34. Dostert C, Petrilli V, Van Bruggen R, et al. Innate immune acti-
vation through Nalp3 inflammasome sensing of asbestos and
silica. Science 2008; 320: 674–677.
35. Duewell P, Kono H, Rayner KJ, et al. NLRP3 inflammasomes
are required for atherogenesis and activated by cholesterol crys-
tals. Nature 2010; 464: 1357–1361.
36. Feldmeyer L, Keller M, Niklaus G, et al. The inflammasome
mediates UVB-induced activation and secretion of interleukin-
1bata by keratinocytes. Curr Biol 2007; 17: 1140–1145.
37. Hise AG, Tomalka J, Ganesan S, et al. An essential role for the
NLRP3 inflammasome in host defense against the human fun-
gal pathogen Candida albicans. Cell Host Microbe 2009; 5:
487–497.
38. Hornung V, Bauernfeind F, Halle A, et al. Silica crystals and
aluminum salts activate the NLRP3 inflammasome through
phagosomal destabilization. Nat Immunol 2008; 9: 847–856.
39. Mariathasan S, Weiss DS, Newton K, et al. Cryopyrin activates
the inflammasome in response to toxins and ATP. Nature 2006;
440: 228–232.
40. Martinon F, Petrilli V, Mayor A, et al. Gout-associated uric acid
crystals activate the NLRP3 inflammasome. Nature 2006; 440:
237–241.
41. Willingham SB, Bergstralh DT, O’Connor W, et al. Microbial
pathogen-induced necrotic cell death mediated by the inflamma-
some component CIAS1/cryopyrin/NLRP3 and ASC. Cell Host
Microbe 2007; 2: 147–159.
42. Bruey JM, Bruey-Sedano N, Luciano F, et al. Bcl-2 and Bcl-XL
regulate proinflammatory caspase-1 activation with NALP1. Cell
2007; 129: 45–56.
43. Faustin B, Lartigue L, Bruey JM, et al. Reconstituted NALP1
inflammasome reveals two-step mechanism of caspase-1 activa-
tion. Mol Cell 2007; 25: 713–724.
44. Broz P, Ruby T, Belhocine K, et al. Caspase-11 increases suscep-
tibility to Salmonella infection in the absence of caspase-1.
Nature 2012; 490: 288–291.
45. Kayagaki N, Warming S, Lamkanfi M, et al. Non-canonical
inflammasome activation targets caspase-11. Nature 2011; 479:
117–121.
46. Sollberger G, Strittmatter GE, Kistowska M, et al. Caspase-4 is
required for activation of inflammasomes. J Immunol 2012; 188:
1992–2000.
47. Akhter A, Caution K, Abu Khweek A, et al. Caspase-11 pro-
motes the fusion of phagosomes harboring pathogenic bacteria
with lysosomes by modulating actin polymerization. Immunity
2012; 37: 35–47.
48. Schroder K and Tschopp J. The inflammasomes. Cell 2010; 140:
821–832.
49. Mehta VB, Hart J and Wewers MD. ATP stimulated release of
IL-1b and IL-18 required priming by LPS and is independent of
caspase-1 cleavage. J Biol Chem 2001; 276: 3820–3826.
50. Ferrari D, Pizzirani C, Adinolfi E, et al. The P2X7 receptor: a
key player in IL-1 processing and release. J Immunol 2006; 176:
3877–3883.
51. Cruz CM, Rinna A, Forman HJ, et al. ATP activates a reactive
oxygen species-dependent oxidative stress responses and secre-
tion of proinflammatory cytokines in macrophages. J Biol
Chem 2007; 282: 2871–2879.
Huang et al. 15

XML Template (2013) [21.10.2013–2:55pm] [1–16]//blrnas3/cenpro/ApplicationFiles/Journals/SAGE/3B2/INIJ/Vol00000/130129/APPFile/SG-INIJ130129.3d (INI) [PREPRINTER stage]
52. Coutinho-Silva R, Correa G, Sater AA, et al. The P2X7 receptor
and intracellular pathogens: a continuing struggle. Purinergic
Signal 2009; 5: 197–204.
53. Liu H, Bowes RC 3rd, van de Water B, et al. Endoplasmic reticu-
lum chaperones GRP78 and calreticulin prevent oxidative stress,
Ca2+disturbances, and cell death in renal epithelial cells. J Biol
Chem 1997; 272: 21751–21759.
54. Hayashi T, Saito A, Okuno S, et al. Damage to the endoplasmic
reticulum and activation of apoptotic machinery by oxidative
stress in ischemic neurons. J Cereb Blood Flow Metab 2005; 25:
41–53.
55. He S, Yaung J, Kim YH, et al. Endoplasmic reticulum stress
induced by oxidative stress in retinal pigment epithelial cells.
Graefes Arch Clin Exp Ophthalmol 2008; 246: 677–683.
56. Hitomi J, Katayama T, Eguchi Y, et al. Involvement of caspase-4
in endoplasmic reticulum stress-induced apoptosis and Ab-induced cell death. J Cell Biol 2004; 165: 347–356.
57. Yoshida H, Haze K, Yanagi H, et al. Identification of the
cis-acting endoplasmic reticulum stress response element respon-
sible for transcriptional induction of mammalian Glc-regulated
proteins. Involvement of basic leucine zipper transcription fac-
tors. J Biol Chem 1998; 273: 33741–33749.
58. Diez E, Balsinde J, Aracil M, et al. Ethanol induces release of
arachidonic acid but not synthesis of eicosanoids in mouse peri-
toneal macrophages. Biochim Biophys Acta 1987; 921: 82–89.
59. Stancovski I and Baltimore D. NF-kappaB activation: the I
kappaB kinase revealed? Cell 1997; 91: 299–302.
60. Guha M and Mackman N. LPS induction of gene expression in
human monocytes. Cell Signal 2001; 13: 85–94.
61. Rao KM. MAP kinase activation in macrophages. J Leukoc Biol
2001; 69: 3–10.
62. Angel P and Karin M. The role of Jun, Fos and the AP-1 com-
plex in cell-proliferation and transformation. Biochim Biophys
Acta 1991; 1072: 129–157.
63. Whitmarsh AJ and Davis RJ. Transcription factor AP-1 regula-
tion by mitogen-activated protein kinase signal transduction
pathways. J Mol Med 1996; 74: 589–607.
64. Medvedev AE, Kopydlowski KM and Vogel SN. Inhibition of
lipopolysaccharide-induced signal transduction in endotoxin-tol-
erized mouse macrophages: dysregulation of cytokine, chemo-
kine, and toll-like receptor 2 and 4 gene expression. J Immunol
2000; 164: 5564–5574.
65. Johansen C, Moeller K, Kragballe K, et al. The activity of cas-
pase-1 is increased in lesional psoriatic epidermis. J Invest
Dermatol 2007; 127: 2857–2864.
66. Ranganathan AC, Zhang L, Adam AP, et al. functional coupling
of p38-induced up-regulation of Bip and activation of RNA-
dependent protein kinase-like endoplasmic reticulum kinase to
drug resistance of dormant carcinoma cells. Cancer Res 2006;
66: 1702–1711.
67. Schindler JF, Monahan JB and Smith WG. p38 pathway kinases
as anti-inflammatory drug targets. J Dent Res 2007; 86: 800–811.
68. Ushio S, Namba M, Okura T, et al. Cloning of the cDNA for
human IFN-gamma-inducing factor, expression in Escherichia
coli, and studies on the biologic activities of the protein.
J Immunol 1996; 156: 4274–4279.
69. Cheng J, Imanishi H, Morisaki H, et al. Recombinant HBsAg
inhibits LPS-induced COX-2 expression and IL-18 production by
interfering with the NFkappaB pathway in a human monocytic
cell line, THP-1. J Hepatol 2005; 43: 465–471.
70. Bauernfeind F, Horvath G, Stutz A, et al. Cutting edge: NF-
kappaB activating pattern recognition and cytokine receptors
license NLRP3 inflammasome activation by regulating NLRP3
expression. J Immunol 2009; 187: 613–617.
71. Liao PC, Chao LK, Chou JC, et al. Lipopolysaccharide/
adenosine triphosphate-mediated signal transduction in the regu-
lation of NLRP3 protein expression and caspase-1-mediated
interleukin-1b secretion. Inflamm Res 2013; 62: 89–96.
72. Zhang R and Li S. COX-2 as a novel target of CRF family pep-
tides’ participating in inflammation. Biochem Biophys Res
Commun 2009; 382: 483–485.
73. Park JY, Pillinger MH and Abramson SB. Prostaglandin E2 syn-
thesis and secretion: the role of PGE2 synthases. Clin Immunol
2006; 119: 229–240.
74. Greenhough A, Smartt HJ, Moore AE, et al. The COX-2/PGE2
pathway: key roles in the hallmarks of cancer and adaptation to
the tumour microenvironment. Carcinogenesis 2009; 30: 377–386.
75. Huang RY and Chen GG. Cigarette smoking, cyclooxygenase-2
pathway and cancer. Biochim Biophys Acta 2011; 1815: 158–169.
76. Leiro J, Alvarez E, Arranz JA, et al. Effects of cis-resveratrol on
inflammatory murine macrophages: antioxidant activity and
down-regulation of inflammatory genes. J Leukoc Biol 2004;
75: 1156–1165.
77. Barbera-Cremades M, Baroja-Mazo A, Gomez AI, et al. P2X7
receptor-stimulation causes fever via PGE2 and IL-1b release.
FASEB J 2012; 26: 2951–2962.
78. Orallo F. Comparative studies of the antioxidant effects of cis-
and trans-resveratrol. Curr Med Chem 2006; 13: 87–98.
79. Yokouchi M, Hiramatsu N, Hayakawa K, et al. Involvement of
selective reactive oxygen species upstream of proapoptotic
branches of unfolded protein response. J Biol Chem 2008; 283:
4252–4260.
80. Kuznetsov G, Bush KT, Zhang PL, et al. Perturbations in mat-
uration of secretory proteins and their association with endoplas-
mic reticulum chaperones in a cell culture model for epithelial
ischemia. Proc Natl Acad Sci U S A 1996; 93: 8584–8589.
81. Schmidt RL and Lenz LL. Distinct licensing of IL-18 and IL-1bsecretion in response to NLRP3 inflammasome activation. PLoS
One 2012; 7: e45186.
82. Wenzel E and Somoza V. Metabolism and bioavailability of
trans-resveratrol. Mol Nutr Food Res 2005; 49: 472–481.
83. Walle T, Hsieh F, DeLegge MH, et al. High absorption but very
low bioavailability of oral resveratrol in humans. Drug Metab
Dispos 2004; 32: 1377–1382.
16 Innate Immunity 0(0)
Copyright © 2022 FDOKUMEN




















