
Chitinase genes expressed by infective larvae of the filarial nematodes, Acanthocheilonema viteae...
13
Molecular and Biochemical Parasitology 75 (1996) 207-219 MOLECULAR i!iKHEmcA PARAsIToLoGy Chitinase genes expressed by infective larvae of the filarial ne- matodes, Acanthocheilonema viteae and Onchocerca volvulus 1 Yang Wu”, Ralph Adamb, Steven A. Williams”, Albert E. Bianco”.* “Liverpool School of Tropical Medicine, Liverpool L3 5QA, UK bDepartmenr of Zoology, University of Hohenheim, Stuttgart, German) ‘Department of Biological Sciences, Smith College, Northampton, MA. USA Received 13 May 1995; revised 23 October 1995; accepted 2 November 1995 Abstract Stage-specific products of 220 and 75 kDa were identified by metabolic labelling of infective larvae of the filarial nematode Acanthocheilonema viteae in ticks. Synthesis was temperature sensitive, occuring at 27°C but not at 37°C. These products were secreted 3-6 days after leaving the vector during post-infective development, but subsequent expression was not detected. The smaller protein with a pl of 6.2, was purified by two-dimensional electrophoresis and the N-terminal amino acid sequence was derived. This provisionally identified the protein as a chitinase, which was confirmed biochemically by glycol-chitin substrate gel electrophoresis. The polymerase chain reaction was used to amplify a product from a cDNA library of A. viteae infective larvae. The nucleotide sequence codes for a putative signal peptide of 20 amino acids and a mature protein of 504 residues (M, 56 kDa), exhibiting 69% identity (81% similarity allowing for conservative substitutions) with the MFI chitinase described from microfilariae of Brugiu malayi [21]. N-linked glycosylation may account for some, or all, of the discrepancy in M, between the predicted polypeptide and the native parasite product (75 kDa). Primers based on the A. uiteae sequence were used to amplify a related sequence from a cDNA library of Onchocerca volvulus infective larvae. The 0. volvulus cDNA codes for a 20-amino acid signal peptide followed by 477 residues with an IV, of 54 kDa, and shares 67% identity with the A. viteae chitinase (80% similarity allowing for conservative substitutions) and 69% identity with the B. rnalqi MFl molecule. It is proposed that chitinases expressed by infective stages of these filarial nematodes may play a role in ecdysis during post-infective development. Keywords: Acanthocheilonema viteae; Onchocerca volvulus; Infective larvae; Secretion: Chitinase; Gene sequence Abbreviations: mf, microfilariae; L3, third-stage larva; L4, fourth stage larva; PCR, polymerase chain reaction * Corresponding author. Tel.: + 44 151 7089393: Fax: + 44 151 7091079; E-mail: [email protected]. 1. Introduction ’ Note: Nucleotide sequence data reported in this paper have been submitted to the GenBankTM data base with the accession numbers L42010 (A. uiteae) and L42021 (0. v011u1us1. Filarial nematodes are highly specialized and successful parasites of arthropods and vertebrates. In spite of differences amongst species in their 0166-6851/96/$15.00 C 1996 Elsevier Science B.V. All rights reserved SSDI 0166-6851(95)02529-4
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Chitinase genes expressed by infective larvae of the filarial nematodes, Acanthocheilonema viteae...

Molecular and Biochemical Parasitology 75 (1996) 207-219
MOLECULAR
i!iKHEmcAL PARAsIToLoGy
Chitinase genes expressed by infective larvae of the filarial ne- matodes, Acanthocheilonema viteae and Onchocerca volvulus 1
Yang Wu”, Ralph Adamb, Steven A. Williams”, Albert E. Bianco”.*
“Liverpool School of Tropical Medicine, Liverpool L3 5QA, UK bDepartmenr of Zoology, University of Hohenheim, Stuttgart, German)
‘Department of Biological Sciences, Smith College, Northampton, MA. USA
Received 13 May 1995; revised 23 October 1995; accepted 2 November 1995
Abstract
Stage-specific products of 220 and 75 kDa were identified by metabolic labelling of infective larvae of the filarial nematode Acanthocheilonema viteae in ticks. Synthesis was temperature sensitive, occuring at 27°C but not at 37°C. These products were secreted 3-6 days after leaving the vector during post-infective development, but subsequent expression was not detected. The smaller protein with a pl of 6.2, was purified by two-dimensional electrophoresis and the N-terminal amino acid sequence was derived. This provisionally identified the protein as a chitinase, which was confirmed biochemically by glycol-chitin substrate gel electrophoresis. The polymerase chain reaction was used to amplify a product from a cDNA library of A. viteae infective larvae. The nucleotide sequence codes for a putative signal peptide of 20 amino acids and a mature protein of 504 residues (M, 56 kDa), exhibiting 69% identity (81% similarity allowing for conservative substitutions) with the MFI chitinase described from microfilariae of Brugiu malayi [21]. N-linked glycosylation may account for some, or all, of the discrepancy in M, between the predicted polypeptide and the native parasite product (75 kDa). Primers based on the A. uiteae sequence were used to amplify a related sequence from a cDNA library of Onchocerca volvulus infective larvae. The 0. volvulus cDNA codes for a 20-amino acid signal peptide followed by 477 residues with an IV, of 54 kDa, and shares 67% identity with the A. viteae chitinase (80% similarity allowing for conservative substitutions) and 69% identity with the B. rnalqi MFl molecule. It is proposed that chitinases expressed by infective stages of these filarial nematodes may play a role in ecdysis during post-infective development.
Keywords: Acanthocheilonema viteae; Onchocerca volvulus; Infective larvae; Secretion: Chitinase; Gene sequence
Abbreviations: mf, microfilariae; L3, third-stage larva; L4, fourth stage larva; PCR, polymerase chain reaction
* Corresponding author. Tel.: + 44 151 7089393: Fax: + 44 151 7091079; E-mail: [email protected].
1. Introduction
’ Note: Nucleotide sequence data reported in this paper
have been submitted to the GenBankTM data base with the accession numbers L42010 (A. uiteae) and L42021 (0.
v011u1us 1.
Filarial nematodes are highly specialized and
successful parasites of arthropods and vertebrates.
In spite of differences amongst species in their
0166-6851/96/$15.00 C 1996 Elsevier Science B.V. All rights reserved
SSDI 0166-6851(95)02529-4

208 Y. Wu et al. J Molecular and Biochemical Parasitology 75 (1996) 207-219
host specificities and tissue tropisms, all members of the Filarioidea share the common requirement to undergo host-host transmission as infective, third-stage (L3) larvae. It can be expected that the transfer between host environments has exerted powerful adaptive selection on these organisms at this point in their development. Indeed, the differ- entiation of L3 larvae is tightly regulated by ex- trinsic factors, presumably of host origin. since intermoult growth is interrupted in the infective stages awaiting egress from the vectors.
Patterns of gene expression in L3 larvae may therefore shed light on important questions con- cerning the acquisition of infectivity by pre-infec- tive stages. However, difficulties in the laboratory maintenance of many filarial species, coupled with the inherent complexity of these organisms, have limited the rate of progress in our understanding of their developmental biology. Recently, there have been a number of advances in the methods available to investigate stage-specific protein syn- thesis and transcription in the larval stages of filariae. These studies have begun to reveal the dynamic, and sometimes subtle, nature of the changes that occur in gene expression during early development [I -61.
Previously, we have described a method for studying the kinetics of protein synthesis in devel- oping larvae of Onchocerca by micro-injecting radio-labelled amino acids into parasitized blackflies. This is necessary for the investigation of biosynthesis in vector-stage parasites, because methods for the in vitro cultivation of microfilar- iae through to L3 larvae are not available. How- ever, infective larvae of Onchocerca and the rodent filaria Acanthocheilonema viteae may be grown to the fourth-stage in culture [7,8], enabling post-infective stages to be metabolically labelled in vitro. Larvae labelled in the vectors and trans- ferred to isotope-free medium have also been used to study events triggered by parasite transmission.
Here we describe the use of these metabolic labelling techniques to identify a 75-kDa protein from infective larvae of A. viteae. Expression is temperature sensitive and stage-specific, ceasing in post-infective larvae that secrete the protein be- fore the third moult. N-terminal amino acid se- quencing of this product revealed it to be derived
from a chitinase gene. We report the cloning and sequencing of this larval chitinase, together with a related cDNA from infective stages of the human parasite, Onchocerca volvulus.
2. Material and methods
2.1. Parasites and vectors
Acanthoeheilonema (formerly Dipetalonema) viteae was maintained in Mongolian jirds (Meri- ones unguiculatus) and the argasid tick, Or- nithodorus tartakovskyi, as described [9]. Ticks were held at 2628°C and 80% relative humidity: at this temperature, infective larvae developed from microfilariae in approximately one month. L3 larvae were obtained by dissection in RPM1 1640 medium containing antibiotics (200 IU peni- cillin, 100 pg streptomycin ml - ‘) and serum (10% newborn calf serum). Jirds received subcutaneous inoculations of up to 100 L3 larvae and developed patent infections in approximately 2 months. De- veloping larvae and adults were recovered at ne- cropsy from subcutaneous tissues at various intervals [lo]. Microfilariae were obtained from blood drawn by cardiac puncture under terminal anaesthesia. Parasites were isolated from the blood by nuclepore filtration, washed in RPM1 medium and concentrated by centrifugation (600 g x 15 min) and resuspension.
2.2. In vitro culture of infective larvae
Development of A. viteae larvae from ticks to post-infective L3 stages was induced by in vitro cultivation using the cell-free system of Franke and Weinstein [7]. Infected ticks were surface-ster- ilized in 1% benzalkonium chloride (Calbiochem) for 60 s, followed by three washes in phosphate- buffered saline with antibiotics (1000 IU peni- cillin, 1000 pg streptomycin ml-‘). Larvae were recovered by dissection in IMDM/NCTC medium containing serum and antibiotics (40% Iscove’s Modified Dulbecco Medium (Gibco), 40% NCTC 135 medium (Gibco), 20% heat-inactivated foetal calf serum, 100 IU penicillin, 100 pg streptomycin ml - I). After washing by sedimentation and resus-

Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219 209
pension in fresh medium, larvae were dispensed into 24-well Linbro tissue culture plates and incu- bated in an atmosphere of 5% CO2 in air. Cul- tures were maintained at either 37°C or 27°C.
2.3. Metabolic label&g procedures
A. uiteae larvae were metabolically labelled within the tissues of ticks by the micro-injection of [“Slmethionine, using modifications of the pro- cedure of Bianco et al. [I]. Only ticks that had been infected more than 28 days previously and already contained infective larvae were used in these experiments, Inoculations of 5-10 PCi [“?S]methionine 0800 Ci mM ~ ‘, Amersham) in less than 1 ~11 of diluent (20 mM potassium ac- etate, 0.1% (v/v) 2-mercaptoethanol) were admin- istered, using a finely extruded capillary tube coupled to a perforated bulb (Drummond micro- cap assemblies). Ticks were restrained with for- ceps and inoculated through the lateral margin, under the low power of a dissecting microscope. Larvae were recovered by dissection 3 days later and washed by sedimentation and resuspension in three changes of medium.
Labelled L3 larvae from the vector were placed in culture for various periods in the absence of radioisotope at temperatures that do (37°C) or do not (27°C) stimulate post-infective development. Supernatant medium was obtained from serum- free parasite cultures held at 37°C and established at 1000 parasites ml ’ . This was checked visually for the absence of larvae and centrifuged at 26 000 g for 20 min to remove any particulate material. Proteins were precipitated with 0.1% (w/v) sodium deoxycholate and 2.5 volumes of acetone at - 80°C over 16 h. To examine protein synthesis in post-infective stages, unlabelled L3 larvae were recovered from ticks, or later developmental forms from jirds (see above). These were cultured in medium supplemented with 200 PCi [35S]methionine ml ~ ’ and maintained for 2 or 3 days at 37°C.
2.4. SDS-PAGE and fluorograph?
For one dimensional SDS-PAGE [I 11, proteins were extracted from larvae by boiling for 5 min in
electrophoresis sample buffer (3% (w/v) SDS, 62 mM Tris-HCl, pH 6.8, 15% (v/v) glycerol) con- taining 5% 2-mercaptoethanol. Proteins precipi- tated from supernatant medium of parasite cultures were re-solubilized directly in reducing sample buffer and boiled. All preparations were centrifuged for 10 min at 16 000 g to remove insoluble material, and electrophoresed at 20 mA on 8-20% gradient or 7% homogeneous slab gels with molecular mass markers (M,.: 200 kDa, myosin; 116 kDa, P-galactosidase; 97 kDa, phos- phorylase b; 67 kDa, bovine serum albumin; 43 kDa, ovalbumin; 30 kDa, carbonic anhydrase; 20 kDa, soybean trypsin inhibitor; 14 kDa, a-lactal- bumin). As required, liquid scintillation counting permitted the various samples to be adjusted to provide equivalent numbers of trichloroacetic acid-precipitable cpm per lane. For two-dimen- sional electrophoresis, samples were solubilised in lysis buffer (9.5 M urea, 5% 2-mercaptoethanol, 2% NP40, 2% ampholines [in proportion I:4 for pH 5-7:3.5-10 range]), aided by brief sonication. After centrifugation at 16 000 g to remove insolu- ble material, samples were fractionated by IEF [12], followed by 7% SDS-PAGE. Gels were fixed and stained with Coomassie Brilliant Blue, im- mersed in AmplifyTM (Amersham) for 30 min and dried for fluorography.
2.5. Protein pur$cation and amino acid sequencing
Two-dimensional electrophoresis was used to purify a 75-kDa protein of A. uiteae L3 larvae. Extracts corresponding with 200 unlabelled larvae and 20 labelled parasites were loaded and elec- trophoresed on each of 22 first and second dimen- sion gels. Autoradiographs were used to locate the position of the 75-kDa protein. The appropriate part of each gel was excised, the dried spots were pooled and then equilibrated in buffer (20% glyc- erol, 0.125 M Tris-HCl, pH 6.8, O.l’% SDS, 1 mM EDTA) with one change over 1 h. The equilibrated material was loaded onto a 1.5-mm SDS-PAGE minigel, electrophoresed at 30 mA and transferred by electroblotting onto polyvinyli- dene difluoride membrane (Applied Biosystems), using Tris-glycine buffer [13]. The membrane was

210 Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219
stained with Coomassie Brilliant Blue to locate and excise the protein band. N-terminal amino acid sequencing was performed by Edman degra- dation, using a model 473A automated protein sequencer (Applied Biosystems).
2.6. Polymerase chain reaction (PCR) amplifications and DNA sequencing
cDNA libraries prepared from mRNA of in- fective L3 larvae were constructed in /2Uni-ZAP XR (Stratagene) for A. oiteae (University of Hohnheim, Germany) and 0. volvuZus (Smith College, USA). Bacteriophage from each of these libraries was amplified by the sloppy plate method [14] and lambda DNA was prepared for use as template in gene-specific PCR amplifica- tions [14]. For the amplification of A. uiteae chitinase, a sense PCR primer (5’-GGGTAT- GTTCGTGGATGTTACTAC-3’) was designed based on the N-terminal sequence of the A. viteae 75-kDa product (see Fig. 5). Codon usage tables for 0. volvulus [ 151 and Brugia malayi [ 161 were employed in design of the oligonucleotide sequence. For the amplification of 0. volvulus chitinase, a sense primer (5’-GCACTCA- CATTCTTTATGCATTCG-3’) was designed based on a region of the A. viteae chitinase se- quence (Fig. 5). One or other of the gene-specific primers and an anti-sense primer based on the T7 promoter sequence (5’-TAATACGACTCAC- TATAGGG-3’) in /iUni-ZAP were used in PCR. Amplification products were ligated directly into the TA cloning vector, pCRI1 (Invitrogen) and double-stranded sequencing was performed by the dideoxy nucleotide chain termination method [17], using Sequenase version 2.0 T7 DNA poly- merase (US Biochemical, Cleveland, USA). Nested sets of oligonucleotide primers were used in sequencing reactions to generate contigs and determine the nucleotide sequence in both direc- tions. PCR reactions were repeated with the /ZUni-ZAP DNA templates and new sets of primers designed to overlap the original priming site and extend the sequences 5’ of the N-termi- nus determined from protein sequencing. These included a sense primer based on the T3 pro- moter sequence in 1Uni-ZAP (5’-ATTAACCCT-
CACTAAAGGGA-3’) and antisense primers based on gene-specific internal sites predicted by DNA sequencing (5’-GATGATGTCTCGGTC- GATGTTTCA-3’ for A. viteae; 5’-TRTTA- CACGTGAGTACATYCC-3’ for 0. volvulus; see Fig. 5). Deduced amino acid sequences were pre- dicted using the GeneJockey program from Bio- Soft (Cambridge, UK). BLAST N and BLAST P programs [18] were used to search the computer databases for sequence similarities.
2.7. Chitinolytic assay
Chitinase activity in parasite extracts was mea- sured by the method of glycol chitin substrate gel electrophoresis [ 191. Extracts of larvae were pre- pared in sample buffer [ll], without 2-mercap- toethanol or boiling, and subjected to SDS-PAGE on 8-20% gradient gels containing 0.01% (w/v) glycol chitin. Following elec- trophoresis, gels were immersed in 1% (v/v) Tri- ton X-100 and incubated overnight at 37°C. The gels were stained for glycoi chitin with caicofluor white M2R (Sigma) and destained in water, us- ing ultraviolet light to excite the fluorochrome and reveal dark bands of lysis corresponding with chitinolytic activity. Purified hen egg white lysozyme (lo-50 ng) was included as a positive control [ 191.
2.8. Deglycosylation of parasite products
Metabolically labelled L3 larvae of A. viteae were heated to 100°C for 5 min in 10 ~1 of buffer containing 0.5% SDS, 10 mM EDTA, 10 mM 2-mercaptoethanol, 100 mM Tris-HCl pH 7.0. After cooling, 15~1 of 2% NP-40 in 100 mM Tris-HCl, pH 7.0, and 5 units of peptide N-gly- cosidase F enzyme (PNGase F, Boehringer) were added. Samples were incubated for 16 h at 37°C. Control reactions were run in parallel, from which the enzyme was omitted. For the analysis of reaction products, 15 ~1 of 4 x electrophore- sis sample buffer was added and the material was heated to 100°C for 5 min. SDS-PAGE and fluorography were performed as described above.
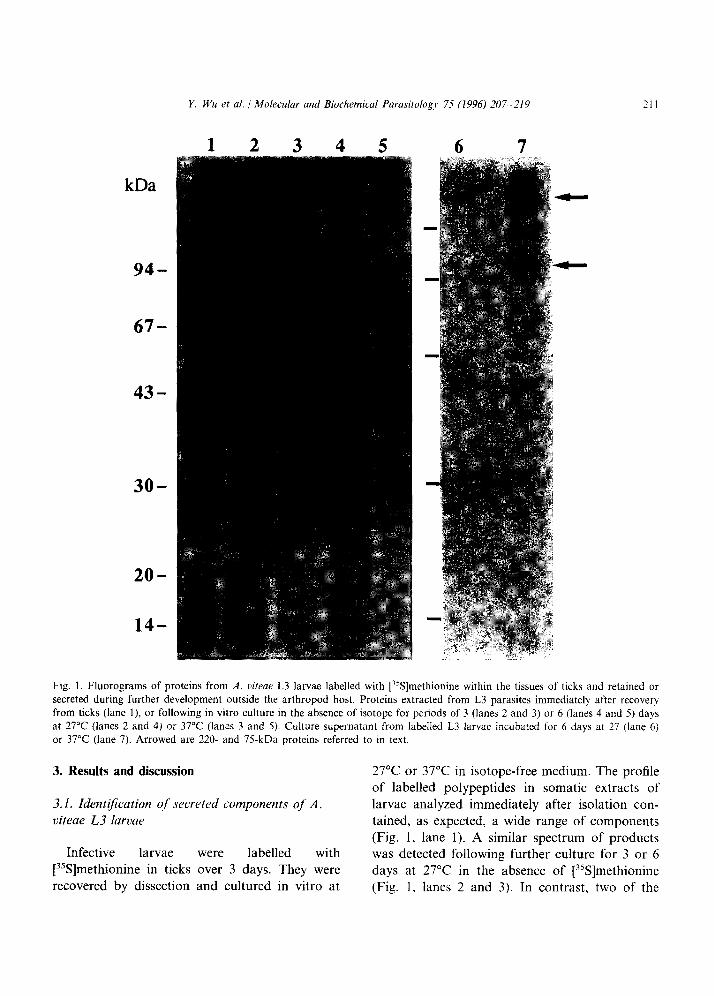
Y. Wu et al. 1 Molecular and Biochemical Parasitolog?, 75 (1996) 207-219 211
kDa
Fig. I. Fluorograms of proteins from A. uiteae L3 larvae labelled with [%]methionine within the tissues of ticks and retained or secreted during further development outside the arthropod host. Proteins extracted from L3 parasites immediately after recovery from ticks (lane I), or following in vitro culture in the absence of isotope for periods of 3 (lanes 2 and 3) or 6 (lanes 4 and 5) days at 27°C (lanes 2 and 4) or 37°C (lanes 3 and 5). Culture supernatant from labelled L3 larvae incubated for 6 days at 27 (lane 6) or 37°C (lane 7). Arrowed are 220- and 75-kDa proteins referred to in text.
3. Results and discussion
3.1. IdentlJhtion of secreted components of A. viteae L3 larvue
Infective larvae were labelled with
[35S]methionine in ticks over 3 days. They were
recovered by dissection and cultured in vitro at
27°C or 37°C in isotope-free medium. The profile
of labelled polypeptides in somatic extracts of
larvae analyzed immediately after isolation con-
tained, as expected, a wide range of components
(Fig. 1, lane 1). A similar spectrum of products
was detected following further culture for 3 or 6
days at 27°C in the absence of [35S]methionine
(Fig. 1, lanes 2 and 3). In contrast, two of the

212 Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219
12 345 678 910
kDa
30-
20-
14-
Fig. 2. Fluorograms of [35S]methionine-labelled proteins synthesized by A. uiteae parasites at various points throughout the life-cycle. Pulse-labellings were performed over I-3-day periods. L3 larvae Iabelled in the vector at 27°C (lane 1) or during post-infective culture in vitro at 37°C (lane 2). Larvae labelled in vitro at 37°C after development for 5 (lane 3) or 21 (lanes 4 and 5) days in jirds. Worms recovered on day 21 could be separated into male (lane 4) and female (lane 5) worms. Developing embryos (lane 6) adult males (lane 7) or adult females (lane 8) recovered from jirds and labelled in vitro at 37°C. Microfilariae purified from blood and labelled in vitro at 27°C (lane 9) or 37°C (lane 10). Arrowed on the left is a 75-kDa protein referred to in text.
most intensely labelled constituents of M, 220 and 75 kDa were selectively depleted from the somatic extracts of parasites that had been cul- tured at 37°C (Fig. 1, lanes 3 and 5). This was evident within 3 days of incubation, and by 6 days both polypeptides were undetectable. Both proteins were present in supernatant medium from cultures maintained at 37°C (Fig. 1, lane 7), but at greatly reduced levels at 27°C (Fig. 1, lane 6). The temperature dependency of secretion suggests that it may be triggered by post-infec- tive development, because only those parasites maintained at the higher temperature are stimu- lated to undergo differentiation and moulting to L4 larvae [7].
3.2. Stage-specificity of expression of the 220- and 75kDa products
Secretion of the 220- and 75-kDa proteins by A. viteae post-infective larvae might represent a dis- crete event, the start of sustained secretion with ongoing synthesis, or a discontinuous process as- sociated with the lead up to moulting. To discrim- inate amongst these possibilities, various developmental stages of A. viteae were pulse-la- belled with [35S]methionine in vitro to determine the kinetics of de novo synthesis of these molecules (Fig. 2). L3 larvae pulse labelled in vitro at 27’C, so as to provide no stimulus for further development, continued to elaborate both the 220- and 75-kDa products (Fig. 2, lane 1) as

Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219 213
we had previously demonstrated in ticks (Fig. 1. lane 1). However, maintenance of the same larvae at 37°C resulted in an abrupt cessation in the synthesis of these proteins (Fig. 2, lane 2). Metabolic labellings were also carried out with developmental stages recovered from jirds. Worms recovered after 5 days (i.e., 48 h before the third moult [20]), 21 days (i.e., 48 h before the fourth moult [ZOJ) or 60 days (i.e., following mat- uration to adults [20]) of infection were labelled for 24 h in cultures maintained at 37°C. Develop- ing embryos from fecund females were isolated from the uteri after labelling for independent analysis. Microfilariae obtained from the blood were labelled in vitro at either 37°C or 27°C to reflect temperature conditions experienced before and after transmission from the vertebrate to invertebrate host. In none of these stages was there clear evidence of expression of the 75-kDa protein (Fig. 2, lanes 3-10). A polypeptide of estimated IM, 75 kDa was present in microfilariae, although this was expressed at 37°C and not at 27”C, contrasting with the temperature-sensitive synthesis of the 75-kDa protein in L3 stages (Fig. 2, lanes 9, 10). Data on synthesis of the 220-kDa protein were less easy to interpret because several products co-migrated in this region of the gel.
3.3. Pur@ication of the 7_5-kDa product and derivation of N-terminal sequence
Synthesis and secretion of the 75-kDa product of L3 larvae was studied by two-dimensional elec- trophoresis as a prelude to its purification. Lar- vae were metabolically labelled in ticks and examined directly (Fig. 3A), or further cultured in isotope-free medium for 3 days at 37°C (Fig. 3B) which had been established to trigger secre- tion of the 75-kDa protein (Fig. 1). In this pulse- chase experiment, a polypeptide of 75 kDa and pZ 6.2 was found to be selectively depleted from somatic larval extracts following in vitro cultiva- tion (Fig. 3A and B, arrowed). Loss of this molecule was not observed in larvae incubated at 27°C (data not shown). Unlabelled L3 larvae from ticks were also pulse-labelled in vitro at 27°C (Fig. 3C) and 37°C (Fig. 3D). A polypep- tide of the same apparent molecular mass and
isoelectric point was specifically synthesized only at the lower temperature (Fig. 3C and D, down- ward arrows), confirming our earlier observations based on one-dimensional electrophoresis (Fig. 2). The same temperature-sensitivity appeared to govern synthesis (Fig. 3C and D, upward arrows) and secretion (Fig. 3A and B, upward arrows) of the 220-kDa protein, which migrated as a smear with a pZ value in the range of 7.0-7.8.
Two-dimensional electrophoresis was used to isolate the 75-kDa product as described in Mate- rials and methods. From a total of 2500 L3 larvae fractionated on 22 gels, 0.9 pmol of protein was purified f’rom which the N-terminal protein sequence XYVRGFYYTNXAQYR was obtained.
3.4. The 75-kDa product of L3 larvae is u chitinase
The N-terminal amino acid sequence was used to search the computer databases for homologous protein sequences using BLAST P [18]. This re- vealed a striking similarity with the N-terminus of an endochitinase described by Fuhrmann et al. [21] from Brugia maluyi (GYVRGCYYT- NWAQYR). Two isoforms of the B. malayi chiti- nase have been described of similar molecular mass (70 and 75 kDa), but unlike the A. viteae product they are reported to be stage-specific products of microfilariae. Accordingly, we used glycol-chitin substrate gel electrophoresis [ 193 to determine. whether the 75-kDa product of A. viteae L3 larvae has functional chitinolytic activ- ity. The results of this experiment are shown in Fig. 4. Somatic extracts of L3 larvae taken di- rectly from ticks contained molecules with dis- tinct chitin degrading activity that migrated at an A4, of 75 kDa (Fig. 4). The chitinolytic activity was lost if extracts were prepared from larvae that had been cultured at 37°C for 3 days after recovery from ticks (not shown). However, it was retained in larvae that were cultured at 27°C. These features are consistent with the 75-kDa protein defined by our earlier metabolic labelling experiments. No chitinolytic activity was seen that co-migrated with the 220-kDa product.
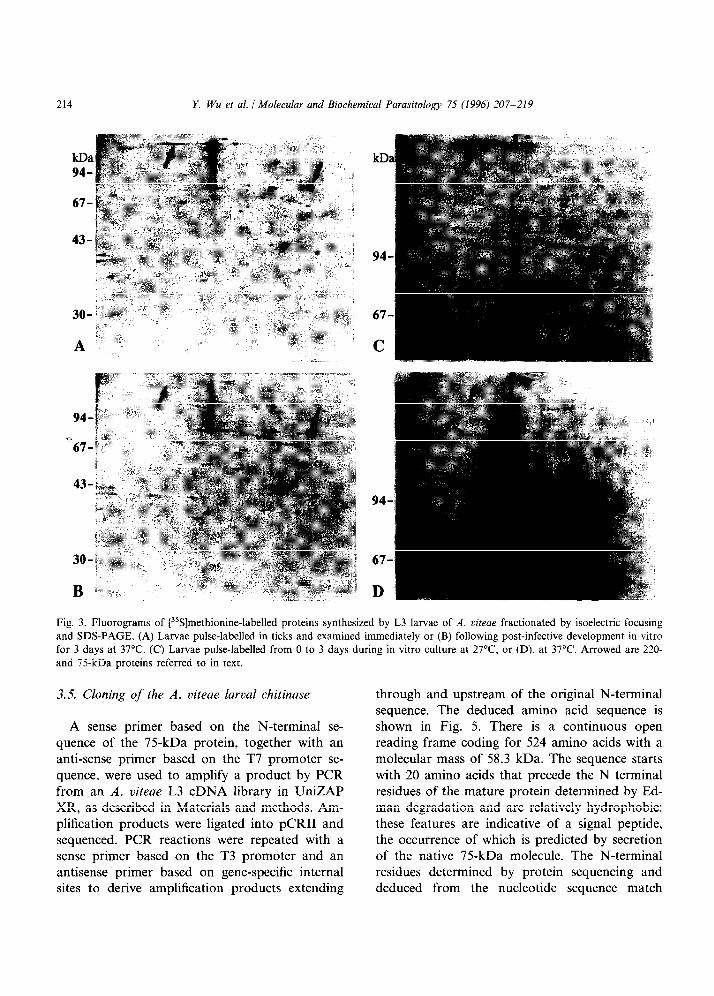
214 Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219
67
D
Fig. 3. Fluorograms of [35S]methionine-labelled proteins synthesized by L3 larvae of A. uiteae fractionated by isoelectric focusing and SDS-PAGE. (A) Larvae pulse-labelled in ticks and examined immediately or (B) following post-infective development in vitro for 3 days at 37°C. (C) Larvae pulse-labelled from 0 to 3 days during in vitro culture at 27”C, or (D), at 37°C. Arrowed are 220- and 75-kDa proteins referred to in text.
3.5. Cloning of the A. viteae larval chitinase
A sense primer based on the N-terminal se- quence of the 75-kDa protein, together with an anti-sense primer based on the T7 promoter se- quence, were used to amplify a product by PCR from an A. viteae L3 cDNA library in UniZAP XR, as described in Materials and methods. Am- plification products were ligated into pCRI1 and sequenced. PCR reactions were repeated with a sense primer based on the T3 promoter and an antisense primer based on gene-specific internal sites to derive amplification products extending
through and upstream of the original N-terminal sequence. The deduced amino acid sequence is shown in Fig. 5. There is a continuous open reading frame coding for 524 amino acids with a molecular mass of 58.3 kDa. The sequence starts with 20 amino acids that precede the N terminal residues of the mature protein determined by Ed- man degradation and are relatively hydrophobic: these features are indicative of a signal peptide, the occurrence of which is predicted by secretion of the native 75-kDa molecule. The N-terminal residues determined by protein sequencing and deduced from the nucleotide sequence match

Y. Wu et al. / Molecular and Biochemical Parasitologjj 75 (1996) 207-219 ?I5
closely: However, there was one discrepancy at cysteine-6 assigned as phenylalanine in the origi- nal N-terminal sequencing. The remaining protein is relatively hydrophilic and has an estimated M, of 56.1 kDa. There is a single site conforming with the motif for N-linked glycosylation (NDT, see Fig. 5) [22], seven sites of potential myristyla- tion [23] and 19 sites of possible casein kinase II, tyrosine kinase, or protein kinase phosphorylation [24-261 (not shown). The open reading frame terminates in a TGA stop codon followed by a short 3’ untranslated region (50 nucleotides). A polyadenylation signal (AATAAA) occurs 10 nu- cleotides upstream of the poly-(A) tail (see Gen- Bank accession L42010). Sequence identity at the amino acid level (excluding the signal peptide) of the A. viteae protein with B. mnlayi MFl chitinase was 69%: allowing for conservative substitutions, similarity rose to 81%.
1 2
kDa
Fig. 4. Glycol-chitin substrate gel electrophoresis of a PBS-ex-
tract of A. uiteae infective larvae (lane 2). Arrowed is a 75kDa
protein exhibiting chitinolytic activity. Hen egg white lysozyme
was included as a positive control (lane 1).
3.6. A. viteae lurval chitinase is glycosylated
The size of the polypeptide predicted by the open reading frame of the A. viteae chitinase (56 kDa) is significantly smaller than the native protein estimated by SDS-PAGE (75 kDa). Since the amino acid sequence contained a putative site of N-linked glycosylation (Fig. 5), there was the possibility that this discrepancy might be ex- plained by addition of carbohydrate side chain(s). To investigate this, A. viteae L3 larvae were metabolically labelled in ticks. The parasite ex- tract was treated with PGNase F to cleave N- linked sugars, and the digestion products were analyzed alongside undigested extract by SDS- PAGE (Fig. 6). PGNase treatment resulted in a significant diminution in the relative amount of native molecule migrating at 75 kDa. In parallel, a band of approximately 56 kDa was detected (Fig. 6 lane 2) consistent with the size of the predicted polypeptide backbone derived from the gene sequence. Further experiments will be re- quired to confirm that this relates to N-linked glycosylation via the single predicted site at as- paragine-399.
3.7. Cloning of a related larval chitinase from 0. volvzllus
Owing to similarities in A. viteue larval chiti- nase and the B. maluyi MFl molecule, it was possible to use the aligned sequences to identify conserved regions for the construction of PCR primers. A sense primer based on the amino acid sequence CTHILYAFA in the A. viteae enzyme, together with an anti-sense primer based on the T7 promoter sequence, were used to amplify a product from an 0. volvulus L3 cDNA library constructed in /. Uni-ZAP XR. Amplification products were ligated into pCRI1 and sequenced. PCR reactions were repeated with a sense primer based on the T3 promoter and an antisense primer based on a gene-specific internal site to derive amplification products extending through and upstream of the original sense primer site. The deduced amino acid sequence is shown in Fig. 5. There is a continuous open reading frame coding for 497 amino acids with a molecular mass

216 Y. Wu et al. / Molecular and Biochemical Parasiiology 75 (1996) 207-219
o.vol~uh~ MRIGAMLIPFIILGNAI-IAYGWRGCWTNWAQYRQGEGKFLPEDIPKGLGTHILYAFAKVDQSGTSLPFEWNDEDT~ 79
A.vireae .NWI-.ITL....A...TVVN I . . . . . . . . . . . . . . . . . . . . .._................... EK.. .MA........E. 79 B.malayi .NRTTLILF....S.T.TVIH..........,...,D . . . . . . . GN..N..............EL.D.K.........E. 80
o.~oh’~hb~ SK GMYSRVTKLKEM)PEMKILLSYGGYNFGSSTFTAIRNRAEKRKHFIKSAIAFLRKNKFM;FDFDWEYPIGMAQEYAKC 159
A.viteae . . . . . . ..I..R....TL................A..AKS........Q..TT....H......L.....T.V.E.H... 159 B.malayi . . . . ..A....R.TN.GL.V...........AI..G.AKS.Q.TER............N.....L.....V.V.E.H... 160
O.VO~&S ~V~EEAKKSDSEQLLLTAAVSAGKIfTIDQSYNVSLGENFDLLSLMSYDFHGSW~LHAKLHPTKGETSGT 239
A.viieae .E...A.......E.GKQ............M...E........KSL...F.....L.....K.....GE.R..ER..... 239 Bmalayi .EA..T...... .T.GKQR...........G...G....E...K.....F.....L.....K.....G........V..I 240
o.~ohhS GIFNTEFi'ANYWL SKGMPKQKIIIGIPTYGRG~LRDSSKTTIGAEGISPSSPSTTNPAGGTAAYWEICKYLKEGGKETI 319
A.viteae .VC.........AE.............A.S.....SNP.E.A.....DR......................V.......V 319
Bmulayi . . . . . . . ..D..A......E.......M.AQ....DNP.E.A...AASR...A.K........S...............V 320
o.Voh’Uk4S DEQGVGACMVQGSQWYGYDNEETIRMKMRWLKEKGYGGAFIWTLDEDDFKGTSCGEGPYPLLSAINHELKGEATATTRSL 3%'
A.viteae .KKS.. .Y..K.D..H........KI..K....ED.....M..............K......N...NG.ES.Q.PS..AV 399
B.malayi HQE.. ..Y..K.D............I..K.............A......T.K...K.... ..N..SS..E..-------- 3%
o.VohhS RTT--------ITQSSTIGSTKFETTTTASEITKNNKIKTTTTIAVEPTGRSSDIK----------------CPESFGLFR 455 A.viteae PE.TEDTEVEAM.EAPGTANGTEME.AEMP.T.EDIE .E.T.AAPETTEGTEVETTTSWKPGEE...PD...P 479
B.malayi ---~IT--.EEPS.TE.EAYE.DETE.TSE~~AYD.DETEETSET.ATTYDTDETE--------GOF...RD...P 459
o.voh’uh~ HPNDCHLFIHCAHDHPYVKLCPPNTFFNDKIKVCDHFGEC-DE-- 497 A.viteae .HS.... ..N..NNY.HIME..VG...D.T....NYMRNAp.TCK 524 Bmalayi . . T . . . . ..Q..NNIA..MQ..AT.....A......MTNAP.TCI 504
Fig. 5. Deduced amino acid sequences of cDNAs of chitinase genes from 0. volvulus (GenBank accession L42021) and A. ciieae (L42010) infective larvae, aligned with the MFI chitinase gene from B. malayi microfilariae [21]. Numbering of the sequences begins from the start methionine. Residues identical to the 0. voluuius sequence are marked with a dot, while substitutions are shown in single letter code. Gaps introduced to maximize alignment are indicated by a dash. Putative signal peptide sequences are in italics. The N-terminal residues of the mature protein in A. viteae determined by amino acid sequencing are indicated by bold underlining. Residues corresponding with sites of annealing of sense (double underlining) and antisense (single underlining) primers that were used in conjunction with vector sequences during polymerase chain reaction amplifications of cDNA are marked. Asparagine residues occurring within the N-linked glycosylation concensus sequence are shown in bold. A region of sequence divergence referred to in text is indicated by stippled underlining.
of 55.7 kDa. The first 20 amino acids have the appearance of a signal peptide, since they are relatively hydrophobic and are followed immedi- ately by residues that align with the A. uiteae chitinase sequence. The remaining protein is mostly hydrophilic and has an estimated M, of 53.5 kDa. There is a single site conforming with the motif for N-linked glycosylation (NWS see Fig. 5) [22], seven sites of potential myristylation [23] and 16 sites of possible casein kinase II, tyrosine kinase, or protein kinase phosphorylation [24-261 (not shown). The open reading frame terminates in a TAG stop codon followed by a 3’ untranslated region (58 nucleotides) and poly-(A)
tail (see GenBank accession L42021). Overall se- quence identity (excluding the signal peptide) of the 0. volvulus and A. uiteae chitinases was 67%: allowing for conservative substitutions, this rose to 80% similarity.
3.8. Comparative primary structures of the jilarial
chitinase genes
An alignment of the deduced amino acid se- quences of the 0. volvulus, A. viteae and published B. malayi chitinases is shown in Fig. 5. The partial sequence of a chitinase gene from Wucjhereria bancrofti [27] was included in the comparison, but

Y. Wu et al. 1 Molecular and Biochemical Parasitology 75 (1996) 207-219 217
is not shown in the figure owing to the limited extent of homology. Alignments of the chitinase sequences from B. malayi and W. bancrofti with those from a more diverse range of organisms (from bacteria to yeast) have previously been published [21,27]. Each of the sequences in Fig. 5 has a putative signal peptide of 20 (0. uolvulus/A. uiteae) or 21 (B. malayi) amino acids: these share some similarity with one another, with seven iden- tical residues and five conservative substitutions,
kDa
94-
67-
43-
30-
20-
14-
Fig. 6. Fluorogram of [“SS]methionine-labelled proteins syn- thesized by L3 larvae of A. uiteae in ticks, treated with peptide N-glycosidase F (lane 2) or with buffer lacking enzyme (lane 1) to reveal proteins with carbohydrate side-chains attached via N-linked glycosylation. Arrowed are 75- and 56-kDa proteins referred to in text.
although their relatedness is markedly less than the amino acid sequences of the mature protein. Downstream of the signal peptide, the amino acid identity across all three sequences is 60% (72% including conservative substitutions). By compari- son, only 17% of residues from the W. bancrofti sequence (43% allowing for conservative substitu- tions) could be matched, the greatest similarity occurring between amino acids 158 and 232 (as numbered in Fig. 5).
Chitinases of bacteria and yeast are thought to possess four domains (including the signal pep- tide), the most conserved of which is the catalytic domain at the N-terminus of the molecule [28,29]. Another is the serine/threonine-rich region to- wards the C-terminal end, which appears to ex- hibit considerable degeneracy amongst the filarial chitinases based on the alignments in Fig. 5 (un- derlined residues). In the B. ma&i MFl se- quence, it has been suggested that the high PEST (proline, glutamic acid, serine, threonine) score [30] in this part of the molecule may be significant in the binding of caIcium, which is known to be important in activation of the MFl product in microfilariae [2 1,3 11. Possibly, therefore, this de- generacy has arisen in response to a requirement to achieve post-translational control over activa- tion of these enzymes, which may vary amongst species or stage. The extreme C-terminal region of chitinase in yeast is predicted to comprise the fourth domain, the high affinity chitin-binding domain [28]: Here the filarial chitinases exhibit a high degree of homology (Fig. 5).
One of the first descriptions of filarial chitinases was that from females of Onchocerca gibsoni, in which the enzyme was proposed to have a role in the formation of the chitinous egg shell and in hatching [32,33]. Parasite chitinases have also been implicated in determining infectivity for their mosquito hosts: both malaria and Brugia (MFl) chitinases are believed to be secreted as a mecha- nism to penetrate the peritrophic membrane, ei- ther by direct enzymatic action, or by competitive inhibition of mosquito lectins released as a defen- sive measure [21,34]. In addition, antibodies against the MFl chitinase mediate clearance of microfilaraemia from experimentally infected jirds, suggesting that the enzyme may be accessi-

218 Y. Wu et al. / Molecular and Biochemical Parasitology 75 (1996) 207-219
ble to immune attack [21,35]. The 43-kDa chiti- nase-like molecule from W. buncrofti is preferen- tially recognized by sera from putative immune individuals exposed to filariasis. This reinforces the notion that filarial chitinases may play a role in the induction of host immunity [27,36]. It would be interesting to determine whether immu- nization with the A. viteae or 0. volvulus larval chitinases interferes with the transmission and/or establishment of infective larvae.
The function of the larval chitinases in A. viteae and 0. volvulus is as yet unknown. They may aid in egress of larvae from the mouth-parts of the vector, although this seems unlikely in view of the timing of secretion of the A. viteae product. More likely is that these enzymes may participate in ecdysis by modifying the L3 cuticle as a prelude to moulting. In A. uiteae, the third moult occurs 7 days after transmission of infective larvae, so secretion of the chitinase must be an early event in the processes leading to moulting if it does have this role. Interestingly, we were unable to observe de novo synthesis of the chitinase by post-infec- tive larvae, including parasites approaching the third and fourth moults. If it does participate in ecdysis, then it may be that the L3 cuticle has distinct biochemical properties requiring the ac- tion of a chitinolytic enzyme, possibly as a conse- quence of it being evolved for life in both the invertebrate and vertebrate host.
Acknowledgements
We appreciate the assistance of Professor Richard Lucius in whose laboratory the A. viteae cDNA library was constructed and Dr Rosalind Jenkins for critical comments and aid in prepara- tion of the manuscript. We thank Professor Mike Ferguson for providing a protocol for N-gly- canase digestion. The 0. VOZVUZUS infective larvae cDNA library was prepared on behalf of the Edna McConnell Clark Foundation with larvae supplied by Drs Milan Trpis and Sara Lustigman. N-terminal amino acid sequencing was performed by Dr John Fox of Alta Bioscience, University of Birmingham, UK. This work was supported by a grant from the Wellcome Trust.
References
[1] Bianco, A.E., Robertson, B.D., Kuo, Y.-M., Townson, S. and Ham, P.J. (1990) Developmentally regulated expres- sion and secretion of a polymorphic antigen by Onchocerca infective-stage larvae. Mol. Biochem. Para- sitol. 39, 2033212.
[2] Scott, A.L., Ibrahim, MS. and Tamashiro, W.K. (1990) Surface associated antigens of second, third and fourth stage larvae of Dirofilaria immitis. Acta Trop. 47, 339- 353.
[3] Frank. G.R. and Grieve, R.B. (1991) Metabolic labelling of Dirojiilaria immitis 3rd~stage and 4th~stage larvae and their excretory-secretory products. J. Parasitol. 77, 950- 956.
[4] Ibrahim, M-S., Richie, T.L. and Scott, A.L. (1992) Sur- face-associated antigens of Brugia malayi L2 and L3 parasites during vector-stage development. Mol. Biochem. Parasitol. 52, 97-l 10.
[5] Martin, S.A.M., Thompson, F.J. and Devaney, E. (1995) The construction of spliced leader cDNA libraries from the filarial nematode Brugia malayi. Mol. Biochem. Para- sitol. 70, 241-245.
[6] Yenbutr, P. and Scott, A.L. (1995) Molecular cloning of a serine proteinase inhibitor from Brugia malayi. Infect. Immun. 63, 1745-1753.
[7] Franke, E.D. and Weinstein, P.P. (1984) In vitro cultiva- tion of Dipetalonema viteae 3rd- stage larvae - evalua- tion of culture media, serum, and other supplements. J. Parasitol. 70, 618-628.
[8] Lok, J.B., Pollack, R.J., Cupp, E.W., Bernardo, M.J., Donnelly, J.J. and Albiez, E.J. (1984) Development of Onchocerca lienalis and 0. volvulus from the third to fourth larval stage in vitro. Tropenmed. Parasitol. 35, 2099211.
[9] Worms, M.J.. Terry, R.J. and Terry, A. (1961) Dipetalonema vitei: filarial parasite of the jird, Meriones libycus. I. Maintenance in the laboratory. J. Parasitol. 47, 963-970.
[IO] Eisenbeiss, W.F., Apfel, H. and Meyer, T.F. (1991) Pro- tective immunity linked with a distinct developmental stage of a filarial parasite. J. Immunol. 152, 7355742.
[11] Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of head of bacteriophage T4. Nature 227. 680-685.
[12] O’Farrell, P.H. (1975) High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 250, 40077 4021.
[13] Towbin, H., Staehelin, T. and Gordon, J. (1979) Elec- trophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350-4354.
1141 Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) In: Molecular Cloning. A Laboratory Manual. 2nd edition. (Nolan, C., ed.), Vol. I. New York: Cold Spring Harbor Laboratory Press.

Y. Wu et al. 1 Molecular and Biochemical Parasitology 75 (1996) 207-219 319
u51
1161
[I71
[181
[I91
PO1
PII
WI
~231
~241
P51
WI
Unnasch, T.R., Katholi. CR. and Coate, L.M. (1992) Onchocerca volvulus: frequency of codon usage. Exp. Par- asitol. 75, 457-459. Hammond, M.P. (1994) Codon usage and gene organiza- tion in Brugia. Parasitol. Res. 80, 173-175. Sanger, F., Nicklen, S. and Coulson, A.R. (1977) DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 546335467. Altschul, S.F., Gish. W., Miller, W., Myers, E.W. and Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215. 403-410. Trudel, J. and Asselin, A. (1989) Detection of chitinase activity after polyacrylamide gel electrophoresis. Anal. Biochem. 178, 362-366. Johnson. M.H., Orhiel, T.C. and Beaver, P.C. (1974) Diperalonerna uiteae in the experimentally infected jird, Meriones unguiculatus. 1. Insemination, development from egg to microfilaria, reinsemination and longevity of mated and unmated worms. J. Parasitol. 60, 302-309. Fuhrman. J.A.. Lane, W.S.. Smith. R.F., Piessens, W.F. and Perler. F.B. (1992) Transmission-blocking antibodies recognize microfilarial chitinase in brugian lymphatic filariasis. Proc. Natl. Acad. Sci. USA 89, 154881552. Marshall, R.D. (1972) Glycoproteins. Annu. Rev. Biochem. 41, 673-702. Towler, D.A.. Gordon, J.I., Adams, S.P. and Glaser, L. (1988) The biology and enzymology of eukaryotic protein acylation. Annu. Rev. Biochem. 57, 69-99. Cooper, J.A., Esch, F.S.. Taylor, S.S. and Hunter, T. (1984) Phosphorylation sites in enolase and lactate dehy- drogenase utilized by tyrosine protein kinase in vivo and in vitro. J. Biol. Chem. 259. 7835-7841. Kishimoto, A., Nishiyama, K., Nakanishi, H., Uratsuji, Y., Nomura, H., Takeyama, Y. and Nishizuka, Y. (1985) Studies on the phosphorylation of myelin basic protein by protein kinase C and adenosine 3’:5’-monophosphate-de- pendent protein kinase. J. Biol. Chem. 260. 12492- 12499. Pinna, L.A. (1990) Casein kinase 2 - an eminence-grise in cellular regulation. Biochim. Biophys. Acta 1054, 267- 284.
~271
[281
1291
[301
1311
1321
[331
t341
[351
[361
Raghavan. N., Freedman, D.O.. Fitzgerald, P.C.. Un- nasch, T.R., Ottesen, EPA. and Nutman, T.B. (1994) Cloning and characterization of a potentially protective chitinase-like recombinant antigen from Wuchereria ban- crofti. Infect. Immun. 62, 1901-1908. Kuranda, M.J. and Robbins. P.W. (1991) Chitinase is required for cell separation during growth of Saccha- romyces cereuisiae. J. Biol. Chem. 266, 19758- 19767. Watanabe, T., Suzuki, K., Oyanagi, W., Ohnishi. K. and Tanaka, H. (1990) Gene cloning of chitinase Al from Bacillus circulans WL-12 revealed its evolutionary rela- tionship to Serratia chitinase and to the type 111 homol- ogy units of fibronectin. J. Biol. Chem. 265, 15659915665. Rogers, S.. Wells, R. and Rechsteiner, M. (1986) Amino acid sequences common to rapidly degraded proteins - the PEST hypothesis. Science 234, 364-368. Fuhrman, J.A. and Piessens, W.F. (1989) A stage-specific calcium-binding protein from microfilariae of Brugia malayi (Filariidae). Mol. Biochem. Parasitol. 35, 249- 258. Brydon, L.J.. Gooday, G.W.. Chappell, L.H. and King, T.P. (1987) Chitin in egg shells of Onchocerca gibsoni and Onchocerca ooloulus. Mol. Biochem. Parasitol. 25, 267 272. Gooday, G.W., Brydon, L.J. and Chappell, L.H. (1988) Chitinase in female Onchocerca gibsoni and its inhibition by allosamidin. Mol. Biochem. Parasitol. 29, 223-225. Shahabuddin, M., Toyoshima, T.. Aikawa, M. and Kaslow, D.C. (1993) Transmission-blocking activity of a chitinase inhibitor and activation of malarial parasite chitinase by mosquito protease. Proc. Natl. Acad. Sci. USA 90, 4266-4270. Canlas M. Wadee, A., Lamontagne, L. and Piessens. W.F. (1984) A monoclonal antibody to surface antigens on microfilariae of Brugia mala.vi reduces microfilaremia in infected jirds. Am. J. Trap. Med. Hyg. 33. 420-424. Freedman, D.O., Nutman, T.B. and Ottesen, E.A. (1989) Protective immunity in Bancroftian filariasis: Selective recognition of a 43 kD larval stage antigen by infection- free individuals in an endemic area. J. Clin. Invest. 83, 14-22.










