Central Australian waterbodies: the importance of permanence in a desert landscape
19
ARTICLE IN PRESS Review Central Australian waterbodies: The importance of permanence in a desert landscape J.B. Box a, , A. Duguid a , R.E. Read b , R.G. Kimber c , A. Knapton b , J. Davis d , A.E. Bowland e a Biodiversity Conservation Unit, Department of Natural Resources, Environment and the Arts, P.O. Box 1120, Alice Springs, NT 0870, Australia b Natural Systems Division, NorthernTerritory Government Department of Natural Resources Environment and the Arts, Alice Springs, NT, Australia c P.O. Box 2436, Alice Springs, NT, Australia d School of Environmental Science, Murdoch University, Western Australia, Australia e Wildlife Management Division, Parks and Wildlife Service of the Northern Territory, Palmerston, NT, Australia article info Article history: Received 9 July 2007 Received in revised form 24 January 2008 Accepted 27 February 2008 Keywords: Arid zones Central Australia Desert wetlands Groundwater Springs abstract Central Australia (CA) has an arid environment characterised by low and unpredictable rainfall, high temperatures and high evaporation rates. Within this desert context reliable water features, sustained mainly by natural groundwater discharge, provide distinct and isolated habitats for both aquatic and terrestrial species, even where these features are only minimally inundated. CA water features are varied, and include springs, waterholes and seepages that provide refuge for relict, endemic, and widely dispersed species confined to discontiguous habitats. They have been well known to Aboriginal people for thousands of years, and are often sites of great cultural significance. In spite of their biological, cultural and economic importance, the physical and ecological characters of many waterbodies are not well known. CA has experienced some of the most rapid rates of warming observed on the Australian continent. Successfully managing the con- sequences of climate change in CA will depend, in part, on developing a better understanding of the factors that influence long-term biodiversity in these aquatic systems. Because isolated CA waterbodies are at the forefront of continental warming, they could provide insights into how climate change will potentially impact other arid zone aquatic ecosystems. & 2008 Elsevier Ltd. All rights reserved. Contents 1. Introduction ......................................................................................... 1396 2. Factors that influence water permanence in CA ............................................................. 1397 2.1. Geography and climate .......................................................................... 1397 2.2. Water-related features ........................................................................... 1398 2.2.1. Stored surface waters..................................................................... 1398 2.2.2. Groundwater ........................................................................... 1399 3. Knowledge of the location and physical character of CA waterbodies ........................................... 1401 3.1. Historical Aboriginal knowledge of CA waters ........................................................ 1401 3.2. An overview of European explorers’ records of CA waters ............................................... 1403 3.3. Current knowledge of long-lasting CA water features .................................................. 1404 Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/jaridenv Journal of Arid Environments 0140-1963/$ - see front matter & 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.jaridenv.2008.02.022 Corresponding author. Tel.: +618 89518224; fax: +61889518790. E-mail address: [email protected] (J.B. Box). Journal of Arid Environments 72 (2008) 1395–1413
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Central Australian waterbodies: the importance of permanence in a desert landscape

ARTICLE IN PRESS
Contents lists available at ScienceDirect
Journal of Arid Environments
Journal of Arid Environments 72 (2008) 1395– 1413
0140-19
doi:10.1
� Cor
E-m
journal homepage: www.elsevier.com/locate/jaridenv
Review
Central Australian waterbodies: The importance of permanence in adesert landscape
J.B. Box a,�, A. Duguid a, R.E. Read b, R.G. Kimber c, A. Knapton b, J. Davis d, A.E. Bowland e
a Biodiversity Conservation Unit, Department of Natural Resources, Environment and the Arts, P.O. Box 1120, Alice Springs, NT 0870, Australiab Natural Systems Division, Northern Territory Government Department of Natural Resources Environment and the Arts, Alice Springs, NT, Australiac P.O. Box 2436, Alice Springs, NT, Australiad School of Environmental Science, Murdoch University, Western Australia, Australiae Wildlife Management Division, Parks and Wildlife Service of the Northern Territory, Palmerston, NT, Australia
a r t i c l e i n f o
Article history:
Received 9 July 2007
Received in revised form
24 January 2008
Accepted 27 February 2008
Keywords:
Arid zones
Central Australia
Desert wetlands
Groundwater
Springs
63/$ - see front matter & 2008 Elsevier Ltd.
016/j.jaridenv.2008.02.022
responding author. Tel.: +618 89518224; fax
ail address: [email protected] (J.B. B
a b s t r a c t
Central Australia (CA) has an arid environment characterised by low and unpredictable
rainfall, high temperatures and high evaporation rates. Within this desert context reliable
water features, sustained mainly by natural groundwater discharge, provide distinct and
isolated habitats for both aquatic and terrestrial species, even where these features are
only minimally inundated. CA water features are varied, and include springs, waterholes
and seepages that provide refuge for relict, endemic, and widely dispersed species
confined to discontiguous habitats. They have been well known to Aboriginal people for
thousands of years, and are often sites of great cultural significance. In spite of their
biological, cultural and economic importance, the physical and ecological characters of
many waterbodies are not well known. CA has experienced some of the most rapid rates
of warming observed on the Australian continent. Successfully managing the con-
sequences of climate change in CA will depend, in part, on developing a better
understanding of the factors that influence long-term biodiversity in these aquatic
systems. Because isolated CA waterbodies are at the forefront of continental warming,
they could provide insights into how climate change will potentially impact other arid
zone aquatic ecosystems.
& 2008 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1396
2. Factors that influence water permanence in CA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1397
2.1. Geography and climate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1397
2.2. Water-related features . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1398
2.2.1. Stored surface waters. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1398
2.2.2. Groundwater . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1399
3. Knowledge of the location and physical character of CA waterbodies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1401
3.1. Historical Aboriginal knowledge of CA waters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1401
3.2. An overview of European explorers’ records of CA waters. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1403
3.3. Current knowledge of long-lasting CA water features . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1404
All rights reserved.
: +618 89518790.
ox).

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131396
4. Biological attributes. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1404
4.1. Fauna . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1405
4.1.1. Fishes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1405
4.1.2. Macroinvertebrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1406
4.2. Flora . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1407
4.3. Ecological considerations. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1408
5. Activities that can potentially affect CA waterbodies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1408
5.1. Non-native plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1409
5.2. Feral animals and domestic stock . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1409
5.3. Water withdrawals and the creation of artificial wetlands . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1409
5.4. Climate change. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1410
6. Research needs. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1410
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1410
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1411
1. Introduction
The importance of reliable surface water in arid environments has been long recognised, but more so in terms of humanor animal uses than broader ecological significance. Globally the cultural, hydrologic, and biological values of aridwaterbodies are not well understood by resource managers, users, or researchers (Myers and Resh, 1999; Sada and Sharpe,2004), perhaps because they are often small, isolated features in expansive desert landscapes. In inland Australia, theimportance of sites with long-lasting surface waters is thought to far exceed their geographic area (Hatton and Evans,1998). Arid-zone springs in the western US have been referred to as, ‘‘precious jewels of the desert’’ (http://www.dri.edu/Home/Features/text/0705_springs.htm), a reflection of their fragility as well as ecologic importance (Sada and Pohlmann,2006). In CA many waterbodies are also culturally precious because they have deep ceremonial, economic, and socialsignificance for Aboriginal people.
The values traditionally ascribed to wetlands globally also apply to most arid-zone wetlands, and includeeconomic, cultural, aesthetic, scientific, environmental, and conservation importance (Fisher et al., 1972; Myersand Resh, 1999; Williams, 1999). Arid-zone waterbodies, due to their isolation, often contain high endemismand can be biological hotspots that contribute to regional aquatic biodiversity in disproportion to their size (Sada et al.,2005; Shepard, 1993; Williams, 1999). They also can be important areas for terrestrial animals and plants (Gillis,1991; Gubanich and Panik, 1986). Because they provide reliable drinking water, arid waterbodies are often heavilyused by humans and/or their livestock, and historically these uses were rarely balanced against ecological needs orconsiderations (Sada and Pohlmann, 2006; Shepard, 1993). In the western US, for example, native species that rely onspring-fed aquatic systems have suffered higher extinction rates than species found in any other arid land type (Sada andVinyard, 2002). When wetlands in arid zones are degraded by human activities, the impacts may be irreversible (Williams,1999).
CA, despite its arid environment, contains a diverse array of aquatic habitats, including permanent, temporary, salineand freshwater environments (Duguid et al., 2005). Virtually all sites in CA with long-lasting surface water have somecomponent of groundwater discharge. Groundwater-fed systems in CA include spring-fed pools and runs, riverinewaterholes, and areas such as seepages where surface waters can be minimal but long lasting. Groundwater can alsoinfluence more temporary water features, such as freshwater swamps and saline playas.
Few broad-scales efforts have been made to sample CA waterbodies, and a general inventory of CA wetlandswas not made until 2001 (Duguid et al., 2005). This lack of effort may be due to the common perception that waterresources in CA are sparse (Davis, 1996) or alternatively, could be due to a lack of research capacity (e.g., the population ofthe Northern Territory (NT) is fewer than 210,000 people, with most residing in the capital city ca. 1000 km to the north ofCA). However, surveys of some of the more prominent waterbodies (e.g., Davis, 1995, 1996; Davis et al., 1993) suggest thatthese features are relatively unscathed by European settlement and that a unique aquatic fauna is present. If so, then CAwaterbodies can potentially serve as model systems to investigate: (1) the ecological role of unaltered isolated wetlands inarid zones, (2) the evolutionary responses, including allopatric speciation, to long-term habitat fragmentation, (3) theimportance of groundwater inputs to isolated aquatic ecosystems, and (4) the ecological responses to climate change inarid wetlands, as CA has experienced some of the strongest effects of global warming thus far recorded on the continent.
The objectives of this paper are to review the historical and current knowledge of waterbodies in CA, with an emphasison the physical and biological attributes of permanent water features. While some discussion of ephemeral waterbodiesand groundwater-dependent ecosystems (GDEs) are included, our focus is mainly on long-lasting surface waters, becausein CA permanent and semi-permanent springs and waterholes are of particular biological, ecological and culturalsignificance. While acknowledging the significant early exploration, discovery, use and naming of inland waters byAboriginal people, a detailed treatment of these issues is beyond the scope of this paper. However, a brief overview ofhistorical Aboriginal knowledge of CA waterbodies is presented. Activities that can impact waterholes and research needsare also discussed.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1397
2. Factors that influence water permanence in CA
2.1. Geography and climate
This paper focuses on the arid zone south of latitude 201S in the NT, hereafter termed CA (Fig. 1). Much of this region isflat with sparse, unconnected drainages. However, low relief ranges (100–800 m) are also abundant, and significantlyinfluence permanent water features through a myriad of factors, including groundwater discharge and recharge. Many CAsprings are found in these ranges and most river drainages originate from them.
The broad landscape of CA is considered ancient in a global context. The region has not been glaciated since at least thePermian Period; however, Quaternary global glacial cycles have strongly influenced palaeo-climates over the past tens tohundreds of thousands of years, causing variations in temperature and rainfall patterns (Allan and Lindesay, 1998).Consequently, fluvial processes have occurred more frequently in wetter periods while aeolian processes, including sanddune formation, have dominated dry periods (Wasson, 1984).
Fig. 1. CA, arbitrarily defined here as the part of the Northern Territory south of latitude 201S.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131398
The present climate is characterised by low and unpredictable rainfall and high temperatures and evaporationrates. For example, at Barrow Creek (Fig. 2) the mean annual rainfall from 1874 to 2004 was 316 mm while the meanannual pan evaporation was 3100 mm (O’Grady et al., 2007). Recently CA has experienced some of the most rapidclimate warming observed on the Australian continent, and temperature trend maps suggest CA is an area of particularlystrong warming trends when measured over most periods, especially those starting prior to 1950 (Bureau of Meteorology,2007).
2.2. Water-related features
2.2.1. Stored surface waters
CA waterbodies are mostly temporary and include freshwater lakes, clay pans, isolated rockholes, a variety of freshwaterswamps, riverine waterholes, and saline playas (Duguid et al., 2005) (Table 1). CA rivers are temporary, mostly isolatedfrom each other, and terminate in saline playas or flood out into swamps or sandy deserts (Duguid et al., 2005). All CA riversflow only after rain events, except in a few areas where springs contribute significant discharge to create short (e.g., tens ofmetres) runs. In rare cases heavy rains can contribute to river flow lasting weeks or months (e.g., upper and mid-Finke Riverin 2001).
Although in other parts of arid Australia riverine waterholes were identified as unique ecosystems almost entirelydependent on groundwater (Hatton and Evans, 1998), waterholes in CA are mostly inundated from stored surface water.There is no groundwater discharge at the majority of riverine waterholes in CA, which number in the thousands and aremainly temporary water features. Nevertheless some permanent springs and permanent or semi-permanent pools withinriver channels occur in CA, and these channel features are highly significant (Davis, 1996; Duguid et al, 2005; Spencer, 1896;Zietz, 1896).
Fig. 2. List of place names for CA waterbodies referred to in the text, most of which were named by local Aboriginal people or early European explorers.
ARTICLE IN PRESS
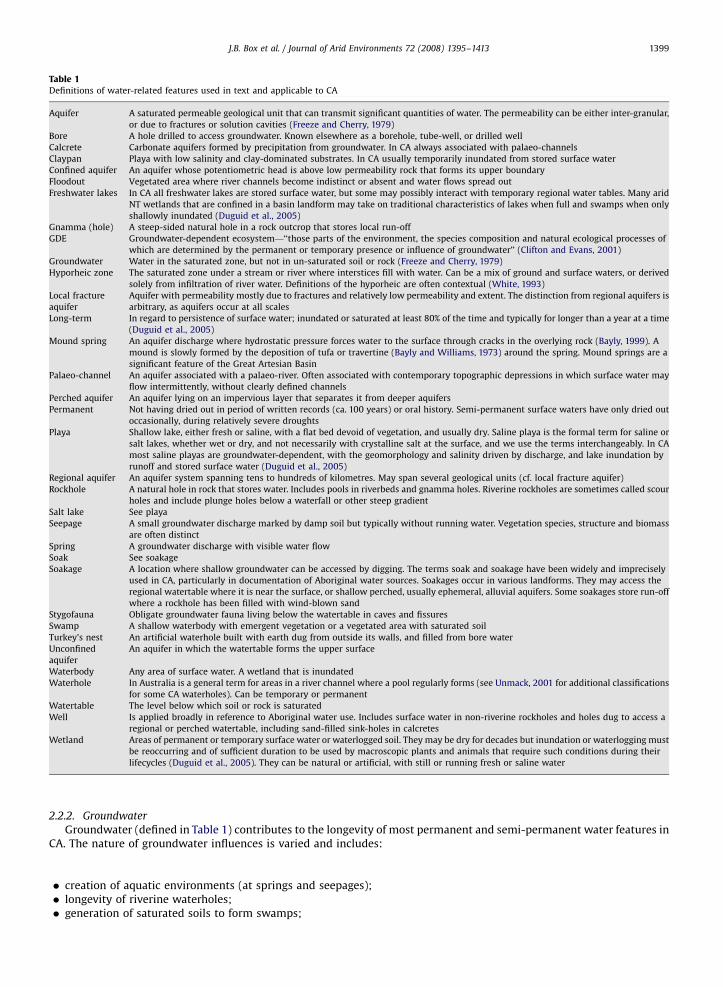
Table 1Definitions of water-related features used in text and applicable to CA
Aquifer A saturated permeable geological unit that can transmit significant quantities of water. The permeability can be either inter-granular,
or due to fractures or solution cavities (Freeze and Cherry, 1979)
Bore A hole drilled to access groundwater. Known elsewhere as a borehole, tube-well, or drilled well
Calcrete Carbonate aquifers formed by precipitation from groundwater. In CA always associated with palaeo-channels
Claypan Playa with low salinity and clay-dominated substrates. In CA usually temporarily inundated from stored surface water
Confined aquifer An aquifer whose potentiometric head is above low permeability rock that forms its upper boundary
Floodout Vegetated area where river channels become indistinct or absent and water flows spread out
Freshwater lakes In CA all freshwater lakes are stored surface water, but some may possibly interact with temporary regional water tables. Many arid
NT wetlands that are confined in a basin landform may take on traditional characteristics of lakes when full and swamps when only
shallowly inundated (Duguid et al., 2005)
Gnamma (hole) A steep-sided natural hole in a rock outcrop that stores local run-off
GDE Groundwater-dependent ecosystem—‘‘those parts of the environment, the species composition and natural ecological processes of
which are determined by the permanent or temporary presence or influence of groundwater’’ (Clifton and Evans, 2001)
Groundwater Water in the saturated zone, but not in un-saturated soil or rock (Freeze and Cherry, 1979)
Hyporheic zone The saturated zone under a stream or river where interstices fill with water. Can be a mix of ground and surface waters, or derived
solely from infiltration of river water. Definitions of the hyporheic are often contextual (White, 1993)
Local fracture
aquifer
Aquifer with permeability mostly due to fractures and relatively low permeability and extent. The distinction from regional aquifers is
arbitrary, as aquifers occur at all scales
Long-term In regard to persistence of surface water; inundated or saturated at least 80% of the time and typically for longer than a year at a time
(Duguid et al., 2005)
Mound spring An aquifer discharge where hydrostatic pressure forces water to the surface through cracks in the overlying rock (Bayly, 1999). A
mound is slowly formed by the deposition of tufa or travertine (Bayly and Williams, 1973) around the spring. Mound springs are a
significant feature of the Great Artesian Basin
Palaeo-channel An aquifer associated with a palaeo-river. Often associated with contemporary topographic depressions in which surface water may
flow intermittently, without clearly defined channels
Perched aquifer An aquifer lying on an impervious layer that separates it from deeper aquifers
Permanent Not having dried out in period of written records (ca. 100 years) or oral history. Semi-permanent surface waters have only dried out
occasionally, during relatively severe droughts
Playa Shallow lake, either fresh or saline, with a flat bed devoid of vegetation, and usually dry. Saline playa is the formal term for saline or
salt lakes, whether wet or dry, and not necessarily with crystalline salt at the surface, and we use the terms interchangeably. In CA
most saline playas are groundwater-dependent, with the geomorphology and salinity driven by discharge, and lake inundation by
runoff and stored surface water (Duguid et al., 2005)
Regional aquifer An aquifer system spanning tens to hundreds of kilometres. May span several geological units (cf. local fracture aquifer)
Rockhole A natural hole in rock that stores water. Includes pools in riverbeds and gnamma holes. Riverine rockholes are sometimes called scour
holes and include plunge holes below a waterfall or other steep gradient
Salt lake See playa
Seepage A small groundwater discharge marked by damp soil but typically without running water. Vegetation species, structure and biomass
are often distinct
Spring A groundwater discharge with visible water flow
Soak See soakage
Soakage A location where shallow groundwater can be accessed by digging. The terms soak and soakage have been widely and imprecisely
used in CA, particularly in documentation of Aboriginal water sources. Soakages occur in various landforms. They may access the
regional watertable where it is near the surface, or shallow perched, usually ephemeral, alluvial aquifers. Some soakages store run-off
where a rockhole has been filled with wind-blown sand
Stygofauna Obligate groundwater fauna living below the watertable in caves and fissures
Swamp A shallow waterbody with emergent vegetation or a vegetated area with saturated soil
Turkey’s nest An artificial waterhole built with earth dug from outside its walls, and filled from bore water
Unconfined
aquifer
An aquifer in which the watertable forms the upper surface
Waterbody Any area of surface water. A wetland that is inundated
Waterhole In Australia is a general term for areas in a river channel where a pool regularly forms (see Unmack, 2001 for additional classifications
for some CA waterholes). Can be temporary or permanent
Watertable The level below which soil or rock is saturated
Well Is applied broadly in reference to Aboriginal water use. Includes surface water in non-riverine rockholes and holes dug to access a
regional or perched watertable, including sand-filled sink-holes in calcretes
Wetland Areas of permanent or temporary surface water or waterlogged soil. They may be dry for decades but inundation or waterlogging must
be reoccurring and of sufficient duration to be used by macroscopic plants and animals that require such conditions during their
lifecycles (Duguid et al., 2005). They can be natural or artificial, with still or running fresh or saline water
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1399
2.2.2. Groundwater
Groundwater (defined in Table 1) contributes to the longevity of most permanent and semi-permanent water features inCA. The nature of groundwater influences is varied and includes:
�
creation of aquatic environments (at springs and seepages); � longevity of riverine waterholes; � generation of saturated soils to form swamps;
ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131400
�
generation of mesic micro-climates, notably in sheltered rocky areas; � contribution of mineral salts to aquatic environments and to soils; � formation of hypersaline environments and associated saline playas; � occasional formation of mounds due to a combination of mineral salts deposited from groundwater, and theaccumulation of organic matter and wind-blown soils;
� growth of terrestrial vegetation that uses groundwater and � contribution to subterranean environments including wet caves, aquifers and hyporheic zones.CA contains four major sedimentary basins (Fig. 3), and some of these basins have several distinct aquifers, both regionaland local. Groundwater is stored and transmitted in aquifers, which can be in rock or regolith (Jones and Quinlan, 1962) andcan be unconfined or confined by overlying impervious rock (Lloyd and Jacobson, 1987). In CA both local and regionalaquifers can influence permanent surface waters, while perched aquifers are uncommon and are typically temporarysources of groundwater. Regional aquifers in CA can span tens to hundreds of square kilometres and support someimportant permanent water features. Many regional aquifers in arid Australia are recharged from temporary inundations inrivers, swamps and lakes (Bowler, 1986) while others are predominantly recharged from rivers and river floodouts(e.g., Harrington, 1999).
Local aquifers, with flow paths that may be restricted to a few hundred metres, are also highly significant in CA. Themajority of springs in CA discharge from fractured rock aquifers of sandstone, fissured limestone or quartzite, and areusually located near the base of ranges. Localised fractured rock aquifers are often recharged after rain events by acombination of creek flow and infiltration directly into the ground.
Groundwater residence times in CA range from less than a year to tens of thousands of years (Cresswell et al., 1999a, b;Harrington, 1999; Jacobson et al., 1989), depending on the geological setting, and are a reflection of the prevailing aridityand the underlying hydrogeology and landforms of the region (Jacobson, 1996). Discharge from CA springs can varymarkedly over time, depending on the magnitude and timing of recharge events, and some may dwindle to seepages ordisappear completely between events. Surface flows can also be strongly influenced by evapotranspiration rates associatedwith ambient temperatures and vegetation.
Water chemistry is also strongly influenced by groundwater, and varies greatly from waterbody to waterbody. The lowconductivities (o100 mScm�1) measured at some CA springs represent some of the freshest sites recorded on the Australiancontinent (Davis, 1997). The low ionic values indicate both a lack of soluble salts in the aquifers (sandstones and quartzites)
Fig. 3. The major geological basins of CA that influence both regional and local aquifers found in the study area.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1401
and limited evaporative concentration. In contrast, the groundwater in sub-surface brine pools and springs associated withplayas may be nearly saturated with sodium chloride (e.g., 350,000–400,000 mg/L dissolved solids near Curtin Springs)(Jacobson, 1996), and develop when salts, both from rainfall and leached from sedimentary rocks, are concentrated interminal basins.
The terms soak and soakage have been widely used in CA, particularly by early European explorers and in thedocumentation of Aboriginal water sources (Table 1). Soakages occur in various landforms including dune swales andstreambeds. When relatively deep holes are dug to access a watertable the term ‘well’ is often used (e.g., Hercus and Clarke,1986) (Table 1). However, in descriptions of Aboriginal water use, the terms ‘well’ and ‘‘native well’’ are applied broadly,and can include non-riverine rockholes and shallow soaks.
3. Knowledge of the location and physical character of CA waterbodies
We summarise knowledge of the location and physical properties of long-lasting CA waterbodies into three culturaldomains: traditional Aboriginal, early European exploration and contemporary western knowledge. Whether individualwaterbodies were permanent was critical to how they were used by humans and biota, and we regard surface waters aspermanent if they have not dried up in oral or written history, or in living memory. Semi-permanent sites are those wherewater persists for years and which only dry out during severe droughts.
3.1. Historical Aboriginal knowledge of CA waters
Many waterbodies in CA have deep ceremonial, economic, and social significance for Aboriginal people. It is bothexplicit and implicit in the accounts of all of the earliest European explorers that, prior to their arrival, any waters that they‘‘discovered’’ had been used and intellectualised for thousands of years by Aboriginals. Indeed, it may be said thatAboriginal people had an encyclopaedic knowledge of all waters within their own traditional country, and often a goodknowledge of waters in the countries of their neighbours. This included recording and mapping within their oral traditionsas well as active management (e.g., ‘‘brush matting’’ or covering waterholes with dead branches) of many sites. Permanentsources of water were of particular importance and were essential for survival during severe droughts (Bayly, 1999; Gould,1969a) and most are still considered culturally significant.
Various aspects of traditional Aboriginal knowledge of water resources were reviewed by Magarey (1895), who notedthat knowledge about how to locate ‘‘life-saving’’ water was crucial to the ‘‘Australian aborigine, dwelling—in realmswhere water is scarce. This knowledge has passed down from generation to generation, and, having come [to also live in thearid lands], the white man, too, has inherited much of the treasured lore.’’ Magarey (1895), and later others (e.g., Bayly,1999; Charnley, 1954) categorised sources of drinking water, including soakages, native dams, claypans, riverinewaterholes, springs, small rockholes, crevices and gnamma holes (Table 1). Water or moisture was also obtained from rainwater in tree hollows, excavated tree roots, water-holding frogs (e.g., Cyclorana spp.), dew, tree bark, and edible tubers(e.g., Ipomoea spp.). Various aspects of traditional Aboriginal knowledge of water resources were reviewed by Bayly (1999),Charnley (1954), Gould (1969b), Kavanagh (1984), Kimber (1985, 1990, 2001), Magarey (1895), Noble and Kimber (1999),and Terry (1937).
Comments made by Magarey (1895) and Schulze (1891) indicate an early appreciation of the assistance (sometimesfreely volunteered and sometimes under duress) given to explorers and pioneer frontiersmen by traditional Aboriginalcustodians of different arid regions. However, based on the records of explorers and other early travellers, informationabout the flora and fauna associated with waterholes was patchy, tended to be very general, and rarely were Aboriginalspecies’ names recorded. In 1891 Louis Schulze, a missionary at Hermannsburg, was one of the first Europeans to recordspecific details of Arrernte (i.e., the local Aboriginal people) aquatic knowledge, by noting that there were at least nineArrernte names for Finke River fishes and one species of crayfish (Table 2).
As had been the case elsewhere in Australia, the missionaries’ drive to learn the local language, so that they couldtranslate the bible and convert Aboriginal people to Christianity, resulted in this kind of species knowledge beingincreasingly well-recorded, although in most cases the Arrernte names for species were generally given in association withthe common, rather than the scientific, names of the particular flora and fauna. Similarly, although early CA explorers andother travellers recorded various Aboriginal words for water and water-related features (e.g., see Giles, 1889; Gillen, 1995;Gosse, 1874) they do not greatly assist in scientific understandings of CA waterbodies.
Aboriginal knowledge of waterbodies is substantially restricted to oral tradition and anthropological reports, and themajority of knowledge is not in the public domain. In the southwest NT, for example, a few senior Aboriginal people wereable to name many soakages, cave pools, river pools and seepages that were subsequently listed in a public report (Toyneet al., 1996) but were not located on topographic maps. Only a ‘‘fraction’’ of these features were known to governmentagencies (Hatton and Evans, 1998), were documented in the scientific literature, and most still are not included ontopographic or hydrological maps (e.g., Wischusen, 1998). This disparity between current western knowledge andAboriginal knowledge of the locations of water sources is indicative of the dearth of current scientific understanding aboutwaterbodies across CA.

ARTICLE IN PRESS
Table 2Significant waterbodies named and observations made by early European explorers
Explorer/
surveyor
Expedition
dates
Significant waterbodies/entities named Water-related observations
John McD.
Stuart
1860– 1862 Finke River, Hamilton Spring, Hugh River, Mount Denison,
Mount Leichhardt, Annas Reservoir, Bonney Creek, Owen
Springs
First to note ‘‘new parrots’’, native wells, fish, reeds, rushes,
ducks, polygonum and cucumber plant associated with
water; first account of a soakage
J.W.
Waterhouse
1860– 1862 First person, as ‘‘naturalist’’ to Stuart’s expedition, with
scientific understanding to travel through CA. Made notes
of water permanency and aquatic species (e.g., snails,
mussels, crabs)
John Ross 1870– 1871 Todd River, Waite River, Stirling Creek First to record crabs (in a Stirling Creek tributary) and
mosquitoes as associated with any waters in CA; first to
mention coolabahs; first to emphasize tea-trees and
limestone good water indicators; mentioned cycad palms
common in gorges and high up in ranges
W.W. Mills 1871 Alice Spring (sic)
Ernest Giles 1872– 1873 Ehrenberg Range, Hull River, Lake Amadeus, Palmer River,
Middleton’s Fish Ponds, Docker River, Bloods Range
Petermann Ranges
Discovered the Glen of Palms and relict red cabbage palms;
first to record the Arrente name for the Finke River
(Larapinta—‘‘river [of] salt’’); gives first general
descriptions of insects associated with waters and their
near vicinity; noted deliberate Aboriginal burning took
place near waters, and that Aboriginal art was found in
rock-shelters near certain waters
W.C. Gosse 1873 Ayers Rock [Uluru], Reynolds Range, Cockatoo Creek,
Lander River, Maggie Spring [Mutitjulu Waterhole]
At Uluru recorded that ‘‘carpee’’ (now kapi) was the
Aboriginal word for water at that locality
P.E.
Warburton
1873 Central Mount Wedge, Glen Elder [Pulka Karrinya], Glen
Hughes, Ethel Creek, Mary Springs, Waite Creek, Eva
Springs [now Spring]
H.V. Barclay 1876 Jervois Range, Plenty River
F. Scarr 1878 Reported that the mudlark was ‘‘the only reliable water
indicating bird I know of.’’ Mentioned their camp was ‘‘in a
state of siegey [by] numberless rats,’’ one of the first
mentions of rats in CA
C. Winnecke 1877 Painta Springs, Bob Spring, Hale River
C. Winnecke 1879– 1880 Scarr Hill, Stokes Creek, Mt. Hogarth, Duffield Creek First reference to yabbies; first to comment on freshwater
mussels, and noted that in some Aboriginal camps they
lived ‘‘principally on fresh water mussels.’’ Learned that
‘‘coketa’’ was the word for water in the Sandover River
area. First to mention corellas and shells (i.e., soil of the
swampsy.covered with innumerable very friable shells’’)
C. Winnecke 1883 Adam Ranges, Mount Tietkens, Hay River Noted that Coonamucka Swamp, a year-long lasting water
‘‘studded with box trees,’’ was a place of ‘‘curious and rare
plants,’’ including a ‘‘carnivorous’’ plant
C. Chewings 1881– 1886 First to record details of the extensive Finke River
catchment area. Over a 5-year period, made more detailed
and accurate assessments of whether individual
waterbodies were permanent than previous explorers
W.H. Tietkens 1889 Gill Creek, Cleland Hills, Lake MacDonald First to clearly state the differences in geography and
vegetation on either side of Lake Amadeus. First to describe
aspects of the lake’s surface waters and salt crust.
Describes springs on the Erldunda cattle station
Horn
Expedition
1894 Led by the indefatigable surveyor-explorer Charles
Winnecke. First designated scientific expedition to CA. The
five scientists on the expedition collected over 170
previously unknown species of plants and animals.
Scientists on the expediton were also the first to put the
water supplies into a geological and geographical
perspective
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131402

ARTICLE IN PRESS
Table 2 (continued )
Explorer/
surveyor
Expedition
dates
Significant waterbodies/entities named Water-related observations
A.A. Davidson 1898–1890 Helped clarify relationships between permanence and fish
persistence. Noted three fish species—Hytrl’s catfish
(Neosilurus hyrtlii), spangled grunter (Leiopotherapon
unicolour), banded grunter (Amniataba percoides) co-
occurred in Bonney Creek.
Louis Schulze 1891 Missionary at Hermannsburg who was one of the first to
give not only a better appreciation of how the Finke River’s
waters increased in salinity during drought, but also an
early record of Arrernte (i.e., the local Aboriginal people)
knowledge of fish
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1403
3.2. An overview of European explorers’ records of CA waters
The first written descriptions of CA waterbodies are the records of early European explorers (Table 2). Initially mostexplorers’ notes on CA waterbodies were in regard to the provision of drinking water for members of the expedition andtheir animals. This is not surprising, in that they were attempting to traverse and map the maximum amount of countryand plot its nature with regards to mining, pastoral, and other interests of the predominantly European colonisers. Waterswere important in allowing them to progress, especially permanent or long-lasting waters that were readily accessible tohorses and camels. Although most explorers noted whether they considered particular waterbodies to be permanent ortemporary, their initial assessments of specific waterbodies (e.g., permanent or temporary; large or small) were influencedby climatic factors such as time of year and whether they travelled during droughts or after good rains. The difficulty theyhad in determining permanence is not surprising and contemporary scientific knowledge of hydro-period and hydrologicaldynamics is still sparse for many waterbodies.
On April 4, 1860 John McDouall Stuart recorded the first sighting of a CA waterbody by a European—the Finke River. Helater named other major creeks, springs, and waterbodies in CA (Hardman, 1975) (Table 2). Although Stuart initiallythought that the majority of these waterbodies were permanent, on subsequent trips he discovered that most had dried.Stuart also noted many native wells, rockholes and other catchments during his travels through this area, but consideredonly two to be permanent. Later explorers (i.e., Murray, 1907; Winnecke, 1882) correctly noted that these two waterbodiesalso dried out after prolonged drought.
In 1872 Ernest Giles followed the Finke River beyond Stuart’s farthest point (Giles, 1889). Giles added considerabledescription to the nature and distribution of waters, named a number of major CA waterbodies, both permanent andtemporary (Table 2), and was the first to record the presence of red cabbage palms (Livistona mariae subsp. mariae) in CA, agroundwater-dependent endemic subspecies. In 1873 William Gosse documented the presence of additional waterbodiesin southwest CA, including those at Uluru (Ayers Rock). Both Gosse and Giles travelled after extensive rains, and manymajor waters were later found to be temporary.
In the 1870s several other explorers discovered and named significant waterbodies in CA (Table 2). Many of these newlynamed ‘‘springs’’ were later found to revert to soakages in dry periods (Winnecke, 1882), including the one in the ToddRiver after which Alice Springs, the largest city in CA, was named.
Charles Chewings’ multiple explorations of the entire Finke River complex added many additional details about thewater resources of CA (Chewings, 1886) (Table 2). White (1914) later qualified some of Chewings’ observations on waterpermanence, and cautioned that during the drought of 1914 Running Waters, a normally large (44 km) series of connectedpools on the Finke River, had become a ‘‘small, putrid pool’’ and that only nearby Boggy Hole was a reliable source ofsurface water. Other explorers also added to the overall geographical and hydrological knowledge of CA (Table 2) andhelped to further clarify whether particular waterbodies were permanent.
While many explorers made records of the local flora and fauna, most were generalists who described the more obviousaspects; the presence of grasses, bushes, birds, fish, trees, etc.; made comments on a core number of plants and animalsthat were well-known as a result of previous knowledge (e.g., river red gums (Eucalyptus camaldulensis), mulga (Acacia
aneura), mallee (Eucalyptus spp.), wallabies (Petrogale spp.), zebra finches (Taeniopygia guttata)); made detailed amateurdescriptions of striking new plants or animals; and sometimes collected specimens so that scientists could accuratelydescribe them. Other than an occasional mention of fish and/or crabs and frogs, they rarely mentioned anything in regardsto the aquatic fauna of CA waterbodies. It was not until the Horn Expedition of 1894, the first scientific expedition to CA,that details of the flora and fauna of CA waterbodies were documented by professional scientists (see Morton andMulvaney, 1996).
Stuart and other European explorers used the presence of certain animals or plants, as well as the presence of Aboriginalcamps or tracks, as indicators of where water was present and/or permanent. For example, although Stuart mentioned thatgums (Eucalyptus spp.), ti-trees (Melaleuca spp.) and knotweed (Polygonum spp. or Persicaria lapathifolia) were good

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131404
indicators of water, he regarded only rushes, which presumably included bullrushes (Typha domingensis and Juncus spp.)and reeds (Phragmites australis), as evidence of permanent water (Hardman, 1975). While other plants, such as samphire(Halosarcia spp.) around saline playas, cane grass (Eragrostis australasica) at claypans, ‘‘water-weed’’ in rockholes, and nativefigs (Ficus spp.) in rocky areas, were sometimes mentioned by later explorers to indicate water was present, all agreed withStuart that rushes and reeds were significant indicators of permanence, even though during droughts reeds and rushescould die at the surface and water was not always visible or accessible. Similarly, Stuart noted that the presence ofAboriginal camps and tracks could indicate that water was nearby, but did not indicate permanence, as Aboriginal peoplemoved camp as waters dried up.
Various birds were mentioned by Stuart and all other explorers of CA as good indicators of water, but Stuart onlyconsidered ‘‘diamond birds’’ (zebra finches) as ‘‘a sure sign of the proximity of water.’’ Zebra finches, however, can obtainwater from areas that are not accessible to humans, and Scarr (1878) suggested that zebra finches were not entirely reliablewater indicators, and that in his opinion the ‘‘mudlark’’ [magpie-lark, Grallina cyanoleuca] was the ‘‘only reliable waterindicating bird.’’ Regardless, the presence of these species does not necessarily indicate the presence of permanent water.
The majority of explorers also mentioned the presence of fish (usually in general terms) when they were sighted.Initially many explorers thought that the presence of fish indicated that a waterbody was permanent, but on subsequenttrips were to find these same waterholes had dried. Davidson (1905) was the first to note that permanent headwaters wereessential to the persistence of fish in a particular drainage. He also went into more detail than most explorers by noting thatthree species of fish co-occurred in a single drainage (Table 2).
Little mention was generally made of aquatic invertebrates, although flies, mosquitoes, and sometimes ants were oftennoted in the vicinity of water. Ross (1870–1871) was the first to associate mosquitoes with water, and to record crabs(Holthusiana transversa) from CA. In 1872–1873 Giles (1889) provided the first general descriptions of insects associatedwith waters and their near vicinity—flies, ants, and ‘‘large and small red hornets.’’ These invertebrates, however, were notconsidered to be reliable indicators of the presence of permanent water.
3.3. Current knowledge of long-lasting CA water features
It is probable that both Aboriginal and ‘European’ observational knowledge of the location and persistence of waterholesand springs has diminished over the past 50 or so years, as people spend less time travelling the country on foot orhorseback and natural waterbodies have been replaced by bores as the primary sources of drinking water for humans andcommercial grazing herds.
In general, current knowledge of the location of long-lasting CA water features is incomplete. The number of knownsprings, seepages and waterholes in CA believed to be permanent (and probably groundwater-fed) are grouped by geo-hydrological characters and presented in Table 3. This table was compiled from a variety of sources, including a detailed1948 map of the northwest portion of the Finke River catchment. This map is unique in that it combined observations by itsauthors (a pastoralist and a mission worker) with information obtained from Aboriginal people. This map locates andnames a great many (46) springs and associated waterholes, a majority of which (31) were not found recorded in otherwritten or database records, but have been subsequently verified. Less confidence can be placed in modern topographicmaps, as many waterbodies called ‘spring’ were named by early European explorers for features now regarded aswaterbodies with no demonstrable groundwater discharge. Additional waterbodies may be known but unrecorded, assome landholders are reluctant to disclose where springs are located for fear of attracting campers and bushwalkers.
Some artificial ‘permanent’ aquatic habitats also occur in CA. A small number of dams are reputed to never dry out dueto natural springs below the water surface (Table 3). The most dramatic artificial wetland in CA was created byuncontrolled Great Artesian Basin (GAB) bore flow in the Simpson Desert (McDills No. 1 Bore) where a large lake (ca. 1 kmlong) existed for several decades due to uncontrolled discharge. Although it is now much smaller following control works itcontinues to provide an isolated aquatic habitat. Many elevated earth impoundments (‘‘turkey’s nests’’) are also kept‘permanently’ inundated by bore water, as are a few town and community sewage ponds and some mine tailings dams.
Interestingly, there are relatively few mound springs (Table 1) in CA, which are a well-known feature of the GAB in SouthAustralia and Queensland. Two factors probably inhibit mound spring formation in CA. First, most of the springs in CA thathave relatively high discharge volumes also have low conductivities, which is not conducive to the formation of mounds.Second, most springs are in drainage channels with flows that are sufficiently frequent and energetic to erode anyconcretions that could form a mound.
4. Biological attributes
Water longevity undoubtedly influences the aquatic biodiversity of CA. Permanent waterbodies serve as sourcepopulations for species to recolonise non-permanent sites, especially after prolonged droughts (Bunn et al., 2006). Somewaterbodies in CA also provide refugia for plants and animals that were probably more widely distributed during less aridpaleo-climates (Chippendale, 1963; Latz, 1996; Morton et al., 1995; Spencer, 1896; Tate, 1896a).

ARTICLE IN PRESS
Table 3Number of permanent or long-lasting CA water features based on groundwater discharge
No. Comment
Aquifer—localised discharge
Spring—permanent/semi-permanent surface flow 26
Spring—permanent/semi-permanent seepage 32
Spring—intermittent surface flow 20
Spring—minor seepage 4
Discharge—subsurface 3
Discharge (Spring)—unknown type 91
Waterhole sustained by spring 24 Includes springs/seepages adjacent to waterhole
and those hidden below the water surface
Dam sustained by spring 3 Artificial water source
Bore flow to ‘permanent’ artificial lake 2 Artificial water source
Total 205
Exposed watertable
Channel waterhole sustained by watertable 9 The surface water of these waterholes is
sustained by groundwater but in contrast to
springs, the discharge is not under pressure.
Eight on Mid-Finke River, 1 on Frew River
Total ‘point’ locations 214
Aquifer—broad-area discharge
Salt playas Very numerous
Saline swamps Some
Features were categorized using a variety of published and unpublished sources of information, including topographic, geological and hydrogeology
maps; hydrological and botanical databases; various hydrological and biological survey reports; as well as miscellaneous reports, journal papers and
historical maps of surface waters within the study area. Many of these water features are included on Fig. 3 (under GDEs).
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1405
4.1. Fauna
The aquatic fauna of CA is poorly known, although one of the earliest biological collections was made in the early 1860sduring the Stuart Expedition. A larger, more comprehensive biological survey was made during the Horn ScientificExpedition of 1894, but since then few broad-scale sampling efforts have been made. Recent collecting efforts in CA includesurveys of some permanent waterbodies in the West MacDonnell and George Gill Ranges (e.g., Davis, 1995, 1996; Daviset al., 1993), and an inventory of wetlands in the arid NT that included observations and limited collections of birds, fishesand invertebrates (Duguid et al., 2005). Additional, albeit disparate, collections have also been made through communityoutreach programs or other projects, but any data generated from these efforts are generally non-existent or unavailable.
Collections of macroinvertebrates and fishes from isolated waterbodies in CA suggest that an aquatic fauna is presentthat is distinct from other arid Australian waterbodies (Bayly, 2001; Blackburn, 1896; Davis, 1996; Davis et al., 1993; Duguidet al., 2005; Glover and Sim, 1978; Tate, 1896b; Unmack, 2001; Zietz, 1896). For example, the fishes of CA include at leastthree endemic species (Unmack, 1995), and may include additional, undescribed species (Duguid et al., 2005). A smallrockhole near Papunya, sampled after flooding, contained an aquatic invertebrate fauna that was distinct from similarhabitats in southwestern Australia and included undescribed species (Bayly, 2001). Davis et al. (1993) speculated that theisolation of waterbodies in CA, as well as their unique hydrological and physio-chemical conditions, has likely contributedto allopatric speciation. Invertebrates (e.g., the dragonfly, Hemicordulia flava) and fish species endemic to CA support thisassertion, as does the aquatic fauna of the George Gill Range (Davis et al., 1993), which was found to be distinct from otherpermanent waterbodies in Australia.
4.1.1. Fishes
The fish fauna of CA includes 14 native species of which three are endemic to CA (Wager and Unmack, 2000). Anadditional one to three species, introduced from other parts of Australia, may also be extant (Duguid et al., 2005). Naturalfish populations in CA are virtually restricted to river systems and associated in-channel waterholes that persist after short-lasting and infrequent flow events. Occasionally fish also occur in temporary swamps and lakes which are filled by largeriver flows, and fish (e.g., the golden perch, Macquaria spp.) native to other areas of Australia have been introduced into asmall number of artificial dams (Duguid et al., 2005).
The relationship between fishes and groundwater in CA mirrors the relationship between waterhole persistence andgroundwater, and it is thought that fish populations persist in arid river systems through two main mechanisms (Unmack,

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131406
2001). First, fish persist in permanent and semi-permanent riverine waterholes of which most, if not all, are partiallysustained by groundwater. Second, fish may persist in river systems without permanent waterholes if there is a geographicspread of long-lasting waterholes that never dry out at the same time, because even in a drought some part of the systemmay receive rainfall and runoff. In addition, at least one species, the spangled perch (Leiopotherapon unicolour), may persistin wet sand when surface waters have dried up (Peter Latz and Scott McConnell, pers. comm.), although it is not knownhow long they can persist in that state.
In the more arid southern part of CA we consider permanent waterholes to be the main mechanism by which fishpopulations persist. This is most notable in regards to the Finke River system, where nine fish species occur, including threeendemic to the system (Wager and Unmack, 2000). The Finke has been isolated from the rest of the Lake Eyre system forthousands of years, although it is possible that rare connections have occurred during mega-floods in the past hundreds ofyears (e.g., see Bourke, 1998; Duguid et al., 2005; Pickup, 1991). The Finke River system is one of the better studied in CA,and contains two of the largest springs known in the arid NT (Illara and Running Waters) (Duguid et al., 2005). These andother springs help sustain long-lasting waterholes in the Finke where fish populations persist.
The most species diverse river of CA is the Georgina, which includes 11 fish species (Wager and Unmack, 2000). In mostyears the Georgina River is connected to the Diamantina River of central Queensland, which sometimes flows into LakeEyre (Fig. 2). Therefore, the Georgina River at times connects to the rivers that drain much of the eastern portion of theAustralian arid zone. There are many large waterholes (several 41 km in length) in the main channels of the Georgina Riverwhere it dissects heavy clay plains, but these waterholes are probably not permanent (Randal, 1978). Unlike the Finke River,large springs and shaded gorges are not significant features of the Georgina River. However, groundwater contributes towaterhole permanence in the arid CA portion of the catchment (Duguid et al., 2005), including the Toko Range(6 fish species) (Fogarty et al., 1995) and the Sandover River system (3 fish species) (Gibson et al., 1989). These types ofspring-fed pools may be important drought refugia for fishes in the Georgina catchment. It is likely that fish persistence inthe system is facilitated by its extremely wide drainage area, its intermittent connection with other river systems, andrelatively predictable monsoon-influenced summer rainfalls in parts of the catchment.
Many CA river systems contain only a single fish species, the spangled perch, which tolerates a range of water qualities(e.g., high salinity, low dissolved oxygen) and has an exceptional ability to disperse into shallow and/or temporary waters(e.g., Duguid et al., 2005; Wager and Unmack, 2000). Fish assemblages in several other river systems are more diverse,notably the Frew River (5 species) and Whistleduck Creek (7 species). Both rivers have relatively small catchments with fewdocumented springs, but contain large waterholes (4100 m long; some 41 km) that are moderately deep (44 m).Although some of these waterholes are presumed to be permanent (e.g., Davidson, 1905), there is insufficientdocumentation to assess this, or the mechanisms by which fish persist in these drainages.
4.1.2. Macroinvertebrates
It is difficult to critically examine the aquatic macroinvertebrate biodiversity of CA waterbodies because of the lack oftaxonomic knowledge for most of the major faunal groups that are found in CA. Of the approximately 150 records of aquaticmacroinvertebrate occurrences from CA that were examined, 42% of the animals collected were identified to species, whilean additional 15% were marked as potentially new or undescribed species. Almost 40% of animals collected were notidentified to species, either because the taxonomic expertise was not available to make an accurate identification, orbecause the specimens examined did not match existing descriptions. For example, Davis et al. (1993) identified to genus83% of the invertebrates collected (95 taxa) from the George Gill Range, but only 45% to species, mainly because theanimals collected were taxonomically distinct from previously described species. It is difficult, therefore, to estimate whatpercentage of the CA fauna consists of rare and/or endemic species. Recent biological surveys have significantly increasedboth the total number of species as well as the number of new and/or endemic species in other parts of Australia, such asthe wheatbelt region of Western Australia (Pinder et al., 2004).
This lack of taxonomic resolution for CA aquatic macroinvertebrates is not confined to aquatic insects. Freshwatermolluscs, for example, are a diverse group of considerable ecological importance and are critically imperilled globally(Lydeard et al., 2004). They are poorly known both taxonomically and biologically, and few people can accurately identifythem to species (Lydeard et al., 2004; Ponder, 1997). None of the bivalves or gastropods collected by Davis et al. (1993) fromthe George Gill Range was identified to species, although specimens were examined by malacologists familiar with theAustralian freshwater molluscan fauna. Analogous to other aquatic taxonomic groups, systematic, ecologic and biologicstudies are ‘‘urgently needed’’ to facilitate the conservation of freshwater molluscs in Australia (Ponder and Walker, 2003).Additional and unique faunal groups also live only in subterranean aquatic habitats (e.g., stygofauna and animals found inhyporheic zones) (Boulton et al., 2003; Humphreys, 2001; Malard et al., 2002). Although these faunal groups are, in general,poorly known in CA (Boulton and Hancock, 2006; Hatton and Evans, 1998), recent studies suggest that a number of newand/or endemic species are present (Balke et al., 2004; Humphreys, 2006; Ponder and Walker, 2003; Watts andHumphreys, 2006). The conservation of aquatic macroinvertebrates in CA, many of which are short-range endemics, ishampered by this lack of taxonomic information (Harvey, 2002).
Total aquatic macroinvertebrate species richness (4130, from records examined by the authors) and average number ofspecies found per site (from 4 to 64) (from Davis, 1995; Davis et al., 1993) for CA waterbodies are similar to richnessestimates from waterbodies (temporary and permanent) in other regions of Australia (Boulton and Suter, 1986; Davis et al.,2006; Pinder et al., 2000), and are generally higher than estimates for Mojave Desert springs in the USA (Sada et al., 2005).

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1407
Davis et al. (1993) noted that species richness in the George Gill Range was comparatively high due to habitat diversity andto the adaptability of species to the unique conditions of CA waterbodies. However, species composition differed from otherregions in that no stoneflies (Plecoptera), water slaters (Isopoda) or scuds (Amphipoda) were recorded (Davis et al., 1993).Stoneflies may be absent in CA because water temperatures exceed their thermal tolerances, while the latter groups may beabsent because of lower inputs of riparian plant material. The composition of the CA aquatic macroinvertebrate fauna canbe considered unique to CA (Davis et al., 1993), especially as it differs markedly from the fauna of the specialised moundsprings of the GAB, which contain a number of endemic macroinvertebrate species (Ponder, 2004). To persist, the CAaquatic macroinvertebrate fauna must tolerate conditions associated with increasing aridity of the region, includingwarmer waters, episodic flow regimes and a lack of allocthonous organic inputs associated with sparser riparianvegetation.
The invertebrate fauna of Australian saline playas has been well studied (for a summary see Williams, 1998a) but fewdata are from CA. This is in contrast to the extensive investigations made of the hydrogeology (e.g., Jacobson, 1996) andbiology (e.g., Timms, 2007) of saline playas in other parts of Australia. It is possible that the invertebrate fauna of CA salineplayas is similar to those studied elsewhere in the Australian arid zone, such as Lake Eyre. However, future work is neededto test that assertion.
Although the majority of macroinvertebrate work in CA has focused on permanent waterbodies, it is not clear whichspecies are obligate groundwater species. Some groups of aquatic macroinvertebrates, including snails (gastropods), seedshrimps (ostracods), and beetle larvae (e.g., water penny, Sclerocyphon fuscus) may depend on long-lasting waterbodiesbecause they are confined to discontinuous habitats and have short-range dispersal abilities. However, relationshipsbetween waterbody permanence and aquatic macroinvertebrate persistence are not well known and may prove difficult toascertain. For example, in a long-term (over 20 years) study of 21 springs in the western US, spring invertebrateassemblages differed greatly between springs, and two taxa—flatworms (Turbellaria) and stoneflies—were consistentlyabsent from springs that were not permanent through drought years (Erman, 2002). Other taxa (e.g., molluscs), whilefound in permanent springs, were possibly absent from some springs due to other factors.
4.2. Flora
The vegetation associated with long-lasting water features in CA is varied, which is not surprising given that thesefeatures encompass a variety of habitats, from large waterholes to seepages. Individual plant species may be categorisedaccording to factors such as fidelity to particular habitats (e.g., aquatic, riparian, clay soil, etc.) and reliance on groundwater.A list of vascular wetland plants (Duguid et al., 2005) and rare plants (White et al., 2000) in CA, both of which drew on arich heritage of botanical study, provide a basis for assessing the relationship between plant species and permanent waterfeatures.
Most species of riparian and swamp plants in CA are widespread, common, and not obviously associated withgroundwater discharge or permanent water features. Riparian woodlands are mostly dominated by river red gums and to alesser extent, coolabahs (Eucalyptus coolabah and E. victrix) as well as other trees and shrubs. Although not welldocumented, soakages in CA rivers probably do not contain a particular suite of species.
Although it is not known to what extent most woody riparian species use groundwater in CA, two rare and endemic CAriparian species likely depend on groundwater. Red cabbage palms endemic to the Palm Valley area of CA are consideredclimatic relicts whose distribution is highly disjunct (41000 km separation) from other, closely related, cabbage palms(White et al., 2000). Without groundwater input they would have disappeared with the onset of aridity during thePleistocene Epoch (Wischusen et al., 2004). Similarly, the desert bottlebrush (Melaleuca faucicola), endemic to theMacDonnell Ranges Bioregion is mainly found in shady gullies and gorges where springs or seepages occur (Duguid et al.,2005).
Thirty-nine aquatic and semi-aquatic plant species are known to occur in CA (Duguid et al., 2005), but onlya few of these have a strong fidelity to sites which are known to be permanent. For example, the tassel sedge(Carex fascicularis) is restricted to two sheltered springs and is highly disjunct from other Australian populationsthat occur in more mesic climates. In contrast the club rush (Schoenoplectus littoralis) predominantly occurs in riverinewaterholes with groundwater discharge, but also sporadically occurs in temporary, non-permanent waterbodies.Most other aquatic and semi-aquatic species are well adapted to cycles of wetting and drying and do not depend onpermanent waters. The common reed (Phragmites australis) is an exception, in that it tolerates periodic drying butpredominantly occurs in areas of known groundwater discharge. Possibly its root system requires perpetually damp soil inorder to survive and resprout when areas are re-flooded. Other aquatic species, including bullrushes (Typha spp.) andrushes (Juncus spp.), are typical of long-term pools but also occur in more temporary waters, and are not reliable indicatorsof groundwater.
Some herbaceous species require permanently saturated soils that are created by springs or seepages, but these plantsmay not require inundation. The species in this group are all rare in CA, disjunct from more abundant populations in moremesic areas, and may be relics of wetter palaeo-climates (Chippendale, 1963; Latz, 1996). For example, 11 fern species areknown from just five CA ranges, in places where springs/seepages occur in relatively shaded topographic positions. Four ofthese relict fern species occur in environments that are effectively small swamps, subject to brief inundations during rain

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131408
events, while the other seven typically occur on rock ledges or cracks where there is minimal soil present, but freegroundwater is provided (White et al., 2000).
The flora and fauna of saline playas and saline springs, as was generally recognised by early explorers, is markedlydifferent from other water features (Duguid et al., 2005; Williams, 1998b; Williams and Kokkinn, 1988). The majority ofsaline playas are so salty that their lake beds are devoid of vascular plants, with the exception of the tuberous tassel (Ruppia
tuberosa), an aquatic plant which often occurs when playas are filled. Most saline playas, however, are fringed by suites ofsalt-tolerant plants. While not all these species are restricted to groundwater discharge features, many are, and the vastmajority of saline habitats in CA are associated with groundwater discharge.
4.3. Ecological considerations
It is increasingly clear that isolated arid wetlands are habitats for rare and endemic species (Erman, 1996; Erman andNagano, 1992). It has also been suggested that the aquatic fauna of at least some CA waterbodies has remained relativelyunchanged since European settlement (Davis, 1996), particularly waterbodies located in inaccessible (i.e., to humans, feralanimals) upper sections of gorges. These waterbodies also often lack fish, both native and non-native, which in some casescan exert significant predation pressure on native invertebrate species. These observations are significant, in that in otherparts of Australia the unsustainable extraction of artesian water and other impacts have probably led to the extinction ofmany aquatic species, although these extinctions have not been well documented (Lydeard et al., 2004; Ponder and Walker,2003). If permanent waterbodies in CA are ecologically intact and contain a diverse and endemic fauna, then thesewaterbodies are probably ecologically and evolutionarily important (Davis et al., 1993). Whether many other waterbodiesin CA have remained relatively intact will not be known until additional biological surveys are conducted.
Semi-permanent and permanent wetlands serve as source populations for the re-colonisation of more temporary,nearby waterbodies, and provide refugia for rare or relict species during prolonged droughts. For example, permanentwaterbodies serve as source populations from which fishes recolonise after drought (Glover, 1982; Spencer, 1896; Unmack,2001). Insects capable of active dispersal, such as water beetles (hydrophilids), diving beetles (dytiscids), water boatmen(corixids), and backswimmers (notonectids), emigrate from temporary wetlands to more permanent waters whentemporary pools dry up (Bayly, 1997). Insects and other invertebrates that can not actively disperse may perish whentemporary waterholes dry out (Stanley et al., 1994), but often have life history strategies (e.g., eggs that are resistant todesiccation, aestivation) that allow them or their progeny to persist in temporary wetlands.
Because permanent wetlands serve as refugia, they tend to have higher species diversity than more ephemeralwaterholes. In isolated wetlands in southern New Hampshire, both amphibian and macroinvertebrate assemblages werepositively correlated with hydroperiod length and wetland size (Baber et al., 2004). In CA Davis et al. (1993) found a similarpattern, in that less permanent waterbodies contained fewer species than waterbodies with longer hydroperiods. The largeexpanse of area between some temporary and permanent waterbodies in CA may preclude the dispersal of animalsbetween the two habitat types. For example, an isolated temporary waterbody sampled by Bayly (2001) did not containinsects that normally actively retreat to more permanent waterbodies during dry periods. These examples suggest that theconservation of permanent waterbodies could protect more native biological diversity than conserving temporarywaterbodies (Baber et al., 2004), in part because the permanence and constancy of individual waterbodies have beenshown to be positively correlated with species richness (e.g., Erman and Erman, 1995).
Although empirical data are lacking for most taxa, isolated waterbodies in CA probably contain species that are uniquedue to long-term hydrologic isolation and the presence of relic and endemic species, such as at Palm Valley in the FinkeGorge National Park. Chippendale (1963) regarded some of the CA ranges as ‘relict’ areas, ‘‘where plants of a higher rainfallperiod have survived.’’ Davis et al. (1993) and Davis (1997) reached a similar conclusion regarding the aquatic invertebratefauna of the George Gill and West MacDonnell Ranges, and suggested that the presence of endemics at these sites wasevidence that long-term isolation had facilitated speciation.
5. Activities that can potentially affect CA waterbodies
Alterations to inland waters can impact their biota disproportionately to other ecosystems, especially if thosewaterbodies contain localised, endemic species (Hutchings and Ponder, 1999). The aquatic flora and fauna of long-lastingwaterbodies in CA include a number of species that are either endemic or relic. In both cases, the ability of these plantsand/or animals to disperse into new areas is limited. Therefore activities that negatively impact CA waterbodies mayfacilitate species declines, more so than in other areas, by the presence of an inordinate number of taxa that have short-range dispersal abilities, such as fishes, hydrobiid snails and some aquatic insect larvae (e.g., water pennies).
The potential threats to wetlands in the arid NT were reviewed by Duguid et al. (2005). Activities that can potentiallyimpact isolated waterbodies in CA include drinking, fouling and grazing by livestock and feral animals, road construction,non-native plants, groundwater extraction, pollution (e.g., garbage, pesticides), tourism, mining, and long-term changes inweather patterns, such as those predicted to occur with global climate change. These impacts, either singly or jointly, canaffect temperature regimes, sedimentation processes, riparian vegetation, chemical concentrations, solar radiation (Ermanand Erman, 1995) and evaporation rates, which in turn can potentially make waterbodies unsuitable for animals and/or

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1409
human usage. The following effects have been documented when wetlands are degraded (modified from the US EPA, 2002):(1) the number of native species declines, (2) the number of specialised taxa or guilds declines, (3) the percentage oftrophic and habitat specialists declines, (4) the total number of individual animals declines, (5) the percentage of largeindividuals, as well as size classes, declines, (6) the percentage of non-native species increases, (7) the percentage ofpresumed tolerant species increases, (8) the percentage of trophic and habitat generalists increases, and (9) the percentageof individual animals with anomalies or birth defects increases. Some of the major activities that can impact CAwaterbodies are discussed below.
5.1. Non-native plants
The spread of non-native plant species may be one of the greatest threats to the biota of wetlands in CA and at least 44significant weed species have been reported from the region (Duguid et al., 2005). Non-native plants can replace nativevegetation, which in turn can impact native animals that rely on native plants for food and/or habitat. For example, in manywetlands buffel grass (Cenchrus ciliaris) and couch grass (Cynodon dactylon) dominate understoreys (Duguid et al., 2005)and have displaced yalka (Cyperus bulbosus) at some sites, a food source for native animals, including the endangered bilby(Macrotis lagotis) (Peter Latz., pers. comm.). Couch grass may also have displaced the endangered native dwarf desert spike-rush (Eleocharis papillosa) in some areas (e.g., see Duguid et al., 2005). Non-native plants can also impact ecosystem-levelprocesses by changing soil salinity (e.g., the athel pine (Tamarix aphylla) in the Finke River drainage), soil nutrientcomposition, evaporation rates, watertable levels, groundwater recharge rates, and the geomorphology of waterbodiesthrough erosional processes (Gordon, 1998; Griffin et al., 1989; Maitre, 2004; Zhang and Schilling, 2006).
5.2. Feral animals and domestic stock
Introduced herbivores that can potentially impact CA waterbodies include cattle, camels, horses and donkeys, andhistorically included rabbits, sheep and goats. Rabbits especially, because of their historical high numbers in CA, can havean impact on isolated wetlands through intense grazing on riparian vegetation, but their numbers in CA have dropped morethan 80% since the introduction of rabbit calicivirus disease in the mid-1990s (Edwards et al., 2002).
Introduced herbivores can impact waterbodies by trampling, grazing, fouling, muddying, destabilising, and drinking.The environmental impacts of feral animals on waterbodies in other areas of Australia have been well documented(for a review see Norris and Low, 2005), and in CA overgrazing by domestic and feral animals was considered a majorconservation threat (Storrs and Finlayson, 1997). Camels, for example, are widespread and abundant (at least 300,000animals) throughout arid Australia, their population is predicted to double approximately every 8 years (Edwards et al.,2004a, b), and their impacts can be considerable (Norris and Low, 2005). In South Australia, for example, camels dranksome waterbodies dry, occasionally fell into and died in waterholes, and caused eutrophication and siltation, all of whichmade these waterbodies unusable by native birds and mammals (Copley et al., 2003). In CA feral camels have damagedwaterbodies that were previously inaccessible to most feral or domestic stock, including culturally significant springs nearUluru (J. Brim Box, pers. obs.).
Domestic stock, mainly cattle, can have similar impacts, and in some situations are not in managed herds and areeffectively feral. The number of CA waterbodies that have been fenced or are slated to be fenced to exclude feral ungulatesand domestic stock is generally low, but is increasing. However, data are lacking on the effectiveness of these projects inprotecting or restoring ecosystem health. It is particularly difficult and expensive to protect waterbodies from camels,which are not stopped by conventional stock fences.
5.3. Water withdrawals and the creation of artificial wetlands
In CA present-day human activities are sustained by groundwater extractions and the damming of surface waters.Pastoral expansion in inland Australia in the 20th century was made possible by the use of artificial water sources such asbores, dams, wells and troughs (Landsberg et al., 1997). In areas of inland Australia where productive rangelands occur, theaverage distance between artificial water sources is generally less than 10 km (James et al., 1999).
Artificial water sources have, in some cases, created wetland environments that are similar, at least in some regards(e.g., vegetation types), to natural waterholes. Three direct effects of artificial water sources were noted by James et al.(1999); the creation of wetlands that supported some native species of plants and animals; the range expansion andincrease in abundance of native animals that required a reliable supply of drinking water; and an increase in the breedingranges of some species of terrestrial invertebrates. However, the following negative impacts were also noted, mainlybecause most artificial water points were also used by feral and domestic grazing animals as well as non-native predatorssuch as cats and foxes (James et al., 1999); an increase in the predation of medium-sized native mammals that may havecontributed to subsequent extinctions; the extirpation of ground-dwelling bird species and range reduction of other birdspecies due to habitat degradation; and changes in the distribution and abundance of some groups of terrestrial insects.
In other parts of Australia, impacts to the aquatic fauna from groundwater extractions have been well documented(e.g., Humphreys and Harvey, 2001; Ponder, 1997). For example, in the GAB the unsustainable extraction of artesian water

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131410
has probably led to local extinctions (Lydeard et al., 2004), and it is possible that stock trampling, fouling, and grazingexacerbated these extinction events. In CA, some springs have been used for stock or domestic water, either by piping wateraway or by cattle and horses directly using the spring and associated pools. Data on the impacts of groundwater extractionsin CA are limited, and in most cases major groundwater extractions occur only near larger towns and communities and in afew irrigation areas.
5.4. Climate change
The projected impacts of climate change in Australia and New Zealand, and apropos to CA, include a more El Nino-likeprecipitation pattern and an increase in heavy precipitation events (IPCC, 2002). This may result in increased groundwaterrecharge, a possible change in statistical frequency of stream flows, and longer dry periods between flows. The specificimpacts that anthropogenic climate change will have on CA waterbodies can only be speculated on. Predicting or managingthe consequences of climate change on the biota of CA waterbodies is not yet possible, but developing a betterunderstanding of the factors that influence long-term biodiversity in these aquatic systems will be a cornerstone to such anendeavour.
If human-mediated climate change does occur, changes in hydrological conditions could result in a net decreasein species diversity, as has occurred in other isolated wetlands (Erman and Erman, 1995). Species of aquaticmacroinvertebrates in CA have survived a drying climate for about 18,000 years, and would potentially need to adapt tofurther fluctuations in water temperatures, episodic flow regimes, and changes in allocthonous organic inputs associatedwith changes in riparian vegetation.
Species that may be more at risk from global climate change include those with limited ranges, narrow habitatrequirements, and poor dispersal abilities (IPCC, 2002). Some plants and animals found in CA waterbodies share thosecharacteristics, and these taxa are probably at an increased risk of extirpation if significant climate change occurs,especially those species endemic to CA. The remoteness of most wetlands in CA may limit the migration of new speciesmore suitable to the new conditions into these areas, resulting in a net loss of biodiversity.
6. Research needs
Inventories of the aquatic fauna of CA are limited, and many areas of potentially high biological significance (e.g., springsof the Dulcie and Chewings Ranges) have not been extensively surveyed. Surveys of biological diversity provide a baselineby which future surveys can be compared, and can be used to document changes in habitat quality and availability, as wellas species richness and diversity. For example, Davis (1996) compared modern records of the aquatic fauna of sites in CA tospecimens collected during the Horn expedition in 1894. The similarity between records for some taxonomic groupssuggests the aquatic fauna of these waterbodies has remained relatively unchanged over a hundred-year period, especiallyat those sites where human and introduced animal impacts were minimal. Inventories where specimens are collected forgenetic analysis are especially needed, as molecular tools can provide a nexus to examine gene flow and substantiate thelevel of allopatric speciation that is suspected to have occurred in CA. In addition, improved ecological understandingrequires improved knowledge of waterbody hydrology, including groundwater discharge and recharge, patterns ofinundation and drying, and changes in water depth and chemistry.
Whether an isolated wetland is protected is often determined by whether that waterbody provides a service(e.g., supports tourism or recreational uses, serves as a water source for livestock, or harbours a species of interest) (Erman,2002). Wetlands that are ecologically and/or culturally important may not be protected if the services they provide are notdeemed important or of value, or if their ecological or cultural values are unknown or unrecognised. In addition, thebiological and physical characteristics of many arid wetlands are not known because they have not been adequatelysurveyed. For example and analogous to CA, thousands of springs occur throughout the western US, but many have notbeen inventoried because they are small and difficult to find (Sada and Pohlmann, 2006). Only recently have studiesconfirmed the significant endemism that exists in dryland wetlands in other regions of Australia and internationally(Hershler et al., 2002; Pinder et al., 2004; Williams, 1999). Baseline surveys of the biota of isolated CA waterbodies are alsoneeded, especially from those ranges where permanent waterbodies are known to occur, and in light of the potential forglobal climate change and human-mediated impacts.
Acknowledgements
The Northern Territory Government (NTG) Department of Natural Resources, Environment and the Arts providedfunding for this work. Many years of data collection by NTG water and biological survey staff contributed to the underlyingdata and various current staff provided encouragement and advice. We thank Graham Ride for providing a copy of Bowmanand Scherer’s 1948 map, and Kathy McConnell for digitally capturing site information from it. Blair Trewin providedguidance on how to interpret climate change data from CA. We thank Glenn Edwards, Peter Barker, and three anonymousreviewers for helpful comments made on earlier versions of this paper.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1411
References
Allan, R., Lindesay, J., 1998. Past climates of Australasia. In: Hobbs, J.E., Lindesay, J.A., Bridgman, H.A. (Eds.), Climates of the Southern Continents, Present,Past and Future. Wiley, Chichester, pp. 208–247.
Baber, M.J., Fleishman, E., Babbitt, K.J., Tarr, T.L., 2004. The relationship between wetland hydroperiod and nestedness patterns in assemblages of larvalamphibians and predatory macroinvertebrates. Oikos 107, 16–27.
Balke, M., Watts, C.H.S., Cooper, S.J.B., Humphreys, W.F., Vogler, A.P., 2004. A highly modified stygobitic diving beetle of the genus Copelatus (Coleoptera,Dytiscidae): taxonomy and cladistic analysis based on mtDNA sequences. Systematic Entomology 29, 59–67.
Bayly, I.A.E., 1997. Invertebrates of temporary waters in gnammas on granite outcrops in Western Australia. Journal of the Royal Society of WesternAustralia 80, 167–172.
Bayly, I.A.E., 1999. Review of how indigenous people managed for water in desert regions of Australia. Journal of the Royal Society of Western Australia 82,17–25.
Bayly, I.A.E., 2001. Invertebrate occurrence and succession after episodic flooding of a central Australian rock-hole. Journal of the Royal Society of WesternAustralia 84, 29–32.
Bayly, I.A.E., Williams, W.D., 1973. Inland Waters and their Ecology. Longman, Melbourne.Blackburn, T., 1896. Coleoptera. In: Spencer, W.B. (Ed.), Report on the Work of the Horn Scientific Expedition to Central Australia. Melville, Mullen and
Slade, Melbourne, pp. 254–261.Boulton, A.J., Hancock, P.J., 2006. Rivers as groundwater-dependent ecosystems: a review of degrees of dependency, riverine processes and management
implications. Australian Journal of Botany 54, 1–12.Boulton, A.J., Suter, P.J., 1986. Ecology of temporary streams—an Australian perspective. In: De Deckker, P., Williams, W.D. (Eds.), Limnology in Australia.
CSIRO, Melbourne, pp. 313–327.Boulton, A.J., Humphreys, W.F., Eberhard, S.M., 2003. Imperilled subsurface waters in Australia: biodiversity, threatening processes and conservation.
Aquatic Ecosystem Health and Management 61, 41–54.Bourke, M.C., 1998. Fluvial geomorphology and paleofloods in arid central Australia (Ph.D. Thesis abstract). Quaternary Australasia 16, 31–32.Bowler, J.M., 1986. Spatial variability and hydrologic evolution of Australian lake basins: analogue for Pleistocene hydrologic change and evaporite
formation. Palaeogeography, Palaeoclimatology, Palaeoecology 54, 21–41.Bunn, S.E., Thoms, M.C., Hamilton, S.K., Capon, S.J., 2006. Flow variability in dryland rivers: boom, bust and the bits in between. River Research and
Applications 22, 179–186.Bureau of Meteorology, 2007. Trend Maps—Australian Climate Variability and Change /http://www.bom.gov.au/cgi-bin/silo/reg/cli_chg/trendmaps.cgiS.Charnley, W., 1954. Water in the desert. Walkabout July 1, 1954, 29–30.Chewings, C., 1886. The sources of the Finke River. The Adelaide Observer May 29, 1886 May 29, 1886, 41.Chippendale, G.M., 1963. The relic nature of some central Australian plants. Transactions of the Royal Society of Australia 86, 31–34.Clifton, R., Evans, C., 2001. Environmental water requirements of groundwater dependent ecosystems. Environmental Flows Initiative Technical Report
No. 2. Commonwealth of Australia, Canberra.Copley, P.B., Baker, L.M., Nesbitt, B.J., Foulkes, J.N., 2003. Mammals. In: Robinson, A.C., Copley, P.B., Canty, P.D., Baker, L.M., Nesbitt, B.J. (Eds.),
A Biological Survey of the Anangu Pitjantjatjara Lands, South Australia, 1991–2001. Department for Environment and Heritage, Adelaide,pp. 197–242.
Cresswell, R.G., Jacobson, G., Wischusen, J., Fifield, L.K., 1999a. Ancient groundwaters in the Amadeus Basin, Central Australia: evidence from the radio-isotope 36Cl. Journal of Hydrology 223, 212–220.
Cresswell, R., Wischusen, J., Jacobson, G., Fifield, K., 1999b. Assessment of recharge to groundwater systems in the arid southwestern part of the NorthernTerritory, Australia, using chlorine-36. Hydrogeology Journal 7, 393–404.
Davidson, A.A., 1905. Journal of Explorations in Central Australia, 1898–1900. Government Printer, Adelaide.Davis, J.A., 1995. The aquatic ecosystems of the West MacDonnell Ranges, Northern Territory, with particular reference to the aquatic invertebrate fauna.
Report to the Parks and Wildlife Commission of the Northern Territory, Alice Springs.Davis, J.A., 1996. Aquatic ecosystems in central Australia: comparison of recent records of fishes and invertebrates with those of the Horn Expedition. In:
Morton, S.R., Mulvaney, D.J. (Eds.), Exploring Central Australia: Society, the Environment and the 1894 Horn Expedition. Surrey Beatty and Sons,Chipping Norton, pp. 282–286.
Davis, J.A., 1997. Conservation of aquatic invertebrate communities in central Australia. Memoirs of the Museum of Victoria 56, 491–503.Davis, J.A., Harrington, S.A., Friend, J.A., 1993. Invertebrate communities of relict streams in the arid zone: the George Gill Range, Central Australia.
Australian Journal of Marine and Freshwater Research 44, 483–505.Davis, J., Horwitz, P., Norris, R., Chessman, B., McGuire, M., Sommer, B., 2006. Are river bioassessment methods using macroinvertebrates applicable to
wetlands? Hydrobiologia 572, 115–128.Duguid, A., Barnetson, J., Clifford, B., Pavey, C., Albrecht, D., Risler, J., McNellie, M., 2005. Wetlands in the arid Northern Territory, vol. 1. A Report to the
Australian Government Department of the Environment and Heritage on the inventory and significance of wetlands in the arid NT. Northern TerritoryGovernment Department of Natural Resources, Environment and the Arts, Alice Springs.
Edwards, G.P., Dobbie, P.W., Berman, D.McK., 2002. Population trends in European rabbits and other wildlife of central Australia in the wake of rabbithaemorrhagic disease. Wildlife Research 29, 557–565.
Edwards, G.P., Pople, A., Saalfeld, K., Caley, P., 2004a. Feral mammals in Australia’s rangelands: future threats and the role of monitoring programmes inmanagement strategies. Austral Ecology 29, 40–50.
Edwards, G.P., Saalfield, K., Clifford, B., 2004b. Population trend of camels in the Northern Territory, Australia. Wildlife Research 31, 509–511.Erman, N.A., 1996. Status of aquatic invertebrates. In: Sierra Nevada Ecosystem Project: Final report to Congress, vol. II. Assessments and Scientific Basis for
Management Options. University of California, Davis, Centers for Water and Wildland Resources, Davis, pp. 987–1008 (Chapter 35).Erman, N.A., 2002. Lessons from a long-term study of springs and spring invertebrates (Sierra Nevada, California, USA) and implications for conservation
and management. In: Conference Proceedings. Spring-fed Wetlands: Important Scientific and Cultural Resources of the Intermountain Region. DesertResearch Institute, Reno also at /http://www.wetlands.dri.eduS.
Erman, N. A., Erman, D.C., 1995. Spring permanence, species richness and the role of drought. In: Ferrington Jr., L.C. (Ed.), Biodiversity of Aquatic Insectsand Other Invertebrates in Springs. Journal of the Kansas Entomological Society 68 (Suppl., Special Publication No. 1), 50–64.
Erman, N.A., Nagano, C.D., 1992. A review of the California caddisflies (Trichoptera) listed as candidate species on the 1989 federal ‘‘Endangered andThreatened Wildlife and Plants; Animal Notice of Review’’. California Fish and Game 78, 45–56.
Fisher, C.D., Lingren, E., Dawson, W.R., 1972. Drinking patterns and behavior of Australian desert birds in relation to their ecology and abundance. Condor74, 111–136.
Fogarty, M., Fuller, D., McAdam, D., O’Neil, P.C.W., Walsh, S., 1995. Ranger training/flora and fauna survey of the Toko Ranges, Northern Territory 8–20October 1995. Parks and Wildlife Commission of the Northern Territory, Alice Springs.
Freeze, R.A., Cherry, J.A., 1979. Groundwater. Prentice-Hall, Englewood Cliffs, NJ.Gibson, D.F., Kube, P.D., Ginns, A.J., Cole, J.R., Parsons, D.J., 1989. A resource survey of the Dulcie Ranges, Northern Territory. Report to the Conservation
Commission of the Northern Territory, Alice Springs.Giles, E., 1889. Australia Twice Traversed. Sampson, Low, Marston, Searle and Rivington, London.Gillen, R.S. (Ed.), 1995. F.J. Gillen’s First Diary 1875. Wakefield Press, Kent Town.Gillis, A.M., 1991. Should cows chew cheatgrass on common lands? Bioscience 41, 668–675.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–14131412
Glover, C.J.M., 1982. Adaptations of fishes in arid Australia. In: Barker, W.R., Greenslade, P.J.M. (Eds.), Evolution of the Flora and Fauna of Arid Australia.Peacock Publications, Adelaide, pp. 241–246.
Glover, C.J.M., Sim, T.C., 1978. A survey of Australian ichthyology. The Australian Zoologist 19, 245–256.Gordon, D.R., 1998. Effects of invasive, non-indigenous plant species on ecosystem processes: lessons from Florida. Ecological Applications 8, 975–989.Gosse, W.C., 1874. W.C. Gosse’s Explorations, 1873. Libraries Board of South Australia, Adelaide.Gould, R.A., 1969a. Subsistence behaviour among the Western Desert Aborigines of Australia. Oceania 39, 253–274.Gould, R.A., 1969b. Yiwara: Foragers of the Australian Desert. Collins, Melbourne.Griffin, G.F., Stafford Smith, D.M., Morton, S.R., Allen, G.E., Masters, K.A., Preece, N., 1989. Status and implications of the invasion of tamarisk (Tamarisk
aphylla) on the Finke River, Northern Territory, Australia. Journal of Environmental Management 29, 297–315.Gubanich, A.A., Panik, H.R., 1986. Avian use of waterholes in Pinyon-Juniper. In: Everett, R.L. (Ed.), Proceedings of the Pinyon-Juniper Conference, USDA
Forest Service General Technical Report INT-215, pp. 534–540.Hardman, W. (Ed.), 1975. The Journals of John McDouall Stuart. Libraries Board of South Australia, Adelaide.Harrington, G.A., 1999. Recharge mechanisms and chemical evolution in an arid groundwater system, Central Australia. Ph.D. Thesis, School of Chemistry,
Physics and Earth Sciences, Flinders University, Adelaide.Harvey, M.S., 2002. Short-range endemism among the Australian fauna: some examples from non-marine environments. Invertebrate Systematics 16,
555–570.Hatton, T., Evans, R., 1998. Dependence of ecosystems on groundwater and its significance to Australia. Occasional Paper No. 12/98, Land and Water
Resources Research and Development Corporation, Canberra.Hercus, L., Clarke, P., 1986. Nine Simpson Desert wells. Archaeology in Oceania 21, 51–62.Hershler, R., Madsen, D.B., Currey, D. (Eds.), 2002. Great Basin aquatic systems history. Smithsonian Contributions to the Earth Sciences No. 33.
Washington, D.C.Humphreys, W.F., 2001. Groundwater calcrete aquifers in the Australian arid zone: the context to an unfolding plethora of stygal biodiversity. In:
Humphreys, W.F., Harvey, M.S. (Eds.), Subterranean Biology in Australia 2000. Records of the Western Australian Museum 64, pp. 63–83.Humphreys, W.F., 2006. Aquifers: the ultimate groundwater dependent ecosystems. In: Eamus, D. (Ed.), Special Edition on Groundwater Dependent
Ecosystems. Australian Journal of Botany, vol. 54, pp. 115–132.Humphreys, W.F., Harvey, M.S. (Eds.), 2001. Subterranean Biology in Australia 2000. Records of the Western Australian Museum 64, pp. 1–242.Hutchings, P.A., Ponder, W.F., 1999. Workshop: criteria for assessing and conserving threatened invertebrates. In: Ponder, W.F., Lunney, D. (Eds.),
The other 99 percent. The Conservation and Biodiversity of Invertebrates. Transactions of the Royal Zoological Society of NSW, Mosman,pp. 264–315.
IPCC, 2002. Climate change and biodiversity. In: Gitay, H., Suarez, A., Watson, R. Dokken, D. (Eds.), IPPC Technical Paper V. WMO/UNEP Publication also at/http://www.ipcc.ch/pdf/technical-papers/climate-changes-biodiversity-en.pdfS.
Jacobson, G., 1996. The interrelationship of hydrogeology and landform in central Australia. In: Morton, S.R., Mulvaney, D.J. (Eds.), Exploring CentralAustralia: Society, the Environment and the 1894 Horn Expedition. Surrey Beatty and Sons, Chipping Norton, pp. 249–266.
Jacobson, G., Lau, G.C., McDonald P.S., Jankowski, J., 1989. Hydrogeology and groundwater resources of the Lake Amadeus and Ayers Rock region, NorthernTerritory. Australian Bureau of Mining and Mineral Resources, Geology and Geophysics Bulletin, p. 230.
James, C.D., Landsberg, J., Morton, S.R., 1999. Provision of watering points in the Australian arid zone: a review of effects on biota. Journal of AridEnvironments 41, 87–121.
Jones, N.O., Quinlan, T., 1962. An outline of the water resources of the Alice Springs area. In: Perry, R.A. (Ed.), General Report on Lands of the Alice SpringsArea, Northern Territory, 1956–7. Land Research Series No. 6. CSIRO, Melbourne.
Kavanagh, B.L., 1984. Survival Water in Australia’s Arid Lands. Australian National University, Canberra.Kimber, R.G., 1985. Resource use and management in central Australia. Australian Aboriginal Studies 1984, 12–23.Kimber, R.G., 1990. Hunter gatherer demography: the recent past in central Australia. In: Meehan, B., White, N. (Eds.), Hunter–Gatherer Demography Past
and Present. Oceania Monograph 39. University of Sydney, Sydney, pp. 160–170.Kimber, R.G., 2001. Australian Aboriginals’ perceptions of their desert homelands, Parts 1 and 2. Arid Lands Newsletter, The Deserts in Literature II, No. 50,
November–December 2001.Landsberg, J., James, C.D., Morton, S.R., Hobbs, T., Stol, J., Drew, A., Tongway, H., 1997. The effects of artificial sources of water on rangeland biodiversity.
Final Report to the Biodiversity Convention and Strategy Section of the Biodiversity Group, Environment Australia (Biodiversity Technical Paper No. 3),Canberra.
Latz, P.K., 1996. Knowledge of relict plants in central Australia: a measure of botanical research during the last 100 years. In: Morton, S.R., Mulvaney, D.J.(Eds.), Exploring Central Australia: Society, the Environment and the 1894 Horn Expedition. Surrey Beatty and Sons, Chipping Norton, pp. 225–229.
Lloyd, J.W., Jacobson, G., 1987. The hydrogeology of the Amadeus Basin, Central Australia. Journal of Hydrology 93, 1–21.Lydeard, C., Cowie, R.H., Ponder, W.F., Bogan, A.E., Bouchet, C.S.A., Cummings, K.S., Frest, T.J., Gargominyh, O., Herbert, D.G., Hershler, R., Perez, K.E., Roth, B.,
Seddon, M., Strong, E.E., Thompson, F.G., 2004. The global decline in nonmarine molluscs. BioScience 54, 321–330.Magarey, A.T., 1895. Australian Aborigines’ water-quest. Proceedings of the Royal Geographical Society of Australasia (South Australian Branch) 3, 67–82.Maitre, D.C.le, 2004. Predicting invasive species impacts on hydrological processes: the consequences of plant physiology for landscape processes. Weed
Technology 18, 1408–1410.Malard, F., Tockner, K., Dole-Olivier, M.J., Ward, J.V., 2002. A landscape perspective of surface–subsurface hydrological exchanges in river corridors.
Freshwater Biology 47, 621–640.Morton, S.R., Mulvaney, D.J. (Eds.), 1996. Exploring Central Australia: Society, The Environment and the 1894 Horn Expedition. Surrey Beatty and Sons,
Chipping Norton, Sydney.Morton, S.R., Short, J., Barker, R.D., 1995. Refugia for Biological Diversity in Arid and Semi-arid Australia. Biodiversity Series, Paper No. 4. Biodiversity Unit,
Department of Environment, Sport and Territories, Canberra.Murray, W.R., 1907. Record of prospecting operations in the country South-West from Barrow Creek and the Davenport ranges—1906. In: South Australia
Parliamentary Proceedings, Adelaide.Myers, M.J., Resh, V.H., 1999. Spring-formed wetlands of the arid west. Islands of aquatic invertebrate biodiversity. In: Batzer, D.P., Radar, R.B., Wissiner,
S.A. (Eds.), Invertebrates in Freshwater Wetlands of North America: Ecology and Management. Wiley, New York, pp. 811–828.Noble, J.C., Kimber, R.G., 1999. On the ethno-ecology of mallee root-water. Aboriginal History 21, 170–202.Norris, A., Low, T., 2005. Review of the Management of Feral Animals and Their Impact on Biodiversity in the Rangelands: A Resource to Aid NRM Planning.
Pest Animal Control CRC Report 2005. Pest Animal Control CRC, Canberra.O’Grady, A.P., Cook, P.G., Duguid, A., Knapton, A., Fass, T., 2007. Ti Tree Basin, NT—groundwater dependence of terrestrial vegetation. In: Howe, P. (Ed.),
Project REM1—A Framework for Assessing Environmental Water Requirements for Groundwater Dependent Ecosystem. Report 2. Field Studies. Landand Water Australia, Braddon, pp. 12–49.
Pickup, G., 1991. Event frequency and landscape stability on the floodplain systems of arid Central Australia. Quaternary Science Reviews 10, 463–473.Pinder, A.M., Halse, S.A., Shiel, R.J., McRae, J.M., 2000. Granite outcrop pools in south-western Australia: foci of diversification and refugia for aquatic
invertebrates. Journal of the Royal Society of Western Australia 83, 149–161.Pinder, A.M., Halse, S.A., McRae, J.M., Shiel, R.J., 2004. Aquatic invertebrate assemblages of wetlands and rivers in the wheatbelt region of Western
Australia. Records of the Western Australian Museum Supplement No. 67, pp. 7–37.Ponder, W.F., 1997. Conservation status, threats and habitat requirements of Australian terrestrial and freshwater Mollusca. Memoirs of the Museum of
Victoria 56, 421–430.

ARTICLE IN PRESS
J.B. Box et al. / Journal of Arid Environments 72 (2008) 1395–1413 1413
Ponder, W.F., 2004. Endemic aquatic macroinvertebrates of artesian springs of the Great Artesian Basin—progress and future directions. Records of theSouth Australian Museum Monograph Series 7, 101–110.
Ponder, W.F., Walker, K.F., 2003. From mound springs to mighty rivers: the conservation status of freshwater molluscs in Australia. Aquatic EcosystemHealth and Management 6, 5–18.
Randal, M.A., 1978. Hydrogeology of the South-Eastern Georgina Basin and environs, Queensland and Northern Territory. Geological Survey of QueenslandPublication No. 366, Geological Survey of Queensland, Sydney.
Sada, D.W., Pohlmann, K.F., 2006. Mojave Inventory and Monitoring Network Spring Survey protocols: Level I and Level II. Draft Report to the US NationalPark Service, Reno.
Sada, D.W., Sharpe, S.E. (Eds.), 2004. Conference Proceedings. Spring-fed Wetlands: Important Scientific and Cultural Resources of the IntermountainRegion. Desert Research Institute, Reno /http://www.wetlands.dri.eduS.
Sada, D.W., Vinyard, G.L., 2002. Anthropogenic changes in historical biogeography of Great Basin aquatic biota. In: Hershler, R., Madsen, D.B., Currey, D.(Eds.), Great Basin Aquatic Systems History. Smithsonian Contributions to the Earth Sciences No. 33, pp. 277–293.
Sada, D.W., Fleishman, E., Murphy, D.D., 2005. Associations among spring-dependent aquatic assemblages and environmental and land use gradients in aMojave Desert mountain range. Diversity and Distributions 11, 91–99.
Scarr, F., 1878. The Field Books of Frank Scarr on an Expedition West of the Herbert River, Queensland, in 1878. Queensland State Archives, Brisbane.Schulze, L., 1891. The aborigines of the upper and middle Finke River: their habits and customs, with introductory notes on the physical and natural-
history features of the country. Royal Society of South Australia, Transactions and Proceedings and Report 14 (1891), 210–246.Shepard, W.D., 1993. Desert springs—both rare and endangered. Aquatic Conservation: Marine and Freshwater Ecosystems 3, 351–359.Spencer, B. (Ed.), 1896. Report on the Work of the Horn Scientific Expedition to Central Australia, vol. 4. Melville, Mullin and Slade, Melbourne.Stanley, E.H., Buschman, D.L., Boulton, A.J., Grimm, N.B., Fisher, S.G., 1994. Invertebrate resistance and resilience to intermittency in a desert stream.
American Midland Naturalist 131, 288–300.Storrs, M.J., Finlayson, M., 1997. Overview of the conservation status of wetlands of the Northern Territory. Supervising Scientist Report 116, Darwin.Tate, R., 1896a. Botany. In: Spencer, W.B. (Ed.), Report on the Work of the Horn Scientific Expedition to Central Australia. Part 2. Melville, Mullen and Slade,
Melbourne, pp. 94–117.Tate, R., 1896b. Mollusca. In: Spencer, W.B. (Ed.), Report on the Work of the Horn Scientific Expedition to Central Australia. Part 2. Melville, Mullen and
Slade, Melbourne, pp. 181–226.Terry, M., 1937. Sand and Sun. Michael Joseph Ltd., London.Timms, B.V., 2007. The biology of the saline lakes of central and eastern inland of Australia: a review with special reference to their biogeographical
affinities. Hydrobiologia 576, 27–37.Toyne, P., Japanangka Granites, R., Zimran, S., 1996. Towards a Bicultural Water Program in Central Australia: Consultations with Aboriginal Community
Groups. Publication of the Australian Government Publishing Service, Canberra.Unmack, P.J., 1995. Desert fishes down under. In: Proceedings of the Desert Fishes Council, 1994 Symposium, vol. 26, pp. 71–95.Unmack, P.J., 2001. Fish persistence and fluvial geomorphology in central Australia. Journal of Arid Environments 49, 653–669.US EPA, 2002. Methods for Evaluating Wetland Condition: Introduction to Wetland Biological Assessment. Office of Water, US Environmental Protection
Agency, Washington, DC (EPA-822-R-02-014).Wager, R., Unmack, P.J., 2000. Fishes of the Lake Eyre Catchment of Central Australia. Queensland Department of Primary Industries, Sydney.Wasson, R.J., 1984. Late Quaternary palaeoenvironments in the Australian desert dunefields. In: Vogel, J.C. (Ed.), Late Cainozoic Palaeoclimates of the
Southern Hemisphere. Balkema, pp. 419–432.Watts, C.H.S., Humphreys, W.F., 2006. Twenty-six new Dytiscidae (Coleoptera) of the genera Limbodessus Guignot and Nirripirti Watts and Humphreys,
from underground waters in Australia. Transactions of the Royal Society of South Australia 130, 123–185.White, D.S., 1993. Perspectives on defining and delineating hyporheic zones. Journal of the North American Benthological Society 12, 61–69.White, S., 1914. Into the dead heart: an ornithological trip through Central Australia. W.K. Thomas and Company, Adelaide.White, M., Albrecht, A., Duguid, A., Latz, P., Hamilton, M., 2000. Plant species and sites of botanical significance in the southern bioregions of the Northern
Territory; vol. 1: significant vascular plants. Report to the Australian Heritage Commission from the Arid Lands Environment Centre, Arid LandsEnvironment Centre and the Parks and Wildlife Commission of the Northern Territory, Alice Springs.
Williams, W.D., 1998a. Biodiversity in salt lakes. In: McComb, A.J., Davis, J.A. (Eds.), Wetlands for the Future. Contributions from INTECOL’s V InternationalWetlands Conference. Gleneagles Publishing, Australia, pp. 161–166.
Williams, W.D., 1998b. Salinity as a determinant of the structure of biological communities in salt lakes. Hydrobiologia 381, 191–201.Williams, W.D., 1999. Conservation of wetlands in drylands: a key global issue. Aquatic Conservation: Marine and Freshwater Ecosystems 9, 517–522.Williams, W.D., Kokkinn, M.J., 1988. The biogeographical affinities of the fauna in episodically filled salt lakes: a study of Lake Eyre South, Australia.
Hydrobiologia 158, 227–236.Winnecke, C., 1882. Herbert River and north-eastern exploring expedition. Parliamentary Papers South Australia No. 121, pp. 7–31.Wischusen, J., 1998. Hydrogeology of the Yuendumu-Papunya-Kintore Region Northern Territory (notes to accompany the Western Water Study 1:500000
Major Aquifer Systems Map), Record 1998/31. Australian Geological Survey Organization, Canberra.Wischusen, J.D.H., Fifield, L.K., Cresswell, R.G., 2004. Hydrogeology of palm valley, central Australia: a Pleistocene flora refuge? Journal of Hydrology 293,
20–46.Zhang, Y.K., Schilling, K.E., 2006. Effects of land cover on water table, soil moisture, evapotranspiration, and groundwater recharge: a field observation and
analysis. Journal of Hydrology 319, 328–338.Zietz, A., 1896. Pisces. In: Spencer, B. (Ed.), Report on the Work of the Horn Scientific Expedition to Central Australia. Part 2. Melville, Mullen and Slade,
Melbourne, pp. 176–180.