
Canonical germinal center B cells may not dominate the memory response to antigenic challenge
13
International Immunology, Vol. 13, No. 5, pp. 643–655 © 2001 The Japanese Society for Immunology Canonical germinal center B cells may not dominate the memory response to antigenic challenge Yi-Feng Lu, Mallika Singh and Jan Cerny Department of Microbiology and Immunology, University of Maryland School of Medicine, 655 West Baltimore Street, BRB 13-15, Baltimore, MD 21201, USA Keywords: antibody-forming cells, bone marrow, somatic hypermutation, spleen, V H genes Abstract Spleen and bone marrow (BM) are the major sites of antibody production and anamnestic response in systemically immunized mice. We examined the VDJ segment repertoire of antibody plaque- forming cells (APFC) in those two sites in the course of antibody responses to the hapten nitrophenyl (NP). Individual IgG APFC expressed any one of 10 V H segments of the V186.2/V3 (J558) gene family: 186.2, 102, 23, C1H4, 165.l, CH10, 3, 593.3, 24.8 and 671.5. The majority of cells in both spleen and BM expressed the V186.2 gene joined to a D segment with Tyr95. During a 2-month period after a single immunization, the V186.2 ⍣ APFC in BM accumulated 3 times as many somatic mutations than splenic APFC (average 8.5 versus 3 mutations/V H ); this process was T h dependent as shown by in vivo depletion of CD4 ⍣ lymphocytes. However, the V186.2 ⍣ APFC in both spleen and BM shared a recurrent W33L replacement, indicating their common origin from germinal centers. The APFC expressing the other (analogue) V H segments were evenly represented in the spleen and BM, but they accumulated few, if any, mutations. The anamnestic V186.2 ⍣ APFC were highly mutated both in the spleen and BM; they represented a new and unexpected clonotype. The V/D segments were joined by Gly95 instead of Tyr95, the W33L was absent and a new shared K58R replacement appeared. The APFC expressing the ‘analogue’ V H genes comprised ~20% of the anamnestic response and did not accumulate more mutations, but their affinities were in the range of the memory V186.2 ⍣ cells. These data suggest that the late primary and secondary responses to a hapten may be born by different B cell lineages, and that some clonotypes may reach the memory pool without an extensive mutation and expansion. Introduction Following primary immunization with a T-dependent antigen, rational explanation for the finding, made in several laborat- ories, that the primary antibody responses and the anamnestic antibody-forming cells and memory B cells evolve in different compartments of lymphatic tissues (1), and perhaps from response are dominated by different clonotypes (4–8). This scenario also predicts that the repertoire of memory antibody separate precursor cell lineages (2). The antibody-secreting plasmacytes, which produce the bulk of primary antibody, response reflects that of the GC B cells (9–11); however, this proposal deserves further scrutiny. differentiate from B cells in the T cell-rich areas of periarteriolar lymphatic sheaths, whereas memory B cells originate in The repertoire of progenitor memory B cells has been studied extensively in splenic GC during the primary response germinal centers (GC) that form transiently after the immuniza- tion (1). It is now widely accepted that the GC B cells, which to a hapten, (4-hydroxy-3-nitrophenyl)acetyl (NP), in mice with IgH b allotype. The heavy chain V regions (V H ) of NP-binding Ig undergo rounds of somatic hypermutations in their rearranged BCR genes followed by affinity-based selection, are progen- molecules in these animals are encoded by several germline segments of the V186.2/V3 subfamily of the J558 family itors of both cells that produce antibody with increasing affinity during the advancement of immune response and the memory (12–17). The repertoire of NP-reactive GC B cells can be summarized as follows: B cells that give rise to the rapid anamnestic antibody response upon antigen recall (3). This model provides a (1) The majority of expressed BCR are encoded by V H 186.2/ Correspondence to: J. Cerny Transmitting editor: C. Terhorst Received 2 September 2000, accepted 5 February 2001
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Canonical germinal center B cells may not dominate the memory response to antigenic challenge

International Immunology, Vol. 13, No. 5, pp. 643–655 © 2001 The Japanese Society for Immunology
Canonical germinal center B cells may notdominate the memory response to antigenicchallenge
Yi-Feng Lu, Mallika Singh and Jan Cerny
Department of Microbiology and Immunology, University of Maryland School of Medicine, 655 WestBaltimore Street, BRB 13-15, Baltimore, MD 21201, USA
Keywords: antibody-forming cells, bone marrow, somatic hypermutation, spleen, VH genes
Abstract
Spleen and bone marrow (BM) are the major sites of antibody production and anamnestic responsein systemically immunized mice. We examined the VDJ segment repertoire of antibody plaque-forming cells (APFC) in those two sites in the course of antibody responses to the haptennitrophenyl (NP). Individual IgG APFC expressed any one of 10 VH segments of the V186.2/V3(J558) gene family: 186.2, 102, 23, C1H4, 165.l, CH10, 3, 593.3, 24.8 and 671.5. The majority of cellsin both spleen and BM expressed the V186.2 gene joined to a D segment with Tyr95. During a2-month period after a single immunization, the V186.2� APFC in BM accumulated 3 times as manysomatic mutations than splenic APFC (average 8.5 versus 3 mutations/VH); this process was Thdependent as shown by in vivo depletion of CD4� lymphocytes. However, the V186.2� APFC inboth spleen and BM shared a recurrent W33L replacement, indicating their common origin fromgerminal centers. The APFC expressing the other (analogue) VH segments were evenly representedin the spleen and BM, but they accumulated few, if any, mutations. The anamnestic V186.2� APFCwere highly mutated both in the spleen and BM; they represented a new and unexpectedclonotype. The V/D segments were joined by Gly95 instead of Tyr95, the W33L was absent and anew shared K58R replacement appeared. The APFC expressing the ‘analogue’ VH genes comprised~20% of the anamnestic response and did not accumulate more mutations, but their affinities werein the range of the memory V186.2� cells. These data suggest that the late primary and secondaryresponses to a hapten may be born by different B cell lineages, and that some clonotypes mayreach the memory pool without an extensive mutation and expansion.
Introduction
Following primary immunization with a T-dependent antigen, rational explanation for the finding, made in several laborat-ories, that the primary antibody responses and the anamnesticantibody-forming cells and memory B cells evolve in different
compartments of lymphatic tissues (1), and perhaps from response are dominated by different clonotypes (4–8). Thisscenario also predicts that the repertoire of memory antibodyseparate precursor cell lineages (2). The antibody-secreting
plasmacytes, which produce the bulk of primary antibody, response reflects that of the GC B cells (9–11); however, thisproposal deserves further scrutiny.differentiate from B cells in the T cell-rich areas of periarteriolar
lymphatic sheaths, whereas memory B cells originate in The repertoire of progenitor memory B cells has beenstudied extensively in splenic GC during the primary responsegerminal centers (GC) that form transiently after the immuniza-
tion (1). It is now widely accepted that the GC B cells, which to a hapten, (4-hydroxy-3-nitrophenyl)acetyl (NP), in mice withIgHb allotype. The heavy chain V regions (VH) of NP-binding Igundergo rounds of somatic hypermutations in their rearranged
BCR genes followed by affinity-based selection, are progen- molecules in these animals are encoded by several germlinesegments of the V186.2/V3 subfamily of the J558 familyitors of both cells that produce antibody with increasing affinity
during the advancement of immune response and the memory (12–17). The repertoire of NP-reactive GC B cells can besummarized as follows:B cells that give rise to the rapid anamnestic antibody
response upon antigen recall (3). This model provides a (1) The majority of expressed BCR are encoded by VH186.2/
Correspondence to: J. Cerny
Transmitting editor: C. Terhorst Received 2 September 2000, accepted 5 February 2001

644 Delineation of memory B cells
DFL16.1/JH2 rearrangements (10,18–20) with canonical fusion and hybridoma selection or FACS that may introducea sampling bias. Using this approach, we aimed to analyzeTyr95 that is considered critical for NP binding (10).
(2) In most studies (18,20), but not all (10), the CDR3 appear the repertoire of memory effector cells in comparison with GClineage cells that produce high-affinity antibody during theto have little junctional diversity.
(3) The V186.2� GC B accumulate somatic point mutations advancement of primary response. Furthermore, memoryresponses were examined not only in the spleen, but also into an average of 3–5 mutations/VH (10,18–20). This
includes a recurrent point mutation in VH position 33 that bone marrow (BM), because the latter is a major site of rapidanamnestic responses (27–29). The antigen boost may havereplaces Trp (W) with Leu (L) in a majority of recovered
cells (10,19). This mutation alone increases the affinity of initiated a rapid migration of activated memory cells fromperipheral lymphatics to BM, where they differentiate intoV186.2–DFL16.1–JH2(Vλ1) antibody for binding to NP by
10-fold (21) indicating a selection of GC B cells into the plasmacytes (29). Another hypothesis is that BM may becolonized by memory B cells and/or their progenitors whichhigher-affinity memory cell population.
(4) In addition to the V186.2 clonotypes, the NP-specific GC give rise to antibody-forming cells in situ after secondaryimmunization (27,28). The latter scenario is consistent withcontained a variable portion of B cells expressing other
VH segments of the V186.2/V3 gene family, such as the observation that GC B cells home to BM upon adoptivetransfer into naive recipients (30). Finally, the question arisesV102, V23, VC1H4, V165.1, CH10, V3, V24.8 and V593
(17,19,20). These have been dubbed ‘analogue’ genes whether compartmentalization of memory cells is a randomor selective process. If it is selective, the memory responses(20) due to their sequence homology to the dominantly
expressed V186.2 segment (22). The analogue gene in the spleen and BM could have different repertoires.rearrangements collectively account for ~65% of all clonesrecovered from GC at 6 days post-immunization, but they
Methodsbecame rare by the second week (20).Does the repertoire of memory anti-NP antibody mirror that Mice, antigens and immunization
of GC B cells? The answer may depend on the experimentalC57BL/6 and C57BL/6-scid mice (age 6–8 weeks) weredesign and method. Studies (14,16) have shown that Bpurchased from the Jackson Laboratory (Bar Harbor, ME) orcell hybridomas generated from mice undergoing secondarythe Charles River Laboratory (Wilmington, MA) and maintainedresponses bear certain characteristics of GC cells: the majorityin sterile microisolator cages (Lab Products, Maywood, NJ).of the clones expressed the VH186.2 gene segmentsNP or its analogue (4-hydroxy-5-iodo-3-nitrophenyl)acetylrearranged to DFL16.1 genes that contained, on average,(NIP) (Cambridge Research Biochemical, Cambridge, UK)4–5 mutations/VH including the W33L replacement. Neverthe-were conjugated at various substitution ratios to keyholeless, the secondary clones tended to have different CDR3limpet hemocyanin, chicken γ-globulin (CGG) (both fromrearrangements in which the nucleic acid deletions at the DJSigma, St Louis, MO) or BSA (Amersham Life Science,boundary were more extensive (14,16) compared to those inCleveland, OH) as described (31). Mice were immunized withthe GC B cells (10,18–20). This repertoire pattern suggestsa single i.p. injection of 10 µg of an antigen in alum andthat precursors of memory B cells represented only a smallchallenged with 10 µg of a soluble antigen in PBS, i.p.fraction of the GC cell population. However, these studies
used complicated experimental designs that could bias theT cell depletion in in vivoresults. The secondary responses were elicited respectivelyAnti-CD4 mAb GK1.5 (ATCC, Rockville, MD) was producedin mice treated neonatally with an anti-idiotypic antibody (16)as ascitic fluid and the IgG was purified on a HiTrap Proteinand in a single mouse that received two consecutive antigenG column (Amersham Pharmacia Biotech, Piscataway, NJ).boosts within 3 weeks after the priming (14). In contrast,Mice were injected i.v. with 1–2 mg IgG in 0.25 ml of PBS onSiekevitz et al. (23) generated secondary anti-NP hybridomasdays 18, 23, 32 and 42 after the immunization with NP-CGG;upon an adoptive transfer of antigen-primed cells followedcontrol animals received 1 mg of purified rat IgG (Sigma ) onby challenge with a anti-idiotype as a surrogate antigen. Thethe same days .secondary clones expressing the VH186.2 gene had fewer
average mutations (~1.5 mutations/VH) than would bePreparation of NIP-red blood cellsexpected if they originated from GC, but many of them hadSheep blood cells in Alsever’s solution (Colorado Serum,nucleotide deletions in D segments similar to the afore-Denver, CO) were washed in DPBS and resuspended to 10%mentioned hybridomas from other studies (14,16). Neither of(v/v) in 0.1 M bicarbonate buffer, pH 9.4. Succinimide esterthese studies assessed the role of GC analogue B cells (20)of NIP (Cambridge), dissolved in dimethylformamide (Sigma),in the secondary response.was added to a final concentration of 25 µg/ml of sheepThe aim of the present study was to examine the repertoirered blood cells and the mixture was incubated at roomof NP-specific anamnestic responses elicited in primed, intacttemperature for 30–60 min. Cells were then washed andanimals with a low dose of soluble antigen boost. Withinresuspended to 20% (v/v) in DPBS.2 days the memory B cells (but not the naive lymphocytes)
differentiate into effector IgG antibody-producing cells (24,25)APFC assaythat are readily identified in lymphocyte suspension ex vivo,
using a modified antibody plaque-forming cell (APFC) assay Lymphocytes producing the NP-specific IgG antibody weredetected with a modified hemolytic plaque assay (26) using(26). The rearranged Ig genes from individual APFC can be
amplified and sequenced without the need for either cell sheep red blood cells coupled with hapten. Antibodies pro-

Delineation of memory B cells 645
Table 1. Detection of NP-specific IgG APFC
Cell linea No. of plaques in agarose platesb with antibody added
None Anti-IgM Anti-IgG Anti-IgM � Anti-IgG
B1-8 150, 140 15, 0 120, 90 2, 5B1-8γ1 0, 0 0, 1 113, 127 119, 131B1-8 � B1-8γ1c 170, 165 25, 0 297, 277 147, 107
aB1-8 is a NP-specific IgM hybridoma (12) and B1-8γ1 is a NP-specific IgG1-producing transfectoma (35).bPlates (in duplicates) contained 1 � 103 cells from the indicated line.cMixture of 103 cells from each line.
duced early in the response to NP bind more avidly to NIP, binding activity to both NP-BSA and NIP-BSA. Hybridomaswere cloned by limiting cell dilution at 0.3 cell/well and thewhereas the secondary antibodies bind to NP and NIP withpositive clones were expanded. mAb were purified fromsimilar affinities (32,33). In order to capture cells producingculture supernatants using Protein A–Sepharose columnsboth types of antibody molecules, we performed the plaque(Pharmacia Biotech, Uppsala, Sweden); their H chain andassay with sheep red blood cells conjugated to NIP. PetriL chain isotypes were determined by standard ELISA withdishes (10 cm diameter) were pre-coated with 5 ml of 0.5%NP-BSA and NIP-BSA as antigens in solid phase. The platesagarose (Gibco, Grand Island, NY). A mixture containingwere probed with goat anti-mouse Ig isotype-specific anti-2.5 ml of 0.3% agarose in Basal Medium Eagle (Gibco) keptbodies, and anti-κ and anti-λ chain antibodies (all fromin a 46°C waterbath, 100 µl of lymphocyte suspension andSouthern Biotechnology Associates). The VDJ segments from100 µl of NIP-red blood cells, with or without 25 µl ofthe clones were characterized by PCR amplification andantibody mixture containing goat anti-mouse IgM (Southernsequencing as described below.Biotechnology Associates, Birmingham, AL) and rabbit anti-
The affinities of mAb were estimated in two ways. Theirmouse IgG (Sigma), was poured on the solid agarose baseKa values for NP and NIP haptens were measured byand the dishes were incubated at 37°C/5% CO2 for 2 h. Thefluorescence quenching in a Shimadzu RF-5301 fluoro-plates were then covered with 3 ml of diluted guinea pigspectrophotometer using monovalent haptens NP- andcomplement (Gibco) and incubated for an additional 2 h toNIP-aminocaproric acid (Cambridge Research) as described
make the hemolytic plaques visible. The hemolytic plaque (35,36). Relative affinities were also determined by the bindingassay readily detects the IgM-producing cells; however, the of mAb to NP-BSA conjugates substituted with differentIgG-producing cells can be detected with an optimal concen- amounts of haptenic groups (37). Purified mAb at concentra-tration of exogenous anti-IgG antibody (26). Our assay was tions from 0.3 l to 3 µg/ml were tested by ELISA for bindingcalibrated for the detection of IgG using stable cells lines B1- to NP3-BSA and NP18-BSA. Increasing ratio of binding calcu-8 (34) and B1-8γ1 (35) that produce anti-NP antibody of IgM lated as OD450NP3-BSA/OD450NP18-BSA was indicative ofand IgG1 isotype respectively. The agarose plates contained increasing affinity of mAb to NP. A similar ELISA protocol wasa mixture of anti-mouse Ig antibodies that inhibited the IgM- used to determine the heteroclicity of the NP-induced mAb,producing cells and facilitated the IgG producers (Table 1). i.e. their preferential binding to NIP (32). The mAb were
For enumeration of APFC, 8–20�105 cells were plated per allowed to react with NP18-BSA and NIP24-BSA and thedish in duplicate. For the isolation of a single APFC, the binding reaction � OD450NIP/OD450NP was calculated. Ratioscell suspension was highly diluted in order to increase the ~1.0 indicate a lack of heteroclitic reactivity.probability of sampling one cell. We plated 1�104 splenocytes
Recovery and molecular analysis of GC B cells(estimated to be 5�103 B cells) and 3–4�104 BM cellsSpleens were removed from mice on days 12 after primary(estimated as 3–4�103 B cells), which yielded �1 plaqueimmunization, frozen sections were prepared for immunohisto-per dish. The center of each plaque, containing a cell clearlychemistry, and NP-reactive GC B cells were identified by dualvisible under a dissecting microscope, was aspirated in astaining with peanut agglutinin and NIP-BSA as previouslyvolume of ~0.4 mm3 of agarose. From the total number ofdescribed (38). Cells (~100) from individual GC were micro-cells plated in 2.5 ml volume, we calculate that each sampledissected (20,38) and transferred into microcentrifuge tubescontained approximately one B cell. On average, 30–60% offor PCR DNA amplification. The first-round PCR productsamples yielded a PCR product using the specific primerswas re-amplified with the nested primers shown below, with(see below). In contrast, no products were obtained fromadditional recognition sequences for restriction enzymes (17).
random sampling of the red blood cells lawn. The PCR product was digested with the restriction enzymes,ligated to a plasmid, cloned in competent Escherichia coliHybridomas and affinity measurementsand sequenced as described previously (17).
Spleen cells were harvested from mice at day 4 after theAmplification and sequencing of VDJ DNA recovered fromsecondary challenge with NP-CGG and fused with an SP2/0individual APFCmyeloma cell line (ATCC), using polyethylene glycol (average
mol. wt 1450) (Sigma) according to standard procedures. The individual NIP�, IgG APFC were picked using 50-µldisposable micropipettes (Fisher Scientific, Pittsburgh, PA)Supernatants from fusion wells were screened by ELISA for

646 Delineation of memory B cells
and transferred into a 0.2 ml microcentrifuge tube with 2.5 µlof 10�PCR buffer, 0.5 µl of 20 mg/ml proteinase K (BoehringerMannheim, Mannheim, Germany), 0.5 µl of 5% Tween 20 and21.5 µl of dH2O. The tubes were incubated at 56°C for 1 hand the proteinase K was then inactivated at 96°C for 10 min.DNA amplification was carried out by two rounds of PCRusing pairs of nested primers. The initial round of amplificationused primers 5�-CCTGACCCAGATGTCCCTTCTTCTCCAGC-AGG-3� and 5�-GGGTCTAGAGGTGTCCCTAGTCCTTCAT-GACC-3�, corresponding to the V186.2 genomic DNA 5�transcription start site sequence and to the intron JH2sequence respectively. Using the Expand High Fidelity PCRkit (Boehringer Mannheim), the crude cell lysate was mixedin a 50 µl reaction volume with 350 µM dNTP, 0.4 µM of eachprimer and 1.0 µl of Expand High Fidelity polymerase. It wasthen amplified by one cycle of 95°C 2 min/55°C 4 min/72°C2 min and then 40 cycles of 95°C 1 min/55°C 1 min/72°C 2min in a GeneAmp 2400 PCR System (Perkin Elmer). Reaction Fig. 1. Kinetics of IgG APFC in the spleen (s) and bone marrow (u) of
mice immunized with 10 µg of NP-CGG in alum, i.p. (1o immunization).mixture (2 µl) from the first-round PCR was amplified for anCohorts of primed mice received a secondary (2o) challenge of 10additional 40 cycles (94°C 30 s/55°C 1 min/68°C 1 min) usingµg of soluble NP-CGG on days 35 and 60, and the anamnestic APFCthe nested primers 5�-CTAGAATTCAGGTCCAACTGCAGCA- responses in the spleen (d) and BM (j) were determined 4 days
GCC-3�, complementary to the initial 20 nucleotides of the after the boost.V186.2/V3 gene family; and 3� primer 5�-GGGTCTA-GAGGTGTCCCTAGTCCTTCATGACC-3� complementary tothe JH2 gene segment. The second reaction mixture (5 µl)
Resultswas electrophoresed on a 1% agarose gel for identificationof amplified DNA (~350 bp fragment). The PCR product,
Distinctive repertoires of late APFC in the spleen and boneisolated with the QIAquick gel extraction kit (Qiagen, Hilden,
marrowGermany), was directly sequenced by the Biopolymer Labor-
The kinetics of appearance of hapten-specific, IgG-producingatory of the University of Maryland School of Medicine usingAPFC in the spleen and bone marrow in mice immunizedan automatic DNA sequencing system (Applied Biosynthesis).systematically (i.p.) with NP-CGG in alum is shown in Fig. 1.DNA codons are numbered according to Kabat et al. (39).This pattern of antibody-producing cells is familiar from severalrecent studies (40–42). The splenic APFC reached the peakAssignment of DNA sequences to germline VH genesat days 7–10 after the immunization and then declined to a
Germline sequences of individual VH segments of V186.2/V3 low, steady level between days 35 and 65. In contrast, specificgene family differ from one another by 2–30 nucleotide APFC did not appear in bone marrow until the second weekresidues (22). The APFC that utilized V segments 102, 23, after priming and their number increased through the 2-monthC1H4, 165.1, CH10, 3, 593.3, 24.8 and 671.5 contained few, period of observation (Fig. 1). Cells persisting in the spleenif any, somatic mutations (see Results), and, therefore, the and bone marrow for 2 months after the immunization will besequences were easily matched to their respective germline referred to as ‘late’ APFC.counterparts. In contrast, the sequences representing the VDJ rearrangements in a small sample of early splenicV186.2 gene differed from their presumptive germline counter- APFC (day 7) involved predominately the V186.2 segmentpart by up to 20 nucleotides indicating that the V segments without somatic mutations (Table 2). At the later stage of thewere either somatically mutated or that they were derived response (days 35–65), about half of the splenic APFC usedfrom a different germline gene. Distinction was possible the V186.2 gene and the remaining cells expressed ninebecause the nucleotide differences between the recovered different analogue genes that were evenly representedsequences and the V186.2 germline sequence typically fell (Table 2). (See Methods for description of V gene assignment.)into the positions that are shared by members of the V186.2/ Nucleotide substitutions were present in 30–40% of VH genesV3 gene family and spared those positions that characterize with an average of 3.0 mutations/V186.2 gene and 1.8individual germline genes. Thus codons nos 11, 20, 43, 50, mutations/analogue V gene. The frequency of mutants and74 and 91 (and others) that distinguish the 186.2 gene were the average mutations in the rearranged segments of thenever mutated. Moreover, when mutation did occur in a codon V186.2/V3 gene family in the late splenic APFC are thus verythat defines the germline V186.2 gene it spared the key similar to that found in the splenic GC B cells in NP-CGG-residue. An example is given with codon 58 which is AAG in immunized mice (Table 2, column 2) (10,17,18,20). However,V186.2 and AAC in all other members of this gene family. the late APFC in bone marrow showed a different repertoireMost 186.2 sequences from anamnestic response shared the (Table 2). In a sample that was comprised of 12 distinctpoint mutation AAG → AGG, whereas the third germline G clonotypes from six animals (Table 2, footnote), it was foundresidue was never changed. These patterns allowed an that 90% of V186.2� APFC were mutated with an average of
8.5 mutations/V gene, which is nearly a 3-fold increase overunambiguous classification of V186.2 mutants.

Delineation of memory B cells 647
Table 2. VH gene usage and somatic mutations in NP-reactive B cells during the primary responsea
VH gene Spleen Bone marrow
GC B cells (day 12)b Early APFC (day 7)c Late APFC (days 35–65)c Late APFC (days 35–65)c
Mutated/ Average Mutated/ Average Mutated/ Average Mutated/ Averagetotald mutations/VH totald mutations/VH totald mutations/VH totald mutations/VH
186.2 32/35 (91%) 2.8 (1–8)e 0/9 0 6/14 (43%) 3.0 (2–6)e 17/19 (89%) 8.5 (2–20)e
R/Sf: CDR1�2 3:1 4.5:1 5.6:1FWR 1.7:1 1:1 2.8:1
Analogue102 3/6 2.5 0/1 0 2/3 3.0 1/2 3.023 9/16 2.0 – 1/3 2.0 3/6 1.0C1H4 8/10 1.9 – 0/2 0/3165.1 2/2 2.3 – 1/3 1.0 0/1CH10 – – 0/1 0/13 1/1 4.0 – 0/1 0/2593.3 – – 1/4 1.0 0/224.8 1/1 2.0 – 1/3 1.0 –671.5 – 0/1 0 0/1 –
analogue, Σ � 24/36 (67%) 2.2 6/21 (29%) 1.8 4/17 (24%) 1.5
aMice were immunized with 10 µg of NP-CGG in alum i.p. and sacrificed on the indicated days.bVDJ segments were recovered by microdissection of GC (50–100 cells) from frozen splenic sections (see Methods) followed by DNA
amplification, cloning and sequencing. The sample represents seven GC from three mice.cIndividual anti-NP IgG APFC were analyzed by direct sequencing of the PCR product of DNA amplification. The late splenic APFC were
sampled from five individual mice and represent 12 independent clonotypes according to distinct V(D)J rearrangements. The BM APFC wereisolated from six mice (five of them were the same as the donors of the splenic cells) and represent 12 distinct clonotypes.
dProportion of cells containing VH segments with one or more nucleotide substitutions among the total number of analyzed cells.eRange of mutations among individual VH186.2 genes.fRatio of replacement/silent mutations in the CDR1 � 2 and framework regions.
the mutation frequency found either in GC B cells or in from Th-depleted mice was comparable to the control groupin respect to the dominance of V186.2 gene rearrangements,late splenic APFC. The dramatic difference between the
distribution of mutations among the V186.2� APFC in the prevalence of W33L replacement and high ratios of R/Smutations in CDR1 � 2; however, the average frequency ofspleen and those in the bone marrow is shown in Fig. 2(a).
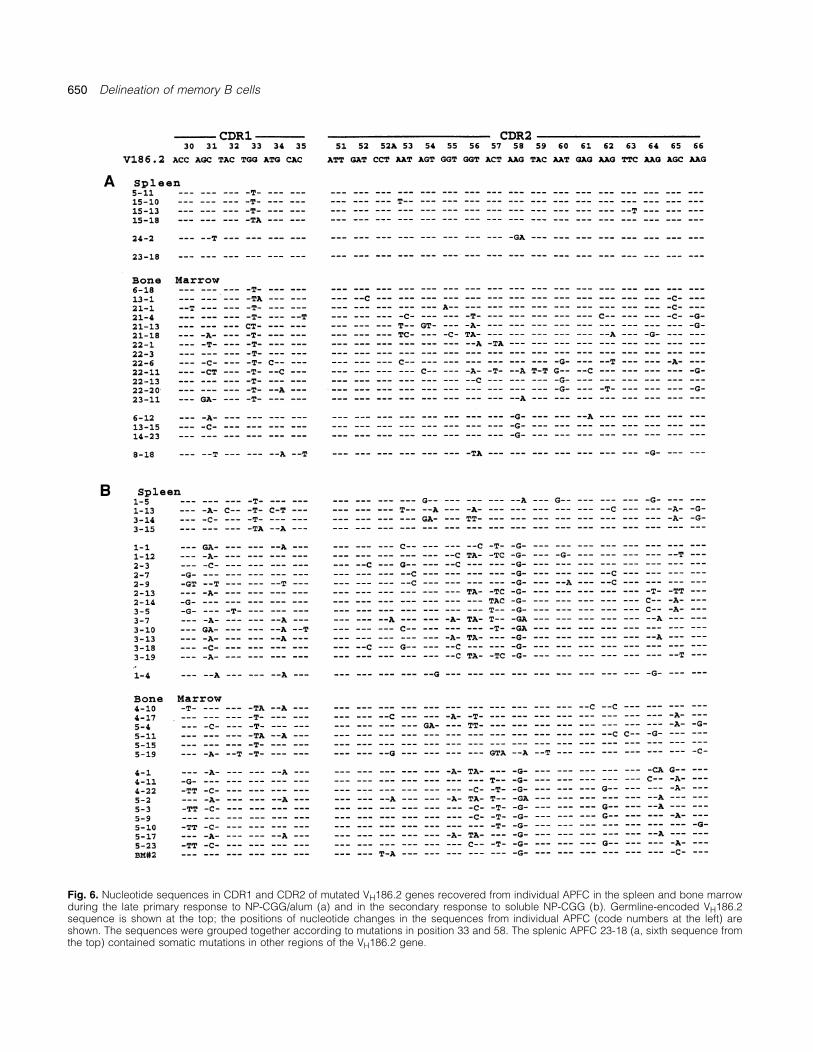
Most of the V186.2� APFC mutants in the spleen and BM mutations/VH was reduced by half, to the levels found in GCB cells. Indeed, the Th cell depletion selectively decreasedcarried the W33L and used the canonical CDR3 with Tyr95
(see Figs 6a and 7a), which affirms their relatedness to the BM APFC with �7 mutations/VH, whereas the cells with 1–6mutations/VH were equally frequent in both Th-depleted andGC B cell lineage, yet the APFC in BM accumulated more
somatic mutations. control animals (Fig. 4).Despite the high mutation frequency in the V186.2� cells,
Repertoire shift in V186.2� memory APFCthe BM APFC expressing the analogue VH genes were eitherunmutated or contained only 1–3 mutations/gene (Table 2, Next we examined whether the repertoire detected in an
anamnestic response to NP is similar to that of GC memoryV102 and V23), a frequency comparable to that of GC B cellsand the late splenic APFC. Thus the high number of mutations B cells and their late APFC progeny. Cohorts of mice were
challenged with 10 µg of soluble NP-CGG on days 35 andin BM APFC is peculiar to the dominant V186.2 clonotype.65 after the priming, and NP-specific IgG APFC were sampled
Somatic repertoire of late APFC in BM is T cell dependent 4 days after the boost (Fig. 1). The data from both animalgroups (days 35 and 65) are presented together becauseInasmuch as somatic hypermutation is driven by Th cells
(43,44), it was of interest whether the appearance of the their anamnestic responses were comparable. Eighty percentof secondary APFC expressed the V186.2 gene while thehighly mutated BM APFC was T cell dependent. CD4�
lymphocytes were depleted with repeated injections of anti- remaining 20% used various analogue VH genes (Table 4).Somatic mutations were found in all V186.2� cells in bothCD4 mAb GK1.5 beginning on days 18 after the immunization
with NP-CGG, i.e. at the time of diminishing GC reaction spleen and BM, with an average of 11 mutations/VH (Table 4,see the footnote for details on sampling). In contrast to the(42,45). The treatment reduced the numbers of CD3�CD4�
lymphocytes to �0.1% in both spleen and BM until days 35, late primary APFC, there was no difference in the distributionof mutations among the secondary APFC in the two anatomicwhen they slowly begun to rise (Fig. 3). As shown in Table 3,
the repertoire of the late NP-specific IgG APFC in the BM sites (Fig. 2b). To our surprise the secondary APFC population

648 Delineation of memory B cells
was dominated by a clonotype that has been rarely detected replaced lysine (K) with arginine (R) in CDR2 position 58 inup to 80% of cells (Figs. 5 and 6). Remarkably, the K58Rbefore in the GC B cell lineage. Typical NP-reactive V186.2�
B cells that emerge from the primary GC have the W33L mutants always carried Gly95, which joined the V186.2 genereplacement (10,19), and this motif is also found in ~70% oflate primary APFC in both spleen and BM (Figs 5 and 6).Among the memory APFC, however, this mutation declinedto ~30% while a new recurrent mutation appeared that
Fig. 3. Depletion of CD4� T cells in the spleen (d) and bone marrow(j). Mice that were immunized with NP-CGG/alum (day 0) receivedfour injections of anti-CD4 mAb (1–2 mg each, i.v.) on days 18, 24,32 and 41 (arrows). CD3�CD4� lymphocytes were enumerated byFig. 2. Distribution of somatic mutations in the rearranged VH186.2
genes among hapten-specific APFC in the spleen (empty bars) and flow cytometry (two animals/interval). Horizontal bars indicate thepercentages of CD3�/4� lymphocytes in the spleen (solid) and BMBM (stripped bars) at the (a) late primary responses (days 35–65)
after immunization and (b) anamnestic response (day 4) after the (dashed) of control animals injected with normal rat IgG (averagefrom four mice with individual range shown by the vertical bars).antigen boost).
Table 3. Somatic mutations in NP-specific APFC in bone marrow upon T cell depletiona
GK1.5 anti-CD4 Rat IgG (control)
Day 37 Day 63 Total Day 37 Day 63 Total
VH186.2 n � 13 8 21 17 20 37average mutations/VH 4.0 5.4 4.7 8.6 7.7 8.2W→L position 33 83% 87% 85% 72% 60% 60%R/S, CDR1 � 2 �12 6.7 11.0 5.6 9.5 6.8FWR 3.0 3.6 3.3 1.1 1.8 1.4
VH analogue n � 8 13mutations/VH 1.7 5.5R/S, CDR1 � 2 – 2.0FWR 1.0 3.0
aAverage numbers of NP-specific IgG APFC in BM at 37 and 63 days respectively were 20 and 35 APFC/106 cells in mice treated with anti-CD4 as compared to 42 and 88 APFC/106 cells in the control group. Two to three mice were analyzed at each time point.

Delineation of memory B cells 649
to either DFL16-1 or DSP2-5/7 segments (Fig. 7). The use of binding antibody molecule that preclude the co-existence ofboth mutations in V186.2, or that the two clonotypes, Leu33/Gly95 instead of Tyr95 must have developed from junctional
diversity during V–D–J rearrangements. The V186.2 sequence Tyr95 and Arg58/Gly95, represent two separate pathways ofmemory maturation and selection.contains a mutational hot-spot consensus motif AAGTA (46,47)
in position 58–59; however, that position 58 mutation is alwaysRepertoire of analogue VH genes in memory anti-NP response.A → G (Fig. 6) argues for antigen selection of the mutated
cells. Thus the patterns of recurrent mutations and VD junc- Various analogue VDJ rearrangements were found in thesecondary APFC in the spleen as well as in BM but unliketions identify two functionally distinct populations of V186.2�
memory B cells: (i) Leu33/Tyr95 (Lys58) GC B cells and the heavily mutated V186.2 genes, the analogue VH segmentsstill contained only 1–3 mutations/VH (Table 4), which repres-progenitors of late APFC, and (ii) Arg58/Gly95 (Trp33) progen-
itors of anamnestic APFC. ents no increase over the average mutations in GC B cellsWe noted with interest that, without exception, the W33L
and K58R mutations in V186.2 were exclusive of one anotheramong 70 APFC analyzed in this study (Fig. 6). We speculatethat there are either structural constraints imposed on the NP-
Fig. 5. Frequencies of W33L (plain bars) and K58R (hatched bars)mutations in VH186.2 gene in late primary (1o) and in secondary (2o)responses to NP-CGG. Each bar represents the proportion (%) ofFig. 4. Distribution of somatic mutations in VH186.2 genes in bone
marrow APFC from CD4 T cell-depleted (hatched bars) and control APFC carrying the respective mutation in the spleen (light bars) andBM (dark bars).(empty bars) mice.
Table 4. VH repertoire and somatic mutations in secondary APFCa
VH gene Splenic APFC Bone marrow APFC
Mutated/total (%)b Average mutations/VH Mutated/total (%)b Average mutations/VH
186.2 18/18 (100%) 11.5 (7–23)c 16/16 (100%) 11.2 (2–18)c
R/Sd: CDR 1�2 6:1 8.3:1FWR 3:1 2.6:1
Analogue102 2/2 3.0 1/1 3.0C1H4 1/1 2.0 –CH10 – 1/1 1.0593.3 1/2 1.0 –
analogue, Σ � 4/5 (80%) 2.0 2/2 (100%) 2.0
aMice were immunized with 10 µg of NP-CGG in alum and challenged 5–8 weeks later with 10 µg of soluble NP-CGG (see Fig. 1). Theywere sacrificed 4–5 days after the challenge and examined for NP-specific IgG APFC (seven mice). Spleens from two animals were pooledfor hybridoma production.
bProportion of cells containing VH segments with one or more nucleotide substitutions among the total number of analyzed cells. SplenicAPFC were obtained from two mice and represent at least 10 distinct V(D)J clonotypes. The BM APFC were isolated from three mice andrepresent at least six independent clonotypes. The size of V(D)J repertoire is probably an underestimate because these are heavily mutatedsequences. In some cases, it was difficult to determine whether the nucleotide differences in CDR3 represent somatic events orjunctional diversity.
cRange of mutations among individual VH186.2 genes.dReplacement/silent mutation ratio.
650 Delineation of memory B cells
Fig. 6. Nucleotide sequences in CDR1 and CDR2 of mutated VH186.2 genes recovered from individual APFC in the spleen and bone marrowduring the late primary response to NP-CGG/alum (a) and in the secondary response to soluble NP-CGG (b). Germline-encoded VH186.2sequence is shown at the top; the positions of nucleotide changes in the sequences from individual APFC (code numbers at the left) areshown. The sequences were grouped together according to mutations in position 33 and 58. The splenic APFC 23-18 (a, sixth sequence fromthe top) contained somatic mutations in other regions of the VH186.2 gene.

Delineation of memory B cells 651
Fig. 7. Structures of CDR3 of rearranged VH186.2 genes from Fig. 4. The names of corresponding D genes are shown in the right column.
(Table 2). The patterns of recurrent somatic mutations could A transition in the GGCT mutational hot-spot (46,47) in position26, which replaces Gly with Asp, suggesting an antigennot be assessed because individual analogue VH gene
mutants were recovered too infrequently throughout this study, selection process among the V102� B cells even though themutation is in FWR2. [A different interpretation is given towith the exception of the V102 gene (Fig. 8). Seven out of
nine V102 sequences from unrelated APFC contained a G → V102 position 10 where we found A instead of G in every

652 Delineation of memory B cells
Fig. 8. Recurrent nucleotide substitutions in rearranged VH102 genes recovered from GC, APFC (pooled data from primary and secondaryresponses) and secondary hybridomas from NP-CGG-immunized mice. The nucleotide sequences in framework regions 1–3 and CDR2 ofgermline-encoded VH102 are shown on the top.
sequence, including those that did not contain any other proceed along distinct pathways of repertoire maturation andnucleotide substitution (Fig. 8). We believe that A in position differentiation. The predominant cell population, which uses10 represents the correct germline sequence of this gene]. the VH186.2–DFL16-1/2 rearrangement with the canonical
Some of the analogue VH sequences recovered from sec- Tyr95 joint and accumulates somatic mutations including theondary APFC were unmutated, raising the possibility that the recurrent W33L, apparently migrates from GC to the spleenrespective APFC originated from naive B cells in the presence and BM, and differentiates into high-affinity antibody-produ-of carrier-primed Th cells, rather than from true memory cing cells at later stages of the immune response (42,48,49).B cells. In order to gain further insight into this problem, Here we show that somatic diversification of the V186.2/Tyr95/the repertoire of analogue VH gene-expressing B cells was Leu33 cell lineage is tissue specific. The APFC in BM evolvedexamined by construction of hybridomas. Mice immunized from highly mutated precursors, whereas the splenic APFCwith NP-CGG/alum were boosted 60 days later with soluble did not accumulate mutations above the level seen in the GCantigen and the splenocytes were immortalized at days 4 B cells. The primary GC in the spleen cease by 4–5 weeksafter the antigen boost. Several stable, NP-binding hybridoma after the immunization with antigen in alum (42,45), butclones were obtained that expressed respectively VH102, perhaps some B cells continue to mutate in the post-GCCH10 and 23 (Table 5). The binding affinities of these mAb phase of the response (48). The distinct repertoire of BMwere comparable to the affinities of the V186.2� secondary effector cells could indicate that only the highly mutated Bhybridomas and at least 1 log higher than that of the V186.2�
cells home to BM and differentiate in to APFC. Alternatively,mAb B1-8γ1 (Table 5) that is representative of the primary and more likely, the progeny of GC B cells circulate throughgermline-encoded anti-NP antibody (35). The fine specificity spleen and BM, and the latter environment provides signal(s)of the secondary analogue mAb is also comparable to that
for their continuing somatic mutation and differentiation toof secondary canonical mAb: they bind NP and NIP equallyAPFC in situ, as has been proposed for human BM (50). Suchwell, whereas the primary, unmutated V186.2� antibodiessignal(s) could be generated by BM CD3�CD4� lymphocytes,show a preferential binding to NIP (Table 5) (35). Althoughwhich are unusual in that the majority has a memory/activatedthe sample is small, it is noticeable that the affinities of thephenotype (51). Indeed, depletion of CD4� lymphocytesanalogue mAb to NP did not correlate with the extent of3 weeks after the immunization selectively eliminated themutation in VH, and that even unmutated mAb may have anhighly mutated APFC in BM (Fig. 4).affinity that is typical of secondary response (cf. the V23 mAb
Although the principal GC B cell lineage, V186.2/Tyr95/11-2-9 and 60-2-10 and the VCH10 mAb G6-18-6 and G6-Leu33, does contain precursor memory cells (14,16), the brisk18-10 in Table 5).anamnestic response to a low-dose, soluble antigen boost isdominated by a different population of effector V186.2� APFCDiscussionthat have a distinct N residue Gly95, germline-encoded Trp33and the K58R replacement. B cell clones with Gly95 haveThe model of antibody response to NP demonstrates how
primary splenic GC spawn multiple B cell populations that rarely been recovered from NP-specific GC (10,18,48) sug-

Delineation of memory B cells 653
Table 5. NP-reactive secondary mAb expressing different VH genes
VH gene Hybridoma H chain isotype Mutations/VH Affinity
Ka (M–7)a NP3:NP18 bindingb NIP:NP bindingc
NP NIP
23 11-2-9 γ1 1 0.6 0.5 �1.0 NT60-2-10 γ1 0 0.5 0.6 �1.0
CH10 G6-18-6 γ1 5 1.4 2.0 1.0 0.5G6-18-10 γ1 0 0.4 0.8 0.8 0.7
102 E8-11-1 γ1 6 1.3 0.7 1.0 1.7G6-18-13 γ1 3 1.0 1.4 0.5 0.8
186.2 G-6-6 γ1 7 2.3 1.6 0.8 0.6B-8-2 γ2b 10 2.5 2.5 0.4 0.9
B1-8γ1 (control)d γ1 0 14.0 1.3 �0.01–0.08 �10
aAffinity was measured by fluorescence quenching using monovalent NP and NIP haptens (see Methods).bRelative binding of mAb to NP3-BSA and NP18-BSA was determined by ELISA.cRelative binding of mAb to NIP24-BSA and NP18-BSA in ELISA. The control, primary mAb, B1-8γ1, shows the typical preferential binding to NIP.dB1-8γ1 is a prototype of primary anti-NP antibody encoded by unmutated VH 186.2 gene in canonical VDJ rearrangement (35).
gesting that the V186.2�/Gly95/Arg58 memory cell subset unmutated. As stated earlier, we cannot rule out that theunmutated APFC were generated from naive rather thanundergoes a post-GC expansion and maturation to become
a ‘higher order of memory’ cell (52) in the spleen and BM. memory B cells. However, we were able to obtain secondaryhybridomas that used analogue VH genes with few or noFurokawa et al. (53) recently found that the V186.2/Gly95
mAb (Gly99 in their numbering system) reached higher affinit- somatic mutations, which had affinities that are representativeof the memory repertoire to NP (Table 5). We hypothesize thaties than the V186.2/Tyr95 mAb did during the affinity matura-
tion of anti-NP response. Although the authors did not mention VH analogue clones may reach the memory pool withoutmutation and affinity maturation, consistent with the predictionit, all of their V186.2/Gly95 shared the K58R and none had
the W33L, as is the case of APFC in our study. We propose (54) that the germline-encoded affinities of certain BCR arenot increased by introduction of somatic mutations.that K58R is a key mutation for selection of the highest affinity
memory B cells in the post-GC stage of anti-NP response. The molecular and functional heterogeneity of memoryB cells has been best demonstrated by Berek, Milstein andThe third population of NP-reactive GC/memory B cells is
composed of diverse clonotypes expressing at least 10 co-workers, in studies of hybridomas from mice that wererepeatedly immunized with a hapten, oxazolone (4,55,56).closely related VH analogue genes which, as a group, account
for almost half of APFC in the late primary response in the The secondary and tertiary responses were dominated bydistinct cell lineages, depending on immunization protocol.spleen and BM, and a variable portion of the anamnestic
response. These clonotypes were observed in earlier studies, Berek and Milstein (56) proposed that co-evolution of function-ally diverse memory B cell clones is made possible becausebut their potential was not appreciated. Weiss and Rajewsky
(9) recovered a large number of rearranged VH analogue of their compartmentalization, either by antigen presentation,anatomical separation or cell kinetics. The NP-specific mem-genes at 6 weeks after immunization with NP-CGG, many of
which were said to contain somatic mutations. Others have ory borne by the VH186.2 clonotypes exemplifies the paradigmof repertoire maturation based on selection of higher-affinity Bdescribed NP-binding hybridomas after secondary immuniza-
tions which expressed several analogue VH genes (14,23). In cell mutants during the late primary and secondary responses(55,57,58). On the other hand, the behavior of NP-reactiveone study (23), the anamnestic response to NP was initiated
with anti-idiotype, which led the authors to postulate that the VH analogue clones is reminiscent of the responses in whichthe antibody affinities improve little (59) or not at all (60,61)high percentage of NP-binding hybridomas with analogue VH
genes resulted from a triggering of naive B cells. A most after the repeated immunization. We propose that the evolutionof distinct phenotypes of memory B cells may depend on theconvincing demonstration of the existence of analogue VH
memory cells was provided by Decker et al. (43) in their initial affinities of the respective BCR for NP and on the signalsfrom Th cells. The latter is suggested by the observationsstudy of secondary and tertiary responses to NP in splenic
fragments. Between 20 and 50% of fragments that produced that the expansion of NP-specific V186.2 clones is T cell-dependent: the analogue VH clonotypes become prevalentthe secondary or tertiary anti-NP antibody were occupied
by analogue VH gene clonotypes that contained somatic when T cell help is either decimated by advanced age (38)or by-passed altogether by immunization with a T-independentmutations, which were also recovered from APFC in our study,
including V593, 24.8, 23 and 165.1. Here we show that the form of NP (13). We may reasonably postulate that the affinityof BCR influences the concentration of MHC–peptide ligandslate primary and secondary APFC expressing analogue VH
do not accumulate somatic mutations above the range found on the B cells, which has been shown to influence the qualityof T cell help (62,63). We envision that the NP-reactive clonesin the GC B cell progenitors, and that many of them remain

654 Delineation of memory B cells
response to the hapten NP uses a large repertoire of heavy chainexpressing analogue VH genes solicit a level of ‘help’ thatgenes. Cell 43:715.results in an early exit from the GC (20,30,45), and limited
14 Blier, P. R. and Bothwell, A. 1987. A limited number of B cellhypermutation and expansion. On the other hand, the V186.2�
lineages generate the heterogeneity of a secondary Immuneclones may experience a prolonged interaction with Th cells response. J. Immunol. 139:3996.
15 Hawkins, R. E. and Winter, G. 1992 Cell selection strategies forleading to extensive proliferation and accumulation of somaticmaking antibodies from variable gene libraries: trapping themutations.memory pool. Eur. J. Immunol. 22:867.
16 Cumano, A. and Rajewsky, K. 1986. Clonal recruitment andsomatic mutation in the generation of immunologic memory to theAcknowledgementshapten NP. EMBO J. 5:2459.
17 Nie, X. Basu, S. and Cerny, J. 1997. Immunization with immuneThis work was supported in part by US Public Health Service GrantsAG-08193 and PO1AG10207. The authors thank Martin Flajnik for complex alters the repertoire of antigen-reactive B cells in thecritical review and editing of the manuscript, and June Green for germinal centers. Eur. J. Immunol. 27:3517.illustrations and manuscript preparation. 18 Jacob, J., Kelsoe, G., Rajewsky, K. and Weiss, U. 1991. Intraclonal
generation of antibody mutants in germinal centers. Nature354:389.
19 McHeyzer-William, G. C., Nossal, G. J. and Lalor, P. A. 1991.AbbreviationsMolecular characterization of single memory B cells. Nature
APFC antibody plaque-forming cells 350:502.BM bone marrow 20 Jacob, J., Przylepa, J., Miller, C. and Kelsoe, G. 1993. In situCGG chicken γ-globulin studies of the primary immune response to (4-hydroxy-3-GC germinal center nitrophenyl) acetyl. III. The kinetics of V-region mutation andNIP iodonitrophenyl selection in germinal centers B cells. J. Exp. Med. 178:1293.NP nitrophenyl 21 Allen, D., Simon, T., Sablitsky, F., Rajewsky, K. and Cumano, A.
1988. Antibody engineering for the analysis of affinity maturationof an anti-hapten response. EMBO J. 7:1995.
References 22 Gu, H., Tarlington, D., Muller, W., Rajewsky, K. and Forster, I.1991. Most peripheral B cells in mice are ligand selected. J. Exp.1 Thorbecke, G. J. 1990. Focusing: the dilemma of interpretingMed. 173:1357.sharp images on a blurred background. J. Immunol. 145:2779.
23 Siekevitz, M., Kocks, C., Rajewsky, K. and Dildrop, R. 1987.2 Linton, P.-J., Decker, D. and Klinman, N. R. 1989. PrimaryAnalysis of somatic mutation and class switching in naive andantibody-forming cells and secondary B cells are generated frommemory B cells generating adoptive primary and secondaryseparate precursor cell subpopulations. Cell 59:1049.responses. Cell 48:757.3 Gearhart, P. J. 1993. Somatic mutation and affinity maturation. In
24 Jacobson, E. B. and Thorbecke, G. J. 1968. Relationship ofPaul, W. E., ed., Fundamental Immunology, p. 865. Raven Press,germinal centers in lymphoid tissue to immunological memory.New York.IV. Formation of 19S and 7S antibody by splenic white and red4 Berek C., Griffiths, G. M. and Milstein, C. 1985. Molecular eventspulp during secondary response in vitro. Lab. Invest. 19:635.during maturation of the immune response to oxazolone. Nature
25 Liu, V.-J., Zhang, J., Lane, P. J. L., Chan, E. Y.-T. and MacLennan,316:412.I. C. M. 1991. Sites of specific B cell activation in primary and5 Metzger, D. W., Miller, A. and Sercarz, E. E. 1981. Idiotypicsecondary responses to T cell-dependent and T cell-independentrepertoire of anti-hen eggwhite lysozyme antibodies probed withantigens. Eur. J. Immunol. 21:2951.hybridomas: selection after immunization of Idx marker common
26 Dresser, D. W. and Wortis, H. H. 1965. A localized homolysis into antibodies of distinct epitope specificity. J. Exp. Med. 154:701.gel method for detection of cells producing 7S antibody: use of6 Busto, P., Gerstein, R., Dupre, L., Giorgetti, C. A., Selsing, E. andantiglobulin serum to detect cells producing antibody with lowPress, J. L. 1987. Molecular analysis of heavy and light chainshemolytic efficiency. Nature 208:858.used by primary and secondary anti-(T,G)-A-L antibodies
27 Benner, R., Meima, F., Vander Meulen, G. M. and van Muiswinkel,produced by normal and xid mice. J. Immunol. 139:608.W. B. 1974. Antibody formation in bone marrow. I. Evidence for7 Kavaler, J., Caton, A. J., Staudt, L. M. and Gerhard, W. 1991. Adevelopment of plaque-forming cells in situ. Immunol. 26:247.B cell population that dominates the primary response to influenza
28 Benner, R. and Haaijman, J. J. 1980. Aging of lymphoid systemvirus hemagglutinin does not participate in the memory response.at the organ level with special reference to the bone marrow asEur. J. Immunol. 21:2687.site of antibody production. Dev. Comp. Immunol. 4:591.8 Minnerath, J. M., Wakem, L. P., Comfort, L. L., Sherman, E. and
29 Benner, R., Van Ovdenaren, A. and de Rviter, H. 1977. AntibodyJemmerson, R. 1995. The BALB/c mouse B-cell response toformation in novel bone marrow. IX. Peripheral lymphoid organspigeon cytochrome c initiates as a heteroclitic response specificare involved in the initiation of bone marrow antibody formation.for the self antigen mouse cytochrome c. Proc. Natl Acad. Sci.Cell. Immunol. 34:125.USA 92:12379.
30 DiLosa, R. M., Maeda, K., Masuda, A., Szakal, A. K. and Tew,9 Weiss, U. and Rajewsky, K. 1990. The repertoire of somaticJ. G. 1991. Germinal center B cells and antibody production inantibody mutants accumulating in the memory compartment afterthe bone marrow. J. Immunol. 146:4071.primary immunization is restricted through affinity maturation
31 Weinberger, J. Z., Green, M. I., Benacerraf, B. and Dorf, M. E.and mirrors that expressed in the secondary response. J. Exp.1979. Hapten-specific T cell responses to 4-hydroxy-3-nitrophenylMed. 172:1681.acetyl. I. Genetic control of delayed-type hypersensitivity by V10 McHeyzer-Williams, M. G., McLean, M. J., Lalor, P. A. and Nossal,and I-A region genes. J. Exp. Med. 149:1136.G. J. V. 1993. Antigen driven B cell differentiation in vivo. J. Exp.
32 Imanishi, T. and Makela, O. 1973. Strain differences in fineMed. 178:295.specificity of mouse anti-hapten antibodies. Eur. J. Immunol.11 Weiss, U, Zoebelein, R. and Rajewsky, K. 1992. Accumulation3:323.of somatic mutants in the B cell compartment after primary
33 Makela, O. and Karjalainen, K. 1978. A Mendelian idiotype isimmunization with a T cell-dependent antigen. Eur. J. Immunol.demonstrable in the heteroclitic anti-NP antibodies of the mouse.22:511.Eur. J. Immunol. 8:105.12 Bothwell, A. L. M., Paskind, M., Reth, M., Imanishi-Kari,
34 Reth, M., Hammerling, G. J. and Rajewsky, K. 1978. Analysis ofT. Rajewsky, K. and Baltimore, D. 1981. Heavy chain variablethe repertoire of anti-NP antibodies in C57BL/6 mice by cellregion contribution to the NPb family of antibodies: somaticfusion. I. Characterization of antibody families in the primary andmutation evident in a γ 2a variable region. Cell 24:625.
13 Maizels, N. and Bothwell, A. 1985. The T-cell-independent immune hyperimmune responses. Eur. J. Immunol. 8:393.

Delineation of memory B cells 655
35 Dal Porto, J. M., Haberman, A. M., Schlomchik, H. J. and Kelsoe, and antibody-forming cell compartments in the primary immuneresponse. EMBO J. 16:2996.G. 1998. Antigen drives very low affinity B cells to become
49 Tao, W., Hardardottir, F. and Bothwell, A. L. M. 1993. Extensiveplasmacytes and enter germinal centers. J. Immunol. 161:5373.somatic mutation in the Ig heavy chain V genes in a late primary36 Eisen, H. N. and McGuigan, J. E. 1971. Quenching of antibodyanti-hapten immune response. Mol. Immunol. 30:593.fluorescence by haptens; a method for determining antibody–
50 Paramithiotis, E. and Cooper, M. D. 1997. Memory B lymphocytesligand affinity. In Williams, C. A. and Chase, M. W., eds, Methodsmigrate to bone marrow in humans. Proc. Natl Acad. Sci. USAin Immunology and Immunochemistry, vol. 3, p. 395. Academic94:208.Press, New York.
51 Price, P. W. and Cerny, J. 1999. Characterization of CD4� T cells37 Herzenberg, L. A., Black, S. J., Tokuhisa, T. and Herzenberg,in mouse bone marrow. I. Increased activated/memory phenotypeL. A. 1980. Memory B cells at successive stage of differentiation.and altered TCR Vβ repertoire. Eur. J. Immunol. 29:1051.Affinity maturation and the role of IgD receptors. J. Exp. Med.
52 Klinman, N. R. 1997. The cellular origins of memory B cells.151:1071.Semin. Immunol. 9:241.38 Yang, X., Stedra, J. and Cerny, J., 1996. Relative contribution of
53 Furukawa, K., Akasako-Furutawa, A., Shirai, H., Nakamura, H.T and B cells to hypermutation and selection of the antibodyand Azuma, T. 1999. Junctional amino acids determine therepertoire in germinal centers of aged mice. J. Exp. Med. 183:959.maturation pathway of an antibody. Immunity 11:329.39 Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S. and Foeller,
54 Shannon, M. and Mehr, R. 1999. Reconciling repertoire shift withC. 1991. Sequences of Proteins of Immunological Interest. NIHaffinity maturation: the role of deleterious mutations. J. Immunol.publ. no. 91. US DHHS, Bethesda, MD.162:3950.40 Manz, R. A., Lohning, M., Cassese, G., Thiel, A. and Radbruch,
55 Berek, C. and Milstein, C. 1987. Mutation drift and repertoire shiftA. 1998. Survival of long-lived plasma cells is independent ofin the maturation of the immune response. Immunol. Rev. 96:23.antigen. Int. Immunol. 10:1703.
56 Berek, C. and Milstein, C. 1988. The dynamic nature of the41 Slifka, M. K., Anha, R., Whitmire, J. K. and Ahmed, R. 1998.antibody repertoire. Immunol. Rev. 105:5.Humoral immunity due to long-lived plasma cells. Immunity 8:363.
57 Rajewsky, K., Forster, I. and Cumano, A. 1987. Evolution and42 Takahashi, Y., Dutta, P. R., Cerasoli, D. M. and Kelsoe, G. 1998. somatic selection of the antibody repertoire in the mouse.In situ studies of the primary immune response to (4-hydroxy-3- Science 238:749.nitrophenyl)acetyl. V. Affinity maturation develop in two stages of 58 Manser, T., Wysocki, L. J., Margolies, M. N. and Gefter, M. L.clonal selection. J. Exp. Med. 187:885. 1987. Evolution of antibody variable region structure during the
43 Decker, D.J., Linton, P.-J., Zaharevitz, S., Biery, M., Gingeras, immune response. Immunol. Rev. 96:141.T. R. and Klinman, N. R. 1995. Defining subsets of naive and 59 Goldbaum, F. A., Cauerhff, A., Velikovsky, C. A., Llera, A. S.,memory B cells based on the ability of their progeny to somatically Riottot, M.-M. and Poljak, R. J. 1999. Lack of significant differencesmutate in vitro. Immunity 2:195. in association rates and affinities of antibodies from short-term and
44 Kallborg, E., Jainandunsing, S., Gray, D. and Leanderson, T. long-term responses to hen egg lysozyme. J. Immunol. 162:6040.1996. Somatic mutation of immunoglobulin genes in vitro. 60 Newman, M. A., Mainhart, C. S., Mallett, C. P., Lavoie T. B. andScience 271:1285. Smith-Gill, S. J. 1992. Patterns of antibody specificity during the
45 Jacob, J. and Kelsoe, G. 1991. In situ studies of the primary BALB/c immune response to hen eggwhite lysozyme.immune response to (4-hydroxy-3-nitrophenyl)acetyl. I. The J. Immunol. 149:3760.architecture and dynamics of responding cell populations. J. Exp. 61 Roost, H. P., Backman, M. F., Haag, A., Kalinke, U., Pliska, V.,Med. 173:1165. Hengartner, H. and Zinkernagel, F. R. 1995. Early high-affinity
46 Betz, A. G., Neuberger, M. S. and Milstein, C. 1993. Discriminating neutralizing anti-viral IgG responses without further overallintrinsic and antigen-selected mutational hot spots in improvements of affinity. Proc. Natl Acad. Sci. USA 92:1257.immunoglobulin V genes. Immunol. Today 14:405. 62 Stockinger, B., Zal, T., Zal, A. and Grey, D. 1996. B cells solicit
47 Rogozin, I. B. and Kolehanov, N. A. 1992. Somatic their own help from T cells. J. Exp. Med. 183:891.hypermutagenesis in immunoglobin genes. II. Influence of 63 Murray, J. S., Pfeiffer, C., Madri, J. and Bottomly, K. 1992. Majorneighboring base sequences on mutagenesis. Biochim. Biophys. histocompatibility complex (MHC) control of CD4 T cell subsetActa 1171:11. activation. II. A single peptide induces either humoral or cell-
48 Smith, K. G. C., Light, A., Nossal, G. J. V. and Tarlinton, D. 1997. mediated responses in mice of distinct MHC genotype. Eur. J.Immunol. 22:559.The extent of affinity maturation differs between the memory




















