
Viruses of hyperthermophilic archaea: entry and egress from ...
Upload
independentCategory
view
3download
0
Ca2+-mediated exocytosis of subtilisin-like protease1: a key step in egress of Plasmodiumfalciparum merozoites
Shalini Agarwal, Maneesh Kumar Singh, Swati Garg,Chetan E. Chitnis* and Shailja Singh*Malaria Group, International Centre for GeneticEngineering and Biotechnology (ICGEB), New Delhi,India.
Summary
Egress of Plasmodium falciparum merozoites fromhost erythrocytes is a critical step in multiplicationof blood-stage parasites. A cascade of proteolyticevents plays a major role in degradation of mem-branes leading to egress of merozoites. However,the signals that regulate the temporal activationand/or secretion of proteases upon maturation ofmerozoites in intra-erythrocytic schizonts remainunclear. Here, we have tested the role of intracel-lular Ca2+ in regulation of egress of P. falciparummerozoites from schizonts. A sharp rise in intrac-ellular Ca2+ just before egress, observed by time-lapse video microscopy, suggested a role forintracellular Ca2+ in this process. Chelation of intra-cellular Ca2+ with chelators such as BAPTA-AMor inhibition of Ca2+ release from intracellularstores with a phospholipase C (PLC) inhibitorblocks merozoite egress. Interestingly, chelationof intracellular Ca2+ in schizonts was also foundto block the discharge of a key protease PfSUB1(subtilisin-like protease 1) from exonemes ofP. falciparum merozoites to parasitophorousvacuole (PV). This leads to inhibition of processingof PfSERA5 (serine repeat antigen 5) and a blockin parasitophorous vacuolar membrane (PVM)rupture and merozoite egress. A complete under-standing of the steps regulating egress of P. falci-parum merozoites may provide novel targets fordevelopment of drugs that block egress and limitparasite growth.
Introduction
The human malaria parasite Plasmodium falciparumcauses the most virulent form of malaria leading to tre-mendous morbidity and mortality (Richards and beeson,2009). All the clinical symptoms of malaria are attributedto the blood stage of the parasite life cycle in which theparasite invades and multiplies within host erythrocytes(Miller et al., 2002a; Cowman and Crabb, 2006). Follow-ing invasion, single-nucleated ring-stage parasites multi-ply within a growth-permissive parasitophorous vacuole(PV) that is enclosed by parasitophorous vacuolar mem-brane (PVM) (Dluzewski et al., 1992; Lingelbach andJoiner, 1998). Daughter merozoites exit from such matureschizonts upon rupture of the PVM and erythrocyte mem-brane and reinvade other erythrocytes to establish a newcycle of infection (Lew, 2005; 2011). The process by whichP. falciparum merozoites exit from infected host erythro-cytes is known as egress. Egress is a tightly regulatedmulti-step process, which involves rapid degradation ofboth the limiting membranes, PVM and erythrocyte mem-brane, leading to release of merozoites (Winograd et al.,1999; Glushakova et al., 2005; 2010; Dvorin et al., 2010;Taylor et al., 2010).
Proteases play a major role in degradation of both PVMand erythrocyte membrane during egress (Salmon et al.,2001; Wickham et al., 2003; Arastu-Kapur et al., 2008;Chandramohanadas et al., 2009; Yeoh et al., 2009). Deg-radation of these limiting membranes requires variousparasite-encoded proteases such as the cytoskeleton-degrading protease falcipain-2 (Dhawan et al., 2003;Sijwali and Rosenthal, 2004; Sijwali et al., 2006), plas-mepsin II (Liu et al., 2005) and a family of nine putativepapain-like proteases called serine repeat antigens(SERA 1–9) (Aoki et al., 2002; Miller et al., 2002b).PfSERA5, an abundant protease of PV, has been shownto be involved in egress of P. falciparum merozoites and isessential for parasite growth (Delplace et al., 1988; Aokiet al., 2002; Miller et al., 2002b). The temporal processingof PfSERA5 late in schizogony is required for the suc-cessful exit of merozoites from PVM and host erythro-cytes (Delplace et al., 1988; Li et al., 2002; Arastu-Kapuret al., 2008; Blackman, 2008; Yeoh et al., 2009). Asubtilisin-family serine protease, PfSUB1, localized inapical organelles of merozoites referred to as exonemes,
Received 14 August, 2012; revised 30 October, 2012; accepted 23November, 2012. *For correspondence. E-mail [email protected];[email protected]; Tel. (+91) 11 2674 1358; Fax (+91) 112674 2316.
Cellular Microbiology (2012) doi:10.1111/cmi.12086
© 2012 Blackwell Publishing Ltd
cellular microbiology

has been reported to mediate processing of PfSERA5(Arastu-Kapur et al., 2008; Blackman, 2008; Yeoh et al.,2009). The signalling events that trigger the secretionof PfSUB1 from exonemes to the PV enabling theprocessing of PfSERA5 and egress of merozoites arenot known.
Elevation of free cytosolic calcium (Ca2+), referred tohere as intracellular Ca2+, has been shown to triggersecretion of proteins from micronemes of P. falciparummerozoites and sporozoites as well as Toxoplasmagondii tachyzoites (Carruthers et al., 1999; Carruthersand Sibley, 1999; Ono et al., 2008; Singh et al., 2010).In this study, we tested whether a rise in levels ofintracellular Ca2+ also regulates discharge of effectormolecules such as PfSUB1 from exonemes of P. falci-parum merozoites leading to egress from mature sch-izonts. We monitored real-time levels of intracellular Ca2+
during late schizogony by time-lapse video microscopy.A significant increase in Ca2+ was observed in schizontsjust before egress suggesting that intracellular Ca2+
might play a role in merozoite egress. Chelation ofextracellular Ca2+ with EGTA had no effect on mero-zoite egress. Chelation of intracellular Ca2+ by treatmentwith BAPTA-AM and treatment with phospholipase C(PLC) inhibitor U73122, which inhibits release of Ca2+
from endoplasmic reticulum (ER), inhibited merozoitesegress. Importantly chelation of intracellular Ca2+ in sch-izonts blocked secretion of PfSUB1 from exonemes tothe PV and inhibited the processing of PfSERA5 andrupture of PVM. This report identifies for the first timethat intracellular Ca2+ levels regulate discharge of a dis-tinct set of apical organelles referred to as, ‘exonemes’,which plays a critical role in merozoite egress frommature schizonts.
Results
Measurement of intracellular Ca2+ levels during egressof P. falciparum merozoites from host erythrocytes
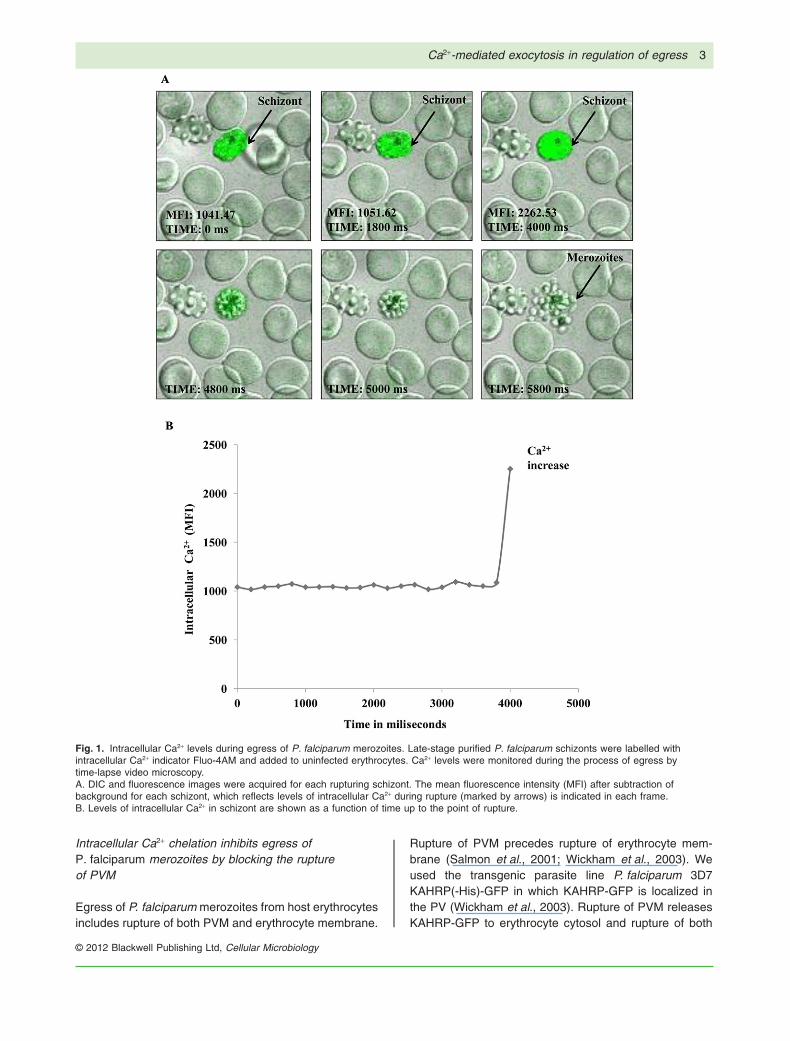
We have investigated real-time intracellular Ca2+ levelsduring egress of P. falciparum merozoites from hosterythrocytes using time-lapse video microscopy. PurifiedP. falciparum schizonts were loaded with Fluo-4AM andobserved under confocal microscope equipped with atemperature-controlled stage and mixed gas environ-ment chamber. Differential interference contrast (DIC)and fluorescence images were captured simultane-ously. Mature schizonts with well-defined symmetricallyarranged merozoites around condensed haemozoin wereobserved before egress (Fig. 1A; Videos S1A–C, S2A–C,S3A–C and S4A–C) as reported earlier (Glushakovaet al., 2010). Few seconds before release, a twofoldincrease in mean fluorescence intensity (MFI) of intracel-
lular Ca2+ in schizonts was observed (Fig. 1A and B;Fig. S1A–C; Videos S1C, S2C, S3C and S4C). In the finalstep, cluster of merozoites with high levels of intracellularCa2+emerged from the ruptured erythrocyte (Fig. 1A,Videos S1A and C, S2A and C, S3A and C, and S4Aand C).
Elevated intracellular Ca2+ levels are required for egressof P. falciparum merozoites
An increase in intracellular Ca2+ late in schizogony priorto egress indicates that Ca2+ might play an essentialrole in egress of P. falciparum merozoites. To study thepotential roles of extracellular Ca2+ and intracellular Ca2+
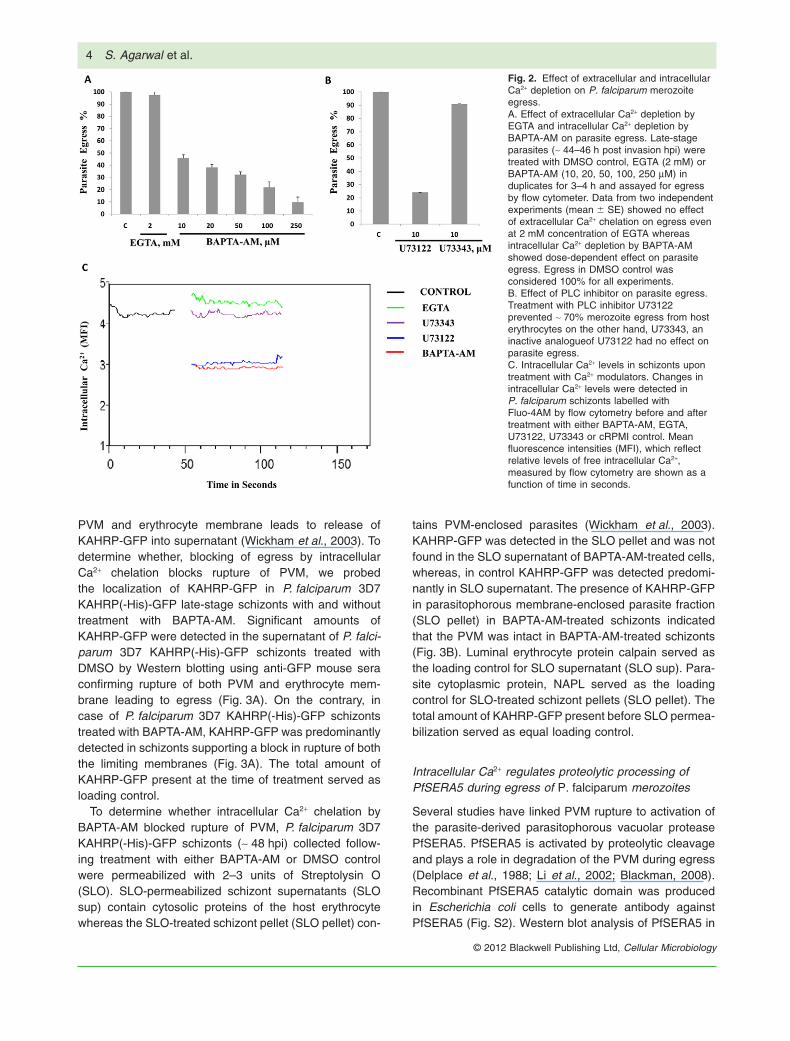
in the parasite egress process, aliquots of late-stageschizonts [~ 44–46 h post invasion (hpi)] were treatedwith either extracellular Ca2+ chelator EGTA or intracel-lular Ca2+ chelator BAPTA-AM, and assayed for egressby detecting rupture of schizonts by flow cytometry.EGTA did not affect merozoite egress from host eryth-rocytes (Fig. 2A). Unlike EGTA, BAPTA-AM inhibitedmerozoite egress from host erythrocytes in a dose-dependent manner (Fig. 2A).
Egress of merozoites from host erythrocytes in absenceof free extracellular Ca2+ indicates that Ca2+ released fromthe ER might be the source of Ca2+ required for egress.Activation of PLC leads to inositol trisphosphate (IP3)generation which in turn triggers release of Ca2+ from theER in P. falciparum-infected erythrocytes (Ross et al.,1989; Beraldo and Garcia, 2005; Vaid et al., 2008; Singhet al., 2010; Alves et al., 2011). To test whether Ca2+
released from the ER upon PLC activation is the source ofintracellular Ca2+ that triggers egress, schizonts weretreated with U73122 (10 mM), a specific inhibitor of PLCand assayed for egress. Treatment of P. falciparum sch-izonts with U73122 prevents merozoite egress from hosterythrocytes (Fig. 2B). On the other hand, pre incubationof P. falciparum schizonts with U73343 (10 mM) an inac-tive analogue of U73122, has no effect on parasite egress(Fig. 2B).
Intracellular Ca2+ levels were measured in P. falci-parum schizonts after treatment with EGTA, BAPTA-AM,U73122, U73343 or DMSO control. On treatment ofschizonts with BAPTA-AM and U73122 the level ofintracellular Ca2+ was reduced ~1.5-fold compared withintracellular Ca2+ levels in control P. falciparum schizontsresuspended in DMSO (Fig. 2C). Intracellular Ca2+
levels in schizonts treated with EGTA and U73343 werefound to be similar to intracellular Ca2+ levels in controlP. falciparum schizonts resuspended in DMSO (Fig. 2C).The reduction in level of Ca2+ in schizonts followingtreatment with PLC inhibitor U73122 suggests that theER is the source of free intracellular Ca2+ that leads toegress.
2 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
Intracellular Ca2+ chelation inhibits egress ofP. falciparum merozoites by blocking the ruptureof PVM
Egress of P. falciparum merozoites from host erythrocytesincludes rupture of both PVM and erythrocyte membrane.
Rupture of PVM precedes rupture of erythrocyte mem-brane (Salmon et al., 2001; Wickham et al., 2003). Weused the transgenic parasite line P. falciparum 3D7KAHRP(-His)-GFP in which KAHRP-GFP is localized inthe PV (Wickham et al., 2003). Rupture of PVM releasesKAHRP-GFP to erythrocyte cytosol and rupture of both
Fig. 1. Intracellular Ca2+ levels during egress of P. falciparum merozoites. Late-stage purified P. falciparum schizonts were labelled withintracellular Ca2+ indicator Fluo-4AM and added to uninfected erythrocytes. Ca2+ levels were monitored during the process of egress bytime-lapse video microscopy.A. DIC and fluorescence images were acquired for each rupturing schizont. The mean fluorescence intensity (MFI) after subtraction ofbackground for each schizont, which reflects levels of intracellular Ca2+ during rupture (marked by arrows) is indicated in each frame.B. Levels of intracellular Ca2+ in schizont are shown as a function of time up to the point of rupture.
Ca2+-mediated exocytosis in regulation of egress 3
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
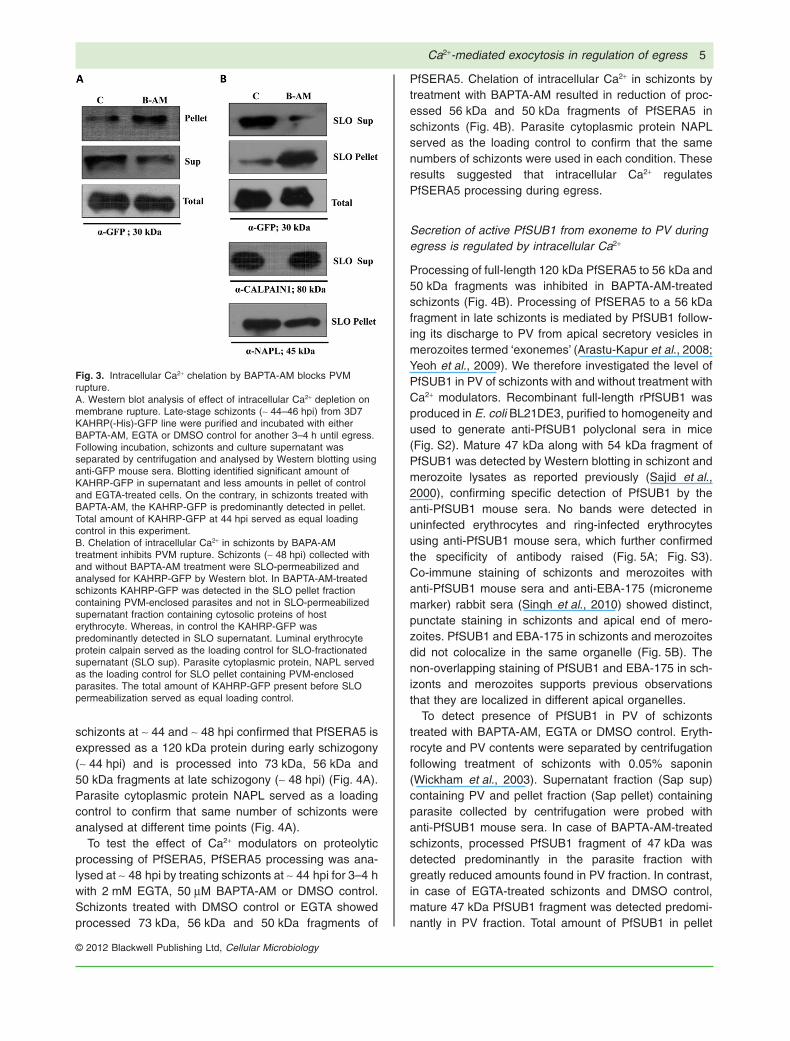
PVM and erythrocyte membrane leads to release ofKAHRP-GFP into supernatant (Wickham et al., 2003). Todetermine whether, blocking of egress by intracellularCa2+ chelation blocks rupture of PVM, we probedthe localization of KAHRP-GFP in P. falciparum 3D7KAHRP(-His)-GFP late-stage schizonts with and withouttreatment with BAPTA-AM. Significant amounts ofKAHRP-GFP were detected in the supernatant of P. falci-parum 3D7 KAHRP(-His)-GFP schizonts treated withDMSO by Western blotting using anti-GFP mouse seraconfirming rupture of both PVM and erythrocyte mem-brane leading to egress (Fig. 3A). On the contrary, incase of P. falciparum 3D7 KAHRP(-His)-GFP schizontstreated with BAPTA-AM, KAHRP-GFP was predominantlydetected in schizonts supporting a block in rupture of boththe limiting membranes (Fig. 3A). The total amount ofKAHRP-GFP present at the time of treatment served asloading control.
To determine whether intracellular Ca2+ chelation byBAPTA-AM blocked rupture of PVM, P. falciparum 3D7KAHRP(-His)-GFP schizonts (~ 48 hpi) collected follow-ing treatment with either BAPTA-AM or DMSO controlwere permeabilized with 2–3 units of Streptolysin O(SLO). SLO-permeabilized schizont supernatants (SLOsup) contain cytosolic proteins of the host erythrocytewhereas the SLO-treated schizont pellet (SLO pellet) con-
tains PVM-enclosed parasites (Wickham et al., 2003).KAHRP-GFP was detected in the SLO pellet and was notfound in the SLO supernatant of BAPTA-AM-treated cells,whereas, in control KAHRP-GFP was detected predomi-nantly in SLO supernatant. The presence of KAHRP-GFPin parasitophorous membrane-enclosed parasite fraction(SLO pellet) in BAPTA-AM-treated schizonts indicatedthat the PVM was intact in BAPTA-AM-treated schizonts(Fig. 3B). Luminal erythrocyte protein calpain served asthe loading control for SLO supernatant (SLO sup). Para-site cytoplasmic protein, NAPL served as the loadingcontrol for SLO-treated schizont pellets (SLO pellet). Thetotal amount of KAHRP-GFP present before SLO permea-bilization served as equal loading control.
Intracellular Ca2+ regulates proteolytic processing ofPfSERA5 during egress of P. falciparum merozoites
Several studies have linked PVM rupture to activation ofthe parasite-derived parasitophorous vacuolar proteasePfSERA5. PfSERA5 is activated by proteolytic cleavageand plays a role in degradation of the PVM during egress(Delplace et al., 1988; Li et al., 2002; Blackman, 2008).Recombinant PfSERA5 catalytic domain was producedin Escherichia coli cells to generate antibody againstPfSERA5 (Fig. S2). Western blot analysis of PfSERA5 in
Fig. 2. Effect of extracellular and intracellularCa2+ depletion on P. falciparum merozoiteegress.A. Effect of extracellular Ca2+ depletion byEGTA and intracellular Ca2+ depletion byBAPTA-AM on parasite egress. Late-stageparasites (~ 44–46 h post invasion hpi) weretreated with DMSO control, EGTA (2 mM) orBAPTA-AM (10, 20, 50, 100, 250 mM) induplicates for 3–4 h and assayed for egressby flow cytometer. Data from two independentexperiments (mean � SE) showed no effectof extracellular Ca2+ chelation on egress evenat 2 mM concentration of EGTA whereasintracellular Ca2+ depletion by BAPTA-AMshowed dose-dependent effect on parasiteegress. Egress in DMSO control wasconsidered 100% for all experiments.B. Effect of PLC inhibitor on parasite egress.Treatment with PLC inhibitor U73122prevented ~ 70% merozoite egress from hosterythrocytes on the other hand, U73343, aninactive analogueof U73122 had no effect onparasite egress.C. Intracellular Ca2+ levels in schizonts upontreatment with Ca2+ modulators. Changes inintracellular Ca2+ levels were detected inP. falciparum schizonts labelled withFluo-4AM by flow cytometry before and aftertreatment with either BAPTA-AM, EGTA,U73122, U73343 or cRPMI control. Meanfluorescence intensities (MFI), which reflectrelative levels of free intracellular Ca2+,measured by flow cytometry are shown as afunction of time in seconds.
4 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
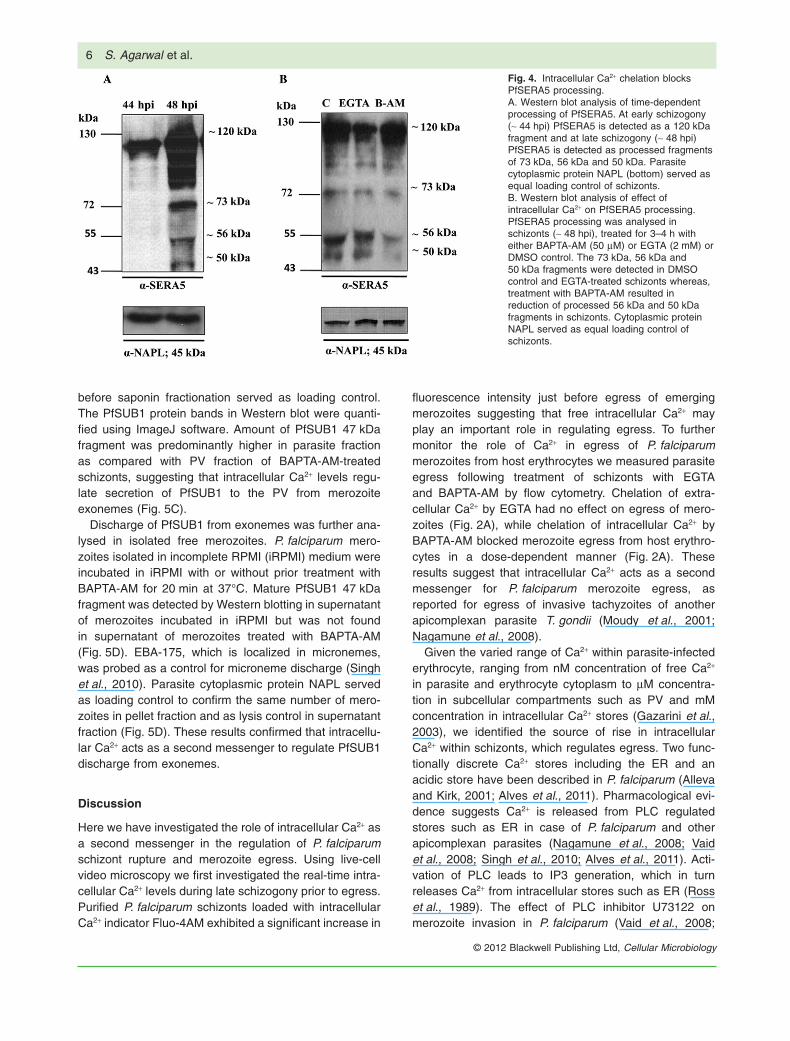
schizonts at ~ 44 and ~ 48 hpi confirmed that PfSERA5 isexpressed as a 120 kDa protein during early schizogony(~ 44 hpi) and is processed into 73 kDa, 56 kDa and50 kDa fragments at late schizogony (~ 48 hpi) (Fig. 4A).Parasite cytoplasmic protein NAPL served as a loadingcontrol to confirm that same number of schizonts wereanalysed at different time points (Fig. 4A).
To test the effect of Ca2+ modulators on proteolyticprocessing of PfSERA5, PfSERA5 processing was ana-lysed at ~ 48 hpi by treating schizonts at ~ 44 hpi for 3–4 hwith 2 mM EGTA, 50 mM BAPTA-AM or DMSO control.Schizonts treated with DMSO control or EGTA showedprocessed 73 kDa, 56 kDa and 50 kDa fragments of
PfSERA5. Chelation of intracellular Ca2+ in schizonts bytreatment with BAPTA-AM resulted in reduction of proc-essed 56 kDa and 50 kDa fragments of PfSERA5 inschizonts (Fig. 4B). Parasite cytoplasmic protein NAPLserved as the loading control to confirm that the samenumbers of schizonts were used in each condition. Theseresults suggested that intracellular Ca2+ regulatesPfSERA5 processing during egress.
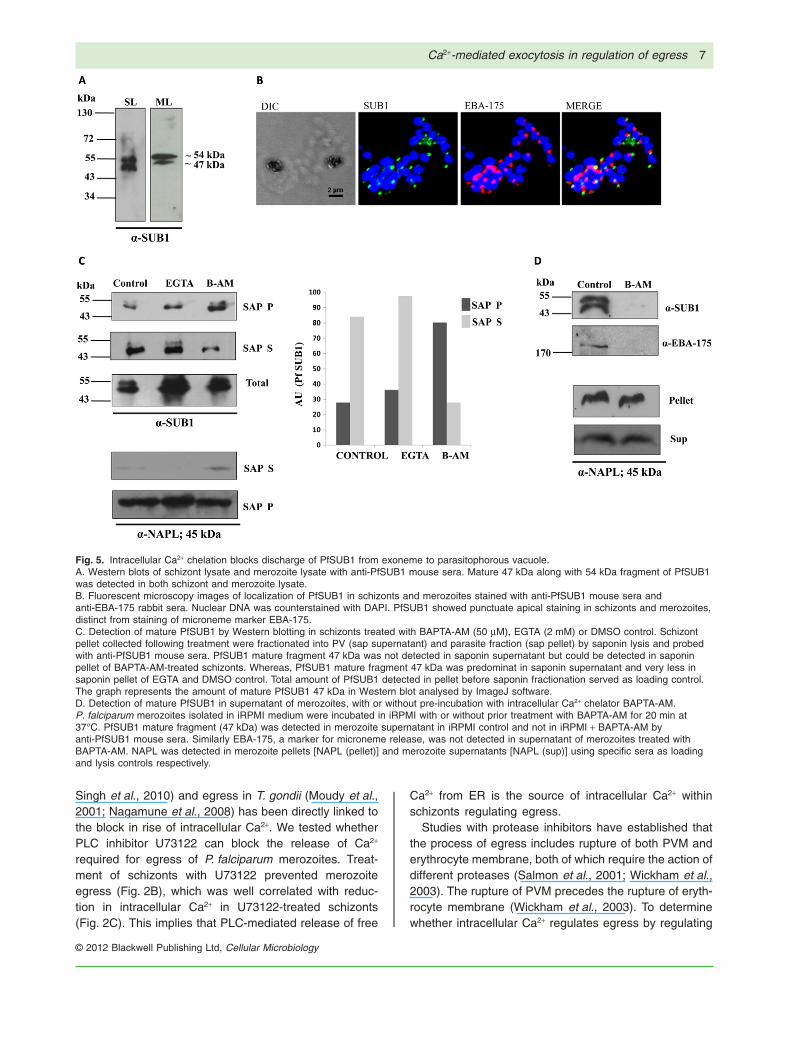
Secretion of active PfSUB1 from exoneme to PV duringegress is regulated by intracellular Ca2+
Processing of full-length 120 kDa PfSERA5 to 56 kDa and50 kDa fragments was inhibited in BAPTA-AM-treatedschizonts (Fig. 4B). Processing of PfSERA5 to a 56 kDafragment in late schizonts is mediated by PfSUB1 follow-ing its discharge to PV from apical secretory vesicles inmerozoites termed ‘exonemes’ (Arastu-Kapur et al., 2008;Yeoh et al., 2009). We therefore investigated the level ofPfSUB1 in PV of schizonts with and without treatment withCa2+ modulators. Recombinant full-length rPfSUB1 wasproduced in E. coli BL21DE3, purified to homogeneity andused to generate anti-PfSUB1 polyclonal sera in mice(Fig. S2). Mature 47 kDa along with 54 kDa fragment ofPfSUB1 was detected by Western blotting in schizont andmerozoite lysates as reported previously (Sajid et al.,2000), confirming specific detection of PfSUB1 by theanti-PfSUB1 mouse sera. No bands were detected inuninfected erythrocytes and ring-infected erythrocytesusing anti-PfSUB1 mouse sera, which further confirmedthe specificity of antibody raised (Fig. 5A; Fig. S3).Co-immune staining of schizonts and merozoites withanti-PfSUB1 mouse sera and anti-EBA-175 (micronememarker) rabbit sera (Singh et al., 2010) showed distinct,punctate staining in schizonts and apical end of mero-zoites. PfSUB1 and EBA-175 in schizonts and merozoitesdid not colocalize in the same organelle (Fig. 5B). Thenon-overlapping staining of PfSUB1 and EBA-175 in sch-izonts and merozoites supports previous observationsthat they are localized in different apical organelles.
To detect presence of PfSUB1 in PV of schizontstreated with BAPTA-AM, EGTA or DMSO control. Eryth-rocyte and PV contents were separated by centrifugationfollowing treatment of schizonts with 0.05% saponin(Wickham et al., 2003). Supernatant fraction (Sap sup)containing PV and pellet fraction (Sap pellet) containingparasite collected by centrifugation were probed withanti-PfSUB1 mouse sera. In case of BAPTA-AM-treatedschizonts, processed PfSUB1 fragment of 47 kDa wasdetected predominantly in the parasite fraction withgreatly reduced amounts found in PV fraction. In contrast,in case of EGTA-treated schizonts and DMSO control,mature 47 kDa PfSUB1 fragment was detected predomi-nantly in PV fraction. Total amount of PfSUB1 in pellet
Fig. 3. Intracellular Ca2+ chelation by BAPTA-AM blocks PVMrupture.A. Western blot analysis of effect of intracellular Ca2+ depletion onmembrane rupture. Late-stage schizonts (~ 44–46 hpi) from 3D7KAHRP(-His)-GFP line were purified and incubated with eitherBAPTA-AM, EGTA or DMSO control for another 3–4 h until egress.Following incubation, schizonts and culture supernatant wasseparated by centrifugation and analysed by Western blotting usinganti-GFP mouse sera. Blotting identified significant amount ofKAHRP-GFP in supernatant and less amounts in pellet of controland EGTA-treated cells. On the contrary, in schizonts treated withBAPTA-AM, the KAHRP-GFP is predominantly detected in pellet.Total amount of KAHRP-GFP at 44 hpi served as equal loadingcontrol in this experiment.B. Chelation of intracellular Ca2+ in schizonts by BAPA-AMtreatment inhibits PVM rupture. Schizonts (~ 48 hpi) collected withand without BAPTA-AM treatment were SLO-permeabilized andanalysed for KAHRP-GFP by Western blot. In BAPTA-AM-treatedschizonts KAHRP-GFP was detected in the SLO pellet fractioncontaining PVM-enclosed parasites and not in SLO-permeabilizedsupernatant fraction containing cytosolic proteins of hosterythrocyte. Whereas, in control the KAHRP-GFP waspredominantly detected in SLO supernatant. Luminal erythrocyteprotein calpain served as the loading control for SLO-fractionatedsupernatant (SLO sup). Parasite cytoplasmic protein, NAPL servedas the loading control for SLO pellet containing PVM-enclosedparasites. The total amount of KAHRP-GFP present before SLOpermeabilization served as equal loading control.
Ca2+-mediated exocytosis in regulation of egress 5
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
before saponin fractionation served as loading control.The PfSUB1 protein bands in Western blot were quanti-fied using ImageJ software. Amount of PfSUB1 47 kDafragment was predominantly higher in parasite fractionas compared with PV fraction of BAPTA-AM-treatedschizonts, suggesting that intracellular Ca2+ levels regu-late secretion of PfSUB1 to the PV from merozoiteexonemes (Fig. 5C).
Discharge of PfSUB1 from exonemes was further ana-lysed in isolated free merozoites. P. falciparum mero-zoites isolated in incomplete RPMI (iRPMI) medium wereincubated in iRPMI with or without prior treatment withBAPTA-AM for 20 min at 37°C. Mature PfSUB1 47 kDafragment was detected by Western blotting in supernatantof merozoites incubated in iRPMI but was not foundin supernatant of merozoites treated with BAPTA-AM(Fig. 5D). EBA-175, which is localized in micronemes,was probed as a control for microneme discharge (Singhet al., 2010). Parasite cytoplasmic protein NAPL servedas loading control to confirm the same number of mero-zoites in pellet fraction and as lysis control in supernatantfraction (Fig. 5D). These results confirmed that intracellu-lar Ca2+ acts as a second messenger to regulate PfSUB1discharge from exonemes.
Discussion
Here we have investigated the role of intracellular Ca2+ asa second messenger in the regulation of P. falciparumschizont rupture and merozoite egress. Using live-cellvideo microscopy we first investigated the real-time intra-cellular Ca2+ levels during late schizogony prior to egress.Purified P. falciparum schizonts loaded with intracellularCa2+ indicator Fluo-4AM exhibited a significant increase in
fluorescence intensity just before egress of emergingmerozoites suggesting that free intracellular Ca2+ mayplay an important role in regulating egress. To furthermonitor the role of Ca2+ in egress of P. falciparummerozoites from host erythrocytes we measured parasiteegress following treatment of schizonts with EGTAand BAPTA-AM by flow cytometry. Chelation of extra-cellular Ca2+ by EGTA had no effect on egress of mero-zoites (Fig. 2A), while chelation of intracellular Ca2+ byBAPTA-AM blocked merozoite egress from host erythro-cytes in a dose-dependent manner (Fig. 2A). Theseresults suggest that intracellular Ca2+ acts as a secondmessenger for P. falciparum merozoite egress, asreported for egress of invasive tachyzoites of anotherapicomplexan parasite T. gondii (Moudy et al., 2001;Nagamune et al., 2008).
Given the varied range of Ca2+ within parasite-infectederythrocyte, ranging from nM concentration of free Ca2+
in parasite and erythrocyte cytoplasm to mM concentra-tion in subcellular compartments such as PV and mMconcentration in intracellular Ca2+ stores (Gazarini et al.,2003), we identified the source of rise in intracellularCa2+ within schizonts, which regulates egress. Two func-tionally discrete Ca2+ stores including the ER and anacidic store have been described in P. falciparum (Allevaand Kirk, 2001; Alves et al., 2011). Pharmacological evi-dence suggests Ca2+ is released from PLC regulatedstores such as ER in case of P. falciparum and otherapicomplexan parasites (Nagamune et al., 2008; Vaidet al., 2008; Singh et al., 2010; Alves et al., 2011). Acti-vation of PLC leads to IP3 generation, which in turnreleases Ca2+ from intracellular stores such as ER (Rosset al., 1989). The effect of PLC inhibitor U73122 onmerozoite invasion in P. falciparum (Vaid et al., 2008;
Fig. 4. Intracellular Ca2+ chelation blocksPfSERA5 processing.A. Western blot analysis of time-dependentprocessing of PfSERA5. At early schizogony(~ 44 hpi) PfSERA5 is detected as a 120 kDafragment and at late schizogony (~ 48 hpi)PfSERA5 is detected as processed fragmentsof 73 kDa, 56 kDa and 50 kDa. Parasitecytoplasmic protein NAPL (bottom) served asequal loading control of schizonts.B. Western blot analysis of effect ofintracellular Ca2+ on PfSERA5 processing.PfSERA5 processing was analysed inschizonts (~ 48 hpi), treated for 3–4 h witheither BAPTA-AM (50 mM) or EGTA (2 mM) orDMSO control. The 73 kDa, 56 kDa and50 kDa fragments were detected in DMSOcontrol and EGTA-treated schizonts whereas,treatment with BAPTA-AM resulted inreduction of processed 56 kDa and 50 kDafragments in schizonts. Cytoplasmic proteinNAPL served as equal loading control ofschizonts.
6 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
Singh et al., 2010) and egress in T. gondii (Moudy et al.,2001; Nagamune et al., 2008) has been directly linked tothe block in rise of intracellular Ca2+. We tested whetherPLC inhibitor U73122 can block the release of Ca2+
required for egress of P. falciparum merozoites. Treat-ment of schizonts with U73122 prevented merozoiteegress (Fig. 2B), which was well correlated with reduc-tion in intracellular Ca2+ in U73122-treated schizonts(Fig. 2C). This implies that PLC-mediated release of free
Ca2+ from ER is the source of intracellular Ca2+ withinschizonts regulating egress.
Studies with protease inhibitors have established thatthe process of egress includes rupture of both PVM anderythrocyte membrane, both of which require the action ofdifferent proteases (Salmon et al., 2001; Wickham et al.,2003). The rupture of PVM precedes the rupture of eryth-rocyte membrane (Wickham et al., 2003). To determinewhether intracellular Ca2+ regulates egress by regulating
Fig. 5. Intracellular Ca2+ chelation blocks discharge of PfSUB1 from exoneme to parasitophorous vacuole.A. Western blots of schizont lysate and merozoite lysate with anti-PfSUB1 mouse sera. Mature 47 kDa along with 54 kDa fragment of PfSUB1was detected in both schizont and merozoite lysate.B. Fluorescent microscopy images of localization of PfSUB1 in schizonts and merozoites stained with anti-PfSUB1 mouse sera andanti-EBA-175 rabbit sera. Nuclear DNA was counterstained with DAPI. PfSUB1 showed punctuate apical staining in schizonts and merozoites,distinct from staining of microneme marker EBA-175.C. Detection of mature PfSUB1 by Western blotting in schizonts treated with BAPTA-AM (50 mM), EGTA (2 mM) or DMSO control. Schizontpellet collected following treatment were fractionated into PV (sap supernatant) and parasite fraction (sap pellet) by saponin lysis and probedwith anti-PfSUB1 mouse sera. PfSUB1 mature fragment 47 kDa was not detected in saponin supernatant but could be detected in saponinpellet of BAPTA-AM-treated schizonts. Whereas, PfSUB1 mature fragment 47 kDa was predominat in saponin supernatant and very less insaponin pellet of EGTA and DMSO control. Total amount of PfSUB1 detected in pellet before saponin fractionation served as loading control.The graph represents the amount of mature PfSUB1 47 kDa in Western blot analysed by ImageJ software.D. Detection of mature PfSUB1 in supernatant of merozoites, with or without pre-incubation with intracellular Ca2+ chelator BAPTA-AM.P. falciparum merozoites isolated in iRPMI medium were incubated in iRPMI with or without prior treatment with BAPTA-AM for 20 min at37°C. PfSUB1 mature fragment (47 kDa) was detected in merozoite supernatant in iRPMI control and not in iRPMI + BAPTA-AM byanti-PfSUB1 mouse sera. Similarly EBA-175, a marker for microneme release, was not detected in supernatant of merozoites treated withBAPTA-AM. NAPL was detected in merozoite pellets [NAPL (pellet)] and merozoite supernatants [NAPL (sup)] using specific sera as loadingand lysis controls respectively.
Ca2+-mediated exocytosis in regulation of egress 7
© 2012 Blackwell Publishing Ltd, Cellular Microbiology

rupture of PVM, we used the transgenic P. falciparum line3D7 KAHRP(-His)-GFP in which KAHRP-GFP is localizedin the PV and is released to the erythrocyte cytosol fol-lowing rupture of PVM and is finally released in the culturesupernatant following rupture of erythrocyte membrane.KAHRP-GFP accumulates in the PV following treatmentof the 3D7 KAHRP(-His)-GFP schizonts with BAPTA-AMindicating that the PVM is intact in these schizonts(Fig. 3B). Several studies have linked the processingof the parasite-derived parasitophorous vacuolar pro-tease PfSERA5 with proteolysis of PVM during egress(Delplace et al., 1988; Blackman, 2008). PfSERA5 isexpressed as a 120 kDa precursor protein that is furtherprocessed to 73 kDa, 56 kDa and finally to 50 kDa formthat is released during host cell rupture (Li et al., 2002).We found that inhibition of P. falciparum merozoite egressby BAPTA-AM treatment resulted in inhibition of PfSERA5processing to 56 and 50 kDa fragments (Fig. 4B). Sincewe observed PfSERA5 processing being blocked at the56 kDa intermediate which has a proteolytic role in egress(Blackman, 2008), and PfSUB1 mediates this processing(Arastu-Kapur et al., 2008; Yeoh et al., 2009) we furthertested levels of PfSUB1 in the PV in BAPTA-AM-treatedschizonts.
Previous studies suggested that PfSUB1 resides ina set of apical subcellular secretory vesicles termed‘exonemes’, which are distinct from other apicalorganelle such as micronemes, rhoptries and densegranules (Arastu-Kapur et al., 2008; Yeoh et al., 2009).Western blot analysis of mature bands of PfSUB1 in thePV following treatment of schizonts with either BAPTA-AM, EGTA or DMSO control, showed a reduction inPfSUB1 mature fragment (~ 47 kDa) in PV of BAPTA-AM-treated schizonts (Fig. 5C). This suggested that che-lation of intracellular ca2+ blocks the secretion of activePfSUB1 from exonemes to the PV which in turn inhibitsprocessing of PfSERA5. This result demonstrates thatrise in intracellular Ca2+ triggers secretion of PfSUB1from a distinct set of apical organelles referred to asexonemes which is a critical step in P. falciparum mero-zoites egress.
The Ca2+-dependent discharge of PfSUB1 fromexonemes was further validated in isolated P. falciparummerozoites, where, PfSUB1 was detected in supernatantsof merozoites resuspended in iRPMI but, not in mero-zoites pre-incubated with BAPTA-AM (Fig. 5D). Treatmentwith BAPTA-AM also inhibited secretion of micronemeprotein EBA-175 from P. falciparum merozoites. Theseresults suggest that exoneme discharge is regulated byintracellular Ca2+ as shown previously for discharge ofmicronemes (Singh et al., 2010). A clear understanding ofthe steps that regulate merozoite egress at the molecularlevel can lead to the identification of novel drug targets toinhibit parasite growth.
Experimental procedures
Parasite lines and in vitro culture
Plasmodium falciparum 3D7 was cultured in O+ erythrocytes, withRPMI 1640 (Invitrogen, USA) supplemented with 25 mM Hepes(Invitrogen, USA), 4.5 mg ml-1glucose (Sigma, USA), 0.1 mMhypoxanthine (Invitrogen, USA), 25 mg ml-1 gentamicin (Invitro-gen, USA) and 0.5% AlbuMax I (Invitrogen, USA), according tomethods described earlier (Trager and Jensen, 1976). Parasiteculture was maintained in mixed gas environment (5% O2, 5%CO2 and 90% N2). Parasites were synchronized by sorbitol treat-ment at ring stage. The synchronized ring-stage parasites werecultivated to yield schizonts (~ 44–46 hpi) and used for egressexperiments. P. falciparum 3D7 KAHRP(-His)-GFP (Kindlyprovided by A.F. Cowman, University of Melbourne, Melbourne3050, Australia) was also maintained as described above andused for egress experiments.
Imaging Ca2+ fluxes during P. falciparum merozoitesegress from host erythrocyte by live-cellvideo microscopy
Purified P. falciparum schizonts were loaded with 10 mM intrac-ellular Ca2+ indicator Fluo-4AM in complete RPMI (cRPMI) for20 min at 37°C in mixed gas environment, mixed with unin-fected erythrocytes in a Dvorak chamber and observed with aconfocal microscope (Zeiss LSM510) equipped with atemperature-controlled stage. Laser excitation was provided ata wavelength of 488 nm and fluorescent emissions were col-lected at wavelengths above 515 nm using 505–550 band passfilter. DIC and fluorescence images were captured duringegress of P. falciparum merozoites using a 63¥, 1.4 NA lens.Twelve-bit time-lapse images of egress events were capturedand MFI, which reflects relative levels of free intracellular Ca2+
within regions of interest containing mature schizont in the focalplane was determined after background subtraction usingImage Pro software (Singh et al., 2010). MFI for Ca2+ levels inschizonts till the frame of egress was then plotted as a functionof time in seconds.
Parasite egress assay
To assess the effect of Ca2+ modulating agents on merozoiteegress, aliquots of late-stage schizonts (~ 44–46 hpi) at afinal haematocrit of 0.5% and parasitaemia of 8–10% weretreated with 2 mM EGTA [Ethyleneglycol-bis(b-aminoethyl)-N,N,N′,N′-tetracetic Acid], 10, 20, 50, 100, 250 mM BAPTA-AM[1,2-bis(o-Aminophenoxy)ethane-N,N,N′,N′-tetraaceticacid Tetra(acetoxymethyl) Ester] and 10 mM PLC inhibitor U73122 and 10mM U73343, an inactive analogue of U73122 (Calbiochem, USA)in duplicate wells. DMSO-treated schizonts were used as con-trols for each experiment. Following treatment with Ca2+ modu-lating agents, P. falciparum schizonts were incubated for 3–4 h toallow egress and remaining parasites were then collected,washed and stained with ethidium bromide (10 mg ml-1). Sch-izonts collected at initial 0 h time point were also stained. Theethidium bromide-stained parasites were analysed by flowcytometry on FACSCalibur (Becton Dickinson, USA) using Cel-lQuest software. Fluorescence signal (FL-2) was detected with
8 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology

590 nM band pass filter using an excitation laser of 488 nmcollecting typically 10 0000 cells per sample. Following acquisi-tion data were analysed using FlowJo software (Tree Star,Ashland) by determining the proportion of FL-2-positive cellsrepresenting schizonts. Parasite egress % was calculated as theper cent of schizonts ruptured in treatment and control during theincubation time as compared with the initial number of schizontsat 0 h, using the formula described below. Parasite egress % wasthen plotted considering per cent of schizonts ruptured in controlas 100% egress.
Parasite Egress I T I C% = −( ) −( )[ ]100
I: % of schizonts before treatment; T: % of schizonts aftertreatment with inhibitors; C: % of schizont after treatmentwith DMSO control.
Measurement of intracellular Ca2+ in P. falciparumschizonts on treatment with Ca2+ modulating agents byflow cytometry
Intracellular Ca2+ levels in P. falciparum schizonts labelledwith Fluo-4AM after treatment with EGTA, BAPTA-AM, U73122,U73343 or DMSO control were measured by flow cytometry withsome modifications of the method described previously for meas-urement of Ca2+ levels in merozoites (Singh et al., 2010; Singhand Chitnis, 2013). P. falciparum schizonts (~ 46–48 hpi) wereisolated using Miltenyi Biotec magnetic separator (MiltenyiBiotec, USA) and loaded with 10 mM Fluo-4AM for 20 min at 37°Cin mixed gas environment. Fluo-4AM loaded schizonts were thenincubated with either 2 mM EGTA (Calbiochem, USA), 50 mMBAPTA-AM (Calbiochem, USA), 10 mM U73122 or 10 mMU73343 (Calbiochem, USA) for 50–60 min at 37°C. Fluores-cence signal (Fl-1) was then measured on flow cytometer (FAC-SCalibur, Becton Dickinson, USA), with a 430/30 nm band passfilter using an excitation laser of 488 nm for a period of 1 min.
Following acquisition, data were analysed using FlowJo soft-ware (Tree Star, Ashland) by determining the proportion of FL-1-positive cells. Kinetics of MFI for FL-1-positive cells, whichreflects intracellular Ca2+ levels in schizonts, was plotted againsttime for each treatment conditions as described previously (Singhand Chitnis, 2013).
SLO fractionation and immunoblot analysis
Plasmodium falciparum 3D7 KAHRP(-His)-GFP schizonts (~ 44–46 hpi) were purified on a magnetic column (Miltenyi Biotec,USA) and incubated with either 50 mM BAPTA-AM or DMSOcontrol for 3–4 h to allow egress. Following treatment, pelletcontaining schizonts and culture supernatant were separatedby centrifugation. Samples were resuspended in SDS-PAGEsample buffer and separated on 12% SDS-PAGE reducing gelsand transferred to 0.45 mm nitrocellulose membrane (Amersham,USA), blocked in 5% skimmed milk in PBS overnight at 4°C andprobed with anti-GFP mouse sera (Roche, USA) at 1:1000 dilu-tion and horseradish peroxidase-conjugated goat anti-mouse IgGantibodies (Sigma Aldrich, USA) at 1:2000 dilution. The blot wasdeveloped using ECL Plus Western Blotting Detection Reagents(GE Healthcare, USA) using manufacturer’s protocol. The sch-izonts at the time of treatment (~ 44 hpi) were also collected andprobed for total amount of KAHRP-GFP.
For detection of KAHRP-GFP released to erythrocyte cytosol,BAPTA-AM treated as well as untreated schizonts (~ 48 hpi)were incubated with 3–4 haemolytic units of SLO in iRPMI for6 min at room temperature as described previously (Ansorgeet al., 1996; Wickham et al., 2003). SLO-permeabilized superna-tant containing cytosolic fraction of host erythrocyte and SLOpellet containing PVM-enclosed parasites were separated bycentrifugation and analysed by Western blot as described above.Rabbit sera against luminal erythrocyte protein calpain1 (CAPN1;Sigma, USA) was used at a dilution of 1:500 as control forSLO-fractionated erythrocyte cytosol (SLO sup). Rabbit seraagainst the P. falciparum cytoplasmic protein NAPL was used ata dilution of 1:2000 as a control for parasite in SLO pellet(Wickham et al., 2003; Chandra et al., 2005).
Cloning, expression and purification of PfSERA5 andPfSUB1 and generation of specific antisera
PfSERA5 catalytic domain (amino acid residues 578–828) wasPCR-amplified from cDNA of P. falciparum 3D7 parasites byusing the primer pair: forward 5′-GGCGCGGATCCGAAAATAATTGTATATCTAATCTTCAAGTTG-3′ and reverse 5′-GGCCGCTCGAGCTAATTATTCATAGGTAAATCAACATTGAATATAAC-3′and cloned in pProExHTb vector (Invitrogen, USA) betweenBamHI and XhoI sites and transferred to E. coli BL21-DE3(Novagen, USA) to express rPfSERA5 as N-terminal 6-His tagprotein. The recombinant E. coli cells were grown to mid-logphase and expression of rPfSERA5 was induced at OD600 of 0.5with 1 mM isopropyl b-D-1-thiogalactopyranoside (IPTG) for 4 h.E. coli cells were collected by centrifugation, lysed by sonicationand N-terminal 6-His tagged rPfSERA5 was purified from thesolubilized inclusion bodies by metal affinity chromatographyusing Ni-NTA matrix (Qiagen, the Netherlands) to homogeneityand used for immunization.
A codon optimized synthetic gene encoding the PfSUB1(amino acid residues 27–688) fused to a C-terminal 6-His tag wascloned in the expression vector pET28a and transferred to E. coliBLR-DE3 (Novagen, USA). The recombinant E. coli cells weregrown to mid-log phase and expression of rPfSUB1 was inducedat OD600 of 0.7 with 1 mM IPTG for 4 h. E. coli cells were col-lected by centrifugation, lysed by sonication and rPfSUB1 waspurified from the solubilize inclusion bodies by metal affinity chro-matography using Ni-NTA matrix (Qiagen, the Netherlands) andused for immunization.
To generate PfSERA5 and PfSUB1 antisera, 5- to 6-week-oldfemale BALB/c mice were immunized with 25 mg of his-fusionrPfSERA5 and rPfSUB1 emulsified in Freund’s complete adju-vant on day 0 followed by two boosts emulsified with Freund’sincomplete adjuvant on days 28 and 56. The animals were bledfor serum collection on day 70.
Immunofluorescence assay
For detection of PfSUB1 by immunofluorescence assays (IFA),synchronized P. falciparum cultures containing schizonts andmerozoites were smeared on glass slides, dried, fixed withmethanol and probed with anti-PfSUB1 mouse sera diluted 1:200and anti-EBA-175 (microneme marker) (Sim et al., 1992), rabbitsera diluted 1:50. Slides were washed three times with PBS andincubated for 1 h with Alexa 488-conjugated goat anti-mouse IgG
Ca2+-mediated exocytosis in regulation of egress 9
© 2012 Blackwell Publishing Ltd, Cellular Microbiology

antibodies diluted 1:200 or Alexa-Fluor 594-conjugated goat anti-rabbit IgG antibodies diluted 1:500 and mounted with DAPI Anti-fade reagent (Molecular Devices, USA) for nuclear staining.Images were acquired in confocal microscope (A1R, Nikon,Japan) using NIS element software.
Detection of PfSERA5 processing byimmunoblot analysis
Plasmodium falciparum 3D7 schizonts at ~ 44 hpi and ~ 48 hpiwere probed for SERA5 processing. Schizonts were purified on amagnetic column (Miltenyi Biotec, USA) at ~ 44 hpi and ~ 48 hpi,collected by centrifugation and resuspended in SDS-PAGEsample buffer. Proteins were separated on 12% SDS-PAGEreducing gels and subjected to immunoblotting using anti-PfSERA5 mouse sera at 1:500 dilution and horseradishperoxidase-conjugated goat anti-mouse IgG antibodies (SigmaAldrich, USA) at 1:2000 dilution. To detect the effect of Ca2+
modulators on PfSERA5 processing, purified schizonts (~ 44 hpi)were treated with either 2 mM EGTA, 50 mM BAPTA-AM orDMSO control for another 3–4 h and analysed at (~ 48 hpi) byWestern blotting for PfSERA5 processing. Anti-NAPL rabbit serawere used at a dilution of 1:2000, as a control for equal numberof schizonts in each condition.
Detection of PfSUB1 by immunoblot analysis
Plasmodium falciparum schizonts (~ 48 hpi), ring-stage parasites(~ 14–16 hpi) and merozoites were collected by centrifugation asdescribed previously (Singh et al., 2010). Schizonts, merozoites,rings and uninfected erythrocytes were then resuspended inSDS-PAGE sample buffer and separated on 12% SDS-PAGEreducing gels and subjected to immunoblotting using anti-PfSUB1 mouse sera at 1:500 dilution and horseradishperoxidase-conjugated goat anti-mouse IgG antibodies (SigmaAldrich, USA) at 1:2000 dilution.
To detect the effect of Ca2+ modulators on PfSUB1 dischargefrom exoneme to PV, purified schizonts (~ 44–46 hpi) weretreated with 2 mM EGTA, 50 mM BAPTA-AM or DMSO controlfor another 3–4 h. Schizonts were then collected and fraction-ated by 0.05% saponin into PV (saponin sup) and parasite(saponin pellet). Saponin-fractionated sup and pellet as well asschizonts before saponin fractionation for each condition wereanalysed by Western blotting for the presence of PfSUB1 asdescribed above.
For analysis of discharge of PfSUB1 in merozoites, P. falci-parum merozoites were isolated and incubated for 20 min at37°C with either 50 mM BAPTA-AM or DMSO control in iRPMI.Supernatants were separated by centrifugation and used fordetection of exoneme protein PfSUB1, using anti-PfSUB1 mousesera or microneme protein EBA-175 using anti-EBA-175 rabbitsera (Singh et al., 2010). Anti-NAPL rabbit sera were used at adilution of 1:2000 as loading and lysis control for merozoitepellets and merozoite supernatants for each condition.
Acknowledgements
This work was supported by Innovative Young BiotechnologyAward (IYBA) grant, from the Department of Biotechnology (DBT)Govt. of India, Program Support Grant from DBT, MalSig and
EVIMalaR grants from the European Commission. S.S. is arecipient of the Innovative Young Biotechnologist Award. C.E.C.is a recipient of the TATA Innovation Fellowship from DBT. Theauthors thank: Dr Alan Cowman, Walter and Eliza Hall Institutefor Medical Research, Melbourne for P. falciparum 3D7(-His)-GFP parasite line; Dr Amit Sharma, ICGEB, New Delhi for anti-NAPL sera and Prof. Virander S. Chauhan, Director, ICGEB, forproviding PfSERA-5 construct used for PfSERA5 protein produc-tion. We also thank ICGEB for access to confocal microscopyfacility and IYBA grant support for upgrading confocal micro-scope with live-cell imaging facility. S.A. was supported by DBTand M.K.S. was supported by IYBA JRF.
References
Alleva, L.M., and Kirk, K. (2001) Calcium regulation in theintraerythrocytic malaria parasite Plasmodium falciparum.Mol Biochem Parasitol 117: 121–128.
Alves, E., Bartlett, P.J., Garcia, C.R., and Thomas, A.P.(2011) Melatonin and IP3-induced Ca2+ release from intra-cellular stores in the malaria parasite Plasmodium falci-parum within infected red blood cells. J Biol Chem 286:5905–5912.
Ansorge, I., Benting, J., Bhakdi, S., and Lingelbach, K. (1996)Protein sorting in Plasmodium falciparum-infected redblood cells permeabilized with the pore-forming proteinstreptolysin O. Biochem J 315: 307–314.
Aoki, S., Li, J., Itagaki, S., Okech, B.A., Egwang, T.G., Mat-suuoka, H., et al. (2002) Serine repeat antigen (SERA5) ispredominantly expressed among the SERA multigenefamily of Plasmodium falciparum and the acquired antibodytiters correlate with serum inhibition of the parasite growth.J Biol Chem 277: 47533–47540.
Arastu-Kapur, S., Ponder, E.L., Fonovic, U.P., Yeoh, S.,Yuan, F., Fonovic, M., et al. (2008) Identification of pro-teases that regulate erythrocyte rupture by the malariaparasite Plasmodium falciparum. Nature Chem Biol 4:203–213.
Beraldo, F.H., and Garcia, C.R.S. (2005) Products of tryp-tophan catabolism induce Calcium release and modulatethe cell cycle of Plasmodium falciparum malaria parasites.J Pineal Res 39: 224–230.
Blackman, M.J. (2008) Malarial proteases and host cellegress: an ‘emerging’ cascade. Cell Microbiol 10: 1925–1934.
Carruthers, V.B., and Sibley, L.D. (1999) Mobilization ofintracellular Calcium stimulates microneme discharge inToxoplasma gondii. Mol Microbiol 31: 421–428.
Carruthers, V.B., Giddings, O.K., and Sibley, L.D. (1999)Secretion of micronemal proteins is associated withToxoplasma invasion of host cells. Cell Microbiol 1: 225–236.
Chandra, B.R., Olivieri, A., Silvestrini, F., Alano, P., andSharma, A. (2005) Biochemical characterization of the twonucleosome assembly proteins from Plasmodium falci-parum. Mol Biochem Parasitol 142: 237–247.
Chandramohanadas, R., Davis, P.H., Beiting, D.P., Harbut,M.B., Darling, C., Velmourougane, G., et al. (2009) Api-complexan parasites co-opt host calpains to facilitate theirescape from infected cells. Science 314: 794–797.
Cowman, A.F., and Crabb, B.S. (2006) Invasion of red bloodcells by malaria parasites. Cell 124: 755–766.
10 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology

Delplace, P., Bhatia, A., Cagnard, M., Camus, D., Colombet,G., Debrabant, A., et al. (1988) Protein p126: a parasito-phorous vacuole antigen associated with the release ofPlasmodium falciparum merozoites. Biol Cell 64: 215–221.
Dhawan, S., Dua, M., Chishti, A.H., and Hanspal, M. (2003)Ankyrin peptide blocks falcipain-2-mediated malaria para-site release from red blood cells. J Biol Chem 278: 30180–30186.
Dluzewski, A.R., Mitchell, G.H., Fryer, P.R., Griffiths, S.,Wilson, R.J., and Gratzer, W.B. (1992) Origins of the para-sitophorous vacuole membrane of the malaria parasite,Plasmodium falciparum, in human red blood cells. J CellSci 102: 527–532.
Dvorin, J.D., Martyn, D.C., Patel, S.D., Grimley, J.S., Collins,C.R., Hopp, C.S., et al. (2010) A plant-like kinase in Plas-modium falciparum regulates parasite egress from eryth-rocytes. Science 328: 910–912.
Gazarini, M.L., Thomas, A.P., Pozzan, T., and Garcia, C.R.S.(2003) Calcium signaling in a low Calcium environment:how the intracellular malaria parasite solves the problem.J Cell Biol 161: 103–110.
Glushakova, S., Yin, D., Li, T., and Zimmerberg, J.(2005) Membrane transformation during malaria parasiterelease from human red blood cells. Curr Biol 15: 1645–1650.
Glushakova, S., Humphrey, G., Leikina, E., Balaban, A.,Miller, J., and Zimmerberg, J. (2010) New stages in theprogram of malaria parasite egress imaged in normal andsickle erythrocytes. Curr Biol 20: 1117–1121.
Lew, V.L. (2005) Malaria: endless fascination with merozoiterelease. Curr Biol 15: R760–R761.
Lew, V.L. (2011) Malaria: surprising mechanism of merozoiteegress revealed. Curr Biol 21: R314–R316.
Li, J., Matsuoka, H., Mitamura, T., and Horii, T. (2002) Char-acterization of proteases involved in the processing ofPlasmodium falciparum serine repeat antigen (SERA). MolBiochem Parasitol 120: 177–186.
Lingelbach, K., and Joiner, K.A. (1998) The parasitophorousvacuole membrane surrounding Plasmodium and Toxo-plasma: an unusual compartment in infected cells. J CellSci 111: 1467–1475.
Liu, J., Gluzman, I.Y., Drew, M.E., and Goldberg, D.E. (2005)The role of Plasmodium falciparum food vacuole plasmep-sins. J Biol Chem 280: 1432–1437.
Miller, L.H., Baruch, D.I., Marsh, K., and Doumbo, O.K.(2002a) The pathogenic basis of malaria. Nature 415: 673–679.
Miller, S.K., Good, R.T., Drew, D.R., Delorenzi, M., Sanders,P.R., Hodder, A.N., et al. (2002b) A subset of Plasmodiumfalciparum SERA genes are expressed and appear to playan important role in the erythrocytic cycle. J Biol Chem277: 47524–47532.
Moudy, R., Manning, T.J., and Beckers, C.J. (2001) The lossof cytoplasmic potassium upon host cell breakdown trig-gers egress of Toxoplasma gondii. J Biol Chem 276:41492–41501.
Nagamune, K., Moreno, S.N., Chini, E.N., and Sibley, L.D.(2008) Calcium regulation and signaling in apicomplexanparasites. Subcell Biochem 47: 70–81.
Ono, T., Cabrita-Santos, L., Leitao, R., Bettiol, E., Purcell,L.A., Diaz-Pulido, O., et al. (2008) Adenylyl cyclase alpha
and cAMP signaling mediate Plasmodium sporozoiteapical regulated exocytosis and hepatocyte infection. PLoSPathog 4: e1000008.
Richards, J.S., and Beeson, J.G. (2009) The future foblood-stage vaccines against malaria. Immunol Cell Biol87: 377–390.
Ross, C.A., Meldolesi, J., Milner, T.A., Satoh, T., Supat-tapone, S., and Synder, S.H. (1989) Inositol 1,4,5-trisphosphate receptor localized to endoplasmic reticulumin cerebellar Purkinje neurons. Nature 339: 468–470.
Sajid, M., Withers-Martinez, C., and Blackman, M.J. (2000)Maturation and specificity of Plasmodium falciparumsubtilisin-like protease-1, a malaria merozoite subtilisin-likeserine protease. J Biol Chem 275: 631–641.
Salmon, B.L., Oksman, A., and Goldberg, D.E. (2001)Malaria parasite exit from the host erythrocyte: a two-stepprocess requiring extraerythrocytic proteolysis. Proc NatlAcad Sci USA 98: 271–276.
Sijwali, P.S., and Rosenthal, P.J. (2004) Gene disruption con-firms a critical role for the cysteine protease falcipain-2 inhemoglobin hydrolysis by Plasmodium falciparum. ProcNatl Acad Sci USA 101: 4384–4389.
Sijwali, P.S., Koo, J., Singh, N., and Rosenthal, P.J. (2006)Gene disruptions demonstrate independent roles for thefour falcipain cysteine proteases of Plasmodium falci-parum. Mol Biochem Parasitol 150: 96–106.
Sim, B.K., Toyoshima, T., Haynes, J.D., and Aikawa, M.(1992) Localization of the 175-kilodalton erythrocytebinding antigen in micronemes of Plasmodium falciparummerozoites. Mol Biochem Parasitol 51: 157–159.
Singh, S., and Chitnis, C.E. (2013) Flow cytometry-basedmethods for measurement of cytosolic calcium and surfaceprotein expression in Plasmodium falciparum merozoites.Methods Mol Biol 923: 281–290.
Singh, S., Alam, M.M., Pal-Bhowmick, I., Brzostowski, J.A.,and Chitnis, C.E. (2010) Distinct external signals triggersequential release of apical organelles during erythrocyteinvasion by malaria parasites. PLoS Pathog 6: e1000746.
Taylor, H.M., McRobert, L., Grainger, M., Sicard, A., Dluze-wski, A.R., Hopp, C.S., et al. (2010) The malaria parasitecyclic GMP-dependent protein kinase plays a central rolein blood-stage schizogony. Eukaryot Cell 9: 37–45.
Trager, W., and Jensen, J.B. (1976) Human malaria parasitesin continuous culture. Science 193: 673–675.
Vaid, A., Thomas, D.C., and Sharma, P. (2008) Role ofCa2+/CaM-PfPKB signaling pathway in erythrocyte inva-sion by Plasmodium falciparum. J Biol Chem 283: 5589–5597.
Wickham, M.E., Culvenor, J.G., and Cowman, A.F. (2003)Selective inhibition of a two step egress of malaria para-sites from the host erythrocyte. J Biol Chem 278: 37658–37663.
Winograd, E., Clavijo, C.A., Bustamante, L.Y., and Jaramillo,M. (1999) Release of merozoites from Plasmodiumfalciparum-infected erythrocytes could be mediated by anon-explosive event. Parasitol Res 85: 621–624.
Yeoh, S., O’Donnell, R.A., Koussis, K., Dluzewski, A.R.,Ansell, K.H., Osborne, S.A., et al. (2009) Subcellular dis-charge of a serine protease mediates release of invasivemalaria parasites from host erythrocytes. Cell 131: 1072–1083.
Ca2+-mediated exocytosis in regulation of egress 11
© 2012 Blackwell Publishing Ltd, Cellular Microbiology

Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Videos S1–S4. Intracellular Ca2+ levels during egress of P. fal-ciparum merozoites from host erythocytes. P. falciparum late-stage schizonts were labelled with Fluo-4AM and added toerythrocytes. Both DIC and fluorescence images were acquiredsimultaneously on a Zeiss LSM510 confocal microscope with a1.4 NA lens, using 488 nm excitation at low power to minimizebleaching and cell damage. Videos S1A, S2A, S3A, S4A: mergeof the DIC and fluorescence images are shown. Videos S1B,S2B, S3B, S4B: DIC images are shown. Videos S1C, S2C, S3C,S4C: fluorescence images are shown.Fig. S1. A–C. Analysis of intracellular Ca2+ levels during egressof P. falciparum merozoites. Late-stage purified P. falciparumschizonts were labelled with Fluo-4AM and added to uninfectederythrocytes. Intracellular Ca2+ levels were monitored in P. falci-parum schizonts during egress by time-lapse video microscopy.Selected pairs of merge of both DIC and fluorescence images areshown. Mean fluorescence intensities (MFI) after background
subtraction is reported for schizonts from three independentexperiments corresponding to Videos S2, S3 and S4 from whichmerozoites were released following its rupture. An increasein intracellular Ca2+ just before egress was observed in eachschizonts.Fig. S2. Expression and purification of the recombinantrPfSERA5 and rPfSUB1 proteins. The amino acid region (E578 toN828, within catalytic domain) of PfSERA5 and amino acid region(E27 to H688, full-length excluding signal sequence) of PfSUB1was expressed and purified to homogeneity.A. The purified protein of PfSERA5 was analysed on a SDS-PAGE and detected in immunoblot using anti-His tag antibody.B. The purified protein of PfSUB1 was analysed on a SDS-PAGEand detected in immune-blots using an anti-His tag antibody.Fig. S3. Western blot analysis to confirm the specificity of anti-PfSUB1 mouse sera. Western blot of lysates prepared fromring-infected erythrocytes (~ 14–16 hpi) and schizont-infectederythrocytes (~ 48 hpi) as well as of uninfected erythrocytes, withanti-PfSUB1 mouse sera. Mature 47 kDa along with 54 kDa frag-ment of PfSUB1 detected only in schizont-infected erythrocyteslysate.
12 S. Agarwal et al.
© 2012 Blackwell Publishing Ltd, Cellular Microbiology
Copyright © 2022 FDOKUMEN




















