
C5L2: a controversial receptor of complement anaphylatoxin, C5a
10
The FASEB Journal • Review C5L2: a controversial receptor of complement anaphylatoxin, C5a Rui Li, 1 Liam G. Coulthard, 1 M. C. L. Wu, Stephen M. Taylor, and Trent M. Woodruff 2 School of Biomedical Sciences, University of Queensland, Brisbane, Queensland, Australia ABSTRACT C5a is the paramount proinflammatory mediator of the complement cascade, and has been previously thought to act only through a single, G-pro- tein-coupled, C5a receptor (C5aR; also termed CD88). In 2000, a second C5a receptor, C5L2 (previously known as GPR77), was discovered; yet, despite 12 yr of intensive research, its biological, or pathophysiological, function is both enigmatic and controversial. Unlike C5aR, this receptor does not couple to G proteins, and early studies promoted the hypothesis that C5L2 func- tions as a decoy receptor. However, recent data have provided other evidence for more complicated and conflicting interactions between C5L2 and other inflam- matory mediators. C5L2 has been recently demon- strated to physically interact with both C5aR and -ar- restin to negatively regulate C5aR signaling toward an anti-inflammatory manner, and to reduce pathology, in several disease models in vivo. In direct contrast, other groups have demonstrated that C5L2 stimulation caused release of HMGB1 both in vitro and in vivo, and enhanced pathology in sepsis models, suggesting a clear proinflammatory signaling role. These astoundingly contradictory data challenge our precepts and compli- cate the foundational bases for the possible targeting of C5L2 as a therapeutic option in inflammatory dis- ease. C5L2 may be the great masquerader in comple- ment biology; its function dependent on the cell type, species, and disease context. Because of these unusual and unforeseen complexities, we present the current state of knowledge on C5L2 structure, expression and, most controversially, its putative functions.—Li, R., Coulthard, L.G., Wu, M. C. L., Taylor, S. M., Woodruff, T. M. C5L2: a controversial receptor of complement anaphylatoxin, C5a. FASEB J. 27, 000 – 000 (2013). www.fasebj.org Key Words: inflammation therapeutics The complement system plays a pivotal role in immunity, and is often described as the first line of defense against pathogens. At the apex of the comple- ment zymogen cascade lies C5, a 190-kDa precursor to the effector molecules, C5a and C5b. C5b has a well- defined function in initiating the formation of the membrane attack complex, and C5a historically been described as a trigger for the cellular immune response, signaling through its receptor, C5a receptor 1 (C5aR) (1). Through C5aR, C5a is a potent leukocyte chemoat- tractant and mediates inflammatory responses, and as such, is involved in many diseases including sepsis, rheumatoid arthritis, asthma, ischemia-reperfusion in- juries, and inflammatory bowel disease (2). However, recently a second receptor for C5a has been discovered that does not fit neatly with the perceived functions of C5a, generating controversy in the literature. This second receptor, C5a-like receptor 2 (C5L2), remains a source of intrigue for the complement research com- munity. We now attempt to sift through the reports, mine the frustrations of the frustrated, cut through the mists of ambiguities, and peer into the enlightened darkness of the past, to provide a small beacon of light and hope for the next wave of research into the most recent of many enigmas of complement. Succinctly: What is C5L2, why is it there, what does it do, and why do we care? STRUCTURE C5L2 was first cloned in 2000, and it has closest homology with C5aR (also known as CD88; ref. 3). The following year, it was determined that C5L2 is encoded by GPR77 in humans on chromosome 19, q13.33-13.34, located downstream of the gene encoding C5aR; in 2002, it was confirmed that C5L2 binds C5a with high affinity (4). C5L2 binds to C5a with the same high affinity as C5aR (K d 2.5 nm), but to its degradation fragment, C5a desArg , with a much higher affinity (K d 12 nm) than C5aR (K d 660 nm). C3a desArg , also 1 These authors contributed equally to this work. 2 Correspondence: School of Biomedical Sciences, Univer- sity of Queensland, St Lucia, QLD, 4072, Australia. E-mail: [email protected] doi: 10.1096/fj.12-220509 Abbreviations: ASP, acylation-stimulating protein; C5aR, C5a receptor 1; C5L2, C5a-like receptor 2; GPCR, G-protein- coupled receptor; GRK, G-protein receptor kinase; HMGB1, high mobility group box protein 1; IL-6, interleukin-6; IFN-, interferon-; LPS, lipopolysaccaride; SIP, spingosine-1-phos- phate; Sphk1, sphingosine kinase 1; TLR4, Toll-like receptor 4; TNF, tumor necrosis factor 1 0892-6638/13/0027-0001 © FASEB The FASEB Journal article fj.12-220509. Published online December 13, 2012.
Transcript of C5L2: a controversial receptor of complement anaphylatoxin, C5a

The FASEB Journal • Review
C5L2: a controversial receptor of complementanaphylatoxin, C5a
Rui Li,1 Liam G. Coulthard,1 M. C. L. Wu, Stephen M. Taylor,and Trent M. Woodruff2
School of Biomedical Sciences, University of Queensland, Brisbane, Queensland, Australia
ABSTRACT C5a is the paramount proinflammatorymediator of the complement cascade, and has beenpreviously thought to act only through a single, G-pro-tein-coupled, C5a receptor (C5aR; also termed CD88).In 2000, a second C5a receptor, C5L2 (previouslyknown as GPR77), was discovered; yet, despite 12 yr ofintensive research, its biological, or pathophysiological,function is both enigmatic and controversial. UnlikeC5aR, this receptor does not couple to G proteins, andearly studies promoted the hypothesis that C5L2 func-tions as a decoy receptor. However, recent data haveprovided other evidence for more complicated andconflicting interactions between C5L2 and other inflam-matory mediators. C5L2 has been recently demon-strated to physically interact with both C5aR and �-ar-restin to negatively regulate C5aR signaling toward ananti-inflammatory manner, and to reduce pathology, inseveral disease models in vivo. In direct contrast, othergroups have demonstrated that C5L2 stimulationcaused release of HMGB1 both in vitro and in vivo, andenhanced pathology in sepsis models, suggesting a clearproinflammatory signaling role. These astoundinglycontradictory data challenge our precepts and compli-cate the foundational bases for the possible targetingof C5L2 as a therapeutic option in inflammatory dis-ease. C5L2 may be the great masquerader in comple-ment biology; its function dependent on the cell type,species, and disease context. Because of these unusualand unforeseen complexities, we present the currentstate of knowledge on C5L2 structure, expression and,most controversially, its putative functions.—Li, R.,Coulthard, L.G., Wu, M. C. L., Taylor, S. M., Woodruff,T. M. C5L2: a controversial receptor of complementanaphylatoxin, C5a. FASEB J. 27, 000–000 (2013).www.fasebj.org
Key Words: inflammation � therapeutics
The complement system plays a pivotal role inimmunity, and is often described as the first line ofdefense against pathogens. At the apex of the comple-ment zymogen cascade lies C5, a 190-kDa precursor tothe effector molecules, C5a and C5b. C5b has a well-defined function in initiating the formation of themembrane attack complex, and C5a historically beendescribed as a trigger for the cellular immune response,signaling through its receptor, C5a receptor 1 (C5aR)(1). Through C5aR, C5a is a potent leukocyte chemoat-tractant and mediates inflammatory responses, and assuch, is involved in many diseases including sepsis,rheumatoid arthritis, asthma, ischemia-reperfusion in-juries, and inflammatory bowel disease (2). However,recently a second receptor for C5a has been discoveredthat does not fit neatly with the perceived functions ofC5a, generating controversy in the literature. Thissecond receptor, C5a-like receptor 2 (C5L2), remains asource of intrigue for the complement research com-munity. We now attempt to sift through the reports,mine the frustrations of the frustrated, cut through themists of ambiguities, and peer into the enlighteneddarkness of the past, to provide a small beacon of lightand hope for the next wave of research into the mostrecent of many enigmas of complement. Succinctly:What is C5L2, why is it there, what does it do, and whydo we care?
STRUCTURE
C5L2 was first cloned in 2000, and it has closesthomology with C5aR (also known as CD88; ref. 3). Thefollowing year, it was determined that C5L2 is encodedby GPR77 in humans on chromosome 19, q13.33-13.34,located downstream of the gene encoding C5aR; in2002, it was confirmed that C5L2 binds C5a with highaffinity (4). C5L2 binds to C5a with the same highaffinity as C5aR (Kd�2.5 nm), but to its degradationfragment, C5adesArg, with a much higher affinity(Kd�12 nm) than C5aR (Kd�660 nm). C3adesArg, also
1 These authors contributed equally to this work.2 Correspondence: School of Biomedical Sciences, Univer-
sity of Queensland, St Lucia, QLD, 4072, Australia. E-mail:[email protected]
doi: 10.1096/fj.12-220509
Abbreviations: ASP, acylation-stimulating protein; C5aR,C5a receptor 1; C5L2, C5a-like receptor 2; GPCR, G-protein-coupled receptor; GRK, G-protein receptor kinase; HMGB1,high mobility group box protein 1; IL-6, interleukin-6; IFN-�,interferon-�; LPS, lipopolysaccaride; SIP, spingosine-1-phos-phate; Sphk1, sphingosine kinase 1; TLR4, Toll-like receptor4; TNF, tumor necrosis factor
10892-6638/13/0027-0001 © FASEB
The FASEB Journal article fj.12-220509. Published online December 13, 2012.

referred to as acylation-stimulating protein (ASP), hasbeen identified to have a direct interaction with C5L2on adipocytes and macrophages, which may contributeto the inflammatory-adipose phenotype (5). However,these results are controversial, since it has been shownthat C5L2 does not bind C3a or ASP/C3adesArg, and it isunlikely to be directly related to lipid and carbohydratemetabolism (6). This viewpoint is currently unresolved,and there are no corroborating studies outside theresearch group making these claims. How this plays outremains to be seen.
C5L2 is a 337-aa, 7-transmembrane protein that isgrouped in the G-protein family of receptors, togetherwith C5aR. C5L2 shares 38% sequence homology withC5aR and is conserved across mammalian species (Fig. 1);however, C5L2 exhibits sequence differences in thehighly conserved DRY motif located at amino acidresidues 137–139. The DRY motif is an importantregion of the third intracellular loop of G-protein-coupled receptors (GPCRs), and enables coupling of Gproteins (7). In C5L2, the DLC sequence of the thirdintracellular loop appears to be evolutionarily linked tothe conserved DRY sequence of other GPCRs. In sup-port of this conclusion, mutating the leucine residue ofthe DLC sequence to an arginine has been shown topromote intracellular calcium release in C5L2-trans-fected RBL cells (8). There is also an apparent lack ofan NPXXY motif, which has previously been demon-strated to be essential for phospholipase C and mito-gen-activated protein kinase activation in formyl pep-tide receptor signaling, a group of phylogenetically
related proteins (9). In addition. C5L2 has a shortenedthird, typically G-protein-binding, intracellular loop(Fig. 1). These differences have all contributed to theoriginal prevailing hypothesis that C5L2 is a nonsignal-ing decoy receptor (2).
On the extracellular domains, C5L2 has a similarpattern of tyrosine and acidic N-terminal residues toC5aR, which have been shown to be a major feature ofextracellular binding of C5a (10). C5L2 also sharessimilarities with C5aR in the number of charged andhydrophobic residues in the loops and transmembraneregions, which are involved in interaction with the Cterminus of C5a (11). Specifically, there is conservationof tyrosine residues 10 and 13 at the N terminus, whichhave been shown to be critical for C5a binding by C5aR(2). However, mutation of these residues in C5L2diminishes the affinity for C5adesArg, but not C5a (10).Affinity for C5a is reduced in the presence of mutationsto aspartic acid residues 22, 18, and 25, of which D18was demonstrated to also be of importance in C5a-C5aRcoupling (10).
EXPRESSION
C5L2 is generally seen to be expressed in similar tissueand cellular locations to C5aR, in agreement with theiradjacent location on the genome. Currently, there arevery few natural cells expressing just one of the tworeceptors; for whatever reasons, they are generallynatively expressed in tandem. The expression of C5L2
Figure 1. Amino acid sequence alignment of C5L2. Human (top), mouse(middle), and rat (bottom) were aligned using ClustalW2 (EuropeanBioinformatics Institute). Transmembrane (TM) regions as well as intracel-lular (IC) and extracellular (EC) loops are annotated. Asterisks indicateconserved residues. Regions identified as important for G-protein couplingin other GPCRs, but mutated in C5L2, are annotated as # (DRY region) and## (NPXXY region). Numbers at left indicate the residue number.
2 Vol. 27 March 2013 LI ET AL.The FASEB Journal � www.fasebj.org
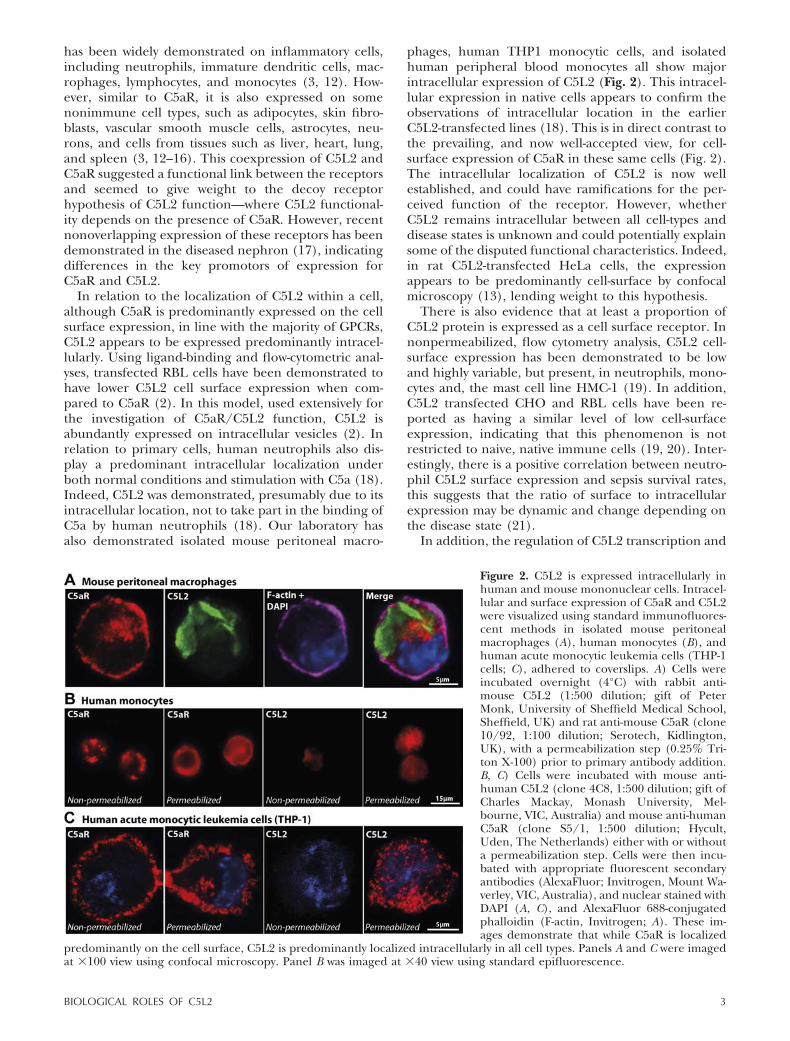
has been widely demonstrated on inflammatory cells,including neutrophils, immature dendritic cells, mac-rophages, lymphocytes, and monocytes (3, 12). How-ever, similar to C5aR, it is also expressed on somenonimmune cell types, such as adipocytes, skin fibro-blasts, vascular smooth muscle cells, astrocytes, neu-rons, and cells from tissues such as liver, heart, lung,and spleen (3, 12–16). This coexpression of C5L2 andC5aR suggested a functional link between the receptorsand seemed to give weight to the decoy receptorhypothesis of C5L2 function—where C5L2 functional-ity depends on the presence of C5aR. However, recentnonoverlapping expression of these receptors has beendemonstrated in the diseased nephron (17), indicatingdifferences in the key promotors of expression forC5aR and C5L2.
In relation to the localization of C5L2 within a cell,although C5aR is predominantly expressed on the cellsurface expression, in line with the majority of GPCRs,C5L2 appears to be expressed predominantly intracel-lularly. Using ligand-binding and flow-cytometric anal-yses, transfected RBL cells have been demonstrated tohave lower C5L2 cell surface expression when com-pared to C5aR (2). In this model, used extensively forthe investigation of C5aR/C5L2 function, C5L2 isabundantly expressed on intracellular vesicles (2). Inrelation to primary cells, human neutrophils also dis-play a predominant intracellular localization underboth normal conditions and stimulation with C5a (18).Indeed, C5L2 was demonstrated, presumably due to itsintracellular location, not to take part in the binding ofC5a by human neutrophils (18). Our laboratory hasalso demonstrated isolated mouse peritoneal macro-
phages, human THP1 monocytic cells, and isolatedhuman peripheral blood monocytes all show majorintracellular expression of C5L2 (Fig. 2). This intracel-lular expression in native cells appears to confirm theobservations of intracellular location in the earlierC5L2-transfected lines (18). This is in direct contrast tothe prevailing, and now well-accepted view, for cell-surface expression of C5aR in these same cells (Fig. 2).The intracellular localization of C5L2 is now wellestablished, and could have ramifications for the per-ceived function of the receptor. However, whetherC5L2 remains intracellular between all cell-types anddisease states is unknown and could potentially explainsome of the disputed functional characteristics. Indeed,in rat C5L2-transfected HeLa cells, the expressionappears to be predominantly cell-surface by confocalmicroscopy (13), lending weight to this hypothesis.
There is also evidence that at least a proportion ofC5L2 protein is expressed as a cell surface receptor. Innonpermeabilized, flow cytometry analysis, C5L2 cell-surface expression has been demonstrated to be lowand highly variable, but present, in neutrophils, mono-cytes and, the mast cell line HMC-1 (19). In addition,C5L2 transfected CHO and RBL cells have been re-ported as having a similar level of low cell-surfaceexpression, indicating that this phenomenon is notrestricted to naive, native immune cells (19, 20). Inter-estingly, there is a positive correlation between neutro-phil C5L2 surface expression and sepsis survival rates,this suggests that the ratio of surface to intracellularexpression may be dynamic and change depending onthe disease state (21).
In addition, the regulation of C5L2 transcription and
Figure 2. C5L2 is expressed intracellularly inhuman and mouse mononuclear cells. Intracel-lular and surface expression of C5aR and C5L2were visualized using standard immunofluores-cent methods in isolated mouse peritonealmacrophages (A), human monocytes (B), andhuman acute monocytic leukemia cells (THP-1cells; C), adhered to coverslips. A) Cells wereincubated overnight (4°C) with rabbit anti-mouse C5L2 (1:500 dilution; gift of PeterMonk, University of Sheffield Medical School,Sheffield, UK) and rat anti-mouse C5aR (clone10/92, 1:100 dilution; Serotech, Kidlington,UK), with a permeabilization step (0.25% Tri-ton X-100) prior to primary antibody addition.B, C) Cells were incubated with mouse anti-human C5L2 (clone 4C8, 1:500 dilution; gift ofCharles Mackay, Monash University, Mel-bourne, VIC, Australia) and mouse anti-humanC5aR (clone S5/1, 1:500 dilution; Hycult,Uden, The Netherlands) either with or withouta permeabilization step. Cells were then incu-bated with appropriate fluorescent secondaryantibodies (AlexaFluor; Invitrogen, Mount Wa-verley, VIC, Australia), and nuclear stained withDAPI (A, C), and AlexaFluor 688-conjugatedphalloidin (F-actin, Invitrogen; A). These im-ages demonstrate that while C5aR is localized
predominantly on the cell surface, C5L2 is predominantly localized intracellularly in all cell types. Panels A and C were imagedat �100 view using confocal microscopy. Panel B was imaged at �40 view using standard epifluorescence.
3BIOLOGICAL ROLES OF C5L2

translation has been demonstrated to be related to C5astimulation. It has been shown that C5a decreases C5L2protein in rat polymorphonuclear cell lysates duringsepsis (21). This control of expression is likely a directproduct of the C5a-C5L2 axis, as the presence orabsence, of C5aR appears to have little influence onexpression of C5L2 (21). In support of this, no signifi-cant changes in C5L2 expression have been foundbetween wild-type and C5aR�/� mouse neutrophils(22). Another important regulator of C5L2 expressionis the gram-negative bacterial cell coat component,lipopolysaccharide (LPS). LPS is a potent ligand forToll-like receptor 4 (TLR4), which is of considerableinterest to complementologists for its potential signal-ing interactions with C5aR (23). In peripheral bloodmononuclear cells, LPS has been demonstrated toincrease C5aR-mediated responsiveness to C5a via thedown-regulation of C5L2 expression (24).
The up-regulation of C5L2 expression is regulatedby factors such as interferon-� (IFN-�), noradrena-line, dibutyryl-cAMP insulin, and thiazolidinedionein various cell types (6, 15, 25, 26). In addition, C5L2expression also appears to be dependent on theactivity of sphingosine kinase 1 (Sphk1), an enzymeresponsible for generating the signaling lipid, sphin-gosine-1-phosphate (S1P), which is a key mediator inphagocyte function (27–29). Mice deficient in Sphk1have reduced C5L2 expression when compared towild-type mice on the same background, and have nomacrophage Erk1/2 phosphorylation in response toC5a. The researchers postulated that inflammatorycytokines and C5a activate Sphk1, which can gener-ate S1P during inflammation. S1P then increasesexpression of C5L2 to negatively regulate inflamma-tion (30). Estrogen receptor agonists also increaseC5aR mRNA expression, whereas the regulation ofC5L2 is dependent on the type of agonist. Oestradiolincreased C5L2 expression, whereas a selective phar-macological agonist of the estrogen receptor alphaagonist resulted in the down-regulation of C5L2mRNA (31).
MOLECULAR FUNCTION
The physiological role of C5L2 is still both enigmaticand controversial; three conflicting observations aboutthe function of C5L2 have been presented since itsdiscovery in 2000 (Fig. 3). Two hypotheses describe ananti-inflammatory role for C5L2 in the immune system(18, 20), whereas a third provides evidence for C5L2acting as a proinflammatory entity, triggering release ofcytokines, such as the damage-associated high mobilitygroup box protein 1 (HMGB1; ref. 22). These activitiesseem mutually exclusive; however, we believe that notonly is this not necessarily so, but quite apposite. Theevidence for each of the postulated functions is de-scribed below.
Anti-inflammatory role as a recycling decoy receptor
The decoy receptor model for C5L2 was initially de-scribed, based on the lack of structural motifs known toparticipate in G-protein coupling (4, 8). However,Scola and colleagues in 2009 provided the first physicalevidence for this model in transfected CHO, HL-60,Hela, and RBL cells (20). The researchers noted C5L2-mediated, clathrin-dependent internalization of C5aand C5adesArg in differentiated HL-60 and HeLa cells.This effect was not inhibited by a C5aR antagonist,strongly suggesting that ligand degradation occurs afteruptake by C5L2. Using human PMNs, C5L2 was con-firmed to be the main receptor of C5adesArg. Theexperiments suggested that C5L2 functioned by remov-ing active complement fragments from the extracellu-lar environment, preventing C5aR activation (20). Inthis model, the binding of C5a to C5L2 induces inter-nalization of the complex and endosomal degradationof C5a; C5L2 is then returned to the cell surface. Theanti-inflammatory actions of this model depend onC5L2 sequestering the available C5a (and C5adesArg) toprevent the proinflammatory interaction between C5aand C5aR.
Anti-inflammatory role via �-arrestin pathway
Bamberg and colleagues (18) developed another anti-inflammatory model of C5L2 function, whereby C5L2complexes with �-arrestin to negatively regulate inflam-mation. While antibody blockade of C5L2 induced anincreased level of C5a-mediated chemotaxis andERK1/2 phosphorylation, there was no influence onligand uptake or C5aR endocytosis in human PMNs.This result contrasts with the previous hypothesis thatC5L2 acts as a decoy receptor. Bamberg and colleagues(18) demonstrated that C5L2 functions independentlyof calcium mobilization, and inhibits the C5aR-medi-ated response to C5a through interactions with �-arres-tin. It was proposed that binding of C5a to C5aRactivates intracellular C5L2 and triggers phosphoryla-tion of both receptors by G-protein receptor kinases(GRKs), facilitating their association with �-arrestin.The C5aR-�-arrestin complex activates ERK1/2, but theC5L2-�-arrestin complex results in ERK1/2 inhibition.The net signal is determined by the summation of thetwo competing pathways (18).
The evidence for this hypothesis is supported by theintracellular localization of C5L2. Bamberg and col-leagues (18) used human PMNs to demonstrate acontrasting expression pattern for C5aR and C5L2;C5aR is localized to the cell membrane, whereas C5L2is expressed predominantly intracellularly. Indeed, inthe transfected RBL and CHO cells used to support thedecoy receptor hypothesis, the majority of C5L2 ap-pears perinuclear, which remains unexplained in thedecoy receptor model (20). C5aR and C5L2 were alsodemonstrated to colocalize after administration of C5a.However the investigators used a high dose of C5a (100nM) to induce colocalization, despite demonstrating
4 Vol. 27 March 2013 LI ET AL.The FASEB Journal � www.fasebj.org

that C5L2 negatively regulates C5a induced ERK1/2phosphorylation at 1 nM. This is an important discrep-ancy to highlight, as the investigators rely heavily oncolocalization of C5aR/C5L2/�-arrestin to supporttheir hypothesis. Other studies have also investigatedC5L2/�-arrestin colocalization in different cell typeswith contrasting results, suggesting that this hypothesismay not be globally applicable (20, 32, 33). However, arecent study has supported this hypothesis throughdemonstrating green fluorescent protein-labeled �-ar-restin-2 localization to cytoplasmic vesicles on stimula-tion with C5a, demonstrating an EC50 for this effect of2 nM. In addition, these researchers demonstrated thatthe �-arrestin coupling was independent of any G-pro-tein coupling (34). Although there have been previousnonsupportive studies for this hypothesis, these studies
focused their attention on �-arrestin-1, rather than�-arrestin-2 (20).
Proinflammatory role through intracellular signaling,and release of cytokines [interlukin-6 (IL-6), tumornecrosis factor � (TNF-�), and HMGB1]
In direct contrast to both the decoy receptor and�-arrestin models of C5L2 function, a proinflammatoryfunction for C5L2 has been postulated. It has beennoted by several groups that the cytokine expressionprofile of wild-type and C5L2�/� neutrophils differswhen exposed to C5a. C5L2 appears to be the trigger inthe C5a-induced up-regulation of IL-6, TNF-�, andcomplement receptor 3 in neutrophils (13). C5L2�/�
neutrophils also demonstrate impaired C3aR/C5aR-
Figure 3. Three conflicting mechanisms for C5L2 function. Each row of panels represents a separate mechanism of C5L2function. Top panels: decoy receptor model. 1) C5L2, on the cell surface, binds C5a, preventing C5a-C5aR interaction. 2) C5L2cannot couple G proteins; no second messenger signaling occurs. 3) C5L2 internalizes on binding C5a. 4) C5 is degraded inintracellular vesicles. 5) C5L2 returns to the cell surface. Middle panels: �-arrestin complex. 1) C5L2 is located intracellularly,C5a binds C5aR at the cell surface. 2) On C5a binding, a C5aR/G-protein/C5L2 complex is formed, causing phosphorylationof the C terminus of both receptors. 3) Phosphorylation allows �-arrestin binding. 4) Complex of receptors and �-arrestin isinternalized in early recycling endosome. 5) Presence of �-arrestin prevents MAPK pathway signaling through C5aR. Bottompanels: signaling. 1, 2) C5L2, on the cell surface, competes with C5aR for C5a binding. 3) Binding of C5a to C5L2 results inMAPK phosphorylation independent of C5aR action. 4) MAPK pathway signals through unknown mechanism to the nucleus.5) Signaling results in HMGB1 release from nucleus. The top two rows, decoy receptor and �-arrestin, are hypothesized to beanti-inflammatory in action, whereas, the bottom row, signaling, is hypothesized to be proinflammatory in action.
5BIOLOGICAL ROLES OF C5L2

induced ERK1/2 and AKT activation, suggesting thatC5L2 is a positive modulator for both C5a- and C3a-induced responses (12).
Perhaps the most compelling evidence of a proin-flammatory function for C5L2 comes from the cecal-ligation and puncture model of sepsis. C5L2�/� micehad reduced mortality when compared to wild-typelittermates, and significantly reduced plasma inflamma-tory cytokine concentrations (22). The function ofC5L2 in this system was postulated to be as a trigger forthe release of cytokines, including transcription factorand proinflammatory mediator HMGB1, from circulat-ing leukocytes. Rittirsch and colleagues (22) used in-hibitors of MEK1/2, JNK1/2, and p38 to attenuate therelease of HMGB1 in peritoneal macrophages. In con-trast to the �-arrestin model of C5L2 activity, these datasuggest that the release of HMGB1 via C5L2 signaling isdependent on MAPK activity, an intriguing findinggiven that C5L2 lacks the traditional intracellular moi-eties that permit G-protein coupling. In addition, theHMGB1 release in C5L2�/� macrophages was alsoattenuated when stimulated with LPS, indicating thatLPS signaling via Tlr4 depends on the integrity ofC5L2. In addition, deficiency of C5L2 leads to a re-duced inflammatory cell infiltration in vivo and C5a-induced downstream signaling in cells. C3a-inducedsignaling and F-actin formation were reduced in C5L2�/�
cells. Also, defective in vivo inflammatory responses andhematopoietic cell regeneration in C5L2�/� mice werereported (12). Other observations also support a pro-inflammatory function for C5L2; for instance, C5L2-deficient mice are protected from the development ofairway hyperresponsiveness mucus production in twomodels of experimental allergic asthma (35).
This seemingly proinflammatory role for C5L2 iscontradictory to the observations in C5L2-knockoutmice, where the biological activity of the C5a/C5aR axisis enhanced both in vivo and in vitro. In studies innondisease models, the biological role of C5L2 thusappears to be limiting the proinflammatory response tothe anaphylatoxin. Other studies demonstrate that up-regulation of C5L2 may be of benefit in inflammatorystates driven by C5a, including sepsis, asthma, cysticfibrosis, and chronic obstructive lung disease (36).Deficiency of C5L2 results in reduced inflammatory cellinfiltration, suggesting that C5L2 is critical for optimalC5a-mediated cell infiltration in certain in vivo settings.Furthermore, like mice incapable of C3a/C3aR signal-ing, C5L2-deficient mice are hypersensitive to LPS-induced septic shock, show reduced ovalbumin (OVA)-induced airway hyperresponsiveness and inflammation,and are mildly delayed in hematopoietic cell regener-ation after �-irradiation (12).
Therefore, the question remains, how to integratethese seemingly conflicting observations to develop acomplete understanding of C5L2. At least in part, someanswers may lay in the various models used to investi-gate the function of this protein (which may indeed becentral to the lack of consensus surrounding the func-tions of C5L2). The models using transfected CHO and
RBL cell lines to investigate C5L2 function, althoughuseful for investigating ligand-receptor interactions,come with major caveats with regard to function. Forinstance, the expression of the putative second messen-ger for C5L2, �-arrestin2, may be lower or absent fromthese cell lines.
C5L2 PHYSIOLOGY AND PATHOPHYSIOLOGY
C5a has been implicated in many diseases, such assepsis (37), rheumatoid arthritis (38), asthma andallergy (36, 39–41), ischemia-reperfusion injuries (42–44), inflammatory bowel disease (45–47), atherosclero-sis (48), and neurodegeneration (16, 49–51). However,many of these studies have focused on C5aR withoutspecifically investigating the role of C5L2. Modulatingthe actions of C5L2 may yet prove to be a usefultherapeutic tool. However, the function of C5L2 indisease is still a highly debated topic. The function ofthis receptor, as to whether it attenuates or propagatesinflammation, varies between disease states, and evenbetween cell types in the same disease.
Sepsis
In mouse models of sepsis, blocking C5L2 producedincreased levels of proinflammatory mediators, IL-6and TNF-� (13). Moreover, rat C5L2 was shown to haveanti-inflammatory properties, as its reduction resultedin increased NOS2 and NFkB gene expression (15). Inaddition, higher levels of C5L2 in PMNs from sepsispatients who survived were found in comparison tononsurvivors, a finding which appears to cement ananti-inflammatory role for C5L2 in sepsis (21).
However, there is evidence that the organ dysfunc-tion associated with sepsis may be mediated, in part,through C5L2. In rats, 24 h after CLP there is signifi-cant apoptosis of adrenomedullary cells, as assessed bythe TUNEL technique. These effects were reversed bydual blockade of the C5a receptors, C5aR and C5L2,however the researchers did not attempt blockade ofeach receptor independently (52). Since blockade ofboth C5a receptors abolished adrenomedullary apopto-sis in vivo, it was suggested that C5aR and C5L2 may bepromising pharmacological targets in the clinical set-ting of sepsis. Although, such targeting of C5L2 mayexacerbate the other sequalae of sepsis that C5L2 hasbeen demonstrated to mitigate.
To add to the confusion of the specific role for C5L2in sepsis, there is the observation that C5L2-knockoutmice have increased survival in midgrade sepsis (22).This series of experiments demonstrated that C5L2 wasrequired for release of the proinflammatory mediator,HMGB1, in the presence of both C5a and LPS. Inaddition, HMGB1 release from C5aR�/� macrophageswas diminished in the presence of inhibitors toMEK1/2, JNK1/2, and Akt, indicating that C5L2 isinvolved in intracellular signal transduction in thismodel. However, in high-grade sepsis, only the block-
6 Vol. 27 March 2013 LI ET AL.The FASEB Journal � www.fasebj.org

ade of signaling from both C5aR and C5L2 improvedsurvival, perhaps suggesting that the severity of thedisease affects the functions of C5L2.
Metabolism
C5L2-knockout mice demonstrate insulin resistanceand high proinflammatory serum cytokines whenplaced on a high-fat/high-sucrose diet (53). There issome evidence that this diabetogenic phenotype is theresult of a disrupted interaction between C5L2 andASP, which is also known as C3adesArg, the breakdownproduct of C3a. The ASP/C5L2 interaction may induceadipocyte differentiation, and it is suggested that thedown-regulation of C5L2 in adipocytes contributes toinsulin resistance. Interestingly, progesterone has beendemonstrated to down-regulate C5L2 in both matureadipocytes and preadipocytes, providing an interestingangle for research into gestational diabetes (54). Inthese studies, C5L2 has been reported to have a highbinding affinity for both human and mouse C5L2 (Kd118 and 31 nM, respectively); however, this directinteraction has been disputed by other groups thathave been unable to demonstrate any interaction be-tween C5L2 and ASP (5, 6).
Despite the controversy over the putative interactionbetween C5L2 and ASP, both proteins have beendemonstrated to affect metabolism at the level of thecentral nervous system. Both C5L2 and C5aR are pres-ent in the anterior pituitary gland, and C5a was dem-onstrated to stimulate adrenocorticotropic hormonerelease; however, whether this effect is through C5aR orC5L2 is unclear (55). In addition, administration ofASP into the third ventricle promotes an anorexigenicstate through modulation of neuropeptide Y and pro-opiomelanocortin expression (56).
Asthma
Although C5aR has previously been demonstrated topromote the development of asthma (40), little atten-tion has been given to C5L2. However, Zhang andcolleagues (35) show that C5L2-knockout mice areprotected from the development of allergic asthma.This protection was determined to be due to C5L2expression on dendritic cells promoting the inflamma-tory T-cell phenotype of asthma. Interestingly, thealtered cytokine production of C5L2 dendritic cells wasalso present in vitro when cells were stimulated withhouse dust-mite allergen, suggesting that the role ofC5L2 in this model may not be through the directbinding of C5a. Rather, C5L2 may act as a modulator ofother pattern recognition receptors to promote theasthma phenotype.
CONCLUSIONS
The actions of C5L2 in health and disease are not onlyfar from conclusive, the field is in considerable and
seemingly paralyzed disarray. In contrast to the well-documented, global proinflammatory role for C5aR,relentlessly emerging evidence strongly indicates thatthe situation for C5L2 is much more complex, or at thevery least, bewildering. There may be no such thing asa unifying theory for C5L2 function and its role invarious contrived experimental and clinical situations;indeed, there is no reason, a priori, to so suppose. Threeseparate models of C5L2 receptor function in comple-ment biology currently exist; two anti-inflammatorymodels and one proinflammatory model. Indeed, thethree models of C5L2 function need not be mutuallyexclusive; all three rely on evidence gathered fromseparate species, cell types, and experimental models.The anti-inflammatory models of C5L2 function havebeen originally based on observations made withoutimmune challenge, whereas the proinflammatory hy-pothesis has its major genesis in observations of C5L2function in septic animals. In addition, the originalobservation regarding the lack of G-protein-couplingmotifs is not unprecedented, for in recent years otherGPCR-classed receptors have been found to signal viapathways other than coupling G proteins, such as�2-adrenergic and AT1a receptors (57). C5L2 may turnout to be the great masquerader of complement-medi-ated inflammology, its function dependent on the celltype or disease present.
Based on the multifarious described roles for C5L2,the therapeutic targeting of C5L2 (be it for activationor inhibition), becomes a more difficult endeavor. Wehave to realize the proper functional role of C5L2 inhost immunity and disease pathology before rationallytargeting it for future clinical treatment. If C5L2 has ananti-inflammatory role, increasing its expression oractivity may be beneficial. However, if it plays a proin-flammatory role, such as suggested in sepsis, thenspecific C5L2 inhibitors or antibodies may bring bene-fits. Perhaps it has all these roles in some situations;perhaps it has some of these roles in all situations.
To date, there is no selective activator of C5L2available, although recent preliminary findings withour collaborating researchers have identified potentialselective C5L2-activating peptides (58), which are cur-rently under further investigation in our laboratories.Polyclonal (22) and monoclonal (18) antibodies havebeen developed to target mouse and human C5L2,respectively. In addition, the C5a mutant A8�71-73,originally reported as an inhibitor of C5aR, is alsoknown to inhibit C5L2 (19). At the pharmacologicallevel, C5L2 is also a difficult receptor to target usingclassical methods; the intracellular location of C5L2means that ligand access is diminished. The lack ofG-protein coupling, clearly defined signaling, and con-sequences of activation also make C5L2 therapeuticscreening a challenge. In addition, C5L2 may also befunctionally dependent on other receptors, meaningthat therapeutics developed from transfected cell linesmay not translate well into the clinical scenario.
Quo vadis. The puzzle with C5L2 is not that it cannotbe effectively studied, but rather that it can be, and has
7BIOLOGICAL ROLES OF C5L2

been, for more than a decade, with the full armamen-tarium of the tools of modern biology, at every level ofinvestigation: molecular, cellular, tissue, and wholeanimal. The puzzle is that it is still such a puzzle. Unlikethe classical receptor for C5a, C5aR, which is bothhistorically and universally acknowledged to be of aproinflammatory bent, the specious nature of C5L2 hasengendered confusion within the community of adepts.This quagmire leads us to propose the following sce-nario: it will remain the proverbial riddle for theproverbial near future, but the reasons for this areneither complex, nor controversial. There have beenmany reports now, from many reputable complementlaboratories, on C5L2. It can be reasonably said thatthere are diverse views and that these are sometimesseemingly contradictory, but this is not so unusual, norso unexpected. But, is this normally the case for a singlediscrete receptor, especially in light of its hugely stud-ied cognate primary receptor, regarding which there islittle disagreement as to its fundamental raison d’être?No, it is not; C5L2 seems different in this way. Pharma-cologically, single receptors have clearly restricted, andsomewhat ideologically comforting, functions, albeitwith varying degrees of cellular expression or locales:C5L2 is apparently breaking free from this ideology.How can one receptor do obverse things in diverse cellsor diverse systems? Because it can—or because it must?
Perhaps we can offer a conciliatory viewpoint forC5L2. It is a strange receptor, in that its behavior seemsso different when studied by different groups in differ-ent systems at different times. But, perhaps, this isindeed the reality, and our simple minds seek a simpleexplanation which is simply not yet available. Or,perhaps it is the Janus receptor, the ultimate masquer-ader at the complement ball, there to confuse andtaunt us to a new level of understanding. Certainly, it isquite unlikely to have evolved for this reason, so thechallenge for us is now clear. We need to clean up thismess—possibly of our own making, and resulting fromour limitations—and resolve whether the situation ismore complex than we allow, or whether we are merelyasking the wrong questions. Neither situation carrieswith it a mutual exclusivity, but nor does it offer a clearset of experiments to solve our dilemma. Rarely has thefunction of a receptor been so troublesome, and it isespecially galling is that it is happening now, when wehave so many technically sophisticated investigativetools to employ in our quest to comprehend. This is theboth our conundrum and our dilemma. Complementis never easy, and C5L2 has made it less easy.
The situation is truly more complicated than we areused to, for we have been led astray by the boss receptorfor C5a, with its relatively simplistic and easily compre-hensible functions, so that we feel dismayed and con-fused that there is no similarly easy nor shining path forclarifying the proper place of this infuriating molereceptor. We are used to receptors (like C5aR) havingstupefyingly clear roles in multiple systems, and we aredisappointed if new receptors like C5L2 do not fall intoour little boxes, because it is all so new and we are not
mentally prepared for the new. Is this because it is arogue mutated GPCR that is no longer coupled to Gproteins for signal transduction? Has this then skewedits function over evolutionary time in all the variouscells and species, so that we now are all at sea in ourprimitive attempts to unravel its place in complementbiology? Perhaps so, or even, probably, more so. Un-comfortably, we do not know, for we have been caughtunawares, and we have now not the mentation, orperhaps force of will, to proceed clearly apace in anygiven direction. We need a new and fresh perspective,one that does not easily gather at cozy internationalmeetings.
The case now seems to us to be this: C5L2 mayindeed have many different roles and functions inmany different systems, cells, tissues, organs, and/orspecies. This is an uncomfortable place to inhabit, forthis will make it difficult to do both predictable andpredictive research, and to reach an enabling consen-sus about C5L2 in the foreseeable future. So rarely isthe challenge so clear, or so great, by a new ligand-binding site for C5a. (Dare we yet call it a receptorwithout clear agreement as to its signaling proper-ties?) However, we seem to agree that all the signs arethat C5L2 is an important player in C5a biology, andthe portents are that agents modulating C5L2 recep-tors may well have therapeutic carry; we just can’t saywhat right now these agents are— or may be, norwhat utility they may have. Agonists, maybe; antago-nists, perhaps—these are the challenges, both prox-imal and distal: They are multifarious, and guidancewill neither come soon nor easily, but we believe thearea to be both intrinsically interesting and worthy ofpursuit, and it could well reap clinical dividends inthe not-so-soon future. The place of C5L2 in comple-ment pathophysiology is yet to be unraveled from itstangled skein, but it certainly seems a worthy andchallenging target at which to aim, if we have arenewed force of clear and determined intent, not tobe defeated by this arriviste.
The authors thank the National Health and Medical Re-search Council of Australia (project grant APP1021413), andthe Australian Research Council (Future Fellowship FT110100332),for supporting their research into C5L2. The authors alsothank Prof. Charles Mackay (Monash University, Melbourne,VIC, Australia) and Dr. Peter Monk (University of SheffieldMedical School, Sheffield, UK) for the gift of the mouse andhuman C5L2 antibodies used in Fig. 2 of this review.
REFERENCES
1. Woodruff, T. M., Nandakumar, K. S., and Tedesco, F. (2011)Inhibiting the C5-C5a receptor axis. Mol. Immunol. 48, 1631–1642
2. Monk, P. N., Scola, A.-M., Madala, P., and Fairlie, D. P. (2007)Function, structure and therapeutic potential of complementC5a receptors. Br. J. Pharmacol. 152, 429–448
3. Ohno, M., Hirata, T., Enomoto, M., Araki, T., Ishimaru, H., andTakahashi, T. A. (2000) A putative chemoattractant receptor,C5L2, is expressed in granulocyte and immature dendritic cells,but not in mature dendritic cells. Mol. Immunol. 37, 407–412
8 Vol. 27 March 2013 LI ET AL.The FASEB Journal � www.fasebj.org

4. Cain, S. A., and Monk, P. N. (2002) The orphan receptor C5L2has high affinity binding sites for complement fragments C5aand C5a des-Arg(74). J. Biol. Chem. 277, 7165–7169
5. Cui, W., Simaan, M., Laporte, S., Lodge, R., and Cianflone, K.(2009) C5a- and ASP-mediated C5L2 activation, endocytosis andrecycling are lost in S323I-C5L2 mutation. Mol. Immunol. 46,3086–3098
6. Johswich, K., Martin, M., Thalmann, J., Rheinheimer, C., Monk,P. N., and Klos, A. (2006) Ligand specificity of the anaphyla-toxin C5L2 receptor and its regulation on myeloid and epithe-lial cell lines. J. Biol. Chem. 281, 39088–39095
7. Prossnitz, E. R., Gilbert, T. L., Chiang, S., Campbell, J. J., Qin, S.,Newman, W., Sklar, L. A., and Ye, R. D. (1999) Multipleactivation steps of the N-formyl peptide receptor. Biochemistry 38,2240–2247
8. Okinaga, S., Slattery, D., Humbles, A., Zsengeller, Z., Morteau,O., Kinrade, M. B., Brodbeck, R. M., Krause, J. E., Choe, H. R.,Gerard, N. P., and Gerard, C. (2003) C5L2, a nonsignaling C5Abinding protein. Biochemistry 42, 9406–9415
9. He, R., Browning, D. D., and Ye, R. D. (2001) Differential rolesof the NPXXY motif in formyl peptide receptor signaling. J.Immunol. 166, 4099–4105
10. Scola, A.-M., Higginbottom, A., Partridge, L. J., Reid, R. C.,Woodruff, T., Taylor, S. M., Fairlie, D. P., and Monk. P. N.(2007) The role of the N-terminal domain of the complementfragment receptor C5L2 in ligand binding. J. Biol. Chem. 282,3664–3671
11. Buck, E., Bourne, H., and Wells, J. A. (2005) Site-specificdisulfide capture of agonist and antagonist peptides on the C5areceptor. J. Biol. Chem. 280, 4009–4012
12. Chen, N. J., Mirtsos, C., Suh, D., Lu, Y. C., Lin, W. J., McKerlie,C., Lee, T., Baribault, H., Tian, H., and Yeh, W. C. (2007) C5L2is critical for the biological activities of the anaphylatoxins C5aand C3a. Nature 446, 203–207
13. Gao, H., Neff, T. A., Guo, R. F., Speyer, C. L., Sarma, J. V.,Tomlins, S., Man, Y., Riedemann, N. C., Hoesel, L. M., Younkin,E., Zetoune, F. S., and Ward, P. A. (2005) Evidence for afunctional role of the second C5a receptor C5L2. FASEB J. 19,1003–1005
14. Kalant, D., MacLaren, R., Cui, W., Samanta, R., Monk, P. N.,Laporte, S. A., and Cianflone, K. (2005) C5L2 is a functionalreceptor for acylation-stimulating protein. J. Biol. Chem. 280,23936–23944
15. Gavrilyuk, V., Kalinin, S., Hilbush, B. S., Middlecamp, A.,McGuire, S., Pelligrino, D., Weinberg, G., and Feinstein, D. L.(2005) Identification of complement 5a-like receptor (C5L2)from astrocytes: characterization of anti-inflammatory proper-ties. J. Neurochem. 92, 1140–1149
16. Woodruff, T. M., Costantini, K. J., Crane, J. W., Atkin, J. D.,Monk, P. N., Taylor, S. M., and Noakes, P. G. (2008) Thecomplement factor C5a contributes to pathology in a rat modelof amyotrophic lateral sclerosis. J. Immunol. 181, 8727–8734
17. Van Werkhoven, M. B., Damman, J., Daha, M. R., Krikke, C., vanGoor, H., van Son, W. J., Hillebrands, J. L., van Dijk, M. C., andSeelen, M. A. (2013) Novel insights in localization and expres-sion levels of C5aR and C5L2 under native and post-transplantconditions in the kidney. Mol. Immunol. 53, 237–245
18. Bamberg, C. E., Mackay, C. R., Lee, H., Zahra, D., Jackson, J.,Lim, Y. S., Whitfeld, P. L., Craig, S., Corsini, E., Lu, B., Gerard,C., and Gerard, N. P. (2010) The C5a receptor (C5aR) C5L2 isa modulator of C5aR-mediated signal transduction. J. Biol. Chem.285, 7633–7644
19. Otto, M., Hawlisch, H., Monk, P. N., Müller, M., Klos, A., Karp,C. L., and Köhl, J. (2004) C5a mutants are potent antagonists ofthe C5a receptor (CD88) and of C5L2: position 69 is the locusthat determines agonism or antagonism. J. Biol. Chem. 279,142–151
20. Scola, A.-M., Johswich, K.-O., Morgan, B. P., Klos, A., and Monk,P. N. (2009) The human complement fragment receptor, C5L2,is a recycling decoy receptor. Mol. Immunol. 46, 1149–1162
21. Huber-Lang M., Sarma, J. V., Rittirsch, D., Schreiber, H., Weiss,M., Flierl, M., Younkin, E., Schneider, M., Suger-Wiedeck, H.,Gebhard, F., McClintock, S. D., Neff, T., Zetoune, F., Bruckner,U., Guo, R. F., Monk, P. N., and Ward, P. A. (2005) Changes inthe novel orphan, C5a receptor (C5L2), during experimentalsepsis and sepsis in humans. J. Immunol. 174, 1104–1110
22. Rittirsch, D., Flierl, M. A., Nadeau, B. A., Day, D. E., Huber-Lang, M., Mackay, C. R., Zetoune, F. S., Gerard, N. P., Cianf-lone, K., Köhl, J., Gerard, C., Sarma, J. V., and Ward, P. A.(2008) Functional roles for C5a receptors in sepsis. Nat. Med. 14,551–557
23. Peng, Q., Li, K., Wang, N., Li, Q., Asgari, E., Lu, B., Woodruff,T. M,, Sacks, S. H., and Zhou, W. (2009) Dendritic cell functionin allostimulation is modulated by C5aR signaling. J. Immunol.183, 6058–6068
24. Raby, A. C., Holst, B., Davies, J., Colmont, C., Laumonnier, Y.,Coles, B., Shah, S., Hall, J., Topley, N., Köhl, J., Morgan, B. P.,Labéta, M. O. (2011) TLR activation enhances C5a-inducedpro-inflammatory responses by negatively modulating the sec-ond C5a receptor, C5L2. Eur. J. Immunol. 41, 2741–2752
25. Huber-Lang, M., Sarma, J. V., Zetoune, F. S., Rittirsch, D., Neff,T. A., McGuire, S. R., Lambris, J. D., Warner, R. L., Flierl, M. A.,Hoesel, L. M., Gebhard, F., Younger, J. G., Drouin, S. M., Wetsel,R. A., and Ward, P. A. (2006) Generation of C5a in the absenceof C3: a new complement activation pathway. Nat. Med. 12,682–687
26. MacLaren, R., Kalant, D., and Cianflone, K. (2007) The ASPreceptor C5L2 is regulated by metabolic hormones associatedwith insulin resistance. Biochem. Cell Biol. 85, 11–21
27. Liu, H.-B., Cui, N.-Q., Wang, Q., Li, D.-H., and Xue, X.-P. (2008)Sphingosine-1-phosphate and its analogue FTY720 diminishacute pulmonary injury in rats with acute necrotizing pancreati-tis. Pancreas 36, e10–e15
28. McVerry, B. J., and Garcia, J. G. N. (2004) Endothelial cellbarrier regulation by sphingosine 1-phosphate. J. Cell. Biochem.92, 1075–1085
29. McVerry, B. J., Peng, X., Hassoun, P. M., Sammani, S., Simon,B. A., and Garcia, J. G. (2004) Sphingosine 1-phosphate reducesvascular leak in murine and canine models of acute lung injury.Am. J. Respir. Crit. Care Med. 170, 987–993
30. Bachmaier, K., Guzman, E., Kawamura, T., Gao, X., and Malik,A. B. (2012) Sphingosine kinase 1 mediation of expression ofthe anaphylatoxin receptor C5L2 dampens the inflammatoryresponse to endotoxin. PLoS ONE 7, e30742
31. Farkas, I., Sárvári, M., Aller, M., Okada, N., Okada, H., Likó, I.,and Liposits, Z. (2012) Estrogen receptor alpha and betadifferentially mediate C5aR agonist evoked Ca(2�)-influx inneurons through L-type voltage-gated Ca(2�) channels. Neuro-chem. Int. 60, 631–639
32. Cui, W., Lapointe, M., Gauvreau, D., Kalant, D., and Cianflone,K. (2009) Recombinant C3adesArg/acylation stimulating pro-tein (ASP) is highly bioactive: a critical evaluation of C5L2binding and 3T3-L1 adipocyte activation. Mol. Immunol. 46,3207–3217
33. Riedemann, N. C., Guo, R. F., Hollmann, T. J., Gao, H., Neff,T. A., Reuben, J. S., Speyer, C. L., Sarma, J. V., Wetsel, R. A.,Zetoune, F. S., and Ward, P. A. (2004) Regulatory role of C5a inLPS-induced IL-6 production by neutrophils during sepsis.FASEB J. 18, 370–372
34. Van Lith, L. H. C., Oosterom, J., Van Elsas, A., and Zaman,G. J. R. (2009) C5a-stimulated recruitment of beta-arrestin2 tothe nonsignaling 7-transmembrane decoy receptor C5L2. J.Biomol. Screen. 14, 1067–1075
35. Zhang, X., Schmudde, I., Laumonnier, Y., Pandey, M. K., Clark,J. R., König, P., Gerard, N. P., Gerard, C., Wills-Karp, M., andKöhl, J. (2010) A critical role for C5L2 in the pathogenesis ofexperimental allergic asthma. J. Immunol. 185, 6741–6752
36. Gerard, N. P., Lu, B., Liu, P., Craig, S., Fujiwara, Y., Okinaga, S.,and Gerard, C. (2005) An anti-inflammatory function for thecomplement anaphylatoxin C5a-binding protein, C5L2. J. Biol.Chem. 280, 39677–39680
37. Huber-Lang, M. S., Younkin, E. M., Sarma, J. V., McGuire, S. R.,Lu, K. T., Guo, R. F., Padgaonkar, V. A., Curnutte, J. T.,Erickson, R., Ward, P. A. (2002) Complement-induced impair-ment of innate immunity during sepsis. J. Immunol. 169, 3223–3231
38. Woodruff, T. M., Strachan, A. J., Dryburgh, N., Shiels, I. A.,Reid, R. C., Fairlie, D. P., Taylor, S. M. (2002) Antiarthriticactivity of an orally active C5a receptor antagonist againstantigen-induced monarticular arthritis in the rat. ArthritisRheum. 46, 2476–2485
39. Abe, M., Shibata, K., Akatsu, H., Shimizu, N., Sakata, N.,Katsuragi, T., and Okada, H. (2001) Contribution of anaphyla-
9BIOLOGICAL ROLES OF C5L2

toxin C5a to late airway responses after repeated exposure ofantigen to allergic rats. J. Immunol. 167, 4651–4660
40. Baelder, R., Fuchs, B., Bautsch, W., Zwirner, J., Köhl, J., Hoy-mann, H. G., Glaab, T., Erpenbeck, V., Krug, N., and Braun, A.(2005) Pharmacological targeting of anaphylatoxin receptorsduring the effector phase of allergic asthma suppresses airwayhyperresponsiveness and airway inflammation. J. Immunol. 174,783–789
41. Lambrecht, B. N. (2006) An unexpected role for the anaphyla-toxin C5a receptor in allergic sensitization. J. Clin. Invest. 116,628–632
42. Arumugam, T. V., Shiels, I. A., Strachan, A. J., Abbenante, G.,Fairlie, D. P., and Taylor, S. M. (2003) A small molecule C5areceptor antagonist protects kidneys from ischemia/reperfu-sion injury in rats. Kidney Int. 63, 134–142
43. Proctor, L. M., Arumugam, T. V., Shiels, I., Reid, R. C., Fairlie,D. P., and Taylor, S. M. (2004) Comparative anti-inflammatoryactivities of antagonists to C3a and C5a receptors in a rat modelof intestinal ischaemia/reperfusion injury. Br. J. Pharmacol. 142,756–764
44. Woodruff, T. M., Arumugam, T. V., Shiels, I. A., Reid, R. C.,Fairlie, D. P., and Taylor, S. M. (2004) Protective effects of apotent C5a receptor antagonist on experimental acute limbischemia-reperfusion in rats. J. Surg. Res. 116, 81–90
45. Woodruff, T. M., Arumugam, T. V., Shiels, I. A., Reid, R. C.,Fairlie, D. P., and Taylor, S. M. (2003) A potent human C5areceptor antagonist protects against disease pathology in a ratmodel of inflammatory bowel disease. J. Immunol. 171, 5514–5520
46. Johswich, K., Martin, M., Bleich, A., Kracht, M., Dittrich-Breiholz, O., Gessner, J. E., Suerbaum, S., Wende, E.,Rheinheimer, C., and Klos, A. (2009) Role of the C5a receptor(C5aR) in acute and chronic dextran sulfate-induced models ofinflammatory bowel disease. Inflamm. Bowel Dis. 15, 1812–1823
47. Jain, U., Woodruff, T. M., and Stadnyk, A. W. (2012) The C5areceptor antagonist PMX205 ameliorates experimentally in-duced colitis associated with increased IL-4 and IL-10. Br. J.Pharmacol. doi:10.1111/j.1476-5381.2012.02183.x
48. Manthey, H. D., Thomas, A. C., Shiels, I. A., Zernecke, A.,Woodruff, T. M., Rolfe, B., and Taylor, S. M. (2011) Comple-ment C5a inhibition reduces atherosclerosis in ApoE�/� mice.FASEB J. 25, 2447–2455
49. Woodruff, T. M., Crane, J. W., Proctor, L. M., Buller, K. M.,Shek, A. B., de Vos, K., Pollitt, S., Williams, H. M., Shiels, I. A.,
Monk, P. N., and Taylor, S. M. (2006) Therapeutic activity ofC5a receptor antagonists in a rat model of neurodegeneration.FASEB J. 20, 1407–1417
50. Fonseca, M. I., Ager, R. R., Chu, S. H., Yazan, O., Sanderson,S. D., LaFerla, F. M., Taylor, S. M., Woodruff, T. M., and Tenner,A. J. (2009) Treatment with a C5aR antagonist decreasespathology and enhances behavioral performance in murinemodels of Alzheimer’s disease. J. Immunol. 183, 1375–1383
51. Pavlovski, D., Thundyil, J., Monk, P. N., Wetsel, R. A., Taylor,S. M., and Woodruff, T. M. (2012) Generation of complementcomponent C5a by ischemic neurons promotes neuronal apo-ptosis. FASEB J. 26, 3680–3690
52. Flierl, M. A., Rittirsch, D., Chen, A. J., Nadeau, B. A., Day, D. E.,Sarma, J. V., Huber-Lang, M. S., and Ward, P. A. (2008) Thecomplement anaphylatoxin C5a induces apoptosis in adreno-medullary cells during experimental sepsis. PLoS ONE 3, e2560
53. Fisette, A., Munkonda, M. N., Oikonomopoulou, K., Pagli-alunga, S., Lambris, J. D., and Cianflone, K. (2012) C5L2receptor disruption enhances the development of diet-inducedinsulin resistance in mice. [E-pub ahead of print] Immunobiologydoi:10.1016/j.imbio.2012.04.001
54. Wen, Y., Wang, H. W., Hu, X. F., Cianflone, K., Wei, J., Xia, Z.,and Li, R. Z. (2008) [Role of progesterone in acylation stimu-lating protein-receptor C5L2 pathway in adipocytes and preadi-pocytes]. Zhonghua Yi Xue Za Zhi 88, 114–118
55. Francis, K., Lewis, B. M., Monk, P. N., and Ham, J. (2008)Complement C5a receptors in the pituitary gland: expressionand function. J. Endocrinol. 199, 417–424
56. Roy, C., Roy, M. C., Gauvreau, D., Poulin, A. M., Tom, F. Q.,Timofeeva, E., Richard, D., and Cianflone, K. (2011) Acuteinjection of ASP in the third ventricle inhibits food intake andlocomotor activity in rats. Am. J. Physiol. Endocrinol. Metab. 301,E232–E241
57. Shukla, S. D., Velazquez, J., French, S. W., Lu, S. C., Ticku,M. K., and Zakhari, S. (2008) Emerging role of epigenetics inthe actions of alcohol. Alcohol. Clin. Exp. Res. 32, 1525–1534
58. Monk, P. N., Bellows-Peterson, M. L., Smadbeck, J., Floudas,C. A., Kieslich, C. A., Morikis, D., and Woodruff, T. M. (2012)De novo protein design of agonists and antagonists of C5areceptors. Immunobiology 217, 1162–1163
Received for publication October 25, 2012.Accepted for publication November 19, 2012.
10 Vol. 27 March 2013 LI ET AL.The FASEB Journal � www.fasebj.org




















