
BOLETÍN CIENTÍFICO CENTRO DE MUSEOS MUSEO DE ...
266
BOLETÍN CIENTÍFICO CENTRO DE MUSEOS MUSEO DE HISTORIA NATURAL Vol. 21 No. 2 SCIENTIFIC BULLETIN MUSEUM CENTER NATURAL HISTORY MUSEUM Vol. 21 No. 2 bol.cient.mus.his.nat. Manizales (Colombia) Vol. 21 No. 2 266 p. julio-diciembre de 2017 ISSN 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of BOLETÍN CIENTÍFICO CENTRO DE MUSEOS MUSEO DE ...

Boletín CientífiCoCentro de Museos
Museo de Historia natural Vol. 21 no. 2
sCientifiC Bulletin MuseuM Center
natural History MuseuMVol. 21 no. 2
bol.cient.mus.his.nat. Manizales (Colombia) Vol. 21 No. 2 266 p. julio-diciembre de 2017
ISSN 0123-3068(Impreso)
ISSN: 2462-8190 (En línea)



ISSN 0123–3068 (Impreso)ISSN: 2462-8190 (En línea)-Fundada en 1995-Nueva periodicidad semestralTiraje 150 ejemplaresVol. 21 No. 2, 266 p.julio-diciembre, 2017Manizales - Colombia
Boletín CientífiCoCentro de Museos
Museo de Historia natural
Felipe César Londoño López Orlando Londoño BetancourtLuisa Fernanda Giraldo ZuluagaAura Liliana Gaviria GiraldoAndrés Felipe Betancourth LópezMarco Tulio Jaramillo SalazarOlga Lucía Hurtado
rectorVicerrector académico
Vicerrectora de investigaciones y PostgradosVicerrectora administrativa
Vicerrector de Proyeccióndecano facultad de Ciencias exactas y naturales
Centro de Museos
Revista especializada en estudios de Historia Natural y áreas biológicas afines.
Boletín Científico Centro de Museos
Museo de Historia natural
Ricardo Walkerinvestigador, fundador Boletín Científico Museo de Historia natural, universidad de Caldas
Luis Carlos Pardo-Locarnoingeniero agronómo, Ph.d., Msc., Ciat Palmira, ValleJohn Harold CastañoMsc. Programa Biología,universidad de Caldas
Luís M. Constantino entomólogo, Msc., Centro de investigaciones para el café -CeniCafÉ -
Cristobal Ríos M.Biólogo, Centro de ecologíaiViC, Caracas, Venezuela
Gabriel Jaime Castañoingeniero forestal, departamento de recursos naturales y Medio ambiente, universidad de Caldas
Ricardo Álvarez LeónBiólogo Marino, Msc. fundación Maguaré, Manizales
Cómite editorialÁngel L. Viloria Biólogo-Zoólogo, Ph.d., Centro de ecología, iViC, Venezuela
Tomasz Pyrczentomólogo, Ph.d., Museo de Zoología universidad Jaguellónica, Polonia
Zsolt BálintBiologo, Ph.d., Museo de Historia natural de Budapest, Hungría
Carlos López Vaamondeingeniero agrónomo, entomólogo, Msc., Ph.d., Bsc. Colegio imperial de londres, uK
George BeccaloniZoologo, Ph.d., Bsc.- Colegio imperial de londres, uK
Olaf Hermann H. MielkeZoólogo, Ph.d., departamento de Zoología, universidad federal de Paraná, Brasil
Roger Royentomólogo, Ph.d., Museo de Historia natural de París, francia
Cómite internacional
Julián A. Salazar E.Médico Veterinario & Zootecnista (MVZ). universidad de Caldas, Centro de Museos.
director
Publindex Categoría A2Zoological RecordScieloIndex CopernicusScopus
indexada por

Juan David Giraldo MárquezJuan David López GonzálezCamilo Sepúlveda Betancurth Silvia L. SpaggiariCarlos Fernando Nieto BetancurCarlos Eduardo Tavera Pinzón
Comité técnico de apoyo a la ediciónCoordinador Comité técnico
diagramaciónCorrección de estilo
traducción de resúmenes al inglésimplementación metodología scielo
soporte tecnológico
Vicerrectoría de investigaciones y PostgradosUniversidad de Caldas Sede CentralCalle 65 No. 26-10275(+6) 8781500 ext. 11222 - [email protected]@ucaldas.edu.coManizales – Colombia
Ventas, suscripciones y Canjes
dirección:apartado aéreo:
teléfonos:e-mail:
Universidad de CaldasVicerrectoría de Investigaciones y Postgrados
edición
Avispa Conura de la familia Chalcididae parasitando una larva de Leptinotarsa-Coleoptera. Foto: Francisco Arango
El contenido de esta publicación puede reproducirse citando la fuente.
La responsabilidad de lo expresado en cada artículo es exclusiva del autor y no expresa ni compromete la posición
de la revista.
PatrocinadoresUniversidad de Caldas Vicerrectoría de Investigaciones y Postgrados
Centro de MuseosFacultad de Ciencias Exactas y Naturales
América PintorescaCréditos Separadores
http://boletincientifico.ucaldas.edu.cositio Web
fotografía Portada


BotÁniCaBotany
Contenido
ConserVaCiÓn y BiodiVersidaden aGroeCosisteMas
Conservation and Biodiversity in Agroecosystems
bol.cient.mus.his.nat. Manizales (Colombia) Vol. 21 No. 2 266 p. julio-diciembre de 2017
ISSN 0123-3068(Impreso)
ISSN: 2462-8190 (En línea)
CatÁloGo de la flora (anGiosPerMas) del PÁraMo de BiJaGual, BoYaCÁ, ColoMBia
CataloGue of tHe flora (anGiosPerMae) of tHe PÁraMo de BiJaGual, BoYaCÁ, ColoMBia
Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón
15
PriMer reGistro de Hydnopolyporus fimbriatus (Cooke) d.a. reid (PolYPorales: MeriPilaCeae) Para el dePartaMento de CundinaMarCa, ColoMBia
first rePort of Hydnopolyporus fimbriatus (Cooke) d.a. reid (PolYPorales: MeriPilaCeae) for tHe dePartMent of
CundinaMarCa, ColoMBia
Andrea Castiblanco-Z., César Augusto Pinzón-o., Jonás Pinzón-o.
30
anÁlisis de la ContaMinaCiÓn Por ColiforMes terMotolerantes en el estuario del río
ranCHería, la GuaJira (ColoMBia)
analYsis of ContaMination BY tHerMotolerant ColiforM BaCteria in tHe estuarY of tHe ranCHeria riVer, la GuaJira
(ColoMBia)
Geomar Molina-Bolívar, Iris Jiménez-Pitre
41
Control alternatiVo de Diaphorina citri KuWaYaMa (HeMiPtera: liViidae) utiliZando Caldo
sulfoCÁlCiCo
alternatiVe Control of Diaphorina citri KuWaYaMa (HeMiPtera: liViidae) usinG liMe sulPHur
Ana María Restrepo-García, Alberto Soto-Giraldo51

ZooloGía VerteBradosVertebrate Zoology
enferMedades en ViVeros CoMerCiales de Musa aaB ‘doMiniCo HartÓn’ en el dePartaMento de
Caldas, ColoMBia
diseases in CoMMerCial nurseries of Musa aaB ‘doMiniCo HartÓn’ in tHe dePartMent of Caldas, ColoMBia
Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada
61
diVersidad de las MosCas de las frutas (diPtera: tePHritidae) Y sus Parasitoides en siete
MuniCiPios del dePartaMento de nariÑo
diVersitY of fruit-flies (diPtera: tePHritidae) and tHeir Parasitoids in seVen MuniCiPalities of tHe dePartMent
of nariÑo
María Isabel Cruz B, Tito Bacca, Nelson A. Canal81
BioloGía reProduCtiVa de Sicalis flaveola (aVes: tHrauPidae) en Cali, ColoMBia
reProduCtiVe BioloGY of Sicalis flaveola (aVes: tHrauPidae) in Cali, ColoMBia
Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto
101
Bird asseMBle assoCiated to tWo landsCaPe units in a troPiCal drY forest
ensaMBle de aVes asoCiado a dos unidades de PaisaJe en un BosQue seCo troPiCal
Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo,
Alan Giraldo115

estudios de orGanisMos del BosQue HuMedo troPiCal
Organisms studies of the tropical rainforest
el BosQue de BaVaria (VillaViCenCio, Meta), re-Visitado: nueVos reGistros aÑadidos a la lista
General de esPeCies (lePidoPtera: rHoPaloCera)
tHe BaVaria forest (VillaViCenCio, Meta) reVisited: neW reCords added to tHe CHeCKlist of sPeCies
(lePidoPtera: rHoPaloCera)
Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl
147
una nueVa esPeCie de Pharneuptychia forster, 1964 del Área natural ÚniCa los estoraQues –norte de santander, ColoMBia– (lePidoPtera:
nYMPHalidae: satYrinae)
a neW sPeCies of Pharneuptychia forster, 1964 froM tHe estoraQues natural area, nortH of santander-
ColoMBia (lePidoPtera: nYMPHalidae: satYridae)
efraín Reinel Henao-Bañol, luis Hernando Meneses
173
ZooloGía inVerteBradosInvertebrate Zoology
dietas VeGetales en el desarrollo larVal de dos esPeCies de Morpho (lePidoPtera: nYMPHalidae,
satYrinae, MorPHini) en CautiVerio, en iQuitos, loreto, PerÚ
VeGetaBle diets on tHe larVal deVeloPMent of tWo Morpho sPeCies (lePidoPtera: nYMPHalidae, satYrinae,
MorPHini) in CaPtiVitY in iQuitos, loreto, Peru
Joel Vásquez, Wenceslao Peña, Ricardo Zárate, Julio Pinedo, Rocío Correa, Juan José Palacios,
Juan José Bellido, Juan José Ramírez, Gerardo lamas133
nueVo reGistro de CorYPHorus (inseCta: ePHeMeroPtera) Y notas adiCionales soBre su
MorfoloGía
neW reCord of CorYPHoridae (inseCta: ePHeMeroPtera) and additional notes on its MorPHoloGY
Tatiana Cárdenas lópez, Ximena ospina londoño, Miguel Rodríguez Zambrano, María Paula Rozo, Rodrigo Ivan Romero Zúñiga, lucimar Gomes Dia
184

noVedades en Historia naturalnatural HistorY neWs
instruCCiones a los autoresautHor Guidelines
245
255
autoresautHors
251
distriBuCiÓn PotenCial de Oxysternon conspicillatum (WeBer, 1801) en diferentes esCenarios de CaMBio
CliMÁtiCo en ColoMBia
Potential distriBution of Oxysternon conspicillatum (WeBer, 1801) in different CliMate CHanGe sCenarios in ColoMBia
Bedir Germán Martínez-Quintero, Alejandro echeverri-Rubio, Fabián Guillermo Gaviria-ortiz
190
neW sPeCies of tHe Genus Trigonospila PoKornY (diPtera: taCHinidae) Parasitises adult Compsus
sp. sCHoenHerr (ColeoPtera: CurCulionidae) in ColoMBia
nueVa esPeCie del GÉnero Trigonospila PoKornY (diPtera: taCHinidae) Para ColoMBia, Parasitando adultos de Compsus sp. sCHoenHerr (ColeoPtera: CurCulionidae)
Natalia Vinasco, luis Fernando Vallejo, Alberto Soto
207
Sphaeropthalma pinedai sp. nov.: PriMer reGistro de un Mutílido BraQuíPtero Para aMÉriCa del sur
(HYMenoPtera: Mutillidae)
Sphaeropthalma pinedai sp. nov.: first reCord of BraCHYPterous Mutillid for soutH aMeriCa (HYMenoPtera: Mutillidae)
Roberto A. Cambra, Diomedes Quintero A.
220
diVersidad de esCorPiones (CHeliCerata) en el dePartaMento del atlÁntiCo, ColoMBia
diVersitY of sCorPions (CHeliCerata) in tHe dePartMent of atlÁntiCo, ColoMBia
eider Sará-C., eduardo Flórez-D., Neis Martínez-H.
231

“Un hombre sin pasiones, está tan cerca de la estupidez que solo le falta abrir la boca para caer en ella”
Séneca (4 A.C- 65)


BotÁniCaBotany


Boletín CientífiCoCentro de Museos
Museo de Historia natural
CatÁloGo de la flora (anGiosPerMas) del PÁraMo de BiJaGual, BoYaCÁ, ColoMBia*
Merly Yenedith Carrillo Fajardo1, María eugenia Morales-Puentes2, Pablo Andrés Gil-leguizamón3
resumen
Se elaboró un catálogo de las angiospermas del páramo de Bijagual con información de las especies, distribución local y nacional. Se registraron 35 familias, 73 géneros y 120 especies.
Palabras clave: alta montaña, distribución, diversidad, plantas vasculares.
CataloGue of tHe flora (anGiosPerMae) of tHe PÁraMo de BiJaGual, BoYaCÁ, ColoMBia
abstract
An angiosperms catalogue of the Bijagual moor was prepared with the information of the species, and their local and national distribution. Thirty-five families, 73 genera and 120 species were recorded.
Key words: high mountain, distribution, diversity, vascular plants.
* FR: 24-I-17. FA: 28-VIII-17.1 Sistemática Biológica, Herbario UPTC, Universidad Pedagógica y Tecnológica de Colombia & Universidad de Córdoba. Colombia. E-mail: [email protected] ORCID: 0000-0003-0725-57032 Sistemática Biológica, Herbario UPTC, Escuela de Ciencias Biológicas, Facultad de Ciencias, Universidad Pedagógica y Tecnológica de Colombia. Tunja, Colombia. E-mail: [email protected] ORCID: 0000-0002-5332-99563 Sistemática Biológica, Herbario UPTC, Escuela de Ciencias Biológicas, Facultad de Ciencias, Universidad Pedagógica y Tecnológica de Colombia. Tunja, Colombia. E-mail: [email protected] ORCID: 0000-0002-9414-8991
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 15-29. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: CARRILLO, M.Y., MORALES-PUENTES, M.E. & GIL-LEGUIZAMÓN, P.A., 2017.- Catálogo de la flora (angios-permas) del páramo de Bijagual, Boyacá, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 15-29. DOI: 10.17151/bccm.2017.21.2.1

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón16
introduCCiÓn
En Colombia los páramos corresponden al 3% del territorio, equivalente a 2’907.137 ha, es decir, el 50% de las coberturas de este ecosistema a nivel mundial (SARMIENTO et al., 2013). El conocimiento de la vegetación paramuna colombiana tiene sus inicios con las investigaciones de CUATRECASAS (1934), desde entonces, el trabajo en los páramos ha sido continuo, dado que aún existen lugares que no han sido reconocidos florísticamente (PÉREZ-MORENO et al., 2016).
La flora paramuna presenta altos niveles de endemismo (90%) y al menos el 60% del inventario del país (HERNÁNDEZ et al., 1992; MIRANDA-ESQUIVEL et al., 2002; DÍAZ-GRANADOS et al., 2005; RANGEL-CH., 2007; ARELLANO-P. & RANGEL-CH., 2010; RIVERA-OSPINA & RODRÍGUEZ, 2011; MADS, 2012; SARMIENTO & LEÓN, 2015). El páramo de Bijgual es reconocido como un conector biológico de los ecosistemas de alta montaña al sur de Boyacá (CIRI, 2003; CORPOCHIVOR, 2006; UNIVERSIDAD MILITAR NUEVA GRANADA & CORPOCHIVOR, 2008; CARRILLO-FAJARDO, 2013). El objetivo de este catálogo es dar a conocer la flora vascular de este ecosistema estratégico para el país.
Área de estudio
El macizo de Bijagual está ubicado en la Provincia de páramos de la Cordillera Oriental colombiana (LONDOÑO et al., 2014), en jurisdicción de los municipios de Viracachá, Ciénega, Ramiriquí y Tibaná, sobre el costado occidental de Zetaquira y oriental de Rondón (Boyacá); cuenta con una extensión de 8.605 ha, distribuidas entre 2400-3400 (±3530) m, con ecosistemas de bosque alto andino y páramo (MORALES-PUENTES et al., 2012). Las precipitaciones anuales varían entre 1000-2500 mm y tienen distribución monomodal entre abril y noviembre, con un leve descenso en agosto y septiembre; la temperatura es de 13-17ºC y varía según la altitud, también la humedad relativa es alta, aunque no hay datos exactos (SIMA et al., 1996).
MÉtodos
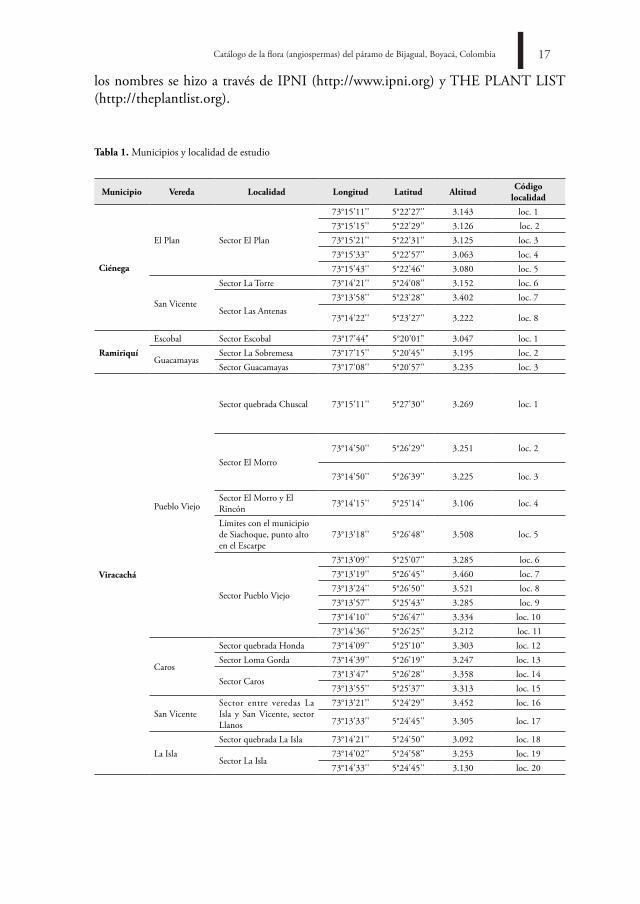
trabajo de campo. Se realizaron colecciones generales en 31 sitios de Bijagual en los municipios de Viracachá, Ramiriquí y Ciénega, en coberturas de frailejonal, pajonal y arbustal, en un rango altitudinal entre 3000-3521 m (Tabla 1) en época seca y lluviosa.
fase de laboratorio. Se llevó a cabo en el Herbario UPTC, y se siguió el protocolo de herborización. La determinación se realizó con bibliografía especializada y comparación en herbarios: UPTC, HECASA y FMB, en línea COL (http://www.biovirtual.unal.edu.co), MO (http://www.tropicos.org), NY (http://www.nybg.org) y KEY (http://www.kew.org). Asimismo, se contó con la asesoría de especialistas. La verificación de
Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 17
los nombres se hizo a través de IPNI (http://www.ipni.org) y THE PLANT LIST (http://theplantlist.org).
tabla 1. Municipios y localidad de estudio
Municipio Vereda localidad longitud latitud altitud Código localidad
Ciénega
El Plan Sector El Plan
73°15'11'' 5°22'27'' 3.143 loc. 173°15'15'' 5°22'29'' 3.126 loc. 273°15'21'' 5°22'31'' 3.125 loc. 373°15'33'' 5°22'57'' 3.063 loc. 473°15'43'' 5°22'46'' 3.080 loc. 5
San Vicente
Sector La Torre 73°14'21'' 5°24'08'' 3.152 loc. 6
Sector Las Antenas73°13'58'' 5°23'28'' 3.402 loc. 7
73°14'22'' 5°23'27'' 3.222 loc. 8
ramiriquí
Escobal Sector Escobal 73°17'44" 5°20'01" 3.047 loc. 1
GuacamayasSector La Sobremesa 73°17'15'' 5°20'45'' 3.195 loc. 2Sector Guacamayas 73°17'08'' 5°20'57'' 3.235 loc. 3
Viracachá
Pueblo Viejo
Sector quebrada Chuscal 73°15'11'' 5°27'30'' 3.269 loc. 1
Sector El Morro73°14'50'' 5°26'29'' 3.251 loc. 2
73°14'50'' 5°26'39'' 3.225 loc. 3
Sector El Morro y El Rincón 73°14'15'' 5°25'14'' 3.106 loc. 4
Límites con el municipio de Siachoque, punto alto en el Escarpe
73°13'18'' 5°26'48'' 3.508 loc. 5
Sector Pueblo Viejo
73°13'09'' 5°25'07'' 3.285 loc. 673°13'19'' 5°26'45'' 3.460 loc. 773°13'24'' 5°26'50'' 3.521 loc. 873°13'57'' 5°25'43'' 3.285 loc. 973°14'10'' 5°26'47'' 3.334 loc. 1073°14'36'' 5°26'25'' 3.212 loc. 11
Caros
Sector quebrada Honda 73°14'09'' 5°25'10'' 3.303 loc. 12Sector Loma Gorda 73°14'39'' 5°26'19'' 3.247 loc. 13
Sector Caros73°13'47" 5°26'28'' 3.358 loc. 1473°13'55'' 5°25'37'' 3.313 loc. 15
San VicenteSector entre veredas La Isla y San Vicente, sector Llanos
73°13'21'' 5°24'29'' 3.452 loc. 16
73°13'33'' 5°24'45'' 3.305 loc. 17
La IslaSector quebrada La Isla 73°14'21'' 5°24'50'' 3.092 loc. 18
Sector La Isla73°14'02'' 5°24'58'' 3.253 loc. 1973°14'33'' 5°24'45'' 3.130 loc. 20

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón18
elaboración del catálogo. La información incluida presenta las monocotiledóneas y luego las dicotiledóneas. Las familias están organizadas en orden alfabético, según CRONQUIST (1988) y APG IV (2016) con la información del orden, y sí existe, la modificación a nivel de familia. Asimismo, la citación de cada familia está acompañada por el autor, el año de publicación y el número de géneros y especies que se registran en el área de estudio; nombre de la especie, sinónimo o basónimo (cuando sea necesario); hábito de crecimiento según FONT QUER (2000); distribución nacional sobre las tres cordilleras –Oriental (Or.), Central (C.) y Occidental (Oc.)– según BERNAL et al. (2016), consultas virtuales (SIB, COL, MO y NY) y las colecciones FMB, HECASA y UPTC; rango altitudinal; material revisado, departamento (Boyacá), municipio (Ciénega, Ramiriquí y Viracachá), localidad incluye coordenadas y altitud (Tabla 1), fecha de colección y número del colector principal.
resultados
En este catálogo se registran 35 familias, 73 géneros y 120 especies.
Catálogo de la flora vascular del páramo de Bijagual (Sistema de clasificación: 1) CRONQUIST, 1988. 2) APG IV, 2016).
Monocotiledónea
BroMeliaCeae Juss., 1789, Bromeliales¹, Poales² (1 género/2 especies).Puya goudotiana Mez. Hierba. En los Andes (Or.), 2760-3550 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1408 (UPTC). Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1771 (UPTC). Puya. sp. Hierba. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1701 (UPTC).CYPeraCeae Juss., 1789, Cyperales¹, Poales² (4/4).Carex cf. confertospicata Boeckeler. Hierba. En los Andes (Or. y C.), 2880-3725 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1823 (UPTC).Cyperus rufus Kunth. Hierba. En los Andes (Or.), 2250-3500 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1827 (UPTC).Oreobolus goeppingeri Suess. Hierba. Boyacá, Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1762 (UPTC). Rhynchospora ruiziana Boeckeler. Hierba. En los Andes (Or., C. y Occ.), 1200-4000 m. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2636 (UPTC).erioCaulaCeae Martinov, 1820, Eriocaulales¹, Poales² (1/3).Paepalanthus columbiensis Ruhland. Hierba. En los Andes (Or. y C.), 1900-3600 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1694 (UPTC); loc. 15, 3 abr 2012, Gil-l., et al. 1733 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1777 (UPTC);

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 19
loc. 4, 23 ago 2012, Gil-l., et al. 2616 (UPTC). P. pilosus (Kunth) Kunth. Hierba. En los Andes (Or. y C.). 2550-3300 m. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2637 (UPTC).JunCaCeae Juss., 1789, Juncales¹, Poales² (1/2).Juncus effusus L. Hierba. En los Andes (Or., C. y Occ.), 1800-3950 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1821 (UPTC). J. liebmanni var. quitensis (Buchenau) Balslev. Hierba. En los Andes (Occ.), 2500-2830 m. Boyacá, Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1773 (UPTC). liliaCeae Dumort., 1829, Liliales¹ y ² (2/3).Bomarea hirsuta (Kunth) Herb. Trepadora. En los Andes (Or., C. y Occ.), 1700-4000 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2613 (UPTC).B. setacea (Ruiz & Pav.). Hierb. Trepadora. En los Andes (Or., C. y Occ.), 1700-4200 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1920 (UPTC); Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1706 (UPTC); loc. 7, 22 ago 2012, Gil-l., et al. 2597 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1719 (UPTC). Eccremis coarctata (Ruiz & Pav.) Baker. Hierba. En los Andes (Or.), 1990-3100 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1828 (UPTC).orCHidaCeae Juss. 1789, Orchidales¹, Asparagales² (5/12).Cyrtochilum ramosissimum (Lindl.) Dalström. sinónimos: odontoglossum ramosissimum Lindl. Epífita. Andes (Or.), 2300-2600 m. Boyacá, Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1731 (UPTC).Epidendrum decurviflorum Schltr. Epífita. Andes (Or., C. y Occ.), 2000-3360 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1683 (UPTC); loc. 9, 3 abr 2012, Gil-l., et al. 1756 (UPTC). E. aff. frutex Rchb. f. Epífita. En los Andes (Or., C. y Occ.), 2500-3899 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1919 (UPTC). Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1730 (UPTC).Epidendrum sp. 1. Hierba. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1923 (UPTC).Epidendrum sp. 2. Hierba. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1691 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1716 (UPTC).Epidendrum sp. 3. Epífita. Boyacá, Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1758 (UPTC).Epidendrum sp. 4. Hierba. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2598 (UPTC).Epidendrum sp. 5. Hierba. Boyacá, Ciénega, loc. 7, 24 ago 2012, Gil-l., et al. 2660 (UPTC).Fernandezia lanceolata (L.O. Williams) Garay & Dunst. Epífita. En los Andes (Or.),

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón20
2700-3000 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 1855 (UPTC); Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1330 (UPTC).Odontoglossum lindenii Lindl. Epífita. En los Andes (Or.), 1866-3990 m. Boyacá, Viracachá, loc. 6, 6 dic 2012, Gil-l., et al. 1761 (UPTC). Pterichis aff. galeata Lindl. Hierba. Andes (C. y Occ.), 2600-4180 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1824 (UPTC). Pterichis sp. Hierba. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2642 (UPTC).PoaCeae Barnhart, 1828, Cyperales¹, Poales² (7/8).Agrostis perennans (Walter) Tuck. Hierba. En los Andes (Or., C. y Occ.), 1750-4500 m. Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2584 (UPTC).Anthoxanthum odoratum L. Hierba. En los Andes (Or., C. y Occ.), 1800-4100 m. Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2586 (UPTC).Calamagrostis effusa (Kunth) Steud. Hierba. En los Andes (Or., C. y Occ.), 1760-4718 m. Boyacá, Ciénega, loc. 7, ago 24 2012, Gil-l., et al. 2658 (UPTC). ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2011 (UPTC); Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1699 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1769 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1776 (UPTC); loc. 16, 23 ago 2012, Gil-l., et al. 2635 (UPTC). C. cf. recta Trin. ex Steud. Hierba. En los Andes (Or., C. y Occ.), 3172-4250 m. Boyacá, Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1742 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1772 (UPTC).Chusquea tessellata Munro. Hierba. Andes (Or., C. y Occ.), 1900-4350 m. Boyacá, Ciénega, loc. 8, 24 ago 2012, Gil-l., et al. 2662 (UPTC); loc. 7, 24 ago 2012, Gil-l., et al. 2656 (UPTC); loc. 6, 3 abr 2012, Gil-l., et al. 1397 (UPTC). ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2016 (UPTC); loc. 3, 3 may 2012, Gil-l., et al. 1940 (UPTC). Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1734 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1707 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1698 (UPTC); loc. 7, 22 ago 2012, Gil-l., et al. 2596 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1712 (UPTC). Cortaderia nitida (Kunth) Pilg. Hierba. En los Andes (Or., C. y Occ.) 2500-4000 m. Boyacá, Ciénega, loc. 7, 24 ago 2012, Gil-l., et al. 2655 (UPTC); loc. 6, 3 abr 2012, Gil-l., et al. 1402 (UPTC). ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1944 (UPTC). Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1740 (UPTC), loc. 20, 15 abr 2012, Gil-l., et al. 1825 (UPTC). Neurolepis sp. Hierba. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2634 (UPTC).Paspalum sp. Hierba. Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2585 (UPTC).

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 21
sMilaCaCeae Vent., 1799, Liliales¹ y ² (1/1).Smilax sp. Hierba. Boyacá, Ciénega, loc. 5, 25 ago 2012, Gil-l., et al. 2663 (UPTC).
Dicotiledónea
aPiaCeae Lindl., 1836, Apiales¹ y ² (1/1).Eryngium humile Cav. Hierba. En los Andes (Or., C. y Occ.), 2120-4500 m. Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2581 (UPTC).araCeae Juss., 1789, Arales¹, Alismataceae² (1/1).Anthurium oxybelium Shott. Epífita. En los Andes (Or., C. y Occ.), 1350-3860 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1414 (UPTC). araliaCeae Juss., 1789, Apiales¹ y ² (1/1).Oreopanax integrifolius Cuatrec. Arbusto. En los Andes (Or.), 2200-2300 m. Boyacá, Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1316 (UPTC). asteraCeae Bercht. & J. Presl, 1820, Asterales¹ y ² (7/21).Ageratina elegans (Kunth) R.M. King & H. Rob., Arbusto. En los Andes (Or.), 2400-3380 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1702 (UPTC).A. killipii H. Rob., Arbusto. En los Andes (Or.), 2800-3200 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1399 (UPTC). A. theaefolia (Benth.) R.M. King & H. Rob. Arbusto. En los Andes (Or. y C.), 2100-4100 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1697 (UPTC); loc. 10, 21 ago 2012, Gil-l., et al. 2587, 2590 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1720 (UPTC).A. tinifolia (Kunth) R.M. King & H. Rob. Arbusto. En los Andes (Or., C. y Occ.), 1750-3900 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1404 (UPTC). Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2614 (UPTC); loc 9, 3 abr 2012, Gil-l., et al. 1755 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1727 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1723 (UPTC).Diplostephium alveolatum Cuatrec. Arbusto. En los Andes (Or.), 3310-4430 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2608 (UPTC).D. dentatum Cuatrec. Arbusto. En los Andes (Or.), 2880-2880 m. Boyacá, Viracachá, loc. 10, 21 ago 2012, Gil-l., et al. 2588 (UPTC).D. floribundum (Benth.) Wedd. Arbusto. En los Andes (Or., C. y Occ.), 2380-3800 m. Boyacá, Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1749 (UPTC).D. oblongifolium Cuatrec., Arbusto. Andes (Or.), 2770-3100 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1819 (UPTC).D. phylicoides (Kunth) Wedd. Arbusto. Andes (Or.), 2770-3100 m. Boyacá, Ciénega, loc. 4, 11 jul 2012, Gil-l., et al. 2575 (UPTC); loc. 6, 3 abr 2012, Gil-l., et al. 1398 (UPTC). Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2644 (UPTC); loc. 13, 21 ago

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón22
2012, Gil-l., et al. 2580 (UPTC); loc. 15, 3 abr 2012, Gil-l., et al. 1736 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1684 (UPTC); loc. 9, 3 abr 2012, Gil-l., et al. 1751 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1714 (UPTC).Espeletia argentea Bonpl. Hierba. Andes (Or.), 2700-3900 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1822 (UPTC). E. boyacensis Cuatrec. Hierba. Andes (Or.), 2345-3865 m. Boyacá, Viracachá, loc. 18, 12 jul 2012, Gil-l., et al. 2577 (UPTC); loc. 2, 21 abr 2012, Gil-l., et al. 2347 (UPTC). E. cayetana (Cuatrec.) Cuatrec. Hierba. Andes (Or.), 3500-3650 m. Boyacá, Viracachá, loc. 5, 22 ago 2012, Gil-l., et al. 2603 (UPTC).E. cf. corymbosa Bonpl. Hierba. Andes (Or.), 2600-3500 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2601 (UPTC). E. murilloi Cuatrec. Arbusto. Andes (Or.), 2700-3700 m. Boyacá, Ciénega, loc. 1, 25 ago 2012, Gil-l., et al. 2666 (UPTC); loc. 6, 3 abr 2012, Gil-l., et al. 1409 (UPTC). ramiriquí, loc. 2, 3 sep 2012, Gil-l., et al. 2671 (UPTC). Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1732 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1696 (UPTC); loc. 20, 15 abr 2012, Gil-l., et al. 1826 (UPTC); loc 19, 14 abr 2012, Gil-l., et al. 1764 (UPTC); loc. 7, 22 ago 2012, Gil-l., et al. 2602 (UPTC); loc. 3, 21 abr 2012, Gil-l., et al. 2348 (UPTC). ramiriquí, loc. 1, 5 may 2012 (UPTC). Espeletia sp. Hierba. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2599 (UPTC).Munnozia senecionidis Benth. Trepadora. Andes (Or., C. y Occ.), 1000-3820 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1837 (UPTC).Pentacalia guadalupe (Cuatrec.) Cuatrec. Arbusto. Andes (Or.), 2120-4040 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1921 (UPTC). Viracachá, loc. 14, 3 abr 2012, Gil-l., et al. 1748 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1708 (UPTC).P. pulchella (Kunth) Cuatrec. Arbusto. Andes (Or., C. y Occ.), 1900-3850 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1922 (UPTC).P. tolimensis (Sch. Bip. ex Wedd.) Cuatrec. Arbusto. Andes (Or., C. y Occ.), 2330-4180 m. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2647 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1728 (UPTC).P. tolimensis var. colombiana (Cuatrec.) S. Díaz. Arbusto. Andes (Or., C. y Occ.), 1750-4300 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2612 (UPTC).P. trichopus (Benth.) Cuatrec. Arbusto. Andes (Or., C. y Occ.), 2800-3930 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1407 (UPTC). Pentacalia sp. Arbusto. Boyacá, Ciénega, loc. 8, 24 ago 2012, Gil-l., et al. 2661 (UPTC).Scrobicaria ilicifolia (L. f.) B. Nord. Arbusto. Andes (Or.), 2600-3910 m. Boyacá,

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 23
Viracachá, loc. 13, 21 ago 2012, Gil-l., et al. 2579 (UPTC).Senecio subruncinatus Greenm. Hierba. Andes (Or. y C.), 3000-3800 m. Boyacá, Ciénega, loc. 2, 25 ago 2012, Gil-l., et al. 2665 (UPTC); loc. 7, 24 ago 2012, Gil-l., et al. 2659 (UPTC).BeGoniaCeae C. Agardh., 1824, Violales¹, Cucurbitales² (1/1). Begonia ferruginea L. f. Hierba. Andes (Or. y C.), 1700-4000 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1935 (UPTC).BerBeridaCeae Juss., 1789, Ranunculales¹ y ² (1/2).Berberis petriruizii L.A. Camargo. Arbusto. Andes (Or.), 3000-3550 m. Boyacá, Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1752 (UPTC). B. cf. muiscarum L.A. Camargo. Hierba. Andes (Or.), 2900-2900 m. Boyacá, Ciénega, loc. 7, 24 ago 2012, Gil-l., et al. 2657 (UPTC). CaMPanulaCeae Juss., 1789, Campanulales¹, Asterales² (1/1).Siphocampylus retrorsus Vatke & E. Wimm. Hierba. Andes (Or., C. y Occ.), 1000-3200 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1937 (UPTC); loc 3, 3 may 2012, Gil-l., et al. 1924 (UPTC).CHlorantHaCeae R. Br. ex Sims, 1820, Piperales¹, Chlorantales² (1/2).Hedyosmum crenatum Occhioni. Arbusto. Andes (Or., C. y Occ.), 1990-3350 m. Boyacá, Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1372 (UPTC).H. parvifolium Cordem., Arbusto. Andes (Or.), 2600-3400 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1885 (UPTC).CletHraCeae Klotzsch, 1851, Ericales¹ y ² (1/1).Clethra fimbriata Kunth. Arbusto. Andes (Or. y C.), 1990-3900 m. Boyacá, Viracachá, loc. 13, 21 ago 2012, Gil-l., et al. 2578 (UPTC); loc. 4, 23 ago 2012, Gil-l., et al. 2650 (UPTC).ClusiaCeae Juss., 1789 (Theales1, Hypericum hace parte de Clusiaceae; alpighiales2, Clusiaceae e Hypericaceae son familias independientes en el mismo orden) (1/5). Hypericum laricifolium Juss. Hierba. Andes (Or., C. y Occ.), 1700-4500 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1704 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1774 (UPTC).H. mexicanum L. Hierba. Andes (Or., C. y Occ.), 1700-4110 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1926 (UPTC). Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2646 (UPTC); loc. 9, 3 abr 2012, Gil-l., et al. 1750 (UPTC).H. myricariifolium Hieron. Hierba. Andes (Or.), 2482-4120 m. Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2583 (UPTC).H. lycopodioides Triana & Planch. Arbusto. Andes (Or.), 2730-4000 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1400 (UPTC). ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1925 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1703

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón24
(UPTC); loc. 20, 15 abr 2012, Gil-l., et al. 1811 (UPTC).CunoniaCeae R. Br., 1814, Rosales¹, Oxalidales² (1/4).Weinmannia fagaroides Kunth. Árbol. Andes (Or. y C.), 2600-4000 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1410 (UPTC). Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1812 (UPTC). W. reticulata Ruiz & Pav. Árbol. Andes (Or., C. y Occ.), 1800-3100 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1931 (UPTC). W. rollottii var. rollottii Killip. Árbol. Andes (Or. y C.), 2600-3300 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1411 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1690 (UPTC). W. tomentosa L. f. Árbol. Andes (Or., C. y Occ.), 2000-3624 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1939 (UPTC). Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1726 (UPTC).eriCaCeae Juss., 1789, Ericales¹ y ² (5/12).Bejaria resinosa Mutis ex. L. f. Arbusto. Andes (Or. y C.), 1750-3800 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1685 (UPTC); loc. 20, 15 abr 2012, Gil-l., et al. 1818 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1713 (UPTC).Disterigma alaternoides (Kunth) Nied. Arbusto. Andes (Or., C. y Occ.), 1200-3650 m. Boyacá, ramiriquí, loc 3, 3 may 2012, Gil-l., et al. 1938 (UPTC). Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1739 (UPTC); loc. 15, 3 abr 2012, Gil-l., et al. 1738 (UPTC); loc. 10, 21 ago 2012, Gil-l., et al. 2594 (UPTC). D. empetrifolium (Kunth) Drude. Hierba. Andes (Or., C. y Occ.), 2070-4300 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1928 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1686 (UPTC); loc. 14, 3 abr 2012, Gil-l., et al. 1745 (UPTC); loc. 16, 23 ago 2012, Gil-l., et al. 2641 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1768 (UPTC).Gaultheria anastomosans (L.f.) Kunth. Arbusto. Andes (Or., C. y Occ.), 1990-4020 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1943 (UPTC); loc. 3, 3 may 2012, Gil-l., et al. 1932 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1688 (UPTC); loc. 7, 22 ago 2012, Gil-l., et al. 2607 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1721 (UPTC).G. erecta Vent. Hierba. Andes (Or., C. y Occ.), 1500-3731 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1406 (UPTC). ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1941 (UPTC). Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1767 (UPTC).G. rigida Kunth. Hierba. Andes (Or., C. y Occ.), 1800-3600 m. Boyacá, Viracachá, loc. 16, 23 ago 2012, Gil-l., et al. 2639 (UPTC).Macleania rupestris (Kunth) A.C. Sm. Arbusto. Andes (Or., C. y Occ.), 1990-3990 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1929 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1700 (UPTC); loc. 20, 15 abr 2012, Gil-l., et al.

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 25
1816 (UPTC); loc. 10, 21 ago 2012, Gil-l., et al. 2589 (UPTC).Pernettya prostrata (Cav.) DC. sinónimo: Gaultheria mysinoides Kunth. Arbusto. Andes (Or., C. y Occ.), 1720-4525 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1401, 1405 (UPTC). Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1766 (UPTC); loc. 19, 14 abr 2012, Gil-l., et al. 1765 (UPTC); loc 20, 15 abr 2012, Gil-l., et al. 1810 (UPTC); loc. 7, 22 ago 2012, Gil-l., et al. 2609 (UPTC).faBaCeae Lindl., 1836, Fabales¹ y ² (1/1).Lupinus dotatus C.P. Sm. Hierba. Andes (Or.), 3000-3000 m. Boyacá, Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2617 (UPTC).GentianaCeae Juss., 1789, Gentianales¹ y ² (3/4).Gentianella corymbosa (Kunth) Weaver & Ruedenberg. Hierba. Andes (Or., C. y Occ.), 2600-4100 m. Boyacá, Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2615 (UPTC).Halenia asclepiadea (Kunth) G. Don. Hierba. Andes (Or., C. y Occ.) y SNSM, 2600-4000 m. Boyacá, Viracachá, loc. 14, 3 abr 2012, Gil-l., et al. 1746 (UPTC). H. campanulata Cuatrec. Hierba. Andes (Or., C. y Occ.), 3300-4500 m. Boyacá, Viracachá, loc. 17, 23 ago 2012, Gil-l., et al. 2649 (UPTC). Macrocarpaea sp. Arbusto. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1942 (UPTC).GeraniaCeae Juss., 1789, Geraniales¹ y ² (1/1).Geranium cf. alonsoi Aedo. Hierba. Andes (Or.). Boyacá, Viracachá, loc. 11, 21 ago 2012, Gil-l., et al. 2582 (UPTC).GrossulariaCeae Juss., 1789, Rosales¹, Saxifragales ² (2/2).Escallonia myrtilloides L. f. Arbusto. Andes (Or., C. y Occ.) y SNSM, 2700-3930 m. Boyacá, Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2651 (UPTC). Ribes sp. Arbusto reclinado. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2605 (UPTC).MelastoMataCeae Juss., 1789, Myrtales¹ y ² (5/10).Bucquetia glutinosa (L. f.) D.C. Arbusto. Andes (Or.), 2100-4104 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1396 (UPTC). Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1722 (UPTC). Castratella piloselloides Naudin. Hierba. Andes (Or., C. y Occ.), 2120-4000 m. Boyacá, Ciénega, loc. 3, 25 ago 2012, Gil-l., et al. 2664 (UPTC). Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1770 (UPTC).Miconia cataractae Triana. Arbusto. Andes (Or. y C.), 2420-3900 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 1992 (UPTC). Viracachá, loc. 6, 6 dic 2012, Gil-l., et al. 1760 (UPTC).M. ligustrina (Sm.) Triana. Arbusto. Andes (C. y Or.), 2050-3700 m. Boyacá,

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón26
ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2008 (UPTC); loc. 3, 3 may 2012, Gil-l., et al. 1936 (UPTC). Viracachá, loc. 9, 3 abr 2012, Gil-l., et al. 1753 (UPTC); loc. 10, 21 ago 2012, Gil-l., et al. 2592 (UPTC).M. salicifolia (Bonpl. ex Naudin) Naudin. Arbusto. Andes (Or.), 2500-4200 m. Boyacá, Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2618 (UPTC).M. aff. cleefii L. Uribe. Arbusto. Andes (Or.), 3320-3900 m. Boyacá, Ciénega, loc. 7, 24 ago 2012, Gil-l., et al. 2654 (UPTC).M. chionophila Naudin. Hierba. Andes (Or., C. y Occ.), 2880-4250 m. Boyacá, Viracachá, loc. 8, 22 ago 2012, Gil-l., et al. 2604 (UPTC). Monochaetum myrtoideum Naudin. Arbusto. Andes (Or.), 1650-4375 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1814 (UPTC).Tibouchina grossa (L. f.) Cogn. Árbol. Andes (Or., C. y Occ.), 1850-4500 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1412 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1693 (UPTC). MYrsinaCeae R. Br., 1810, Primulales¹, Ericales² (1/1).Geissanthus andinus Mez. Arbusto. Andes (Or., C. y Occ.), 1825-3700 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 1998 (UPTC); loc. 3, 3 may 2012, Gil-l., et al. 1933 (UPTC). Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1689 (UPTC); loc. 1, 12 abr 2012, Gil-l., et al. 1725 (UPTC).MYrtaCeae Juss., 1789, Myrtales¹ y ² (1/1).Myrcianthes rhopaloides (Kunth) McVaugh. Arbusto. Andes (Or. y C.), 1600-3000 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2006 (UPTC). Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2653 (UPTC).onaGraCeae Juss., 1789, Myrtales¹ y ² (Un género-una especie).Fuchsia canescens Benth. Hierba. Andes (Or., C. y Occ.), 1025-3830 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2610 (UPTC); loc. 6, 6 dic 2012, Gil-l., et al. 1759 (UPTC).PolYGalaCeae Hoffmanns & Link, 1809, Polygalales¹, Fabales² (1/1).Monnina salicifolia Ruiz & Pav. Arbusto. Andes (Or., C. y Occ.) y SNSM, 1600-3700 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2012 (UPTC).PolYGonaCeae Juss., 1789, Polygonales¹, Caryophyllales² (1/1).Muehlenbeckia tamnifolia (Kunth) Meisn. Bejuco. Andes (Or., C. y Occ.) y SNSM, 1200-4500 m. Boyacá, ramiriquí, loc. 1, 5 may 2012, Gil-l., et al. 2004 (UPTC). Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1724 (UPTC).rosaCeae Juss., 1789, Rosales¹ y ² (3/4).Hesperomeles goudotiana (Dence.) Killip. Arbusto. Andes (Or. y Occ.), 2400-3950 m. Boyacá, Viracachá, loc. 10, 21 ago 2012, Gil-l., et al. 2591 (UPTC).H. obtusifolia (Pers.) Lindl. Arbusto. Andes (Or., C. y Occ.), 2000-4020 m. Boyacá,

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 27
Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1729 (UPTC).Lachemilla aphanoides (Mutis ex L. f.) Rothm. Hierba. Andes (Or. y C.), 2300-4050 m. Boyacá, Viracachá, loc. 19, 14 abr 2012, Gil-l., et al. 1775 (UPTC).Rubus compactus Benth. Hierba. Andes (Or., C. y Occ.), 2000-3700 m. Boyacá, Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1601 (UPTC).ruBiaCeae Juss., 1789, Rubiales¹, Gentianales² (4/5).Arcythophyllum nitidum (Kunth) Schltdl. Arbusto. Andes (Or., C. y Occ.), 1500-4500 m. Boyacá, Ciénega, loc. 6, 3 abr 2012, Gil-l., et al. 1413 (UPTC). ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1930 (UPTC). Viracachá, loc. 14, 3 abr 2012, Gil-l., et al. 1744 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1687 (UPTC); loc. 20, 15 abr 2012, Gil-l., et al. 1815 (UPTC). Galium corymbosum Ruiz & Pav. Hierba. Andes (Or. y C.), 2600-4000 m. Boyacá, Viracachá, loc. 10, 21 ago 2012, Gil-l., et al. 2595 (UPTC).G. hypocarpium (L.) Endl. ex Griseb. Hierba. Andes (Or., C. y Occ.) y SNSM, 1300-3900 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1927 (UPTC). Viracachá, loc. 20, 15 abr 2012, Gil-l., et al. 1813 (UPTC).Nertera granadensis (Mutis ex L. f.) Druce. Hierba. Andes (Or., C. y Occ.) y SNSM, 1300-4300 m. Boyacá, Viracachá, loc. 12, 12 abr 2012, Gil-l., et al. 1705 (UPTC); loc. 20, 15 abr 2012, Gil-l., et al. 1817 (UPTC).Palicourea aschersonianoides (Wernham) Steyerm. Hierba. Andes (Or. y C.), 1750-3750 m. Boyacá, ramiriquí, loc. 3, 3 may 2012, Gil-l., et al. 1877 (UPTC).sCroPHulariaCeae Juss., 1789, Scrophylariales¹, Lamiales² (3/3).Alonsoa meridionalis (L.f.) Kuntze. Hierba. Andes (Or., C. y Occ.) y SNSM, 1370-4000 m. Boyacá, Viracachá, loc. 4, 23 ago 2012, Gil-l., et al. 2619 (UPTC).Bartsia santolinifolia (Kunth) Benth. Hierba. Andes (Or. y C.), 2560-4335 m. Boyacá, Viracachá, loc. 15, 3 abr 2012, Gil-l., et al. 1737 (UPTC); loc. 12, 12 abr 2012, Gil-l., et al. 1710 (UPTC); loc. 16, 23 ago 2012, Gil-l., et al. 2638 (UPTC). Castilleja fissifolia L. f. Hierba. Andes (Or., C. y Occ.) y SNSM, 1300-5000 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2606 (UPTC).sYMPloCaCeae Desf., 1820, Ebenales¹, Ericales² (1/1).Symplocos aff. cundinamarcensis B. Ståhl. Arbusto. Andes (Or.), 2800-3310 m. Boyacá, Viracachá, loc. 7, 22 ago 2012, Gil-l., et al. 2611 (UPTC).WinteraCeae R. Br. ex Lindl., 1830, Magnoliales¹, Canellales² (1/1).Drimys granadensis L.f. Árbol. Andes (Or.), 2900-2900 m. Boyacá, Viracachá, loc. 1, 12 abr 2012, Gil-l., et al. 1378 (UPTC). La diversidad florística se ve reflejada a través de los inventarios, que son un insumo básico para el estudio de la diversidad funcional, así como la taxonómica, filogenética, entre otros; sin dejar de lado los trabajos en biogeografía de los ecosistemas estratégicos,

bol.c
ient
.mus
.hist
.nat
. 21
(2) j
ulio
- di
ciem
bre
2017
. 15-
29Merly Yenedith Carrillo Fajardo, María eugenia Morales-Puentes, Pablo Andrés Gil-leguizamón28
como es el páramo; y la comprensión de las dinámicas de los servicios ecosistémicos que ofertan estas importantes áreas de conservación (PÉREZ-MORENO et al., 2016).
aGradeCiMientos
A la Universidad Pedagógica y Tecnológica de Colombia por la beca DIN 04 de 2012, a través de la Dirección de Investigaciones (DIN); así mismo, al Convenio de Colaboración DHS No. 5211071 entre Ecopetrol S.A. & UPTC (201-2011). A los Herbarios UPTC, COL, FMB y HECASA y a los botánicos: S. Díaz-Piedrahíta+, B.V. Rodríguez-Cabeza, J. Aguilar, M. Quijano, J.C. Farfán, O. Rivera, R. Sánchez y F. Cardona, por la colaboración en las determinaciones y corroboraciones del material. Al Grupo Sistemática Biológica por el apoyo en el trabajo de campo, laboratorio y trámites administrativos, y a Z. Carrillo-Fajardo y C.C. Vidal-Pastrana por la ayuda en la sistematización de la información.
referenCias
APG IV, 2016.- An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the linnean Society.
ARELLANO-P., H. & RANGEL-CH., J.O., 2010.- Fragmentación y estado de conservación en páramos de Colombia. 479-542 (en) RANGEL-CH., J.O. (ed.) Colombia diversidad biótica X: Cambios global (natural) y climático (antrópico) en el páramo colombiano. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
BERNAL, R., GRADSTEIN, R. & CELIS, M. (eds.), 2016.- Catálogo de las plantas y líquenes de Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
CARRILLO-FAJARDO, M.Y., 2013.- Fitogeografía de la flora paramuna del Macizo de Bijagual, Boyacá-Colombia: Tesis, Universidad Pedagógica y Tecnológica de Colombia, Facultad de Ciencias, Tunja.
CIRI, F., 2003.- Investigación, planificación y desarrollo en zonas ecoturísticas. Caso de estudio: Plan de desarrollo ecoturístico para la zona de los macizos de Mamapacha y Bijagual (Boyacá). Turismo y Sociedad, 2: 83-90.
COL - Herbario Nacional Colombiano, Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, 2004 y continuamente actualizado.- Colecciones en Línea. Publicado en Internet. http://www.biovirtual.unal.edu.co
CORPOCHIVOR, 2006.- el plan de ordenamiento y manejo de la cuenca del río Aragoa - síntesis ambiental. Boyacá, Colombia.CRONQUIST, A., 1988.- The evolution and classification of flowering plants. The New York Botanical Garden, Bronx, New York, USA.CUATRECASAS, J., 1934.- Observaciones geobotánicas en Colombia. Trabajos del Museo Nacional de Ciencias Naturales Serie
Botánica. Madrid, 27: 144.DÍAZ-GRANADOS, M.A., NAVARRETE-GONZÁLEZ, J.D. & SUÁREZ-LÓPEZ, T., 2005.- Páramos: Hidrosistemas sensibles.
Revista de Ingeniería, 22: 64-75.FONT QUER, P., 2000.- Diccionario de Botánica. Península, Barcelona.HERNÁNDEZ, J., HURTADO, A., ORTIZ, R. & WALSHBURGER, T., 1992.- Unidades biogeográficas de Colombia: 105-152
(en) G. HALFFTER (ed.) la diversidad biológica de Iberoamérica. Instituto de Ecología, México. IPNI - The International Plants Name Index, 2011.- http://www.ipni.org/KEY - Royal Botanical Gardens, 2013.- http://www.kew.orgLONDOÑO, C., CLEEF, A. & MADRIÑAN, S., 2014.- Angiosperm flora and biogeography of the paramo region of Colombia,
Northern Andes. Flora, 209: 81-87.MADS (Ministerio de Ambiente y Desarrollo Sostenible), 2012.- Política nacional para la gestión integral de la biodiversidad y servicios
ecosistémicos (PNGIBSe). República de Colombia.MIRANDA-ESQUIVEL, D., RANGEL-CH, O. & ROA-FUENTES, L., 2002.- Endemismo en páramos colombianos y el análisis
de parsimonia de endemismos (PAE): 1: 253-266 (en) JARAMILLO, C., CASTAÑO-URIBE, C., ARJONA-HINCAPIÉ, F., RODRÍGUEZ, J.V. & DURÁN, C.L. (eds.) Memorias Congreso Mundial de Páramos. Paipa, Boyacá.
MORALES-PUENTES, M.E., GIL-LEGUIZAMÓN, P.A., SIMBAQUEBA-GUTIÉRREZ, A.L. & CARRILLO-FAJARDO, M.Y., 2012.- Informe final convenio de colaboración DHS N°5211071: Biodiversidad y objetos de conservación de los ecosistemas estratégicos de Sucuncuca y Bijagual (Provincia de lengupá) Boyacá. Universidad Pedagógica y Tecnológica de Colombia, Tunja.
PÉREZ-MORENO, H., MARÍN, C. & LEÓN, O., 2016.- Cambios en las coberturas paramunas (en) GÓMEZ, M.F., MORENO, L.A., ANDRADE, G.I. & RUEDA, C. (ed.). Biodiversidad 2015. estado y tendencias de la biodiversidad continental de Colombia. Instituto Alexander von Humboldt, Bogotá.
RANGEL-CH., J.O., 2007.- La región paramuna en Colombia y en la Serranía de Perijá: 1-18. (en) RANGEL-CH, J.O. (ed.) Colombia, Diversidad Biótica V. la Alta Montaña de la Serranía de Perijá. Instituto de Ciencias Naturales, Universidad Nacional

Catálogo de la flora (angiospermas) del páramo de Bijagual, Boyacá, Colombia 29
de Colombia, Bogotá.RIVERA-OSPINA, D. & RODRÍGUEZ, C., 2011.- Guía divulgativa de criterios para la delimitación de páramos de Colombia.
Ministerio de Ambiente, Vivienda y Desarrollo Territorial e Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
SARMIENTO, C., CADENA, C., SARMIENTO, M. ZAPATA, J. & LEÓN, O., 2013.- Aportes a la conservación estratégica de los páramos de Colombia: Actualización de la cartografía de los complejos de páramo a escala 1:100.000. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
SARMIENTO, C., & LEÓN, O. (eds.), 2015.- Transición bosque-páramo. Bases conceptuales y métodos para su identificación en los Andes colombianos. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
SIB - Sistema de Información sobre Biodiversidad de Colombia, 2014.- http://maps.sibcolombia.netSIMA (Sistemas Integrados de Manejo Ambiental), CORPOCHIVOR & CORPOBOYACÁ, 1996.- Plan de manejo ambiental
páramo de Bijagual. Santa Fe de Bogotá.The Plant List, A working list of all plant species, 2013.- http://www.theplantlist.org/ NY - The New York Botanical Garden. Virtual Herbarium, 2013.- http://www.nybg.org/UNIVERSIDAD MILITAR NUEVA GRANADA & CORPOCHIVOR, 2008.- Páramo de Bijagual (en) Contrato Interadministrativo
Nº043/2008 para la “complementación y actualización del estudio sobre el estado del área de páramos en los sectores de Bijagual, Mamapacha, Cristales y Castillejo, en la jurisdicción de Corpochivor y formular un Plan de Manejo Ambiental para dichas áreas”. Boyacá, Colombia.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
PriMer reGistro de Hydnopolyporus fimbriatus (Cooke) d.a. reid (PolYPorales: MeriPilaCeae) Para el
dePartaMento de CundinaMarCa, ColoMBia*
Andrea Castiblanco-Z.1, César Augusto Pinzón-o.2, Jonás Pinzón-o.3
resumen
Hydnopolyporus fimbriatus es un hongo lignícola reportado en Colombia por primera y última vez en 1930 para el departamento del Valle del Cauca. Durante un rastreo realizado en el campus de la Universidad Nacional de Colombia, sede Bogotá, se registró la presencia de la especie, con lo cual se expande su distribución para el departamento de Cundinamarca. Se recolectaron y examinaron 3 basidiomas siguiendo los protocolos establecidos en la literatura especializada. Se aporta información micológica sobre características macroscópicas y microscópicas, distribución actual, época de colecta y sustrato de crecimiento de los basidiomas.
Palabras clave: Colombia, Hydnopolyporus, nueva distribución, Polyporales, Universidad Nacional de Colombia.
first rePort of Hydnopolyporus fimbriatus (Cooke) d.a. reid (PolYPorales: MeriPilaCeae) for tHe dePartMent of
CundinaMarCa, ColoMBia
abstract
Hydnopolyporus fimbriatus is a lignicolous fungus reported in Colombia for the first and last time in 1930 for the department of Valle del Cauca. During the collection at the campus of Universidad Nacional de Colombia, in Bogotá, the presence of the species was recorded with which its distribution for the department of Cundinamarca expands. Three basidiome were collected and examined following the protocols established by the specialized literature. Mycological information is provided on macroscopic and microscopic characteristics, current distribution, time of collection and substrate for growth of the basidiome.
Key words: Colombia, Hydnopolyporus, new distribution, Polyporales, Universidad Nacional de Colombia. * FR: 18-IX-2016. FA: .3-VI-20171 Licenciada en Biología. Universidad Pedagógica Nacional. Departamento de Biología. Línea de Investigación Ecología de Sistemas Acuáticos de la Región Andina. Grupo de Investigación CASCADA. Bogotá, Colombia. Autor para correspondencia. E-mail: [email protected] ORCID: 0000-0002-8232-90742 Licenciado en Biología. Profesor Universidad Pedagógica Nacional. Herbario (UPN). Departamento de Biología. Línea de Investigación Enseñanza y Aprendizaje de la Botánica. Bogotá, Colombia. E-mail: [email protected]: 0000-0002-6704-51373 Student of College of the Atlantic. Program Human Ecology. Bachelor of Science. COA. Maine. USA. E-mail: [email protected]
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 30-37. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: CASTIBLANCO-Z., A., PINZÓN-O., C.A. & PINZÓN-O., J., 2017.- Primer registro de Hydnopolyporus fimbria-tus (Cooke) D.A. Reid (Polyporales: Meripilaceae) para el departamento de Cundinamarca, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 30-37. DOI: 10.17151/bccm.2017.21.2.2

Primer registro de Hydnopolyporus fimbriatus (cooke) D.A. Reid (polyporales: meripilaceae)... 31
introduCCiÓn
La familia Meripilaceae Jülich (Polyporales) comprende especies de hongos lignícolas con amplia distribución en diferentes biomas terrestres (RAYNER, 1995). A estos organismos se les ha reconocido una ecología relevante dentro de los ecosistemas forestales, ya que dinamizan la liberación y reciclaje de nutrientes en el fenómeno de descomposición de detritos (RUIZ & VARELA, 2006) al producir degradación de la celulosa y la lignina (RAYNER, 1995) mediante pudrición blanca (GILBERTSON & RYVARDEN, 1986, 1987; RYVARDEN, 1991).
La familia está representada por nueve géneros: Antrodia P. Karst, Grifola Gray, Hen-ningsia Möller, Meripilus Karst, Physisporinus Karst, Diachanthodes Singer, Abortiporus Murrill, Rigidoporus Murrill, e Hydnopolyporus D.A. Reid (KIRK et al., 2001). Los cuatro últimos se encuentran reportados en Colombia, distribuidos en 10 especies (VASCO-PALACIOS & FRANCO-MOLANO, 2013). Hydnopolyporus representado por Hydnopolyporus fimbriatus (Fr.) fue reportado por CHARDON & TORO (1930) e Hydnopolyporus palmatus (Hook.) O. Fidalgo por CHARTON & TORO (1930) y DENNIS (1970). Estas especies corresponden a dos de las tres especies conocidas en el mundo (KIRK et al., 2001). H. fimbriatus posee basidiomas anuales, sésiles, solitarios a cespitosos, con píleos flabeliformes, irregulares a espatulados que forman rosetas, posee superficie himenial variable y un sistema hifal monomítico (GILBERTSON & RYVARDEN, 1986).
Esta especie fue registrada por primera vez en el departamento del Valle del Cauca en los años 30 (CHARDON & TORO, 1930) y de acuerdo a VASCO-PALACIOS & FRANCO-MOLANO (2013) hasta la fecha no se conocen nuevos reportes en el país. Por lo tanto, el objetivo del presente artículo es ampliar la distribución de H. fimbriatus para el departamento de Cundinamarca a partir de basidiomas recolectados en el campus de la Universidad Nacional de Colombia, sede Bogotá. MÉtodos
Área de estudio
La zona de muestreo se ubica en la Ciudad Universitaria, Universidad Nacional de Colombia, sede Bogotá, a 2600 msnm, coordenadas 4°38’04,05” N y 74°05’09,06” O, departamento de Cundinamarca, Colombia (Figura 1). El campus se constituye de 121,35 hectáreas, con predominio de un paisaje urbano alternado con potreros (INFANTE-BETANCOUR et al., 2008).

Andrea Castiblanco-Z., César Augusto Pinzón-o., Jonás Pinzón-o.32bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 3
0-37
Distribución de Hydnopolyporus fimbriatus para Colombia, registrada por CHAR-TON & TORO (1930) y VASCO-PALACIOS & FRANCO-MOLANO (2013) ( ). Nuevo registro para el departamento de Cundinamarca ( ). B. Universidad Nacional de Colombia. Área de colecta ( ).
figura 1.a.
La zona presenta periodos secos y de lluvias con precipitación promedio anual de 806,3 mm3, con la mayoría de lluvias concentradas en los meses de febrero, marzo, octubre y noviembre. La temperatura media anual corresponde a 14,1°C, con una máxima de 20,0°C y una mínima de 9,0°C (IDEAM, 2007). El área muestra variedad de espe-cies arbóreas y arbustivas, tanto nativas como foráneas, representadas por Ceroxylum quindiuense H. Wendl ex H. Karst, Croton bogotanus Cuatrec., Quercus humboldtii Bonpl., Tibouchina lepidota Bonpl., Inga ornata Kunth, Schinus molle L., Tecoma stans (L.) Juss. ex Kunth, Sambucus nigra L., Cupressus lusitánica Mill, Cotoneaster pannosus Franch. (INFANTE-BETANCOUR et al., 2008).
Muestreo
Los basidiomas se recolectaron aplicando un muestreo oportunista en zonas verdes aledañas al Parque Humboldt, en las facultades de Odontología, Ciencias Humanas, Medicina, Medicina Veterinaria y de Zootecnia. Las fructificaciones se fotografiaron y describieron macroscópicamente de acuerdo a ZALAMEA (2000) y WU et al.

Primer registro de Hydnopolyporus fimbriatus (cooke) D.A. Reid (polyporales: meripilaceae)... 33
(2004). Para la evaluación microscópica se obtuvieron cortes a mano alzada de los basidiomas con ayuda de un estereoscopio AmScope SE305R-PZ y se observaron con un microscopio eléctrico Olympus CX3 previa aplicación de KOH al 5% y Rojo Congo a las muestras. Las características macro y microscópicas fueron analizadas para identificación siguiendo a FIDALGO (1963), REID (1976), GILBERTSON & RYVARDEN (1986), TEIXEIRA (1994, 1995), LEAL & GUGLIOTTA (2008) y LÓPEZ & GARCÍA (2011). Todos los ejemplares fueron depositados en la colección del Herbario de la Universidad Pedagógica Nacional, sede Bogotá. Para la citación del nombre científico se siguió el sistema de clasificación Index Fungorum.
resultados
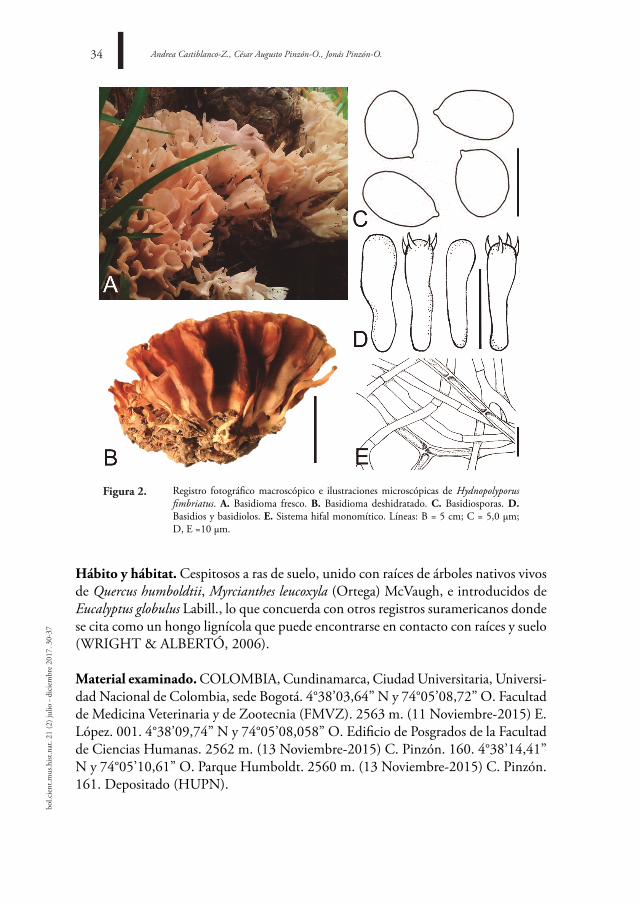
Hydnopolyporus fimbriatus (Cooke) D.A. Reid, Persoonia 2(2): 151 (1962) Figura. 2
= Microporus fimbriatus (Cooke) Kuntze, Revis. gen. pl. (Leipzig) 3(2): 496 (1898).= Coltricia perennis var. fimbriata (Cooke) Bondartsev, Trut. Grib evrop. Chasti SSSR Kavkaza [Bracket Fungi europ. U.S.S.R. Caucasus] (Moscow-Leningrad): 417 (1953).= Abortiporus fimbriatus (Cooke) O. Fidalgo & M. Fidalgo, Arq. Mus. Nac. Rio de Janeiro 43: 158 (1957).= Hydnopolyporus fimbriatus f. fimbriatus (Cooke) D.A. Reid, Persoonia 2(2): 151 (1962).
Basidioma: 5,6-7,8 x 3,5-4,5 cm, cespitoso e imbricado, sésil a estipitado, forman-do rosetas constituidas por varios píleos flebaliformes a espatulados que confluyen y adhieren por la base o estípite. Píleo: espatulado o flabelado, dividido en lóbulos múltiples, algunos forman anostomosis con píleos vecinos, superficie del píleo glabra a pubescente, concéntricamente zonada y longitudinalmente estriado, entero o fimbria-do, consistencia correosa a cartilaginosa, blanquecino a blanco amarillento, ocráceo; color café a café claro en la zona marginal en ejemplares secos. Himenóforo: liso, en algunas zonas con estructuras semejantes a papilas, concoloro con el píleo. Contexto: hasta 0,2 cm, delgado, blanco, ligeramente elástico, fibroso. olor: fúngico. sabor: no registrado. Basidios: 16-27 x 3,5-5,1 µm, cilíndricos a ligeramente clavados, efibulados en base, 4-esterigmatados, pared delgada, hialinos en KOH. Basidiolos: 17,2-25,7 x 4,1-4,6 µm, espatulados, pared delgadas, hialinos en KOH. Basidiosporas: 3,8-5,2 x 2,8-3,5 µm, elipsoides a ovoides, pared delgada, lisas, inamiloides, hialinas en KOH. sistema hifal: monomítico, 3,1-9,8 µm de diámetro, hifas generativas anchas o delgadas, ramificadas, septadas, efibuladas, hialinas, paredes delgadas, hialinas en KOH. reacciones macroquímicas: negativo a KOH al 5% en todas las estructuras.
Andrea Castiblanco-Z., César Augusto Pinzón-o., Jonás Pinzón-o.34bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 3
0-37
Registro fotográfico macroscópico e ilustraciones microscópicas de Hydnopolyporus fimbriatus. a. Basidioma fresco. B. Basidioma deshidratado. C. Basidiosporas. d. Basidios y basidiolos. e. Sistema hifal monomítico. Líneas: B = 5 cm; C = 5,0 µm; D, E =10 µm.
figura 2.
Hábito y hábitat. Cespitosos a ras de suelo, unido con raíces de árboles nativos vivos de Quercus humboldtii, Myrcianthes leucoxyla (Ortega) McVaugh, e introducidos de eucalyptus globulus Labill., lo que concuerda con otros registros suramericanos donde se cita como un hongo lignícola que puede encontrarse en contacto con raíces y suelo (WRIGHT & ALBERTÓ, 2006).
Material examinado. COLOMBIA, Cundinamarca, Ciudad Universitaria, Universi-dad Nacional de Colombia, sede Bogotá. 4°38’03,64” N y 74°05’08,72” O. Facultad de Medicina Veterinaria y de Zootecnia (FMVZ). 2563 m. (11 Noviembre-2015) E. López. 001. 4°38’09,74” N y 74°05’08,058” O. Edificio de Posgrados de la Facultad de Ciencias Humanas. 2562 m. (13 Noviembre-2015) C. Pinzón. 160. 4°38’14,41” N y 74°05’10,61” O. Parque Humboldt. 2560 m. (13 Noviembre-2015) C. Pinzón. 161. Depositado (HUPN).

Primer registro de Hydnopolyporus fimbriatus (cooke) D.A. Reid (polyporales: meripilaceae)... 35
distribución. Hydnopolyporus fimbriatus presenta una amplia distribución. Se difunde en zonas tropicales y subtropicales del Nuevo Mundo (WRIGHT & ALBERTÓ, 2006), siendo común para áreas tropicales del continente americano (REID, 1962; FIDALGO, 1963). La especie se ha registrado en Brasil (FIDALGO & FIDALGO, 1957; RICK, 1960; LEAL & GUGLIOTTA, 2008; GIBERTONI & DRECHSLER-SANTOS, 2010) y México (RAYMUN DO & VALENZUELA, 2003; LÓPEZ & GARCÍA, 2011; MEDINA-JARITZ et al., 2012). En Colombia únicamente se había reportado para el Valle del Cauca (CHARDON & TORO, 1930). En esta ocasión, se amplía su distribución para el departamento de Cundinamarca, 87 años después de haber sido encontrada por primera vez en el país.
observaciones. Las descripciones de los basidiomas colombianos concuerdan muy bien con las aportadas por GILBERTSON & RYVARDEN (1986), WRIGHT & ALBERTÓ (2006), LEAL & GUGLIOTTA (2008) y LÓPEZ & GARCÍA (2011). Macroscópicamente H. fimbriatus se caracteriza por presentar basidiomas cespitosos con píleos abanicados unidos que forman rosetas, con superficie himenial lisa. A nivel microscópico muestra un sistema hifal monomítico y basidiosporas globosas a subglobosas, características que delimitan la especie (GILBERTSON & RYVAR-DEN, 1986). Macroscópicamente podría confundirse con Hydnopolyporus palmatus, sin embargo, esta última presenta un sistema hifal trimítico (REID, 1976; LÓPEZ & GARCÍA, 2011) que lo diferencia de H. fimbriatus con sistema hifal monomítico.
disCusiÓn
Se ha reportado la proliferación de hongos en ambientes perturbados o transformados por acción humana (STAMETS, 1996), como lo son las áreas urbanas. Sin embargo, en Colombia, la diversidad de hongos en estas zonas no ha sido bien documentada. Tan solo 38 especies son citadas en la literatura, 4 de ellas colectadas en el campus de la Universidad Nacional de Colombia, sede Bogotá, por PULIDO (1983), 6 en zonas residenciales de la ciudad de Medellín y del Jardín Botánico “Joaquín Antonio Uribe” (ANÓNIMO, 2000) y 28 reportadas por LÓPEZ-QUINTERO et al. (2011) en el campus de la Universidad de Antioquia, Medellín.
El campus de la Universidad Nacional de Colombia, sede Bogotá, muestra predominio de lo urbano, no obstante, aún conserva zonas con una composición de especies herbáceas, arbóreas y arbustivas que generan un gran potencial para el desarrollo de una amplia diversidad de organismos, como es el caso de los hongos lignícolas. Por lo tanto, se debe asegurar la existencia de estas zonas como áreas que representan la biodiversidad de la ciudad.
Al realizar nuevos muestreos, tanto en las zonas donde se observaron y recolectaron algunos basidiomas de H. fimbriatus como en áreas aledañas, no se encontraron nuevos

Andrea Castiblanco-Z., César Augusto Pinzón-o., Jonás Pinzón-o.36bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 3
0-37
hallazgos de la especie. Esto pudo deberse a la constante actividad de poda de pasto y tala de árboles que se ha generado en los últimos años en el campus. Teniendo en cuenta que esta especie se ubica sobre las raíces de los árboles y muy próximo al suelo, el corte de vegetación y de madera elimina al basidioma de su sustrato, lo que impide la permanencia y actividad ecológica del hongo. De tal forma, es necesario evaluar el impacto que pueden tener la poda y tala en el reconocimiento de la biodiversidad de la Ciudad Universitaria, más aun cuando se trata de esta especie fúngica de desarrollo anual con una actividad ecológica relevante en el proceso de descomposición.
La biodiversidad de la Ciudad Universitaria es aún desconocida. En este caso, se reporta a H. fimbriatus, una especie registrada en Colombia por primera y única vez hace 87 años. Con este registro se amplía la distribución de la especie para el departamento de Cundinamarca y se aumenta a 39 el número de hongos que proliferan en ambientes de alta actividad antrópica. Es necesario continuar estudiando la micota de los paisajes urbanos ya que estas áreas podrían presentar un gran número de especies, como se espera sea el caso de la Ciudad Universitaria. Así, en la medida en que se generen más investigaciones micológicas, mayor cantidad de registros nuevos serán generados y nuevas distribuciones serán conocidas. aGradeCiMientos
A Edwin Alejandro Casas López, estudiante de Medicina Veterinaria de la Universidad Nacional de Colombia, por colaborar en la colecta de los especímenes. Al Herbario de la Universidad Pedagógica Nacional (HUPN), sede Bogotá, por el préstamo de material bibliográfico. referenCias
ANÓNIMO, 2000.- Hongos y musgos del Valle de Aburrá. Medellín (Colombia). Metropolitana del Valle de Aburrá, Subdireccional Ambiental, Medellín.
CHARDON, C.C. & TORO, R.A., 1930.- Mycological exploration of Colombia. Journal of the Department of Agriculture of Porto Rico, 14: 195-369.
DENNIS, R.W.G., 1970.- Fungus flora of Venezuela and adjacent countries. Kew Bulletin, Additional Series III, 3: 1-531.FIDALGO, O., 1963.- Studies on the type species of Hydnopolyporus. Mycologia, 55: 713-727.FIDALGO, O. & FIDALGO, M.E.P.K., 1957.- Revisão de Fungi São Paulensis. Arquivos do Museu Nacional Rio de Janeiro, 43: 157-188.GIBERTONI, T.B. & DRECHSLER-SANTOS, E.R., 2010.- Lignocellulolytic Agaricomycetes from the Brazilian Cerrado biome.
Mycotaxon, 111: 87-90.GILBERTSON, R.L. & RYVARDEN, L., 1986.- North American Polypores. Vol 1. Fungiflora, Oslo.GILBERTSON, R.L. & RYVARDEN, L., 1987.- North American Polypores. Vol 2. Fungiflora, Oslo. IDEAM (INSTITUTO DE HIDROLOGÍA, METEOROLOGÍA Y ESTUDIOS AMBIENTALES), 2007.- estudios de la
caracterización climática de Bogotá y cuenca alta del Río Tunjuelo. IDEAM, Bogotá.INFANTE-BETANCOUR, J., JARA-MUÑOZ, A. & RIVERA-DÍAZ, O., 2008.- Árboles y arbustos más frecuentes de la Universidad
Nacional de Colombia, sede Bogotá. Universidad Nacional de Colombia, Bogotá. KIRK, P.M., CANNON, P.F., DAVID, J.C. & STALPERS, J.A., 2001.- Ainsworth & Bisby’s Dictionary of Fungi. 9th ed. CAB
International, Wallingford.LEAL, G.R. & GUGLIOTTA, A.M., 2008.- Criptógamos do Parque Estadual das Fontes do Ipiranga, São Paulo, SP. Fungos, 9:
Meripilaceae. Hoehnea, 35 (1): 99-110. LÓPEZ, A.R. & GARCÍA, J.A., 2011.- Hydnopolyporus fimbriatus. Funga veracruzana, 125: 1-4. Instituto de Investigaciones Fores-
tales, Universidad Veracruzana.

Primer registro de Hydnopolyporus fimbriatus (cooke) D.A. Reid (polyporales: meripilaceae)... 37
çLÓPEZ-QUINTERO, C.A., VASCO-PALACIOS, A.M. & FRANCO-MOLANO, A.E., 2011.- Nuevos registros de macromicetes de Colombia I. Macromicetes recolectados en zonas urbanas de Medellín (Antioquia). Act Bio, 33 (95): 261-274.
MEDINA-JARITZ, N.B., PALACIOS-PACHECO, M.R. & VALENZUELA-GARZA, R., 2012.- Adiciones al conocimiento de los hongos poliporoides de Chiapas. Act. Bot. Mex, 101: 95-126.
PULIDO, M.M., 1983.- estudios en Agaricales Colombianos: los hongos de Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
RAYMUNDO, T. & VALENZUELA, V., 2003.- Los poliporáceos de México VI. Los hongos poliporoides del estado de Oaxaca. Polibotánica, 16: 79-112.
RAYNER, A.D.M., 1995.- Fungi, a vital component of ecosystem function in Woodland (en) ALLSOPP, D., COLWELL, R.R. & HOWKSWORTH, D.L. (eds.) Microbial Diversity and ecosystem Function. CAB International, Cambridge.
REID, D.A., 1962.- Notes on fungi which have been referred to the Thelephoraceae senso lato. Persoonia, 2 (2): 109-170. REID, D.A., 1976.- Notes on polypores. 2. Mem. NY Bot. Gard., 28 (1): 179-198.RICK, J., 1960.- Basidiomycetes Eubasisii in Rio Grande do Sul, Brasília 4. Meruliaceae, Polyporaceae, Boletaceae. Iheringia, Série
Botânica, 7: 193-295.RUIZ, A. & VARELA, A., 2006.- Nuevos registros de Aphyllophorales (Basidiomicota) en bosque montano húmedo y de niebla de
Colombia. Caldasia, 28 (2): 259-266. RYVARDEN, L., 1991.- Genera of Polypores: nomenclature and taxonomy. Fungiflora, Oslo.STAMETS, P., 1996.- Psilocybin Mushrooms of the World. Ten Speed Press Berkeley, California. TEIXEIRA, A.R., 1994.- Genera of Polyporaceae: an objective approach. Chácara Botânica, São Paulo.TEIXEIRA, A.R., 1995.- Método para estudo das hifas do basidiocarpo de fungos poliporáceos. Instituto de Botânica, São Paulo. VASCO-PALACIOS, A.M. & FRANCO-MOLANO, A.E., 2013.- Diversity of Colombian Macrofungi. (Ascomycota-Basidiomycota).
Mycotaxon, 121: 1-58. WRIGHT, J.E., & ALBERTÓ, E., 2006.- Hongos: Guía de los hongos la región Pampeana II. Hongos sin laminillas. L.O.L.A., Buenos Aires.WU, Q., W.U., THIERS, B.M. & PFISTER, D.H., 2004.- Preparation, preservation and use of Fungal specimens in herbaria: 23-
36 (en) MUELLER, G., BILLS, G. & FOSTER, M. (eds.) Biodiversity of Fungi: Inventory and Monitoring Methods. Elsevier Academic Press, London.
ZALAMEA, M., 2000.- estudio de la comunidad de hongos lignícolas en dos estadios sucesionales de un bosque alto andino: Tesis, Universidad Nacional de Colombia, Facultad de Ciencias, Bogotá.


ConserVaCiÓn y BiodiVersidaden aGroeCosisteMas
ConserVation and BiodiVersityin aGroeCosysteMs


Boletín CientífiCoCentro de Museos
Museo de Historia natural
anÁlisis de la ContaMinaCiÓn Por ColiforMes terMotolerantes en el estuario del río ranCHería,
la GuaJira (ColoMBia)*
Geomar Molina-Bolívar1, Iris Jiménez-Pitre2
resumen
El estuario del río Ranchería es una fuente hídrica de gran importancia para el municipio de Riohacha; formado por diferentes cuerpos de agua constituye un sistema dinámico y complejo que abastece tanto a centros poblados como rurales. Uno de los criterios más usados para la determinación de la calidad del agua es la medición de Coliformes Termotolerantes (CTT). Con el objetivo de estudiar la distribución espaciotemporal de estos elementos se colectaron muestras de agua en cinco zonas diferentes y se analizó el contenido de CTT mediante la técnica de tubos múltiples (NMP/100 ml). Los muestreos se realizaron cada 15 días, entre septiembre de 2014 y agosto de 2015, en los cuales no se establecieron diferencias estadísticamente significativas en el contenido de CTT entre los sitios, con excepción de EM (igualdad de varianza; ρ = 0,018), pero sí entre los periodos de lluvia y sequía (ρ = 0,027; ρ = 0,029) y en el cuarto trimestre en VF (ρ = 0,000), los mayores registros se encuentran relacionados con DB y CG, así como en las zonas más cercanas a los centros urbanos o de explotación agrícola.
Palabras clave: calidad del agua, coliformes termotolerantes, río Ranchería, distribución espacial.
analYsis of ContaMination BY tHerMotolerant ColiforM BaCteria in tHe estuarY of tHe ranCHeria
riVer, la GuaJira (ColoMBia)
abstract
The estuary of the Rancheria River is a water source of great importance for the municipality of Riohacha. Formed by different bodies of water, constitutes a dynamic and complex system that supplies both, urban centers and rural areas. One of the the most commonly used criteria for determining the quality of water is the measurement of Thermotolerant Coliforms (TC). With the objective of studying the spatiotemporal distribution of these elements, water samples were collected in five different areas and the content of TtC was analyzed using the multiple-tube technique (MPN/100 ml). Samplings were taken every fifteen days between September 2014 and August 2015, in which no statistically significant differences were found in the content of TtC between the sites where samples were taken, with the exception of EM (equality of variance; p = 0.018); but significant differences were found between the periods of rain and drought (ρ = 0.027; p = 0.029) and in the fourth quarter in VF (ρ = 0.000). The highest records are related to DB and CG, as well as in the areas closer to either urban centers or to agricultural exploitation centers.
Key words: water quality, fecal coliforms, Ranchería River, spatial distribution.
* FR: 17-06-2017. FA: 08-08-2017.1 Investigador Asociado, Universidad de La Guajira, Grupo de Investigación BIEMARC. Riohacha, Colombia. Autor para correspondencia. E-mail: [email protected] Investigador Sénior, Universidad de La Guajira, Grupo de Investigación BIEMARC. Riohacha, Colombia.
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 41-50. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: MOLINA-BOLÍVAR, G. & JIMÉNEZ-PITRE, I., 2017. - Análisis de la contaminación por Coliformes Termotol-erantes en el estuario del río Ranchería, La Guajira (Colombia). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 41-50. DOI: 10.17151/bccm.2017.21.2.3

Geomar Molina-Bolívar, Iris Jiménez-Pitre42bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 4
1-50
introduCCiÓn
El río Ranchería es uno de los cuerpos de agua más importantes del departamento de La Guajira en Colombia, nace en la laguna de Chirigüa a 3700 m.s.n.m., drena sus aguas por la vertiente oriental de la Sierra Nevada de Santa Marta, en sentido suroeste-noreste, y desemboca en el mar Caribe (RESTREPO-LÓPEZ et al., 2015). El delta estuarino se encuentra bajo un régimen climático bimodal está compuesto por lagunas, manglares y los brazos Calancalá y El Riíto, ubicados sobre el límite nororiental de la ciudad de Riohacha, forman la desembocadura principal del río donde ocurren diferentes procesos biológicos.
La desembocadura de ambos brazos y la mayor parte de la ribera de la laguna de Buenavista están conformadas por zonas de manglar (DÍAZ, 2001), lo anterior resalta la jerarquía del delta del río Ranchería, al abarcar áreas de influencia sobre manglares, litoral costero, lagunas y charcas temporales (PROYECTO PROLAGUNAS, 2001). Con un recorrido de 248 km, esta vertiente comparte ictiofauna con los sistemas de los ríos Magdalena y Catatumbo, este último perteneciente a la cuenca del Lago Maracaibo (MÓJICA et al., 2006). En estos ecosistemas marinos costeros varios factores contribuyen a su deterioro, entre los que se encuentran la sedimentación, anomalías climáticas, acumulación de contaminantes y basuras, interrupción de los flujos bioenergéticos, sobreexplotación de los recursos, utilización de artes de pesca altamente destructivas y actividades turísticas (GARAY et al., 2001), estos procesos de deterioro han venido ocurriendo desde épocas ancestrales, favorecidos por la acción antrópica (VÉLEZ & AGUIRRE, 2016).
Por la constante descarga en el brazo El Riíto de una serie de alcantarillas que sirven de drenaje a las calles del Distrito Turístico y Cultural de la ciudad de Riohacha en la época de lluvias, y porque durante todo el año existe comunicación con un cuerpo de agua pequeño que recoge las aguas residuales (POLANÍA et al., 2006), este sistema estuarino es un ecosistema altamente cambiante de naturaleza dinámica y abierta, en el que, gracias al intercambio de diferentes arroyuelos y el mar, tiene lugar una serie de procesos químicos y biológicos que permiten una alta productividad. Por tanto, los aspectos relacionados con la acumulación de materiales, producto de las aguas residuales y alcantarillado urbano, deben ser monitoreados a fin de mantener el equilibrio ecológico de la zona.
El cambio en los patrones hidrológicos de los ríos del Caribe colombiano se ha caracterizado por el fortalecimiento de la señal oscilatoria cuasi-decadal. La superposición de los principales componentes oscilatorios y el incremento pronunciado de los caudales después del año 2000 (RESTREPO et al., 2014), señalan incrementos promedios anuales hasta de un 17% en el caudal, con respecto a los promedios de largo plazo, lo cual tiene impacto sobre las dinámicas ecológicas en los cauces de

Análisis de la contaminación por coliformes termotolerantes en el estuario del río ranchería, la Guajira (Colombia) 43
agua afectados. Estos cambios han sido pobremente explorados, no poseen referentes sobre las tendencias en los elementos físicos, químicos y biológicos, lo cual resalta la importancia de un monitoreo periódico de la calidad del agua.
La contribución de los centros urbanos en el incremento de Coliformes Termotolerantes (CTT) en el agua de zonas marino-costeras es un factor de riesgo significativo para la creación de problemas de salud pública (RAMOS et al., 2008). A través de la utilización de herramientas moleculares se ha confirmado la presencia de escherichia coli asociada a aguas marino-costeras del noreste de Venezuela (MARTÍNEZ et al., 2015). Utilizando la técnica del Número Más Probable (NMP/100ml) para analizar el contenido de CTT en aguas superficiales de ríos de Perú, se determinó que el contenido es variable en función de la zona de muestreo, estando algunas zonas por encima del límite permitido por la legislación nacional peruana (TRUJILLO & GUERRERO, 2015). En el caso Colombiano, se ha estudiado que una contribución importante en el estudio de la microbiología diferencial en las aguas se dio en el río Combeima, principalmente en el contenido de coliformes totales hacia la época seca, lo cual señala la importancia de las descargas de aguas sin tratar por parte de los centros poblados rurales y explotaciones pecuarias, aun cuando el contenido de CTT encontrado no aumente significativamente.
Para facilitar la interpretación, comparación temporal y espacial de los datos, se utilizan expresiones matemáticas que representan a todos los parámetros posibles para evaluar el recurso hídrico y cuya aplicación es específica para cada región o fuente en particular, denominados índices de calidad de agua o bien índices de contaminación (SAMBONI et al., 2007). Un estudio de calidad del agua superficial de la microcuenca La Brava, en el departamento de Huila en Colombia, determinó que el contenido de CTT era significativo pero menor que en otras zonas más cercanas a centros poblados, mientras que en las zonas rurales de alta actividad pecuaria uno de los principales factores de contaminación son las heces de los animales en la mayoría de los casos, las cuales son vertidas en los cauces sin procesamiento alguno. En cualquier caso, dependiendo de la localidad y la época del año, aunado a la actividad antrópica, el contenido de contaminación microbiana es dinámico, por lo que requiere de su constante monitoreo (ANGARITA & ORTIZ, 2014). Teniendo en cuenta lo anteriormente mencionado, el objetivo del presente trabajo fue analizar la distribución espacial y estacional de CTT en el estuario del río Ranchería como elemento biológico fundamental para la estimación de la calidad del agua.
Materiales Y MÉtodos
En la zona del estuario del río Ranchería, ubicado en el departamento de la Guajira en Colombia, se realizaron 18 muestreos entre septiembre de 2014 y agosto de 2015, realizados en cinco localidades: Desembocadura (DB), El Mirador (EM), Villa Fátima (VF), Cangrejito (CG), Valle de los Cangrejos (VC) (Figura 1).

Geomar Molina-Bolívar, Iris Jiménez-Pitre44bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 4
1-50
Ubicación del área de estudio con sitios de muestreos (brazos El Riíto y Calancalá).figura 1.
Las muestras de agua superficial fueron colectadas de forma manual, siguiendo el procedimiento para análisis bacteriológico. Las muestras fueron recogidas en frascos esterilizados de 250 ml de capacidad, los cuales fueron abiertos y cerrados bajo el agua, a fin de evitar contaminación de las muestras.
Se analizaron los CTT (NMP/100 ml) siguiendo la metodología de tubos múltiples de la técnica del NMP por su exactitud, rapidez y por el recuento directo de colonias. La enumeración de CTT por NMP/100 ml se realiza en una etapa. Los resultados se expresan como colonias de CTT por 100 ml (NMP/100 ml). La calidad química del agua se evaluó en función de lo establecido en el Decreto 1575 del 9 de mayo de 2007 (MINISTERIO DE LA PROTECCIÓN SOCIAL, 2007) y en la Resolución 2115 de 22 junio de 2007 (MINISTERIO DE LA PROTECCIÓN SOCIAL & MINISTERIO DE AMBIENTE, VIVIENDA Y DESARROLLO TERRITORIAL, 2007).
El estudio siguió un modelo estadístico completamente aleatorizado, con los sitios de muestreo como tratamiento y el contenido de CTT como la variable dependiente. Inicialmente los datos se transformaron con Log10, para posteriormente realizar el análisis los datos mediante estadística descriptiva para establecer la calidad de la información obtenida, para lo cual se evaluó el comportamiento de estadísticos de posición, tendencia central, dispersión y forma. Con la prueba de Tukey se realizó
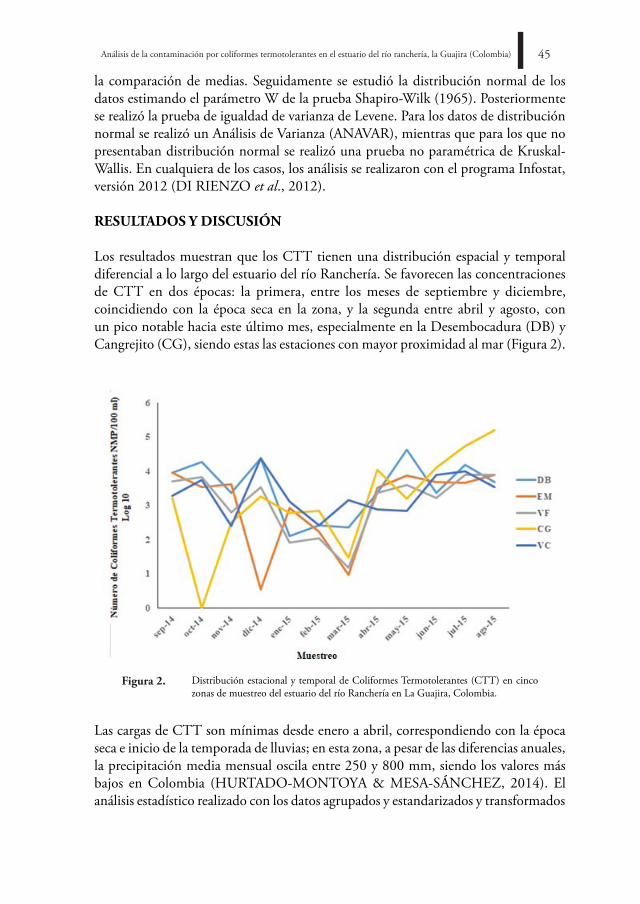
Análisis de la contaminación por coliformes termotolerantes en el estuario del río ranchería, la Guajira (Colombia) 45
la comparación de medias. Seguidamente se estudió la distribución normal de los datos estimando el parámetro W de la prueba Shapiro-Wilk (1965). Posteriormente se realizó la prueba de igualdad de varianza de Levene. Para los datos de distribución normal se realizó un Análisis de Varianza (ANAVAR), mientras que para los que no presentaban distribución normal se realizó una prueba no paramétrica de Kruskal-Wallis. En cualquiera de los casos, los análisis se realizaron con el programa Infostat, versión 2012 (DI RIENZO et al., 2012).
resultados Y disCusiÓn
Los resultados muestran que los CTT tienen una distribución espacial y temporal diferencial a lo largo del estuario del río Ranchería. Se favorecen las concentraciones de CTT en dos épocas: la primera, entre los meses de septiembre y diciembre, coincidiendo con la época seca en la zona, y la segunda entre abril y agosto, con un pico notable hacia este último mes, especialmente en la Desembocadura (DB) y Cangrejito (CG), siendo estas las estaciones con mayor proximidad al mar (Figura 2).
Distribución estacional y temporal de Coliformes Termotolerantes (CTT) en cinco zonas de muestreo del estuario del río Ranchería en La Guajira, Colombia.
figura 2.
Las cargas de CTT son mínimas desde enero a abril, correspondiendo con la época seca e inicio de la temporada de lluvias; en esta zona, a pesar de las diferencias anuales, la precipitación media mensual oscila entre 250 y 800 mm, siendo los valores más bajos en Colombia (HURTADO-MONTOYA & MESA-SÁNCHEZ, 2014). El análisis estadístico realizado con los datos agrupados y estandarizados y transformados
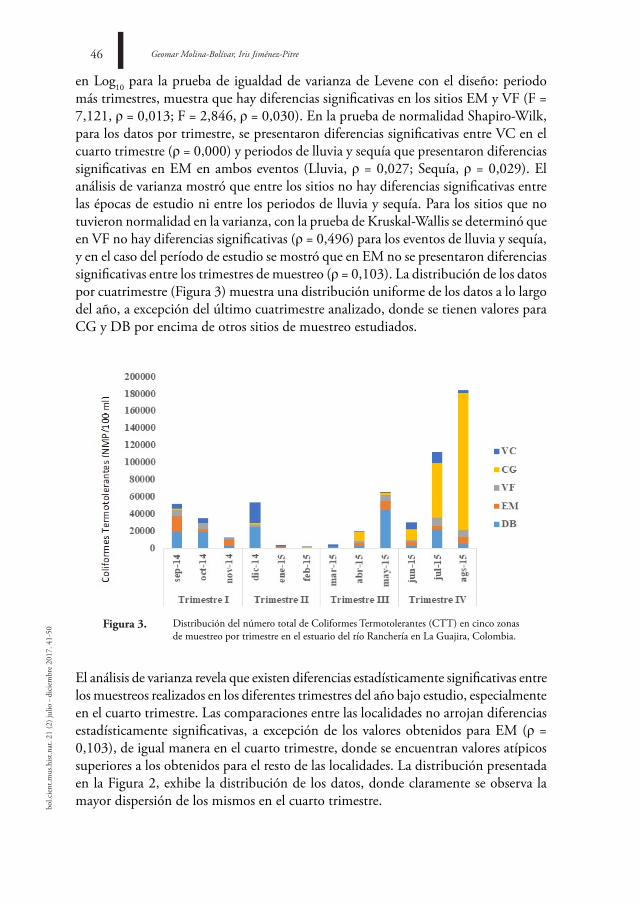
Geomar Molina-Bolívar, Iris Jiménez-Pitre46bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 4
1-50
en Log10 para la prueba de igualdad de varianza de Levene con el diseño: periodo más trimestres, muestra que hay diferencias significativas en los sitios EM y VF (F = 7,121, ρ = 0,013; F = 2,846, ρ = 0,030). En la prueba de normalidad Shapiro-Wilk, para los datos por trimestre, se presentaron diferencias significativas entre VC en el cuarto trimestre (ρ = 0,000) y periodos de lluvia y sequía que presentaron diferencias significativas en EM en ambos eventos (Lluvia, ρ = 0,027; Sequía, ρ = 0,029). El análisis de varianza mostró que entre los sitios no hay diferencias significativas entre las épocas de estudio ni entre los periodos de lluvia y sequía. Para los sitios que no tuvieron normalidad en la varianza, con la prueba de Kruskal-Wallis se determinó que en VF no hay diferencias significativas (ρ = 0,496) para los eventos de lluvia y sequía, y en el caso del período de estudio se mostró que en EM no se presentaron diferencias significativas entre los trimestres de muestreo (ρ = 0,103). La distribución de los datos por cuatrimestre (Figura 3) muestra una distribución uniforme de los datos a lo largo del año, a excepción del último cuatrimestre analizado, donde se tienen valores para CG y DB por encima de otros sitios de muestreo estudiados.
Distribución del número total de Coliformes Termotolerantes (CTT) en cinco zonas de muestreo por trimestre en el estuario del río Ranchería en La Guajira, Colombia.
figura 3.
El análisis de varianza revela que existen diferencias estadísticamente significativas entre los muestreos realizados en los diferentes trimestres del año bajo estudio, especialmente en el cuarto trimestre. Las comparaciones entre las localidades no arrojan diferencias estadísticamente significativas, a excepción de los valores obtenidos para EM (ρ = 0,103), de igual manera en el cuarto trimestre, donde se encuentran valores atípicos superiores a los obtenidos para el resto de las localidades. La distribución presentada en la Figura 2, exhibe la distribución de los datos, donde claramente se observa la mayor dispersión de los mismos en el cuarto trimestre.
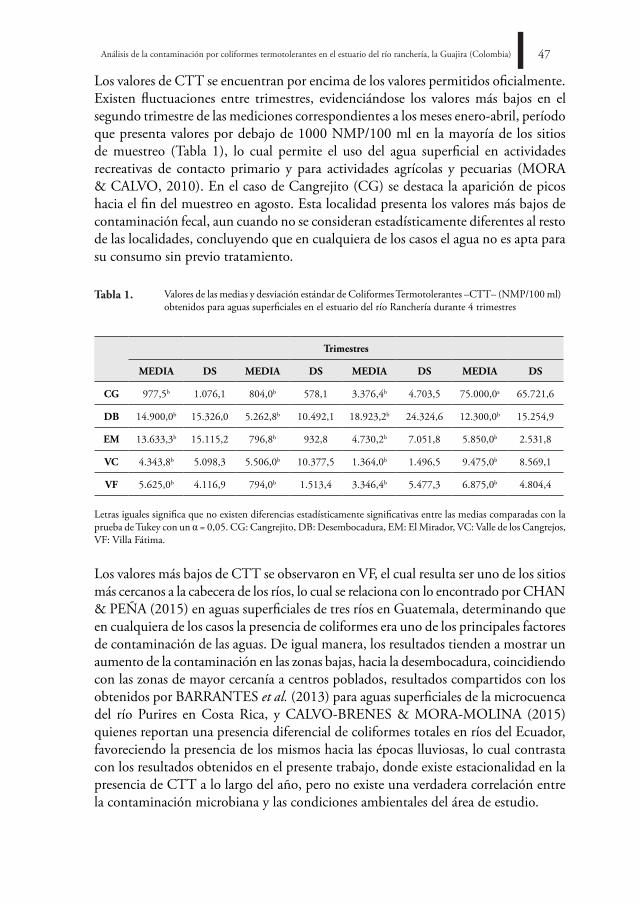
Análisis de la contaminación por coliformes termotolerantes en el estuario del río ranchería, la Guajira (Colombia) 47
Los valores de CTT se encuentran por encima de los valores permitidos oficialmente. Existen fluctuaciones entre trimestres, evidenciándose los valores más bajos en el segundo trimestre de las mediciones correspondientes a los meses enero-abril, período que presenta valores por debajo de 1000 NMP/100 ml en la mayoría de los sitios de muestreo (Tabla 1), lo cual permite el uso del agua superficial en actividades recreativas de contacto primario y para actividades agrícolas y pecuarias (MORA & CALVO, 2010). En el caso de Cangrejito (CG) se destaca la aparición de picos hacia el fin del muestreo en agosto. Esta localidad presenta los valores más bajos de contaminación fecal, aun cuando no se consideran estadísticamente diferentes al resto de las localidades, concluyendo que en cualquiera de los casos el agua no es apta para su consumo sin previo tratamiento.
Valores de las medias y desviación estándar de Coliformes Termotolerantes –CTT– (NMP/100 ml) obtenidos para aguas superficiales en el estuario del río Ranchería durante 4 trimestres
tabla 1.
trimestres
Media ds Media ds Media ds Media ds
CG 977,5b 1.076,1 804,0b 578,1 3.376,4b 4.703,5 75.000,0a 65.721,6
dB 14.900,0b 15.326,0 5.262,8b 10.492,1 18.923,2b 24.324,6 12.300,0b 15.254,9
eM 13.633,3b 15.115,2 796,8b 932,8 4.730,2b 7.051,8 5.850,0b 2.531,8
VC 4.343,8b 5.098,3 5.506,0b 10.377,5 1.364,0b 1.496,5 9.475,0b 8.569,1
Vf 5.625,0b 4.116,9 794,0b 1.513,4 3.346,4b 5.477,3 6.875,0b 4.804,4
Letras iguales significa que no existen diferencias estadísticamente significativas entre las medias comparadas con la prueba de Tukey con un α = 0,05. CG: Cangrejito, DB: Desembocadura, EM: El Mirador, VC: Valle de los Cangrejos, VF: Villa Fátima.
Los valores más bajos de CTT se observaron en VF, el cual resulta ser uno de los sitios más cercanos a la cabecera de los ríos, lo cual se relaciona con lo encontrado por CHAN & PEÑA (2015) en aguas superficiales de tres ríos en Guatemala, determinando que en cualquiera de los casos la presencia de coliformes era uno de los principales factores de contaminación de las aguas. De igual manera, los resultados tienden a mostrar un aumento de la contaminación en las zonas bajas, hacia la desembocadura, coincidiendo con las zonas de mayor cercanía a centros poblados, resultados compartidos con los obtenidos por BARRANTES et al. (2013) para aguas superficiales de la microcuenca del río Purires en Costa Rica, y CALVO-BRENES & MORA-MOLINA (2015) quienes reportan una presencia diferencial de coliformes totales en ríos del Ecuador, favoreciendo la presencia de los mismos hacia las épocas lluviosas, lo cual contrasta con los resultados obtenidos en el presente trabajo, donde existe estacionalidad en la presencia de CTT a lo largo del año, pero no existe una verdadera correlación entre la contaminación microbiana y las condiciones ambientales del área de estudio.
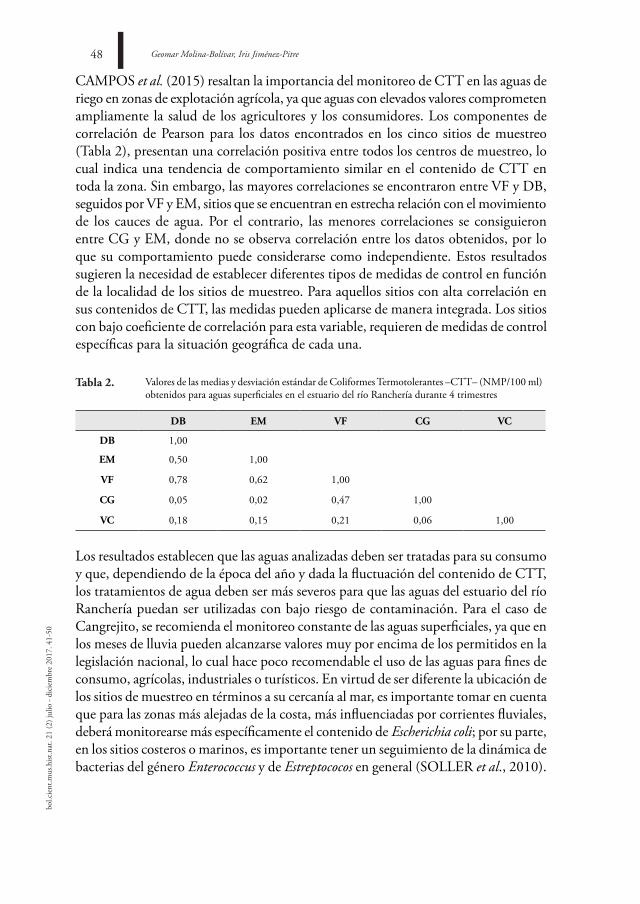
Geomar Molina-Bolívar, Iris Jiménez-Pitre48bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 4
1-50
CAMPOS et al. (2015) resaltan la importancia del monitoreo de CTT en las aguas de riego en zonas de explotación agrícola, ya que aguas con elevados valores comprometen ampliamente la salud de los agricultores y los consumidores. Los componentes de correlación de Pearson para los datos encontrados en los cinco sitios de muestreo (Tabla 2), presentan una correlación positiva entre todos los centros de muestreo, lo cual indica una tendencia de comportamiento similar en el contenido de CTT en toda la zona. Sin embargo, las mayores correlaciones se encontraron entre VF y DB, seguidos por VF y EM, sitios que se encuentran en estrecha relación con el movimiento de los cauces de agua. Por el contrario, las menores correlaciones se consiguieron entre CG y EM, donde no se observa correlación entre los datos obtenidos, por lo que su comportamiento puede considerarse como independiente. Estos resultados sugieren la necesidad de establecer diferentes tipos de medidas de control en función de la localidad de los sitios de muestreo. Para aquellos sitios con alta correlación en sus contenidos de CTT, las medidas pueden aplicarse de manera integrada. Los sitios con bajo coeficiente de correlación para esta variable, requieren de medidas de control específicas para la situación geográfica de cada una.
Valores de las medias y desviación estándar de Coliformes Termotolerantes –CTT– (NMP/100 ml) obtenidos para aguas superficiales en el estuario del río Ranchería durante 4 trimestres
tabla 2.
dB eM Vf CG VC
dB 1,00
eM 0,50 1,00
Vf 0,78 0,62 1,00
CG 0,05 0,02 0,47 1,00
VC 0,18 0,15 0,21 0,06 1,00
Los resultados establecen que las aguas analizadas deben ser tratadas para su consumo y que, dependiendo de la época del año y dada la fluctuación del contenido de CTT, los tratamientos de agua deben ser más severos para que las aguas del estuario del río Ranchería puedan ser utilizadas con bajo riesgo de contaminación. Para el caso de Cangrejito, se recomienda el monitoreo constante de las aguas superficiales, ya que en los meses de lluvia pueden alcanzarse valores muy por encima de los permitidos en la legislación nacional, lo cual hace poco recomendable el uso de las aguas para fines de consumo, agrícolas, industriales o turísticos. En virtud de ser diferente la ubicación de los sitios de muestreo en términos a su cercanía al mar, es importante tomar en cuenta que para las zonas más alejadas de la costa, más influenciadas por corrientes fluviales, deberá monitorearse más específicamente el contenido de escherichia coli; por su parte, en los sitios costeros o marinos, es importante tener un seguimiento de la dinámica de bacterias del género enterococcus y de estreptococos en general (SOLLER et al., 2010).

Análisis de la contaminación por coliformes termotolerantes en el estuario del río ranchería, la Guajira (Colombia) 49
Si bien en la mayoría de los muestreos los valores son significativamente menores, los mismos sobrepasan lo recomendado por organismos reguladores internacionales, como la Agencia de Protección Ambiental en EE.UU. (USEPA por sus siglas en inglés) y el Ministerio de Salud de Canadá (Health Canada), los cuales establecen como valores máximos 126 NMP/100 ml en el caso de USEPA y 200 NMP/100 ml en Canadá (LÈVESQUE & GAUVIN, 2007; USEPA, 2012). Asimismo, es importante resaltar que los puntos de muestreo con mayor contaminación se ubicaron en las zonas más bajas del estuario donde existe una mayor densidad poblacional. Esta información es importante para el desarrollo de acciones de control específicas para cada una de estas zonas geográficas, con el fin de evitar el efecto negativo de fuentes de contaminación que pueden afectar la calidad de las aguas y, por ende, la salud de las comunidades ubicadas en su cercanía.
ConClusiones
El contenido de Coliformes Termotolerantes (CTT) en el estuario del río Ranchería en La Guajira, Colombia, fue diferente en los cinco sitios de muestreo, con características estacionales diferenciales. Según las pruebas de normalidad no se encontraron diferencias estadísticamente significativas entre los sitios analizados. Excepto en VC en el trimestre cuarto (ρ = 0,000) y en EM los periodos de lluvia y sequía (ρ = 0,027; ρ = 0,029). Se favoreció el contenido de CTT en la zona de Cangrejito hacia el cuarto trimestre, donde se observaron valores atípicamente altos. Los menores valores se encontraron en VF, correspondiendo al sitio más cercano a la cabecera de los ríos, donde el recorrido del agua había sido menor. No se encontró correlación entre el contenido de CTT entre los sitios, con excepción de CG con DB y EM (ρ = 0,05; ρ = 0,02). El agua presenta condiciones de calidad no apta para el consumo humano, ni se recomienda su uso para el riego agrícola, requiriendo de tratamientos específicos para tal fin.
aGradeCiMientos
Los autores desean expresar su agradecimiento a la Universidad de La Guajira y a la Corporación Autónoma de La Guajira (CORPOGUAJIRA), por su apoyo financiero y logístico para la realización de esta investigación.
referenCias
ANGARITA, W. & ORTIZ, E., 2014.- Caracterización fisicoquímica y microbiológica del agua en la micro cuenca la Brava, la cual abastece el acueducto independiente Adamiuain en el sector norte de la ciudad de ocaña: Tesis, Universidad Francisco de Paula Santander, Facultad de Ciencias Agrarias y del Ambiente, Ocaña.
BARRANTES, K., CHACÓN, L.M., SOLANO, M. & ACHÍ, R., 2013.- Contaminación fecal del agua superficial de la microcuenca del río Purires, Costa Rica, 2010-2011. Revista de la Sociedad Venezolana de Microbiología, 33: 40-45.
CALVO-BRENES, G. & MORA-MOLINA, J., 2015.- Evaluación de la calidad del agua en los ríos Tigre y Rincón de la península de Osa en dos períodos de tiempo distintos. Tecnología en Marcha, 28 (3): 55-63.

Geomar Molina-Bolívar, Iris Jiménez-Pitre50bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 4
1-50
CAMPOS, C., CONTRERAS, A.M. & LEIVA, F.R., 2015.- Evaluación del riesgo sanitario en un cultivo de lechuga (lactuca sativa) debido al riego con aguas residuales sin tratar en el Centro Agropecuario Marengo (Cundinamarca, Colombia). Biosalud, 14 (1): 69-78.
CHAN, M. & PEÑA, W., 2015.- Evaluación de la calidad del agua superficial con potencial para consumo humano en la cuenca alta del Sis Icán, Guatemala. Cuadernos de Investigación UNeD, 7 (1): 19-23.
DÍAZ, K., 2001.- estudio de la regeneración natural del manglar en el Riíto y el Valle de los Cangrejos, delta del río Ranchería. Universidad de La Guajira, Riohacha.
DI RIENZO, J.A., CASANOVES, F., BALZARINI, M.G., GONZÁLEZ, L., TABLADA, M. & ROBLEDO, C.W., 2012.- InfoStat versión 2012. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
GARAY, J., MARÍN, B., RAMÍREZ, B., VÉLEZ, A., TRONCOSO, W., LOZANO, H., ACOSTA, J., CADAVID, B., LANCHEROS, A., MEDINA, O. & RONDÓN, M., 2001.- Diagnóstico y evaluación de la calidad ambiental marina en el Caribe y Pacífico Colombiano. Red de vigilancia para la conservación y protección de las aguas marinas y costera de Colombia – REDCAM. Informe técnico final, INVEMAR. Santa Marta.
HURTADO-MONTOYA, A.F. & MESA-SÁNCHEZ, O.J., 2014.- Reanalysis of monthly precipitation fields in Colombian territory. DYNA, 81 (186): 251-258.
LÈVESQUE, B. & GAUVIN, D., 2007.- Microbiological guideline values for recreational bathing in Canada: time for change? Can J Infect Dis Med Microbiol, 2: 153-7.
MARTÍNEZ, R.E., VILLALOBOS, L.B. & CASTILLO, L.K., 2015.- escherichia coli diarreogénicas procedentes de aguas marinas recreacionales caracterizadas por reacción en cadena de la polimerasa. Revista Científica, FCV-lUZ, XXV (3): 248-254.
MINISTERIO DE LA PROTECCIÓN SOCIAL, 2007.- Decreto 1575 del 9 de mayo de 2007. Por el cual se establece el Sistema para la Protección y Control de la Calidad del Agua para Consumo Humano. Diario Oficial 46623. Imprenta Nacional de Colombia, Bogotá.. Recuperado de http://www.ins.gov.co/sivicap/Normatividad/Decreto%201575%20de%202007.pdf
MINISTERIO DE LA PROTECCIÓN SOCIAL & MINISTERIO DE AMBIENTE, VIVIENDA Y DESARROLLO TERRITORIAL, 2007.- Resolución 2115 de 22 junio de 2007. Por medio de la cual se señalan características, instrumentos básicos y frecuencias del sistema de control y vigilancia para la calidad del agua para consumo humano. Diario Oficial 46679. Imprenta Nacional de Colombia, Bogotá. Recuperado de http://www.ins.gov.co/sivicap/Normatividad/Resoluci%C3%B3n%202115%20de%202007.pdf
MOJICA, J.I., CASTELLANOS, C., SÁNCHEZ-DUARTE, P. & DÍAZ, C., 2006.- Peces de la cuenca del río Ranchería, La Guajira, Colombia. Biota Colombiana, 7: 129142.
MORA, J. & CALVO, G., 2010.- Estado actual de contaminación con coliformes fecales de los cuerpos de agua de la Península de Osa. Tecnología en Marcha, 23 (5): 34-40.
POLANÍA, J., OROZCO-TORO, C.A. & ÁNGEL, I.F., 2006.- Delta del río Ranchería (La Guajira, Colombia): caudal, salinidad y transporte de sólidos y su posible influencia sobre composición y estructura de los manglares. Actual Biol, 28 (84): 27-37.
PROYECTO PROLAGUNAS, 2001.- Protección y recuperación de humedales costeros del Caribe colombiano. Universidad Jorge Tadeo Lozano, Bogotá.
RAMOS, L., VIDAL, L., VILARDI, S. & SAAVEDRA, L., 2008.- Análisis de la contaminación microbiológica (Coliformes totales y fecales) en la Bahía de Santa Marta, Caribe Colombiano. Act. Biol. Colomb., 13 (3): 87-98.
RESTREPO, J.C., ORTIZ, J.C., PIERINI, J., SCHROTTKE, K., MAZA, M. & OTERO, L., 2014.- Freshwater discharge into the Caribbean Sea from the Rivers of Northwestern South America (Colombia): Magnitude, variability and recent changes. Journal of Hydrology, 509: 266-281.
RESTREPO-LÓPEZ, J.C., ORTIZ-ROYERO, J.C., OTERO-DÍAZ, L. & OSPINO-ORTIZ, S.R., 2015.- Transporte de sedimentos en suspensión en los principales ríos del Caribe colombiano: magnitud, tendencias y variabilidad. Revista de la Academia Colombiana de Ciencias exactas, Físicas y Naturales, 39 (153): 527-546.
SAMBONI, N.E., CARVAJAL, Y. & ESCOBAR, J.C., 2007.- Revisión de parámetros físico-químicos como indicadores de calidad y contaminación del agua. Ingeniería e Investigación, 27 (3): 172-181.
SOLLER, J., SCHOEN, M.E., BARTRAND, T., RAVENSCROFT, J.E. & ASHBOLT, N.J., 2010.- Estimated human health risks from exposure to recreational waters impacted by human and non-human sources of fecal contamination. Water Res, 44: 4674-91.
TRUJILLO, G.P. & GUERRERO, A.M., 2015.- Caracterización físico-química y bacteriológica del agua marina en la zona litoral costera de Huanchaco y Huanchaquito, Trujillo, Perú. ReBIol, 35 (1): 23-33.
UNITED STATES ENVIRONMENTAL PROTECTION AGENCY (USEPA), 2012.- Recreational Water Quality Criteria. Recuperado de http:// water.epa.gov/scitech/swguidance/standards/criteria/health/recreation/index.cfm
VÉLEZ, C. & AGUIRRE, N., 2016.- Influencia del río Atrato en el Golfo de Urabá durante el holoceno tardío, Mar Caribe colombiano. Bol. Invest. Mar. Cost., 45 (1): 73-97.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
Control alternatiVo de Diaphorina citri KuWaYaMa (HeMiPtera: liViidae) utiliZando Caldo
sulfoCÁlCiCo*
Ana María Restrepo-García1, Alberto Soto-Giraldo2
resumen
La concentración letal (CL) es usada para estimar la toxicidad de plaguicidas a artrópodos. Sin embargo, la CL es una medida incompleta de los efectos de los productos sobre poblaciones, pues analiza solamente la mortalidad como parámetro de toxicidad. Se sabe que individuos que sobreviven a la exposición a plaguicidas pueden sufrir efectos subletales. En este trabajo fue estudiado en laboratorio los efectos letales y subletales del caldo sulfocálcico en el crecimiento poblacional de Diaphorina citri. Los efectos letales fueron estimados a través de test de toxicidad aguda con obtención de curvas dosis-respuesta y sus respectivas concentraciones letales, y los efectos subletales a través de la estimativa de la tasa instantánea de crecimiento poblacional (ri). Las concentraciones letales y subletales del caldo sulfocálcico para D. citri fueron de 0,57 y 0,38% del producto, respectivamente, lo que indica que podría utilizarse para el control del fitófago.
Palabras clave: azufre, Huanglongbing, Candidatus liberibacter, productos alternativos.
alternatiVe Control of Diaphorina citri KuWaYaMa (HeMiPtera: liViidae) usinG liMe sulPHur
The lethal concentration (LC) is used to estimate the toxicity of pesticides in arthropods. However, LC is an incomplete measure of the effects of these products on populations, since it only examines mortality as a parameter of toxicity. It is known that individuals which survive exposure to pesticides may suffer sublethal effects. Therefore, the lethal and sublethal effects of lime sulfur spray on Diaphorina citri was studied in the laboratory. Lethal effects were estimated through tests of acute toxicity to obtain dosage-response curves and their respective lethal concentrations and sublethal effects by estimating the instantaneous population growth rate (ri). The lethal and sublethal concentrations of lime Sulphur for D. citri were 0.57 and 0.38% of the product respectively, indicating that it could be used to control this phytophagous.
Key words: sulfur, Huanglongbing, Candidatus liberibacter, alternative products.
* FR:20-V-2017 . FA: 20-IX-20171 Ingeniera Agrónoma. Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] Ph.D. Departamento de Producción Agropecuaria, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0002-9727-8919
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 51-60. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:RESTREPO-GARCÍA, A.A. & SOTO-GIRALDO, A., 2017.- Control alternativo de Diaphorina citri kuwayama (Hemiptera: Liviidae) utilizando caldo sulfocálcico. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 51-60. DOI: 10.17151/bccm.2017.21.2.4

Ana María Restrepo-García, Alberto Soto-Giraldo52bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 5
1-60
introduCCiÓn
La producción mundial de cítricos es superior a los 100 millones de ton anuales; es el grupo de frutales más cultivado en Colombia para el consumo interno, con una participación de 1’681.877 ton, y es el segundo en área cultivada después del banano (ORDUZ & MATEUS, 2012; DANE, 2016). En el país, el cultivo de los cítricos posee 160.408 has plantadas, lo que equivale al 12% de la superficie nacional dedicada al cultivo de frutas, de las cuales el 46,8% se encuentran distribuidas en los departamentos de Caldas, Santander, Tolima, Antioquia y Cauca (DANE, 2016).
Los cítricos, como todas las especies cultivadas, son susceptibles al ataque de insectos, ácaros, hongos, virus, bacterias y nematodos fitoparásitos que afectan la cantidad y calidad de las cosechas. El ataque de insectos al cultivo no radica únicamente en el daño directo que ocasiona, sino también en la capacidad que tienen de transmitir enfermedades de una planta a otra, resultando en pérdidas económicamente significativas, como es el caso del psílido asiático Diaphorina citri Kuwayama (Hemiptera: Liviidae), una de las plagas más devastadoras que atacan el cultivo de los cítricos (HALBERT & MANJUNATH, 2004; FERNÁNDEZ & MIRANDA, 2005; ALEMÁN et al., 2007; ORTEGA et al., 2013), por ser el vector de la bacteria Candidatus liberibacter que causa la enfermedad denominada Huanglongbing (HLB) o enverdecimiento de los cítricos (CHUNG & BRLANSKY, 2005; GRAFTON et al., 2006).
Cuando los árboles se enferman por la presencia de la bacteria, el fruto sufre diversas afectaciones que interfieren en la calidad para su posterior comercialización (RESTREPO-GARCÍA et al., 2016); el diámetro se reduce 17%, no presenta un color uniforme, el brillo baja 13%, se pierde 7% de jugo, mientras que el ácido se incrementa 29% (ROGERS, 2011). En el año 2015 se estableció la emergencia fitosanitaria en el territorio nacional por la presencia de adultos de D. citri infectados con dicha bacteria (ICA, 2015); y en 2016 se declaró en cuarentena fitosanitaria a los departamentos de La Guajira y Atlántico por presencia de la bacteria en material vegetal (ICA, 2016a, 2016b).
El manejo de la enfermedad está dirigido a controlar el insecto vector, ya que esta no tiene cura; para ello, los agricultores utilizan principalmente plaguicidas de síntesis química, lo que puede ocasionar problemas de selección de individuos resistentes debido al uso continuo de ingredientes activos, reducción o supresión de especies benéficas, alta toxicidad de los productos a los aplicadores, presencia de residuos en los alimentos y en el medio ambiente, fitotoxicidad, entre otros (FILGUEIRA, 2000; EASTERBROOK et al., 2001; FRAGOSO et al., 2002; ALEMÁN et al., 2007; PICANÇO et al., 2007).

Control alternativo de diaphorina citri kuwayama (hemiptera: liviidae) utilizando caldo sulfocálcico 53
Una alternativa viable a los problemas generados por el uso excesivo de estos productos es la utilización de métodos de control que deben priorizar la seguridad ambiental y social, y que sean eficientes en el control del insecto (FAJARDO et al., 2013). En la búsqueda de tales métodos, productos alternativos a los químicos convencionales como los caldos fitoprotectores han sido usados por productores de hortalizas para el control de plagas y enfermedades, especialmente en los sistemas de producción ecológicos y orgánicos (CAMPANHOLA & BETTIOL, 2003; VENZON et al., 2007; FAJARDO et al., 2013); dentro de estos, se encuentra el caldo sulfocálcico, que es obtenido por el tratamiento térmico del azufre y la cal (ABBOT, 1945). Esta mezcla es conocida, principalmente, debido a su acción fungicida (SMILANICK & SORENSON, 2001; MONTAG et al., 2005), y también es utilizada como acaricida e insecticida (WARDLOW & LUDLAM, 1975; GUERRA, 1985; PENTEADO, 2000; CHAGAS et al., 2001; GUIRADO, 2001; OCETE et al., 2003; VENZON et al., 2007; SOTO, 2009; SOTO et al., 2011; FAJARDO et al., 2013). El efecto tóxico del caldo sulfocálcico a los insectos se da por la reacción de los compuestos de este sobre la planta, con el agua y el gas carbónico, resultando en gas sulfídrico y azufre coloidal (ABBOT, 1945).
En este trabajo se evaluó el efecto insecticida del caldo sulfocálcico sobre el psílido asiático de los cítricos D. citri en condiciones controladas. Inicialmente, se evaluó la toxicidad letal a través de la estimativa de la concentración letal (CL) del producto; sin embargo, como la CL es una medida incompleta de los efectos de los productos sobre poblaciones, pues analiza solamente la mortalidad como parámetro de toxicidad (STARK & BANKS, 2003), se evaluaron también los efectos subletales de los productos a través de la estimativa de la tasa instantánea de crecimiento poblacional (ri) (STARK & RANGUS, 1994).
Materiales Y MÉtodos
El estudio se realizó en el Centro de Investigación y Cría de Enemigos Naturales de la Universidad de Caldas, ubicado en el municipio de Manizales (coordenadas: 05°03’ N - 75°29’ W). Los especímenes de D. citri fueron criados en plantas de Swinglea glutinosa de 40 cm de longitud, acondicionadas en jaulas de madera de 1 m x 1 m x 1 m, forradas con tela muselina (Figura 1A). La cría se inició con ninfas y adultos colectados en campo en la granja Montelindo de la Universidad de Caldas (Figuras 1B y C). Posteriormente, fueron estimadas las concentraciones letales del caldo sulfocálcico sobre ninfas de D. citri. Las concentraciones evaluadas fueron seleccionadas a través de bioensayos preliminares, se situaron entre el límite inferior, donde el producto no causó mortalidad, y el límite superior de respuesta, donde causó 100% de mortalidad.

Ana María Restrepo-García, Alberto Soto-Giraldo54bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 5
1-60
A B C
Establecimiento de cría de D. citri. a: Jaulas. B y C: Adultos y ninfas de D. citri en brotes de Swinglea glutinosa. (Fotografías: A.M. Restrepo).
figura 1.
El efecto letal y subletal del caldo sulfocálcico sobre D. citri fue estudiado de acuerdo con metodología adaptada de PENTEADO (2000), manejando tratamiento térmico del azufre y cal virgen, utilizando para cada litro de agua: 250 g de azufre y 125 g de cal virgen (Figura 2A y B). La concentración obtenida de caldo sulfocálcico fue de 29° Baumé.
Caldo sulfocálcico. a: Preparación del producto. B: Medición de los grados Baumé. (Fotografías: A.M. Restrepo).
figura 2.
Para los bioensayos de concentración-respuesta se seleccionaron ramas de cítricos, las cuales se infestaron con ninfas de D. citri en todos los estados de desarrollo (Figura 3A). Los brotes fueron depositados en recipientes trasparentes de 11,5 cm de largo por 11 cm de diámetro (capacidad de 30 oz) y al interior de ellos se colocó una base de espuma humedecida, después fueron expuestos a la aplicación directa con el producto a diferentes concentraciones (0,15; 0,25; 0,35; 0,45 y 0,5%) (Figura 3B). También, se utilizó un testigo absoluto y por cada tratamiento se realizaron 5 repeticiones.
A B

Control alternativo de diaphorina citri kuwayama (hemiptera: liviidae) utilizando caldo sulfocálcico 55
A B C
Montaje del experimento. a: Ramas infestadas. B: Aplicación de los tratamientos. C: Unidades experimentales en incubadora. (Fotografías: A.M. Restrepo).
figura 3.
Los recipientes se cubrieron con tela muselina para facilitar la aireación y se ubicaron en una incubadora marca Scientific a temperatura de 25°C±2°C y 60±10% HR (Figura 3C). La mortalidad fue evaluada 24 h después de la aplicación de los productos, y las curvas de concentración-mortalidad fueron estimadas por el análisis de Probit (FINNEY, 1971).
La evaluación de la acción subletal de los productos sobre las ninfas de D. citri fue aplicada utilizando la misma metodología citada para la evaluación de la acción letal de los productos. Esta se realizó a través de la estimativa de la tasa instantánea de crecimiento (ri), por la siguiente fórmula (STARK & BANKS, 2003):
ri = ln (nf / n0)/ Δt
Donde Nf es el número final de individuos, N0
es el número inicial de individuos, Δt
es la variación de tiempo (duración del experimento: 15 días). El valor positivo de ri
significa que la población está en crecimiento; ri = 0, indica que la población está
estable, mientras que un valor negativo de ri señala que la población está en descenso
y en vías de extinción. El análisis de regresión se aprovechó para la evaluación de ri en
función de las concentraciones utilizadas.
resultados Y disCusiÓn
En la Toxicidad letal del caldo sulfocálcico fueron obtenidas la Cl50 de 0,38 y la Cl95
de 0,57% de concentración del producto a 29º B (Tabla 1).
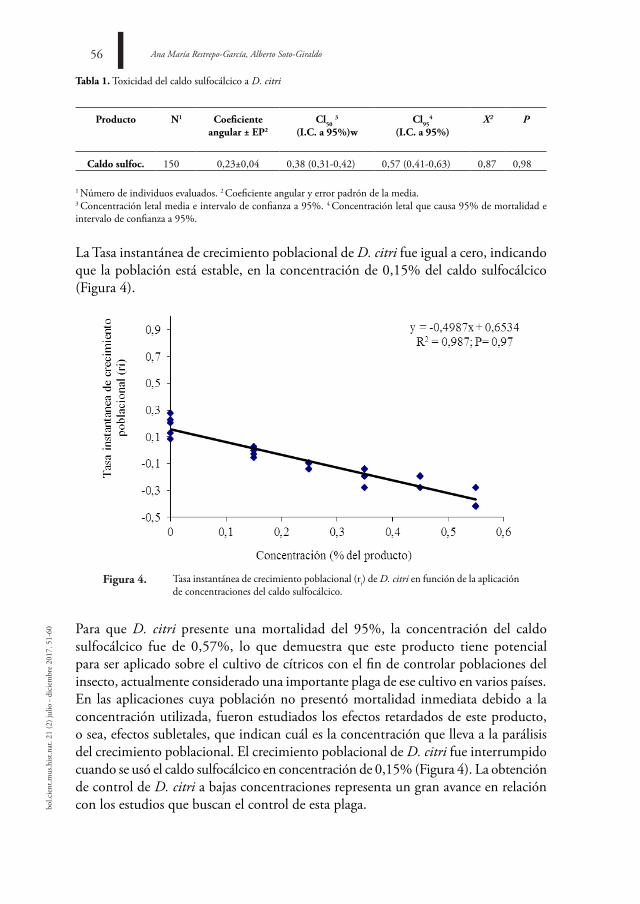
Ana María Restrepo-García, Alberto Soto-Giraldo56bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 5
1-60
tabla 1. Toxicidad del caldo sulfocálcico a D. citri
Producto n1 Coeficienteangular ± eP2
Cl50 3
(i.C. a 95%)wCl95
4
(i.C. a 95%)X2 P
Caldo sulfoc. 150 0,23±0,04 0,38 (0,31-0,42) 0,57 (0,41-0,63) 0,87 0,98
1 Número de individuos evaluados. 2 Coeficiente angular y error padrón de la media. 3 Concentración letal media e intervalo de confianza a 95%. 4 Concentración letal que causa 95% de mortalidad e intervalo de confianza a 95%.
La Tasa instantánea de crecimiento poblacional de D. citri fue igual a cero, indicando que la población está estable, en la concentración de 0,15% del caldo sulfocálcico (Figura 4).
Tasa instantánea de crecimiento poblacional (ri) de D. citri en función de la aplicación de concentraciones del caldo sulfocálcico.
figura 4.
Para que D. citri presente una mortalidad del 95%, la concentración del caldo sulfocálcico fue de 0,57%, lo que demuestra que este producto tiene potencial para ser aplicado sobre el cultivo de cítricos con el fin de controlar poblaciones del insecto, actualmente considerado una importante plaga de ese cultivo en varios países. En las aplicaciones cuya población no presentó mortalidad inmediata debido a la concentración utilizada, fueron estudiados los efectos retardados de este producto, o sea, efectos subletales, que indican cuál es la concentración que lleva a la parálisis del crecimiento poblacional. El crecimiento poblacional de D. citri fue interrumpido cuando se usó el caldo sulfocálcico en concentración de 0,15% (Figura 4). La obtención de control de D. citri a bajas concentraciones representa un gran avance en relación con los estudios que buscan el control de esta plaga.

Control alternativo de diaphorina citri kuwayama (hemiptera: liviidae) utilizando caldo sulfocálcico 57
Durante las diferentes lecturas del experimento se observó que la aplicación del producto fitoprotector interfirió en la metamorfosis del psílido (Figuras 5A y B) así como en su alimentación, lo cual ocasionó la muerte del insecto a las pocas horas (Figura 5C). Resultados similares se han obtenido con el uso de bioplaguicidas, donde los efectos de las dosis subletales de los productos en la población se ven manifestados a través de la reducción en el periodo de vida, disminución de la fertilidad, reducción de la fecundidad, trastornos en la muda, cambios en el comportamiento y fagorrepelencia (STARK et al., 1992; BANCHIO et al., 2003; MANN et al., 2011; CAZARES et al., 2014; ORTEGA et al., 2014). Esto demuestra la importancia de utilizar concentraciones subletales de los productos alternativos para el manejo de D. citri, ya que la reducción en la población de adultos que se asientan, alimentan y ovipositan en la panta, reduce la probabilidad de propagación del HLB (OUYANG et al., 2013).
Interferencia del caldo sulfocálcico en el metabolismo de D. citri. a y B: Trastornos en la muda de ninfas y adultos. C: Mortalidad de adulto a las pocas horas de emergencia. (Fotografías: A.M. Restrepo).
figura 5.
La decisión de usar concentraciones letales o subletales de los productos alternativos depende de la población plaga al momento de la aplicación. El producto caldo sulfocálcico, en dosis subletales, presenta, normalmente, selectividad para los enemigos naturales (DIMETRY et al., 1993; SCHMUTTERER, 1997; CASTIGLIONI et al., 2002; SOTO et al., 2011). Se debe tener un conocimiento técnico sobre el producto que se va a utilizar, con el fin de que se obtenga un control satisfactorio de las poblaciones de plagas, de manera que no afecte a los enemigos naturales asociados a estas (SOTO et al., 2011).
Con relación a la concentración del caldo sulfocálcico requerida para el control de D. citri, las concentraciones letales y subletales fueron menores que las recomendadas y utilizadas en sistemas de cultivos orgánicos. Estas concentraciones oscilan de 2 a
A B B

Ana María Restrepo-García, Alberto Soto-Giraldo58bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 5
1-60
4% del producto, que posee una densidad de 29 a 320 Baumé (PENTEADO, 2000; D’ANDRÉA, 2001), aproximadamente 0,58 a 1,28% de polisulfatos de calcio en el producto asperjado. Es así como este producto representa una alternativa para el control de plagas, especialmente cuando los agrotóxicos no son permitidos.
El hecho de que D. citri se presente como altamente susceptible, no implica que este producto pueda ser aplicado indiscriminadamente en concentraciones más altas que las recomendadas como resultados de experimentos científicos en campo o en invernadero, ya que se pueden presentar problemas de fitotoxicidad, además de seleccionar biotipos resistentes al producto. PEÑA et al. (2013) evaluaron sobre el cultivo de fríjol el caldo sulfocálcico para el control de Aphis gossypii (Hemiptera: Aphididae), encontrando mortalidad del 95% y 50% de la población a concentraciones del 0,64% y 0,25% del producto, respectivamente. FAJARDO et al. (2013) evaluaron el efecto de dicho producto sobre Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) en tomate, encontrado efecto letal y subletal a concentraciones del 1,13% y 0,40% del producto, respectivamente. En experimentos realizados en laboratorio e invernadero, utilizando el caldo sulfocálcico en concentración de 0,35% (30º Baumé), se comprobó la eficiencia del producto en el control del ácaro oligonychus ilicis (Acari: Tetranychidae) (TUELHER, 2006).
La necesidad de encontrar productos alternativos a los agrotóxicos convencionales con potencial preventivo y curativo contra las plagas y que no presenten efectos dañinos al medio ambiente, al consumidor y al productor, viene creciendo en todo el mundo. Por presentar esas características, el caldo sulfocálcico puede ser considerado como importante alternativa para uso en la agricultura tradicional y orgánica. La aplicación del caldo sulfocálcico en las concentraciones adecuadas representa una alternativa viable para el control de D. citri sobre plantas de cítricos, tanto cuando se aplican en las dosis letales como en las dosis subletales. En la utilización de dosis subletales es importante considerar el tiempo para la acción insecticida de esos productos.
ConClusiones
El uso del caldo sulfocálcico representa una alternativa viable a los plaguicidas sintéticos. Los resultados presentados demuestran que, a bajas concentraciones, controla a ninfas de D. citri en plantas de cítricos.
aGradeCiMientos
A la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas por la financiación de la investigación.

Control alternativo de diaphorina citri kuwayama (hemiptera: liviidae) utilizando caldo sulfocálcico 59
referenCias
ABBOT, C.E., 1945.- The toxic gases of lime-sulfur. Journal economic entomology, 38 (5): 618-620.ALEMÁN, J., BAÑOS, H. & RAVELO, J., 2007.- Diaphorina citri y la enfermedad huanglongbing: una combinación destructiva
para la producción citrícola. Revista Protección Vegetal, 22 (3): 154-165.BANCHIO, E., VALBDARES, G.D., PALACIOS, S. & CARPINELLA, C., 2003.- Effects of Melia azadarach (Meliaceae) fruit
extract on the leaf miner liriomyza huidobrensis (Diptera: Agromyzidae) assessment in laboratory and field experiments. Annual Biology, 143: 187-193.
CAMPANHOLA, C. & BETTIOL, W., 2003.- Panorama sobre o uso de agrotóxicos no Brasil: 13-51 (en) CAMPANHOLA, C. & BETTIOL, W. (eds.) Métodos alternativos de controle fitossanitário. Embrapa Meio Ambiente, Jaguariúna.
CASTIGLIONI, E., VENDRAMIM, J.D. & TAMAI, M.A., 2002.- Evaluación del efecto tóxico de extractos acuosos y derivados de meliáceas sobre Tetranychus urticae (Koch) (Acari: Tetranychidae). Agrociencia, VI (2): 75-82.
CAZARES, N., VERDE, M., LÓPEZ, J. & ALMEYDA, I., 2014.- Evaluación de diferentes extractos vegetales contra el psílido asiático de los cítricos Diaphorina citri (Hemiptera: Liviidae). Revista Colombiana de entomología, 40 (1): 67-73.
CHAGAS, P.R.R., TOKESHI, H. & ALVES, M.C., 2001.- Efficiency of lime sulfur in the control of the two-spotted mite in papaya in conventional and organic (Bokashi- EM) systems: 255-258 (en) Conference on Kyusei Nature Farming, 6. Pretoria. Proceedings Japan/ Atami: INFRC.
CHUNG, K. & BRLANSKY, R., 2005.- Citrus Diseases exotic to Florida; Huanglongbing (citrus greening). Fact Sheet PP-210. Plant Pathology Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences. University of Florida. Recuperado de http://polk.ifas.ufl.edu/hort/documents/publications/Citrus%20Greening.pdf
D’ANDREA, P.A., 2001.- Aspectos práticos e tendências no uso dos fertiprotetores: 97-101 (en) HEIN, M. (org.) Resumos do 10
encontro de Processos de Proteção de Plantas: controle ecológico de pragas e doenças. Agroecológica, Botucatu. DANE, 2016.- 3er Censo Nacional Agropecuario. GIT Área de Comunicación - Departamento Administrativo Nacional de
Estadística (DANE), Bogotá. Recuperado de https://www.dane.gov.co/files/images/foros/foro-de-entrega-de-resultados-y-cierre-3-censo-nacional-agropecuario/CNATomo2-Resultados.pdf
DIMETRY, N.Z., AMER, S.A.A. & REDA, A.S., 1993.- Biological activity of two neem seed kernel extracts against the two-spotted spider mite Tetranychus urticae Koch. Journal of Applied entomology, 116: 308-312.
EASTERBROOK, M.A., FITZGERALD, J.D. & SOLOMON, M.G., 2001.- Biological control of strawberry tarsonemid mite Phytonemus pallidus and two-spotted spider mite Tetranychus urticae on strawberry in the UK using species of Neoseiulus (Amblyseius) (Acari: Phytoseiidae). experimental and Applied Acarology, 25: 25-36.
FAJARDO, S.C., SOTO, A. & KOGSON, J.F., 2013.- Eficiencia de productos alternativos contra Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 17 (1): 91-97.
FERNÁNDEZ, M. & MIRANDA, I., 2005.- Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte I: Características morfológicas, incidencia y enemigos naturales asociados. Rev Protección Veg., 20 (1): 27-31.
FILGUEIRA, F.A.R., 2000.- Novo manual de olericultura: agrotecnologia moderna na produção e comercialização de hortaliças. UFV, Viçosa. FINNEY, D.J., 1971.- Probit analysis. Cambridge University Press, Cambridge.FRAGOSO, D.B., GUEDES, R.N.C., PICANÇO, M.C. & ZAMBOLIM, L., 2002.- Insecticide use and organophosphate resistance
in the coffee leaf miner leucoptera coffeella (Lepidoptera: Lyonetiidae). Bulletin of entomological Research, 92: 203-212.GRAFTON, E., GODFREY, K., ROGERS, M., CHILDERS, C. & STANSLY, P., 2006.- Asian Citrus Psyllid. ANR Publications
8205. Recuperado de http://anrcatalog.ucanr.edu/pdf/8205.pdf GUERRA, M.S., 1985.- Receituário caseiro: alternativa para o controle de pragas e doenças de plantas cultivadas e seus produtos. EMATER,
Brasília.GUIRADO, N., 2001.- Extrato de plantas no controle da leprose dos citros: 147-159 (en) HEIN, M. (org.) Primer encontro de
Processos de Proteção de Plantas: Controle ecológico de pragas e doenças. Agroecológica, Botucatu.HALBERT, S.E. & MANJUNATH, K.L., 2004.- Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A
literature review and assessment of risk in Florida. Florida entomologist, 87: 330-353.ICA, 2015.- Resolución N° 00002390. Por medio de la cual se declara el estado de emergencia fitosanitaria en el territorio nacional
por la presencia de adultos de Diaphorina citri infectados con la bacteria de la enfermedad del HLB de los cítricos. (pp. 1-7).ICA, 2016a.- Resolución N° 00004713. Por medio de la cual se declara en cuarentena fitosanitaria el departamento de La Guajira,
por la presencia de la plaga denominada Huanglongbing (HLB) de los cítricos. ICA, 2016b.- Resolución N° 00010508. Por medio de la cual se declara en cuarentena fitosanitaria el departamento del Atlántico,
por la presencia de la plaga denominada Huanglongbing (HLB) de los cítricos. MANN, R.S., ROUSEFF, R.L., SMOOT, J.M., CASTLE, W.S. & STELINSKI, L.L., 2011.- Sulfur volatiles from Allium spp. affect
Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), response to citrus volatiles. Bulletin of entomological Research, 101: 89–97.
MONTAG, J., SCHREIBER, L. & SCHONHERR, J., 2005.- An in vitro study on the infection activities of hydrated lime and lime sulphur against apple scab (Venturia inaequalis). Journal of Phytopathology, 153: 485-491.
OCETE, R., LÓPEZ, M.A., DANCSHÁZY, Z., OCETE, M.E., PÉREZ, M.A., KAJATI, I. & RULL, G., 2003.- IPM-sustainable tests on two apple tree pests, eriosoma lanigerum Hausm (Homoptera: Aphididae) and Aphis pomi De Geer (Homoptera: Aphididae), carried out in La Rioja (Spain). Boletín de Sanidad Vegetal, Plagas. Ministerio de Agricultura, Pesca y Alimentación. p.35-45.
ORDUZ, J. & MATEUS. D., 2012.- Generalidades de los cítricos y recomendaciones agronómicas para su cultivo en Colombia: 49-88 (en) GARCÉS, L.F. (ed.) Cítricos: cultivo, poscosecha e industrialización. Serie Lasallista Investigación y Ciencia. Corporación Universitaria Lasallista, Caldas - Ministerio de Agricultura y Desarrollo Rural (MADR) - Universidad de Antioquia, Itagüí.
ORTEGA, L., VILLEGAS, A., RAMÍREZ, A. & MENDOZA, E., 2013.- Abundancia estacional de Diaphorina citri (Hemiptera: Liviidae) en plantaciones de cítricos en cazones, Veracruz, México. Acta Zoológica Mexicana, 29 (2): 317-333.

Ana María Restrepo-García, Alberto Soto-Giraldo60bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 5
1-60
ORTEGA, L.; MENDOZA, E., LUGO, G. & ALMADA, V., 2014.- Toxicidad de extractos de “venadillo” (Swietenia humilis Zucc.) en adultos y ninfas de Diaphorina citri (Hemiptera: Liviidae). entomología Mexicana, 1: 746-749.
OUYANG, G., FANG, X., LU, H., ZHOU, X., MENG, X., YU, S., GUO, M. & XIA, Y., 2013.- Repellency of five mineral oils against Diaphorina citri (Hemiptera: Liviidae). Florida entomologist, 96 (3): 974-982.
PENTEADO, S.R., 2000.- Controle alternativo de pragas e doenças com as caldas bordalesa, sulfocálcica e Viçosa. Buena Mendes Gráfica e Editora, Campinas.
PEÑA, M.J., CASTRO, J.C. & SOTO, A., 2013.- Evaluación de insecticidas no convencionales para el control de Aphis gossypii Glover (Hemiptera: Aphididae) en fríjol. Revista U.D.C.A. Actualidad y Divulgación Científica, 16 (1): 131-138.
PICANÇO, M.C., SOTO, A., BACCI, L., FIDELIS, E.G., SILVA, G.A. & DE SENA, M.E., 2007.- Controle biológico das principais pragas de hortaliças no Brasil: 505-537 (en) ZAMBOLIM, L. (ed.) Manejo integrado de doenças e pragas hortaliças. UFV, Viçosa.
RESTREPO-GARCÍA, A.M., ARIAS-ORTEGA, P.L. & SOTO-GIRALDO, A., 2016.- Primer reporte de Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae) en el departamento de Caldas, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 20 (1): 204-210.
ROGERS, M.E., 2011.- Dice experto estadounidense: “Si el HLB llega a Colombia, debe usarse insecticida”. Frutas y Hortalizas, revista de la asociación hortofrutícola de Colombia, 20: 18-21.
SCHMUTTERER, H., 1997.- Insect growth-disrupting and fecundity reducing ingredients from the neem and chinaberry trees: 119-170 (en) MORGAN, E.D. & MANDAVA, N.B. (eds.) CRC Handbook of natural pesticides. CRC series in naturally occurring pesticides, Florida.
SMILANICK, J.L. & SORENSON, D., 2001.- Control of postharvest decay of citrus fruit with calcium polysulfide. Postharves Biology and Technology, 21: 157-168.
SOTO, A., 2009.- Control del ácaro oligonychus ilicis con calda sulfocálcica. Agronomía, 17 (1): 7-11.SOTO, A., VENZON, M. & PALLINI, A., 2011.- Integración de control biológico y de productos alternativos contra Tetranychus
urticae (Acari: Tetranychidae). Revista U.D.C.A. Actualidad y Divulgación Científica, 14 (1): 23-29.STARK, J.D. & BANKS, J.E., 2003.- Population – level effects of pesticides and other toxicants on arthropods. Annual Review of
entomology, 48: 505- 519. STARK, J.D. & RANGUS, T., 1994.- Lethal and sublethal effects of the neem insecticide, Margosan-O, on pea aphid. Journal of
Pest Science, 41: 155-160.STARK, J.D., VARGAS, R.I., MESSING, R.H. & PURCELL, M., 1992.- Effects of cyromazine and diazinon on three economically
important Hawaiian tephritid fruit flies (Diptera: Tephritidae) and their endoparasitoids (Hymenoptera: Braconidae). Journal of economic entomology, 85: 1687-1694.
TUELHER, E.S., 2006.- Toxicidade de bioprotetores da cafeicultura orgânica sobre o ácaro-vermelho do cafeeiro oligonychus ilicis e o ácaro predador Iphiseiodes zuluagai: Dissertação de Mestrado, Universidade Federal de Viçosa, Viçosa.
VENZON, M., PALLINI, A., FADINI, M.A.M., OLIVEIRA, H., MIRANDA, V.S. & DE ANDRADE, A.P.S., 2007.- Controle alternativo de ácaros em hortaliças: 607-625 (en) ZAMBOLIM, L. (ed.) Manejo integrado de doenças e pragas hortaliças. UFV, Viçosa.
WARDLOW, L.R. & LUDLAM, F.A.B., 1975.- Biological studies and chemical control of brown scale (Parthenolecanium corni (Bouché)) on Red Currant. Plant Pathology, 24: 213-216.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
enferMedades en ViVeros CoMerCiales de Musa aaB ‘doMiniCo HartÓn’ en el dePartaMento de Caldas,
ColoMBia*
Yohana Maritza Viveros Folleco1, Óscar Adrián Guzmán Piedrahita2, Bernardo Villegas estrada3
resumen
La propagación vegetativa de plátano (Musa AAB Simmonds) mediante cormos o plántulas infectadas ha sido la principal responsable de la diseminación de problemas fitopatológicos. Por ello, el objetivo de este estudio fue identificar las enfermedades presentes en cormos y plántulas de viveros comerciales de plátano (Musa AAB) ‘Dominico Hartón’ en el departa-mento de Caldas, Colombia. Los muestreos se realizaron en cinco viveros: dos ubicados en el municipio de Chinchiná, y los demás en los municipios de Risaralda, Neira y Victoria. En cada vivero se recolectó el 10% de las plántulas y se les diagnosticó las enfermedades ocasionadas por nematodos, bacterias, virus y hongos en los laboratorios de Fitopatología y Nematología de la Universidad de Caldas. En las plántulas y en los cormos, de cuatro de los cinco viveros estudiados, se extrajeron e identificaron nematodos de los géneros Radopholus, Meloidogyne y Helicotylenchus, mientras que el género Pratylenchus se encontró en un solo vivero. En los viveros de Chinchiná se encontró la bacteriosis causada por Dickeya chrysanthemi con 8% de incidencia, y el rayado del banano – Banana streak badnavirus con 20% de incidencia. En los diferentes órganos de la planta solamente se encontraron enfermedades fungosas en las hojas como las sigatokas negra y amarilla, con 28 y 11% de incidencia, respectivamente, y las manchas foliares ocasionadas por los hongos Cordana spp. y Colletotrichum spp., con incidencia menor a 5%. Los anteriores resultados demostraron que el material de siembra comercial de plátano Dominico Hartón contaminado está contribuyendo a la diseminación de estos problemas fitosanitarios, amenazando los cultivos de musáceas de esta región y del país, debido al daño que ocasionan en los tejidos de las plantas y las pérdidas en rendimiento.
Palabras clave: Musa, nematodos fitoparásitos, bacterias, virus.
* FR: 6-VI-2017. FA: 22-IX-2017.1 Candidata a Magíster en Fitopatología. Programa de Maestría en Fitopatología, Facultad de Ciencias Agropecua-rias, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0003-3495-98882 M.Sc. Profesor Asociado, Programa de Maestría en Fitopatología, Facultad de Ciencias Agropecuarias, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0001-1724-23043 M.Sc. Profesor Asociado, Programa de Maestría en Fitopatología, Facultad de Ciencias Agropecuarias, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0003-4441-5278
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 61-80. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:VIVEROS, Y.M., GUZMÁN, Ó.A. & VILLEGAS, B., 2017.- Enfermedades en viveros comerciales de Musa AAB ‘Dominico Hartón’ en el departamento de Caldas, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 61-80. DOI: 10.17151/bccm.2017.21.2.5

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada62bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
diseases in CoMMerCial nurseries of Musa aaB ‘doMiniCo HartÓn’ in tHe dePartMent of Caldas,
ColoMBia
abstract
Vegetative propagation of plantain (Musa AAB Simmonds) by infected corms or seedlings has been the main responsible for the dissemination of phytopathological problems. Therefore, the objective of this study was to identify the diseases that affect corms and seedlings in com-mercial nurseries of plantain (Musa AAB) ‘Dominico Hartón’ in the Department of Caldas, Colombia. Samplings were carried out in five nurseries, two located in the municipality of Chinchiná, and the others, in the municipalities of Risaralda, Neira and Victoria. In each nursery, 10% of the seedlings were collected and diseases caused by nematodes, bacteria, viruses and fungi were diagnosed in the Plant Pathology and Nematology laboratories of Universidad de Caldas. Nematodes of the genus Radopholus, Meloidogyne and Helicotylenchus were extracted and identified in the seedlings and corms of four of the five nurseries studied while the genus Pratylenchus was found in a single nursery. In the nurseries in Chinchiná, the incidence of bacteriose caused by Dickeya chrysanthemi was 8%, and Banana streak badnavirus had 20% incidence. In the different organs of the plant, only fungal diseases were found in leaves such as black and yellow Sigatoka, with 28% and 11% incidence respectively, and leaf spot caused by fungi Cordana spp. and Colletotrichum spp., with an incidence less than 5%. The forego-ing results demonstrated that the contaminated commercial of plantain planting material is contributing to the dissemination of these phytosanitary problems, threatening Musa crops of this region and the country due to the damage they cause in plant tissues and yield losses.
Key words: Musa, plant parasitic nematodes, bacteria, virus.
introduCCiÓn
Alrededor del mundo, las plantaciones de plátano (Musa AAB Simmonds) han sido tradicionalmente establecidas mediante material de propagación vegetativo como cormos o plántulas, los cuales han sido los principales responsables de la diseminación a gran escala principalmente de nematodos endoparásitos migratorios como Radopholus similis Cobb (Thorne) y Pratylenchus coffeae (Zimmermann) Filipjev & Schuurmans Stekhoven (LOOS, 1961; SARAH et al., 1996; MARTÍNEZ et al., 2004; GOWEN et al., 2005; GUZMÁN, 2011; CARDONA & GUZMÁN, 2013), bacterias como Ralstonia solanacearum raza 2 Smith causante del moko o maduraviche (GRANADA, 2003; GÓMEZ et al., 2005; GENIN & DENNY, 2012) y Dickeya chrysanthemi (BURKHOLDER) SAMSON, causante de la pudrición acuosa del pseudotallo (NAGARAJ et al., 2012; VAN VAERENBERGH et al., 2012), virus como el rayado del banano – Banana streak badnavirus (BSV) y el Mosaico del pepino – Cucumber

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 63
mosaic cucumovirus (CMV) (DANIELLS et al., 1995; LOCKHART & JONES, 1999; ROOSSINCK, 1999; PLOETZ, 2004; MARTÍNEZ, 2005; VILLEGAS et al., 2011) y el hongo Fusarium oxysporum f. sp. cubense (E.F. Smith) Snyder & Hansen, causante del marchitamiento por Fusarium (PLOETZ, 2006; ICA, 2012a). De igual manera, las plántulas de plátano, además de diseminar los patógenos antes mencionados, también pueden diseminar los hongos que producen la Sigatoka negra, causada por Mycosphaerella fijiensis Morelet estado anamorfo: Paracercospora fijiensis (Morelet) Deighton y la Sigatoka amarilla, causada por Mycosphaerella musicola Leach, estado anamorfo: Pseudocercospora musae (Zimm.) Deighton (FULLERTON, 1994; ICA, 2012b). Tanto las sigatokas, como las enfermedades antes mencionadas, pueden afectar el crecimiento y la producción de las especies del género Musa en todos sus estados de desarrollo, siendo indispensable el cuidado fitosanitario de los cormos o rizomas debido a que son el material de siembra más usado en viveros comerciales (SARAH et al., 1996; DICKSON & DE WAELE, 2005; COTO, 2009).
En Colombia, enfermedades como el moko y el marchitamiento por Fusarium se encuentran dentro de las enfermedades en alerta fitosanitaria y control oficial por el Instituto Colombiano Agropecuario –ICA– (resoluciones 03330 de 2013 y 2398 de 2011, respectivamente), debido a que pueden ser transmitidas por semilla asexual, siendo esta la principal forma de dispersión entre las regiones productoras de plátano y banano (ICA, 2012a; ÁLVAREZ, 2013).
Una de las herramientas preventivas, que tiene el agricultor frente a la amenaza de los anteriores problemas fitosanitarios en sus cultivos, es la adquisición de material vegetal sano, ya que este determina el éxito del cultivo en cuanto a sanidad, costos de producción, rendimientos y homogeneidad en la producción. Teniendo en cuenta la importancia del material de siembra sano en las nuevas plantaciones de musáceas, y con el propósito de conocer el estado sanitario de los cormos y las plántulas que son adquiridas por los agricultores, el objetivo de este estudio fue identificar las enfermedades presentes en el material de siembra de viveros comerciales de plátano (Musa AAB Simmonds) ‘Dominico Hartón’ del departamento de Caldas, Colombia.
Materiales Y MÉtodos
localización. Los muestreos se realizaron en cinco viveros, ubicados en cuatro municipios del departamento de Caldas, Colombia, así: dos viveros en Chinchiná (altitud 1310 m, humedad relativa 78,3% y temperatura promedio 21,1ºC), uno en Risaralda (altitud 1119 m, humedad relativa 78,7% y temperatura promedio 23,2ºC), otro en Neira (altitud 1427 m, humedad relativa 81,3% y temperatura promedio 19,5ºC) y el último en Victoria (altitud 758 m, humedad relativa 88,3% y temperatura promedio 24,3ºC), los cuales comercializan cormos y plántulas de plátano (Musa AAB) ‘Dominico Hartón’.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada64bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
tamaño y recolección de las muestras. La cantidad (número) de cormos o plántulas muestreada en el vivero de cada municipio correspondió a una muestra de 1 a 10% de la población con base en la siguiente fórmula (OSPINA, 2001):
n =
Z 2. p. q. N
N. e 2 + Z 2. p. q
Donde: n = Tamaño de la muestra, Z = 1,96 (tabla de distribución normal para el 95% de confiabilidad y 5% de error), N = Universo (cormos y plántulas), p = 0,05 (probabilidad a favor), q = 1 – p = 0,95, e = 0,05 (Error).
En los cinco viveros se recolectó el 1% de los cormos del cultivo comercial y el 10% de muestras de plántulas en condiciones de almácigo. En todos los casos, las muestras se recolectaron haciendo un recorrido en zigzag. Para el vivero 1 de Chinchiná se colectaron 36 cormos entre 200 y 250 g del cultivo en campo y 21 plántulas de almácigo en bolsas de 20 x 25 cm, mientras que en el vivero 2 de Chinchiná se tomaron 32 cormos de 400 g y 30 plántulas sembradas en bolsas de 20 x 25 cm. En el vivero de Risaralda se colectaron 28 cormos de 400 g y 33 plántulas en bolsa de 20 x 23 cm, en el vivero Neira se muestrearon 24 cormos de 300 g y 19 plántulas sembradas en bolsas de 20 x 25 cm, en el vivero de Victoria se muestrearon 22 cormos de 400 g y 41 plántulas sembradas en bolsas de 17 x 23 cm. A todos los cormos, según información de los propietarios del vivero, se les realizó desinfección con carbofuran (Furadan® 330 SC).
Cada una de las muestras fue depositada en bolsas plásticas sellables, las cuales se rotularon con: nombre del propietario, vivero, vereda, municipio y tipo de material vegetal.
descripción de principales actividades agronómicas realizadas a las plantas en condiciones de almácigo y campo en los viveros de cada municipio
1) Chinchiná 1. El cultivo de plantas madre de donde se extraen los colinos proviene de lotes comerciales de la misma finca con buena producción. Las plantas se fertilizan al momento de la siembra con gallinaza y cada 45 días con Urea (60% de N) + DAP: Fosfato diamónico (48% de P2O5), y el manejo de arvenses se realiza con machete entre las calles. Las semillas para los almácigos, bolsas de 20 x 25 cm, se desinfectan con carbofuran (Furadan® 330 SC). El suelo para el llenado de las bolsas se solariza y se mezcla con gallinaza. En el almácigo, cuando las plántulas tienen cuatro hojas, se fertilizan con Urea + DAP al igual que a los 30 días después de siembra. Posteriormente, cada 30 o 45 días se aplica triple 15 (15% de N, P y K, respectivamente) después de haber realizado un manejo manual de arvenses. Las sigatokas se manejan con deshoje y despunte sanitario.

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 65
2) Chinchiná 2. Las plantas madre de plátano Dominico Hartón con las que se inició el vivero provenían de material in vitro. En condiciones de campo, realizan tratamiento a los cormos antes de la siembra desinfectándolos con carbofuran (Furadan® 330 SC), y adicionan cal agrícola y materia orgánica. La primera fertilización se realiza con fosfato diamónico (DAP) cuando las plantas tienen cinco hojas, la segunda a los 45 días con triple 15 (N-P-K) y luego cada 30 o 45 días hasta la venta. El manejo de las sigatokas es realizado con deshojes y despuntes y aplicaciones de Propiconazol; y el manejo de arvenses con machete entre las calles, cada vez antes de aplicar el fertilizante. En almácigo se utilizan bolsas de 20 x 25 cm. El suelo es solarizado y mezclado con gallinaza al momento de la siembra; cada 30 días se aplica Urea + DAP. A los cormos, una vez arrancados, se les hace inmersión en solución de carbofuran (Furadan® 330 SC).
3) risaralda. El vivero inició con material in vitro. En condiciones de campo, realizan tratamiento de cormos con carbofuran (Furadan® 330 SC). La primera fertilización se hace con DAP cuando las plantas tienen cinco hojas, la segunda y cada 30 días con Urea + DAP + Fertibanano (18N-6P-28K-2MgO-2S); también, cada 45 días se fertiliza el retorno con triple 15 + Fertibanano. El manejo de sigatokas se realiza con Mancozeb en dosis de 1,8 L/ha, y el manejo de arvenses con machete, entre las calles, cada 45 días, y luego se aplica el fertilizante. En condiciones de almácigo, las plántulas de vivero se siembran en bolsas de 20 x 23 cm. Se realiza solarización del suelo y al momento de la siembra se mezcla con gallinaza + micorrizas, la desinfección de cormos se hace con carbofuran (Furadan® 330 SC) y cada 30 días se aplica DAP y triple 15. El manejo de arvenses se realiza manualmente, cada que se aplica el fertilizante o cuando se aplica riego.
4) neira. El origen de la semilla fue de plántulas de Dominico Hartón in vitro certificadas como material libre de enfermedades y ahora se extraen colinos de cada cepa inicial que fue sembrada en condiciones de campo, donde se realizan varias fertilizaciones: la primera al momento de la siembra con materia orgánica + cal dolomita; la segunda, cuando las plántulas tienen cuatro hojas, con Urea + DAP; la tercera a los 30 o 45 días después de la siembra con triple 15 + DAP o con triple 15 + Fertibanano. También, se realiza fertilización del retorno con materia orgánica + Biofertmex. El manejo de sigatokas es realizado con deshoje y despunte; además de aplicaciones de Tebuconazole y Pyrimethanil. El manejo de arvenses se hace con machete entre las calles, cada vez que se aplica el fertilizante. En condiciones de almácigo, se realiza solarización del suelo y al momento de la siembra se mezcla con gallinaza + micorrizas, cada 30 días se aplica DAP + triple 15, además de Beauveria spp. y Trichoderma spp. La desinfección de cormos se hace con carbofuran (Furadan® 330 SC) y yodo agrícola. El manejo de arvenses es manual cuando se aplica el fertilizante o riego.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada66bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
5) Victoria. El cultivo inició con plántulas in vitro certificadas como material libre de enfermedades y ahora obtienen colinos de cada cepa inicial. En condiciones de campo se hace tratamiento de cormos presiembra con carbofuran (Furadan® 330 SC) – Yodo Biomex Di-31 – Biofertmex. La fertilización la realizan al momento de siembra con materia orgánica: la primera, cuando las plántulas tienen cinco hojas, con Urea + DAP + Cazumag-p (15%P2O5-15%CaO-4%S-20%MgO-32,2%SiO2); la segunda a los 30 o 45 días después de la siembra con Urea + DAP + Cazumag-p (1) + Fertibanano (1). Asimismo, también fertilizan cada 30 o 45 días con triple 15 + Cazumag-p (1) + Fertibanano (1) aplicándolo al hijo de retorno con materia orgánica + Biofertmex. El manejo de las sigatokas se hace mediante deshoje y despunte; al igual que con la aplicación de Mancozeb 1,5 L/ha. También, se hace control biológico de insectos plaga con Beauveria spp. y Trichoderma spp. El manejo de arvenses se realiza con machete entre las calles, cada vez que se aplica el fertilizante. En condiciones de almácigo se realiza solarización del suelo y al momento de la siembra se mezcla con cascarilla de arroz, relación 2:1, aplicación de micorrizas y desinfección de cormos con la mezcla: Carbofuran – Yodo Biomex Di-31 – Biofertmex. Al momento de la siembra aplican gallinaza + micorrizas, y cada 30 días aplican DAP + triple 15 y los hongos controladores biológicos Beauveria spp. y Trichoderma spp., y realizan control manual de arvenses cuando se aplican fertilizantes.
Procedimientos para la identificación de los agentes causantes de enfermedades. El diagnóstico fitosanitario de las muestras recolectadas se realizó en los laboratorios de Fitopatología y Nematología del Departamento de Producción Agropecuaria de la Facultad de Ciencias Agropecuarias de la Universidad de Caldas.
a. extracción e identificación de nematodos fitoparásitos. A cada una de las plántulas recolectadas del almácigo se le lavaron las raíces con agua corriente. Luego, con la ayuda de un cuchillo, se cortaron en trozos de 1 cm de longitud y se mezclaron para luego pesar 30 g. Así mismo, a cada uno de los cormos recolectados del campo se le realizó limpieza sanitaria, que consistió en hacer cortes o pelado con un cuchillo, alrededor de los primeros 10 cm del tejido externo de ellos, y finalmente, se cortaron trozos transversales de 1 cm de longitud y se mezclaron hasta completar 30 g.
La extracción de nematodos se realizó modificando el procedimiento de JENKINS (1964) con base en el uso de tamices y el principio de flotación de ellos en azúcar. Después de la extracción, los nematodos se lavaron con abundante agua y se transfirieron 15 ml de la suspensión a una caja Petri.
La cuantificación de nematodos se realizó utilizando la metodología de FIGUEROA (1990), utilizando un estereoscopio Leica EZ4-D y una cámara de lectura. El procedimiento se repitió tres veces. Los datos obtenidos se promediaron y transformaron a cantidad (#) de nematodos en 100 g de raíces y cormos.

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 67
Para la identificación de los nematodos se transfirieron 20 individuos de la suspensión a un portaobjetos para observarlos a través del objetivo 40X, comparándolos con las claves taxonómicas de THORNE (1961), MAGGENTI et al. (1987), MAI et al. (1996), SIDDIQI (2000), PERRY & MOENS (2006), EISENBACK & HUNT (2009) y GUZMÁN (2016). Finalmente, se calcularon los promedios del número de nematodos fitoparásitos en 100 g de raíces y cormos, al igual que los géneros de estos.
B. aislamiento e identificación de bacterias fitopatógenas. Para determinar la presencia de bacterias fitopatógenas en los haces vasculares, se hicieron cortes de pseudotallo o de raíz de 1-2 cm de largo de una plántula muestreada. Posteriormente, se realizaron observaciones de flujo bacterial siguiendo la metodología de FRENCH & HEBERT (1980), en la cual el tejido se colocó suspendido en posición horizontal, en la superficie del agua de un tubo de ensayo. Este se dejó allí por 5 min para posteriormente realizar la observación de posible flujo bacteriano. Adicionalmente, a las muestras que resultaron positivas para el flujo bacterial se les realizó la tinción de Gram con base en la metodología de FRENCH & HEBERT (1980).
Finalmente, las bacterias que resultaron Gram negativas se aislaron y purificaron en medio de cultivo según la metodología de BOTERO et al. (2013), a partir de porciones de tejido con pudrición acuosa, como las calcetas, de las cuales se escogieron fracciones de 5 x 5 cm incluyendo tejido sano y tejido enfermo, las cuales se dividieron en subfracciones de 0,5 x 0,5 cm con una cuchilla de bisturí previamente desinfestada con el fuego de un mechero, para luego ser maceradas en una caja Petri que contenía 5 ml de agua estéril. Luego se obtuvo la suspensión bacterial, de la cual se tomó 1 ml y mediante diluciones se tomó una muestra de la dilución de 10-5 y se sembró en medio de cultivo Agar Nutritivo, AN, (3 g extracto de carne, 2,5 g glucosa, 15 g de agar / 1 L de agua) en condiciones estériles. También se realizaron siembras de las colonias en diferentes medios de cultivo para caracterizar las bacterias con base en su morfología y reacciones bioquímicas siguiendo el esquema de SCHAAD (1988). Finalmente, para el reconocimiento de R. solanacearum, causante del moko del plátano y banano, se utilizaron tirillas ImmunoStrip® Rs (Agdia, 2016). De la muestra se tomó una sección de aproximadamente 6,5 cm2, se introdujo en la bolsa de extracción, se siguió el protocolo (AGDIA, 2015) y por último se retiró la tira de prueba y se interpretaron los resultados.
Una vez identificadas las bacterias fitopatógenas, se determinó la incidencia aplicando así: cantidad (#) de plantas con síntomas / cantidad (#) de plantas muestreadas y multiplicado por 100.
C. identificación de virus fitopatógenos. En todos los viveros, del 10% de las plántulas muestreadas de cada uno de los almácigos, se evaluó la incidencia de enfermedades virales; se recolectaron porciones de la parte media de la hoja más joven

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada68bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
para ser procesadas y sometidas a los métodos de diagnóstico de los causantes como se describen a continuación:
· detección del virus del mosaico del pepino – Cucumber mosaic cucumovirus (CMV) mediante la técnica de immunostrip® de agdia: Se siguió el protocolo de Agdia Incorporated (http://www agdia.com), como se indicó anteriormente (AGDIA, 2015).
· detección del virus del rayado del banano – Banana streak badnavirus (BsolV) con elisa de triple anticuerpo (tas-elisa): Se utilizaron platos de poliestireno para microtitulación, con dos repeticiones para cada muestra al igual que para los controles. Se siguió el procedimiento según el fabricante (DSMZ, 2015). La lectura de los resultados se realizó con promedios de las dos repeticiones. Se asumió como parámetro de clasificación de muestras positivas, cuando el valor de la absorbancia fuera dos veces mayor que la media de los controles negativos (MATTHEWS, 1991).
d. aislamiento e identificación de hongos fitopatógenos. Este procedimiento se realizó con base en la metodología propuesta por CASTAÑO-ZAPATA & DEL RÍO (1997), de la siguiente manera: inicialmente, las muestras de hojas, raíces, cormos y pseudotallos con síntomas de enfermedad, se llevaron al estereoscopio Leica EZ4-D para observar la posible presencia de estructuras reproductivas de hongos. Cuando estaban presentes en alguno de los tejidos, se realizaron raspados superficiales y cortes con un bisturí en las zonas afectadas para remover el micelio con una aguja de disección, y posteriormente con la ayuda de lactofenol al 0,05% (20 g de fenol cristalino + 20 cm3 de ácido láctico + 20 cm3 de glicerina + 20 cm3 de agua destilada, y azul de algodón al 5% en agua) se colocaron las estructuras fúngicas en una lámina portaobjetos con una gota de lactofenol y se cubrió con un cubreobjetos para observar al microscopio de luz con el objetivo 40X. La identificación de los hongos se realizó con base en las claves taxonómicas de hongos desarrolladas por BARNETT & HUNTER (1998) y CASTAÑO-ZAPATA (2015). Posteriormente, se determinó su incidencia (%) de las enfermedades fungosas con la misma fórmula descrita anteriormente para calcular la incidencia de enfermedades bacterianas. Para determinar la(s) especie(s) de Mycosphaerella causante(s) de la(s) sigatoka(s) se utilizó la metodología de impronta descrita por AGUIRRE et al. (2003). También, se determinó el índice de severidad de la(s) sigatoka(s) con los diagramas estándares de STOVER (1971) modificados por CRAENEN (1998).

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 69
resultados Y disCusiÓn
enfermedades causadas por nematodos fitoparásitos
En las plántulas y en los cormos de cuatro de los cinco viveros estudiados, se encontraron e identificaron nematodos de los géneros Radopholus, Meloidogyne y Helicotylenchus, mientras que el género Pratylenchus se encontró en un solo vivero (Figuras 1 y 2).
Las mayores poblaciones de nematodos fueron del género Radopholus en las raíces de las plántulas de plátano Dominico Hartón de los viveros de Risaralda, Chinchiná 1 y 2 y Neira con 3.978, 2.792, 2.246 y 1.368 nematodos/100 g, respectivamente, seguidas por las de Meloidogyne con 537 y 1.251 nematodos/100 g en los viveros Chinchiná 2 y Neira. Las poblaciones del género Helicotylenchus fueron menores con valores inferiores a 612 individuos/100 g (Figura 1). El género Pratylenchus solo se encontró en el vivero Chinchiná 2 con 257 nematodos/100 g (Figura 1). En los cormos de las plántulas, al igual que en suelo, se encontraron menores cantidades de nematodos que en las raíces para los géneros antes mencionados, cuyos valores fueron menores a 988 individuos/100 g de raíces, cormo o suelo (Figura 1).
También, es de destacar la ausencia de Radopholus, Meloidogyne y Pratylenchus en el vivero del municipio de Victoria (Figura 1), lo cual evidencia el adecuado manejo sanitario del material vegetal comercializado a los agricultores de la región, ya que no se encontraron los fitonematodos más devastadores y ampliamente distribuidos en los cultivos de musáceas como Radopholus similis y Pratylenchus coffeae, los cuales, por su hábitat alimenticio de endoparásitos migratorios, se diseminan fácilmente en los cormos o semillas (GOWEN et al., 2005; GUZMÁN, 2011).
En las raíces retiradas de los cormos de plátano Dominico Hartón de los viveros ubicados en Chinchiná, Risaralda y Neira, se encontró que el género Radopholus tuvo las mayores poblaciones con valores entre 2.477 y 8.534 nematodos/100 g de raíces, seguido por los géneros Meloidogyne y Helicotylenchus con valores inferiores a 1.895 individuos/100 g de raíces (Figura 2). En el vivero Chinchiná 2 solo se encontró el género Pratylenchus con valores de 3.381 y 305 individuos/100 g de cormo y raíces, respectivamente (Figura 2). Así mismo, del tejido de los cormos pelados del plátano Dominico Hartón de los mismos viveros, se aislaron en mayor población los nematodos de los géneros Radopholus y Meloidogyne, endoparásitos migratorios y sedentarios, respectivamente (Figura 3). Estos resultados concuerdan con los síntomas primarios en las raíces y cormo tales como lesiones de color rojizo o marrón que fueron característicos de los nematodos endoparásitos migratorios (Figura 3), al igual que nudos o agallas ocasionadas por los endoparásitos sedentarios (Figura 4).

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada70bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
En este estudio, los nematodos endoparásitos migratorios presentaron la mayor población, contraria a la de los endoparásitos sedentarios y semiendoparásitos migratorios; debido a estas características alimenticias, los primeros pueden movilizarse desde las raíces hasta el cormo, conllevando a su diseminación de un lugar a otro a través de los cormos contaminados, de ahí la importancia de realizar la limpieza sanitaria de estos antes de la siembra en el sitio definitivo (ALARCÓN & CASTAÑO, 2006; GUZMÁN et al., 2012). Los cormos o rizomas pueden transportar con mayor facilidad los nematodos endoparásitos migratorios como Radopholus, Pratylenchus y Helicotylenchus; mientras que Meloidogyne, por su condición de endoparásito sedentario, es diseminado con mayor facilidad a través de las plántulas de almácigo. Aunque producir semilla de musáceas in vitro es uno de los métodos de manejo más importante para liberar material de siembra de estos fitonematodos (FIGUEROA, 1990; ÁLVAREZ et al., 2013), este método es de poco uso en Colombia, principalmente para áreas pequeñas de cultivo.
Con base en las poblaciones de nematodos encontradas en los viveros de plátano y banano del departamento de Caldas, los géneros de fitonematodos más comunes fueron: Radopholus, Meloidogyne, Helicotylenchus y Pratylenchus. Resultados que coinciden con los obtenidos por ARAYA (2004) en estudios realizados en cultivos de musáceas en Centroamérica, donde estos parásitos son los más comunes.
El nivel poblacional de los nematodos que causan daño económico en cultivos establecidos de musáceas es muy variable debido al material vegetal, las condiciones edafoclimáticas y a otros factores, pero se ha evaluado bajo una población dada, por ejemplo, Radopholus con 10.000 nematodos/100 g de raíces (TARTÉ & PINOCHET, 1981), Meloidogyne con 5.000 nematodos/100 g de raíces (DAVIDE & MARASIGAN, 1985), Pratylenchus con 8.000 nematodos/100 g de raíces (Rodríguez et al., 2001) y Helicotylenchus con 50.000 nematodos/100 g de raíces (SIKORA & SCHLOSSER, 1973). Con el parámetro general de 10.000 Radopholus/100 g de raíces se toman decisiones para su manejo en las plantaciones de banano en América Central y Ecuador (CHÁVEZ-VELAZCO et al., 2009). Este no debería considerarse como parámetro de manejo, por lo que se hace necesario estudiar los niveles de daño económico en contextos regionales (PINOCHET, 1988). En el caso de viveros, el nivel de nematodos fitoparásitos debe ser de cero.
En la zona de estudio se evidenció el desconocimiento generalizado de los nematodos fitoparásitos por parte de los propietarios de los viveros. Esta situación, ha contribuido a la diseminación generalizada de los nematodos fitoparásitos en toda la región donde se realizó el estudio, haciendo la excepción del vivero del municipio de Victoria donde no se encontraron los fitonematodos más importantes en este cultivo. Esto se debió a que el propietario estableció el cultivo del plátano Dominico Hartón en un lote donde no se habían sembrado musáceas anteriormente y a que el material de siembra
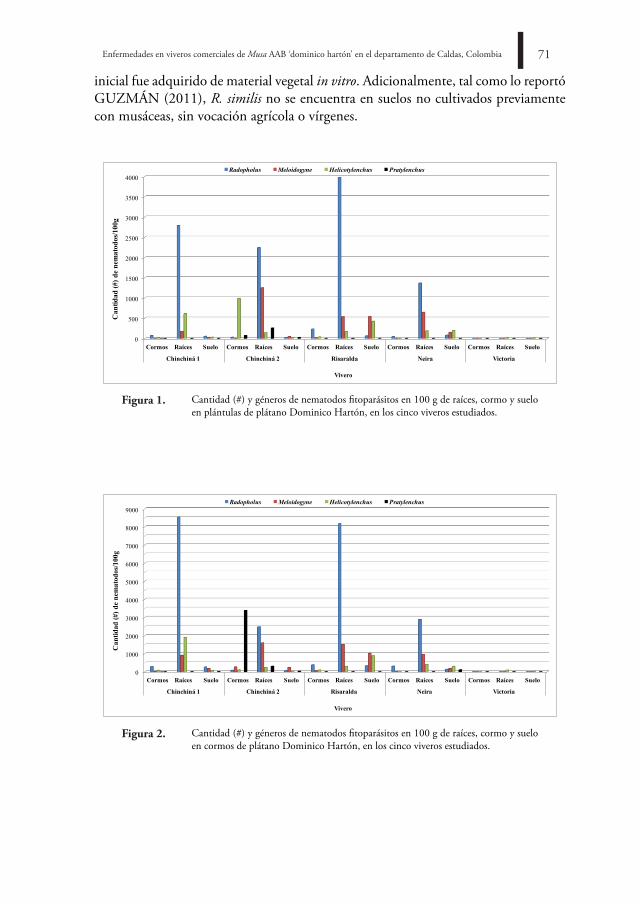
Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 71
inicial fue adquirido de material vegetal in vitro. Adicionalmente, tal como lo reportó GUZMÁN (2011), R. similis no se encuentra en suelos no cultivados previamente con musáceas, sin vocación agrícola o vírgenes.
0
500
1000
1500
2000
2500
3000
3500
4000
Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo
Chinchiná 1 Chinchiná 2 Risaralda Neira Victoria
Can
tidad
(#) d
e ne
mat
odos
/100
g
Vivero
Radopholus Meloidogyne Helicotylenchus Pratylenchus
Cantidad (#) y géneros de nematodos fitoparásitos en 100 g de raíces, cormo y suelo en plántulas de plátano Dominico Hartón, en los cinco viveros estudiados.
figura 1.
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo Cormos Raíces Suelo
Chinchiná 1 Chinchiná 2 Risaralda Neira Victoria
Can
tidad
(#) d
e ne
mat
odos
/100
g
Vivero
Radopholus Meloidogyne Helicotylenchus Pratylenchus
Cantidad (#) y géneros de nematodos fitoparásitos en 100 g de raíces, cormo y suelo en cormos de plátano Dominico Hartón, en los cinco viveros estudiados.
figura 2.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada72bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
Raíces y cormos de plátano Dominico Hartón parasitados por nematodos fitoparásitos. a. Raíces con lesiones de color marrón o rojizas ocasionadas por Radopholus similis. B y C. Cormo y fragmento de cormo con lesiones de color rojizo y necróticas ocasionadas por R. similis. d. Raíces y fragmento de raíces con nudos o agallas producidas por Meloidogyne spp.
figura 3.
Nematodos fitoparásitos encontrados en los cormos y plántulas de plátano Dominico Hartón. a. Hembra adulta de Radopholus similis en el objetivo 40X. B. Hembra adulta de Helicotylechus spp. en 40X. C. Estado juvenil (J2) de Meloidogyne spp. en 40X. d. Hembras adultas de Pratylenchus spp. en 40X. e. Hembra adulta de Aphelenchoides spp. en 10X. f. Hembra adulta de Tylenchus spp. en 10X.
figura 4.

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 73
enfermedades causadas por bacterias fitopatógenas
En los viveros Chinchiná 1 y Chinchiná 2 se encontraron plántulas de plátano Dominico Hartón con pudrición acuosa del pseudotallo, con olor fétido y presencia de exudado bacteriano, principalmente en las calcetas y cormos. En algunas plántulas, con estados avanzados de la enfermedad, el patógeno ocasionó necrosis del pseudotallo. De esta pudrición se obtuvieron bacterias con forma de bacilo, Gram (-) y según las pruebas bioquímicas se identificó como Dickeya chrysanthemi, con incidencia del 8 y 4%, respectivamente (Figura 5).
Procedimiento de identificación de las bacterias. a. Fragmento de calceta en tubo de ensayo con agua destilada estéril emitiendo flujo bacteriano. B. Células bacterianas Gram negativas. C. Filancia de bacterias en prueba KOH. d. Crecimiento de la bacteria Dickeya chrysanthemi en Agar Nutritivo.
figura 5.
Según ARANZAZU et al. (2002), un factor predisponente de esta enfermedad es el desbalance nutricional de las plantas, especialmente por deficiencias de boro y potasio; además, entre los factores que aumentan la severidad de la enfermedad están los períodos de sequía, alternados con fuertes lluvias. FERNÁNDEZ & LÓPEZ (1970) consideran que una de las causas de su diseminación en el cultivo es la falta de selección de semilla y de desinfestación de herramientas, siendo el agricultor el principal diseminador de la bacteria al no realizar las prácticas antes mencionadas. Otros factores predisponentes para la enfermedad son los destronques inmediatos al pseudotallo después de la cosecha y la carencia de semilla de buena calidad. Adicionalmente, BELALCÁZAR (1991) afirmaron que la alta incidencia de insectos, como Metamasius hemipterus sericeus, incrementa la dispersión de la bacteria en campo.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada74bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
En el vivero Chinchiná 1 también se encontraron plántulas de plátano Dominico Hartón con síntomas de clorosis y marchitamiento de hojas, síntomas característicos del moko o maduraviche ocasionado por Ralstonia solanacearum raza 2. A los cormos y pseudotallos de las plántulas se les realizaron cortes transversales y longitudinales sin observar síntomas de necrosis u obstrucciones de los haces vasculares, y tampoco exudados bacterianos (GRANADA, 2003; GÓMEZ et al., 2005). Luego de utilizar el Kit rápido de reconocimiento de R. solanacearum en el tejido vegetal con ImmunoStrip-Rs® de Agdia® (AGDIA, 2016), se descartó la presencia de esta bacteria. Los síntomas de clorosis y marchitamientos pudieron ser ocasionados por deficiencias nutricionales o estrés hídrico debido al fenómeno de El Niño que se presentó en Colombia durante el estudio.
En los demás viveros, después de realizar la prueba antes mencionada en plántulas de plátano y banano, no se detectó presencia de la bacteria. La ausencia de R. solanacearum raza 2 es un aspecto positivo de los cinco viveros estudiados.
enfermedades causadas por virus
Después de usar ImmunoStrip® (AGDIA, 2015) para detección de CMV en plántulas de plátano Dominico Hartón con síntomas como mosaicos, clorosis intervenal, detención del crecimiento y necrosamiento de la hoja cigarro, el resultado fue negativo, por lo tanto, estos síntomas no están asociados a esta enfermedad en ninguno de los cinco viveros evaluados. Por lo tanto, estos síntomas pueden relacionarse con otros factores como estrés hídrico, fitotoxicidad por algún plaguicida aplicado o efecto de la presencia de nematodos fitoparásitos en las raíces.
Después de aplicar la técnica TAS-ELISA, el BSOLV fue detectado en las plántulas de plátano Dominico Hartón del vivero Chinchiná 1 con una incidencia de 20% (Tabla 1). A pesar de que las plántulas recolectadas en este vivero no tenían retraso en el crecimiento, en ellas se observaron síntomas como rayado clorótico en los espacios intervenales (Figura 6A), deformación y arrugamiento con presencia de rayas cloróticas paralelas a la nervadura central de las hojas (Figura 6B).
Los síntomas antes descritos, coinciden con los reportados por VILLEGAS et al. (2011) y LÓPEZ-CARDONA et al. (2014) en plantas de Dominico Hartón infectadas con BSOLV, que presentan síntomas de rayado en hojas, líneas paralelas a la nervadura central, distorsión de la filotaxia de la hoja, mostrando el síntoma típico de hojas en forma de abanico. Así mismo, según MARTÍNEZ (2005), en plantas con menos de dos meses de edad se observó desarrollo rápido de los síntomas virales en las hojas número 4 y 5.
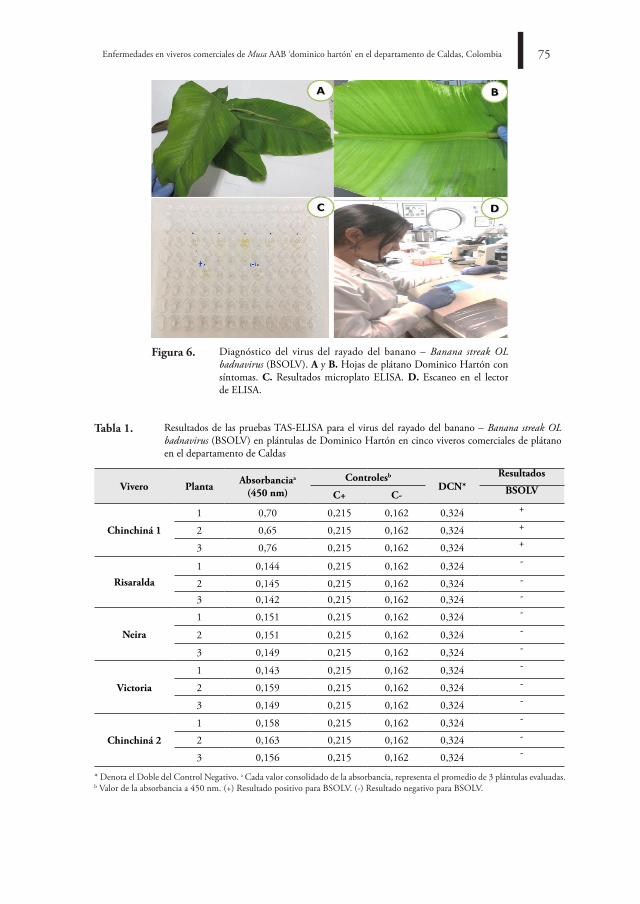
Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 75
Diagnóstico del virus del rayado del banano – Banana streak ol badnavirus (BSOLV). a y B. Hojas de plátano Dominico Hartón con síntomas. C. Resultados microplato ELISA. d. Escaneo en el lector de ELISA.
figura 6.
Resultados de las pruebas TAS-ELISA para el virus del rayado del banano – Banana streak ol badnavirus (BSOLV) en plántulas de Dominico Hartón en cinco viveros comerciales de plátano en el departamento de Caldas
tabla 1.
Vivero Planta absorbanciaa
(450 nm)Controlesb
dCn*resultados
C+ C- BsolV
Chinchiná 1
1 0,70 0,215 0,162 0,324 +
2 0,65 0,215 0,162 0,324 +
3 0,76 0,215 0,162 0,324 +
risaralda
1 0,144 0,215 0,162 0,324 -
2 0,145 0,215 0,162 0,324 -
3 0,142 0,215 0,162 0,324 -
neira
1 0,151 0,215 0,162 0,324 -
2 0,151 0,215 0,162 0,324 -
3 0,149 0,215 0,162 0,324 -
Victoria
1 0,143 0,215 0,162 0,324 -
2 0,159 0,215 0,162 0,324 -
3 0,149 0,215 0,162 0,324 -
Chinchiná 2
1 0,158 0,215 0,162 0,324 -
2 0,163 0,215 0,162 0,324 -
3 0,156 0,215 0,162 0,324 -
* Denota el Doble del Control Negativo. a Cada valor consolidado de la absorbancia, representa el promedio de 3 plántulas evaluadas. b Valor de la absorbancia a 450 nm. (+) Resultado positivo para BSOLV. (-) Resultado negativo para BSOLV.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada76bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
enfermedades causadas por hongos fitopatógenos
De todos los tejidos analizados (raíces, cormos, pseudotallos y hojas) solamente se encontraron enfermedades fungosas en las hojas de las plántulas de plátano Dominico Hartón. Las enfermedades más comunes fueron las sigatokas negra y amarilla, con una incidencia del 28 y 11%, respectivamente, en el vivero Chinchiná 1, en comparación con las plántulas de los demás viveros cuya incidencia estuvo entre 0 y 8% (Tabla 2). Las especies de Mycosphaerella que ocasionan las sigatokas fueron corroboradas con la prueba de la impronta descrita por AGUIRRE et al. (2003). En ambos viveros de Chinchiná, la severidad de las sigatokas negra y amarilla fue del 50 y 33%, respectivamente (datos no mostrados).
Los anteriores resultados demostraron que los viveros ubicados en el municipio de Chinchiná presentaron la mayor incidencia de las sigatokas por contar con condiciones climáticas favorables para el desarrollo de la enfermedad. La Sigatoka negra se puede presentar y establecer donde se cultiven plátano y banano, especialmente destructiva sobre variedades susceptibles en regiones cálidas y húmedas localizadas por debajo de los 550 msnm y con un promedio de temperatura entre 25 y 28°C (MERCHÁN, 1998).
Los síntomas de la Sigatoka negra varían en función del estado de desarrollo de la planta, la susceptibilidad del hospedante y la severidad del ataque. Según MERCHÁN (1998), en las plántulas de plátano las lesiones son anormalmente grandes, ovales a circulares, lo cual concuerda con lo encontrado en plántulas de Dominico Hartón de los dos viveros de Chinchiná, donde la enfermedad se reconoció por la gran cantidad de rayas o estrías y manchas definidas de color café a negro que cubrían el área foliar desde la tercera hoja más joven hacia abajo (ARANZAZU et al., 2002), lo cual fue corroborado con la prueba de la impronta descrita anteriormente por AGUIRRE et al. (2003).
La Sigatoka amarilla presentó mayor incidencia en las plántulas de los viveros Chinchiná 1 y Chinchiná 2, ubicados por encima de los 1200 msnm, lo que coincide con BELALCÁZAR (1991), quienes encontraron una relación estrecha del cultivo con la enfermedad por encima de esta altitud. En ausencia de prácticas de control, se estima que las hojas tan pronto como emergen son infectadas por ascosporas en temporada lluviosa y por conidios en época seca, situación que se presenta en estos viveros donde hay deficiencias de prácticas preventivas para el manejo de la enfermedad; esto ligado a las condiciones ambientales propicias que generan ciclos permanentes de la enfermedad, ya que el periodo de incubación de estos hongos es de aproximadamente 29 días (BELALCÁZAR et al., 2000).
Otra enfermedad que se diagnosticó en las hojas de las plántulas de plátano Dominico Hartón fue la mancha Cordana (Figura 7), con una incidencia menor a 5% en los viveros Chinchiná y Victoria, mientras que en los demás viveros no se presentó (Tabla 2).
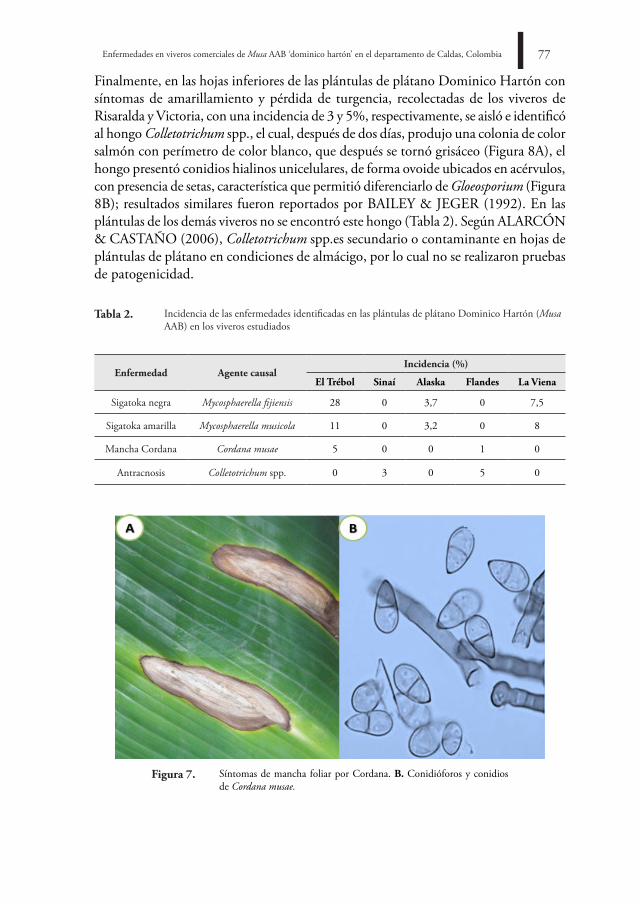
Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 77
Finalmente, en las hojas inferiores de las plántulas de plátano Dominico Hartón con síntomas de amarillamiento y pérdida de turgencia, recolectadas de los viveros de Risaralda y Victoria, con una incidencia de 3 y 5%, respectivamente, se aisló e identificó al hongo Colletotrichum spp., el cual, después de dos días, produjo una colonia de color salmón con perímetro de color blanco, que después se tornó grisáceo (Figura 8A), el hongo presentó conidios hialinos unicelulares, de forma ovoide ubicados en acérvulos, con presencia de setas, característica que permitió diferenciarlo de Gloeosporium (Figura 8B); resultados similares fueron reportados por BAILEY & JEGER (1992). En las plántulas de los demás viveros no se encontró este hongo (Tabla 2). Según ALARCÓN & CASTAÑO (2006), Colletotrichum spp.es secundario o contaminante en hojas de plántulas de plátano en condiciones de almácigo, por lo cual no se realizaron pruebas de patogenicidad.
Incidencia de las enfermedades identificadas en las plántulas de plátano Dominico Hartón (Musa AAB) en los viveros estudiados
tabla 2.
enfermedad agente causalincidencia (%)
el trébol sinaí alaska flandes la Viena
Sigatoka negra Mycosphaerella fijiensis 28 0 3,7 0 7,5
Sigatoka amarilla Mycosphaerella musicola 11 0 3,2 0 8
Mancha Cordana Cordana musae 5 0 0 1 0
Antracnosis Colletotrichum spp. 0 3 0 5 0
Síntomas de mancha foliar por Cordana. B. Conidióforos y conidios de Cordana musae.
figura 7.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada78bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
a. Colletotrichum sp. en el medio de cultivo PDA. B. Conidióforos con conidios hialinos ovoides teñidos con el colorante azul de lactofenol y presencia de setas.
figura 8.
ConClusiones
En las plántulas y cormos de plátano Dominico Hartón y banano Gros Michel de los viveros evaluados, las enfermedades más frecuentes fueron las ocasionadas por nematodos fitoparásitos de los géneros Radopholus, Pratylenchus, Meloidogyne y Helicotylenchus, y las sigatokas negra y amarilla. Con menor incidencia se encontraron la pudrición acuosa del pseudotallo, mancha Cordana, el virus OL del rayado del banano, marchitamientos y antracnosis, resultados que demuestran cómo el material de siembra comercial está contribuyendo a la diseminación de estos problemas fitosanitarios.
Los nematodos fitoparásitos, especialmente los endoparásitos migratorios como Radopholus similis y Pratylenchus spp., están siendo diseminados por los agricultores debido a que se encontraron en el 80% del material de siembra de los viveros del departamento de Caldas, Colombia, amenazando los cultivos de musáceas de esta región y del país debido al daño que ocasionan en las raíces y las pérdidas en rendimiento.
El rendimiento de los cultivos de plátano y banano del departamento de Caldas, Colombia, está siendo amenazado por la comercialización de material de siembra contaminado en los viveros comerciales; debido posiblemente al desconocimiento de estas enfermedades o a su mal manejo, conllevando a la diseminación de problemas fitosanitarios y al empobrecimiento de los agricultores. Por lo tanto, es necesario continuar realizando estudios regionales sobre el estado fitosanitario de las especies

Enfermedades en viveros comerciales de Musa AAB ‘dominico hartón’ en el departamento de Caldas, Colombia 79
vegetales que se comercializan en los viveros para poder predecir daños y determinar cuándo se debe aplicar una medida de manejo.
aGradeCiMientos
A la Universidad de Caldas, y al Fondo de Ciencia, Tecnología e Innovación del Sistema General de Regalías por la financiación de esta investigación en el marco del proyecto “Implementación de estrategias para el fortalecimiento tecnológico de la actividad agrícola y de transformación del plátano en Caldas”.
referenCias
AGDIA, 2015.- Pathogen Tests - ImmunoStrip Tests® for Cucumber mosaic virus (CMV).- www.agdia.comAGDIA, 2016.- Pathogen Tests – ImmunoStrip Tests® for Ralstonia solancearum (Rs). www.agdia.comAGUIRRE, M., CASTAÑO-ZAPATA, J. & ZULUAGA, L., 2003.- Método rápido de diagnóstico de Mycosphaerella musicola
Leach y M. fijiensis Morelet, agentes causantes de las Sigatokas amarilla y negra. Rev. Acad. Colomb. Cienc., 27 (105): 619-623.ALARCÓN, R. & CASTAÑO, J., 2006.- Reconocimiento fitosanitario de las principales enfermedades del plátano Dominico Hartón
(Musa spp., Simmonds). Agronomía, 14 (1): 65-80.ÁLVAREZ, A.M., 2013.- Diversity and diagnosis of Ralstonia solanacearum: 437-447 (en) ALLEN, C., PRIOR, P. & HAYWARD,
A.C. (eds.) Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex. APS Press, St. Paul, MN, USA.ÁLVAREZ, E., CEBALLOS, G., GAÑÁN, L., RODRÍGUEZ, D., GONZÁLEZ, S. & PANTOJA, A., 2013.- Producción de material
de ‘siembra’ limpio en el manejo de las enfermedades limitantes del plátano. Centro Internacional de Agricultura Tropical (CIAT). Publicación CIAT No. 384.
ARANZAZU, F., VALENCIA, J., ARCILA, M., CASTRILLÓN, C., BOLAÑOS, M., CASTELLANOS, P., PÉREZ, J. & RODRÍGUEZ, J., 2002.- el cultivo de plátano: manual técnico. Corpoica, Manizales.
ARAYA, M., 2004.- Los fitonematodos del banano (Musa AAA Subgrupo Cavendish cultivares Grande Naine, Valery y Williams) su parasitismo y combate: 84-105 (en) XVI Reunión Internacional Acorbat. Publicación especial. Oaxaca, México.
BAILEY, J. & JEGER, M., 1992.- Colletotrichum. Biology, pathology and control. CAB International, Wallingford, UK.BARNETT, H.L., & HUNTER, B., 1998.- Illustrated Genera of Imperfect Fungi. 4th ed. APS Press, St. Paul, Minn. BELALCÁZAR, S., 1991.- El cultivo del plátano (Musa AAB Simmonds) en el trópico. Manual de asistencia técnica No. 50. Armenia,
Colombia.BELALCÁZAR, S., MERCHÁN, V., MAYORGA, M., LONDOÑO, M., PULIDO, J., GARCÍA, F., DE POLANÍA, I., LEÓN,
G. & VALENCIA, J., 2000.- Plagas y enfermedades del plátano. Boletín de Sanidad Vegetal. Bogotá.BOTERO, O., CASTAÑO-ZAPATA, J., SALDARRIAGA, C. & CASTRO, T., 2013.- Manual práctico de bacteriología vegetal.
Primera edición. Universidad de Caldas. Manizales. CARDONA, L. & GUZMÁN, O., 2013.- Mecanismos de diseminación de nematodos fitoparásitos en plátano (Musa acuminata
Simmonds) grupo AAB, cultivariedad Dominico Hartón. Agron., 21 (1): 26-36.CASTAÑO-ZAPATA, J., 2015.- Principios básicos de hongos fitopatógenos. Editorial Universidad de Caldas, Manizales.CASTAÑO-ZAPATA, J. & DEL RÍO, M. L., 1997.- Manual para el diagnóstico de hongos, bacterias, virus y nematodos fitopatógenos.
Zamorano Academic Press, Honduras.CHÁVEZ-VELAZCO, C., SOLÓRZANO-FIGUEROA, F. & ARAYA-VARGAS, M., 2009.- Relación entre nematodos y la
productividad del banano (Musa AAA) en Ecuador. Agronomía Mesoamericana, 20 (2): 351-360.COTO, J., 2009.- Guía para multiplicación rápida de cormos de plátano y banano. Centro de Comunicación Agrícola de la Fundación
Hondureña de Investigación Agrícola (FHIA), La Lima, Cortés, Honduras, C.A.CRAENEN, K., 1998.- Technical manual on black sigatoka disease of banana and plantain. International Institute of Tropical Agriculture,
Ibadan-Nigeria.DANIELLS, J., THOMAS, J.E. & SMITH, M., 1995.- Seed transmission of banana streak virus confirmed. Infomusa, 4 (1): 7.DAVIDE, R.G. & MARASIGAN, L.Q., 1985.- Yield loss assessment and evaluation of resistance of banana cultivars to the nematodes
Radopholus similis Thorne and Meloidogyne incognita Chitwood. Phil Agri., 68: 335-349.DICKSON, D.W. & DE WAELE, D., 2005.- Nematode parasites of peanut (en) LUC, M., SIKORA, R. & BRIDGE, J. (eds.) Plant
Parasitic Nematodes in Subtropical and Tropical Agriculture. 2nd Edition. CAB International, London.DSMZ-GERMAN COLLECTION OF MICROORGANISMS AND CELL CULTURES, 2015.- https://www.dsmz.de/catalogues/
catalogue-plant-viruses-and-antisera.htmlEISENBACK, J.D. & HUNT, D.J., 2009.- Chapter 16: General morphology: 18-54 (en) PERRY, R.N., MOENS, M. & STARR,
J.L. (eds.) Root-knot Nematodes. CABI Publishing, Wallingford, UK.FERNÁNDEZ, B. & LÓPEZ, S., 1970.- Pudrición acuosa del pseudotallo del plátano (Musa paradisica) causada por erwinia
paradisiaca, n. sp. Cenicafé, 21 (1): 1-44.FIGUEROA, M., 1990.- Dinámicas poblacionales de cuatro géneros de nematodos parásitos en plátano (Musa AAB, subgrupo plátano
Cv currare). ASBANA, 14 (33): 5-7.

Yohana Maritza Viveros Folleco, Óscar Adrián Guzmán Piedrahita, Bernardo Villegas estrada80bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 6
1-80
FRENCH, E.R. & HEBERT, T., 1980.- Métodos de investigación fitopatológica. Instituto Interamericano de Ciencias Agrícolas, Costa Rica.
FULLERTONH, R. A., 1994.- Sigatoka leaf diseases. In: Compendium of Tropical Fruit Diseases. Ploetz, R.C. et al. (Editors). The American Phytopathological Society, St. Paul, Minnesota. Pp. 12-14.
GENIN, S. & DENNY, T., 2012.- Pathogenomics of the Ralstonia solanacearum species complex. Annual Review of Phytopathology, 50: 67-89.
GÓMEZ, G., ÁLVAREZ, E. & LLANO, E., 2005.- Identificación y caracterización del agente causal del Moko de plátano Ralstonia solanacearum raza 2 provenientes de plantaciones afectadas en Colombia. Fitopatología Colombiana, 28 (2): 71-75.
GOWEN, S., QUÉNÉHERVÉ, P. & FOGAIN, R., 2005.- Chapter 16: Nematodes parasites of bananas and plantains: 611-643 (en) LUC, M., SIKORA, R. & BRIDGE, J. (eds.) Plant Parasitic Nematodes in Subtropical and Tropical Agriculture. 2nd Edition. CAB International, London.
GRANADA, G.A., 2003.- Manejo integrado del Moko (Ralstonia solanacearum Raza 2) en cultivos de banano y plátano. Centro de Investigación del Banano-CENIBANANO. Boletín Técnico No. 2: 6-12.
GUZMÁN, O.A., 2011.- El nematodo barrenador (Radopholus similis [Cobb] Thorne) del banano y plátano. Revista luna Azul, 32: 137-153.
GUZMÁN, O.A., 2016.- Manual para la identificación de nematodos fitoparásitos. Primera edición. Manizales, Caldas. 134p.GUZMÁN, O.A., CASTAÑO, J. & VILLEGAS, B., 2012.- Efectividad de la sanidad de cormos de plátano Dominico Hartón
(Musa AAB Simmonds), sobre nematodos fitoparásitos y rendimiento del cultivo. Rev. Acad. Colomb. Cienc., 36 (138): 45-55.ICA (INSTITUTO COLOMBIANO AGROPECUARIO), 2012a.- Manejo fitosanitario del cultivo del plátano (Musa spp.) - Medidas
para la temporada invernal. Produmedios, Bogotá.ICA (INSTITUTO COLOMBIANO AGROPECUARIO), 2012b-. Manejo de las principales plagas y enfermedades del cultivo del
plátano. Produmedios, Bogotá. JENKINS, W.R., 1964-. A rapid centrifugal flotation technique for separating nematodes from soil. Plant Disease Reporter, 48 (9): 692.LOCKHART, B.E.L. & JONES, D.R., 1999.- Banana streak virus: 263-274 (en) JONES, D.R. (ed.) Diseases of Banana, Abacá and
enset. CAB Publishing, Wallingford, UK.LOOS, A.C., 1961.- Eradication of the burrowing nematode, Radopholus similis from bananas. Plant Disease Reporter, 45 (6): 457-461.LÓPEZ-CARDONA, N., VILLEGAS-ESTRADA, B. & ARANGO-ISAZA, R., 2014.- Evaluación de incidencia y pérdidas
ocasionadas por virus que afectan cultivos de plátano y banano (Musa spp.) en la zona central cafetera. Agronomía, 22 (1): 22-35.MAGGENTI, A., LUC, M., RASKI, D., FORTUNER, R. & GERAERT, E., 1987.- A reappraisal of Tylenchina (Nemata). 2.
Classification of the suborder Tylenchina (Nemata: Diplogasteria). Revue Nématol, 10 (2): 135-142.MAI, W.F., MULLIN, P.G. & LYON, H.H., 1996.- Plant Parasitic Nematodes: A Pictorial Key to Genera. Fifth Edition. Cornell
University Press, Ithaca, NY.MARTÍNEZ, G., 2005.- Los virus del plátano en la zona cafetera: su influencia en la viabilidad económica del cultivo: 214-219 (en)
Memorias II Seminario Internacional Sobre Producción, Comercialización e Industrialización de Plátano. Manizales, Colombia.MARTÍNEZ, G., TREMONT, O. & HERNÁNDEZ, J., 2004.- Manual técnico para la propagación de Musáceas. Revista CeNIAP
HoY, 4.MATTHEWS, R.E.F., 1991.- Plant Virology. 3 ed. Academic press, New York.MERCHÁN, V.M., 1998.- Manejo de problemas fitosanitarios del cultivo del plátano en la zona central cafetera: 177-192 (en)
Seminario Internacional sobre Producción de Plátano. Armenia, Quindío.NAGARAJ, M.S., UMASHANKAR, N., PALANNA, K.B. & KHAN, A., 2012.- Etiology and management of Tip-over disease of
banana by using biological agents. International Journal of Advanced Biological Research, 2 (3): 483-486.OSPINA, D., 2001.- Introducción al muestreo. Universidad Nacional de Colombia – Unilibros, Bogotá.PERRY, R. & MOENS, M., 2006.- Plant nematology. London: CAB International. 447p.PINOCHET, J., 1988.- Manejo de nematodos en viveros forestales: 66-72 (en) Memorias Seminario de Manejo Integrado de Plagas
(1986, San José, C.R.). PLOETZ, R., 2004.- Enfermedades y plagas: Un análisis de su importancia y manejo. InfoMusa, 13 (2): 11-16.PLOETZ, R.C., 2006.- Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense.
Phytopathology, 96: 653-656. ROOSSINCK, M.J., 1999.- Cucumoviruses (Bromiviridae)-general features: 315-320 (en) GRANOFF, A. & WEBSTER, R.G. (eds.)
encyclopedia of virology. 2nd ed. Academic Press, San Diego.SARAH, J.L., PINOCHET, J. & STANTON, J., 1996.- El nematodo Barrenador del Banano Radopholus Similis Cobb. Plagas de
Musa - Hoja Divulgativa No. 1.SCHAAD, R., 1988.- Laboratory guide for identification of plant pathogenic bacteria. The American Phytopathological Society
(APS), Minnesota. 253 p.SIDDIQI, M.R., 2000.- Tylenchida: Parasites of Plants and Insects. 2nd Edition. CABI Bookshop, Wallingford, UK.SIKORA, R.A. & SCHLOSSER, E., 1973.- Nematodes and fungi associated with root systems of bananas in a state of decline in
Lebanon. Plant Disease Reporter, 57: 615-618.STOVER, R.H., 1971.- A proposed international scale for estimating intensity of banana leaf spot. Tropical Agriculture, 48: 185-196.TARTÉ, R. & PINOCHET, J., 1981.- Problemas nematológicos del banano: contribuciones recientes a su conocimiento y combate.
INIBAP, Panamá.THORNE, G., 1961.- Principles of Nematology. McGraw-Hill - Book Company, New York.VAN VAERENBERGH, J., BAEYEN, S., DE VOS, P. & MAES, M., 2012.- Sequence diversity in the Dickeya fliC gene: Phylogeny
of the Dickeya genus and TaqMan® PCR for ‘D. solani’, New Biovar 3 variant on potato in Europe. PloS oNe, 7 (5): e35738. http://dx.doi.org/10.1371/journal.pone.0035738.
VILLEGAS, B., CAÑAS, G.P. & LÓPEZ N., 2011.- enfermedades virales de los cultivos de plátano y banano en el eje Cafetero, diagnóstico y manejo. Universidad de Caldas - Editorial Tizán, Manizales.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
diVersidad de las MosCas de las frutas (diPtera: tePHritidae) Y sus Parasitoides en siete MuniCiPios
del dePartaMento de nariÑo*
María Isabel Cruz B.1, Tito Bacca2, Nelson A. Canal 3
resumen
El objetivo de este trabajo fue contribuir al conocimiento de la diversidad de moscas de las frutas (Diptera: Tephritidae) y de sus enemigos naturales en el departamento de Nariño. Se colectaron 14.005 frutos correspondientes a 16 especies de plantas hospederas, pertenecientes a 10 familias botánicas (Rubiaceae, Rosaceae, Myrtaceae, Fabaceae, Anacardiaceae, Rutaceae, Cucurbitaceae, Annonaceae, Solanaceae y Caricaceae), de las cuales todas presentaron infestación por moscas de las frutas o parasitoides, a excepción de la familia Annonaceae. Las especies de moscas de las frutas recuperadas en este estudio correspondieron a Anastrepha complejo fraterculus, Anastrepha striata, Anastrepha obliqua, Anastrepha grandis y Ceratitis capitata, especies conocidas por su polifagia y gran capacidad de adaptación. Se obtuvieron 242 individuos, pertenecientes a siete especies de parasitoides: Doryctobracon crawfordi, Doryctobracon zeteki, Utetes anastrephae, Microcrasis sp., Bracon sp., Torymus sp., e Ichneumonidae sp.1. Los parasitoides se obtuvieron de nueve hospederos correspondientes a café, guayaba, níspero, hobo, feijoa, naranja, mango, durazno y guama. La mayor frecuencia de ocurrencia de parasitoides se presentó en Doryctobracon crawfordi, con valores de 84,1, 76,7 y 40% en café, guayaba y hobo, respectivamente. Utetes anastrephae obtuvo porcentajes de 5,7 y 60% en café y hobo. También se presentó parasitismo en moscas presentes en las familias botánicas Rubiaceae, Rosaceae, Myrtaceae, Fabaceae, Anacardiaceae y Rutaceae.
Palabras clave: insectos plaga de frutas, controladores biológicos, Diptera, Hymenoptera.
diVersitY of fruit-flies (diPtera: tePHritidae) and tHeir Parasitoids in seVen MuniCiPalities of tHe de-
PartMent of nariÑo
The objective of this work was to contribute to the knowledge of the diversity of fruit flies (Diptera: Tephritidae) and their natural enemies in the department of Nariño. A total of 14,005 fruits from 16 host plant species were collected, belonging to ten botanical families (Rubiaceae, Rosaceae, Myrtaceae, Fabaceae, Anacardiaceae, Rutaceae, Cucurbitaceae, Annonaceae, Solanaceae and Caricaceae), all of which showed infestation by fruit flies or
* FR: 3-III-2017. FA: 3-X-2017.1 Magíster en Producción de Cultivos. Facultad de Ciencias Agrícolas, Universidad de Nariño. Pasto, Colombia. E-mail: [email protected] Profesor Titular, Facultad de Ingeniería Agronómica, Universidad del Tolima. Ibagué, Colombia. E-mail: [email protected] ORCID: 0000-0001-1724-23043 Profesor Asociado, Facultad de Ingeniería Agronómica, Universidad del Tolima. Ibagué, Colombia. E-mail: [email protected] ORCID: 0000-0003-4024-8687
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 81-98. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:CRUZ, M.I., BACCA, T. & CANAL, N.A., 2017.- Diversidad de moscas de las frutas (Diptera: Tephritidae) y sus parasitoides en siete municipios del departamento de Nariño. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 81-98. DOI: 10.17151/bccm.2017.21.2.6

María Isabel Cruz B., Tito Bacca, Nelson A. Canal82bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
parasitoids, except for the Annonaceae family.The species of fruit flies collected in this study were Anastrepha fraterculus complex, Anastrepha striata, Anastrepha obliqua, Ceratitis capitata and Anastrepha grandis species known for their polyphagia and great adaptability. A total of 242 individuals belonging to seven species of parasitoids, Doryctobracon crawfordi, Doryctobracon zeteki, Utetes anastrephae, Microcrasis sp., Bracon sp., Torymus sp., and Ichneumonidae sp.1 were obtained. Parasitoids were collected from nine plant hosts corresponding to coffee, guava, loquat, hobo, guava, orange, mango, peach and guama. The highest frequency of parasitoids occurrence was presented in Doryctobracon crawfordi, with values of 84.1, 76.7 and 40% in coffee, guava and hobo respectively. Utetes anastrephae was 5.7% in coffee and 6.0% in hobo. Parasitism was also present in Rubiaceae, Rosaceae, Myrtaceae, Fabaceae, Anacardiaceae and Rutaceae.
Key words: fruit insect pests, biological control agents, Diptera, Hymenoptera.
introduCCiÓn
El comercio mundial de frutas y hortalizas ha crecido positivamente en los últimos años, en gran parte por el cambio en los hábitos de consumo de las personas, cuyas tendencias son cada vez más saludables. En la última década en Colombia, la superficie y producción hortifrutícola cultivada también ha crecido constantemente con una tasa de 3% en promedio, para el 2016 se sembraron 987 mil hectáreas con una producción de 10,4 millones de toneladas (ASOHOFRUCOL, 2016).
Entre las principales limitantes de la producción de los frutales en Colombia están los problemas fitosanitarios, donde las moscas de las frutas tiene gran importancia por la grandes pérdidas ocasionadas, especialmente el género Anastrepha Schinner, 1868 (CANAL, 2010; CASTAÑEDA et al. 2010). El Instituto Colombiano Agropecuario (ICA) ha reportado cerca de 52 especies de moscas de las frutas, donde Anastrepha complejo fraterculus, Anastrepha grandis, Anastrepha obliqua, Anastrepha serpentina, Anastrepha striata y Ceratitis capitata o mosca del mediterráneo se destacan por ser de importancia cuarentenaria (ICA, 2012). El estado larval de estas moscas provoca daño físico directo a la parte comestible de las frutas, e indirectamente permite la contaminación con hongos y bacterias, y en ocasiones puede causar la caída del fruto. En Colombia se reportan pérdidas en promedio entre el 30-40% de la producción de frutales, pero puede llegar al 70% cuando no se aplica ningún manejo de la plaga. Inclusive, cuando el daño no es significativo desde el punto de vista agronómico, el acceso a los mercados internacionales por medidas cuarentenarias es prohibido (ICA, 2012).
El primer paso para plantear medidas de manejo eficiente de plagas es el conocimiento de la diversidad de las especies plagas, y cuando el interés es fomentar el control

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 83
biológico el primer paso es conocer la diversidad de enemigos naturales (ALUJA et al., 2003; CARVALHO et al., 2010; CASTAÑEDA et al., 2010). En Colombia son escasos los estudios sistemáticos tendientes al conocimiento de la diversidad de Anastrepha y más escasos aún son los trabajos que estudian la diversidad de parasitoides de moscas de las frutas (CASTAÑEDA et al., 2010; RUIZ et al., 2013; SAAVEDRA-DÍAZ et al., 2017). En el departamento de Nariño apenas sí se conoce el trabajo de PORTILLA et al. (1995), quienes estudiaron las moscas de las frutas y parasitoides asociados a café en algunos municipios del departamento.
El papel de los parasitoides del orden Hymenoptera tiene reconocida importancia en el control de moscas de las frutas. Las investigaciones realizadas dan indicios de la riqueza y diversidad de estos biocontroladores en diversos países en los cuales se ha trabajado en forma continua en la identificación y conocimiento de especies nativas e introducidas (OVRUSKI et al., 2000; NÚÑEZ et al., 2004a). En Colombia las especies de parasitoides que atacan las moscas de la familia Tephritidae y su distribución son ampliamente desconocidas, a pesar de que se han identificado 22 especies pertenecientes a 16 géneros y 5 familias (RUIZ-HURTADO et al., 2013).
Teniendo en cuenta los anteriores antecedentes, se tuvo como objetivo del presente trabajo identificar las moscas y parasitoides de moscas de las frutas (Diptera: Tephritidae) asociados a especies frutales en siete municipios del departamento de Nariño.
Materiales Y MÉtodos
lugares de muestreo. El estudio se realizó en siete municipios del departamento de Nariño, Colombia: La Unión, Buesaco, Arboleda, Imués, Tangua, Yacuanquer y Taminango (Tabla 1). En cada municipio se seleccionaron dos fincas al azar por muestreo, y la frecuencia de recolección de frutos fue cada 15 días de enero a diciembre de 2014.
Lugares donde se recolectaron las muestras de frutos en el departamento de Nariño, Colombia.tabla 1.
Municipios Veredas altura msnm (mínima – máxima)
Arboleda Las Palmas, Rosa Florida, El Olivo, El Empate, San Vicente 1.423 – 1.900
Yacuanquer Minda, Inantás Bajo, Argüello Alto, Tasnaque, La Cocha, Chapacual, Zaragoza 1.814 – 2.264
Buesaco Higuerones, Pajajoy, Sumapaz, Villamoreno 1.702 – 2.261Tangua El Obraje, Tapialquier Bajo, Tablón Obraje, El Yunguillo 1.885 – 2.183
La Unión La Betulia, Chilcal Bajo, La Playa, Juan Solarte, Pedrera A, La Cañada, Chaguarurco, El Mayo 1.192 – 1.849
Imués Pilcuán, El Pedregal, Santa Rosa 1.806 – 2.127Taminango Panoya, Guayacanal, Paso Feo, Alto Don Diego 1.333 – 1.842

María Isabel Cruz B., Tito Bacca, Nelson A. Canal84bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
Los frutos se recolectaron directamente de la planta, según su disponibilidad y dependiendo de las épocas de maduración de cada fruta, se seleccionaron frutos totalmente maduros o sobremaduros con signos de daño por mosca de las frutas. Se tomaron frutos de varias especies, hasta reunir entre 0,5-1 kg o un número determinado de frutos por muestra por cada especie de planta. Las muestras se tomaron completamente al azar, y cada muestra se introdujo en bolsas plásticas con todos los datos requeridos para su identificación. Posteriormente las muestras se empacaron, etiquetaron y guardaron en una caja de icopor, inmediatamente después de terminada la labor de muestreo del día. Las muestras se trasladaron al Laboratorio de Entomología de la Universidad de Nariño.
Procesamiento de los frutos en laboratorio. Los frutos se pesaron y posteriormente según su grado de madurez se extrajeron las larvas. Los frutos menos maduros se dejaron en cámaras de cría para maduración de frutos y desarrollo de las larvas. Las larvas que abandonaron los frutos se ubicaron en cámaras de pupación. Las cámaras de maduración consistieron en cajas plásticas (26 x 26 x 8 cm), con fondo de papel absorbente que fue humedecido diariamente con agua destilada, evitando así la deshidratación de los frutos y la mortalidad de las larvas. La cámara fue cubierta con una tapa con malla de tul para su aireación. Los frutos se dejaron allí de tres a 10 días, para permitir el desarrollo de huevos y larvas. Las cámaras de maduración se revisaron periódicamente, con el objeto de visualizar la madurez de los frutos y determinar si se requería hacer su disección.
disección de frutos. Los frutos se disecaron con la ayuda de un cuchillo para los cortes gruesos y de un bisturí para los cortes delicados. Para la extracción de larvas del tercer instar (L3) se usaron pinzas blandas o pinceles, estas se pasaron a las cámaras de pupación, con el fin de permitir la continuación de su ciclo biológico y obtener los adultos para su identificación.
Cámaras de pupación. Las larvas de tercer instar y las pupas se colocaron dentro de frascos de vidrio con 15 g de vermiculita esterilizada y humedecida en relación 1:3 (p/v) con agua destilada, hasta obtener el empupamiento y alcanzar el estado adulto. Una vez emergieron los adultos, se dejaron dentro de las cámaras entre dos y tres días para que fijaran adecuadamente los colores. Los adultos se alimentaron con una solución azucarada. Después de dos a tres días de emergidos los adultos, se pasaron a frascos con alcohol al 70%, para su conservación e identificación taxonómica, estas muestras se marcaron con los mismos datos de colección de las frutas.
identificación de especies
identificación de plantas hospederas. Para la identificación de las plantas hospederas desconocidas se colectaron partes de las plantas como ramas, tallos, hojas, flores y

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 85
frutos. La identificación se realizó con el apoyo del Herbario de la Universidad de Nariño.
identificación de especies de moscas de las frutas y parasitoides. Las moscas que emergieron fueron identificadas por medio de caracteres morfológicos del tórax, alas y ovipositor, usando las claves taxonómicas de KORYTKOWSKI (2009). Los parasitoides fueron identificados utilizando la clave de WHARTON & YODER (2015).
análisis de datos. Con los datos obtenidos en la fase de laboratorio se calculó el nivel de infestación de las muestras de frutos (I), el porcentaje de parasitismo (%P) y la frecuencia de ocurrencia de especies de parasitoides (%F), siguiendo la metodología de BARANOWSKI et al. (1993), UCHÔA et al. (2003), NÚÑEZ et al. (2004a) y TAIRA et al. (2013), utilizando las siguientes fórmulas:
índices de diversidad. Para evaluar la abundancia y diversidad de los parasitoides colectados durante 12 meses de muestreo en siete municipios del departamento de Nariño se utilizaron los índices de diversidad de Simpson, Shannon-Wiener, Margalef y de Brillouin (VILLAREAL et al., 2006).
resultados Y disCusiÓn
reconocimiento de moscas de las frutas. Se capturaron 1.381 moscas pertenecientes a cinco especies, cuatro del género Anastrepha, y Ceratitis capitata, todas previamente reportadas para el departamento de Nariño por PORTILLA et al. (1994, 1995) (Tabla 2): Anastrepha complejo fraterculus (41,6%), Anastrepha striata (35,3%), Anastrepha obliqua (10,4%), Ceratitis capitata (12,5%) y Anastrepha grandis (0,14%) (Tabla 2). La abundancia entre los municipios osciló entre 317 moscas en el municipio de Arboleda a 28 moscas en el municipio de Taminango, localizados los dos en el norte del departamento de Nariño (Tabla 2). El género Anastrepha es un taxón neotropical que cuenta con más de 300 especies (NORRBOM et al., 2015) y se distribuyen desde el nivel del mar hasta los 2.500 m de altitud (NÚÑEZ, 1981; CASTAÑEDA et al., 2010). En la presente investigación A. complejo fraterculus, A. striata, A. obliqua

María Isabel Cruz B., Tito Bacca, Nelson A. Canal86bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
y A. grandis fueron encontradas entre los 1.192 y 2.264 msnm (Tabla 2 y Figura 1), siendo que las tres primeras especies se encuentran ampliamente distribuidas en todos los municipios y en la mayoría de las altitudes evaluadas. A. striata se encuentra por debajo de los 2.000 m y A. grandis solo fue encontrada a 1.434 m (Figura 1). Las especies predominantes, A. fraterculus s.l. y A. striata son las especies predominantes a estas altitudes en otros levantamientos (YEPES & VÉLEZ, 1989; NUÑEZ et al., 2004a; MARTÍNEZ & SERNA, 2005; CASTAÑEDA et al., 2010). Anastrepha fraterculus se distribuye entre 1.432 y 2.264 m de altitud, como ha sido reportado para el morfotipo andino de este complejo (CANAL et al., in press). A. striata está ampliamente distribuida en Colombia desde el nivel del mar hasta los 2.000 m. Como resultado de levantamientos sistemáticos a diferentes altitudes, CASTAÑEDA et al. (2010) y RUIZ-HURTADO et al. (2013) indican que en Colombia A. obliqua se distribuye en regiones bajas, por debajo de los 1.500 m de altitud, sin embargo, en este trabajo y en Antioquia sobre la Cordillera Occidental (ARÉVALO et al. 1999) esta especie fue encontrada sobre los 2.000 m. RUIZ-ARCE et al. (2012) y CASTAÑEDA et al. (2015) refieren variabilidad genética y morfométrica de la especie, sugiriendo la existencia de especies crípticas; en Colombia existe variabilidad entre las poblaciones de los ríos Magdalena y Cauca; es probable que esta variabilidad se refleje también en la biología.

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 87
Dist
ribuc
ión,
frec
uenc
ia (%
) y n
úmer
o de
esp
ecie
s mos
cas d
e la
s fru
tas y
sus p
aras
itoid
es c
olec
tada
s dur
ante
doc
e m
eses
de
mue
streo
en
siete
mun
icip
ios d
el
depa
rtam
ento
de
Nar
iño.
tabl
a 2.
Mun
icip
ios
alt
ura
msn
m
(mín
ima
– m
áxi-
ma)
% d
e la
frec
uenc
ia (f
) de
espe
cies
de
para
sito
ides
(núm
ero
de in
divi
duos
)%
de
la fr
ecue
ncia
(f) d
e es
peci
es d
e m
osca
s(n
úmer
o de
indi
vidu
os)
D. c
raf
U. a
nD
. ze
t.M
ic.
Bra
.ic
hTo
rA
. fra
A. s
tA
. ob
A. g
rC
. ca
Arbo
leda
1.42
3 –
1.90
025
,74
(52)
64,2
9 (9
)0
75
(3)
00
047
,6
(151
)26
,1 (8
3)24
,6
(78)
0,63
(2
)0,
94 (3
)
Yacu
anqu
er1.
814
– 2.
264
23,2
7 (4
7)7,
14
(1)
00
00
039
,1
(119
)37
,8 (1
15)
8,8
(27)
014
,1 (4
3)
Bues
aco
1.70
2 –
2.26
116
,34
(33)
010
0(4)
010
0 (1
)10
0 (1
)0
47,1
3 (1
15)
47,9
(117
)4,
5 (1
1)0
0,4
(1)
Tang
ua1.
885
– 2.
183
16,3
4 (3
3)0
025
(1
)0
010
0 (1
6)29
,4
(55)
12,8
(24)
00
57,7
(1
08)
La U
nión
1.19
2 –
1.84
99,
41 (1
9)21
,43
(3)
00
00
013
,6
(23)
70,2
(118
)16
,0
(27)
00
Imué
s1.
806
– 2.
127
8,42
(17)
00
00
00
68,4
(9
1)23
,3 (3
1)0,
7 (1
)0
7,5
(10)
Tam
inan
go1.
333
– 1.
842
0,50 (1)
7,14
(1
)0
00
00
75 (2
1)0
00
25(7
)
tota
l10
0 (2
02)
100
(14)
100(
4)10
0 (4
)10
0 (1
)10
0 (1
)10
0 (1
6)10
0 (5
75)
100
(488
)10
0 (1
44)
100
(2)
100
(172
)
f83
,47
5,78
1,65
1,65
0,41
0,41
6,61
41,6
335
,33
10,4
20,
1012
,44
Mos
cas d
e las
frut
as: A
. fra
= A
. fra
terc
ulus
, A. s
t = A
. stri
ata,
A. o
b =
A. ob
liqua
, A. g
r = A
. gra
ndis,
C. c
a =
C. c
apita
ta. P
aras
itoid
es: D
. cra
f = D
oryc
tobr
acon
craw
ford
i, D
. zet
= D
. zet
eki,
U. a
n =
Ute
tes a
nastr
epha
e, M
ic.=
Micr
ocra
sis sp
., Br
a.=
Brac
on sp
., Ic
h =
Ichn
eum
onid
ae sp
. 1, T
or =
Tor
ymus
sp.
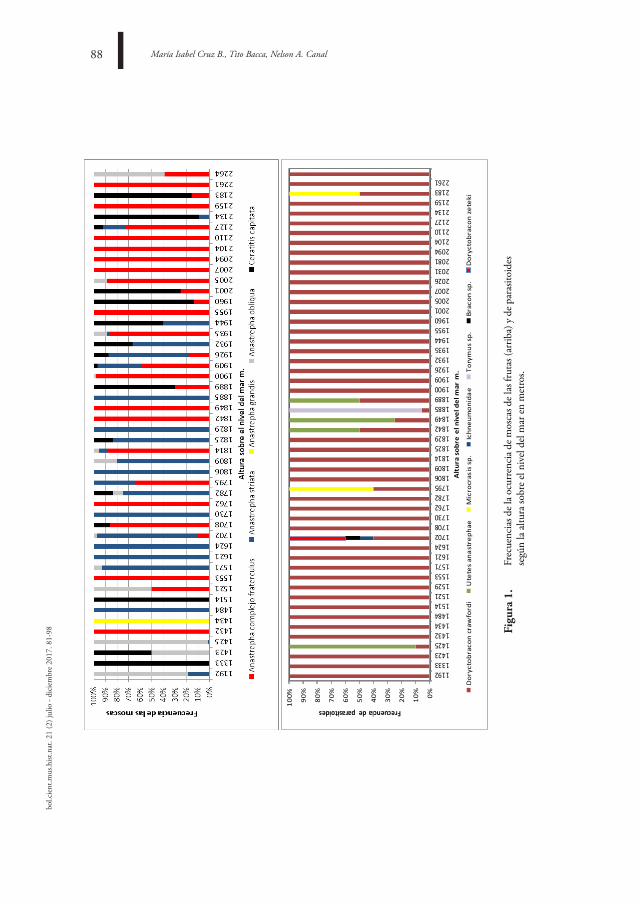
María Isabel Cruz B., Tito Bacca, Nelson A. Canal88bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
0%10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
119213331423142514321434148415141521152915531571162116241702170817301762178217951806180918141825182918421849188518891900190919261932193519441955196020012005200720262031208120942104211021272134215921832261
Frecuencia de parasitoides
Alt
ura
sobr
e el
niv
el d
el m
ar m
.
Dor
ycto
brac
on c
raw
ford
iU
tete
s an
astr
epha
eM
icro
cras
is s
p.Ic
hneu
mon
idae
Tory
mus
sp.
Brac
on s
p.D
oryc
tobr
acon
zet
eki
Frec
uenc
ias d
e la o
curr
enci
a de m
osca
s de l
as fr
utas
(arr
iba)
y d
e par
asito
ides
se
gún
la a
ltura
sobr
e el
niv
el d
el m
ar e
n m
etro
s.fi
gura
1.

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 89
Con relación a Ceratitis capitata, de acuerdo a los datos del ICA (2012), en los resultados de vigilancia fitosanitaria sobre moscas de las frutas en el departamento de Nariño, en la ruta de monitoreo establecida entre el paso fronterizo Puente Internacional de Rumichaca, en el municipio de Ipiales hasta el municipio de Tangua, esta especie se reporta desde los 1.797 a 1.917 msnm, de los 2.000 m de altura en adelante no se reportan capturas, sin embargo en este trabajo la distribución altitudinal para C. capitata estuvo entre los 1.333 y 2.183 msnm.
De acuerdo a ALUJA et al. (2003), A. complejo fraterculus, A. striata y A. obliqua son especies conocidas dentro del género por su polifagia y gran capacidad de adaptación. En el estudio realizado por CASTAÑEDA et al. (2010), el rango altitudinal para el género Anastrepha estuvo entre los 300 y 2.500 msnm. Las especies predominantes correspondieron a A. striata, A. fraterculus y A. obliqua. Del mismo modo otros autores, como GIRALDO et al. (2008) en el Valle del Cauca y KING (2005) en Tolima, reportaron la presencia de estas mismas especies en sus trabajos, por lo tanto no es extraño que hayan sido las especies con mayor presencia en este estudio. De acuerdo a ZUCCHI (2000), el primer factor que debe incidir sobre la distribución altitudinal de las especies de Anastrepha es la presencia de su hospedero, lo que concuerda con lo planteado por CASTAÑEDA et al. (2010), quienes también afirman que la presencia de ciertas especies de moscas de las frutas en un determinado rango altitudinal puede deberse a la disponibilidad de la planta huésped, por lo tanto sugieren para el estudio de esta plaga la búsqueda de hospederos alternativos, con el fin de conocer la relación planta-hospedero, y determinar la secuencia temporal de la presencia de la plaga y de este modo tener una idea real de la diversidad de especies en la región.
relación entre hospederos y moscas de las frutas. Se colectaron 14.005 frutos correspondientes a 16 especies de plantas hospederas, pertenecientes a 10 familias botánicas, de las cuales todas presentaron infestación por moscas de las frutas, a excepción de la familia Annonaceae (Tablas 3 y 4). Entre las nueve familias botánicas hospederas de mosca, el 80% de infestación se restringe a Rubiaceae, Rosaceae y Myrtaceae (Tablas 3 y 4). Las especies vegetales Coffea arabica, perteneciente a la familia Rubiaceae y Acca sellowiana, de la familia Myrtaceae, fueron las plantas hospederas que mostraron mayor presencia de moscas de las frutas, con cuatro especies identificadas: A. complejo fraterculus, A. obliqua, A. striata y C. capitata, mientras que la especie eriobotrya japonica (Rosaceae) fue hospedera de tres especies: A. complejo fraterculus, A. striata y C. capitata, al igual que la especie Psidium guajava (Myrtaceae) que resultó afectada por la presencia de A. complejo fraterculus, A. striata y A. obliqua. Por su parte, la especie Prunus pérsica (Rosaceae) presentó solo dos especies: A. complejo fraterculus y A. obliqua (Tabla 3).
En las familias Fabaceae, Anacardiaceae, Rutaceae y Cucurbitaceae se encontró un número menor de moscas de las frutas, las cuales fluctuaron entre una y dos especies,
María Isabel Cruz B., Tito Bacca, Nelson A. Canal90bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
tampoco fue posible observar afectación en la familia Annonaceae, sin embargo en las familias Solanaceae y Caricaceae a pesar de haberse presentado infestación en los frutos no se obtuvieron adultos de moscas de las frutas (Tabla 3).
Las especies vegetales Spondias mombin (Anacardiaceae) y Citrus sinensis (Rutaceae) presentaron la presencia de A. striata y A. obliqua, mientras que Mangifera indica (Anacardiaceae), Citrus nobilis (Rutaceae) y Cucurbita sp. (Cucurbitaceae) solo presentaron una especie de mosca, A. striata, C. capitata y A. grandis, respectivamente (Tabla 3).
SAAVEDRA-DÍAZ et al. (2017) colectaron e individualizaron frutos para estudiar las relaciones tróficas entre los frutos hospederos, las moscas de las frutas y sus parasitoides. Ellos encontraron que especies de Anastrepha en bosque seco tropical tienden a no compartir hospedero y que cuando lo hacen, normalmente no comparten el mismo fruto. En este estudio, especies polífagas como A. obliqua, A. striata y A. serpentina apenas ocasionalmente usaron hospederos diferentes a sus preferidos. En este trabajo no se individualizaron frutos, pero se observó una preferencia de las especies por hospederos específicos.
Frutales hospederos de las diferentes especies de moscas tefrítidas con su respectivo nivel de infestación en siete municipios del departamento de Nariño
tabla 3.
familia botánica especie de planta nombre
comúnn° de frutos
n° de larvas
nivel de infes. (i)
n° de moscas
especies moscas de las frutas
Rubiacea Coffea arabica Café 12.362 1.118 0,09 569 A. fraterculus, A. striata, A. obliqua, C. capitata
Rosaceae Prunus persica Durazno 12 166 13,83 31 A. fraterculus, A. obliqua
Rosaceae eriobotrya japonica Níspero 969 668 0,69 195 A. fraterculus, A. striata,
C. capitata
Myrtaceae Acca sellowiana Feijoa 87 287 3,30 216 A. fraterculus, A. striata, A. obliqua, C. capitata
Myrtaceae Psidium guajava Guayaba 204 637 3,12 212 A. fraterculus, A. striata, A. obliqua
Fabaceae Inga sp. Guama 13 85 6,54 58 A. striataAnacardiaceae Spondias mombin Hobo 187 150 0,80 67 A. striata, A. obliquaAnacardiaceae Mangifera indica Mango 8 12 1,50 6 A. striata
Rutaceae Citrus nobilis Mandarina 33 54 1,64 31 A. striata, A. obliquaRutaceae Citrus sinensis Naranja 23 5 0,22 2 C. capitata Rutaceae Citrus aurantifolia Lima 17 0 0 0 0 especies
Cucurbitaceae Cucurbita sp. Zapallo 3 3 1,0 2 A. grandisSolanaceae Solanum quitoense Lulo 66 1 0 0 0 especiesCaricaceae Carica papaya Papaya 9 23 2,56 0 0 especies
total 13.993 3.209 X = 2,52 1.389 5 especies
Estudios realizados por ALUJA et al. (2003) han permitido demostrar que las especies polífagas usan diferentes hospederos como estrategia ecológica para mantener poblaciones significativas en épocas de poca presencia de frutos, e incluso comparten hospederos con otras especies polífagas.

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 91
Según NÚÑEZ et al. (2004a), CASTAÑEDA et al. (2010), SARMIENTO et al. (2012) y RUIZ et al. (2013), A. striata está asociada con plantas de la familia Myrtaceae, pero en el Neotrópico se ha encontrado que afecta 18 especies de plantas de 7 familias y se ha reportado en zonas altas ocasionalmente en frutos de café y hobo; apenas un reporte de Ecuador refiere esta especie parasitando Inga (NORRBOM, 2004). A. fraterculus ataca 80 especies de plantas pertenecientes a 20 familias (NORRBOM, 2004). Según SARMIENTO et al. (2012) y SAAVEDRA-DÍAZ et al. (2017), A. obliqua ha sido reportada utilizando Psidium guajava como hospedero en otros trabajos pero con baja infestación; estos autores argumentan que P. guajava no es un hospedero de uso común en Colombia, pero que la polifagia de la especie le permite usarlo ocasionalmente.
En lo referente a la presencia de A. grandis, esta especie fue recuperada de un hospedero de la familia Cucurbitaceae (Tabla 3), lo que concuerda con lo reportado por CASTAÑEDA et al. (2010) en el departamento del Tolima. Dichos autores argumentan que su presencia se debe posiblemente a que esta especie presenta una distribución altitudinal entre los 950 y los 1.600 msnm. En este trabajo esta especie fue recuperada a los 1.434 msnm en el municipio de Arboleda y solo fueron colectados dos individuos (Tabla 2).
De acuerdo a LIQUIDO et al. (1991), C. capitata dentro de la familia Tephritidae es una de las especies más polífagas; según estos autores se han reportado más de 350 especies de hospedantes reales o potenciales para esta especie. En el departamento de Nariño, C. capitata fue reportada por PORTILLA et al. (1994) en cultivos de café. MORALES et al. (2004) y GUZMÁN (2010) reportaron a C. capitata atacando a: Acca sellowiana, eriobotrya japonica, Coffea arabica y Citrus nobilis.
nivel de infestación de frutos (i). El nivel de infestación total de frutos durante la investigación fue de 2,52 larvas/fruto, valor que se obtuvo de los frutos pertenecientes a nueve familias botánicas. Las especies vegetales Prunus persica e Inga sp. reportaron los niveles más altos, con valores de 13,83 y 6,54, respectivamente (Tabla 3). El resto de especies de frutos presentaron niveles de infestación que fluctuaron entre 0,02 a 3,3 (Tabla 3). En frutos de café fue donde se obtuvo el mayor número de larvas, consecuente con el mayor número de frutos colectados (Tabla 3). En algunas plantas el número de frutos disponibles fue escaso (durazno, zapallo, chirimoya, tomate de árbol, papaya y mango). La abundancia y calidad de estos frutos en campo los convirtió en hospederos alternantes en la zona de estudio, pudiendo afectar la infestación en frutos, lo cual a su vez puede variar con la especie de tefrítido (MALAVASI & MORGANTE, 1980). Por lo tanto, el factor que influye en gran medida en la disminución de la infestación es la mayor disponibilidad de frutas que lleva a la distribución homogénea de la oviposición (NÚÑEZ et al., 2004b). Según PROKOPY (1984), dependiendo de la densidad de adultos en el ambiente y la susceptibilidad del fruto a la oviposición,

María Isabel Cruz B., Tito Bacca, Nelson A. Canal92bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
los niveles de daño son variables en cada hospedante y permiten el desarrollo de la larva. En la escogencia del fruto como sitio de oviposición actúan diversos estímulos (forma, color, contenido de nutrientes, tamaño, madurez, entre otros), todos estos factores generan el comportamiento de oviposición y final aceptación del fruto.
Porcentaje y frecuencia de parasitismo en moscas de las frutas. Se obtuvieron 241 individuos parasitoides de las especies: Doryctobracon crawfordi, Doryctobracon zeteki, Utetes anastrephae, Microcrasis sp. y Bracon sp., pertenecientes a la familia Braconidae, un individuo de la familia Torimidae (Torymus sp.) y un individuo de la familia Ichneumonidae (Tabla 2). D. carwfordi, U. anastrephae y Microcrasis ya habían sido reportados previamente en el departamento de Nariño por PORTILLA et al. (1994) en frutos infestados de café. Entretanto, parasitoides del género Bracon o de la familia Torymidae no habían sido registrados en Colombia; los otros parasitoides son registros nuevos para Nariño. Según RUIZ et al. (2013), en Colombia se conocen cinco familias, 16 géneros y 24 especies de controladores biológicos de tefrítidos.
Los parasitoides fueron obtenidos en nueve frutos hospederos (Tablas 4 y 5). Autores como STECK et al. (1986) y ALUJA et al. (1990) han realizado estudios para determinar las tasas naturales de parasitismo en áreas naturales y cultivadas, catalogando porcentajes de parasitismo altos (alrededor del 56%) y bajos (5%), dependiendo del tamaño del fruto, siendo mayor en frutos pequeños. Según lo anterior, el porcentaje de parasitismo de este estudio en la mayoría de los casos fue medio porque estuvo entre 15,46 a 27,06 y bajo para el caso de la familia Fabaceae. (Tabla 5).
Especies de moscas de las frutas por cada familia de hospedero con el respectivo número de parasitoides emergidos y el porcentaje de parasitismo
tabla 4.
familia botánicano. de moscas
% de parasitismoAnastrepha spp. C. capitata
Rubiacea 411 158 15,46Rosaceae 216 2 27,06
Myrtaceae 418 10 17,75Fabaceae 58 0 1,724
Anacardiaceae 73 0 21,91Rutaceae 31 2
Cucurbitaceae 2 0total 1.209 172 X = 14,99
En Brasil, GUIMARÃES et al. (2004) sostienen que las frutas de las familias Myrtaceae y Anacardiaceae son más atractivas a los parasitoides, datos que están acorde con los valores de parasitismo obtenidos para estas familias en esta investigación. Adicionalmente se pudo observar que se presentó relación entre los mayores porcentajes de parasitismo y la diversidad de frutos de menor tamaño (Tablas 3 y 4).

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 93
Doryctobracon crawfordi fue el parasitoide más abundante, presentó alta frecuencia de ocurrencia en café, guayaba y hobo con valores de 84,1, 76,7 y 40%, respectivamente (Tabla 5), y fue la única especie emergida en durazno, níspero, feijoa, guama, mango y naranja (Tabla 2). Este parasitoide fue hallado en Arboleda con 25,74% (52 individuos), en Yacuanquer con 23,74% (47 individuos), en Buesaco y Tangua con 16,34% (33 individuos cada uno), en La Unión con 9,41% (19 individuos), en Imués con 8,42% (17 individuos) y en Taminango con 0,50% (1 individuo) (Tabla 2). D. crawfordi es la especie predominante en la zona media altitudinal de las cordilleras colombianas (RUIZ-HURTADO et al., 2013).
El total de individuos colectados de Utetes anastrephae en esta investigación fue de 14, fue recuperado en café y hobo, y los porcentajes de frecuencia de ocurrencia fueron 5,7 y 60%, respectivamente (Tabla 5). Esta especie parasitoide fue hallada en los municipios de Arboleda con 64,2% (9 individuos), La Unión con 21,43% (3 individuos), Yacuanquer y Taminango con 7,14% (1 individuo) respectivamente (Tabla 2), y hallado en especies de Anastrepha complejo fraterculus, Anastrepha striata, Anastrepha obliqua y Ceratitis capitata. Este parasitoide fue colectado entre los 1.425 y 1.889 m de altitud; fue la segunda especie en abundancia y normalmente es colectado en todos los levantamientos de parasitoides de moscas de las frutas (RUIZ-HURTADO et al., 2013). Los otros parasitoides se encontraron en un rango altitudinal a entre los 1.600 y 1.900 msnm (Figura 1).
En Coffea arabica, en el municipio de Buesaco, se obtuvieron cuatro ejemplares de D. zeteki y un ejemplar de Bracon sp. (Tabla 5), colectados junto a Anastrepha complejo fraterculus, Anastrepha striata, Anastrepha obliqua y Ceratitis capitata. D. zeteki fue el parasitoide más común en el Valle del Cauca, pero provenían en su mayoría de la región de Buenaventura y también fue el parasitoide más común en áreas húmedas de Panamá (CARREJO & GONZÁLEZ, 1999; MEDIANERO et al., 2006). El género Bracon es megadiverso y dos especies han sido reportadas parasitando tefritídeos, pero ninguna de ellas en América Latina (WHARTON & YODER, 2015).
Los parasitoides Bracon sp., Microcrasis sp., Ichneumonidae sp.1 y Torymus sp. tuvieron una frecuencia de ocurrencia correspondiente a 21,9, 5,7, 4,5 y 1,4%, respectivamente (Tabla 5). El torímido fue colectado en el municipio de Tangua en Psidium guajava y recuperado de las especies de mosca Anastrepha complejo fraterculus, Anastrepha striata y Anastrepha obliqua (Tablas 2 y 5). Según WHARTON & YODER (2015), una especie de Torimidae del género Microdontomerus ha sido reportada parasitando moscas del género Bactorcera en Sur África; estos autores y OVRUSKI et al. (2000) no hacen referencia a colectas de torímidos en América. GRISSELL (2005) reporta parasitoides torímidos atacando tefrítidos en asteráceas, la biología y taxonomía del género Torymus es escasa y requiere ser estudiada.
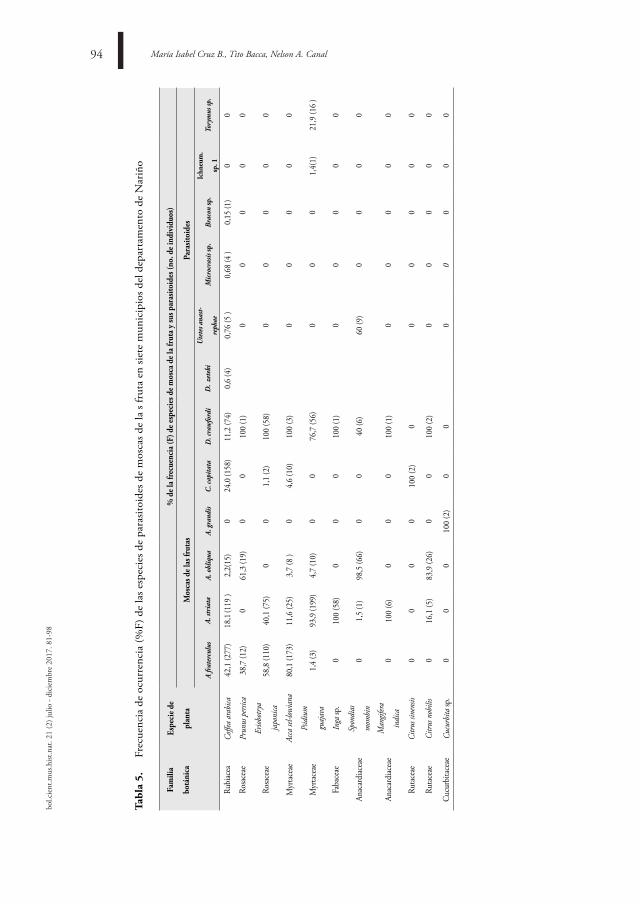
María Isabel Cruz B., Tito Bacca, Nelson A. Canal94bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
tabl
a 5.
F
recu
enci
a de
ocu
rren
cia
(%F)
de
las e
spec
ies d
e pa
rasi
toid
es d
e m
osca
s de
la s
frut
a en
siet
e m
unic
ipio
s del
dep
arta
men
to d
e N
ariñ
o

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 95
Se hallaron 4 individuos de Microcrasis sp. en los municipios de Arboleda y Tangua (Tabla 5), este parasitoide fue recolectado junto a Anastrepha complejo fraterculus, Anastrepha striata, Anastrepha obliqua y Ceratitis capitata (Tabla 5), y la especie botánica hospedera fue Coffea arabica. Microcrasis sp. hace parte de la subfamilia Alysiinae, la cual es una de las más ricas de Braconidae con más de 1.900 especies descritas a nivel mundial (WHARTON & YODER, 2015). El parasitoide Microcrasis sp. fue reportado por PORTILLA et al. (1994) en el departamento de Nariño, en moscas de las frutas en café en los municipios de Tangua y Yacuanquer, lo que concuerda con lo encontrado en este estudio en donde este parasitoide fue colectado también en el municipio de Tangua y Arboleda. Del mismo modo, RUIZ-HURTADO et al. (2013) en dos áreas cafeteras del departamento del Tolima reportaron la presencia de este parasitoide en su investigación; SARMIENTO et al. (2012), en tres localidades de la vertiente occidental de la Cordillera Oriental colombiana, identificaron a Microcrasis sp. parasitando moscas de las frutas en café, y GUARÍN & LEÓN (2002) y NÚÑEZ et al. (2004a) también hallaron parasitoides de este género en la provincia de Vélez (Santander).
Algunos géneros de ichneumonidos han sido colectados de puparios de Tephritidae (WHARTON & YODER, 2015). Respecto al ejemplar Ichneumonidae sp.1, fue hallado en Psidium guajava, en el municipio de Buesaco, sin embargo debido a la baja cantidad de individuos no fue posible establecer una relación directa con alguna de las especies de mosca en donde fue recuperado (Anastrepha complejo fraterculus, Anastrepha striata y Anastrepha obliqua).
índices de abundancia y diversidad. De acuerdo al valor obtenido en el índice de Simpson (0,70), este indica que existe poca diversidad de la comunidad de parasitoides. Es decir, si se toman dos individuos al azar, existe alta probabilidad de que estos sean de la misma especie. El índice Shannon-Wiener presentó un valor de -0,67, el cual se considera bajo, si se tiene en cuenta que en la mayoría de los ecosistemas naturales este varía entre 0,5 y 5, situación que se explica debido a que de las seis especies identificadas en el estudio, Doryctobracon crawfordi fue la que presentó el mayor número de individuos capturados, y por lo tanto fue la especie que más influyó en la investigación. Por su parte, el índice de Margalef obtuvo un valor de 0,91, valor que al encontrarse por debajo de 2, determinó que la biodiversidad estimada de los parasitoides de moscas de las frutas identificados en siete municipios muestreados fue baja. Para evaluar cómo estaban representadas las especies de parasitoides con relación al número total de individuos, se utilizó el índice de Brillouin, cuyo valor obtenido fue de 0,63, lo que indica que la relación entre los 242 individuos de todas las especies de parasitoides colectados y el número de individuos de cada una fue baja.

María Isabel Cruz B., Tito Bacca, Nelson A. Canal96bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
ConClusiones
Las especies de moscas de las frutas recuperadas en este estudio correspondieron a Anastrepha complejo fraterculus, Anastrepha striata, Anastrepha obliqua, Anastrepha grandis y Ceratitis capitata, siendo Anastrepha complejo fraterculus la especie más abundante. Los municipios Arboleda, Yacuanquer, Buesaco y Tangua fueron las localidades donde las especies de moscas de las frutas presentaron mayor abundancia durante los meses de muestreo; las familias Rubiaceae, Rosaceae y Myrtaceae presentaron el mayor número de especies de moscas de las frutas afectando frutos. Para el departamento de Nariño se reportaron siete parasitoides de moscas de las frutas: Doryctobracon crawfordi, D. zeteki, Utetes anastrephae, Microcrasis sp., Bracon sp., Torymus sp., e Ichneumonidae sp.1, dentro de los cuales los tres últimos son nuevos reportes para este departamento y Colombia, lo que demuestra la diversidad de este grupo en esta zona del país. En cuanto a los índices de diversidad, los valores obtenidos fueron bajos, esto es debido posiblemente a las bajas abundancias de los parasitoides respecto a Doryctobracon crawfordi, ya que esta especie obtuvo 202 individuos sobre el total de parasitoides obtenidos que fue de 242 individuos, por lo tanto es importante tener en cuenta que este tipo de índices se ve influenciado por el tamaño de muestra.
referenCias
ALUJA, M., GUILLÉN, J., LIEDO, P., CABRERA, M., RÍOS, E., DE LA ROSA, G., CELEDONIO, H. & MOTA, D., 1990.- Fruit infesting Tephritids (Diptera: Tephritidae) and associated parasitoids in Chiapas, México. entomophaga, 35 (1): 28-39.
ALUJA, M., RULL, J., SIVINSKI, J., NORRBOM, A., WHARTON, R., MACÍAS, R., DÍAZ-FLEISCHER, F. & LÓPEZ, M., 2003.- Fruit flies of the genus Anastrepha (Diptera: Tephritidae) and associated native parasitoids (Hymenoptera) in the tropical rainforest biosphere reserve of Montes Azules, Chiapas, Mexico. environmental entomology, 32 (6): 1377-1385.
ARÉVALO, P.E., RESTREPO, A.A. & ALEIZA, G.M.A. 1997.- Las moscas de las frutas del género Anastrepha Schiner (Diptera: Tephritidae) en el departamento de Antioquia-Colombia. pp. 271-282. (en): Memorias Aconteceres entomológicos para Comprender los Insectos y estudiarlos. Medellín, Colombia.
ASOHOFRUCOL (ASOCIACIÓN HORTIFRUTÍCOLA DE COLOMBIA), 2016.- Balance del sector hortifrutícola en 2016. Recuperado de http://www.asohofrucol.com.co/interna.php?cat=3&scat=45&act=1
BARANOWSKI, R., GLENN, H. & SIVINSKI, J., 1993.- Biological control of the Caribbean fruit fly (Diptera: Tephritidae). The Florida entomologist, 76 (2): 245-251.
CANAL, N., GALEANO-OLAYA, P., CASTAÑEDA, M., in press.- Phenotypic structure of Colombian populations of Anastrepha fraterculus complex (Diptera: Tephritidae). Florida entomologist.
CANAL, N.A. 2010.- New species and records of Anastrepha Schiner (Diptera: Tephritidae) from Colombia. Zootaxa, 2425: 31-44.CARREJO, S. N. & GONZÁLEZ. R. O. R.1993.- Una nueva especie de Anastrepha Schiner, 1868 (Diptera: Tephritidae) de Colombia.
Boletín del Museo entomológico de la Universidad del Valle, 1(2): 47-53.CARVALHO, R. DA S., SOARES F., W. DOS S. & RITZINGER R., 2010.- Umbu-cajá como repositório natural de parasitoide
nativo de mosca-das-frutas. Notas Científicas. Pesquisa Agropecuária Brasileira Agropecuárias, 45 (10): 1222-1225.CASTAÑEDA, M., OSORIO, A., CANAL, N. & GALEANO, P., 2010.- Especies, distribución y hospederos del género Anastrepha
Schiner en el departamento del Tolima, Colombia. Agronomía Colombiana, 28 (2): 265-271. CASTAÑEDA, M., SELIVON, D., HERNÁNDEZ-ORTIZ, V., SOTO, A. & CANAL, N., 2015.- Morphometric divergence in
populations of Anastrepha obliqua (Diptera, Tephritidae) from Colombia and some Neotropical locations (en) DE MEYER, M., CLARKE, A., VERA, M. & HENDRICHS, J. (eds.) Resolution of Cryptic Species Complexes of Tephritid Pests to enhance SIT Application and Facilitate International Trade. ZooKeys, 540: 61-81.
GIRALDO, M., HERBERTH, M., & ZUCCHI, R., 2008.- Monitoreo de moscas de las frutas (Diptera: Tephritidae) en nueve municipios, Valle del Cauca (en) Resúmenes XXXV Congreso de la Sociedad Colombiana de entomología. Socolen, Cali. p. 89.
GRISSELL, E. 1995.- Toryminae (Hymenoptera: Chalcidoidea: Torymidae). A redefinition, generic classification, an annotated world catalog of species. Memoirs on entomology International, 2:1-470.
GUARÍN, G. & LEÓN, G., 2002.- Reconocimiento, distribución temporal y espacial de moscas de las frutas (DIP: Tephritidae) y sus parasitoides en guayaba (Psidium guajava l) y café (Coffea arabica l) en tres municipios de la Provincia de Vélez (Santander) Colombia: Tesis, Escuela de Biología, U.P.T.C., Tunja.

Diversidad de las moscas de las frutas (diptera: tephritidae) y sus parasitoides en siete municipios de Nariño 97
GUIMARÃES, J., DE SOUZA, M., RAGA, A. & ZUCCHI, R., 2004.- Levantamento e interações tritóficas de Figitídeos (Hymenoptera: Eucoininae) parasitoides de larvas frugívoras (Diptera) no Brasil. Arquivos do Instituto Biológico, 71: 51-56.
GUZMÁN, R., 2010.- Mosca del mediterráneo Ceratitis capitata (Wiedemann). Ficha técnica. Colegio de Postgraduados. Dirección General de Sanidad Vegetal - Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. México. p. 41.
ICA (INSTITUTO COLOMBIANO AGROPECUARIO), 2012.- Boletín Epidemiológico. Resultados de Vigilancia Fitosanitaria sobre Moscas de las Frutas en el departamento de Nariño. Primer Trimestre. Dirección Técnica de Epidemiología y Vigilancia Fitosanitaria.
KING, W., 2005.- Detección y determinación taxonómica de moscas de las frutas en el departamento del Tolima (en) Resúmenes XXXII Congreso de la Sociedad Colombiana de entomología. Socolen, Bogotá. p. 106.
KORYTKOWSKI, C., 2009.- Manual para la identificación de Moscas de las Frutas genero Anastrepha Schiner, 1868. Universidad de Panamá, programa de Maestría en Entomología. Vicerrectoría de Investigación y Postgrado. Panamá.
LIQUIDO, N., SHINODA, L. & CUNNINGHAM, R., 1991.- Host plants of the Mediterranean fruit by (Diptera: Tephritidae): an annotated world review. Miscellaneous Publications of the entomological Society of America, 77: 1-52.
MALAVASI, A. & MORGANTI, J., 1980.- Biología da moscas-frutas (Diptera: Tephritidae). II. Índices de infestação em diferentes hospedeiros e localidades. Revista Brasil Biologia, 40 (1): 17-24.
MARTÍNEZ, A. J. O. & SERNA, S. J. F. 2005.- Identificación y localización geográfica de especies Anastrepha (Diptera: Tephritidae) en Cundinamarca (Colombia). Agronomía Colombiana, 23(1): 102-111.
MEDIANERO, E., KORYTKOWSKI, C., CAMPO, C. & DE LEÓN, C., 2006.- Hymenoptera parasitoides asociados a Anastrepha (Diptera: Tephritidae) en Cerro Jefe y Altos de Pácora, Panamá. Revista Colombiana de entomología, 32 (2): 136-139.
MORALES, P., CERMELI, M., GODOY, F. & SALAS, B., 2004.- Lista de hospederos de la mosca del Mediterráneo Ceratitis capitata Wiedemann (Diptera: Tephritidae) basada en los registros del Museo de Insectos de Interés Agrícola del INIA CENIAP. entomotropica, 19 (1): 51-54.
NORRBOM, A. 2004.- BioSystematic Database of World Diptera (BDWD). Recuperado de URL: < http://www.sel.barc.usda.gov/diptera/tephriti/tephriti.htm>.
NORRBOM A., RODRIGUEZ E., STECK G., SUTTON B. & NOLAZCO, N., 2015. New species and host plants of Anastrepha (Diptera: Tephritidae) primarily from Peru and Bolivia. Zootaxa, 4041 (1): 1-94.
NÚÑEZ, L., 1981.- Contribución al reconocimiento a las moscas de las frutas (Díptera: Tephritidae) en Colombia. Revista ICA, 16 (4): 173-179.
NÚÑEZ, L., GÓMEZ, R., GUARÍN, G. & LEÓN, G., 2004a.- Moscas de las frutas (Diptera: Tephritidae) y parasitoides asociados con Psidium guajava L. y Coffea arabica L. en tres municipios de la Provincia de Vélez (Santander, Colombia). Parte 2: Identificación y evaluación de parasitoides del Orden Hymenoptera. Revista Corpoica, 5 (1): 13-21.
NÚÑEZ, L., GÓMEZ, R., GUARÍN, G. & LEÓN, G., 2004b.- Moscas de las frutas (Diptera: Tephritidae) y parasitoides asociados con Psidium guajava L. y Coffea arabica L. en tres municipios de la Provincia de Vélez (Santander, Colombia). Parte 1: Índices de infestación y daño por moscas de las frutas (Diptera: Tephritidae). Revista Corpoica, 5 (1): 5-12.
OVRUSKI, S., ALUJA, M., SIVINSKI, J. & WHARTON, R., 2000.- Hymenopteran parasitoids on fruit-infesting Tephritidae (Diptera) in Latin America and the southern United States: diversity, distribution, taxonomic status and their use in fruit fly biological control. Integrated Pest Management Reviews, 5 (2): 81-107.
PORTILLA, M., GONZÁLEZ, G. & NÚÑEZ, L., 1994.- Infestación, reconocimiento e identificación de moscas de las frutas y sus enemigos naturales en café. Revista Colombiana de entomología, 20 (4): 261-266.
PORTILLA, M., GONZÁLEZ, G. & NÚÑEZ, L., 1995.- Evaluación y descripción del daño ocasionado por Ceratitis capitata al café. Revista Colombiana de entomología, 21 (1): 15-24.
PROKOPY, R., 1984.- Getting to know a fruit fly. Journal of Georgia entomology Society, 17 (9): 30-36.RUIZ-ARCE, R., BARR, N., OWEN, C., THOMAS, D. & MCPHERON, B., 2012.- Phylogeography of Anastrepha obliqua inferred
with mtDNA sequencing. Journal of economic entomology, 105: 2147-2160.RUÍZ-HURTADO, F., RAMÍREZ, J., ROJAS, B., GALEANO, P & CANAL, N. 2013.- Diversidad de parasitoides (Hymenoptera)
de moscas frugívoras (Diptera: Tephritoidea) en dos áreas cafeteras del departamento del Tolima, Colombia. Revista Tumbaga. 8(11): 29-53.
SAAVEDRA-DÍAZ, J., GALEANO-OLAYA, P. & CANAL, N., 2017.- Relaciones ecológicas entre frutos hospederos, moscas frugívoras y parasitoides en un fragmento de bosque seco tropical. Revista de Ciencias Agrícolas, 34 (1): 32-49.
SARMIENTO, C., AGUIRRE, H. & MARTÍNEZ-A., J., 2012.- Anastrepha (Diptera: Tephritidae) y sus asociados: Dinámica de emergencia de sus parasitoides en frutos de tres especies de plantas. Boletín del Museo de entomología de la Universidad del Valle, 13 (1): 25-32.
STECK, G., GILSTRAP, E., WHARTON, R. & HART, W., 1986.- Braconid parasitoids of Tephritidae (Diptera) infesting coffee and other fruits in West-central Africa. entomophaga, 31 (1): 59-67.
TAIRA, T., ABOT, A., NICÁCIO, J., UCHÔA, M., RODRIGUES, S. & GUIMARÃES, J., 2013.- Fruit flies (Diptera, Tephritidae) and their parasitoids on cultivated and wild hosts in the Cerrado-Pantanal ecotone in Mato Grosso do Sul, Brazil. Revista Brasileira de entomologia, 57 (3): 300-308.
UCHÔA, M., MOLINA, R., OLIVEIRA, I., ZUCCHI, R., CANAL, N. & DIAS, N., 2003.- Larval endoparasitoids (Hymenoptera) of frugivorous flies (Diptera, Tephritoidea) reared from fruits of the cerrado of the State of Mato Grosso do Sul, Brazil. Revista Brasileira de entomologia, 47 (7): 181-186.
VILLAREAL, H., ÁLVAREZ, M., CÓRDOBA, S., ESCOBAR, F., FAGUA, G., GAST, F., MENDOZA, H., OSPINA, M. & UMAÑA, A., 2006.- Métodos para el análisis de datos: una aplicación para resultados provenientes de caracterizaciones de biodiversidad (en) Manual de métodos para el desarrollo de inventarios de biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
WHARTON, R. & YODER, M., 2015.- Parasitoids of Fruit-Infesting Tephritidae. Recuperado de http://paroffit.org/public/site/paroffit/home/key

María Isabel Cruz B., Tito Bacca, Nelson A. Canal98bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 8
1-98
YEPES, R. F. & VÉLEZ, A. R.1989.- Contribución al conocimiento de las moscas de las frutas (Tephritidae) y sus parasitoides en el departamento de Antioquia. Revista Facultad Nacional de Agronomía, 42(2): 73-98.
ZUCCHI, R.A., 2000.- Espécies de Anastrepha, sinonímias, plantas hospedeiras e parasitoides: 41-48 (en) MALAVASI, A. & ZUCCHI, R.A. (eds.) Moscas-das-frutas de importância econômica no Brasil. Conhecimento básico e aplicado. Ribeirao Preto, Holos, Brasil.

ZooloGía VerteBradosVertebrate Zoology


Boletín CientífiCoCentro de Museos
Museo de Historia natural
BioloGía reProduCtiVa de Sicalis flaveola (aVes: tHrauPidae) en Cali, ColoMBia*
Camilo espinosa1, lorena Cruz-Bernate2, Guillermo Barreto3
resumen
El Sicalis Coronado (Sicalis flaveola) es una especie principalmente granívora, común en hábitats abiertos, anidador secundario de cavidades y también usa nidos abandonados de otras especies. El conocimiento sobre la historia natural de la especie proviene principalmente de Argentina y Brasil; sin embargo, acerca de Sicalis flaveola flaveola distribuida en Colombia y Venezuela sólo se conocen algunos registros de tamaño de nidada. Entre enero de 2013 y mayo de 2015 se estudió la biología reproductiva de S. f. flaveola en el campus de Meléndez en la Universidad del Valle, Cali, Colombia. Se instalaron 94 cajas nido para propiciar mayor recurso de anidamiento y se hicieron cuatro recorridos semanales en busca de nidos activos y revisión de cajas nido. El tamaño de postura fue de 3,46 huevos, el promedio de polluelos que nacen es de tres con una proporción de sexo 1:1. La incubación estuvo a cargo de la hembra mientras el cuidado de los polluelos fue efectuado por ambos miembros de la pareja. El éxito reproductivo de la especie al momento de abandonar el nido fue de 65,8%. Se encontraron diferencias con las subespecies de las latitudes del sur de Suramérica, con un menor rango de tamaño de postura y con períodos más largos de incubación y permanencia de polluelos en el nido. Sin embargo, en aspectos como el cuidado paternal durante la incubación y el cubrimiento de los polluelos, son similares.
Palabras clave: cajas nido, latitud, ecología urbana, tamaño de postura, tasa de crecimiento, trópico, Thraupidae.
reProduCtiVe BioloGY of Sicalis flaveola (aVes: tHrauPidae) in Cali, ColoMBia
abstract
The Saffron Finch (Sicalis flaveola) is a mainly grain-eating species adapted to open habitats and a secondary cavity nester that also uses abandoned nests from other species. Knowledge about the natural history of the species comes mainly from Argentina and Brazil. However, only some data on the clutch size of S. f. flaveola distributed in Colombia and Venezuela were available. The reproductive biology of S. f. flaveola was conducted on the Meléndez Campus of * FR: marzo 31, 2017. FA: abril 11, 2017.1 BSc. Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle. Cali, Colombia. E-mail: [email protected] ORCID: 0000-0002-1538-97552 M.Sc. Profesor Asistente. Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle. Cali, Colombia. E-mail: [email protected] ORCID: 0000-0002-6854-88523 Ph.D. Profesor Titular. Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle. Cali, Colombia. E-mail: [email protected] ORCID: 0000-0003-2568-2629
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 101-114. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:ESPINOSA, C., CRUZ-BERNATE, L. & BARRETO, G., 2017.- Biología reproductiva de Sicalis flaveola (Aves: Thraupidae) en Cali, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 101-114. DOI: 10.17151/bccm.2017.21.2.7

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto102bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
introduCCiÓn
El Sicalis Coronado (Sicalis flaveola) es una especie principalmente granívora de la familia Thraupidae (KLICKA et al., 2007). Se distribuye en Suramérica, y se reconocen cinco subespecies: S. f. pelzeni (Argentina, Bolivia, Brasil, Paraguay, Uruguay y Chile), S. f. brasiliensis (Brasil), S. f. flaveola (Colombia y Venezuela), S. f. valida (Ecuador y Perú) y S. f. koenigi (Argentina) (RISING et al., 2011). En Colombia, su distribución inicial, se reporta en la región Caribe, oriente de los Andes desde Arauca hasta Meta y oriente del Vichada; pero luego fue reportada en el Valle del Cauca, como aves que escaparon de jaulas o expansión de su rango geográfico y los primeros registros datan de 1971 (HILTY, 1985).
Esta especie, común en áreas abiertas y semiabiertas con intervención humana, como parques y cultivos (MARCONDES-MACHADO, 1988), presenta una dieta principalmente granívora aunque incluye también frutos e insectos que obtiene en el suelo, estrato herbáceo, ramas y follaje hasta los 5 m de altura (ZOTTA, 1940; DE LA PEÑA & PENSEIRO, 2003; DE LA PEÑA, 2011). Se alimenta en grupos donde la mayoría de los miembros son juveniles (MARCONDES-MACHADO, 1997).
Los primeros aportes sobre biología reproductiva de la especie provienen de Brasil, a partir de cinco nidos en cautiverio con los cuales se determinó época reproductiva y tamaño de postura (BUSCHINELLI, 1971). Posterior a ésto, se encuentran trabajos sobre la biología reproductiva de la especie pero sólo en zonas templadas del sur como en Argentina, Chile y en áreas subtropicales de Brasil (MANSON, 1985; QUIROGA et al., 2006; PALMERIO & MASSONI, 2009). Es considerada una especie socialmente monógama, aunque se han reportado dos casos de poligínia en Brasil (MARCONDES-MACHADO, 1982). Construye su nido en forma de taza al interior de cavidades en árboles seniles, en estructuras antrópicas como lámparas,
Universidad del Valle in Cali, Colombia, between January 2013 and May 2015. Ninety-four nesting boxes were installed to provide additional nesting resource and four transects were done weekly in search of active nests and checking the nest boxes. Clutch size was 3.46 eggs, hatched chick average was three and gender proportion was 1:1. Incubation was carried out by the female while caring for the chicks was done by the couple. The reproductive success of the species at the moment of leaving the nest was 65.8%. Differences were found with the subspecies of southern latitudes of South America with a smaller range of posture and longer incubation and nesting stay periods. However, in aspects such as parental care during incubation and nesting of the chicks, they were similar.
Key words: clutch size, growth rate, latitude, life history, nesting boxes, urban ecology, Thraupidae.

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 103
al interior de nidos abandonados de otras especies (ejemplo: nidos de horneros) y en cajas nido (MANSON 1985; MARCONDES-MACHADO, 1988; PALMERIO & MASSONI, 2009). El tamaño de la nidada en Argentina y Chile puede variar entre 2 y 6 huevos (MANSON, 1985; DE LA PEÑA, 1995; QUIROGA et al., 2003; PALMERIO, 2012; OROZCO et al., 2016) y la época reproductiva en la zona templada del sur es entre octubre y marzo (PALMERIO, 2012).
A pesar de su amplia distribución en Suramérica y de la existencia de información sobre la reproducción de la especie en algunos países, aún se presentan vacíos sobre la biología y el comportamiento reproductivo en el extremo norte de Suramérica. En Colombia, por ejemplo, existen sólo algunas observaciones de nidos, avistamientos esporádicos y reporte de época reproductiva con tres nidos (HILTY & BROWN, 2001). En Venezuela sólo existe información sobre tamaño de la postura y huevos (RAMO & BUSTO, 1984; CRUZ & ANDREWS, 1989). Así, llenar los vacíos de información planteados permite contribuir a esclarecer conceptos sobre las estrategias de vida y presiones selectivas a las cuales se ven sometidas algunas de las especies en la región tropical. Trabajos sobre la historia natural de las especies son fundamentales para el entendimiento de su ecología conductual y ocupan, cada vez, un mayor posicionamiento en el campo de la ornitología debido a la implicación que tienen sus aportes en la conservación (STUTCHBURY & MORTON, 2001). A pesar de su importancia, aún es precario el conocimiento sobre algunas especies, en especial, aquellas más comunes que ocupan ambientes urbanos (MURGUI & HEDBLOM, 2017), como es el caso del Sicalis Coronado. El objetivo de este estudio es, por lo tanto, describir por primera vez los principales aspectos de la biología reproductiva de S. f. flaveola, en Cali, Colombia, con énfasis en estacionalidad de la época reproductiva, tamaño de postura, periodo de incubación, cuidado paternal y desarrollo de polluelos.
MÉtodos
Área de estudio
La investigación se llevó a cabo entre enero de 2013 y mayo de 2015 en el campus Meléndez de la Universidad del Valle (3°38’25,6’’ N, 76°19’13,6’’ W, 990 msnm), municipio de Santiago de Cali, departamento del Valle del Cauca, Colombia. De acuerdo con las características climáticas, la región se clasifica como Bosque Seco Tropical (bs-T), según el sistema de Holdridge (ESPINAL, 1967); con una temperatura promedio de 24,1°C y una humedad relativa promedio de 73%. La precipitación promedio anual es de 1471 mm, con dos picos de lluvia, entre marzo y mayo y de septiembre a noviembre (RIVERA, 2006). El campus tiene un área aproximada de 100 ha, de las cuales 9,5 ha están ocupadas por edificaciones, 44 ha por árboles, 45,5 ha por prados y pastizales y 1 ha por dos lagos (ARDILA-TÉLLEZ & CRUZ-BERNATE, 2014).

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto104bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
Métodos de campo
Época reproductiva. Se instalaron 94 cajas nido (dimensiones: 25 cm de alto, 13 cm de ancho y 10 cm de profundidad) con un orificio de entrada de 4 cm de diámetro y techo móvil para facilitar su revisión. Se ubicaron en toda el área de estudio, en árboles a 2 m de altura sobre el suelo, con una distancia mínima de separación entre ellas de 30 m y se revisaron cada tres días. El área de estudio se recorrió dos veces por semana, para un total de 232 recorridos adicionales, con el objeto de encontrar nidos silvestres. A estos nidos se les determinó la altura sobre el suelo y el volumen de la cavidad, teniendo en cuenta que correspondieron a nidos en lámparas o cercos con formas ortoédricas y cilíndricas. Se determinó el número de nidos activos (con huevos o polluelos) por mes, y se estableció la época reproductiva para la especie.
Postura, incubación y permanencia de los polluelos en el nido. Los nidos activos tuvieron seguimiento y observación diaria de dos a tres horas continuas a una distancia de 20 m entre las 8:00 y 18:00 h. Se registró la actividad que realizó cada miembro de la pareja durante dichos periodos de observación. De cada nido se registraron las variables: tamaño y medidas de la postura, duración del período de incubación, y permanencia de los polluelos en el nido (desde la eclosión del primer huevo hasta el abandono del nido por el último volantón). El período de incubación se estimó como el tiempo transcurrido desde que la hembra comienza a incubar hasta la eclosión del último huevo. Los huevos fueron medidos y pesados el mismo día en que fueron puestos. Se determinó el tipo de eclosión que presentaron los nidos; considerándose sincrónica aquella postura donde todos los huevos eclosionaron el mismo día y asincrónica cuando tomó varios días.
desarrollo de polluelos. Se registró la tasa de crecimiento de los polluelos, con base en la medición diaria de cuatro caracteres morfológicos desde el momento del nacimiento hasta el abandono del nido. Los caracteres morfológicos fueron: masa corporal medida con una balanza digital (± 0,01 g), largo del pico, tarso-metatarso y ala medidos con un calibrador digital (± 0,01 mm). La longitud del ala se tomó desde la articulación hasta el extremo de la novena pluma primaria.
anillamiento y éxito reproductivo. Para el reconocimiento individual, los adultos reproductivamente activos y los polluelos fueron marcados mediante anillos tarsales de colores dispuestos en combinaciones únicas. Los adultos se capturaron con redes de niebla instaladas en la vecindad inmediata de los nidos y con el uso de una puerta móvil instalada en la entrada de las cajas nido. El éxito reproductivo se midió como el número de polluelos que llegaron al momento de emplumar en relación con el total de polluelos nacidos, y el éxito de anidamiento se midió como el número de nidos en que al menos un polluelo alcanzó la edad de emplumar.

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 105
determinación molecular del sexo. La proporción secundaria de sexos de los polluelos producidos por la especie se determinó mediante el método molecular propuesto por ELLEGREN (1996). Para este análisis se tomaron muestras de sangre (10 µl) en polluelos y adultos de la vena braquial y se conservaron en “Tarjetas FTA” Whatman. Se empleó el lisado celular con la resina de intercambio iónico “Chelex 100” para liberar el ADN y se cuantificó con Thermo ScientificTM NanoDrop 2000. La PCR se realizó con base en lo descrito por FRIDOLFSSON & ELLEGREN (1999) utilizando los iniciadores 2550F - 2718R. Los productos amplificados fueron separados en geles de agarosa al 1% y coloreados con bromuro de etidio (SAMBROOK & RUSSELL, 2001).
análisis de datos. Para determinar si existían diferencias significativas en el número de nidos activos entre las temporadas reproductivas anuales se empleó una prueba de Bondad de ajuste (ZAR, 2010). El volumen de la cavidad donde fueron construidos los nidos silvestres se calculó con base en la fórmula matemática establecida para el cilindro y el ortoedro. Se calculó la ganancia de masa y talla diaria de los polluelos con la resta de las medidas tomadas entre días consecutivos. Se utilizó un modelo lineal generalizado mixto para analizar si el tiempo diario dedicado por las hembras al cubrimiento de los polluelos variaba significativamente con el transcurrir de los días. Las variables “día” (discreta) y “tiempo de permanencia en el nido” (continua) fueron consideradas factores fijos, mientras que la variable “nido” (discreta) fue aleatoria. Se consideró como “día 1” el día que comenzó la eclosión. Para el análisis de los datos se usó el programa estadístico R (R CORE TEAM, 2017).
resultados
nido
Se registraron en total 18 nidos (nueve en cajas nido y nueve en lámparas y cercos). La época reproductiva para la especie es continua durante el año, no se encontraron diferencias significativas en el número de nidos activos entre los meses de mayor reproducción (χ2 = 0,875 gl = 2, p = 0,001) y se observaron volantones vocalizando y solicitando alimento a lo largo del año (Fig. 1). El volumen de los nidos en lámparas y cercos varió de 654,23 cm3 a 14064,91 cm3 (media = 2790,15, DE = 4258,22, n = 9) y la altura sobre el suelo en que se hallaron fue de 1,63-2,94 m (media = 2,4 m, DE = 0,50, n = 9).
Postura
Los huevos fueron de color crema con vetas marrones, con un peso de 1,6-2,5 g (media = 2,1 g, DE = 0,23, n = 52), largo de 19,26-22,8 mm (media = 20,08 mm, DE = 1, n = 52) y ancho de 13-15,46 mm (media = 14,55 mm, DE = 0,63, n = 52). El tamaño de la postura fue de 3-4 huevos (media = 3,46, DE = 0,51, n = 15) (Fig. 2). La postura fue seriada, es decir que ponen un huevo por día.

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto106bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
Distribución de la frecuencia de nidos activos por mes del Sicalis Coronado (Sicalis faveola flaveola) durante el periodo enero 2013-mayo 2015 en el campus de la Universidad del Valle, Cali, suroccidente de Colombia.
figura 1.
Nido del Sicalis Coronado (Sicalis flaveola flaveola) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia. a) Taza y huevos. b) Utilización de cajas nido.
figura 2.
incubación
Según la eclosión, los huevos no se incuban siempre desde el primer día y la incubación puede comenzar cualquier día hasta que se pone el último huevo. Se observaron cinco nidos con eclosión sincrónica y seis con eclosión asincrónica. La duración del período de incubación fue de 14-19 días (media = 16, DE = 1, n = 6) y la hembra es la única encargada de esta actividad. Del tiempo de observación, la hembra dedicó el 53,1% a la incubación y el resto del tiempo el nido permaneció solo (Fig. 3a). El macho acompañó a la hembra durante la incubación, vocalizando cerca al nido, y también durante los vuelos que ella ejecutaba al llegar o salir del mismo.

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 107
Permanencia de polluelos en el nido
El cuidado de los polluelos (cubrimiento y alimentación) fue compartido parcialmente por los miembros de la pareja, ya que la hembra es la única que cubre los polluelos. Ambos miembros de la pareja alimentaron los polluelos, pero presentaron diferencias en sus patrones de inversión paternal. El tiempo que las hembras dedicaron a los polluelos en el interior del nido disminuyó significativamente a medida que avanzó el desarrollo de los polluelos (Tabla 1 y Fig. 3a). El número promedio de visitas al nido efectuadas por las hembras fue de 1,62±0,68 visitas/h, con duración de 1-68 min (media = 5, DE = 7), mientras que para los machos fue de 0,89±0,42 visitas/h (Fig. 3b), con una duración menor a un minuto. La permanencia de los polluelos en el nido fue de 15-19 días (media = 18,25, DE = 1, n = 8).
Incubación y cuidado de polluelos del Sicalis Coronado (Sicalis faveola flaveola) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia. a) Porcentaje de tiempo diario dedicado por la hembra a incubación y cuidado de polluelos. b) Promedio de visitas diarias por ambos miembros de la pareja durante la permanencia de polluelos en el nido.
figura 3.

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto108bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
Resultado del Modelo lineal generalizado mixto usado para evaluar variación del tiempo de permanencia en el nido que efectuó la hembra durante el cuidado de los polluelos de Sicalis Coronado (Sicalis faveola flaveola) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia
tabla 1.
Valor Error estándar Grados de libertad
t-valor p-valor
Intercepto 35 3,6 46 9,7 0,001Día -2,68 0,41 46 -6,47 0,001
desarrollo de polluelos
Los polluelos nacieron ciegos, con un fino plumón gris en las regiones lumbar, cefálica y humeral. En el cuarto día salieron los cañones de las primarias, el día siguiente abrieron los ojos y al séptimo día se observaron los cañones largos de las plumas de las alas (Fig. 4). El peso de los polluelos presentó un incremento de 1,37±0,19 g (n = 35) hasta el octavo día, después fue de 0,36±0,28 g/día. Los polluelos fueron capaces de volar fuera del nido desde el decimocuarto día de edad, cuando el largo de la novena primaria alcanzó 41,91±3,7 mm, y el peso fue de 15,04±1,25 g, que correspondieron al 62,14% y 69,18% de las medidas del adulto, respectivamente. El crecimiento del tarso fue de 1,38±0,38 mm/día hasta el noveno día, después fue de 0,16±0,19 mm/día y al momento de abandonar el nido el tarso midió 20,52±1,33 mm. El crecimiento del pico fue constante con una tasa de 0,35±0,15 mm durante todo el desarrollo; al momento de abandonar el nido el pico alcanzó 8,85±0,53 mm de largo (Fig. 5).
Éxito reproductivo y determinación molecular del sexo
En 18 nidos se encontraron 56 huevos, de los cuales 43 (76,7%) llegaron hasta el momento de la eclosión y 38 (67,8%) eclosionaron exitosamente. De los 18 nidos, cuatro (22,2%) no pasaron del periodo de incubación y en 11 nidos al menos un polluelo alcanzó la edad del primer vuelo, dando un éxito de anidamiento de 61,1%. El éxito reproductivo, medido como el número de polluelos que llegaron al momento del éxodo (25) en relación con el total de polluelos nacidos (38), fue del 65,8%.
Se determinó el sexo con métodos moleculares en la totalidad de los polluelos que llegaron a la edad de volantón (n = 16). Los fragmentos amplificados del intrón del gen CHD1 estaban dentro del rango de valores reportados en la literatura (CHD1Z = 600-650 pb y CHD1W = 400-450 pb) (Fig. 6). Se encontró que no existe una diferencia significativa en el número de polluelos que llegan a la edad del éxodo según el sexo (siete machos vs. nueve hembras); la proporción secundaria de sexos fue de 1:1 (χ2 = 0,25 gl = 1, p = 0,001).

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 109
Progresos morfológicos durante el crecimiento de los polluelos del Sicalis Coronado (Sicalis faveola flaveola) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia. a) Polluelo de un día de edad. b) Siete días. c) 10 días. d) 12 días. e) 15 días. f) Adulto.
figura 4.
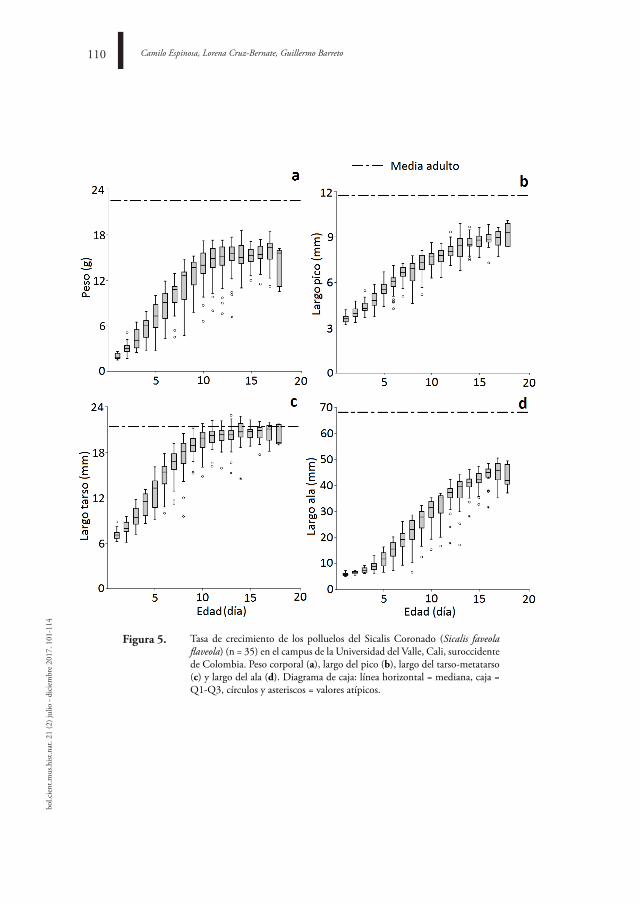
Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto110bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14 Tasa de crecimiento de los polluelos del Sicalis Coronado (Sicalis faveola flaveola) (n = 35) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia. Peso corporal (a), largo del pico (b), largo del tarso-metatarso (c) y largo del ala (d). Diagrama de caja: línea horizontal = mediana, caja = Q1-Q3, círculos y asteriscos = valores atípicos.
figura 5.

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 111
Gel de agarosa al 1% con los productos de amplificación del intrón comprendido entre el nucléotido 2550 y 2718 del gen CHD1 del Sicalis Coronado (Sicalis flaveola flaveola) en el campus de la Universidad del Valle, Cali, suroccidente de Colombia. De izquierda a derecha: marcador de peso molecular de 100 pb. H = hembras con dos bandas: CHD1Z y CHD1W. M = machos con una banda: CHD1Z.
figura 6.
disCusiÓn
En el caso del Sicalis Coronado, el tamaño de la nidada hallado en este estudio para la subespecie S. f. flaveola (3,46 huevos) es similar al reportado para la misma subespecie en Venezuela, en donde se halló un promedio de huevos por postura de 3,6 (CRUZ & ANDREWS, 1989) y 3,75 huevos (RAMO & BUSTO, 1984), con un rango de 3 a 5 huevos. Estos resultados difieren ligeramente de la subespecie S. f. pelzeni, para la cual se registró un tamaño de postura de 4,0 (QUIROGA et al., 2003), 5,0 (DE LA PEÑA, 1995), 3,87 (PALMERIO, 2012), 4,22 (MANSON, 1985) y 4,1 huevos (OROZCO et al., 2016), con rangos de 2 a 6 huevos. Los datos encontrados en este trabajo apoyan lo reportado en paserinos, donde el tamaño de las nidadas es menor en los trópicos que en zonas templadas (MARTIN et al., 2006), con posturas de 2-3 huevos en el neotrópico vs. 4-5 huevos en la zona templada del hemisferio norte (MARTIN, 1996). Diferencias en tamaños de nidada pueden reflejar variaciones en presiones selectivas que tienen las especies, además de poder influir en rasgos demográficos y de vulnerabilidad de las poblaciones (SAETHER et al., 1996; SAETHER & BAKKE, 2000).
Una explicación a la diferencia en el tamaño y el rango de las posturas, puede ser la duración de las estaciones en el hemisferio sur (WINKLER et al., 2014). Se conoce que, al inicio de la temporada reproductiva, existen mejores condiciones ambientales

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto112bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
y se incrementan las probabilidades de encontrar parejas disponibles de mejor calidad (YOUNG, 1994; HANSSON et al., 2000; WINKLER et al., 2014). El tamaño de las posturas tiende a variar a lo largo de la temporada reproductiva en las zonas templadas, donde el número de huevos por postura disminuye a medida que avanza la época reproductiva (WINKLER et al., 2014). En las especies del trópico las posturas no varían en gran medida, ya que las condiciones ambientales son relativamente estables. En S. f. flaveola se encontró que las posturas fueron de tres huevos entre enero y septiembre, y entre octubre y diciembre –la época de mayor pluviosidad donde las condiciones ambientales son las más favorables– las posturas fueron en su mayoría de cuatro huevos, evidenciando un esfuerzo reproductivo regulado a lo largo del año con una variación mínima.
El 28% de los nidos en S. f. flaveola (información de este estudio más datos no publicados) no pasaron el periodo de incubación debido a depredación, mientras que en S. f. pelzeni el porcentaje fue de 35% (PALMERIO, 2012). Estos valores, sugieren que las presiones sobre el nido, durante los períodos de postura e incubación en el Sicalis Coronado, se encuentran en un rango cercano entre el hemisferio sur y el trópico; lo que concuerda con lo reportado por MARTIN et al. (2006), quienes no encontraron diferencias significativas en la depredación de nidos entre el trópico, hemisferio sur y el hemisferio norte.
Los cuidados de los padres, en el periodo de incubación y de polluelos, traen un riesgo alto; generan un mayor gasto energético y exposición a los depredadores que comprometen la supervivencia de la pareja (GHALAMBOR & MARTIN, 2001; MARTIN, 2002). El período de incubación en S. flaveola para Colombia fue de 16 días, mientras que lo reportado para Argentina fue de 12,95 (n = 104) (PALMERIO & MASSONI, 2009, 2011), 13,3 (n = 34) (MANSON, 1985) y 16 días (n = 5) (QUIROGA et al., 2003), y en Chile de 11,1 días (n = 7) (OROZCO et al., 2016). Excepto por lo mencionado por QUIROGA et al. (2003), el período de incubación es más largo en el trópico y concuerda con lo encontrado en otras especies de paserinos (MARTIN, 2002). Esta diferencia se podría explicar por el menor tiempo dedicado a la incubación por parte de las hembras en el trópico (MARTIN, 2002). Se considera que existe una relación inversa entre el porcentaje diario de atención al nido por parte de los adultos y la duración total del periodo de incubación (GHALAMBOR & MARTIN, 2001; MARTIN, 2002). Aunque el riesgo sobre la nidada aumenta con periodos de incubación más largos en el trópico, se compensaría por la posibilidad de futuras reproducciones de los adultos (MARTIN, 2002), ya que las especies del trópico tienen una mayor esperanza de vida.
La duración del período de cuidado de los polluelos difiere entre las subespecies, en S. f. flaveola fue de 18,25 días mientras que en S. f. pelzeni fue de 14 (MANSON, 1985; PALMERIO & MASSONI, 2009; OROZCO et al., 2016). Esto apoyaría la

Biología reproductiva de Sicalis flaveola (aves: Thraupidae) en Cali, Colombia 113
hipótesis que algunas especies del trópico presentan períodos de cuidado de polluelos más largos que en las zonas templadas (STUTCHBURY & MORTON, 2001). Aparentemente, un período más largo de permanencia de los polluelos en el nido les confiere ventajas en términos de mayor tiempo para el crecimiento en la longitud del ala, que a su vez contribuirá con un mejor desempeño durante el vuelo después de emplumar (MARTIN, 2015). Aún con las diferencias encontradas entre el Sicalis Coronado de Argentina y Colombia, estas no se reflejan en el éxito de anidamiento; en S. f. pelzeni fue del 55% (PALMERIO, 2012) y para S. f. flaveola fue del 61,1%.
Las teorías clásicas consideran aspectos como: limitación de alimentos, depredación de nidos y la mortalidad de adultos, para explicar la variación en la biología reproductiva de las aves en diferentes latitudes (MARTIN, 2004). Sin embargo, la limitación de alimentos y la depredación no han explicado el tamaño de la postura de muchas especies tropicales (MARTIN, 1996, 2004; MARTIN el al., 2000). Los trabajos de historia natural se han centrado en las diferencias en el tamaño de la postura. Pero otros aspectos, como los aquí descritos, también deben continuarse en investigación para conocer los mecanismos que subyacen en estos procesos diferenciadores entre especies.
Se ha planteado que las especies del trópico son similares a las del hemisferio sur y difieren en las estrategias de vida de las especies del hemisferio norte. En el caso del Sicalis Coronado (S. f. flaveola) la incubación está a cargo de la hembra, y el cuidado de los polluelos antes del abandono del nido es efectuado por la pareja; similar a lo que ocurre con S. f. pelzeni. Pero también existen diferencias: la subespecie en el Valle del Cauca presenta un rango de postura menor, y períodos más largos de incubación y permanencia de polluelos en el nido.
aGradeCiMientos
A Delly Brigitte Tosse del Laboratorio de Genética Humana (LGMH) por su acompañamiento. A Humberto Álvarez Ph.D. por sus consejos y revisión del proyecto, a Wilmar Torres M.Sc. por su asesoría estadística. A Martín Quiroga Ph.D., Andrés Palmerio Ph.D., Sandra Valderrama Ph.D., Karpar Delhey Ph.D. y Vladimir Rojas Ph.D. que mejoraron la calidad del manuscrito. Al Departamento de Biología y al postgrado de Ciencias-Biología de la Universidad del Valle (Cali, Colombia) por su apoyo al proyecto.
referenCias
ARDILA-TÉLLEZ, J.D. & CRUZ-BERNATE, L., 2014.- Aspectos ecológicos de las aves migratorias neárticas en el campus de la Universidad del Valle. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 18: 93-108.
BUSCHINELLI, A., 1971.- Contribuição ao estudo da biologia do canário da terra. Cienc Cult. (São Paulo), 23: 163-166. CRUZ, A. & ANDREWS, R.W., 1989.- Observations on the breeding biology of passerines in a seasonally flooded savanna in
Venezuela. Wilson Bull., 101: 62-76.DE LA PEÑA, M.R., 1995.- Ciclo Reproductivo de las Aves Argentinas. Univ. Nacional Litoral, Santa Fe, Argentina.

Camilo espinosa, lorena Cruz-Bernate, Guillermo Barreto114bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
01-1
14
DE LA PEÑA, M.R., 2011.- Observaciones de campo en la alimentación de las aves. Rev. FAVe, 15: 99-107.DE LA PEÑA, R. & PENSEIRO, J.F., 2003.- Contribución de la flora en los hábitos alimentarios de las aves en un bosque del centro
de la provincia de Santa Fe, Argentina. ornitol Neotrop, 14: 499-513.ELLEGREN, H., 1996.- First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc
Biol Sci, 263 (1367): 1635-1641.ESPINAL, L.S., 1967.- Visión ecológica del departamento del Valle del Cauca. Univ. del Valle, Santiago de Cali, Colombia.FRIDOLFSSON, A.K. & ELLEGREN, H., 1999.- A simple and universal method for molecular sexing of non-ratite birds. J Avian
Biol, 30: 116-121.GHALAMBOR, C.K. & MARTIN, T.E., 2001.- Fecundity-survival trade-offs and parental risk-taking in birds. Science, 292: 494-497.HANSSON, B., BENSCH, S. & HASSELQUIST, D., 2000.- The quality and the timing hypotheses evaluated using data on great
reed warblers. oikos, 90: 575-581.HILTY, S.L., 1985.- Distributional changes in the Colombian avifauna: a preliminary blue list. ornithol. Monogr., 36: 1000-1012.HILTY, S.L. & BROWN, W.L., 2001.- Guía de las aves de Colombia. American Bird Conservancy, USA.KLICKA, J., BURNS, K. & SPELLMAN, G.M., 2007.- Defining a monophyletic Cardinalini: a molecular perspective. Molecular
phylogenetics and evolution, 45 (3): 1014-1032.MANSON, P., 1985.- The nesting biology of some passerines of Buenos Aires, Argentina. ornithol. Monogr., 36: 954-972.MARCONDES-MACHADO, L.O., 1982.- Poliginia em Sicalis flaveola brasiliensis (Gmelin, 1789) (Passeriformes, Emberizidae).
Rev. Bras. Zool., 1: 95-99.MARCONDES-MACHADO, L.O., 1988.- Experiência de repovoamento com Sicalis flaveola brasiliensis (Gmelin, 1789) (Passeriformes,
Emberizidae) em área destinada à pecuária leiteira. Rev. Bras. Zool., 5: 193-200. MARCONDES-MACHADO, L.O., 1997.- Comportamento social de Sicalis flaveola brasiliensis em cativeiro (Passeriformes,
Emberizidae). Iheringia, Sér. Zool., 82: 151-158.MARTIN, T.E., 1996.- Life history evolution in tropical and south temperate birds: what do we really know? J. Avian Biol., 27: 263-272.MARTIN, T.E., 2002.- A new view of avian life-history evolution tested on an incubation paradox. Proc Biol Sci, 269 (1488): 309-316.MARTIN, T.E., 2004.- Avian life-history evolution has an eminent past: does it have a bright future? Auk, 121: 289-301.MARTIN, T.E., 2015.- Age-related mortality explains life history strategies of tropical and temperate songbirds. Science, 349: 966-970.MARTIN, T.E., MARTIN, P.R., OLSON, C.R., HEIDINGER, B.J. & FONTAINE, J.J., 2000.- Parental care and clutch sizes in
North and South American birds. Science, 287 (5457): 1482-1485.MARTIN, T.E., BASSAR, R.D., BASSAR, S.K., FONTAINE, J.J., LLOYD, P., MATHEWSON, H.A., NIKLISON, A.M. &
CHALFOUN, A., 2006.- Life‐history and ecological correlates of geographic variation in egg and clutch mass among passerine species. evolution, 60: 390-398.
MURGUI, E. & HEDBLOM, M., 2017.- ecology and conservation of birds in urban environments. Springer, Cham, Switzerland.OROZCO, P.M, SANTILLÁN, M.Á., BRAGAGNOLO, L.A., REBOLLO, M.E., LÓPEZ, F.G. & MARTÍNEZ, P.A., 2016.-
Aportes a la biología reproductiva del Chirigüe Azafrán (Sicalis flaveola) en cajas nido en un bosque semiárido del centro de Argentina. Rev. Chil. ornitol., 22 (2): 165-170.
PALMERIO, A.G., 2012.- Maduración tardía del plumaje y costo reproductivo en el Jilguero Dorado Sicalis flaveola: Tesis, Univ. Buenos Aires, Buenos Aires, Argentina.
PALMERIO, A.G. & MASSONI, V., 2009.- Reproductive biology of female Saffron Finches does not differ by the plumage of the mate. Condor, 111: 715-721.
PALMERIO, A.G. & MASSONI, V., 2011.- Parental care does not vary with age‐dependent plumage in male Saffron Finches Sicalis flaveola. Ibis, 153: 421-424.
QUIROGA, M., DEL BARCO, O. & AGOSTELLI, F., 2003.- First approaches to de reproductive biology of Sicalis flaveola Birds: Emberizidae at the alluvial valley of Paraná River, Argentina. Rev. FAVe, 2: 35-40.
QUIROGA, M., DEL BARCO, O., SAIGO, G. & AGOSTELLI, F., 2006.- Biología reproductiva del Jilguero Dorado (Sicalis flaveola) (Aves: Emberizidae) en la Reserva Universitaria El Pozo El Pozo (Santa Fe, Argentina). Fabicib, 10: 61-68.
R CORE TEAM, 2017.- R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
RAMO, C. & BUSTO, B., 1984.- Nidificación de los passeriformes en los Llanos de Apure (Venezuela). Biotropica, 16 (1): 59-68.RISING, J., JARAMILLO, A., COPETE, J.L., MADGE, S. & RYAN, P., 2011.- Family Emberizidae (Buntings and New World
Sparrows): 428-876 (en) DEL HOYO, J., ELLIOT, A. & CHRISTIE A. (eds.) Handbook of the Birds of the World. Volume 16: Tanagers to New World Blackbirds. Lynx Editions, Barcelona.
RIVERA, H.F., 2006.- Composición y estructura de una comunidad de aves en un área suburbana en el occidente colombiano. ornitol. Colomb., 4: 22-38.
SAETHER, B.E. & BAKKE, Ø., 2000.- Avian life history variation and contribution of demographic traits to the population growth rate. ecology, 81 (3): 642-653.
SAETHER, B.E., RINGSBY, T.H. & RØSKAFT, E., 1996.- Life history variation, population processes and priorities in species conservation: towards a reunion of research paradigms. oikos, 77 (2): 217-226.
SAMBROOK, J. & RUSSELL, D.W., 2001.- Molecular cloning: A laboratory manual. Cold spring Harbor Laboratory Press, New York.STUTCHBURY, B.J. & MORTON, E.S., 2001.- Behavioral ecology of tropical birds. Academic Press, San Diego.YOUNG, B.E., 1994.- Geographic and seasonal patterns of clutch-size variation in house wrens. Auk, 111: 545-555.WINKLER, D.W., RINGELMAN, K.M., DUNN, P.O., WHITTINGHAM, L., HUSSELL, D.J., CLARK, R.G. et al., 2014.-
Latitudinal variation in clutch size–lay date regressions in Tachycineta swallows: effects of food supply or demography? ecography, 37: 670-678.
ZAR, J.H., 2010.- Biostatistical analysis. Prentice Hall, New Jersey.ZOTTA, A., 1940.- Lista sobre el contenido estomacal de las aves argentinas. Hornero, 7: 402-411.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
Bird asseMBle assoCiated to tWo landsCaPe units in a troPiCal drY forest*
Juan Pablo Sánchez-Montenegro1,2, Katherine Pérez-Castillo1,2, Mario Fernando Garcés-Restrepo2, Alan Giraldo2
abstract
The transformation of ecosystems through human activities can affect the biological and ecological processes of bird species, and can even lead to groups of species becoming isolated due to deforestation, causing a decrease in genetic diversity and the extirpation of species. To evaluate the composition and structure of bird communities associated with the tropical dry forest ecosystem in the Guarinocito jurisdiction (La Victoria, Caldas) we identified two landscape units: a forest area (dry forest matrix) and an interfered area (matrix of lands), presenting human intervention such as roads and grassland systems. Point counts of 10 min long were carried out in each unit to survey birds between 06:00 and 09:00 hours and between 16:00 and 18:00 hours. Mist nets were set up to complete the species record. Three four-day long sampling trips were carried out in April 2013, November 2013, and October 2014 to consider the seasonal variability that affects the environmental conditions in the study area. The sampling representativeness was evaluated using non-parametric estimates. All observed or captured individuals were assigned to a trophic guild in order to evaluate the community structure in the two established landscape units. A total of 1,005 individuals from 36 families and 127 species were recorded. The most abundant species were Thraupis episcopus (7.36%) and Sicalis flaveola (6.77%). Sample representativeness ranged from 68.18% (Jack 2) to 86.03% (Bootstrap). Although no significant differences between the two landscape units were detected (THutchinson=1.59, p=0.81), species diversity was higher in the forested area than in the intervened area. The dominant trophic guild in both landscape units was that of insectivorous birds. No significant differences in bird community trophic structure were found between the two landscape units (X2=14.60, p=0.33). The size, shape, or intervention history of the fragments could have resulted in a lack of differences in the structure and composition of the avifauna in the two landscape units.
Key words: Dry forest, birds, assemblage, composition, structure.
* FR: 25-XI-2016 . FA: 22-IV-20171 Universidad del Valle, Departamento de Biología. Programa Académico de Biología. Cali, Colombia. E-mail: [email protected], [email protected] Universidad del Valle, Departamento de Biología. Grupo de Investigación en Ecología Animal. Cali, Colombia. E-mail: [email protected], [email protected], ecología.animal@correounivalle. edu.co
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 115-130. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:SÁNCHEZ-MONTENEGRO, J.P., PÉREZ-CASTILLO, K., GARCÉS-RESTREPO, M.F. & GIRALDO, A., 2017.- Bird assemble associated to two landscape units in a tropical dry forest. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 115-130. DOI: 10.17151/bccm.2017.21.2.8

116 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
ensaMBle de aVes asoCiado a dos unidades de PaisaJe en un BosQue seCo troPiCal
resumen
La transformación de los ecosistemas por la actividad humana puede llegar a afectar los procesos biológicos y ecológicos de las especies de aves, incluso puede conllevar a que grupos de especies queden aisladas por matrices de desforestación, causando disminución en la diversidad genética y extirpaciones. Con el propósito de evaluar la composición y estructura de la comunidad de aves asociadas al ecosistema de bosque seco tropical en la vereda Guarinocito (La Victoria, Caldas), se identificaron dos unidades de paisaje: zona boscosa (matriz de bosque seco) y zona intervenida (matriz de terrenos), que presentan intervención humana, como carretera y sistemas de pastizales. En cada unidad se establecieron puntos de conteo de 10 minutos de duración en donde se realizaron censos de aves entre las 06:00 – 09:00 horas y entre las 16:00 – 18:00 horas, y se dispusieron redes de niebla para complementar el registro de especies. Se realizaron tres jornadas de muestreo con una duración de cuatro días durante abril de 2013, noviembre de 2013 y octubre de 2014 para incorporar la variación climática estacional que afecta las condiciones ambientales de la zona de estudio. Se evaluó la representatividad del muestreo mediante el uso de estimadores no paramétricos. Todos los individuos observados o capturados fueron asignados a un gremio trófico con el objetivo de evaluar la estructura de la comunidad entre las unidades de paisaje establecidas. Se registraron 1005 individuos de 36 familias y 127 especies, siendo Thraupis episcopus (7,36%), Sicalis flaveola (6,77%) las especies más abundantes. La representatividad del muestreo estuvo entre el 68,18% (Jack 2) y el 86,03% (Bootstrap). No se detectaron diferencias significativas en la diversidad entre las dos unidades de paisaje (THutchinson=1,59; p = 0,81), sin embargo la diversidad de especie fue mayor en la zona boscosa que en la zona intervenida. El gremio trófico dominante en ambas unidades de paisaje fue el de las aves insectívoras. No se encontraron diferencias significativas en la estructura trófica de la comunidad de aves entre las dos unidades de paisaje (X2=14,60; p=0,33). El tamaño, forma o historia de intervención de los fragmentos puede estar generando que no exista diferencia en estructura y composición en la avifauna en las diferentes unidades del paisaje.
Palabras clave: Bosque seco, aves, ensamblaje, composición, estructura.
introduCtion
Birds play important roles in ecosystems, such as in the control of plagues, pollination, and seed dispersal, and are therefore a taxonomic group of critical importance in natural restoration processes for forest conservation (SODHI et al., 2011). The ecological functions of birds are even more relevant now that habitat transformations due to human activities, such as forestry, mining, and agriculture, are some of the main threats to the biological diversity of the planet (JANTZ et al., 2003; SODHI et al., 2011; PATTEN & SMITH-PATTEN, 2012).

117Bird assemble associated to two landscape units in a tropical dry forest
One of the most threatened ecosystems in the tropics is dry forest (TDF), due to soil fertility issues (PORTILLO-QUINTERO et al., 2013; BANIN et al., 2014; CAO et al., 2015; TRILLERAS et al., 2015). Great swaths of this ecosystem have been transformed into cultivation areas and human settlements (JANZEN, 1988; ARCILA et al., 2012; MARINARO et al., 2014; GASPARRI et al., 2015). The valley of the upper Magdalena River, on the western and eastern foothills of the Eastern and Central cordilleras of the Colombian Andes, consists of tropical dry forest (HOLDRIDGE, 1967). This is an area of lowlands with notable ecosystem mosaics (ACOSTA-GALVIS et al., 2006).
The geographic valley of the Magdalena River, along with the tropical dry forests of the Caribbean region of Colombia, are considered focal areas of endemism for birds associated with the Neotropical dry forest (CRACRAFT, 1985; STOTZ et al., 1996; STATTERSFIELD et al., 1998; MYERS et al., 2000). Information on bird species of the tropical dry forests of Colombia has been based mainly on inventories and lists of birds from specific regions or locations (NARANJO, 1992; STILES et al., 1995; ESTELA & LÓPEZ-VICTORIA, 2005; LOSADA-PRADO et al., 2005. There are few studies that describe the ecology of the avifauna and species dominance, or specific analyses of the bird community of this ecosystem (e.g. LOSADA-PRADO & MOLINA-MARTÍNEZ, 2012).
The composition and structure of bird communities is influenced by historical processes that have marked their geographical distribution (GASTON & BLACKBURN, 2000), as well as by recent ecological processes such as human disturbance (SODHI et al., 2011). The latter factor influences habitat preferences, behavior, and the reproductive success of species. However, response differs depending on the level of transformation and on the structure and composition of remaining habitats (DURÁN & KATTAN, 2005). When the levels of alteration are elevated, the remaining forest cover does not differ in structure and composition from the human matrix (RENGIFO, 1999).
The present study was carried out to evaluate and compare the composition, diversity, and trophic structure of the bird assemblage in two landscape units of a tropical dry forest located in the jurisdiction of Guarinocito, La Victoria, Caldas, Colombia. The objective was to identify whether the habitat modifications caused by human intervention (mining and livestock farming) have caused a decrease in the structure and composition of the bird community associated with the remaining forest cover.
Materials and MetHods
Three sampling periods of five days each, were carried out between April 2013 and October 2014. Sampling occurred in the “Hacienda La Española” (5°21´51.9´´ N, 74°47´45.7´´ W), jurisdiction of Guarinocito, municipality of La Victoria, Caldas, Colombia (Figure 1). The area corresponds to tropical dry forest, typical of the

118 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
Magdalena Valley (DEVIA et al., 2003). The altitude above sea level ranges between 240 and 280 m. There is a bimodal precipitation regime, with four seasons corresponding to two rainy seasons (May-June and September-November) and two dry seasons (January-April and July-August). The average annual precipitation is 1590 mm and the average temperature is 28.2ºC (CORPOCALDAS, 2001).
Geographical localization of Hacienda La Española in Caldas department. (Source: Google Earth Pro ®).
figure 1.
Two contrasting landscape units were identified in the study area. One intervened area comprised 75 ha destined to livestock farming) with a vegetation dominated by shrubs and grasses, wetlands (swamps and oxbow lakes), and mining activities (stone extraction and artisanal gold extraction), and a 90 ha of forested area corresponding to dry forest fragments that are in natural regeneration.
The avifauna present in each landscape unit was evaluated using point counts and mist netting. The sampling effort was equal in each unit, with the same number of point counts and mist nets. Sampling was carried out twice per day, between 06:00 and 10:00, and between 15:30 and 18:30. Each point count consisted of an observer using binoculars and recording birds seen within a previously established 50 m radius during 10 min. Point count stations were located 100 to 200 m apart (RALPH et

119Bird assemble associated to two landscape units in a tropical dry forest
al., 1996). The species, number of individuals, and type of substrate on which the observed specimen was at the time of observation were recorded. Mist nets were 12 m long by 3 m high with 16 mm mesh size, and were used as a complementary method to increase the probability of recording fast-flying species. Three double nets were located randomly within the study area each sampling day, covering a total of 72 m2. Nets were checked every hour from 07:00 until 18:00 hours. Specimen identification was carried out in the field following the taxonomic guides of HILTY & BROWN (1986) and considering the geographic distribution described by the IUCN (2014).
A sampling unit was defined as all bird records obtained during two points counts as well as records from mist nets, establishing in this way 30 sampling units for each landscape unit during the entire study period. Species accumulation curves were constructed using this information. Sample representativeness was determined based on the species richness estimators Chao2, Bootstrap, and first- and second-order Jackknife (MORENO, 2001), which were calculated using the program Estimates® v8.2.0. (COLWELL, 2012). The Shannon entropy was calculated as 1D=exp(H’) (JOST, 2006) to obtain the true diversity value, and Simpson´s dominance index (Ds) was calculated as D_s=1-λ (BROWER & ZAR, 1984) for each landscape unit. The diversity between landscape units was compared with a t test modified by Hutcheson (tH) (HUTCHESON, 1970; MAGURRAN, 2004), and dominance was compared using a computational resampling technique “Bootstrap” (KREBS, 1999). Rarefaction curves were also constructed to compare species richness between landscape units. Abundance data was transformed using the function log10(x+1) to reduce the effect of very abundant species, and the degree of similarity in assemblage structure was compared using the Bray-Curtis index calculated from the transformed abundance data. These analyses were carried out using the software PAST® version 2.17 (HAMMER et al., 2001).
The trophic structure analysis was performed by classifying each species according to its diet. A total of 11 trophic guilds were identified as follows. F: frugivores, I: insectivores, N: nectarivores, S: seed-eaters, C: carnivores, FI: frugivores and insectivores, FS: frugivores and seed-eaters, NI: nectarivorous and insectivores, CI: carnivores and insectivores, FNI: frugivores, nectarivores, and insectivores, FCI: frugivores, carnivores, and insectivores, and SIF: seed-eaters, insectivores, and frugivores. The omnivorous guild was ignored as it does not provide a clear representation of resource use (RIVERA-GUTIÉRREZ, 2006). Although trophic guild classification was performed following guidelines by PEARSON (1975) and STILES & ROSSELLI (1998), food capture techniques or foraging height were not considered. The proportion of each trophic guild within each landscape unit was calculated, and landscape units were compared by performing a contingency table analysis using a Pearson´s X2 test (PLACKETT, 1983).

120 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
results
A total of 122 points counts were performed and 4,392 m of mist nets were installed. This sampling effort resulted in 1,005 records, corresponding to 127 species distributed in 18 orders and 42 families. The most abundant species were Thraupis episcopus (7.5%), Sicalis flaveola (6.8%), Forpus conspicillatus (5.4%), Pitangus sulphuratus (3.6%), Columbina talpacoti (3.4%), egretta thula (3.1%), and Tyrannus melancholicus (2.5%).
There was a total of 722 birds detected in the intervened area, belonging to 94 species, 16 orders, and 33 families. The following species had the highest number of records in this landscape unit: S. flaveola (9.4%), T. episcopus (8.2%), F. conspicillatus (6.78%), C. talpacoti (4.6%), and e. thula (4.3%). There was a total of 283 birds detected in the forested area, belonging to 79 species, 15 orders, and 31 families. The following species had the highest number of records in this landscape unit: Myiothlypis fulvicauda (17.6%), Coereba flaveola (5.6%), T. episcopus (5.3%), Catharus ustulatus (4.6%), and Troglodytes aedon (4.2%) (Table 1).
Of the 127 species recorded in the study area, 49 species were recorded only in the intervened landscape unit, of which six species were migratory (Himantopus mexicanus, Hirundo rustica, Machetornis rixosa, Porphyrio martinicus, and Tringa flavipes), and one species was endemic (Myiarchus apicalis). On the other hand, 34 species were recorded only in the forested landscape unit, of which only Parkesia noveboracensis was migratory. One migratory species was recorded in both the intervened and forested areas (Actitis macularia), and a vulnerable endemic species (ortalis columbiana) was recorded in both landscape units (Table 1).
Between 68.2% and 86.0% of potential bird species from the Hacienda La Española were recorded during sampling. Between 65.7% and 85.4% of potential species were recorded in the intervened landscape unit, whereas between 58.9 and 82.8% of potential species were recorded in the forested landscape unit (Table 2). There were no significant differences between the two landscape units in the degree of uncertainty associated with the identity of the species to which a randomly selected individual was assigned (THutchinson=1.59, p=0.81). However, species richness was lower in the forested area (79 species) than in the intervened area (95 species), and dominance was slightly higher in the intervened area (p value Ds bootstrap = 0.029). True diversity was 50.9 effective species in the forested area, whereas true diversity was 45.6 effective species in the intervened landscape unit. The true diversity estimate for the entire study area was 62.2 effective species (Table 3), with a degree of similarity between the two units in assemblage composition of 43.47%.
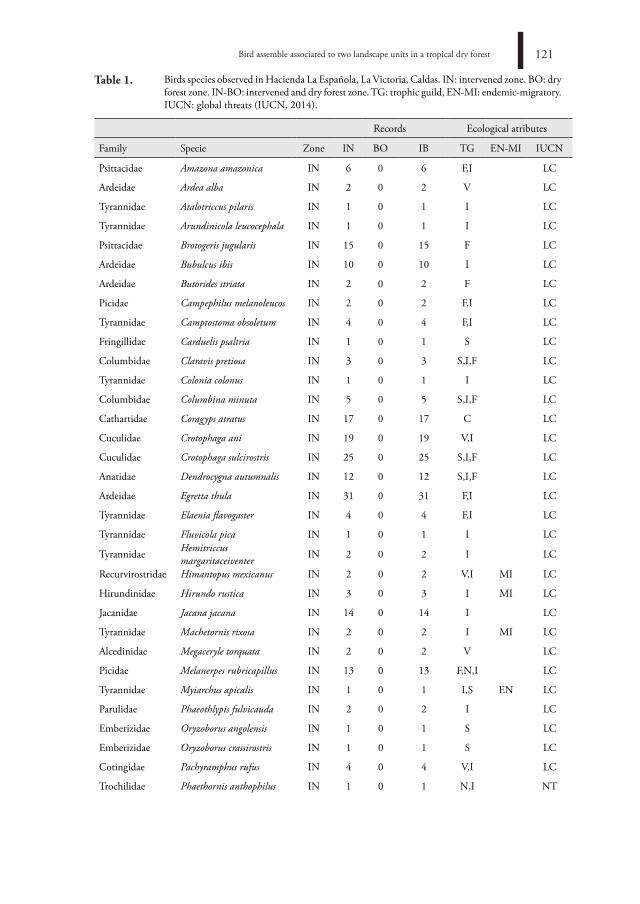
121Bird assemble associated to two landscape units in a tropical dry forest
Birds species observed in Hacienda La Española, La Victoria, Caldas. IN: intervened zone. BO: dry forest zone. IN-BO: intervened and dry forest zone. TG: trophic guild, EN-MI: endemic-migratory. IUCN: global threats (IUCN, 2014).
table 1.
Records Ecological atributes
Family Specie Zone IN BO IB TG EN-MI IUCN
Psittacidae Amazona amazonica IN 6 0 6 F,I LC
Ardeidae Ardea alba IN 2 0 2 V LC
Tyrannidae Atalotriccus pilaris IN 1 0 1 I LC
Tyrannidae Arundinicola leucocephala IN 1 0 1 I LC
Psittacidae Brotogeris jugularis IN 15 0 15 F LC
Ardeidae Bubulcus ibis IN 10 0 10 I LC
Ardeidae Butorides striata IN 2 0 2 F LC
Picidae Campephilus melanoleucos IN 2 0 2 F,I LC
Tyrannidae Camptostoma obsoletum IN 4 0 4 F,I LC
Fringillidae Carduelis psaltria IN 1 0 1 S LC
Columbidae Claravis pretiosa IN 3 0 3 S,I,F LC
Tyrannidae Colonia colonus IN 1 0 1 I LC
Columbidae Columbina minuta IN 5 0 5 S,I,F LC
Cathartidae Coragyps atratus IN 17 0 17 C LC
Cuculidae Crotophaga ani IN 19 0 19 V,I LC
Cuculidae Crotophaga sulcirostris IN 25 0 25 S,I,F LC
Anatidae Dendrocygna autumnalis IN 12 0 12 S,I,F LC
Ardeidae egretta thula IN 31 0 31 F,I LC
Tyrannidae elaenia flavogaster IN 4 0 4 F,I LC
Tyrannidae Fluvicola pica IN 1 0 1 I LC
Tyrannidae Hemitriccus margaritaceiventer IN 2 0 2 I LC
Recurvirostridae Himantopus mexicanus IN 2 0 2 V,I MI LC
Hirundinidae Hirundo rustica IN 3 0 3 I MI LC
Jacanidae Jacana jacana IN 14 0 14 I LC
Tyrannidae Machetornis rixosa IN 2 0 2 I MI LC
Alcedinidae Megaceryle torquata IN 2 0 2 V LC
Picidae Melanerpes rubricapillus IN 13 0 13 F,N,I LC
Tyrannidae Myiarchus apicalis IN 1 0 1 I,S EN LC
Parulidae Phaeothlypis fulvicauda IN 2 0 2 I LC
Emberizidae oryzoborus angolensis IN 1 0 1 S LC
Emberizidae oryzoborus crassirostris IN 1 0 1 S LC
Cotingidae Pachyramphus rufus IN 4 0 4 V,I LC
Trochilidae Phaethornis anthophilus IN 1 0 1 N,I NT
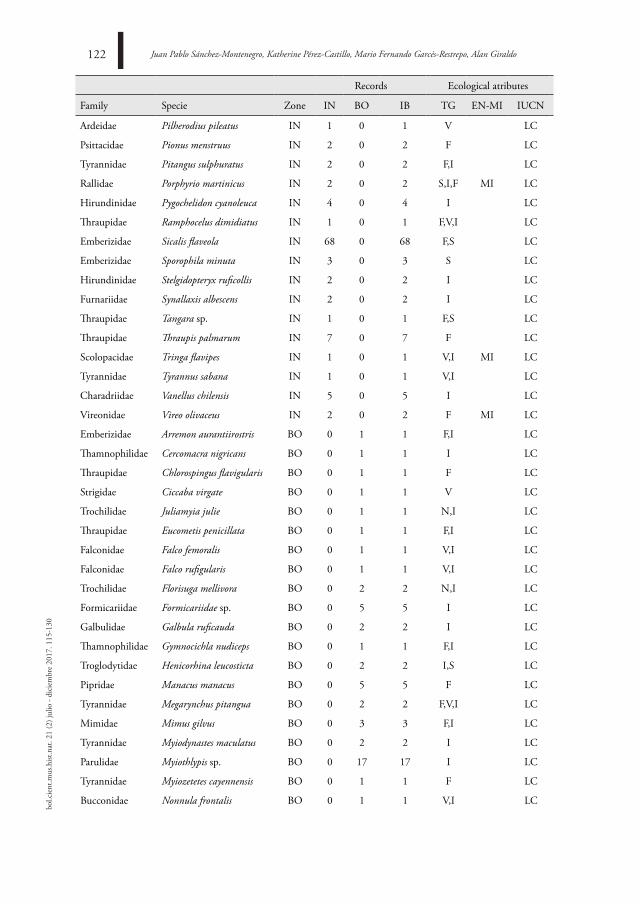
122 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
Records Ecological atributes
Family Specie Zone IN BO IB TG EN-MI IUCN
Ardeidae Pilherodius pileatus IN 1 0 1 V LC
Psittacidae Pionus menstruus IN 2 0 2 F LC
Tyrannidae Pitangus sulphuratus IN 2 0 2 F,I LC
Rallidae Porphyrio martinicus IN 2 0 2 S,I,F MI LC
Hirundinidae Pygochelidon cyanoleuca IN 4 0 4 I LC
Thraupidae Ramphocelus dimidiatus IN 1 0 1 F,V,I LC
Emberizidae Sicalis flaveola IN 68 0 68 F,S LC
Emberizidae Sporophila minuta IN 3 0 3 S LC
Hirundinidae Stelgidopteryx ruficollis IN 2 0 2 I LC
Furnariidae Synallaxis albescens IN 2 0 2 I LC
Thraupidae Tangara sp. IN 1 0 1 F,S LC
Thraupidae Thraupis palmarum IN 7 0 7 F LC
Scolopacidae Tringa flavipes IN 1 0 1 V,I MI LC
Tyrannidae Tyrannus sabana IN 1 0 1 V,I LC
Charadriidae Vanellus chilensis IN 5 0 5 I LC
Vireonidae Vireo olivaceus IN 2 0 2 F MI LC
Emberizidae Arremon aurantiirostris BO 0 1 1 F,I LC
Thamnophilidae Cercomacra nigricans BO 0 1 1 I LC
Thraupidae Chlorospingus flavigularis BO 0 1 1 F LC
Strigidae Ciccaba virgate BO 0 1 1 V LC
Trochilidae Juliamyia julie BO 0 1 1 N,I LC
Thraupidae eucometis penicillata BO 0 1 1 F,I LC
Falconidae Falco femoralis BO 0 1 1 V,I LC
Falconidae Falco rufigularis BO 0 1 1 V,I LC
Trochilidae Florisuga mellivora BO 0 2 2 N,I LC
Formicariidae Formicariidae sp. BO 0 5 5 I LC
Galbulidae Galbula ruficauda BO 0 2 2 I LC
Thamnophilidae Gymnocichla nudiceps BO 0 1 1 F,I LC
Troglodytidae Henicorhina leucosticta BO 0 2 2 I,S LC
Pipridae Manacus manacus BO 0 5 5 F LC
Tyrannidae Megarynchus pitangua BO 0 2 2 F,V,I LC
Mimidae Mimus gilvus BO 0 3 3 F,I LC
Tyrannidae Myiodynastes maculatus BO 0 2 2 I LC
Parulidae Myiothlypis sp. BO 0 17 17 I LC
Tyrannidae Myiozetetes cayennensis BO 0 1 1 F LC
Bucconidae Nonnula frontalis BO 0 1 1 V,I LC
123Bird assemble associated to two landscape units in a tropical dry forest
Records Ecological atributes
Family Specie Zone IN BO IB TG EN-MI IUCN
Caprimulgidae Nyctidromus albicollis BO 0 2 2 I LC
Rallidae Pardirallus maculatus BO 0 1 1 V,I LC
Columbidae Patagioenas subvinacea BO 0 2 2 S LC
Trochilidae Phaethornis striigularis BO 0 1 1 N,I LC
Trochilidae Phaethornis longirostris BO 0 1 1 N,I LC
Tyrannidae Phyllomyias nigrocapillus BO 0 1 1 I LC
Tyrannidae Poecilotriccus sylvia BO 0 1 1 I LC
Ramphastidae Pteroglossus torquatus BO 0 1 1 F,V,I LC
Cardinalidae Saltator striatipectus BO 0 1 1 F,I LC
Parulidae Parkesia noveboracensis BO 0 1 1 I MI LC
Troglodytidae Thryothorus leucotis BO 0 2 2 I LC
Tyrannidae Tyrannus tyrannus BO 0 4 4 F,I LC
Dendrocolaptidae Xiphorhynchus guttatus BO 0 1 1 I LC
Columbidae Zenaida auriculata BO 0 2 2 S LC
Scolopacidae Actitis macularius IN-BO 2 1 3 I MI LC
Trochilidae Amazilia saucerrottei IN-BO 1 3 4 N,I LC
Trochilidae Amazilia tzacatl IN-BO 1 2 3 N,I LC
Psittacidae Amazona autumnalis IN-BO 2 2 4 F,I LC
Parulidae Basileuterus rufifrons IN-BO 1 2 3 S,I,F LC
Cathartidae Cathartes aura IN-BO 3 1 4 S LC
Turdidae Catharus ustulatus IN-BO 3 13 16 C LC
Alcedinidae Chloroceryle amazona IN-BO 1 8 9 V LC
Alcedinidae Chloroceryle americana IN-BO 2 1 3 V LC
Trochilidae Chlorostilbon mellisugus IN-BO 3 5 8 N,I LC
Coerebidae Coereba flaveola IN-BO 5 16 21 N LC
Picidae Colaptes punctigula IN-BO 2 2 4 I LC
Columbidae Columbina passerina IN-BO 10 4 14 S,I,F LC
Columbidae Columbina talpacoti IN-BO 33 1 34 S,I,F LC
Cuculidae Crotophaga major IN-BO 15 5 20 V,I LC
Tinamidae Crypturellus soui IN-BO 3 10 13 F LC
Cuculidae Cyanocorax affinis IN-BO 2 1 3 F LC
Cuculidae Cyanolyca pulchra IN-BO 12 1 13 F,V,I LC
Thraupidae euphonia laniirostris IN-BO 24 3 27 F,I LC
Psittacidae Forpus conspicillatus IN-BO 49 5 54 F,I LC
Columbidae Geotrygon montana IN-BO 1 1 2 S LC
Trochilidae Glaucis hirsutus IN-BO 2 11 13 N LC
124 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
Records Ecological atributes
Family Specie Zone IN BO IB TG EN-MI IUCN
Icteridae Icterus nigrogularis IN-BO 1 1 2 F,I LC
Columbidae leptotila verrauxi IN-BO 8 2 10 F LC
Falconidae Milvago chimachima IN-BO 5 2 7 V LC
Tyrannidae Mionectes oleagineus IN-BO 6 3 9 F LC
Tyrannidae Mionectes olivaceus IN-BO 4 8 12 F LC
Thamnophilidae Myrmornis torquata IN-BO 16 2 18 I,S LC
Cracidae ortalis columbiana IN-BO 4 5 9 F EN VU
Columbidae Patagioenas cayennensis IN-BO 13 9 22 F LC
Tyrannidae Phaeomyias murina IN-BO 1 4 5 V,I LC
Cuculidae Piaya cayana IN-BO 13 10 23 I LC
Cardinalidae Piranga rubra IN-BO 29 7 36 F,I LC
Accipitridae Rupornis magnirostris IN-BO 3 3 6 V LC
Thamnophilidae Thamnophilus multistriatus IN-BO 1 1 2 V,I LC
Thraupidae Tangara inornata IN-BO 12 2 14 F,S LC
Cuculidae Tapera naevia IN-BO 1 2 3 I LC
Thraupidae Thraupis episcopus IN-BO 59 15 74 F LC
Tyrannidae Todirostrum cinereum IN-BO 9 5 14 I LC
Troglodytidae Troglodytes aedon IN-BO 6 12 18 I LC
Turdidae Turdus ignobilis IN-BO 4 11 15 F,I LC
Tyrannidae Tyrannus melancholicus IN-BO 23 2 25 F,I LC
Emberizidae Volatinia jacarina IN-BO 8 4 12 F,S LC
Tyrannidae Zimmerius viridiflavus IN-BO 2 1 3 I LC
Efficiency of sampling effort in Hacienda La Española (HE) and each landscape unittable 2.
HE %
BO %
IN%
Chao 2 79.90 69.90 77.44
Jack 1 74.46 68.24 73.19
Jack 2 68.12 58.89 65.75
Bootstrap 86.03 82.76 85.46
BO: dry forest. IN: intervened zone.
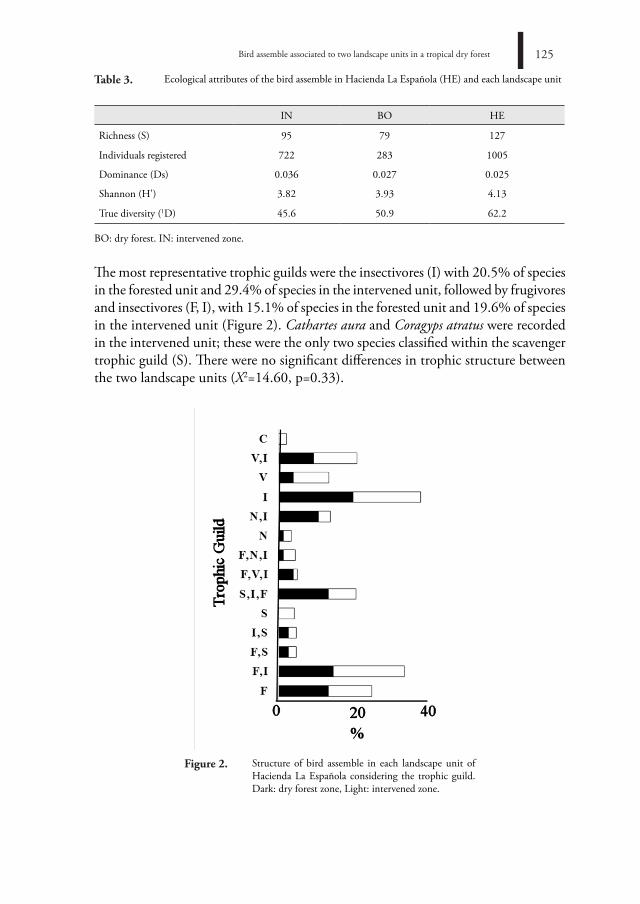
125Bird assemble associated to two landscape units in a tropical dry forest
Ecological attributes of the bird assemble in Hacienda La Española (HE) and each landscape unit table 3.
IN BO HE
Richness (S) 95 79 127
Individuals registered 722 283 1005
Dominance (Ds) 0.036 0.027 0.025
Shannon (H') 3.82 3.93 4.13
True diversity (1D) 45.6 50.9 62.2
BO: dry forest. IN: intervened zone.
The most representative trophic guilds were the insectivores (I) with 20.5% of species in the forested unit and 29.4% of species in the intervened unit, followed by frugivores and insectivores (F, I), with 15.1% of species in the forested unit and 19.6% of species in the intervened unit (Figure 2). Cathartes aura and Coragyps atratus were recorded in the intervened unit; these were the only two species classified within the scavenger trophic guild (S). There were no significant differences in trophic structure between the two landscape units (X2=14.60, p=0.33).
Structure of bird assemble in each landscape unit of Hacienda La Española considering the trophic guild. Dark: dry forest zone, Light: intervened zone.
figure 2.

126 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
disCussion
Despite being mobile organisms, birds are sensitive to changes in habitat structure (CLEARY et al., 2005; DÍAZ et al., 2005). For this reason, they constitute excellent models to understand the response of wild animal communities to environmental modifications, especially those generated by human activities (MACGREGOR-FORS & SCHONDUBE, 2011). The ability of birds to move through landscapes probably results in the persistence of populations in fragmented habitats such as those found in the study area, and can modulate the structure and composition of bird assemblages at a given location (GILLIES & CLAIRE, 2010).
The dry forest fragments found in the Hacienda la Española are small and far from each other (VARGAS-FIGUEROA et al., 2016), and birds probably move between fragments looking for the food resources they need (e.g. MERRIAM, 1984; BAUDRY & MERRIAM, 1988; OPDAM, 1990; TAYLOR et al., 1993; BEIER & NOSS, 1998; RENGIFO, 1999; FAHRIG, 2003; URIARTE et al., 2011). Due to the size and shape of fragments, there could be an enhanced edge effect, which facilitates the occurrence of generalist species in the forest, and would result in bird assemblages with similar diversity attributes (BENETTE & SAUNDERS., 2010; MORTELLITI et al., 2010; PINO et al., 2010; SMITH et al., 2011).
Other elements that can modulate local diversity of bird species in a fragmented habitat such as the dry forest of the Hacienda la Española are microclimatic conditions and habitat structural complexity. According to COLLINGE (1996), when habitats become fragmented the microclimatic conditions are altered drastically, affecting significantly the plant and animal communities. These microclimatic conditions are generally defined by the vegetation structure and local topography (SUGGITT et al., 2011). In thermal conditions, such as those found in the study area, the expected microclimatic differences between forested and intervened areas can be a determinant factor for bird diversity, as suggested by PATTENT et al. (2012) in a study carried out in lowland forests of Belize and Costa Rica.
Habitat structural complexity tends to decrease in fragmented forests due to the use of tree species from the understory by local residents, which leads to a simplification of the habitat that affects local species diversity (TEWS et al., 2004). Plant species typical of open areas were observed inside the forest fragments of the study area, which suggests that these fragments are a mix of the original forest and secondary growth (RENGIFO, 1999; DURÁN & KATTAN, 2005), where habitat structure has been simplified.
It has been widely demonstrated that the establishment of livestock farming and/or mining exploitation within dry forest matrices has a negative effect on the abundance and structure of bird assemblages (NARANJO, 1992; SAAD & PETIT, 1992; ESTRADA

127Bird assemble associated to two landscape units in a tropical dry forest
& COATES-ESTRADA, 2005; TSCHARNTKE et al., 2008). Areas with livestock farming and mining exploitation are inadequate habitats for forest species, due to their low structural complexity and low availability of food resources (MARTIN, 1984; ESTRADA et al., 1997; HUGHES et al., 2002; TSCHARNTKE et al., 2008). The bird assemblages that are found in landscapes under pressure from human activities usually lack species that are sensitive to perturbations, as those species are usually the first to become extinct locally due to processes caused by humans such as the transformation or degradation of the habitat (STOTZ et al., 1996; PETIT & PETIT, 2003).
The lack of difference in trophic structure between the assemblages of the two evaluated landscape units in the Hacienda La Española is probably the result of a lack of difference in food resource availability, or of the species using both landscape units to obtain resources (e.g. BLAKE & LOISELLE, 2001; HERZOG & KESSLER, 2002). The differential effect of species recorded in only one landscape unit would therefore be reduced, because the feeding preferences of species that use both landscape units would result in a decrease in the functional differences of the bird assemblages found in the two units. However, it should be noted that despite a lack of differences in diversity or trophic composition between the assemblages, the similarity percentage was low (<50%), which suggests that there are differences in structure between the assemblages.
Moreover, the species recorded in the Hacienda La Española during this study cor-respond to species frequently found in areas with high human perturbation, and the presence of threatened species was low. It is therefore necessary to incorporate other variables in future studies, such as the climatic season or the migratory patterns of spe-cies (SKIRVIN, 1981; PETIT & PETIT, 2003; MOORE et al., 2004; GÓMEZ et al., 2015), information that would provide a better idea of the effect of human perturbation on the dry forests of the Magdalena Valley. This kind of evidence is a key factor for the planning of conservation strategies for the tropical dry forest remnants, one of the natural elements of highest conservation interest for national as well as international scientific communities in the Neotropic region (CEBALLOS & GARCÍA, 1995; TURNER & CORLETT, 1996; SÁNCHEZ-AZOFEIFA et al., 2005; PORTILLO-QUINTERO & SÁNCHEZ-AZOFEIFA, 2010; GARCÍA et al., 2014; PRIETO-TORRES et al., 2016).
aCKnoWledGeMent
We thank Santiago García Revelo for his great help during field work. Wilmar Bolívar and Ángela María González provided complementary information on the environmental conditions in the study area. This study was co-financed by the Biology Department of the Universidad del Valle and by the research project “Implementación de indicadores biológicos como herramienta para evaluar cambios en la integridad ecológica de los fragmentos de bosque seco tropical presentes en Victoria - La Dorada, Caldas” CI7945, Univalle-Corpocaldas-Ecopetrol (Convenio de colaboración No. 5212085/2013).

128 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
referenCes
ACOSTA-GALVIS, A.R., HUERTAS-SALGADO, C. & RADA, M., 2006.- Aproximación al conocimiento de los anfibios en una localidad del Magdalena medio (Departamento de Caldas) Colombia. Revista de la Academia Colombiana de Ciencias exactas, Físicas y Naturales, 30 (115): 291- 303.
ARCILA, A.M., VALDERRAMA, C. & CHACÓN, P., 2012.- Estado de la fragmentación del bosque seco de la cuenca alta del río Cauca. Biota Colombiana, 13 (2): 86-101.
BANIN, L., LEWIS, S.L., LÓPEZ‐GONZÁLEZ, G., BAKER, T.R., QUESADA, S.A., CHAO, K.-J., et al., 2014.- Tropical forest wood production: a cross‐continental comparison. Journal of ecology, 102 (4): 1025-1037.
BAUDRY, J. & MERRIAM, G., 1988.- Connectivity and connectedness: functional versus structural patterns in landscapes: 23-28 (in) SCHREIBER, K.F. (ed.) Connectivity in landscape ecology. Proceedings of the 2nd International Seminar of the “International Association for Landscape Ecology.” Miinstersche Geographische Arbeiten 29, Miinster, Germany.
BEIER, P. & NOSS, R., 1998.- Do habitat corridors provide connectivity? Conservation biology, 12: 1241-1252.BENNETE, A.F. & SAUNDERS, D.A., 2010. Habitat fragmentation and landscape change. 88-106. In: SODHI, H. S. & EHRLICH,
P. R. (eds) Conservation Biology for All . Oxford University Press, Oxford. BLAKE, J.G. & LOISELLE, B.A., 2001.- Bird assemblages in second-growth and old-growth forests, Costa Rica: perspectives from
mist nets and point counts. Auk, 118 (2): 304-326. BROWER, J.E. & ZAR, J.H., 1984.- Field and laboratory Methods for general ecology. 2a. Ed. WCB Publishers.CAO, S., YU, Q., SÁNCHEZ-AZOFEIFA, A., FENG, J., RIVARD, B. & GU, Z., 2015.- Mapping tropical dry forest succession
using multiple criteria spectral mixture analysis. ISPRS Journal of Photogrammetry and Remote Sensing, 109: 17-29.CEBALLOS, G. & GARCÍA, A., 1995.- Conserving neotropical biodiversity: the role of dry forests in western Mexico. Conservation
biology, 9 (6): 1349-1353.CLEARY, D.F.R., GENNER, M.J., BOYLE, T.J.B., SETYAWATI, T., ANGRAETI, C.D. & MENKEN, S.B.J., 2005.- Associations
of bird species richness and community composition with local and landscape-scale environmental factors in Borneo. landscape ecology, 20: 989-1001.
COLWELL, R., 2012.- User’s guide to estimates. http:// viceroy.eeb.uconn.edu/estimatesCOLLINGE, S.K., 1996.- Ecological consequences of habitat fragmentation: implications for landscape architecture and planning.
landscape and urban planning, 36 (1): 59-77. CRACRAFT, J., 1985.- Historical biogeography and patterns of differentiation within South American avifauna: areas of endemisms.
ornithological monographs, 36: 49-84.DEVIA, C.A., MAYA, D.L., CORDOBA, L.M., VILLA, L.A. & HERAZO, B., 2003.- Valle del río Cimitarra. Pontificia Universidad
Javeriana, Bogotá, Colombia.DÍAZ, I.A., ARMESTO, J.J., REID, S., SIEVING, K.E. & WILLSON, M.F., 2005.- Linking forest structure and composition: avian
diversity in successional forests of Chiloé Island, Chile. Biological conservation, 123 (1): 91-101. DURÁN, S.M. & KATTAN, G.H., 2005.- A Test of the utility of exotic tree plantation for understory birds and food resources in
the Colombian Andes. Biotropica, 37 (1): 129-135.ESTELA, F.A. & LÓPEZ-VICTORIA. M., 2005.- Aves de la parte baja del río Sinú, Caribe colombiano: inventario y ampliaciones
de distribución. Boletín de investigaciones marinas y costeras, 34 (1): 7-42. ESTRADA, A. & COATES-ESTRADA, R., 2005.- Diversity of Neotropical migratory landbirds species assemblages in forest fragments
and man-made vegetation in Los Tuxtlas, Mexico. Biodiversity and conservation, 14: 1719-1734 ESTRADA, A., COATES-ESTRADA, R. & MERITT, D.A., 1997.- Anthropogenic landscape changes and avian diversity at Los
Tuxtlas, Mexico. Biodiversity and conservation, 6: 19-43. FAHRIG, L., 2003.- Effects of habitat fragmentation on biodiversity. Annual review of ecology, evolution and systematics, 34: 487-515. GARCÍA, H., CORZO, G., ISAACS, P. & ETTER, A., 2014.- Distribución y estado actual de los remanentes del bioma de Bosque
Seco Tropical en Colombia: insumos para su gestión: 228-251 (in) PIZANO, C. & GARCÍA, H. (eds.) el Bosque Seco Tropical en Colombia. Instituto de Investigaciones Biológicas Alexander von Humboldt (IAvH), Bogotá.
GASPARRI, N.I., GRAU, H.R., & SACCHI, L.V., 2015.- Determinants of the spatial distribution of cultivated land in the North Argentine dry Chaco in a multi-decadal study. Journal of arid environments, 123: 31-39.
GASTON, K.J. & BLACKBURN, T.M., 2000.- Pattern and Process in Macroecology. Blackwell Publishing Company, Malden, MA, USA.GILLIES, C.S. & ST CLAIR, C.C., 2010.- Functional responses in habitat selection by tropical birds moving through fragmented
forest. Journal of applied ecology, 47 (1): 182-190.GÓMEZ, C., GÓMEZ-BAHAMÓN, V., CÁRDENAS-ORTIZ, L. & BAYLY, N.J., 2015.- Distribution of Nearctic-Neotropical
migratory birds along a South American elevation gradient during spring migration. The Wilson journal of ornithology, 127 (1): 72-86.
HAMMER, O., HARPER, D.A.T. & RYAN, P.D., 2001.- PAST: Paleontological statistics software package for education and data analysis. Paleontología electrónica, 4 (1): 9.
HERZOG, S.K. & KESSLER, M., 2002.- Biogeography and composition of dry forest bird communities in Bolivia. Journal of ornithology, 143 (2): 171-204.
HILTY, S.L. & BROWN, W.L., 1986.- A guide to the birds of Colombia. Princeton University Press, New Jersey, USA.HOLDRIDGE, L.R., 1967.- life Zone ecology. Tropical Science Center, San José, Costa Rica.HUGHES, J.B., DAILY, G.C. & EHRLICH, P.R., 2002.- Conservation of tropical forest birds in countryside habitats. ecological
letters, 5 (1): 121-129. HUTCHESON, K., 1970.- A test for comparing diversities based on the Shannon formula. Journal of theoretical biology, 29: 151-154.IUCN, 2014.- The IUCN Red list of Threatened Species. http://www.iucnredlist.org/JANTZ, S.M., BARKER, B., BROOKS, T.M., CHINI, L.P. & FAHRIG, L., 2003.- Effects of habitat fragmentation on biodiversity.

129Bird assemble associated to two landscape units in a tropical dry forest
Annual review of ecology, evolution, and systematics, 34: 487-515. JANZEN, D., 1988.- Tropical Dry Forest: The most threatened major tropical ecosystem. National Forum on Biodiversity. Washington,
D.C. 130-137 pp.JOST, L., 2006.- Entropy and diversity. oikos, 113: 363-375.KREBS, C.J. 1999.- ecological methodology. Benjamin/Cummings, Menlo Park, California. LOSADA-PRADO, S., CARVAJAL-LOZANO, A.M. & MOLINA-MARTÍNEZ, Y.G., 2005.- Listado de especies de aves de la
cuenca del río Coello (Tolima, Colombia). Biota Colombiana, 6 (1): 101-115.LOSADA-PRADO, S. & MOLINA-MARTÍNEZ, Y., 2012.- Avifauna del Bosque Seco Tropical en el departamento del Tolima
(Colombia): análisis de la comunidad. Caldasia, 33 (1): 271-294.MACGREGOR‐FORS, I. & SCHONDUBE, J.E., 2011.- Use of tropical dry forests and agricultural areas by Neotropical bird
communities. Biotropica, 43 (3): 365-370.MAGURRAN, A., 2004.- Measuring biological diversity. Blackwell publishing, Malden, MA, USA.MARINARO, S., GRAU, H.R., MACCHI, L. & ZELAYA, P.V., 2014.- Land tenure and biological communities in dry Chaco forests
of northern Argentina. Journal of arid environments, 123: 60-67.MARTIN, T.E., 1984.- Impact of livestock grazing on birds of a Colombian cloud forest. Tropical ecology, 25: 158-171.MERRIAM, G., 1984.- Connectivity: a fundamental characteristic of landscape pattern: 5-15 (in) BRANDT, J. & AGGER, P. (eds.)
Methodology in landscape ecological research and planning. Volume 1. Theme 1: landscape ecological concepts. Roskilde University Centre, Roskilde, Denmark.
MOORE, J.E., SCHEIMAN, D.M. & SWIHART, R.K., 2004.- Field comparison of removal and modified double-observer modeling for estimating detectability and abundance of birds. Auk, 121 (3): 865-876.
MORENO, C.E., 2001.- Métodos para medir la biodiversidad. Vol. 1. M&T–Manuales y Tesis SEA, Zaragoza.MORTELLITI, A., FAGIANI, S., BATTISTI, C., CAPIZZI, D. & BOITANI, L., 2010.- Independent effects of habitat loss, habitat
fragmentation and structural connectivity on forest‐dependent birds. Diversity and distributions, 16 (6): 941-951.MYERS, N., MITTERMEIER, R.A., MITTERMEIER, C.G., DA FONSECA, G.A. & KENT, J., 2000.- Biodiversity hotspots for
conservation priorities. Nature, 403 (6772): 853-858.NARANJO, L.G., 1992.- Estructura de la avifauna en un área ganadera en el Valle del Cauca, Colombia. Caldasia, 17 (1): 55-65.OPDAM, P., 1990.- Dispersal in fragmented populations: the key to survival. 3-17 (in) BUNCE, R.G.H. & HOWARD, D.C. (eds.)
Species dispersal in agricultural habitats. Belhaven Press, London, UK.PATTEN, M.A. & SMITH-PATTEN, B.D., 2012.- Testing the microclimate hypothesis: Light environment and population trends
of Neotropical birds. Biological Conservation, 155: 85-93.PEARSON, D.L., 1975.- The relation of foliage complexity to ecological diversity on three Amazonian bird communities. Condor,
77: 453-466. PETIT, L.J. & PETIT, D.R., 2003.- Evaluating the importance of human‐modified lands for Neotropical bird conservation.
Conservation biology, 17 (3): 687-694.PINO, P.J., PÖYRY, J., RAATIKAINEN, K.M., SANG, A., STEFANESCU, C., TEDER, T., ZOBEL, M. & STEFFAN-DEWENTER,
I., 2010.- Habitat fragmentation causes immediate and time‐delayed biodiversity loss at different trophic levels. ecological letters, 13 (5): 597-605.
PLACKETT, R.L., 1983.- Karl Pearson and the chi-squared test. International statistical review, 51 (1): 59-72.PORTILLO-QUINTERO, C.A. & SÁNCHEZ-AZOFEIFA, G.A., 2010.- Extent and conservation of tropical dry forests in the
Americas. Biology conservation, 143 (1): 144-155.PORTILLO-QUINTERO, C., SÁNCHEZ-AZOFEIFA, A. & DO ESPIRITO-SANTO, M.M., 2013.- Monitoring deforestation
with MODIS Active Fires in Neotropical dry forests: An analysis of local-scale assessments in Mexico, Brazil and Bolivia. Journal of arid environment, 97: 150-159.
PRIETO-TORRES, D.A., NAVARRO‐SIGÜENZA, A.G., SANTIAGO‐ALARCÓN, D. & ROJAS‐SOTO, O.R., 2016.- Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation. Global change biology, 22 (1): 364-379.
RALPH, C.J., GEUPEL, G.R., PYLE, P., MARTIN, T.E., DESANTE, D.F. & MILÁ, B., 1996.- Manual de métodos de campo para el monitoreo de aves terrestres. Gen. Tech. Rep. PSW-GTR-159. Albany, CA, U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station.
RENGIFO, L., 1999.- Effect of Natural and Anthropogenic Landscape Matrices on the Abundance of Subandean Bird Species. ecological applications, 11 (1): 14-31.
RIVERA-GUTIÉRREZ, F., 2006.- Composición y estructura de una comunidad de aves en un área suburbana en el suroccidente Colombiano. ornitología colombiana, 4: 28-38.
SAAD, V.A. & PETIT, D.R., 1992.- Impact of pasture development on winter bird communities in Belize, Central America. Condor, 94: 66-71.
SÁNCHEZ‐AZOFEIFA, G.A., QUESADA, M., RODRÍGUEZ, J.P., NASSAR, J.M., STONER, K.E., CASTILLO, A. et al., 2005.- Research Priorities for Neotropical Dry Forests. Biotropica, 37 (4): 477-485.
SKIRVIN, A.A., 1981.- Effect of time of day and time of season on the number of observations and density estimates of breeding birds. Studies in avian biology, 6: 271-274.
SMITH, A.C., FAHRIG, L. & FRANCIS, C.M., 2011.- Landscape size affects the relative importance of habitat amount, habitat fragmentation, and matrix quality on forest birds. ecography, 34 (1): 103-113.
SODHI, N.S., ŞEKERCIOĞLU, C.H., BARLOW, J. & ROBINSON, S.K., 2011.- effects of Habitat Fragmentation on Tropical Birds. In Conservation of Tropical Birds, Wiley-Blackwell, England, Oxford.
STATTERSFIELD, A., CROSBY, M.J., LONG, A.J. & WEGE, D.C., 1998.- endemic bird areas of the world: priorities for biodiverstiy conservation. BirdLife International, Cambridge, Reino Unido.
STILES, F.G., BOHÓRQUEZ, C.I., CADENA, C.D., DE LA ZERDA, S., HERNÁNDEZ, M. ROSSELLI, L., et al., 2000.- Aves

130 Juan Pablo Sánchez-Montenegro, Katherine Pérez-Castillo, Mario Fernando Garcés-Restrepo, Alan Giraldobo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
15-1
30
de la sabana de Bogotá. Asociación Bogotana de Ornitología, Bogotá.STILES, F.G. & ROSSELLI, L., 1998.- Inventario de las aves de un bosque alto andino: comparación de dos métodos. Caldasia,
20: 29-43. STILES, F.G., TELLERÍA, J.L. & DÍAZ, M., 1995.- Observaciones sobre la composición, ecología y zoogeografía de la avifauna de
la Sierra de Chiribiquete, Caquetá, Colombia. Caldasia, 17 (82-85): 481-500.STOTZ, D.F., FITZPATRICK, J.W., PARKER III, T.A. & MOSKOVITZ, D.K., 1996.- Neotropical Birds: ecology and Conservation.
University of Chicago Press, Chicago, Illinois, USA.SUGGITT, A.J., GILLINGHAM, P.K., HILL, J.K., HUNTLEY, B., KUNIN, W.E., ROY, D.B. & THOMAS, C.D., 2011.- Habitat
microclimates drive fine‐scale variation in extreme temperatures. oikos, 120 (1): 1-8. TAYLOR, P.D., FAHRIG, L., HENEIN, K. & MERRIAM, G., 1993.- Connectivity is a vital element of landscape structure. oikos,
68: 571-573.TEWS, J., BROSE, U., GRIMM, V., TIELBÖRGER, K., WICHMANN, M.C., SCHWAGER, M. & JELTSCH, F., 2004.- Animal
species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography, 31 (1): 79-92.
TRILLERAS, J.M., JARAMILLO, V.J., VEGA, E.V. & BALVANERA, P., 2015.- Effects of livestock management on the supply of ecosystem services in pastures in a tropical dry region of western Mexico. Agriculture, ecosystem and environment, 211: 133-144.
TSCHARNTKE, T., SEKERCIOGLU, C.H., DIETSCH, T.V., SODHI, N.S., HOEHN, P. & TYLIANAKIS, J.M., 2008.- Landscape constraints on functional diversity of birds and insects in tropical agroecosystems. ecology, 89 (4): 944-951.
TURNER, I.M. & CORLETT, R.T., 1996.- The conservation value of small, isolated fragments of lowland tropical rain forest. Trends in ecology and evolution, 11 (8): 330-333.
URIARTE, M., ANCIÃES, M., DA SILVA, M.T., RUBIM, P., JOHNSON, E. & BRUNA, E.M., 2011.- Disentangling the drivers of reduced long-distance seed dispersal by birds in an experimentally fragmented landscape. ecology, 92 (4): 924-937.
VARGAS-FIGUEROA, J.A., GONZÁLEZ-COLORADO, A.M., BARONA-CORTÉS, E., BOLÍVAR-GARCÍA, W. & GIRALDO, A., 2016.- Composición y estructura vegetal de fragmentos de bosque seco tropical y de dos zonas con actividad antrópica en la Dorada-Victoria, Caldas. Revista Ciencias, 20(2): 13-60.

estudios de orGanisMos del BosQue HuMedo troPiCal
Organisms studies of the tropical rainforest


Boletín CientífiCoCentro de Museos
Museo de Historia natural
dietas VeGetales en el desarrollo larVal de dos esPeCies de Morpho (lePidoPtera: nYMPHalidae,
satYrinae, MorPHini) en CautiVerio, en iQuitos, loreto, PerÚ*
Joel Vásquez1, Wenceslao Peña1, Ricardo Zárate2, Julio Pinedo3, Rocío Correa1, Juan José Palacios2, Juan José Bellido4, Juan José Ramírez5, Gerardo lamas5
resumen
Se determinó el efecto de dietas vegetales en el desarrollo larval de dos especies de mariposas diurnas en cautiverio en el Centro de Investigaciones Allpahuayo del Instituto de Investigaciones de la Amazonía Peruana (IIAP), Iquitos, Perú. Para evaluar sus ciclos biológicos se emplearon 20 larvas recién emergidas de cada una de las especies, de forma individual, y para examinar el efecto de diferentes dietas en su desarrollo larval se empleó un diseño completamente al azar, con dos tratamientos y diferentes repeticiones (n1 = 31 y n2 = 14). Se reporta que la dieta con el hospedero Arachis pintoi fue óptima para las especies Morpho menelaus occidentalis y Morpho helenor theodorus, que completaron sus ciclos biológicos respectivamente en 97,51±7,98 días (n = 15), con 75% de supervivencia, y 73,22±7,26 días (n = 14), con una supervivencia de 70%. M. helenor theodorus completó su ciclo biológico con su hospedero natural Platymiscium stipulare en 75,84±10,19 días (n = 10), siendo su ciclo biológico más prolongado y con supervivencia más baja (50%). Las plantas hospederas Vigna aff. candida y Desmodium adscendens no fueron favorables para las dos especies de Morpho, ya que las larvas murieron en el primer estadio. En M. helenor theodorus no existe relación entre el sexo y la dieta respecto a la longevidad (prueba Chi cuadrado). Las hembras vivieron en promedio 53,76 días y los machos 25,45 días.
Palabras clave: Morpho menelaus occidentalis, Morpho helenor theodorus, plantas hospederas, ciclo de vida.
1 Programa de Investigación en Cambio Climático Desarrollo Territorial y Ambiente, Instituto de Investigaciones de la Amazonía Peruana. Iquitos, Perú. E-mail: JJP: [email protected]; RZ: [email protected] Facultad de Agronomía, Universidad Nacional de la Amazonía Peruana. Iquitos, Perú. E-mail: [email protected] 3 Unidad de Comunicación e Información, Instituto de Investigaciones de la Amazonía Peruana. Iquitos, Perú. E-mail: [email protected] 4 Museo de Historia Natural, Universidad Nacional Mayor de San Marcos. Lima, Perú. E-mail: [email protected] 5 Disponible en http://www.ebay.com/bhp/morpho-menelaus - consulta: 27 abril de 2017.
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 133-146. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:VÁSQUEZ, J., PEÑA, W., ZÁRATE, R., PINEDO, J., CORREA, R., PALACIOS, J.J., BELLIDO, J.J., RAMÍREZ, J.J. & LAMAS, G., 2017.- Dietas vegetales en el desarrollo larval de dos especies de Morpho (lepidoptera: nymphalidae, satyrinae, morphini) en cautiverio, en Iquitos, Loreto, Perú. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 133-146. DOI: 10.17151/bccm.2017.21.2.9

Joel Vásquez et al.134bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
introduCCiÓn
En la búsqueda de alternativas de aprovechamiento sostenible de los recursos naturales es prioritario el desarrollo de trabajos de investigación que permitan implementar alternativas de manejo de la biodiversidad. En tal sentido, una de las alternativas para el uso y conservación de los recursos naturales es el negocio comercial de mariposas, debido a que estos insectos son difíciles de sobreexplotar, es un recurso renovable y en cambio los bosques son fáciles de destruir (ORSAK, 1993; CONSTANTINO, 1996; MORENO, 1998; SALAZAR et al., 2010). Teóricamente es fácil lograr la reproducción de un insecto si se cuenta con los recursos mínimos, tanto humanos como tecnológicos; pero cabe aclarar que no es fácil garantizar el crecimiento y desarrollo normal del individuo adulto recién eclosionado si se desconocen los factores limitantes de sus necesidades nutricionales (RAMÍREZ, 1994). Se puede afirmar que bajo condiciones de cautiverio un proceso de cría de mariposa debe mejorar los índices de postura, eclosión larval, desarrollo larval y emergencia del adulto, ya que las especies de mariposas, al ser trabajadas como animales susceptibles a domesticación,
VeGetaBle diets on tHe larVal deVeloPMent of tWo Morpho sPeCies (lePidoPtera: nYMPHalidae, satYrinae,
MorPHini) in CaPtiVitY in iQuitos, loreto, Peru
abstract
The effect of vegetable diets on the larval development of two daytime butterflies in captivity at the Allpahuayo Research Center of the Instituto de Investigaciones de la Amazonía Peruana (IIAP), Iquitos, Peru, was determined. To evaluate their biological cycles, 20 freshly emerged larvae of each species were used individually and, to estimate the effect of different diets in their larval development, a completely randomized design was used with two treatments and different replicates (n1 = 31 and n2 = 14). It was reported that the diet with the host plant Arachis pintoi was optimal for both Morpho menelaus occidentalis and Morpho helenor theodorus species, which completed their biological cycles in, 97.51±7.98 days (n = 15), with 75% sur-vival, and 73.22±7.26 days (n = 14), with a survival rate of 70% respectively. Morpho helenor theodorus completed its biological cycle on its natural host plant Platymiscium stipulare in 75.84±10.19 days (n = 10), being its biological cycle longer with a lower survival rate (50%). The host plants Vigna aff. candida and Desmodium adscendens were not adequate for the two species of Morpho, since the larvae died in the first stage. In Morpho helenor theodorus, there is no relationship between gender and diet in relation to longevity (Chi square test). Females lived in average 53.76 days and males 25.45 days.
Key words: Morpho menelaus occidentalis, Morpho helenor theodorus, host plants, life cycle.

Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 135
no estarían sujetas a la competencia por alimento, no existirían depredadores, ni modificaciones de factores como luz, ruido, sombra, temperatura o humedad, que afectan considerablemente el ciclo biológico de estos insectos y habría control sanitario y manejo de parámetros reproductivos (RAMÍREZ, 1994). Las mariposas del género Morpho son muy cotizadas en el mercado internacional para la elaboración de artesanías con precios que oscilan entre US$ 8,00 a 75,00 (Ebay, 2017). En el Perú, GALLUSER et al. (2010) estudiaron el desarrollo y polimorfismo de M. (Iphimedeia) telemachus (Linnaeus, 1758) utilizando como planta hospedera natural a Abuta grandifolia (Menispermaceae). RAMÍREZ et al. (2014) reportaron el ciclo biológico de M. cisseis phanodemus Hewitson, 1869, también en su hospedero natural A. grandifolia. RUIZ et al. (2015) estudiaron el ciclo biológico de M. helenor theodorus Fruhstorfer, 1907, utilizando Arachis pintoi (Fabaceae) como una planta alterna. VÁSQUEZ et al. (2017) reportaron las plantas hospederas de M. helenor theodorus y M. menelaus occidentalis C. Felder & R. Felder, 1862, las mismas que presentan lento crecimiento y producen escaso follaje imposibilitando desarrollar su crianza sostenible en condiciones de cautiverio. Una alternativa fue la utilización de plantas de corto periodo vegetativo y suficiente follaje, que permitan alimentar las larvas hasta completar su ciclo de vida en cautiverio, por lo que fue necesario investigar el efecto de dos dietas alternas en el desarrollo larval de dos especies de mariposas diurnas: M. menelaus occidentalis y M. helenor theodorus.
MÉtodoloGía
Área de estudio
El presente trabajo se desarrolló de febrero a diciembre de 2016, en el Centro de Investigaciones Allpahuayo (CIA) del Instituto de Investigaciones de la Amazonía Peruana (IIAP), que está ubicado a 26,5 km de Iquitos, en la carretera Iquitos-Nauta (Figura 1), con coordenadas 03°57’S, 73°26’ W. El CIA está limitado por la carretera Iquitos-Nauta y el río Nanay, y presenta precipitaciones mayores a 2400 mm anuales, distribuidas de tal forma que no hay una estación seca definida; no obstante, la estación más lluviosa se extiende de noviembre a mayo. La humedad atmosférica oscila entre 80 y 90% y la temperatura media anual excede los 24°C.
recolección de adultos
Para la recolección de mariposas adultas de M. menelaus occidentalis y M. helenor theodorus se utilizó la técnica del cebo, que consiste en poner rodajas de plátano maduro macerado en jugo de caña de azúcar por un lapso de tres días. Tales cebos fueron colocados al interior del bosque, en la punta de una estaca de 1,20 m, lo que atraía las mariposas y cuando estas se alimentaban fueron recolectadas empleando una red entomológica. Las mariposas capturadas se colocaron en sobres entomológicos

Joel Vásquez et al.136bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
para ser transportadas y liberadas en un mariposario de 250 m2 con 3 m de altura, que contenía sus plantas hospederas, Vigna aff. candida (Fabaceae) para M. menelaus occidentalis (FIGURA 6C) y Platymiscium stipulare (Fabaceae) para M. helenor theodorus (RUIZ et al., 2015; VÁSQUEZ et al., 2017). La alimentación de los adultos fue realizada cada tres días, utilizando rodajas de plátanos maduros fermentados en jugo de caña de azúcar. Luego de la oviposición los huevos fueron recolectados para iniciar los experimentos.
Ubicación del Centro de Investigaciones Allpahuayo, Loreto, Perú.figura 1.
tratamientos en estudio
Para el presente estudio se establecieron seis tratamientos, los cuales se detallan en la Tabla 1.
unidades experimentales
Para la evaluación del ciclo biológico se utilizaron 20 larvas (UE = unidad experimental) recientemente emergidas, totalizando 120 larvas. Para el caso de la evaluación del efecto de las dietas sobre el desarrollo larval de M. helenor theodorus se utilizaron diferentes números de repeticiones para A. pintoi (n = 31) y para P. stipulare (n = 14).

Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 137
Tratamientos de dietas vegetales para dos especies de Morphotabla 1.
tratamiento dietas y especiesT1T2T3T4T5T6
A. pintoi con M. menelaus occidentalisD. adscendens con M. menelaus occidentalisV. aff. candida con M. menelaus occidentalis (testigo)A. pintoi con M. helenor theodorusD. adscendens con M. helenor theodorusP. stipulare con M. helenor theodorus (testigo)
evaluación del ciclo biológico
Previa a la evaluación del ciclo biológico, se hizo un ensayo preliminar de aceptación alimentaria con las dietas alternas en cautiverio. Luego se recolectaron los huevos recientemente ovipositados sobre sus plantas hospederas en el mariposario, los que fueron transportados al laboratorio, donde se registró la emergencia de las larvas. Luego las larvas se confinaron en envases de cría de 1 litro en forma individual y se prepararon pequeños floreros con las hojas de cada planta hospedera en estudio. Se registró el número de estadios larvales, y el periodo de duración de cada estadio larval, prepupa y pupa. Para calcular el porcentaje de sobrevivencia larval, se aplicó la siguiente formula.
# LV % SV= ------------ x 100 # TLCtto
donde: % SV: Porcentaje de supervivencia de larvas. # LV: Número de larvas vivas.# TLCtto: Número de larvas criadas según tratamiento.
evaluación de las dietas alimenticias en el desarrollo larvario
Se registró el peso inicial y final del alimento con una frecuencia diaria, durante todo el desarrollo larvario hasta el estado de pupa. En cada estadio larvario inmediatamente después de cada muda se registró con una regla graduada los datos de la longitud larval, y el ancho de la cápsula cefálica se midió con una regla micrométrica. Para registrar el peso de la larva se utilizó una balanza electrónica en gramos y solo se aplicó a larvas a partir del cuarto estadio debido a que las larvas en estadios anteriores fueron muy pequeñas.

Joel Vásquez et al.138bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
relación sexo y dieta sobre la longevidad de los adultos de Morpho helenor theodorus
Para determinar la relación del sexo y la dieta sobre la longevidad de los adultos obtenidos de las larvas sometidas a las dos dietas (A. pintoi y P. stipulare), después de su nacimiento los adultos fueron marcados y liberados en el mariposario, registrándose el tiempo de vida (en días) de cada uno, tanto machos (FIGURA 6D) como hembras, que fueron alimentados con rodajas de plátano fermentado en jugo de caña de azúcar, con una frecuencia interdiaria.
determinación taxonómica de las plantas hospederas
Las muestras botánicas fueron recolectadas de acuerdo a JUDD et al. (1999) y su determinación taxonómica se realizó por medio de las claves de GENTRY (1993), VÁSQUEZ (1997), RIBEIRO et al. (1999) y VÁSQUEZ & ROJAS (2004); también se utilizó la información disponible en Internet de los herbarios del Missouri Botanical Garden (http://tropicos.org/ - consultado en enero 2017) y del Field Museum of Natural History (http://fm1.fieldmuseum.org/vrrc/ - consultado en enero 2017). Las muestras entomológicas están depositadas en el laboratorio de crianza de mariposas del IIAP y las botánicas en el Herbario Iquitos (HIQ) del IIAP.
análisis de datos
Para el análisis de variancia (ANOVA), se utilizó el programa estadístico SPSS-21 con un nivel de significancia del 95% para determinar el efecto de la dieta vegetal sobre el desarrollo larval de M. helenor theodorus. Para estimar la relación de sexo y dieta sobre la longevidad de los adultos en días, se aplicó la prueba estadística de Chi cuadrado.
resultados
Ciclo biológico de M. menelaus occidentalis en días
En la Figura 2 se muestra que M. menelaus occidentalis no logró adaptase a D. adscendens (Fabaceae) ni a su hospedero natural V. aff. candida. Las larvas del primer estadio se alimentaron de las plantas pero ninguna logró pasar al segundo estadio, en cambio con A. pintoi lograron completar su ciclo biológico en 97,51±7,98 días.
supervivencia larval de Morpho menelaus occidentalis
La Figura 3 muestra que las larvas de M. menelaus occidentalis no lograron adaptarse completamente a D. adscendens, sobreviviendo solo por 4,0±0,76 días. Con V. aff. candida sobrevivieron solo por 6,00±1,21 días (primer estadio). En cambio, con A. pintoi (FIGURA 6A) alcanzaron el 80% de supervivencia, y 75% alcanzó el estadio de pupa.
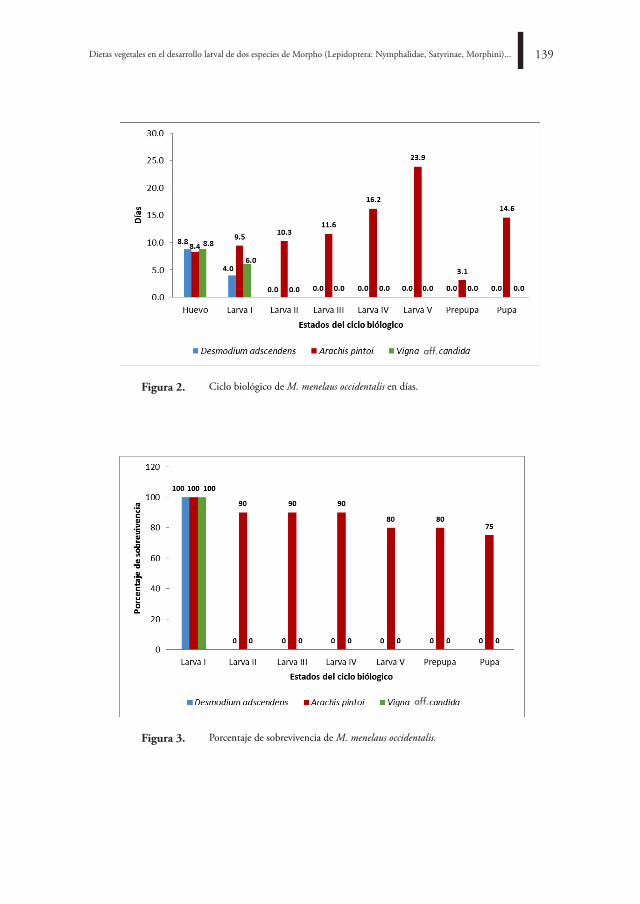
Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 139
Ciclo biológico de M. menelaus occidentalis en días.figura 2.
Porcentaje de sobrevivencia de M. menelaus occidentalis.figura 3.
aff.
aff.

Joel Vásquez et al.140bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
duración del ciclo biológico de Morpho helenor theodorus en días
La Figura 4 muestra que M. helenor theodorus no logró adaptase a D. adscendens. Las larvas del primer estadio se alimentaron de la planta pero ninguna logró pasar al segundo estadio. En cambio, con A. pintoi lograron completar su ciclo biológico en 73,22±7,26 días y con P. stipulare (testigo) alcanzaron 75,84±10,19 días. El comportamiento es bastante estable en los diferentes estadios larvales y en las fases de prepupa y pupa. Las discrepancias, en el número de días de la duración del ciclo biológico no son relevantes. El mayor tiempo de la fase larval se observa en el estadio V, siendo más prolongado con P. stipulare (testigo) que con A. pintoi.
Porcentaje de sobrevivencia de M. menelaus occidentalis.figura 4.
supervivencia larval de M. helenor theodorus
La Figura 5 muestra que las larvas de M. helenor theodorus no lograron alimentarse completamente de D. adscendens, sobreviviendo 3,5±0,74 días, luego dejaron de comer, excretando líquido se acortaron y murieron sin alcanzar el segundo estadio. En cambio, la mayor supervivencia se obtuvo con A. pintoi, y el 70% de las larvas logró alcanzar las etapas de prepupa y pupa, mientras que con P. stipulare (testigo) la supervivencia larval fue menor (50%) y todas las larvas alcanzaron la etapa de pupa.
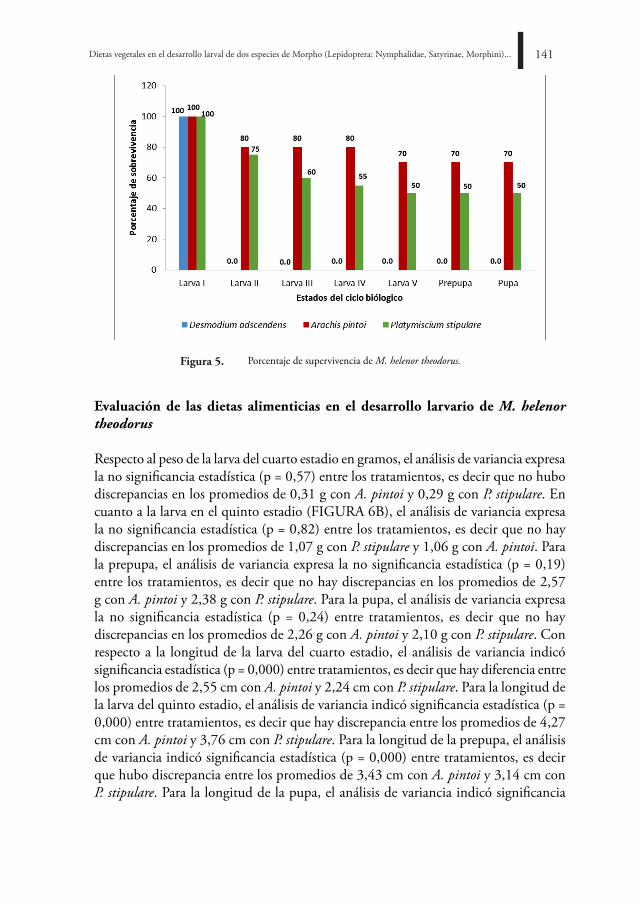
Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 141
Porcentaje de supervivencia de M. helenor theodorus.figura 5.
evaluación de las dietas alimenticias en el desarrollo larvario de M. helenor theodorus
Respecto al peso de la larva del cuarto estadio en gramos, el análisis de variancia expresa la no significancia estadística (p = 0,57) entre los tratamientos, es decir que no hubo discrepancias en los promedios de 0,31 g con A. pintoi y 0,29 g con P. stipulare. En cuanto a la larva en el quinto estadio (FIGURA 6B), el análisis de variancia expresa la no significancia estadística (p = 0,82) entre los tratamientos, es decir que no hay discrepancias en los promedios de 1,07 g con P. stipulare y 1,06 g con A. pintoi. Para la prepupa, el análisis de variancia expresa la no significancia estadística (p = 0,19) entre los tratamientos, es decir que no hay discrepancias en los promedios de 2,57 g con A. pintoi y 2,38 g con P. stipulare. Para la pupa, el análisis de variancia expresa la no significancia estadística (p = 0,24) entre tratamientos, es decir que no hay discrepancias en los promedios de 2,26 g con A. pintoi y 2,10 g con P. stipulare. Con respecto a la longitud de la larva del cuarto estadio, el análisis de variancia indicó significancia estadística (p = 0,000) entre tratamientos, es decir que hay diferencia entre los promedios de 2,55 cm con A. pintoi y 2,24 cm con P. stipulare. Para la longitud de la larva del quinto estadio, el análisis de variancia indicó significancia estadística (p = 0,000) entre tratamientos, es decir que hay discrepancia entre los promedios de 4,27 cm con A. pintoi y 3,76 cm con P. stipulare. Para la longitud de la prepupa, el análisis de variancia indicó significancia estadística (p = 0,000) entre tratamientos, es decir que hubo discrepancia entre los promedios de 3,43 cm con A. pintoi y 3,14 cm con P. stipulare. Para la longitud de la pupa, el análisis de variancia indicó significancia

Joel Vásquez et al.142bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
estadística (p = 0,000), con discrepancia entre los promedios de 2,70 cm con A. pintoi y 2,55 cm con P. stipulare. Para el peso del alimento consumido (en gramos) en el quinto estadio, el análisis de variancia expresó la no significancia estadística (p = 0,017) entre tratamientos, es decir que no hay discrepancia en los promedios de 15,75 g con P. stipulare y 14,59 g con A. pintoi. Para la longevidad de los adultos de ambos sexos, el análisis de variancia indicó la no significancia estadística (p = 0,37) entre tratamientos, no habiendo discrepancia entre los promedios de 37,4 días (n = 14) con A. pintoi y 31,9 días (n = 9) con P. stipulare.
B
C D
a) Larva del quinto estadio de Morpho menelaus occidentalis sobre hojas de Ara-chis pintoi (JV). B) Larva del quinto estadio de Morpho helenor theodorus sobre hoja de Platymiscium stipulare (WP). C) Adulto hembra de Morpho menelaus occidentalis. d) Adulto macho de Morpho helenor theodorus (WP). (Fotografías tomadas por JV = Joel Vásquez y WP = Wenceslao Peña).
figura 6.
relación sexo y dieta sobre la longevidad de los adultos de M. helenor theodorus
Las hembras vivieron en promedio 53,76 días (n = 8) y los machos 25,42 días (n = 15) con ambas dietas. Los adultos de ambos sexos obtienen mayor longevidad con A. pintoi para ambos sexos, y para ambas dietas la longevidad es mayor en las hembras. Según la prueba de Chi cuadrado (X2 = 0,00123) se acepta que no existe relación entre sexo y dieta con respecto a la longevidad.
A

Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 143
disCusiÓn
M. menelaus occidentalis no se adaptó a V. aff. candida bajo condiciones de cautiverio, las hojas de la planta se resecaron rápidamente, y las larvas murieron en el primer estadio, similar a lo reportado por GALLUSSER et al. (2010), que reportaron alta mortalidad (60%) en larvas del primer estadio de M. (Iphimedeia) telemachus criados en su hospedero natural bajo condiciones de cautiverio. Según MULANOVICH (2007), en muchos casos las mariposas ovipositan en algunas plantas hospederas cuya toxicidad es alta y, por lo tanto, el nivel de supervivencia de las orugas es bajo. El hospedero alterno Desmodium adscendens resiste la desecación y sus hojas permanecen frescas en condiciones de cautiverio, sin embargo las larvas mueren en el primer estadio; es posible que esto se deba a que la planta contiene saponinas (ALLUÉ-CREUS, 1999), compuestos altamente tóxicos para los animales (VALLE, 2000). En cambio, con A. pintoi las larvas se adaptaron al alimento y lograron completar su ciclo biológico en 97,51±7,98 días. En forma similar, YOUNG (1974) reportó la adaptación alimentaria de Morpho peleides Kollar, 1849, a una dieta alterna, Arachis hypogaea, completando el ciclo en 104,8 días en Costa Rica. M. helenor theodorus tampoco se adaptó a D. adscendens, las larvas murieron en el primer estadio. Con A. pintoi completaron el ciclo biológico en 73,2±8,34 días, un periodo menor en 2,67 días con respecto a su hospedero natural P. stipulare (75,87±9,12 días); igualmente, mostraron mayor supervivencia (70%) que con el hospedero natural (50%). Datos similares reportó YOUNG (1974), quien indicó una diferencia de 0,8 días menor en el ciclo biológico de M. peleides en A. hypogaea (104,8 días) en contraste a Mucuna urens (105,6 días). RUIZ et al. (2015) reportaron datos similares para la misma subespecie (M. h. theodorus) estudiada en la misma zona de Loreto con A. pintoi, obteniendo un ciclo biológico de 68,11±1,85 días, con una diferencia de 5,09 días con respecto a los resultados presentados aquí. Dicha diferencia, se asume, debida a que el presente estudio fue llevado a cabo entre noviembre y enero con una fluctuación de temperatura ambiental de 25 a 27°C, mientras el estudio de RUIZ et al. (2015) se efectuó entre febrero y julio con una temperatura ambiental de 27 a 29°C. La duración del ciclo biológico varía de acuerdo a la temperatura, a menor temperatura el ciclo se prolonga y a mayor temperatura el ciclo se acorta, como lo reportó HURTADO (2009), quien indica un periodo de 36,15 días a 28°C y de 26,15 días a 31°C para Utetheisa ornatrix venusta (Lepidoptera: Arctiinae). En los tratamientos que incluyeron dietas alimenticias, el peso de la larva de M. helenor theodorus en el cuarto y quinto estadio no fue significativo (A. pintoi con 2,26 g y P. stipulare con 2,10 g), estos resultados nos permiten inferir que esta modesta diferencia puede deberse a que la dieta alterna de A. pintoi es rica en porcentaje proteico, con 16,2% (RINCÓN, 1999). La longitud de los estadios larvales IV y V, prepupa y pupa en M. helenor theodorus, fue altamente significativa estadísticamente, la dieta alterna de A. pintoi indica superioridad sobre la longitud de los estadios larvales, prepupa y pupa en contraste con P. stipulare; esta discrepancia puede deberse a los nutrimentos de las plantas y en su gran mayoría por

Joel Vásquez et al.144bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
su composición química y nutricional (JAENIKE, 1990; SILVA, 2005; CLARO & RUIZ, 2010), lo que explicaría que la riqueza proteica es un factor favorable en una dieta alimenticia, es por eso que las larvas que consumieron A. pintoi, con 16,2% de riqueza proteica, registraron incrementos de pesos y longitudes mayores con respecto a las larvas que consumieron P. stipulare, con solo 1,54%, siendo A. pintoi una planta que mantiene altos niveles nutricionales inclusive a las 12 semanas de rebrote (ROJAS, 2007). En nuestro estudio se resalta el efecto del contenido proteico del alimento frente a la cantidad consumida.
El peso del alimento consumido en el quinto estadio en M. helenor theodorus no fue significativo. La discrepancia de pesos pudiera deberse a la consistencia fisiológica de las plantas alimenticias, A. pintoi se mantiene más turgente y P. stipulare se reseca más rápido. Tal como indican HOLLOWAY et al. (1992), la pérdida de agua en el alimento perjudica el desarrollo del insecto.
Las diferencias en longevidad del adulto de M. helenor theodorus no fueron significativas entre A. pintoi (37,4 días) y P. stipulate (31,9 días); pudiera esperarse que algunas especies han evolucionado para pasar de una dieta polífaga a dietas monófagas, oligófagas o estenófagas, según nivel de especialización (ANDREWS & QUEZADA, 1989), lo que permite alcanzar mayor longevidad. Asimismo, se podría indicar que esta diferencia de longevidad de adultos en relación a las dietas alimenticias se deba al aspecto crítico en el ciclo de vida de la mariposa, que es la habilidad de la hembra de ovipositar y de la oruga de alimentarse de una planta hospedera en particular (MULANOVICH, 2007). En nuestro estudio, las orugas alimentadas con A. pintoi en condiciones de cautiverio estarían prolongando favorablemente la duración de vida de los adultos.
La relación de sexo y dieta con la longevidad del adulto en M. helenor theodorus, según la prueba de Chi cuadrado nos indica que no hay dependencia. Las hembras de M. helenor theodorus viven en promedio 52,75 días y los machos 25,1 días, tal como lo ratifican RUIZ et al. (2015). Las hembras presentan mayor longevidad con respecto a los machos, sin embargo la longevidad de los insectos adultos en cautiverio no solo está influenciada por la alimentación en sus estados larvales sino también por la temperatura ambiental y la alimentación del adulto (SOTO & IANNACONE, 2008).
ConClusiones
M. menelaus occidentalis no completó el ciclo biológico en D. adscendens y V. aff. candida. Esta especie completó el ciclo biológico con la dieta alterna de A. pintoi en 97,51±7,98 días con una supervivencia larval de 80%. Para M. helenor theodorus, Desmodium adscendens no fue adecuada como dieta alterna; completó su ciclo biológico en A. pintoi (73,22±7,26 días) con una supervivencia de 70%, y en P. stipulare (75,84±10,19 días) con una supervivencia de 50%.

Dietas vegetales en el desarrollo larval de dos especies de Morpho (Lepidoptera: Nymphalidae, Satyrinae, Morphini)... 145
El peso de las larvas de quinto estadio de M. helenor theodorus no fue influenciado por A. pintoi (1,06 g) o P. stipulare (1,07 g). La longitud de los estadios larvales, prepupa y pupa, fue influenciada por A. pintoi (4,27 cm) y P. stipulare (3,76 cm). La longevidad del adulto no fue influenciada por ninguna de las dos dietas. El peso del alimento consumido en el quinto estadio no fue significativo, y A. pintoi se mantuvo más turgente que P. stipulare.
De las dietas estudiadas, se concluye que A. pintoi resultó favorable como dieta alterna para la crianza sostenible en cautiverio de las dos especies de Morpho.
aGradeCiMientos
Este trabajo fue financiado por el “Programa Nacional de Innovación para la Competitividad y Productividad, Innóvate-Perú” a través del proyecto “Modelos tecnológicos de crianza de 10 especies de mariposas diurnas para su aprovechamiento en bionegocios en la Región Loreto”, conducido por el Programa de Investigación en Biodiversidad Amazónica del Instituto de Investigaciones de la Amazonía Peruana. Los autores agradecen a Percy Huiñapi, Johnny Callirgos y Percy Ruiz por su apoyo durante los muestreos de campo, y a Manuel Martin Brañas por la revisión del manuscrito.
referenCias
ALLUÉ-CREUS, P., 1999.- Desmodium adscendens Swartz (Papilionáceas). Natura Medicatrix, 49 (1): 42-46.ANDREWS, K.L. & QUEZADA, J.R., 1989.- Manejo integrado de plagas insectiles en la agricultura: estado actual y futuro. Escuela
Agrícola Panamericana, Departamento de Protección Vegetal, Zamorano.CLARO, R. & RUIZ, N., 2010.- Aceptación de una dieta artificial por larvas de la mariposa Battus polydamas polydamas (Lepidoptera:
Papilionidae). Acta Biol. Colomb., 15 (1): 47-62.CONSTANTINO, L.M., 1996.- Ciclos de vida y plantas hospederas de lepidópteros diurnos con potencial económico en condiciones
de colinas bajas del Chocó biogeográfico: 75-86 (en) OSORIO, H. (ed.) Memorias del II seminario sobre investigación y manejo de fauna para la construcción de sistemas sostenibles. CIPAV, Cali.
GALLUSSER, S., RAMÍREZ, C. & BLANDIN, P., 2010.- Observaciones sobre el desarrollo y polimorfismo de Morpho (Iphimedeia) telemachus (Linnaeus, 1758) en el noreste Peruano (Lepidoptera, Nymphalidae, Morphinae). Bull. Soc. entomol. Fr., 115 (1): 5-15.
GENTRY, A., 1993.- A field guide to the families and genera of woody plants of northwest South America (Colombia, ecuador, Perú) with supplementary notes on herbaceous taxa. Conservation International, Washington DC.
HOLLOWAY, G.J., BRAKEFIELD, P.M., KOFMAN, S. & WINDIG, J.J., 1992.- An artificial diet for butterflies, including Bicyclus species, and its effect on development period, weight and wing pattern. J. Res. lepid, 30 (1-2): 121-128.
HURTADO, L., 2009.- Descripción del ciclo biológico de Utetheisa ornatrix venusta (Dalm) (Lepidoptera: Arctiidae) con una dieta artificial. Centro Agrícola, 36 (4): 79-84.
JAENIKE, J., 1990.- Host specialization on phytophagous insects. Ann. Rev. ecol. Syst., 21 (1): 243-273.JUDD, W.S., CAMPBELL, C.S., KELLOG, E.A., STEVENS, P.F. & DONOGHUE, M.J., 1999.- Plant systematics: A phylogenetic
approach. Sinauer, Sunderland, MA.MORENO, R., 1998.- Análisis económico del proyecto de fauna: cría de mariposas. Instituto Von Humboldt, Bogotá.MULANOVICH, A., 2007.- Guía para el manejo sustentable de las mariposas en el Perú. Instituto de Investigaciones de la Amazonía
Peruana, Iquitos.ORSAK, L., 1993.- Killing butterflies to save butterflies: a tool for tropical forest conservation in Papua New Guinea. News lepid.
Soc., 35 (3): 71-80.RAMÍREZ, C., GALLUSSER, S., LACHAUME, G. & BLANDIN, P., 2014.- The ecology and life cycle of the Amazonian Morpho
cisseis phanodemus Hewitson, 1869, with a comparative review of early stages in the genus Morpho (Lepidoptera: Nymphalidae: Morphinae). Trop. lepid. Res., 24 (2): 67-80.
RAMÍREZ, J., 1994.- Manejo de fauna silvestre y los límites previsibles de la sustentabilidad: 1-23 (en) Seminario de investigación y manejo de fauna para el desarrollo de sistemas sostenibles de producción en el trópico, Pontificia Universidad Javeriana, Cali.
RIBEIRO, J.E.L.S., HOPKINS, M.J.G., VICENTINI, A., SOTHERS, C.A., COSTA, M.A.S., BRITO, J.M., SOUZA, M.A.D., MARTINS, L.H.P., LOHMANN, L.G., ASSUNÇÃO, P.A.C.L., PEREIRA, E.C., SILVA, C.F., MESQUITA, M.R. &

Joel Vásquez et al.146bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
33-1
46
PROCÓPIO, L.C., 1999.- Flora da Reserva Ducke. Guia de identificação das plantas vasculares de uma floresta de terra-firme na Amazônia Central. Instituto Nacional de Pesquisas da Amazônia, Manaus.
RINCÓN, C.A., 1999.- Maní forrajero (Arachis pintoi) la leguminosa para sistemas sostenibles de producción agropecuaria. Información Técnica Corpoica, 3 (24): 2-8.
ROJAS, A., 2007.- Ventajas y limitaciones para el uso del maní forrajero perenne (Arachis pintoi) en la ganadería tropical: 88-99 (en) 9° Seminario de manejo y utilización de pastos y forrajes en sistemas de producción animal. Asociación Venezolana de Producción Animal, Barquisimeto.
RUIZ, E., VÁSQUEZ, J., ZÁRATE, R. & PINEDO, J., 2015.- Aspectos biológicos de Morpho helenor theodorus (Fruhstorfer) (Lepidoptera: Nymphalidae; Morphinae) y Mechanitis polymnia (Linnaeus), (Lepidoptera: Nymphalidae; Ithomiinae), en la Amazonia baja del Perú. Folia Amazónica, 24 (1): 45-54.
SALAZAR J.A., VARGAS, J.I., MORA, A.M. & BENAVIDES, J., 2010.- Identificación preliminar de los Rhopalocera que habitan el Centro Experimental Amazónico (C.E.A.) Mocoa-Putumayo y algunas especies aptas para criar en cautiverio (Insecta: Lepidoptera). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 14 (1): 150-188.
SILVA, K.L., 2005.- Interações evolutivas entre borboletas da tribo Troidini (Papilionidae, Papilioninae) e suas plantas hospedeiras no gênero Aristolochia (Aristolochiaceae): Tesis, Instituto de Biologia, Universidade Estadual de Campinas, São Paulo.
SOTO, J. & IANNACONE, J., 2008.- Efecto de dietas artificiales en la biología de adultos de Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae). Acta Zool. Mex., 24 (2): 1-22.
VALLE, P., 2000.- Toxicología de alimentos. Instituto Nacional de Salud Pública, Centro Nacional de Salud Ambiental, México.VÁSQUEZ, J., ZÁRATE, R. HUIÑAPI, J., PINEDO, J., RAMÍREZ, J.J., LAMAS, G. & VELA, P., 2017.- Plantas alimenticias de
19 especies de mariposas diurnas (Lepidoptera) en Loreto, Perú. Rev. Peru. Biol. 24 (1): 35-42.VÁSQUEZ, R., 1997.- Flórula de las reservas biológicas de Iquitos, Perú. Missouri Botanical Garden, St. Louis.VÁSQUEZ, R. & ROJAS, R., 2004.- Plantas de la Amazonía peruana clave para identificar las familias de Gymnospermae y
Angiospermae. Arnaldoa, 13 (1): 1-261.YOUNG, A.M., 1974.- The rearing of the neotropical butterfly Morpho peleides (Nymphalidae) on peanuts. J. lepid. Soc., 28 (2): 90-99.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
el BosQue de BaVaria (VillaViCenCio, Meta), reVisitado: nueVos reGistros aÑadidos a la lista General de esPeCies (lePidoPtera: rHoPaloCera)*
Julián A. Salazar-e.1, Gregory Nielsen2, Vaclav Pacl3
resumen
Este trabajo pretende ampliar el listado de mariposas diurnas que habitan en Bavaria, un sec-tor de piedemonte ubicado en la vertiente este de la Cordillera Oriental, en el departamento del Meta, mediantes recientes registros de géneros y especies que no se conocían para el lugar. Lo anterior permite afirmar que dicha región es la mejor muestreada en mariposas de toda la Cordillera Oriental alcanzando un total de 473 especies.
Palabras clave: Colombia, Meta, lista de especies, biodiversidad.
tHe BaVaria forest (VillaViCenCio, Meta) reVisited: neW reCords added to tHe CHeCKlist of sPeCies (lePi-
doPtera: rHoPaloCera)
abstract
This work aims to broaden the list of daytime butterflies that live in Bavaria, a foothills area located on the east gradient of the Eastern Cordillera, in the department of Meta, using recent records of genera and species that were not known for the place. This allows to state that this area is the best sampled region in butterflies throughout the Eastern Cordillera reaching a total of 473 species.
Key words: Colombia, Meta, species checklist, biodiversity.
* FR: 22-VI-2016 . FA: .10-V-20171 MSc.,Museo de Historia Natural Universidad de Caldas.de Caldas, Manizales, [email protected] ORCID: 0000-0003-2268-78032 B.S., Aquapro, Km 6 Villaviencio, Meta, [email protected] ORCID: 0000-0003-0728-60593 MSc, Investigador Independiente, [email protected] ORCID: 0000-0002-8183-1417
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 147-169. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR:SALAZAR-E., J.A., NIELSEN, G. & PACL, V., 2017-. El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies (Lepidoptera: Rhopalocera). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 147-169. DOI: 10.17151/bccm.2017.21.2.10

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl148bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
introduCCiÓn
El estudio de las mariposas, relativo a la realización de inventarios o censos con la identidad de especies que habitan diversas regiones, es crucial para conocer la Lepidopterofauna de Colombia y de otros lugares (SALAZAR, 1995; VARGAS, 2003; CAMERO & CALDERÓN, 2007; RODRÍGUEZ-TORRES, 2010; BOOM et al., 2013; GRANOBLES et al., 2013; RODRÍGUEZ & MILLER, 2013; GARCIA-SALIK et al., 2014; GAVIRIA & HENAO, 2014; VARGAS & SALAZAR, 2014). La Cordillera Oriental constituye el enclave montañoso de los Andes más grande y extenso del país. Se extiende en dirección suroeste-nordeste desde el Macizo Colombiano en el departamento del Cauca hasta la Serranía de Perijá en La Guajira, con alturas promedio de 3000 msnm. En este sentido y dada su alta complejidad biológica, esta región ha sido objeto de diversas prospecciones faunísticas dedicadas a mariposas diurnas, en especial en zonas de piedemonte (ABADÍA, 1913; APOLINAR, 1915a, 1915b; FASSL, 1918; BÜRGER, 1919; ANDRADE & AMAT, 1996; FAGUA, 1999; FRAIJA & FAJARDO, 2006; SALAZAR et al., 2010; CASTILLO et al., 2011; GAMBOA & ROMERO, 2014; URBANO et al., 2014).
Partiendo de este propósito, el presente trabajo pretende ampliar la lista preliminar que, sobre este llamativo grupo de insectos, publicara SALAZAR et al. (2003) para un pequeño sector boscoso ubicado muy al pie de la ciudad de Villavicencio, en la vertiente este de la Cordillera Oriental, amén de hacer algunas observaciones adicionales; corrección de identidades y divulgación ilustrada de las mariposas que se encuentran en el bosque de Bavaria y zonas circundantes.
Material Y MÉtodos
El retorno de las exploraciones, para registrar otras especies no incluidas por SALAZAR et al., (2003) en el bosque de Bavaria, se ha realizado por las siguientes personas y fechas: enero 2-11 de 2004 (J. Salazar & J.I. Vargas); abril de 2007 (V. Pacl); septiembre de 2008 (V. Pacl); diciembre 1-7 de 2009 (J. Salazar & J.I. Vargas); junio de 2009 (V. Pacl); marzo 30, julio 3, agosto 30, septiembre 8 y octubre 10 de 2011 (G. Nielsen); mayo 15-24 de 2013 (J. Salazar & G. Nielsen); octubre 25-27 de 2013 (V. Pacl); enero 18-21 de 2015 (V. Pacl); febrero 15 y 29 de 2015 (V. Pacl). Asimismo, se tuvo en cuenta un material antiguo recogido por D. Acosta y nuevamente revisado como parte de sus propias colectas en el lugar. Las diversas capturas se lograron con redes entomológicas de cabo corto y tres redes tipo “tropical net” con palos de aluminio de 3,80 m de longitud. Se emplearon trampas Van Someren-Rydon (RYDON, 1964) suspendidas en el estrato medio alto del bosque, en especial en las inmediaciones de las represas, cebadas con banano o pescado en descomposición. Como complemento se visitaron igualmente otras áreas de piedemonte, relacionadas con el bosque de Bavaria para verificar la presencia de especies registradas, como Chirajara, Pipiral, Las Brisas, río Ocoa, La Cuncia, ríos Acacias y Ariari, los caños Marayal, Camba y La Esmeralda en las vecindades del municipio de San Juan de Arama, Meta.

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 149
En la clasificación del material obtenido usamos los trabajos de SEITZ (1924), LEWIS (1975), SMART (1975), D`ABRERA (1981, 1984, 1987, 1988, 1994, 1995, 2001), JENKINS (1983, 1984, 1985a, 1985b, 1987, 1990), LECROM et al. (2002), LECROM et al. (2004), LAMAS (2004), MIELKE (2005), RODRÍGUEZ et al. (2010), SALAZAR et al. (2010), CONSTANTINO et al. (2013) y las ilustraciones digitales del BOA, encontradas en www.butterfliesofamerica.com. Para la parte botánica se consultaron los estudios de ESPINAL & MONTENEGRO (1963), CUATRECASAS (1989) y CARVAJAL et al. (2007).
Área de estudio
El bosque de Bavaria (Figura 1) se encuentra a cinco minutos de la ciudad de Villavicencio, Meta, en las vecindades del antiguo depósito de la cervecera Bavaria, ubicado en el margen izquierdo de la vía que lleva al municipio de Restrepo, y poco antes de atravesar el puente sobre el río Guatiquía. Sus coordenadas son 04°09’12” de latitud Norte y 73°38’06” de longitud Oeste y a una altura que oscila entre los 467 y 651 msnm, con temperatura de 27ºC promedio y una precipitación media anual de 3.663 mm (IGAC, 1996). El bosque forma parte de un fragmento selvático de piedemonte transicional entre la parte montañosa y los llanos contiguos. La extensión explorada fue de 5 ha y ecológicamente pertenece a la formación vegetal de bosque muy húmedo tropical que, según ESPINAL & MONTENEGRO (1963), ocupa el costado este de la Cordillera Oriental hasta la frontera con Ecuador en el sur.
Vista aérea satelital del bosque de Bavaria con el río Guatiquía a la derecha y la ciudad de Villavicencio (Meta) al pie. Ilustración: G. Nielsen.
figura 1.

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl150bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
Topográficamente el bosque es de buena pendiente, con árboles que alcanzan los 25 metros de altura o más. Alberga dos quebradas: La Rochela, ubicada al lado del camino principal, y Susumuco, cuyas represas pequeñas regulan el nivel de agua que antes se usaba para la fabricación de cerveza. Los lugares de muestreo fueron los siguientes:
A. Borde del bosque: Constituye la floresta que limita con el muro de la fábrica y separada por un camino carreteable que comienza a ascender después de una hora de caminada hasta tornarse estrecho y pedregoso al culminar en el potrero de la vereda La Bendición. El dosel inicial es de unos 15 metros con predominancia de carboneros (Albizzia spp.).
B. entradas: Cuando el camino se sumerge en el bosque cerrado, al llegar a un enorme tanque de almacenamiento, una entrada primaria aparece a unos 80 metros escoltada por un tubo de agua que se desprende del camino y acaba en la quebrada La Rochela. Unos 250 metros más arriba aparece una segunda entrada que va a la represa, con presencia en el sotobosque de abundantes Calathea y Psychotria, entre otros.
C. represas: Sitios donde se permaneció la mayor parte del tiempo y se probaron las trampas cebadas Van Someren-Rydon en un claro de luz y vegetación circundante en las orillas dominadas por palmas de los géneros Jessonia y Phytelephas. Una segunda represa se encuentra alejada a 40 minutos de camino por una trocha cerrada en la actualidad.
D. interiores de bosque: Con zonas de sotobosque y bosque cerrado que ambientan el camino que lleva a las represas. De acuerdo con CARVAJAL et al. (2007), algunos de los elementos florísticos de esta área de piedemonte lo conforman: Pourouma minor Benoist (Cecropiaceae), Stylogyne longifolia Mart. ex. Miq. (Myrsinaceae), Carludovica plicata Klotzch (Ciclantaceae), Ficus spp. (Moraceae), Guarea sp. (Meliaceae), Anthurium sp. (Araceae), Heliconia spp. (Heliconiaceae), Bathysa bracteosa Well. (Rubiaceae), Tapirira guianensis Aul. (Anacardiaceae), Swartzia trianae Benth. (Fabaceae), Cyclanthus bipartitus Port. (Cyclanthaceae), Psychotria sp. (Rubiaceae) y palmas de Socratea exorrhiza Mart., Syagrus sancona Karst. y Jessonia spp.(Arecaceae).
E. Cuchilla del bosque: Incluye la parte más alta del lugar (650-700 msnm), cuyo acceso, bastante pendiente, se hace por un desecho del camino ubicado en el margen izquierdo que parte de la primera finca del sector. Este sube hasta dos torres de conexión eléctrica separadas por dos promontorios emergentes que fueron explorados por uno de nosotros (JS). La vegetación es aquí de bajo porte (3 a 5 metros de altura), dominado por plantas como Ficus sp. (Moraceae), Heliconia sp. (Heliconiaceae), Cecropia sp. (Cecropiaceae), Pteridium aquilinum (Polypodiaceae), Clusia sp. y Achillea sp.

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 151
HallaZGos
SALAZAR et al. (2003) concluían en su momento que la fauna de mariposas estaba conformada por 392 especies, pertenecientes a 221 géneros y citadas en el listado final. Se especulaba además que dicha cifra podría llegar hasta las 400 especies con la inclusión de nuevos registros para el sector. Ahora, con estos nuevos aportes de capturas, el total aumenta a 436 especies, casi todas confinadas en las inmediaciones de las represas. Dentro de los hallazgos más notables fueron: una pareja de los Papiliónidos Pt. zagreus bacchus (Fldr.) ♂ y Pterourus zagreus zagreus (Dbl. ) ♀ capturada en plena cópula, lo que induce a creer que estamos frente a una sola especie altamente polimórfica, registros inusuales de las especies Callicore hesperis (Gdt.), C. pitheas (Latr.), Adelpha pione (Hew.), Prepona laertes (Hbn), Battus ingenuus chalceus (Dyar), Dynamine myrson (Bates); abundantes poblaciones de Methona confusa (Hew.) que anteriormente no se conocían, y el extraordinario descubrimiento del ninfálido pantropical Hypolimnas misippus (L.) de origen africano y de rara presencia en el norte de Sudamérica (LOSADA, 2013). Se suma a lo anterior la confirmación de registros en Bavaria para las especies Prepona eugenes simois (Fldr.), Morpho telemachus iphiclus (Fldr.), Coenophlebia archidona (Hew.), eunica sydonia (Gdt.), e. sophonisba agele (Seitz), Doxocopa agathina (Cr.), Bussa ematheon (Cr.), entre algunas; en contraste con la ausencia notable de otras como Mimoides ariarathes evagorides (Brown & Lamas), Morpho marcus intermedia (Kaye), Brassolis sophorae ardens (Stichel), eurytides dolicaon deileon (Fldr.) y Hamadryas chloe (Stoll).
CoMentarios adiCionales
Respecto a la concentración de especies, SALAZAR et al. (2003) ya habían ofrecido una serie de someros comentarios comparando la riqueza de este sector con respecto a más censos de mariposas realizados para otros piedemontes de la Cordillera Oriental (FAGUA, 1999; FAGUA et al., 1999; ARIAS & HUERTAS, 2001; PÉREZ & FAGUA, 2002). Ahora con 436 especies, el bosque de Bavaria, ubicado en estas montañas de los Andes, constituye uno de los más diversos. No obstante, han aparecido otros estudios donde hacen listados de mariposas como en FRAIJA & FAJARDO (2006), quienes realizaron listados de especies para cinco localidades de piedemonte en el departamento del Meta (Loma Linda, Buenavista, Restrepo, San Antonio y Cafam), registrando 45 especies pertenecientes a 35 géneros. CASTILLO et al. (2011) inventariaron una reserva del municipio de El Calvario (quebrada la Honda, afluente del río Guatiquía), logrando censar 40 especies en 29 géneros. URBANO et al. (2014) más al norte, en el piedemonte Llanero del Casanare, seleccionaron 7 localidades entre los 530 y 1100 msnm. Los resultados, basados en mediciones de diversidad, identificaron 126 pertenecientes a 92 géneros de las 175 especies, ya conocidas para este departamento (YARA-ORTIZ et al., 2011). Finalmente, MESA & GUERRA (2013), en una interesante tesis de grado, listan y figuran 167 especies de 106 géneros

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl152bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
que vuelan entre La Cuncia y La Concepción, dos localidades situadas más al sur, a 7 km de Villavicencio.
Buena parte de las especies reportadas en estos trabajos se registraron para el bosque de Bavaria, excepto las siguientes cuya presencia se espera para el lugar: nymphaloidea- Callicore pygas cyllene (Hewitson, 1847), Paulogramma pyracmon peristera (Hewitson, 1853), Hamadryas fornax (Hübner, 1823), H. februa ferentina (Godart, 1824), H. feronia (Linnaeus, 1758), Marpesia berania (Hewitson, 1852), Panacea procilla (Hewitson, 1854), Chlosyne lacinia (Geyer, 1837), Dynamine vicaria (Bates, 1865); Papilionoidea- Parides aeneas bolivar (Hewitson, 1850) y Parides eurimedes agathocles (Kollar, 1850). Si bien las especies Chlosyne poecile (Felder, 1867), Ch. hipodrome (Geyer, 1837), Sais browni (Takahashi, 1979), S. rosalia (Cramer, 1777) y Caerois chorinaeus (Fabricius, 1775) no son conocidas aquí, se encuentran asociadas a los ambientes de sabana y bosque de galería relativamente próximos al piedemonte.
Hay que mencionar, igualmente, una contribución importante realizada por estudiantes de la Universidad Pedagógica y Tecnológica de Colombia liderados por H. Pulido (GÓMEZ et al., 2010) al muestrear 274 especies del bosque de Bavaria y el Jardín Botánico de Villavicencio, usando redes manuales de colecta y trampas Van Someren-Rydon cebadas con varios tipos de señuelos. Utilizando varios estimadores de diversidad (Sobs, Chao), predicen la posible riqueza de esos lugares en mariposas, con 394 especies, cifra casi igual a las 392 originalmente listadas en SALAZAR et al. (2003). En tanto, el otro estimador (Chao) supone 636 frente a las 473 especies logradas ahora, de hecho ellos afirman en la discusión que las 274 especies representan un 8,3 por ciento de las 3274 reportadas por ANDRADE (2002) para el país, y un 42,6 por ciento de las 642 conocidas en el departamento del Meta. Enfatizan en aquellas que habitan las partes bajas de la Cordillera Oriental, habitando las vecindades de la ciudad de Villavicencio al acercarse al 43 por ciento del total de especies registradas. Si bien ellos profundizan en la riqueza basados en estimadores, no especifican cuántas especies se recogieron en ambos lugares por separado, lo que hubiese facilitado un reconocimiento taxonómico detallado de las mariposas que habitan el bosque de Bavaria.
ColofÓn
El bosque de Bavaria incluido en Villavicencio, así como otros ubicados en el piedemonte este de la Cordillera Orienta; los bosques fragmentados de los ríos Guayuriba y Ariari hasta San Juan de Arama, constituyen mosaicos ecológicos complejos que tienen una biodiversidad insospechada en mariposas y otros organismos, debido a la relativa proximidad de la majestuosa Serranía de la Macarena por un lado y del Parque Nacional Natural Chiribequete por el otro, merecen ser estudiados con urgencia dada la alta concentración de endemismos y especies nuevas que aumentarían

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 153
sin duda el valor biológico de su biota y de los innumerables servicios ambientales que prestarían al ser humano presente en esas regiones.
resultados
apéndice i. lista actualizada de especies y subespecies de rhopalocera registradas para el bosque de Bavaria (basada en SALAZAR et al. 2003)
Abreviación*: Nuevo registro
FAMILIA/ TAXON PaPilionoidea/ PaPilionidaeBattus crassus lepidus (C. & R. Felder, 1861) Battus lycidas (Cramer, 1777) Battus belus varus (Kollar, 1850)* Battus polydamas polydamas (Linnaeus, 1758) eurytides serville acritus (R. & J., 1906) eurytides dolicaon deileon (C. & R. Felder, 1865) Heraclides androgeus androgeus (Cramer, 1775) Heraclides isidorus nymphius (R. & J., 1906) Heraclides anchisiades anchisiades (Esper, 1788)* Heraclides thoas nealces (C. & R., 1906) Heraclides paeon thrason (C. & R. Felder, 1865) Mimoides ariarathes evagorides (B. & Lamas, 1994) Mimoides phaon therodamas (C. & R. Felder, 1865)* Neographium agesilaus agesilaus (Guer-Men., 1835) Parides anchises nielseni (Boll. & Salz., 2001) Parides lysander lysander (Cramer, 1775) Parides neophilus olivencius (Bates, 1861) Parides sesostris sesostris (Cramer, 1779) Parides erithalion browni (Lc. Const. & Salazar, 2001) Protesilaus protesilaus protesilaus (L., 1758)* Pterourus menatius coroebus (C.&R. Felder, 1861) Pterourus zagreus zagreus (Doubleday, 1847) Pterourus zagreus f. bachus (C. & R. Felder, 1865) Pieridae dismorphiinaeDismorphia crisia fedora (Lucas, 1852) Dismorphia amphione praxinoe (Doubleday, 1844)* Moschoneura pinthous methymna (Godart, 1819)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl154bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
enantia lina spp. Pseudopieris viridula viridula (C. & R. Felder, 1861) ColiadinaeAnteos menippe (Hübner, 1819) eurema albula albula (Cramer, 1775) eurema xanthochlora (Kollar, 1850) Phoebis sennae marcelina (Cramer, 1777) Phoebis philea philea (L., 1763) Phoebis argante (Fabricius, 1775) leucidia brephos (Hübner, 1809) Aphrissa boisduvalii (C. & R. Felder, 1861) PierinaeArchonias brassolis critias (C & R. Felder, 1859) Catasticta sisamnus bithyna (Röber, 1924) Glutophrissa drusilla (Cramer, 1777) Itaballia demophile (L., 1763) Melete lycimnia ssp.* Perrhybris lorena (Hewitson, 1852) nYMPHaloidea/ HeliconiinaeDione juno juno (Cramer, 1777) Dryas iulia iulia (Fabricius, 1775) eueides aliphera (Godart, 1819) eueides tales cognata (Weymer, 1890) eueides isabella (Stoll, 1781)* Heliconius erato hydara (Hewitson, 1867) Heliconius ethilla metalilis (Butler, 1873) Heliconius heurippa (Hewitson, 1854) Heliconius numata messene (Felder, 1862) Heliconius numata euphone (Felder, 1862) Heliconius numata aristiona (Hewitson, 1853)* Heliconius ismenius (Latreille, 1817) Heliconius melpomene (Linnaeus, 1758) Heliconius sara sara (Fabricius, 1793) Heliconius wallacei (Reak, 1866) Heliconius xanthocles melete (Felder, 1865) Heliconius hecale ithaca (Felder, 1862) laparus doris viridana (Stichel, 1906) Philaethria dido dido (Linnaeus, 1763) Philaethria pygmalion (Fruhst., 1912)

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 155
danainaelycorea halia atergatis (Doubleday, 1847)* lycorea ilione lamirus (Latreille, 1817)* Danaus plexippus megalippe (Hübner, 1826)* acraeinaeActinote anteas (Doubleday, 1847)*
ithomiinaeAeria eurimedia (Cramer, 1777)* Ceratinia tutia poecila (Bates, 1862) Callithomia lenea ssp. Dircenna loreta (Haensch, 1903) Dircenna dero euchytma (Felder, 1865) episcada polita polita (Weymer, 1899)* Godyris dircenna (Felder, 1865)* Godyris zavaleta zavaleta (Hewitson, 1855) Hypoleria sarepta sidonia (Hewitson, 1857) Hypoleria (aff.) lavinia rhene (G. & S., 1878) Hypoleria ocalea (Doubleday, 1847)* Hypoleria lavinia (Hewitson, 1855) * Hypoleria alema alema (Hewitson, 1857)* Hyposcada illinissa sinilia (H-Schff., 1865) Hypothiris ninonia fimbria (Hewitson, 1855) Hypothiris ninonia antonina (Staudinger, 1884) Hypothiris semifulva fulminans (Butler, 1873) Hypothiris fluonia rowena (Hewitson, 1857) Hypothiris leprieuri (Feisth., 1835) Hypothiris euclea intermedia (Bulter, 1873) Heterosais giulia giulia (Hewitson, 1865) Hyaliris ocna (H-Schff., 1865) Ithomia iphianasa iphianassa (Dbl. & Hew., 1847) Ithomia terra oenanthe (Weymer, 1899) Ithomia agnosia (Hewitson, 1855)* Methona confusa confusa (Butler, 1833)* Mechanitis polymnia veritabilis (Butler, 1873)* Mechanitis mazaeus mazaeus (Hewitson, 1860) Mechanitis mazaeus messenoides (Felder, 1865) Mechanitis mazaeus deceptus (Butler, 1873) Melinaea menophilus hicetas (G. & S., 1879) Melinaea marsaeus messenina (Felder, 1865)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl156bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
Napeogenes inachia johnsoni (Fox & real, 1971) Napeogenes pharo peranthes (Bates, 1862)* Napeogenes apulia apulia (Hewitson, 1858)* oleria onega ilerda (Hewitson, 1854) oleria gunilla lubilerda (Haensch, 1905)* Pseudoscada timna (Hewitson, 1855)* Pteronymia veia (Hewitson, 1853)* Pteronymia primula (Bates, 1862)* Pteronymia sp.* Thyridia psidii psidii (Linnaeus, 1758)* MorphinaeAnthirraea philoctetes philaretes (Felder, 1862) Morpho marcus intermedia (Kaye, 1917) Morpho telemachus iphiclus (Felder, 1862) Morpho achilles patroclus (Felder, 1861) Morpho achilles phokylides (Fruhstorfer, 1912) Morpho menelaus occidentalis (Felder, 1862) BrassolinaeBrassolis sophorae luridus (Stichel, 1902) Caligo eurilochus livius (Staudinger, 1886) Caligo idomeneus (Linnaeus, 1758) Caligo illioneus illioneus (Cramer, 1775) Catoblepia berecynthia berecynthia (Cramer, 1777) opsiphanes cassina numatius (Fruhstorfer, 1912)* opsiphanes invirae intermedius (Stichel, 1902) opsiphanes quiteria phylas (Fruhstorfer, 1912)
limenitidinaeAdelpha iphiclus iphiclus (Linnaeus, 1758) Adelpha epione agilla (Fruhstorfer, 1907) Adelpha pleasure pseudomalea (Hall, 1938) Adelpha cocala cocala (Cramer, 1779) Adelpha boreas boreas (Buller, 1866) Adelpha naxia naxia (Felder, 1867) Adelpha lycorias lara (Hewitson, 1850) Adelpha cytherea olbia (Felder, 1867) Adelpha mesentina chancha (Staudinger, 1886) Adelpha pollina pollina (Furhstorfer, 1915) Adelpha serpa celerio (Bates, 1864) Adelpha malea aethalia (Felder, 1867)

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 157
CharaxinaeAgrias amydon bogotana (Fruhstorfer, 1895) Agrias claudina intermedius (Fassl, 1912) Archaeoprepona demophon (Linnaeus, 1758) Archaeoprepona demophoon gulina (Fruhst., 1904) Archaeoprepona demophoon andicola (Fruhstorfer, 1904)* Archaeoprepona amphimachus (Fabricius, 1775) Archaeoprepona licomedes licomedes (Creamer, 1777) Consul fabius (Cramer, 1776) Consul panariste panariste (Hewitson, 1856)* Hypna clytemnestra negra (Felder, 1862) Cymatogramma glauce glauce (Felder, 1862) Cymatogramma xenocles xenocles (West., 1850) Cymatogramma arginussa onophis (Geyer, 1861) Coenophlebia archidona (Hewitson, 1862) Fountainea ryphea (Cramer, 1775) Fountainea eurypyle (Felder, 1882) Memphis pseudiphis (Staudinger, 1887) Memphis philumena philumena (Doubl., 1849) Memphis offa offa (Druce, 1897) Memphis acidalia acidalia (Hübner, 1819) Memphis acidalia memphis (Felder, 1867)* Pseudocharaxes xenocrates (Westw., 1850) Prepona laertes demodice (Godart, 1824)* Prepona laertes philipponi (Le Moult, 1932) Prepona pylene simois (Felder, 1867) Prepona pheridamas (Cramer, 1777) Siderone galanthis thebais (Felder, 1862) Zaretys itys (Cramer, 1777)
nYMPHaloidea (sensu lato)Antanassa drusilla (Fabricius, 1861) Anartia amathea (Linnaeus, 1758) Anartia jatrophae (Linnaeus, 1763)* Biblis hyperia (Cramer, 1779) Callicore lyca bella (Röber, 1924) Callicore astarte stratiotes (Felder, 1861) Callicore hesperis (Guer-Men., 1844)* Callicore pitheas (Latreille, 1813)* Catacore kolyma (Hewitson, 1852) Castilia eranites (Hewitson, 1857) Castilia ofella (Hewitson, 1864)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl158bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
Catonephele numilia numilia (Cramer, 1775) Catonephele acontius acontius (Linnaeus, 1777) Catonephele salambria (Felder, 1861) Colobura dirce (Linnaeus, 1758) Diaethria clymena (Cramer, 1775) Diaethria gabaza (Hewitson, 1855) Doxocopa zunilda felderi (G. & S., 1884) Doxocopa agathina (Cramer, 1777) Doxocopa laurentia seraphina (Hübner, 1825) Dynamine agacles core (Röber, 1915) Dynamine anubis pieridoides (Felder, 1867) Dynamine racidula racidula (Felder, 1852) Dynamine gisella (Hewitson, 1857) Dynamine myrson (Doubleday, 1849)* eresia clio clio (Linnaeus, 1758)* eresia eunice gudruna (Röber, 1913) eunica concordia (Hewitson, 1852) eunica phasis (Felder, 1862) eunica sophonisba agele (Seitz, 1915) eunica sydonia sydonia (Godart, 1824) eunica norica norica (Hewitson, 1852) eunica eurota eurota (Cramer, 1775)* eunica volumna celma (Hewitson, 1852)* epiphile lampethusa zipa (Mengel, 1899) Hamadryas amphinome amphinome (L., 1767)* Hamadryas arinome arinome (Lucas, 1853) Hamadryas laodamia saurites (Fruhst., 1916) Hamadyas chloe chloe (Stoll, 1787) Haematera pyrame pyrame (Hübner, 1819)* Historis orion dious (Lamas, 1995) Historis acheronta acheronta (Fab., 1775)* Hypanartia lethe (Fab., 1793) Hypolimnas misippus (L., 1764)* Junonia evarete (Cramer, 1779) Marpesia petreus petreus (Cramer, 1776) Marpesia chiron marius (Cramer, 1779) Nessaea hewitsoni hewitsoni (Felder, 1859) Nica flavilla canthara (Doubleday, 1849)* Panacea prola prola (Doubleday, 1848) Peria lamis (Cramer, 1779) Pyrrhogyra amphiro amphiro (Bates, 1865) Pyrrhogyra crameri nautaca (Fruhst., 1908)*

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 159
Siproeta stelenes meridionalis (Fruhst., 1909) Smyrna blomfildia (Fab., 1781) Tigridia acesta fulvescens (Butler, 1873) Temenis laothoe hondurensis (Fruhst., 1907) Temenis laothoe f. violetta (Cramer, 1777) Temenis pulchra pallidior (Oberthür, 1901) Tegosa anieta (Hewitson, 1852) Victorina elissa (Hübner, 1819) satyrinaeChloreuptychia arnaca (Fabricius, 1776) Chloreuptychia herseis (Godart, 1824) Cithaerias phantoma (Fassl, 1922) euptychia mollina (Hübner, 1813) euptychia sp. erichtoides antonina (Felder, 1867) Haetera piera negra (Felder, 1862) Hermeuptychia hermes (Fabricius, 1775) Magneuptychia newtoni (Hall, 1939) Magneuptychia ocypete (Fabricius, 1776) Manataria hercynia maculata (Hoppffr., 1874)* Pareuptychia ocirrhoe ocirrhoe (Fab., 1776) Pareuptychia lydia (Cramer, 1777)* Pierella lamia chalybea (Godman, 1905) Posttaygetis penela penelea (Cramer, 1777) Taygetis cleopatra (Felder, 1867) Taygetis laches laches (Fab., 1793) Taygetis mermeria mermeria (Cramer, 1776)* Taygetis rufomarginata (Staudinger, 1888) Taygetis thamyra (Cramer, 1779)* Cepheuptychia cephus (Fabricius, 1775)* riodinidaeAlesa amesis (Cramer, 1777) Alesa prema (Godart, 1824)* Ancyluris aulestes jocularis (Stichel, 1909) Ancyluris aulestes callaghani (Lamas, Const. & Salz, 2102)* Ancyluris meliboeus meliboeus (Fab., 1776)* Ancyluris meliboeus eudaemon (Stichel, 1910)* Ancyluris etias (Saunders, 1859)* Amarynthis meneria (Cramer, 1776) Anteros formosus (Cramer, 1777)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl160bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
Anteros acheus (Stoll, 1781) Argyrogrammana stilbe (Godart, 1824) Brachyglenis esthema (Felder, 1862) Calephelis velutina (G. & S., 1878) Calospila emylius (Cramer, 1775) Chamaelimnas briola (Bates, 1868) Chorinea octavius (Fab., 1787) Crocozona coecias arcuata (Godman, 1903) Cremna actoris (Cramer, 1776) Cyrenia martia martia (Westwood, 1851) emesis cypria (Felder, 1861) emesis lucinda (Cramer, 1775) emesis cerea (Linnaeus, 1767) emesis melancholica (Stichel, 1916) eurybia halimede (Hübner, 1807) eurybia donna (Felder, 1862) eurybia molochina hyacinthina (Stich., 1910) eurybia patrona (Weymer, 1875) eurybia dardus (Fab., 1787) eurybia dardus annulata (Stich., 1910) eurybia annius ssp. euselasia eumenes (Hewitson, 1853) euselasia issoria (Hewitson, 1869) euselasia orfita (Cramer, 1777 euselasia hygenius (Stoll, 1787) euselasia uzita (Hewitson, 1853) euselasia artos (H-Schaff., 1853) euselasia eugeon (Hewitson, 1856) euselasia fervida hahneli (Staudinger, 1888) euselasia eulione (Hewitson, 1856) euselasia toppini (Sharpe, 1915) euselasia mys (H-Schaff., 1853) euselasia midas (Fab., 1775) euselasia teleclus (Stoll, 1787) euselasia corduena corduena (Hewitson, 1874) euselasia labdacus labdacus (Stoll, 1870) euselasia eucrates (Hewitson, 1872) euselasia albomaculiga (Callaghan, 1999) euselasia eumedia (Hewitson, 1853) euselasia uria angustifasciata (Lathy, 1926)* eunogyra satyrus (Westwood, 1851) Detritivora cleonus (Stoll, 1781)

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 161
Hades noctula (Westwood, 1851) Hyphilaria parthenis (Westwood, 1851) Isapis argyrtus (Doubleday, 1777) Ithomeis aurantiaca corena (Felder, 1862)* Ihomiola floralis cascella (Hewitson, 1870)* lasaia agesilas agesilas (Latreille, 1809) lasaia pseudomeris (Clench, 1972)* leucochimona icare icare (Hübner, 1819) Melanis marathon assimulata (Stichel, 1910) Melanis electron (Fab., 1793) Melanis smithiae xarifa (Hewitson, 1853) Menander hebrus (Cramer, 1775) Menander pretus picta (G. & S., 1886) Mesene leucophrys (Bates, 1868) Mesene nola (Cramer, 1777) Mesosemia machaera (Hewitson, 1860) Mesosemia cippus (Hewitson, 1859) Mesosemia synnephis (Stichel, 1909) Mesosemia zanoa orthia (Stichel, 1915) Mesosemia philocles jeziela (Butler, 1869) Mesosemia magete humboldtiana (Rod.Sal & Const, 2009)* Mesosemia thymetus thymetina (Bulter, 1869)* Metacharis regalis (Butler, 1867) Monethe albertus albertus (Felder, 1862) Nymphidium cachrus (Stoll, 1787) Nymphidium leucosia (Hübner, 1806) Nymphidium plinthobaphis (Stichel, 1910) Nymphidium fulminans (Bates, 1868) Nymphidium lysimon (Stoll, 1790) Nymphidium caricae (Linnaeus, 1758) Nymphidium baeotia (Hewitson, 1853)* Nymphidium omois (Hewitson, 1865)* Napaea eucharila (Bates, 1867)* Parcella amarynthina (Felder, 1865) Panaropsis thyatira (Hewitson, 1853)* Pheles strigosa (Staudinger, 1876) Perophthalma tullius (Fab., 1767) Pirascca tyriotes (G. & S., 1878) Riodina lysippus (Linnaeus, 1758) Rhetus periander (Cramer, 1777) Rhetus arcius (Linnaeus, 1763) lyropteryx apollonia (Westwood, 1851)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl162bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
Semomesia croesus lacrimosa (Stichel, 1915) Sarota chrysus (Stoll, 1781) Sarota gyas (Cramer, 1775) Sarota lasciva (Stichel, 1911) Setabis myrtis (Westwood, 1851) Setabis lagus ssp.* Symmachia basilissa (Bates, 1868) Symmachia rubina (Bates, 1866) Symmachia tricolor (Hewitson, 1867) Symmachia batesi (Staudinger, 1887)* Syrmatia lamia (Bates, 1868) Synargis abaris (Cramer, 1776) Synargis calyce (Felder, 1862) “Siseme” pedias (Godman, 1903)* Terathopthalma phelina (Felder, 1862) Theope barea (G. & S., 1878) Theope pedias (H-Schaff., 1853) Theope terambus (Godart, 1824) Theope nycteis (Westwood, 1851) Theope philotes (Westwood, 1851) Theope azurea (Bates, 1868)* Theope syngenes ssp.* Xenandra pelopia (Druce, 1890) Xynias lithosina (Bates, 1868) lYCaenidaeArcas imperialis (Cramer, 1775) Arawacus aetolus (Sulzer, 1776) Arawacus dolylas (Cramer, 1776) Allosmaitia myrtusa (Hewitson, 1867) Brevianta ematheon (Cramer, 1777) Calycopis pisis (G. & S., 1887) Calycopis cerata (Hewitson, 1877) Calycopis calus (Godart, 1824) Calycopis lerbela (Field, 1967) Chalybs janias (Cramer, 1779) Celmia celmus (Cramer, 1775) Contrafacia imma (Pritw., 1865) Cyanophrys pseudolongula (Clench, 1944) Cupathecla cupentus (Stoll, 1781) evenus regalis (Cramer, 1775) Gargina gargophia (Hewitson, 1877)

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 163
Janthecla leea (Robb. & Venables, 1991) Janthecla sista (Hewitson, 1867) Michaelus jebus (Godart, 1824) Nicolaea besidia (Hewitson, 1868) ostrinotes halciones (Butler & Druce, 1872) Panthiades bitias (Cramer, 1777) Panthiades aeolus (Fab., 1775) Pseudolycaena marsyas (Linnaeus, 1758) Paiwarria telemus (Cramer, 1775)* Terenthina terentia (Hewitson, 1868) Theritas lisus (Stoll, 1790) Theritas hemon (Cramer, 1775) Theritas pegeus (Hewitson, 1865) Theritas mavors (Hübner, 1818) Thestius pholeus (Cramer, 1777) Tmolus mutina (Hewitson, 1867) Theclopsis lydus (Hübner, 1819)
HesPeriidae (sensu lato) Aides aegita (Hewitson, 1866) Achlyodes mithridates thraso (Hbn., 1831) Anthoptus epictetus (Fab., 1793) Anisochoria pedaliodina polysticta (Mabille, 1877)* XAntigonus nearchus (Latreille, 1817)* Apaustus gracilis (Felder, 1867) Asphita agenoria (Hewitson, 1876) Astraptes hopfferi (Plotz, 1881) Astraptes enotrus (Cramer, 1781) Astraptes alardus (Stoll, 1790) Astraptes galesus (Mabille, 1888) Astraptes aulestes (Cramer, 1781) Astraptes creteus (Cramer, 1780) Augiades crinisus (Cramer, 1780) Autochton neis (Geyer, 1832) Bolla cupreiceps (Mabille, 1891) Calpodes ethlius (Stoll, 1782) Camptopleura theramenes (Mabille, 1877) Cobalus sp.1 Cyclosemia anastomosis (Mabille, 1878) Celaenorrhinus eligius (Stoll, 1781)* Dubiella fiscella (Hewitson, 1877) Dyscophellus euribates (Stoll, 1782)

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl164bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
ebusus ebusus (Stoll, 1780) entheus priassus (Linnaeus, 1758) entheus genthius (Mabille, 1777) entheus sp. elbella azeta (Hewitson, 1866) elbella intersecta (Mabille, 1869) epargyreus exadeus (Cramer, 1779) eracon paulinus (Stoll, 1782) euphyes sp. eutychide sp. Helias cama (Evans, 1953)* Heliopetes alana (Reak, 1868) Hyalothyrus neleus (Linnaeus, 1758) Jemadia gnethus (Fab., 1781)* Marela tamyroides (Felder, 1867)* Moeris striga (Geyer, 1832) Myscelus pardalina (Felder, 1867) Mylon pelopidas (Fab., 1793) Milanion hemes (Cramer, 1772) Nascus broteas (Cramer, 1870) Nisoniades ephora (H-Schaff., 1870) Naevolus orius (Mabille, 1883) Nyctelius nyctelius (Latreille, 1824) Noctuana noctua (Felder, 1867) Noctuana stator (Godman, 1899)* orthos gabina (Godman, 1901) Papias phaeomelas (Hübner. 1831) Papias subcostulata (H-Schaff., 1870) Perichares seneca crotona (Hewitson, 1866) Perichares adela (Hewitson, 1867)* Pellicia dimidiata (H-Schaff., 1870) Phareas coeleste (Westwood, 1852) Phanus vitreus (Stoll, 1780) Paracarystus menestries (Ltareille, 1824) Phytonides crameri (Mab. & Boullet, 1917) Poligonus leo (Gmelin, 1790) Potamanaxas sp. Pyrgus adepta (Plotz, 1884)* Passova passova styx (Möschler, 1875) Pseudonascus paulliniae (Sepp, 1842)* Quadrus cerealis (Stoll, 1782) Quasimellana eulogius (Plotz, 1882) Saliana salius (Cramer, 1775)

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 165
Saliana fischer (Latreille, 1824)* Saliana triangularis (Kaye, 1914)* Saliana fusta (Evans, 1955)* Staphylus sp. Talides sergestus (Cramer, 1775) Thracides cilissa (Hewitson, 1867)* Tarsoctenus praecia rufibasis (Boullet, 1910) Typhedanus crameri (McHenry, 1960) Urbanus doryssus (Swainson, 1833) Urbanus dorantes (Stoll, 1790) Virga sp. Vehilius sp. Vettius triangularis (Hübner, 1831) Vettius phyllus (Cramer, 1777)*
aGradeCiMientos
En especial a David Acosta Silva por ceder amablemente su archivo fotográfico con especies concernientes de la región, a Dora Acosta y José Ignacio Vargas en la colaboración con las capturas. Al profesor Hannier Pulido y los estudiantes Camilo A. Gómez Jiménez, Naisla T. Manrique, Oscar E. Garzón, Oscar F. Moreno y William Bravo Pedraza, todos ellos de la Universidad Pedagógica y Tecnológica de Colombia, por el envío del informe no publicado sobre las especies de Bavaria y del Jardín Botánico de Villavicencio. Literatura e identificación adicional fueron debidas en su momento a Les Catchick, Sonia Cárdenas y José I. Vargas.
referenCias
ABADÍA, G., 1913.- Excursión Científica. Bol. Soc. Cienc. Nat. Inst. la Salle, 1 (2): 58-62.ANDRADE, M.G., 2002.- Biodiversidad de las Mariposas (Lep. Rhop.) de Colombia: 153-172 (en) COSTA, VANIN, LOBO &
MELIC (eds.) PrIBeS-SeA. Monografías Tercer Milenio, Zaragoza, España.ANDRADE, M.G. & AMAT, G., 1996.- Un estudio regional de las mariposas altoandinas en la Cordillera Oriental (en) ANDRADE,
AMAT & FERNÁNDEZ (eds.) Insectos de Colombia, estudios escogidos. Ac. Col. Cienc. Nat., 10: 149-180 + figs. APOLINAR, M., 1915a.- Datos para la Entomología. Bol. Soc. Cienc. Nat. Inst. la Salle, 3 (9): 136-139.APOLINAR, M., 1915b.- Miscelánea, capturas interesantes. Bol. Soc. Cienc. Nat. Inst. la Salle, 3 (9): 141-144.ARIAS, J.J. & HUERTAS, B.C., 2001.- Mariposas diurnas de la Serranía de los Churumbelos, Cauca. Distribución altitudinal y
diversidad de especies (Lep. Rhop. ). Rev. Col. ent., 27 (3-4): 169-176.BOOM, C., SEÑA, L., VARGAS, M. & MARTÍNEZ, N., 2013.- Mariposas Hesperioidea y Papilionoidea (Ins. Lep.) en un fragmento
de bosque seco tropical, Atlántico, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 17 (1): 149-167.BÜRGER, O., 1919.- Reissen eines Naturforshers in Tropischen Südamerika: 469 pp. Leipzig.CAMERO, E. & CALDERÓN, A., 2007.- Comunidad de mariposas diurnas (Lep. Rhop.) en un gradiente altitudinal del cañón del
Río Combeima-Tolima, Colombia. Acta Biol. Col., 12 (2): 95-110.CARVAJAL, L., PATARROYO, J., PUENTES, D. & VALERO, M.J., 2007.- Caracterización florística y estructural del piedemonte
llanero, primera parte.: 130 pp. Cormacarena, Gobernación del Meta, U. Distrital Francisco José de Caldas.CASTILLO, R., CABRERA, D. & LOMBANA, E., 2011.- Caracterización preliminar de la Lepidopterofauna (Ins. Rhop.) presente
en la reserva privada refugio del oso de anteojos (Calvario-Meta, Colombia). Cultura Científica, 28-35 + figs. CONSTANTINO, L. SALAZAR, J.A. & RODRÍGUEZ, G., 2013.- Estudio sobre el género Ancyluris Hübner, 1819 en Colombia
y descripción de nuevos taxa (Lep. Riodinidae). Bol. Mus. Hist. Nat. U. de Caldas, 16 (2): 209-235 + figs.CUATRECASAS, J., 1989.- Aspectos de la vegetación natural de Colombia. Pérez-Arbelaezia, 2 (8): 155-284.D´ABRERA, B., 1981-2001.- Butterflies of the Neotropical Región, 7 parts. Lansdowne & Hill House, Victoria.

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl166bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
ESPINAL, L.S. & MONTENEGRO, E., 1963.- Formaciones vegetales de Colombia (Memoria explicativa sobre el mapa ecológico). IGAC. 201 pp.
FAGUA, G., 1999.- Variación de mariposas y hormigas de un gradiente altitudinal de la Cordillera Oriental: 317-362 (en) Insectos de Colombia, II.
FAGUA, G., AMARILLO, A. & ANDRADE, M.G., 1999.- Mariposas como indicadores del grado de intervenci6n en la cuenca del Río Pato, II: 285-315 (en) Insectos de Colombia.
FASSL, A.H., 1918.- Die Vertikale verbreitung der Lepidopterenin der Columbischen Ost-Cordillere. ent. Rundsch., 35 (1): 1-4, 30-31, 11: 48-50.
FRAIJA, N. & FAJARDO, G., 2006.- Caracterización de la fauna del orden Lepidoptera (Rhop.) en cinco diferentes localidades de los llanos orientales colombianos. Acta Biol. Col., 11 (1): 55-68.
GAMBOA, W. & ROMERO, L., 2014.- Caracterización entomológica de las especies predominantes en el área boscosa de la Unad Cead, Acacias, Meta: 80 pp. + figs.: Tesis, Ing. For., Univ. Nacional Abierta y a Distancia (UNAD).
GARCIA-SALIK, L., CARNEIRO, E., DOLIBAINA, R., DIAS, F.M., RIBEIRO, L., CASAGRANDE, M.M. & MIELKE, O., 2014.- Borboletas de la Estacao Ecológica do Caiuá (Lep. Hesp. Pap.). SHIlAP, Revta lepid., 42 (166): 265-280.
GRANOBLES, O., PEÑUELA, M., OSPINA, K. & GIRALDO, A., 2013.- Diurnal Butterflies (Lep. Rhop.) from corregimiento Santa Inés, Yumbo-Colombia. Ambiente y Sostenibilidad, 3: 15-25 + figs.
GAVIRIA, F. & HENAO, E., 2014.- Diversidad de mariposas diurnas (Hesp-Pap.) en tres estados sucesionales de un bosque húmedo premontano bajo, Tuluá, Valle del Cauca. Rev. Fac. Cienc. U. Nac. de Col., Medellín, 3 (2): 49-80.
GÓMEZ, J., MANRIQUE, N., GARZÓN, O., MORENA, O. & BRAVO, W., 2010.- Aproximación a la diversidad de la fauna de lepidópteros diurnos del Jardín Botánico y Reserva de Bavaria, Villavicencio, Meta, Colombia. Inf. 31 pp. + figs. Universidad Pedagógica y Tecnológica de Colombia.
IGAC, 1996.- Diccionario Geográfico de Colombia. II: 1026., III: 1364-1369. Instituto Geográfico Agustín Codazzi, Bogotá.JENKINS, D.W., 1983-1990.- Revision of Neotropical Nymphalidae. Bull. Allyn Mus., (varios números).LAMAS, G., (ed.), 2004.- Atlas of the Neotropical lepidoptera, Checklist: Part 4ª, Hesperioidea-Papilionoidea: 439 pp. ATL, Scient.
Publ. Gainesville.LE CROM, J.F., CONSTANTINO, L.M. & SALAZAR, J.A., 2002.- Mariposas de Colombia. Tomo I. Papilionidae: 112 pp. + pls.
Carlec, Ltda.LE CROM, J.F., LLORENTE, J., CONSTANTINO, L.M., & SALAZAR, J.A., 2004.-Mariposas de Colombia. Tomo II. Pieridae:
112 pp. + pls. Carlec, Ltda.LEWIS, H.L., 1975.- las Mariposas del Mundo: 23-88. Omega, Barcelona.LOSADA, M.E., 2013.- Hypolimnas misippus (Linnaeus, 1764) (Lep. Nymph.), una especie de distribución pantropical presente en
Venezuela. entomotropica, 28 (3): 237-241 + figs.MESA, D.L. & GUERRA, M.A., 2013.- Mariposas del corregimiento la Concepción, Villavicencio, Meta (lep. Pap.) una lista anotada:
Tesis, Biol., Un. Pedagógica Nacional. 112 pp. + figs.MIELKE, O.H.H., 2005.- Catalogue of the American Hesperioidea. 6 vols.: 1536 pp. Sociedade Brasileira de Zoologia. NIELSEN, G. & SALAZAR, J.A., 2014.- On the presence of certain rare species of Riodinidae (Lep.Papilionoidea) in a forest
fragment on the Eastern Slope of the colombian Cprdillera Oriental and a a new regional list. Bol. Cien. Mus. Hist. Nat. u. de Caldas, 18 (2): 203-226 + figs.
PÉREZ, G. & FAGUA, G., 2002.- Distribución geográfica de Papilionoideos (Lep.) del flanco oriental de la Cordillera Oriental colombiana. entomólogo, 30 (94): 38.
RODRÍGUEZ-TORRES, D., 2010.- Caracterizaciones biológicas en la hacienda Macondo (Mapiripán, Meta). orinoquia 14 sup., (1): 18-27.
RODRÍGUEZ, G., CONSTANTINO, L. & SALAZAR, J.A., 2010.- Estudio sobre las especies colombianas de Anteros Hübner [1819] (Lep: Riodinidae). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 14 (1): 221-251 + figs.RODRIGUEZ, G. & MILLER, H., 2013.- Inventario preliminar de los Rhopalocera de Mitú, Vaupés, Colombia (Ins. Lepidoptera). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 17 (1) : 196-218 + figs.
RYDON, A.H.B., 1964.- Notes on the use of butterfly traps in East Africa. J. lepid. Soc., 18 (1): 51-58.SALAZAR, J.A., 1995.- Lista preliminar de las mariposas que habitan el departamento del Putumayo. Notas sobre su distribuci6n
en la zona Andina. Colombia Amazónica, 8 (1): 11-69.SALAZAR, J.A., GIRALDO, M. & VARGAS, J.I., 2003.- Más observaciones sobre la concentración de mariposas territoriales en
cumbres de cerros colombianos y especies residentes en el bosque de Bavaria, Villavicencio, Meta, su distribución espacial y trofismo. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 7: 255-317 + figs.
SALAZAR, J.A., RODRÍGUEZ, G. & CONSTANTINO, L., 2010.- Contribución al conocimiento del género Mesosemia Hübner [1819] en Colombia y descripción de nuevos taxa (Lepidoptera: Riodinidae).
SEITZ, A., 1924.- (editor) Die Gross-Schmettterlinge er Erde, V. Exostische fauna, pls., Alfred Kernen, Stuttagart.SMART, P., 1975.- The International Butterfly Book: 275 pp. + pls. Salamander Books and Crowell comp., London-NY.URBANO, P., MUNÉVAR, J., MAHECHA, O. & HINCAPIÉ, E., 2014.- Diversidad y estructura de las comunidades de Lepidoptera
en la zona del ecotono entre el piedemonte llanero y la sabana inundable en Casanare-Colombia (Lep. Pap.). SHIlAP, Revta lepid., 42 (167): 433-437.
VARGAS, J.I., 2003.- Mariposas diurnas de Punta Soldado, Buenaventura, Valle. Contribución a su Historia Natural. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 7: 195-207 + figs.
VARGAS, J.I. & SALAZAR, C.A., 2014.- Biodiversidad y mariposas en una región del alto Chocó, San José del Palmar, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 18 (1): 259-284 + figs.
YARA-ORTIZ, C. PEÑA, J. & URBANO, P., 2011.- Biodiversidad del Casanare: 286 pp.+ figs. Gobernación de Casanare, WWF, Colombia, Bogotá.

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 167
fig. 1. Morpho telemachus iphiclus ♂. fig. 2. Morpho achilles patroclus ♂. fig. 3. Morpho achilles phokylides ♂. fig. 4. Morpho marcus intermedia ♂. fig. 5. Morpho menelaus occidentalis ♂. fig. 6. Antirrhaea philoctetes philaretes ♂.

Julián A. Salazar-e., Gregory Nielsen, Vaclav Pacl168bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
47-1
69
fig. 7. Nessaea hewitsoni ♂. fig. 8. Hypolimnas misippus ♂. fig. 9. Caligo idomeneus ♀. fig. 10. Caligo eurilochus livius ♂. fig. 11. Agrias amydon bogotana ♂. fig. 12. Agrias claudina intermedius ♂. fig. 13. Prepona pylene simois ♂. fig. 14. Archaeoprepona licomedes licomedes ♀.

El bosque de Bavaria (Villavicencio, Meta), revisitado: nuevos registros añadidos a la lista general de especies... 169
fig. 15. Parides anchises nielseni ♂. fig. 16. Coenophlebia archidona ♂. fig. 17. Pterourus zagreus f. bacchus ♂. fig. 18. eueides tales cognata ♂. fig. 19. Methona confusa confusa ♂. fig. 20. Hamadryas chloe chloe ♂. fig. 21. Brevianta ematheon ♂. fig. 22. Semomesia croesus lacrimosa ♂. fig. 23. Cyrenia martia ssp. ♂. fig. 24. Callicore lyca bella ♂. (Láminas prep. G. Nielsen).


ZooloGía inVerteBradosInvertebrate Zoology


Boletín CientífiCoCentro de Museos
Museo de Historia natural
una nueVa esPeCie de Pharneuptychia forster, 1964 del Área natural ÚniCa los estoraQues –norte de santander, ColoMBia– (lePidoPtera: nYMPHalidae:
satYrinae)*
efraín Reinel Henao-Bañol1, luis Hernando Meneses2
abstract
Se describe una nueva especie para el género Pharneuptychia Forster, 1964 con base en ejem-plares colectados en el proyecto “Caracterización de la entomofauna del Área Natural Única Los Estoraques (ANULE)”. La especie propuesta es hasta el momento propia del área de estudio, sin embargo, varias especies similares no descritas son encontradas en cercanías hacia el límite con el departamento de Santander y zonas limítrofes con Venezuela. Esta especie es propia de áreas abiertas del Bosque seco en la provincia de Ocaña, Norte de Santander, Colombia.
Key words: mariposas, Pharneuptychia, nueva especie, Norte de Santander, Colombia
a neW sPeCies of Pharneuptychia forster, 1964 froM tHe estoraQues natural area, nortH of santander-
ColoMBia (lePidoPtera: nYMPHalidae: satYridae)
resumen
A new species is described for the genus Pharneuptychia Forster, 1964 based on specimens collected in the project “Characterization of the entomofauna of the Área Natural Unica Los Estoraques (ANULE)”. The proposed species is so far typical from the study area, but several similar species not described are found close to the border with the department of Santander and areas bordering Venezuela. This species is characteristic of open areas of the Dry Forest in the province of Ocaña, Norte de Santander, Colombia.
Palabras clave: Butterflies, Colombia, euptychiina, Estoraques, New species, Norte de Santander, Pharneuptychia
* FR 2-V-2017: . FA: 4-IX-2017.1 Doctor en Ciencias-Biología. Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. E-mail: [email protected] ORCID: 0000-0002-5482-61452 Ingeniero Agrónomo. Parques Nacionales Naturales de Colombia, Área Natural Única Los Estoraques. Norte de Santander, Colombia. E-mail: [email protected] ORCID: 0000-0002-4905-390X
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 173-183. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: HENAO-BAÑOL, E.R. & MENESES, L.H., 2017.- Una nueva especie de Pharneuptychia Forster, 1964 del Área Natural Única Los Estoraques –Norte de Santander, Colombia– (Lepidoptera: Nymphalidae: Satyrinae). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 173-183. DOI: 10.17151/bccm.2017.21.2.11

efraín Reinel Henao-Bañol, luis Hernando Meneses174bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
73-1
83
introduCCiÓn
Los Euptychiina son una de las subtribus más ricas en especies de los Satyrinae que contienen más de 400 especies, distribuidas principalmente en la región Neotropical (LAMAS & VILORIA, 2004), agrupados en 47 géneros, 14 de los cuales son monotípicos (MURRAY & PROWELL, 2005; MARÍN et al., 2011).
Habitan principalmente bosque de tierras bajas por debajo de los 1000 m de altitud, con la excepción quizás del género Forsterinaria Grey, 1973 (PEÑA & LAMAS 2005; PULIDO & ANDRADE 2008) y algunas especies aisladas de otros géneros. Los estudios filogenéticos a nivel molecular de PEÑA et al., (2006), utilizando un extenso muestreo en Satyrinae, encontraron que los Euptychiina son un grupo monofiletico (PEÑA et al., 2010, MARÍN et al., 2017), dentro del cual se ubica el género Pharneuptychia FORSTER (1964), el cual es un pequeño grupo de mariposas que cuenta con seis especies (LAMAS 2004, WARREN et al., 2016), sin subespecies y varias por describir. Son características de hábitats secos o semidesérticos en áreas abiertas propias de pastizales o pajonales.
Las alas presentan una forma semicuadrada con cuatro ocelos en AP y de uno a tres en AA. La mayoría de las especies presentas manchas con escamas iridiscentes de color amarillo o plata brillante.
FORSTER (1964), propuso el género donde destaca la forma de las alas (con excep-ción de la especie P. innocentia (C. Felder & R. Felder, 1867), que es semiredonda), desarrollo de ocelos en AA y AP y en un ocelo notorio oscuro en APD, las valvas amplias o anchas y Gnathos delgado.
Pharneuptychia amerita un revisión sistemática, ya que su monofilia no ha sido evalua-da y se desconoce las relaciones filogenéticas de las especies, al observar la constancia de algunos caracteres de alas y de los órganos genitales es posible que sea un grupo parafiletico, como lo encontrado por NAKAHARA et al., 2016 desde el punto de vista molecular. En la actualidad se lleva a cabo un estudio sobre la filogenia molecular y la historia biogeográfica para este género y otros géneros cercanos (E. BARBOSA et al., in prep.)
Las especies descritas en este género han sido citadas de Argentina, Brasil, Bolivia y Venezuela siendo propias de la región neotropical sudamericana, con un registro en la isla de Chacachacare en la republica de Trinidad y Tobago (COCK, 2014). El género en Colombia ha sido citado por MARÍN & SOTO (2009) para Antioquia en el Municipio de Santa Fé de Antioquia y SÁNCHEZ & GANTIVA (2014) para el Cerrejón (La Guajira).
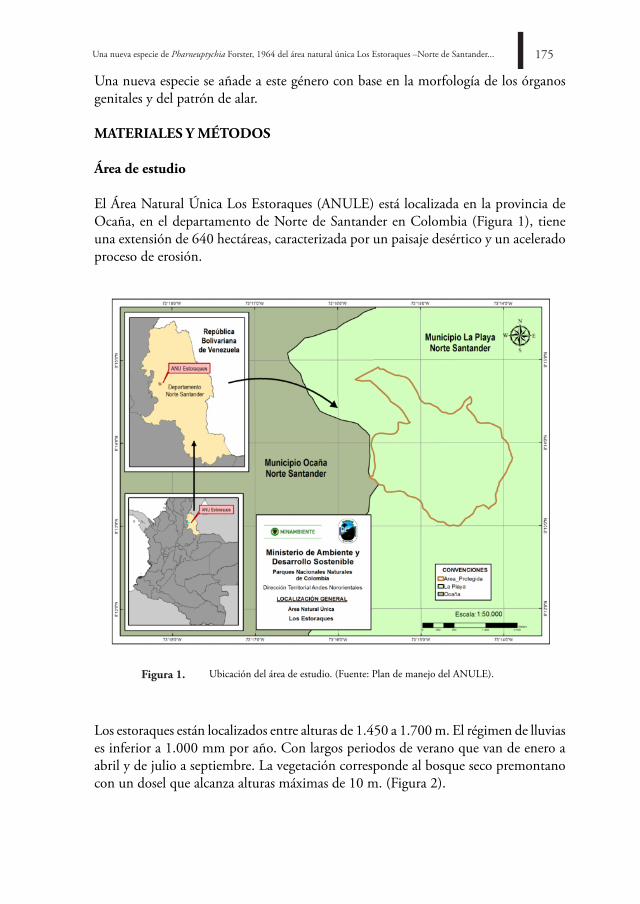
Una nueva especie de Pharneuptychia Forster, 1964 del área natural única Los Estoraques –Norte de Santander... 175
Una nueva especie se añade a este género con base en la morfología de los órganos genitales y del patrón de alar.
Materiales Y MÉtodos
Área de estudio
El Área Natural Única Los Estoraques (ANULE) está localizada en la provincia de Ocaña, en el departamento de Norte de Santander en Colombia (Figura 1), tiene una extensión de 640 hectáreas, caracterizada por un paisaje desértico y un acelerado proceso de erosión.
Ubicación del área de estudio. (Fuente: Plan de manejo del ANULE). figura 1.
Los estoraques están localizados entre alturas de 1.450 a 1.700 m. El régimen de lluvias es inferior a 1.000 mm por año. Con largos periodos de verano que van de enero a abril y de julio a septiembre. La vegetación corresponde al bosque seco premontano con un dosel que alcanza alturas máximas de 10 m. (Figura 2).

efraín Reinel Henao-Bañol, luis Hernando Meneses176bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
73-1
83
Panorámica del Área Natural Única los Estoraques.figura 2.
MetodoloGía
Los ejemplares recolectados de esta nueva especie se obtuvieron bajo el proyecto “Caracterización de la entomofauna del Área Natural Única Los Estoraques (ANULE)”, mediante el uso de red entomológica y trampas Van Someren-Rydon con fruta en fermentación.
Las fotografías fueron tomadas con cámara digital Panasonic Lumix DMC-FH1 y la ilustración de la venación fue elaborada empleando cámara lucida convencional a 9x.
Los ejemplares empleados para este estudio fueron depositados en la colección del área natural (CANULE) y duplicados (paratipos) fueron depositados en el Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN-MHN-L) y la colección personal Efraín Henao (CEH-085).
Los órganos genitales fueron preparados con métodos estandarizados y de ilustración. El tamaño de la escala es de 1 mm o 1 cm según sea el caso. Abreviaturas empleadas en algunos casos: AD: Ala dorsal; AV: Ala ventral; AAD: Ala anterior dorsal; APD: Ala posterior dorsal; AAV: Ala anterior ventral; APV: Ala posterior ventral. D: Dorsal. V: Ventral. El Tamaño se da como la medida del ala anterior en mm. Gen: Órganos genitales; N: Número de ejemplares examinados.

Una nueva especie de Pharneuptychia Forster, 1964 del área natural única Los Estoraques –Norte de Santander... 177
resultados
Pharneuptychia Forster, 1964
Pharneuptychia estoraquensis Henao & Meneses n. sp. Figuras 3-5.
diagnosis
Macho. Difiere de los machos de sus congéneres P. pharnabazos (Bryk, 1953) y P. boliviana (Hayward, 1957) por la presencia de seis ocelos en la APV (en ejemplares muy longevos o desgastados se puede difuminar o perder el primero y último ocelo), generalmente los primeros cinco ocelos con dos pequeñas pupilas de color gris claro brillante (plateadas), el último reducido a una pequeña mancha casi imperceptible de pequeñas escamas de color amarillo, en algunos ejemplares completamente desvanecido o ausente.
descripción. Macho. Tamaño de 30-32 mm. Figuras 4 (A-B).
Cabeza. Marrón oscura con escamas crema por debajo de los ojos. Ojos labros de color marrón oscuro brillante. Palpos prominentes y largos, de color marrón oscuro, densamente pilosos ventralmente con largos pelos negros y crema; predominan los pelos crema. Antenas de color marrón en la parte dorsal y pequeñas manchas laterales de escamas de color crema en cada segmento en la región ventral y cerca de ¾ de la longitud.
Tórax. Negro brillante y densamente peludo de color gris. Patas anteriores cubiertas con pelos grisáceos; las medias y posteriores de color marrón claro y jaspeado. Abdomen. Región dorsal negra y con pelos de color gris, ventralmente peludo de color crema.
Superficie dorsal. AA y AP de color marrón oscuro a través de toda la superficie alar. AA hasta con cuatro ocelos, poco notorios o imperceptibles de color amarillo, el segundo ocelo más grande siempre visible, rodeado de un círculo delgado amarillo con fondo negro y una pupila plateada. En el AA se nota una línea marrón oscura submarginal ondulada y línea marrón oscura marginal. La venación de las alas (Figura 3) presenta un particular engrosamiento corto en la región externa de la celda discal que se observa externamente como si fuera una deformación.

efraín Reinel Henao-Bañol, luis Hernando Meneses178bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
73-1
83
Ilustración de la venación de Pharneuptychia estoraquensis n. sp.figura 3.
Superficie ventral. Superficie del AA de color marrón claro con escamas amarillas, color grisáceo hacia ápice y borde de ala con pequeñas escamas de color gris brillante. En AP las dos líneas marrones oscuras mencionadas se repiten y las líneas comprendidas entre la región postbasal y submarginal y submarginal y marginal son delgadas y de color marrón oscuras (Figuras 4 A-B).
Hembra. Tamaño 33-34 mm. Figuras 4 (C-D), 6.
Similar al macho pero con más presencia de escamas grises en la región anal del APV y presencia de línea medial marrón oscura discreta o poco notoria. Ocelos, dibujos y líneas más intensos, con pupilas gris-azuladas brillantes.

Una nueva especie de Pharneuptychia Forster, 1964 del área natural única Los Estoraques –Norte de Santander... 179
Pharneuptychia estoraquensis n. sp. Macho (Holotipo). a: dorsal. B: ventral. Hembra (Alotipo). C: dorsal. d: ventral.
figura 4.
Órganos genitales
Macho (Figura 5). Vista lateral del tegumen en forma cóncava; uncus largo, recto con terminación en forma de punta; gnathos largo, delgado y ligeramente curvo en dirección hacia arriba. Apéndices angulares anchos y cortos en vista lateral, saccus largo y más grueso en la base. Valva amplia y ancha con porción distal redondeada y corta; aedeago largo y delgado con una región membranosa desde el extremo hacia la región media con porción anterior ancha que se abre antero-dorsalmente hacia la región media.

efraín Reinel Henao-Bañol, luis Hernando Meneses180bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
73-1
83
Pharneuptychia estoraquensis n. sp. a: cápsula genital vista lateral. Aedeago. B: vista lateral. C: vista dorsal.
figura 5.
Hembra (Figura 6). Papilas pequeñas, ductus burse largo y delgado, corpus burse globoso con dos hileras de signos delgados que llegan a la mitad del corpus Espiráculo en la región dorso-apical. Lamella antevaginal presente con la expansión lateral no fusionada con el tergo VIII.
Pharneuptychia estoraquensis n. sp. Hembra (Alotipo). a: vista lateral. B: vista ventral.
figura 6.

Una nueva especie de Pharneuptychia Forster, 1964 del área natural única Los Estoraques –Norte de Santander... 181
Material estudiado
Material estudiado: HolotiPo. Macho con la siguiente etiqueta: / HOLOTIPO / COLOMBIA, Norte de Santander, La playa de Belén, ANU Los Estoraques, vereda la Honda, Filo los Espinos, Quebrada Platanillo,08°14’10,5”N73°14’54,8”W 1570m, 16-Dic-2016, E. Henao. V. García & R. Peñaranda Legs. (CANULE)/Holotipo Pharneuptychia estoraquensis n. sp. Henao & Meneses. det. 2017. alotiPo: COLOMBIA, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, 08°13’10,3”N 73°14’32,4 W 1520 m, 20-oct-2015, E. Henao & Funcionarios Legs. (CANULE)/Gen. EH-003. ParatiPos: 2 machos, Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, 08°13’48,5”N 73°14’03.3”W 1500 m, 21-oct-2015, E. Henao & Funcionarios Legs., Colombia, Norte de San-tander, La playa de Belén, ANU Los Estoraques, vereda la Honda, Filo los Espinos, Quebrada Platanillo,08°14’54,8”N73°14’54,8”W 1570m, 23-ago-2016, E. Henao & Funcionarios (MHN-ICN-L); 1 machos, Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, Finca San Antonio-Los Espinos, 08°14’10,6”N73°14’25,2”W 1500 m, 20-oct-2015, E. Henao & Funcionarios Legs., 1 hembra, Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero sitio Myrtaceas 08°13’48,5”N73°14’03,3”W 1500 m 3-Nov-2016 (CEH-085); 1 macho, COLOM-BIA, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero sitio Myrtaceas, 08°13’48,5”N73°14’03,3”W 1500 m 23-ago-2016, E. Henao & Funcionarios Legs., (CANULE); 1 macho, Colombia, Norte de Santander, La Playa de Belén, ANU Los Estoraques, Quebrada la Honda, camino bocatoma, 08°14’29,3”N 73°15’40,0”W 1700m 22-oct.2016, E. Heno & Funcionarios Legs., (CANULE); 1 macho, Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, 08°13’10,3”N 73°14’32,4 W 1520 m, 20-oct-2016, E. Henao & Funcionarios Legs., (CANULE); 1 macho Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, 08°14’10,6”N 73°14’05,4” W 1500 m, 20-oct-2016, E. Henao & Funcionarios Legs., (CANULE); 1 macho Colombia, Norte de Santander, La playa de Belén, ANU Los Estoraques, Vereda Rosa Blanca, Sector Cenicero, 08°13’48,5”N 73°14’03,3”W 1700 m, 03-nov-2016, E. Henao & Funcionarios Legs., 1 macho, mismos datos, fecha y colectores (CANULE).
localidad tipo: COLOMBIA, Norte de Santander, La Playa de Belén, ANU Los Estoraques, Vereda La Honda, Filo los Espinos, Quebrada Platanillo, 08°14’10,5”N 73°14’54,8”W 1570 m.
etimología. Esta especie es nombrada estoraquensis, referente al sitio de recolección de los ejemplares en el Área Natural Única Los Estoraques, la cual está protegida

efraín Reinel Henao-Bañol, luis Hernando Meneses182bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
73-1
83
mediante el Sistema de Parques Naturales de Colombia y es uno de los lugares más bellos por su conformación geomorfológica y paisajística.
disCusiÓn
Debido a que el género Pharneuptychia (Foster, 1964) es un pequeño grupo de mariposas que cuenta con seis especies descritas (WARREN et al., 2016) y cuya distribución ha estado enfocada a la parte sur de América especialmente en Brasil y países cercanos, no es difícil detectar una nueva especie con base en los patrones biogeográficos. La única especie con una distribución similar a la nueva especie corresponde a Pharneuptychia innocentia (C. Felder & R. Felder, 1867), pero los patrones de coloración, maculación y órganos genitales indican que no son especies cercanas e incluso podrían pertenecer a géneros diferentes.
La especie Pharneuptychia boliviana (Hayward, 1957), especie similar a la especie propuesta, presenta grandes diferencias en los patrones de maculación y en los órganos genitales, especialmente en la forma de la valva y el aedeago como también acontece con Pharneuptychia pharnabazos (Bryk, 1953) que adicionalmente se diferencia de la nueva especie por presentar ocelos de mayor tamaño y la presencia de cuatro ocelos en el APV.
observaciones de comportamiento y hábitat
Esta especie fue observada en zonas abiertas de bosque seco en cercanías de pastizales-pajonales nativos especialmente entre las 9:00 a 15:00 horas, con vuelo suave, rasante o bajo, asociada a satyrinos de los géneros Hermeuptychia Forster, 1964, Pareuptychia Forster, 1964, Cissia Doubleday, 1848 e Yphthimoides Forster, 1964.
Aunque esta especie sólo ha sido registrada en el área de estudio, existe la probabili-dad de ocurrencia en otras zonas, debido a las características del área, incluso podría existir la posibilidad de extenderse a las zonas de frontera con nuestro país hermano Venezuela, en áreas de bosque seco tropical.
aGradeCiMientos
ERHB agradece al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, especialmente al profesor Gonzalo Andrade-C., por apoyar las actividades de investigación y recibir algunos paratipos. A los funcionarios del área, Víctor García, Ronal Peñaranda, Gustavo Gómez y Elibardo Bayona, quienes apoyaron las actividades de campo, su disposición son ejemplo a seguir. A Julián Adolfo Salazar por el apoyo incondicional y comentarios al manuscrito. Agradecimientos especiales a los doctores Thamara Zacca y André Victor Lucci Freitas, quienes confirmaron la

Una nueva especie de Pharneuptychia Forster, 1964 del área natural única Los Estoraques –Norte de Santander... 183
nueva identidad. A los evaluadores que con sus sugerencias y comentarios ayudaron a mejorar este trabajo.
referenCias
COCK, M. J., 2014.- An updated and annotated checklist of the larger butterflies (Papilionoidea) of Trinidad, West Indies: Papilionidae, Pieridae and Nymphalidae. A Journal of World Insect Systematics. Insecta Mundi. 0353: 1–14
FORSTER, W., 1964.- Beitrage zur kenntnis der insektenfauna Boliviens XIX Lepidoptera III. Satyridae. Veröffentlichungen der staatssammlung Manchen 8: 51-188. pls. 27-35.
LAMAS, G., 2004.– Atlas of Neotropical Lepidoptera. Checklist: Part 4A. Hesperioidea - Papilionoidea: XXXVI +439 pp. Association for Tropical Lepidoptera, Gainesville.
LAMAS, G. & VILORIA A. L., 2004.- Tribe Satyrini, Subtribe Erebiina; Subtribe Hypocystina; Subtribe incertae sedis. In G. Lamas (ed.), Checklist: Part 4A. Hesperioidea-Papilionoidea. In J. B. Heppner (ed.), Atlas of Neotropical Lepidoptera. Vol. 5A:215-217, 224. Gainesville: Assoc. Trop. Lepid.
MARÍN, M. A.; URIBE, S. I., 2009.- Actualización sobre Euptychiina (Lepidoptera: Satyrinae) representadas en la colección del Museo Entomológico Francisco Luis Gallego. Boletín del Museo Entomológico Francisco Luis Gallego 1 (2): 22-32.
MARÍN, M. A., PEÑA, C., URIBE, S. I. & FREITAS, A. V. L. 2017. Morphology agrees with molecular data: phylogenetic affinities of Euptychiina butterflies (Nymphalidae: Satyrinae). Syst Entomol, 42: 768–785.
NAKAHARA, S. VEGA., G. & WILLMOTT K. R., 2016.- Description of a new species of euptychia Hübner, 1818 (Lepidoptera: Nymphalidae: Satyrinae) from the western Andes. Zootaxa 3-4184 (2):358-366.
PEÑA, C. & LAMAS, G., 2005.- Revision of the butterfly genus Forsterinaria Gray, 1973 (Lepidoptera: Nymphalidae, Satyrinae). Revista Peruana de Biología 12 (1): 5–48.
PEÑA, C., WAHLBERG, N., WEINGARTNER, E., KODANDARAMAIAH, U., NYLIN, S., FREITAS, A. V. L., BROWER, A. V. Z., 2006.- Higher level phylogeny of satyrine butterflies (Lepidoptera: Nymphalidae) based on DNA sequence data. Molecular Phylogenetics and Evolution, 40(1): 29-49. 40: 29-49.
PLAN DE MANEJO. AREA NATURAL UNICA LOS ESTORAQUES (PMANULE), 2016. - Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales. Territorial Andes Nororientales. La Playa de Belén, Norte de Santander, 224 p.
SANCHEZ, A. P & GANTIVA C.H. Entomofauna. Hormigas, mariposas y escarabajos. Pag. 89-120 En: Báez, L. y F. Trujillo (Eds.)., 2014.- Biodiversidad en Cerrejón. Carbones de Cerrejón, Fundación Omacha, Fondo para la Acción Ambiental y la Niñez. Bogotá, Colombia. 352 p.
WARREN A. D, DAVIS K. J, STANGELAND E. M, PELHAM J. P, GRISHIN N. V., 2016.- Illustrated Lists of American Butterflies. http://www.butterfliesofamerica.com [accessed Sept 2, 2017]

Boletín CientífiCoCentro de Museos
Museo de Historia natural
nueVo reGistro de Coryphorus (inseCta: ePHeMeroPtera) Y notas adiCionales soBre su
MorfoloGía*
Tatiana Cárdenas lópez1, Ximena ospina londoño2, Miguel Rodríguez Zambrano2, María Paula Rozo3,
Rodrigo Ivan Romero Zúñiga4, lucimar Gomes Dias5
resumen
Se presenta la distribución actual de la familia Coryphoridae con dos nuevos registros del género Coryphorus para el departamento del Putumayo, Colombia. Adicionalmente, se muestran detalles de las espinas y setas de las ninfas mediante microscopía electrónica (SEM), y se describen variaciones morfológicas de la genitalia del adulto.
Palabras clave: Coryphoridae, C. aquilus, cuenca Amazónica, efímera, insecto acuático.
neW reCord of CorYPHoridae (inseCta: ePHeMeroPtera) and additional notes on its
MorPHoloGY
abstract
The current distribution of the Coryphoridae family and two new records of the Coryphorus genus from the Department of Putumayo in Colombia are presented. Additionally, details of the spines and setae of the nymphs are shown by scanning electron microscopy (SEM) and morphological variations of the adult genitalia are described.
Key words: Coryphoridae, C. aquilus, Amazonian basin, ephemeral, aquatic insect.
* FR: 2-VII-2017. FA: .30-VIII-20171 B.Sc. Facultad de Ciencias Exactas y Naturales, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0003-4363-361X2 Programa de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected], ORCID: 0000-0003-1288-2867 [email protected] ORCID: 0000-0002-5857-99973 M.Sc. Universidade Federal do Rio de Janeiro. Rio de Janeiro, Brasil. E-mail: [email protected] ORCID: 0000-0003-2792-20914 M.Sc. Grupo de Investigación SENAGROQUIN-SENA, Quindío. Armenia, Colombia. E-mail: [email protected] ORCID: 0000-0002-4209-80215 D.Sc. Facultad de Ciencias Exactas y Naturales, Grupo de Investigación BIONAT: Biodiversidad y Recursos Naturales, Universidad de Caldas. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0001-6480-7688
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 184-189. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: CÁRDENAS-LÓPEZ, T., OSPINA, X., RODRÍGUEZ, M., ROZO, M.P., ROMERO, R.I. & DIAS, L.G, 2017.- Nuevo registro de Coryphorus (Insecta: Ephemeroptera) y notas adicionales sobre su morfología. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 184-189. DOI: 10.17151/bccm.2017.21.2.12

Nuevo registro de Coryphorus (Insecta: Ephemeroptera) y notas adicionales sobre su morfología 185
introduCCiÓn
La familia Coryphoridae es monotípica y está representada por el género Coryphorus con la especie, Coryphorus aquilus (PETERS, 1981; MOLINERI et al., 2001). La localidad tipo de la especie está ubicada en Brasil y Colombia, departamentos del Amazonas y Meta, respectivamente (PETERS, 1981). Actualmente, el género Coryphorus está registrado para diferentes localidades de Brasil (Amazonas, Mato Grosso y Amapá) (PETERS 1981; SHIMANO et al., 2011; BELMONT et al., 2015), Colombia (Amazonas y Meta) (PETERS, 1981; MOLINERI et al., 2001), Venezuela y Guyana Francesa (MOLINERI et al., 2011).
Tanto la biología como la taxonomía y distribución de esta monotípica familia son muy poco conocidas y se necesitan más estudios que proporcionen información sobre los vacíos en el conocimiento del grupo. El objetivo de este trabajo fue ampliar la distribución de la familia y realizar los nuevos registros de la especie para Colombia.
Materiales Y MÉtodos
Las ninfas se recolectaron mediante coladores manuales y redes surber en las quebradas Canalendre y Aguas Negras de los municipios de Mocoa y Puerto Asís, departamento del Putumayo, Colombia, respectivamente. En esta última localidad se colectaron adultos con trampas de luz, entre las 4:00 y 6:00 am. El material recolectado se almacenó en alcohol a 96% y se depositó en la Colección Entomológica del Programa de Biología de la Universidad de Caldas (CEBUC) y en la Colección Entomológica del Departamento de Zoología, Universidade Federal do Rio de Janeiro (DZRJ). Las fotografías fueron tomadas con un estereomicroscopio marca Carl Zeiss Stemi 508 con Axiocam ERc5. Las imágenes de barrido fueron tomadas con un Microscopio Electrónico de Barrido (SEM) marca FEI Quanta 250. La confirmación de la identificación de los individuos fue realizada mediante la revisión y comparación con la descripción original de la ninfa (PETERS, 1981) y del adulto (MOLINERI et al., 2001).
Material examinado
12 ninfas. Colombia, Putumayo, Puerto Asís, Quebrada Aguas Negras. N 0º31.60’19’’, O 76º31.63’76’’, altura: 250 m. 17/XII/2015. Colectores: Lucimar Gomes Dias, Ximena Ospina, Miguel Rodríguez, Rodrigo Romero-Z., María Paula Rozo y Tatiana Cárdenas-L. [CEBUC]. 15 adultos machos, igual al anterior [5 en CEBUC y 10 en DZRJ]. 1 ninfa, igual al anterior, excepto: Mocoa, Río Canalendre. N 1º7.1’98’’, O 76º37’43.04’’, altura: 543 m. 15/XII/2015 [CEBUC].

Tatiana Cárdenas lópez et al.186bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
84-1
89
resultados Y disCusiÓn
En total se recolectaron 13 ninfas y 15 adultos de Coryphorus. Una ninfa para Mocoa y 12 ninfas y 15 adultos para Puerto Asís. Las ninfas fueron encontradas en las zonas de las quebradas con baja velocidad del agua y presencia de barrancos con raíces asociadas. Los adultos fueron colectados solamente en horas de la madrugada (entre 4:00 y 6:00 am), y no en horas de la noche como lo reporta MOLINERI et al., (2001), quienes observaron dos horarios de vuelo para este género (entre 4:00 - 6:00am y 18:00 – 20:00pm).
Los caracteres observados para la identificación del género (PETERS, 1981; MOLINERI et al., 2001) fueron: en la ninfa: ojos pedunculados (Figs. 1A, 2A), presencia de tubérculos pares en la región dorsal de la cabeza (Figs. 2A, 3E) y simples en la región dorsal del tórax y abdomen (Figs. 2A-B), branquias operculares subovales con setas en los bordes (Fig. 2B), proyección de los tergos abdominales II-V curvada dorsalmente (Figs. 2A-B). En los adultos: región costal de las alas anteriores de coloración gris intenso y región cubital y anal de coloración hialina (Fig. 1B) y para la forma de la genitalia del macho, fueron encontradas algunas variaciones en la región apical del pene, diferente a lo descrito por MOLINERI et al., (2001). Adicionalmente, las imágenes mostradas con microscopía electrónica, proporcionan un mayor detalle de las pequeñas espinas distribuidas en todo el cuerpo (Fig. 2 y 3) y las setas del margen posterior de los tergos abdominales, las cuales son relativamente largas, gruesas y curvadas, características que no fueron descritas por PETERS (1981). Las variaciones encontradas, pueden indicar que se trata de una nueva especie de Coryphorus, sin embargo, análisis moleculares están siendo realizados para corroborar las variaciones morfológicas aquí reportadas.
Este nuevo registro para Colombia es importante, ya que el departamento del Putumayo ha sido poco estudiado y está localizado en una región biogeográfica muy diversa, ubicada al occidente de la Amazonía colombiana, convirtiéndose en una zona de interés científico para el estudio de insectos acuáticos.

Nuevo registro de Coryphorus (Insecta: Ephemeroptera) y notas adicionales sobre su morfología 187
a. Ninfa de Coryphorus: hábito dorsal. B. Adulto macho de Coryphorus: hábito lateral.
figura 1.
Ninfa de Coryphorus (SEM). a: hábito dorsal. B: abdomen en vista dorsal.
figura 2.

Tatiana Cárdenas lópez et al.188bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
84-1
89
Ninfa de Coryphorus (SEM). C: detalles del tórax. d: detalles de los tergos. e: detalle de la cabeza. f: uña anterior.
figura 3.
Distribución actual de Coryphorus Peters 1981 para Suramérica.figura 4.

Nuevo registro de Coryphorus (Insecta: Ephemeroptera) y notas adicionales sobre su morfología 189
aGradeCiMientos
A Gustavo Bolaños del Instituto de Investigaciones en Estratigrafía –IIES–, por las fotos de microscopía electrónica y al Laboratorio de Colecciones Biológicas de la Universidad de Caldas por la infraestructura. A Diego Martínez por la elaboración del mapa.
referenCias
BELMONT, E.L., CRUZ, P.V. & HAMADA, N., 2015.- A new species of Tricorythopsis Traver, 1958 (Leptohyphidae) and occurrence of Pannota (Insecta: Ephemeroptera) species in Amapá state, Brazil. Zootaxa, 4007 (1): 104-112.
MOLINERI, C., PETERS, J.C. & ZÚÑIGA, M.C., 2001.- A new family, Coryphoridae (Ephemeroptera: Ephernerelloidea), and description of the winged and egg stages of Coryphorus. Insecta Mundi, 15 (2): 1-7.
MOLINERI, C., GRILLET, M.E., NIETO, C., DOMINGUEZ, E. & GUERRERO, E., 2011.- New species and records for the mayfly families Caenidae, Leptohyphidae and Coryphoridae (Ephemeroptera, Pannota) from Venezuelan Guayana’s Uplands. Zootaxa, 2750: 39-50.
PETERS, W.L., 1981.- Coryphorus aquilus a new genus and species of Tricorythidae from the Amazon Basin (Ephemeroptera). Aquatic Insects, 3 (4): 209-217.
SHIMANO, Y., SALLES, F.F. & CABETTE-RAMOS, H.S., 2011.- Ephemeroptera (Insecta) ocorrentes no Leste do Estado do Mato Grosso, Brasil. Biota Neotrop, 11 (4): 239-254.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
distriBuCiÓn PotenCial de Oxysternon conspicillatum (WeBer, 1801) en diferentes esCenarios de CaMBio
CliMÁtiCo en ColoMBia*
Bedir Germán Martínez-Quintero1, Alejandro echeverri-Rubio2, Fabián Guillermo Gaviria-ortiz3
resumen
Los escenarios de cambio climático son herramientas útiles para la evaluación de impactos, iniciativas de adaptación y mitigación, y creación de modelos climáticos; estos se proyectan en el tiempo para predecir la unidad de hábitat en que se encuentran las especies. Adicional-mente, los modelos de distribución de especies estiman el área de ocupación de un organismo con potencial bioindicador como los escarabajos coprófagos, entre ellos la especie oxysternon conspicillatum con amplia distribución geográfica en Colombia. El propósito de este trabajo fue determinar la distribución potencial de o. conspicillatum en un escenario pesimista (A2) y uno optimista (B1) de cambio climático en Colombia para los años 2050 y 2080. Para el desarrollo del modelo se recolectó información publicada y/o depositada en diferentes colec-ciones del país, de manera tal que se obtuvieron 427 registros, distribuidos en 11 fuentes de información. La especie se registró en 18 departamentos, donde el Eje Cafetero fue el que presentó mayor disponibilidad de datos geográficos. Se encontró que la probabilidad de ocupación de o. conspicillatum en la Cordillera Oriental disminuye drásticamente para estos dos años. Este resultado concuerda con lo observado en otros grupos bioindicadores como las plantas y las mariposas. o. conspicillatum es una especie ampliamente distribuida en Colombia, sin embargo, para los años 2050 y 2080 bajo dos escenarios de cambio climático (A2 y B1) presentará una disminución considerable de su presencia a nivel nacional.
Palabras clave: área de ocupación, cambio climático, modelación, MaxEnt, bioindicador.
* FR: 14-IV-2017. FA: .3-IX-20171 Biólogo. Línea de investigación en Desarrollo Sostenible y Medio Ambiente, Universidad de Manizales. Manizales, Colombia. E-mail: [email protected] (autor de correspondencia). ORCID: 0000-0002-5798-73312 Doctorando, Universidad Nacional de Colombia. Profesor Instructor, Universidad de Manizales, Centro de Inves-tigación en Medio Ambiente y Desarrollo. Manizales, Colombia. E-mail: [email protected] Msc en Ciencias Biológicas (Entomología), Universidade Federal do Paraná. E-mail: [email protected]
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 190-206. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: MARTÍNEZ-QUINTERO, B.G., ECHEVERRI-RUBIO, A. & GAVIRIA-ORTIZ. F.G., 2017.- Distribución potencial de Oxysternon conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático en Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 190-206. DOI: 10.17151/bccm.2017.21.2.13

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 191
Potential distriBution of Oxysternon conspicillatum (WeBer, 1801) in different CliMate CHanGe sCenarios
in ColoMBia
abstract
Climate change scenarios are useful tools for impact assessment, adaptation and mitigation initiatives, and climate modeling wich are projected over time to predict the unit of habitat in which the species are. In addition, the species distribution models estimate the area of oc-cupation of an organism with bioindicator potential such as dung beetles, including the spe-cies oxysternon conspicillatum with a wide geographic distribution in Colombia. The purpose of this work was to determine the potential distribution of o. conspicillatum in a pessimist (A2) and an optimist (B1) climate change scenario in Colombia in the years 2050 and 2080. For the development of the model, published and/or deposited in different collections of the country samples was collected so that 427 records were obtained, distributed in 11 sources of information. The species was recorded in 18 departments from which the Coffee Triangle was the area that showed greater availability of geographic data. It was found that the probability of occupation of o. conspicillatum in the Eastern Cordillera will decrease drastically for these two years. This result coincides with what was observed in other bioindicator groups such as plants and butterflies. o. conspicillatum is a widely distributed species in Colombia. However, for the years 2050 and 2080 under the two climate change scenarios (A2 and B1) it wil show a considerable decrease in its presence at the national level
Key words: area of occupation, climate change, modeling, MaxEnt, bioindicator.
introduCCiÓn
Colombia es un país megadiverso, sin embargo, de acuerdo al índice de vulnerabilidad al cambio climático para la región de América Latina y el Caribe, se encuentra en una categoría de riesgo alto (CAF, 2014). El cambio climático, sus efectos sobre el ciclo del agua y la pérdida de hábitat están estrechamente ligados a la reducción de la biodiversidad; es altamente probable que la transformación en los regímenes de precipitación e incremento de la temperatura incidan directamente en el tamaño, fisiología, estructura, abundancia y distribución de una amplia variedad de organismos (BÖHNING et al., 2008). Estos factores suponen un cambio en las interacciones entre las especies y su distribución dentro de los ecosistemas, entre otros (IPCC, 2007). El resultado de la coacción de dichos factores tiene repercusiones en la calidad de los servicios ambientales suministrados por los ecosistemas (IPCC, 2007).
Actualmente, la implementación de modelos para predecir la respuesta de las especies, ecosistemas y paisajes, son una tendencia en estudios de ecología y conservación. Para establecer el efecto del cambio climático se han desarrollado herramientas

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz192bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
predictivas denominadas escenarios de cambio, basadas en las emisiones de gases de efecto invernadero a largo plazo, que son un conjunto de imágenes sobre lo que podría suceder en el futuro, útiles para evaluar la manera como podrían contribuir las fuerzas determinantes tanto sociales como naturales en las emisiones futuras (IPCC, 2000). La información que se produce es apropiada para la proyección de modelos del clima, la valoración de los impactos y para las estrategias de adaptación y mitigación (IPCC, 2000, 2002).
Por otra parte, la respuesta biótica al cambio climático ha sido estudiada en varios niveles de organización, desde fenología de organismos, hasta rangos de distribución de las poblaciones y ensamblajes de comunidades, considerando al cambio climático como aspecto importante en la extinción de especies (PARMESAN & YOHE, 2003; WILSON et al., 2007). Sin embargo, gran parte de estos estudios se restringen a países del hemisferio norte y por lo general son a largo plazo o monitoreos, por lo cual no existe un buen número de datos en el planeta que permitan dar una aproximación sobre las posibles respuestas de los organismos, asimismo son escasas las investigaciones basadas en la distribución actual de los mismos al cambio climático (WILSON et al., 2007). Por consiguiente, conocer la distribución de las especies y sus factores limitantes proporciona argumentos sólidos en la implementación de estrategias de conservación de la biodiversidad (BROWN, 1995) y más cuando se implementa en grupos con potencial bioindicador, es una herramienta fundamental en las propuestas de conservación y biodiversidad dentro de escenarios de cambio climático, apoyando la toma de decisiones sobre estrategias a seguir a largo plazo.
Dentro de estos modelos se encuentra el software gratuito MaxEnt (método de máxima entropía), empleado para realizar predicciones o inferencias a partir de información incompleta (VALDERRAMA, 2013). Este modelo de distribución de especies es una caracterización de las condiciones ambientales adecuadas para una especie, y permite identificar dónde están espacialmente ubicados los sitios aptos para el desarrollo de la especie investigada. Por lo tanto, este método puede ser utilizado en numerosas áreas dentro de diversas problemáticas; entre sus variadas aplicaciones se encuentran la restauración ecológica, la priorización de zonas para iniciativas de conservación biológica, la evaluación de patrones de propagación de especies invasivas y modelaciones de efectos del cambio climático sobre los ecosistemas (MORALES, 2012).
Aunque para muchos de los insectos estas herramientas son poco exploradas, los escarabajos coprófagos (Scarabaeinae) son un grupo de insectos con características ecológicas que los hacen importantes indicadores ecológicos de perturbación antrópica (HALFFTER & MATTHEWS, 1966; HANSKI & CAMBEFORT, 1991, VULINEC, 2000), entre ellas, son sensibles a la perturbación debido a que son afectados directamente por los cambios en las variables microclimáticas, temperatura,

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 193
humedad o características del suelo (NICHOLS et al., 2007), por lo tanto, los escarabajos coprófagos son un excelente grupo indicador de salud de los ecosistemas (VULINEC, 2000).
El papel de los coprófagos va más allá de la indicación; aportan en diferentes servicios ecosistémicos tales como ciclaje de nutrientes, aireación del suelo, dispersión secundaria de semillas, entre otros (NICHOLS et al., 2007), pero realmente es poco lo que se conoce acerca de la respuesta de los coprófagos a la heterogeneidad espacial de los paisajes y al cambio climático (SPECTOR, 2006; NICHOLS et al., 2007).
Después de evidenciar que el cambio climático tiene una afectación sobre la diversidad y que grupos bioindicadores como los escarabajos coprófagos son excelentes indicadores de la salud de un ecosistema, el objetivo de este trabajo fue determinar la distribución potencial de una especie de escarabajo coprófago (oxysternon conspicillatum) en diferentes escenarios de cambio climático en Colombia, en dos épocas diferentes en el futuro, en los años 2050 y 2080.
Materiales Y MÉtodos
El modelo de distribución potencial de o. conspicillatum se realizó con una buena cantidad de datos de registro de presencia en las diferentes regiones del país, asimismo se garantizó que la toma de datos proviene de fuentes de información confiables depositada en colecciones. Posteriormente, las variables analizadas fueron llevadas a una escala de observación de los datos de forma precisa y con una cobertura considerada pertinente por los autores del presente documento. Finalmente, para establecer la confiabilidad del modelo fue necesario tener conocimientos básicos de la biología, la ecología, la información bibliográfica de la especie y de la fauna asociada a la misma, que permitió comparar la realidad de los datos actuales con los posibles modelos.
Los criterios bajo los cuales se seleccionó o. conspicillatum (Figura 1), como especie para modelar su ocupación en el tiempo y evaluar su respuesta de distribución frente al cambio climático en Colombia, se dividen en tres aspectos: i) potencial indicador de perturbación, ii) amplia distribución geográfica y iii) taxonomía bien definida.
Para este análisis, se usaron todos los registros de o. conspicillatum, depositados en el portal de datos del Sistema de Información de Biodiversidad de Colombia (SIB) y en registros encontrados en trabajos publicados en el país. Se tabularon en Excel, fueron depurados y se utilizaron los que estaban correctamente georreferenciados, es decir los de alta confiabilidad, dada por aquellos registros donde las coordenadas de latitud y longitud eran veraces y se ubicaban en las zonas indicadas, esto se corroboró en el portal Google earth.

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz194bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
Especie de o. conspicillatum, hembra y macho. (Tomado de CULTID et al., 2012).
figura 1.
determinación de la distribución potencial actual
La distribución potencial de o. conspicillatum se realizó empleando MaxEnt (Maximum entropy Modelling of Species Geographic Distributions), el cual aplica el principio de la máxima entropía para calcular la distribución de especies más probable y más uniforme posible para un taxón (VALDERRAMA, 2013). El resultado es un valor de idoneidad de hábitat en función de las características ambientales de los puntos de presencia que se introduzcan en el modelo (PHILLIPS et al., 2006, PHILLIPS & DUDÍK, 2008). MaxEnt realiza la aproximación al nicho ecológico de los taxones estudiados, a partir del modelo climático global WorldClim, el cual se basa en 19 variables climáticas más altitud (Tabla 1), derivadas de la temperatura y la precipitación, para identificar los valores abióticos que limitan su distribución (AUSTIN, 2002) y además se le pueden incorporar variables de diferentes fuentes como cobertura vegetal, altitud, entre otras.

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 195
tabla 1. Variables extraídas de WorldClim
Variables climáticas
bio1 = Temperatura media anual bio11 = T° media del trimestre más calientebio2 = Rango T° media diurna (Tmax-Tmin) bio12 = Precipitación total anualbio3 = Isotermalidad (bio1/bio7) × 100 bio13 = Precipitación del mes más húmedobio4 = Estacionalidad de T° (DS × 100) bio14 = Precipitación del mes más secobio5 = T° máxima del mes más caliente bio15 = Estacionalidad de la precipitaciónbio6 = T° mínima del mes más frío bio16 = Precipitación trimestre más húmedobio7 = Rango de T° anual (bio5-bio6) bio17 = Precipitación del trimestre más secobio8 = T° media del trimestre más húmedo bio18 = Precipitación del trimestre más calientebio9 = T° media del trimestre más seco bio19 = Precipitación del trimestre más caliente bio10 = T° media del trimestre más frío Alt = Altitud
Modelación de la distribución futura de O. conspicillatum bajo un escenario de cambio climático
Para los modelos predictivos se utilizaron los datos de BioClim con una resolución 2,5 minutos, implementados con el modelo MRI-CGCM3, el cual reproduce el clima medio global, incluyendo la variación estacional en diversos aspectos de la atmósfera y los océanos; considerando el clima simulado evaluando el fenómeno de El Niño y la Oscilación del Sur y el Ártico, y del Antártico (YUKIMOTO et al., 2012). Los escenarios utilizados fueron A2 y B1 (Tabla 2), extremos para el cambio climático global, siendo el A2 el pesimista, donde el planeta está fragmentado y no sustentable, con concentraciones de gases de efecto invernadero mayores. El escenario B1 es un escenario optimista donde el planeta es integrado y sustentable con mayor uso de energías renovables y menor concentración de gases de efecto invernadero (RASKIN et al., 2005).
Se realizaron mapas de distribución potencial actual (2016) y futura (2050 y 2080) en los diferentes escenarios, para conocer los cambios en las áreas potenciales de distribución de o. conspicillatum bajo cada modelo. Estos fueron utilizados para análisis descriptivo y en el programa Qgis 2.2. se establecieron diferencias en el tamaño de distribución y las regiones más afectadas. Finalmente, para determinar la pérdida o ganancia de área entre años y escenarios, se tomó como referencia por decisión de los autores el 80% de la probabilidad de ocurrencia de o. conspicillatum en el país (mayor idoneidad de hábitat), donde todos los valores iguales o por encima de este límite fueron los utilizados para calcular el área.
Los datos de o. conspicillatum se encontraron registrados en 11 fuentes de información, seis artículos científicos (MEDINA & KATTAN, 1996; HUERTAS et al., 2003; EDMONDS & ZÍDEK, 2004; GARCÍA & PARDO, 2004; ESPARZA & AMAT,

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz196bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
2007; CONCHA-LOZADA et al., 2010), cuatro colecciones de proyectos que aún no están publicadas (CR-EC 2004; CR-S 2007; CRCUT 1 2008; ESCOBAR-IAvH-Cham) y en el portal del Sistema de Información Biológica (SIB).
tabla 2. Descripción de los escenarios de cambio climático. (Tomado de IPCC, 2000).
escenario Características distintivas
A2: Describe un mundo heterogéneo
la autosuficiencia y la conservación de las identidades locales. las pautas de fertilidad en el conjunto de las regiones convergen muy lentamente, con lo que se obtiene una población mundial en continuo crecimiento. el desarrollo económico está orientado básicamente a las regiones, y el crecimiento económico por habitante así como el cambio tecnológico están más fragmentados y son más lentos que en otras líneas evolutivas.
B1: Describe un mundo convergente
Tiene una misma población mundial que alcanza un máximo hacia mediados del siglo y desciende posterior-mente, como en la línea evolutiva A1, pero con rápidos cambios de las estructuras económicas orientados a una economía de servicios y de información, acompañados de una utilización menos intensiva de los materiales y de la introducción de tecnologías limpias con un aprovechami-ento eficaz de los recursos. en ella se da preponderancia a las soluciones de orden mundial encaminadas a la sostenibilidad económica, social y medioambiental, así como a una mayor igualdad, pero en ausencia de inicia-tivas adicionales en relación con el clima.
resultados Y disCusiÓn
Se obtuvo un total de 427 registros de o. conspicillatum con datos de georreferenciación correctos para Colombia, distribuidos en 18 departamentos del país. Amazonas es el departamento con mayor número de registros con un total de 140, sin embargo, la región del Eje Cafetero es la zona donde se presenta mayor número de registros en distintas localidades, estos valores soportan la amplia distribución geográfica de la especie en el país. En términos generales, esta especie es ubicua en Colombia, tiene alta presencia en las tres cordilleras andinas y en la Amazonía, de tal manera que se encuentra en un rango entre los 0 m a los 3000 m de altitud como lo registra

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 197
EDMONDS & ZÍDEK (2004), por lo que puede estar afectada por los factores de cambio climático que se presenten en el territorio nacional. o. conspicillatum presenta la particularidad de estar presente en zonas abiertas (fuera de los bosques) en la región andina y restringida a zonas boscosas en la región de la Amazonía, esto hace que sea una especie de distribución geográfica y requerimientos ecológicos amplios (especie generalista).
La distribución potencial actual de o. conspicillatum presenta un modelo bien ajustado y confiable, con un AUC de 0,985. Las variables ambientales que influyen directamente sobre el modelo de distribución actual son la isotérmica (bio 3) con un 30,4% y la estacionalidad de la temperatura (bio 4) con un 20,9%, ambas variables derivan de la temperatura de las zonas en que se ha registrado esta especie. Además, los datos ecológicos obtenidos en otros proyectos de algunos individuos en campo sustentan la idea de que la temperatura es la variable que presenta mayor peso en la actividad diaria de las poblaciones de o. conspicillatum, lo que posiblemente pueda generar una presión en la distribución de esta especie en el tiempo.
De acuerdo a las distribuciones potenciales obtenidas, o. conspicillatum posee zonas climáticamente favorables en gran parte del territorio colombiano, asociada con altitudes entre los 1000 m y 2500 m. El número de celdas de estas distribuciones potenciales con respecto al número de localidades donde existen presencias conocidas, pueden ser superiores a las 100 veces. Asimismo, en otros grupos de insectos, en países donde la tradición taxonómica es menor y donde hay una menor diversidad entomológica (ROCCHINI et al., 2011), es muy probable que los registros de o. conspicillatum presenten evidentes sesgos ambientales y espaciales, de modo que la distribución “real” de esta especie sea mucho más amplia que la conocida. Las zonas potenciales detectadas con el modelo deben considerarse como territorios de interés, no obstante, es importante tener en cuenta que posiblemente existan limitantes de dispersión, distribución y otros factores que tienen la capacidad de limitar las localidades que supuestamente son favorables climáticamente (WILLIAMS et al., 2009).
Las mediciones potenciales donde podría registrarse o. conspicillatum, tanto en la actualidad como en un futuro, indican que actualmente esta especie ocupa mayor área en las tres cordilleras con respecto al resto del país (Figura 2). Mientras que para un escenario de cambio climático A2 y B1, para los años 2050 y 2080, se encontró que en las cordilleras Oriental y Central disminuye representativamente con respecto a la Cordillera Occidental en la que aumenta (Figuras 3 y 4).
Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz198bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
Distribución geográfica potencial de o. conspicillatum en la actualidad.
figura 2.
Distribución potencial de o. conspicillatum bajo un escenario A2 (izquierda) y B1 (derecha) en el año 2050.
figura 3.

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 199
Estos datos concuerdan con lo reportado por otros autores en diferentes grupos bioindicadores; se ha encontrado que varias especies de plantas disminuyen su área de ocupación bajo el mismo escenario de cambio climático, siendo la Cordillera Oriental la más afectada en cuanto a riqueza de especies y reducción del área de ocupación (TRIVIÑO, datos no publicados). Además, TRIVIÑO y colaboradores, en un trabajo no publicado, encontraron que la población de mariposas se ve mayormente afectada en la Cordillera Oriental bajo un escenario A2 en el año 2050, por consiguiente este grupo se va a ver limitado en esta zona del país.
Esta simulación permite establecer que el cambio climático tendrá una alta influencia en la distribución de o. conspicillatum en un futuro, debido a que esta especie va a estar ausente en gran parte del territorio nacional, afectando posiblemente el equilibrio ecológico por la pérdida del servicio ecosistémico prestado por la misma. Por otra parte, se proyecta que para el 2080 habrá un pequeño incremento en la probabilidad de ocurrencia de esta especie en el Caribe colombiano. No obstante, la presencia en esa zona puede estar limitada por factores de competencia de la especie Digitonthophagus gazella, especie introducida que habita en la región del Caribe (NORIEGA et al., 2012).
Distribución potencial de o. conspicillatum bajo un escenario A2 (izquierda) y B1 (derecha) en el año 2080.
figura 4.

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz200bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
Es alarmante el efecto de la ausencia de o. conspicillatum como especie generalista en cuanto a la pérdida de servicios ecosistémicos, debido a sus características ecológicas, puede suponerse resiliente a cualquier perturbación ambiental, lo que hace necesario investigaciones enfocadas a encontrar cuál es la tendencia de diversos organismos en respuesta al cambio climático. Estos resultados conllevan a proponer como estrategias de conservación a corto, mediano y largo plazo, el monitoreo de paisajes inmersos en la Cordillera Oriental y generar planes de manejo que permitan mitigar los efectos del cambio climático en esta cordillera.
Adicionalmente, determinar la distribución potencial de una especie indicadora como o. conspicillatum permitirá complementar los datos registrados en términos de variables ambientales a futuro y suministrar herramientas para determinar zonas de protección y manejo especial para afrontar los cambios en el clima por lo menos en un periodo de 60 años. Esta herramienta de predicción ayuda a generar nichos ecológicos fundamentales, los cuales pueden ser complementados con otros aspectos ecológicos y/o antrópicos como: relación con otras especies (mutualismo), modelación de cambios de usos del suelo, lo que podría modificar los arreglos de los elementos del paisaje donde se encuentra o. conspicillatum y así alterar posibles rutas de desplazamiento para responder a los cambios futuros.
En cuanto al área afectada en términos de ocurrencia de la especie en estudio con una probabilidad superior al 80% de registro en un lugar, en específico en Colombia, el área que cubriría sería cerca de 93.500 km2 que corresponden al 8,2% del territorio nacional (Tabla 3, Figura 5). Pese a que este porcentaje es bajo, para una especie con restricciones fisiológicas de temperatura, humedad, altitud entre otras, la idoneidad del hábitat donde se puede registrar es alta, restringiéndose principalmente a las cordilleras y en pequeñas proporciones del Amazonas. Además, a nivel nacional el porcentaje de desaparición de la especie en el corte del 80% no pasa el 1%, lo que posiblemente explique la alta presencia de esta especie en gran parte del territorio nacional.
Áreas de probabilidad superior al 80% de ocurrencia de o. conspicillatum en Colombia. Porcentajes del área de idoneidad de hábitat vs. área total de Colombia y área total de probabilidad actual. Todos los valores bajo los dos escenarios de cambio climático en los años 2050 y 2080.
tabla 3.
Escenarios - año Área Hábitat Idóneo(km2)
Presencia vs. Total(%)
Presencia vs. Actual(%)
Actual 93470,35 8,2A2 2050 81161,85 7,1 86,8A2 2080 78836,31 6,9 84,3B1 2050 88450,01 7,8 94,6

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 201
Distribución geográfica potencial de o. conspicillatum en la actualidad con una probabilidad de ocurrencia superior al 80%.
figura 5.
Al encontrar que los hábitats idóneos para la ocurrencia de o. conspicillatum son las tres cordilleras, se revisó puntualmente el área de pérdida o ganancia en cada cordillera y así se definió si hay o no ganancias en los diferentes escenarios durante el cambio climático en los periodos de 2050 y 2080. Se encontró que la Cordillera Oriental será la que presente mayor afectación sobre los hábitats donde hay mayor probabilidad de ocurrencia de este escarabajo, asimismo en un escenario A2 en 2050 disminuirá el área en 84% aproximadamente y en 2080 para este mismo escenario será una disminución del 52% (Tabla 4). Para el escenario B1 en esta misma cordillera, en 2050 se verá una pérdida de área en aproximadamente 60%, pero para el año 2080 esta será casi igual a la del escenario A2, con cerca del 52% del área perdida para este año (Figuras 6 y 7).
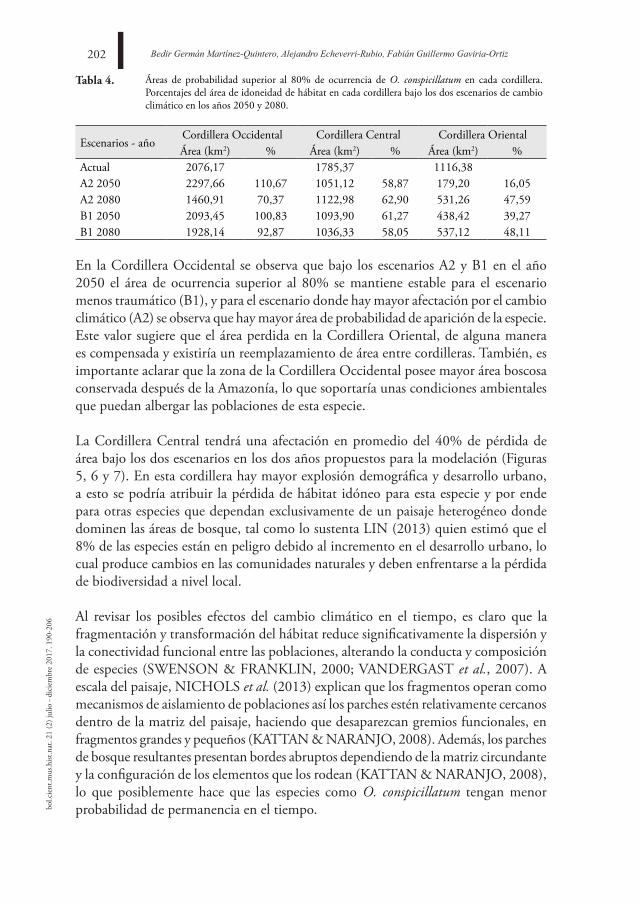
Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz202bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
Áreas de probabilidad superior al 80% de ocurrencia de o. conspicillatum en cada cordillera. Porcentajes del área de idoneidad de hábitat en cada cordillera bajo los dos escenarios de cambio climático en los años 2050 y 2080.
tabla 4.
Escenarios - añoCordillera Occidental Cordillera Central Cordillera OrientalÁrea (km2) % Área (km2) % Área (km2) %
Actual 2076,17 1785,37 1116,38A2 2050 2297,66 110,67 1051,12 58,87 179,20 16,05A2 2080 1460,91 70,37 1122,98 62,90 531,26 47,59B1 2050 2093,45 100,83 1093,90 61,27 438,42 39,27B1 2080 1928,14 92,87 1036,33 58,05 537,12 48,11
En la Cordillera Occidental se observa que bajo los escenarios A2 y B1 en el año 2050 el área de ocurrencia superior al 80% se mantiene estable para el escenario menos traumático (B1), y para el escenario donde hay mayor afectación por el cambio climático (A2) se observa que hay mayor área de probabilidad de aparición de la especie. Este valor sugiere que el área perdida en la Cordillera Oriental, de alguna manera es compensada y existiría un reemplazamiento de área entre cordilleras. También, es importante aclarar que la zona de la Cordillera Occidental posee mayor área boscosa conservada después de la Amazonía, lo que soportaría unas condiciones ambientales que puedan albergar las poblaciones de esta especie.
La Cordillera Central tendrá una afectación en promedio del 40% de pérdida de área bajo los dos escenarios en los dos años propuestos para la modelación (Figuras 5, 6 y 7). En esta cordillera hay mayor explosión demográfica y desarrollo urbano, a esto se podría atribuir la pérdida de hábitat idóneo para esta especie y por ende para otras especies que dependan exclusivamente de un paisaje heterogéneo donde dominen las áreas de bosque, tal como lo sustenta LIN (2013) quien estimó que el 8% de las especies están en peligro debido al incremento en el desarrollo urbano, lo cual produce cambios en las comunidades naturales y deben enfrentarse a la pérdida de biodiversidad a nivel local.
Al revisar los posibles efectos del cambio climático en el tiempo, es claro que la fragmentación y transformación del hábitat reduce significativamente la dispersión y la conectividad funcional entre las poblaciones, alterando la conducta y composición de especies (SWENSON & FRANKLIN, 2000; VANDERGAST et al., 2007). A escala del paisaje, NICHOLS et al. (2013) explican que los fragmentos operan como mecanismos de aislamiento de poblaciones así los parches estén relativamente cercanos dentro de la matriz del paisaje, haciendo que desaparezcan gremios funcionales, en fragmentos grandes y pequeños (KATTAN & NARANJO, 2008). Además, los parches de bosque resultantes presentan bordes abruptos dependiendo de la matriz circundante y la configuración de los elementos que los rodean (KATTAN & NARANJO, 2008), lo que posiblemente hace que las especies como o. conspicillatum tengan menor probabilidad de permanencia en el tiempo.
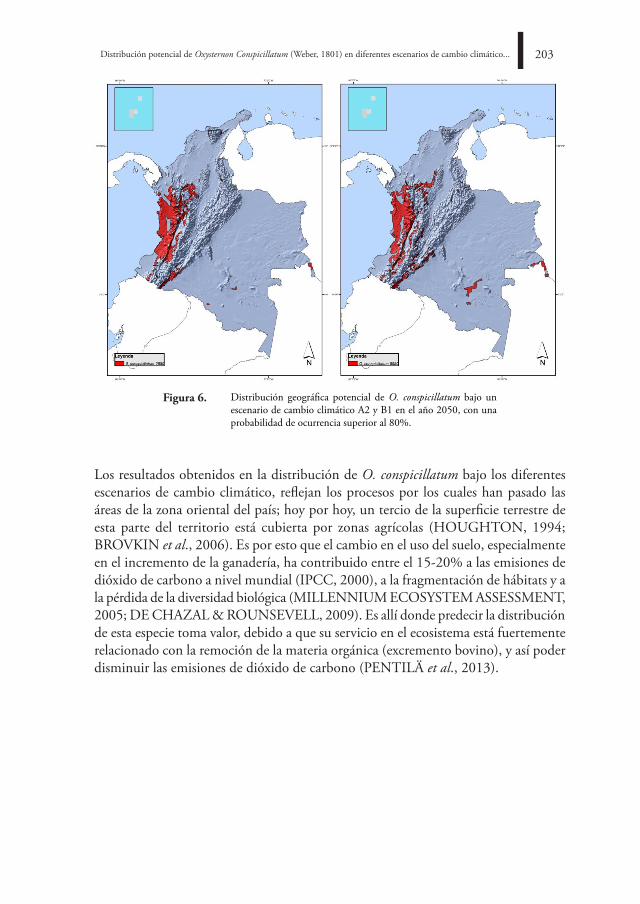
Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 203
Distribución geográfica potencial de o. conspicillatum bajo un escenario de cambio climático A2 y B1 en el año 2050, con una probabilidad de ocurrencia superior al 80%.
figura 6.
Los resultados obtenidos en la distribución de o. conspicillatum bajo los diferentes escenarios de cambio climático, reflejan los procesos por los cuales han pasado las áreas de la zona oriental del país; hoy por hoy, un tercio de la superficie terrestre de esta parte del territorio está cubierta por zonas agrícolas (HOUGHTON, 1994; BROVKIN et al., 2006). Es por esto que el cambio en el uso del suelo, especialmente en el incremento de la ganadería, ha contribuido entre el 15-20% a las emisiones de dióxido de carbono a nivel mundial (IPCC, 2000), a la fragmentación de hábitats y a la pérdida de la diversidad biológica (MILLENNIUM ECOSYSTEM ASSESSMENT, 2005; DE CHAZAL & ROUNSEVELL, 2009). Es allí donde predecir la distribución de esta especie toma valor, debido a que su servicio en el ecosistema está fuertemente relacionado con la remoción de la materia orgánica (excremento bovino), y así poder disminuir las emisiones de dióxido de carbono (PENTILÄ et al., 2013).

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz204bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
Distribución geográfica potencial de o. conspicillatum bajo un escenario de cambio climático A2 y B1 en el año 2080, con una probabilidad de ocurrencia superior al 80%.
figura 7.
A partir de estos resultados se pueden implementar planes de conservación de ecosistemas como el monitoreo de especies (caso o. conspicillatum), y demuestra las ventajas e importancia que tienen registros geográficos de las especies e incentiva su uso. Adicionalmente, se pueden lanzar hipótesis espaciales en diferentes escenarios a escala temporal y espacial, no obstante es importante mencionar que estos modelos tienen limitaciones que no permiten incorporar variables de interacciones bióticas, procesos históricos, historia natural, capacidad de dispersión y barreras biogeográficas de las especies.
ConClusiones
Actualmente o. conspicillatum es una especie ampliamente distribuida en Colombia, sin embargo, para los años 2050 y 2080 bajo dos escenarios de cambio climático (A2 y B1) presentará una disminución considerable de su presencia a nivel nacional. Bajo cualquiera de los escenarios planteados (A2 y B1) en los dos años modelados (2050 y 2080), a nivel nacional existirán afectaciones en el área de uso de esta especie. No obstante, a escala de las cordilleras, la que más afectación tendrá por el cambio climático será la Cordillera Oriental que disminuirá en más del 80% su idoneidad de hábitat para o. conspicillatum.

Distribución potencial de oxysternon Conspicillatum (Weber, 1801) en diferentes escenarios de cambio climático... 205
Es inevitable que ocurra el nuevo cambio climático como consecuencia de diferentes factores, entre ellos las actividades antrópicas y que a su vez están afectando inminentemente la biodiversidad, exponiendo la reducción de las poblaciones de especies importantes y sensibles (o. conspicillatum) que proporcionan servicios ambientales a la sociedad. Además, las herramientas predictivas como MaxEnt no incorporan todos los factores que describan el establecimiento y distribución de las especies, sin embargo, son de gran aplicabilidad para los planes de conservación a futuro.
aGradeCiMientos
Los autores agradecen a WCS/Programa Colombia por permitir el acceso a la información de o. conspicillatum. Especial agradecimiento al doctor Carlos A. Cultid-Medina por suministrar información de esta especie y a la bióloga Patricia E. Jiménez-Pérez por su ayuda en la edición del documento.
referenCias
AUSTIN, M.P., 2002.- Spatial prediction of species distribution: an interface between ecological theory and statistical modelling. ecol. Model., 157: 101-118.
BÖHNING, G.K., JETZ, W. & SCHAEFER, H.C., 2008.- Impact of climate change on migratory birds: community reassembly versus. Glob. ecol. Biogeogr., 17 (1): 38-49.
BROVKIN, V., CLAUSSEN, M., DRIESSCHAERT, E., FICHEFET, T., KICKLIGHTER, D., LOUTRE, M.F & SOKOLOV, A., 2006.- Biogeophysical effects of historical land cover changes simulated by six Earth system models of intermediate complexity. Climate Dynamics, 26 (6): 587-600.
BROWN, J.H., 1995.- Macroecology. University of Chicago Press, Chicago.CAF. 2014. Índice de vulnerabilidad y adaptación al cambio climático en la región de América Latina y el Caribe. Caracas: CAF.
Retrieved from http://scioteca.caf.com/handle/123456789/517 CONCHA-LOZADA, C.M, GALLEGO, M.C. & PARDO-LOCARNO, L.C., 2010.- Fragmentación de ecosistemas montanos e
impactos estructurales y poblacionales sobre la comunidad de escarabajos coprófagos (Col. Scarabaeinae) en el alto río Cauca, Popayán, Colombia. Bol. Cient. Mus. Hist. Nat., 14: 43-55.
CULTID, A.C., MEDINA, C.A., MARTÍNEZ, B.G., ESCOBAR, A.F., CONSTANTINO, L.M. & BETANCUR, N., 2012.- escarabajos Coprófagos (Scarabaeinae) del eje Cafetero: Guía para el estudio ecológico. WCS Cali, Colombia.
DE CHAZAL, J. & ROUNSEVELL, M., 2009.- Land-use and climate change within assessments of biodiversity change: A review. Glob. environ. Chang., 19: 306-315.
EDMONDS, W.D. & ZÍDEK, J., 2004.- Revision of the Neotropical dung beetle genus oxysternon (Scarabaeidae: Scarabaeinae: Phanaeini). Folia Heyrovskyana, Supplementum, 11: 1-58.
ESPARZA, A.C. & AMAT G., 2007.- Composición y riqueza de escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) en un gradiente altitudinal de selva húmeda tropical del Parque Nacional Natural Catatumbo – Barí (Norte de Santander), Colombia. Actual Biol., 29: 181-192.
GARCÍA, J.C. & PARDO, L.C., 2004.- Escarabajos Scarabaeinae saprófagos (Coleoptera: Scarabaeidae) en un bosque muy húmedo premontano de los Andes Occidentales Colombianos. ecol. apl., 3 (2): 59-63.
HALFFTER, G. & MATHEWS, G., 1966.- The Natural History of Dung Beetles of the Subfamily Scarabaeinae (Coleoptera, Scarabaeinae). Fol. entomol. Mex., 12-14: 1-312.
HANSKI, I. & CAMBEFORT, Y., 1991.- Dung beetle ecology. Princeton University Press, Princeton, New Jersey.HOUGHTON, R.A., 1994.- The worldwide extent of land-use change. Bioscience, 44: 305-313.HUERTAS, B., ARIAS, J. & PARDO, L.C., 2003.- Estudio preliminar de los escarabajos coprófagos (Coleoptera: Scarabaeidae) de
la Serranía de Los Churumbelos, Cauca (Expedición Colombia 1998). Bol. Cient. Mus. Hist. Nat., 7: 215-218.IPCC (INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE), 2007.- Synthesis Report. Contribution of Working
Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change Geneva, Switzerland.IPCC (INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE) - WORKING GROUP III, 2000.- escenario de emisiones:
resumen para responsables de políticas. Grupo Intergubernamental de Expertos sobre el Cambio Climático, Ginebra.IPCC (INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE) - WORKING GROUP II, 2002.- Cambio climático y
biodiversidad. IPCC, Geneva, Switzerland. KATTAN, G. & NARANJO, L.G., 2008.- Regiones biodiversas, herramientas para la planificación de sistemas regionales de áreas protegidas.
WCS Colombia, Fundación EcoAndina, WWF Colombia.

Bedir Germán Martínez-Quintero, Alejandro Echeverri-Rubio, Fabián Guillermo Gaviria-Ortiz206bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 1
90-2
06
LIN, B., 2013.- Creating Sustainable Cities – What can we learn from each other? Urban land use change and human-environmental well-being. Research Scientist, 09 August 2013.
MEDINA, C.A. & KATTAN, G., 1996.- Diversidad de coleópteros coprófagos (Scarabaeidae) en la Reserva Forestal de Escalerete. Cespedecia, 21(68): 89-102.
MILLENNIUM ECOSYSTEM ASSESSMENT, 2005.- ecosystems and Human Well-being: Biodiversity Synthesis. World Resources Institute, Washington, DC.
MORALES, N., 2012.- Modelos de distribución de especies: Software Maxent y sus aplicaciones en Conservación. Rev. Cons. Amb.., 2 (1): 1-3.
NICHOLS, E., LARSEN, T., SPECTOR, S., DAVIS, A.L., ESCOBAR, F., DÁVILA, M. & VULINEC, K., 2007.- The Scarabainae Research Network. Global dung beetles response to tropical forest modification and fragmentation: A quantitative literature review and meta-analysis. Biol Cons., 137: 1-19.
NICHOLS, E., URIARTE, M., BUNKER, D.E., FAVILA, M.E., SLADE, E.M., VULINEC, K. & SPECTOR, S.H., 2013.- Trait‐dependent response of dung beetle populations to tropical forest conversion at local and regional scales. ecology, 94 (1): 180-189.
NORIEGA, J.A., MORENO, J., OTAVO, S. & CASTAÑO, E., 2012.- New departmental records for Digitonthophagus gazella (Coleoptera: Scarabaeidae) in Colombia. Acta biol. Colomb., 17 (1): 201-204.
PARMESAN, C. & YOHE, G., 2003.- A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421 (6918): 37-42.
PENTILÄ, A., SLADE, E.M., SIMOJOKI, A., RIUTTA, T., MINKKINEN, K. & ROSLIN, T., 2013.- Quantifying Beetle-Mediated Effects on Gas Fluxes From Dung Pats. PloS oNe, 8: 71454.
PHILLIPS, S.J., ANDERSON, R.P. & SCHAPIRE, R.E., 2006.- Maximum entropy modeling of species geographic distributions. ecol. Model., 190: 231-259.
PHILLIPS, S. & DUDÍK, M., 2008.- Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. ecography, 31; 161-175.
RASKIN, P., MONKS, F., RIBEIRO, T., VUUREN, D.V. & ZUREK, M., 2005.- Global Scenarios in Historical Perspective: 35-44. Cap. 2. (en) MILLENNIUM ECOSYSTEM ASSESSMENT, ecosystems and Human Well-being: Scenarios. Island Press, Washington, DC.
ROCCHINI, D., LOBO, J.M., JIME, A., BACARO, G. & CHIARUCCI, A., 2011.- Accounting for uncertainty when mapping species distributions: the need for maps of ignorance. Prog. in Phys. Geogr., 35: 211-226.
SPECTOR S. 2006. Scarabaeinae dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) an invertebrate focal taxon for biodiversity research and conservation. The Coleopterist Bulletin 5: 71-83.
SWENSON, J.J. & FRANKLIN, J., 2000.- The effects of future urban development on habitat fragmentation in the Santa Monica Mountains. landscape ecol., 15: 713-730.
VALDERRAMA, W., 2013.- Modelo de Nicho Ecológico mediante el método de máxima entropía (MaxEnt) para distribución de rabia silvestre en animales, transmitida por el murciélago Desmodus rotundus en Perú. Boletín epidemiológico SeNASA/, Mes 10, semanas 40-44.
VANDERGAST, A.G., BOHONAK, A.J., WEISSMAN, D.B. & FISHER, R.N., 2007.- Understanding the genetic effects of recent habitat fragmentation in the context of evolutionary history: phylogeography and landscape genetics of a southern California endemic Jerusalem cricket (Orthoptera: Stenopelmatidae: Stenopelmatus). Molecular ecology, 16(5): 977-992.
VULINEC, K., 2000.- Dung beetles (Coleoptera: Scarabaeida e), monkeys, and conservation in Amazonia. Fla. entomol., 83 (3): 229-241.
WILSON, R.D, TRUEMAN, J.W.H, WILLIAMS, S.E. & YEATE, D.K., 2007.- Altitudinally restricted communities of Schizophoran flies in Queensland’s Wet Tropics: vulnerability to climate change. Biodivers. Conserv., 16 (11): 3163-3177.
WILLIAMS, J.N., SEO, C., THORNE, J., NELSON, J.K., ERWIN, S., O’BRIEN, J.M. & SCHWARTZ, M.W., 2009.- Using species distribution models to predict new occurrences for rare plants. Div. Dist., 15: 565-576.
YUKIMOTO, S., ADACHI, Y., HOSAKA, M., SAKAMI, T., YOSHIMURA, H., HIRABARA, M., TANAKA, T.Y., SHINDO, E., TSUJINO, H., DEUSHI, M., MIZUTA, R., YABU, S., OBATA, A., NAKANO, H., KOSHIRO, T., OSE, T. & KITOH, A., 2012.- A new global climate model of Meteorological Research Institute: MRI-CGCM3–model description and basic performance. J Meteorol Soc Jpn., 90A: 23-64.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
neW sPeCies of tHe Genus Trigonospila PoKornY (diPtera: taCHinidae) Parasitises adult Compsus sp. sCHoenHerr
(ColeoPtera: CurCulionidae) in ColoMBia*
Natalia Vinasco1, luis Fernando Vallejo2, Alberto Soto3
abstract
Trigonospila unicaldasi new specie, is described for the first time for science. In order to differentiate it from other rarely occurring species in the Americas, key characteristics of the adult were identified through scanning electron microscopy. The species was found parasitising adult weevils of the genus Compsus sp. (Coleoptera: Curculionidae) associated with citrus crops in the central coffee-growing region of Colombia. The interesting symbiotic phenomenon of control described in this paper is considered unusual in the biology of Tachinidae given that only 5-10% of the family parasitise their hosts at the adults stage.
Key words: parasitism, adults, scanning electron microscope, description.
nueVa esPeCie del GÉnero Trigonospila PoKornY (diPtera: taCHinidae) Para ColoMBia, Parasitando adultos de Compsus sp. sCHoenHerr (ColeoPtera: CurCulionidae)
resumen
Trigonospila unicaldasi nueva especie, es registrada por primera vez para la ciencia con el propósito de diferenciarla de otras especies que son de escasa ocurrencia en las Américas. Para lograr este objetivo se consideraron las estructuras clave del adulto utilizando la técnica de Microscopía Electrónica de Barrido (MEB) para su completa descripción. La especie se encontró parasitando adultos de Compsus sp. (Coleoptera: Curculionidae) asociados a cultivos de cítricos en la zona cafetera central de Colombia. El interesante fenómeno simbiótico de control mencionado en este documento se considera un caso poco usual en la biología de los Tachinidae debido a que solo del 5% al 10% de estos parasitan especies en estado adulto.
Palabras clave: parasitismo, adultos, Microscopía Electrónica de Barrido, descripción.
* FR: . FA: .1 Ingeniera Agrónoma. Universidad de Caldas. Facultad de Ciencias Agropecuarias. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0002-179316752 Biólogo, M.Sc., Ph.D. Universidad de Caldas. Departamento de Producción Agropecuaria. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0002-0610-12703 I.A., M.Sc., Ph.D. Universidad de Caldas. Departamento de Producción Agropecuaria. Manizales, Colombia. E-mail: [email protected] ORCID: 0000-0002-9727-8919
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 207-219. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: VINASCO, N., VALLEJO, L.F. & SOTO, A., 2017.- New species of the genus Trigonospila Pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr (Coleoptera: Curculionidae) in Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 207-219. DOI: 10.17151/bccm.2017.21.2.14

Natalia Vinasco, luis Fernando Vallejo, Alberto Soto208bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
introduCtion
Trigonospila Pokorny 1886 (Tachinidae: Blondeliini) is a cosmopolitan genus, present in the Old World in Eurasia, Australia, Oriental and Afrotropical region. In the New World is represented for three from the north of Mexico (O’HARA & WOOD, 2004) and four in South America, of which, two were found on larvae of Crambidae and Pyralidae (Lepidoptera), reared in Guanacoste, Costa Rica (JANZEN & HALLWACHS, 2005). Recently, FLEMING et al. (2015) describe three new species of the genus Trigonospila, and describe the male of Trigonospila panamensis (Townsend), all reared from wild-caught caterpillars collected from Área de Conservación Guanacaste, in northwestern Costa Rica. BYUN & HAN (2010) differentiated 18 nominal species (3 Palaearctic, 3 Oriental, 5 Australasian, 4 Afrotropical, and 3 Nearctic), with two species of the families Oecophoridae and Gelechiidae (Lepidoptera) parasitised by Trigonospila brevifacies (Crosskey 1973), and the Asian beetle Plesiophthalmus nigrocyaneus (Motschulsky) (Tenebrionidae) as host to Trigonospila vittigera (Coquillett). T. brevifacies was introduced to Australia and northern New Zealand to control the tortricid pest epiphyas postvittana (Walker) between 1967 and 1973, and in southern New Zealand in 1999 (SHAW et al., 2001). The species is known to oviposit directly between the head and thorax of lepidopteran larvae, where the newly hatched larvae of the parasitoid complete their whole life cycle (GREEN, 1984). Parasitism on adult insect hosts is poorly known in Tachinid flies (BROWN et al., 2010). T. unicaldasi represents a promising parasitoid for the control of Compsus sp. (ARNETT et al., 2002), an important economic species, detected since 1939, mainly in citrus crops in the departments of Quindío, Caldas, Valle, Tolima, Antioquia and Risaralda in Colombia, causing considerable damage to foliage and tree roots (PÁSSARO, 2012).
This study describes a new species of the genus Trigonospila found parasitising adults of the citrus weevil, Compsus sp. in Colombia. Using images obtained from scanning electron microscopy we describe the male holotype and differential female characteristics which allow the species to be described as Trigonospila unicaldasi.
Materials and MetHods
The description follows the format of BYUN & HAN (2010) which includes several morphometric ratios: frons-head ratio (frons/head width); eye ratio (length/width); arista-antenna ratio (arista length/antenna length excluding arista); wing-thorax ratio (wing length/thorax length); abdomen ratio; genital differences. The structures morphological in this study was based conform to the keys found in WOOD (1985) and WOOD & ZUMBADO (2010).

New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 209
Collection of entomological material: Twenty insects were taken from hosts Citrus limon (lemon) trees, Rubus glauca (andean blackberry) bushes and Passiflora molissima (curuba) plants, grown in an experimental plot of the Granja Tesorito, belonging to the Universidad de Caldas. The university farm is located 6 km along the highway between Manizales and Bogotá in Valle de Maltería. The specimens were collected using the umbrella method, which consists of putting a plastic under each plant and then shaking it so that the insects fall on it facilitated by Compsus sp. adults thanatosis behaviour, or apparent death some of which had previously been parasitised by T. unicaldasi. The specimens were collected in plastic containers, measuring 20 x 15 x 6 cm, and labelled with the following information: date, host plant, altitude above sea level, quantity per host plant and name of collector. The material was taken to the Entomological Laboratory of the Universidad de Caldas (LEUC) to be disinfected (washed with distilled water, 3% sodium hypochlorite and 75% ethanol for 5 minutes). Fifteen specimens of Compsus sp. adults were placed on sand and filter paper inside sterilised plastic containers measuring 15 x 15 x 10 cm. From this moment on the parasitised specimens were monitored every 48 hours in order to obtain immature stages of T. unicaldasi. Pupae of T. unicaldasi were transferred to a cage designed for manipulating live insects (BIOQUIP® reference 1450TC). Small recipients with cotton wool soaked in water and 10% honey solution were installed for flies to feed on until adult specimens were obtained. Twenty individuals were chosen from the whole population (10 males and 10 females) for analysis with scanning electron microscopy.
scanning electron microscope analysis: Specimens of T. unicaldasi were taken to the Electron Microscope Laboratory of the Universidad de Caldas where samples were placed on plates and photographed with an FEI- QUANTA 200 electron microscope and processed in Microsoft Office 2010. Dimensions given for the anatomical structures are the mean of measurements taken from 10 male and 10 female specimens, obtained via scanning electron microscopy. Ten of the specimens were deposited in the Laboratory of Entomology at the Faculty of Agricultural Sciences, Universidad de Caldas, Manizales, Colombia.
results
Trigonospila unicaldasi Vinasco, Vallejo and soto, sp. nov. (Figs. 1-11)
type material: Holotype labelled as “Colombia, department of Caldas, municipality of Manizales, Granja Tesorito, Valle de Maltería, Universidad de Caldas, 2280 m, January 20, 2011, in adults of Compsus sp. feeding on Citrus limon L. (lemon), Alberto Soto Giraldo”. Original label in Spanish: “Colombia, departamento de Caldas, Municipio de Manizales, Granja Tesorito, Valle de Maltería, Universidad de Caldas 2280 m, enero 20 de 2011, en adultos de Compsus sp. alimentándose de hojas Citrus limon L. (limón), Alberto Soto Giraldo”.

Natalia Vinasco, luis Fernando Vallejo, Alberto Soto210bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
Holotype deposited at the Entomology Laboratory Collection, Universidad de Caldas (LEUC), Manizales, Colombia.
description of the male holotype
diagnosis: Terminology used by BYUN & HAN (2010) and the morphological description by Wood (1985) and WOOD & ZUMBADO (2010) of members of the genus Trigonospila is based on a combination of the following characters: transverse body bands, compound eyes with proclinate and reclinate orbital setae, upper frontal setae, ocellar setae, parafacial setae, facial ridge, vibrissae and subvibrissae, epistomal suture, gena, antennae, flagellum, flagellomere, arista, thoracic segments, wing venation, distribution of setae on legs (Fig. 1). Measurements are given in millimetres.
Trigonospila unicaldasi, adult male. Lateral view.figure 1.
Male: Body pale yellow with vertical brown bands on thorax and horizontal bands on abdomen; wing length: 1.12-1.54; thorax length: 1.09-0.91; body length, from cephalic capsule to posterior end of abdomen 3.0-3.82; mid abdominal zone 1.14-1.53. Semicircular compound eyes with minute setae 0.01in length and alternating with ommatidia. Eye ratio 0.47-0.82, arista-antenna ratio 0.53-0.86; gena-eye ratio 0.09; exterior setae present; occelar seta 0.29, as long as outer vertical seta; postocellar

New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 211
seta 0.07; paravertical seta short but different to anterior, almost 0.14, as long as ventral outer seta; ocellar triangle; bare lunule; orbital plate yellow, same shade as dorso, with wide, long setae; orbital seta markedly smaller than frontal setae; parallel setae at least twice as long as inner; divergent ocellar setae; dark, bare frontal vitta; dark brown arista with 3 aristomere; scape very short and wide, pedicel slender, first flagellomere markedly wide and long; facial carina wide, concave, as long as length of first flagellomere.
Head (Figs. 2-3). Yellow-brown in colour, dimensions: 1.25 frontal width, and 0.96 laterally. Parafacial bare, paler than other regions around compound eye; postocular wide in relation to eye margin; occiput with small setae; postcranium with one row of black setae, longer than postocular row, rest of the postcranium covered with small, pale setae. Postgena short, 0.08 in height, moderately swollen; genal setae differentiated and genal dilation not well developed; prominent vibrissa, markedly wide at base, sufficiently long to overlap, and project over epistome (Fig. 2), supravibrissa also evident. Supravibrissal setulae absent; 4-6 subvibrissal setae; mouthparts with lightly clavate palps and with small setae (Fig. 3); prementum dark brown, with small setae; labellum paler and covered with abundant setae.
Head of male Trigonospila unicaldasi. Frontal view (note appearance of lunule).
figure 2.
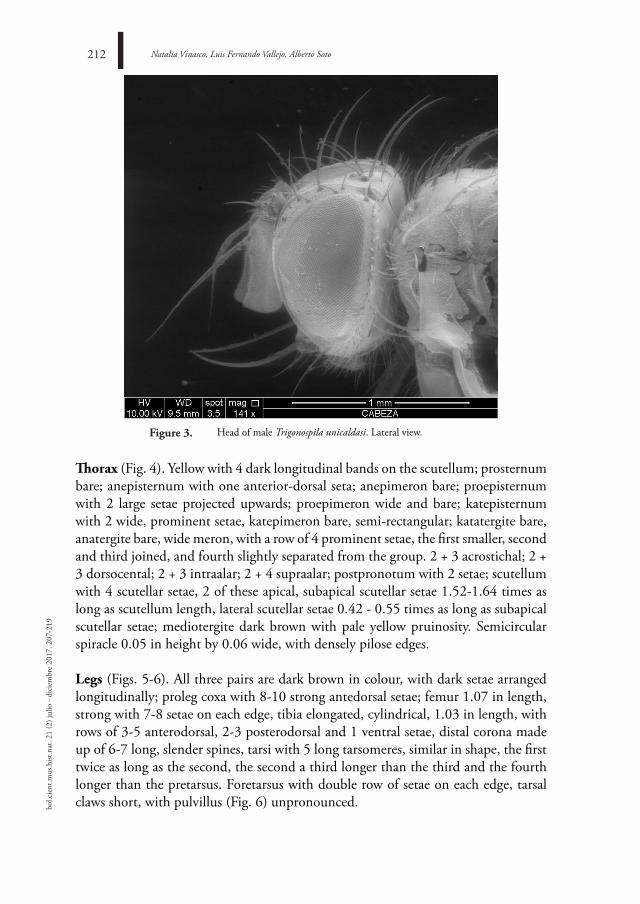
Natalia Vinasco, luis Fernando Vallejo, Alberto Soto212bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
Head of male Trigonospila unicaldasi. Lateral view.figure 3.
Thorax (Fig. 4). Yellow with 4 dark longitudinal bands on the scutellum; prosternum bare; anepisternum with one anterior-dorsal seta; anepimeron bare; proepisternum with 2 large setae projected upwards; proepimeron wide and bare; katepisternum with 2 wide, prominent setae, katepimeron bare, semi-rectangular; katatergite bare, anatergite bare, wide meron, with a row of 4 prominent setae, the first smaller, second and third joined, and fourth slightly separated from the group. 2 + 3 acrostichal; 2 + 3 dorsocental; 2 + 3 intraalar; 2 + 4 supraalar; postpronotum with 2 setae; scutellum with 4 scutellar setae, 2 of these apical, subapical scutellar setae 1.52-1.64 times as long as scutellum length, lateral scutellar setae 0.42 - 0.55 times as long as subapical scutellar setae; mediotergite dark brown with pale yellow pruinosity. Semicircular spiracle 0.05 in height by 0.06 wide, with densely pilose edges.
legs (Figs. 5-6). All three pairs are dark brown in colour, with dark setae arranged longitudinally; proleg coxa with 8-10 strong antedorsal setae; femur 1.07 in length, strong with 7-8 setae on each edge, tibia elongated, cylindrical, 1.03 in length, with rows of 3-5 anterodorsal, 2-3 posterodorsal and 1 ventral setae, distal corona made up of 6-7 long, slender spines, tarsi with 5 long tarsomeres, similar in shape, the first twice as long as the second, the second a third longer than the third and the fourth longer than the pretarsus. Foretarsus with double row of setae on each edge, tarsal claws short, with pulvillus (Fig. 6) unpronounced.

New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 213
Thorax of male Trigonospila unicaldasi. Lateral view.figure 4.
Legs of male Trigonospila unicaldasi. General view of the pro, meso and meta legs.
figure 5.
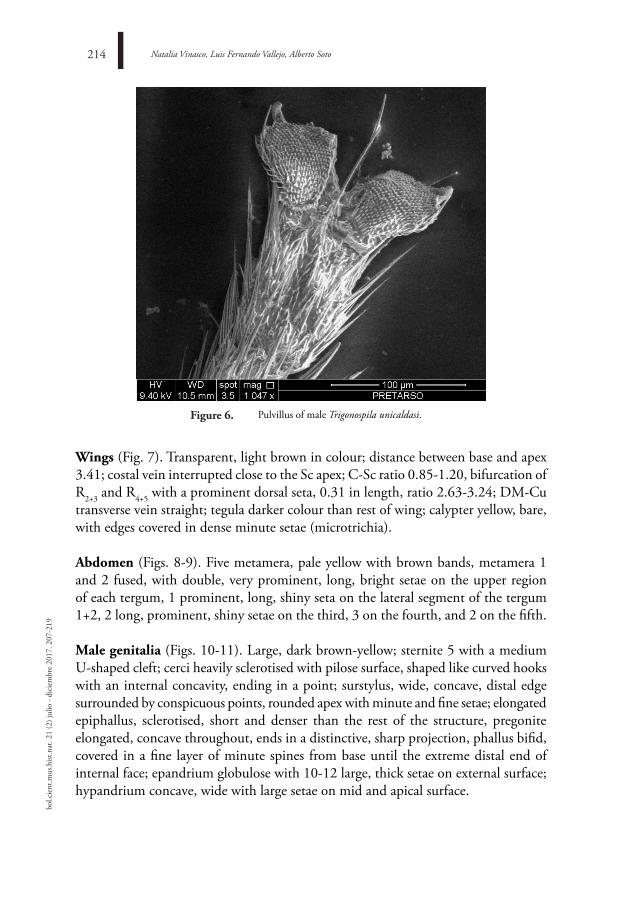
Natalia Vinasco, luis Fernando Vallejo, Alberto Soto214bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
Pulvillus of male Trigonospila unicaldasi. figure 6.
Wings (Fig. 7). Transparent, light brown in colour; distance between base and apex 3.41; costal vein interrupted close to the Sc apex; C-Sc ratio 0.85-1.20, bifurcation of R2+3 and R4+5 with a prominent dorsal seta, 0.31 in length, ratio 2.63-3.24; DM-Cu transverse vein straight; tegula darker colour than rest of wing; calypter yellow, bare, with edges covered in dense minute setae (microtrichia).
abdomen (Figs. 8-9). Five metamera, pale yellow with brown bands, metamera 1 and 2 fused, with double, very prominent, long, bright setae on the upper region of each tergum, 1 prominent, long, shiny seta on the lateral segment of the tergum 1+2, 2 long, prominent, shiny setae on the third, 3 on the fourth, and 2 on the fifth.
Male genitalia (Figs. 10-11). Large, dark brown-yellow; sternite 5 with a medium U-shaped cleft; cerci heavily sclerotised with pilose surface, shaped like curved hooks with an internal concavity, ending in a point; surstylus, wide, concave, distal edge surrounded by conspicuous points, rounded apex with minute and fine setae; elongated epiphallus, sclerotised, short and denser than the rest of the structure, pregonite elongated, concave throughout, ends in a distinctive, sharp projection, phallus bifid, covered in a fine layer of minute spines from base until the extreme distal end of internal face; epandrium globulose with 10-12 large, thick setae on external surface; hypandrium concave, wide with large setae on mid and apical surface.
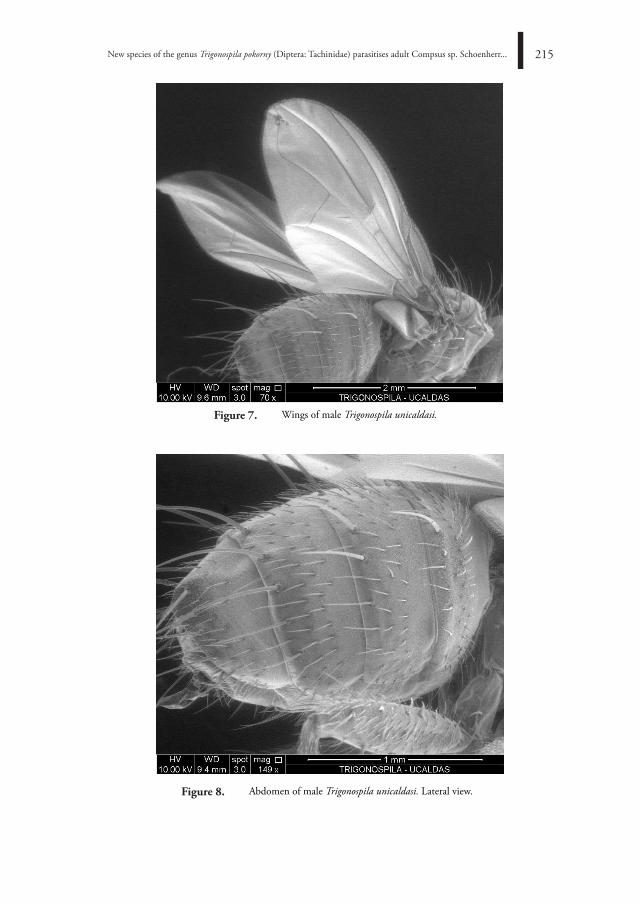
New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 215
Wings of male Trigonospila unicaldasi.figure 7.
Abdomen of male Trigonospila unicaldasi. Lateral view.figure 8.
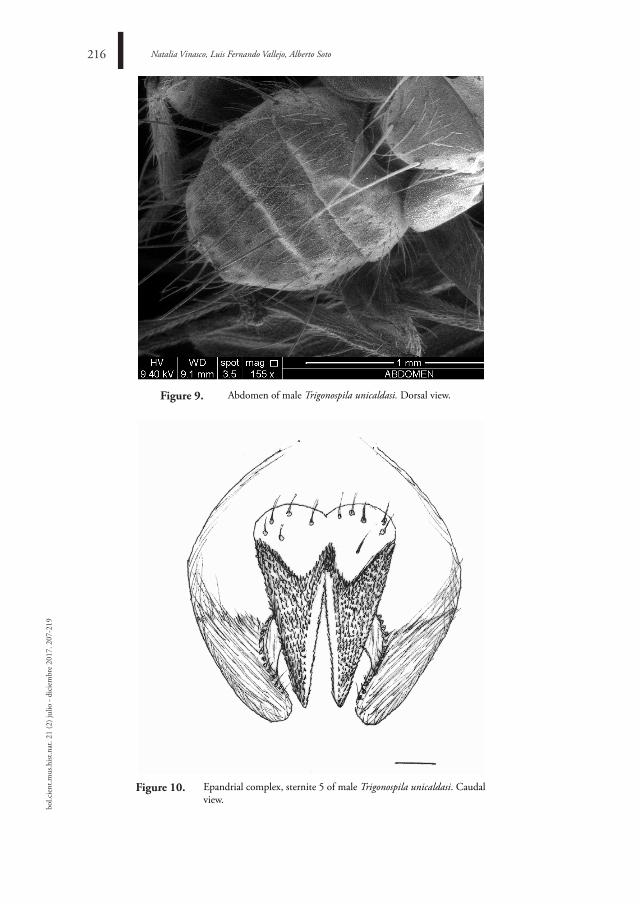
Natalia Vinasco, luis Fernando Vallejo, Alberto Soto216bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
Abdomen of male Trigonospila unicaldasi. Dorsal view.figure 9.
Epandrial complex, sternite 5 of male Trigonospila unicaldasi. Caudal view.
figure 10.

New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 217
female. Similar to males except for genital structure. Body length from cephalic capsule to posterior end of abdomen 3.41-5.0; mid abdominal zone 1.34-1.82; wing length 3.445; thorax length 1.199; frons-head ratio 0.70-0.94; gena-eye ratio 0.174; from 9 to 10 frontal setae; 3 orbital setae, 0.50-0.81 times as long as inner vertical seta; 1 reclinate orbital seta, 0.54-0.75 times as long as inner vertical seta. Thorax and abdomen as described for male except for female terminalia.
Material examined. Male holotype: Colombia, Caldas, km 6 on Manizales – Bogotá Highway, in Valle de Maltería, 5°06′ N, 75°33′ W, 2160, immature stages of Trigonospila sp collected from Compsus sp. in plantations of Passiflora mollisima, Rubus glauca aned Citrus limon x limon 1, 2011, col. A. Soto. G (LEUC). Paratypes: same as holotype: 5 adult males, 5 adult females.
etymology. The specific epithet ‘unicaldasi’ is in homage to the Universidad de Caldas, institution supporting this research and where the authors are based. Furthermore, the parasi.0toid was found for the first time within the university farms.
Geographic distribution. Colombia, Caldas, Manizales. Pre-montane Forest (bh-PM) according to the classification of life zones by HOLDRIDGE (1967).
Epandrial complex, sternite 5 of male Trigonospila unicaldasi. Lateral view.
figure 11.

Natalia Vinasco, luis Fernando Vallejo, Alberto Soto218bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
07-2
19
Host. Compsus sp. (Schoenherr) (Coleoptera: Curculionidae).
Biology. Adult Compsus sp. parasitised by T. unicaldasi shows slow behaviour and a dark colouration at the end of its abdomen (Fig. 9). At an average temperature of 18°C and relative humidity of 75%, adult females survive for an average of 35±10 days (n = 20) and adult males for an average of 25±11 days (n = 23). Sex ratio in adults is 1:2 (females:males) for 78% (n = 47) of emergent adults.
associated plants. Although it is known that Compsus sp. attack several species of plants, especially citrus fruits, the specimens were collected mainly on P. mollisima, where the largest number of parasitised adults was found, possibly due to the availability of nectar from its flowers serving as a food source for adults of T. unicaldasi.
observations. As stated above, the genus Trigonospila is poorly known in the Neotropical region and is not comprehensively described in literature. As a result, species are very difficult to identify with certainty. T. unicaldasi has been compared with some Trigonospila species housed at the Canadian National Collection of Insects Agriculture and Agri-Food. Dr James O’Hara, (Systematic Entomology, Eastern Cereal and Oil Seed Centre, Ottawa, Canada) identified and placed these samples within the genus after comparisons with specimens from the abovementioned collection. In order to put a name to this potential biological control agent, both its identification by O’Hara and the presence of this species in the Neotropics is sufficient to be able to name this a new species to science.
disCussion
The genus Trigonospila has been poorly studied around the world, with most studies concerning the Neartic, Paleartic and Asia. The first record for Colombia comes from adult citrus weevils, Compsus sp., parasitised by T. unicaldasi, in a pre-montane forest. The weevils were observed to reduce their movement before mature larvae of the parasitoid emerged through the exterior anal canal or through the cervical membrane, decapitating the host insect. Most of the time, parasitoid larvae emerge after the weevil has died and move slowly until finding a suitable place in which to pupate (SOTO, 2002). The parasitoids obtained belong to the Diptera order, Tachinidae family, Exoristinae subfamily and Blondellini tribe (SOTO & OCAMPO, 2011).
T. unicaldasi represents a promising biological control agent for adult Compsus (Coleoptera: Curculionidae) given that this coleopteran has become a limiting pest for the citrus fruit industry due to inappropriate management by farmers in certain agricultural areas of the country. The pest has caused a reduction in vigour and productivity of citrus trees; increased their susceptibility to radicular fungal attacks;

New species of the genus Trigonospila pokorny (Diptera: Tachinidae) parasitises adult Compsus sp. Schoenherr... 219
and, in many cases, their death (CORRALES, 2002). Furthermore, is it unusual to find a Tachinid parasitising coleopteran adults, rather, it is more common to find them parasitising immature stages, thus this represents a novel and interesting relationship between parasitoid and host in the field (BROWN et al., 2010).
This study contributes to knowledge of the family Tachinidae and principally, the genus Trigonospila, poorly known in Latin America. Also, it aims to resolve the confusion arisen between the genera Trigonospila and lyxophaga, especially concerning differences between the two, such as their parasitism mechanisms, morphotypical dimensions, antennal segments, genital differences and phenotypic characteristics. These characteristics have been observed using scanning electron microscopy, which guarantees the veracity of, and complements, morphological studies.
aCKnoWledGeMents
We are very grateful to Office of the Vice-rectorate for Research at the Universidad de Caldas and INNPULSA for funding this research; also, to the researchers who took part in the study and all those who commented on this paper.
referenCes
ARNETT, R.H., THOMAS, M.C., SKELLEY, P.E. & FRANK, J.H., 2002.- American Beetles: Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton.
BROWN, B.V., BORKENT, A., CUMMING, J.M., WOOD, D.M., WOODLEY, N.E. & ZUMBADO, M.A., 2010.- Manual of Central American Diptera: Volume 2. NRC Research Press.
BYUN, H.W & HAN, H.Y., 2010.- Taxonomic Review of the Genus Trigonospila Pokorny (Diptera: Tachinidae: Blondeliini) in Korea. Korean J. Syst. Zoo., 26 (3): 243- 249.
CORRALES, G.A., 2002.- Manual Ilustrado para la producción de Cítricos. Asocítricos. Produmedios, Bogotá D.C.FLEMING, A., WOOD, D., JANZEN, D., HALLWACHS, W. & SMITH, M., 2015.- Three new species of Trigonospila Pokorny
(Diptera: Tachinidae), from Área de Conservación Guanacaste, northwestern Costa Rica, with a key for their identification. J. Biod. Data, 3: 1-32.
GREEN, O.R., 1984.- New Zealand host and locality records for an introduced totricid parasite, Trigonospila brevifacies (Hardy) (Diptera: Tachinidae). New Zealand ent., 8: 69-71.
HOLDRIDGE, L.R., 1967.- life Zone ecology. Tropical Science Center, San José, Costa Rica. JANZEN, D.H. & HALLWACHS, W., 2005.- Dynamic database for an inventory of the macrocaterpillar fauna, and its food plants
and parasitoids of the Área de Conservación Guanacaste (ACG), northwestern Costa Rica. University of Pennsylvania. Available: http://janzen.sas.upenn.edu/
O’HARA, J.E. & WOOD, D.M., 2004.- Catalogue of the Tachinidae (Diptera) of America north of Mexico. Memoirs on entomology International, 18 (4): 410 pp.
PÁSSARO, C.P., 2012.- Cítricos: cultivo, poscosecha e industrialización. Serie Lasallista Investigación y Ciencia. Corporación Universitaria Lasallista, Caldas.
SHAW, P.W., LO, L.P. & WALLIS, D.R., 2001.- Recent introduction and reestablishment of the leafroller parasitoid Trigonospila brevifacies (Hardy) (Diptera: Tachinidae) in Nelson. New Zealand P. Prot., 54: 33-36.
SOTO, G.A., 2002.- Nuevo registro de parasitoide del picudo de los cítricos (Compsus n. sp.) en Colombia. Bol. Fito., 60: 1-2.SOTO, G.A. & OCAMPO, A., 2011.- Estudio preliminar de Trigonospila sp. (Diptera: Tachinidae), Parasitoide de Compsus viridilineatus
(Coleoptera: Curculionidae). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 15 (1): 150-159. WOOD, D.M., 1985.- A taxonomic conspectus of the Blondeliini of North and Central America and the West Indies (Diptera:
Tachinidae). Memoirs of the entomological Society of Canada, 132: 1-132.WOOD, D.M. & ZUMBADO, M.A., 2010.- Tachinidae (tachinid flies, parasitic flies): 1343-1417 (in) BROWN, B.V., BORKENT,
A., CUMMING, J.M., WOOD, D.M., WOODLEY, N.E. & ZUMBADO, M.A. (eds.), Manual of Central American Diptera. 2. NRC Research Press, Ottawa, xvi + 715-1442 pp.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
Sphaeropthalma pinedai sp. nov.: PriMer reGistro de un Mutílido BraQuíPtero Para aMÉriCa del sur
(HYMenoPtera: Mutillidae)*
Roberto A. Cambra1, Diomedes Quintero A.2
resumen
Sphaeropthalma pinedai sp. nov. (Hymenoptera: Mutillidae), especie endémica de Chile, se describe e ilustra de ambos sexos. S. pinedai representa el primer registro para América del Sur de una especie de Mutillidae con machos braquípteros. Se presenta un listado de especies de Mutillidae en América con machos braquípteros o ápteros.
Palabras clave: Sphaeropthalminae, especie nueva, taxonomía, Mutillidae nocturna, Neotropical.
Sphaeropthalma pinedai sp. nov.: first reCord of BraCHYPterous Mutillid for soutH aMeriCa
(HYMenoPtera: Mutillidae)
abstract
Sphaeropthalma pinedai sp. nov. (Hymenoptera: Mutillidae), an endemic species of Chile is described and illustrated by gender. S. pinedai represents the first record for South America of a Mutillid species with brachypterous males. A list of Mutillidae species from America with brachypterous or apterous males is given.
Key words: Sphaeropthalminae, new species, taxonomy, nocturnal Mutillidae, Neotropical.
* FR: 10-IV-2017 . FA: 25-VIII-2017.1 Museo de Invertebrados G. B. Fairchild, Universidad de Panamá. Panamá, República de Panamá. Correo para correspondencia - E-mail: [email protected] ORCID: 0000-0002-3799-47102 Museo de Invertebrados G. B. Fairchild, Universidad de Panamá. Panamá, República de Panamá. E-mail: [email protected] ORCID: 0000-0002-2961-2196
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 220-230. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: CAMBRA, R.A. & QUINTERO, D., 2017.- Sphaeropthalma pinedai sp. nov.: primer registro de un mutílido braquíptero para América del Sur (Hymenoptera: Mutillidae). Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 220-230. DOI: 10.17151/bccm.2017.21.2.15

Sphaeropthalma pinedai sp. nov.: Primer registro de un mutílido braquíptero para América del sur... 221
introduCCiÓn
En machos de avispas mutílidas la reducción de alas (braquíptero) o ausencia de ellas (áptero) es una condición poco común. Machos de mutílidos ápteros o braquípteros para América se han registrado en los siguientes siete géneros: Chilemutilla Cambra & Quintero, 2007, Dasymutilla Ashmead, 1899, limaytilla Casal, 1964, Myrmilloides Andre, 1903, Morsyma Fox, 1899, Sphaeropthalma Blake, 1871 y Stethophotopsis Pitts, 2000 (BLAKE, 1872; MICKEL, 1936; KROMBEIN, 1940; SCHUSTER, 1945; PITTS & MCHUGH, 2000; MANLEY & PITTS, 2002; CAMBRA & QUINTERO, 2007). MANLEY & PITTS (2002) informan conocer que individuos de Sphaeropthalma unicolor (Cresson) y odontophotopsis melicausa melicausa (Blake) mastican sus alas para desprenderlas, sin brindar más detalles de tales observaciones.
El presente trabajo tiene la finalidad de describir el primer macho de Mutillidae braquíptero para América del Sur y presentar un listado de todas las especies de mutílidos con machos braquípteros o ápteros registrados para América.
Materiales Y MÉtodos
La mayoría de los especímenes fueron capturados con seis trampas de caída o pitfall traps que fueron colocadas debajo de un arbusto de Baccharis linearis (Asteraceae). Las trampas consistieron de vasos plásticos de 8 cm de diámetro y 10 cm de profundidad, algunos con líquido azucarado hasta la mitad y otros vacíos. Las seis trampas fueron colocadas en conjunto, separadas entre sí por escasos centímetros. Se pusieron en el suelo barreras como guías para que los ejemplares se toparan con estas y caminaran hacia las trampas. Las trampas fueron revisadas diariamente durante los días 27, 28 y 29 de noviembre de 2015. Dos hembras fueron recolectadas de forma manual cuando caminaban durante el día, aproximadamente a las 14:00 horas, en un sendero arenoso (Fig. 1).
Las fotografías de los especímenes fueron obtenidas con una cámara digital olympus Stylus, utilizando un estereomicroscopio LEICA M 165C, y realizando el procesamiento de imágenes con el programa ArcSoft PhotoStudio. Las mediciones fueron hechas con un micrómetro adjunto a un lente ocular del estereomicroscopio.
Las siguientes abreviaciones son utilizadas en la descripción del material: T = terguito, S = esternito. Los especímenes se encuentran depositados en la colección del Museo de Invertebrados G. B. Fairchild, Universidad de Panamá (MIUP).

Roberto A. Cambra, Diomedes Quintero A.222bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
20-2
30
Aspectos del hábitat de S. pinedai en Valparaíso, Chile.figuras 1-2.
resultados Y disCusiÓn
Sphaeropthalma pinedai Cambra y Quintero sp. nov.
Holotipo macho. Longitud total 9,4 mm. Coloración: cabeza, mesosoma y primer segmento metasomal rojos; segmentos metasomales dos a siete negros; mandíbulas amarillas, excepto extremo distal que es negro; antenas, tégulas, patas y espolones tibiales amarillos. Cabeza: contorno redondeado (Fig. 6), cubierta con setas blancas, braquiplumosas, erectas y semierectas; vértex, frente y genas con punteado mediano, separado en vértex y genas, más apretado en la frente; distancia desde ocelo posterior al margen interno del ojo 2,9 veces el diámetro mayor del ocelo; carena escrobal ausente; tubérculo antenal liso, sin protuberancia lamelada; escapo con una carenita longitudinal en su mitad apical; primer segmento del flagelo 0,88 veces tan largo como el segundo segmento; mandíbula con su ápice bidentado (Fig. 8), en su reborde dorsal basal con una carena elevada, base del borde ventral con laminilla bien desarrollada (Fig. 9); clípeo ligeramente proyectado y cóncavo en el borde anterior medial, lado de la concavidad distintivamente angulado (Fig. 8). Mesosoma: con setas

Sphaeropthalma pinedai sp. nov.: Primer registro de un mutílido braquíptero para América del sur... 223
blancas braquiplumosas, largas o medianas, erectas y semierectas; dorso del pronoto, mesoescuto y escutelo con punteado grande, profundo y apretado, algunos punteados confluentes; pronoto liso lateralmente, ángulo humeral con una carena transversa en su parte anterior; dorso del propodeo con reticulado grande (Fig. 7); mesopleura lisa, excepto su borde posterior con reticulación mediana; metapleura y área lateral del propodeo lisos; mesoescuto con notaulo profundo y se extiende desde el margen anterior hasta el margen posterior del mesoescuto; mesoesterno simple, sin espina o elevación transversa delante del pliegue de la coxa media; coxas sin carenas o procesos; braquíptero, alas anteriores y posteriores (Figs. 3, 7) muy cortas, ambas extendiéndose hasta el margen anterior del propodeo. Metasoma: T1 con constricción posterior, peciolado (Fig. 3), sin fusionarse uniformemente con T2; T1 es 1,2 veces más largo que su ancho máximo; T1 y S1 con setas blancas braquiplumosas, largas y erectas, excepto ápice de T1 con banda transversa de setas plumosas; T2 a T6 y S2 a S6 con setas blancas, braquiplumosas largas, o plumosas medianas, las setas plumosas son más abundantes en las franjas apicales; T7 glabro, excepto cuarto basal con setas blancas braquiplumosas y margen apical con una banda transversa de setas blancas; T1 con punteado mediano y apretado, excepto el ápice con punturas pequeñas esparcidas; T2 con punteado muy pequeño y esparcido, excepto cerca del borde apical y lateral con punteado mediano y cercanos entre sí, T3 a T6 con punteado pequeño y muy separado; T7 con punteado mediano en su base, área pigidial lisa y sin carena lateral; S1 sin punteado distintivo, en sus tres quintos basales con dos quillas elevadas y muy cercanas, en sus dos quintos apicales divergen y son distintivamente menos elevadas; S7 (Fig. 10) con punteado mediano, sin carena o quillas, el borde apical convexo, sin emarginación medial en donde es ligeramente angulado; T2 y S2 con línea de felpa lateral. Genitalia: parámero (Figs. 13, 14, 15, 16) vista lateral, ligeramente curvo hacia arriba, ancho en su base y adelgazándose suavemente hacia el ápice, terminando en punta, con setas largas en su base, las cuales se hacen más cortas hacia el ápice; cuspis cerca de 2,0 veces el largo del digitus (Fig. 16), ancho en su mitad basal para luego angostarse en su mitad apical, con setas largas; digitus de ancho uniforme, poco piloso; valva peniana (Figs. 11, 12) con una espina aguda apical y una proyección digitiforme subapical, margen dorsal apical con al menos dos setas y el margen ventral, inmediatamente por debajo de la proyección digitiforme, con siete setas.
alotipo hembra (Figs. 4, 5). Longitud total 5,7 mm. Coloración: cabeza, mitad basal de la mandíbula, mesosoma y metasoma rojos; antenas y patas amarillas excepto tibias dos y tres marrón; mitad apical de la mandíbula negra. Cabeza: contorno redondeado, cubierta con setas blancas, braquiplumosas, erectas y semierectas; vértex, frente y genas con punteado mediano, apretadas y separadas en el vértex y frente, confluentes en las genas; carenas escrobal y genal ausentes; tubérculos antenales simples, sin elevaciones laminares o tuberculares; escapo no carenado; primer segmento del flagelo 1,2 veces tan largo como el segundo segmento; mandíbula bidentada, el diente dorsal preapical muy pequeño, borde ventral con dentículo basal bien desarrollado; clípeo con carena

Roberto A. Cambra, Diomedes Quintero A.224bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
20-2
30
transversa arqueada. Mesosoma: piriforme en vista dorsal (Fig. 4), más ancho en el área pronotal para luego angostarse hacia el espiráculo propodeal, margen lateral del propodeo casi recto; dorso del mesosoma con tres carenas transversas sobre la mitad posterior, estas muy bajas y ligeramente convexas anteriormente, la carena próxima al propodeo es más pequeña; dorso del propodeo con reticulaciones medianas en su mitad anterior, la mitad posterior mayormente lisa; dorso del mesosoma con setas doradas braquiplumosas y plumosas, largas o medianas, erectas y semierectas; área lateral del mesosoma con setas braquiplumosas, largas y esparcidas; dorso del pronoto, mesonoto y metanoto con punteado grande, profundo y apretado; pronoto con una quilla transversa latero-anterior; mesopleuron, metapleuron y área lateral del propodeo lisos, sin punteado distintivo; coxas sin carenas o procesos. Metasoma: T1 con constricción posterior, débilmente nodoso y corto, sin fusionarse uniformemente con T2; T1 y S1 con setas blancas braquiplumosas, largas y erectas, excepto ápice de T1 con banda transversa de setas plumosas; T2 y S2 mayormente con setas doradas braquiplumosas y plumosas entremezcladas con unas pocas setas braquiplumosas rojas a casi negras; ápice de T2 a T5 y S2 a S6 con banda transversa, densa, de setas blancas plumosas; T6 sin setas, excepto su borde basal lateral con setas blancas plumosas; T1 con punteado pequeño mediano y esparcido; S1 sin punteado distintivo; T2 y S2 con punteado mediano y separados entre sí; T3 a T5 y S3 a S5 con punteado pequeño y esparcido; S1 sin punteado distintivo, con una gran elevación longitudinal medial y corta; T2 con línea de felpa lateral, S2 sin línea de felpa lateral; T6 con área aplanada, delimitada por carena lateral que alcanza el margen posterior; área pigidial con esculturación granulosa poco perceptible y entremezclada con unas pocas estrías longitudinales cortas.
diagnosis. Macho: Se diferencia de las demás especies de Sphaeropthalma por el siguiente conjunto de caracteres morfológicos: braquíptero; mandíbula con su ápice bidentado, su reborde dorsal basal con una carena elevada y la base del borde ventral con laminilla; notaulo presente, que se extiende desde el margen anterior hasta el margen posterior del mesoescuto; morfología de los parámeros, volsella (cuspis, digitus) y valvas penianas.
Hembra: Se diferencia por el siguiente conjunto de caracteres morfológicos: carena escrobal ausente, mandíbula con su ápice bidentado y el borde ventral basal con dentículo; clípeo con carena transversa arqueada; mesosoma piriforme; pronoto con fina carena curva delimitando su margen posterior; dorso del mesosoma con tres carenas transversas sobre la mitad posterior; T1 con constricción posterior, nodoso y corto; T6 con área pigidial delimitada por carena lateral.
Variación intraespecífica: Machos, longitud corporal de 4,5 a 9,4 mm (Media 7,3 mm, desviación estándar 1,5 mm, n = 9). Hembras, longitud corporal de 4,5 a 5,7 mm (Media 4,9 mm, desviación estándar 0,38 mm, n = 13). Una hembra presenta en

Sphaeropthalma pinedai sp. nov.: Primer registro de un mutílido braquíptero para América del sur... 225
el pronoto una fina carena curva que delimita su margen posterior; otras dos hembras presentan esta carena discontinua y casi ausente. La coloración puede variar ligeramente en machos y hembras. Un macho tiene el metasoma en gran parte rojo, con pocas áreas rojo oscuro. Las hembras pueden presentar las patas totalmente amarillas, el T2 con su mitad basal roja y la mitad distal negra o rojo oscuro. Estas ligeras diferencias en coloración las consideramos variaciones intraespecíficas, lo cual es común en muchas especies de mutílidas (WILSON & PITTS, 2009; BERGAMASCHI et al., 2012).
Sphaeropthalma pinedai. 3. Holotipo macho, habitus, vista dorso-lateral. 4-5. Alotipo hembra, habitus. 4. Vista dorsal. 5. Vista lateral. Escala = 1,0 mm.
figuras 3-5.
notas taxonómicas: Cinco especies de Sphaeropthalma han sido registradas para Chile (NONVEILLER, 1990): S. chilicola Andre, 1904, macho, S. gayi Mickel, 1937, macho, S. tenuiventris (Spinola, 1851), macho, S. testaceipes André, 1898, macho, y S. tetragonodera (Spinola, 1851), hembra, esta última erróneamente mencionada como macho en NONVEILLER (1990: 72).
Hemos examinado (24 febrero de 2003) un espécimen hembra de S. tetragonodera (Spinola), depositado en el Museo Nacional de Historia Natural de París, el cual
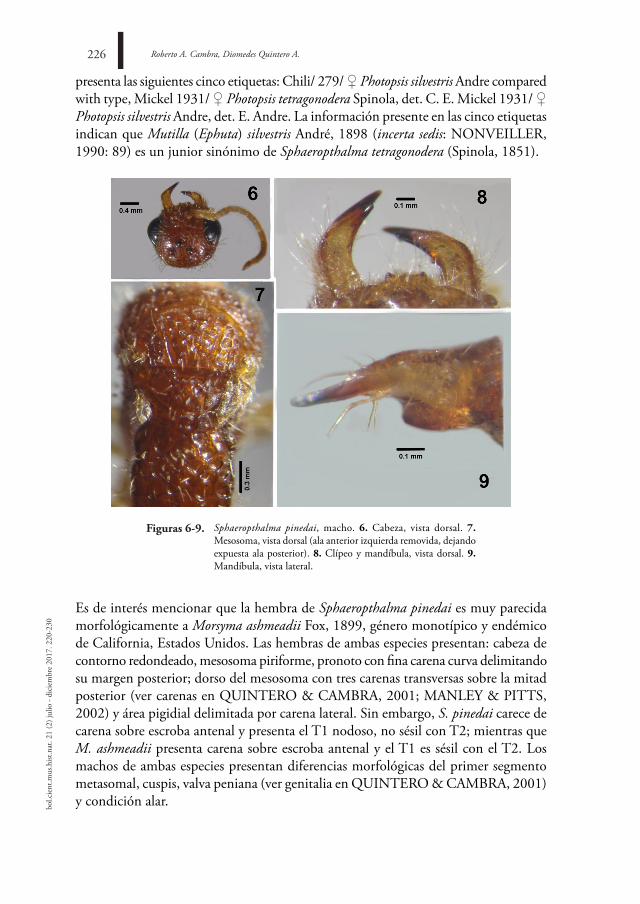
Roberto A. Cambra, Diomedes Quintero A.226bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
20-2
30
presenta las siguientes cinco etiquetas: Chili/ 279/ ♀ Photopsis silvestris Andre compared with type, Mickel 1931/ ♀ Photopsis tetragonodera Spinola, det. C. E. Mickel 1931/ ♀ Photopsis silvestris Andre, det. E. Andre. La información presente en las cinco etiquetas indican que Mutilla (ephuta) silvestris André, 1898 (incerta sedis: NONVEILLER, 1990: 89) es un junior sinónimo de Sphaeropthalma tetragonodera (Spinola, 1851).
Sphaeropthalma pinedai, macho. 6. Cabeza, vista dorsal. 7. Mesosoma, vista dorsal (ala anterior izquierda removida, dejando expuesta ala posterior). 8. Clípeo y mandíbula, vista dorsal. 9. Mandíbula, vista lateral.
figuras 6-9.
Es de interés mencionar que la hembra de Sphaeropthalma pinedai es muy parecida morfológicamente a Morsyma ashmeadii Fox, 1899, género monotípico y endémico de California, Estados Unidos. Las hembras de ambas especies presentan: cabeza de contorno redondeado, mesosoma piriforme, pronoto con fina carena curva delimitando su margen posterior; dorso del mesosoma con tres carenas transversas sobre la mitad posterior (ver carenas en QUINTERO & CAMBRA, 2001; MANLEY & PITTS, 2002) y área pigidial delimitada por carena lateral. Sin embargo, S. pinedai carece de carena sobre escroba antenal y presenta el T1 nodoso, no sésil con T2; mientras que M. ashmeadii presenta carena sobre escroba antenal y el T1 es sésil con el T2. Los machos de ambas especies presentan diferencias morfológicas del primer segmento metasomal, cuspis, valva peniana (ver genitalia en QUINTERO & CAMBRA, 2001) y condición alar.
Sphaeropthalma pinedai sp. nov.: Primer registro de un mutílido braquíptero para América del sur... 227
Sphaeropthalma pinedai, macho. 10. Hipopigio (S7). 11. Valva peniana completa, vista lateral, cara externa. 12. Valva peniana, área distal mostrando detalles de proyecciones y setas, ea = espina aguda apical, pd = proyección digitiforme subapical.
figuras 10-12.
etimología: Especie dedicada a Cristian Pineda, recolector del holotipo.
Material examinado (9 ♂, 13 ♀): Holotipo ♂: Chile, Provincia Valparaíso, Comuna de Quintero, Ritoque, 32°48’55,8”S 71°31’57,1”W, 27-29 nov. 2015, col. C. Pineda. Alotipo hembra y paratipos: mismos datos de recolecta que el holotipo, 8 ♂, 11 ♀. Chile, Provincia Huasco: Llano del Algarrobal, 1 nov. 1991, col. J. E. Barriga, 1 ♀; Vista Alegre, 6 nov. 1991, col. J. E. Barriga, 1 ♀.
Los especímenes machos presentan las alas extendidas, la cutícula y genitalia están totalmente endurecidas y desarrolladas, indicando que no son tenereos.
descripción del hábitat: Los especímenes de Ritoque fueron recolectados en una terraza costera, que se encuentra a 1,0 km de distancia del mar. La vegetación es muy escasa, con herbazal bajo y con la presencia predominante de arbustos de Baccharis linearis (Asteraceae), conocida comúnmente como “romerillo” (Fig. 2). Esta planta se encuentra en terrenos áridos y degradados. Aunque no se observaron especímenes de S. pinedai alimentándose de esta planta, existe la posibilidad de que ella sea fuente alimenticia de la avispa mutílida.
Roberto A. Cambra, Diomedes Quintero A.228bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
20-2
30
Sphaeropthalma pinedai, genitalia, macho. 13. Vista dorso lateral. 14. Vista ventral. 15. Paramere, vista lateral, cara externa. 16. Paramere = p, cuspis = c, digitus = d, vista lateral, cara interna. Escala = 0,2 mm.
figuras 13-16.
Comentarios: Sphaeropthalma pinedai es la primera especie braquíptera registrada para América del Sur. Mutílidos machos braquípteros o ápteros son una condición extremadamente rara. En América, la familia Mutillidae está constituida por dos subfamilias: 1) Sphaeropthalminae, conformada por 55 géneros (LELEJ & BROTHERS, 2008; WILLIAMS & PITTS, 2009; WILLIAMS et al., 2011; BARTHOLOMAY et al., 2015), de los cuales solo siete géneros tienen machos con representantes braquípteros o ápteros (Tabla 1). 2) Mutillinae, representada en América por cinco géneros (BROTHERS, 2006), ninguno de ellos con machos braquípteros o ápteros. La Myrmosidae, de distribución holártica, era considerada una subfamilia de Mutillidae, pero análisis moleculares y morfológicos la ubican como una familia aparte (PILGRIM et al., 2008). Todas las especies de Mutillidae en América, con representantes machos braquípteros o ápteros, habitan en zonas desérticas o relativamente áridas de solo tres países: Estados Unidos, México y Chile (Tabla 1).

Sphaeropthalma pinedai sp. nov.: Primer registro de un mutílido braquíptero para América del sur... 229
tabla 1. Especies de Mutillidae en América con machos braquípteros o ápteros
especie Condición alar País referenciaChilemutilla aptera Cambra & Quintero (2007)
áptero Chile CAMBRA & QUINTERO (2007)
Dasymutilla hector (Blake, 1871)
braquíptero o alado USA MICKEL (1936)
Dasymutilla waco (Blake, 1871)
braquíptero o alado USA MICKEL (1936)
Limaytilla sp. áptero Chile CAMBRA & QUINTERO (2007)
Myrmilloides grandiceps (Blake, 1872)
braquíptero USA BLAKE (1872), SCHUSTER (1945), MANLEY & PITTS (2002)
Morsyma ashmeadii fox, 1899
áptero o alado USA FOX (1899), KROMBEIN (1940) , QUINTERO & CAMBRA (2001), PITTS (2007)
Sphaeropthalma brachyptera schuster, 1945
braquíptero o alado U S A , México
SCHUSTER (1945)
Sphaeropthalma arnalduri Pitts, 2010
áptero PITTS et al. (2010)
Sphaeropthalma pinedai Cambra & Quintero
braquíptero Chile Presente trabajo
Stethophotopsis maculata Pitts, 2000
braquíptero USA PITTS & MCHUGH (2000)
aGradeCiMientos
A Cristian Pineda por la donación de los especímenes, fotografías e información sobre características biológicas del hábitat donde se capturaron los especímenes, y a Kevin A. Williams, Department of Food & Agriculture, California, por los comentarios y sugerencias para mejorar este trabajo.
referenCias
BARTHOLOMAY, P.R., WILLIAMS, K.A., LUZ, D.R. & MORATO, E.F., 2015.- Frigitilla gen. nov., a new genus of Amazonian Mutillidae (Hymenoptera). Zootaxa, 3957: 49-58.
BERGAMASCHI, A.C.B., CAMBRA, R.A., BROTHERS, D.J. & MELO, G.A.R., 2012.- lynchiatilla Casal, 1963 (Hymenoptera: Mutillidae): a new species from Brazil associated with Paroxystoglossa spiloptera Moure (Hymenoptera: Apidae: Halictinae), and notes on other species. Zootaxa, 3548: 55-64.
BLAKE, C.A., 1872.- Additions to the “Synopsis of North America Mutillidae”. Transactions of the American entomological Society, 4: 71-76.
BROTHERS, D.J., 2006.- Mutillidae: 586-594 (en) HANSON, P. & GAULD, I. (eds.) Hymenoptera de la Región Tropical. Memoirs of the American entomological Institute, 77: 1-994.
CAMBRA, R.A. & QUINTERO A., D., 2007.- Chilemutilla, a new genus of Mutillidae (Hymenoptera) from Chile, and the description of the first wingless mutillid male from South America. Transactions of the American entomological Society, 133 (1): 167-180.
FOX, W.J., 1899.- The North American Mutillidae. Transactions of the American entomological Society, 25: 219-300.KROMBEIN, K.V., 1940.- Studies in the Tiphiidae (Hymenoptera Aculeata). IV. A Revision of the Myrmosine of the New World
with a discussion of the Old World species. Transactions of the American entomological Society, 65: 415-465.

Roberto A. Cambra, Diomedes Quintero A.230bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
20-2
30
LELEJ, A. & BROTHERS, D.J., 2008.- The genus-group names of Mutillidae (Hymenoptera) and their type species, with a new genus, new name, new synonymies, new combinations and lectotypifications. Zootaxa, 1889: 1-79.
MANLEY, D.G. & PITTS, J.P., 2002.- A key to genera and subgenera of Mutillidae (Hymenoptera) in America North of Mexico with description of a new genus. Journal of Hymenoptera Research, 11 (1): 72-100.
MICKEL, C.E., 1936.- New species and records of Nearctic mutillid wasps of the genus Dasymutilla (Hymenoptera). Annals of the entomological Society of America, 29 (1): 29-60.
NONVEILLER, G., 1990.- Catalogue of the Mutillidae, Myrmosidae and Bradynobaenidae of the Neotropical Region including Mexico: 1-149 (en) VAN ACHTERBERG, C. (ed.) Hymenopterorum Catalogus (nova editio). SPB Academic Publishing, The Hague (The Netherlands).
PILGRIM, E.M., VON DOHLEN, C.D. & PITTS, J.P., 2008.- Molecular phylogenetics of Vespoidea indicate paraphyly of the superfamily and novel relationships of its component families and subfamilies. Zoologica Scripta, 37: 539-560.
PITTS, J.P., 2007.- Description of the female of Sphaeropthalma galapagensis Williams and notes on Morsyma Fox and Caenotilla Pitts and Manley (Hymenoptera: Mutillidae). The Pan-Pacific entomologist, 83 (2): 95-100.
PITTS, J.P. & MCHUGH, J.V., 2000.- Stethophotopsis, a new genus of Sphaeropthalmini (Mutillidae: Sphaeropthalminae) with a brachypterous male from Arizona. Journal of Hymenoptera Research, 9 (1): 29-33.
PITTS, J.P., WILSON, J.S., WILLIAMS, K.A. & BOEHME, N.F., 2010.- Nocturnal velvet ant males (Hymenoptera: Mutillidae) of Deep Canyon, California including four new species and a fifth new species from Owens Lake Valley, California. Zootaxa, 2553: 1-34.
QUINTERO, D. & CAMBRA, R.A., 2001.- On the endemic genus Morsyma Fox (Hymenoptera: Mutillidae: Sphaeropthalminae), with a description of the previously unknown female. Transactions of the American entomological Society, 127 (4): 451-458.
SCHUSTER, R.M., 1945.- An interesting new brachypterous species of Photopsis (Hymenoptera, Mutillidae). Pan-Pacific entomologist, 21 (4): 149-151.
WILLIAMS, K.A. & PITTS, J.P., 2009.- Eight new species of lomachaeta Mickel and the synonymy of Smicromutilla Mickel (Hymenoptera: Mutillidae). Journal of Hymenoptera Research, 18 (2): 227-243.
WILLIAMS, K.V., BROTHERS, D.J. & PITTS, J.P., 2011.- New species of Tobantilla Casal, 1965 and a new genus and species, Gogoltilla chichikovi gen. et sp. nov., from Argentina (Hymenoptera: Mutillidae). Zootaxa, 3064: 41-68.
WILSON, J.S. & PITTS, J.P., 2009.- Species boundaries of Sphaeropthalma unicolor (Hymenoptera: Mutillidae): is color useful for differentiating species? Journal of Hymenoptera Research, 18 (2): 212-226.

Boletín CientífiCoCentro de Museos
Museo de Historia natural
diVersidad de esCorPiones (CHeliCerata) en el dePartaMento del atlÁntiCo, ColoMBia*
eider Sará-C.1, eduardo Flórez-D.2, Neis Martínez-H.3
resumen
Se determinó la variación de la diversidad de escorpiones en siete fragmentos de Bosque Seco Tropical (BST) en el departamento del Atlántico. Los métodos de captura utilizados fueron cernido de hojarasca, agitación de follaje y captura manual nocturna con luz UV. En cada transecto se realizó un esfuerzo de dos noches y tres días. Se recolectaron 516 ejemplares representados en siete especies, seis géneros y tres familias (Buthidae, Chactidae y Hormuridae). Buthidae fue la más diversa y abundante con cuatro géneros, cinco especies y 486 individuos, destacándose la especie Tityus (Archaeotityus) tayrona cuya presencia fue constante en todos los fragmentos. El fragmento de bosque de la Reserva Campesina La Sierra (R.C.S) presentó la mayor diversidad con seis especies, seis géneros y tres familias, lo cual se debe posiblemente a la oferta de recursos y microhábitats. Con los resultados se logra ampliar el área de distribución de las especies T. (Atreus) aff. asthenes y Chactas brevicaudatus, además se encontró que en el departamento está representado el 60% de las familias y el 42,8% de los géneros registrados para Colombia, lo que indica que los relictos de BST en este departamento mantienen una fauna importante de los escorpiones registrados para Colombia.
Palabras clave: Buthidae, diversidad, Hormuridae, Tityus (Archaeotityus) tayrona.
diVersitY of sCorPions (CHeliCerata) in tHe dePartMent of atlÁntiCo, ColoMBia
abstract
The variation of the diversity of scorpions in seven fragments of Tropical Dry Forest (TDF) in the Department of Atlántico was determined. The sampling was conducted by fallen leaves hoeing, foliage agitation and manual capture at night with UV light. An effort of two nights and three days was made in each transect. A total of 516 specimens representing seven species,
* FR: 1-V-2017 . FA: .10-VIII-20171 Biólogo. Semillero de Investigación de Artrópodos ¨NEOPTERA¨ del Caribe colombiano, Grupo de Investigación Biodiversidad del Caribe colombiano, Universidad del Atlántico. Barranquilla, Colombia. E-mail: [email protected]: 0000-0001-5555-46902 Doctor en Ciencias- Biología. Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. E-mail: [email protected] ORCID: 0000-0001-1724-23043 Estudiante de Doctorado en Ciencias-Biología. Grupo de Investigación Biodiversidad del Caribe colombiano, Departamento de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia. E-mail: [email protected]
bol.cient.mus.hist.nat. 21 (2), julio-diciembre, 2017. 231-244. ISSN: 0123-3068 (Impreso) ISSN: 2462-8190 (En línea)
CÓMO CITAR: SARÁ-C., E., FLÓREZ-D., E. & MARTÍNEZ-H., N., 2017.- Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 21 (2): 231-244. DOI: 10.17151/bccm.2017.21.2.16

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.232bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
six genera and three families (Buthidae, Chactidae and Hormuridae) were collected. The most diverse and abundant family was Buthidae with four genera and 486 individuals, standing out the species Tityus (Archaeotityus) tayrona whose presence was constant in the seven fragments. The fragment of forest Reserva Campesina La Sierra (R.C.S) presented the greatest diversity with six species, six genera and three families, which is possibly due to the abundance of resources and microhabitats. With the results, the area of distribution of the species Tityus (Atreus) aff. asthenes and Chactas brevicaudatus expanded. In addition, it was found that 60% of the families and 42.8% of the genera reported for Colombia are represented in the Department of Atlántico which indicates that the relics of TDF in this Department maintain a significant fauna of the scorpions registered for the country.
Key words: Buthidae, diversity, Hormuridae, Tityus (Archaeotityus) tayrona.
introduCCiÓn
La fragmentación del Bosque Seco Tropical (BST) pone en grave riesgo a la biodiversidad asociada y por lo tanto, a los procesos ecológicos que aseguran la funcionalidad de este ecosistema (SÁNCHEZ-AZOFEIFA et al., 2005; PIZANO & GARCÍA, 2014). Aunque el BST se encuentra en un estado crítico de fragmentación y degradación, con un porcentaje muy pequeño de representatividad dentro de las áreas del Sistema Nacional de Áreas Protegidas en Colombia; es uno de los ecosistemas más amenazados y menos estudiados (PIZANO & GARCÍA, 2014). En la actualidad, este ecosistema en la región Caribe colombiana está representado por fragmentos inmersos en una matriz compuesta por cultivos, potreros y cercas vivas (RANGEL-ACOSTA & MARTÍNEZ-HERNÁNDEZ, 2017), lo cual muy probablemente genera variaciones en la estructura de la vegetación, composición faunística y disponibilidad de recursos. Estas alteraciones en el BST traen como consecuencia pérdida de especies residentes y/o colonización de nuevos hábitats, dentro de los cuales están los asentamientos humanos; que en el caso de los escorpiones se podrían convertir en un problema de salud pública (GÓMEZ & OTERO, 2007).
En Colombia se han realizado diversos estudios a los escorpiones (FLÓREZ, 1990, 2001; LOURENÇO & FLÓREZ, 1989, 1990; LOURENÇO, 1991, 1992, 1994a, 1994b, 1997, 2002; FLÓREZ & SÁNCHEZ, 1995; BOTERO-TRUJILLO & FAGUA, 2007; BOTERO-TRUJILLO, 2008; TERUEL & GARCÍA, 2008a, 2008b; TERUEL & RONCALLO, 2008, 2010; BOTERO-TRUJILLO & FLÓREZ, 2011; ARMAS et al., 2012), en donde la mayoría de los trabajos centran su esfuerzo en conocer la distribución y riqueza de estos arácnidos. Hace poco se empezó a abordar este tema en la costa Caribe colombiana, destacándose el estudio realizado por GÓMEZ et al. (2013), donde se describe la composición de la escorpiofauna del departamento de Sucre; mientras que BRAVO & RODIÑO (2013) analizaron la variación de la

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 233
escorpiofauna para algunos parches de BST en el departamento de Córdoba y SARÁ-CASTILLO & MARTÍNEZ-HERNÁNDEZ (2017) realizaron la distribución y taxonomía de los escorpiones del departamento del Atlántico. Adicionalmente, BOTERO-TRUJILLO (2008) registró por primera vez algunos bútidos para el departamento del Atlántico y LOURENÇO (1991) hizo referencia a la composición de la escorpiofauna de la Sierra Nevada de Santa Marta (SNSM) y algunos departamentos aledaños a ella (LOURENÇO & FLÓREZ, 1990; LOURENÇO, 1997).
Sin embargo, esos trabajos son insuficientes para generar una información completa sobre el estado actual de la biodiversidad de este orden en el BST, más específicamente en el departamento del Atlántico; donde las expansiones de las fronteras ganaderas y agrícolas han provocado la reducción de este ecosistema a pequeños fragmentos inmersos en matrices de uso ganadero y agrícola.
Teniendo en cuenta lo anterior y con el fin de contribuir al conocimiento de la biodiversidad del orden Scorpiones en el país y en este tipo de ecosistema, así como el papel que juegan los fragmentos de BST en el mantenimiento y conservación de este grupo de arácnidos, se analizó la variación espacial de la diversidad en este departamento. La información generada en este trabajo será útil para implementar planes de manejo y conservación de los fragmentos de BST y futuras investigaciones sobre este grupo de arácnidos.
Materiales Y MÉtodos
Área de estudio
El presente estudio se llevó a cabo en siete fragmentos de BST (R.C.M, C.S.L, R.C.S, D.M.I.L, R.B, C.U.A y Piojó) en el departamento del Atlántico (Figura 1, Tabla 1). Escogidos porque presentaron relictos pequeños con vegetación típica de bosque seco, donde predominan las familias Fabaceae, Malvaceae, Cactaceae, entre otras. Con especies arbóreas como Bursera simarouba (indio encuero), Spondia mombin (hobo), Ficus elastica (caucho), Anacardium excelsum (carcolí), Sterculia apetala (camajón), Cedrela odorata (cedro), Coccoloba uvifera (uvito), Gliricidia sepium (matarratón). Además, se tuvieron otros criterios como accesibilidad y localización en el departamento.
En general, esta región presenta dos periodos climáticos. La mayor concentración de lluvias se presenta en mayo y octubre, y la menor en enero, febrero y marzo, con un valor promedio anual de 990 m. La mayor concentración de lluvias se presenta en mayo y octubre, y la menor en enero, febrero y marzo (RANGEL-CH. & CARVAJAL-COGOLLO, 2012).
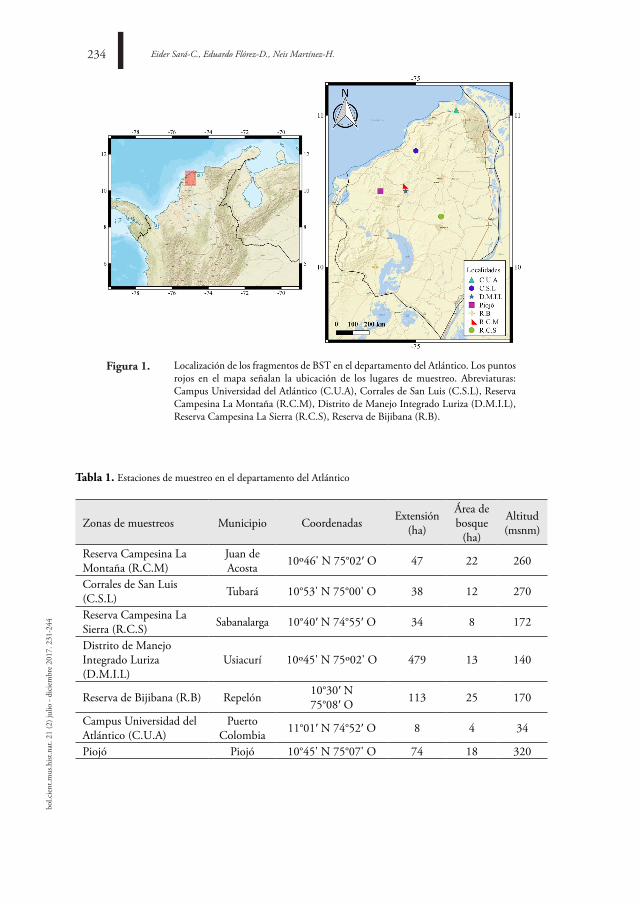
eider Sará-C., eduardo Flórez-D., Neis Martínez-H.234bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
Localización de los fragmentos de BST en el departamento del Atlántico. Los puntos rojos en el mapa señalan la ubicación de los lugares de muestreo. Abreviaturas: Campus Universidad del Atlántico (C.U.A), Corrales de San Luis (C.S.L), Reserva Campesina La Montaña (R.C.M), Distrito de Manejo Integrado Luriza (D.M.I.L), Reserva Campesina La Sierra (R.C.S), Reserva de Bijibana (R.B).
figura 1.
tabla 1. Estaciones de muestreo en el departamento del Atlántico
Zonas de muestreos Municipio Coordenadas Extensión(ha)
Área de bosque
(ha)
Altitud(msnm)
Reserva Campesina La Montaña (R.C.M)
Juan de Acosta 10º46' N 75°02′ O 47 22 260
Corrales de San Luis(C.S.L) Tubará 10°53' N 75°00' O 38 12 270
Reserva Campesina La Sierra (R.C.S) Sabanalarga 10°40′ N 74°55′ O 34 8 172
Distrito de Manejo Integrado Luriza (D.M.I.L)
Usiacurí 10º45' N 75º02' O 479 13 140
Reserva de Bijibana (R.B) Repelón 10°30′ N75°08′ O 113 25 170
Campus Universidad del Atlántico (C.U.A)
Puerto Colombia 11°01′ N 74°52′ O 8 4 34
Piojó Piojó 10°45' N 75°07' O 74 18 320

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 235
Métodos y diseño de muestreo
El trabajo de campo se realizó desde enero de 2012 hasta julio de 2013. Por cada fragmento se realizaron dos muestreos, uno en época seca y otro durante las lluvias. La inversión de tiempo por muestreo fue de dos noches y tres días en cada fragmento. La unidad muestral consistió en un transecto de 500 m de largo por 50 m de ancho, donde se marcaron 10 puntos distanciados 50 m aproximadamente. En cada punto se delimitó un diámetro de 40 m para realizar los siguientes métodos de colecta directa:
Captura Manual con Luz UV (C.M.UV): consiste en recolectar con ayuda de pinzas específicas para escorpiones, buscando debajo de las piedras, ranuras, madrigueras, hojarasca, troncos, con una inversión de 15 minutos por punto para un total de 150 minutos/noche. Esta actividad se realizó entre las 7 p.m. y las 10 p.m.
Agitación de Follaje (A.F): se utilizó una red entomológica con un diámetro de 50 cm en tela y un mango de 50 cm. La red se colocó debajo de ramas secas, arbustos, nidos y troncos y posteriormente se realizaron golpes con un madero con el fin de que cayeran los diferentes especímenes con hábitos arborícolas. Para esto se realizaron tres sesiones, cada una de 20 golpes para un total de 60 golpes por punto.
Cernido de Hojarasca (C.H): consistió en recolectar en un recipiente plástico 10 L de hojarasca que fueron cernidos en un tamizador Winkler con diámetro de 16 mm, devolviendo la hojarasca sobrante al bosque. En el campo, el material cernido fue revisado en bandejas plásticas de color blanco con el fin de capturar especímenes con ayuda de pinzas.
Preservación e identificación de los especímenes
Las muestras se preservaron en alcohol al 70% y se trasladaron hasta las instalaciones del Laboratorio de Zoología de Invertebrados, ubicado en la Universidad del Atlántico. Los individuos recolectados en las expediciones se identificaron hasta especie con la ayuda de las claves propuestas por LOURENÇO (1998) y FLÓREZ (2001). Las identificaciones se confirmaron haciendo revisiones del material depositado en la colección de escorpiones del Instituto de Ciencias Naturales (ICN) de la Universidad Nacional de Colombia, sede Bogotá, y depositados en el Museo de Zoología de la Universidad del Atlántico y la Colección de Aracnología del ICN.
análisis de los datos
Se determinó el número de especies por técnica y fragmentos. Para la estimación de la cobertura del muestreo por fragmento se utilizó el estimador de cobertura recomendado por CHAO & JOST (2012):

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.236bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
Donde f1 y f2 son el número de especies con uno y dos individuos en la muestra respectivamente, y n es el número de individuos. La cobertura del muestro varía entre 1-100% cuando este toma valores de ≈ 100%, con respecto a la técnica de captura.
La composición fue obtenida mediante la determinación taxonómica de las especies de escorpiones, y para la riqueza y diversidad de escorpiones entre los fragmentos de BST se utilizó un análisis de Interpolación y Extrapolación propuesto por CHAO et al. (2014), utilizando los números de Hill (HILL, 1973) en unidades del número efectivo de especies siguiendo lo propuesto por JOST (2006). Se calcularon los valores de diversidad verdadera: 0D (riqueza especies), 1D (el exponencial del índice de Shannon) y 2D (el inverso del índice de Simpson) (JOST, 2006). El análisis fue realizado con el script iNeXT para R Studios (CHAO et al., 2014) y ejecutado en el programa R versión 3.1.1 (R DEVELOPMENT CORE TEAM, 2014).
Teniendo en cuenta la composición de especies y su respectiva abundancia por fragmentos, se realizó un análisis de agrupamiento por pares mediante el método de promedios no ponderados (UPGMA), realizándose el agrupamiento por fragmento por medio del índice de similitud de Jaccard (CHAO et al., 2014). Esto con el fin de observar porcentajes de similitud que existen entre las estaciones de muestreo en términos de composición de especies. Los análisis se realizaron con el programa PRIMER 6.0 (CLARKE & WARWICK, 2001).
resultados
Composición de especies
Se capturaron 516 individuos entre adultos y juveniles, distribuidos en siete especies, seis géneros y tres familias. La familia con mayor riqueza y abundancia fue Buthidae con cinco especies (71,43%) y 486 individuos. La familia que siguió en abundancia fue Hormuridae con 28 individuos, seguido de Chactidae con dos individuos (0,39%) (Tabla 2).
Tityus (Archaeotityus) tayrona Lourenço presentó una alta dominancia con 437 individuos (84,69%), mientras que Rhopalurus laticauda Thorell, Centruroides edwardsii (Gervais) y Chactas brevicaudatus (Karsch) fueron las menos frecuentes con 5, 4 y 2 individuos respectivamente y solo fueron registradas en el fragmento R.C.S (Tabla 2, Figura 2). En el caso de Ananteris columbiana Lourenço, presentó un bajo número de capturas (26 individuos), pero se encontró en seis de las siete localidades de muestreo (Tabla 2).

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 237
Especies comunes y poco frecuentes en los fragmentos de BST en el departamento del Atlántico. a: T. (Archaeotityus) tayrona. B: R. laticauda. C: C. edwardsii, d: T. (Atreus) aff. Asthenes.
figura 2.
tabla 2. Variación de la abundancia y diversidad de escorpiones por fragmentos de BST en el departamento del Atlántico
familia especies r.C.M C.s.l r.C.s d.M.i.l r.B C.u.a Piojó total
Buthidae Ananteris columbiana 8 1 3 2 9 3 0 26
Centruroides edwardsii 0 0 4 0 0 0 0 4
Rhopalurus laticauda 0 0 5 0 0 0 0 5
Tityus (Atreus) aff. asthenes 0 5 0 0 7 0 2 14
Tityus (Archaeotityus) tayrona 112 40 44 39 137 22 43 437
Chactidae Chactas brevicaudatus 0 0 2 0 0 0 0 2
Hormuridae opisthacanthus elatus 5 0 6 14 0 0 3 28
Número de individuos 125 46 64 55 153 25 48 516
Diversidad de orden 0 (0D) 3 3 6 3 3 2 3 7
Diversidad de orden 1 (1D) 1,50 1,56 3,02 2,04 1,50 1,44 1,50
Diversidad de orden 2 (2D) 1,24 1,30 2,02 1,76 1,24 1,27 1,24
Las abreviaturas se describen en la Tabla 1.

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.238bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
riqueza y abundancia por técnica de captura
La cobertura del muestreo fue alta para cada uno de los fragmentos estudiados (100%) para los tres órdenes (0D, 1D, 2D) de la diversidad analizados, lo que demuestra que el esfuerzo y los métodos de captura utilizados permitieron obtener una buena estimación de la diversidad de cada uno de los sitios. Se observó que la técnica más eficiente fue captura manual con luz UV, método que permitió capturar todas las especies registradas y 407 individuos (Tabla 3). Por otro lado, con las técnicas agitación de follaje y cernido de hojarasca (A.F y C.H), se lograron recolectar 109 individuos (65 y 44, respectivamente) distribuidos en dos especies (T. tayrona y A. columbiana) (Tabla 3).
tabla 3. Riqueza y abundancia de escorpiones por técnica de captura en el área de estudio
Especies/Técnicas C.M.uV a.f C.HAnanteris columbiana 16 5 5Centruroides edwardsii 4Chactas brevicaudatus 2opisthacanthus elatus 28Rhopalurus laticauda 5Tityus (Atreus) aff. asthenes 14Tityus (Archaeotityus) tayrona 338 60 39Número de especies 7 2 2Número de individuos 414 65 44
Abreviaturas: Captura Manual con Luz UV (C.M.UV), Agitación de Follaje (A.F) y Cernido de Hojarasca (C.H).
Variación de la riqueza y diversidad por fragmento
Con el análisis de Interpolación y Extrapolación se determinaron cambios a nivel de fragmentos en cuanto a la riqueza y diversidad de escorpiones (Tabla 2). Con relación a la riqueza (0D), el mayor valor (6 especies) se observó en R.C.S y el menor (2 especies) en C.U.A. Estos mismos fragmentos también presentaron los mismos resultados para la diversidad 1D. En el caso de la diversidad 2D, el menor valor (1,24 especies efectivas) se registró en la R.C.M y el mayor (2,02 especies efectivas) en la R.C.S (Tabla 2).
agrupación entre las estaciones de muestreos
Teniendo en cuenta el análisis de similitud con el índice Jaccard (CHAO et al., 2014), se formaron tres grupos (Figura 3). El primero conformado por el fragmento de la R.C.M y R.B con una similitud del 87%; el segundo conformado por los fragmentos R.C.S, D.M.I.L, C.S.L y Piojó, con una similitud del 78%. El tercer grupo conformado

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 239
por el fragmento C.U.A, el cual presentó una similitud del 60% con el resto de las estaciones de muestreo.
Agrupamiento de las localidades de muestreos utilizando el índice de similitud de Jaccard. Abreviaturas: Campus Universidad del Atlántico (C.U.A), Corrales de San Luis (C.S.L), Distrito de Manejo Integrado Luriza (D.M.I.L), Reserva Campesina La Sierra (R.C.S), Reserva Campesina La Montaña (R.C.M), Reserva de Bijibana (R.B).
figura 3.
disCusiÓn
Los datos obtenidos muestran resultados similares a otros estudios realizados en la región Caribe y otras regiones del país (AGUDELO, 2004; FLÓREZ, 2004; BRAVO & RODIÑO, 2013; GÓMEZ et al., 2013). Se reportan las especies con importancia médica T. (Atreus) aff. asthenes y C. edwardsii (GÓMEZ & OTERO, 2007; CUPITRA-VERGARA et al., 2015). Aunque ya se han realizado registros de especies, principalmente de la familia Buthidae (BOTERO-TRUJILLO, 2008), el presente estudio estableció los primeros datos de riqueza y distribución de escorpiones en fragmentos de BST en el departamento del Atlántico y se confirma la presencia de las especies A. columbiana, C. edwardsii, R. laticauda, T. (Archaeotityus) tayrona, y se amplió el rango de distribución de Ch. brevicaudatus, T. (Atreus) aff. asthenes y o. elatus.
Por otro lado, se observó un mayor número en la diversidad de géneros y especies de escorpiones en los fragmentos de BST en el departamento del Atlántico, en comparación con otros inventarios del orden realizados en la región Caribe (BRAVO & RODIÑO, 2013; GÓMEZ et al., 2013), debido a un mayor esfuerzo y cobertura del área de muestreo; así como la focalización de las faenas de campo en las dos épocas

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.240bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
(seca y lluvias) en un ecosistema que presenta estacionalidad marcada de acuerdo a los niveles de precipitación.
La alta diversidad de Buthidae se puede atribuir a que esta familia es la de mayor distribución en Colombia y la más diversa en este grupo de arácnidos (FLÓREZ, 2001). La alta dominancia de T. (Archaeotityus) tayrona se debe posiblemente a la preferencia de la especie por los hábitats secos y se ha adaptado a un modo de vida con cierto grado de tolerancia a ecosistemas que se encuentran muy perturbados y deteriorados (FLÓREZ, 2004). Por otro lado, autores como GÓMEZ & OTERO (2007) sugieren que T. (Archaeotityus) tayrona podría estar dentro de las especies de la familia Buthidae que necesitan hábitats, microhábitats y ambientes específicos, pero los resultados obtenidos en este estudio sugieren que esta especie tolera ambientes modificados con alto grado de intervención antrópica, tal como los fragmentos C.U.A y C.S.L.
Para realizar un juicio sobre el bajo número de capturas del resto de las especies se hace necesario realizar un estudio mucho más detallado sobre la disponibilidad de recursos y la adaptabilidad de estas especies en los fragmentos de BST; o si su baja abundancia es un efecto de las técnicas de captura y el muestreo. Para esto último, se requiere concentrar la intensidad y cantidad de muestreos para aumentar la probabilidad de capturas de estas especies. Por otro lado, la baja abundancia R. laticauda y C. edwardsii en este tipo de fragmentos, se puede atribuir a que son especies que no se adaptan con facilidad a ambientes intervenidos como lo registran MANZILLA & DE SOUSA (2003) y CUPITRA-VERGARA et al. (2015).
En el caso de Ch. brevicaudatus había sido registrada como endémica de la SNSM (LOURENÇO, 1991, 1997), sin embargo FLÓREZ (2013) determinó su rango de distribución para los departamentos de Bolívar y Magdalena, y con este estudio se amplía su distribución para el departamento del Atlántico. Este mismo autor menciona que el hábitat natural es el bosque seco xerofítico o sub-xerofítico, extendiéndose desde las zonas litorales en los departamentos anteriormente mencionados, hasta zonas más interiores como el norte de Antioquia (Caucasia) (FLÓREZ, 2013).
Recientemente, BOHÓRQUEZ-GÓMEZ (2016) realizó la revisión de los Tityus del subgénero Atreus para Colombia y no fue posible dilucidar la identidad de Tityus aff. asthenes, debido a que no se pudo revisar el material tipo. Los caracteres morfológicos de los especímenes capturados en este trabajo concuerdan en su gran mayoría con la diagnosis que propone BOHÓRQUEZ-GÓMEZ (2016) para T. aff. asthenes. Se registró para los departamentos de Antioquia, Cauca, Chocó, Nariño y Valle del Cauca en un rango altitudinal entre 1-440 m (BOHÓRQUEZ-GÓMEZ, 2016); ampliándose su distribución para el departamento del Atlántico.

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 241
El fragmento de bosque localizado en la Reserva Campesina La Sierra (R.C.S) presentó la mayor riqueza de escorpiones, de las cuales tres solo fueron exclusivas para este lugar (R. laticauda, C. edwardsii y Ch. brevicaudatus). Esto se atribuye posiblemente a que en este parche de bosque se observó una quebrada que pasa la mayor parte del tiempo con un pequeño cauce de agua, lo que podría brindar una mayor disponibilidad de microhábitats y condiciones microclimáticas de temperatura y humedad optimas que pueden incidir en la composición de escorpiones y la disponibilidad de presas. Adicional a lo anterior, es probable que en este lugar se presente un efecto refugio por la pérdida de conectividad con otros fragmentos de bosque y por la reducción del área por el incremento de las actividades antrópicas en la zona; pero con los resultados obtenidos es difícil demostrar esta hipótesis. La baja riqueza específica y genérica de la estación C.U.A, se debe a que solo se registraron las especies A. columbiana y T. (Archaeotityus) tayrona, predominando notablemente esta última. Esto se debe a que esta estación, aunque tienes zonas con vegetación nativa de BST, presenta una alta intervención antrópica; donde es muy frecuente la construcción de caminos carreteables, zonas de viveros, centros educativos y urbanizaciones, así como la expansión de cementerios adyacentes al fragmento.
El estimador de cobertura (C ̂n) para los fragmentos de BST en el departamento del Atlántico presentó una elevada estimación de la riqueza, demostrándose que el esfuerzo y la variabilidad de técnicas fueron efectivas para determinar la diversidad de escorpiones en la zona de estudio. El éxito de las lámparas UV en el estudio de escorpiones es ampliamente conocido, siendo el método más usado en la captura de estos arácnidos, razón por la cual se esperaba que C.M.UV fuera la técnica más eficiente en el presente estudio. La técnica de Agitación de Follaje (A.F) se implementó por primera vez para el estudio de escorpiones y demostró ser una técnica complementaria para la captura de estos artrópodos en BST. Esta se empleó debido a que permite revisar arbustos, enredaderas y palizadas de difícil acceso en el BST; donde en muestreos previos se observó con gran frecuencia la presencia de escorpiones en estos microhábitats. Aunque C.H fue la técnica de la que se obtuvo menor número de capturas, demostró ser una técnica eficiente debido a que permite la captura de individuos que se refugian en el suelo, mantillo y la hojarasca. Por último, es importante resaltar que para un aumento en el porcentaje de capturas de escorpiones se debe implementar este tipo de técnicas en próximos estudios en el bosque seco, debido a la complementariedad entre cada una de ellas.
A pesar de que la captura de escorpiones con trampas de caída es ampliamente conocida y utilizada, en el presente estudio no se incluyó debido a que en muestreos previos se registraron resultados pobres en número de individuos capturados. Posiblemente lo accidentado del terreno en la mayoría de la estaciones sea una de las causas de estos resultados, pero es una técnica que ha arrojado resultados positivos en otros lugares y podría ser utilizada en futuros estudios en el área.

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.242bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
La alta similitud (60 y 87%) en la composición de especies entre las estaciones de muestreo, se debe a que presentan fauna de escorpiones muy parecida; lo cual se puede atribuir a que presentan un mismo origen geológico y en algún momento estuvieron conectados o hicieron parte de un solo bosque. En el caso de T. (Archaeotityus) tayrona, o. elatus y A. columbiana que son las especies mayormente compartidas entre las estaciones, son especies comunes en ambientes intervenidos (FLÓREZ, 2004) como los fragmentos de BST en el departamento del Atlántico.
La importancia de conocer la distribución de los escorpiones radica en la capacidad de adaptación de estos y el riesgo clínico que pueden representar (GÓMEZ & OTERO, 2007; CUPITRA-VERGARA et al., 2015). Cuando convergen alto grado de toxicidad del veneno y alto grado de adaptabilidad a medios cambiantes es cuando se presentan graves problemas de escorpionismo en una zona geográfica (LOURENÇO & CLOUDSLEY-THOMPSON, 1996; GÓMEZ & OTERO, 2007). Por esta razón, se hace necesario conocer la diversidad y la ecología de estos artrópodos y su posible uso en la biotecnología, debido a que hay especies reportadas en este estudio con importancia médica. De esta manera, un adecuado conocimiento de la distribución, diversidad y variabilidad de estos arácnidos, permitirá formular políticas de educación y prevención de accidentes con escorpiones que atiendan adecuadamente a la población más vulnerable (CUPITRA-VERGARA et al., 2015) y de paso contribuir a la conservación de la biodiversidad en el Caribe colombiano.
El registro del 60% de las familias y el 40% de los géneros conocidos para Colombia en siete zonas con diferente grado de perturbación en el departamento del Atlántico, puede interpretarse como un registro de alta diversidad biológica para este grupo en esta región. Por otro lado, más allá del registro de especies, se evidencia la necesidad de realizar estudios que muestren el estado actual del BST en el departamento del Atlántico y cómo las actividades antrópicas afectan la dinámica de este ecosistema; utilizando como modelo la diversidad de este grupo de arácnidos. Adicional a lo anterior, el presente trabajo constituye una línea base importante sobre la diversidad de escorpiones en el Caribe colombiano, ya que aporta una información ecológica valiosa sobre la estructura de la comunidad de estos arácnidos en un ecosistema que se encuentra en un estado crítico de fragmentación y degradación, y con un porcentaje muy pequeño de representatividad dentro de las áreas del Sistema Nacional de Áreas Protegidas en Colombia.
ConClusiones
Esta investigación representa el primer estudio de escorpiones en Bosque Seco Tropical del departamento de Atlántico. La escorpiofauna del BST en este departamento está compuesta por tres familias (Buthidae, Hormuridae y Chactidae), seis géneros (Ananteris, Chactas, Centruroides, opisthacanthus, Rhopalurus y Tityus) y siete especies,

Diversidad de escorpiones (Chelicerata) en el departamento del Atlántico, Colombia 243
de las cuales T. (Archaeotityus) tayrona es la especie con más amplia distribución y abundancia, encontrándose en todas las estaciones de muestreo. La estación con mayor diversidad (seis de las siete especies capturadas) fue la Reserva Campesina La Sierra (R.C.S), fragmento donde se observó mayor oferta de recursos, disponibilidad de hábitats y condiciones microclimáticas que influyen en un mayor número de especies.
aGradeCiMientos
A los integrantes del semillero de Investigación de Artrópodos “NEOPTERA” del Caribe colombiano, en especial a Leonel Martínez y a Eduardo Villareal por su colaboración en la fotografía de las especies y el diseño del mapa. A la Vicerrectoría de Investigación y Proyección Social y a la Facultad de Ciencias Básicas de la Universidad del Atlántico, por facilitar los permisos de recolecta y el uso del Laboratorio de Zoología para la determinación taxonómica de los escorpiones. Al biólogo Renzy Medrano por la ayuda en la recolecta de los especímenes. Al Instituto de Ciencias Naturales (ICN) de la Universidad Nacional de Colombia, sede Bogotá, por su apoyo y colaboración en la identificación de los especímenes.
referenCias
AGUDELO, C., 2004.- estudio preliminar de los escorpiones (Arachnida: Scorpionida) del Caquetá: Tesis, Universidad de la Amazonia, Facultad de Ciencias Básicas, Florencia.
ARMAS, L.F., SARMIENTO, D.L. & FLÓREZ, E., 2012.- Composición del género Centruroides Marx, 1890 (Scorpiones: Buthidae) en Colombia, con la descripción de una nueva especie. Boletín Sociedad entomológica Aragonesa, 50: 105-114.
BOHÓRQUEZ-GÓMEZ, R.M., 2016.- Revisión taxonómica y distribución geográfica de las especies de Tityus, Subgénero Atreus, (Scorpiones, Buthidae) presentes en Colombia: Tesis, Universidad Nacional de Colombia, Facultad de Ciencias, Bogotá.
BOTERO-TRUJILLO, R., 2008.- The scorpion genus Ananteris in Colombia: comments on the taxonomy and description of two new species (Scorpiones, Buthidae). The Journal of Arachnology, 36: 287-299.
BOTERO-TRUJILLO, R. & FAGUA, G., 2007.- Additions to the knowledge of the geographical distribution of some Colombian Scorpions (Buthidae: Ananteris, Rhopalurus, Tityus). Revista Ibérica de Aracnología, 14: 129-134.
BOTERO-TRUJILLO, R. & FLÓREZ, E., 2011.- A revisionary approach of Colombian Ananteris (Scorpiones, Buthidae): two new species, a new synonymy, and notes on the value of trichobothria and hemispermatophore for the taxonomy of the group. Zootaxa, 2904: 1-44.
BRAVO, E. & RODIÑO, I., 2013.- escorpiofauna (Arachnida: Scorpionida) en algunos relictos de Bosque Seco en el Departamento de Córdoba-Colombia: Tesis, Universidad de Córdoba, Facultad de Ciencias Básicas e Ingenierías, Montería.
CHAO, A., GOTELLI, N.J., HSIEH, T.C., SANDER, E.L., MA, K.H., COLWELL, R.K. & ELLISON, A.M., 2014.- Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. ecological Monographs, 84: 45-67.
CHAO, A. & JOST, L., 2012.- Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. ecology, 93: 2533-2547.
CLARKE, K.R. & WARWICK, R.M., 2001.- Change in marine communities: an approach to statistical analysis and interpretation. 2nd ed. PRIMER-E Ltd, Plymouth. Inglés, United Kingdom.
CUPITRA-VERGARA, N.I., CUBIDES-CUBILLOS, S., SALDARRIAGA-CÓRDOBA, M.M. & ESTRADA-GÓMEZ, E., 2015.- Distribución de Centruroides edwardsii (GERVAIS, 1843) en el departamento en Antioquia, Colombia. Acta biológica Colombiana, 20: 207-215.
FLÓREZ, E., 1990.- Escorpiones de Colombia. Catálogo de especies Cespedesia, 16/17 (57-58): 117-127.FLÓREZ, E., 2001.- Escorpiones de la familia Buthidae (Chelicerata: Scorpiones) de Colombia. Biota colombiana, 2: 25-30.FLÓREZ, E., 2004.- Escorpiones del chocó Biogeográfico/Costa Pacífica: 127-132 (en) RANGEL, J.O. (ed.) Colombia Diversidad
Biótica IV. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.FLÓREZ, E., 2013.- Estudio taxonómico, análisis filogenético y distribucional del género Chactas (Scorpiones, Chactidae): Tesis,
Universidad Nacional de Córdoba, Facultad de Ciencias, Argentina.FLÓREZ, E. & SÁNCHEZ, H., 1995.- Diversidad de los arácnidos de Colombia. Aproximación inicial: 327-372 (en) RANGEL, J.O
(ed.) Colombia Diversidad Biótica I. Instituto de Ciencias Naturales, Universidad Nacional de Colombia - INDERENA, Bogotá.GÓMEZ, A.D., MARTÍNEZ, A.J., MENDOZA, P.H., ÁLVAREZ, G.D. & RUIZ, P.S., 2013.- Registro de Escorpiones (Chelicerata:

eider Sará-C., eduardo Flórez-D., Neis Martínez-H.244bo
l.cie
nt.m
us.h
ist.n
at. 2
1 (2
) jul
io -
dici
embr
e 20
17. 2
31-2
44
Scorpiones) para el Departamento de Sucre, Colombia. Revista Colombiana Ciencia Animal, 5: 150-153.GÓMEZ, J. & OTERO, R., 2007.- Ecoepidemiología de los escorpiones de importancia médica en Colombia. Revista Facultad
Nacional de Salud Pública, 25: 50-60.HILL, M.O., 1973.- Diversity and Evenness: A Unifying Notation and Its Consequences. ecology, 54: 427-432.JOST, L., 2006.- Entropy and diversity. oikos, 113: 363-375.LOURENÇO, W.R., 1991.- Les scorpions de Colombie, II. Les faunes des régions de Santa Marta et de la Cordillère Orientale.
Approche biogéographique. Senckenbergiana Biologica, 71: 275-288.LOURENÇO, W.R., 1992.- Biogéographie évolutive, écologie et les stratégies biodémographiques chez les scorpions neotropicaux.
Scorpions Biogeographica, 67: 171-190.LOURENÇO, W.R., 1994a.- Biogeographic patterns of tropical South American scorpions. Studies on Neotropical Fauna and
environment, 29: 219-231.LOURENÇO, W.R., 1994b.- Scorpions Chelicerata de Colombie, VI: Quatre nouvelles espèces de Buthidae des régions Amazonienne,
Sud-pacifique et de la Cordillère Orientale. Revista de la Academia Colombiana de Ciencias exactas, Físicas y Naturales, 19: 387-392.LOURENÇO, W.R., 1997.- Synopsis de la faune de scorpions de Colombie, avec des considérations sur la systématique et la
biogéographie des espèces. Revue Suisse de Zoologie, 104: 61-94.LOURENÇO, W.R., 1998.- Panbiogeographie, les distributions disjointes et le concept de famille relictuelle. Scorpions Biogeographica,
74: 133-144LOURENÇO, W.R., 2002.- Scorpion in Amazonian Arachnida and Myriapoda: 399-438 (en) ADIS, J. (ed.) Amazonian Arachnida
and Myriapoda. Pensoft, Sofia-Moscow.LOURENÇO, W.R. & CLOUDSLEY-THOMPSON, J.L., 1996.- Effects of human activities on the environment and on the
distribution of dangerous species of scorpions: 49-60 (en) BON, C. & GOYFON, M. (eds.) envenomings and their treatments. París, Francia.
LOURENÇO, W.R. & FLÓREZ D.E., 1989.- Los escorpiones (Chelicerata) de Colombia. I. La fauna de la Isla Gorgona. Aproximación biogeográfica. Caldasia, 16 (76): 66-70
LOURENÇO, W.R. & FLÓREZ, E., 1990.- Scorpions (Chelicerata) de Colombie IV. Biogéographie et diversité biologique des scorpions de Colombie avec des commentaires sur les refuges Quaternaires. Scorpions Biogeographica, 66 (2): 65-74.
MANZANILLA, J. & DE SOUSA, L., 2003.- Ecología y distribución de Rhopalurus laticauda Thorell, 1876 (Scorpiones: Buthidae) en Venezuela. Saber, 15: 3-14.
PIZANO, C. & GARCÍA, H., 2014.- el Bosque Seco Tropical en Colombia. Instituto de Investigación de Recursos Biológico Alexander von Humboldt, Bogotá.
R DEVELOPMENT CORE TEAM. 2014.- R: A language and environment for Statistical Computing. R Foundation for Statistical Computing, Viena (Austria). Disponible en: http://www.R-project.org
RANGEL-ACOSTA, J.L. & MARTÍNEZ-HERNÁNDEZ, N.J., 2017.- Comparación de los ensamblajes de escarabajos copro-necrófagos (Scarabaeidae: Scarabaeinae) entre fragmentos de bosque seco tropical y la matriz adyacente en el departamento del Atlántico-Colombia. Revista Mexicana de Biodiversidad, 88 (2): 389-401.
RANGEL-CH., J.O. & CARVAJAL-COGOLLO, J.E., 2012.- Clima de la Región Caribe Colombiana: 67-129 (en) RANGEL-CH., J.O. (ed.) Colombia Diversidad Biótica XII: la región Caribe de Colombia. Universidad Nacional de Colombia, Bogotá.
SÁNCHEZ-AZOFEIFA, G.A., QUESADA, M., RODRÍGUEZ, J.P., NASSAR, J.M., STONER, K.E., CASTILLO, A. et al., 2005.- Research priorities for Neotropical dry forests. Biotropica, 37: 477-485.
SARÁ-CASTILLO, E. & MARTÍNEZ-HERNÁNDEZ, N.J., 2017.- Fauna de escorpiones (Arachnida: Scorpiones) en Fragmentos de Bosque Seco Tropical (BST), del Departamento del Atlántico, Colombia. Editorial Académica Española.
TERUEL, R. & GARCÍA, L.F., 2008a.- Rare or poorly known scorpions from Colombia. I. Redescription of Tityus macrochirus Pocock 1897 (Scorpiones: Buthidae). euscorpius, 63: 1-11.
TERUEL, R. & GARCÍA, L.F., 2008b.- Rare or poorly known scorpions from Colombia. II. Redescription of Tityus columbianus (Thorell 1876). euscorpius, 64: 1-14.
TERUEL, R. & RONCALLO, C., 2008.- Rare or poorly known scorpions from Colombia. III. On the taxonomy and distribution of Rhopalurus laticauda Thorell, 1876 (Scorpiones: Buthidae), with description of a new species of the genus. euscorpius, 68: 1-12.
TERUEL, R. & RONCALLO, C., 2010.- Rare or poorly known scorpions from Colombia. IV. Additions, synonymies and new records (Scorpiones: Buthidae, Scorpionidae). euscorpius, 105: 1-15.

Boletín CientífiCoCentro de Museos
Museo de Historia naturalnoVedades en Historia natural
a.- exposición temporal “enraiZaMiento: tHe CoMMon floW - el fluJo CoMun”
Se trata de una singular exhibición que coordinó la Dra. Olga Lucía Hurtado en el marco del evento: XVi festival internacional de la imagen, y lanzada durante el mes de junio de 2017 en el Centro de Museos por el artista caldense Luis Camargo, de gran trayectoria internacional, quien hace una relación entre el conocimiento cultural y la realidad de la naturaleza, ejemplificada con ciertas imágenes basadas en la geometría fractal. Asimismo, refiere a los flujos de la naturaleza presentes en los ríos, el corazón humano y el mismo universo recordando la frase “Todo está en todo, y nada es completo sin todo lo demás”, y mostrando un orden en las formas aparentemente caóticas de la naturaleza. Dicha percepción, según el artista, fue la inspiración para desarrollar tales flujos en una relación coherente entre las partes y el todo. Detalles: [email protected]
b.- ilustración adicional de material comparativo con entidades conocidas de riodinidae para el artículo de nuevas especies y subespecies del Boletín 21 (1) de 2017
En la pasada edición del Boletín se tuvo la oportunidad de describir varias especies de ilustrados de izquierda a derecha Riodinidae, una singular familia de pequeñas mariposas diurnas casi exclusivas del Neotrópico. En dicho trabajo, 7 entidades fueron descritas como nuevas para Colombia, a saber: Ithomeis eulema caucana n. ssp., Minotauros confusa n. sp., Panaropsis semiota remedios n. ssp., Pachytone erebia thalassina n. ssp., Calospila metaensis n. sp., Melanis pixe pseudoalena n. ssp.,

246
y Emesis brimo insulicola n. ssp. Ahora bien, cada una de ellas fue comparada con especies relativas ya conocidas para dilucidar mejor su nuevo estatus específico con los siguientes ejemplos aquí: Ithomeis eulema eulema (Hewitson, 1870), Hypophylla florus (Staudinger, 1887), Panara phereclus (Linnaeus, 1758), Crocozona pheretima (C. & R. Felder, 1865), Calospila rhesa (Hewitson, 1858), Melanis pixe corvina Stichel 1910), entre otras. Detalles: Julián A. Salazar E., [email protected]
Ithomeis eulema eulema (Hew.) Hypophylla florus (Stgr.)
Panara phereclus (L.) Crocozona pheretima (Fldr.)
Calospila rhesa (Hew.) Melanis pixe corvina (Stich.)

247
c.- depredación sobre Alesa amesis (Cramer, 1777) (lep., riodinidae)
Las arañas de la familia Salticidae, importantes controladores biológicos muy comunes en nuestras zonas tropicales, son depredadoras altamente especializadas en la captura de presas sin acudir a las telarañas como método de retención. La mayoría de ellas por su tamaño pequeño cazan en especial dípteros, himenópteros, homópteros y otros órdenes gracias a su gran agilidad para el salto, la astucia y su excelente visión. Ocasionalmente depredan mariposas, como en este caso una hembra del riodinido neotropical Alesa amesis (Cr.), especie de amplia distribución amazónica que frecuenta inflorescencias de diversas plantas y arbustos. En este caso una araña, posiblemente de la subtribu Freyina (Salticinae: Aelurillini) (W. Galvis com. pers.), es la protagonista del evento observado en Mocoa, Putumayo. Detalles y fotografia: [email protected]
d.- un singular ginandromorfo bilateral de Alesa amesis (Cramer, 1777) (lep., riodinidae)
El ginandromorfismo bilateral se deriva de los vocablos griegos gine: femenino, andro: masculino y morfo: forma. Es una anomalía cromosómica que ha sido documentada ampliamente por la ciencia y frecuente, mayormente, en la clase Insecta, en especial Lepidoptera y otros organismos. En consecuencia, los ginandromorfos presentan carecterísticas reunidas de ambos sexos, por lo general evidenciadas en diferentes partes el cuerpo. Cuando cada parte se distribuye equitativamente, o sea que cada mitad corresponde a un sexo diferente, se habla de que es una anomalía bilateral. Como ejemplo, ilustramos un individuo de Alesa amesis (Cr.) fotografiado en las selvas del Perú, que tiene el lado derecho correspondiente al macho y el lado izquiedo a la hembra e incluso, más espectacular, el abdomen, en el cual se nota la perfecta división de los anillos para cada sexo compartido. Este fenómeno es excepcional en la familia Riodinidae y rara vez documentado entre sus especies. Detalles: foto y registro David Geale, [email protected]

248
e.- Setabis rhodinosa (stichel, 1911) y Setabis preciosa (stichel, 1929), dos nuevos registros de especies para la fauna colombiana (lepidoptera: riodinidae)
Estas dos raras especies amazónicas de Setabis Westwood, 1851 fueron descritas por el naturalista alemán Hans Ferdinand Emil Julius Stichel en las décadas de 1910 y 1920 respectivamente, por ejemplares tipo femeninos procedentes de Ecuador (Sarayacu) y Perú (Pebas). Hasta el presente, y por lo que sabemos, no se han vuelto a registrar en listados o inventarios recientes de mariposas neotropicales publicados por diversos autores. Sin embargo, su presencia en Colombia era de esperarse, por lo que se confirma con la captura de dos ejemplares hembras que son ilustrados aquí. Datos: Setabis rhodinosa (Stich.); Colombia, Putumayo: Mocoa-Reserva Paway, a 590 m, ♀, 28-II-2017, J. Salazar leg. (CJS). S. preciosa (Stich.); Colombia, Meta: San Juan de Arama, a 350 m., ♀, 16-V-2015, J. Salazar leg. (CJS). Detalles: [email protected]
Setabis rhodinosa (Stich.) ♀ Setabis preciosa (Stich.) ♀

249
f.- el Boletín Científico de nuevo en Categoría a2 de Colciencias
El Boletín Científico de la Universidad de Caldas, órgano divulgativo de las diversas investigaciones en Historia Natural del alma máter desde 1996, ha estado clasificado en las categorías de Publindex C y B a partir del volumen 12 de 2008 y en A2 desde el volumen 13 de 2009, hasta el presente. Ahora, en julio 13 de 2017, se anunció de nuevo su renovación en dicha categoría en la convocatoria No. 768 de 2016, etapa II “clasificación oficial”, gracias a la denodada labor editorial, de los autores y de la Oficina de Revistas Científicas por un lado, y por estar incluido en el cuartil (Q2) del Índice bibliográfico sCiMaGo Journal ranK-sJr, por el otro. Este índice internacional es una medida de influencia científica de las revistas académicas, que explica tanto el número de citas recibidas por una revista como el prestigio de las revistas de donde proceden dichas citas. En tal sentido, el Boletín Científico aparece en el puesto 22 dentro de las primeras 50 publicaciones nacionales con el cuartil 2, uno de los más altos, que supera incluso al de otras revistas como Caldasia, Agronomía Colombiana y Acta Biológica Colombiana que tienen cuartil 3, y a la Revista Colombiana de entomologia con el cuartil 4. Este honroso lugar es el primero entre la revistas científicas de la Universidad de Caldas, junto a la revista KEPES con igual cuartil pero posicionada en el puesto 67 de dicho índice en cuestión. El editor agradece todo el apoyo recibido de la Vicerrectoría de Investigaciones y Posgrados, de la Oficina de Revistas Científicas con su personal a cargo y de la Facultad de Ciencias Exactas, Físicas y Naturales, que ayudaron en gran medida a tan enorme logro. Detalles: [email protected]

250
g.- relación de material adicional faltante en artículo
En el pasado Boletín 20 (2) de 2016, el presente autor junto a Alfonso Villalobos publicamos el estudio “Morfología descriptiva de los genitales masculinos en algunas especies neotropicales pertenecientes a la tribu Anaeini (Lep. Charaxidae)” pp. 199-224. Si bien se consideraron las genitalias de 15 especies, faltó la relación de dos de ellas referidas al material examinado para esa ocasión y es como sigue (en asterisco * los seleccionados):
Especie: Annagrapha polyxo (Druce) (Lámina fig. 7 en el artículo, p. 223), cantidad: 10 ♂♂, procedencia, PerÚ: Loreto, Iquitos-Pebas, Picuroyacu*, 150 m, 15-III-2011. Loreto, Centro Fuente-Río Momón, XII-2010. Loreto-Carbajal-Río Itaya, M. Dottax col. ColoMBia: Meta, Sierra de la Macarena, 500 m, 26-III-1970, E. W. Schmidt-Mumm leg.
Especie: Annagrapha dia divina (Röber) (Lámina fig. 8 en el artículo, p. 223), cantidad: 4 ♂♂, procedencia, PerÚ: Junín, Satipo-Kuvikiari*, 1200 m, XII-2009. Junín, Satipo, Shima, M. Dottax leg. (Detalles: [email protected]).

251autores
Mely Yenedith Carrillo Msc, Universidad de Córdoba, Monteria, Colombia. Correo electrónico: [email protected] ORCID: 0000-0003-0725-5703
Maria Eugenia Morales Puentes.PhD, Universidad Pedagojica y Tecnologica de Colombia, Tunja, Boyacá. Correo electrónico: [email protected]. ORCID: 0000-0002-5332-9956
Pablo Andrés Gil Leguizamón. Msc. Universidad Pedagojica y Tecnologica de Colombia, Tunja, Boyacá. Correo electrónico: [email protected]. ORCID: 0000-0002-9414-8991
Andrea Castiblanco, Bióloga. Universidad Pedagógica Nacional, Bogotá. Correo electrónico: [email protected]. ORCID: 0000-0002-8232-9074
César Augusto Pinzón. Biólogo, Universidad Pedagógica Nacional, Bogotá. Correo electrónico: [email protected]. ORCID: 0000-0002-6704-5137
Jonás Pinzón. PhD. Colegio Atlantic, COA, USA. Correo electrónico: [email protected]
Ana María Restrepo. Pregrado, Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected], ORCID: No dispone
Alberto Soto Giraldo. PhD. en Entomología. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-9727-8919
Yolanda Maritza Viveros. Ingeniería Agronómica. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-3495-9888
Oscar Adrián Guzmán. Msc. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0001-1724-2304
Bernardo Villegas Estrada. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-4441-5278
Maria Isabel Cruz. Particular, Ibagué, Colombia. Correo electrónico: [email protected]. ORCID: No dispone
Tito Bacca. PhD. Entomologia, Universidad del Tolima, Ibagué. Correo electrónico: [email protected]. ORCID: 0000-0001-1724-2304
Nelson Canal. Ingeniería Agronómica. Universidad del Tolima, Ibagué. Correo electrónico: [email protected]. ORCID: 0000-0003-4024-8687
Camilo Espinosa. Biólogo. Universidad del Valle, Cali, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-1538-9755
Lorena Cruz Bernate. Msc., Universidad del Valle, Cali, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-6854-8852
Guillermo Barreto. PhD. Universidad del Valle, Cali, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-2568-2629
Alan Giraldo. Especialista en Biología. Universidad del Valle. Cali, Valle, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0001-9182-888x

252
Christine Margareth Hahn. MVZ. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-0018-4642
Marco Tulio Jaramillo. Msc. Ingeniería Ambiental. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0001-6688-3318
José Neis Martínez. Msc. Ciencias Biológicas. Universidad del Atlántico. Barranquilla, Atlántico, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-7815-1437
Joel Vásquez. Msc., Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: [email protected], ORCID: 0000-0002-0584-7310
Wenceslao Peña. ecólogo, Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: no dispone, ORCID: 0000-0003-3903-9861
Ricardo Zarate. Msc., Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: [email protected] ORCID: 0000-0002-5904-8020
Julio Pinedo. Msc., Universidad Nacional de la Amazonia peruana, correo electrónico: no dispone, ORCID: 0000-0001-6784-0803
Rocío Correa, Doctorado, Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: no dispone, ORCID: 0000-0002-8002-4963
Juan José Palacios. Ingeniero Geografo, Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: [email protected]., ORCID: 0000-0002-0479-5053
Juan José Bellido. Msc., Instituto de Investigaciones de la Amazonia peruana, Iquitos, correo electrónico: [email protected]., ORCID: 0000-0003-3463-4661
Juan José Ramirez. Biólogo, Universidad Nacional Mayor de san Marcos, Lima, Peru. Correo electrónico: [email protected] ORCID: 0000-0001-5760-5172
Gerardo Lamas. Doctorado, Universidad Nacional Mayor de san Marcos, Lima, Peru. Correo electrónico: [email protected], ORCID: 0000-0002-3664-6730
Julián A. Salazar E. MVZ., Msc. Universidad de Caldas, Manizales, Colombia, correo electrónico. Julianmantisgmail.com. ORCID: 0000-0003-2268-7803
Vaclav Pacl. Msc., Escuela de Bellas Artes, Cali, Colombia. Correo electrónico: improbahotmail.com. ORCID: 0000-0002-8183-1417
Greg Nielsen. B.S. Piscicola Aquapro, Villaviencio, Meta. Correo electrónico: [email protected]. ORCID numero 0000-0003-0728-6059
Natalia Vinasco. Ingeniera Agrónoma, Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected] ORCID: 0000-0002-17931675
Luis Fernando Vallejo. PhD. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-0610-1270
Roberto Cambra. Doctorado, Universidad de Panamá, Panamá. Correo electrónico: [email protected] ORCID: 0000-0002-3799-4710

253
Diomedes Quintero. Doctorado. Universidad de Panamá, Panamá. Correo electrónico: [email protected]. ORCID: 0000-0002-2961-2196
Tatiana Cárdenas. Bióloga, Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-4363-361X
Ximena Ospina. Bióloga. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-1288-2867
Miguel Zambrano. Biólogo. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-5857-9997
María Rozo. Bióloga. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-2792-2091
Rodrigo Romero. Msc. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-4209-8021
Lucimar Dias Gomes. Doctorado. Universidad de Caldas, Manizales, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0001-6480-7688
Bedir Martinez. Msc. Universidad de Caldas, Manizales, Colombia. Correo electronico: [email protected]: 0000-0002-5798-7331
Alejandro Rubio. Biólogo, Universidad de Caldas, Manizales, Colombia. Correo electrónico: no dispone, ORCID: no dispone
Fabian Gaviria. Msc, Universidad de Paraná, Brasil. Correo electrónico: [email protected] ORCID: no dispone
Eider Sara. Biólogo, Universidad del Atlántico, Barranquilla, Colombia. Correo electrónico: [email protected] ORCID: 0000-0001-5555-4690
Eduardo Flórez. Doctorado, Universidad Nacional de Colombia, Bogotá. Correo electrónico: [email protected] ORCID: 0000-0001-1724-2304
Efrain Reinel Henao. Msc, PhD, ICN, Universidad Nacional de Colombia, Bogotá. Correo electrónico: [email protected] ORCID: 0000-0002-5482-6145
Luis H. Meneses. Biólogo, Parques Nacionales, ANULE,Bucaramanga, Colombia. Correo electrónico: [email protected] ORCID: 0000-0002-4905-390X
Mario Garcés. Msc., Universidad del Valle, Cali. Correo electrónico: [email protected] ORCID: 0000-0002-9937-7902
Katherine Pérez. Bióloga, Universidad del Valle, Cali. Correo electrónico: [email protected] ORCID: 0000-0001-5320-0346
Juan Pablo Sanchez. . Biólogo, Universidad del Valle, Cali. Correo electrónico: [email protected] ORCID: 0000-0002-4667-2422
Juan Mauricio Posada-Herrera. Biólogo. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-2436-4995

Jorge Luis Rangel Acosta. Magíster en Biología. Universidad del Atlántico. Barranquilla, Atlántico, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-1837-8696
Ana María Restrepo García. Ingeniera Agrónoma. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-9596-320X
Karen Ríos. Pregrado MVZ, Medicina Veterinaria y Zootecnia U. Cooperativa de Colombia. Ibagué, Tolima, Colombia. ORCID: 0000-0002-8528-8052
David Sanín. Máster en Biología Vegetal. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-0680-5531
Julio Andrés Sierra-Giraldo. Biólogo. Universidad de Caldas. Manizales, Caldas, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0002-3016-102X
Derly Suárez. Pregrado MVZ. U. Cooperativa de Colombia. Ibagué, Tolima, Colombia. ORCID: 0000-0003-2905-948X
Ana María Torres-Mejía. Msc. Medicina de la Conservación. Posgrado Regional en Ciencias Veterinaria Tropicales (PCVET), Universidad Nacional de Costa Rica. Lagunillas, Heredia, Costa Rica. Correo electrónico: [email protected]. ORCID: 0000-0001-9497-1825
Juan Fernando Vélez García. Magíster en Ciencias Veterinarias. Universidad del Tolima. Ibagué, Tolima, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0003-1517-2916
Melissa Abud Hoyos. Magister en Ciencias, Biología, Universidad del Valle, Cali, Valle, Colombia. Correo electrónico: [email protected]. ORCID: 0000-0001-5055-5256

255
el Boletín CientífiCo del Centro de Museos de la Universidad de Caldas es una revista especializada para la divulgación de trabajos resultantes de la investigación en Historia natural relacionada en ciencias biológicas afines. La revista se publica dos veces al año con las siguientes temáticas: Artículos principales (hasta 30 páginas manuscritas) relacionados con Mastozoología, Ornitología, Herpetología, Entomología, Botánica y Conservación.
Artículos enfocados a la conservación de colecciones biológicas o a diversos aspectos de Museología en Historia Natural.
Comunicaciones cortas como revisiones de libros, reconocimientos de trayectoria profesional, anuncios de cursos y eventos que tengan que ver con Historia Natural.
Sin excepción todo artículo presentado en este boletín debe ser original y no publicado en ninguna otra revista o medio electrónico existente.
Pautas Generales
Los artículos pueden ser escritos en español o inglés y serán publicados en el idioma en que sean enviados, pero no se aceptarán aquellos que estén publicados o sometidos a otra revista.
Los manuscritos serán revisados por, al menos uno o dos especialistas en el tema y podrán consultar con el director para su aceptación o no en la revista. Aquellos artículos ceñidos fielmente a las instrucciones indicadas aquí serán más favorecidos para su publicación.
Se debe enviar un original y dos copias de los trabajos, con letra de tamaño grande (12 o 14 puntos), alineados a la izquierda, en papel tamaño carta, con márgenes de 3 cm en todos los lados y a doble espacio (incluyendo título, palabras clave, resúmenes, textos, tablas y bibliografía).
Además se debe enviar el material en medio magnético (Disquete, Disco Compacto) y grabado en archivo MS-Word. En lo posible, todos los manuscritos deben llevar un título, un resumen y un índice de palabras clave en español (ideal si se incluye una versión en inglés), excepto en los de reconocimiento. No es indispensable que las páginas vayan numeradas.
Cuando los trabajos sean resultado de investigaciones sobre especies en particular que necesiten ejemplares testigo de colecciones, es importante indicar el lugar (museo, herbario o institución ojalá registrada ante el Instituto Alexander Von Humboldt) donde se encuentren depositados los especímenes. Así mismo, se debe indicar el colector o colectores, las fechas de captura y los sitios de recolección estrictamente. Igualmente, al referir material conservado odepositado en el Museo de Historia de Natural, Universidad Caldas debe citarse esta institución abreviadamente (MHN-UC) que lo diferencia del material contenido en el Museo de Historia Natural, Universidad del Cauca, puesto que tiene siglas abreviadas parecidas (MHN-UCC) y que han sido empleadas de modo confuso en otros números del boletín.
instruCCiones a los autores

256
de los artículos principales
El manuscrito de los artículos principales o el relacionado con la conservación de colecciones biológicas se debe dividir del siguiente modo secuencial: el título, el resumen y las palabras clave debajo de cada resumen, los nombres y apellidos de los autores omitiendo el segundo apellido o mencionándolo con la letra inicial, la dirección institucional de cada autor y el correo electrónico para correspondencia, la introducción, la metodología y los materiales empleados, los resultados, su discusión, los agradecimientos, la bibliografía citada en el texto, las tablas y la figuras respectivas usadas.
De las abreviaciones y estilo Los nombres científicos de especies se escriben en itálica. Es recomendable escribir el nombre completo por primera vez en el texto, pero en adelante se abrevian en lo posible. Por ninguna razón se deben dividir las palabras en el margen derecho en ningún lugar del manuscrito.
Se debe usar el sistema métrico y sus abreviaciones pata todas las medidas y utilizar cifras decimales que en los artículos en español van separadas con comas y en los de inglés con puntos (ej: 0,010 y 0.010).
de las referencias
Los nombres de los autores consultados en las referencias y que son mencionados en el texto deben citarse con letras mayúsculas y bajo las siguientes indicaciones o ejemplos:
“MARTÍNEZ (2000)” si el nombre del autor es parte de la oración, y “(MARTÍNEZ, 2000)”si no lo es“(MUÑOZ, 1999, 2000)” para dos artículos de un autor citados a la vez.“(MUÑOZ, 1999a, 1999b)” para dos artículos por el mismo autor en el mismo año“ (ALBERICO et al., 2000; BERTH et al., 2001; CALLEJAS et al., 2002; JORDAN et al., 2003; SALAZAR et al., 2003)”, para dos o más publicaciones de autores diferentes.
Deberán citarse en orden alfabético y luego cronológico o cuando aparezca un grupo de citas bibliográficas del mismo autor en años distintos. Los diversos autores se separan por punto y coma. Cuando la referencia corresponda a una cita de dos autores, los apellidos se conectan por el símbolo “&” y si la publicación es de más autores, debe mencionarse únicamente el primer autor en mayúsculas seguidos por “ et al “: “(MUÑOZ et al., 2001)”.
Se pueden mencionar datos no publicados, comunicaciones personales o por vía electrónica y artículos recientemente aceptados, en este caso deben enviar copia de la carta de aceptación al director de la revista para su verificación.
La lista bibliográfica mencionada al final del artículo debe ser en orden alfabético, con base en el primer apellido de los autores siempre en mayúsculas y luego el cronológico, siguiendo los siguientes ejemplos:

257
artículo publicado en revista
PECK, S.B. & FORSYTH, A., 1982.- Composition, structure and competitive behavior in a guild of Ecuadorian rain forest dung beetles (Coleoptera: Scarabaeidae). Canadian J. Zool., 60 (7): 1624-1634.
Capítulo en volumen editado
LÓPEZ, H. & MONTENEGRO, O., 1993.- Mamíferos no voladores de Carpanta: 165-187 (en) ANDRADE, G.I (ed.) Carpanta: Selva Nublada y Páramo. Fundación Natura, Bogotá.
Citación de libro
HILTY, S.L. & BROWN, W.L., 1986.- A Guide to the Birds of Colombia. Princeton Un. Press.
Citación de tesis
ÁLVAREZ, J.A., 1993.- Inventario de las mariposas (lepidoptera, Rhopalocera), con anotaciones ecológicas para dos zonas del departamento de Risaralda, Colombia: Tesis, Universidad Nacional de Colombia, Facultad de Ciencias, Bogotá.
En lo posible se deben abreviar los nombres de las revistas citadas y escribirlos en itálica, así mismo el título de los libros pero estos se escriben completamente sin abreviar. Para efectos de esta revista en la bibliografía o cuando se cite el Boletín Científico Museo de Historia Natural, Universidad de Caldas debe abreviarse como “Bol. Cient. Mus. Hist. Nat. U. de Caldas”.
de las comunicaciones cortas
En ellas se incluye las Novedades en Historia Natural, las Revisiones Bibliográficas, los homenajes biográficos etc. que pueden escribirse sin el uso de subtítulos en la introducción, materiales o métodos, resultados o discusión.
tablas
Deben citarse en el texto, identificadas y enumeradas consecutivamente con números arábigos. El encabezamiento debe ser conciso y descriptivo e ir sobre éstas. Las abreviaturas o símbolos se deben explicar al pie de la tabla. Las tablas deben salvarse como parte del texto y estar separadas en páginas diferentes después de las referencias bibliográficas. Deben explicarse por sí solas y no duplicar lo planteado en el texto. Es imprescindible digitarlas todas de acuerdo con el menú del Word ¨Insertar Tabla¨, en celdas distintas (no se aceptarán cuadros con valores separados por la herramienta ENTER o colocadas como figura).

258
figuras
Incluyen gráficos y fotografías, los cuales deben citarse apropiadamente en el texto y estar identificados y enumerados consecutivamente, usando números arábigos. El título de la figura debe ser conciso y descriptivo y se debe colocar debajo de la figura. Es importante cerciorarse de que cada gráfico o figura estén citados en el texto. Si se utilizaron figuras, datos publicados o inéditos provenientes de otra fuente, deben contener la fuente de donde fueron extraídos y referenciarla.
Las unidades, letras, números y símbolos deben ser claros y uniformes en todas las ilustraciones y de tamaño suficiente para que sigan siendo legibles, incluso después de la reducción necesaria para su publicación. Los títulos y explicaciones detalladas se deben incluir en los pies o epígrafes y no sobre las propias ilustraciones.
Los puntos de las curvas en los gráficos estadísticos se deben representar con marcadores contrastantes como círculos, cuadrados, triángulos o rombos (rellenos o vacíos). Así mismo, las curvas se deben identificar, de forma tal que el exceso de información no comprometa la comprensión del gráfico.
En el caso de gráficos de barras, se deben usar diferentes efectos de relleno (puntillados, líneas horizontales, verticales, diagonales, etc.). Se deben evitar los colores grises ya que dificultan la visualización en la impresión y no se debe usar fuente de letra en negrilla en las figuras.
Para enviar las figuras en medio digital se deben escanear a una resolución de mínimo 300 dpi, o se pueden digitalizar por medio de una cámara fotográfica.
de la Correspondencia
Todo material para publicar debe enviarse a:Comité EditorialBoletín Científico Museo de Historia NaturalCentro de Museos, Universidad de CaldasApartado aéreo 275, Manizales- ColombiaCarrera 23 No 58-65 Sede PalograndeTelefax 8851374E-mail: [email protected]
Los manuscritos que no se ciñan a las normas indicadas arriba se devolverán a sus autores con recomendaciones pertinentes para una adecuada publicación. A cada autor a quien se le publique se le enviarán copias del Boletín Científico del Museo de Historia Natural correspondientes al número donde aparece su trabajo publicado.
El Boletín Científico se encuentra indexado en los siguientes abstracts internacionales:
Biosis: Biological Abstract y Zoological Record Bibliography of Neotropical Butterflies (Association For Tropical Lepidoptera)- AbstractsIcom: Conseil International des Musées- Abstract

259
The Boletín CientífiCo del Centro de Mueseos of the Universidad de Caldas is a specialized journal, whose purpose is to disclose research results in natural HistorY related to similar biological sciences. The journal is published twice a year with the following topics: Main articles (up to 30 printed pages) related to Mastozoology, Ornithology, Herpetology, Entomology, Botany, and Conservation.
Articles aimed at the conservation of biological collections or at diverse aspects of Museology in Natural History.
Short communications such as book revisions, acknowledgements of professional trajectory, course and event announcements related to Natural History.
Without exceptions, all articles presented to this journal should be original. They must not be published in another journal or electronic means.
General Guidelines
The articles can be written in Spanish or English and will be published in their original language. However, articles that have been published in other journals will not be included.
The texts will be revised by at least one or two specialists and their concept will influence the director’s decision regarding publication. The articles the strictly follow these indications will be favored.
The original, two copies, a CD copy of the text with letter size 12 or 14, letter size pages, with 3 cm margins on all sides and doubled spaced (including title, key words, abstract, texts, tables and bibliography) in MSWord format should be sent.
Whenever possible, all of the texts should have a title, abstract and key words in both Spanish and English, except in the acknowledgments texts. When the texts are research results on particular species that require witness samples of collections, it is important to indicate the place (museum, herbarium or institution, preferably one that is registered in the Alexander Von Humboldt Institute) where the specimens are deposited. The collectors, the capturing dates and the recollection sites should also be indicated.
In addition, when referring to conserved or deposited material from the Museum of Natural History of the Universidad de Caldas, the institution should be cited using the abbreviation (MHN-UC), which differentiates it from the material found at the Museum of Natural History, Universidad del Cauca (MHN-UCC), to clear up the confusion, since they have been used interchangeably in previous volumes of the journal.
Main articles Main articles and those related to the conservation of biological collections should have the following order: title, abstract and key words, authors’ full names,
autHor Guidelines

260
institutional address and electronic mail address, introduction, methodology and materials, results, discussion, acknowledgments, bibliography, tables and graphs.
abbreviations and style
The scientific names of the species should be written in italics. It is advisable to write the complete name the first time it appears in the text, but from then on it should be abbreviated.
Words should not be divided anywhere in the text. The metric system and its abbreviations must be used for all measurements, separating the decimal numbers with periods.
Bibliography
The name of the authors consulted in the bibliography and that are mentioned in the text, should be cited in capital letters and under the following specifications:
MARTINEZ (2000), if the author’s name is part of the sentence, and (MARTINEZ, 2000) if not.(MUÑOZ, 1999, 2000) for two articles of the same author cited simultaneously.(MUÑOZ, 1999a, 1999b) for two articles of the same author from the same year.(ALBERICO et al., 2000; BERTH et al., 2001; CALLEJAS et al., 2002; JORDAN et al., 2003; SALAZAR et al., 2003), for two or more publications of different authors. They should be cited in alphabetical order, first, and then in chronological order. The diverse authors should be separated by semicolons.
When the reference corresponds to a citation of two authors, the last names should be connect by the “&” symbol, and if the publications belong to more than two authors, the last name of the first author goes in capital letters followed by “et al”: (MUÑOZ, et al., 2001).
Unpublished information, personal communications, electronic mails, and recently accepted articles can be mentioned. In the last case, the acceptance letter should be sent to the director for its verification.
The bibliographical list mentioned a the end of the article should be in alphabetical order according to the last name of the authors, always in capital letters and then in chronological order, as in the following examples:
Journal article
PECK, S.B. & FORSYTH, A., 1982.-Composition, structure and competitive behavior in a guild of Ecuadorian rain forest dung beetles (Coleoptera: Scarabaeidae). Canadian J. Zool., 60 (7):1624-1634.

261
Book chapter
LÓPEZ, H. & MONTENEGRO, O., 1993-Non—flying mammals of Carpanta: 165-187 (in) ANDRADE, G.I. (ed.) Carpanta: Selva Nublada y Páramo. Fundación Natura, Bogotá.
Book
HILTY, S.L. & BROWN, W.L., 1986-A Guide to the Bird of Colombia. Princeton Un. Press.
Thesis
ÁLVAREZ, J.A., 1993-Butterfly (lepidoptera, Rhopalocera) inventory, with ecological notes for two zones of the department of Risaralda, Colombia: Thesis, Universidad Nacional de Colombia, Science Faculty, Bogotá.
Whenever possible the name of the cited journals should be abbreviated and written in italics, additionally, the book titles should be written completely, without any abbreviations. When citing the Boletín Científico Museo de Historia Natural, Universidad de Caldas should be abbreviated as follows: “Bol. Cient. Mus. Hist. Nat. U de Caldas”.
short communications
These include novelties in Natural History, Bibliographical Revisions, Bibliographical Tributes, which can be written without subtitles (introduction, materials and methods, results, discussion).
Charts
They must be mentioned in the text, identified and enumerated consecutively with Arabic numbers. The heading must be concise and descriptive and must be placed above the charts. The abbreviations or symbols must be explained below the chart. The charts must be saved as part of the text and be separated on different pages after the bibliographical references. They must explained themselves, and not repeat what was mentioned in the text. It is essential that they are created with the Word menu “Insert Chart”, with different cells (charts with values separated by the ENTER key or as figures will not be accepted).
figures
They include graphs and photographs, which must be mentioned appropriately in the text. They must be identified and enumerated consecutively, using Arabic numbers. The title of the figure must be concise and descriptive and it should be placed underneath the figure.

262
It is important to make sure that each graph or figure is mentioned in the text. If figures, published or unpublished data originating from another source were used, they must contain the original source, and it must be referenced.
The units, letters, numbers and symbols must be clear and uniform in all the illustrations and of a size big enough for their legibility, even after the necessary reduction made for their publication. The titles and detailed explanations must be included underneath, and not in the illustrations themselves.
The points of the curves in statistical graphs should be represented with contrasting markers such as circles, squares, triangles or rhombuses (filled or empty). Additionally, the curves should be clearly identified, so that the excess of information does not jeopardize the understanding of the graph.
In the case of bar graphs, different filling effects must be used (dotted or horizontal, vertical, diagonal lines, etc.). The gray colors should be avoided since they hinder the visualization of the printed form, and bold letters should not be used in the figures.
In order to send the figures in digital form, they should be scanned in a resolution of minimum 300 dpi, or whenever possible, digitized by means of a camera.
Correspondence
Materials can be sent to: Comité EditorialBoletín Científico Museo de Historia NaturalCentro de Museos, Universidad de CaldasA. A. 275, Manizales, Caldas, ColombiaCarrera 23 # 58-65 Sede PalograndeTelefax: 8851374E-mail: [email protected]
The texts that do not follow the indicated norms will be returned to their authors with the appropriate comments for its publication.
Each author whose article is published will receive a copy of the Boletín Científico del Museo de Historia Natural Journal, corresponding to the number in which it is included.
The Boletín Científico Journal is indexed in the following international abstracts:
Biosis: Biological Abstract and Zoological Record.Bibliography of Neotropical Butterflies (Association for Tropical Lepidoptera)-Abstracts.Icom: Conseil International des Musées- Publication Abstract.

forMato de susCriPCiÓn Vicerrectoría de Investigaciones y Postgrados Universidad de Caldas. Calle 65 N° 26 - 10
A.A. 275 Manizales - ColombiaTel: 8781500 ext 11222
emails: [email protected] [email protected]
nombre / Name
Cédula / Identificatión number
dirección / Address
Ciudad / City
departamento / State
País / Country
teléfono / Phone Number
Profesión / Profession
Institución / Employer
dirección de envío /Mailing Address
suscriptores nacionales por un año. (2) ejemplares
Último ejemplar recibido / last issue mailed:
Año/Year Volumen/Volume Número/Number Fecha / Date
Código Postal /Zip Code
Boletín CientífiCoCentro de Museos
Museo de Historia natural



Esta revista se terminó de imprimiren el mes de noviembre de 2017 en los talleres de Editorial Matiz
Manizales - Colombia




















