
Sistemas de assentamentos no Sítio Salvaterra 2, Baixo Rio Itapecuru, Bacabeira – Maranhão


Meio Físico, Diversidade Biológica eSociocultural do Baixo Rio Negro,
Amazônia Central
2005

PRESIDENTE DA REPÚBLICA
MINISTRO DA CIÊNCIA E TECNOLOGIA
DIRETOR DO INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA
Editor-Chefe
Luís Inácio Lula da Silva
Sérgio Machado Rezende
José Antônio Alves Gomes
George Henrique Rebêlo
Editores-AdjuntosAntonio Carlos Webber
Efrem Jorge Gondim Ferreira
Esta obra foi publicada com o apoio financeiro:INPA, CNPq, FAPEAM e CEULM/ULBRA
Av. André Araújo, 2936 Caixa Postal, 478CEP:69060-001, Manaus AM, Brasil.
Tel: 55(92)642-3438 Fax: 55(92) 643-3223site: / e-mail:www.inpa.gov.br [email protected]
EDITORA

(Organizadores)
Edinaldo Nelson Santos-Silva Fábio Marques AprileVeridiana Vizoni Scudeller Sérgio Melo
2005
Meio Físico, Diversidade Biológica eSociocultural do Baixo Rio Negro,
Amazônia Central

Copyright 2005 Instituto Nacional de Pesquisas da Amazônia
Revisão ortográfica:H. Dias
Projeto gráfico e capa:Áttema Design Editorial
Diagramação e arte final:
Denis Maerlant / FALPE Comunicação
Fotolitos e impressão: Jacaré Gráfica e EditoraRua Paulo Eduardo de Lima, 1106Conj. Renato Souza Pinto II Cidade Nova IManaus - AM
©
www.attema.com.br
www.falpe.com.br
Conselho Nacional de DesenvolvimentoCientífico e Tecnológico
Financiadores:Parceiro:
BioTupé: meio físico, diversidade biológica e socio-cultural / Edinaldo Nelson dos Santos-Silva...[ .],organizadores. Manaus : INPA, 2005.246p.ISBN 85-211-0023-X1.Águas pretas 2. Biodiversidade Amazônia 3.Recursos naturais - Amazônia I. Santos-Silva,Edinaldo Nelson
et al
©
Proc. 55.1361/01-8
Temático 958/2003
PPI.1-3540

Prefácio ................................................................ IX
Introdução ................................................................ XI
Revisores ................................................................ XXIII
CAPÍTULO 2 Variáveis limnológicas: contribuição ao estudoespaço-temporal de águas pretas amazônicas ....... 19
CAPÍTULO 3 Limnologia do Lago Tupé: dinâmicaespaço-temporal do oxigênio dissolvido .............. 35
CAPÍTULO 4 Considerações sobre a geoquímica e dinâmicasedimentar do Lago Tupé ............................... 69
Meio Físico
CAPÍTULO 1 Modelos geomorfológicos para o Lago Tupé .......... 3Fábio Marques APRILE Assad José DARWICH&
Assad José DARWICH; Fábio Marques APRILEBarbara Ann ROBERTSON
Assad José DARWICH; Fábio Marques APRILE;Barbara Ann ROBERTSON Luiz Fernando ALVES
Fábio Marques APRILE; Assad José DARWICH José Carlos RAPOSO
&
&
&
Sumário

Diversidade Biológica
CAPÍTULO 5 Fitoplâncton ................................................ 87Sérgio MELO, Sérgio R. M. REBELO, Karla F. SOUZA, Mariângela MENEZESLezilda C. TORGAN
&
CAPÍTULO 6 Desmídias com ocorrência planctônica ................ 99
CAPÍTULO 7 Distribuição vertical do zooplâncton e sua relaçãocom as variáveis ambientais ............................. 109
CAPÍTULO 8 As esponjas do Lago Tupé ............................... 123
CAPÍTULO 9 Diversidade de macromicetos lignolÍticos ............. 135
CAPÍTULO 10 O uso da fauna pelos moradores da ComunidadeSão João e Colônia Central .............................. 141
CAPÍTULO 11 Estrutura fitossociológica de uma floresta de terrafirme na Amazônia Central .............................. 155
CAPÍTULO 12 Composição da Entomofauna Aquática em Igarapésda Reserva de Desenvolvimento Sustentável Tupé,Manaus, AM ................................................. 169
CAPÍTULO 13 Diversidade e composição da ictiofauna do LagoTupé ......................................................... 181
CAPÍTULO 14 Bancos de dados sobre biodiversidade na Amazônia:a experiência do Projeto Biotupé ...................... 199
Sérgio MELO, Sérgio R. M. REBELO, Karla F. SOUZA, Climéia C. SOARESMaria da Graça SOPHIA
Daniel PREVIATTELLI; Edinaldo Nelson dos SANTOS-SILVAAssad José DARWICH
Cecilia VOLKMER-RIBEIRO Franciely Benetti de ALMEIDA
André Luis WILLERDING, Marcerla Farias BARBOSA Thaís CamposBURLAMAQUI
Adriana Kulaif TERRA George Henrique REBÊLO
Karen Souza DINIZ Veridiana Vizoni SCUDELLER
Maria José do Nascimento LOPES
Mauro José CAVALCANTI
&
&
&
&
&
&
Maria Gercilia M. SOARES Kedma Cristine YAMAMOTO&

Diversidade Sociocultural
CAPÍTULO 15 Histórias: narrativas e depoimentos dascomunidades São João do Tupé e Colônia Central ... 217
enrique REBÊLO; Adriana Kulaif TERRA;Viviane MariaGuedes LAYME Tábita Maciel AMORIMGeorge H
&
CAPÍTULO 16 A visão vernacular e o desafio cultural paraConstruções em mutirão .................................. 239Leandro GASPARINI André Munhoz de Argollo FERRÃO&


As abordagens científicas dos biomas e ecossistemas amazônicos como objetosinterdisciplinares, são recentes. Na tradição científica brasileira privilegiou-se, até os anos 70, osinventários do meio físico por áreas de saber e/ou disciplinas específicas. Assim, a vida animal evegetal foi classificada segundo padrões especializados da botânica, entomologia, zoologia, etc. Afloresta e a malha hídrica, como unificador do conhecimento sobre o trópico úmido, apenasrecentemente passam a ser estudadas como um todo orgânico. Esta atitude científica induz à outrasintersecções, agora entre as ciências da natureza e da sociedade.
O estudo denominado “Biotupé: meio físico, diversidade biológica e sociocultural,realizado sob a responsabilidade do Instituto Nacional de Pesquisas da Amazônia - INPA e CentroUniversitário Luterano de Manaus - CEULM/ULBRA, se inscreve neste enfoque inter etransdisciplinar. As representações científicas produzidas em conjunto vêm apresentandoexcelentes resultados para compreensão sistêmica do trópico úmido, para a explicação dadiversidade física e da especificidade de funcionamento de cada ecossistema em particular. Nosestudos deste tipo residem enormes esforços para estabelecer relações causais e de determinaçãoentre fenômenos físicos de várias ordens, que buscam compreender processos de formação eevolução da natureza. O lugar e a interferência da ação humana nesses processos é uma dimensãoessencial para a apreensão da diversidade adaptativa da condição humana.
A ampliação das bases explicativas, compreensivas e descritivas, por si só, se constitui emproblema metodológico com implicações no discurso que as narrativas científicas têm dificuldadede coordenar. Imaginemos pois, a ordem e a densidade de questões, quando abordagens das ciênciasda natureza e das ciências da sociedade encontram-se sob um mesmo objetivo: o conhecimentorecíproco entre as intervenções e o ambiente físico em uma região particular. Os movimentos e ostempos de ocupação e usos, organização do espaço e os processos e ciclos da natureza são dinâmicose nem sempre harmonizam a história e a descoberta científica. Na verdade, em se tratando deAmazônia, o conhecimento tradicional de Povos e Nações têm precedência sobre a classificação daciência ocidental. Mediante diferentes formas de adaptabilidade os povos daAmazônia ocuparam asplanícies, várzeas, montanhas e manejaram a vida vegetal e animal em diferentes intervenções. As
locus
Prefácio

interações que se processaram entre a ação humana e a evolução natural são muito importantespara as percepções contemporâneas da Amazônia. Diríamos mesmo, que são fundamentais paraarticulação de hipóteses e questões de pesquisa acerca da origem, evolução e manutenção dosambientes da região.
O Tupé caracteriza uma região lacustre, composta de 05 cursos d'agua (igarapés), ligados aoRio Negro por um canal. Às margens do lago Tupé há populações recentes, etnias remanejadas, epovos originários do Alto Rio Negro. À esta diversidade de ocupação e uso do espaço físico, dediferentes momentos históricos, e de conhecimento, o INPA e CEULM/ULBRA pesquisam ageomorfologia, os aspectos limnológicos, a especificidade da dinâmica do lago em suas variáveisgeoquímica e sedimentar. A preeminência destes estudos para a compreensão da região é auto-explicável. Todas as possibilidades de adensamento demográfico, de arranjos e cadeias produtivasdevem, em tese, adequar-se às variáveis ambientais do ordenamento territorial da Amazônia.Mesmo que assim não fosse, é possível afirmar que este estudo pode ser fio condutor de novasabordagens da imensa bacia do Rio Negro, a partir de uma de suas amostras.
O Tupé é também um povoamento recente de novos ribeirinhos, migrantes de vários outrosrecantos da Amazônia e do Brasil, que utilizam as margens do Rio Negro como foco turístico e delazer. O impacto negativo dessas atividades sobre a vida natural é visível ainda que necessite de sermedido, classificado e cientificamente explicado. As atividades de campo captam,simultaneamente, variáveis físicas, ambientais e culturais para a compreensão do “Biotupé”. Olago, a praia, os igarapés, o canal de ligação com o Rio Negro são mais do que ângulos privilegiadosde acesso à fruição estática da paisagem. São ambientes produtores de vida no trópico úmido e,portanto, são unidades de reprodução da vida física, social e cultural, muito além de simplesrecursos para os usos da sobrevivência econômica.
O olhar interdisciplinar engatinha na escolha de variáveis compreensivas e explicativas quedão ao “Biotupé”a força dos estudos pioneiros. Ações de pesquisas deste porte constituem umachave para o acesso ao conhecimento da Amazônia profunda, além da superfície descritiva doslevantamentos.
Profª Drª Marilene Corrêa da Silva Freitas
Doutora em ciências sociais e professora do Departamento deCiências Sociais da Universidade Federal do Amazonas desdefevereiro de 1979. Atualmente dirige a Secretaria de Ciência eTecnologia do Estado doAmazonas.
X

Introdução
Reserva de DesenvolvimentoSustentável do Tupé: característicasgerais
O Estado do Amazonas possui uma das maiores áreas de florestas e águas continentais domundo. Estes ambientes abrigam uma elevada diversidade de organismos terrestres e aquáticos,cujo conhecimento restringe-se a grupos taxonômicos específicos e, ou, regiões restritas. Visandopreservar, manejar e, ou, usar de modo sustentável os recursos naturais, diversas regiões do Estadotêm sido demarcadas como área de proteção ambiental, seja no âmbito federal, estadual oumunicipal. Dentre estas destacam-se as Reservas de Desenvolvimento Sustentável (RDS), que sãoáreas de proteção ambiental associadas ao conceito de uso sustentável dos recursos naturais. Estasáreas abrigam a população local que continuam a usar os recursos naturais, no entanto, de forma a
Veridiana Vizoni SCUDELLER
Fábio Marques APRILE
1
2
Sérgio MELO
Edinaldo Nelson dos SANTOS-SILVA
2
2
Mestrado em Biotecnologia e Recursos Naturais UEA e Coordenação de Biologia - Ceulm/Ulbra. Av.Solimões, 2 Japiim II. Manaus AM;
Coordenação de Pesquisas em Biologia Aquática INPA. Av. André Araújo, 2936, CP 470. Manaus AM
1
2
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

XII
Scudeller et al.
básico de preservar a natureza e, aomesmo tempo, assegurar as condiçõese os meios necessários para areprodução e a melhoria dos modos eda qualidade de vida e exploração dosrecursos naturais das populaçõestradicionais, bem como valorizar,conservar e aperfeiçoar o conhecimen-to e as técnicas de manejo doambiente, desenvolvidas por estas
populações (Art. 1 ).
A RDS Tupé localiza-se na margemesquerda do rio Negro, a Oeste deManaus distante aproximadamente25km em linha reta do centro dacidade, a uma altitude média 20ma.n.m. Apresenta atualmente osseguintes limites: começa naconfluência do rio Negro com a
margem direita do igarapé Tatu (03
03'02,241”S e 60 17'46,121”W),seguindo por este até a sua nascente
(03 01'18,293”S e 60 19'10,903”W);desse ponto, segue por uma linha retano sentido sul/norte até o igarapé
Acácia (02 58'03,139”S e 60 19'10,405”W), daí, por uma linha mediana,até a confluência com o igarapé
Tarumã-Mirim (02 57'25,023”S e 6012'45,624”W), e por fim segue pelamargem direita do igarapé Tarumã-
Mirim até a sua foz com o rio Negro (03
01'42,851”S e 60 10'30,770”W), seguindo pela sua margem esquerda, até oponto inicial do igarapé Tatu (Fig. 1).
o
o
Localização
o
o o
o o
o o
o
o -
preservar e manter a biodiversidade. Entre as RDS doEstado do Amazonas, a Reserva de DesenvolvimentoSustentável Tupé (RDS Tupé) ocupa uma posição única,por ser a maior unidade de proteção do município deManaus com cerca de 12.000ha.
Ao longo dos últimos quinze anos, visando a proteçãoambiental da região do lago Tupé, o poder públicomunicipal instituiu os seguintes instrumentos legais:
a) em 1990, a Praia do Tupé foi declarada comoÁrea de Relevante Interesse Ecológico pela Lei Orgânicado Município de Manaus (LOMAM);
b) em 1995, pela Lei Municipal Nº 321, foi criadaa Unidade Ambiental do Tupé (UNA-Tupé) definindo-setambém os limites territoriais dessa área, integrante dosistema municipal de unidades de conservação;
c) em 1999 o Decreto Municipal No 4.581, de18.06 instituiu aquela área como “espaço territorial derelevante interesse ecológico”, e atribuiu à SecretariaMunicipal de Desenvolvimento e Meio Ambiente(SEDEMA) a gestão ambiental daquela localidade;
d) ainda em 1999, por meio da Portaria No18/99, de 24.06, a SEDEMA instituiu o Regulamento daÁrea de Relevante Interesse Ecológico do Tupé (ARIE -Tupé), definindo os limites espaciais da mesma, bemcomo, estabelecendo diretrizes e estratégias para aimplantação e o funcionamento dessa unidadeambiental;
e) em 2002, pela Lei Nº 671/02 e atendendo aoque estabelece a legislação federal referente ao SistemaNacional de Unidades de Conservação (SNUC), a UNA-Tupé foi reenquadrada como Reserva de Desenvolvimen-to Sustentável Tupé - RDS Tupé.
f) em 2005, através do Decreto Nº 8044, assinadopelo prefeito de Manaus em 25.08 foi finalmente criada aReserva de Desenvolvimento Sustentável do Tupé (nessedocumento denominada REDES do Tupé) com área totalde 11.973ha e perímetro de 47.056m, com o objetivo
Histórico da criação da RDS Tupé

XIII
Reserva de desenvolvimento sustentável do Tupé: características gerais
Histórico do sistemade classificação davegetação
O projeto Radam Brasil (1978) tevecomo objetivo estudar, em escalaregional, a área da conhecida“Amazônia Legal”, retificando limites
e conceituando novamente o recobrimento vegetal da“afamada Hiléia brasileira de Humboldt”. A partir deparâmetros climáticos, pesquisadores do projeto RadamBrasil (1978) definiram três tipos de vegetação florestal:ombrófilo denso, de 0 a 30 dias secos; ombrófilo abertode 30 a 60 dias secos; e estacional semidecidual, de 60 a90 dias secos. No entanto, usando parâmetrosfisionômico-ecológico das formações procurou-seatualizar um sistema universal de classificação das
Figura 1. Limites e localização das comunidades existentes na RDS Tupé e no seu entorno.
M03.550 1.775 3.550
11
10
5
6
34
2
1
79
A
B
Rio Negro
8
Reserva de Desenvolvimento Sustentável do Tupé
Trilhas
Perímetro
Comunidades
Legenda 1. Agrovila2. São Sebastião3. Ebenezér4. Julião5. Fátima6. Livramento
7. São João do Tupé8. Central9. Tatulândia10. Arara11 Bela Vista
A. Trilha CentralB. Trilha da Cachoeira

formações vegetais.Para Veloso Góes-Filho (1982), a região da Floresta
Ombrófila Densa é a região realmente hileiana. Para osautores, foi esta área equatorial que Humboldt chamoude “Hyleia” e que Martius designou “Nayades”. Para aregião de Manaus foram determinadas três regiõesfitoecológicas evidenciadas pelas classes de formaçõesde Campinarana, da Floresta Densa Tropical(reconhecida popularmente como Floresta de TerraFirme) e da Floresta Aberta Tropical, além das áreas deacumulação inundáveis (Radam Brasil, 1978).
ARegião Ecológica da Floresta Densa Tropical recobre153.688km de área e compreende seis sub-regiões, cadauma posicionada em gradientes litológicos emorfológicos característicos. De acordo com o RadamBrasil (1978), não foram observadas diferenças muitosignificativas na composição florística de cada umadelas, isto dentro dos limites que a escala dos trabalhospermitiu. Os autores reconheceram que as espéciesdominantes entre as árvores altas pertencem às famíliasLeguminosae, Lecythidaceae e Sapotaceae em primeirolugar, correspondendo a 40,8% respectivamente deárvores com diâmetros superiores a 10cm.
Grande parte da Amazônia, 65% da área total, érecoberta por floresta de terra firme, ou Floresta DensaTropical, caracterizada pela elevada riqueza ediversidade de espécies (Prance, 1987; Guillaumet,1987). O termo geral da floresta de terra firme englobavários tipos de vegetação que variam segundotopografia, localização geográfica, geologia, entreoutros fatores.
Segundo Radam Brasil (1978), as áreas deacumulação inundáveis, anteriormente denominadasSub-Região das Áreas de Acumulação Inundáveis,constituem-se de entidades dinâmicas (graminóide,arbustiva e arbórea) em nítida fase de sucessão.Ocorrem em áreas aluviais com sedimentos do
&
Floresta Densa Tropical
Áreas deAcumulação Inundáveis
2
Quaternário, onde a inundaçãoperiódica condiciona e seleciona asdiferentes formações. Parecemevidentes os reflexos da água decolorações diferentes. Para Coelho
. (1976) Radam Brasil (1978)“uma característica marcante é acoloração da água presente nos riosonde predominam os depósitosarenosos, a água toma uma coloraçãoescura, proveniente de ácidos húmicoslixiviados por ação de água das chuvasque provocam a acidez da água a pontode, em alguns rios, atuar como inibidorda vida aquática; já nos rios de águabarrenta com grande profusão desedimentos em suspensão, a vidaaquática é intensa e variada emespécies”. Tal fato justifica, regional-mente o uso dos termos “famintos”para designar os rios de água preta e“ricos” para os de água branca (Sioli,1984). Evidentemente, observa-se onítido reflexo de tais característicassobre a vegetação das planíciesaluviais, de acordo com materialdepositado”.
As florestas inundadas apresentamcaracterísticas estruturais e florísticasdistintas das matas de terra firmeresultantes de diferenças geomorfoló-gicas e hidrológicas, como flutuaçãodo nível dos rios, duração do períodode inundação, variação do relevo,entre outras (Prance, 1979; Ayres,1986). Segundo Nelson Oliveira(1999) existe uma situação de totaldesconhecimento florístico e da estru-turação da comunidade vegetal dasflorestas periodicamente inundáveis,visto que menos de 0,0003% desta áreajá foi amostrada em estudos anterio-
et
al apud
&
XIV
Scudeller et al.

res. Sabe-se de apenas poucosinventários florestais em igapó noBrasil, a maioria concentrada na regiãodas proximidades de Manaus (Rodri-gues, 1961; Keel Prance, 1979;Revilla, 1981; Piedade, 1985; Worbes,1986 Ferreira, 1997; Ferreira,1991). No entanto, o estudo maispróximo geograficamente da bacia dolago Tupé é o de Ferreira (1991), noigarapé Tarumã-Mirim.
Nas planícies aluviais ocorrem comalta freqüência as espécies
(H.B.K.) Muell. Arg.(seringueira), spp. espp. (ucuubas), (L.)Gaertn. (sumaúma), L.(açacu), com destaque nos aluviõesmarginais ao rio Negro e tributários, asespécies Meg.(louro-inhamuí),
Camb. (jacareúba) eHub. ( aparajuba) (Radam
Brasil, 1978). Para os autores, osmaiores gregarismos de
H.B.K. verificam-se no relevocolinoso dos interflúvios tabularespróximos aos grandes lagos do sistemafluviolacustre do rio Solimões e, ainda,nos interflúvios de três afluentes do rioNegro que são os rios Demini, Jufari eBranco.
&
apud
Hevea
brasiliensis
Virola Iryanthera
Ceiba pentandra
Hura creptans
Ocotea barcelensis
Callophyllum brasi-
liense Manilkara
amazonica M
Bertholletia
excelsa
Caracterização dastrilhas utilizadas peloprojeto Biotupé
Trilha da Cachoeira
A trilha da Cachoeira possui uma
extensão aproximada de 3.800m. Seu início estálocalizado na sede da Comunidade São João do Tupé(03 2'50”S e 60 15'21”W), e, seu ponto final em umacachoeira situada à 03 02'02”S e 60 16'33”W.
se estende por -
o o
o o
o
o o
o
o
o
o o
A cachoeira, que dá o nome à trilha, apresenta umapequena queda de 1,5m , gerada por pequenas falhas nasrochas areníticas da região.
A trilha terrenos de platôs e vertentes. Em seu percurso prevalece solo arenoso e,pontualmente, nas áreas de platô observa-se latossolosargilosos e lateritos. A trilha é praticamente toda plana,exceto entre os km 1,3-1,5, onde corta um pequenoigarapé e existe uma inclinação do terreno acentuada(ca. 45 ).
A cobertura vegetal da trilha da Cachoeiracaracteriza-se por floresta alta, aberta e subosquevariando de aberto com palmeiras na porção maispróxima à sede da comunidade a fechado nas demaispartes da trilha.
A trilha Central possui uma extensão aproximada de2.000m. Seu início está localizado na margem direita doigarapé do Tupé (02 59'54,8”S e 60 16'17,4”W), e, seuponto final no limite do terreno de um morador daComunidade Colônia Central à 03 00'43,2”S e60 16'02,5”W.
A trilha se inicia em um terreno com um acliveacentuado (ca. 50 ) nos primeiros 100m. A partir daí,segue o tempo todo no divisor de águas (plana). Em seupercurso prevalece solos argilosos.
A cobertura vegetal da trilha Central caracteriza-sepor floresta alta, aberta e subosque aberto compalmeiras.
A trilha da Copaíba possui uma extensão aproximadade 1.000m. Seu início está localizado na margem direitado igarapé do Tupé (03 00'46”S e 60 02'15”W), próximoao Sítio do Pica Pau Amarelo (sítio de um comunitário daSão João) e, seu ponto final no limite do terreno da
Trilha Central
Trilha da Copaíba
XV
Reserva de desenvolvimento sustentável do Tupé: características gerais

Comunidade Colônia Central, próximo ao barracãocomunitário, à 03 00'45,5”S e 60 16'05,6”W.
Esta trilha foi aberta para facilitar o acesso aobarracão comunitário da Central. É uma trilha que,segundo relatos, já existia, mas que havia sidoabandonada há muito tempo. A trilha se inicia em umterreno com um forte aclive (ca. 50 ) nos primeiros100m. A partir daí, segue no platô (divisor de águas),sendo bastante plana. Em seu percurso prevalece soloargiloso e argilo-arenoso.
A cobertura vegetal da trilha Central caracteriza-sepor floresta alta, fechada e subosque aberto.
Os ambientes aquáticos da RDS Tupé compreendemprincipalmente alguns lagos e igarapés, no entanto, osestudos limnológicos na RDS concentram-se no lagoTupé, um lago de águas pretas (lago de Ria), localizadona margem esquerda do rio Negro (Fig. 1) com o qualmantêm-se conectado durante todo o ano, mas noperíodo de seca acentuada não sofre sua influência pois,o afluxo de água ocorre apenas do lago para o rio. No seutrecho mais profundo, o lago apresenta profundidadevariando entre 4,5m (seca) e 15m (cheia) dependo damagnitude do pulso de inundação.
Os estudos realizados no lago Tupé evidenciaram umaárea superficial de 68ha, comprimento de 3km, umalargura máxima de 300m, profundidade máxima de 15m,e reduzidos valores de pH (4,4-5,8) com concentraçãomédia de oxigênio dissolvido de 6mg.L podendoapresentar, no entanto, com condições de anoxia nascamadas mais profundas (Rai Hill, 1981a, 1981b,1984). Estes autores, com base em microorganismos ecaracterísticas físico-químicas, classificaram o lagocomo distrófico, oligomítico. Em relação aosinvertebrados aquáticos, apenas uma publicação édisponível sobre a fauna de macrobentos (Reiss, 1977),trabalho no qual o autor registra uma comunidadecomposta por poucos organismos, tanto nos aspectos
o o
o
Histórico dos estudoslimnológicos
-1
&
qualitativos quanto nos quantitativos,considerando a zona profunda do lago.
Somente no final dos anos 80iniciaram-se os estudos sobre as algasdo lago Tupé. Cronberg (1987) em umapublicação de revisão sobre crisofíceasde regiões tropicais apresenta 10táxons com ocorrência neste lago,entre estes uma nova variedade,
var. Cronberg,cuja localidade típica é o lago Tupé. E,no final da década de 90, Meyer .(1997) ao proporem uma nova espéciede Dinophyceae (
B. Meyer), registraram suaocorrência nas águas do Tupé. Noentanto, nenhum trabalho é conhecidoabordando a comunidade fitoplanctônica como um todo. Santos (1983)estudando as comunidades de peixescapturadas no lago Tupé e no rio Negroatravés de pesca experimental encontrou diferenças nas estruturas dessascomunidades. Em relação aos demaisgrupos biológicos não se têm conhecimento do seu estudo no lago Tupé, queembora localizado a poucos quilômetros de Manaus pouco se conhece sobrea biologia dos seus organismos eecologia deste ambiente.
A principal característica climáticada área abrangida pela Folha AS.20Manaus (Radam Brasil, 1978) é a quasetotal ausência de período seco, excetono setor E/NE, onde este apresenta-securto, com apenas um mês.
O clima da área é classificadosegundo Köppen (1948) como do tipo“Am” quente e constantemente úmido
Synura spinulosa striata
et al
Peridiniopsis
amazonica
-
-
-
-
Aspectos climáticos
XVI
Scudeller et al.

em sua quase totalidade, muitoembora possua uma estação secapouco pronunciada no setor leste ondeo clima é classificado como do tipo“Aw” (quente e ligeiramente seco). Aconstância anual da temperatura émarcada por uma amplitude térmicaque não ultrapassa os 5ºC (RadamBrasil, 1978).
A homogeneidade espaço-tempo-ral que ocorre na região Amazônica emrelação à temperatura não ocorre coma pluviosidade. A área compreendidapelo município de Manaus possui totaispluviométricos elevados, com distri-buição mensal mais regular no centro-oeste e com estações mais marcadas aleste. A estação menos chuvosaverifica-se entre julho e setembro naslocalidades do setor leste e desetembro a novembro nas do setorcentro-oeste. A estação mais chuvosano setor leste ocorre de dezembro aabril e de março a maio no centro-oeste da Folha. Segundo estudosrealizados por Figueroa Nobre(1990), a região onde está localizada aRDS Tupé encontra-se entre asisopletas 2.000 e 2.200mm e apresentaprecipitação anual máxima de2.210mm. A marcha mensal daprecipitação pluviométrica em Manausrevela os primeiros quatro meses doano com pluviosidade mais intensa, emcontraste com os meses de agosto asetembro, onde normalmente aprecipitação é inferior a 60mm.
Precipitação Pluviométrica
&
Temperatura doAr
Solos
A temperatura média registrada nas estaçõesmeteorológicas de Manaus é superior a 26ºC. Os mesesmais quentes do ano ocorrem de agosto a novembro,durante a primavera austral, coincidindo com apassagem do sol do Hemisfério Norte para o Sul. Nesteperíodo, a temperatura média em Manaus atinge 28ºC.Os meses menos quentes ocorrem durante o invernoaustral, Manaus apresenta temperaturas mínimas emtorno de 23ºC e as mínimas absolutas de 19ºC a 20ºC. Aamplitude térmica no setor centro-oeste, onde estáinserida a RDS (bacia do rio Negro), está em torno de 7ºCe no setor centro-sul (bacia do Solimões) é mais elevada(10Cº). Esta diferença, no entanto, parece não refletirdiretamente na fisiologia da vegetação.
ARDS Tupé situa-se em áreas das Formações Solimõese Barreiras com relevos tabuliformes, ondulado edissecado em cristas e colinas. Os solos predominantessão do tipo Podzólico Vermelho Amarelo, LateritaHidromórica e LatossoloAmarelo (Radam Brasil, 1978).
Na R Tupé estão inseridas seis comunidades, das quais o Projeto Biotupé atua em duas: São
João do Tupé e Colônia Central .De uma forma geral, as comunidades da RDS
Tupé podem ser descritas como povoados ou grupos deunidades residenciais compostas de várias famílias,distribuídas de forma irregular, organizadas emassociações criadas para cooperar sobre decisõesrelacionadas à educação e economia.
A comunidade São João situa-se na porta de entradapara a RDS Tupé, mais precisamente às margens do rioNegro e lago Tupé, sendo limitada por praia, mata deigapó e terra firme. Suas residências estão à beira dolago ou agrupadas na pequena vila que está se
População
DS(Fig. 1)
XVII
Reserva de desenvolvimento sustentável do Tupé: características gerais

estruturando nas proximidades da praia do Tupé.A comunidade Colônia Central situa-se em área
de terra firme da RDS Tupé, sendo seu acesso realizadoatravés da trilha Central, trilha dos Índios e trilha daCopaíba, utilizadas por moradores e pesquisadores doprojeto Biotupé, que têm respectivamente 2km, 2km e1km de extensão (Fig. 1).
Após conversas com os comunitários tivemosevidências que a ocupação demográfica da área deabrangência do projeto vem se dando ao longo deaproximadamente 40 anos (na Comunidade São João) e15 anos (na Colônia Central) (vide Rebelo Kulaif, 2005,neste volume).
Segundo levantamento socioeconômicopreliminar de 2002, a comunidade São João do Tupécompreendia 31 famílias e a comunidade Colônia Central24. O tamanho médio das famílias da comunidade SãoJoão é muito similar ao das observadas na Central,aproximadamente quatro pessoas por família. Asmaiores famílias compostas por 11 pessoas foramobservadas na Central. Mas famílias grandes assim foramrelativamente raras em ambas comunidades. Famílias deuma pessoa foram mais freqüentes em ambascomunidades, mas na Central, somadas, havia maisfamílias de 2-4 pessoas.
Quanto ao lugar de origem, a maioria dosentrevistados em ambas comunidades é amazonense,cerca de 17% do total de entrevistados nasceram emManaus, e entre os amazonenses entrevistados naCentral, metade eram indígenas do alto Rio Negro(Desãna, Tukano e Tuyuka), que migraram recentemente(depois deste levantamento eles se deslocaram maisuma vez, se mudando para a São João). Somado aosnascidos em outros estados da região (Pará, Roraima), hámais nortistas na São João, que na Central. Na Central hámais nordestinos (Maranhão, Piauí), do que na São João(Ceará). Amazônidas e migrantes partilham um modo devida ligado à agricultura familiar, extração de produtosda floresta, caça e pesca.
Ao serem indagados sobre a realização deatividades produtivas de subsistência, 57% das famíliasresidentes na comunidade São João afirmam
&
desenvolver alguma atividadeprodutiva. Destes 57%, são as seguintesatividades produtivas de geração derenda realizadas: 38% fruticultura(abacaxi, cupuaçu, banana e manga);21% roça (mandioca: produção defarinha); 13% avicultura (galinha); 8%artesanato (palha, plástico, bijuteria);4% plantas medicinais (remédiosalternativos); 4% apicultura (criaçãode abelhas com extração do mel); 4%exploram barracas na praia e 8% nãoresponderam à indagação. 40% dasfamílias de moradores que nãoexploram os recursos naturais obtêmrenda do serviço público (SEDEMA,SEMSA, SEMED, SUSAM, Polícia Militardo Estado) e 3% não responderam sobreesse aspecto.
O mesmo foi perguntado aosresidentes da comunidade ColôniaCentral e os dados deste levantamentopor família foram os seguintes: 83% dasfamílias realizam atividades produti-vas de exploração dos recursosnaturais e 17% dos residentes não seencontravam nas residências. Do totalde 83%, as atividades econômicas degeração de renda desenvolvidas são asseguintes: 50% fruticultura (cupuaçu32%, abacaxi 8%, coco 2%, abacate 2%,café 2%, açaí 2% e pupunha 2%); roça46% (mandioca para exploração deseus derivados: farinha, tucupi, gomae tapioca); avicultura 2% (galinha) efeijão 2%.
Quanto à pergunta: se gosta-riam de realizar atividades produtivasde exploração dos recursos naturaispara geração de renda e quaisatividades seriam essas, o resultadotambém foi diferenciado. Na comuni-
XVIII
Scudeller et al.

dade São João os resultados obtidosforam os seguintes: 90% responderamque sim, 7% não respondeu ao quesito eapenas 3% responderam que nãopretendem ou não querem desenvol-ver atividades produtivas. Dos 90% queresponderam positivamente: 33% dasfamílias gostariam de desenvolveratividades voltadas para a agricultura(roça e feijão); 27% à criação deanimais (galinha, porco e cabra); 17%ao comércio (mercearias e minimercados); 10% exploração da praia(barracas de bebidas e alimentos erestaurantes); 3% à pesca; 7% nãoresponderam e 3% responderam emoutras atividades.
As perspectivas em relação àsatividades produtivas por parte dasfamílias de moradores da comunidadeColônia Central, segundo os resultadosobtidos são as seguintes: 83% dasfamílias afirmaram que gostariam derealizar atividade produtiva, os demais17% não foram encontrados em suasresidências. Dos 83% das famílias quedemonstram interesse em realizaratividades econômicas de geração derenda 53% preferem feijão e roça demandioca; 13% optam pela fruticultura(cupuaçu, abacaxi, coco, açaí); 9% gos-tariam de desenvolver a piscicultura(criação de peixes em cativeiro); 4%são pela criação de animais (galinha eporco); e 4% se referiram a outrasatividades.
Na São João poucos praticam aagricultura hoje em dia. A disponibili-dade da praia durante grande parte doano (meses além das praias de verão dorio Negro), o crescimento da comuni-dade em torno da “infraestrutura
turística” presente, combinado com a facilidade deacesso ajudam a explicar este desenvolvimento. Mesmocom a presença de fontes permanentes de água paracultivo, o solo na São João é considerado fraco paraagricultura pelos seus atuais moradores, além dapresença das saúvas (citada por muitos moradores).Enquanto na Central as condições ecológicas atuais(cultivo sobre solo florestal) combinadas com a demandapor farinha e frutas pela comunidade São João e pela“infraestrutura turística” estimulam o desenvolvimentoda agricultura de pequena escala, além do pequenocomércio com Manaus.
As dificuldades de acesso e de transporterepresentam os maiores empecilhos para o escoamentoadequado da produção, havendo um alto índice deperda, principalmente de cupuaçu, pois no período desafra, esses problemas se impõem e a falta de condiçõespara transportar a produção ou ainda de transformá-lalocalmente em polpa ou em outros derivados acabamcausando seguidos prejuízos e desânimo nos produtores.Soma-se a esses problemas a total ausência de energiaelétrica, o que impede o armazenamento e obeneficiamento da produção, obrigando os produtores arealizarem a comercialização “ ” das frutas.
Um fator determinante para o agravamento dascondições de vida e de não utilização adequada dosrecursos naturais tanto na comunidade Colônia Central,quanto da comunidade São João é a constatação de que,apesar de terem força de vontade, há a ausência de umaorganização comunitária que possa desenvolver nos seusintegrantes um sentimento, uma visão de mundo e umaconsciência de vida coletiva. Iniciar esse processo deconstrução da convivência comunitária, a partir dospróprios atores sociais envolvidos é um desafio e tambémuma necessidade na busca do respeito ao meio ambientee proporcionar a perspectiva de melhoria na qualidadede vida dessas populações. É o desenvolvimentohistórico dos conceitos que envolvem a relação homem enatureza.
in natura
XIX
Reserva de desenvolvimento sustentável do Tupé: características gerais

Bibliografia Citada
Ayres, J. M. 1986. The conservation status of the whiteuakari. , (7): 22-26.
Cronberg, G. 1987. Scaled chrysophytes from thetropics. , 95: 191-232.
Ferreira, L. V. 1991. O efeito do período de inundação nazonação de comunidades, fenologia e regeneraçãoem uma floresta de igapó na Amazônia Central.Dissertação de Mestrado, Programa de PósGraduação-Fundação Universidade do Amazonas,Manaus,Amazonas. 161 pp.
Ferreira, L.V. 1997. Effects of the duration of flooding onspecies richness and floristic composition in threehectares in the Jaú National Park in floodplain florestin Central Amazonia. ,6: 1353-1363.
Figueroa, S.N., Nobre, C.A. (1990). Precipitationdistribution over Central and Western Tropical SouthAmerica.
, 5(6):36-45.Guillaumet, J. L. 1987. Some structural aspects of the
Forest. , 43:241-20.Keel, S. H. K. Prance, G. T. 1979. Studies on the
vegetation of a White-sand black-water igapó (rioNegro, Brasil). , 9:645-655.
Köppen, W. 1948. ; con un estúdio de losclimas de la tierra. Fonde de Cultura Económica,México.
Meyer B., Rai, H., Cronberg, G. 1997. The thecalstructure of spec. nov.(Dinophyceae), a new cist-producing freshwaterdinoflagellate from Amazonian floodplain lakes.
, 65 (1-4): 365-375.Nelson, B.W.; Oliveira, A.A. 1999. Avaliação e ações
prioritárias para a conservação do bioma FlorestaAmazônicas. Área Botânica. In: MMA.
PROBIO-Programa Nacional daDiversidade Biológica, MMA, Brasília.
Piedade, M.T.F. 1985. Ecologia e biologia reprodutiva deMart. (Palmae) como exemplo
Primate conservation
Nova Hedwigia
Biodiversity and Conservation
Climanálise: Boletim de Monitoramento e
Análise Climática
Experientia
ActaAmazonicaClimatologia
Peridiniopsis amazonica
Nova Hedwigia
Ações
Prioritárias para a Conservação da Biodiversidade da
Amazônia.
Astrocaryum jauary
&
XX
Scudeller et al.
de população adaptada às áreasi n u n d á v e i s d o r i o N e g r o(b l a ckwate r ) . D i s s e r t a ção(mestrado). Manaus.
Prance, G. T. (eds.).
Clarendon Press, Oxford,U. K. p. 28-45.
Prance, G. T. 1987. Biogeography ofthe Neotropical plantas. In:Whitmore, T. E.
Prance, G.T. 1979. Notes on thevegetation of Amazonia III. Theterminology of Amazon foresttypes subject to inundation.
, 31: 26-38.RADAM BRASIL, 1978. Levantamento
de Recursos Naturais. Folha SA. 20M a n a u s . R i o d e J a n e i r o ,Departamento Nacional deProdução Mineral.
Rai, H. Hill, G. 1981a. Physical andchemical studies of lago Tupé; acentral Amazonian black water,“Ria Lake”.
, 66(1):37-82.Rai, H. Hill, G. 1981b. Bacterial
biodynamics in Lago Tupé, acentralAmazonian black water “rialake”. .58(4): 420-468.
Rai, H. Hill, G. 1984. Microbiology ofAmazonian waters. In: Sioli, H.(Ed).
. Dr. W.Junk Publ., Dordrecht. p. 413-441.
Reiss, F. 1977. Qualitative andquantitative investigations onmacrobenthic fauna of CentralAmazon lakes. I. Lago Tupé, a blackwater lake on the lower Rio Negro.
, 6(2):203-235.
Biogeography and
quaternary history in tropical
America.
Brittonia
Int. Rev. Ges.
Hydrobiol.
Arc. Hydrobiol. Suppl
The Amazon; Limnology and
landscape ecology of a mighty
tropical river and its basin
Amazoniana
&
&
&
&

Revilla, J.D.C. 1981. Aspectosflorísticos e fitossociológicos defloresta inundável (igapó) PraiaGrande, rio Negro, Amazonas,Brasil. Dissertação (mestrado).Manaus INPA.
Rodrigues, W.A. 1961. Aspectosfitossociológicos das caatingas dorio Negro.
, sérieBotânica, 15:1-41.
Santos, G.M. 1983. Aspectosecológicos da pesca experimental
Boletim do Museu
Paraense Emilio Goeldi
em sistemas “lacustres” e fluvial no baixo rio Negro.Anais do
. ManausAM. p. 367-391.Sioli, H.1984. Introduction: History of the discovery of
the Amazon and research of Amazonian waters andlandscapes. p. 1-13. In: Sioli, H. (ed.)The Amazon:Limnology and Landscape Ecology of a MightyTropical River and its Basin. Monographiae Biologicae56. Dr. W. Junk Publishers, Dordrecht, TheNetherlands.
Veloso, H.P.; Góes-Filho, L. 1982. A vegetação daAmazônia. Silvicultura em São Paulo, 16(1): 333-346.
III Congresso Brasileiro de Engenharia de
Pesca CONBEP
XXI
Reserva de desenvolvimento sustentável do Tupé: características gerais


ADALBERTO PERDIGÃO PACHECO DE TOLEDO
ADEMIR CASTRO E SILVA
ANA CARLA DOS SANTOS BRUNO
Universidade Federal de São Carlos, Centro de Ciências Biológicas e da Saúde, Departamento deHidrobiologia.Via Washington Luiz, km 235MonjolinhoCEP 13565-905 - São Carlos, SP - BrasilCaixa-Postal: 676Telefone: (16) 3351-8383 Fax: (16) 3351-8310
Escola Superior de Tecnologia (EST/UEA), Departamento de Engenharia Florestal.Avenida Darcy Vargas, 1.200ChapadaCEP 69055-020 - Manaus, AM - BrasilTelefone: (92) 3236-1470 Fax: (92) 3236-1470
Instituto Nacional de Pesquisas da Amazônia, Núcleo de Pesquisas em Ciências Humanas eSociais.Av. André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - BrasilTelefone: (92) 3643-3371
Revisores

XXIV
CLAUDIA PEREIRA DE DEUS
EDINEA MASCARENHAS DIAS
EDUARDO COUTO DALCIN
Instituto Nacional de Pesquisas daAmazônia, Coordenação de Pesquisasem Biologia Aquática.Avenida André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - BrasilCaixa-Postal: 478Telefone: (92) 3643-3292Fax: (92) 3643-3292
Instituto Nacional de Pesquisas daAmazônia, Núcleo de Pesquisas emCiências Humanas e Sociais.Av. André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - BrasilTelefone: (92) 3643-3371 Ramal: 3371
Instituto Nacional de Pesquisas daAmazônia.Programa de Pesquisas emBiodiversidade/Núcleo deBiogeoinformáticaAv. André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM Brasil
EFREM JORGE GONDIM FERREIRAInstituto Nacional de Pesquisas daAmazônia, Coordenação de Pesquisasem Biologia Aquática.Avenida André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - BrasilCaixa-Postal: 478Telefone: (92) 3643-3234Fax: (92) [email protected]
ANA MARIA LEAL-ZANCHET
ÂNGELO MARTINS DE SOUZA JÚNIOR
ANTÔNIO CARLOS DEMANBORO
BÁRBARA ANN ROBERTSON
BEATRIZ RONCHI-TELES
Universidade do Vale do Rio dos Sinos, Centro deCiências da Saúde, Instituto de Pesquisas de Planárias.Av. Unisinos, 950Cristo ReiCEP 93022-000 - São Leopoldo, RS - BrasilCaixa-Postal: 275Telefone: (51) 5903333 Ramal: 1271 Fax: (51) 5908122
Universidade Estadual Paulista Júlio de Mesquita Filho,Instituto de Geociências e Ciências Exatas de RioClaro.Departamento de GeografiaRio Claro, SP Brasil
Pontifícia Universidade Católica de Campinas,Faculdade de Engenharia Ambiental.Campus I - Cidade UniversitáriaCEP 13084-900 - Campinas, SP - Brasil
Instituto Nacional de Pesquisas da AmazôniaCoordenação de Pesquisas em Biologia Aquática,Laboratório de Plâncton.Avenida André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - Brasil - Caixa-Postal: 479Telefone: (92) 3643-3383 Fax: (92) 3643-3324
Instituto Nacional de Pesquisas da Amazônia.
André Araújo 2936 - AleixoCEP 69060-001 - Manaus, AM - Brasil - Caixa-Postal: 478Telefone: (92) 3643-3202 Fax: (92) 3643-3332
demanboro@puc_campinas.edu.br
Coordenação de Pesquisas em Entomologia

XXV
ELSA RODRIGUES HARDY
GILMAR WANZELLER SIQUEIRA
ILSE WALKER
IRINEU BIANCHINI JÚNIOR
Instituto Nacional de Pesquisas daAmazôniaCoordenação de Pesquisas emBiologia Aquática, Laboratório dePlâncton.Avenida André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - Brasil -Caixa-Postal: 479Telefone: (92) 3643-3260Fax: (92) 3643-3292
Universidade Federal do Pará, Centrode Ciências Exatas e Naturais,Departamento de Química.Avenida Augusto Corrêa, 01,LAQUANAM/UFPAGuamáCEP 66075-100 - Belém, PA - BrasilTelefone: (91) 31832136 Ramal: 2136
Instituto Nacional de Pesquisas daAmazônia, Coordenação de Pesquisasem EcologiaAv. André Araújo, 2936 AleixoCEP 69060-001 Caixa Postal 478Manaus- AM
Universidade Federal de São Carlos,Centro de Ciências Biológicas e daSaúde, Departamento deHidrobiologia.Via Washington Luiz, km 235MonjolinhoCEP 13565-905 - São Carlos, SP -Brasil - Caixa-Postal: 676Telefone: (16) 3351-8383 Fax: (16)3351-8310
LIDIAMAR BARBOSA DE ALBUQUERQUE
LILIANA RODRIGUES
MARIA ANA FARINACCIO
Universidade Católica Dom Bosco, Museu Dom Bosco.Rua Barão do Rio Branco, 1843CEP 79002-173 - Campo Grande, MS - BrasilTelefone: (67) 312-6491
Universidade Estadual de Maringá, Centro de CiênciasBiológicas, Departamento de Biologia.UEM - Avenida Colombo, 5790 Bloco G-90Jardim UniversitárioCEP 87020-900 - Maringá, PR - BrasilTelefone: (44) 261-4629
Universidade de São PauloDepartamento de Botânica - IBUSPRua do Matão, 277 - Caixa Postal, 11461CEP 05422-970 - Cidade Universitária - ButantÃ, SP
LUIZ FERNANDO ALVES
MARCOS CALLISTO DE FARIA PEREIRA
Instituto Nacional de Pesquisas da AmazôniaCoordenação de Pesquisas em Biologia Aquática.Avenida André Araújo, 2936AleixoCEP 69060-001 - Manaus, AM - Brasil - Caixa-Postal: 479Telefone: (92) 3643-3137 Fax: (92) 3643-3224
Universidade Federal de Minas GeraisInstituto de Ciências BiológicasDepartamento de Biologia Geral.UFMG, ICB, Av Antonio Carlos, CP. 486PampulhaCEP 30161-970 - Belo Horizonte, MG - BrasilCaixa-Postal: 486Telefone: (31) 3499-2595 Fax: (31) 3499-2567

XXVI
PEDRO AUGUSTO SUAREZ MERA
SANDRA DO NASCIMENTO NODA
VERA LUCIA RAMOS BONONI
VICTOR PY-DANIEL
Instituto Nacional de Pesquisas daAmazôniaCoordenação de Pesquisas emBiologia Aquática,Laboratório de Plâncton.Avenida André Araújo, 2936 - AleixoCEP 69060-001 - Manaus, AM - Brasil -Caixa-Postal: 479Telefone: (92) 3643-3138 -
Fundação Universidade do Amazonas,Faculdade de Ciências Agrárias,Departamento de CiênciasFundamentais e DesenvolvimentoAgrícola.Avenida Gal. Rodrigo Otávio Jordão,Campus UniversitárioJapiimCEP 69077-000 - Manaus, AM - BrasilTelefone: (92) 3644-2354
Intituto de Botânica, Seção deMicologia e Liquenologia.Av. Miquel Estefano, 3687Água FundaCEP 04301-012 - São Paulo, SP - BrasilTelefone: (011) 5073-6300
Instituto Nacional de Pesquisas daAmazônia, Coordenação de Pesquisasem Ciências da Saúde, Laboratório deEtnoepidemiologia.Av. André Araújo 2936AleixoCEP 69060-001 - Manaus, AM - BrasilCaixa-Postal: 478Telefone: (92) 3643-3232
MARIA DAS GRAÇAS MACHADO DE SOUZA
PAULO ROBERTO DUARTE LOPES
Universidade de Brasília, ICB, Departamento deBotânica, Campus Universitário Darcy RibeiroAsa Norte - CP 04457CEP 70910-970 - DF - BrasilTelefone: (61) 307-2282 Ramal: 24
Universidade Estadual de Feira de Santana,Departamento de Ciências Biológicas, Laboratório deIctiologia.Campus Universitário Km 03, BR-116Novo HorizonteCEP 44031-460 Feira de Santana, BA-Brasil
MARLENE SOFIA ARCIFA
NEUSA HAMADA
NÚBIA ABRANTES GOMES
Universidade de São Paulo, Faculdade de FilosofiaCiências e Letras de Ribeirão Preto, Departamento deBiologia.Av. Bandeirantes, 3900Monte Alegre14040901 - Ribeirão Preto, SP - BrasilTelefone: (16) 602-3832Fax: (16) 602-3832
Instituto Nacional de Pesquisas da Amazônia.Coordenação de Pesquisas em EntomologiaAv. André Araújo, 2936AleixoCaixa Postal 47869060-001 - Manaus, AM - BrasilTelefone: (92) 36433369
Universidade Federal de Roraima, Centro de CiênciasBiológicas e da Saúde, Departamento de Biologia.Av. Cap. Ene Garcez, 2413Aeroporto69301207 - Boa Vista, RR - BrasilTelefone: (95) 6213176 Fax: (95) 6238584

XXVII
WALDECIRO COLAÇOUniversidade Federal de Pernambuco,Centro de Tecnologia, Departamentode Energia Nuclear.Cidade UniversitáriaCEP 50740-540 - Recife, PE - BrasilTelefone: (81) 3271-8252 Ramal: 355Fax: (81) [email protected]

1CAPÍTULO 1 - Modelos geomorfológicos para o Lago Tupé
Fábio Marques APRILE Assad José DARWICH&
CAPÍTULO 2 - Variáveis limnológicas: contribuição ao estudo espaço-temporal de águaspretas amazônicas
CAPÍTULO 3 - Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigêniodissolvido
CAPÍTULO 4 - Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
Assad José DARWICH; Fábio Marques APRILE Barbara Ann ROBERTSON
Assad José DARWICH; Fábio Marques APRILE; Barbara Ann ROBERTSONLuiz Fernando ALVES
Fábio Marques APRILE; Assad José DARWICH José Carlos RAPOSO
&
&
&
Meio Físico
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),


Meio FísicoCapítulo 1
Modelos geomorfológicos para oLago Tupé
RESUMO - O objetivo deste trabalho foi estudar a geomorfologia do lago Tupé, tendo comoproposta apresentar um modelo morfométrico para a bacia, baseado em aferiçõesrealizadas no período de seca do ciclo hidrológico 2003/2004. Os estudos incluíram a árealivre do lago, cinco igarapés de floresta e o canal de conexão com o rio Negro. O lago estáconectado ao rio por um canal de cerca de 20m de largura, 0,5m de profundidade e 150mde comprimento. A amplitude média de variação do nível da água no canal é semelhante àdo rio acima da cota 19m a.n.m. (acima do nível do mar). Os resultados da análisemorfométrica revelaram que o lago apresenta uma seção transversal com padrão tendendoao triangular, com formato de cunha irregular “V” deslocado para a margem direita.Durante a fase de seca a área do espelho de água foi de 66,9ha, e o volume deaproximadamente 1.440.260m .
PALAVRAS-CHAVE: geomorfologia, batimetria, geometria hidráulica, análise areal, lagoTupé.
3
Fábio Marques APRILE
Assad José DARWICH
Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Biologia AquáticaAv. André Araújo 2936, Caixa Postal 470 Manaus, AM, 69060-001
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

4
Aprile Darwich&
formas relacionadas com o escoamen-to das águas dos lagos.Ao contrário dosrios, os lagos na maioria das vezes nãoagem como transportadores de grandequantidade de materiais intemperiza-dos.
A geomorfologia lacustre ainda épouco estudada, no entanto, sabe-seque o transporte de material clásticolimita-se a curtas distâncias, quandode origem autóctone, em decorrênciada alta taxa de sedimentação. Já omaterial de origem alóctone chegaatravés do sistema fluvial com altaenergia, podendo ser distribuído portoda área lacustre.
Os lagos funcionam como canaisacumuladores, sendo que o fluxo dematerial clástico para o lago faz parteintegrante do ciclo hidrológico local.No caso do lago Tupé, tanto as caracte-rísticas físicas quanto as químicas ebiológicas, estão diretamente associa-das ao pulso de inundação do rioNegro. Neste caso, a alimentação dolago se processa preferencialmenteatravés de águas superficiais prove-nientes do rio Negro e dos igarapés defloresta, e em menor grau através deáguas subterrâneas e pelas águas daschuvas. Pelo fato do lago comounidade de sistema estar em contínuainteração com o meio, pode-se dizerque as condições climáticas, a litologiae a cobertura vegetal controlam amorfogênese das vertentes e, por suavez, o tipo de carga detrítica fornecidapelo sistema.
Todos os acontecimentos queocorrem na bacia de drenagemrepercutem de forma direta ou indire-ta no lago. Sendo assim, o estudo dos
Introdução
A geomorfologia é a ciência que estuda as formas derelevo de um ecossistema. Nesse estudo estão incluídosnão apenas os aspectos morfológicos como também osprocessos atuantes. Segundo Christofoletti (1997) oestudo morfológico ainda não evoluiu o suficiente parapermitir uma classificação aceitável em função dessecritério, muito embora já seja possível utilizarclassificações para vertentes, redes de drenagem,lineamento de canais, dunas e áreas costeiras. Emcontraposição, o estudo dos processos atuantes nosistema está bastante desenvolvido, já sendo possívelestabelecer uma classificação lógica e exeqüível dosfatos geomorfológicos através do estudo da dinâmicaregional.
Os primeiros estudos geomorfológicos que se têmnotícia remontam da antigüidade, tendo o primeirogrande avanço durante os séculos XV e XVI. No Brasil aevolução do conhecimento geomorfológico é recente. Apartir da década de 1940 o conhecimento geomorfológi-co do território brasileiro evoluiu de maneira rápida,com o surgimento de várias publicações nas áreas degeologia e geografia, destacando-se os estudos deMartonne (1943, 1944), Ruellan (1944, 1950 e 1952),Freitas (1951), Ab’Saber (1956, 1958, 1964), Bigarella(1961, 1964), Carvalho (1965) e Christofoletti (1977,1978). No campo geológico com ênfase para os processosde dinâmica sedimentar destacaram-se os trabalhos deSuguio (1973, 1994). Especificamente na área do lagoTupé, Rai Hill (1981) apresentaram os primeiros dadoshidrológicos e morfológicos sobre a bacia.
Com o passar do tempo, a geomorfologia no Brasilpassa a adotar novas linhas de pesquisa dentro dageomorfologia climática, envolvendo a elaboração eaplicação de modelos para explicar a evolução dasfeições típicas do relevo atual, que segundo CunhaGuerra (1998) caracteriza-se por um reafeiçoamentocontínuo da superfície e por uma sobreposição de formassobre uma estrutura morfológica preexistente, seja denatureza morfoestrutural seja morfoclimática. Ageomorfologia lacustre estuda os processos e define as
&
&

5
cursos de água que compõem a baciado lago Tupé só pode ser realizado emfunção da perspectiva global dosistema hidrográfico. O objetivo destetrabalho foi estudar o lago Tupé no seuaspecto geomorfológico, tendo comoproposta apresentar um modelomorfométrico e um mapa escalonadopara o lago.
Este estudo foi desenvolvido nolago Tupé durante o ciclo hidrológico2003/2004. Técnicas bastante divulga-das de análise morfométrica foramaplicadas segundo procedimentosmetodológicos descritos por Carvalho(1965), Maier (1983), Costa (1991) eChristofoletti (1997). O lago Tupé foidelimitado com auxílio de um GPSGarmin 12x e a partir de consultas emcartas e fotos aéreas. Os estudosgeomorfológicos incluíram a área livredo lago, os principais igarapés defloresta (igarapé Helena, das Pedras,da Cachoeira, Monteiro e Terra Preta) eo canal de comunicação com o rioNegro. Para a análise areal foramaplicados transectos dividindo o lagoem seções transversal, longitudinal ediagonal. Réguas foram instaladas empontos específicos do lago paraacompanhar a variação da coluna deágua em função do ciclo hidrológico.Para a análise batimétrica foi utilizadafita graduada 100 0,01m. Para definiros vários processos atuantes nageomorfologia do lago, foi feito umestudo da granulometria dos solos esedimentos do sistema através de
Metodologia
+
Modelos geomorfológicos para o Lago Tupé
técnicas de peneiramento e separação fracionada.Maiores detalhes da descrição metodológica sãoapresentados em Aprile . (2005, neste volume). Apartir dos resultados foi elaborado um modelomorfométrico para o lago. Um mapa foi digitalizado comauxílio dos software’s MaxiCad, Surfer eArcview.
Os primeiros estudos morfométricos relacionandolargura e profundidade em rios do centro oeste dosEstados Unidos foram desenvolvidos por LeopoldMaddock (1953). Para os autores, tais variáveis físicasestão diretamente ligadas à forma e tamanho do canal, eapresentam aplicabilidade em locais de margens bemdefinidas. Estudos posteriores de Leopold Wolman(1957, 1960) e Wolman Leopold (1957) revelaram queos eventos de magnitude moderada e de ocorrênciafreqüente podem controlar a forma de um canal.Tomando o lago Tupé como um grande canal receptor deáguas e material clástico, nota-se que sua morfologia ébem definida, sendo que o pulso de inundação associadoà variação pluviométrica são diretamente responsáveispela oscilação do nível de água do lago. A Figura 1apresenta a variação de nível da água do rio Negro e dolago Tupé durante o ciclo hidrológico de 2003,detalhando além disso, o início da enchente do cicloseguinte e a seca de 2002.
et al
Resultados e Discussão
Geometria Hidráulica
&
&&
Os registros diários de variação de nível do rio Negrodesde 1902 realizados no Porto de Manaus indicam que ascurvas de variação do pulso de inundação são sempremonomodais, semelhantes à da Figura 1, e comamplitudes em torno de 10m. Nesse período, o nível dorio apresentou cota máxima de 29,69m a.n.m., em junhode 1953, e mínima de 13,64m a.n.m., no final de outubrode 1963. Verifica-se portanto, que a amplitude total dopulso de inundação, no último trecho do rio Negro,alcançou cerca de 16m durante o período de registros.Naqueles ciclos hidrológicos, as amplitudes totais dospulsos foram, respectivamente, de 12,62m e 13,65m.

6
Essas variações têm estreita relação com a variação denível no lago Tupé por sua ligação com o rio Negro, amínima foi de 17,07m em 31 de outubro de 1953 emáxima de 27,31m em 17 de junho de 1963.
O lago Tupé está conectado ao rio por um canal decerca de 20m de largura, por 0,5m de profundidade e150m de comprimento durante a fase de seca (águasbaixas) e a amplitude média de variação de nível ésemelhante à do rio acima da cota 19m a.n.m. Quando onível da água está abaixo da cota 19m a.n.m não háentrada de água no lago pelo rio. Ocorre nesta fase,afluxo de água do lago ao rio proveniente,principalmente, da influência dos igarapés de florestaque convergem ao lago e da precipitação. No pico dacheia, em média, o nível rio atinge valores em torno dacota 28m a.n.m. e o lago apresenta profundidades cercade 15m em sua região central e de 10m na região decabeceira. Apesar da grande flutuação anual de nível, asoutras medidas lineares (comprimento e largura) e desuperfície (área livre) aumentam muito pouco quandocomparadas às de lagos de várzea, nos quais os desníveisda bacia são quase sempre muito pequenos. No Tupé, aocontrário, além do desnível, a bacia do lago apresentamargens íngremes, com vale em forma de “V”. Por isso, aárea inundável da bacia lacustre é pequena e a área livre
Figura 1. Nível da água no rio Negro medido no Porto de Manause no lago Tupé, na Estação Central (ET10). De outubro de 2002 amarço de 2004.
é exatamente a mesma área limitadapelas margens, visto não haverpresença de macrófitas aquáticas quepossam diminuir a superfície livre dolago.
Na foz do lago há bancos de areiacom altitude máxima em torno da cota30m a.n.m. bloqueando a bacia dolago, o que provavelmente pressionoua formação de um canal escavado até acota 19m a.n.m na parte lateral dobloqueio. Com isto, mesmo que o rioatinja cotas menores que 19m, o quefreqüentemente acontece no períodode seca, o lago permanece comprofundidade máxima cerca de 4,5m.Nesta fase, predomina as condiçõeshidroquímicas das águas dos igarapés eda precipitação pluviométrica.Durante todo o período de medições noPorto de Manaus (101 anos), em 26anos a cota mínima esteve acima de19m, sendo que oito dessas estiveramacima de 21m, com máxima de 21,84mem 1974. Observa-se que em mais de100 anos de medições, cerca de 74,3%dos períodos de águas baixas a cota dorio esteve bem abaixo de 19m. Épossível que o percentual real demáximas e mínimas durante aexistência do lago esteja próximodesses observados e ainda assim ocanal do lago está em torno da cota 19.Por isso, em primeiro lugar pode-sesupor que, a afluência do lago ao riotenha sido sempre muito pequena, aponto de não escavar abaixo da cotaatual do canal e, em segundo lugar,que o banco de areia na foz do lagotenha sido, de fato, depositado pelorio em conseqüência de eventosgeológicos remotos, como sugeriu
Aprile Darwich&
Pro
fundid
ade
do
lago
Tupé
(m)
13
16
19
22
25
28
31
(ano)
Nív
eld
'água
rio
Negro
(metr
os
a.n
.m.)
-1,5
1,5
4,5
7,5
10,5
13,5
16,5nível máximo 2003rio = 28,27m a.n.m.
lago = 14,5m
Nível d'água rio Negro Nível d'água lago Tupé
nível mínimo 2002rio = 17,19m a.n.m.
lago = 4,5m
out/02
nov
dez
jan/0
3
fev
mar
abr
mai
jun jul
ago
set
out
nov
dez
jan/0
4
fev
mar
nível mínimo 2003rio = 19,01m a.n.m.
lago = 4,5m

7
Fittkau (1974).Segundo Riccomini (2000) do
ponto de vista geológico, a morfologiados canais é o principal atributoconsiderado na classificação dossistemas hídricos. A morfologia doscanais lacustres e fluviais é controladapor uma série de fatores autocíclicoscomo a carga de sedimentostransportada, largura e profundidadedo canal, velocidade do fluxo,declividade, rugosidade do leito ecobertura vegetal nas margens e,alocíclicos como as variáveis climáticas e geológicas. No Tupé, esseseventos são bem definidos e têminfluência marcante sobre a hidroquímica do lago. Embora com decliveacentuado em direção ao rio, mas comvazão reduzida, o canal estreito epouco escavado contribui para oisolamento do lago. As margensíngremes e a floresta circundante oprotegem dos ventos dificultando acirculação da coluna de água. A cargade sedimentos é consideravelmentereduzida e caracterizada, ora pelomaterial do rio Negro, quando as águasdesse rio predominam no lago, ora pelomaterial transportado pelos igarapésde floresta que caracterizam o lago nosperíodos de enchente e vazante, emespecial no período de águas baixas(início da enchente ou final davazante). Em todas as situações omaterial em suspensão é muito fino esua quantidade extremamente reduzida, em torno de 10mg.L- . Esta situa-ção contribui para a elevada transpa-rência da água, que é maior no períodode isolamento do lago (Secchi de até1,5m) quando toda a massa aquática
et al.
-
-
-1
Figura 2. Perfil longitudinal do lago Tupé para o período de seca2003 – 2004.
Apesar da Bacia Amazônica consistir de umatopografia relativamente plana, com fracos declives, osformadores do rio Negro nascem em áreas acidentadasdo território nacional (Planalto das Guianas), a 2.100mde altitude (Cunha Guerra, 1998). Essa característicacontribui para impedir o acúmulo do material
&
encontra-se com luz fotossintéticamente ativa.O fluxo e o material sedimentar são dois elementos
fundamentais na estruturação do sistema de geometriahidráulica em cursos aluviais (Christofoletti, 1997). Cadaum desses elementos pode ser caracterizado a partir devariáveis mensuradas nas seções transversal,longitudinal e diagonal do lago, dando subsídios para aapresentação de um modelo mais confiável damorfologia do sistema lacustre estudado.
Entre a linha da costa, margem esquerda do rioNegro, e o ponto mais interno do lago, acima dosigarapés da Helena (EI2) e das Pedras (EI3), se estendeuma zona de transição, cujo gradiente batimétricoaumenta de forma irregular em direção ao rio (Fig. 2),tendo como trecho mais profundo a área que vai daconfluência entre os braços maior (longitudinal) e menor(transversal) do lago, onde está localizada a estação demonitoramento denominada estação central (ET10), e oponto mais próximo do canal de conexão com o rioNegro.
Modelos geomorfológicos para o Lago Tupé
0
-1
-2
-3
-4
-5
-6
-0,69
-3,60
-5,63
-0,71 (ET1)rio Negro
Pro
f.(M
)
ET5
ET10
decliv. max.

8
sedimentar mais fino ao longo do leito principal do rioNegro.
A mobilização de sedimentos lacustres dependediretamente da velocidade orbital (velocidade dedesprendimento e transporte), da granulometria, pesoespecífico, forma dos sedimentos e ângulo de inclinaçãodo sistema (declividade). O alto declive do lagoproporciona um aumento da velocidade das águas,principalmente das camadas mais profundas, originandoum padrão de transporte sedimentar, com áreas deinundação alimentadas pelas águas do rio durante osperíodos de enchente e cheia. A declividade total dolago, calculada para o trecho localizado entre os pontosmais extremos deste, como mostra a Figura 2, foi de 1,9m.km . A declividade máxima, observada entre ospontos de amostragem ET1 e ET5, foi de 4,1 m.km . Emlocais sem um aporte significativo de sedimentoslamosos, oriundos dos igarapés de floresta e solosmarginais, a mobilização dos sedimentos envolve otransporte predominantemente de sedimentosarenosos, como ocorre no rio Negro e no canal deconexão com o lago.
As lamas são depositadas nas profundidades em que avelocidade de corrente não mais exerce sua ação deselecionamento sedimentar. O ponto máximo desedimentação das partículas finas no lago Tupé temocorrido nas proximidades da estação ET10, onde seencontra a maior profundidade do lago. Um maiordetalhamento dos processos que envolvem a dinâmicasedimentar lacustre é abordado em Aprile . (2005,neste volume).
Considerando o canal principal de escoamento de umlago, este em decorrência da sua formação geológica eda ação das águas, pode apresentar três formas padrãode seção transversal: o tipo parabolóide, trapezoidal etriangular (Fig. 3). Em regiões de alta declividade, há umpredomínio dos canais trapezoidais. Já em regiões combaixa energia potencial, o predomínio são das seçõestriangulares. Os resultados da análise morfométricarevelaram que o lago Tupé, apresenta uma seçãotransversal com padrão tendendo ao triangular, comformato de cunha irregular ou “V”, deslocado para a
-1
-1
et al
margem direita como mostra a Figura4. A declividade mais acentuada namargem direita está diretamenteassociada a formação geológica daregião. O lago encontra-se inseridoentre dois vales, com solos argilosos eareno-argilosos e uma fina camada deglay húmico nas direções norte enoroeste, além de uma extensaformação arenosa (bancos de areia) nolimite da sua foz com o rio Negro.Segundo Fittkau (1974), durante oPleistoceno havia uma imensa área decachoeiras ao norte do lago, de ondegrande quantidade de areia teria sidotransportada em direção ao rio, sendodepositada, principalmente, ondehoje é a foz do lago Tupé. De acordocom o autor ainda, a deposição dessematerial teria sido decisiva naformação de parte da região ocidentaldo rio Negro. Considerando-se estaafirmação pode-se observar que a áreade deposição referida estende-se,subindo o rio Negro pela margemesquerda, desde a praia da PontaNegra passando pela foz dos igarapésTarumã e Tarumã-Mirim até o lagoTupé. Além disso, no mesmo trecho dorio, pela margem direta, há extensasfaixas de deposições arenosas seme-lhantes às da margem esquerda, talcomo também registrado à montantedessa área do rio Negro.
A formação em cunha pressupõeque no passado, antes da deposição deareia na foz do lago, a região do lagoapresentava alta velocidade decorrente, devido a um grande desnívelno relevo. Em decorrência da grandevelocidade de fluxo, os processoserosivos eram intensos, com grande
Aprile Darwich&

9
modelo, que estão representadas na Figura 4. A primeiraunidade denominada interflúvio e declive cominfiltração, refere-se aos processos pedogenéticosassociados com movimento vertical da água superficiale, com a eluviação mecânica e química pelo movimentolateral da água subsuperficial. A segunda é a escarpa,cuja inclinação acentuada e abrupta favorece ointemperismo mecânico e químico. A terceira unidade édefinida como declive intermediário, onde pode ocorrertransporte de material pelos movimentos coletivos dossolos, ação da água e formação de terracetes. O quarto éo sopé coluvial, onde ocorre reposição de material pelosmovimentos coletivos e escoamento superficial,transporte de material, reptação e formação de cones dedejeção. Por fim, a quinta unidade é formada por umdeclive aluvial com deposição aluvial, processosoriundos do movimento da coluna de água associado aotransporte de material para jusante em direção a Et10.
Figura 4. Modelo de seção transversal do lago Tupé para operíodo de seca 2003/2004 e os processos geomorfológicosdominantes. (A) interflúvio e declive com infiltração; (B)escarpa; (C) declive intermediário; (D) sopé coluvial e (E)declive aluvial e leito central.
quantidade de material clástico sendoretirado e transportado das margens,moldando o relevo atual. Ainda hoje seencontra pequenas cachoeirasinterligadas aos igarapés de floresta,com leito de rochas sedimentaresmoldadas pela ação das águas. RaiHill (1981) sugerem que a morfologiado lago Tupé, que segundo os autorestem um aspecto côncavo, é resultadode duas frentes: a expansão fluvial queelevou os níveis do leito do rio,provocando um aumento dos processosde sedimentação, associada a forçados ventos. Segundo os autores aformação do lago é de um períodogeologicamente recente.
&
Figura 3. Formatos padrão de seçãotransversal de um lago. (A) parabolóide,(B) trapezoidal e (C) triangular.
De forma geral, os vários modelosp ropos to s em geomor fo log i aapresentam relações que envolvemrelevo, geologia e clima, assim comosuas interações com o intemperismo,os solos, processos erosivos e dinâmicahídrica.
A partir do modelo de vertenteapresentado por Dalrymple .(1968) foi esquematizado cinco dasnove unidades hipotéticas descritas no
et al
No aspecto geomorfológico, os igarapés apresentamvariações no relevo. O igarapé da Helena, o mais àmontante, e o igarapé Terra Preta, localizado no braçotransversal, apresentam um formato próximo ao modelotrapezoidal, com uma inclinação mais acentuada namargem esquerda. Já os igarapés das Pedras, Cachoeira
Modelos geomorfológicos para o Lago Tupé
A
A
A
BB
CC
D DE
B
C
d
Margem EsquerdaMargem Direita
d
d

10
e Monteiro, diferentemente, apresentam umamorfologia que tende ao modelo triangular, comdeformações. O igarapé da Cachoeira é o que melhor seadequa a esse modelo, apresentando seções íngremespróximo ao leito e interflúvio igualmente inclinado nasmargens (Figs. 5a-5e).
O lago Tupé tem um papel de escoadouro das águas esedimentos em suspensão provenientes dos igarapés defloresta para o rio Negro durante os períodos de vazantee seca. Já nos períodos de enchente e cheia, o lagotorna-se um acumulador de águas pretas provenientes dorio Negro. Essa dinâmica fluvio-lacustre é grandementefavorecida pela geomorfologia da região.
A Tabela 1 apresenta os resultados da análisemorfométrica do lago Tupé. O lago Tupé apresenta umformato em "t", com um braço maior ou longitudinal deaproximadamente 2.504m e um braço menor outransversal de 1.149m de comprimento. A larguramáxima foi observada exatamente na confluência dosdois braços, com aproximadamente de 211 a 250m. Alargura mínima foi determinada no extremo oposto dafoz do lago, acima da estação ET1, com valores próximosde 32m. A partir dos resultados da análise morfométricado lago Tupé chegou-se a um valor aproximado da área doespelho de água de 66,9ha, e um volume para o períodode seca de aproximadamente 1.440.260m . Rai Hill(1981), estudando alguns parâmetros morfométricospara o lago ao longo de um ciclo hidrológico completo,encontraram uma área aproximada de 68ha e um volume2.570.000m , sendo este volume baseado naprofundidade média para o período de cheia.Comparando os dois resultados, observa-se que emtermos de área, o lago não varia muito entre a seca e a
AnálisesAreal e Batimétrica
3
3
&
cheia, contudo, o volume aumenta nomesmo período cerca de 56%. Essefenômeno é característico de lagosencrostados em regiões de vale. Dessemodo, não é possível obter umarelação direta entre a área e o volume,sendo mais confiável uma relaçãoentre este último com a profundidademédia.
Apesar dos processos morfodinâmi-cos atuais não serem intensos, fatocomprovado pela baixa dissipaçãoenergética proveniente das ondas eventos, e pela reduzida troca desedimentos entre a praia, localizadano canal, e a zona submersa do lago, aprofundidade do lago é bastantevariada, fundamentalmente pelainfluência do pulso de inundação.Durante o período de seca aprofundidade média do lago é de 2,1m,enquanto que na cheia fica em tornode 11,1m. Na estação ET10, aprofundidade varia dentro de um ciclohidrológico com mínima de 5,1 noperíodo de seca e máxima de 15mdurante a cheia.
A partir da elaboração detransectos ao longo da bacia do lagoTupé (Fig. 6), foi realizada a análisebatimétrica do lago, dos igarapésHelena, das Pedras, da Cachoeira,Monteiro e Terra Preta, além do canalde conexão com o rio Negro,apresentados nas Figuras 7 e 8,respectivamente.
Aprile Darwich&

11
Modelos geomorfológicos para o Lago Tupé
Figura 5. Modelo de seção transversal para osprincipais igarapés constituintes do lago Tupé para operíodo de seca 2003/2004. (M.E. = margemesquerda; M. D. = margem direita)
Igarapé da Helena
Igarapé do MonteiroIgarapé da Cachoeira
Igarapé Terra Preta
Igarapé das PedrasM.E.
M.E.M.E.
M.E.
M.E.M.D.
M.D.M.D.
M.D.
M.D.

12
Tabela 1. Medidas morfométricas do lago Tupé obtidas durante o período de seca 2003/2004.
Setor do Lago Setor dos Igarapés
comprimento máximo (m) 2.504 EI2 comprimento máximo (m) 117,4
comprimento mínimo (m) 1.149 EI2 largura média (m) 29,6
largura máxima (m) 250,1 EI2 profundidade média(m) 0,4
largura mínima (m) 32,4 EI3 comprimento máximo (m) 162,1
largura média (m) 97,4 EI3 largura média (m) 36,4
diagonal maior (m) 1.014 EI3 profundidade média(m) 0,3
diagonal menor (m) 148,5 EI4 comprimento máximo (m) 274
EI4 largura média (m) 37,8
profundidade máxima (m) 5,6 EI4 profundidade média(m) 0,6
profundidade mínima (m) 0,1 EI6 comprimento máximo (m) 257,8
profundidade média(m) 2,1 EI6 largura média (m) 50,9
profundidade relativa (m) 2,2 EI6 profundidade média(m) 0,6
EI8 comprimento máximo (m) 157,9
declividade total (m/km) 1,9 EI8 largura média (m) 18,1
declividade máxima (m/km) 4,1 EI8 profundidade média(m) 0,6
talude da seção ET10 14,9 Setor do Canal (EC11)
área da seção ET10 (m2) 383,7 comprimento máximo (m) 315
perímetro molhado ET10 (m) 151,4 largura máxima (m) 41,2
raio hidráulico ET10 (m) 2,5 largura mínima (m) 15,1
largura média (m) 26,5
área de superfície (ha) 66,9 profundidade máxima (m) 1,4
volume estimado (m3) 1.440.260 profundidade mínima (m) 0,1
tipo de bacia "Ria" profundidade média(m) 0,5
Aprile Darwich&

13
Figura 6. Mapa da bacia do lago Tupé com os transectos aplicados para as análises morfométrica ebatimétrica.
Modelos geomorfológicos para o Lago Tupé
c9
8 43 2
Canal700 x
TSR
Q
EI3
EI4
EI2
aa
a
bb
a
ba
b
B
P
0 135 270m
O
N
SL
K
EI6
1
2
J
I HG
FE
D
EI8
CBA
Canal
NM
c7
c6c5
c1
W E

-0,80-1,20
-3,22
-5,63
-2,68
-5,53
-2,91
-6
-5
-4
-3
-2
-1
0P
rof.
(m)
AB
-5,61
-2,45
-1,13
-0,28
-5,14
-0,95
-2,55-2,10
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
CD
-5,08
-2,68
-0,18-1,10
-5,00
-3,39-3,65
-1,70
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
EF
-3,76
-2,19
-1,16
-0,98-1,21
-1,12
-0,92
-3,21
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
KL
-0,88-1,21-1,11
-0,87-0,69-0,41
-3,60
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)MN
-0,35 -0,47-0,61
-2,02
-1,17
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
OP
M.E. M.D. M.E. M.D.
-2,16
-1,51
-3,68
-5,00
-1,80-2,61
-1,21
-4,81
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
GH
-0,48 -0,79
-2,61
-4,15
-0,87
-3,00
-1,16
-3,60
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
IJ
-0,21-0,75-0,62-0,35
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
QR
-0,70-0,44-0,33 -0,61
-6
-5
-4
-3
-2
-1
0
Pro
f.(m
)
ST
Figura 7 Análise batimétrica dolago Tupé (período de seca2003/2004).
.
(M.E. = margemesquerda; M. D. = margem direita)
14
Aprile Darwich&

-0,41-0,54
-0,12 -0,22
-0,84
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
Pro
f.(m
)
a
b
EI2
-0,06 -0,34 -0,36
-0,53
-0,38
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
Pro
f.(m
)
a
b
EI3
-0,05
-0,66-0,78
-0,72-0,61
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
Pro
f.(m
)
a
b
EI4
-2,04
-0,23-0,04
-0,74-0,67
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
Pro
f.(m
)
a
b
EI6
-0,3-0,3-0,1
-0,9
-0,5-0,5
-1,4
-1,1
-0,5
-0,1
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
M. E. M. D.
Pro
f.(m
)
EC11
-1,02
-0,52-0,08
-0,18
-1,42
-2,1
-1,8
-1,5
-1,2
-0,9
-0,6
-0,3
0,0
M. E. M. D.
Pro
f.(m
)
a
b
EI8
Figura 8 Análise batimétrica dos igarapés de floresta e do canal do lago Tupé (período de seca2003/2004).
.(M.E. = margem esquerda; M. D. = margem direita)
15
Modelos geomorfológicos para o Lago Tupé

16
Aprile Darwich&
Geol. Sur. Prof. Paper
U. S. Geol. Sur. Prof.
Paper .
Geol. Soc. America Bulletin
.Geoecologia,
hidrografia, hidroquímica, clima e
processos antrópicos da bacia do rio
Jacaré Pepira - SP
Revista Brasileira
de Geografia
Revista Brasileira
de Geografia
Int.
Revue ges. Hydrobiol
Decifrando a Terra
.
Revista Brasi leira de
Geografia
XVI Congresso
Internacional de Geografia
.
252, Washington,EUA.
Leopold, L. B.; Wolman, M. G. 1957. Riverchannel patterns: braided, meanderingand straight.
282 B, Washington, EUA p. 39-85.
Leopold, L. B.; Wolman, M. G. 1960. Rivermeanders.71 p. 769-794.
Maier, M. H. 1983.
. Tese de Doutorado,Universidade Federal de São Carlos, SãoCarlos, SP, 303pp.
Martonne, E. de 1943. Problemasmorfológicos do Brasil tropicalatlântico (1 parte).
, Rio de Janeiro, 5(4):532-550.
Martonne, E. de 1944. Problemasmorfológicos do Brasil tropicalatlântico (2 parte).
, Rio de Janeiro, 6(2):222-238.
Rai, H.; Hill, G. 1981. Physical andchemical studies of lago Tupé; a CentralAmazonian Black Water,, Ria Lake.
., 66(1):37-82.Riccomini, C., Giannini, P. C. F.; Mancini, F.
2000. Rios e processos aluviais. In:Teixeira, W.; Toledo, M. C. M.;Fairchild, T. R.; Taioli, F. (eds.).
. , Oficina de TextosSão Paulo p. 191-214.
Ruellan, F. 1944. Evolução geomorfológicada baía de Guanabara e das regiõesviz inhas.
, 6(4):445-508.Ruellan, F. 1950. Les surfaces d’erosion de
la région sud-orientale du plateaucentral brésilien. In:
, Lisboa,Portugal p.659-673.
o
o
Bibliografia Citada
Ab’Saber, A. N. 1956. Etat actuel dês connaissances sur lesniveaux d’erosion et les surfaces d’aplanissement du Brésil.In: 18 , Rio de Janeiro1956, Report New York, Union Geographie Internationale,5:7-27.
Ab’Saber, A. N. 1958. A geomorfologia no Brasil., Campinas, São Paulo, 1(2):1-8.
Ab’Saber, A. N. 1964. O relevo brasileiro e seus problemas. In:, vol. I, Cia. Editora Nacional, São
Paulo p.135-250.Aprile, F. M., Darwich, A. J.; Raposo, J. C. 2005. Considerações
sobre a geoquímica e dinâmica sedimentar do lago Tupé. In:Santos-Silva, E. N.; Aprile, F. M.; Scudeller, V. V.; Melo, S.(Orgs.),
. EditoraINPA, Manaus,AM.
Bigarella, J. J. 1961. Considerações sobre a geomorfogênese daSerra do Mar no Paraná. ,Curitiba, PR, (4-5):94-125.
Bigarella, J. J. 1964. Variações climáticas no Quaternário esuas implicações no revestimento florístico no Paraná.
, Curitiba, PR, (10-11):211-213.
Carvalho, L. 1965. Curso de hidrobiologia e barragens: cheias.Fortaleza, Universidade Federal do Ceará, 64pp.
Christofoletti, A. 1977. As tendências atuais da geomorfologiano Brasil. , Campinas, São Paulo,17(33):35-91.
Christofoletti, A. 1978. Aspectos da análise sistêmica emgeografia. , São Paulo, 3(6):1-31.
Christofoletti, A. 1997. . 5 reimpressão, EditoraEdgard Blücher Ltda, São Paulo, 188pp.
Costa, A. F. 1991. ,UFRPE, Imprensa Universitária, Recife 297pp.
Cunha, S. B. da; Guerra,A. J. T. (orgs.) 1998.. , Bertrand Brasil ed., Rio de Janeiro 392pp.
Dalrymple, J. B.; Blong, R. J.; Conacher, A. J. 1968. Ahypothetical nine unit land surface model.
, 12(1):60-76.Fittkau, E. J. 1974. Zur ökologischen Gliederung Amazoniens.
1. Die erdgeschichtliche Entwicklung Amazoniens., 5:77-134.
Freitas, R. O. 1951. Ensaio tectônico do Brasil., 13:171-222.
Leopold, L. B.; Maddock, T. 1953. The hidraulic geometry ofstream channels and some physiographic implications.
Congrés International de Geographie
Notícia
Geomorfológica
Brasil: a terra e o homem
.
BioTupé: meio físico, diversidade biológica e
sociocultural do baixo rio Negro, Amazônia central
Boletim Paranaense de Geografia
Boletim Paranaense de Geografia
Notícia Geomorfológica
GeografiaGeomorfologia
Introdução à ecologia das águas doces
Geomorfologia do
Brasil
Zeitschrift für
Geomorphologie
AmazonianaRevista
Brasileira de Geografia
U. S.

17
Modelos geomorfológicos para o Lago Tupé
Ruellan, F. 1952. Alguns aspectos do relevono planalto central do Brasil.
, Na., 2:17-28.Sugu io , K . 1973 .
. E d . E d g a r dBlucher/EDUSP, São Paulo 110p.
Assoc.
Geog. BrasilI n t rodução à
s e d i m e n t o l o g i a
.
Suguio, K. 1994.. 4 ed., , Editora Edgard
Blücher, São Paulo 500p.Wolman, M. G.; Leopold, L. B. 1957. River flood plains:
some observations on their formation.282 C, Washington, EUA, p. 87-109.
Rochas sedimentares. propriedades,
gênese, importância econômica
.
U. S. Geol. Sur.
Prof. Paper


Variáveis limnológicas: contribuição aoestudo espaço-temporal de águas pretasamazônicas
RESUMO - Durante o ciclo hidrológico 2002/2003 foram realizadas amostragens em 12 pontos decoleta ao longo da bacia do lago Tupé, incluindo os igarapés mais importantes da bacia, o meio dacalha do rio Negro e o canal de conexão entre o rio e o lago. Foram mensuradas algumas variáveisambientais, como: transparência, temperatura da água, saturação do oxigênio, oxigênio dissolvido,condutividade elétrica e pH. Com o objetivo de evidenciar as diversas características espaço-temporais presentes no lago, foram aplicadas técnicas de Análise de Agrupamento através daligação pela média não ponderada com o coeficiente de associação da distância euclidiana aoquadrado. As maiores amplitudes de variação dos parâmetros limnológicos ocorreram durante aseca. Esta fase destacou-se ainda por apresentar as maiores concentrações de oxigênio dissolvido,em oposição aos períodos de cheia e vazante. De modo geral, os igarapés de floresta apresentaramum comportamento bastante homogêneo, principalmente com relação à temperatura da água(próxima de 28,6ºC na seca e 28,5ºC na cheia) e condutividade (5,9 S.cm na seca e 10,3 S.cm na� �
-1 -1
Meio FísicoCapítulo 2
Assad José DARWICH
Fábio Marques APRILE
Barbara Ann ROBERTSON
Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Biologia AquáticaAv. André Araújo 2936, Caixa Postal 470 Manaus, AM, 69060-001
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

Darwich et al.
20
cheia). Os resultados da análise de agrupamento mostraram que a estação localizada no rio Negroisolou-se das demais estações, revelando um comportamento hidroquímico diferenciado.
PALAVRAS-CHAVE: limnologia,Amazônia Central, Rio Negro, águas pretas, lago Tupé.
Introdução
Grande parte dos 7,5 milhões de km da BaciaAmazônica está situada na zona equatorial quente eúmida, com imensa heterogeneidade na vegetação,geologia, clima e na qualidade das águas naturais. Oslagos dessa região são influenciados pelo suprimentoperiódico de substâncias solúveis e de matériainorgânica particulada (Darwich, 1995), comparticipação decisiva do pulso de inundação como forçacontroladora da biota (Junk ., 1989). Variaçõesespaço-temporais, ciclos sazonais e diários de variáveislimnológicas em lagos amazônicos podem apresentargrande variação e o conhecimento dessas variações podecontribuir para o entendimento de padrões de mistura eestratificação nesses ambientes. A dinâmica dessesprocessos é responsável pela distribuição de calor,nutrientes, íons e gases dissolvidos na coluna de água.Por isso, no estudo dessas variáveis é importanteconsiderar a fase do ciclo hidrológico, a hora do dia, olocal e a profundidade da medida, bem como ascondições climáticas e a localização geográfica doambiente.
Estudos sobre mudanças sazonais e diárias devariáveis físico-químicas e químicas em lagos de várzeaamazônicos, têm mostrado a imensa variabilidadehidroquímica entre eles e a importância do pulso deinundação no comportamento dessa variabilidade(Schmidt, 1972a,b; Junk, 1973; Furch ., 1983;
Melack Fisher, 1983; Furch, 1984a,b; Darwich, 1995;
Kern Darwich, 1997; Darwich ., 2003).Particularmente, lagos amazônicos de áreas inundáveisna bacia de rios de água preta estão submetidos à forçase interações semelhantes. No entanto, a resposta do
2
et al
et al
et al
&
&
ambiente à estas forças parece
também ter relação com a localização
do lago em relação a margem do rio e a
proteção dos ventos, o que pode
implicar em padrões diferenciados de
mistura da coluna de água. Dessa
forma, lagos da bacia do rio Negro
estão sujeitos à estas condições, além
da variação de nível do rio e da
afluência pela bacia lacustre. A
contribuição de pequenos igarapés de
floresta e da precipitação pluviométri
ca sobre toda a bacia do lago, exerce
maior ou menor importância de acordo
com a fase do ciclo hidrológico.
Estudos limnológicos na bacia do lago
Tupé têm mostrado ser este um lago
com reduzida concentração de íons
dissolvidos, baixos valores de pH e de
condutividade elétrica, de difícil
circulação e com estratificação
térmica e de oxigênio, pelo menos em
alguma fase do ciclo (Reiss, 1977; Rai
Hill, 1981; Darwich ., 2003).Com base nesse conhecimento e no
fato de ser o Tupé um lago de área
inundável com margens íngremes,
submetido durante a maior parte do
ciclo à flutuação de nível do rio Negro,
estudou-se a sua variação hidroquími-
ca de forma a registrar padrões sazo-
nais de variáveis limnológicas durante
um ciclo hidrológico.
-
et al
&

21
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas
Figura 1: Mapa do lago Tupé (Manaus, AM) com ospontos de amostragem.
Tabela 1: Descrição dos pontos de amostragem no lago Tupé,igarapés e rio Negro com suas respectivas coordenadasgeográficas.
ET1 03º01'33,5"S;60º15'58,0" W lago Tupé cabeceira maior
EI2 03º01'45,4"S;60º15'50,3" W igarapé da Helena
EI3 03º01'47,7"S;60º16'1,6" W igarapé das Pedras
EI4 03º01'58,7"S;60º16'1,2" W igarapé da Cachoeira
ET5 03º02'17,5"S;60º15'44,0" W lago Tupé entre Cachoeira e Monteiro
EI6 03º02'26,0"S;60º15'42,0" W igarapé do Monteiro
ET7 03º02'17,1"S;60º15'45,3" W lago Tupé cabeceira menor
EI8 03º02'10,3"S;60º14'57,5" W igarapé Terra Preta
ET9 03º02'28,0"S;60º15'12,0" W lago Tupé braço menor
ET10 03º02'35,4"S;60º15'17,5" W lago Tupé estação central
EC11 03º02'47,8"S;60º15'12,8" W canal de conexão
EN12 03º02'46,5"S;60º15'13,1" W rio Negro em frente ao lago
Ponto Coordenadas Descrição
Material e Métodos
Foram realizadas amostragens em12 pontos de coleta distribuídos daseguinte forma: cinco no lago, uma nadesembocadura de cada um dos cincoigarapés mais importantes da bacia,uma no canal de ligação entre o rio e olago e outra no meio da calha do rioNegro, à montante do canal do lago(Fig. 1 e Tab. 1). As amostragens foramrealizadas trimensalmente, conside-rando os períodos de seca (dezem-bro/2002), enchente (março/ 2003),cheia (junho/03) e vazante (setem-bro/03).
As amostras de água foramcoletadas com uma garrafa de Ruttner.A transparência da água foi medidacom um disco de Secchi ( = 30cm), eos resultados foram utilizados para ocálculo da zona eufótica. As determinações limnológicas tiveram por baseas recomendações do ProgramaBiológico Internacional para ambientes aquáticos (Golterman ., 1978;Wetzel & Likens, 2000). As medidas detemperatura (°C), saturação dooxigênio (%) e oxigênio dissolvido(mg.L ) foram obtidas através deleitura direta com um oxímetro digitalmodelo Oxi-197. A condutividadeelétrica (mS.cm ) e o potencial hi-drogeniônico foram medidos compotenciômetros digitais WTW LF-197 epH-197, respectivamente. As medidasno perfil vertical foram realizadas acada 0,5m nos primeiros cinco metros,e depois de metro em metro até ofundo. Foi aplicada a Análise deAgrupamento (Cluster), com oobjetivo de evidenciar as diversas
�
-
-
et al
-1
-1
características espaço-temporais presentes no lago. Ométodo empregado foi o agrupamento através daLigação pela Média Não Ponderada, descritoprimeiramente por Sneath Sokal (1973) Statsoft(1997), com o coeficiente de associação da distânciaeuclidiana ao quadrado.
& apud

22
Resultados e Discussão
A profundidade do lago Tupé varia de acordo com onível do rio Negro. Quando o nível do rio está abaixo dacota 19m a.n.m., não há qualquer influência deste sobreo lago, sendo que as águas que correm pelo canal deconexão passam a ser exclusivamente oriundas doecossistema lacustre, procedentes dos igarapés defloresta. Apesar da vazão da água do lago para o rio, onível do lago permanece entre 4,5 e 5,0m no período deseca. Nesta época, a pequena variação na profundidadedo lago é o resultado do equilíbrio entre a precipitação ea contribuição de pequenos igarapés da bacia do lago
a evaporação e o escoamento pelo canal deligação com o rio. No pico da cheia (cota do rio em tornode 29m a.n.m.) a profundidade do lago na estaçãocentral varia entre 14 e 15m.
Fundamentalmente, essas modificações ambientaissão responsáveis pelas principais mudançashidroquímicas no lago Tupé. O pulso de inundaçãoinfluencia diretamente a transparência da água do lago,a qual pode atingir valores de até 1,5m em uma coluna deágua de 4,5m. Dessa forma, o lago apresenta uma zonaeufótica assim como os igarapés, nos quais atransparência eqüivale à profundidade máxima, com0,6m em média. Na cheia os igarapés apresentaramprofundidade média de 10m com transparência entre 1,1e 1,3m, enquanto que o lago e o rio apresentamtransparências equivalentes e em torno de 1,0 m.
Os resultados da análise limnológica são referentes amedições realizadas durante os períodos de enchente(março), cheia (junho), vazante (setembro) e seca(dezembro) do ciclo 2003. Os resultados apresentadosreferem-se às medidas de tendência central na coluna deágua para as variáveis temperatura, saturação dooxigênio, oxigênio dissolvido, condutividade elétrica epH (Figs. 2 a 6 e Tabs. 2 a 6).
As temperaturas médias determinadas ao longo dasestações do lago Tupé e rio Negro para o ciclo hidrológico2002/2003 estão apresentadas na Tabela 2 e Figura 2. Naseca a temperatura variou de 24,8ºC a 32,0ºC, sendo que
versus
a temperatura das águas dos igarapésficou sensivelmente abaixo datemperatura das águas do lago. Amaior variação nesse período foidetectada na estação central (ET10)com uma amplitude de 4,5ºC. Apesardisso, os desvios padrão ficaramabaixo de 1,6, mostrando que talamplitude não foi significativamentemarcante. Na fase de enchente atemperatura variou entre 27,8ºC e30,9ºC, uma diferença de 3,1ºC. Amaior amplitude foi de 3,0ºC,detectada na estação ET1, e a menor0,1ºC na estação localizada no rioNegro (EN12). Na cheia a temperaturamédia da coluna de água variou de24,4ºC a 32,0ºC. Nesse período demaior contribuição das águas do rioNegro para o lago, ocorreram asmaiores amplitudes médias datemperatura. A maior diferença entrea máxima e a mínima foi detectada noigarapé das Pedras (EI3), com umaamplitude de 6,3ºC. A menoramplitude foi determinada no rioNegro, com temperatura média de30,0ºC ± 0,1, mostrando um padrãobastante homogêneo de suas águaspara esta variável. Na vazante atemperatura variou de 27,2ºC a33,6ºC. A maior variação pontual6,1ºC, foi detectada na estação EC11,no canal que liga o lago ao rio Negro,seguida pela estação central com5,6ºC. Considerando o ciclo hidrológi-co como um todo, as temperaturasmais elevadas foram determinadasdurante a fase de enchente, seguidapela fase de vazante.
O oxigênio dissolvido indica o graude arejamento da água. É um impor-
Darwich et al.
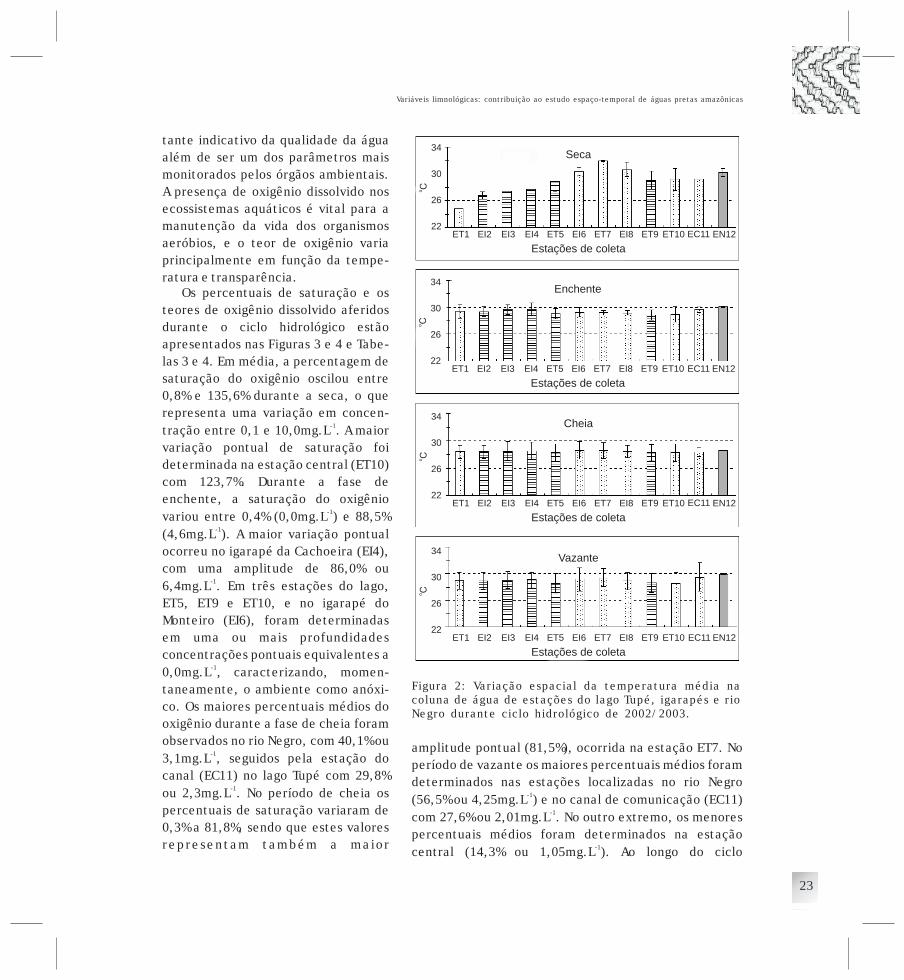
23
tante indicativo da qualidade da águaalém de ser um dos parâmetros maismonitorados pelos órgãos ambientais.A presença de oxigênio dissolvido nosecossistemas aquáticos é vital para amanutenção da vida dos organismosaeróbios, e o teor de oxigênio variaprincipalmente em função da tempe-ratura e transparência.
Os percentuais de saturação e osteores de oxigênio dissolvido aferidosdurante o ciclo hidrológico estãoapresentados nas Figuras 3 e 4 e Tabe-las 3 e 4. Em média, a percentagem desaturação do oxigênio oscilou entre0,8% e 135,6% durante a seca, o querepresenta uma variação em concen-tração entre 0,1 e 10,0mg.L . A maiorvariação pontual de saturação foideterminada na estação central (ET10)com 123,7%. Durante a fase deenchente, a saturação do oxigêniovariou entre 0,4% (0,0mg.L ) e 88,5%(4,6mg.L ). A maior variação pontualocorreu no igarapé da Cachoeira (EI4),com uma amplitude de 86,0% ou6,4mg.L . Em três estações do lago,ET5, ET9 e ET10, e no igarapé doMonteiro (EI6), foram determinadasem uma ou mais profundidadesconcentrações pontuais equivalentes a0,0mg.L , caracterizando, momen-taneamente, o ambiente como anóxi-co. Os maiores percentuais médios dooxigênio durante a fase de cheia foramobservados no rio Negro, com 40,1% ou3,1mg.L , seguidos pela estação docanal (EC11) no lago Tupé com 29,8%ou 2,3mg.L . No período de cheia ospercentuais de saturação variaram de0,3% a 81,8%, sendo que estes valoresrepresentam também a maior
-1
-1
-1
-1
-1
-1
-1
amplitude pontual (81,5%), ocorrida na estação ET7. Noperíodo de vazante os maiores percentuais médios foramdeterminados nas estações localizadas no rio Negro(56,5% ou 4,25mg.L ) e no canal de comunicação (EC11)com 27,6% ou 2,01mg.L . No outro extremo, os menorespercentuais médios foram determinados na estaçãocentral (14,3% ou 1,05mg.L ). Ao longo do ciclo
-1
-1
-1
Figura 2: Variação espacial da temperatura média nacoluna de água de estações do lago Tupé, igarapés e rioNegro durante ciclo hidrológico de 2002/2003.
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas
Seca
Estações de coleta
34
30
26
22ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Enchente
Estações de coleta
34
30
26
22ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Cheia
Estações de coleta
34
30
26
22ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Vazante
Estações de coleta
34
30
26
22ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12

24
ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
24,8 26,8 27,4 27,7 28,8 30,4 32 30,7 29,1 29,2 29,2 30,2
0 0,6 0 0 1,3 0,6 0,1 1,1 1,3 1,6 0 0,6
0 2,1 0 0 4,6 2,1 0,2 3,5 4,6 5,6 0 2,1
24,8 26,4 27,4 27,7 27,1 29,9 31,9 29,9 27,5 27,3 29,2 29,8
24,8 27,2 27,4 27,7 30,9 30,8 32 31,4 31,4 31,8 29,2 31,2
0 0,8 0 0 3,8 0,9 0,1 1,5 3,9 4,5 0 1,4
29,4 29,3 29,6 29,7 29 29,3 29,2 29,2 28,8 28,9 29,7 30,1
1 0,7 0,7 1 0,8 0,7 0,3 0,3 0,8 1,1 0,5 0,1
3,3 2,4 2,4 3,2 2,6 2,3 1,1 1,1 2,7 3,8 1,6 0,2
27,8 28,3 28,5 28,4 28 28,3 28,5 28,6 27,8 27,8 29 30
30,8 30,5 30,6 30,9 30 30 29,4 29,4 29,7 30,7 30,3 30,1
3 2,2 2,1 2,5 2 1,7 0,9 0,8 1,9 2,9 1,3 0,1
28,4 28,4 28,5 28,6 28,3 28,7 28,6 28,5 28,3 28,3 28,3 28,6
1 0,9 1,4 1,2 1,3 1,3 1,1 0,9 1,2 1,3 0,6 0
3,4 3,3 5,1 4,3 4,6 4,6 4 3,3 4,2 4,6 2,3 0,1
27,2 27,2 24,4 27,2 26,9 27,2 27,3 27,2 27 26,9 27,2 28,6
30,4 30,6 30,7 32 32 32 31,7 30,6 31 31,6 29 28,7
3,2 3,4 6,3 4,8 5,1 4,8 4,4 3,4 4 4,7 1,8 0,1
29 28,9 29 29 28,6 29,2 29,4 28,9 28,6 28,5 29,5 30
1,3 1,2 1,3 1,2 1,4 1,7 1,4 1,3 1,4 1,7 2,1 0,1
4,5 4,3 4,4 4,2 5 5,9 4,7 4,6 4,9 5,9 7,1 0,3
27,6 27,6 27,6 27,6 27,2 27,5 27,9 27,5 27,3 27,2 27,5 29,9
30,7 30,6 30,6 30,6 30,7 31,8 31,3 30,8 31,1 32,8 33,6 30,1
3,1 3 3 3 3,5 4,3 3,4 3,3 3,8 5,6 6,1 0,2
C.V. = coeficiente de variação
variação
desvio
C.V.
mínimo
máximo
variação
Vazante
média
desvio
C.V.
mínimo
máximo
variação
Cheia
média
desvio
C.V.
mínimo
máximo
variação
Enchente
média
desvio
C.V.
mínimo
máximo
Seca
média
Tabela 2: Variação da temperatura média na bacia do lago Tupé e rio Negro.
Darwich et al.

25
Figura 3: Variação espacial do percentual desaturação do oxigênio médio na coluna de água deestações do lago Tupé, igarapés e rio Negrodurante ciclo hidrológico de 2002/2003.
Figura 4: Variação espacial do oxigênio dissolvidomédio na coluna de água de estações do lago Tupé,igarapés e rio Negro durante ciclo hidrológico de2002/2003.
em oposição aos períodos de cheia e vazante, queapresentaram os menores teores dessa variável.
Os valores de condutividade elétrica medidosdurante o ciclo hidrológico estão apresentados na Figura5 e Tabela5. Em valores médios, durante a fase de seca acondutividade foi maior na estação ET1 com 8,6µS.cm ,e menor na estação ET7 com 3,7µS.cm . Esses valorescorresponderam também a maior amplitude de variaçãoda condutivida de no período. A maior variação pontualocorreu na estação ET10 com 4,6µS.cm . Os igarapés, de
-1
-1
-
-1
hidrológico as maiores amplitudes devariação ocorreram no período deseca, sendo que a maior variaçãopontual foi observada na estação ET10(123,7%) e, a maior variação espacialocorreu entre as estações ET7 e ET10,com uma amplitude de 134,8% ou9,9mg.L . O período de seca foimarcado por apresentar as maioresconcentrações de oxigênio dissolvido,
-1
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas
Seca
Estações de coleta
150
130
100
70
40
10ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Enchente
Estações de coleta
150
130
100
70
40
10ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Cheia
Estações de coleta
150
130
100
70
40
10ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Vazante
Estações de coleta
150
130
100
70
40
10ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Seca
Estações de coleta
16
12
8
4
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Enchente
Estações de coleta
16
12
8
4
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Cheia
Estações de coleta
16
12
8
4
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Vazante
Estações de coleta
16
12
8
4
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12

ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
85,5 99,4 91,6 96,5 78 105,9 135,5 109 83,7 65 92,2 97,9
0,1 6,9 0,6 0,5 29,4 0,8 0,1 4 17,5 52,1 1,4 8,2
0,1 7 0,7 0,5 37,7 0,8 0,1 3,6 20,9 80,1 1,5 8,3
85,4 94,5 91,1 96,1 40,4 105,3 135,4 106,2 61,4 0,8 91,2 90,3
85,5 104,3 92 96,8 111,3 106,5 135,6 111,8 103,7 124,5 93,2 109,2
0,1 9,8 0,9 0,7 70,9 1,2 0,2 5,6 42,3 123,7 2 18,9
46 45,5 53,7 50,6 30,7 34,9 46,5 43,8 31,6 25,8 42,8 82,2
23,8 32,6 24,1 36,7 31,2 35,6 22,5 24,3 31,3 30,1 13,7 1,2
51,8 71,6 44,9 72,5 101,4 102 48,5 55,4 99,1 116,6 32,1 1,4
16 2,7 16,4 1,9 0,5 0,5 4,2 12 0,5 0,4 22,5 80,7
78,5 78,7 80,1 87,9 73,4 76,5 61,8 88,5 67,9 67,9 58,1 83,4
62,5 76 63,7 86 72,9 76 57,6 76,5 67,4 67,5 35,6 2,7
17,6 23,5 22,5 20,1 18,8 21,9 25,1 20,1 20,7 17,8 29,8 40,1
21,8 22,2 25,7 26,9 24 29,1 24,6 23,9 26,5 23,9 24,3 0,2
123,7 94,4 114,4 133,5 127,9 133,1 98,1 119,3 128 134,2 81,5 0,5
1 0,9 0,9 0,9 1 0,9 0,3 0,7 0,7 0,8 0,8 39,9
66,7 72,6 67,7 80,2 73,2 81,1 81,8 76,5 77 74 59,1 40,4
65,7 71,7 66,8 79,3 72,2 80,2 81,5 75,8 76,3 73,2 58,3 0,5
19,7 19,3 21,3 18,8 17,4 20,5 25,6 19,9 16,3 14,3 27,6 56,5
27,5 28,6 27,7 26 26,4 29 35,3 31,1 26,5 25,8 32,8 0,4
139,3 148,2 130,4 138,8 151,9 141 137,7 156,6 162,6 180,3 118,8 0,7
1,2 1,2 1,3 1,3 1,2 1,2 1,2 1,2 1,2 1,1 1,2 55,8
67,2 69,5 63,5 61,2 62,5 68,5 80 77,2 74 74,7 80,6 56,9
66 68,3 62,2 59,9 61,3 67,3 78,8 76 72,8 73,6 79,4 1,1
C.V. = coeficiente de variação
máximo
variação
média
desvio
C.V.
mínimo
máximo
variação
Vazante
média
desvio
C.V.
mínimo
máximo
variação
Cheia
média
desvio
C.V.
mínimo
máximo
variação
Enchente
média
desvio
C.V.
mínimo
Seca
Tabela 3: Variação da percentagem de saturação do oxigênio média na bacia do lago Tupé e rio Negro.
26
Darwich et al.

27
ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
7,1 7,9 7,3 7,6 6 8 10 8,1 6,4 4,9 7,1 7,4
0 0,5 0 0 2,1 0 0 0,2 1,2 3,8 0 0,5
0,4 6,7 0,3 0,5 35,8 0,2 0,3 2,3 19,1 78,9 0,5 7,2
7,1 7,6 7,2 7,6 3,1 8 9,9 8 4,9 0,1 7,1 6,9
7,1 8,3 7,3 7,6 8,5 8 10 8,3 7,7 9,1 7,2 8,1
0 0,8 0 0 5,4 0 0 0,3 2,8 9,1 0 1,2
3,6 3,4 4 3,8 2,3 2,6 3,5 3,1 2,4 1,9 3,3 6,2
2 2,5 1,8 2,7 2,3 2,7 1,7 1,4 2,4 2,2 1,1 0,1
57,4 72,9 45,5 72,5 102,6 102,7 47,5 46,7 99,4 117,2 31,7 1,1
0,6 0,2 1,2 0,1 0 0 0,5 0,9 0 0 1,6 6,1
5,9 5,9 6 6,5 5,5 5,8 4,7 4,6 5,2 5,1 4,3 6,2
5,3 5,7 4,7 6,4 5,5 5,8 4,2 3,7 5,1 5 2,7 0,1
1,3 1,7 1,7 1,5 1,4 1,6 1,8 1,5 1,5 1,3 2,3 3,1
1,7 1,7 1,9 2 1,8 2,1 1,8 1,8 2 1,8 1,9 0
126,8 95,7 113,5 132,3 127,4 132 97,8 118 127,9 132,6 81,1 0,4
0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 3,1
5,1 5,6 5,1 5,9 5,3 5,9 6 5,5 5,7 5,4 4,5 3,1
5 5,5 5 5,8 5,3 5,8 5,9 5,4 5,6 5,3 4,5 0
1,5 1,4 1,6 1,4 1,3 1,5 1,9 1,5 1,2 1 2 4,3
2,1 2,1 2,1 2 2 2,1 2,6 2,3 2 1,9 2,4 0
138,6 148 133,4 138,7 152,3 139,6 138,3 155,7 162 179,9 117,3 0,8
0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 4,2
5,1 5,1 4,8 4,6 4,7 5 5,9 5,7 5,5 5,4 5,7 4,3
5 5 4,7 4,5 4,6 4,9 5,8 5,6 5,4 5,3 5,6 0,1
C.V. = coeficiente de variação
máximo
variação
média
desvio
C.V.
mínimo
máximo
variação
Vazante
média
desvio
C.V.
mínimo
máximo
variação
Cheia
média
desvio
C.V.
mínimo
máximo
variação
Enchente
média
desvio
C.V.
mínimo
Seca
Tabela 4: Variação da concentração de oxigênio dissolvido média na bacia do lago Tupé e rio Negro.
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas

28
modo geral, apresentaram as menoresamplitudes para essa variável, e umcomportamento mais homogêneo, fatoconfirmado pelos menores valores dedesvio padrão (Tab. 4). Na enchente, amaior concentração média foi aferidano rio Negro (EN12) com 8,2µS.cm , e amenor no igarapé da Cachoeira (EI4)com 7,1µS.cm . A amplitude devariação nesse período foi de6,1µS.cm a 8,5µS.cm , e a maioramplitude pontual foi determinada naestação EI4 2,3µS.cm . Durante a fasede cheia, a maior condutividade médiafoi determinada, mais uma vez, no rioNegro 10,8µS.cm . Nesse período, acondutividade elétrica variou emmédia de 9,1µS.cm (EI4) à 11,4µS.cmnas estações EI8 e ET10. A maiorvariação pontual ficou por conta daestação EI8 com 1,7µS.cm . Na fase devazante, o menor valor médio foidetectado na estação ET1 com7,9µS.cm e o maior valor na estaçãoEI8 com 10,3µS.cm . A amplitude devariação durante a vazante ficou entre6,8µS.cm e 12,6µS.cm . A maiorvariação pontual ocorreu no igarapéTerra Preta (EI8) com 5,3µS.cm .Durante todo o ciclo hidrológico, acondutividade apresentou umavariação pouco significativa, comdesvios-padrão sempre inferiores a 2,5(Tab.5).
Os valores de pH medidos durante ociclo hidrológico estão apresentadosna Figura 6 e Tabela 6. Durante a fasede seca o pH apresentou umaamplitude que variou de 3,89 a 5,95.Nesse período, o maior valor médio foimedido no rio Negro (EN12) 5,23, e omenor valor médio na estação próxima
-1
-1
-1 -1
-1
-1
-1 -1
-1
-1
-1
-1 -1
-1
Figura 5: Variação espacial da condutividade média nacoluna de água de estações do lago Tupé, igarapés e rioNegro durante ciclo hidrológico de 2002/2003.
no canal de conexão entre o rio e o lago, estação EC11,com 3,95. Os valores não apresentaram grandesvariações espaço-temporais, sendo que a maior variaçãopontual foi encontrada na estação EN12 com 0,99. Nafase de enchente a variação espacial ficou entre 3,05,aferido na ET10, e 4,67 no igarapé da Cachoeira (EI4). Amaior variação pontual foi encontrada na ET10 com 1,36.
O lago Tupé apresenta características típicas de águas
Darwich et al.
Seca
Estações de coleta
15
12
9
6
3
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Enchente
Estações de coleta
15
12
9
6
3
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Cheia
Estações de coleta
15
12
9
6
3
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Vazante
Estações de coleta
15
12
9
6
3
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12

29
ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
8,6 6,5 8,45 5,45 5,7 4,25 3,7 4,7 5,49 6,02 4,85 7,42
0 0,7 0,1 0,1 1,3 0,1 0 0,1 0,9 1,8 0,1 0,8
0 10,9 0,8 1,3 23,4 1,7 0 3 16,5 30 1,5 10,8
8,6 6 8,4 5,4 4,4 4,2 3,7 4,6 4,6 3,9 4,8 6,4
8,6 7 8,5 5,5 8 4,3 3,7 4,8 6,9 8,5 4,9 8,1
0 1 0,1 0,1 3,6 0,1 0 0,2 2,3 4,6 0,1 1,7
7,24 7,31 7,13 7,13 7,44 7,38 7,23 7,5 7,63 7,78 7,8 8,15
0,8 1 0,8 1 0,5 0,7 0,5 0,4 0,4 0,3 0,1 0,1
10,6 13,5 11,1 14,1 6,7 10,1 7,2 5,1 4,9 4,4 1,2 0,7
6,3 6,3 6,3 6,1 6,7 6,5 6,9 7,1 7,2 7,3 7,7 8,1
8,4 8,5 8,3 8,4 8 8,4 8,2 7,9 8,3 8,2 7,9 8,2
2,1 2,2 2 2,3 1,3 1,9 1,3 0,8 1,1 0,9 0,2 0,1
10,35 10,29 10,17 10,08 10,34 10,27 10,7 10,61 10,4 10,62 10,27 10,84
0,4 0,3 0,4 0,4 0,5 0,5 0,4 0,4 0,4 0,4 0,5 0,1
4,1 3,3 4,4 3,9 4,8 4,5 3,8 3,4 4,2 3,5 5,1 1,2
9,5 9,4 9,4 9,1 9,8 9,4 9,6 9,7 9,7 10,1 9,7 10,6
11,1 10,8 10,6 10,5 11,2 10,9 11,1 11,4 11,2 11,4 11,1 11
1,6 1,4 1,2 1,4 1,4 1,5 1,5 1,7 1,5 1,3 1,4 0,4
7,9 8,2 8,3 8,57 8,94 8,97 9,43 10,34 9,61 9,31 8,84 8,5
0,7 0,9 0,9 1,1 1,1 1,3 2,1 2,2 1,4 1,3 1,4 0,1
8,5 10,6 11 12,7 12,1 14,4 22,8 21 14,3 13,5 15,3 0,7
6,8 6,8 7 7,1 7,2 7 6,8 7,3 7,2 7 7,1 8,4
8,6 9 9,3 10,1 10 10,2 12 12,6 11,2 10,9 10,5 8,6
1,8 2,2 2,3 3 2,8 3,2 5,2 5,3 4 3,9 3,4 0,2
C.V. = coeficiente de variação
máximo
variação
média
desvio
C.V.
mínimo
máximo
variação
Vazante
média
desvio
C.V.
mínimo
máximo
variação
Cheia
média
desvio
C.V.
mínimo
máximo
variação
Enchente
média
desvio
C.V.
mínimo
Seca
Tabela 5: Variação da condutividade elétrica média na bacia do lago Tupé e rio Negro.
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas

Figura 6: Variação espacial do pH médio na coluna de águade estações do lago Tupé, igarapés e rio Negro duranteciclo hidrológico de 2002/03.
pretas com pH ácido (Rai Hill, 1981).Na cheia o maior pH médio foi aferidona estação ET7 com 4,31, e o menorvalor na estação ET10 com 3,93. Amaior amplitude encontrada foi de3,42 para 4,72, sendo que a maiorvariação pontual foi de 1,20 na estaçãoET1. O período de vazante apresentouvalores de pH que variaram, em média,de 4,15 a 5,02. O maior valor médio depH foi determinado na estação doigarapé da Helena (EI2) com 4,71. Já amaior variação pontual ocorreu naestação a montante da central naestação ET5 com 0,76. As maioresvariações do pH ocorreram entre osperíodos de enchente e cheia,enquanto que durante a vazante o pHse apresentou dentro da faixa do pH4,0 em todas as estações. Os desvios-padrão calculados para o ciclo ficaramsempre abaixo de 0,4 (Tab. 6),indicando uma boa representatividadedos valores médios para cada estação.
Os resultados da Análise deAgrupamento para as variáveis limno-lógicas estão apresentados na Figura 7.A classificação hierárquica evidencioudois grandes grupos de pontos deamostragem. Destacaram-se com fortesimilaridade um primeiro grupo, quereúne os pontos dos igarapés defloresta (igarapé da Helena, dasPedras, da Cachoeira, do Monteiro eTerra Preta) associados ao ponto ET1, eaos pontos ET7 e EC11, que apesar demostrarem-se mais isolados, foramagregados a esse grupo a uma distânciamatricial inferior a 2,9 unidades. Oagrupamento das estações localizadasnos igarapés revela uma homogeneida-de das condições limnológicas para
&
esse tipo de sistema hídrico. No entanto, chama-se aatenção para a forte similaridade espacial da estaçãoET1, localizada no extremo superior do braço maior dolago, com os pontos dos igarapés. Provavelmente, pelasua distância das demais estações do lago, a estação ET1apresenta características químicas e físico-químicasmais próximas de águas de igarapés de floresta. Em uma
30
Darwich et al.
Seca
Estações de coleta
6
4
2
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Enchente
Estações de coleta
6
4
2
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Cheia
Estações de coleta
6
4
2
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
Vazante
Estações de coleta
6
4
2
0ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12

ET1 EI2 EI3 EI4 ET5 EI6 ET7 EI8 ET9 ET10 EC11 EN12
4,01 4,22 4,2 4,57 4,37 4,83 5,1 4,58 4,49 4,54 3,95 5,23
0 0,2 0,1 0 0,3 0 0 0,1 0,2 0,2 0 0,4
0,2 4,9 1,3 0,3 7,3 0,1 0,1 1,7 4,3 5,3 1,1 7,8
4 4,07 4,16 4,56 3,89 4,82 5,09 4,52 4,19 4,26 3,92 4,96
4,01 4,36 4,24 4,58 4,8 4,83 5,1 4,63 4,78 4,98 3,98 5,95
0,01 0,29 0,08 0,02 0,91 0,01 0,01 0,11 0,59 0,72 0,06 0,99
4,06 4,14 4,12 4,29 4,14 4,24 4,25 4,25 4,07 3,9 4 3,78
0,3 0,3 0,3 0,3 0,3 0,2 0,1 0,1 0,3 0,4 0,2 0,2
8 7,2 7,5 6,7 6,2 5,5 3,5 3,5 7,3 11,4 5,7 5,6
3,67 3,65 3,59 3,88 3,51 3,89 4,01 4,02 3,28 3,05 3,74 3,43
4,63 4,47 4,57 4,67 4,48 4,61 4,48 4,47 4,46 4,41 4,27 3,96
0,96 0,82 0,98 0,79 0,97 0,72 0,47 0,45 1,18 1,36 0,53 0,53
4,16 4,21 4,26 4,24 4,08 4,25 4,31 4,23 4,18 3,93 4,15 4,16
0,4 0,3 0,3 0,3 0,3 0,3 0,2 0,3 0,3 0,4 0,3 0,3
8,7 8,1 7,8 6,6 8,6 6,6 5,6 6,4 8 9 7,8 8,4
3,5 3,64 3,66 3,72 3,48 3,73 3,81 3,73 3,6 3,42 3,69 3,43
4,7 4,66 4,72 4,58 4,51 4,63 4,62 4,59 4,64 4,45 4,59 4,44
1,2 1,02 1,06 0,86 1,03 0,9 0,81 0,86 1,04 1,03 0,9 1,01
4,61 4,71 4,67 4,64 4,58 4,65 4,54 4,69 4,7 4,67 4,68 4,38
0,2 0,2 0,1 0,2 0,3 0,1 0,2 0,2 0,2 0,2 0,2 0,2
4,1 3,7 2,9 3,8 5,5 2,4 4,4 3,2 3,7 3,6 3,4 3,5
4,31 4,5 4,53 4,37 4,15 4,46 4,22 4,49 4,37 4,34 4,41 4,16
4,93 5,02 4,91 4,91 4,91 4,79 4,72 4,93 4,94 4,94 4,84 4,57
0,62 0,52 0,38 0,54 0,76 0,33 0,5 0,44 0,57 0,6 0,43 0,41
C.V. = coeficiente de variação
máximo
variação
média
desvio
C.V.
mínimo
máximo
variação
Vazante
média
desvio
C.V.
mínimo
máximo
variação
Cheia
média
desvio
C.V.
mínimo
máximo
variação
Enchente
média
desvio
C.V.
mínimo
Seca
Tabela 6: Variação do pH médio na bacia do lago Tupé e rio Negro.
31
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas

primeira avaliação dos resultados,pode se entender que as estaçõesamostrais localizadas no canal deconexão (EC11) e no extremo superiordo braço menor do lago (ET7), foraminclusas neste grupo pela fortesimilaridade entre si ou com os demaiselementos, más o conhecimento dascaracterísticas geomorfológicas daregião, descritas em Aprile .(2005, neste volume), mostraexatamente o contrário, que taispontos são bastante distintos em suascondições ambientais. Dessa forma,conclui-se que a agregação dessespontos baseia-se não em suasimilaridade, mas no fato destes sediferenciarem em termos limnológicosdas demais estações do lago. Com umadistância matricial inferior a 6,9, osegundo grupo destaca-se pelaagregação dos pontos ET5, ET9 e ET10.Trata-se de um grupo formado pelasestações localizadas na área maisaberta do lago, com maior circulaçãodas águas, e maiores profundidadesmédias. Nesse grupo está incluído aestação Et10, denominada de estaçãocentral pela sua eqüidistância dosextremos do lago. A estação EN12isolou-se das demais estações, essefenômeno é explicado pelo fato de queessa estação é a única com caracterís-ticas lóticas, que a diferenciam dasdemais estações monitoradas.
et al
Figura 7: Dendograma de Similaridade Espacial dasestações de amostragem do lago Tupé para o ciclohidrológico de 2002/03.
32
Darwich et al.
Conclusões
De modo geral, ao longo do ciclo hidrológico asmaiores amplitudes de variação dos parâmetroslimnológicos ocorreram durante o período de seca. Operíodo de seca destacou-se por apresentar as maioresconcentrações de oxigênio dissolvido, em oposição aosperíodos de cheia e vazante. As estações localizadas nosigarapés de floresta apresentaram um comportamentobastante homogêneo, principalmente com relação asvariáveis temperatura e condutividade elétrica, fatoevidenciado na análise de agrupamento. O sistemalêntico apresenta características típicas de águas pretascom pH ácido e baixa condutividade. Os resultados daanálise de agrupamento mostraram que a estaçãolocalizada no rio Negro (EC12) isolou-se das demaisestações, revelando um comportamento hidroquímicodiferenciado das estações do lago Tupé.
Similaridade Espacial
EN12 ET10 ET5 ET7 EC11 EI6 EI8 EI2 EI3 ET1
Dis
tânci
aE
ucl
idia
na
d=25,830
25
20
15
10
5
0
d=14,7
d=6,2
d=2,9

33
Bibliografica Citada
Aprile, F. M., Darwich, A. J. 2005. Modelosgeomorfológicos para o lago Tupé In:Santos-Silva, E. N.; Aprile, F. M.;Scudeller, V. V.; Melo, S. (Orgs.),
. Editora INPA,Manaus,AM.
Darwich, A. J. 1995.
. Tese deDoutorado. Instituto Nacional dePesquisas da Amazônia/ Universidadedo Amazonas (INPA/UA), Manaus (AM).327pp.
Darwich, A. J.; Aprile, F. M.; Robertson, B.A. 2003. Estratificação térmica em umlago de água preta amazônico: umaquestão equatorial.
- CBL 2003.Universidade Federal de Juiz de ForaUFJF. Juiz de Fora (MG), 20-25.julho.2003. CD-ROM, N.39.
Furch, K. 1984a. Seasonal variation of themajor cation content of the várzea-lake Lago Camaleão, middle Amazon,Brazil, in 1981 and 1982.
:1288-1293.
Furch, K. 1984b. Water chemistry of theAmazon basin. The distribution ofchemical elements among freshwaters,In: H. Sioli (ed.).
. Dr.W. Junk Publ. Dordrecht, Boston,Lancaster. P.167-199.
Furch, K.; Junk, W. J.; Dieterich, J.;Kochert, N. 1983. Seasonal variation in
BioTupé: meio físico, diversidade
biológica e sociocultural do baixo rio
Negro Amazônia central
Processos de
decompos ição de Echinochloa
polystachya (H. B. K.) Hitchcock
(Gramineae = Poaceae), capim semi-
aquático da várzea amazônica
IX Congresso
Brasileiro de Limnologia
Verh.
Internat. Verein. Limnol., 22
The Amazon:
Limnology and landscape ecology of a
mighty tropical river and its basin
the major cation ( Na, K, Mg and Ca) content of the Water ofLago Camaleão, in Amazonian floodplain-Lake nearManaus, Brazil. , 8(1):75-89.
Golterman, H. L.; Clymo, R. S.; Ohnstad, M. A. 1978.. 2nd ed.
IBP Handbook, nº 8. Blackwell Scientific Publications,Oxford. Edinburgh London Melbourne. 214pp.
Junk, W. J. 1973. Investigations on the ecology and production-biology of the "floating meadows" (
) on the Middle Amazon. II. The aquaticfauna in the root zone of floating vegetation. ,4(1):9-102.
Junk, W. J.; Bayley, P. B.; Sparks, R. E. 1989., In: D. P. Dodge (ed.).
Proceedings of the International Large River Symposium.Can. Spec. Publ. Fish.Aquat. Sci. 106.p. 110-127.
Kern, J.; Darwich, A. J. 1997. Nitrogen turnover in the várzea.In: Junk, W. J. (ed).
. Springer-Verlag, BerlinHeidelberg. Ecological Studies, Vol. 126. Cap. 6; p.119-135.
Melack, J. M.; Fisher, T. R. 1983.. Arch. Hydrobiol.,
98(4): 422-442.Rai, H.; Hill, G. 1981. Physical and chemical studies of lago
Tupé; a Central Amazonian Black Water,, Ria Lake.., 66(1):37-82.
Reiss, F. 1977. Qualitative and quantitative investigations onthe macrobenthic fauna of Central Amazon Lakes. I. LagoTupé, a black water lake on the lower Rio Negro.
, 2:203-235.Schmidt, G. W. 1972a. Seasonal changes in water chemistry of a
tropical lake (Lago do Castanho,Amazonia, SouthAmerica)., 18:613-621.
Schmidt, G. W. 1972b. Chemical properties of some waters inthe tropical rain-forest region of Central Amazonia alongthe new road Manaus-Caracaraí. , 3(2):199-207.
Statsoft, Inc. 1997. Computer programmanual. Tulsa, OK, EUA.
Wetzel, R. G.; Likens, G. E. 2000. . W. B.Saunders Co. Philadelphia. 357pp.
AmazonianaMethods
for physical and chemical analysis of fresh waters
Paspalo-
Echinochloetum
Amazoniana
The flood pulse
concept in river-foodplain systems
The Central Amazon Floodplain:
Ecology of a Pulsing System
Diel variations and ecological
implications in Amazon foodplain lakes
Int.
Revue ges. Hydrobiol
Amazoniana
Verh. Internat. Verein. Limnol.
Amazoniana
Statistica for Windows
Limnolgical Analysis
Variáveis limnológicas: contribuição ao estudo espaço-temporal de águas pretas amazônicas


RESUMO - Neste capítulo são comentadas características físicas, químicas e físico-químicasda substância água, sua importância ecológica, assim como, das substâncias dissolvidas nomeio hídrico, particularmente o oxigênio dissolvido. Nesse contexto, são discutidos osteores de O dissolvido na coluna de água do lago Tupé, variações espaciais, nictemerais,sazonais, estratificações, fluxos laminares, supersaturação e déficit de O durante o ciclohidrológico 2001/2002. São feitas comparações com o O que aflui do rio Negro e dosprincipais igarapés de floresta à bacia lacustre. As medidas de oxigênio dissolvido nos
2
2
2
Meio FísicoCapítulo 3
Assad José DARWICH
Luiz Fernando ALVES
Fábio Marques APRILE
Barbara Ann ROBERTSON
Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Biologia AquáticaAv. André Araújo 2936, Caixa Postal 470 Manaus, AM, 69060-001
Limnologia do Lago Tupé: dinâmicaespaço-temporal do oxigênio dissolvido
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

36
diferentes ambientes evidenciaram, em todas a fases do ciclo hidrológico, elevadasconcentrações de O nas camadas mais superficiais do lago e um déficit considerável deoxigênio no hipolímnio. Na região central do lago, a área mais profunda da bacia lacustre,ficou caracterizada a existência de oxiclina com perfil clinogrado clássico. A análisenictemeral do oxigênio na região central mostrou a existência de uma acentuadaestratificação no teor de saturação desse gás durante o período pesquisado. O pulso deinundação apresentou participação decisiva sobre a dinâmica do oxigênio nos ambienteslacustre e fluvial
PALAVRAS-CHAVE: água, substâncias dissolvidas, oxigênio, estratificação, fluxo laminar,igarapé, lago,Amazônia.
2
Introdução
ASubstância Água
A abundância de água é a característica fundamentaldo planeta Terra e, a água doce é o tipo mais diretamenteenvolvido com as atividades e necessidades básicas doHomem. Em qualquer análise sobre os recursoscontinentais dessa substância deve-se considerar que aágua potável é um recurso finito, utilizado pelahumanidade que cresce exponencialmente.
Para o entendimento da estrutura e função das águasdoces é necessário o conhecimento de característicasfundamentais da substância água. Em particular, aspropriedades de expansão térmica e densidade comoconseqüência do arranjo atômico dos átomos dehidrogênio e oxigênio que formam a molécula de água,bem como das ligações entre moléculas, o que conduz aestados tanto ionizados quanto covalentes mantendo-sea integridade da água como substância.
A água dissocia-se em ligações heteropolares, o que afaz um excelente solvente, no entanto, longe se ser osolvente universal. A maior parte das propriedades daágua está em função da temperatura, pressão esubstâncias dissolvidas. Algumas são de interesseecológico marcante. Sua elevada(calor específico a 1 atm a 15°C é 4,1855 J.g = 1 cal.g , émenor apenas que o do Li, H e NH líquido) e o alto
(79kcal.L ) e (537kcal.L a100°C) têm efeito termostático e regulador (Margalef,
capacidade calorífica
calor
de fusão de evaporação
-1 -1
-1 -13
1986). Estas propriedades proporcio-nam um ambiente hídrico muito maisestável que o ambiente terrestre. Issoocorre porque são necessáriasquantidades relativamente elevadasde energia, em forma de calor, pararomper as ligações de hidrogênio naágua em sua forma líquida. Por isso, asflutuações de temperatura na água sedão de forma gradual e os extremosestacionais e diurnos são pequenos emcomparação com os habitats aéreos eterrestres (Margalef, 1983; Wetzel,1993). O calor específico do gelo é ametade (0,5cal.g ) do da água. Comoresultado disso, o gelo se forma demaneira relativamente rápida aoesfriar-se a água a 0°C (Hutchinson,1967, 1975a,b; Wetzel, 1993). Dessaforma, a água necessita perderpequenas quantidades de calor(80cal.g ), para que suas moléculas seunam formando gelo, em comparaçãocom a quantidade de energianecessária (540cal.g ) para romper asligações de hidrogênio durante aevaporação. Ao contrário, o gelo fundecom relativa rapidez e requer menorquantidade de energia para voltar aoestado líquido, que a necessária para
-1
-1
-1
Darwich et al.

aumentar a tempera-tura da água(Wetzel, 1993).
A elevada(em unidades c.g.s. é 88,0 a 0°C e80,36 a 20°C) da água deve-se àestrutura assimétrica de sua moléculae explica a facilidade com que seionizam os sais dissolvidos na água,pois a atração entre os íons éinversamente proporcional à constante dielétrica. A , relativamente elevada (1x10 Pas à 20°C),diminui conforme aumenta atemperatura (a 0°C é 1,8x10 Pas). De0°C a 25°C, a viscosidade cai àmetade. Esta propriedade temimportância na locomoção dosorganismos, fazendo com que aresistência por fricção ao deslocamen-to de um organismo seja 100 vezesmaior na água do que no ar,dependendo da área da superfícieexposta, da velocidade e temperatura,e da quantidade de sais dissolvidos.Assim como a viscosidade, a
, também diminui conforme aumenta a temperatura,(72,76dina.cm a 20°C, a maior detodos os líquidos à temperaturaambiente, com exceção do Hg), e,juntas, influenciam a locomoção de
As singularidades na relação entre o(do vapor é
1729,6cm .g a 100°C e 1atm) e atemperatura regulam a circulaçãovertical e a estabilidade nos lagos emares. A forte absorção de radiação degrandes comprimentos de onda, quecontrasta com a maior transparência àluz, influi sobre a estratificaçãotérmica e sobre a distribuição dasplantas (Margalef, 1986).
volume específico3 -1
constante dielétrica
-
viscosidade -
tensão
superficial -
-3
-3
-1
organismos aquáticos. A tensão superficial da águadiminui com a presença de compostos orgânicos, sejameles de origem alóctone (e.g., agrotóxicos) ouproduzidos durante a fotossíntese por algas eangiospermas submersas ou por morte e lise de células. Atensão superficial da água pode sustentar organismosgrandes como girinos e coleópteros, porém podeprejudicar outros como cladóceras que uma vez sobre apelícula superficial da água não podem, por si só,submergir ao seu habitat (Wetzel, 1993). Algunsorganismos estão adaptados a viver nessa película, umhabitat especial na interface ar-água. O conjunto dessacomunidade chama-se nêuston.
Uma das características de particular importânciapara a manutenção da vida no meio hídrico é a
da água que é 775 vezes maior que adensidade do ar à pressão e temperatura padrões (0°C e760mmHg). Isto proporciona uma maior flutuabilidadeaos organismos e um menor gasto de energia paramanterem sua posição no meio hídrico. Conseqüênciasimportantes na estrutura interna de organismos quevivem no meio aquático são as modificações (e.g.,redução) nos tecidos de sustentação de animais de águadoce, especialmente de invertebrados e em plantasvasculares.
densidade
A densidade da água sofre variação singular emfunção da temperatura e este fato tem importânciaecológica considerável sobre a circulação, a estabilidadee o congelamento da água dos lagos, rios, mares etc. Aágua atinge sua densidade máxima à 3,94°C (igual a1,000000g.cm a 1atm). Acima e abaixo dessatemperatura (até 0°C), há expansão molecular dasubstância água com conseqüente diminuição dadensidade a uma taxa progressivamente crescente devalor (Fig. 1). A mudança de densidade ocorre porque asvariações de temperatura modificam as distânciasintermoleculares e os movimentos de reorientação etranslação das moléculas de água. Por isso, a 0°C e a1atm, a densidade da água no estado líquido é0,9998679g.cm e a do gelo é igual a 0,9168g.cm emconseqüência do aumento do espaço entre as moléculasda substância água ao congelar-se. Essa diferença faz
-3
-3 -3
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
37

com que o gelo seja cerca de 9% mais leve do que a água azero grau centígrado. Por isso, o gelo flutua. Assim, noinverno rigoroso de regiões temperadas, o congelamentoda água nos lagos, rios etc. ocorre primeiramente nasuperfície e o estrato congelado tende a se espessar àmedida que diminuem as temperaturas. Dessa forma, ogelo flutua sobre a água mais “quente” e mais densa.
A diminuição da densidade à medida que atemperatura se distancia de 3,94°C, como foi dito,provoca diferenças de densidades que são pequenasentre um grau e outro, mas altamente significativas dospontos de vista físico-químico, químico, biológico e eco-fisilológico. Verifica-se, entretanto, que essasdiferenças de densidade da água por grau são cada vezmaiores à medida que as temperaturas se distanciam de3,94°C. Assim, a variação da densidade com atemperatura não é linear, visto que na faixa detemperaturas altas, uma pequena diferença detemperatura já provoca uma grande diferença dedensidade. Dessa forma, a análise da “diferença dedensidade por grau descendente” (DDD) (Fig. 1) éaltamente importante para o entendimento deestratificações e misturas da coluna de água de lagos.Sabe-se que para misturar líquidos de densidadesdiferentes necessita-se de trabalho (energia) que éproporcional à diferença de densidade entre eles. Damesma forma, para se misturar massas de água comdensidades diferentes necessita-se de forças quedeverão ser tanto maiores quanto maiores forem asdiferenças de densidades entre as massas.Apartir dessesdados verifica-se que a quantidade de trabalhonecessária para misturar massas “estratificadas” deágua, p. ex., a temperaturas de 32 e 33°C é 40 vezesmaior que a necessária para misturar as mesmas massasentre 4 e 5°C. Entre 29 e 30°C o trabalho necessário é 37vezes maior.
O ponto fundamental dessa variação de densidadecom a temperatura é verificar que, na regiãoAmazônica,pequenas diferenças de temperatura podem estabelecerestratificações químicas (e.g., oxiclinas) muito estáveis.Na estação central do lago Tupé a coluna de águaapresentou amplitude térmica de cerca de 6 graus
centígrados no período de águasbaixas, a seca (de 27 a 33°C), e quasetrês no período de cheia (de 27 a30°C). Nessa faixa de temperatura acoluna de água na seca é cerca de 2,1vezes mais estável do que na cheia. Seesses dados forem comparados comvariações de temperaturas de regiãotemperada, verifica-se que a coluna deágua do Tupé estaria quase 7 (6,7)vezes mais estável na seca do que amesma coluna de água entre 4 e 10°C(mesma amplitude térmica). Na cheia,a coluna de água do Tupé estaria maisestável ainda (cerca de 12,3 vezes) doque em uma faixa equivalente detemperatura na região temperada(entre 4 e 7°C).
Quanto à distribuição de oxigêniodissolvido em um lago de bacia isoladadessa região equatorial, a estabilidadeverificada na coluna de água do Tupépode conduzir à conseqüênciasnegativas, como anoxia permanente.Sob esta condição o O produzido nazona eufótica torna-se impedido deatingir as camadas mais profundas dolago porque a difusão é lenta e oconsumo é grande, tornando o seuhipolímnio pouco propício aosorganismos aeróbicos. Como adensidade também aumenta de formaaproximadamente linear com oaumento das concentrações de saisdissolvidos, a permanência dessacondição em uma região rica em saisdissolvidos, poderia conduzir o lago àuma meromixia permanente. Aavaliação estática das curvas deoxigênio no Tupé sugere a condição demeromixia, no entanto, a contribuiçãopermanente de pequenos afluentes
2
38
Darwich et al.

Figura 1. Influência da temperatura sobre a densidade da água: importância na circulação eestratificação térmica de lagos.
(igarapés) promove um fluxo cons-tante nas camadas mais profundas demaior densidade. Além disso, areduzida quantidade de sais dissolvi-dos (expressa pelos reduzidos valoresda condutividade elétrica) nas águasdesses afluentes, são fatores antagôni-cos a este tipo de estratifica-ção,ainda que as curvas de O sejamsempre do tipo clinogrado. Isto podeser explicado pelo maior consumo deoxigênio nas camadas mais profundas,em conseqüência de processos de
2
decomposição do material orgânico oriundo,principalmente, do escoa-mento superficial da floresta.
Wetzel (1993), comenta que de forma específica, ascaracterísticas da água regulam eficazmente ometabolismo dos lagos e que as modificações do meioaquático se produzem como resposta às variaçõesclimáticas e geográficas. Salienta o autor, que aspeculiares propriedades de expansão térmica, o altocalor específico e as características líquido-sólidas daágua formam um ambiente estratificado que controlaeficientemente a dinâmica química e biótica dos lagos.
A água, enfim, proporciona um meio temperado noqual as flutuações extremas de suas características
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
39
Densidade versus temperatura
0 10 20 30 40
0
1
2
3
4
5
10
15
20
25
30
34
Te
mp
era
tura
(°C
)
A . Diferença de densidade (g/cm)3
vezes 105 /°C abaixando (DDDx10 ).5
DDDx100000
Densidade versus temperatura
0,99310
0,99410
0,99510
0,99610
0,99710
0,99810
0,99910
1,00010
0 10 20 30 40
B. Variação da densidade (g/cm3) da
água com a temperatura (°C)
De
nsi
da
de
(g/c
m3 )
dens

químicas, físicas e físico-químicas, em especial atemperatura, são menos drásticas do que no meio aéreo.Algumas propriedades da água (e.g., variação detemperatura e de densidade com a temperatura), unidasa um grau de viscosidade relativamente elevado, têmpossibilitado o desenvolvimento de um grande númerode adaptações bióticas, que têm servido para sustentar aprodutividade aquática.
Todas as particularidades da água como substânciaestão relacionadas às suas características físicas,químicas e físico-químicas peculiares e intrínsecas dessasubstância como conseqüência de sua estrutura atômicae ligações, bem como pela associação de suas moléculasnas fases sólida, líquida e gasosa. Certamente por isso, aágua tornou-se a essência da vida como a conhecemos naTerra e domina por completo a composição química detodos os organismos.
Amatéria viva é fortemente hidratada e grande partedas reações bioquímicas características da vida serealiza entre as substâncias dissolvidas no meio aquoso.Estas substâncias foram e continuam sendo adicionadasao meio aquático, basicamente por meio da circulaçãoglobal da substância água, tornando as águas dedeterminadas regiões caracterizadas de acordo com asespécies químicas nelas contidas.
A água de todos os compartimentos no planeta Terracircula e se renova em tempos estimados para cadacompartimento. Wetzel (1993), apresenta dados sobreos tempos de renovação das águas. O tempo para osoceanos, calculado com base na evaporação líquida, foiestimado em 37 mil anos. No outro extremo, o tempo derenovação da água dos rios está estimado em cerca de 20dias para os grandes rios que deságuam nos oceanos. Riosmenores apresentam tempos mais reduzidos. A águasempre volta aos compartimentos por precipitaçãopluviométrica completando o ciclo nos mares. Ao voltar,a água absorve parte da terra. Por dissolução, erosão eescoamento adquirem os seus componentes dissolvidos.Estes são essencialmente os mesmos em todos oscompartimentos (oceanos, gelo polar, água subterrânea,
Espécies Químicas Dissolvidas na Água
umidade do solo e subsolo, lagos, rios,vapor atmosférico etc.), contudo, asconcentrações são muito variáveis porrazões diversas, e.g., diferençasgeológicas nas bacias de drenagens esazonalidade.
Como conseqüência da grandevariabilidade química e do teor dosconstituintes adicionados, os com-ponentes das águas naturais podem serdivididos em 5 classes, classificadoscomo 1) íons e compostos inorgânicosdissolvidos, 2) compostos inorgânicosparticulados, 3) compostos orgânicosdissolvidos, 4) materiais orgânicosparticulados e 5) gases dissolvidos(Golterman ., 1978). Os principaisfatores que determinam as quantida-des, tipo e origem de materialdissolvido e em suspensão são: o clima,o relevo, e o tipo de rochas queconstituem as bacias de drenagem.
De particular importância nalimnologia são os íons e compostosinorgânicos dissolvidos (1) e os gases(5). Os primeiros podem serconvenientemente divididos emconstituintes principais, constituintessecundários, elementos-traço.
Os constituintes principais sãoaqueles elementos ou compostos que,freqüentemente ocorrem em maioresquantidades nas águas naturais. Aconcentração dos constituintesnumericamente mais importantes (Na+ K + Ca + Mg + Cl + HCO + SO =)varia de 1,0 a 10meq.L e em conjuntosão os principais responsáveis pelacondutividade da água e pelasalinidade de águas interiores (Stumm
Morgan, 1981). São também citadoscomo os principais cátions e ânions os
et al
+
+ 2+ + - - -
-1
23 4
&
40
Darwich et al.

íons de Fe e Mn , e carbonatos (CO =)em águas com elevados valores de pH(Hutchinson, 1975b; Golterman .,1978). Têm importância na produtividade global dos ecossistemas aquáticos e no metabolismo de certasbactérias, capazes de obter energia apartir de formas reduzidas de algunselementos (Fe e Mn ) (LampertSommer, 1997). Os componentes destegrupo têm origem, principalmente, nointemperismo das rochas e dos solos dabacia de drenagem e de acumulaçãodos corpos de água, além da precipitação pluviométrica e da contribuiçãoantrópica.
Os constituintes secundáriosocorrem em menores quantidades,com concentrações entre menos de1mg/L e poucos mg/L, destacando-secompostos do N e suas frações (NH +;NO -; NO -); do P e suas frações(ortofosfatos P-PO , e fósforodissolvido) e silicatos (SiO , HSiO -, Si-Si(OH) +), os quais podem ocorrer emmaiores quantidades. Em conjuntotêm importância no metabolismo deecossistemas aquáticos, como naprodutividade primária, no armazena-mento de energia e na formação deestruturas de organismos aquáticos.Têm origem principal na chuva e nafixação molecular (N), nas rochas dabacia de drenagem dos corpos de água(P), na decomposição de minerais(silicatos). Alguns compostos do N(NO -) e P (P-PO ) podem ocorrer emelevadas concentrações em lagos devárzea amazônicos enriquecidos pordecomposição de macrófitas aquáticasno período de águas baixas (Darwich
., 2001). Além disso, podem ser
3+ 2+
3+ 2+
-
3-
3-
3
4
3 2
4
2 3
4
3 4
et al
-
-
et
al
&
adicionados ao meio hídrico por atividadesantropogênicas etc., e provocarem problemas comoeutrofização e poluição.
Os elementos-traço ocorrem em quantidades muitopequenas, freqüentemente em partes por bilhão, emtodos os ambientes. São essenciais no metabolismo deorganismos aquáticos tais como na fotossíntese, cadeiarespiratória, fixação de nitrogênio etc. Golterman(1975), inclui entre eles o Al, B, Fe, Co, Cu, Mo, Mn e Zn,além de Ag, Cd, Cr, Hg, Pb, Sn e outros. Têm origemnatural no intemperismo das rochas e erosão dos solos.Podem ser adicionados ao meio aquático por atividadesindustriais, de mineração, na agricultura, domésticaetc., e dessa forma, em quantidades maiores, tornam-setóxicos aos organismos em geral.
Ainda quanto à separação, divisão dos elementos noambiente aquático, há considerações quanto àquantidade dos elementos ou a importância deles nomeio, podendo-se separá-los ou agrupá-los em macro emicronutrientes. Nessa condição, p.ex., tantoconstituintes secundários quantos os elementos-traçopodem aparecer como micronutrientes. Afora essasconsiderações generalizadas quanto aos teores e aqualidade dos constituintes nos compartimentos, é certoque há considerável variação local dentro de um mesmocompartimento, de uma mesma bacia hidrográfica, deum mesmo lago.
Os gases dissolvidos na água podem vir do ar ou deatividades metabólicas de organismos aquáticos. Os maisimportantes são o oxigênio (O ), o nitrogênio (N ) e o gáscarbônico (CO ), todos com origem atmosférica e,respectivamente, na fotossíntese, na atividadebacteriana e na respiração dos organismos. Além desses,são importantes também o gás sulfídrico (H S) e ometano (CH ), ambos com origem na atividadebacteriana. Todos têm solubilidade influenciada pelatemperatura e pela pressão parcial do gás específico. OCO tem solubilidade também relacionada com o sistemabicarbonato e apresenta fontes adicionais importantescomo a (a) oxidação bacteriana de matéria orgânica,tanto na água como nos sedimentos, a (b) atmosfera echuva, a (c) decomposição e, (d) em certas formaçõesrochosas e em águas subterrâneas.
2
2
2
2
2
4
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
41

Para todos os gases dissolvidos há um equilíbrio pordifusão e a quantidade que se dissolve depende tambémda constante de solubilidade do gás específico.Biologicamente, no entanto, o oxigênio é o gás dissolvidomais importante na água.
O oxigênio dissolvido na água tem origem no processode fotossíntese de algas e macrófitas aquáticas e suapresença no meio hídrico depende, como fonte, dessaprodução e da difusão do ar atmosférico. O ar contémcerca de 20,95% de oxigênio e praticamente todo orestante é nitrogênio, além de uma pequena quantidadede outros gases. No entanto, o oxigênio é mais solúvelque o N e sua concentração no meio aquático podeapresentar considerável variação sazonal, espacial ediária, dependendo do ambiente considerado.Obviamente, o oxigênio é essencial para o metabolismode todos os organismos aquáticos aeróbicos. Por isso, adistribuição do O nos lagos é essencial para se entendera distribuição, a abundância, o comportamento, e ocrescimento fisiológico dos organismos aquáticos, alémde influir até sobre a solubilidade de muitos nutrientesinorgânicos (Wetzel, 1993), o que faz desse gásdissolvido o parâmetro de fundamental importância parao meio hídrico.
A presença do O na água depende da suasolubilidade, a qual varia com a temperatura e a pressãoatmosférica. Consequentemente a solubilidade étambém influenciada pela altitude que se encontra oambiente aquático considerado, pelas condiçõesmeteorológicas e, pela pressão hidrostática a umadeterminada profundidade na coluna de água. Visto quea solubilidade aumenta com a profundidade, quantomaior a profundidade, maior é a capacidade deacumulação de gás no ponto considerado até que seforme bolhas que se deslocam à superfície.
A solubilidade do O diminui conforme aumenta atemperatura e a quantidade máxima para a saturação(14,621mg/L) ocorre a zero grau centígrado (Fig. 2-A).Verifica-se que a 40°C a quantidade de O necessária
Oxigênio Dissolvido
2
2
2
2
2
para a saturação é apenas 43,9%daquela a 0°C. Considerando umatemperatura média de 29°C para aságuas amazônicas, observa-se quepara ocorrer saturação a essatemperatura são necessár ios7,691mg/L, ou seja, cerca de 52,6%daquela a 0°C.A importância ecológicadessas observações é que menoresconcentrações de O na coluna de águade lagos amazônicos próximos ao níveldo mar, representam maiores níveis desaturação desse gás no meio hídrico.Como o lago Tupé encontra-sepraticamente ao nível do mar (ca. de20m a.n.m.) e as temperaturas regis-tradas na sua coluna d'água variaramde 27 a 34°C de 2001 a 2002, nota-seque as concentrações para a saturaçãodo O2 naquele ambiente estiveram emtorno da metade do teor para asaturação a 0°C, respectivamente,54,5% e 48,3% (Fig. 2B). Por isso,pequenas concentrações de Orepresentam elevadas percentagensde saturação nessa região. Dessaforma, como será discutido nestecapítulo, foram registradas até10,81mg O .L a uma temperatura de32,9°C nas camadas superficiais. Estaconcentração corresponde a 148,9% desaturação, o que significa supersatura-ção. Nas camadas mais profundas,abaixo da zona eufótica, asconcentrações de O foram sempremuito baixas e as temperaturas defundo mantiveram-se sempre em tornode 27°C durante todo o ciclohidrológico. Nota-se que a essatemperatura, 1,0mgO /L eqüivale acerca de 13% de saturação e que, doponto de vista funcional, pode
2
2
2
2
2
-1
42
Darwich et al.

representar razoável quantidade deoxigênio para a comunidade aquática.
Nos rios, abaixo de grandescachoeiras o teor de oxigênio na águapode estar aumentado em conseqüência da agitação mecânica, ou comomenciona Likens Ragotzkie (1965,1966), bolhas de ar podem sertransportadas para maiores profundidades, onde, devido à maior pressãohidrostática, a solubilidade do O
-
-
&
2
aumenta. Dados recentes (Darwich ., Nãopublicado) sobre o curso superior do rio Madeiraevidenciaram que a concentração do oxigênio dissolvidona coluna de água está aumentada em cerca de 20% atémesmo cerca de 10 quilômetros à jusante das principaiscachoeiras. Considerando que nesse trecho o rioapresenta grande velocidade de corrente e visibilidadepraticamente nula em conseqüência da grandequantidade de material em suspensão (transparência aodisco de Secchi de 10cm), este oxigênio é certamente
et al
Figura 2. Variação da solubilidade do oxigênio com a temperatura na água pura a 760 mmHg, de 0°C a40°C (lado A) e nas temperaturas registradas no lago Tupé a mesma pressão. De novembro de 2001 asetembro de 2002 (de 27°C a 34°C), caixa pontilhada (lado B em detalhe).
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
43
Saturação do oxigênio versus
temperatura
5
6,5
8
9,5
11
12,5
14
15,5
0 15 30 45
°C
O2
(mg
/L)
A
Saturação do oxigênio: temperatura no
lago Tupé
6,8
7
7,2
7,4
7,6
7,8
8
8,2
26 27 28 29 30 31 32 33 34 35 36
°C
O2
(mg
/L)
B

proveniente da atmosfera em sua quase totalidade e estásendo adicionado ao rio por agitação mecânica nascachoeiras.
Têm-se determinado o metabolismo de certascomunidades de rios, lagos etc., através de medidas devariações diárias do teor de oxigênio na água. Rios nãopoluídos normalmente estão saturados ou ligeiramentesupersaturados de oxigênio e podem apresentar grandesvariações sazonais (Golterman, 1975). No entanto, naAmazônia, grandes rios como o Negro, Solimões e oAmazonas, em excelente estado de conservação, podemapresentar baixas concentrações de oxigênio dissolvidono período de águas altas (cheia). Nesse período Darwich(1995), mediu no rio Solimões/ Amazonas concentraçõescom cer-ca de 1,3mg.L , em torno de 15% de saturaçãona coluna de água. Os teores mais elevados registradosvariaram de 77 a 95% de saturação, no período de seca,com média anual pouco abaixo de 60%. O teor deoxigênio nos grandes rios amazônicos parece serconsideravelmente estável em ciclos diários,independente de sua quantidade na coluna de água comoregistrou Darwich (1995) para o rio Amazonas. Os valoresvariaram com a fase do ciclo hidrológico. Por outro lado,em ambientes lênticos amazônicos a variação éfreqüentemente grande e estreitamente relacionadacom a área do lago, com a fase do ciclo hidrológico, coma hora do dia e com a profundidade da coluna de água,como registrado no lago Tupé. Nesses ambientes,certamente, dois fatores antagônicos importan-tesatuam na disponibilidade do O presente: de um lado adifusão atmosférica e a produtividade primária e, deoutro, a oxidação direta do material orgânico que entrano ambiente, a atividade bacteriana e a respiração dacomunidade aquática.
Os lagos de várzea amazônicos têm poucaprofundidade (cerca de 6-8m no nível médio das águas),quase sempre variando com a flutuação anual do nível daágua dos grandes rios. Através da sua conexão com o rio(paraná, canal de ligação), enchem durante o período desubida das águas desses rios e secam à medida que essaságuas baixam de nível. Independente da sua superfície,alguns secam parcialmente, enquanto outros ficam
-1
2
completamente secos, no nível maisbaixo das águas dos rios que osabastecem. De acordo com Junk (1980)e Furch . (1983), dentro de umaclassificação limnológica esses lagospodem ser colocados entre rios(sistemas abertos) e lagos (sistemasfechados). Como resultado dessamudança periódica, a química da águado lago de várzea é principalmenteinfluenciada pelo suprimento, tam-bém periódico, de substâncias solúveise de matéria inorgânica particulada aqual forma os depósitos de sedimentosaluviais e com isso o sedimento dopróprio lago (Furch ., 1983). Comoparte desse conjunto encontram-se oslagos de várzea em ilhas de várzea,provavel-mente muito mais dinâmicosem suas modificações do que osdemais, visto que, no pico das cheiaspodem adquirir características fluviaispelo transbor-damento dos diqueslaterais. Além disso, estão sujeitos àmodificações geográficas mais acen-tuadas pela queda de diques edeposição anual sedimentos. Todasessas modificações cíclicas exercemparticular influência sobre a concen-tração do oxigênio dissolvido na colunade água dos rios e lagos dessa região.
Estudos sobre mudanças sazonaisde variáveis físico-químicas e químicasem lagos de várzea têm mostrado queeles podem ser quimicamente diferentes entre si; que a qualidade de suaságuas pode ser consideravelmen tediferente da do rio e que um mesmolago, quase sempre, apresentadiferentes padrões no conteúdo deeletrólitos em diferentes anos eperíodos de um mesmo ano (Schmidt,
et al
et al
-
-
44
Darwich et al.

1972a,b; Junk, 1973; Furch .,1983; Darwich, 1995; Kern, Darwich,1997, 1999).
Sabe-se que, independente dasmodificações hidrológicas do rioprincipal, é de fundamental importân-cia a contribuição de pequenosigarapés que fluem da terra firme paraos lagos de várzea localizados nasmargens dos rios. Nos períodos demaior afluência, as águas destesigarapés apresentam elevados teoresde oxigênio dissolvido e grandeescassez de eletrólitos e nutrientes.De um lado, podem diluir o teor dassubstâncias dissolvidas e, de outro,melhorar as condições de oxigênio daságuas dos lagos de várzea. A influênciadessa contribuição sobre o lago é tantomaior quanto maior for o volume daafluência ou, quanto menor for ovolume do lago. Além disso, se o lagoestiver mais próximo da terra firmeessa influência pode ser ainda maiorporque a área de várzea drenada serámenor. Nesse caso, há, portanto,menor possibilidade de consumo de Ono trajeto e menor quantidade dematerial lixiviado do solo devido aomenor percurso na área de várzea ricaem nutrientes. A maior parte dessaágua da terra firme é dependente daschuvas locais e sem correlação com aoscilação do nível da água dos rios. Noslagos de ilhas do Solimões/Amazonas,e em outros, obviamente a água daschuvas nas proximidades, antes dechegar ao lago passa por solos ricos emnutrientes oriundos da deposição desedimentos do rio. No seu trajeto,além do empobrecimento dos soloscom maior quantidade de nutrientes, a
et al
&
2
água favorece os processos de decomposição,principalmente do material vegetal morto provocandolixiviação e liberação de íons e nutrientes que sãotransportados ao lago.
Soma-se a isso, uma série de processos internos dolago, como o desenvolvimento e a decomposição dosprodutores primários e os processos de ressuspensão dossedimentos, que desempenham um papel decisivo noenriquecimento da água. De acordo com Furch .(1983), o conteúdo de elementos e nutrientes de um lagode várzea é muitas vezes atribuído às macrófitasaquáticas e semi-aquáticas cuja produção primária ebiomassa por unidade de área, excede àquela dofitoplâncton.
Os lagos da bacia do rio Negro também estão sujeitosà variação de nível do rio, bem como recebem aafluência de pequenos igarapés de floresta. Da mesmaforma, as águas destes igarapés apresentam-se bemoxigenadas e com baixa concentração de íonsdissolvidos, diluindo ainda mais a água dos lagos. Assimcomo na várzea, sob esse aspecto, a maior parte dessaágua depende das chuvas nas bacias desses igarapés enão da oscilação do nível do rio. A diferença marcante éque, mesmo no trajeto final desses igarapés, onde fluempor solos de áreas inundáveis, não há arraste de solosricos em nutrientes depositados junta-mente com ossedimentos na inundação anterior, como acontece navárzea. Além disso, pode ser desprezível a lixiviação deíons de material vegetal oriundo da decomposição demacrófitas aquáticas que cresceram durante a enchentee cheia, pela quase ausência dessa vegetação na bacia dorio Negro.Aconseqüência imediata desse fato no períodode descida das águas e na seca é a diminuição do teoriônico de suas águas nas camadas mais superficiais,mesmo com a decomposição do material vegetal mortoda floresta transportado ao lago pelas chuvas e igarapés.Ainda como resultado dessa atividade há o transporte deágua com maior concentração de oxigênio dissolvido,pelo baixo consumo desse gás no trajeto até o lago e pelaadição mecânica de O durante o percurso à bacialacustre. Nessa fase o aumento da concentração deoxigênio dissolvido pode também estar favorecido pelo
et al
2
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
45

aumento da transparência dos lagos tornando oambiente mais propício à produtividade primária.
Para o conhecimento ecológico de um sistemaaquático são necessárias informações básicas sobre suadinâmica. Essa dinâmica, de acordo com Ganf Horne(1975, Esteves ., 1988), se encontra emrelação direta com a posição geográfica do ecossistema,tendendo a ser menos evidenciada em latitudesmenores. Desta forma, até mesmo os modelos propostosdentro da própria Amazônia para o entendimento deambientes hídricos devem ser observados especificamente para os locais e épocas onde foram desenvolvidos.Como o Tupé recebe a afluência de pequenos igarapés defloresta e está submetido à flutuação do nível do rioNegro, objetivou-se investigar a dinâmica espaço-temporal do oxigênio dissolvido na água desse lago e nasua afluência durante um ciclo hidrológico. Para melhorentender essa dinâmica, foram medidas variaçõesnictemerais do O na coluna de água do lago no mesmoperíodo, o que poderá fornecer informações valiosassobre o funcionamento e a ecologia de lagos naAmazônia.
O Tupé é um lago de área inundável com margensíngremes em um vale em forma de “V”, bloqueado porbancos de areia na sua foz, com superfícieaparentemente bem protegida dos ventos. Está namargem esquerda do rio Negro, o maior rio de águaspretas do mundo em bom estado de conservação.Apresenta características hidrológicas semelhantes àmaioria dos lagos de área inundável da bacia desse rio.Há presença de estreitas faixas de igapó nas áreas decabeceiras, nas laterais, nos igarapés e vales no entornodo lago. Há também registros de poucos indivíduos de
sp. Durante a enchente, cheia e vazante aqualidade de sua água aproxima-se mais da do rio Negro,visto que acima da cota 19m a.n.m. o rio aflui ao lago eambos apresentam flutuação de nível semelhante. Na
&apud et al
-
Utricularia
2
Material e Métodos
Área de Estudo - Lago Tupé
cheia as profundida-des variam de 10ma 15m. No período de águas baixas(seca), quando a cota do rio é menorque 19m a.n.m., há apenas afluênciado lago ao rio e pode-se dizer que olago isola-se do rio, visto não haverqualquer possibilidade de influênciado rio sobre lago. Nesta fase do ciclohidrológico, sua água assemelha-sequimicamente às águas dos igarapés deterra firme que afluem à bacia do lagoe, à água da precipitação pluviométri-ca. No pico dessa fase sua profundida-de varia de 0,5m nas áreas de cabecei-ra, à 4,5m, em média, na regiãocentral, que é o encontro dos eixosprincipal e secundário do lago. Essedesnível no fundo do lago, dá umdeclive no eixo principal de cerca de1,3m.km das cabeceiras à regiãocentral, maior no eixo secundário.Detalhes sobre a área do lago sãoencontrados em Aprile . (2005a,neste volume).
O teor de oxigênio dissolvido foimedido durante um ciclo hidrológi-coque compreendeu a seca (águasbaixas) de 2001, e a enchente, cheia evazante de 2002. As datas de registrosnesses períodos foram, respectiva-mente, 21 de novembro, 08 de março,09 de junho e 28 de setembro. Asmedidas foram feitas com a finalidadede acompanhar a dinâmica espacial eas variações nictemerais e sazonais dooxigênio na coluna de água do lago.Além disso, a concentração de O foimedida a) na coluna d'água dos cincoprincipais igarapés de floresta que
-1
et al
Período daAmostragem
2
46
Darwich et al.

afluem ao lago; b) no canal de ligaçãodo lago ao rio e, c) no rio Negro,visando quantificar a participação daafluência de acordo com a fase do ciclohidrológico.
O oxigênio foi medido em 12 (doze)estações de coleta: uma no trechoinferior de cada um dos cincoprincipais igarapés; uma no canal deconexão do lago ao rio (EC11); uma norio Negro na parte central da calha dorio à montante do canal (EN12) e, 5(cinco) no lago, sendo duas em cadaum dos dois eixos do lago e uma naparte central de encontro desseseixos. Nesta última, a estação centraldo lago (ET10), foi medida a variaçãonictemeral da concentração dooxigênio dissolvido no mesmo período.Os principais igarapés e as respectivasestações de coleta foram: igarapés daHelena (EI2), das Pedras (EI3), daCachoeira (EI4), do Monteiro (EI6) e daTerra Preta (EI8). As estações no lagocompreenderam, além da estaçãocentral, as estações nas cabeceirasprincipal (ET1) e secundária (ET7), e asestações laterais ET5 e ET9,respectivamente, nos eixos principal esecundário. São estações aproximadamente eqüidistantes da estação ET10,sendo a primeira entre os igarapés daCachoeira e do Monteiro e a outra,abaixo do igarapé da Terra Preta(Darwich ., 2005, neste volume;Aprile ., 2005b, neste volume).
O nível da água no rio Negro varioude 16,81m a.n.m. (metros acima do
Estações de Coleta
Nível da Água
-
et al
et al
nível do mar) na seca de 2001, à 28,91m a.n.m., no picoda cheia de 2002. No período final desse estudo (vazantede 2002), o nível d'água era de 23,25m a.n.m. O nível deenchente no dia de coleta foi de 23,91m. Na seca de 2002(novembro), o nível do rio baixou até a cota 17,19ma.n.m. e pode-se verificar o pulso de inundação do rioNegro (Fig. 3), com curva monomodal caracterís-tica,como as registradas para o rio Amazonas. No período deseca a profundidade do rio na estação EN12 variou emtorno de 30m e a largura entre 3000 e 3500m.Informações adicionais sobre a flutuação do nível das daáguas em outro ciclo hidrológico estão também descritasem Aprile Darwich (2005, neste volume) e Aprile .2005, neste volume).
Durante todo o período dessa pesquisa aprofundidade do lago variou com uma amplitude máximade cerca de 10m. Pode-se verificar que tanto na seca de2001 quanto na de 2002, com diferentes níveis do rio, aprofundidade na estação central do lago (ET10) foisempre a mesma (Fig. 3). A curva indica que na seca de2001, o lago ficou isolado do rio por um período bemmaior, e ainda assim a profundidade máxima do lagopouco oscilou em torno de 4,5m. O fato da profundidadena estação central ser sempre em torno de 4,5m, aindaque o rio esteja muito abaixo da cota 19m a.n.m.,acontece porque o lago “isola-se” do rio abaixo dessacota. A cota 19m a.n.m. (18,7m) corresponde ao nívelaltimétrico do canal de ligação do lago ao rio. Durante oisolamento há apenas afluência do lago ao rio.Aágua queflui é resultante, principalmente, dos principais igarapésacima mencionados e das afluências das cabeceirasprincipal e secundária do lago, além da água deprecipitação pluviométrica diretamente no lago e na suabacia de captação.
Como tem sido dito, o pulso de inundação exerceinfluência marcante sobre as áreas inundáveis naAmazônia. No lago Tupé a variação da amplitude doperíodo do ciclo hidrológico com água abaixo da cota19m tem importância sobre a variação das condiçõeshidroquímicas do lago, porque nesta fase do ciclo não háinfluência do rio sobre a bacia lacustre. Durante esseperíodo as características da água são resultantes daafluência ao lago e dos processos internos na bacia
& et al
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
47

lacustre. Há modificações evidentes em variáveislimnológicas, como a transparência da água,temperatura, produtividade primária, teor de gáscarbônico, potencial hidrogeniônico, condutividadeelétrica, demandas química e bioquímica de oxigênio e,particularmente, na concentração do oxigêniodissolvido, objetivo central dessa abordagem.
A concentração de oxigênio dissolvido foi medida nacoluna de água de todas as estações de amostragem dolago, a cada 0,5m, da superfície à profundidade máxima.No rio Negro as medidas foram, a cada metro, até 10m deprofundidade. Para o estudo da variação nictemeral asmedidas foram realizadas na coluna de água da EstaçãoCentral, em intervalos de três horas, durante 24h, emcada fase do ciclo hidrológico. A escolha da Estação
Medidas do Oxigênio
Central tomou como base a geomorfo-logia do local, de tal maneira que aamostragem coincidisse com a con-fluência dos braços principal (longitu-dinal) e secundário (transversal) dolago, e abrangesse a maior área livre emais profunda da bacia lacustre. Essaregião central é a área de captação detoda a afluência, tanto do rio quantodos igarapés, à bacia lacustre,podendo-se detectar nessa região,grandes transformações no ambienteem estudo. Nesse ponto foramregistradas as maiores profundi-dadesda bacia do lago.
As medidas do oxigênio dissolvidotiveram por base as recomendações doPrograma Biológico Internacional para
Figura 3. Nível da água no rio Negro medido no Porto de Manaus e no lago Tupé, na Estação Central(ET10). De outubro de 2001 a março de 2003.
48
Darwich et al.
Pro
fun
did
ade
lag
oT
up
é(m
)
13
16
19
22
25
28
31
out/
01
nov
d ez
jan/
02 fev
ma
r
ab
r
ma
i
j un
j ul
ag
o
set
o ut
n ov
dez
j an/
03 fev
ma
r
(Meses)
Nív
eld
'ág
ua
rio
Neg
ro
(ma.
n.m
.)
-1,5
1,5
4,5
7,5
10,5
13,5
16,5
nível máximo 2002rio = 28,91m a.n.m.
lago = 14,5m
Nível d'água rio Negro Nível d'água lago Tupé
nível mínimo 2001rio = 16,81m a.n.m.
lago = 4,5m
nível mínimo 2002rio = 17,19m a.n.m.
lago = 4,5m

ambientes aquáticos (Golterman, 1978). Os teores foram obtidos em
mg.L-1 e em percentagem desaturação por meio de leitura diretacom um oxímetro digital WTW modeloOXI-197, com sensibilidade respectivade 0,1 e 0,01. Paralelamente, asmedidas obtidas no potenciômetroforam comparadas com determinaçõesanalíticas pelo método de Winklermodificado segundo recomendaçõesde Mackereth . (1978) e WetzelLikens (2000).
Foi descrito que, o lago Tupéencontra-se praticamente ao nível domar e em uma localização equatorialcom elevadas temperaturas. Como emtemperaturas mais elevadas, menoresconcentrações de O representammaiores percentagens de saturação,uma mesma quantidade em mg.L temsignificados diferentes para regiões delatitudes diferentes. Dessa forma, aimportância fisiológica desse gásdissolvido para a comunidade aquáticadeve estar associada às suas medidasem percentagem de saturação.
Durante o ciclo hidrológico pesqui-sado as camadas mais superficiais dacoluna de água, em torno da zonaeufótica do lago Tupé e principaisafluências, apresentaram elevadasconcentrações de O . Excetuando-se orio Negro, com distribuição homogê-nea na coluna de água, nas camadasmais profundas dos outros ambientesos teores de O foram consideravelmente reduzidos. Houve variação de
et
al.
et al &
2
2
-1
Resultados e Discussão
Oxigênio Dissolvido
2 -
acordo com o local, com a posição no lago e na colunade água, bem como com o período do ciclo. Asvariações sazonais e diárias evidenciaram, em todasas fases, a existência e epilímnio e hipolímnio bemdefinidos. A localização geográfica do lago, seuposicionamento em relação ao rio, suas margensíngremes e com foz bem protegida, além daselevadas temperaturas, são alguns dos fatoresconsiderados de importância quanto à distribuiçãoespacial (longitudinal e vertical), temporal (sazonal)e nictemeral do O , bem como quanto à circulação decamadas naquele ambiente.
As medidas de oxigênio dissolvido nos diferentesambientes evidenciaram, em todas a fases do ciclohidrológico, elevadas concentrações de O nascamadas mais superficiais (Figs. 4 a 7). Verifica-se,entretanto, em todo o ambiente, uma nítidadiferença entre os teores registrados na cheia e nasoutras fases do ciclo. Na cheia, com profundidadesentre 10 e 15m, as maiores concentrações de O nabacia lacustre estiveram em torno de 50% desaturação nas camadas mais superficiais da áreacentral do lago, no rio e no canal do lago (Fig. 6). Nasoutras regiões do lago e nos igarapés os teoressuperficiais oscilaram em torno de 30% de saturação.Na seca, com profundidades entre 0,5m e 5m, asaturação de O dissolvido atingiu seus maioresvalores e manteve-se entre 100% e 150% nas camadassuperficiais, principalmente nas áreas mais rasas e naregião central do lago (Fig. 4). No rio Negro e nasoutras regiões da bacia lacustre a saturaçãomanteve-se entre 80 e 100% de saturação. Nesta fasedo ciclo, nas estações com profundidades até 1,0m,as concentrações foram elevadas, mesmo no fundo.No rio, durante todo o período, praticamente nãoexistiu diferenças entre as concentrações registradasna coluna de água. Ainda nesta fase, apenas naestação Central e nas laterais a esta estação(principal e secundária) foi registrada a existência deepilímnio e hipolímnio bem definidos. Desta forma,
d
2
2
Variação Espaço-Temporal
2
2
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
49

Figura 4. Variação espacial da concentração de oxigênio dissolvido no lago Tupé,margem esquerda do Rio Negro (Manaus-AM) e afluências. Seca 2001.
TupéCabeceira Principal
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Igarapé daHelena
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Igarapé dasPedras
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Igarapé daCachoeira
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
TupéLateral Principal
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Igarapé doMonteiro
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
TupéCentral
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
TupéLateral Secundária
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Igarapé daTerra Preta
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
TupéCabeceira Secudária
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Tupé Canal
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Rio Negro
0,0
1,5
3,0
4,5
0 50 100 150(%)
(m)
Darwich et al.
50

TupéCabeceira Principal
02468
10
0 50 100 150(%)
(m)
Igarapé daHelena
02468
10
0 50 100 150(%)
(m)
Igarapé dasPedras
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé daCachoeira
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéLateral Principal
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé doMonteiro
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéCentral
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéLateral Secundária
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé daTerra Preta
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéCabeceira Secudária
0
2
4
6
8
10
0 50 100 150(%)
(m)
Tupé Canal
0
2
4
6
8
10
0 50 100 150(%)
(m)
Rio Negro
0
2
4
6
8
10
0 50 100 150(%)
(m)
Figura 05. Variação espacial da concentração de oxigênio dissolvido no lago Tupé,margem esquerda do Rio Negro (Manaus-AM) e afluências. Enchente 2002.
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
51

TupéCabeceira Principal
0
3
6
9
12
15
0 50 100 150(%)
(m)
Igarapé daHelena
0
3
6
9
12
15
0 50 100 150(%)
(m)
Igarapé dasPedras
0
3
6
9
12
15
0 50 100 150(%)
(m)
Igarapé daCachoeira
0
3
6
9
12
15
0 50 100 150(%)
(m)
TupéLateral Principal
0
3
6
9
12
15
0 50 100 150(%)
(m)
Igarapé doMonteiro
0
3
6
9
12
15
0 50 100 150(%)
(m)
TupéCentral
0
3
6
9
12
15
0 50 100 150(%)
(m)
TupéLateral Secundária
0
3
6
9
12
15
0 50 100 150(%)(m
)
Igarapé daTerra Preta
0
3
6
9
12
15
0 50 100 150(%)
(m)
TupéCabeceira Secudária
0
3
6
9
12
15
0 50 100 150(%)
(m)
Tupé Canal
0
3
6
9
12
15
0 50 100 150(%)
(m)
Rio Negro
0
3
6
9
12
15
0 50 100 150(%)
(m)
Figura 6. Variação espacial da concentração de oxigênio dissolvido no lago Tupé,margem esquerda do Rio Negro (Manaus-AM) e afluências. Cheia 2002.
Darwich et al.
52

TupéCabeceira Principal
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé daHelena
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé dasPedras
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé daCachoeira
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéLateral Principal
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé doMonteiro
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéCentral
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéLateral Secundária
0
2
4
6
8
10
0 50 100 150(%)
(m)
Igarapé daTerra Preta
0
2
4
6
8
10
0 50 100 150(%)
(m)
TupéCabeceira Secudária
0
2
4
6
8
10
0 50 100 150(%)
(m)
Tupé Canal
0
2
4
6
8
10
0 50 100 150(%)
(m)
Rio Negro
0
2
4
6
8
10
0 50 100 150(%)
(m)
Figura 7. Variação espacial da concentração de oxigênio dissolvido no lago Tupé,margem esquerda do Rio Negro (Manaus-AM) e afluências. Vazante 2002.
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
53

na área central do lago, essas duas regiões ocuparamcerca 20% e 40% da coluna de água, respectivamente. Oepilímnio, supersaturado, e o hipolímnio com teores emtorno de 10% de saturação. No metalímnio a saturação doO diminuiu de cerca de 80 15% de saturação. Assimsendo, na área mais profunda do lago ficou caracterizadaa existência de oxiclina com perfil clinogrado clássico.Esses dados sugerem a existência de elevada produçãoprimária e falta de circulação da coluna de água do lagono período de águas mais baixas. Este fato não implica,necessariamente, em falta de movimentos na coluna deágua. Sugere-se, todavia, a existência de fluxos laminares nos eixos principal e secundário do lago, dascabeceiras ao canal de ligação. Na seca, certamente osfluxos são nas camadas mais superficiais, menos densas,em conseqüência do maior aquecimento superficial e damenor amplitude de temperatura entre pequenascamadas. Esses fluxos são provenientes da afluência àbacia lacustre e devem seguir estratos diferentes deacordo com o nível da água. Durante a época em que nãohá entrada de água do rio ao lago, a principal contribuição de água à bacia lacustre é dos igarapés e das própriascabeceiras, além da pluviométrica. Embora asconcentrações de O2 dessa afluência sejam elevadasnesse período, o hipolímnio da região central apresentateores abaixo de 10% de saturação. Deve ser o resultadoda falta de circulação vertical e do maior consumo porrespiração e decomposição da matéria orgânica que seprecipita no fundo e pode variar em concentração deacordo com a hora do dia.
Na enchente (Fig. 5) e cheia (Fig. 6), os dadosinduzem confirmar a existência de fluxos laminares dascabeceiras à foz do lago em estratos mais profundos dacoluna de água. Em estações à montante da Central, commaior freqüência no eixo longitudinal do que notransversal, foram medidas maiores concentrações deoxigênio em camadas mais profundas de regiões dohipolímnio. Embora sejam períodos de entrada de águado rio com maior teor de oxigênio, não foi registrado, nocanal ou mesmo na estação Central, alterações naquantidade de O que se pudesse concluir em ummovimento de camadas profundas no sentido rio-lago.
2 a
-
-
2
Especialmente na enchente quando ascamadas superficiais do lago parecemestar sendo influenciadas pela entradade água do rio com maior concentraçãode oxigênio. Além disso, em todos osperíodos a mistura na coluna d'águadas estações no lago parece estarrestrita às camadas superficiais, limi-tada, no máximo à zona eufótica comas maiores concentrações. Na enchen-te a quantidade de O esteve entre 100e 140% de saturação nas camadas maissuperficiais de todo o ambiente,inclusive do rio. Os dados de vazantemostraram, apenas na estaçãoCentral, pequeno aumento no teor deoxigênio em camadas do hipolímnio.Nesta fase, o teor de O foi maior doque no período anterior e menor doque na enchente e seca. Nota-seainda, que a região do epilímnio commaiores concentrações de oxigênio foiconsideravelmente reduzida, oscilando em um estrato de cerca de 1,5m.Nela, os teores variaram entre 40 a 80%de saturação. No rio os valores forammenores e relativamente constantesem torno 50% de saturação, semelhan-tes aos verificados na cheia.
O estudo de variação nictemeral nacoluna de água em ambientesaquáticos pode esclarecer fenômenosnão perfeitamente entendidos emanálise sazonal. Em lagos equatoriaiscomo o Tupé, alterações diárias devariáveis ambientais podem ser defundamental importância para oentendimento do comportamento decomunidades aquáticas. Nesta região,
2
2
-
Variação Nictemeral
54
Darwich et al.

onde o fotoperíodo tende à uniformi-dade durante o ano, com amplasvariações diárias de luz e temperatura,permite supor a existência de umagrande amplitude de variação no teorde variáveis como o oxigênio dissolvidona coluna de água.
A análise de variações diáriasdo O na região mais profunda e demaior área livre do lago Tupé, EstaçãoET10, mostrou a existência de acentuada estratificação no teor de saturaçãodesse gás durante um ciclo hidrológico(Figs. 8 a 11). Nesse período, as concentrações variaram da supersaturação no epilímnio, a teores considera-velmente reduzidos e, às vezes, àanoxia na camada mais inferior dohipolímnio. No metalímnio o gradientefoi sempre acentuado, freqüentemen-te partindo da supersa-turação, nolimite inferior do epilímnio, à valoresabaixo de 5% de saturação, no limitesuperior de hipolímnio.
2
-
-
-
Durante as águas baixas (Fig. 8) asmaiores concentrações foram registra-das à tarde (entre 15 e 18h) noprimeiro metro de profundidade.Desse horário até às 6h, houve diminui-ção, com aumento lento até às 12h.Nota-se, entretanto, que a colunad'água permanece estratificada aolongo de 24h, e praticamente anóxicaentre 3m e a profundidade máxima(5m). Vale salientar que mesmodurante o período noturno todo oepilímnio, formado por uma camadaem torno de 1,5m, manteve-se comteores de O acima de 80% desaturação, até mesmo às 6h (88%). Adiminuição no teor de oxigênio nascamadas mais superficiais no período
2
noturno parece estar mais associada à perda de O pordifusão para a atmosfera, do que pelo consumo ematividades respiratórias e de decomposição de matériaorgânica. Isto porque foi muito mais intensa a diminuiçãoda saturação no início da noite (até às 21h), quando oepilímnio estava supersaturado, do que no restante doperíodo no escuro. Nas primeiras horas a saturação foireduzida de 140% a cerca de 100%. Nos horários seguintesa perda foi em torno de apenas 10% e esta sim, pode seratribuída à respiração e oxidação. Por outro lado, adecomposição certamente é mais intensa nas camadasmais profundas, visto que, após as 21h a metade inferiorda coluna de água do lago encontrava-se com menos de1% de saturação de oxigênio. Ressalta-se ainda, que avariação do O no epilímnio sugere circulação apenasnessa camada e intensa produtividade na zona eufóticado lago, principal-mente em sua região mais superficial.Dela, o O pode estar alcançando, por difusão, camadasmais abaixo em direção à zona afótica, em especial noshorários com maiores concentrações de oxigênio.
2
2
2
2A migração de O para o hipolímnio em outras fases dociclo deve estar também favorecida pela afluência, umavez que os dados de temperatura mostraram ser o lagoTupé um ambiente de difícil circulação (Darwich .,2003). Esses autores registraram, na seca, amplitudes devariação da temperatura de até 6°C com menoresvalores em torno de 27°C. Nessa faixa de temperatura adiferença de densidade é consideravelmente grandequando comparada às diferenças de temperaturas emregiões temperadas ou mesmo subtropicais e tropicais.Além disso, e visto que o lago Tupé apresenta-se commargens íngremes e floresta que o protegem dos ventos,a estratificação, tanto térmica quanto do O , parece serbastante estável e duradoura. No entanto, é umaestratificação compatível com atividades biológicas nomeio hídrico, o que pode ser comprovado pela presençade animais planctônicos na coluna d'água. A evidênciados fluxos laminares no lago deve proporcionarcondições vitais nas camadas mais profundas, ainda queseja com baixos valores de pH e reduzidas concentraçõesde oxigênio. Logo, a estratificação no Tupé não significa
et al
2
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
55

15h
117,7
29,616,9
8
148,600,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
18h
11983,5
71,318,8
1,7
135,5
130,7
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
21h
101,6
102,2
39,14,4
0,1
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
24h
98,5
98,788,6
0,5
0,1
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
03h
93,3
92
15,80,6
0
0,2
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
06h
87,8
88,4
0,6
0,1
0
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)P
rofu
ndid
ade
(m)
OD
09h
91,4
90,6
5,30,6
0,2
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD12h
92,7
94,279,7
8,10,40,1
0
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
Figura 8. Variação nictemeral da concentração de oxigênio dissolvido (OD) empercentagem de saturação (%sat). Estação Central (ET10), Lago Tupé, margemesquerda do Rio Negro (Manaus-AM). Período de seca: 20-21.novembro.2001 (de 15 às12h).
Darwich et al.
56
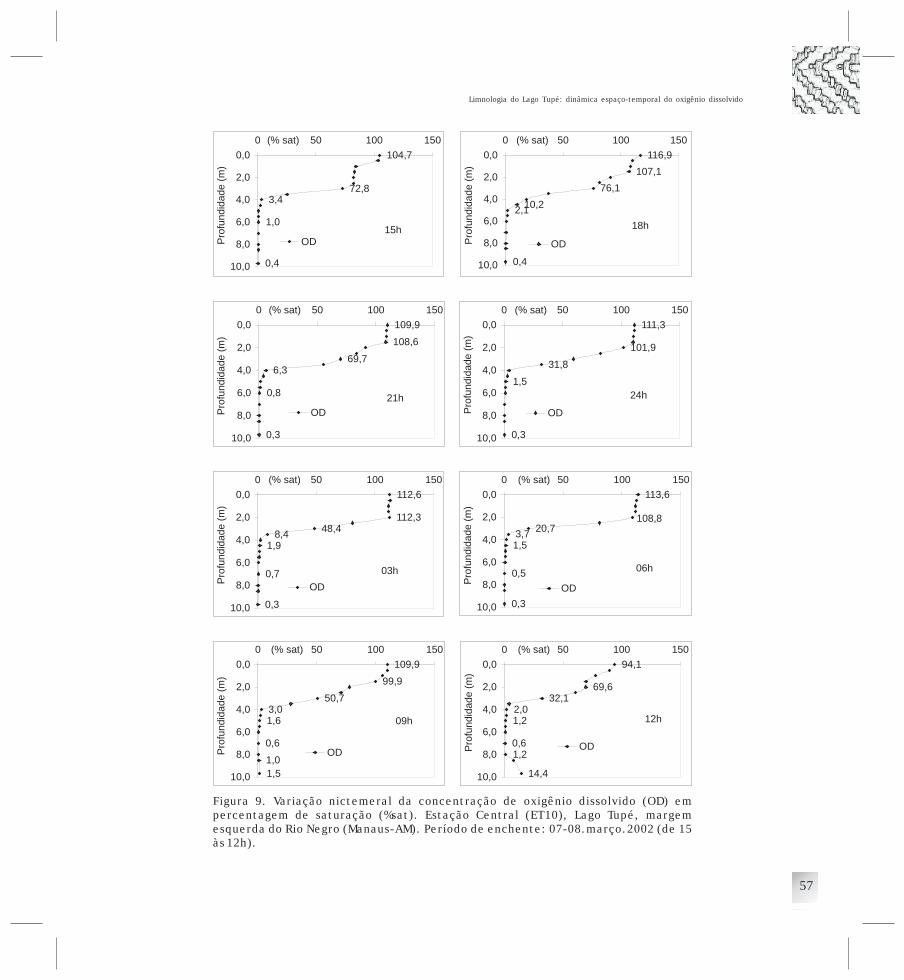
15h
104,7
72,83,4
1,0
0,4
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
18h
116,9
107,1
0,4
10,22,1
76,1
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
21h
109,9
108,6
69,76,3
0,8
0,3
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
24h
111,3
101,9
31,8
1,5
0,3
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
03h
112,6
8,41,9
0,7
0,3
48,4112,3
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
06h
113,6
20,7
1,5
0,5
0,3
108,8
3,7
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)P
rofu
ndid
ade
(m)
OD
09h
109,9
99,9
3,01,6
0,6
1,01,5
50,7
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
12h
94,1
69,6
0,61,2
14,4
1,22,0
32,1
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
Figura 9. Variação nictemeral da concentração de oxigênio dissolvido (OD) empercentagem de saturação (%sat). Estação Central (ET10), Lago Tupé, margemesquerda do Rio Negro (Manaus-AM). Período de enchente: 07-08.março.2002 (de 15às 12h).
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
57

15h
53,1
40,010,6
2,8
0,8
0,3
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
18h
57,1
0,6
20,636,5
1,0
2,3
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
21h
52,8
0,5
10,8
6,3
0,7
1,1
1,6
38,4
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
24h
49,0
48,6
18,7
2,2
0,8
4,1
1,1
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
03h
47,0
15,5
2,3
2,3
3,9
45,2
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
06h
44,3
31,16,2
4,3
2,3
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)P
rofu
ndid
ade
(m)
OD
09h
49,8
38,0
5,6
5,4
7,5
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
12h
44,2
0,6
0,3
2,233,1
0,0
3,0
6,0
9,0
12,0
15,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
Figura 10. Variação nictemeral da concentração de oxigênio dissolvido (OD) empercentagem de saturação (% sat). Estação Central (ET10), Lago Tupé, margemesquerda do Rio Negro (Manaus-AM). Período de cheia: 08-09.junho.2002 (de 15 às12h).
Darwich et al.
58

15h
68,2
5,1
1,8
0,9
1,6
47,14,6
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
18h
68,8
0,7
0,9
3,05,2
43,4
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
21h
68,1
0,8
0,8
1,7
16,93,5
67,80,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
24h
62
3,3
0,6
0,81,6
8,360,9
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
03h
54,8
1,4
0,7
0,8
3,48,953,8
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
O
06h
42,5
1,4
0,8
1,3
4,32,2
38,4
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
09h
50,6
0,8
0,91,74,2
44,6
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
12h
57,7
1,1
46,24,23,2
33,1
0,7
0,0
2,0
4,0
6,0
8,0
10,0
0 50 100 150(% sat)
Pro
fundid
ade
(m)
OD
Figura 11. Variação nictemeral da concentração de oxigênio dissolvido (OD) empercentagem de saturação (% sat). Estação Central (ET10), Lago Tupé, margem esquerdado Rio Negro (Manaus-AM). Período de vazante: 27-28.setembro.2002 (de 15 às 12h).
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
59

necessariamente estagnação da coluna de água.No período de cheia (Fig. 10), assim como na seca, a
oxiclina na coluna de água foi bastante estável, porémcom concentrações de oxigênio bem menores nascamadas mais superficiais, do que àquelas do período deáguas baixas. Além dos menores teores de O noepilímnio, a diferença marcante entre essas duas fasesdo ciclo hidrológico está na amplitude percentual daszonas eufótica e afótica e a localização, do metalímnio.Desta forma, verifica-se que enquanto no período deáguas baixas a zona eufótica representava cerca de 60%da coluna de água, na cheia essa região era de apenas20%. Em ambos os períodos a amplitude dessa zonaoscilou em torno de 3m, maior na seca. A transparênciade Secchi foi sempre um pouco menor no rio. Na seca ometalímnio aparece dentro da zona eufótica e, na cheia,abaixo dessa zona. A conseqüência mais óbvia dessasobservações é a elevada concentração de O na fase deáguas baixas e, provavelmente, o maior consumo deoxigênio no fundo. Esta constatação pode ser o resultadodo próprio aumento da produtividade primária do localem conseqüência da maior disponibilidade de luz, aindaque os nutrientes fossem igualmente disponíveis emambos os períodos. Vale notar que, independente daanálise iônica nas águas do rio Negro e do lago, essasverificações corroboram a afirmação de que no períodode cheia o rio invade o lago. Nessa fase a influênciafluvial pode ser marcante, mas não o suficiente paraquebrar a oxiclina no lago (Fig. 6).
Nos períodos de enchente e vazante que são as fasesintermediárias do ciclo hidrológico, as modificações nadistribuição do oxigênio na coluna de água tambémforam relacionadas com a oscilação no nível da água. Emambos os períodos as profundidades máximas do lagooscilaram em torno de 10m, com o nível do rio em cercade 23,5m a.n.m. Durante essas fases do ciclo, manteve-se a estratificação do O , porém com variação naamplitude do epilímnio, bem oxigenado, e do hipolímniocom reduzidas concentrações de O . O metalímnio variouem localização na coluna d'água. Foram tambémregistrados os fluxos longitudinais detectados emdiferen tes horários da análise nictemeral (Figs. 9 e 11).
2
2
2
2
-
Na enchente (Fig. 9), com exceçãodas medidas às 12h com valores entre80 e 94% de saturação nas camadasmais superficiais, a água estevesupersaturada de O , principalmentedurante o período noturno com teoresacima de 100% até 2m de profundida-de. Verifica-se que a quantidademedida às 6h (114%) foi apenas umpouco menor do que a maior concen-tração de oxigênio nesse período,registrada às 18h (117%). Ao contráriodo que pode ter ocorrido na seca, não épossível falar em grandes perdas de Opara a atmosfera durante a noite naenchente, uma vez que nesse horário aconcentração de oxigênio foi sempremaior com valores acima da saturação(em torno de 110%). Pelas concentra-ções registradas não se pode afirmarque houve redução de oxigênio emconseqüência do maior consumo porrespiração e oxidação da matériaorgânica na fase escura. Certamenteesses processos continuaram ocorren-do em toda a coluna de água e durantetodo o ciclo nictemeral, sendo aexplicação mais provável para amanutenção dos altos níveis de O nolago, a entrada de água do rio pelocanal. Nesse período foram registradaselevadas concentrações de O no rioNegro (120%) e no canal (140%) deligação do lago ao rio. Além disso, nãose pode excluir a possibilidade deprodução de O no escuro pormicroorganismos nesses ambientes.Durante esta pesquisa, experimentoscom demanda bioquímica de oxigênioem água do rio Negro, mostraram, comcerta freqüência, maiores concentra-
2
2
2
2
2
60
Darwich et al.

ções de O após cinco dias de incubaçãono escuro. Afora estas constatações,deve-se ressaltar que no lago, emtodos os momentos, as concentraçõesde O na enchente estiveram acima dasaturação nas camadas mais superfi-ciais, porém menores do que no rio ecanal.
Em qualquer época do ciclohidrológico o canal e o rio apresentamcaracterísticas lóticas, um ambientepouco propício à supersaturação. Porisso, devem ser ressaltadas as elevadasconcentrações de O registradas naenchente e a localização da estação demedidas no rio. Esta estação encontra-se cerca de 40km acima de sua foz e àmontante dela não há acidentesgeográficos (e.g., cachoeiras) quepossam provocar aeração mecânica norio. Considerando que acima e abaixodo ponto de coleta, o rio apresenta-seem alguns trechos com larguras acimade 10km, o acréscimo ou perda de Opoderia ocorrer por ondas emconseqüência de ventos temporáriosna área. Dessa forma, esses eventospoderiam contribuir para aumentardiscretamente o teor de O na água,mas a supersaturação mais evidentedeve ser esperada somente comoresultado de produção por fotossínte-se. Assim sendo, toda a área do cursoinferior do rio Negro deve estarfuncionando como um imenso lago comprodutividade primária fitoplanctôni-ca característica, pelo menos noperíodo de enchente. Medidas isoladasnesta área no início da vazante(18.08.2002) mostraram teores deoxigênio praticamente constantes
2
2
2
2
2
(39% saturação = 3mg.L ), nos primeiros 20m de umacoluna de água em torno de 40m. Nesse período, osvalores de pH foram consideravelmente reduzidos nasmaiores profundidades o que reforça a idéia de umgrande lago com uma zona afótica extremamenteredutora e variável de acordo com a fase do ciclohidrológico. É possível que essa região seja habitada poruma comunidade característica e perfeitamente bemadaptada à elevadas concentrações de substânciashúmicas.
Na vazante a profundidade da coluna de água do lagoe o perfil da distribuição do O durante 24 horas foramsemelhantes aos valores registrados na enchente, noentanto, a amplitude das concentra ções de oxigênio nascamadas mais superficiais foi menor (Fig. 11). Dacomparação entre essas fases verifica-se também quenelas o metalímnio ficou limitado à cerca de 1m deespessura, em torno de 10% da coluna de água. Noentanto, enquanto na enchente o metalímnio localizou-se na zona afótica (entre 3m e 4m), na vazante esteve nomeio da zona com luz fotossinteticamente ativa (entre1m e 2m). A importância dessa verificação é mostrar amaior ou menor participação da água do rio na bacialacustre e a influência do pulso de inundação sobre o teorde oxigênio no lago. Assim, os valores de O nas camadasmais superficiais do lago durante a enchente parecemmais indicar a entrada de água do rio, do querepresentarem uma produção lacustre. Isto porque asconcentrações lacustres são semelhantes às do rio,mesmo no período noturno quando poderia serregistrada queda acentuada no teor de oxigênio nacoluna d'água. Por outro lado, na vazante, ainda que commenores concentrações, a zona eufótica estava maior(em torno de 35% da coluna de água) parecendo indicar amaior participação de águas da afluência dos igarapés dabacia e da precipitação pluviométrica com maiortransparência. Assim, nessa época, o hipolímnio comcerca de 80% da coluna de água ocupou o terço inferiorda zona eufótica e toda a zona afótica do lago. Nesseestágio do ciclo, no qual o lago “caminha” para oisolamento do rio, a zona eufótica vai ampliando-se e asconcentrações de oxigênio vão aumentando. Este
-1
2
-
2
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
61

comportamento parece estar relacionado ao aumento daprodutividade primária do fitoplâncton como resposta aoaumento da transparência da água do lago, e certamenteda disponibilidade de nutrientes. Desta forma, aamplitude das variações sazonal, espaciais e nictemeraisda concentração de oxigênio dissolvido demonstramresponder claramente à entrada e saída de água no lago.De um lado, durante a enchente, cheia e vazante, aconcentração de oxigênio pode estar intimamente ligadaà variação do pulso de inundação do rio Negro e, deoutro, à maior contribui-ção da água dos igarapésdurante o período de águas baixas, ainda que o nível dorio não esteja abaixo da cota 19m acima do nível do mar.
A variação nictemeral média da concentração de Ona coluna de água na região central do lago e os teoresmédios para a coluna d'água em todo o ambienteevidenciam a amplitude das curvas, a variação com operíodo de observação e o padrão fixo de estratificaçãodurante um ciclo hidrológico (Figs. 12 à 14). Na seca,ainda que tenha sido registrada concentrações em tornode 150% de saturação durante o dia, o teor médio porprofundidade aparece ligeiramente menor do que naenchente. Verifica-se desta forma, que os desvios naseca são bem maiores no epilímnio o que demonstra aexistência de picos de produção durante à tarde emenores concentrações ao final do período noturno (Fig.8). Por outro lado, os teores registrados nas camadasmais superficiais durante a enchente, como já descrito,apresentaram menores variações em 24 horas. Este fatoassociado à ocorrência de maiores concentrações,mesmo ao final do período noturno, corrobora aafirmação de que o oxigênio medido no lago nesseperíodo tenha origem principalmente fluvial, comotambém está apresentado na Figura 13, maiorconcentração média na enchente e menor na cheia. Damesma forma, na cheia, com menores concentrações,parece ser importante a influência do rio sobre o lago, noentanto, é possível perceber um discreto declínio dasconcentrações de O nas camadas mais superficiais, aofinal do período noturno (Fig. 12). Esse declínio é muitomais evidente na vazante, período no qual são maiores asconcentrações durante a tarde e o decréscimo noturno
2
2
muito maior do que no períodoanterior. Os desvios nas camadas maissuperficiais e mais produtivas sãotambém maiores, mostrando que olago começa a funcionar independenteda contribuição do rio. É importantesalientar que durante todo o ciclohidrológico a concentração média deO no hipolímnio do lago foi muitobaixa (Figs. 12 e 14). Os elevadosdesvios (e coeficientes de variação)das concentrações diárias porprofundidade, mostram maioresalterações no teor de oxigênio dascamadas mais profundas. Essasvariações, certamente, são o resultadodos fluxos longitudinais, nos eixosprincipal e secundário do lago,transportando maiores teores daafluência à montante da estaçãocentral.
O resfriamento noturno registradoem muitos lagos tropicais eequatoriais, responsável por inversãode massas de água e a conseqüenteinfluência sobre a distribuição denutrientes e organismos planctônicosnesses ambientes, não foi detectadono lago Tupé (Darwich ., 2003). Emum ciclo nictemeral esses autoresregistraram as maiores variações(2,3°C) nas camadas mais superficiaisdurante o período de águas baixas.Essa diferença de temperatura(máxima de 33,1°C) não foi suficientepara provocar homotermia na colunad'água com amplitude de variaçãomáxima de 5,8°C no perfil. Nemmesmo no período de cheia quando asdiferenças foram as menores,certamente como conseqüência daelevada diferença de densidade, entre
2
et al
62
Darwich et al.

SECA 20015 m
0
2
4
6
0 30 60 90 120 150(% sat)
Prof
undi
dade
(m)
de 0 a 148,6 % sat.20-21 novembro
ENCHENTE 20029,7 m
0
2
4
6
8
10
0 30 60 90 120 150(% sat)
Prof
undi
dade
(m)
de 0,3 a 116,9 %sat.07-08 março
CHEIA 200214,5 m
0
4
8
12
16
0 30 60 90 120 150(% sat)
Prof
undi
dade
(m)
de 0,3 à 53,1 %sat.08-09 junho
VAZANTE 20029,6 m
0
2
4
6
8
10
0 30 60 90 120 150(% sat)
Prof
undi
dade
(m)
de 0,7 à 68,8 %sat.27-28 setembro
Figura 12: Variação da concentração média de oxigênio dissolvido (% sat.) nacoluna de água na Estação Central (ET10) do lago Tupé para o ciclo hidrológico2001/2002. Medidas de 3/3 horas das 15 às 12h.
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
63

Seca (água baixa) de 2001
0
30
60
90
120
150
ET1 EI2 EI3 EI4 ET5 EI6 ET10 ET9 EI8 ET7 EC11 EN12
Estações de coleta
O2
%sa
t.
Enchente de 2002
0
30
60
90
120
150
ET1 EI2 EI3 EI4 ET5 EI6 ET10 ET9 EI8 ET7 EC11 EN12
Estações de coleta
O2
%sa
t.
Cheia de 2002
0
30
60
90
120
150
ET1 EI2 EI3 EI4 ET5 EI6 ET10 ET9 EI8 ET7 EC11 EN12
Estações de coleta
O2
%sa
t.
Vazante 2002
0
30
60
90
120
150
ET1 EI2 EI3 EI4 ET5 EI6 ET10 ET9 EI8 ET7 EC11 EN12Estações de coleta
O2
%sa
t.
Figura 13. Variação da concentração média de oxigênio dissolvido (% sat.) nacoluna de água de estações de coleta no lago Tupé (ET), nos principais igarapés(EI) da bacia do lago, no canal de ligação do lago ao rio (EC11) e no rio Negro(EN12). Períodos de seca de 2001 e enchente, cheia e vazante de 2002.
Darwich et al.
64

Oxigênio dissolvido (O2) - ENCHENTE 2002
(horário)15h 18h 21h 24h 3h 6h 9h 12h
0
2
4
6
8
1010
30 30 3020
20 2010
10 1060 60 6050 50 5040 40 4070
70 7080 808090 9090100
100100110 110
110
O2 % saturação
Oxigênio dissolvido (O2) - SECA 2001
(horário)15h 18h 21h 24h 3h 6h 9h 12h
0
1
2
3
4
5
8888
8
2424
24 24
24
1616
16 16
1672 72
72 7272
64 6464 64
64
5656
56 5656
4848
48 4848
4040
40 4040
3232 32 32
3288 88
88 88 88
80 8080 80 80
0
88
0
0
96 96
96112112
104 104128120
120136144
O2 % saturação
Oxigênio dissolvido (O2) - CHEIA 2002
(horário)15h 18h 21h 24h 3h 6h 9h 12h
0
3
6
9
12
15
2020
2020 20
1010
10 10 10
3030
3030 30
4040
4040 40
40
10
5050
10
O2 % saturação
Oxigênio dissolvido (O2)- VAZANTE 2002
(horário)15h 18h 21h 24h 3h 6h 9h 12h
0
2
4
6
8
10
30 30 30 30 3020 20 2020 2010 10 1010 10
40 40 40 40405050 50 50
6060 60
O2 % saturação
Figura 14. Variação nictemeral da concentração de oxigênio dissolvido. Colunade água da Estação Central (ET10), lago Tupé, margem esquerda do rio Negro.Períodos: seca de 2001 (20-21.novembro) e enchente (07-08.março), cheia (08-09.junho) e vazante (27-28.setembro) de 2002.
Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
65

66
Darwich et al.
graus, em temperaturas acima de27°C. Estas temperaturas foram asmenores registradas no lago Tupé. Asconcentrações de oxigênio medidas noTupé durante o período dessa pesquisaconfirmaram estas observações.Durante as fases, como descrito, asconcentrações de O no hipolímnio dolago foram sempre reduzidas. Deve-seressaltar, todavia, que embora o perfilseja estratificado quanto ao teor deoxigênio, a presença dos fluxoslaminares no sentido cabeceira-foz dolago, em decorrência da afluência,deve garantir condições vitais àcomunidade aquática no estrato maisinferior da coluna de água, ainda queseja com baixas concentrações deoxigênio. Além disso, a liberação ecirculação de nutrientes dos sedimen-tos para a coluna de água deve ocorrerpor diferenças de concentração; fluxosmais superficiais da afluência devemgarantir o suprimento de nutrientespara a produtividade primária ou,dependendo da fase do ciclo, dosuprimento direto de nutrientes e Opor meio do rio. Dessa forma, ainvestigação sobre a disponibilidade edinâmica do oxigênio no Tupé confirmaas conclusões de Darwich . (2003)em apresentá-lo como um lago dedifícil circulação, com evidências demisturas apenas nas camadas maissuperficiais em todas as fases do ciclohidrológico pesquisado, o que ocaracteriza, sob esse aspecto, comoum tipo de lago meromítico, come
2
2
et al
stratificação térmica e químicapermanente, diferente de muitos lagospolimíticos amazônicos.
Bibliografia Citada
Aprile, F. M., Darwich, A. J. 2005a. Modelosgeomorfológicos para o lago Tupé In: Santos-Silva, E.N.; Aprile, F. M.; Scudeller, V. V.; Melo, S. (Orgs.),
. Editora INPA, Manaus,AM.Aprile, F. M., Darwich, A. J.; Raposo, J. C. 2005b.
Considerações sobre a geoquímica e dinâmicasedimentar do lago Tupé. In: Santos-Silva, E. N.;Aprile, F. M.; Scudeller, V. V.; Melo, S. (Orgs.),
. INPA, Manaus,AM.Darwich, A. J. 1995.
. Tese de Doutorado. InstitutoNacional de Pesquisas da Amazônia/ Universidade doAmazonas (INPA/UA), Manaus (AM) 327pp.
Darwich, A. J.; Aprile, F. M.; Robertson, B. A. 2003.Estratificação térmica em um lago de água pretaamazônico: uma questão equatorial.
- CBL 2003. UniversidadeFederal de Juiz de Fora UFJF. Juiz de Fora (MG), 20-25.julho.2003. CD-ROM, N.39.
Darwich, A. J.; Aprile, F. M., Robertson, B. A. 2005.Variáveis limnológicas: contribuição ao estudoespaço-temporal de águas pretas amazônicas. In:Santos-Silva, E. N.; Aprile, F. M.; Scudeller, V. V.;Melo, S. (Orgs.),
. INPA, Manaus,AM.Darwich, A.; Kern, J.; Robertson, B.; Souza, E. O. 2001.
Decomposition of and itscontribution to nutrient cycling in a CentralAmazonian floodplain lake.
., 27 (Part 5):2611-2614.Esteves, F. A.; Bozelli, R. L.; Camargo, A. F. M.; Roland,
F.; Thomaz, S. M. 1988. Variação diária (24 horas) detemperatura, O dissolvido, pH e alcalinidade emduas lagoas costeiras do Estado do Rio de Janeiro esuas implicações no metabolismo destesecossistemas. 2:99-127.
Furch, K.; Junk, W. J.; Dieterich, J.; Kochert, N. 1983.
BioTupé: Meio Físico, Diversidade Biológica e
Sociocultural
BioTupé: Meio Físico, Diversidade Biológica e
SocioculturalProcessos de decomposição de
Echinochloa polystachya (H. B. K.) Hitchcock
(Gramineae = Poaceae), capim semi-aquático da
várzea amazônica
.
IX Congresso
Brasileiro de Limnologia
BioTupé: Meio Físico, Diversidade
Biológica e Sociocultural
Echinochloa polystachya
Acta Limnol. Brasil,
Editora
Editora
Verh. Internat. Verein.Limnol
2

Limnologia do Lago Tupé: dinâmica espaço-temporal do oxigênio dissolvido
67
Seasonal variation in the major cation ( Na, K, Mg andCa) content of the Water of Lago Camaleão, inAmazonian floodplain-Lake near Manaus, Brazil.
, 8(1):75-89.Golterman, H. L., 1975. Chemistry. In: Whitton, B. A.
(ed.). . University of California Press,Berkeley and LosAngeles
Golterman, H. L.; Clymo, R. S.; Ohnstad, M. A. 1978.
. 2 ed. IBP Handbook, nº 8. BlackwellScientific Publications, Oxford, Edinburgh, London,Melbourne. 214pp.
Hutchinson, G. E. 1967. . II.Introduction to lake bilogy and the limnoplankton.New York, John Wiley Sons Inc. 1115p.
Hutchinson, G. E. 1975a. . V. I:Geography, physics, and Chemistry. Part 1:Geography and Physics of Lakes. John Wiley Sons.New York. 540pp.
Hutchinson, G. E. 1975b. . V. I:Geography, physics, and Chemistry. Part 2: Chemistryof Lakes. John Wiley Sons. New York. 1015pp.
Junk, W. J. 1973. Investigations on the ecology andproduction-biology of the "floating meadows"(Paspalo-Echinochloetum) on the Middle Amazon. II.The aquatic fauna in the root zone of floatingvegetation. , 4(1):9-102.
Junk, W. J. 1980. Áreas inundáveis - um desafio paraLimnologia. , 10(4):775-795.
Kern, J.; Darwich, A. J. 1997. Nitrogen turnover in thevárzea. In: Junk, W. J. (ed).
. Springer-Verlag, Berlin Heidelberg. Ecological Studies, Vol.126. Cap. 6; p119-135.
Kern, J.; Darwich, A. J. 1999. Studies on the role of N2fixation and denitrification on the Amazonfloodplain. In: Lieberei, R.; Vo, K. Bianchi, H. (eds).
Workshop, December 1998, 275-282pp.Lampert, W.; Sommer, U. 1997.
. Published by OxfordUniversity Press, Inc. Oxford, New York. 382p.
Likens, G. E.; Ragotzkie, R. A. 1965. Vertical water
Amazoniana
River Ecology
,
Methods for physical and chemical analysis of fresh
waters
A treatise on limnology
A treatise on limnology
A treatise on limnology
Amazoniana
ActaAmazonica
The Central Amazon
Floodplain: Ecology of a Pulsing System
Proceedings of the Third SHIFT-Studies on Humam
Impact on Forests and Floodplains in the Tropics -
Limnoecology: the
ecology of lakes and streams
39-80p
nd
&
&
&
&
motionsin a small icecovered lake.., 70:2333-2344.
Likens, G. E.; Ragotzkie, R. A. 1966.Rotary circulation of water in anicecovered lake.
., 16:126-133.Mackereth, F. J. H.; Heron, J.; Talling,
J. F. 1978. Water analysis: somerevised methods for limnologists.Freshwater Biological Association,
nº 8.Windermore Laboratory, Cumbria.121pp.
Margalef, R. 1983. .Ediciones Omega Barcelona.1010pp.
Margalef, R. 1986. . EdicionesOmega. Barcelona. 951pp.
Schmidt, G. W. 1972a. Seasonalchanges in water chemistry of atropical lake (Lago do Castanho,Amazonia, South America).
, 18:613-621.
Schmidt, G. W. 1972b. Chemicalproperties of some waters in thetropical rain-forest region ofCentral Amazonia along the newr o a d M a n a u s - C a r a c a r a í .
, 3(2):199-207.Stumm, W.; Morgan, J. J. 1981.
. 2 ed. John WileySons, New York, 780 pp.
Wetzel, R. G.; Likens, G. E. 2000.. W. B.
Saunders Co. Philadelphia. 357pp.Wetzel, R. G. 1993. .
Serviço de Educação FundaçãoCalouste Gulbenkian Lisboa.919pp.
J. Geophys. Res
Verh. Int. Ver.
Limnol
Scientific Publication
Limnología
,
Ecologia
Verh.
Internat. Verein. Limnol.
Amazoniana
Aquatic chemistry: an introduction
emphasinzing chemical equilibria
in natural waters
Limnolgical Analysis
Limnologia
,
&

Meio FísicoCapítulo 4
Considerações sobre a geoquímica edinâmica sedimentar do Lago Tupé
RESUMO - Os sedimentos refletem a atuação dos parâmetros ambientais por um longoperíodo de tempo, constituindo um registro da dinâmica local. Os sedimentos secaracterizam a partir da sua granulometria, morfometria, análise textural e composiçãomineralógica. Os objetivos principais deste trabalho foram estudar os sedimentos da baciado lago Tupé em seus aspectos granulométrico e textural, identificando as formassedimentares que compõem o leito do lago, identificar os processos erosivos na região docanal de conexão, e apresentar um modelo de fluxo sedimentar para o sistema. Acomposição granulométrica dos sedimentos superficiais da região do lago Tupé variou de96,90% a 99,99% de areia; 0,01% a 2,74% de silte e de 0,0% a 0,38% de argila. Os teores dematéria orgânica oscilaram entre 0,7% e 3,9%. O predomínio das classes de 0,5 a 0,3mm e0,25 a 0,125mm na área do canal, indicam uma influência mais acentuada das correntessobre as partículas mais finas. A estação EC11, localizada no canal de conexão, apresentouum grau de erodibilidade teórico superior a 9.900, um valor muito alto em comparação a
Fábio Marques APRILE
Assad José DARWICH
José Carlos RAPOSO
Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Biologia AquáticaAv. André Araújo 2936, Caixa Postal 470 Manaus, AM, 69060-001
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

região dos igarapés de floresta. No entanto, o alto grau de coesão entre as partículas reduzdrasticamente a probabilidade de erosão na região.
PALAVRAS-CHAVE: dinâmica sedimentar, processos erosivos, granulometria,sedimentologia.
70
Aprile .et al
Introdução
Gênese dos Sedimentos
Os sedimentos são definidos como o materialoriginado da destruição de qualquer tipo de rocha oumaterial de origem biológica, formando substrato nãoconsolidado, geralmente depositado no leito dossistemas hídricos. Segundo Abessa (1996) os sedimentospossuem quatro componentes: fase sólida inorgânica,que inclui fragmento de rochas e grãos mineraisresultantes da erosão terrestre; água intersticial, quepreenche os espaços entre as partículas, podendo chegara 50% do volume, matéria orgânica, que ocupa umpequeno volume, mas é fundamental na regulação dabiodisponibilidade dos elementos, e materiaisantropogênicos, incluindo substâncias químicas.
Do ponto de vista de ciclagem de matéria e fluxode energia, o sedimento é um dos compartimentos maisimportantes dos ecossistemas aquáticos continentais(Esteves, 1998). Sua formação e comportamento sãoresultados integrados de fatores físicos, químicos ebiológicos que ocorrem no lago, os quais por sua vez, sãoparticularmente influenciados pela origem, forma eorientação da bacia de drenagem, pelas condiçõesclimáticas da região, e sua cobertura vegetalpredominante (Coutinho Barbosa, 1986).
Os sedimentos refletem a atuação dosparâmetros ambientais por um longo período de tempo,constituindo um registro da dinâmica local. Nessesentido, é possível revelar informações sobre os animaise vegetais que existiram em uma determinada área,através de fósseis e componentes químicos, identificareventos geológicos pretéritos e, avaliar as várias formase intensidades dos impactos ocorridos em umdeterminado ecossistema, desde que não haja um hiatode tempo geológico significativo. Dessa forma, ossedimentos passaram a ser objeto de estudo daestratigrafia, palinologia, geoquímica e maisrecentemente da paleolimnologia e paleosedimentolo-
&
gia.Os sedimentos podem ser classifi-
cados quanto à composição químicaem sedimentos orgânicos e inorgâni-cos. Os sedimentos orgânicos estãonormalmente localizados nas camadasmais superficiais do leito, sãosedimentos recentes, com grandequantidade de componentes animais evegetais em processo de decomposi-ção. Já os sedimentos inorgânicos sãocaracterizados pelo predomínio desílica, quartzito, argila e elementosmetálicos. Com relação a origem, ossedimentos podem ser autóctones ealóctones. Autóctone, palavra origina-da do grego ( ) que significa por sipróprio ou de si mesmo, é o sedimentoque se formou por processos químicosou biológicos no próprio lugar onde seencontra, não sofrendo nenhum tipode transporte. Alóctone, do grego( ), significa que ou quem não éoriginário do local onde existe ouhabita. Os depósitos alóctones sãoconstituídos por materiais de outrasáreas, exógenos.
Os sedimentos autóctones são umtestemunho das características de umaregião, enquanto que o estudo dossedimentos alóctones, seja através daanálise granulométrica, seja noaspecto textural, permite compreen-der os processos e identificar os fluxosda dinâmica sedimentar.
autós
állos

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
71
Análise Granulométrica
Os sedimentos se caracterizam apartir da sua granulometria, morfome-tria, análise textural e composiçãomineralógica. Dentre as propriedadesfísicas dos sedimentos, o tamanho,forma e arranjo espacial dos compo-nentes mineralógicos são as maisestudadas.
O tamanho das partículas de rochassedimentares detríticas, como as queocorrem na região do lago Tupé,constitui uma propriedade texturalfundamental muito utilizada na classi-ficação dos sedimentos clásticos. ParaAprile (2001) a análise granulométricaconstituí a base para uma descriçãoprecisa dos sedimentos. Através deseus resultados, pode-se obter infor-mações bastante precisas sobre osprocessos físicos atuantes durante asedimentação, além de permitir traçarum fluxo da carga sedimentar naregião pesquisada.
A caracterização granulométrica éfeita pela determinação dasporcentagens em massa dos grãos, emcada classe granulométrica (frações).Vários métodos têm sido utilizadospara relacionar as classes granulomé-tricas com o ambiente deposicional.Udden (1898) foi um dos primeiros adesenvolver uma escala granulométri-ca para sedimentos. Em meados dadécada de 50, Shepard (1954) eShepard Moore (1954) utilizaram umdiagrama triangular para mostrarmisturas entre as frações. Posterior-mente, Folk Ward (1957) e MasonFolk (1958) utilizaram-se de parâme-tros estatísticos para estudar adinâmica sedimentar.
Não existe uma escala universal-mente aceita para a interpretação dasanálises granulométricas, sendo que
&
& &
cada escola, sejaAmericana, Européia ouAsiática, adotasua escala-padrão. Os limites estabelecidos para asvárias classes granulométricas são mais ou menosarbitrários, mas, segundo Wentworth (1933) as principaisclasses granulométricas estariam intimamentecorrelacionadas aos modos fundamentais de transportepor água corrente ou com os diferentes modos dedesintegração da rocha-matriz. Segundo Suguio (1994) aescala de Wentworth (1922) tem sido adotada semmuitas ressalvas pelos sedimentólogos brasileiros. Asescalas classificam as frações sedimentares segundo otamanho da partícula ou intervalo granulométrico (Tab.1).
Tamanho da partícula
(mm)
Descrição
> 256 matacão
256 – 64 calhau
64 – 4,0 seixo
4,0 – 2,0 grânulo
2,0 – 1,0 areia muito grossa
1,0 – 0,50 areia grossa
0,50 – 0,25 areia média
0,25 – 0,125 areia fina
0,125 - 0,062 areia muito fina
0,062 – 0,031 silte grosso
0,031 – 0,016 silte médio
0,016 – 0,008 silte fino
0,008 – 0,004 silte muito fino
< 0,004 argila
Tabela 1. Classificação dos sedimentos baseada nos trabalhosde Wentworth (1922) segundo a análise granulométrica.
Outras classificações sugeridas a partir de estudosdos diâmetros-padrão dos grãos bastante conhecidassão: Krumbein (1936); Krumbein Pettijohn (1938);Krumbein Rasmussen (1941); Young (1966) e Müller(1967).
De acordo com Furtado (1978) a análisegranulométrica reflete diretamente o comportamento
&&

72
hidrodinâmico do ambiente de estudo. Bagnold (1941)utilizou o comportamento hidrodinâmico na definição deareia, concluindo que a areia teria a capacidade deacumulação espontânea, que consiste na utilização daenergia do meio de transporte para reunir os grãosespalhados, deixando a superfície isenta de partículas.Para Maglioca Kutner (1964, 1965) esse comportamen-to hidrodinâmico interfere na variação da concentraçãode matéria orgânica no ambiente. A matéria orgânicacimentante está normalmente associada a fraçõesinferiores a 0,062mm de diâmetro e, pode ser separadapara análise com uso de floculantes comohexametafosfato de sódio, ou líquidos pesados, comobromofórmio (CHBr ) e tetrabromometano (C H Br ).
Esta pesquisa visou estudar o comportamentogeoquímico e sedimentológico dos sedimentossuperficiais da bacia do lago Tupé, levando emconsideração a granulometria, análise textural,processos físicos e químicos atuantes (intemperismo eerosão), gerando um modelo dos processos erosivos e dadinâmica sedimentar para o sistema.
Esta pesquisa foi realizada durante o ciclo hidrológico2003/2004 na área da bacia do lago Tupé,compreendendo doze pontos de amostragem desedimentos no lago, incluindo um ponto no canal deconexão e um no rio Negro. Foram determinadas acomposição granulométrica e análise textural dossedimentos superficiais através das técnicas depeneiramento e separação fracionada, utilizandopeneiras da série TYLER abertura de malhas 2; 1; 0,5;0,3; 0,25; 0,125; 0,063; 0,053; 0,032 e 0,016mm,seguindo a classificação proposta por Shepard (1954) eSuguio (1973). Para determinar a percentagem dasfrações de silte grosso, médio e silte fino associado aargila (< 0,016 ), foi utilizado o procedimento depeneiramento úmido, com lavagem através de águacorrente, desagregando as partículas de menor diâmetroe, determinando a massa seca do material amostradoantes e depois do procedimento metodológico. Sempreque necessário utilizou-se hexametafosfato de sódiocomo agente dispersante. Uma distribuição da classetextural dos sedimentos da bacia do lago foi elaborada
&
3 2 42
Materiais e Métodos
�
através do Diagrama Triangular deShepard (1954). Um estudo daerodibilidade dos solos marginais e ummodelo erosivo para o canal deconexão foi desenvolvido. Para tanto,levou-se em consideração os resulta-dos da composição granulométrica,análise textural, informações sobre aclassificação dos solos superficiais, eestudos sobre o trabalho erosivo e ofluxo de energia envolvido notransporte de material em suspensãona coluna de água. Também foielaborado um modelo da dinâmicasedimentar para a bacia, evidenciandoas influências lacustre e fluvial notransporte de sedimentos.
Como já foi mencionado, existeuma série de métodos de classificaçãogranulométrica. Dentre os maisutilizados está o diagrama triangularde distribuição de Shepard (1954). AFigura 1 apresenta os resultados daanálise granulométrica realizada parao lago Tupé e rio Negro a partir dessemétodo de classificação para o ciclohidrológico 2003/2004.
Através dos resultados foi possívelidentificar a presença de três gruposdistintos de classes sedimentares. Oprimeiro grupo (G1) compreende amaioria dos pontos amostrados, o queem termos de área significa o tiposedimentar predominante no lago.Nesse grupo foram incluídos ossedimentos classificados como areiafina silte argilosa. No segundo grupo(G2) estão as estações situadas nosigarapés de floresta da Helena, das
Resultados eDiscussão
Análise Granulométrica e Textural
Aprile .et al

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
73
Pedras e da Cachoeira, classificadoscomo sedimentos de areia fina siltosa.No terceiro grupo (G3) foram incluídosos pontos do canal de conexão (EC11) eo ponto amostrado no rio Negro, ambosclassificados como sedimentosarenosos. A composição granulométri-ca dos sedimentos superficiais daregião do lago Tupé variou de 96,90% a99,99% de areia; 0,01% a 2,74% de siltee 0,0% a 0,38% de argila. Os teores dematéria orgânica oscilaram entre 0,7%e 3,9%.
A aplicação do método de isovalo-res contínuos permitiu evidenciar trêsestações com predomínio da areiamuito fina associada as frações siltosase argilosa sobre as demais frações, é ocaso das estações ET1, EI6 e ET10. Naestação central (ET10) a concentraçãode partículas mais finas é explicadapela geomorfologia local. O trecho demaior declividade termina próximo aocruzamento dos braços maior e menordo lago, onde se localiza a ET10, dessemodo, com a redução brusca davelocidade de transporte o materialem suspensão sedimenta com maisfacilidade. Os sedimentos do canal deconexão (EC11) e do rio Negro (EN12),como era esperado, apresentaram asmaiores concentrações de areia médiae grossa. O predomínio das classes de0,5 a 0,3mm e 0,25 a 0,125mm na áreado canal, indicam uma influência maisacentuada das correntes sobre aspartículas mais finas, que entram oupermanecem em suspensão, sendocarreadas pelo fluxo contínuo daságuas. A ausência da deposiçãosedimentar nessas estações (EC11 eEN12) reflete a importância dadinâmica fluvial para a região. Outrofato observado é uma faixa crescenteda concentração de argila e argilacoloidal, que vai da entrada do canal
de conexão na direção dos igarapés de floresta, seja emdireção das estações ET7 e EI8 seja para ET5 e EI6 (Fig.2). Essa faixa, bastante nítida, revela umaheterogeneidade dos sedimentos superficiais em relaçãoaos teores de matéria orgânica. Os colóides, sãopartículas com características bem peculiares, quepertencem a um estado da matéria muito estudado emsedimentologia. Trata-se de um complexo argila -composto orgânico que é pesado demais para ter ocomportamento de íon, e fino e leve demais pararespeitar as fórmulas físicas de tensão interna. Em lugarde decantar, este tipo de partícula realiza no fluído ummovimento aparentemente aleatório, denominadomovimento browniano. Acredita-se que no lago Tupé amatéria orgânica associada a argila é provenientediretamente da decomposição e lixiviação daserapilheira no entorno do lago, enquanto que a argila éliberada de determinados solos marginais a partir dosprocessos de intemperismo e erosão.
O tipo de grão presente nos sistemas fluviais elacustres é muito variado, apresentando alterações notamanho, cor e textura, de acordo com as condiçõesgeológicas e climatológicas de cada região. Pettijohn(1940) estudando o tamanho das partículas de materialclástico em mais de 1.000 amostras, observou umadeficiência nas classes de 2mm a 4mm (areia muitogrossa a grânulos) e de 1mm a 2mm (areia grossa a muitogrossa). Muitos autores contudo, não acreditam ser este
Figura 1. Classificação dos sedimentos superficiais do lago Tupée rio Negro para o ciclo hidrológico 2003/2004 com base notrabalho de Shepard (1954).
100%
100%
100%
0%
0%
0%
Silte
Areia
Argila
G1
ET1ET5EI6EI7EI8ET10
EI2EI3Ei4
EC11EN12
G2
G3

74
um fato generalizado. Russell (1968) sugeriu que a causada aparente escassez dos grânulos em depósitos fluviaise lacustres dinâmicos, estaria relacionada ao fato de queestes grãos são transportados mais rapidamente que asareias às quais se acham associados. Então seriam, emconseqüência disso, eliminados dos lagos e rios eacumulados em ambientes marginais (praias) ouestuários. Em estudos sobre os processos sedimentaresna planície costeira do baixo rio Doce, no Estado doEspírito Santo, Aprile . (1998, 2001) encontrarampercentuais de areia grossa e muito grossa na foz do rioDoce que variaram 67,9% (estiagem) a 95,5% (chuvas).Estudos sobre a dinâmica sedimentar local revelaram, noentanto, que esse material não era de origem fluvial,mas sim decorrente da influência marinha na região. Osautores observaram ainda que em virtude da variação dacompetência do rio, o transporte fluvial de grãos élimitado a partículas inferiores a 0,5mm.
A questão do grau de modificação da distribuiçãogranulométrica que pode ser introduzido pelos processos
et al
de transporte ainda não esta suficien-temente entendida. Em geral, oscascalhos e grânulos carreados pelosrios e lagos parecem diminuir detamanho em direção a jusante, emdecorrência da carga de sedimenta-ção. Contudo, quando se estudaregiões de alta energia, como a foz deum rio ou de um grande lago, pode-seobservar exatamente o processoinverso. Essa questão da distribuiçãogranulométrica está associada aquantidade de energia limite necessá-ria para o transporte e/ou precipitaçãodo grão (competência do sistemahídrico), e será abordada maisadiante.
O termo intemperismo é definidocomo o conjunto de processos oumodificações devidos à ação deagentes atmosféricos e biológicos quegeram a destruição física e a decompo-sição química dos minerais e rochas. Osfatores que controlam a ação dointemperismo são o grau de resistênciada rocha mãe a ação do tempo; oclima, que pode apresentar umavariação temporal (sazonalidade)através das chuvas e da temperatura; orelevo, que pode influir no regime deinfiltração e drenagem das águaspluviais e, os ecossistemas terrestres eaquáticos, cuja grande contribuição éo fornecimento de compostosorgânicos para os processos de decom-posição.
Os processos intempéricos atuamatravés de mecanismos modificadoresdas propriedades físicas dos minerais erochas, além de suas característicasquímicas. Em função dos mecanismospredominantes de atuação, sãoclassificados em intemperismo físico
Intemperismo
Figura 2: Isovalores dos percentuais de argila e argila coloidalpresentes nos sedimentos do lago Tupé para ciclo hidrológico2003/2004.
Aprile .et al
Argila+Coloides (%)
0.0 to 0.80.8 to 1.51.5 to 1.91.9 to 3.1
Altasedimentação
ET1
EI2EI3
EI4
N
ET5
EI6
-60.24-60.25
Long. W
-3.05
-3.04
-3.03
La
t.S
-60.26
altasedimentação
baixasedimentação
ET10
ET7
EI8
EC11EN12

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
75
ou mecânico, químico ou biológico,este último ocorre quando a ação físicaou bioquímica de organismos vivos ouda matéria orgânica proveniente desua decomposição participa doprocesso (Suguio, 1994; Toledo .,2000). No ciclo de formação das rochassedimentares o intemperismo tem umpapel fundamental ao lado dasedimentação.
Na região amazônica a ação dointemperismo químico é predominan-temente mais acentuada, devido atemperatura e níveis de chuvas maiselevados. Em regiões como o lagoTupé, onde as rochas sedimentaresentram em contato com as águas derios e lagos de água preta, ocorrereações com os minerais formadoresdas rochas. Essas reações sãoaceleradas pelo fato das águas pretasserem ácidas (pH ~= 4,5) devido adissolução de CO que forma os ácidoscarbônicos, e a presença constante desubstâncias húmicas (ácidos húmicos efúlvicos) oriundas da decomposiçãodos compostos orgânicos. Uma análisequímica das águas dos lagos e rios podedetectar a presença de carbonatos nassuas várias formas, oriundos das rochassedimentares.
Reações de hidratação/hidrólise eoxiredução permitem a dissociaçãodos minerais da rocha mãe. O volumede material dissolvido depende daquantidade de água envolvida noprocesso. A velocidade e sentido dasreações de oxiredução dependem daconcentração de oxigênio dissolvido naágua, e do tamanho da camadaanaeróbica. Tratam-se de reaçõesenvolvendo os íons H+ e OH-, que dessaforma, acabam por modificar o pH doambiente.
Outra reação bastante comumdurante processo de intemperismo é a
et al
2
complexação, que ocorre quando um elemento metálicona sua forma iônica se associa a um composto orgânico.Esta reação ocorre com grande freqüência em áreas demineração, como é o caso do rio Madeira nos trechos altoe médio, onde o mercúrio na sua forma orgânica(metilmercúrio) se associa ao material em suspensão,precipitando na forma de complexo (quelação). Na baciado lago Tupé acredita-se não existir esse tipo de reaçãodurante o processo de intemperismo químico, por setratar de uma região relativamente bem preservada.
Os solos da Amazônia são predominantementeclassificados como oxissolo, um solo típico de regiõestropicais úmidas, altamente intemperizado, ácido epouco fértil, muito similar ao latossolo. Essecompartimento, junto com as rochas, garante através deprocessos químicos o fluxo de óxidos para as águassuperficiais e subterrâneas de toda a região (EMBRAPA,1999 Toledo ., 2000).
Segundo Ruxton (1968) a taxa relativa de mobilidadedos óxidos dos principais elementos químicos das rochas,oriundos da dissociação sedimentar, decresce,geralmente, na seguinte ordem: Ca > Na > Mg > K > Si> Fe > Al . Para o autor, as rochas submetidas aointemperismo químico tendem a perder muito maisóxidos de cálcio, sódio, magnésio e potássio do que ferroe alumínio, o que explicaria o "enriquecimento" dasrochas por estes últimos. Estudos realizados por Darwich(1995) no lago Camaleão, um lago de várzea amazônico,e no rio Solimões/Amazonas, revelaram um padrão dedominância iônica HCO > Ca > Mg > Na > Cl > Kdurante quase todo o ciclo hidrológico. Na região do lagoTupé, Darwich (2003, dados não publicados) encontrou aseguinte relação iônica na coluna de água K > Na > Ca >Mg . A concentração iônica na coluna de água de umdeterminado ecossistema, está diretamente ligada aosprocessos de degradação das rochas e fragmentação dossolos marginais, liberando compostos minerais. Dessemodo, pode-se dizer que a taxa crescente de íons nacoluna de água e inversamente proporcional a da rochamatriz.
A erosão é o processo de desprendimento e arrasteacelerado das partículas do solo causado pela ação daágua e/ou vento. A água é o mais importante agente de
apud et al
2+ +
2+ +
2+ 2+ + +
+ + 2+
2+
2+ + 2+
- -3
Processos Erosivos

76
erosão; chuvas, córregos, rios, todos carregam solo, asondas erosionam as costas dos mares e lagos, de fato,onde há água em movimento, ela está erodindo os seuslimites (Bertoni Lombardi Neto, 1999). A constanteredução da produtividade dos solos tem sido atribuídaprincipalmente à erosão hídrica e ao manejo inadequadodo solo. Outro aspecto de grande relevância é que oaporte de sedimentos oriundos de áreas que sofremerosão promove o assoreamento de rios e lagos,comprometendo a qualidade da água e alterando a vidaaquática, principalmente pela eutrofização (Martins
., 2003).Além do impacto direto da perda de solos para os rios
e lagos em decorrência da erosão, deve-se lembrar quejunto com os solos são carreados compostos orgânicos einorgânicos, que enriquecem o sistema hídrico. Os solosdas regiões tropicais, com as intensas chuvas, estão maissusceptíveis a erosão. A cobertura vegetal é de grandeimportância para a redução das perdas de nutrientes dossolos. Alguns solos erosionam mais que outros, mesmoque a intensidade de chuvas, declividade, coberturavegetal e práticas de manejo sejam as mesmas.
Durante muitas décadas os sedimentólogos têmtentado relacionar a quantidade de erosão medida nocampo com as várias características físicas do solo quepodem ser determinadas em laboratório. Bouyoucos(1935), através de vários experimentos, relacionou aerodibilidade do solo com a sua granulometria, sendodefinida como:
&
etal
direção ao canal (Fig. 5). O transportede partículas através de processoscomo escorregamento ou avalanchacoesiva, característicos de regiões deduna, com fluxo granular clássico(língua de areia) ou ainda cicatrizes deescorregamento produzidas poresforço distensivo, como relataGiannini Riccomini (2000), não sãoobservados na região. No trecho docanal o alto grau de coesão entre aspartículas reduz a ação dos processoserosivos drasticamente. Essa coesãoaparente da areia é controlada peloteor de umidade natural do solo e, éem muito responsável pela retençãodos solos marginais.
Outro fator que os autoresacreditam contribuir para os baixosíndices de erosão no trecho decomunicação rio/lago, é a baixaenergia despendida pelo rio Negrodurante os períodos de enchente evazante. Esse fato pode ser comprova-do observando-se a variação diária donível das águas descrito em AprileDarwich (2005, neste volume). Aságuas do rio Negro tendem a entrar nolago paulatinamente, interferindo omínimo possível no deslocamento daspartículas arenosas presentes àsmargens do canal de conexão.
A erosão em determinada áreafluvial ou lacustre, pode ocorrer emeventos isolados com distribuiçãotemporal discreta. Tais aspectos,associados as condições do meio, emparticular a sazonalidade, dificultam ainterpretação e/ou avaliação do graude alteração das condições naturais.Na região amazônica, em decorrênciado pulso de inundação, que altera ascondições físicas do sistema hídricoseqüencialmente, torna-se difícil acurto prazo detectar o resultado daação erosiva.
&
&
Aplicando a equação ( ) para os pontos amostrados,observa-se que o grau de erodibilidade obtido a partir dasproporções de areia, silte e argila variou de 260 a valoressuperiores a 9.900, este último valor encontrado no canalde conexão rio/lago. Isso mostra que a região do canal deconexão apresenta um grau de erodibilidade teóricomuito alto em comparação a região dos igarapés defloresta. De certo modo, é de se esperar que uma áreadestituída de vegetação e formada por um solotipicamente arenoso, como mostra as Figuras 3 e 4, soframais intensamente a ação do intemperismo e erosão. Noentanto, não é isso que ocorre. A região onde se encontrao canal de comunicação com o rio Negro apresenta umaspecto xeromórfico, com dunas que se inclinam em
i
Aprile .et al
E =% areia + % silte
% argila(i)

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
77
Wischmeier Smith (1958) formu-laram uma equação para determinar aperda de solos pela erosão hídrica,considerando no cálculo da energiacinética total ( ) de uma chuvaerosiva a intensidade da chuva ( ). Aequação pode ser definida como:
&
EcI
conceito de trabalho realizado. Em um sistema nãoconservativo, baseado nos princípios da mecânicaclássica, a energia mecânica ( ) despendida para arealização de um trabalho útil ( ) não é integralmenteconvertida, havendo uma dissipação (
ção ideal, menor será o valor de ,tendendo a zero em uma situação de máximaconservação de energia. Dessa forma tem-se:
EcWs
ωω
). Quanto maisprσximo de uma situa
Figura 3. Detalhe do canal de conexão entre o lago Tupé e o rioNegro.
Figura 4. Canal de conexão com dunas marginais.
A água da chuva exerce sua açãoerosiva sobre o solo mediante oimpacto da gota de chuva, a qual caicom velocidade e energia ( )variável, segundo o seu diâmetro emediante a ação de escorrimento.
O modelo apresentado porWischmeier Smith (1958) é bastanteeficiente em situações onde o regimedas chuvas é bem definido, comoocorre na região sudeste do Brasil, porexemplo. No entanto, na Amazônia,onde as chuvas são bem maisdistribuídas ao longo do ano, umacaracterística de floresta tropicalúmida, e o ciclo hidrológico criacondições bastante dinâmicas nosecossistemas fluviais e lacustres, énecessário levar em conta que osistema não é conservativo.
Baseado nas informações obtidaspela análise granulométrica, foiproposto um modelo de avaliação daperda de solo por erosão hídrica para ocanal de conexão entre o lago Tupé e orio Negro. Foram levados emconsideração o pulso de inundaçãopara a região e o fato do lago ser umsistema lacustre pouco dinâmico.
Para determinar o grau da açãoerosiva em uma determinada área,deve-se levar em consideração o
Ec
&
Modelo Erosivo para o Canal deConexão
Ec = Ws + klog I (ii)
Ec = Ws + Ws = Ec -��� � (1)

78
para uma situação ideal �
de trabalho erosivo ( ), com suasrespectivas perdas. Por fim, deve-seconsiderar o sistema constituído deuma massa ( ) imprimindo-lhe umaforça ( ). Nesse caso, a energiamecânica total ( ) despendida nosistema, é dada pelo somatório dasenergias cinética ( ) e potencial ( ).Tem-se assim:
Ws
MF
ET
Ec Ep
Figura 5. Paisagem que compõe o canal de conexão. Detalhepara a ausência de vegetação do tipo rasteira (gramíneas).
e ético para a realização dotrabalho .
Dividindo a equação 1 por tem-se:
onde = gasto energωWs
Ec
Defini-se assim que é a taxa de eficiência deconversão de energia. A ação erosiva é função da energiacinética pelo tempo, podendo ser expressa a partir dajunção das equações 1 e 2 e sua respectiva derivada.Sendo assim tem-se:
�
Nesta proposição, a ação erosiva em função do tempoé determinada pela entrada de energia ( ) e pela saídaEc
A equação 4 trata da energiadisponível à realização de um trabalhoerosivo ( ), que uma massa de água( ) apresenta quando animada de umavelocidade ( ) por ação da aceleraçãoda gravidade ( ). Essa massa de águapode ser definida como sendoproveniente da precipitação (chuvas)ou escoamento superficial (enxurra-da), cuja propriedade é a desagrega-ção dos solos marginais e conseqüentetransporte de partículas.Associando asequações 2 e 4 para uma energia total,defini-se que o trabalho erosivo é dadopor:
WM
vg
O escorrimento da água nasuperfície do solo é o maior agente detransporte das partículas. A quantida-de de força gerada pela enxurrada érelacionada com a concentração evelocidade com que ela se move emuma declividade. A energia daenxurrada é uma função da massa e davelocidade de escorrimento da água.
A dinâmica energética na produçãode erosão hídrica está associada àdinâmica do ciclo hidrológico,parcialmente convertida em trabalhode desagregação e de transporte departículas de solo. Segundo D'Agostini
Aprile .et al
Ec - Wsk
Limk ���
WsEc
EcEc Ec
�=
Ec�
Ec�- �
WsEc
= 1 -
�WsEc
= para
Ec - �Ec
= � (1) = (2)da Eq.WsEc
Ec
t= (3)
�Ws
Ec�
dEcdt
+��
Ec�
dEcdt
+ M.g.h (4)E = E + E E =r c p r�M-v2
2
+ M.g.h �W = .E W = (S R S �M-v
2
2
+ g.h) (5)W = .M (S M-v
2
2

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
79
(1999) o estado energético seja para aágua da chuva, para a água parada ouem escoamento sobre a superfície dosolo, decorre da manifestação deforças derivadas de relações entremassa.
O ciclo sedimentar clássico constade fases de intemperismo e erosão,transporte, sedimentação (deposi-ção), litificação e metamorfismo (Fig.6). Toda a bacia hidrográfica éresponsável pelo fornecimento detríti-co aos cursos de água que, em conjun-to, torna-se o fenômeno natural demaior ocorrência na esculturação darede de canais e das paisagensencontradas na superfície terrestre(Christofoletti, 1981).
A dinâmica sedimentar estaparticularmente associada às etapasde transporte e sedimentação. Nasáreas continentais, em especial naregião amazônica, estudos dessanatureza ainda são escassos.
A carga de material particulado éformada por partículas de granulome-tria maior, transportadas através dosmecanismos de saltação, arrasto ourastejamento e deslizamento ourolamento. Cada mecanismo apresen-ta um tipo de ação associado a umtamanho de partícula e trajetóriaespecíficos. Em decorrência do volumee da densidade dos grãos, a velocidadeda carga do leito é muito mais lentaque a do fluxo, pois as partículaspodem se deslocar de modointermitente. Já a carga em suspen-são, ocorre quando partículas degranulometria reduzida, como silte eargila, se mantém suspensas pelo fluxoturbulento.
Quanto maior a granulometria de
Dinâmica Sedimentar
um leito sedimentar inconsolidado, maior deverá ser avelocidade mínima necessária para dar início aomovimento dos grãos (valor crítico). Assim sendo, oscascalhos carreados, de forma geral, diminuem detamanho em direção à jusante. Segundo Suguio (1994)isso se deve não apenas a abrasão durante o transportecom conseqüente aumento dos g raus dearredondamento e esfericidade, mas também, pelaredução da competência do sistema hídrico emdecorrência da redução da declividade. O diagramaapresentado na Figura 7 mostra a velocidade mínimanecessária para que ocorra o transporte do materialsedimentar. Nota-se que existe um ponto crítico entre otransporte e a sedimentação do material erodido dossolos marginais. Nesse ponto, partículas de maiordiâmetro podem ora ser depositadas ora ser carreadascom uma pequena variação da velocidade de corrente.
Para que as partículas em movimento sedimentem ou
Figura 6. Ciclo sedimentar simplificado.
(Sedimentar; ígnea; metamórfica)
ROCHAS MATRIZES
INTEMPERISMO
EROSÃO
TRANSPORTE
SEDIMENTAÇÃO
LITIFICAÇÃO E METAMORFISMO

80
vice-versa, deve haver uma relação direta entre avelocidade crítica e o diâmetro das mesmas. Avelocidade crítica necessária para que a partícula deixeo estado de repouso e apresente uma aceleração, éfunção da granulometria do grão. Partículas muito finasnecessitam de uma energia cinética inicial alta paradeixar esse estado de repouso. É o caso dos sedimentosargilosos e silte-argilosos, que apresentam umasuperfície específica muito grande, necessitando, dessaforma, de uma energia cinética bastante elevada paraserem erodidos e por conseguinte transportados. Emoutras palavras, da mesma forma como há umavelocidade mínima necessária para "romper" o estadoinercial de repouso, também há um tamanho mínimonecessário para que ocorra a inclusão da partícula nofluxo turbulento.
No lago Tupé a maior velocidade de corrente éencontrada durante as fases de enchente e vazante.Nesses períodos predomina o processo de transportesedimentar. Já durante os períodos de cheia e seca, porsua vez, há o predomínio dos processos desedimentação. Além disso, dentro das condiçõesmorfométricas do lago, apresentadas por Aprile &Darwich (2005, neste volume), existem trechos de maior
ou menor taxa de sedimentação,dependendo da competência e dadeclividade do lago, como mencionadoanteriormente. A deposição domaterial em suspensão no lago Tupéocorre preferencialmente no eixovertical, promovendo o acúmulogradual e homogêneo dos grãos degranulometria semelhante, e oaumento da espessura da camada desedimentos superficiais do leito. Adeposição por arrasto e rolamentodeposita os sedimentos no sentido datensão tangencial, de forma paralelaao leito sedimentar, permitindo adistribuição das partículas numa formaplana. Os trechos de maior deposiçãosedimentar observados no lago foramprimeiramente no igarapé daCachoeira (EI4) e na estação ET7, e emum segundo momento à jusante daestação ET5 e à montante da estaçãoET10, ambas em trechos de fortedeclividade.
Em um fluxo de baixa viscosidade,como ocorre no curso inferior dosigarapés de floresta, que secomunicam com o lago, não há ou háuma ínfima ação da turbulência.Nesses trechos, há uma tendência daspartículas muito finas a sedimentaremde forma homogênea, formando umaespessa camada de sedimentosrecentes, que mesmo havendo umaruptura do momento de repouso,através de uma força de soerguimento,se esta não se mantiver, o grão tenderáa sedimentar novamente.
Estudos desenvolvidos por Kuenen(1965) e posteriormente por Postma(1967) revelaram que os depósitossedimentares constituídos por grãosmenores que 0,1 mm apresentamgrande força de coesão. Essa forçamanifesta-se em decorrência dagrande diferença entre a velocidade
Aprile .et al
1000
EROSÃO
TRANSPORTE
SEDIMENTAÇÃO
500300200
50
100
3020
10
5321
0,0
01
0,0
02
0,0
03
0,0
05
0,0
1
0,0
20,0
30,0
5
0,1
0,2
0,3
0,5 1 2 3 5
10 20
30
50
100
0,50,30,20,1
log.ve
loci
dade
média
(cm
.s)
-1
Figura 7 Relação velocidade granulometria no ciclosedimentar (Fonte: Suguio, 1994).
. versus

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
81
da corrente necessária para o trans-porte e a velocidade necessária pararecolocá-las em suspensão depois desedimentadas.
Um modelo de dinâmica sedimen-tar para o lago é apresentado na Figura8. Observa-se a partir dos resultadosda análise granulométrica e da taxa deerodibilidade, que a sedimentação deorigem lacustre é predominantementeformada de silte, argila e argila coloi-dal, enquanto que a sedimentaçãofluvial é arenosa. A concentração dematerial em suspensão no lago Tupé éde aproximadamente 10 mg.L , sendoque em alguns pontos esse valor éainda menor, como é o caso do igarapéTerra Preta (EI8) cuja concentraçãochega ao máximo de 4 mg.L .Considerando que a carga de materialem suspensão transportado pelo rioNegro é muito baixa, e que os igarapésde floresta são basicamente de águasclaras, concluí-se que os sedimentosfinos encontrados no leito do lago Tupésão decorrentes, principalmente, daerosão/lixiviação dos solos silte-argilosos e argilo-siltosos da bacia dolago.
O comportamento dinâmico dogrão é uma resposta direta às forçasnele atuantes, e reflete suas caracte-rísticas individuais como forma,tamanho, densidade e rugosidadesuperficial. Nos fluxos de baixaviscosidade, a corrente pode separaros grãos menos densos dos maisdensos. Nos corpos de água preta, ofluxo de baixa viscosidade épredominante, como resultado dabaixa concentração de material emsuspensão existente na coluna deágua. O resultado desse comportamen-
-1
-1
to é uma seleção, de caráter granulométrico emorfométrico, ao longo do leito dos sistemas lacustres efluviais, a exemplo do lago Tupé e do rio Negro.
Ainda são poucos ou quase nenhum os estudosacerca dos efeitos da diminuição granulométrica sobreos parâmetros da distribuição de tamanhos dos grãos, e acorrelação destes com os tipos de fluxo. Estudospreliminares associando o transporte de partículas, seusdiversos tamanhos e a carga de sedimentação, estãosendo desenvolvidos no rio Madeira no trechocompreendido no Estado de Rondônia, pelo autores. Oestudo leva em consideração, entre outros fatores, acarga de material particulado em suspensão na coluna deágua, que pode ser retida a partir de um processo defiltração. Acredita-se que o processo de sedimentação etransporte do material particulado dependa não apenasdo tamanho e natureza do material, mas também, daproporção deste material, do grau de irregularidade doleito do sistema hídrico, e do tempo e distânciaenvolvidos na ação abrasiva.
Figura 8. Modelo de dinâmica sedimentar para o lago Tupé.

82
Echinochloa polystachya (H. B. K.)Hitchcock (Gramineae = Poaceae),capim semi-aquático da várzeaamazônica ,
.
Fundamentos deLimnologia
Journal ofSed. Petrol.
Contribuição aoestudo da sedimentação atual nocanal de São Sebastião, Estado deSão Paulo
.
Decifrando a Terra..
J. Sediment. Petrol
. Sediment.Petrol.
. Tese de DoutoradoInstituto Nacional de Pesquisas daAmazônia/ Universidade doAmazonas (INPA/UA), Manaus (AM)327pp.
Esteves, F. A. 1998.. 2 ed., Rio
de Janeiro 602 p.Folk, R. L.; Ward, W. C. 1957. Brazos
river bar a study in the significanceof grain size parameters.
, 27:3-27.Furtado, V. V. 1978.
. Tese de Doutorado,Instituto Oceanográfico, USP, SãoPaulo, v. 2 102pp.
Giannini, P. C. F.; Riccomini, C. 2000.Sedimentos e processos sedimenta-res. In: Teixeira, W.; Toledo, M. C.M.; Fairchild, T. R. Taioli, F.(eds.). SãoPaulo, Oficina de Textos p. 167-190.
Krumbein, W. C. 1936. The applicationof logarithmic moments to sizef r equency d i s t r i bu t i on o fsediments. .,6:35-47.
Krumbein, W. C.; Rasmussen, W. C.1941. The probable error ofsampling beach sand for heavymineral analysis. J
, 11:10-20.Kuenen, P. H. 1965. Experiments in
Interciência,. p
Krumbein, W. C.; Pettijohn, F. J. 1938.
. Appleton Century Crofts Inc.,New York, 549p.
&
Manual of sedimentary petrogra-phy
Bibliografia Citada
Abessa, D. M. S. 1996.
. Dissertação deMestrado, Instituto Oceanográfico, USP, São Paulo97pp.
Aprile, F. M. 2001.
. Relatório de Pós-Doutorado, Instituto Oceanográfico, USP, São Paulo129pp.
Aprile, F. M.; Shimizu, G. Y.; Lorandi, R.; Silva, L.A. 1998.Lagoas Costeiras do Espírito Santo, Brasil:Caracterização química do sedimento e água destesecossistemas. In: 8 ,São Carlos, SP, 1996. São Carlos, 3:1361-1371.
Aprile, F. M.; Lorandi, R.; Bianchini Jr., I.; Shimizu, G. Y.2001. Tipologia dos ecossistemas lacustres costeirosdo Estado do Espírito Santo, Brasil. , 15(1):17-21.
Aprile, F. M., Darwich, A. J. 2005. Modelosgeomorfológicos para o lago Tupé In: Santos-Silva, E.N.; Aprile, F. M.; Scudeller, V. V.; Melo, S. (Orgs.),
.INPA, Manaus,AM.
Bagnold, R. A. 1941.. Methuen, 265p .
Bertoni, J.; Lombardi Neto, F. 1999.. São Paulo, 4 ed. 355p .
Bouyoucos, G. W. 1935. The clay ratio as a criterion assusceptibility of soils to erosion. J. .,Madison, Wisc., 27:738-741.
Christofoletti, A. 1981. . O canalfluvial. vol. 1, São Paulo 313 p.
Coutinho, M. E.; Barbosa, F. A. R. 1986. Distribuiçãovertical de matéria orgânica, nitrogênio orgânicototal, fósforo total e algumas formas iônicas nossedimentos recentes de 3 lagos de Minas Gerais.
., 1:401-429.D'Agostini, L. R. 1999.
. Editora da UFSC, Florianópolis 131pp.Darwich, A. J. 1995.
Teste de toxicidade de sedimentosda região de Santos-SP-Brasil, (24º S, 46º W),utilizando o anfipoda escavador Tuburenella viscana(crustacea-platyischnopidae)
.
Estudo da dinâmica e do modelo debalanço de massa do carbono no Sistema Estuarino deSantos, São Paulo Brasil
.
Seminário Regional de Ecologia,
Bioikos
BioTupé: Meio Físico, Diversidade Biológica eSociocultural do Baixo Rio Negro, Amazônia Central
The physics of blown sand anddesert dunes
Conservação dosolo
Amer. Soc. Agron
Geomorfologia fluvial
ActaLimnol. Brasil
Erosão. O problema mais que oprocesso .
Processos de decomposição de
UFSCar
Londres. p
Ícone Editora, p
Edgard Blücher, . p
Aprile .et al

Considerações sobre a geoquímica e dinâmica sedimentar do Lago Tupé
83
connection with turbidity currentsand clay suspensions. In: Whitard,W. F. Bradshaw, R. (eds.).
.Butterworth, London p.47-71.
Maglioca, A.; Kutner, A. S. 1964.Conteúdo orgânico dos sedimentosde fundo de Cananéia. São Paulo,Contribuições avulsas do InstitutoOceanográfico de São Paulo.
, 7 15pp.Maglioca, A.; Kutner, A. S. 1965.
Sedimentos de fundo da Enseada doFlamengo. Instituto Oceanográfico,USP, São Paulo,
, 8:1-15.Martins, S. G.; Silva, M. L. N.; Curi, N.;
Ferreira, M. M.; Fonseca, S.;Marques, J. J. G. S. M. 2003. Perdasde solo e água por erosão hídricaem sistemas florestais na região deAracruz (ES). ,27:395-403.
Mason, C. C.; Folk, R. L. 1958.Differentiation of bead, dune andaeolian flat environments by sizeanalysis. Mustang Island.
, 28:211-226.Müller, G. 1967.
(Part I).Hafner Publ. Co., 283 p.
Pettijohn, F. J. 1940. Relativeabundance of size grades of clasticsediments.
, 1940meeting.
&Submarine geology and geophysics
.
SérieOceanog. Física .
Série Oceanog.Física
R. Bras. Ci. Solo
Journ. ofSed. Petrol.
Methods insedimentary petrography
Program Soc. Econ.Paleont. and Mineral
New York. p
Postma, H. 1967. Sediment transport and sedimentationin the estuarine environment. In: Lauff, G. H. (ed.).
., Washington, DC p.158-179.
Russell, R. J. 1968. Where most grains of very coarsesand and fine gravels are deposited. ,11:31-38.
Ruxton, B. P. 1968. Measures of the degree of chemicalweathering of rocks. ., 76:518-527.
Shepard, F. P. 1954. Nomenclature based on sand-silt-clay ratios. ., 24(3):151-158.
Shepard, F. P.; Moore, D. G. 1954. Sedimentaryenvironments differentiated by coarse fractionstudies. . Geol., 38:1792-1802.
Suguio, K. 1973. . Ed.Edgard Blucher/EDUSP, São Paulo 110pp.
Suguio, K. 1994.. 4 ed., Edgard
Blücher, 500 p.Toledo, M. C. M.; de Oliveira, S. M. B.; Melfi, A. J. 2000.
Intemperismo e formação do solo. In: Teixeira, W.;Toledo, M. C. M.; Fairchild, T. R. Taioli, F. (eds.).
. Oficina de Textos,568 p.
Udden, J. A. 1898. Mechanical composition of winddeposits. 1:1-69.
Wentworth, C. K. 1922. A scale of grade and glass termsfor clastic sediments. , 30:377-392.
Wentworth, C. K. 1933. Fundamental limits to the sizesof clastic grains. , 77:633-634.
Wischmeier, W. H.; Smith, D. D. 1958. Rainfall energyand the relationships to soil loss.
, 39:285-291.Young, E. J. 1966. A critique of methods for comparing
heavy mineral suites. ., 36:57-65.
Estuaries. Amer. Assoc. Adv. Sci .
Sedimentology
J. Geol
J. Sedim. Petrol
Bull. Amer. Assoc. Petrol
Introdução à sedimentologia.
Rochas Sedimentares. propriedades,gênese, importância econômica
Decifrando a Terra
Augustana Library Publ.
J. Geol.
Science
Trans. Am. Geoph.Union
J. Sediment. Petrol
São Paulo. p
São Paulo.p
&


2CAPÍTULO 5 - Fitoplâncton
Sérgio MELO, Sérgio R. M. REBELO, Karla F. SOUZA, Mariângela MENEZES2 Lezilda C. TORGAN&
CAPÍTULO 6 - - Desmídias com ocorrência planctônica
CAPÍTULO 7 - Distribuição vertical do zooplâncton e sua relação com as variáveisambientais
CAPÍTULO 8 - As esponjas do Lago Tupé
CAPÍTULO 9 - Diversidade de macromicetos lignolÍticos.
CAPÍTULO 10 - O uso da fauna pelos moradores da Comunidade São João e Colônia Central
CAPÍTULO 11 - Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
CAPÍTULO 12 - Composição da Entomofauna Aquática em Igarapés da Reserva deDesenvolvimento Sustentável Tupé, Manaus, AM
CAPÍTULO 13 - Diversidade e composição da ictiofauna do Lago Tupé
CAPÍTULO 14 - Bancos de dados sobre biodiversidade na Amazônia: a experiência do ProjetoBiotupé
Sérgio MELO, Sérgio R. M. REBELO, Karla F. SOUZA, Climéia C. SOARES Maria da Graça SOPHIA
PREVIATTELLI, Danie; SANTOS-SILVA, Edinaldo Nelson dos DARWICH, Assad José
Cecilia VOLKMER-RIBEIRO Franciely Benetti de ALMEIDA
André Luis WILLERDING, Marcerla Farias BARBOSA Thaís Campos BURLAMAQUI
Adriana Kulaif TERRA George Henrique REBÊLO
Karen Souza DINIZ Veridiana Vizoni SCUDELLER
Maria José do Nascimento LOPES
Mauro José CAVALCANTI
&
&
&
&
&
&
Maria Gercilia M. SOARES Kedma Cristine YAMAMOTO&
DiversidadeBiológica
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),


Diversidade BiológicaCapítulo 5
Fitoplâncton
RESUMO - Com o objetivo de avaliar a composição do fitoplâncton da bacia do lago Tupé,quatro estações neste lago e uma no rio Negro foram amostradas mensalmente durantevinte meses, no período de 2002-2003. Um total de 118 táxons, distribuído em seis divisõestaxonômico foi identificado. A divisão Heterokontophyta, em especial a classeBacillariophyceae (diatomáceas), foi a mais representativa em número de táxons, sendo
o gênero com maior número de espécies. Em relação a distribuição espacial dofitoplâncton não se verificou diferença significativa do número total de táxonsencontrados nas estações de amostragem, como também não foi detectado um nítidopadrão de distribuição dos mesmo ao longo dos períodos hidrológicos analisados. Onzetáxons foram constantes, ou seja, estiveram presentes em pelo menos 70% das amostras
Eunotia
Sérgio MELO , Sérgio R. M. REBELO , Karla F. SOUZA ,
Mariângela MENEZES
Lezilda C. TORGAN
1-4 1-5 1-6
2-7
3-7&
1
2
3
4
5
6
7
Instituto Nacional de Pesquisas da Amazônia - Coordenação de Pesquisas em Biologia AquáticaManaus - AM
Universidade Federal do Rio de Janeiro - Museu Nacional - Departamento de BotânicaRio de Janeiro - RJ
Museu de Ciências Naturais - Fundação Zoobotânica do Rio Grande do SulPorto Alegre - RS
Bolsista PRODOC-CAPES;Bolsista PIBIC-INPA;
Bolsista PIBIC-CNPq/INPA;Bolsista de Produtividade do CNPq.
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

88
Introdução
Os estudos ficológicos da Amazônia tiveram início nofinal da primeira metade do século dezenove porEhrenberg (1843), e segundo Huszar (1994) restringia-se,até o início da década de noventa, a aproximadamente66 trabalhos publicados em periódicos, englobandoestudos taxonômicos, florísticos e/ou ecológicos.Aestesse somam as publicações de Huszar (1996a, 1996b,2000), Sophia Huszar (1996), Huszar Reynolds (1997),Ibañez (1997, 1998), Menezes Huszar (1997), Meyer
. (1997), Kristiansen Menezes (1998), Keppeler .(1999a,b), Melo Huszar (2000), Pascoaloto (2001a,b),Díaz-Castro . (2003), Lopes Bicudo (2003) e Melo
. (2004a,b). Considerando a extensão territorial daAmazônia e a disponibilidade de ambientes aquáticoscontinentais, pode se inferir que é incipiente oconhecimento sobre as algas desta região. Dessa forma,a realização de pesquisas que buscam investigar acomposição taxonômica de algas aquáticas continentaisna Amazônia, bem como os fatores que influenciam nasua ocorrência e distribuição são de grande importânciapara o conhecimento da biodiversidade.
A paisagem amazônica caracteriza-se pelaexuberância da floresta tropical e de seus corpos d'água.Estes últimos apresentam características peculiares quepodem influenciar a ocorrência e desenvolvimento dedeterminados grupos taxonômicos de algas. Entre estascaracterísticas destacam-se os diferentes tipos de águasquanto à coloração (Sioli, 1950) e o pulso de inundação(Junk . 1989). Uma marcada flutuação na densidade e na composição específica do fitoplâncton tem sidoregistrada em estudos sazonais e associada à influênciado pulso de inundação (Ibañez, 1987, 1988; HuszarReynolds 1997; Huszar, 2000). Embora poucos estudostenham se centrado sobre a riqueza de espécies de algasde ambientes de águas pretas, estes têm apontado para
& &&
&&
&
&
etal et al
et alet al
et al , -
uma elevada diversidade taxonômica(Uherkovich, 1976; Uherkovich Rai,1979; Uherkovich, 1984; Melo .,2004a).
Em especial para o lago Tupé, umpequeno lago de águas pretas,localizado próximo a Manaus,destacam-se duas investigaçõesenvolvendo estudos taxonômicos dofitoplâncton. Na primeira, realizadapor Crönberg (1989) foi registrada aocorrência de dez crisofíceas, dasquais destaca-se var.
Cronberg, uma nova variedadedescrita. A segunda investigaçãorealizada por Meyer . (1997)apresenta a descrição de uma novaespécie de dinoflagelado,
B. Meyer, a partir dematerial da região de Manaus,incluindo o do lago Tupé. A ausência deestudos envolvendo um ciclohidrológico, e principalmente, ummaior número de grupos taxonômicos,motivou a realização do presentetrabalho sobre a flora ficológica dabacia do lago Tupé.
No âmbito da presente pesquisa,amostras para análise qualitativa equantitativa do fitoplâncton vêmsendo coletadas desde o início de2002, com o objetivo de conhecer ariqueza de espécies e ecologia destesorganismos. Neste primeiro momento,o objetivo deste trabalho é apresentaruma relação de espécies encontradasno plâncton, exceto desmídias, da
&et al
Synura spinosastriata
et al
Peridinio-psis amazonica
Melo et al.
analisadas ao longo do estudo. Dentre estes destacaram-se Grunow,var. (Ehrenberg) Simonsen, cf.
Stein, Nygaard e f. Ehrenberg.
fitoplâncton, lagos de inundação, águas pretas,Amazônia.
Actinella guianensis
Aulacoseira granulata granulata Peridinium umbonatum
P. gatunense Dinobryon c sertularia
PALAVRAS-CHAVE:

89
bacia do lago Tupé, identificadas apartir de amostras coletadas em cincoestações durante vinte meses,buscando evidenciar a influência dopulso de inundação sobre a distribui-ção temporal e espacial dessacomunidade neste ambiente de águaspretas.
O estudo foi baseado em amostra-gens realizadas na Reserva de Desen-volvimento Sustentável do Tupé (RDSTupé), em quatro estações delimitadasno lago Tupé (ET1, ET5, ET9 e ET10) deforma a tentar obter amostrasrepresentativas de todo o lago, e umaestação na região central do rio Negro(EN12) próximo do lago. O mapa delocalização geográfica da área deestudo e das estações de coletas, bemcomo, o posicionamento geográficodas mesmas é apresentado por Darwich
. (2005, neste volume).A identificação taxonômica do
fitoplâncton foi fundamentada emamostras coletadas através de arrastosverticais e horizontais com rede deabertura de malha de 25 m e fixadasem solução de Transeau (BicudoBicudo, 1970). Amostras de materialvivo de algumas estações foramtambém examinadas. As coletas foramrealizadas mensalmente no período demarço de 2002 a outubro de 2003. No
Material e Métodos
et al
�
&
total 100 amostras foram analisadas em microscópioóptico com aumento de 400 a 1600 vezes. O sistema declassificação adotado para as divisões taxonômicas foibaseado em Lee (1999).
Para cada espécie calculou-se o índice deconstância,modificado de Dajoz (1978), que expressa emforma de percentagem o número de coletas contendo aespécie, através da seguinte equação: C = (p x 100)/P,onde C ≥ 70 representa as espécies constantes; 30 ≤ C <70 representa as espécies freqüentes; 10 ≤ C < 30representa as espécies esporádicas; C < 10 representa asespécies raras; é o número de coletas contendo aespécie e é o número total de coletas.A riqueza especí-fica foi estimada como sendo o número total de táxons.
Valores de saturação de oxigênio, temperatura daágua e pH foram obtidos com medidor portátil WTWMultLine P3 e condutividade elétrica com medidorportátil WTW LF 320, na subsuperfície da coluna d'águano momento da coleta. A transparência da água foiestimada pela profundidade de extinção do disco deSecchi. As coletas abrangeram os quatro períodos dociclo hidrológico, pelo menos uma vez (Tab. 1).
O valor médio da profundidade das estações do lagoTupé variou de 3,6m no período de seca (águas baixas) a11,8m, no período de cheia (águas altas). As condiçõesde temperatura, pH, transparência da água nos quatroperíodos do ciclo hidrológico não apresentaram grandesvariações. Entretanto, observou-se uma variação maisnítida em relação saturação de oxigênio e decondutividade elétrica. A condutividade elétrica da águafoi mais alta (valor médio= 15 Scm ) no período de cheiae mais baixa (valor médio de 5,8 Scm ) no período de
pP
Resultados e Discussão
�
�
-1
-1
Fitoplâncton
Tabela 1. Períodos do ciclo hidrológico e os respectivos meses de coletas de algas planctônicas na RDSTupé, no período de março (M) de 2002 a outubro (O) de 2003.
2002 2003
M A M J J A S O N D J F M A M J J A S O
Enchente (ENC)
Águas altas/Cheia (AA)
Vazante (VAZ)
Águas baixas/Seca (AB)

90
seca, enquanto que a saturação de oxigênio foi maisbaixo (< 38%) no período de cheia e mais alto (>43%) noperíodo de seca. Por outro lado, os períodos de vazante eenchente não demonstraram tendência definida emrelação a essas variáveis. Os valores médios, por
períodos do ciclo hidrológico dasprincipais caracterís ticas limnológicassão apresentados na Tabela 2.
Um total de 118 táxons foi identifi-cado (Tab. 3), distribuído em seis
-
Melo et al.
Tabela 2. Valores médios da profundidade das estações de coletas no lago (m), da profundidadede extinção do disco de Secchi (m), da temperatura da água (°C), da saturação de oxigênio (%),de pH e da condutividade elétrica da água (S cm-1) nos diferentes períodos do ciclo hidrológico(Enchente ENC; Águas altas/Cheia AA; Vazante Vaz e Águas Baixas/seca (AB), de março de2002 a outubro de 2003.
Prof. (m) Secchi (m) Temp.(ºC) OD (%) pH Cond. µScm-1
AA ET1 9.7 1.2 29.9 38.0 4.3 9.0
ET5 13.0 1.3 29.8 37.1 4.3 9.6
ET7 10.7 1.1 29.8 32.3 4.3 10.2
ET10 13.8 1.0 29.7 44.7 4.3 9.8
EN12 - 0.9 28.8 37.1 4.2 10.3
VAZ ET1 5.9 1.1 30.5 54.5 4.4 7.7
ET5 8.5 1.2 30.6 50.6 4.4 7.8
ET7 5.9 1.0 30.7 49.9 4.4 8.0
ET10 10.1 1.1 30.6 51.5 4.5 8.0
EN12 - 1.0 31.8 45.8 4.1 9.5
AB ET1 2.2 1.1 28.8 67.0 3.9 8.6
ET5 4.3 1.3 30.4 73.2 4.4 -
ET7 2.0 1.0 31.2 84.5 5.1 -
ET10 5.8 1.1 30.3 43.6 4.5 -
EN12 - 1.1 31.2 46.2 4.2 7.9
ENC ET1 4.9 1.3 31.0 78.4 4.7 8.0
ET5 - 1.3 30.8 85.1 4.7 9.2
ET7 4.5 1.2 30.1 65.6 4.6 10.1
ET10 9.0 1.2 29.9 73.0 4.8 8.9
EN12 - 1.1 30.1 82.4 5.5 12.0
AA ET1 10.0 1.0 30.5 19.1 4.1 14.1
ET5 12.0 1.1 31.1 22.0 3.9 15.0
ET7 9.5 1.2 30.4 18.4 3.8 23.9
ET10 13.5 1.2 30.0 17.7 3.9 21.7
EN12 - 1.1 30.0 18.6 3.7 21.9
VAZ ET1 8.1 1.1 31.4 57.3 4.1 9.5
ET5 8.0 1.0 30.8 30.4 3.9 9.9
ET7 5.0 1.0 31.2 30.9 3.7 9.7
ET10 8.4 1.2 30.5 31.4 3.9 10.4
EN12 - 1.0 29.4 23.1 3.9 11.5- ausência de dados
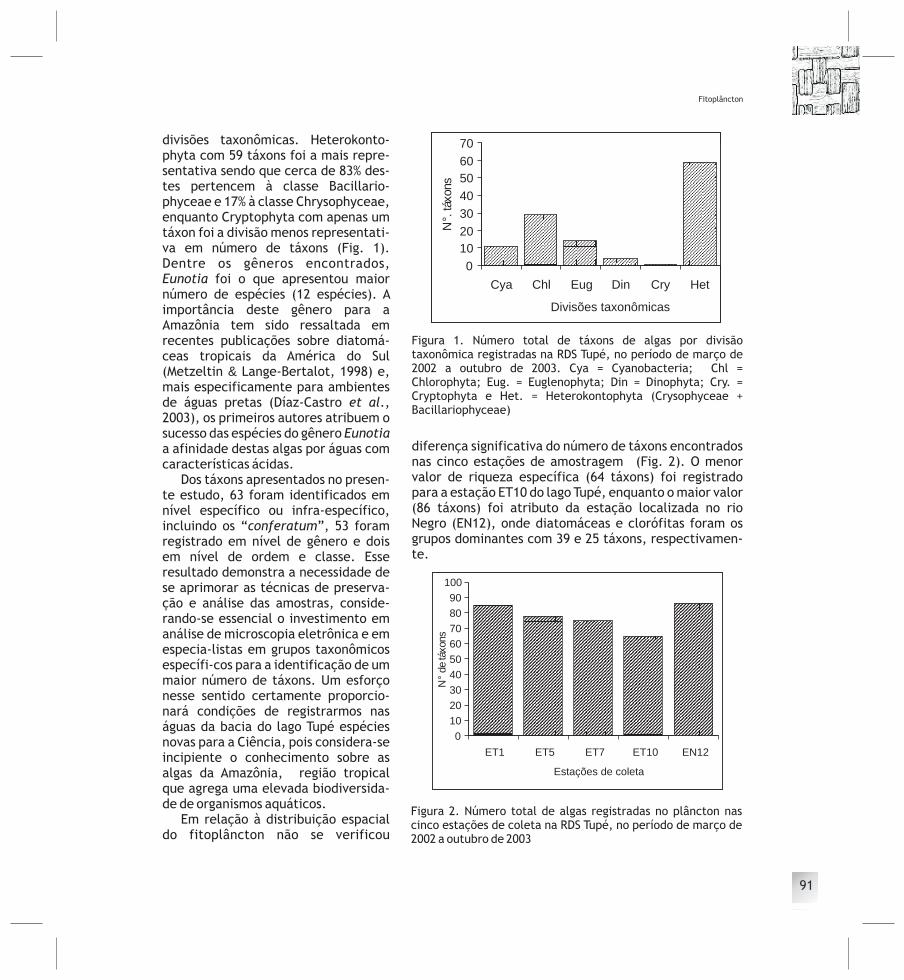
divisões taxonômicas. Heterokonto-phyta com 59 táxons foi a mais repre-sentativa sendo que cerca de 83% des-tes pertencem à classe Bacillario-phyceae e 17% à classe Chrysophyceae,enquanto Cryptophyta com apenas umtáxon foi a divisão menos representati-va em número de táxons (Fig. 1).Dentre os gêneros encontrados,
foi o que apresentou maiornúmero de espécies (12 espécies). Aimportância deste gênero para aAmazônia tem sido ressaltada emrecentes publicações sobre diatomá-ceas tropicais da América do Sul(Metzeltin Lange-Bertalot, 1998) e,mais especificamente para ambientesde águas pretas (Díaz-Castro .,2003), os primeiros autores atribuem osucesso das espécies do gêneroa afinidade destas algas por águas comcaracterísticas ácidas.
Dos táxons apresentados no presen-te estudo, 63 foram identificados emnível específico ou infra-específico,incluindo os “ ”, 53 foramregistrado em nível de gênero e doisem nível de ordem e classe. Esseresultado demonstra a necessidade dese aprimorar as técnicas de preservação e análise das amostras, considerando-se essencial o investimento emanálise de microscopia eletrônica e emespecia-listas em grupos taxonômicosespecífi-cos para a identificação de ummaior número de táxons. Um esforçonesse sentido certamente proporcio-nará condições de registrarmos naságuas da bacia do lago Tupé espéciesnovas para a Ciência, pois considera-seincipiente o conhecimento sobre asalgas da Amazônia, região tropicalque agrega uma elevada biodiversida-de de organismos aquáticos.
Em relação à distribuição espacialdo fitoplâncton não se verificou
Eunotia
et al
Eunotia
conferatum
--
&
diferença significativa do número de táxons encontradosnas cinco estações de amostragem (Fig. 2). O menorvalor de riqueza específica (64 táxons) foi registradopara a estação ET10 do lago Tupé, enquanto o maior valor(86 táxons) foi atributo da estação localizada no rioNegro (EN12), onde diatomáceas e clorófitas foram osgrupos dominantes com 39 e 25 táxons, respectivamen-te.
Fitoplâncton
91
0
1020
30
40
5060
70
Cya Chl Eug Din Cry Het
Divisões taxonômicas
N°.
táxo
ns
Figura 1. Número total de táxons de algas por divisãotaxonômica registradas na RDS Tupé, no período de março de2002 a outubro de 2003. Cya = Cyanobacteria; Chl =Chlorophyta; Eug. = Euglenophyta; Din = Dinophyta; Cry. =Cryptophyta e Het. = Heterokontophyta (Crysophyceae +Bacillariophyceae)
0
10
20
30
40
50
60
70
80
90
100
ET1 ET5 ET7 ET10 EN12
Estações de coleta
N°
detá
xons
Figura 2. Número total de algas registradas no plâncton nascinco estações de coleta na RDS Tupé, no período de março de2002 a outubro de 2003

Táxons/Estações ET1 ET5 ET7 ET10EN12Cyanobacteria
Anabaena sp. E R R EAphanocapsa sp. RChroococcus planctonicuscf. Bethge E EChroococcus sp. R ECoelosphaeropsis .sp REucapsis parallelepipedon (Schmidle) Komárek & Hindák E RGloeocapsa sp. RMerismopedia tenuissimacf. Lemmermann E E E F EMerismopedia sp. RMycrocystis sp. R R E EOscillatoriales sp. E E E EChlorophyta
Ankistrodesmus sp. E E EBotryococcus sp. E E E E RChlorella sp. R RCoelastrum sp. R E R ECoenocystis sp. RDesmodesmus armatuscf. (R. Chodat) Hegewald R E R RDesmodesmus communiscf. (Hegewald) Hegewald E F E E FDesmodesmus opoliensiscf. (P. Richt.) Hegewald E R E R EDesmodesmus sp. E F E E RDictyosphaerium pulchellumcf. Wood F E F F CDictyosphaerium sp. REudorina eleganscf. Ehrenberg E E E F EKirchneriella ianae (Bohl.) Comasd E E E F FKirchneriella obesa (W. West) Schmidle E E E E FMougeotia sp. E E E ROocystis sp. E E E E EPandorina sp. E R R EPediastrum duplex Meyen E R R EPediastrum sp. R R R EScenedesmus acuminatuscf. (Lagerh.) Chodat R R RScenedesmus ellipticus Corda R R RScenedesmus javanensis Chodat E ESphaerocystis planctonicacf. (Korch.) Bourrelly RSpirogyra sp. R R R E
Tabela 3 Lista de táxons de algas registrados no plâncton na RDS Tupé, no período de março de 2002 a outubrode 2003 e o índice de constância de cada espécie: = constantes; = freqüentes; = esporádicas e = raras
.C F E R
continua >
92
Melo et al.

Táxons/Estações ET1 ET5 ET7 ET10EN12Tetraedron minimum (A. Br. ) Hansgirg R E ETetraedron sp. RTetrallanthos lagerheimii Teiling R RVolvox sp. RZygnema .sp R RDinophyta
Gymnodinium sp. E F R F EPeridiniopsis amazonica B. Meyer F F E E EPeridinium gattunense Nygaard C C F C FPeridinium umbonatumcf. Stein C C F C FCryptophyta
Cryptomonas sp. E E REuglenophyta
Euglena acus Ehrenberg E E REuglena allorgei Delflandre E E E REuglena oxyuris Schamarda R EEuglena spirogyra Ehrenberg EEuglena sp.1 REuglena sp.2 ELepocinclis sp. RPhacus longicauda (Ehrenberg) Dujardin R RPhacus orbiculares Hübner E R E EPhacus suecicus Lemmermann R RTrachelomonas armatacf. (E.) Stein RTrachelomonas hispidacf. (Perty) Stein emend Deflandre R R RTrachelomonas . superbacf Swir emend Deflandre RTrachelomonas sp. E E E R EHeterokontophyta (Chrysophyceae)
Dinobryon sertulariacf. Ehrenberg C F C C FEpipyxis sp. R R EMallomonas sp.1 F E E E FMallomonas sp.2 E E E E EMallomonas sp.3 R R R R RMallomonas sp.4 E E E E EMallomonas sp.5 R RMallomonas sp.6 ESynura spinosa striatavar. Cronberg F F F E ESynura sp. R
> continuação
continua >
Fitoplâncton
93

Táxons/Estações ET1 ET5 ET7 ET10EN12Heterokontophyta (Bacillariophyceae)
Actinella brasiliensis Grunow E C E R RActinella guianensis Grunow C R C C CActinella mirabilis Grunow E E C E CAsterionella sp. F F F F CAulacoseira agassiziicf. (Ostenfeld) Simonsen R F E E EAulacoseira ambiguacf. (Grunow) Simonsen R RAulacoseira granulata granulata(Ehrenberg) Simonsen var. F F C C CAulacoseira herzogii (Lemmermann) Simonsen R R F RAulacoseira sp.1 F F F CAulacoseira sp.2 F F FCaloneis sp. REncyonema sp. EEunotia asterionelloides Hustedt E E R E EEunotia bicornigera Metzeltin & Lange Bertalot- E REunotia femoriformis (Patrick) Hustedt R E EEunotia glacialiscf. Meister E F F F FEunotia lineolata HustedtEunotia naegelii Migula E E F REunotia paludosa Grunow E E R R EEunotia subrobusta *Hustedt R EEunotia synedraeformis angustatavar. Hustedt R E E E REunotia trigibbacf. Hustedt REunotia veneriscf. (Kützing) De Toni R REunotia sp. RFragilaria javanicacf. Hustedt R R E E EFrustulia rhomboides (Ehrenberg) De Toni F F F F FFrustulia sp E RGomphonema archaevibrio Lange-Bertalot & Reichardt fo- .
archaevibrio F F E E EG. archaevibrio cuneatumfo. Metzeltin & Lange-Bertalot F F F F EKobayasia sp. E E E FNavicula sp. R R E R ENeidium excisum Kramer & Metzeltin RPinnularia acrosphaeria W. Smith RPinnularia braunii (Grunow) Cleve EPinnularia interruptacf. W. Smith R RPinnularia sp. 1 F E R E E
> continuação
continua >
94
Melo et al.

Táxons/Estações ET1 ET5 ET7 ET10EN12Pinnularia sp.2 EStenopterobia planctonica Metzeltin & Lange-Bertalot R E RStenopterobia sp. ESurirella biseriatacf. Brébisson R ESurirella sp.1 E E R E FSurirella sp.2 E E R ESynedra sp. F F F F FTabellaria sp. E E E E FUrosolenia eriensis eriensis(H.L. Smith) var. Round & Crawford C C F F FUrosolenia erisensis morsavar. (W. West & G. S. West) Torgan F F E E EUrosolenia longiseta F F F F FUrosolenia sp. E E E RBacillariophyceae sp. E E R E*Citada como em Melo (2004a)Eunotia serra et al.
> continuação
O fato da estação no rio Negroapresentar a maior riqueza de espéciespode ser explicado pela suaproximidade do rio Amazonas (ca. de30Km), o que proporciona umrepresamento das águas do rio Negro,em especial no período de águas altas,conferindo a este ambiente lóticocertas características lênticas além dofato de o mesmo poder ter carreadoalgas de outros sistemas aquáticos àmontante. Tais condições são comunspara rios de pequenas correntezas,localizados em grandes planícies, ondeo tempo de residência da massa d'águapode se r su f i c i en te pa ra odesenvolvimento de um verdadeiroplâncton ou ter organismos oriundos dodesprendimento de células bentônicasou de lagos e represas ao longo docurso d'água (Allan, 1995).
Embora a análise da composiçãoespecífica do fitoplâncton, coletadocom rede, possa fornecer algumasinformações importantes sobre opadrão de flutuação temporal dofitoplâncton, diferentemente doesperado, não foi evidenciado um
Fitoplâncton
95
nítido padrão de flutuação temporal no número detáxons (Fig. 3). Maior riqueza específica nos períodos deenchente e seca e menores nos períodos cheia e vazante,conforme evidenciado por Melo (2004) para outroambiente de águas pretas na bacia do rio Negro, não foiconstatado no lago Tupé.
Analisando a ocorrência dos táxons nas estaçõesestudadas ao longo dos vinte meses, onze táxons foramconstantes, estando presentes nas estações em pelomenos 70% das amostras analisadas, e/ou freqüentespresentes entre 30 e 69,9% das amostras analisadas -durante o estudo (Tab. 3), entre os quais destacam-se asdiatomáceas, em especial as espécies
e var. evar. ¸ acompanhados por
cf. , e cf.. Associado a estes táxons foi registrado
elevada riqueza de espécies de desmídias que sãotratadas em literatura como comuns em ambientesoligotróficos e com reduzidos valores de pH (Melo .2005, neste volume).
Em síntese, o presente estudo é pioneiro nosentido de abordar em escala temporal e espacial, pormais de um ciclo hidrológico, o fitoplâncton do lago Tupée rio Negro, onde tem apontado uma elevada riqueza deespécies de algas no plâncton, com uma maiorcontribuição de diatomáceas e desmídias (Melo .2005, neste volume). No entanto, evidencia a
et al.
Actinellaguianensis Aulacoseira granulata granulataUrosolenia eriensis eriensisPeridinium umbonatum P. gatunense Dinobryonsertularia
et al
et al

96
Melo et al.
ET10
0
10
20
30
EN12
0
10
20
30
M A M J J A S O N D J F M A M J J A S OMeses de coleta
ET7
0
10
20
30
ET1
0
10
20
30N
°de
táxo
n sN
°de
táxo
n sN
°de
táxo
n sN
°de
táxo
n sN
°de
táxo
n s
ET5
0
10
20
30
Figura 3. Número total de táxons de algas registradosmensalmente em cada estação de amostragem na RDS Tupé, noperíodo de março de 2002 a outubro de 2003.
necessidade de maiores investimentospara ampliar o conhecimentotaxonômico do fitoplâncton e de suaimportância ecológica na regiãoAmazônica.
Agradecemos a Dra. Ina de SousaNogueira pelo auxílio na identificaçãode algumas espécies de Chlorophyceaee ao laboratório de Ficologia do MuseuNacional-UFRJ e aos diretores doMuseu de Ciências Naturais-FundaçãoZoobotânica do Rio Grande do Sul, quenos disponibilizaram bibliografias eequipamentos para análise do materialcoletado. Este trabalho teve o apoiofinanceiro do CNPq através do ProjetoBioTupé (Proc. 551361/001-8) e dabolsa de RD (Proc.301349/01-0) e de IC(PIBIC/CNPq/INPA), CAPES (PRODOC) eINPA-MCT (PPI 1-3540 e PIBIC/INPA).
Allan, J. D. 1995.
. Chapman Hall ed. 388pp.Bicudo, C. E. M.; Bicudo, R.M.T. 1970.
Coleta e preservação de algas deáguas continentais brasileiras, SãoPaulo. 228pp.
C r o n b e r g , G . 1 9 8 7 . S c a l e dchrysophytes from the tropics.
, 95: 191-232.Darwich, A. J.; Aprile, F. M.;
Robertson, B. A. 2005.
Santos-Silva, E. N.; Aprile, F. M.; Scudeller,V. V. Melo, S.(Orgs.). Biotupé:
Agradecimetos
Bibliografia Citada
Stream Ecology:Structure and function of runningwaters
Nova Hedwigia
Variáveislimnológicas: Contribuição aoestudo espaço-temporal de águaspretas amazônicas. In:
&
&

Fitoplâncton
97
Meio Físico, Diversidade Biológica eSociocultural do Baixo Rio Negro,Amazônia Central. Editora INPA,Manaus-AM.
Díaz-Castro, J. G.; Souza-Mossimann,R. N., Laudares-Silva, R.; Forsberg,B. R. 2003. Composição dacomunidade de diatomáceasperifíticas do Rio Jaú, Amazonas,Brasil. 33 (4): 583-606.
Dajoz, R. 1978. . 3° Ed.Editora Vozes LTDA. Petrópolis-RJ.472pp.
Ehrenberg, C. G. 1843.
.Durckerei der koniglischen, Akad.Wiss. Berlin. p.291-415.
Huszar, V.L.M. 1994.
. Tese. UniversidadeFederal de São Carlos, PPG-ERN,São Carlos, SP 229pp.
Huszar, V.L.M. 2000. . In:Bozelli, R.L., Esteves, F. A.;Roland, F (Eds.). Lago Batata:Impacto e Recuperação de umEcossistema Amazônico. Institutode Biologia-UFRJ e SociedadeBrasileira de Limnologia. P:91-104.
Huszar, V.L.M.; Reynolds, C.S. 1997.Phytoplankton periodicity andsequences of dominance in anAmazonian flood-plain lake (LagoBatata, Pará, Brasil): responses togradual environmental change.
346:169-181.Huszar, V. L. M.; 1996. Floristic
compositionand biogeographicalaspects of the phytoplankton of anAmazonian flooplain lake (LakeBatata, Pará, Brasil).
Acta Amazônica
Ecologia geral,
Verbreitungund Einfluss des mikroscopischenlebens in Sud und Nordmerika
Fitoplâncton deum lago amazônico impactado porrejeito de bauxita (lago Batata,Pará, Brasil): estrutura decomunidade, flutuações espaciaise temporais
.Fitoplâncton
Hydrobiologia,
Acta
Limnologica Brasiliensia
Verh. Internat. VereinLimnol
Hydrobiologia
Spec. Publ.Can. J. Fish.Aquat. Sci ,
Revista Brasileira de Biologia
Revista Brasileira de Biologia
Phycology
Acta Amazonica
J. Plank. Research
Biodiversidade de algas planctônicas do ParqueNacional do Jaú: Janela Seringalzinho
Amazoniana (1/2)
Desmídias com ocorrênciaplanctônica
;do
, 8:127-136Ibañez, M. S. R. 1997. Phytoplankton biomass of a central
Amazonian floodplain lake..,26:605 - 609.
Ibañez, M.S.R. 1998. Phytoplankton composition andabundance of a central Amazonian floodplain lake.
, 362:78-83.Junk, W.P.; Bayley, P.B., Sparks, R.E. 1989. The flood
pulse concept in river floodplain system.. 106: 110-127.
Keppeler, E.C., Lopes, M. R.M. e Lima, C. S. 1999.Ficoflórula do lago Amapá em Rio Branco- Acre, I:Euglenophyceae. ,59(4): 679-686.
Keppeler, E.C., Lopes, M. R.M.; Lima, C. S. 1999.Ficoflórula do lago Amapá em Rio Branco- Acre, IIChlorophyta. , 59(4):687-691.
Lee, R. . 3° edition. Ed. Cambridge UniversyPress., United Kingdom. 614pp.
Lopes, M. R.; Bicudo, C.E.M. 2003. Desmidioflórula deum lago de planície de inundação do Rio Acre, Estadodo Amazonas, Brasil. , 33(2): 167-211.
Melo, S.; Huszar, V.L.M. 2000. Phytoplankton in anAmazonian flood-plain lake (Lago Batata, Brasil): dielvariation and species strategies. ,22(1):63-76.
Melo, S.; Sophia, M.G.; Menezes M.; Souza, C. A. 2004a.
. In: Borges, S.H.; Iwanaga, S.; Durigan, C. C.; Pinheiro, M. R.(Eds).Janelas para a biodiversidade do Parque Nacional doJaú: uma estratégia para o estudo da biodiversidadedaAmazônia. Ed. Ipiranga, Brasília-DF.
Melo, S.; Huszar, V.L.M., Roland, F.; Esteves, F. A.;Bozelli, R. 2004b. Phytoplankton diel variation andvertical distribution in two Amazonian floodplainlakes (Batata Lake and Mussurá Lake, Pará-Brasil)with different mixing regimes. 18 :1-10.
Melo, S.; Rebelo, S. R. M.; Souza, F. S.; Soares, C..;Sophia, M. G. 2005.
. In: Santos-Silva, E. N.; Aprile, F. M.;Scudeller, V. V. Melo, S.(Orgs.). Biotupé: meio físico,diversidade biológica e sociocultural baixo rio
p.83-95.

98
Melo et al.
Negro. INPA, Manaus-AM.Menezes M. Huszar V. L. M. 1997. , a
new species of Chrysophyceae from the Amazonregion, northern Brazil. 85:13-22.
Metzeltin, D.; Lange-Bertalot, H. 1998.I. Lange-Bertalot, H.
(Ed.). Iconographia Diatomologica. Annoted DiatomMicrographs. Vol. 5. Koeltz Scientific Books. 695pp.
Meyer B., Rai, H., Cronberg, G. 1997. The thecalstructure of Peridiniopsis amazonica spec. nov.(Dinophyceae), a new cist-producing freshwaterdinoflagellate fromAmazonian floodplain lakes.
65 (1-4): 365-375.Pascoaloto, D. 2001a. Características ambientais de
cinco igarapés de terra-firme em reservas florestaisno estado do Amazonas e sua relação com
(Batrachospermaceae, Rhodophyta). 31(4): 597-606.
Pascoaloto, D. 2001b. Características físicas e químicasde sete igarapés de terra-firme no Estado do
; Bitrichia amazônica
Algological Studies,
Tropicaldiatoms of South America In:
NovaHedwigia,
Batrachospermum cayennense -ActaAmazônica,
Amazonas e sua relação comspp. (Batrachos
permaceae, Rhodophyta)., sér. Bot.
17(1):3-19.Sophia, M. G.; Huzsar. V. L. M. 1996.
Planktonic desmids of threeAmazonian systems ( Lake Batata,Mussurá and Trombetas River),Pará, Brazil. XIV (1/2) 75-90
Sioli, H. 1950. Das Wasser imAmazona s geb i e t .
., 26:274-280.Uherkóvich, G. 1984. .
In: Sioli, H. (Ed). The Amazon:Limnology and landscape ecologyof a might tropical river and itsbasin. Dr. Junk PublishersDordrecht, London/Lancaster.p.295-310.
Batrachospermum -Bol.
Mus. Para. Emílio Goeldi ,
Amazoniana,:
Fo r s chn .Fortschr
Phytoplankton
,

Diversidade BiológicaCapítulo 6
Desmídias com ocorrência planctônica
RESUMO - Quatro estações de coleta no lago Tupé e uma no rio Negro, próximo ao lagoTupé, foram amostradas em escala mensal durante vinte meses com rede de plâncton,malha de 25mm. No presente trabalho são apresentados os resultados referentes asdesmídias. Um total de 107 táxons foi registrado, os quais estão contidos em 19 gêneros,sendo Meyen, Teiling e Nitsth ex Ralfs os gêneroscom maior riqueza específica. Espacialmente, uma maior riqueza de espécies foiregistrada na estação do rio Negro. Temporalmente foi constatada tendência a maiorriqueza de espécies no período de seca e enchente e menor no período de cheia e vazante.O presente estudo soma evidências sobre a importância das desmídias para ambientesamazônicos e amplia o conhecimento deste grupo de algas para os ambientes de águaspretas, além de ser um dos únicos trabalhos a analisar a composição taxonômica dedesmídias a partir de amostras mensais de águas da região limnética de ambientesamazônicos por mais de um ano de estudo.
PALAVRAS-CHAVE: Desmídias, lago de inundação, águas pretas,Amazônia.
Staurastrum Staurodesmus Closterium
Sérgio MELO , Sérgio R. M. REBELO , Karla F. SOUZA , Climéia C. SOARES
Maria da Graça SOPHIA
1-3 1-4 1-5 1
2&
Instituto Nacional de Pesquisas da Amazônia Coordenação de Pesquisas em Biologia AquáticaManaus - AM
Universidade Federal do Rio de Janeiro / Museu Nacional - Departamento de BotânicaRio de Janeiro - RJ
Bolsista PRODOC - CAPES;Bolsista PIBIC - INPA;
Bolsista PIBIC - CNPq/INPA
2
3
4
5
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

100
Uma importante característica dossistemas aquáticos da Amazôniarefere-se à coloração de suas águas.Com base nesta característica elasforam classificadas por Sioli (1950) emáguas claras, brancas e pretas; estasúltimas caracterizam-se pela escassezde minerais e sólidos em suspensão,porém são ricas em compostosorgânicos coloridos e ácidos. Comoprincipal representante deste tipo deágua destaca-se o rio Negro e a maioriade seus tributários e lagos adjacentes.Pouco, porém, é conhecido sobre ariqueza de espécies e flutuaçãotemporal das desmídias em ambientesde águas pretas da Amazônia. Oconhecimento do fitoplâncton desistemas húmicos na região amazôni-ca, bem como para o restante do Brasilé limitado, contando com poucosestudos, entre estes, destacam-se osapresentados por Melo . (2004), eas publicações de Martins (1980, 1982,1986a, 1986b) que apresentam adesmidioflórula do lago Cristalino,ambiente localizado próximo a Manausna margem direita do rio Negro e dolago São Sebastião localizado a cercade 60Km de Manaus na bacia do rioSolimões, embora apresente águaspretas.
Considerando a grande dimensãogeográfica e grande quantidade ediversidade de ambientes aquáticoscontinentais amazônicos, reveste-sede grande importância ampliar oconhecimento sobre a biodiversidade eecologia de algas planctônicas deambientes de águas pretas, bem comoanalisar o padrão de flutuaçãotemporal e espacial destes organis-mos, buscando apontar possíveisrelações com as variáveis ambientais.Desta forma, o objetivo deste trabalhoé apresentar uma lista de espécies de
et al
Melo et al.
Introdução
No estado do Amazonas, como na Amazônia de ummodo geral, o pulso de inundação (Junk . 1989)exerce importante influência sobre a flutuação temporale espacial do fitoplâncton (Huszar Reynolds, 1997;Ibañez, 1998; Melo Huszar, 2000). Entre estesorganismos destacam-se as desmídias, grupo de algasverdes (Chlorophyta), geralmente registrados em águasácidas e pobres em nutrientes (Margalef, 1983; Moss,1988; Coesel, 1996). Este último autor discutindo sobre abiogeografia de desmídias constatou um gradualaumento no número de táxons a partir das regiõespolares em direção ao equador, onde se concentra umaelevada diversidade morfológica deste grupo deorganismos e que é apontada como a região de origemdestas algas.
O primeiro registro conhecido de desmídias para aregião Amazônica é atribuído a Dickie (1881). Depoisdisto, importantes contribuições ao conhecimento destegrupo taxonômico foram apresentadas a partir dadécada de 40 do século passado. Entre estas, destacam-se as publicações de Grönblad (1945); Förster (1963,1964, 1969, 1974); Scott . (1965);Thomasson (1971,1977); Martins (1980, 1982, 1986a, 1986b); Bittencourt-Oliveira (1993); Sophia Huszar (1996), e maisrecentemente a de Lopes Bicudo (2003). Thomasson(1971) em uma revisão sobre as algas da Amazônia,destaca o grande número de Desmidiaceae presente (960táxons), enquanto Uherkovich (1984) chama a atençãopara o fato de 389 táxons de algas terem sido descritoscomo novos registros para a Ciência a partir de materialdaAmazônia, e deste total 313 são de desmídias.
Embora a maioria dos trabalhos sobre flórulaficológica da Amazônia refira-se às desmídias,isoladamente ou em conjunto com os demais grupos dealgas, estes ainda são relativamente escassos do pontode vista do conhecimento sobre a distribuição geográficadestes organismos na Amazônia, considerando aimensidão territorial e o fato dos trabalhos existentesestarem concentrados em determinadas regiões, fatocorroborado por Lopes Bicudo (2003) que chamaram aatenção para o fato do estudo sobre algas no estado doAmazonas ser demasiadamente restrito a um pequenonúmero de ambientes.
et al
et al
&&
&&
&

desmídias, bem como a distribuiçãodestes organismos em escala temporale espacial na Reserva de Desenvolvi-mento Sustentável do Tupé (RDS Tupé).
Para o presente estudo foramanalisadas amostras coletadas na RDSTupé. Quatro estações foramdelimitadas no lago Tupé (ET1, ET5,ET7, ET10) e uma na região central dorio Negro (EN12) na direção do lago(Darwich . 2005, neste volume).As coletas foram efetuadas em escalamensal no período de março de 2002 aoutubro de 2003, totalizando 100amostras que foram analisadas.
Para identificação do fitoplâncton,amostras foram coletadas através dearrastos verticais e horizontais comrede de abertura de malha de 25m efixadas em solução de Transeau.Amostras de material vivo de algumasestações foram também examinadas.
Foi calculado o índice de constância(modificado de Dajoz, 1978) para cadaespécie, o que expressa em forma depercentagem o número de coletascontendo a espécie. O índice foicalculado pela seguinte equação: =(p x 100)/P, onde
é ; 30 < 70representa as espécies ; 10
< 30 representa as espécies; < 10 representa as
espécies ; é o número de coletascontendo a espécie e é o númerototal de coletas.
O sistema de classificação adotadofoi baseado no apresentado por Förster(1982), que inclui na ordem Zygnem-atales a família Mesotaeniaceae e naordem Desmidiales as famílias Gonato-zygaceae, Peniaceae, Closteriaceae e
Material e Métodos
et al ,
CC
constantes Cfreqüentes
Cesporádicas C
raras pP
≥ 70 representa as
esp cies ≤
≤
Desmidiaceae. Para identificação dos indivíduos foramobservados caracteres morfológicos como o formato,ornamentação e medidas das células. Esses dados foramanalisados com base em literatura especializada.
O ambiente estudado caracterizou-se por apresentarreduzidos valores de pH e condutividade elétrica (Melo
. 2005, neste volume), sendo estas característicaspropícias ao desenvolvimento das desmídias.Contabilizando todas as estações analisadas no períodode 20 meses, foi constatada a ocorrência de 107 táxonsde desmídias na RDS Tupé (Tab. 1), inclusos em 19gêneros, sendo Meyen,Teiling e Nitzsth ex Ralfs os gêneros commaior riqueza de espécies, enquantoKützing, Teiling, (Nag.) ItzigsohnRothe, Brebisson e Bailey comapenas um táxon, foram os que apresentaram menorriqueza específica (Fig. 1). A importância dos gêneros
e em termos deriqueza de espécies nos ambientes amazônicos tem sidoconstatado em outros estudos, como por exemplo Sophia
Huszar (1996); Lopes Bicudo (2003), Melo(2004).
Resultados e Discussão
et al
Staurastrum StaurodesmusClosterium
BambusinaGroenbladia NetriumPenium Triploceras
Staurastrum, Staurodesmus Closterium
et al.
&
& &
Desmídias com ocorrência planctônica
101
0
5
10
15
20
25
30
35
Nºd
etá
xons
Figura 1 Número total de táxons por gênero de desmídiasregistrados na RDS Tupé, no período de junho de 2002 a outubrode 2003. Outros = e
.
Bambusina, Groenbladia, Netrium, PeniumTriploceras
Clo
ster
ium
Cos
mar
ium
Des
mid
ium
Eua
stru
mG
onat
ozig
onH
yalo
thec
aH
aplo
taen
ium
Mic
rast
eria
sO
nych
onem
aP
leur
otae
nium
Spo
ndyl
osiu
mS
taur
astru
mS
taur
odes
mus
Xan
thid
ium
Out
ros

Táxons/estações ET1 ET5 ET7 ET10 EN12
Bambusina borreri (Ralfs) Cleve E F F E F
Closterium closterioidescf. (Ralfs) Louis & Peeters R
Closterium closterioides intermediumvar. (Roy & Bisset) Ruzicka E R R
Closterium cynthia latumvar. (Schmidle) Krieger R
Closterium kuetzingii Brébisson E F E E F
Closterium macilentumcf. Brébisson R
Closterium navicula (Brébisson) Lütkemüller E E E E E
Closterium nematodes Joshua E R R R E
Closterium pronum Brébisson E E
Closterium setaceum Ehrenberg ex Ralfs R
Closterium sp. E R E
Cosmarium contractum Kirchner E E E E E
Cosmarium lobatum Börgesen R
Cosmarium pseudoconnatumcf. Nordstedt E E E E E
Cosmarium subpraemorsumcf. Borge R
Cosmarium sp. R E E R
Desmidium baileyi (Ralfs) Nordstedt R E E E
Desmidium coarctatum cambricumvar. West E E R E E
Desmidium elegans (Raciborski) Grönblad R R E
Desmidium grevilii (Kutzing) De Bary R R R
Desmidium sp. E E
Euastrum evolutum (Nordesdt) W. & G. S. West E R E E E
Euastrum evolutum perornatumvar. Scott & Croasdale R
Euastrum gemmatum monocylumaff. var. (Nordstedt) De Toni E
Euastrum ornans Förster E E E E E
Euastrum sinuosum Lenorm. ex. Archer E E E
Gonatozygon kinahaniicf. (Archer) Rabenhorst R R
Gonatozygon monataenium pilosellumvar. Nordestedt E E E R
Gonatozygon sp. E R E
Groenbladia sp. R R
Haplotaenium bourrellyi (Grönblad & Scott) Bando E E E
Haplotaenium minutum minutumvar. (Ralfs) Bando E E E R
Haplotaenium rectum forsteriivar. Bando R
Haplotaenium rectum rectumvar. (Delponte) Bando R
Hyalotheca dissiliens . dissiliensvar (Smith) Brébisson ex Ralfs R R E
Hyalotheca sp. Ehrenberg E E E
Micrasterias borgei Krieger R R
Tabela 1 Lista de táxons de desmídias registrados na RDS Tupé, no período de junho de 2002 a outubro de 2003 eo índice de constância de cada espécie: = constantes; = freqüentes; = esporádicas e = raras
.C F E R
continua >
102
Melo et al.

Táxons/estações ET1 ET5 ET7 ET10 EN12
Micrasterias laticeps Nordstedt R
Micrasterias mahabuleshwarensis Hobsen R
Micrasterias radiata Hassal R
Micrasterias rotata (Grev.) Ralfs ex Ralfs R E
Micrasterias siolii Scott & Croasdale R
Micrasterias torreyi borgeivar. Förster R R
Netrium oblongumcf. (De Bary) Lütkemüller R R
Onychonema laeve laevevar. Nordstedt F
Onychonema sp. E R R E E
Penium sp. R R
Pleurotaenium coronatum coronatumvar. (Brébisson) Rabenhorst E E E E
Pleurotaenium coronatum fluctuatumvar. W. West R E R
Pleurotaenium tenuissimum (Grönblad & Croasdale) Förster F C C F C
Spondylosium desmidiiforme (Borge) W. & G. S. West R
Spondylosium sp. R E E R E
Staurastrum asteroideum W. & G. S. West R
Staurastrum boergeseniicf. Raciborski R E E
Staurastrum brachiatum brachiatumvar. Ralfs F F F F E
Staurastrum dickiei Ralfs E
Staurastrum elegantissimum brasiliensevar. Förster E E R E F
Staurastrum guentheri Thomasson R E E
Staurastrum hystrixcf. Ralfs R E
Staurastrum iotanumcf. Wolle E
Staurastrum longipescf. Teiling (Nordstedt) Teiling E R E
Staurastrum leptacanthum Nordestedt E
Staurastrum leptacanthum Borgei Förstervar. E
Staurastrum leptocladum Nordstedt E
Staurastrum leptocladum cornutumvar. Wille E F E E F
Staurastrum manfeldtii Delponte E R E
Staurastrum megachanthumcf. Lundell R
Staurastrum pseudosebaldi Wille R
Staurastrum pseudotetracerum (Nordstedt) W. & G. S. West R
Staurastrum quadrangulare longispinumvar. Börgesen E E E R R
Staurastrum quadrangulare prolificumvar. Croasdale E E E E E
Staurastrum quadrinotatum Grönblad F E E R E
Staurastrum radians W. & G. S. West E E
Staurastrum sebaldi Reinsch E F E E E
Staurastrum setigerum Cleve R R E
Staurastrum setigerum . occidentallevar W. & G.S. West R E R R Econtinua >
> continuação
Desmídias com ocorrência planctônica
103

Táxons/estações ET1 ET5 ET7 ET10 EN12
Staurastrum setigerum pectinatumvar. W. & G. S. West R
Staurastrum stelliferumcf. Borge F F E E E
Staurastrum tentaculiferum Borge E F E E R
Staurastrum tetracerum (Kützing) Ralfs R
Staurastrum trifidum inflexumvar. W. & G. S. West R R
Staurodesmus clepsydra obtususvar. (Nordstedt) Teiling R
Staurodesmus convergens (Ehrenberg) Teiling R
Staurodesmus curvatus (Turner) Thomasson E R
Staurodesmus cuspidatus . groenbladiivar Foster E R R E E
Staurodesmus dejectus (Brébisson) Teiling E E
Staurodesmus glaber flexispinumvar. (Förster & Eckert) Teiling R R
Staurodesmus hirundinella (Krieger) Teiling R R R
Staurodesmus lobatus (Börgesen) Bourrelly R R
Staurodesmus mamillatus (Nordstedt) Teiling R R
Staurodesmus spencerianus (Maskell) Teiling R
Staurodesmus subulatus (Kützing) Thomasson E
Staurodesmus triangularis (Lagerhein) Teiling E F F F E
Staurodesmus validus (West) Thomasson E R E E
Triploceras gracile bidentatumvar. Nordstedt R
Xanthidium canadense (Joshua) Forster R
Xanthidium fragile Borge E E E R E
Xanthidium mamillosum (Grönblad) Föster R R E E
Xanthidium sexangulare (Grönblad) Förster R E
Xanthidium siolii Grönblad & Croasdale E R
Xanthidium trilobum Nordstedt R R R
> continuação
104
Melo et al.
Analisando as desmídias das cinco estaçõesamostradas, uma maior riqueza específica foidocumentada para a estação do rio Negro (EN12), e omenor valor foi obtido no ponto de amostragem maispróximo ao canal que conecta o lago Tupé ao rio Negro(ET10), o que pode ser evidenciado na figura 2. Elevadariqueza de espécies em desmídias tem sido associada,entre outros fatores, à presença de macrófitas aquáticas(Brook, 1981). Embora as macrófitas aquáticas sejamraras ou mesmo ausentes, na maior parte do tempo nolago Tupé e no rio Negro, a floresta de igapó pode assumirfunção similar as das macrófitas para as desmídias, fatoque necessita estudos mais específicos.
Temporalmente, foi constatada uma ligeira tendên-cia ao padrão de flutuação sazonal previamente apresen-
tado em outras publicações para aAmazônia que documentaram maiorriqueza de espécie de desmídias nosperíodos de seca (águas baixas) eenchente e menor nos períodos decheia (águas altas) e vazante (Ibañez1997, 1998; Sophia Huszar, 1996;Huszar, 2000). Este fato foi maisevidente na estação EN12 (Fig. 3),onde foram registrados 21 táxons emdezembro (seca) e ausência dedesmídias, no mês de julho 2002(cheia). Tal padrão tem sido associado,entre outros, ao fato de a menorprofundidade da coluna d'água
&

Desmídias com ocorrência planctônica
105
favorecer a circulação e, conseqüente-mente, a permanência e desenvolvi-mento das desmídias no plâncton(Huszar, 1994). Em relação ao lagoTupé isto, no entanto, deve ser melhoravaliado, visto que resultadospreliminares tem apontado para umamarcada estratificação térmica noperíodo de seca (Previattelli ,2005, neste volume), o que indica umanão circulação contínua da massad'água.
Observando os resultados do índicede constância constatou-se que amaioria dos táxons foram raros ouesporádicos, apenas onze destacaram-se por serem freqüentes ou constantesem, pelo menos, uma estação (Tab. 1).E n t r e e s t e s ,
(Grönblad Croasdale)Förster, espécie constante em trêsestações e freqüente nas outras duas,
var.Ralfs, freqüente em
quatro estações e(Ralfs) Cleve e
(Lagerhein) Teiling
et al.
P l e u r o t a e n i u mtenuissimum
Staurastrum brachiatumbrachiatum
Bambusina borreriStaurodesmus
triangularis
&
freqüentes em três estações, foram as espécies quepredominaram nos ambientes estudados.
Em síntese, o presente estudo soma evidências sobrea importância das desmídias para ambientes amazônicose amplia o conhecimento deste grupo de algas para os
Figura 2. Número total de táxons dedesmídias registrados por estação decoleta na RDS Tupé, no período de junho de2002 a outubro de 2003
0
10
20
30
40
50
60
70
80
90
ET1 ET5 ET7 ET10 EN12
Estações de coleta
N°
detá
xons
ET1
0
5
10
15
20
25
M A M J J A S O N D J F M A M J J A S O
ET5
0
5
10
15
20
25
M A M J J A S O N D J F M A M J J A S O
ET7
0
5
10
15
20
25
M A M J J A S O N D J F M A M J J A S O
ET10
0
5
10
15
20
25
M A M J J A S O N D J F M A M J J A S O
EN12
0
5
10
15
20
25
M A M J J A S O N D J F M A M J J A S O
Meses de coleta
Figura 3. Número total de táxons de desmídias registradosmensalmente em cada estação de amostragem na RDS Tupé, noperíodo de junho de 2002 a outubro de 2003
N°
detá
xon s
N°
detá
xon s
N°
detá
xon s
N°
detá
xon s
N°
detá
xon s

106
Melo et al.
Santos-Silva, E. N.; Aprile, F. M.;Scudeller, V. V. Melo, S.(Orgs.).Biotupé: meio físico, diversidadebiológica e sócio-cultural do baixorio Negro, Amazônia central. INPA,Manaus-AM.
Dikie, G. 1881. Notes on algae from theAmazonas and its tributaries.
(108):123-132Coesel, P. F. M. 1996. Biogeography of
desmids. , 336:41-53.
Föster, K, 1963. Desmidiaceen ausBrasilien II. Nord- Brasilies.
., Nova ser., 7(1): 38-92.Föster, K, 1964. Desmidiaceen aus
Brasilien II. Bahya, Goyas, Piauhyund Nord- Brasilies . ,23(3/4): 321-505.
Föster, K. 1969. AmazonischeDesmidieen. 1. Areal Santarém.
, 2: 5-232.Fös ter, K .1974. Amazon i sche
Desmidieen. 2. Areal Maués-nAbacaxis. , 5(2): 135-242.
Föster, K. 1982.
, Band XVI,Teil 8, Halfte 1, 1-593. E.S c h e i z e r b o r t ` s c h eVerlagsbuchhandlung, Stuttgart.543pp.
G r ö n b l a d , R . 1 9 4 5 . D e a l g i sb r a s i l i e n s i b u s p r a e c i p u edesmidiales, in regione inferiores,fluminis Amazonas e professoreAugust Ginsberger (Wien) annoMCMXVII collectis.
. 2(6):1-43.Huszar, V.L.M. 1994.
&
J.Limn. Soc., 18
Hydrobiologia
Revuealgol
Hydrobiologia
Amazoniana
Amazoniana
Das Phytoplankton desSudanessers, Conjugatophyceae,Zygnematales und Desmidiales(excl. Zygnemataceae)
Acta Soc. Sci.Fenn. Sér.B
Fitoplâncton deum lago amazônico impactado porrejeito de bauxita (lago Batata,Pará, Brasil): estrutura decomunidade, flutuações espaciais e
ambientes de águas pretas. Além disso, evidencia aimportância em termos de riqueza de espécies dosgêneros , e , alémde ser um dos únicos trabalhos a analisar a composiçãotaxonômica de desmídias a partir de amostras mensaisde águas da região limnética de ambientes amazônicosdurante mais de um ano de estudo. No entanto, éressaltado o fato de os resultados apresentados nomomento serem provenientes de coletas com rede deplâncton, o que além de selecionar pode concentrar ummaior ou menor volume em função do coletor. Análisedas amostras quantitativas tornam-se necessárias paraum melhor entendimento do padrão de flutuaçãotemporal e espacial das desmídias na RDS Tupé.
Agradecemos aos colegas do laboratório de plânctonda CPBA/INPA e do laboratório de Ficologia do MuseuNacional-UFRJ, que nos disponibiliza-ram bibliografia eequipamentos para análise do material coletado. Estetrabalho teve o apoio financeiro do através doProjeto BioTupe (Proc. 551361/001-8), bolsa de RD(Proc. 301349/01-0) e de IC (PIBIC/CNPq/ INPA),(ProDoc-02) e (PPI 1-3540 e PIBIC/INPA).
Bittencourt-Oliveira, M.C. 1993. Ficoflórula doreservatório de Balbina, Estado do Amazonas, III.Zygnemaphyceae. . 53(3): 477-488.
Brook, A. J. 1981. Oxford,Black. Sci. Publ. (Botanical Monographs, V. 16).
Dajoz, R. 1978. . 3° Ed. Editora VozesLTDA. Petrópolis-RJ. 472pp.
Darwich, A. J.; Aprile, F. M.; Robertson, B. A. 2005.
Staurastrum Staurodesmus Closterium
Revta bras. Biol ,The Biology of Desmids.
Ecologia geral
Variáveis limnológicas: Contribuição ao estudoespaço-temporal de águas pretas amazônicas. In:
Agradecimentos
Bibliografia Citada
CNPq
CAPESINPA-MCT
276pp.

Desmídias com ocorrência planctônica
107
temporais.
Fitoplâncton. In:
Hydrobiologia, .A treatise on
Limnology. Introduction to lakebiology and the limnoplankton
Verh. Internat.Verein Limnol .
Hydrobiologia
Spec.Publ. Can. J. Fish. Aquat. Sci ,
Limnología
Tese. UniversidadeFederal de São Carlos, PPG-ERN,São Carlos, SP. 219pp
Huszar, V.L.M. 2000.L a g o B a t a t a : I m p a c t o eRecuperação de um EcossistemaAmazônico. Ed:Bozelli, R.L.,Esteves, F. A. & Roland, F. In:Bozelli, R. L., Esteves, F. A.Roland, F. (Eds.). Lago Batata:Impacto e recuperação de umecossistema amazônico. p.91-104.
Huszar, V.L.M.; Reynolds, C.S. 1997.Phytoplankton periodicity andsequences of dominance in anAmazonian flood-plain lake (LagoBatata, Pará, Brasil): responses togradual environmental change.
346:169-181Huntchinson, G.E. 1967
.John Willey Sons, New York, v.2.1115pp.
Ibañez, M. S. R. 1997. Phytoplanktonbiomass of a central Amazonianfloodplain lake.
.,26:605 - 609Ibañez, M.S.R. 1998. Phytoplankton
composition and abundance of acentral Amazonian floodplain lake.
, 362:78-83.Junk, W.P.; Bayley, P.B.; Sparks,
R.E;1989. The flood pulse conceptin river floodplain system.
. 106:110-127.
Lopez, M. R.; Bicudo, C.E.M. 2003.Desmidioflórula de um lago deplanície de inundação do Rio Acre,Estado do Amazonas, Brasil. ActaAmazonica, 33(2): 167-211.
Margalef, R. 1983 . Omega,Barcelona.
Martins, D.V. 1980. Contribuição àf ico log ia da Amazônia. 2.
&
&
1010pp.
Desmidiofórula dos lagos Cristalino e São Sebastião,Estado do Amazonas: gêneros filamentosos.
10 (4): 725-741.Martins, D.V. 1982. Contribuição à ficologia daAmazônia.
2. Desmidiofórula dos lagos Cristalino e SãoSebastião, Estado do Amazonas: gêneros
12(2):279-290.
Martins, D.V. 1986a. Meyen (Desmidiaceae)dos lagos Cristalino e São Sebastião, Estado doAmazonas, Brasil. : 15-32.
Martins, D.V. 1986b. Eherenberg,Agardh e Bailey
(Desmidiaceae) dos lagos Cristalino e São Sebastião,Estado do Amazonas, Brasil.: 43-55.
Melo, S.; Huszar, V.L.M. 2000.Phytoplankton in anAmazonian flood-plain lake (Lago Batata, Brasil): dielvariation and species strategies.
(1):63-76.Melo, S.; Sophia, M.G.; Menezes M.; Souza, C. A. 2004.
In: Borges, S.H.; Iwanaga, S.; Durigan, C. C. Pinheiro, M. R.(Eds).Janelas para a biodiversidade do Parque Nacional doJaú: uma estratégia para o estudo da biodiversidadedaAmazônia. Ed. Ipiranga, Brasília-DF.
Melo, S.; Rebelo, S. R. M.; Souza, F. S.; Menezes, M.;Torgan, L. C. 2005. . In: Santos-Silva, E.N.; Aprile, F. M.; Scudeller, V. V. Melo, S.(Orgs.).Biotupé: Meio Físico, Diversidade Biológica eSociocultural no Baixo Rio Negro, Amazônia Central.Editora INPA, Manaus-AM.
Moss, B. 1988.Blackwell Science Publ 276 pp.
Previattelli, D.; Santos-Silva, E. N.; Darwich, A. J. 2005.
Santos-Silva, E. N.;Aprile, F. M.; Scudeller, V. V. Melo, S.(Orgs.).Biotupé: Meio Físico, Diversidade Biológica eSóciocultu ral do Baixo Rio Negro, Amazônia Central.Editora INPA, Manaus-AM.
Rai, H. Hill, G. 1981. Physical and chemical studies ofLago Tupé, a central Amazonian blackwater “ RiaLake”. , 66(1): 37-82.
ActaAmazonica,
Netrium,Closterium, Pleurotaenium. Acta Amazonica,
Staurastrum
Universitas, Ciências, 36Gêneros Euastrum
Micrasterias Triploceras
Universitas, Ciências, 38
J. Plank. Research,22
Biodiversidade de algas planctônicas do ParqueNacional do Jaú: Janela Seringalzinho.
Fitoplâncton
Ecology of freshwaters: man e medium., .
Distribuição vertical do zooplâncton e sua relaçãocom as variáveis ambientais. In:
-
Int. Revue ges. Hidrobiol. Hydrogr.
&
&
&
&
P.83-95.
Oxford

108
Melo et al.
Reynolds,C. S. 1984.Cambridge University Press,
Cambridge. 384pp.Scott, A. M., Grönblard, R., Croasdale, H. & Sioli, H.
1965. Desmids from the Amazon basin, Brasil.69: 1-94.
Sioli, H. 1950. Das Wasser im Amazonasgebiet.26:274-280.
Sophia, M. G. & Huzsar. V. L. M. 1996. Planktonicdesmids of three Amazonian systems ( Lake Batata,Mussurá and Trombetas River), Pará, Brazil.
XIV (1 /2):75-90.Thomasson, K.1971. Amazonian algae. Institut Royal des
Sciences Naturelles de Belgique, 86:1-57.Thomasson, K.1977. Two conspicuous desmids from
Amazonas. , 130: 41-51.Uherkóvich, G. 1984. Phytoplankton. In: Sioli, H. (Ed).
Dr. JunkPublishers.p.295-310
The ecology of freshwaterphytoplankton.
Actabot. Fenn,
Fortschr,
Amazoniana,
Bot. Notser
The Amazon: Limnology and landscape ecology of amight tropical river and its basin.
.

Diversidade BiológicaCapítulo 7
Distribuição vertical do zooplâncton esua relação com as variáveisambientais
RESUMO - A flutuação vertical da densidade zooplanctônica na coluna d'água foi estudadadurante um ciclo nictemeral, no período de seca (novembro de 2001), no Lago Tupé. Dosgrupos considerados, somente os adultos e os copepoditos de(Copepoda, Calanoida), (Cladocera) e(Cladocera) apresentaram um padrão de migração vertical noturna. Os outros grupos nãoapresentaram padrão de migração vertical, tendo no entanto, apresentado distribuiçãoheterogênea, com densidade maior de indivíduos nas camadas mais profundas durantetodo o período. A distribuição dos organismos, de uma maneira geral, está de acordo com ahipótese da mudança na luminosidade solar induzir a migração. Porém há organismos quepossuem um comportamento diferente do esperado e que provavelmente estão sob ainfluência de outros fatores como a distribuição do alimento e a estratificação térmica e de
Aspinus acicularis
Bosmina longirostris Bosminopsis deitersi
Daniel PREVIATTELLI1
Edinaldo Nelson dos SANTOS-SILVA
Assad José DARWICH
2
2&
1
2
Mestrado em Biologia de Água Doce e Pesca Interior do Programa Integrado de Pós-Graduação emBiologia Tropical e Recuros Naturais INPA/UFAM
Bolsista CNPqe-mail: [email protected]
e-mail: [email protected]
Coodenação de Pesquisas em Biologia Aquática, Instituto Nacional de Pesquisas da Amazônia.e-mail: [email protected]
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

oxigênio, que são discutidos.
PALAVRAS-CHAVE: migração vertical nictemeral, zooplâncton, águas pretas, AmazôniaCentral.
110
Previattelli et al.
Introdução
A heterogeneidade na distribuição vertical, e emespecial a migração, é um fenômeno observado emdiferentes organismos zooplanctônicos. SegundoHutchinson (1967), os primeiros registros dessefenômeno foram feitos por Weissmann (1877), Forel(1877), e posteriormente por Pavesi (1882) e Francé(1894). Trabalhos como os de Clarke (1934), Wells(1960), Ferrato (1967) e Green (1967) também trazemdescrições sobre a distribuição heterogênea depopulações zooplanctônicas, incluindo movimentosativos diuturnos na coluna d'água (migração verticalnictemeral), com destaque às possíveis implicaçõesecológicas relacionadas à predação por peixes eciclagem de nutrientes entre diferentes estratos dacoluna d'água.
São reconhecidos diferentes tipos de migraçãovertical nictemeral. A classificação mais usual leva emconta o horário do dia em que há deslocamento dosindivíduos de uma profundidade para outra na coluna d'água: quando há o deslocamento para camadas maissuperficiais e oxigenadas da água durante a noite, dá-seo nome de , que por ser a maisfreqüente é também chamada de ;quando este deslocamento ocorre durante o dia, dá-se onome de (Lampert Sommer, 1997).
A ocorrência de migração vertical é mais facilmenteobservada em lagos estratificados, onde existemdiferenças marcantes entre o epilímnio e o hipolímnio(Margalef, 1983; Wetzel, 1983), sendo que a migraçãonoturna tem uma relação direta com a profundidade,transparência e produtividade do lago (Hutchinson,1967). A migração vertical é também diretamenteassociada à respostas fisiológicas desencadeadas porestímulos ambientais como luz e temperatura. Ashipóteses elaboradas para explicar a migraçãorelacionam esse comportamento à vantagensmetabólicas, em especial para organismos filtradores(Lampert Sommer, 1997; Lampert ., 1988), ou
migração noturnamigração normal
migração reversa
et al
&
&
como uma forma de evitar predadoresque se guiam pela visão (ZaretKerfoot, 1975; Zaret Stuffern, 1976;Arcifa-Zago, 1978), ambas conferindovantagem adaptativa.
Estudos recentes procuraramcompreender a ação desses fatores emconjunto. Dodson (1990) e HanStraškraba (2001) elaboraram modelosmatemáticos que prevêem seucomportamento em função dedeterminadas variáveis ambientais(luz, temperatura, etc.) e biológicas(presença de predadores, movimentovertical de algas, etc.). Autores comoLampert (1989) e Dini Carpenter(1991), realizaram experimentos paraverificar as hipóteses citadas anterior-mente. Gaudy Verriopoulos (2004)usaram esses comportamentos paradescrever as espécies. Hays (2003) fezuma revisão a respeito do significadoadaptativo do deslocamento verticaldo zooplâncton e das conseqüênciasdeste fenômeno para o ecossistema.Seu trabalho aponta grandes diferen-ças no conhecimento em diferentesregiões do globo e faz duas colocaçõesimportantes: que a razão verdadeirapara a migração é a fuga de predadorese, devido aos recentes avanços tecno-lógicos, agora podemos investigar commaior precisão padrões específicos demigração de determinadas espécies,especialmente dos vertebrados.
Mas apesar desse nível elevado deconhecimento, a desigualdade entreas regiões faz com que nas áreas ondehá poucos estudos, trabalhosdescritivos sejam necessários. Oregistro da ocorrência desse fenômeno
&&
&
&
&

Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais
no Brasil até o momento está restrito àpoucos ambientes (Arcifa-Zago, 1978;Nogue i ra Panare l l i , 1997;Perticarrari ., 2003; Perticarrari
., 2004). Na região amazônica emespecial, Fisher . (1983)descreveram o fenômeno em águasbrancas. Brandorff (1978a) fez umestudo com o objetivo de comparar ozooplâncton em águas brancas e águaspretas. Hardy (1980) também fez essacomparação entre os lagos Cristalino eTarumã-Mirim (água preta) e os lagosRedondo, Jacaretinga e Castanho(água branca). Portanto, poucostrabalhos enfocaram a distribuiçãovertical do zooplâncton em águaspretas.
Este estudo teve como objetivoverificar a distribuição vertical dozooplâncton, a ocorrência de migraçãovertical em lagos de águas pretas naAmazônia e apontar algumas de suasimplicações para a dinâmica dessesecossistemas, bem como discutir ainterferência da migração na metodo-logia de amostragem usada nos estu-dos sobre zooplâncton na região.
Por possuir diferentes graustróficos, em função do pulso deinundação, o lago Tupé é caracterizadocomo um lago distrófico, oligotrófico,momentaneamente meso-oligotrófi-co, chegando a ser, durante o períodode águas baixas (seca), meso-eutrófico(Rai Hill, 1982). Durante a seca, háum aumento da concentração denutrientes, especialmente fósforo,devido ao menor volume de água nosistema, alterando assim seu estadotrófico (Rai Hill, 1980; 1981a;1981b). Portanto, neste trabalho,inferências são feitas a respeito dadistribuição vertical do zooplânctondurante o período de águas baixas.Futuros estudos deverão ser realizados
&
&
&
et alet al
et al
para verificar possíveis diferenças entre os outrosperíodos do ciclo hidrológico.
O período de seca de 2001 (novembro), na estaçãoET10 (fig. 1b, Aprile ., 2005, neste volume), foicoletado um volume de 36l de água, em intervalos de 3horas, a cada metro de profundidade, da sub-superfícieaté o fundo do lago (4,5m), utilizando um coletorSchindler/Patalas de 12l. As amostras foram filtradasatravés de uma rede com abertura de malha de 55 m efixadas em formol neutralizado com tetraborato desódio, com uma concentração final de 6%. Os indivíduosforam identificados com o auxílio de literaturaespecializada (Goulden, 1968; Koste, 1978; Negrea,1983; Reid, 1985; Korovchinsky, 1992; Dussart Defaye,1995; Nogrady ., 1995; Segers, 1995; Elmoor-Loureiro, 1997) e contados utilizando-se microscópicoestereoscópico. Foram considerados os seguintesgrupos: Cladocera, Copepoda e Rotifera, assumindo quecada subgrupo possui um comportamento diferente e,conseqüentemente, uma distribuição diferente nacoluna d'água.
Simultaneamente foram medidos , atemperatura, o oxigênio dissolvido, o potencialhidrogeniônico e a condutividade elétrica da água, comodescrito no capítulo de Darwich . (2005).
Os parâmetros físicos e químicos revelaram umaestratificação térmica e de oxigênio dissolvido, com ummetalímnio bastante marcado entre 2,5 e 3,0m (Figs. 1 e2).
Os valores de oxigênio dissolvido e condutividade(Fig. 3) mostraram diferenças entre as profundidades. Ooxigênio dissolvido mostra números semelhantes entre asub-superfície e 2m (zona rica em oxigênio), declínioacentuado entre 2 e 3 m (oxiclina) e entre 3 metros deprofundidade e o fundo do lago uma nova semelhança devalores próximos a zero (zona anóxica). Essaconfiguração assemelha-se com a de um lagomeromítico, como observado por Darwich . (2005,
Materiais e Métodos
Resultados e Discussão
et al
et al
in situ
et al
et al
�
&
111

112
neste volume). Ao longo do ciclonictemeral estudado, flutuações maisacentuadas aparecem entre oshorários diurnos e nas camadassuperficiais da coluna d'água (até 2m).
A concentração hidrogeniônicamostrou pequena alteração entre oshorários e entre as profundidades (Fig.4). No entanto, há uma discretaalteração a 2m de profundidade,diminuindo de 5,0 para 4,5 entre 1 e 4metros, com pico de alteração a 2,5m.
A distribuição das populações nacoluna d'água mostrou-se heterogê-nea, porém, foi observado um padrãogeral. Durante o dia os indivíduostendem a se concentrar nas regiõesmais profundas (abaixo de 2m), edurante a noite distribuem-se maishomogeneamente em toda a colunad'água. Esse comportamento seassemelha com a descrição damigração conhecida como ounormal (Hutchinson, 1967). Essecomportamento é mais claramenteconstatado entre os adultos ecopepoditos de(Copepoda, Calanoida), apresentadonas figuras 5 e 6 e(Cladocera) sendo menos notável em
(Cladocera),como apresentado nas figuras 11 e 12.O fato destas três espécies seremfiltradoras ajuda a explicar o seudeslocamento, que pode estarrelacionado com a otimizaçãometabólica citada por LampertSommer (1997).
A distribuição detambém apresentou uma
distribuição heterogênea, com umnúmero maior de indivíduos entre trêse quatro metros, durante todos oshorários (Fig. 13). Náuplios deCopepoda (Fig. 7) e os Rotiferaencontrados aparentemente não
noturna
Aspinus acicularis
Bosmina longirostris
Bosminopsis deitersi
Holopediumamazonicum
&
Figura 1. Variação vertical nictemeral da saturação do Oxigênio(%), representada por isopletas a intervalos de 20%, emnovembro de 2001.
Figura 2. Variação vertical nictemeral da Temperatura (0 C),representada por isopletas a intervalos de 1 C , em novembrode2001.
o
o
Previattelli et al.

apresentam migração, apesar deterem apresentado uma distribuiçãovertical heterogênea (Fig. 14).
Lass . (2000) realizaramexperimentos com “plankton tower”para verificar a capacidade doscladóceros de ficar em ambienteanóxico, encontrando respostasdiferentes no padrão de migração emdiferentes níveis de concentração deoxigênio no hipolímnio, o que podeexplicar a presença de uma migraçãodiferente encontrada em ambientecom pouco oxigênio.
Han Straškraba (2001) afirmamque o principal fator que induz amigração é a mudança relativa naintensidade luminosa. Mas a foto-resposta é modificada por outrasvariáveis ambientais: pressão depredação, alimento e a própriatemperatura.
Winder . (2003) tambémdiscutiram a relação das condições dealimentação e as grandes profundidades. Kessler (2004) trabalhou com aotimização do “fitness” balanceandoalimento e temperatura em ,encontrando um menor gasto deenergia nos indivíduos em regiões maisfrias da coluna d'água em relação aosque permaneciam em temperaturamais alta.
O fato de não haver mistura dascamadas mais profundas também podeinfluenciar na distribuição, uma vezque se não há mistura de água, não hádeslocamento de matéria ou energiapara os extratos mais superficiais. Issoinfluencia na distribuição do alimentona coluna d'água, fazendo com que boaparte da matéria fique em extratosmais profundos e, conseqüentemente,aqueles organismos que utilizam essematerial. Espécies como
tem um comportamento queparece seguir essa distribuição.
et al
et al
-
Daphnia
H. amazoni-cum
&
Figura 3. Variação vertical nictemeral da Condutividade( S.cm-1), representada por isopletas a intervalos 1 S.cm ,em novembro de 2001.� �
-1
Figura 4. Variação vertical nictemeral do PotencialHidrogeniônico, representada por isopletas a intervalos de 0,1,em novembro de 2001.
113
Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais

114
Figura 5. Distribuição vertical nictemeral de adultos de, representada por isopletas, em novembro de 2001.
Aspinusacicularis
Figura 6. Distribuição vertical nictemeral de copepoditos de, representada por isopletas, em novembro
de 2001.Aspinus acicularis
Previattelli et al.
A. acicularis
A. acicularis

Figura 7. Distribuição vertical nictemeral de nauplius de, representada por isopletas, em novembro
de 2001.Aspinus acicularis
Figura 8. Distribuição vertical nictemeral de adultosCyclopoida, representada por isopletas, em novembro de2001.
115
Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais
A. acicularis
Cyclopoida

116
Figura 9. Distribuição vertical nictemeral de copepoditosCyclopoida, representada por isopletas, em novembro de 2001.
Figura 10. Distribuição vertical nictemeral de nauplius deCyclopoida, representada por isopletas, em novembro de 2001.
Previattelli et al.
horário (h)
horário (h)
Cyclopoida
Cyclopoida

Figura 11. Distribuição vertical nictemeral derepresentada por isopletas, em novembro de
2001.
Bosminalongirostris,
Figura 12. Distribuição vertical nictemeral de, representada por isopletas, em novembro de 2001.
Bosminopsisdeitersi
117
Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais
B. deitersi
B. longirostris
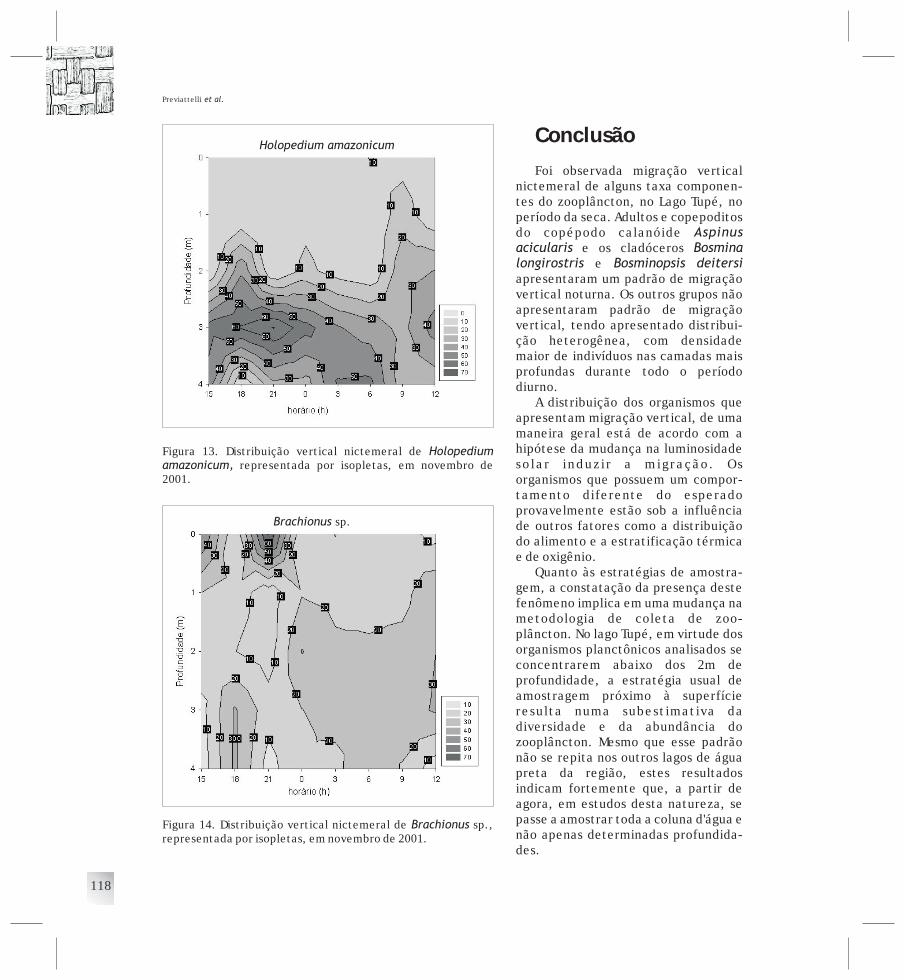
118
Figura 13. Distribuição vertical nictemeral derepresentada por isopletas, em novembro de
2001.
Holopediumamazonicum,
Figura 14. Distribuição vertical nictemeral de sp.,representada por isopletas, em novembro de 2001.
Brachionus
Previattelli et al.
Conclusão
Foi observada migração verticalnictemeral de alguns taxa componen-tes do zooplâncton, no Lago Tupé, noperíodo da seca.Adultos e copepoditosdo copépodo calanóide
e os cladócerose
apresentaram um padrão de migraçãovertical noturna. Os outros grupos nãoapresentaram padrão de migraçãovertical, tendo apresentado distribui-ção heterogênea, com densidademaior de indivíduos nas camadas maisprofundas durante todo o períododiurno.
A distribuição dos organismos queapresentam migração vertical, de umamaneira geral está de acordo com ahipótese da mudança na luminosidadesolar induzir a migração. Osorganismos que possuem um compor-tamento diferente do esperadoprovavelmente estão sob a influênciade outros fatores como a distribuiçãodo alimento e a estratificação térmicae de oxigênio.
Quanto às estratégias de amostra-gem, a constatação da presença destefenômeno implica em uma mudança nametodologia de coleta de zoo-plâncton. No lago Tupé, em virtude dosorganismos planctônicos analisados seconcentrarem abaixo dos 2m deprofundidade, a estratégia usual deamostragem próximo à superfícieresulta numa subestimativa dadiversidade e da abundância dozooplâncton. Mesmo que esse padrãonão se repita nos outros lagos de águapreta da região, estes resultadosindicam fortemente que, a partir deagora, em estudos desta natureza, sepasse a amostrar toda a coluna d'água enão apenas determinadas profundida-des.
Aspinusacicularis Bosminalongirostris Bosminopsis deitersi
Holopedium amazonicum
Brachionus sp.

Academic Publishing, The Hague, 7: 277pp.Elmoor-Loureiro, L. M. A. 1997. Manual de identificação
de cladóceros límnicos do Brasil. .156pp.
Fisher, T. R., Melack, J. M., Robertson, B. A., Hardy, E. R.Alves, L. F. 1983. Vertical distribution of
zooplankton and physico-chemical conditions duringa 24-hour period in a amazon floodplain lake LagoCalado, Brasil. , 13 (3-4): 475-487.
Forel, E. A. 1877. Faune profonde du Léman xxxii. Faunepélagique. , 14: 210-223.
Francé, R. H. 1894. Zur Biologie des Planktons.., 14: 33-38.
Gaudy, R. Verriopoulos, G. 2004. Spatial and seasonalvariations in size, body volume and body proportion(prosome:urosome ratio) of the copepod
in a semi-closed ecosystem (Berre lagoon,western Mediterranean). , 513: 219-229.
Goulden, C. E. 1968. The sistematic and evolution of theMoinidae. , v.58,part 6, 101pp.
Green, J. 1967. The distribution and variation of(Crustacea: Cladocera) in relation to fish
predation in Lake Albert, East Africa. J. Zool., 151:181-189.
Han, B. P. Straškraba, M. 2001. Control mechanisms ofdiel vertical migration: theoretical assumptions. J.Theor. Biol., 210: 305-318.
Hardy, E. 1980. Composição do zooplâncton em cincolagos daAmazônia. 10 (03): 577-609.
Hays, G. C. 2003. A review of adaptative significance andecosystem consequences of zooplankton diel verticalmigrations. , 503: 163-170.
Hutchinson, G. E. 1967. A treatise on limnology. NewYork, John Wiley, v. 2, Introduction to Lake Biologyand the Limnoplankton, John Wiley Sons,New York.1115pp.
Kessler, K. Lampert, W. 2004. Fitness optimization ofin a trade-off between food and
Universa Brasília,
ActaAmazonica
Bull. Soc. Vaud. Sci. Nat.Biol.
Zbl
Acartiatonsa
Hydrobiologia
The American Philosophical Society
Daphnialumholtzi
ActaAmazônica,
Hydrobiologia
Daphnia
Ferrato, A. M. 1967. Notas preliminares sobremigraciones del zooplancton en cuencas isleñas delParana Medio. , 23: 173-188.Acta Zool. Lilloana
&
&
&
&
&
Bibliografia Citada
Arcifa-Zago, M. S. 1978. VerticalMigration ofHerbst, 1967, in AmericanaReservoir, State of São Paulo,Brazil. .Limnol., 20: 1720-1726.
Brandorff, G. O. 1978. Preliminarycomparison of the crustaceanplankton of a wite water and blackwater lake in Central Amazonia.Verh. ., 20:1198-1202.
Clarke, G. 1934. Factors Affecting thevertical distribution of copepods.
, 4 (4): 530-540.
Darwich, A. J.; Aprile, F. M.;Robertson, B. A. Alves, L. F. 2005.Limnologia do lago Tupé: dinâmicaespaço-temporal do oxigêniodissolvido. In: Santos-Silva, E. N.;Aprile, F. M.; Schudeller, V. V.Melo, S. (Eds.).
. Ed. Inpa,AM.
Dini, M. L. & Carpenter, S. R. 1991. Theef fec t o f who le - l ake f i shcommunity manipulations on
migratory behavior., 36, (2): 370-
377.
Dussart, B. H. Defaye, D. 1995.Copepoda: Introduction to theC o p e p o d a . S P B
III Guides to theidentification of the microinvertebrates of continetal waters of theworld editor: H. J. F. Dumont,
Daphnia gessneri
Verh. Internat. Verein
Internat. Verein. Limnol
Ecological Monographs
BioTupé: meiofísico, diversidade biológica esociocultural do baixo rio Negro,Amazônia central
DaphniaLimnol. Oceanogr.
A c a d e m i cPublishing - -
-
SPB
&
&
&
Dodson, S. 1990. Predicting dielvertical migration of zooplankton.
, 35: 1195-1200.Limnol. Oceanogr.
119
Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais

120
temperature. , 140: 381-387.Korovchinsky, N. M. 1992. Sididae Holopediidae
(Crustacea: Daphniiformes).I I I Guides to the identif ication of themicroinvertebrates of continetal waters of the world.editor: H. J. F. Dumont,
n.3, 82p .Koste, W. 1978. Rotatoria Die Rädertiere Mitteleuropas
Ein Bestimmungswerk, begründet von Max VoigtÜberordnung Monogononta. II. Tafelband,
Gebrüder 234pp.Lampert, W.; Schwitt, R. D.; Ruick, P. 1988.
. Bull.Mar. Sci. 43(3): 620-640.
Lampert, W. 1989. The adaptive significance of dielvertical migration of zooplankton.
, 3: 21-27.Lampert, W. Sommer, U. 1997.
. Oxford UniversityPress. New York, Oxford 382pp.
Lass, S., Boersma, M. & Spaak, P. 2000. How do migratingdaphnids cope with fish predation risk in theepilimnion under anoxic conditions in thehypolimnion? 22 (7):1411-1418.
Margalef, R. 1983. Limnología. Edit. Omega. Barcelona.1010pp.
Negrea, S. 1983. Fauna Republicii Socialiste RomâniaCrustacea Cladocera, v.IV, fasc. 12.
, Bucareste.380pp.
Nogrady, T., Pourriot, R. Segers, H. 1995. Rotifera, v. 3:The Notomatidae and: The Scaridiidae.
Guides to the identification of themicroinvertebrates of continetal waters of the worldeditor: H. J. F. Dumont,
n.8, 248p .Nogueira, M. G. Panarelli, E. 1997. Estudo da migração
vertical das populações zooplanctônicas na represade Jurumirim (rio Paranapanema São Paulo, Brasil).
., 9: 55-81.Pavesi, P. 1882. Altra serie di ricerce e studi sulla fauna
pelágica dei laghi italiani., 8: 340-403.
Perticarrari,A.,Arcifa, M. S. Rodrigues, R.A. 2003. Diel
Oecologia
SPBAcademic Publishing
GebrüderBorntraeger,
Verticalimigration of freshwater zooplankton-test of somehypotheses predicting a metabolic advantage
FunctionalEcology
Limnoecology: theecology of lakes and streams
.
Journal of Plankton Research,
EdituraAcademiei Republici Socialiste România
SPB AcademicPublishing
Acta Limnol. Brasil
Att. Soc. Veneto-TorentinaSci. Nat
&
&
&
&
&
Academic Publishing, TheHague, p
Borntraeger, Berlin,
Academic Publishing, TheHague, p
vertical migration of cladocerans ina tropical lake. 11 (1): 15-25.
Perticarrari, A., Arcifa, M. S.Rodrigues, R. A. 2004. Diel verticalmigration of copepods in a Brazilianlake: a mechanism for decreasingrisk of predation?
, 64 (2):289-298.
Reid, W. J. 1985. Chave deidentificação para as espéciescontinentais sulamericanas de vidalivre da ordem Cyclopoida(Crustacea, Copepoda).
, 9: 17-143.Segers, H. 1995. Rotifera, v. 2: The
Lecanidae (Monogononta).Guides to the
identification of the microinvertebrates of continetal waters of theworld editor: H. J. F. Dumont,
n.6 226pp.Weissmann, A. 1877. Das Tierleben im
Bodensee.
Nauplius,
ChaoborusBrazilian Journal of Biology
Bolm.Zool. Univ. S. Paulo
SPBAcademic Publishing
-
Schr. Gesch. Bodensees
&
Rai, H. Hill, G. 1980. Classification ofCentral Amazon lakes on the basisof their microbiological andphysico-chemical characteristics.
, 72: 85-99.Rai, H. Hill, G. 1981a. Physical and
chemical studies of Lago Tupé; aCentral Amazonian black water“Ria Lake”.
., 66 (1): 37-82.Rai, H. Hill, G. 1981b. Bacterial
biodynamics in Lago Tupé, a CentralAmazonian black water “Ria Lake”.
., 58 (4): 420-468.Rai, H. Hill, G. 1982. Distribution of
carbon, chlorophylla and pheo-pigments in the black water lakeecossystem of Central Amazonregion. 82 (1/4):74-87.
Academic Publishing, The Hague,.
&
&
&
&
Hydrobiologia
Int. Revne ges.Hydrobiol
Arch. Hydrobiol
Arch. Hydrobiol,
Previattelli et al.

Umgebund
Limnology..
, 7: 1-31.Wells, L. 1960. Seasonal abundance
and vertical movements ofplanktonic crustacea in LakeMichigan. Fishery Bulletin of theFish and Wildlife Service, 60: 342-369.
Wetzel, R. G. 1983.Saunders Philadelphia, 743pp.
Winder, M., Boersma, M. Spaak, P&
2003. On the cost of vertical migration: are feedingconditions really worse at greater depths?
, 48: 383-393.Freshwater
BiologyZaret, T. M. Kerfoot, W. C. 1975. Fish predation on
: body size selectionvisibility selection. Ecology, 56 (1): 232-237.
Zaret, T. M. Stuffern, J. S. 1976. Vertical migration inzooplankton as a predator avoidance mechanism.
., 21, (6): 804-813.
&
&
Bosmina longirostris versus
Limnol Oceanogr
121
Distribuição vertical do zooplâncton e sua relação com as variáveis ambientais


Diversidade BiológicaCapítulo 8
As esponjas do Lago Tupé
RESUMO - Lagos de águas negras daAmazônia não haviam recebido um estudo de sua faunade esponjas. O levantamento da biodiversidade do Lago Tupé, incluiu o da espongofauna,possibilitando assim aquele estudo.Aamostragem foi realizada em duas ocasiões distintas,ambas na estação seca. Foram ainda extraídas colunas de sedimentos atuais, na porçãocentral do lago, para detecção da contribuição das espículas das esponjas aos sedimentos.As esponjas foram detectadas incrustando a floresta inundada de igapó, dos níveis deimersão mais baixos aos mais altos. A fauna de esponjas levantada destaca-se pelaabundância de espécimes, conspícuos e robustos, particularmente de(Bowerbank, 1864), (Bowerbank, 1867) e BonettoEzcurra de Drago, 1968 (Familia Metaniidae). Essas esponjas apresentam esqueletosextremamente duros, indicando um ambiente de águas altas ricas em sílica. Esponjas deesqueletos mais delicados, particularlmente do gênero (FamíliaSpongillidae) são também abundantes e utilizam substratos mais frágeis como folhas,ramos e caules finos, além da casca das árvores das margens inundadas. Também osafloramentos rochosos situados na boca do lago apresentam conspícuas incrustações deesponjas. Uma característica marcante dessa assembléia de esponjas é a extraordinária
Metania reticulata
Drulia browni Drulia uruguayensis
Trochospongilla
&
Cecilia VOLKMER-RIBEIRO
Franciely Benetti de ALMEIDA&
e-mail: [email protected]
e-mail: [email protected].
Museu de Ciências Naturais, Fundação Zoobotânica do Rio Grande do Sulrua Dr. Salvador França, 1427. 90690-000, Porto Alegre, RS.
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

124
produção estacional de gêmulas, que, nas esponjas da Familia Spongillidae, sedesenvolvem em detrimento das estruturas esqueletais. Por outro lado, o volume notávelde espécimes de esponjas da Familia Metaniidae representa acréscimos estacionais dereduzida espessura, sobre as estruturas esqueletais residuais de décadas de imersõesanteriores. Os sedimentos do lago detêm abundante conteúdo de espículas das esponjaslevantadas.
PALAVRAS CHAVE: esponjas, igapó, lagos de águas pretas, Amazônia Central, sedimentos.
apesar de haver distinguido uma faunade profundidade (no período de águabaixa) e outra litorânea ( no período deágua alta). Assim sendo os poríferos,que constituem uma fauna conspícua eabundante, justamente na faixa deinundação desse lago, haviam sido,até o presente ignoradas. Chauvel
. (1996), no entanto, em estudorealizado no igapó do Rio TarumãMirim, também contribuinte do rioNegro, próximo a Manaus, reconheceram a contribuição que as esponjasaportam, através de suas espículassilicosas, aos sedimentos desse tipo delago, e à circulação da sílica. No casoem pauta as espículas foramidentificadas como pertencentes aespécie (erroneamentegrafado como ), dadohaverem os autores observadoabundância de espécimes dessaespécie incrustando as árvores doigapó.
Batista . (2003), reportam osresultados de estudos detalhados,tanto da qualidade das águas, quantodas comunidades de esponjas dobentos rochoso e dos lagos de várzeado rio Araguaia, portanto, em ambien-tes de água clara. As esponjas conti-nentais são organismos que tem o
et
al
-
Drulia browni
Drullia braunii
et al
Wolkmer Almeida&
Introdução
Os poríferos constituem um componente importantedas comunidades de macro-invertebrados dos ambientesaquáticos daAmazônia (Volkmer-Ribeiro, 1981; Volkmer-Ribeiro Pauls, 2000) com a característica particular depertenceram à fração séssil dessa fauna. As esponjas daRegião Amazônica agrupam-se em assembléias distintasconforme os ambientes aquáticos considerados: ospermanentemente inundados, como fundos de rios e,extensão desses, os lagos de hidrelétricas ou osambientes sujeitos aos pulsos de inundação, como osigapós e lagos de várzea, particularmente da AmazoniaCentral. Nesses últimos à vida das esponjas deve-se a suacapacidade de produzirem gêmulas, corpos dereprodução assexuada, com que as diversas espéciessobrepujam os períodos de exposição aérea nos lagos devárzea e igapós ( Volkmer-Ribeiro, 1981, 1999: Batista
. 2003) Cada gêmula é um dispositivo clonador,constituído de células maternas indiferenciadas eencapsuladas em envoltórios de espongina, comrevestimento de espículas silicosas altamenteespecíficas. Essas células eclodem no período seguintede imersão e diferenciam-se novamente, constituindonovas esponjas sobre o esqueleto silicoso seco daesponja mãe ou sobre qualquer substrato onde as águasaltas ou o fluxo as levem, devido a camada pneumáticade que são geralmente dotadas. Estudos específicos daespongofauna de lagos de águas pretas da Amazôniaainda não haviam sido feitos. No próprio lago Tupé, Reiss(1977), em investigação qualitativa e quantitativa dafauna macrobêntica do lago, não menciona as esponjas,
&
et
al , .

potencial de registrarem informaçõessobre a qualidade das águas e doambiente e, após a morte, remeteremessas informações aos sedimentos,sejam atuais, sub-fósseis ou fósseis,através das espículas, todas silicosas,que ficam aí acumuladas (Harrison,1988; Harrison Warner, 1986; Turcq
. 1998 Sifeddine . 2001).Para efetivação de interpretações decaráter ambiental e paleo-ambiental,propiciadas pelos relictos dessa faunanos sedimentos, é necessário que sedisponham de conhecimentos da faunaatual e se implementem os levantamentos e pesquisas da auto-ecologiadas distintas espécies ou da ecologiade suas assembléias. Água negra naAmazônia é geralmente sinônimo deabundância de determinadas espéciesem detrimento de riquezas específicaselevadas, particularmente quando setrata da Amazônia Central, com seuspulsos de inundação, indutores deseleções marcantes. Por isto as esponjas constituem um item imprescindívelem levantamentos de biocenosesaquáticas nesses ambientes. Por outrolado, poder associar o estudo dessesanimais no lago Tupé, com osparâmetros ambientais que o caracterizam, como oferecido pelos distintoscapítulos deste livro, constitui maisuma ferramenta disponibilizada parainterpretações de paleo-ambientes naAmazônia e, quiçá, na América do Sul.Em razão dessas determinantes foramlevados a efeito dois levantamentosdas esponjas no lago. O primeiro foirealizado pelos participantes daequipe do INPA e, o segundo, pelaautora senior. No primeiro foram
&et al ; et al ,
-
-
-
também colhidas amostras de sedimentos atuais do lago.Os resultados obtidos vem permitir uma primeiracaracteri zação da assembléia de esponjas de um igapó,tanto em termos taxonômicos, quanto espaço/temporais, além de possibilitar um cotejo com levantamentosemelhante, levado a efeito por Batista . (2003),para lagos de várzea do rio Araguaia, portanto de águasclaras.
Foram realizados dois esforços de amostragem deesponjas no Lago Tupé, ambos na estação de águasbaixas, quando os espécimes são facilmentevisualizados, já secos, incrustando a vegetação marginalatingida pela água alta. O primeiro foi levado a efeito noexercício de 2001, pelos participantes da equipe do INPA,atendendo às estações de amostragem estabelecidas ereferenciadas para efeito de estudos e caracterizaçãolimnológica a longo prazo (Aprile ., 2005 nestevolume). Essa amostragem contemplou ainda arecuperação de três colunas de sedimentos na partecentral do lago. As amostras de sedimentos foramtomadas visando a observação microscópico paraconstatação da contribuição das espículas das esponjasaos sedimentos e verificação taxonômica das espéciescontribuintes. O segundo foi realizado no exercício de2004, pela autora senior, visando uma varredura amplados substratos atingidos pelas águas altas em distintospontos das margens do lago.
Foi ainda realizada pela equipe do INPA, através demergulho autônomo, amostragem de esponjas aderidas,abaixo da linha d'água, às pilastras do porto de Manaus,portanto no rio Negro, em ambiente de imersãopermanente.
Todos os materiais, conservados a seco, bem como aspreparações em lâminas, deles resultantes, foramdepositados e catalogados na Coleção de Porífera doMuseu de Ciências Naturais da Fundação Zoobotânica doRio Grande do Sul (MCN-POR). As preparações dedissociação espicular, para fins dos procedimentostaxonômicos identificatórios e que resultaram em
-
-
et al
et al
Material e Método
As esponjas do Lago Tupé
125

126
lâminas permanentes, seguiram Volkmer-Ribeiro (l985).Os mesmos procedimentos foram seguidos com asamostras de sedimentos.
Phylum PoriferaClasse Demospongiae
Gênero Vejdovsky, 1882
T ( Bowerbank, 1863)MCN-POR 6403. Lago Tupé,
margem direita, entrada do igapó. 3º1'58,1"S 60º16'2,5"W. 16.X.2004.; MCN-POR 6538; 6545; 6547, 6568. LagoTupé, margem direita, interior do igapó, 3º1'58,1"S60º16'2,5"W. 16.X.2004; MCN-POR 6554; 6558; 6562,Lago Tupé, 3º2'31,4"S 60º15'17,5"W. 16.X.2004. C.Volkmer Ribeiro Daniel Pimpão leg.
(Bowerbank, 1863)MCN-POR 6405. Lago Tupé,
margem direita, entrada do igapó. 3º1'58,1"S 60º16'2,5"W. 16.X.2004.; MCN-POR 6535; 6542; 6550., 6571. LagoTupé, margem direita, interior do igapó. 3º1'58,1"S60º16'2,5"W. 16.X.2004; MCN-POR 6556; 6561. LagoTupé. 3º2'31,4"S 60º15'17,5"W. 16.X.2004. C. VolkmerRibeiro & Daniel Pimpão leg.; MCN-POR 6136, Coluna 2(0-10cm), 29. IX. 2001. Darwich, Santos Silva Oliveiraleg; MCN-POR 5282. Rio Negro, pilastra no Porto deManaus, esponja coletada abaixo da linha da água(30cm), 11.XII.2001, Darwich, Santos Silva leg.
( Potts, 1882)MCN-POR 6548 Lago Tupé,
margem direita, interior do igapó. 3º1'58,1"S60º16'2,5"W. 16.X.2004; MCN-POR 6555; 6557 Lago Tupé,3º2'31,4"S 60º15'17,5"W. 16.X.2004, C. Volkmer Ribeiro
Resultados
Lista taxonômica
Família Spongillidae
Material examinado:
Material Examinado:
Material examinado:
Trochospongilla
Trochospongilla paulula
Trochospongilla pennsylvanica
rochospongilla gregaria
&
&
&
&
&
&
&
Daniel Pimpão leg.; MCNPOR 6137,Coluna 3 (0-5cm, 5-15cm). 29. IX.2001. Darwich, Santos Silva Oliveiraleg
Gênero Volkmer-Ribeiro, 1970
(Carter,1881)
MCN-POR6402; 6404; 6406. Lago Tupé, margemdireita., entrada do igapó. 3º1'58,1"S60º16'2,5" W. 16.X.2004; MCN-POR6540; 6544; 6549. Lago Tupé, margemdireita., interior do igapó, 3º1'58,1"S60º16'2,5"W.; 16.X.2004. C. VolkmerRibeiro Daniel Pimpão leg.; MCNPOR6135, Coluna 1 (0-10cm, 10-20cm). 29.IX. 2001, Darwich, Santos Silva &Oliveira leg.
(BonettoEzcurra De Drago, 1973)
MCNPOR6136, Lago Tupé, Coluna 2 (0-10cm;10-20cm). 29. IX. 2001, Darwich,Santos Silva Oliveira leg.
( Bonetto &Ezcurra de Drago, 1973)
MCN-POR5375 e 5376: Porto de Manaus( e s p é c i m e a s s o c i a d o a
MCN-POR 5288). MCN-POR 5373 ( espécime associado a
, MCN-POR 5282).
Gênero Gray 1867
(Bowerbank,1863)
.
Familia Potamolepidae
Material examinado:
Material examinado:
Material examinado:
Familia Metaniidae
Oncosclera
Oncosclera spinifera
Oncosclera intermedia
D .
ururugaiensis
T.
paulula
Metania
Metania reticulata
Oncosclera navicella
Wolkmer Almeida&

Material examinado:
-
Material examinado:
-
MCN-POR6397 Lago Tupé, boca. Afloramentorochoso. 3º2'53,6"S 60º15'8,9" W.16.X.2004. MCN-POR 6399-6401. LagoTupé, margem direita., entrada doigapó. 3º1'58,1"S 60º16'2,5" W.16.X.2004. MCN-POR 6541; 6552, 6567,Lago Tupé, margem direita, interior doigapó. 3º1 '58,1"S 60º16 '2,5"W.16.X.2004. MCN-POR 6553; 6559; 6564-66. Lgo Tupé. 3º2'31,4"S 60º15'17,5"W.16.X.2004. C. Volkmer Ribeiro DanielPimpão leg. MCN-POR 5280. Lotecoletado no chão, esponjas caídas nochão seco, ainda não inundado,próximo a Estação Central; MCN-POR5281. Galho à cerca de um metroacima do chão, fora da água. Próximo àEstação Central. MCN-POR 5284. Lotecoletado no chão, esponjas caídas nochão seco, ainda não inundado,próximo a Estação Central. MCN-POR5285. Lote coletado no chão, esponjascaídas no chão seco, ainda nãoinundado, próximo a Estação Central.21. XI. 2001. Darwich, Santos SilvaOliveira leg.
MCN-POR 5286. Igarapé daCachoeira, lote coletado no igapó,cerca de 2m do chão e cerca de 50cmda água; MCN-POR 5287. Lote coletadono igapó, cerca de 2m do chão e cercade 50cm da água. 21. XI. 2001.Scudeller Amaral leg.
Volkmer-Ribeiro,1979
MCN-POR5371, Rio Negro, pilastra no Porto deManaus, esponja coletada abaixo dalinha da água (30 cm), 11.XII. 2001,Darwich, Santos Silva, leg. (associada
&
&
&
Metania subtilis
ao espécime MCN-POR 5282, )
Gênero Gray, 1867(Bowerbank, 1863)
MCN-POR 6536. Lago Tupé,margem direita, interior do igapó. 3º1'58,1"S60º16'2,5"W. 16.X.2004, C. Volkmer Ribeiro DanielPimpão leg. , MCN-POR 5370, 5372, Rio Negro, pilastra noPorto de Manaus, esponja coletada abaixo da linha da
Trochospongilla paulula .
AcalleAcalle recurvataMaterial examinado:
&
Figura 1 Foto de exemplar de “ in situ”incrustanto a vegetação no igapó do Lago Tupé. Foto EdinaldoNelson dos Santos Silva.
. Metania reticulata
As esponjas do Lago Tupé
127

128
água (30 cm), 11.XII. 2001, Darwich, Santos Silva, leg.(associada ao espécime MCN-POR 5282,
)
Gênero Gray, 1867(Bowerbank, 1863)
MCN-POR 6537;6543 Lago Tupé,margem direita, interior do igapó. 3º1'58,1"S60º16'2,5"W. 16.X.2004. C. Volkmer Ribeiro DanielPimpão leg.MCN-POR 5283. Igarapé da Cachoeira, Lotecoletado no chão, esponjas caídas no chão seco, aindanão inundado, próximo a Estação Central , 21. XI. 2001.Scudeller Amaral leg.
Bonetto Ezcurra de Drago,1969
MCN-POR 6389-96; 6398; 6407.Lago Tupé, boca. Afloramento rochoso. 3º2'53,6"S60º15'8,9" W. 16.X.2004. MCN-POR 6539; 6546; 6551.Lago Tupé, margem direita, interior do igapó. 3º1'58,1"S60º16'2,5"W. 16.X.2004. MCN-POR 6560; 6563,6569,Lago Tupé 3º2'31,4"S 60º15'17,5"W. 16.X.2004 C. VolkmerRibeiro Daniel Pimpão leg.; MCNPOR 5288. Rio Negro,pilastra no Porto de Manaus, esponja coletada abaixo da
Trochospongilla
paulula .
DruliaDrulia browni
Drulia uruguayensis
Material examinado:
Material examinado:
&
&
&
&
linha da água (30 cm); MCN-POR 5370 ,pilastra no Porto de Manaus, esponjacoletada abaixo da linha da água (30cm), (associada a MCN-POR 5282,
), 11.XII. 2001,Darwich, Santos Silva, leg.
O levantamento feito permitiuconstatar que há predominância deocupação do ambiente do igapó poresponjas da Familia Metaniidae(gêneros , e ) e,em segundo lugar, da FamiliaSpongillidae (gênero ).Enquanto as esponjas da primeirafamília incrustam os substratos maisreforçados, existentes da boca do lagoaos níveis mais altos de residência daságuas altas, os da segunda ocupamsubstratos mais delicados, comofolhas, chumaços de raízes aéreaspendentes e cascas das árvores eretasou tombadas no chão. A abundânciade gêmulas é uma constante de todasas espécies presentes e, no caso deSpongillidae é notória a redução daformação do esqueleto em favor daprodução de gêmulas (fig. 3 ) em cadaperíodo de imersão.
As esponjas com espécimes maisconspícuos são certamente as daFamilia Metaniidae, devido aconstituição polispicular de suas fibrasesqueletais, compondo assim exem-plares esféricos ou tuberosos de volu-me apreciável, de duros a extrema-mente duros, (particularmente no casode ) e de peso ele-vado, dada a concentração de sílicamineralizada nos esqueletos. Essepeso ocasiona a ruptura dos galhosmais delgados das árvores em que sefixaram, contribuindo para a disper-
Trochospongilla paulula
Metania Drulia Acalle
Trochospongilla
Metania reticulata
Figura 2 Foto de exemplar de , coligida no LagoTupé, Manaus, AM e registrada no presente trabalho com onúmero de catálogo MCN-POR 5283. Foto Vanessa de SouzaMachado.
. Drulia browni
Wolkmer Almeida&

são de suas gêmulas no chão do igapó.É possível perceber que essas crostasnão advém de um único período deimersão e isto se deve, justamente, àdureza dos seus retículos esqueletais,que mantém as gêmulas retidas nocorpo da esponja mãe. Com o próximoperíodo de imersão essas eclodem,constituindo uma nova camada viva,que passa a produzir um novoesqueleto, sobre aquele do períodoanterior. Dada a cor diferenciada daesponja produzida no período deimersão mais recente, percebe-se quea nova camada não corresponde a todasuperfície do período anterior, masconstitui manchas delgadas sobreaquela.
As esponjas da Familia Potamolepi-dae estão representadas no ambientedo igapó por espécimes com reduzidoesqueleto, quando não apenasgêmulas, pertencentes a três espéciesdo gênero , sendo maisfrequente . Mas, mesmoesta, comparece apenas com asgêmulas, no entanto em númerosmuito grandes, constituindo minúsculos domos de cor escura, firmementecolados aos substratos. Aliás, damaneira como foi detectada e descritapor Carter (1881): como sendo agêmula de uma espécie cujo esqueletoera então desconhecido. Nossubstratos rochosos de fundos de riosde toda América do Sul essa esponjaforma crostas espessas e extensas, decor creme, apresentando as gêmulasacima descritas coladas ao substrato eainda gêmulas menores, em grupos, nointerior do esqueleto (TavaresVolkmer-Ribeiro, 1997).Trata-se
Oncosclera
O. navicella
-
&
claramente de esponjas de fundos rochosos que, noigapó, comparecem com suas formas de resistência,devido a não ser esse seu ambiente ideal. Fatosemelhante foi detectado por Batista . (2003) noslagos de várzea do rio Araguaia, com respeito a esponjasda família Potamolepidae.
Aprospecção, efetuada com barco, permitiu levantarda água substratos ainda mergulhados, como galhostombados de árvores ocorrentes no meio do lago. Essescontinham esponjas vivas indicando que a qualidade daágua baixa não é, no igapó, impeditiva ao crescimento daesponja. No entanto convém considerar a possibilidadeque esses galhos pudessem haver caído recentemente, jácom esponjas formadas em níveis anteriormenteexpostos.Aprofundidade a que esses espécimes estavammergulhados era próxima da superfície.
O estudo dos sedimentos mostrou um contribuiçãosignificativa das espículas das esponjas aos mesmos (Fig.4), além de permitir a conferência da composiçãoespecífica da assembléia levantada. Dada a ocorrêncianos sedimento das espículas das gêmulas, além daquelasdo esqueleto, foi possível identificar as espéciescontribuintes e constatar que o levantamento feito
et al
Figura 3. Foto de folha da palmeiraArecaceae coligida no igapódo Lago Tupé, Manaus, AM (MCN-POR 6571) incrustada com aesponja evidenciando apredominância de formação de gêmulas, em detrimento daconstituição do esqueleto. Foto Vanessa de Souza Machado.
Trochospongilla paulula
As esponjas do Lago Tupé
129

130
oferece uma figura bastante real da fauna de esponjasque existe no lago. Houve ocorrência de algumasespículas que indicam, contudo, que mais uma ou duasespécies não foram amostradas. Essas espículas, dadosua raridade, não permitiram entretanto a identificaçãodessas espécies.
As esponjas amostradas nas pilastras do porto deManaus, Rio Negro, constituiram exemplares volumosos,com formas hemisféricas e com esqueletos muitofrágeis. Outras características marcantes a distinguiremos espécimes desse local dos do Lago Tupé foi aocorrência generalizada, nos exemplares do porto deManaus, de epizoísmo entre as distintas espécies ( o queexigiu um tempo bem maior para se chegar àsidentificações), além da quase ausência de gêmulas,indicando que aí as esponjas estão investindo naconstituição do esqueleto, por não estarem enfrentandocondições adversas e que, obviamente, a dispersãodeverá estar se efetivando com a emissão de larvas,resultantes de reprodução sexuada.
Discussão eConclusões
Três espécies de dois gêneros daFamilia Metaniidae Volkmer,
, , ee uma da Família
Spongillidae, ,dominam e tipificam a espongofaunado Lago Tupé. As duas primeiras e aquarta espécie são características deambientes de florestas de várzeasinundadas estacionalmente, identidade que marca, aliás, em toda faixacircum-tropical (Volkmer-Ribeiro,1986) a maioria das espécies e gênerosda Família Metaniidae. As distinçõesmaiores entre os represen tantes dasduas famílias dizem respeito aestrutura dos esqueletos e, dependentes desses, os substratos elegidos paracolonização pelas esponjas. Enquantoas esponjas da Família Metaniidaemostram estruturas esqueletais extremamente duras e cerradas, com asgêmulas retidas nas malhas dessesesqueletos, as da Família Spongillidaecompõem esqueletos frágeis, com asgêmulas soltas e expostas na redeaberta e rudimentar do esqueleto.Esse fato determina padrões dedistribuição bem distintos dessa faunano igapó, as metaniidae utilizandosubstratos fortes, como ramos e galhosmaiores da vegetação, além desubstratos rochosos da boca do lagoenquanto os espongilideos garantempara sí a folhagem e galhos menoressubmersos ou mesmo as cascas dasárvores. Imbricadas nessa distribuiçãoestão as características das gêmulas:
Metania
reticulata Drulia browni Drulia
uruguayensis
Trochospongilla paulula
-
-
-
-
Figura 4. Foto de lâmina permanente, resultante dapreparação de sedimentos atuais do Lago Tupé, evidenciando aabundância de espículas das esponjas. Foto Franciely BenettideAlmeida.
Wolkmer Almeida&

em metaniidae, presas no corpo daesponja mãe, o que redunda emesponjas silicosas volumosas e duras,resultantes de períodos sucessivos esobrepostos de eclosões gemulares ede formação de esqueletos, sustenta-dos por esses substratos mais firmes.No caso dos espongilídeos, as gêmulasdispersam-se facilmente por flutuaçãoe dispensam substratos firmes, já quea adição de temporadas estacionais deprodução redunda em ocupação denovos substratos. O fato é que a adiçãodas duas estratégias de ocupação levaa um ambiente extensivamente coloni-zado pelas esponjas, como foi vistoser o do Lago Tupé. Essa ocupação,amplamente verificada na AmazôniaCentral (Volkmer-Ribeiro, 1981) distri-bui-se tanto no gradiente vertical, desubstratos próximos ao solo, às copasdas árvores mais altas situadas nocentro do lago, quanto no horizontal,espraiando-se pela vegetação dasmargens, até onde as águas das cheiastem seus períodos maiores deresidência.
Houve uma distinção marcanteentre os materiais coligidos no LagoTupé e os amostrado nas pilastras doPorto de Manaus. Os primeirosconstituiram espécimes isolados, semincidência de epizoismo, indicandopreliminarmente uma fauna comausência de competição por substrato.Entende-se esse fato pela disponibili-dade de substratos propiciada pelafloresta marginal inundada do igapó.No porto de Manaus as esponjasencontraram esse substrato disponibi-lizado, de modo mais escasso, pelaspilastras do porto. A escassez, para
Figura 5 Foto do Lago Tupé na ocasião da amostragemrealizada pela autora senior no fim do período de água baixa(outubro de 2004). As manchas negras indicadas pelas flechascorrespondem aos múltiplos espécimes de ede , cujas gêmulas, retidas no esqueleto quesecou, eclodem no seguinte período de águas altas e formamnovas crostas sobre a antiga. Foto Cecilia Volkmer Ribeiro.
.
Metania reticulataDrulia browni
não dizer ausência de gêmulas, nos espécimes do porto,indica uma incidência maior de reprodução/dispersãopor larvas resultantes de reprodução sexuada. Essas,livre-natantes, cumprem a função de, no reduzidotempo de vida que têm, buscar substratos imersos emáguas que apresentam a qualidade de água compatível
As esponjas do Lago Tupé
131

132
com a vida da esponja adulta ( Sara Vacelet, 1973). Aságuas do rio Negro, aliadas às pilastras do porto devemcorresponder a essas necessidades para as esponjas aíocorrentes. Essas podem assim investir em constituiçãoesqueletal permanente, enquanto as do Lago Tupé, dadaa drástica retração do meio líquido na estação da seca,tem de investir na produção de gêmulas.
A prospecção de espículas nos sedimentos,comprovou, mais uma vez a utilidade dessa amostragemindireta, há longo tempo prognosticada e utilizada,tanto para detecção de assembléias atuais (Volkmer-Ribeiro, 1985), quanto pleistocênicas, possibilitandopaleo-interpretações (Harrison, 1988; HarrisonWarner, 1986; Harrison . 1979; Turcq . 1998,Sifeddine . 2001). Os resultados aqui aportadosvem confirmar aqueles obtidos por Chauvel .(1996),no pertinente à abundância das espículas nossedimentos, enfatizando a contribuição importantedessa fauna para a constituição física dos sedimentos dosigapós e no processo de bio-mineralização da sílicaretirada da água. Jorgensen (1944) demonstrou queessas esponjas detém ainda o potencial de retirarem asílica existente em outras fontes, que não a dissolvidana água, como a areia, suas espículas e frústulas dediatomáceas incorporadas aos sedimentos. Assim sendoo componente faunístico espongológico desempenhaum papel importante na geoquímica da água e dossedimentos do lago.
O levantamento efetuado aponta, no conjunto dasespécies detectadas, para uma espongofauna de águasnegras, tão exuberante quanto àquela levantada naságuas claras amazônicas (Batista . 2003). Umaprimeira comparação dos padrões de oxigênio dissolvido,pH e condutividade entre os agora obtidos para as águasaltas no Lago Tupé (Darwich , 2005, neste volume) eaqueles observados nas cheias para os lagos de Várzea doAraguaia, permite distinguir diferenças apreciáveis, comáguas bastante mais ácidas e com baixa condutividade noprimeiro caso, confirmando, aliás, observaçõesaportadas em um número infindo de trabalhos realizadosna Amazônia (Junk Furch, 1985). No entanto essasdiferenças não parecem ser as que mais influem sobre aabundância da fauna espongológica, pelo menos no que
&
&
&
et al , et al
et al ,
et al
et al ,
et al.
respeita a águas pretas e claras, ajulgar-se pelas comparações agorarealizadas e que apontam parasituações similares entre o Lago Tupé eos lagos de Várzea doAraguaia.
As diferenças marcantes daconcentração de bicarbonatos sãocolocadas como um divisor, naAmazônia, (Junk Furch, 1985) entreas águas brancas de um lado e as clarase negras de outro. A similaridade,agora apontada, entre águas negras eclaras, no que tange a abundância dafauna espongológica, vem indicar queesse seja mais um elemento de caráterbiológico a confirmar essa divisão daspropriedades químicas das águas naAmazônia. Restará a viabilização deum levantamento em ambientes deáguas brancas, semelhante ao operadono Araguaia e agora no Tupé, paraefetivar-se a comparação dos resulta-dos e verificar se as diferenças naconcentração de bicarbonatos influemna distribuição também da faunaespongológica.
Por sua vez a abundância deespículas nos sedimentos recomendacautela no uso dessas águas paraatividades de lazer, principalmente noperíodo de águas baixas. As espículas,por movimentação dos sedimentos,podem passar à resuspensão na colunad´água. O contato direto com a pelepode, ocasionalmente, provocardermatites e conjuntivites, pelapenetração das espículas na pele e naconjuntiva. No caso de consumo daágua para ingestão, seja como bebidaou no cozimento de alimentos, aprecaução é idêntica. É óbvio que esserisco ocorrerá nos locais onde ossedimentos acusarem maiores concen-
&
Wolkmer Almeida&

trações das espículas, como aconteceunos locais de recuperação das colunasde sedimento no centro do lago.
Aos Drs. Assad Darwich, EdinaldoNelson dos Santos-Silva, INPA eVeridiana Vizoni Scudeller, CEULM/ULBRA, pelo primeiro esforço deamostragem de esponjas no Lago Tupée no porto do Manaus; ao Dr. EdinaldoNelson dos Santos Silva, pela disponibi-lização de oportunidade e apoio paraque a autora senior levasse a efeito olevantamento abrangente da fauna deesponjas no lago. Ao CNPq., pela bolsade Produtividade em Pesquisa concedi-da à autora senior. À Fundação deAmparo à Pesquisa do Estado do RioGrande do Sul (FAPERGS), pela bolsa deIC concedida à autora junior.
Aprile, F. M.; Darwich, A. J. 2005.Modelos Geomorfológicos para olago tupé, : Santos-Silva, E. N.;Aprile, F. M. Scudeller, V. V.; Melo,S.,
.Ed. Inpa,AM.
Batista, T. C. A.; Volkmer-Ribeiro, C.;Darwich, A.; Alves, L. F. 2003.Freshwater sponges as indicators offloodplain lake environments andof river rocky bottoms in CentralAmazonia. XVII(3/4):525-549.
Agradecimentos
Bibliografia Citada
In
BioTupé: meio físico, diversida-
de biológica e sóciocultural do
baixo rio Negro, Amazônia Central
Amazoniana,
As esponjas do Lago Tupé
133
Carter, H. J. 1881. History and classification of theknown species of Spongilla.
, 5(7): 77-107.Chauvel, A.; Walker, I.; Lucas, Y. 1996. Sedimentation
and pedogenesis in a Central Amazonain Black waterbasin. 33:77-95.
Darwich, A. J.; Aprile, F. M., Robertson, BA. 2005.Variáveis limnológicas: Contribuição ao EstudoEspaço-temporal de Águas Pretas Amazônicas. :Santos-Silva, E. N.; Aprile, F. M. Scudeller, V. V.; Melo,S.,
.Ed. Inpa,AM.
Harrison, F. W. 1988. Utilization of freshwater spongesin paleolimnological studies. In : Gray, J.
. Elsevier, Amsterdam: p. 387-397.Harrison, F. W.; Gleason, P. J.; Stone, P. A. 1979:
Paleolimnology of Lake Okeechobee, Florida: Ananalysis utilizing spicular components of freshwatersponges (Porifera: Spongillidae). ,Philadelphia, 454: 106.
Harrison, F. W.; Warner, B. G. 1986. Fossil fresh-watersponges (Porifera: Spongillidae) from WesternCanada: An Overlooked Group of QuaternaryPleoecological Indicators.
, Lancaster, 105 (2):110-120.
Jorgensen, C. B. 1944. On the spicule formation ofSpongilla lacustris (L.). 1. The dependence of thespicule- formation on the content fo dissolved andsolid silicic acid of the milieu.
, 19: 2-45.Junk, W.; Furch, K. 1985. The physical and chemical
properties of Amazonian Waters and theirRelationship with the Biota, In: Prance, G. T.Lovejoy, T. E. (eds.) ,Pergamon Press, New York: p3-17.
Reiss, F. 1977. Qualitative and quantitativeinvestigations on the macrobenthic fauna of CentralAmazon lakes. 1. Lago Tupé, a black water lake on thelower Rio Negro. , 6 (2): 203-235.
Sara, M.; Vacelet, J. 1973. Ecologie des démosponges.
Annals Magazine of
Natural History
Biogeochemistry,
In
BioTupé: meio físico, diversidade biológica e
sóciocultural do baixo rio Negro, Amazônia Central
Paleolimnology: Aspects of freshwater paleoecology
and biogeography
Notulae naturae
Transactions of the
America Microscopical Society
Det .Kgl. Danske.
Vidensks. Selsk. Biol. Medd.
Amazonia Key Environments
Amazoniana
&

134
Wolkmer Almeida&
Diego State University, San Diego,Cal. USA: p 86-95.
Volkmer-Ribeiro, C. 1985. Esponjas deÁgua Doce. In: Sociedade Brasileirade Zoologia (Ed.),
, São Paulo, 3 :6pp.
Volkmer-Ribeiro, C. 1999. Esponjas.In: Joly, C. A., Bicudo, C. E. de M.
; síntese do conhecimento dofinal do século XX, 4: Invertebradosde água doce. FAPESP São Paulo:p1-9.
Volkmer-Ribeiro, C.; Pauls,S. M. 2000.Esponjas de água Dulce (Porifera:Desmospongiae) de Venezuela.
,Caracas, 20 (1):1-28.
.
Manual de
Técnicas para a preparação de
coleções zoológicas
Biodiversidade do Estado de São
Paulo
,
Acta Biológica Venezuelica
In: P-P. Grassé (ed.): Traité de Zoologie. Vol. III,fasc.I. Masson et Cie, Paris.: P 462-576.
Sifeddine,A.; Martin, L.; Turcq, B.; Volkmer-Ribeiro, C.;Soubiès, F.; Cordeiro, R. C.. Suguio, K. 2001.Variations of the Amazonian rainforest environment:a sedimentological record covering 30,000 years.
,168: 221-235.
Tavares, M.C.M. Volkmer-Ribeiro, C. 1997. Redescriçãodas esponjas de água doce(Carter, 1881) (Potamolepidae) eVolkmer-Ribeiro Maciel, l983 (Spongillidae).
, 5 (1): 97-111.Turcq, B.; Sifeddine, A.; Martin, L.; Absy, M.L.; Soubiès,
F.; Souguio, K.; Volkmer-Ribeiro, C. 1998. AmazonForest fires: a lacustrine report of 7.000 years.
, Stockholm, 27 (2): 139-142.Volkmer-Ribeiro, C. 1981. Porifera. In: S.H. Hurlbert, G.
Rodrigues and N.D. Santos (eds.). San
Palaeogeography, Palaeoclimatology, Palaeoecology
Oncoclera navicella
Spongilla spoliata
Biociências
Aquatic Biota of
Tropical South America, Part 2: Anarthropoda
&
Ambio

Diversidade BiológicaCapítulo 9
Diversidade de macromicetoslignolíticos
RESUMO - A diversidade fúngica de macromicetos lignolíticos que ocorrem nas florestas daReserva de Desenvolvimento Sustentável do Tupé (RDS Tupé) foi estudada. Os fungos sãoessenciais para o meio ambiente florestal, atuando na degradação da madeira morta epromovendo a ciclagem de nutrientes. Além disso, podem ser utilizados na alimentação,como fármacos e na recuperação de áreas contaminadas, sendo importantes componentesbiológicos e ecológicos dos ecossistemas e com bom potencial biotecnológico. Foramencontradas oito espécies:
e, classificadas em seis famílias: Tricholomataceae,
Ganodermataceae, Hymenochaetaceae; Polyporaceae, Schizophyllaceae eAuriculareaceae. Os gêneros mais encontrados na área foram e
.
PALAVRAS-CHAVES: Basidiomicetos, Fungos Lignolíticos, Lago Tupé,Amazônia.
Panus crinitus, Pleorotus flabeliforme, Ganoderma
applanatum, G. lucidum, Phellinus gilvus, Pycnoporus sanguineus, Trametes versicolor
Schizophyllum commune
Ganoderma, Phellinus
Polyporus
André Luis WILLERDING
Marcerla Farias BARBOSAThaís Campos BURLAMAQUI
1
2
2
1
2
Professor do Curso de Ciências Biológicas CEULM/ULBRA
Alunas de graduação do Curso de Ciências Biológicas CEULM/ULBRA.
&
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

136136
Introdução
A natureza e toda sua diversidade genética têm umpotencial valor em função da possibilidade de seu uso nasmais diversas formas. O levantamento da biodiversidadetem sido importante para o conhecimento e a utilizaçãodos recursos genéticos (BDT, 1998 a,b). Neste contexto,diferentes tipos de fungos macromicetos são alvos depesquisa em todo o mundo, e não apenas por suaspropriedades lignolíticas, mas como medicamentos oufontes nutrimentais (Tiedje, 1994; Machado ., 1995;Ferreira, 2000; Martins, 2003).
Os fungos degradadores de madeira, conhecidospopularmente como orelha-de-pau, urupê oucogumelos, são Basidiomicetos superiores (Macromice-tos) e apresentam corpos de frutificação (basidiomas)visíveis a olho nu. Esses fungos são denominados delignolíticos por possuírem a capacidade de degradarlignina presente na madeira. Geralmente sapróbios,podem ser encontrados também como pertófilos, sedesenvolvendo em partes mortas de plantas vivas(Souza, 1980). Assim, são essenciais para o meioambiente florestal, atuando na degradação da madeiramorta e contribuindo para a ciclagem de nutrientes(Sotão ., 2003).
Ao enfraquecerem as árvores velhas, estes fungospromovem a remoção destas a partir do “stand” dafloresta. Isto permite o crescimento de novas árvores epor conseqüência, fazem um papel integral na dinâmicaflorestal, pela ciclagem de nutrientes ou pela promoçãoindireta de clareiras, influenciando na sucessão naturalda floresta (Fidalgo, 1968; Jesus, 1988; Campos, 2000).
Os basidiomicetos podem ser bons indicadoresecológicos, pois suas amplitudes ecológicas resultamnuma distribuição das espécies que refletem ascondições ambientais. Se as condições forem favoráveis,os basidiósporos germinam e o micélio se desenvolve nointerior da madeira. Conseqüentemente, a taxa dedecomposição dos troncos caídos varia conforme o local.Levando em consideração que a excessiva umidade damadeira inibe o desenvolvimento destes fungos pelafalta de oxigênio (Souza, 1980), isto pode explicar abaixa ocorrência nas áreas freqüentemente alagadas,como as matas de igapó existentes na área da RDS Tupé.Quando em terra-firme, ou em áreas abertas, estão
et al
et al
sempre situados na região do troncovoltada para o solo, protegendo-se daluminosidade. Mesmo assim, algumasespécies heliófitas, semi-heliófitas eumbrófilas são encontradas (Texeira,1994). Por tais características, oacompanhamento da ocorrênciadestes fungos em áreas florestais,pode dar uma idéia do nível dedegradação ou de manutenção damanta florestal em um determinadolocal (Texeira, 1994; GuglliottaBononi, 1999).
Atualmente, existem cerca de20.000 espécies de basidiomicetoslignolíticos, a maioria localizada emregiões tropicais e sub-tropicais. NoBrasil existem diversos trabalhos sobrea ocorrência destes fungos, tais comoTheissen (1911), Texeira (1945, 1948),Rick (1963), Fidalgo (1957, 1962, 1965,1979), Bononi (1998), Capelari .(1996), Leite (1990) e Fonseca (1994),todos citados em Jesus (1988). Para aAmazônia, não existem tantostrabalhos assim, mas ocorreram algunsestudos na região, como Fidalgo(1968), Souza (1980), Bononi (1981),Capelari Maziero (1988), Jesus(1995; 1996), Campos (2000), Campos
Cavalcanti (2000) e Sotão .(2003).
O conhecimento da diversidadefúngica de macromicetos degradado-res de madeira que ocorrem na RDSTupé é uma oportunidade paraaumentar o conhecimento dos fungoslignolíticos naAmazônia.
O trabalho de campo foi realizadoao longo do ano de 2003. As coletas domaterial biológico (basidiocarpo)foram realizadas em duas trilhas
&
&
&
et al
et al
Materiais e Métodos
Willerding et al.

Ordem Família Gênero Espécie Espécimes
Tricholomataceae Panus Panus crinitus 3
Pleorotus Pleorotus flabeliforme 2
Pleorotus spp. 1
Schizophyllaceae Schizophyllum Schizophyllum commune 4
Auriculares Auriculareaceae Auricularia Auricularia spp. 2
total 71
existente no local, a Trilha Central e ada Cachoeira, com aproximadamente3Km de comprimento cada (Scudeller,
, 2005, neste volume), observan-do a presença dos fungos nos troncosde árvores caídos ao longo dopercurso. O material coletado foiretirado integralmente do substrato,sendo posteriormente acondicionadosem jornal e etiquetado.
Após a etapa de coleta, o materialfoi posto em estufa e depositado noLaboratório de Microbiologia daCoordenação de Biologia do CentroUniversitário Luterano de Manaus(CEULM-ULBRA). No laboratório foiadotada a metodologia clássica para asobservações dos basidiomicetos comobservações macroscópicas como adescrição da cor, das dimensões e dodetalhamento das faces superior einferior do basidiocarpo, além dasobservações microscópicas para as
et al.
estruturas internas para o auxílio na identificaçãotaxonômica seguindo as técnicas propostas em FidalgoBononi (1984) e Texeira (1993).
Foram encontradas oito espécies:
,classificadas em seis famílias: Tricholomataceae,Ganodermataceae, Hymenochaetaceae; Polyporaceae,Schizophyllaceae e Auriculareaceae. Os gêneros maisencontrados na área foram , e
. A Tabela 1 mostra a classificação taxonômicado material coletado. As figuras 1 e 2 mostram adistribuição de espécimes por família e por gênerorespectivamente.
Adistribuição de fungos basidiomicetos degradadoresde madeira coletados na RDS Tupé parece seguir umapredominância das famílias Polyporaceae, Hymenochae
&
Resultados e Discussão
Panus crinitus,Pleorotus flabeliforme, Ganoderma applanatum, G.lucidum, Phellinus gilvus, Pycnoporus sanguineus,Trametes versicolor e Schizophyllum commune
Ganoderma PhellinusPolyporus
-
Diversidade de macromicetos lignolÍticos
Tabela 1. Grupos taxonômicos do material coletado nas trilhas Central e da cachoeira na RDS Tupé.
137
Aphyllophorales Ganodermataceae Ganoderma Ganoderma applanatum 4
Ganoderma lucidum 5
Ganoderma spp. 3
Hymenochaetacea Phellinus Phellinus gilvus 8
Phellinus spp. 15
Polyporaceae Pycnoporus Pycnoporus sanguineus 2
Trametes Trametes versicolor 3
Agaricales
Polyporus spp. 19Polyoporus

138
Willerding et al.
0 5 10 15 20 25 30
Auriculareaceae
Ganodermataceae
Hymenochaetaceae
2
12
23
24
4
6
Polyporaceae
Schizophyllaceae
Tricholomataceae
espécimes
Figura 1. Distribuição de indivíduos coletados por família de Basidiomicetos nas trilhasCentral e da Cachoeira da RDS Tupé.
Gêneros
2
12
3
23
3
19
2
4
0 5 10 15 20 25
Auricularia
Ganoderma
Panus
Phellinus
Pleorotus
Polyporus
Pycnoporus
Schizophyllum
espécimes
Figura 2. Distribuição de gêneros de basidiomicetos indentificados.
nas trilhas Central eda Cachoeira da RDS Tupé

Diversidade de macromicetos lignolÍticos
139
taceae e Ganodermataceae (Fig. 1),sendo representadas principalmentepelos gêneros , e
. (Fig. 2). Um trabalhorealizado por Jesus Bononi (1991),sobre um levantamento de fungosdegradadores de madeira na área daHidrelétrica de Balbina, no Amazonas,apresentou uma abundância para estesmesmo gêneros e com uma maiorfreqüência para a espécie
. Jesus (1996) apresentaum levantamento sobre fungosdegradadores de madeira para a Ilhade Maracá, em Roraima. Seusresultados mostram uma diversidadede espécies fúngicas quando comparada com os resultados deste trabalho.Com relação às famílias, seu trabalhocita dez famílias para aquela área deRoraima, incluindo as famíliasPolyporaceae, Hymenochaetaceae,Ganodermataceae e Auriculareaceae,(que também são citadas nestetrabalho), com muitos gêneros emcomum, como sp.,
sp., sp.,sp., sp. e sp.Curiosamente, somente uma espécie,
, é encontrada tambémna área da RDS Tupé. Esses resultadosdemonstram o quanto é alta adiversidade dos fungos degradadoresde madeira, e que mesmo sobcondições semelhantes de floresta, aheterogeneidade com relação àsespécies permanece alta de local paralocal.
BDT - Base de Dados Tropical, 1998a.
. Grupo de trabalho
Polyporos PhellinusGanoderma
Schizophy-llum commune
-
TrametesGanoderma Auricularia Panus
Phellinus Polyporus
Phellinus gilvus
Estratégia Nacional de Conserva-ção ex situ
&
Bibliografia Citada
t e m á t i c o : C o n s e r v a ç ã o e x s i t u .I n : h t t p : / / w w w . B d t . o r g . b r/ P u b l i c a c o e s / p o l i t i c a / g t t / g t t 3 .
BDT - Base de Dados Tropical, 1998 b.
. I n :http://www.bdt.org.br/publicacoes/politica/gtt/gtt10.BDT - Base de Dados Tropical, 1998 b.
. I n :http://www.bdt.org.br/publicacoes/politica/gtt/gtt 1 0 .
Bononi, V.L.R. 1981. Alguns basibiomicetos hidinóides daregião amazônica. , 9:17-30.
Campos, E.L. 2000..
Universidade Federal de Pernambuco .85pp.
Campos, E.L.; Cavalcanti, M.A.Q. 2000. Primeiraocorrência de s (Imaz.) Imaz.para o Brasil.Acta bot. Brás. 14(3):263-265.
Capelari, M.; Maziero, R. 1988. Fungos macroscópicos deRondônia, região dos rios Jarú e Ji-Paraná.(15):28-36.
Capelari, M.; Gugliotta, A.M.; Figueiredo, M.B. 1996. Oestudo de fungos macroscópicos no Estado de SãoPaulo. In: Joly, C. A; Bicudo, C.E.M. (orgs.).
. SãoPaulo 79pp.
Ferreira, J. E. F. 2000.. (Comunicação Pessoal).
Fidalgo , M.E.P.K. 1968. Contribuition ti the fungi of MatoGrosso, Brazil. (3):171-219.
Fidalgo, O.; Bononi, V.L.R. 1984..
São Paulo: Instituto de Botânica. Manual nº 45.Guglliotta, A.M.; Bononi, V.L.R.
. Boletim doInstituto de Botânica n 12. São Paulo.
Jesus, M.A. 1988. Basidiomicetos lignocelulolíticos defloresta nativa e de Engel. do ParqueEstadual das Fontes do Ipiranga. São Paulo,20 (½):119-126.
Jesus, M.A. 1995. Contribuition to the knowledge ofwood-rotting fungi in Brazil. I. Occurrrence and
EstratégiaNacional de Diversidade Biológica: Microrganismos eB i o d i v e r s i d a d e d e S o l o s
EstratégiaNacional de Diversidade Biológica: Microrganismos eB i o d i v e r s i d a d e d e S o l o s
RickiaBasidiomycotina de manguezais da
Ilha do Algodoal-Maiandeua, Pará, Brasil,
Phellinus mangrovicu,
Hoehnea,
Biodiversidade do Estado de São Paulo.
Cultivos de cogumelos(Fungicultura)
Rickia,Técnicas de coleta,
preservação e harborizaçào de material botânico
Polyporaceae doParque Estadual da Ilha do Cardoso
.
Pinnus elliottiHoehnea,
Recife(Dissertação de Mestrado).
FAPESP.
1999

140
Sotão, H.M.P.; Campos, E.L.;Gugliotta, A.M.; Costa, S.P.S. 2003.F u n g o s M a c r o s c ó p i c o s :basidiomycetos. : Serra (ed.)2003.
. Belém. . 45-59.Souza, M.A. 1980. O gênero
Quelet (Hymenochaetaceae) naAmazônia Brasileira.
. Instituto Nacional dePesquisas da Amazônia,199pp.
Texeira, A.R. 1993. Chave paraidentificação dos gêneros depolyporaceae com base namorfologia do basidiocarpo.
n8.
Texeira, A.R. 1994. Genera ofpolyporaceae: an objectiveapproach.
ITU. São Paulo.Tiedje, J.M. 1994. Microbial Diversity:
of value to Whom?(60):524-25.
InOs manguezais da costa norte
brasileira .Phellinus
Tese deDoutorado
Boletim do Instituto de Botânica
Boletim da ChácaraBotânica.
ASM News,
MPEG
Manaus.
p.
o
Willerding et al.
distribuition of fungi on different substrates from theManaus region,Amazonas State.
.p.1-17.
Jesus, M.A. 1996. Contribuition the knowledge of wood-rotting fungi in Brazil. II. Checklist of fungi fromMaracá Island, Roraima State. Vol LVII.323-328.
Jesus, M.A.; Bononi, V.L.R. 1991. Fungos em essênciasflorestais da área da Usina Hidrelétrica de Balbina,Presidente Figueiredo, AM.
. Boletim ABPM 71, SãoPaulo.
Martins, E. 2003. Muito além do champignon.(33): 52-53.
Machado; K.M.G.; Silva, W.R.F.; Bononi, V.L.R. 1995.
.Abstracts book of Second Latin AmericaBiodegradation and Bioderioration Symposium. 35pp.
Scudeller, V. V.; Aprile, F. M.; Melo. S.; Santos-Silva, E. N.2005. Reserva de Desenvolvimento Sustentável doTupé: características gerais. : Santos-Silva, E. N.;Aprile, F. M.; Scudeller, V. V.; Melo, S. (Orgs.).
.Ed. INPA,AM.
In: The InternationalResearch group on wood preservative. Helsinque
Mycotaxon,
Associação Brasileira dePreservadores de Madeira
CiênciaHoje
Screening of lignolytic fungi for soil remediation II:prodution of peroxidases and phenoloxidases
In
Biotupé: meio físico, diversidade biológica esociocultural do baixo Rio Negro, Amazônia Central

Diversidade BiológicaCapítulo 10
O uso da fauna pelos moradores daComunidade São João e Colônia Central
RESUMO - As comunidades São João do Tupé e Colônia Central estão localizadas na Reservade Desenvolvimento Sustentável do Tupé, à margem esquerda do rio Negro. As estratégiasde subsistência dessas duas comunidades e os recursos utilizados em relação à pesca, caça,animais de uso medicinal e consumo protéico foram comparadas. A riqueza de espécies émaior na São João, mas essas diferenças não são significativas (t = 1,76;0,1<p<0,05). Os moradores das duas comunidades utilizam 50 etnoespécies: 22 de pesca,14 de caça e 23 como medicinal (oito etnoespécies têm mais de um uso). Os recursos maisutilizados foram: jaraqui ( spp.), tucunaré ( spp.), cará ( ),cutia ( ) e sucuriju ( ). O peixe é usado como alimento eo anzol é o instrumento mais utilizado. O lago Tupé e igarapés próximos são os locaispreferidos de pesca. A caça é menos praticada que a pesca, mas também contribui para oconsumo protéico das comunidades, principalmente da Colônia Central. Os répteis sãomais utilizados como medicinais, sendo suas banhas as frações mais utilizadas para tratar
0.05(2), 200
Semaprochilodus Chicla Cichlidae
Dasyprocta agouti Eunectes murinus
Adriana Kulaif TERRA
George Henrique REBÊLO
1
2
e-mail: [email protected] PCI/INPA
e-mail: [email protected] Nacional de Pesquisas da Amazônia (INPA). Caixa Postal 478, cep 69011-970, Manaus-AM
1
2
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

142
Introdução
As relações do homem com o ambiente secaracterizam por uma combinação de uso e conservaçãoe cada sociedade cria critérios próprios que consagram amaneira pela qual os recursos devem ser utilizados(Morán, 1990). A dicotomia existente entre uso derecursos e a conservação do ambiente traz apreocupação em compreender como ocorre essa relaçãohomem/ambiente em cada cultura (Diegues, 1996).
As pesquisas sobre populações tradicionaisconstroem interfaces entre as ciências sociais e asciências da natureza. Saberes sobre a natureza e asformas de manejo dessas populações são fundamentaispara a conservação da biodiversidade (Castro, 2000).
As populações que vivem no trópico úmidopreservam, mantém e utilizam de forma extrativistaalguns produtos da floresta em que vivem. Não sóconvivem como manejam, quando usam seusconhecimentos sobre exatamente onde e quandoencontrar cada recurso (Rebêlo Galatti, 1995).
O trópico úmido é o bioma terrestre com a maiordiversidade biológica e onde se encontram as maioresextensões de terras não cultivadas, daí a preocupação decomo usar e conservar o trópico úmido em geral e a maiordessas áreas, a Amazônia em particular (Morán, 1990).Na região Amazônica, a biodiversidade é um elementofundamental, de caráter econômico, político e culturalpara as populações locais, pois muitas vezes elarepresenta a única fonte de recursos para a suasobrevivência (Lisboa, 2002).
Os rios de águas pretas, como o rio Negro, e terrasdrenadas por eles, apresentam um baixo potencial desubsistência no que diz respeito à utilização, sendochamados de “rios da fome” (Meggers, 1996). Áreasdrenadas por rios de águas pretas são pobres em pesca ecaça (Janzen, 1974). Nessas áreas de solos brancosarenosos e de origem sedimentar, a produtividadeprimária é baixa, limitada pela baixa disponibilidade de
&
nutrientes no solo (Klinge Ohle,1964). Essa condição tem sidoatribuída a diversos fatores, incluindoos baixos nutrientes dissolvidos e abaixa penetração da luz, que reduz aprodutividade primária. As águaspretas possuem baixo pH, permitindoque o CO e ácidos orgânicos decomplexos húmicos, provenientes dadecomposição da biomassa produzidapela floresta de igapó, atuem no graude acidez da água (Sioli, 1983).Segundo Janzen (1974), essa água decoloração escura, que é pobre emoxigênio dissolvido e material emsuspensão, torna-se inóspito ao peixe.Por isso, Fittkau Klinge (1973)consideram que rios de “águas pretas”podem fluir através de áreas pratica-mente privadas de vida animal.
Devido às pressões ambientaisresultantes nas regiões de águas pretase a essa baixa biomassa animal daregião, seria de se esperar algum graude desnutrição em populaçõesribeirinhas. Mas, as estratégias desobrevivência das populações têm tidoalto grau de sucesso, destacando-se oplantio de mandioca, técnicasespecializadas de pesca para lidar coma baixa produtividade, a organizaçãosocial que dispersa a população empovoados menores que em outrasregiões amazônicas e a distribuição dealimentos entre as famílias (Morán,1990).
Para a maioria das comunidadesamazônicas, a mandioca é a maiorfonte de recursos calóricos (Gross,
&
&
2
Terra Rebêlo&
enfermidades. A maioria dos animais foi utilizada por moradores das duas comunidadessendo que alguns tiveram uso restrito.
Recursos Animais, Reserva de Desenvolvimento Sustentável, AmazôniaCentral, EcologiaAplicada.PALAVRAS CHAVE:

O uso da fauna pelos moradores da Comunidade São João e Colônia Central
1975), mas uma dieta baseada emmandioca, por ser pobre em proteína,precisa ser suplementada com caça,pesca ou coleta (Meggers, 1996). Omaior recurso protéico para oshabitantes do rio Negro é o peixe e noalto rio Negro existem povoados queconsomem por dia duas vezes maispeixes do que caça (Clark, 1987).
Para as populações humanas daregião, a caça constitui uma importan-te fonte de proteína animal (Smith,1976). A persistência da pesca, da caçae coleta, juntamente com a agricultu-ra, entre as populações da Amazônia,sugere que a diversidade de modos desubsistência se combina de maneiraapropriada às suas condições políticas,demográficas e ambientais.
Nas margens do rio Negro, próximoà Manaus, existem comunidades quepossuem vínculo com a cidade e com orio, como a Comunidade São João doTupé e a Colônia Central. A proximida-de da RDS Tupé com a cidade de Manaustem causado um forte aumento daação antrópica através do turismo emesmo pelo crescimento urbano(BioTupé, 2003). Esta combinação defatores pede uma melhor compreensãodos recursos animais utilizados pelascomunidades contribuindo para oplanejamento da área. Este estudovisou identificar e conhecer asestratégias de subsistência de duascomunidades: uma que vive na terrafirme e outra que vive na beira do rio,para subsidiar o manejo da RDS emelhorar a qualidade de vida dosmoradores. O objetivo principal foicomparar os padrões de uso derecursos animais na comunidade SãoJoão e na Colônia Central através daestimativa da diversidade dos recursosanimais utilizados, do levantamentodas espécies de caça, pesca, animais
de uso medicinal e verificação do consumo de proteínaanimal pelos moradores das duas comunidades.
Na Comunidade São João vivem, aproximadamente,31 famílias às margens do lago Tupé e rio Negro. Pescamprincipalmente na seca e caçam pouco. A renda damaioria de seus moradores provém do trabalho na escolada comunidade, serviço público e turismo. Buscam partede seus alimentos em Manaus, principalmente noperíodo da cheia em que a pesca é escassa. A ColôniaCentral consiste em, aproximadamente, 24 famílias, quecostumam pescar nos Igarapés próximos, caçar paraconsumo e fazer roça. Suas rendas provêm de pequenaagricultura (roça) e serviços temporários em Manaus.
Para comparar os padrões de uso de recursos da faunaforam realizadas entrevistas com questionário semi-estruturado (Anexo 1), abordando: etnoespéciesutilizadas, tecnologias empregadas, locais de capturadaou coleta, categorias de uso, preferências alimentares,distribuição dos recursos com famílias vizinhas erecordação de refeições. As participações nasentrevistas foram livres e as respostas dos moradoresforam registradas em caderno de campo e/ou gravador.Foram recordadas refeições: do dia da entrevista e dodia anterior, no café da manhã, almoço e jantar. O caféda manhã não foi incluído na análise, pois segundoMatavele . (1995) no Brasil ele representa alimentoscom baixas calorias.
Os animais foram identificados pelos moradores dascomunidades São João e Central, com uso de materialbibliográfico ilustrado (Ferreira, 1998; Emmons Feer,1990; Amaral, 1978, De Schauensee Phelps, 1978). Osmoradores indicaram os animais que são caçados,pescados, ou que ocorrem na área, na reunião realizadano dia 08.11.2003, na sede da Associação dos Moradoresda Colônia Central (Barracão). As dúvidas sobre aidentificação de alguns animais citados nas entrevistastambém foram esclarecidas e os nomes populares foramassociados aos nomes científicos. Os animais de mais deuma espécie biológica, mas descritos pelos moradorescomo uma única etnoespécie, foram classificadas nonível de gênero. A identificação de etnoespécie queindicou espécies biológicas pertencentes a mais de umgênero foi classificada no nível de família, e assim
Material e Métodos
et al
&&
143

144
sucessivamente.Para estimar a diversidade dos recursos utilizados
pelos moradores, foram calculados os índices de Shannone Simpson (Brower ., 1997), através do programa PC-ORD, versão 4.0. Para comparar a diversidade entre ascomunidades foi feito teste t (Magurran, 1988) e paramedir a similaridade entre as comunidades, foi calculadoo coeficiente de similaridade de Jaccard. (Brower .,1997). Segundo Begossi (1996), índices de diversidadesão utilizados para avaliar intensidade de uso de recursospor populações humanas e permitem comparardiferentes populações em diferentes áreas.
De maio a novembro de 2003 foram entrevistados 17moradores na comunidade São João e 12 na ColôniaCentral, sendo um morador de cada família.
Foram identificadas 50 etnoespécies de animaisutilizados (Anexo 2) em 214 citações de animaispescados, caçados e de uso medicinal.
Os animais mais citados foram 6 peixes: jaraqui( spp.), tucunaré ( spp.), cará( ), traíra ( gr. ), matrinchã( ), pacu ( ); 4mamíferos: cutia ( ), paca (
), tatu ( ), e porcos do mato( spp.); 1 réptil: sucuriju ( ).
A comunidade São João usa mais animais que aColônia Central (40 e 37 etnoespécies, respectivamente,Tab. 1), mas a diferença entre as diversidades (H') não ésignificativa (t = 1,76; 0,1<p<0,05). Oscoeficientes de similaridade de Jaccard (CCJ = 0,54)indicam média similaridade entre os dois locais.
Na comunidade São João, 18 espécies foram citadasuma única vez. Nessa comunidade 16 espécies sãopescadas, 15 são utilizadas como medicinais (mamíferos,répteis, peixes, aves e abelhas Meliponinae) e 12 sãocaças de uso alimentar (mamíferos e aves). Paca, tatu ecutia são caçados e utilizados como alimento emedicinal. Nessa comunidade os animais mais citadosforam: jaraqui, tucunaré, sucuriju e tatu (Anexo 2).
et al
et al
Semaprochilodus CichlaCichlidae Hoplias malabaricusBrycon cephalus Mylossoma aureum
Dasyprocta agouti Agoutipaca Dasypus kappleriTayassu Eunectes murinus
Resultados
Diversidade de animais utilizados
0.05(2), 200
Na Colônia Central foram citados 37espécies de animais utilizados, sendo20 citados uma única vez. Do total, 15são pescados, 15 utilizados comomedicinais (mamíferos, répteis,peixes e aves) e 11 são caçados(mamíferos, aves e répteis). Paca,mucura (Didelphidae), jabuti (
spp.), capivara (), traíra e pirarara
( ) sãoutilizados como alimento e medicinal.Os animais mais citados foram ojaraqui pescado no lago, traírapescada nos igarapés e porcos do matocaçados na floresta e capoeiras.
O número de citações de animaisnas duas comunidades se ajusta aomodelo de distribuição da sérielogarítmica (“log series”), no qualexiste um pequeno número de espéciesabundantes e grande proporção deespécies “raras” (Magurran, 1988).(Fig. 1).
A categoria de uso que apresentoumaior diversidade de etnoespécies foia de animais pescados. (Fig. 2).
O declínio de animais de caça epesca foi percebido por 71% dosmoradores. Já o aumento de animaisfoi percebido por 9% dos moradores,mas 20% disseram que não hádiferenças, como visto nos relatos aseguir:
Geoche-lone HydrochaerishydrochaerisPhractocephalus hemioliopterus
“Antes era mais fácil. A populaçãoaumentou e caça fica mais arisca,distante. Não é o povo que temmatado”. Morador da ColôniaCentral.
“Ficaram mais espertos (a caça).Tem muito vestígio, mas não vê”.Morador da Colônia Central.
“A pesca diminuiu muito. O pessoalpescava muito de bomba. Às vezes
Terra Rebêlo&

motor de pesca e fazia arrastão. Adiminuição do igapó. O pessoalcorta e a erosão vai aterrando oslugares pro peixe ficar”. Morador daComunidade São João.
“A caça diminuiu, o peixe não. Opessoal se sustenta da caça. Elas seafastaram. Ou foram pra outraregião ou sumiu mesmo”. Moradorada Colônia Central.
“Quando chegamos as antasficavam próximas de onde a gentemorava. Tinha muita caça. Hojenão. Acabou porque a procura émuita. Não tem muito peixe, émuita gente pra tirar. O jeito é iratrás da caça. As pessoas têm quecriar mais”. Moradora da ColôniaCentral.
“Tem mais porque as pessoasdeixaram de caçar. Depois daspalestras procuramos outrasobrevivência. Não vamos maisatrás. Quando vê uma (caça) agente pega”. Moradora daComunidade São João.
“O macaquinho continua o mesmo.Peixe, aqui é difícil pegar umpeixe”. Moradora da ComunidadeSão João.
Pesca
A pesca nas Comunidades São Joãoe Central é realizada por 75% dasmulheres e 88,2% dos homens. Osmoradores que pescam utilizam essaatividade somente para consumopróprio e dividem o que pescam comfamílias vizinhas. Um único entrevista-do, da Comunidade São João, possuibarraca na praia do Tupé e compra emManaus o peixe que vende aos turistasnos finais de semana. Segundo estemorador existe pouco peixe na região,sendo difícil pescar para consumo
O uso da fauna pelos moradores da Comunidade São João e Colônia Central
São João Central Total
Diversidade
Riqueza de espécies (S) 40 37 50
Citações (N) 113 94 213
Simpson (D) 0,7655 0,831 0,875
Shannon (H) 1,679 1,899 1,811
Tabela 1. Diversidade de animais utilizados nas ComunidadesSão João e Central da RDS Tupé entre os meses de maio enovembro de 2003.
0
5
10
15
1 6 11 16 21 26 31 36 41 46 51 56 61 66 71 76
Espécie
Ab
un
dân
cia
Figura 1. Animais citados por etnoespécies nas ComunidadesSão João e Central na RDS Tupé.
1615
12
17
15
13
0
2
4
6
8
10
12
14
16
18
Pesca AnimaisMedicinais
Caça
Recursos utilizados
Nú
mero
de
etn
oesp
écie
s
São João
Central
Figura 2. Número de animais por categorias de uso, citadas pormoradores das Comunidades São João e Central da RDS Tupé,entre maio e novembro de 2003.
145

146
Caça
A caça na RDS Tupé ocorre commenos freqüência que a pesca, comoobservado nas bacias de água preta(Morán, 1990). É uma atividademasculina e praticada por 45% doshomens entrevistados. As mulheresnão caçam, mas relataram que aatividade é realizada pelo marido oucompanheiro. Em ambas comunida-des, todos os moradores que nãocaçam recebem parte da caça defamílias vizinhas.
Foram efetuados 49 registros deanimais caçados, que pertencem a 14espécies. As espécies registradas commaior freqüência foram os mamíferosterrestres como cutia (spp.), tatu ( spp.), paca( ) e porcos do mato( spp.). Entre as aves maiscitadas estão o jacu (
) e a nambu ( )(Tab. 3).
Os moradores das duas comunida-des não caçam nos mesmos locais. NaSão João, as atividades de caçacostumam ocorrer próximos às casas ena beira dos igapós. As caças maiscitadas são de menor porte, comocutia, tatu ( spp.), jacu( spp.) e tucano (spp.). Na Central, as caçadascostumam ocorrer em floresta de terrafirme e roçados e entre os animais maiscitados estão os animais de médio egrande porte como porcos do mato( spp.), paca ( ) eveado ( ). Algumasespécies como jabuti (spp.) e gavião ( ) foramcitadas exclusivamente por moradoresda Central e outras como jacu( ), macaco decheiro ( ), macaco
DasyproctaDasypus
Agouti pacaTayassu
Penelopejacquacu Tinamus major
DasypusPenelope Ramphastos
Tayassu Agouti pacaMazama americana
GeocheloneAccipitridae
Penelope jacquacuSaimiri sciureus
próprio e para comercializar nas barracas. Quatroentrevistados (dois da São João e dois da Colônia Central)não costumam pescar e consomem peixe quando trazidode Manaus ou dividido ou recebido de famílias vizinhas.
Os moradores da São João pescam de anzol (10citações), malhadeira (6 citações), tarrafa (5 citações),zagaia (3 citações), caniço (2 citações) e flecha (1citação). E os moradores da Colônia Central pescam deanzol (7 citações), tarrafa (4 citações), malhadeira (3citações), caniço (1 citação) e zagaia (1 citação).
Na São João, 89% dos pescadores pescam no lagoTupé, sendo 62,3% em água aberta e 27,7% no igapó. Osoutros 11% pescam no igarapé da Terra Preta.
Na Colônia Central 58% dos pescadores pescam nosigarapés do Caniço, do Pavão, do Julião e em demaisigarapés próximos. Os outros 42% pescam no lago Tupé.
Foram identificadas 22 etnoespécies de peixes, quecorrespondem a aproximadamente 40 espéciesbiológicas. Duas etnoespécies, o jaraqui e o tucunaréforam as espécies de peixes mais citadas (Tab. 2). Ojaraqui é o peixe preferido de 52% dos moradores. Asimilaridade entre as comunidades, em relação à pesca émediana (CC = 0,5).
A sazonalidade da pesca é conhecida e relatada por95% dos pescadores nas entrevistas. Na estação chuvosa,o rio enche e a disponibilidade de peixes é menor. Já naestação seca, quando o rio vaza, a disponibilidade depeixes aumenta. Na Comunidade São João os peixes maiscitados na seca foram tucunaré, jaraqui, pacu e aracu ena cheia jaraqui, pacu, tucunaré e piranha. Na ColôniaCentral os mais citados na seca são outros peixes, como ocará, a traíra, o jeju e o jaraqui (único peixe que tambémé mais citado na São João), e na cheia a traíra, omatrinchã, o cará e o jeju.
O número de espécies taxonômicas é bem maior que ode etnoespécies. Por exemplo, os aracus (Anostomidae),que foram incluídos nessa análise em uma únicaetnoespécie, pertencem a oito espécies taxonômicasdiferentes. Os carás (Cichlidae), também foramanalisados como uma única etnoespécie, pertencem a 3gêneros e 6 espécies taxonômicas diferentes. Sãonecessários estudos adicionais para podermos afirmarquais espécies são efetivamente utilizadas pelaspopulações locais.
J
Terra Rebêlo&

prego ( ), onça () e mucura ( ) pelos
moradores da São João. A similaridadeentre as comunidades, em relação àcaça é mediana (CC = 0,5).
Os moradores relataram que ascaçadas podem ser intencionais (deespera ou à ponto) ou oportunísticas. Ouso de espingardas para captura daspresas foi citado por 100% doscaçadores. Apenas dois entrevistadoscitaram outros instrumentos de caça,como o rifle 44 (de repetição) e o“buboque” (armadilha de toco).
As atividades de caça têm grandeimportância no inverno, já que nesseperíodo a oferta do pescado é menor.Essa sazonalidade é relatada peloscaçadores, e para 55% deles existemdiferenças entre a abundância dacaça, nas diferentes estações do ano,como pode observado nos relatosabaixo:
Nas entrevistas foram citadas 23espécies de uso medicinal. Quanto àdistribuição taxonômica: 57% répteis,33% mamíferos, 6% peixes, 6%invertebrados e 4% aves. A cobrasucuriju ( ), o jacaré( ), a jibóia (
Cebus apella Panteraonca Didelphidae
Eunectes murinusAlligatoridae Boa
J
“Na chuva é melhor que na seca. Nachuva os animais saem pra comer.Tem mais fruto”. Morador dacolônia Central.
“Tem mais na enchente. Na secaquase não dá”. Moradora daComunidade São João.
“Seca a caçada é mais intensa. Émais fácil sentir o animalchegando”. Morador da ColôniaCentral.
Animais de uso medicinal
O uso da fauna pelos moradores da Comunidade São João e Colônia Central
Nome popular Nome científico São João Central Total
Jaraqui Semaprochilodus spp.(1) 14 9 23
Tucunaré Cichla spp. 12 5 17
Cará Cichlidae (3) 7 6 13
Matrinchã Brycon cephalus 5 3 8
Pacu Mylossoma aureum 5 3 8
Traíra Hoplias gr. malabaricus 1 7 8
Aracu Anostomidae (4) 6 0 6
Jandiá Leiarius marmoratus 2 1 3
Jeju Hoplerythrinus unitaeniatus 0 3 3
Piranha Serrasalmidae (5) 2 1 3
Tambaqui Colossoma macropomum 1 2 3
Curimatã Prochilodus nigricans 1 1 2
Jacundá Crenicichla spp. (6) 1 1 2
Orana Hemiodus ocellatus 2 0 2
Pacu-galo Myleus schomburgki 2 0 2
Sorubim Pseudoplatystoma tigrinum 2 0 2
Bararua Uaru amphiacanthoides 1 0 1
Bodó Liposarcus pardalis 0 1 1
Jatuarana Brycon cephalus 0 1 1
Mapará Hypophthalmus spp. (7) 0 1 1
Peixe liso Siluriforme 0 1 1
Sardinha Triportheus spp. (8) 0 1 1
Total 64 47 111
s=22
(1)(2)(3) sp
(4) aff
(5)
(6) sp(7)
S. taeniurus, S. insignis
C. monoculus, C. temensis
Caquetaia spectabilis, Heros ., Cichlasoma amazonarum,
Aequidens tetramerus, Acaronia nassa, Chaetobranchus
flavescens.
Rhytiodus argenteofocus, R. microlepis, Leporinus fasciatus, L. .
affinis, L. friderici. L. trifasciatus, Schizodon vittatum, S. fasciatum.
S. elongatus, S. Aff. ligenmanni, S. spilopleura, S. rhombeus,
Catoprion mento.
C. reticulata, C. Aff. ornata,Catropion .
H. edenatus, H. marginatus. T. flavus, T. Albus
Tabela 2. Peixes citados pelos moradores das Comunidades SãoJoão e Central da RDS Tupé, entre os meses de maio enovembro de 2003.
147

148
constrictor) foram os animais mais citados. Na São João,o animal mais citado foi a sucuriju (27,5%) e na ColôniaCentral a sucuriju (16,5%) e a jibóia (16,5%). Asimilaridade entre as comunidades é baixa (CC = 0,36).
A banha foi a porção animal mais citada (68%).Também houve relatos do uso de dentes, cabeças, fel(bile), línguas, rabos e urina. As banhas e óleos sãogeralmente usadas topicamente em fricções, pura oumisturada a produtos industrializados (sebo de Holanda).De 13% dos animais citados foram aproveitadas mais deuma fração para a produção de diferentes remédios.Como exemplo, da mucura utiliza-se o fel para cólica demulheres grávidas e a língua no tratamento da asma.
Foram relatados 15 problemas de saúde ouenfermidades, para os quais frações de animais sãoutilizadas no tratamento. Dos animais citados, 50%foram usados no tratamento de mais de uma doença. Asindicações mais comuns de uso de animais medicinais são
J
cicatrizações, doenças de pulmão,ouvido e fígado. Como exemplo, abanha da sucuriju ( )e jibóia são indicadas comocicatrizantes; as banhas de boto (
) e guariba () são usadas para asma; o
rabo do tatu ( spp.) Dente dejacaré ( ) foi citado comoamuleto para afastar cobras (Tab. 4).
Existe relação entre os animais deuso medicinal e alimentar. Dois peixestambém foram mencionados comosendo medicinais e a criaçãodoméstica de galinhas fornecealimento e remédio (Tab. 4). Algumasdas espécies mais citadas, comocobras aquáticas e jacarés, existemem baixas densidades na área, mas ouso medicinal consome pequenasquantidades de banha cada vez. Nãohá caça apenas para fins medicinais,pois a grande maioria dos produtosutilizados é sub-produto da caça parafins alimentares.
Em 56 refeições recordadas foramconsumidos 10 itens alimentares nãoprotéicos (arroz, feijão, macarrão,farofa, farinha, cupuaçu, maionese,pão, batata e cará), e cinco itensprotéicos de origem animal (peixe,galinha/frango, carne de gado, caça eovos) (Tab.5).
Do total de alimentos citados, 57%não eram proteína animal. Osalimentos mais consumidos foram;arroz, café, peixe, feijão, galinha/frango e os produtos derivados damandioca (farinha, tapioca, beiju, péde moleque e chibé). Galinha e ovosprovêm de criação doméstica naColônia Central, enquanto na SãoJoão, esses produtos são comprados
Eunectes murinus
Iniageoffrensis Alouattaseniculus
DasypusAlligatoridae
Consumo de proteína animal
Tabela 3. Caças citadas pelos moradores das Comunidades SãoJoão e Central da RDS Tupé, entre os meses de maio enovembro de 2003.
Nome popular Nome científico São João Central Total
Cutia Dasyprocta agouti 7 5 12
Porco do mato Tayassu spp. (1) 1 7 8
Tatu Dasypus kappleri 5 1 6
Paca Agouti paca 1 4 5
Veado Mazama americana 1 3 4
Jacu Penelope jacquacu 3 0 3
Tucano Ramphastos spp. (2) 2 1 3
Jabuti Geochelone spp. 0 2 2
Nambu Tinamus major 1 1 2
Gavião Accipitridae 0 1 1
Macaco de cheiro Saimiri sciureus 1 0 1
Macaco prego Cebus apella 1 0 1
Onça Pantera onca 1 0 1
Mucura Didelphidae (3) 1 0 1
Total 25 25 50
s = 14
(1)(2)(3)
T. pecari, T, tajacu.
R. ambiguus, R. vitellinus, R. culminatus, R. tucanus.
Didelphis marsupiali, Marmosops incanus.
Terra Rebêlo&

O uso da fauna pelos moradores da Comunidade São João e Colônia Central
No
me
po
pu
lar
No
me
cie
ntí
fic
oP
art
eu
tili
za
da
Us
oS
ão
Jo
ão
Ce
ntr
al
To
tal
Su
curiju
Eu
ne
cte
sm
urin
us
ba
nh
aci
catr
iza
nte
,cu
rag
olp
e8
41
2
Jaca
réA
LL
IGA
TO
RID
AE
(1)
ba
nh
a(1
),d
en
te(2
)(1
)asm
a,
pe
rna
sfr
aca
s,cr
ian
çaq
ue
nã
oa
nd
alo
go
,(2
)afa
sta
cob
ra2
35
Jib
óia
Bo
aco
nstr
icto
rb
an
ha
sara
go
lpe
,in
cha
ço,a
sma
,re
um
atis
mo
,p
ere
bin
ha
,e
nfe
rmid
ad
es
14
5
Pa
caA
go
uti
pa
ca
fel
pro
teto
rp
ara
ma
lária
,fí
ga
do
22
4
Jan
da
íra
Me
lip
on
a.
spp
me
lu
sad
oju
nto
com
tre
voro
xo,
cop
aíb
a,
an
diro
ba
30
3
Jacu
raru
Tu
pin
am
bis
nig
rop
un
cta
tus
ba
nh
ain
flam
açã
o,
do
rd
ed
en
te,
do
rd
eo
uvi
do
12
3
Mu
cura
Did
elp
hid
ae
ma
rsu
ialis
fel(1
),lín
gu
a(2
)(1
)m
ulh
er
grá
vid
añ
sen
tird
or,
(2)
asm
a2
13
Tatu
15
Kg
Da
syp
us
ka
pp
leri
rab
o(1
),b
an
ha
(2)
(1,2
)d
or
de
ou
vid
o3
03
Ga
linh
aG
allu
sg
allu
sb
an
ha
inch
aço
,n
ariz
en
tup
ido
11
2
Tart
aru
ga
Po
do
cn
em
ise
xp
an
sa
ba
nh
acr
em
e,
reu
ma
tism
o1
12
Bo
toro
saIn
iag
eo
ffre
nsis
ba
nh
aa
sma
10
1
Ca
lan
go
TE
IID
AE
cab
eça
asm
a1
01
Ca
piv
ara
Hyd
roch
ae
ris
hyd
roch
ae
ris
ba
nh
are
um
atis
mo
01
1
Ca
rne
iro
Ovis
.sp
seb
oin
cha
ço1
01
Cu
tiaD
asyp
rocta
ag
ou
tiu
rin
ap
ara
rd
eb
eb
er
cach
aça
10
1
Gu
arib
aA
lou
att
ase
nic
ulu
sb
an
ha
asm
a0
11
Jab
uti
Ge
och
elo
ne
spp
.b
an
ha
varize
s0
11
On
çaP
an
tera
on
ca
ba
nh
aa
sma
01
1
Pira
rara
Ph
racto
ce
ph
alu
sh
em
iolio
pte
rus
ba
nh
afe
rid
a,
infla
ma
ção
01
1
Po
raq
uê
Ele
ctr
op
ho
ruse
letr
icu
sb
an
ha
reu
ma
tism
o1
01
Po
rcã
oTa
ya
ssu
pe
ca
rip
resa
pn
eu
mo
nia
,d
err
am
e0
11
Tra
íra
Ho
plia
s.
ma
lab
aricu
sg
rb
an
ha
do
rd
eo
uvi
do
,in
cha
ço0
11
Tota
l2
92
55
4
s=
23
(1)
Caim
an
cro
codilus,P
ale
osucos
palp
ebro
sos,M
ela
nosucos
nig
er
Tabe
la4.
Ani
mai
sde
uso
med
icin
al, s
uasfr
açõe
se
usos
cita
dospe
losm
orad
ores
dasCom
unid
ades
São
João
eCen
tral
daRD
STu
pé.
149

150
Alimento São João Central Total
Arroz 11 12 23
Peixe 8 10 18
Feijão 9 8 17
Galinha / frango 8 6 14
Carne gado 4 7 11
Produtos Mandioca 1 8 9
Frutas 1 5 6
Macarrão 3 2 5
Conserva 2 2 4
Batata 1 1 2
Cará 0 2 2
Caça 0 1 1
Cuz-cuz 0 1 1
Garapa 0 1 1
Ovo 1 0 1
Total 49 66 115
s =15
Tabela 5. Alimentos consumidos pelos moradores dasComunidades São João e Central na RDS Tupé, em 56 refeiçõesamostradas, entre maio e novembro de 2003.
37%
29%
22%
8%
2%
2%
peixe
galinha
carne gado
conserva
caça
ovo
Figura 3. Porcentagem do consumo de proteína animal, em 56refeições amostradas nas Comunidades São João e Central daRDS Tupé entre maio e novembro de 2003.
Terra Rebêlo&
em Manaus. A carne de boi, a conservae o leite também são adquiridos nacidade.
Os alimentos de origem animal maisconsumidos foram os peixes (37 %),frango/galinha (29%) e carne de gado(22%). (Fig. 3). Em 14% das refeiçõesnão foi consumida proteína animal.
A diversidade de alimentos citadosvariou entre os períodos de seca echeia. Na cheia a diversidade de itensprotéicos consumidos foi maior. (Tab.6).
A escassez da pesca na cheia deveser o principal motivo para o aumentono consumo de conserva, ovos e frangoque são trazidos da cidade. O númerode itens sem proteína animal tambémfoi maior na cheia. Do total dealimentos consumidos por moradoresda São João, 38% são trazidos dacidade. Já os moradores da ColôniaCentral consumiram 23% dos alimentostrazidos de Manaus. Em comunidadesdo médio Rio Negro, Silva (2003)sugere que a urbanização promoveu asubstituição do peixe por frango eembutidos, principalmente no períododa cheia.
Neste estudo são analisadassemelhanças e diferenças na utilizaçãode animais pelas Comunidades SãoJoão e Colônia Central e suasestratégias de subsistência.
Considerando as amostras representativas as comunidades São João eColônia Central utilizam aproximadamente o mesmo número de animaiscomo recurso.A riqueza de espécies (s)é maior na São João e a diversidade deanimais utilizados (H) é maior na
Discussão eConclusões
-
-

O uso da fauna pelos moradores da Comunidade São João e Colônia Central
Alimento Cheia Seca
caça 2,17 0,00
carne gado 4,35 11,39
conserva 4,35 2,53
galinha / frango 15,22 8,86
ovo 2,17 0,00
peixe 6,52 17,72
sem proteína animal 65,22 59,49
número de itens 46 79
refeições amostradas 24 32
Índice de diversidade (Simpson) 0,7344 0,677
Tabela 6. Porcentagem de alimentos com proteína animalconsumidos por estação, nas Comunidades São João e ColôniaCentral, em 56 refeições amostradas, entre maio e novembrode 2003.
151
Colônia Central, mas essas diferençasnão são significativas (t 200 =1,76; 0,1<p<0,05).
Apesar das comunidades São João eColônia Central, estarem localizadasem ambientes diferentes (lago e terrafirme) e terem diferentes fontes derenda (serviços públicos e atividadesde roça), a diversidade de animaisutilizados é muito similar. As diferen-ças entre as comunidades são algumasespécies de pesca, caça e medicinais,que são mais, ou até exclusivamentecapturadas por moradores da São Joãoe outras por moradores da Central. Osanimais mais utilizados na São João sãopeixes (jaraqui e tucunaré) animaisque vivem em ambientes semi-aquáticos (sucuriju) ou áreas perturba-das (tatu). Na Central, são animais decaça de maior porte (porcos do mato) epeixes de igarapé (traíra e cará) queestão entre os mais utilizados.
Os pescadores da São João eCentral compartilham o mesmo localde pesca (lago Tupé), por isso váriasespécies são pescadas em comumpelas duas comunidades (jaraqui,tucunaré, cará, matrinchã, pacu,traíra, jandiá, piranha, tambaqui,curimatã e jacundá), mas tambémpescam em locais exclusivos. Como ospescadores da São João pescam noigarapé da Terra Preta, e os pescadoresda Colônia Central, nos igarapés doJulião, Caniço, Pavão e outros igarapéspróximos, alguns peixes são maispescados exclusivamente por morado-res da São João (aracu, orana, pacu-galo, sorubim, bararuá e bodó) eoutras pelos moradores da ColôniaCentral (jeju, bodó, jatuarana,mapará, peixe liso e sardinha).
A caça ocorre com menosfreqüência que a pesca, como nasbacias de água preta (Morán, 1990). Os
0.05(2),
moradores das comunidades São João e Colônia Centralnão caçam nos mesmos locais, mas existem caçascomuns entre as duas comunidades (cutia, porco domato, tatu, paca, veado, tucano e nambu). Alguns forammais caçados por moradores da Colônia Central (jabuti egavião) e outros pelos moradores da São João (jacu,macaco de cheiro, macaco prego, onça e mucura).
Nas duas comunidades, todos os moradores que nãocaçam recebem uma parte da caça de famílias vizinhas.Adivisão de alimentos é uma prática comum entrecomunidades daAmazônia (Murrieta , 1999).
O conhecimento sobre animais de uso medicinalparece ser muito difundido por toda Bacia Amazônica(Branch, 1983; Pinto Maduro, 2003). Frações deanimais têm valor na medicina popular da RDS Tupé, esão produtos da fauna local e de criação doméstica. Osrépteis são de grande importância para as ComunidadesSão João e Central como recurso medicinal.
Nas duas comunidades é utilizada a mesma riqueza deanimais medicinais, mas com pequena similaridade. Otatu, a capivara, o jabuti, a onça, a pirarara, o porco e a
et al.
&

152
Terra Rebêlo&
traíra só foram citados por moradores da ColôniaCentral, e são espécies locais. Enquanto na São João, dosanimais medicinais citados exclusivamente, trêsanimais são de criação, como a jandaíra, galinha ecarneiro e três são de rio, como a tartaruga, o boto rosa eo poraque.
A banha é a fração animal medicinal mais comum eseu uso está relacionado ao tratamento de doenças demaior incidência na região, como cortes, doença depulmão e fígado.
De acordo com as entrevistas realizadas, existe maiornúmero de caçadores na Colônia Central. A densidadedos animais caçados pode ser maior e por isso o consumode animais de caça pode ser maior nessa comunidade,apesar de não aparentar nas refeições.
O peixe é o alimento que mais contribui para oconsumo protéico, sendo que na São João a riqueza depeixes citados foi maior. Como a área de pesca no LagoTupé e igapós é maior que a dos igarapés, adisponibilidade e quantidade de peixes para osmoradores da São João podem ser maiores, contribuindopara um maior consumo em relação à Colônia Central.Nesta comunidade, além da pesca (realizadaprincipalmente nos igarapés), carne de gado e galinhacontribuem mais para o consumo protéico do que a caça.Provavelmente por ser considerada uma atividadeclandestina o consumo de caça foi sub-amostrado narecordação de refeições.
Há consumo freqüente de galinha, frango e carne devaca sugerindo que a dependência dos animais de caçalocais para o consumo protéico é relativamente baixa nasduas comunidades, se comparada a outras populaçõeslocais (Hardesty, 1975). Isto pode ser atribuído aproximidade da RDS com a cidade de Manaus, para onde90% maioria dos entrevistados das duas comunidades sedeslocam pelo menos uma vez por mês.
A variação sazonal no nível do rio afeta mais a pescaque a caça. Na cheia, os peixes se dispersam na áreaalagada e a pesca é mais escassa. O consumo de proteínadiminuiu neste período, mesmo com a compra de frangoe conserva na cidade de Manaus. Esta dificuldade de seobter proteína é comum em rios de água preta, de baixaprodutividade e captura de peixes muito pequenos(Clark Uhl, 1987).
A se julgar pela riqueza e diversidade, ascomunidades São João e Colônia Central utilizam os
&
recursos animais de maneira um poucodiferente, mas essas diferenças nãosão estatisticamente significativas. Opresente estudo sobre consumo dealimentos indica que os moradoresutilizam estratégias que exploram asáreas ao redor da comunidade, mastambém buscam recursos vindos defora.
Agradecemos a todos os moradoresdas comunidades São João e Central,que de alguma forma colaboraram comnosso trabalho, e em especial à Luzia,Bida, Ceará, Araújo, Pedrinho,Domingos, Jaumir, Pepe, Arlindo, Lulu,Hélio, Maricleide, Valente, Socorro,Ana, Lavínia, Joel, Joelma, Cobrão eManoel.
Amaral, A.do. 1978..
Editora da Universidade de SãoPaulo, São Paulo, SP. 246 pp.
Begossi, A. 1996. Use of ecologicalmethods in ethnobotany: diversityindices. . NewYork Botanical Garden, Bronx, NY,50 (3): 280-289.
BioTupé. . 2003.D i s p o n í v e l e m :http://biotupe.inpa.gov.br./.Acesso em 10/07/03.
Branch, L.C.; Silva, M.F. da. 1983. Folkmedicine of Ater do Chão, Pará,Brasil. , 13 (5-6):
Agradecimentos
Bibliografia Citada
Serpentes doBrasil: iconografia colorida
Economic Botany
Projeto BioTupé
Acta Amazonica

O uso da fauna pelos moradores da Comunidade São João e Colônia Central
153
737-797.Brower, J.H.; Zar, C.N.; Von Ende.C.N.
1997.. The
McGraw-Hill Companies, UnitedStates ofAmerica. 273 pp.
Cas t ro , E . 2000 . Ter r i tó r io ,biodiversidade e saberes depopulações tradicionais.Diegues, A.C.
. Hucitec, SãoPaulo, SP. p. 164-182.
Clark, K.; Uhl,C. 1987. Farming,fishing and fire in the history of theupper Rio Negro Region ofVenezuela. , 15(1):1-26.
De Schauensee, R.M.; Phelps, W.H.;Tudor, G. 1978. A guide to the birdsof Venezuela. Princeton UniversityPress, Princeton, New Jersey. 424pp.
Diegues, A. C. 1996.. Hucitec, São
Paulo, SP.169 pp.Emmons, L.H.; Feer, F. 1990.
. The University ofChicago Press, Chicago. 281 pp.
Ferreira, E.J.G. 1998.
. EdiçõesIBAMA, Brasília. 383 pp.
Fitkau, E.J.; Klinge, H. 1973. Onbiomass and tropic structure of theCentral Amazonian rain forestecosystem. 5 (1): 2-14.
Gross, D.R. 1975. Protein capture andcultural development in the
Field and laboratorymethods for general ecology
In:Etnoconservação:
novos rumos para a proteção danatureza nos trópicos
Human Ecology
O mito modernoda natureza intocada
Neotropical rainforest mammals: afield guide
Peixescomerciais do médio Amazonas:região de Santarém, Pará
Biotropicamm,
Amazon Basin. 77: 527-549.
Hardesty, D.L. 1975. The Niche Concept: Suggestions ofIts use in Human Ecology. 3 (2): 71-85.
Janzen, D.H. 1974. Topical Blackwater river, animals andmast fruiting by the Dipterocarpaceae. 6(2): 69-103.
Klinge, H.; Ohle, W. 1994. Chemical properties of riversin the Amazonian area in relation to soil conditions.
XV: 1067-1076.Lisboa, P.L.B. 2002.
. Museu Paranaense Emílio Goeldi, Belém, PA.237pp.
Magurran, A.E. 1988.. University Press, Cambridge. 179 pp.
Matavele, J.E.; Begossi, A.; Habib, M.M. 1995. Animalprotein consumption in a brasilian slum (VilaNogueira, Campinas, SP). ,4(2/3): 173-182.
Meggers, B.J. 1996.. Smithsonian Institution Press,
London. 214 pp.Moran, E. F. 1990.
. Vozes, Petrópolis, RJ. 367p .Murrieta, R.S.S.; Dufour, D.L.; Siqueira, A.D. 1999. Food
consumption and subsistence in three caboclopopulation on Marajó Island, Amazonia, Brazil.
, 27 (3): 455-475.Pinto, A. A. C.; Maduro, C.B. 2003. Produtos e
subprodutos da medicina popular comercializados nacidade de Boa Vista, Roraima. , 33(2): 281- 290.
Rebêlo, G.H.; Galatti. 1995.. CTA, Rio Branco,AC. 48 p .
Sioli, H. 1983.. Vozes, Petrópolis, RJ. 72 p .
Smith, N. J. H. 1976. Utilization of game along Brasil'stransamazon Highway. , 6(4): 455456.
American Anthropologist,
Human Ecology,
Biotropica,
Verh. Internat. Verein. Limnol,Natureza, homem e manejo de
recursos naturais na região de Caxiuanã, Melgaro,Pará
Ecological Diversity ans ItsMeasurements
Journal of Human Ecology
Amazônia: man and culture in acounterfeit paradise
A ecologia humana das populações daAmazônia
Human Ecology
Acta Amazonica
Manejo de fauna emreservas extrativistas
Fundamentos da ecologia da maior regiãode florestas tropicais
Acta Amazonica -
p
p
p


Diversidade BiológicaCapítulo 11
Estrutura fitossociológica de umafloresta de terra firme na AmazôniaCentral
RESUMO - Foi amostrada a estrutura arbórea de 0,5ha de uma floresta de terra firme naReserva de Desenvolvimento Sustentável do Tupé (RDS Tupé), localizada a oeste deManaus, distante aproximadamente 25km em linha reta do centro da cidade (03°02'35"S e60°15'18"W). O trabalho teve por objetivo inventariar, identificar e quantificar adiversidade de espécies arbóreas e lianescentes de uma área de terra firme na RDS Tupé.Foi demarcada uma parcela de 100x50m, dividida em 50 parcelas contíguas. Dentro dasparcelas foram amostrados todos os indivíduos arbóreos e lianescentes com o PAP 30cm,excluindo as palmeiras. Os indivíduos foram plaqueados, seu perímetro mensurado e alturaestimada. Foram amostrados 287 indivíduos distribuídos em 118 morfoespécies,pertencentes a 31 famílias. Dessas, 39% (46) foram identificadas até o nível de espécie;55,08% (65) até o nível de gênero e 4,23% (5) até o nível de família; 1,69% (2)permaneceram indeterminadas. As famílias com maior número de espécies foram:
>
Karen Souza DINIZ1
Veridiana Vizoni SCUDELLER2
e-mail: [email protected] do curso de Licenciatura em Ciências Biológicasdo Centro Universitário Luterano de Manaus (Ceulm/Ulbra)
e-mail: [email protected] em Biotecnologia e Recursos Naturais - UEA e Coordenação de Biologia - Ceulm/Ulbra.
Av. Solimões, 2 Japiim II. Manaus AM.
1
2
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

156
Introdução
A floresta Amazônica é o maior reservatório naturalda diversidade vegetal do planeta, onde cada um de seusdiferentes ambientes florestais possui um contingenteflorístico rico e variado, muitas vezes exclusivo dedeterminado ambiente. As múltiplas interrelações entreseus componentes bióticos e abióticos formam umconjunto de ecossistemas altamente complexos e deequilíbrio ecológico extremamente frágil (OliveiraAmaral, 2004). Segundo Ribeiro . (1999), em cadahectare de floresta existem aproximadamente 300espécies de árvores com mais de 10 cm de diâmetro àaltura do peito (DAP). O conhecimento sobre florestastropicais tem papel fundamental na elaboração deestratégias mundiais para a conservação dabiodiversidade (Lima Guedes-Bruni, 1997).
A análise da estrutura da floresta, ou fitossociologia,ainda é a técnica mais utilizada para se fazer deduçõessobre a origem, características ecológicas esinecológicas, dinâmica e tendência sobre seu futurodesenvolvimento (Ribeiro ., 1999). Para Higuchi
. (1982), o inventário florestal é a primeira etapa parapesquisas referentes à recursos naturais e também paraa tomada de qualquer decisão relacionada ao uso daterra. Segundo Martins (1989), o estudo fitossociológicode uma comunidade vegetal revela as interrelações dasespécies no espaço e no tempo.
Levantamentos estritamente florísticos são muitoúteis para a análise inicial da vegetação de umadeterminada área, pois permitem comparações amplascom um grande número de outros trabalhos. Entretanto,a inclusão de medidas de abundância são necessáriasquando se pretende detalhar as comparações entrevegetações de diferentes áreas, principalmente entre
&
&
et al
et al etal
áreas floristicamente semelhantes(Causton, 1988; Van den Berg, 1995).Tais medidas são interessantestambém, pois provem o conhecimentoda estrutura da vegetação estudada,propiciam subsídios para o manejodesta e constituem a base teórica pararecuperação de áreas similares (Vilela
., 1993; Van den Berg, 1995;Scudeller Martins, 2003). SegundoOliveira Amaral (2004), o estudofitossociológico e florístico dasflorestas de terra firme da regiãoamazônica são essenciais para aconservação de sua elevada diversida-de.
O presente trabalho tem comoobjetivos inventariar, identificar equantificar a diversidade de espéciesarbóreas e lianescentes de uma áreade terra firme na Reserva deDesenvolvimento Sustentável do Tupé(RDS Tupé).
A parcela foi instalada na trilha daCachoeira, localizada na margemdireita do lago Tupé com aproximada-mente 3.800m de extensão (Scudeller
., 2005, neste volume) A áreaselecionada para a instalação daparcela fica a 300 m do ponto inicial datrilha e a 20m em linha reta para ointerior da mata.
et al
et al
&&
Materiais e Métodos
Diniz Scudeller&
Sapotaceae (24 espécies), Caesalpinaceae, Burseraceae e Chrysobalanaceae (9 cada) eMoraceae (8). Essas cinco famílias representam 52,1 % da riqueza local. A densidade totalfoi de 556 ind/ha e a área basal por hectare foi de 25,18m . A média da altura total foi de17,27m (variando de 6 a 40m), do perímetro foi de 65,14cm (30 a 910cm) e o índice dediversidade de Shannon Wiener (H') para espécie foi de 4,214 nats.indivíduos .
Amazônia, fitossociologia, terra firme, Reserva de DesenvolvimentoSustentável do Tupé, estrutura arbórea.
2
-1&
PALAVRAS-CHAVE:

Foi demarcada uma área de100x50m, dividida em 50 parcelascontíguas. Todas as árvores e lianascom PAP (perímetro à altura do peito,ou seja, a aproximadamente 1,3m dosolo) 30cm foram marcadas complaquetas de alumínio numeradas emordem crescente e a sua altura total foiestimada. As amostras botânicas,férteis (flores e/ou frutos) ou estéreis,foram coletadas e submetidas aoprocesso de herborização, seguindo asnormas propostas por Mori .(1989). Também foram anotadascaracterísticas da casca e do alburno(por exemplo: cor, resistência, seiva eodor) com o intuito de auxiliar nadeterminação taxonômica.
As amostras foram identificadasatravés de comparação com as demaisexsicatas do Herbarium G.T. Prance(HGTP) do Ceulm/Ulbra coleção doProjeto Biotupé e do herbário doInstituto Nacional de Pesquisa daAmazônia (INPA), além de consulta abibliografia especializada (e.g. Nee,1995; Gentry, 1996; Ribeiro .,1999). As amostras que não puderamser identificadas ao nível específicoreceberam o código de morfotipo.
Após a identificação, as exsicatasde material estéril foram organizadaspor família e separadas em pastas nacoleção testemunho se utilizandoordem alfabética. O material botânicofértil coletado foi incorporado nacoleção do HGTP, com duplicata noINPA.
Neste estudo, foi adotado acircunscrição proposta por Cronquist(1981), com exceção das Leguminosae,que foram tratadas de acordo comPolhill . (1981). A confirmação dagrafia correta e dos autores dasespécies foi obtida consultando oInternational Plant Names Index
>
et al
et al
et al
(www.ipni.org).Segundo Martins (1989), a estrutura da vegetação é
definida por três componentes, disposição vertical dasespécies (estratificação), disposição horizontal(distribuição espacial) e a abundância de cada espécie.Desse modo, a análise fitossociológica estrutural tevecomo base a dominância absoluta (DoAs = ABs.U/A) e arelativa (DoRs =ABs/ABt.100); a freqüência absoluta (FAs= 100.nAs/nAt) e a relativa (FR = 100.FAs/ áreabasal das espécies (ABs = área basal total (ABt= órmulas citadas porMeira Neto (1991, 1997), Damasceno (1997). O valor deimportância das espécies (VIs = DRs + FRs + DOs) e o decobertura da espécie (VCs = DRs + DoRs), foramcalculados segundo Muller-Dombois Ellenberg (1974) eMartins (1991).
Foram preparadas distribuições de densidade deárvores por classe de perímetro e altura e empregadosintervalos de classe com amplitudes crescentes paracompensar o forte decréscimo da densidade nas classesde tamanho maiores, típico da distribuição em “J”invertido.
Para a determinação da diversidade de espécies foicalculado o Índice de diversidade de Shannon (H'), pelafórmula H'= - úmero deindivíduos da espécie i e N=número total de indivíduosamostrados (Magurran, 1988).
Foram amostrados 287 indivíduos distribuídos em 118morfoespécies, pertencentes a 31 famílias (Tab. 1).Dessas 39% (46) foram identificadas até o nível deespécie, 55,08% (65) até o nível de gênero e 4,23% (5) atéo nível de família; 1,69% (2) permanecemindeterminadas. As famílias com maior número deespécies foram: Sapotaceae (24 espécies),Caesalpinaceae, Burseraceae e Chrysobalanaceae (9cada) e Moraceae (8). Essas cinco famílias representam52,1% da riqueza local.
As famílias mais importantes, em ordem decrescentequanto ao número de indivíduos, foram: Burseraceae
∑
∑
∑
∑
FAt), a
p /4
ABs), foram calculados com as f
(pi)(logpi), onde pi=(ni/N, n=n
2 π), a
&
Resultados e Discussão
Riqueza de espécies
Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
157

158
Diniz Scudeller&
Tabela 1. Relação das famílias e espécies arbóreas e lianescentes presentes na floresta de terra firme da Trilhada Cachoeira (RDS Tupé), com os respectivos nomes populares e hábito.A= arbóreo; L= lianescente.
continua >
FAMÍLIA ESPÉCIE NOME POPULAR HÁBITOANACARDIACEAE Anacardium giganteum Hanck ex Engl. cajuí AANNONACEAE Guatteria foliosa Benth. envireira A
Guatteria sp.1 ─ AGuatteria sp.2 ─ AGuatteria sp.3 envira cascuda AGuatteria sp.4 envira fofa AXylopia crinita R. E. Fr. parviflora AXylopia sp. envira vassourinha A
APOCYNACEAE Aspidosperma sp. pau marfim ABOMBACACEAE Scleronema micranthum Ducke cardeiro ABURSERACEAE Protium apiculatum Swartz. breu vermelho A
Protium sp.1 breu de leite AProtium sp.2 breu pitomba AProtium sp.3 breu sem cheiro AProtium sp.4 breu branco AProtium sp.5 breu de cheiro AProtium sp.6 ─ AProtium sp.7 breu dos outros AProtium sp.8 breu A
CAESALPINACEAE Bauhinia sp.1 ─ LBauhinia sp.2 ─ LCopaifera multijuga Hayne copaíba ADialium guianense (Aubl.) Sandwith juntai pororoca ACaesalpinaceae sp.1 ─ ACaesalpinaceae sp.2 ─ AMacrolobium suaveolens Spruce ex Benth. ─ ASwartzia sp.1 ─ ASwartzia sp.2 ─ ATachigali paniculata Aubl. taxi preto ATachigali .sp taxi vermelho A
CECROPIACEAE Cecropia sciadophylla Mart. imbaúba ACHRYSOBALANACEAE Couepia sp. Castanha de urubu A
Hirtella bicornis Mart. & Zucc. ─ AHirtella rodriguensii Prance ─ AHirtella sp. ─ ALicania hirsuta Prance pajurazinho ALicania octandra Kuntze. caraipé ALicania sp.1 mucucu xiado ALicania sp.2 caraipé ALicania sp.3 mucucu sangue A
CLUSIACEAE Symphonia globulifera L.f. anani ADICHAPETALACEAE Dichapetalaceae sp. ─ AEUPHORBIACEAE Pogonophora sp. manatua A

Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
FABACEAE Bocoa viridiflora(Ducke) R.S.Cowan muirajibóia-preta AMachaerium sp. ─ A
HUMIRIACEAE Endopleura uchi (Huber) Cuatrec. uchi AVantanea sp. uchi preto A
HIPPOCRATEACEAE Salacea multiflora (Lam.) DC xixua LLAURACEAE Dicypellium manausense W.A.Rodrigues louro preto A
Licaria aritu Ducke louro aritu AOcotea sp.1 louro amarelo AOcotea sp.2 louro AOcotea sp.3 louro A
LECYTHIDACEAE Eschweilera amazonica R.Knuth matá-matá AEschweilera sp. matá-matá-amarelo A
MELIACEAE Guarea cinnamomea Harms. jito branco AGuarea kunthiana A. Juss. andirobarana vermelho ATrichilia micrantha Benth. ─ ATrichilia .sp ─ A
MENISPERMACEAE Abuta rufescens Aubl. abuta LMIMOSACEAE Enterolobium schomburgkii Benth. ─ A
Enterolobium sp.1 ─ AEnterolobium sp.2 farveira orelha de
macacoA
Inga longiflora Benth. ingá AParkia panurensis Spruce ex H.C. Hopkins ─ AZygia racemosa (Ducke) Barneby & J.W. Grimes angelim rajado A
MORACEAE Brosimum rubescens Taub. pau rainha ABrosimum parinarioides Ducke amapá ABrosimum sp. ─ AFicus sp. ─ AHelicostylis sp. inharé AMoraceae sp. ─ ANaucleopsis ulei (Warburg) Ducke muiritinga APseudolmedia laevis (Ruiz & Pav.) Macbr. pama A
MYRISTICACEAE Iryanthera paraensis Huber. punaIryanthera elliptica Ducke ucuuba pena AVirola elongata Benth. ucuuba vermelha AVirola michelii Heckel virola A
MYRTACEAE Eugenia sp. goiabinha AMyrcia sp. ─ A
OLACACEAE Heisteria sp. ─ AIndeterminada ─ A
QUIINACEAE Lacunaria grandiflora Ducke lacunária ALacunaria sp. ─ AQuiina negrensis A.C.Sm. moela de mutum A
RUBIACEAE Duroia sp. ─ A
FAMÍLIA ESPÉCIE NOME POPULAR HÁBITO
SAPINDACEAE Talisia sp. pitomba do mato Acontinua >
> continuação
159
160
(49), Sapotaceae (42), Mimosaceae (34), Chrysobalanaceae (28), Lecythidaceae (24) e Myristicaceae (23), representando 69,6% do total amostrado (Tab. 1). Asdemais apresentaram menos que 14 indivíduos cada(Tab. 2). Oliveira (1997), em uma revisão sobre estudosquantitativos em mata de terra firme na Amazônia,detectou que, com raras exceções, entre as famílias commaior densidade e diversidade no dossel das matasprimárias estão Leguminosae ( ), Lecythidaceae, Sapotaceae, Burseraceae, Chrysobalanaceae,Moraceae e Lauraceae. Muitos são os estudos onde estasfamílias estão entre as 10 mais importantes docomponente arbóreo de terra firme. No levantamento da
--
sensu lato -
floresta de terra firme da RDS Tupé,das famílias citadas anteriormente,apenas Lauraceae não apareceu entreas famílias mais importantes, noentanto, a espécie com maior númerode indivíduos e valor de importânciafoi Swartz.
A riqueza de espécies é considerada alta em relação aos demaistrabalhos realizados na Amazônia ondeas plantas possuíam o PAP 30cm.Bastos (1948), ao publicar o seuprimeiro estudo quantitativo de uma
Protium apiculatum-
>
Diniz Scudeller&
SAPOTACEAE Micropholis mensalis (Baehni) Aubrév. abiurana goiabinha AMicropholis casiquiarensis Aubrév. ─ AMicropholis cylindrocarpa (Poepp.) Pierre ─ AMicropholis sp.1 ─ AMicropholis sp.2 ─ AMicropholis sp.3 ─ AMicropholis sp.4 ─ AMicropholis sp.5 ─ AMicropholis sp.6 ─ AMicropholis sp.7 ─ AMicropholis sp.8 ─ AMicropholis sp.9 ─ APouteria eugeniifolia (Pierre) Baehni batinga APouteria sp.1 ─ APouteria sp.2 ─ APouteria sp.3 ─ APouteria sp.4 ─ APouteria sp.5 abiurana vermelha APouteria sp.6 abiurana APouteria sp.7 abiurana APouteria sp.8 ─ APouteria sp.9 ─ APouteria sp.10 ─ APouteria sp.11 ─ A
SIMAROUBACEAE Simaba polyphylla (Cavalcante) Thomas marubá roxo ASTERCULIACEAE Theobroma subincana Mart. capui A
Theobroma sylvestris Aubl. et Mart. cacaui ATILIACEAE Luhea sp. açoita cavalo AVOCHYSIACEAE Vochysia sp. ─ AINDETERMINADA 1 ─ ─ AINDETERMINADA 2 ─ ─ A
FAMÍLIA ESPÉCIE NOME POPULAR HÁBITO> continuação

floresta amazônica, utilizando umaamostragem de 1ha identificou 46morfoespécies entre 124 indivíduos.Silva . (1986) registraram 103espécies em um transecto de 1ha. Asmaiores riquezas de espéciesregistradas para a Amazônia ocidentalforam de aproximadamente 300espécies para DAP 10cm (Gentry,1988; Oliveira, 1997).
De acordo com os resultadosobtidos no levantamento de 0,5ha defloresta de terra firme estima-se que adensidade total de espécies arbóreas elianescentes na RDS Tupé é de 556ind/ha e a área basal por hectare de25,18m . A média total da altura é de17,27m (variando de 6 a 40m), doperímetro foi de 65,14cm (30 a910cm).
Apenas 15 espécies representarammais de 50% dos indivíduos amostradose , 8 8 m o r f o e s p é c i e s e s t ã orepresentadas por apenas umindivíduo. Prance . (1976), Amaral(1996), Oliveira (1997) e OliveiraAmaral (2004) também encontraramde 40 a 60% das espécies consideradas“localmente raras”.
,Spruce ex H.C. Hopkins,
sp 1,R.Knuth,
Benth., (Baehni)Aubrév, (Ducke)Barneby J.W. Grimes, sp 1,
sp 1, Heckel,sp 5,
Ducke e sp 1 contribuíramcom 47,38% do total de indivíduos, eapresentaram mais de 52% do total dadensidade relativa (Tab. 2). A espécie
possui 9,40% dos
et al
et al
Protium apiculatum ParkiapanurensisEschweilera . Eschweileraamazonica Virola elongata
Micropholis mensalisZygia racemosa
Licania .Couepia . Virola michelliPouteria . Iryanthera elliptica
Protium .
Protium apiculatum
>
2
Abundância das espécies
&
&
indivíduos amostrados, seguida por ,com 7,66%. As referidas espécies também apresentaramas maiores dominâncias relativas 9,41 e 7,67,respectivamente (Tab. 2). sp 4 apresentoumenos de 1/3 da densidade relativa da
, mas a dominância relativa foi a maior(33,75%) devido ao perímetro de seus indivíduos ser alto,tendo assim a maior área basal encontrada (26,69%, Tab.2).
Aproximadamente 30% dos indivíduos amostrados seencontram distribuídos na primeira classe de perímetro(30-39,9cm) (Fig. 1).Aproximadamente 50% são menoresque 50cm de perímetro. Cerca de 18,81% dos indivíduosamostrados apresentaram o perímetro maior que 90cm,dentre estes se destaca um indivíduo de sp 4que possui o maior perímetro observado (910cm).Oliveira Amaral (2004) propõem que o comportamentodecrescente da curva (“J” invertido) indica pouca ounenhuma pressão antrópica sobre o ambiente. Mais de50% dos indivíduos (66,55%) tiveram suas alturasestimadas entre 10 e 20m, apenas 3,48% tiveram suasalturas estimadas em mais de 30m (Fig. 2).
As cinco espécies com maior VC foramsp 4,
sp 1 e sp 1, representandoaproximadamente 40% do total amostrado (Tab. 2).
Analisando as espécies amostradas quanto à suaimportância ecológica, das seis figuradas com maior VI,
destacamsp 4, sp 1,
e , representando cerca de30% do total (Tab. 2).
Analisando a figura 3, pode-se inferir que acomunidade vegetal é bastante heterogênea quanto à
Parkia panurensis
Micropholis .Protium
apiculatum
Micropholis .
Micropholis. Parkia panurensis, Protium apiculatum,
Eschweilera . Couepia .
se Protium apiculatum, Parkia panurensis,Micropholis . Eschweilera . Eschweileraamazonica Virola elongata
Estrutura diamétrica
Valor de Cobertura (VC)
Valor de Importância (VI)
Curva cumulativa das espécies
&
Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
161

162
Diniz Scudeller&
Espécies ni np DR (%) DA AB DoA DoR FA FR VC% VI%
Protium apiculatum 27 23 9,41 54 0,71 1,42 2,82 46 49,78 6,70 8,25Parkia panurensis 22 13 7,67 44 2,05 4,09 8,13 26 28,13 8,66 5,85Micropholis sp.4 2 2 0,70 4 6,72 13,44 26,70 4 4,33 15,02 4,22Eschweilera sp. 11 9 3,83 22 1,22 2,44 4,84 18 19,48 4,75 3,75Eschweilera amazonica 13 10 4,53 26 0,40 0,81 1,60 20 21,64 3,36 3,70Virola eleogata 11 9 3,83 22 0,25 0,50 0,99 18 19,48 2,64 3,23Micropholis mensalis 9 8 3,14 18 0,17 0,35 0,69 16 17,31 2,10 2,81Zigia racemosa 8 8 2,79 16 0,18 0,37 0,73 16 17,31 1,93 2,77Licania sp.1 8 7 2,79 16 0,16 0,33 0,65 14 15,15 1,89 2,47Couepia sp. 5 5 1,74 10 1,14 2,27 4,51 10 10,82 3,43 2,27Virola michelli 6 6 2,09 12 0,30 0,59 1,18 12 12,99 1,79 2,16Pouteria sp.5 6 6 2,09 12 0,27 0,55 1,09 12 12,99 1,74 2,15Iryanthera elliptica 5 5 1,74 10 0,32 0,65 1,28 10 10,82 1,66 1,84Protium sp.1 5 5 1,74 10 0,09 0,18 0,35 10 10,82 1,15 1,72Ocotea sp.1 4 4 1,39 8 0,16 0,32 0,64 8 8,66 1,11 1,42Licania hirsuta 4 4 1,39 8 0,14 0,28 0,56 8 8,66 1,07 1,41Pseudolmedia laevis 4 4 1,39 8 0,11 0,22 0,44 8 8,66 1,00 1,40Protium sp.8 4 4 1,39 8 0,10 0,21 0,41 8 8,66 0,99 1,39Protium sp.3 4 3 1,39 8 0,11 0,22 0,44 6 6,49 1,00 1,11Micropholis sp.2 3 3 1,05 6 0,18 0,35 0,70 6 6,49 0,96 1,10Scleronema micranthum 3 3 1,05 6 0,14 0,28 0,57 6 6,49 0,89 1,08Pouteria sp.4 3 3 1,05 6 0,12 0,18 0,47 6 6,49 0,83 1,07Eugenia sp 3 3 1,05 6 0,09 0,02 0,37 6 6,49 0,78 1,05Licania sp.3 3 3 1,05 6 0,07 0,14 0,28 6 6,49 0,73 1,04Protium sp.4 3 3 1,05 6 0,07 0,14 0,27 6 6,49 0,72 1,04Protium sp.2 3 3 1,05 6 0,04 0,09 0,17 6 6,49 0,67 1,03Aspidosperma sp. 2 2 0,70 4 0,38 0,77 1,52 4 4,33 1,22 0,87Licania sp.2 3 2 1,05 6 0,06 0,13 0,26 4 4,33 0,72 0,75Enterolobium sp.1 1 1 0,35 2 0,78 1,56 3,10 2 2,16 1,89 0,75Licaria aritu 2 2 0,70 4 0,10 0,19 0,38 4 4,33 0,59 0,72Guatteria sp.4 2 2 0,70 4 0,08 0,16 0,31 4 4,33 0,55 0,71Pogonophora sp. 2 2 0,70 4 0,05 0,10 0,20 4 4,33 0,49 0,70Guarea cinnamomea 2 2 0,70 4 0,03 0,07 0,13 4 4,33 0,46 0,69Abuta rufescens 2 2 0,70 4 0,03 0,07 0,13 4 4,33 0,46 0,69Simaba polyphylla 2 2 0,70 4 0,03 0,07 0,13 4 4,33 0,46 0,69 continua >
Tabela 2. Relação espécies arbóreas e lianescentes amostradas em 0,5ha de uma floresta de terra firme naTrilha da Cachoeira (RDS Tupé), com os respectivos parâmetros fitossociológicos. ni = número de indivíduosamostrados; np = número de parcelas que a espécie estava presente; DR = densidade relativa expressa emporcentagem; DA= densidade absoluta;AB = área basal; DoA= dominância absoluta; DoR = dominância relativa;FA= freqüência absoluta; FR = freqüência relativa; VC = valor de cobertura; VI = valor de importância.

Espécies ni np DR (%) DA AB DoA DoR FA FR VC% VI%
Hirtella rodriguesii 2 2 0,70 4 0,02 0,05 0,10 4 4,33 0,44 0,68Ficus sp. 2 2 0,70 4 0,02 0,05 0,10 4 4,33 0,44 0,68Salaceae multiflora 2 2 0,70 4 0,02 0,04 0,09 4 4,33 0,43 0,68Talisia sp. 2 2 0,70 4 0,02 0,04 0,09 4 4,33 0,43 0,68Symphonia globulifera 2 2 0,70 4 0,02 0,04 0,08 4 4,33 0,43 0,68Trichilia micrantha 2 2 0,70 4 0,02 0,04 0,08 4 4,33 0,43 0,68Naucleopsis ulei 2 2 0,70 4 0,01 0,03 0,06 4 4,33 0,42 0,68Enterolobium sp.2 1 1 0,35 2 0,51 1,01 2,01 2 2,16 1,29 0,60Pouteria sp.6 1 1 0,35 2 0,32 0,64 1,27 2 2,16 0,89 0,50Vochysia sp. 1 1 0,35 2 0,29 0,59 1,17 2 2,16 0,83 0,49Pouteria sp.9 1 1 0,35 2 0,28 0,57 1,13 2 2,16 0,81 0,48Anacardium giganteum 1 1 0,35 2 0,28 0,56 1,11 2 2,16 0,80 0,48Xylopia sp. 1 1 0,35 2 0,12 0,24 0,48 2 2,16 0,46 0,40Micropholis sp.1 1 1 0,35 2 0,11 0,22 0,44 2 2,16 0,43 0,39Tachigali sp. 1 1 0,35 2 0,10 0,21 0,41 2 2,16 0,42 0,39Olacaceae sp. 1 1 0,35 2 0,10 0,20 0,40 2 2,16 0,41 0,39Micropholis sp.7 1 1 0,35 2 0,08 0,17 0,33 2 2,16 0,37 0,38Hirtella sp. 1 1 0,35 2 0,08 0,15 0,30 2 2,16 0,36 0,37Guatteria sp.3 1 1 0,35 2 0,07 0,14 0,28 2 2,16 0,35 0,37Moraceae sp. 1 1 0,35 2 0,07 0,14 0,28 2 2,16 0,35 0,37Copaifera multijuga 1 1 0,35 2 0,07 0,14 0,27 2 2,16 0,34 0,37Protium sp.5 1 1 0,35 2 0,06 0,13 0,26 2 2,16 0,33 0,37Brosimum parinarioides 1 1 0,35 2 0,06 0,13 0,25 2 2,16 0,33 0,37Micropholis sp.8 1 1 0,35 2 0,06 0,12 0,23 2 2,16 0,32 0,36Heisteria sp. 1 1 0,35 2 0,04 0,09 0,18 2 2,16 0,29 0,36Ocotea sp.3 1 1 0,35 2 0,04 0,08 0,16 2 2,16 0,28 0,36Pouteria sp.8 1 1 0,35 2 0,04 0,08 0,16 2 2,16 0,28 0,36Guatteria sp.2 1 1 0,35 2 0,03 0,07 0,14 2 2,16 0,27 0,35Caesalpinaceae sp.2 1 1 0,35 2 0,03 0,07 0,14 2 2,16 0,27 0,35Micropholis casiquiarensis 1 1 0,35 2 0,03 0,07 0,14 2 2,16 0,27 0,35Ocotea sp.2 1 1 0,35 2 0,03 0,07 0,13 2 2,16 0,26 0,35Indeterminada 1 1 0,35 2 0,03 0,05 0,11 2 2,16 0,25 0,35Xylopia crinita 1 1 0,35 2 0,03 0,05 0,10 2 2,16 0,25 0,35Caesalpinaceae sp.1 1 1 0,35 2 0,02 0,05 0,10 2 2,16 0,25 0,35Guarea kunthiana 1 1 0,35 2 0,02 0,05 0,10 2 2,16 0,25 0,35Guatteria sp.1 1 1 0,35 2 0,02 0,04 0,09 2 2,16 0,24 0,35Macrolobium suaveolens 1 1 0,35 2 0,02 0,05 0,09 2 2,16 0,24 0,35Swartzia sp.2 1 1 0,35 2 0,02 0,05 0,09 2 2,16 0,24 0,35Brosimum rubescens 1 1 0,35 2 0,02 0,04 0,09 2 2,16 0,24 0,35Helicostylis sp. 1 1 0,35 2 0,02 0,04 0,09 2 2,16 0,24 0,35
> continuação
continua >
Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
163

Espécies ni np DR (%) DA AB DoA DoR FA FR VC% VI%
Myrcia sp. 1 1 0,35 2 0,02 0,04 0,09 2 2,16 0,24 0,35Swartzia sp.1 1 1 0,35 2 0,02 0,04 0,08 2 2,16 0,24 0,34Trichilia sp. 1 1 0,35 2 0,02 0,04 0,08 2 2,16 0,24 0,34Protium sp.7 1 1 0,35 2 0,02 0,04 0,07 2 2,16 0,23 0,34Vantanea sp. 1 1 0,35 2 0,02 0,03 0,07 2 2,16 0,23 0,34Pouteria sp.11 1 1 0,35 2 0,02 0,04 0,07 2 2,16 0,23 0,34Pouteria sp.3 1 1 0,35 2 0,02 0,04 0,07 2 2,16 0,23 0,34Licania octandra 1 1 0,35 2 0,02 0,03 0,06 2 2,16 0,22 0,34Machaerium sp. 1 1 0,35 2 0,01 0,03 0,06 2 2,16 0,22 0,34Micropholis sp.3 1 1 0,35 2 0,01 0,03 0,06 2 2,16 0,22 0,34Micropholis sp.9 1 1 0,35 2 0,02 0,03 0,06 2 2,16 0,22 0,34Pouteria sp.10 1 1 0,35 2 0,01 0,03 0,06 2 2,16 0,22 0,34Luhea sp. 1 1 0,35 2 0,02 0,03 0,06 2 2,16 0,22 0,34Protium sp.6 1 1 0,35 2 0,01 0,02 0,05 2 2,16 0,22 0,34Dialium guianense 1 1 0,35 2 0,01 0,03 0,05 2 2,16 0,22 0,34Enterolobium schomburgkii 1 1 0,35 2 0,01 0,03 0,05 2 2,16 0,22 0,34Brosimum sp. 1 1 0,35 2 0,01 0,02 0,05 2 2,16 0,22 0,34Bocoa viridiflora 1 1 0,35 2 0,01 0,02 0,05 2 2,16 0,22 0,34Quiina negrensis 1 1 0,35 2 0,01 0,02 0,05 2 2,16 0,22 0,34Micropholis sp.5 1 1 0,35 2 0,01 0,03 0,05 2 2,16 0,22 0,34Pouteria sp.2 1 1 0,35 2 0,01 0,03 0,05 2 2,16 0,22 0,34Tachigali paniculata 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Hirtella bicornis 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Dichapetalaceae sp. 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Endopleura uchi 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Indeterminada 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Dicypellium manausense 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Inga longiflora 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Duroia sp. 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Micropholis cylindrocarpa 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Pouteria sp.7 1 1 0,35 2 0,01 0,02 0,04 2 2,16 0,21 0,34Guatteria foliosa 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Bauhinia sp.1 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Bauhinia sp.2 1 1 0,35 2 0,01 0,01 0,03 2 2,16 0,21 0,34Cecropia sciadophylla 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Iryanthera paraensis 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Lacunaria sp. 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Lacunaria grandiflora 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Micropholis sp.6 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Pouteria eugeniifolia 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34
> continuação
continua >
164
Diniz Scudeller&

7
84
107
40
1910
0
20
40
60
80
100
120
5-9.9 10-14.9 15-19.9 20-24.9 25-29.9 > 30
Classes de altura (m)
Nú
me
rod
ein
div
ídu
os
90
4839
22 1915
54
0102030405060708090
100
30-39.9 40-49.9 50-59.9 60-69.9 70-79.9 80-89.9 > 90
Classes de perímetro (cm)
nú
mero
de
ind
ivíd
uo
s
Espécies ni np DR (%) DA AB DoA DoR FA FR VC% VI%
Pouteria sp.1 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Theobroma subincana 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Theobroma sylvestris 1 1 0,35 2 0,01 0,02 0,03 2 2,16 0,21 0,34Total = 118 287 50 100 574 20,61 41,24 82,22 526 568,88 100 100
> continuação
composição florística, evidenciandoque apenas 0,5ha não é suficiente paracaracterizar tal formação vegetal. Noentanto, segundo Gray . (2004),alguns autores confundem o conceitode diversidade beta, que significa adiferença na composição das espéciese não no acréscimo do número, quandocompararam doi s ambientes .Acrescido a isso, o índice de Shannonencontrado no presente estudo foi de4,214 nats.indivíduo , consideradorelativamente alto para florestastropicais (Knight, 1975). Portanto, osresultados apresentados no presenteestudo são significativos para inferirsobre a estrutura de uma floresta deterra firme naAmazônia Central.
A floresta de terra firme na RDSTupé apresenta uma alta diversidadede espécies. Em apenas 0,5ha foramamostradas 118 espécies com PAP30cm. As famílias mais importantes emdiversidade, densidade e biomassa nãovariam consideravelmente entre oslevantamentos analisados. Sapota-ceae, Caesalpinaceae, Burseraceae,Chrysobalanaceae e Moraceae são ascinco famílias com maior riqueza deespécies e número de indivíduos porhectare.
As espécies ,sp.1 e
sp.4 são as mais importantes na referida comunidade.
et al
Protium apiculatumParkia panurensis, EschweileraMicropholis -
-1
Conclusões
Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
Figura 1. Distribuição relativa, por intervalo de perímetro, dasárvores e lianas amostradas na floresta de terra firme da Trilhada Cachoeira (RDS Tupé). Os números sobre as barras referem-se à quantidade dos indivíduos amostrados em cada intervalo.
Figura 2. Distribuição relativa, por intervalo de altura, dasárvores e lianas amostradas na floresta de terra firme da Trilhada Cachoeira (RDS Tupé). Os números sobre as barras referem-se à quantidade dos indivíduos amostrados em cada intervalo.
O índice de diversidade de Shannon de 4,214nats.indivíduo , considerado alto no contexto de estudossemelhantes.
-1
165

166
Diniz Scudeller&
020406080
100120140
1 4 7 10 13 16 19 22 25 28 31 34 37 40 43
número de parcelas
nú
mero
de
esp
écie
s
acu
mu
lad
as
Figura 3. Curva cumulativa das espécies com PAP 30cmamostradas na floresta de terra firme da Trilha da Cachoeira(RDS Tupé).
>
Agradecimentos
Bibliografia Citada
Agradecemos a toda equipe do projeto BiotupéVegetação que de uma forma ou de outra contribuiu paraa realização deste trabalho, principalmente na fase delevantamento do campo e ao seu “Zé Lima”parabotânico do INPA que auxiliou na determinaçãotaxonômica dos indivíduos amostrados.
Amaral, I.L. 1996.. Dissertação
de Mestrado, INPA/FUA, Manaus,AM. 121pp.Bastos, A.M. 1948. As matas de Santa Maria de Vila Nova.
, MinistériodaAgricultura, Serviço Florestal, Setor de InventáriosFlorestais, 1: 281-288.
Causton, D. R. 1988..
Unwin Hyman, London. 342pp.Cronquist, A. 1981.
. ColumbiaUniversity 1262pp.
Damasceno, G.A. 1997. Estudo florístico efitossociológico de um trecho de mata ciliar do RioParaguai, Pantanal-MS, e suas relações com regime deinundação. Dissertação de Mestrado, UNICAMP,Campinas, SP. 115pp.
Diversidade florística em floresta deterra firme, na região do rio Urucu AM
Anuário Brasileiro de Economia Florestal
An introduction to vegetationanalysis, principles, practice and interpretation
An integrated system ofclassification of flowering plants
, New York:
Gentry, A.H. 1988. Tree speciesrichness of upperAmazonian forest.
,85: 156-227.
Gentry, A.H. 1996. A field guide to thefamilies and genera of woody plantsof North-West South America(Colombia, Equador, Peru) withspplementary notes on herbaceoustaxa. Conservation Internation:University of Chicago Press,Chicago. 895pp.
Gray, J.S.; Ugland, K.L.; Lambshead,J. 2004. On species accumulationand species-area curves.
., 13: 567-568.Higuchi, N.; Santos, J.; Jardim, F.C.S.
1982. Tamanho de parcelasamostra is para inventár iosflorestais. , 12(1):91-103.
Knight, D.H. 1975. A phytosociologicalanalysis of species-rich tropicalforest on Barro Colorado Island,Panama. ,45: 259-280.
Lima, H.C. de Guedes-Bruni, R. R.1997.
.Jardim botânico, Rio de Janeiro.346pp.
Magurran, A.E. 1988..
Princeton University Press,Princeton, New Jersey. 179pp.
Martins, F.R. 1989. Fitossociologia deflorestas do Brasil: um históricobibliográfico.
, 40: 103-164.Martins, F.R. 1991.
. UNICAMP,Campinas-SP. 146pp.
Meira Neto, A.A. 1991.
Proceedings of the NationalAcademy of Sciences of the USA
GlobalEcol. Biogeogr
Acta Amazonica
Ecological Monographs
;Serra de Macaé de Cima:
D i v e r s i d a d e F l o r í s t i c a eConservação em Mata Atlântica
Ecologicaldiversity and its measurement
Pesquisa, sérieBotânica
Estrutura de umafloresta mesófila
Composiçãoflorística e fitossociológica de
&

Estrutura fitossociológica de uma floresta de terra firme na Amazônia Central
167
Central, Amazonas, Brasil. , 34(1):21-34.
Polhill, R.M.; Raven, P.H.; Stirton, C.H. 1981. Evolutionand systematics of the Leguminosae. In: Polhill, R.M.
Raven, P.H. (eds.) .Part 1. Royal Botanic Garden, Kew. p 1-26.
Prance, G.T.; Rodrigues, W.A.; Silva, M.F. 1976.Inventário florestal de um hectare de mata de terrafirme, km 30 da estrada Manaus-Itacoatiara.
, 6:9-35.Ribeiro, J.E.L. da S.; Hopkins, M. J. G.; Vincentini, A.;
Sothers, C. A.; Costa, M. A. da; Brito, J. M. de; Souza,M. A. D. de; Martins, L. H. P.; Lohmann, L. G.;Assunção, P. A. C. L.; Pereira, E. da C.; Silva, C.F. da;Mesquita, M. R., Procópio, L. C. 1999.
. INPA, Manaus-AM. 798pp.
Silva, M. F.; Rosa, N.A.; Oliveira, J. 1986. EstudoBotânico na área do projeto ferro Carajás. 3.Aspectos florístico da mata do aeroporto da SerraNorte-PA. , 3(1): 1-20.
Van den Berg, E. 1995.
. Dissertação de Mestrado UFLA,Lavras-MG. 73pp.
Acta Amazônica
Advances in legume systematics.
ActaAmazonica
Flora daReserva Ducke: guia de identificação das plantasvasculares de uma floresta de terra firme naAmazônia Central
Bol. Mus. Paraense Emílio Goeldi
Estudo florístico efitossociológico de uma floresta ripária em Itutinga,MG, e análise das correlações entre variáveisambientais e a distribuição das espécies de portearbóreo-arbustivo ,
&
Scudeller, V. V.; Aprile, F. M.; Melo. S.; Santos-Silva, E. N.2005. Reserva de Desenvolvimento Sustentável doTupé: características gerais. : Santos-Silva, E. N.;Aprile, F. M.; Scudeller, V. V.; Melo, S. (Orgs.).Biotupé: meio físico, diversidade biológica esociocultural do baixo Rio Negro, Amazônia Central.Ed. INPA,AM.
In
fisionomias de vegetação deCerrado sensu lato da EstaçãoEcológica de Santa Bárbara(E.E.S.B.) Município de Águas deSanta Bárbara, Estado de SãoPaulo ,
Estudoflorístico, estruturais e ambientaisnos estratos arbóreos e herbáceos-arbustivo de uma florestaestacional semidecidual em Viçosa,MG ,
Manual demanejo do herbário fanerogâmico
Aims and methods ofvegetation ecology. Willey andSons,
Diversidade,e s t r u t u r a e d i n â m i c a d ocomponente arbóreo de umafloresta de terra firme de ManausAmazonas ,
. Dissertação de MestradoUNICAMP, Campinas, SP. 98pp.
Meira Neto, A.A. 1997.
. Tese de Doutorado UNICAMP,Campinas, SP. 154pp.
Mori, S.A.; Silva, L.A.M.; Lisboa, G.;Coradin, L. 1989. 2. ed.
.Centro de Pesquisa do Cacau.
104pp.Mueller-Dombois; D. Elemberg, H.
1974.
New York. 547pp.Nee. M. 1995. Flora preliminar do
Projeto Dinâmica Biológica deFragmentos Florestais. Manaus,AM. 264pp.
Oliveira, A. A. 1997.
. Tese de doutorado. USPSão Paulo. 187pp.
Oliveira, A. N.; Amaral, I.L. 2004.Florística e fitossociologia de umafloresta de vertente na Amazônia
Ilhéus:


Diversidade BiológicaCapítulo 12
Composição da entomofauna aquáticaem igarapés da Reserva deDesenvolvimento Sustentável Tupé,Manaus, AM
RESUMO - AAmazônia é conhecida como o principal centro de biodiversidade do mundo, eos insetos representam o grupo de seres vivos com o maior número de espécies. Nopresente estudo se busca conhecer a composição e padrão de distribuição da entomofaunade dois igarapés de água preta, da Reserva de Desenvolvimento Sustentável Tupé,localizada à aproximadamente 28 km da cidade de Manaus, AM. As amostras foramrealizadas com o auxílio de um quadrado de 25cm nos períodos de enchente, cheia evazante. Em cada período foram tomadas 5 amostras de substrato folhiço em ambientes decabeceira, queda d'água e desembocadura dos dois igarapés. O material coletado foilavado e triado com auxílio de peneiras de 125
és) x 5 (meses de coleta) x 3(pontos de coleta), com cinco repetições. As médias foram comparadas pelo Teste deTukey, ao nível de 5% de probabilidade. Foram obtidos 4534 indivíduos, com ocorrência de20 ordens e/ou famílias no Igarapé das Pedras e 25 ordens e/ou famílias no Igarapé daCachoeira. A maior abundância relativa de insetos ocorreu no Igarapé da Cachoeira, com64%, e no Igarapé das Pedras a abundância foi de 36%, representadas principalmente porDiptera, das famílias Chironomidae e Ceratopogonidae; Trichoptera, pela família
μm. O delineamento experimental adotadofoi o inteiramente ao acaso, em esquema fatorial 2 (igarap
Maria José do Nascimento LOPESe-mail: [email protected]
Instituto Nacional de Pesquisas da Amazônia (INPA),Coordenação de Pesquisas em Entomologia (CPEN).
Avenida André Araújo, 2936, Petrópolis, CP 478, 690011-970, Manaus, AM.
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

Helicopsychidae e, Coleoptera. A maior diversidade de famílias se deu nos ambientes decabeceira e nas queda d'água. A menor diversidade foi registrada na desembocadura. Osvalores de pH, condutividade elétrica e temperatura da água registrada se encontramdentro do padrão representativo para aAmazônia Central.Atualmente, tanto o Igarapé dasPedras quanto o Igarapé da Cachoeira se encontram em bom estado de preservação, sendo,no entanto necessário a manutenção da floresta circundante a fim de preservar a fauna deinvertebrados dos igarapés da área da Reserva e esta importante fonte de água potável.
Insetos aquáticos; ambiente lótico, água preta,Amazônia, Brasil.PALAVRAS-CHAVE:
sendo utilizados como indicadorespara determinar a qualidade doscorpos de água em diferentes regiõesdo mundo (Rosenberg Resh, 1993;Moulton, 1998; Callisto Esteves,1998; Da-Silva, 1998, entre outros). Aeutrofização que ocorre nos corposd'água devido a interferência humanaleva inicialmente a um aumento nadensidade e na produção, acarretandoposteriormente o desaparecimento eextinção das espécies ou depopulações. A crescente destruição dehabitats que vêm ocorrendo naAmazônia tem tornado o trabalho deinventariar e manter materialtes temunho em co leções defundamental importância. No presenteestudo se busca conhecer e inventariara diversidade biológica dos insetosaquáticos que vivem nos pequenosigarapés naturais da Reserva deDesenvolvimento Sustentável do Tupésujeitos à inundação periódica,buscando manter material testemunhoda entomofauna aquática e fornecersubsídios a futuros programas deavaliação do grau de impactoambiental na região.
&&
Introdução
A floresta amazônica, embora seja poucodiferenciada do ponto de vista topográfico e climático,não é homogênea ecologicamente (Fittkau ., 1975).A flutuação anual do nível da água dos rios provoca ainundação das terras mais baixas, em até cerca de 10m,determinando a estrutura de todos os biótopos aquáticose semiaquáticos dos principais rios (Fittkau 1975).
A Amazônia é conhecida como o principal centro debiodiversidade do mundo e os insetos, que sãoconsiderados essenciais para o funcionamento dosecossistemas terrestre e aquático, representa o grupo deseres vivos com o maior número de espécies (Erwin,1983). Estes desempenham importante função no fluxode energia nos ecossistemas, participando ora comoconsumidores ora como decompositores, sendo esteúltimo aspecto, responsável, em parte, por acelerar ociclo de nutrientes (Fonseca Rafael, 1991).Informações sobre a diversidade da entomofaunaaquática na região Amazônica foram levantadas pordiversos autores, entre os quais Patrick . (1966);Fittkau (1967); Fittkau . (1975); Irmler Junk(1982); Nessimian (1985); Walker (1985); Freitas(1994);Walker (1994); Ferreira (1996); Cargnin-Ferreira(1998); Cleto-Filho Walker (2001); Pes, 2001; Couceiro
. (2001); Walker (2003). Ênfases para a entomofaunaaquática da área do lago (ria lake) do Tupé são dadas porFittkau . (1975) e Reiss (1977).
Dentre os insetos aquáticos, objeto deste estudo, osEphemeroptera, Plecoptera, Trichoptera e Diptera,apresentam espécies sensíveis a alterações ambientais,
et al
et al.,
et alet al
et al
et al
&
&
&
170
Lopes

Materiais e Métodos
O estudo foi desenvolvido em doisigarapés de água preta localizados naRese rva de Desenvo l v imentoSustentável do Tupé (RDS Tupé), quesofrem inundação periódica porocasião da enchente dos rios. A RDSTupé está situada a 28 km da cidade deManaus, AM, Brasil. O igarapé daCachoeira, localizado nas coordenadas03° 02' 03.2'' S e 60° 16' 34.6'' W,próximo à Comunidade São João doTupé (CSJT), recebendo visitas debanhistas, principalmente no trechoda queda d'água (cachoeira); e Igarapédas Pedras, que é um igarapé de menorporte, localizado nas coordenadas 03°01' 41.3'' S e 60° 16' 36.5'' W, maisdistante da referida comunidade,recebendo pouca visitação em virtudedo difícil acesso.
As coletas foram efetuadas aolongo de 5 meses abrangendo osperíodos de enchente (abril e maio de2002), cheia (junho 2002) e vazante(julho e agosto de 2002). Em cadaigarapé foram selecionados 3 pontos.O primeiro ponto corresponde a umtrecho de cabeceira em área defloresta de terra firme, de dosselfechado, cujo leito é composto porareia, raízes e folhiço (folhas mortas);o segundo ponto está localizado acerca de 200m à jusante do primeiro,correspondendo a uma área de quedad'água e dossel parcialmente fechado,onde nos períodos de cheia as águasinvadem o canal principal do igarapé echegam até a base da queda d'água; oterceiro ponto está situado noencontro de águas dos igarapés com olago (desembocadura, corresponden-do à zona de igapó). A distância doterceiro ponto em relação ao segundo
variou cerca de 300m. De acordo com Junk Wantzen(2004) o primeiro ponto corresponde a uma área dedrenagem, o segundo ponto representado pelo canal doigarapé e, o terceiro ponto corresponde à zona que sofreinundação periódica.
Considerando que a seleção dos pontos foi feita apartir da queda d'água e o Igarapé da Cachoeira é maisextenso que o Igarapé das Pedras, o ponto cabeceiraescolhido para o Igarapé da Cachoeira corresponde a umtrecho meandrítico, sem muita alteração antrópica, enão a uma nascente como no Igarapé das Pedras.
Mensalmente foram tomadas cinco amostras desubstrato folhiço em cada ponto utilizando um quadradode 25cm. Nos períodos ou pontos em que este substratonão estava disponível em função do aumento do nível daságuas que, por ocasião da cheia cobre o folhiço do leito,foi coletado o substrato que estivesse disponível (como,folhiço flutuante ou o folhiço encontrado aderido àsmargens). O material coletado foi transportado para oLaboratório de Insetos Aquáticos do INPA, lavado empeneiras de 125 foi íada com auxílio deestereomicroscópio e identificada tomando por base ostrabalhos de Merritt Cummins (1979); McCaffertyProvonsha (1983); Fernández Domínguez (2001).
O delineamento experimental adotado foi ointeiramente ao acaso, em esquema fatorial 2 (igarapés)X 5 (meses de coleta) X 3 (pontos de coleta), com cincorepetições. As médias foram comparadas pelo Teste deTukey ao nível de 5% de probabilidade cujos resultadosestão representados nas figuras 1 e 2.
Foram tomados os valores de pH, condutividadeelétrica e temperatura da água no momento das coletasutilizando pHmetro, condutivímetro e termômetroportáteis, respectivamente, a fim de caracterizar osambientes. Nos meses de maio e junho os valores nãoforam registrados.
No Igarapé da Cachoeira os valores de pH variaramentre 3.7 a 6.4; a condutividade elétrica variou entre 6.7e 9.8 água variou entre 25 a
&
& &&
μm, a fauna tr
μ S cm
Resultados
Valores de pH, condutividade elétrica etemperatura da água no momento das coletas
¹ e, a temperatura da
Composição da Entomofauna Aquática em Igarapés da Reserva de Desenvolvimento Sustentável Tupé, Manaus, AM
171

32°C. No Igarapé das Pedras os valores de pH variaramentre 3.2 a 5.4; a condutividade elétrica variou entre 7.5e 11 água variou entre 24 a30°C (Tab. 3).
De todos os ambientes, na desembocadura (igapó)foram registrados os maiores valores de condutividade.
Quanto aos períodos, no mês de abril, em todos ospontos do Igarapé das Pedras, foram registrados osmaiores valores de condutividade elétrica.
Foram coletados 4.534 indivíduos, distribuídos em 8ordens e reconhecidas 20 famílias de insetos (Tabelas 1 e2).
As ordens de insetos identificadas foram:Ephemeroptera, conhecidas vulgarmente comoborboletas de piracema, ou siriruia; Odonata,conhecidas como libélulas ou lavadeiras; Plecoptera,que por serem pouco conhecidos não têm nome vulgar;Hemiptera, que são conhecidos como baratas-d'água,percevejos ou barbeiros; Megaloptera, são insetosgrandes, mas pouco conhecidos; Coleoptera, são osbesouros; Trichoptera, são minúsculas mariposas comasas densamente recobertas por pêlos; Diptera, que sãoos mosquitos e as moscas; Hymenoptera, que são asformigas e Isoptera, cupins caíram acidentalmente nasamostras.
As ordens Coleoptera e Diptera, esta últimarepresentada pelas famílias Chironomidae eCeratopogonidae, foram os grupos que apresentarammaior freqüência (100% e 90%, respectivamente)estando presentes em todos os ambientes (cabeceira,queda d'água e desembocadura) nos dois igarapés. OsDiptera pertencentes às famílias Simuliidae e Tipulidaesó ocorreram no mês de maio (enchente), na cabeceirado Igarapé da Cachoeira, o que corresponde à cerca de3% das amostras.
A ordem Trichoptera, representada pelas famíliasCalamoceratidae, Sericostomatidae, Glossossomatidae,Helicopsychidae, Hydropsychidae, Hydroptilidae,Lepidostomatidae, Leptoceridae, Odontoceridae,Philopotamidae, e Polycentropodidae, foi a ordem queapresentou maior riqueza em número de famílias. Estegrupo ocorreu principalmente nos trechos de cabeceira e
μ S cm¹ e, a temperatura da
Riqueza da Entomofauna do Igarapé da Cachoeira eIgarapé das Pedras
no trecho de queda d'água, estandop r a t i c a m e n t e a u s e n t e n adesembocadura do igarapé.
Aordem Ephemeroptera, represen-tada pelas famílias Caenidae,Leptophlebiidae, Leptohyphidae ePolymitarcyidae, esteve presente emtodos os locais amostrados, com baixafreqüência.
Para o Igarapé das Pedras foramreconhecidas 20 ordens e/ou famíliasde insetos e 23 para o Igarapé daCachoeira.
A maior abundância relativa deinsetos aquáticos ocorreu no Igarapéda Cachoeira, com 64%, e no Igarapédas Pedras a abundância foi de 36%(Fig. 1), representados principalmentepor Diptera, nas famílias Chironomidaee Ceratopogonidae; Trichoptera, pelafamília Helicopsychidae e, Coleopte-ra. A análise de variância mostrouefeito de interação, ou interdepen-dência, entre os fatores estudados:igarapés estudados em relação aosmeses de observação; igarapésestudados em relação aos pontos decoleta; meses de observação emrelação aos pontos de coleta; e,igarapés pelos meses de observação eos pontos de coleta (Tab. 4).
Na interação entre o fator igarapé eos meses de observação se obteveresultado significativo (p<0,01) (Tab.4). No Igarapé da Cachoeira (Fig. 1), severificou a menor ocorrência deinsetos no período de enchente (abril emaio), corresponde a um períodomuito chuvoso com correntezavariável entre os pontos de coleta e apresença de pequena quantidade de
Distribuição de insetos aquáticosnos Igarapés da Cachoeira e dasPedras
172
Lopes

Ordem Abr 2002 Mai 2002 Jun 2002 Jul 2002 Ago 2002
Família Pto 1 Pto 2 Pto 3 Pto 1 Pto 2 Pto 3 Pto 1 Pto 2 Pto 3 Pto 1 Pto 2 Pto 3 Pto 1 Pto 2 Pto 3Ephemeroptera
Caenidae - - - - - - - - + - - - - - -Leptophlebiidae - - - + + - - - - - - - - + -Leptohyphidae - - - - + - - - - - - - - - -
Polymitarcyidae - - - - - - - - + - - - + - -Odonata - - - - - - + + + + + + + - -Plecoptera
Perlidae - + - + - - + + - - - - + - -Hemiptera - - + - - - + + + + - + - - +Megaloptera
Corydalidae - - - - - - + - - - - - - - -Coleoptera + + + + + + + + + + + + + - +Trichoptera
Calamoceratidae - - - + + - + - - + + - + + -Sericostomatidae - - - - - - - - - - - - - - -Glossosomatidae - - - - - - - - - - - - - - -Helicopsychidae + - - + + - + - - + + - - - -
Hydropsychidae - - - + - - - - - - - - - - -Hydroptilidae - + - - - - + + - - - - + - -
Lepidostomatidae - - - - - - - + - - - - - + -Leptoceridae - - - - - - - - - - - - - - -
Odontoceridae + - - + - - + + - - - - - + -Philopotamidae - - - + - - - - - - - - - + -
Polycentropodidae - - - - + - - + + + - - - - -Diptera
Chironomidae + + + + + + + + + + + + + + +Ceratopogonidae + + + + + - + + + + + + + + +
Simuliidae - - - - - - - - - - - - - - -Tipulidae - - - - - - - - - - - - - - -
Diptera (n ident) + + - + + - + - + + + + - + +
Total de famílias 6 6 4 11 9 2 12 10 9 9 7 6 8 8 5
Tabela 1.Dados de Presença (+) e Ausência (-) dos grupos taxonômicos de insetos aquáticos coletados emambiente de cabeceira, em queda d'água e da desembocadura do Igarapé das Pedras, na RDS do TUPÉ,Manaus,Am.
substrato folhiço; e os maiores valoresforam registrados no período de cheia(junho), que corresponde a um períodochuvoso com forte correnteza emtodos os pontos de coleta e a presençade grande quantidade de substratofolhiço; seguido pelo período devazante, que corresponde a umperíodo sem chuvas e com correntezamoderada e a água restrita ao canal doigarapé. No Igarapé das Pedras (Fig. 1)foi verificado a maior ocorrência de
insetos no período da vazante (julho), correspondendo aum período sem chuvas, com água restrita ao canal doigarapé, com folhiço principalmente na área de quedad'água, correnteza variável; e nos períodos de enchentee cheia os valores registrados foram semelhantes entresi. No período da enchente foi registrado correntezaforte e pouco substrato folhiço; no período de cheia foiregistrado correnteza forte e a presença de grandequantidade de areia misturada ao substrato folhiço, oque provavel-mente, tornou o substrato pouco favorávelpara a manutenção dos organismos.
Na interação entre o fator igarapé e os pontos de
Composição da Entomofauna Aquática em Igarapés da Reserva de Desenvolvimento Sustentável Tupé, Manaus, AM
173

174
Lopes
Ord
emA
br
20
02
Ma
i2
00
2J
un
20
02
Ju
l2
00
2A
go
20
02
Fam
ílias
Pto
1P
to2
Pto
3P
to1
Pto
2P
to3
Pto
1P
to2
Pto
3P
to1
Pto
2P
to3
Pto
1P
to2
Pto
3
Ep
he
me
rop
tera
Cae
nida
e-
--
--
+-
--
--
--
--
Lept
ophl
ebiid
ae+
++
++
--
--
--
--
--
Pol
ymita
rcyi
dae
-+
+-
--
--
--
--
+-
-
Od
on
ata
-+
-+
+-
--
-+
+-
++
+P
lec
op
tera
Per
lidae
++
-+
--
--
--
+-
-+
-
He
mip
tera
--
+-
+-
--
--
--
-+
+
Me
ga
lop
tera
Cor
ydal
idae
+-
--
--
--
--
--
--
-
Co
leo
pte
ra+
++
++
-+
++
++
-+
++
Tri
ch
op
tera
Cal
amoc
erat
idae
-+
--
--
-+
-+
--
++
-
Ser
icos
tom
atid
ae-
--
--
+-
--
--
--
--
Glo
ssos
omat
idae
--
--
--
+-
--
--
--
-
Hel
icop
sych
idae
+-
-+
+-
++
-+
+-
++
-
Hyd
rops
ychi
dae
--
--
--
--
--
--
--
Hyd
ropt
ilida
e-
+-
--
--
+-
--
--
+-
Lepi
dost
omat
idae
--
--
--
-+
--
--
-+
-
Lept
ocer
idae
++
-+
+-
--
--
--
++
+
Odo
ntoc
erid
ae-
--
-+
--
+-
--
--
+-
Phi
lopo
tam
idae
--
--
--
--
--
--
+-
-
Pol
ycen
trop
odid
ae-
--
--
--
+-
--
-+
--
Dip
tera C
hiro
nom
idae
++
++
++
++
++
++
++
+
Cer
atop
ogon
idae
++
++
+-
++
+-
++
++
+
Sim
uliid
ae-
--
+-
--
--
--
--
--
Tip
ulid
ae-
--
+-
--
--
--
--
--
Dip
tera
(nid
ent.)
++
++
++
++
++
--
++
+
Tota
lde
fam
ílias
911
811
104
610
46
72
1113
7
Tabe
la2.
Dad
osde
Pres
ença
(+)
eAus
ênci
a(-
)do
sgr
upos
taxo
nôm
icos
dein
seto
saq
uáti
cos
cole
tado
sem
ambi
ente
deca
bece
ira,
emqu
eda
d'ág
uae
dade
sem
boca
dura
doIg
arap
éda
Cac
hoei
ra, n
aRD
Sdo
TU
PÉ, M
anau
s,Am
.

Tabela 3. Valores de pH, condutividade elétrica e temperatura da água registrados nos ambientes de cabeceira(Pto. 1), de queda d'água (Pto. 2) e da desembocadura (Pto. 3) dos Igarapés da Cachoeira e das Pedras, na RDSdo TUPÉ (Manaus,Am) nos meses deAbril, Julho eAgosto de 2002.
Composição da Entomofauna Aquática em Igarapés da Reserva de Desenvolvimento Sustentável Tupé, Manaus, AM
175
Igarapé da Cachoeira Igarapé das Pedras
MesespH Condutividade
μ Scm-1Temperatura
°CpH Condutividade
μ Scm-1Temperatura
°CPto1
Pto2
Pto3
Pto1
Pto2
Pto3
Pto1
Pto2
Pto3
Pto1
Pto2
Pto3
Pt1
Pto2
Pto3
Pto1
Pto2
Pto3
Abril02 4.8 4.8 5.4 7.9 6.7 9.8 25 26 31 5.0 5.4 - 8.4 9.7 11.0 24 24 28
Julho02 6.4 3.9 3.9 8.3 7.4 9.4 27 27 32 4.0 3.6 3.3 8.0 7.7 10.0 26 26 30
Agosto02 4.0 3.9 3.7 8.1 6.8 9.0 25 25 30 3.2 3.9 4.1 8.0 7.5 9.8 27 27 30
0
5
10
15
20
abr mai jun jul ago
Meses de observação (2002)
Nú
mero
de
inseto
sIg. Cachoeira Ig. Pedras
Fígura 1. Efeitos da interação entre os fatores “igarapés” e“meses de observação” sobre o número de insetos na Reservade Desenvolvimento Sustentável do Tupé, Manaus,Am.
coleta se obteve resultado significativo (p<0,01) (Tab.4). De um modo geral os ambientes de cabeceira e os dequeda d'água apresentaram maior diversidade defamílias de insetos. A menor diversida-de de insetos foiregistrada na desembocadura, ou foz (Fig. 2).
Na interação entre o fator igarapé, os meses e ospontos de coleta foi obtido resultado significativo(p<0,05) (Tab. 4).
Discussão
processo de decomposição por ser umambiente de águas com fracacorrenteza, quase parada. Nosambientes de cabeceira e queda d'águaforam registrados valores menores dec o n d u t i v i d a d e , s e n d o e s t e ssemelhantes entre si, devido acorrenteza ser mais forte e com menorquantidade de material orgânico emdecomposição. Segundo Esteves(1998), a variação diária dacondutividade elétrica da águafornece informações a respeito deprocessos importantes nos ecossiste-mas aquáticos, como produçãoprimária (redução dos valores) edecomposição (aumento dos valores).Os valores encontrados nos Igarapés daCachoeira e das Pedras devem estarsendo influenciados pela variaçãodiária da condutividade elétrica daágua.
Quanto aos períodos, no mês deabril, em todos os pontos do Igarapédas Pedras, foram registrados osmaiores valores de condutividadeelétrica devendo estar relacionada àforte chuva que caiu durante todo o dia

0
5
10
15
20
25
cabeceira queda d'água foz
Pontos de observação
Nú
mero
de
inseto
s
I. Cachoeira I. Pedras
Valores de pH, condutividadeelétrica e temperatura da água nomomento das coletas
Os valores de pH, condutividade etemperatura da água registrada paraos Igarapés da Cachoeira e das Pedrasestão dentro do padrão consideradopor Esteves (1998) e Walker (1987)representativo para a AmazôniaCentral. De acordo com Walker (1987)a condutividade elétrica dos igarapésda Amazônia Central varia entre 6 e 12
água e váriassubstancias orgânicas ionizáveis(Esteves, 1998).
Quanto aos ambientes, os maioresvalores de condutividade foramregistrados na desembocadura (igapó)estando, possivelmente, associados ao
μ S cm¹, podendo ser influenciada pelatemperatura, pH da
Figura 2. Efeitos da interação entre os fatores “igarapés” e“pontos de coleta” sobre o número de insetos na Reserva deDesenvolvimento Sustentável do Tupé, Manaus,Am.
176
Lopes
Causa de Variação G.L . S.Q. Q.M . F
Igarapé (A) 1 9.3021 9.3021 4.8684 *
Mês (B) 4 31.6382 7.9095 4.1395 **
Pontos de coleta (C) 2 8.4445 4.2223 2.2098 NS
Interação A x B 4 30.9081 7.7270 4.0440 **
Interação A x C 2 24.8612 12.4306 6.5057 **
Interação B x C 8 126.2111 15.7764 8.2567 **
Interação A x B x C 8 34.4986 4.3123 2.2569 *
(Tratamentos) 29 265.8639 9.1677 -
Resíduo 120 229.2875 1.9107 -
* Nível de significância de 5%** Nível de significância de 1%NS Não significativo nos níveis de 5 e 1%
Tabela 4. Análise de variância referente ao número de insetos observados na Reserva de DesenvolvimentoSustentável do TUPÉ (Manaus,Am), em dois igarapés, durante cinco meses de 2002 e em diferentes pontos deamostragem.

de em termos de famílias em igarapés sem alteraçãoquando comparados com sistemas alterados (Cleto-Filho
Walker, 2001; Couceiro ., 2001). As diferenças nadiversidade de famílias obtidas entre os dois igarapésestudados não são devidas à alterações no sistema e simdevido à disponibilidade de foliço e nível das águas.Podemos considerar que nestes dois igarapés da RDSTupé, a entomofauna se encontra bem representada.
Na interação entre o fator igarapé e os meses deobservação se obteve resultado significativo (p<0,01).De acordo com Irmler (1981) quando o canal do rio oulago inunda, os organismos aquáticos são lavados,podendo também migrar para outra área alagada ouemergir. A menor ocorrência de insetos no período deenchente verificada no Igarapé da Cachoeira pareceestar relacionada com a lavagem e retirada do substratoque ocasionou a migração dos organismos aquáticos. Amaior quantidade de indivíduos obtidos no período dacheia estava relacionada com a ocorrência de grandequantidade de substrato folhiço. Na vazante, se obtevemenor quantidade de indivíduos em função da água estarrestrita ao canal do igarapé e da quantidade de folhiçosubmerso. Segundo Irmler (1981), quando o nível da águacai, os organismos aquáticos se deslocam para os corposd'água permanentes ou mostram adaptações parasobreviver no período seco.
No Igarapé das Pedras foi verificado a maiorocorrência de insetos no período da vazante (julho),devido à água estar restrita ao canal do igarapé e apresença de folhiço principalmente na área de quedad'água; no período da enchente foi registradocorrenteza forte e pouco substrato folhiço e no períodode cheia registrou-se correnteza forte e a presença degrande quantidade de areia misturada ao substratofolhiço, o que provavelmente, tornou o substrato poucofavorável para a manutenção dos organismos. Para osdois igarapés, a ocorrência dos insetos nos períodos davazante, enchente e cheia está relacionada com acorrenteza e disponibilidade de substrato.
Na interação entre o fator igarapé e os pontos decoleta se obteve resultado significativo (p<0,01). De ummodo geral os ambientes de cabeceira, ou área de
& et al
Distribuição de insetos aquáticos nos Igarapés daCachoeira e das Pedras
Composição da Entomofauna Aquática em Igarapés da Reserva de Desenvolvimento Sustentável Tupé, Manaus, AM
177
de coleta. Este fato corrobora aafirmação de Esteves (1998), de que,em um período de 24 horas se podeesperar que ocorram alterações do pHdo meio, alteração na condutividade,que podem interferir na estrutura dascomunidades.
Foram obtidos 4.534 indivíduos,distribuídos em 8 ordens e 26 famíliasde insetos (Tabelas 1 e 2), que éc o n s i d e r a d o o g r u p o m a i srepresentativo, em termos dediversidade e abundância em águascorrentes (Hynes, 1970; KikuchiUieda, 1998).
Para o Igarapé das Pedras foramreconhecidas 22 ordens e/ou famíliasde insetos e 27 para o Igarapé daCachoeira. A maior riqueza em termosde famílias obtida no Igarapé daCachoeira se deve ao fato de se tratarde um igarapé maior, que dispõeconsequentemente, de um maior fluxode água, maior quantidade desubstrato raízes e folhiço submersodisponíveis para a manutenção daentomofauna. Reiss (1977) estudandoa fauna de invertebrados da área doLago do Tupé encontrou indivíduospertencentes aos Trichoptera,Chironomidae, Chaoboridae, Odonata,Ceratopogonidae. De acordo comFittkau . (1975) os insetos dosigarapés de terra firme estão representados por larvas de Trichoptera,Simullidae, Chironomidae, Ephemeroptera, Odonata, Plecoptera, Megaloptera, Coleoptera (Girinidae) eHeteroptera .
Estudos têm mostrado que a faunade insetos apresenta maior diversida-
Riqueza da Entomofauna doIgarapé da Cachoeira e Igarapé dasPedras
&
et al-
--

manuscrito. Aos revisores anônimosque muito contribuíram para arealização deste.
Callisto, M.; Esteves, F.A. 1998.Biomonitoramento da macrofaunabentônica de Chironomidae emdois igarapés amazônicos sobinfluência das atividades de umamineração de bauxita.Nessimian, J.L. A.L.(Eds.).
. Series OecologiaBrasiliensis, vol. V. PPGE-UFRJ. Riode Janeiro, Brasil. p. 299-308.
Cargnin-Ferreira, E. 1998. Fatoresinfluindo na distribuição de gruposfuncionais de macroinvertebradosaquáticos em pequenos tributáriosdo rio Jaú, Amazônia Central.Dissertação de Mestrado, INPA/UA,Manaus. 52pp.
Cleto-Filho, S.E.N.; Walker, I. 2001.Efeitos da ocupação urbana sobre amacrofauna de invertebradosaquáticos de um igarapé da cidadede Manaus/ AM Amazônia Central.
31(1): 69-89.Couceiro, S.R.M.; Hamada, N;
Ferreira, R. 2001.
, comosubsídio para avaliação de impactoambiental. Anais da X Jornada deIniciação Científica, PIBIC/INPA,65-68.
Da-Silva, E. 1998. Estratégias deadaptação das espécies deEphemeroptera às condiçõesambientais da Restinga de Maricá,Estado do Rio de Janeiro.
Bibliografia Citada
In:
Ecologia de InsetosAquáticos
ActaAmazonica,
Estrutura decomunidade de entomofauna emigarapés sob a influência do RioNegro em Manaus, AM
In:
& Carvalho,
178
Lopes
drenagem, e os de queda d'água, apresentaram maiordiversidade de famílias de insetos. A menor diversidadede insetos foi registrada na desembocadura, ou foz. Deacordo com Irmler (1975), em igarapés com elevadaflutuação do nível da água a distribuição daconcentração de oxigênio representa o fator maisimportante para a comunidade bêntica, sendoprovavelmente este o fator responsável pela menordiversidade registrada na desembocadura, ou foz.
De acordo com Junk . (1989) as condiçõeshidrológicas do rio, considerando sua área de drenageme zona sujeita a inundação; e os aspectos biológicos,como a troca de organismos, sua biomassa e energia,devem ser considerados como uma unidade indivisível.Para os Igarapés da Cachoeira e das Pedras, o efeito deinteração, ou interdependência significativo (p<0,05),verificado entre os fatores igarapé, meses e pontos decoleta sugere que estes aspectos também devem serconsiderados como uma unidade indivisível.
Segundo Walker (1990), em igarapés de água preta afauna existente obtém seus nutrientes básicos da própriafloresta, através da serrapilheira (folhiço) caída na água,corroborando os dados obtidos no presente estudo ondea maior quantidade de insetos obtido no período de cheiaestá relacionado a maior quantidade de substratofolhiço. Segundo Walker (1990) o segredo dos igarapés eigapós da Amazônia é que “a fauna está, onde está oalimento”.
Considerando que os ambientes de cabeceira dosigarapés da Amazônia dependem quase queexclusivamente do ambiente terrestre (Sioli, 1971) e obom estado de preservação em que se encontram osIgarapés da Cachoeira e das Pedras, da RDS Tupé, énecessário que se tome medidas para manter e preservara floresta localizada as margens destes igarapés a fim demanter a diversidade da fauna de invertebrados e omodelo hidrográfico natural buscando preservar estaimportante fonte de água potável.
Ao Dr. Sidney Alberto do Nascimento Ferreira peloauxílio na análise estatística dos dados e sugestões ao
et al
Agradecimentos

Composição da Entomofauna Aquática em Igarapés da Reserva de Desenvolvimento Sustentável Tupé, Manaus, AM
179
(Eds.).
. Editora do Instituto Nacional dePesquisas daAmazônia. p.397-402.
Freitas, C.E.C. 1994.
. Dissertação deMestrado INPA/UA, Manaus. 65pp.
Hynes, H.B.N. 1970. . 3ªed. Toronto Press, Toronto. 555pp.
Irmler, U. 1975. Ecological studies of the aquatic soilinvertebrates in three inundation forest of CentralAmazonia. ,V (3): 337-409.
Irmler, U. 1981. Überlebensstrategien von Tieren ims a i s o n a l ü b e r f l u t e t e n A m a z o n i s c h e nÜberschwemmungswald. Zool. Anz. Jena., 2061/2:26-38.
Irmler, U.; Junk, W.J. 1982. The inhabitation ofartificially exposed leaf samples by aquaticmacroinvertebrates at the margin of amazonianinundation forest. , 23: 64-75.
Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The floodconcept in river floodplain system. In: Dodge D P(ed.). Proc. Int. Large River Symp (LARS),
106: 110-127.Junk, W.J.; Wantzen, K.M. 2004. The flood pulse
concept: new aspects, approaches and applicationsan update. . Welcomme, R.L T. Petr (eds.).Proceedings of the Second International Symposiumon the management of large rivers for fisheries,Volume 2. Food and Agriculture Organization &Mekong River Comission. FAO Regional Office for Asiaand the Pacific, Bangkok. RAP Publication 2004/16,p. 117-149. Obtido em 04/03/2005 emhttp://www.lars2.org proceedings.
Kikucki, R.M.; Uieda, V.S. 1998. Composição dacomunidade de invertebrados de um ambiente lóticotropical e sua variação temporal e espacial.Nessimian, J.L. A.L. Carvalho (Eds.).
. Series Oecologia Brasiliensis, vol.V. PPGE-UFRJ. Rio de Janeiro, Brasil. p. 157-173.
McCafferty, W.P.; Provonsha, A.V. 1983.
, Jonesand Bartlett publishers, Inc. Portola Valley, Boston.448pp.
Bases Científicas para estratégias depreservação e desenvolvimento da Amazônia: Fatos ePerspectivas
Comunidades bênticas decachoeiras da bacia do rio Urubuí (Amazonas -Brasil): Persistência e colonização
,The ecology of running waters
Amazoniana
Tropical Ecology
. .Can. Spec.
Pub. Fish.Aquat. Sci.,
In:
In:Ecologia de
Insetos Aquáticos
AquaticEntomology. The fishermen's and ecologists'sillustrated guide to insects and their relatives
&
&
Nessimian, J.L. A.L. Carvalho(Eds.).
. Series OecologiaBrasiliensis, vol. V. PPGE-UFRJ. Riode Janeiro, Brasil. 29-40.
Erwin, T.L. 1983. Beetles and otherinsects of tropical forest canopiesat Manaus, Brazil, sampled byinsecticidal fogging.
.Blacwel l Sc ient i f ic Publ ic.
. p.59-75.Esteves, F.A. 1998.
. Editora Interciência,Rio de Janeiro, Brasil. 602pp.
Fernández, H.R.; Domínguez, E.2001.
. E d i t o r i a lUnivers i tar ia de Tucumán,Tucumán,Argentina. 282p.
Ferreira, A.C.R. 1996.
.Dissertação de Mestrado INPA/UA,Manaus. 76pp.
Fittkau, E.J. 1967..
Vol. 3 (Limnologia): 97-108.
Fittkau, E.J.; Irmler, U.; Junk, W.J.;Reiss, F.; Schmidt, G.W. 1975.Product iv ity, biomass, andpopulation dynamics in Amazonianwater bodies. I
. Springer Verlag,New York, Berlin. p. 289-311.
Fonseca, C.R.V.; Rafael, J.A. 1991.Entomologia Sistemática - Umaferramenta bás ica para odesenvolvimento na Amazônia.Val, A.L., Figliuolo, R., Feldberg, E.
&Ecologia de Insetos
Aquáticos
In:s Tropical rain
forest: Ecology and Management
Fundamentos deLimnologia
Guía para la determinaciónde los artrópodos bentônicoss u d a m e r i c a n o s
Estudo da faunade Perlidae (Plecoptera) em doisigarapés da Amazônia Central
,
On the ecology ofamazonian rainforest streamsAtas do Simpósio sobre a BiotaAmazônica,
n:Tropical Ecological
Systems. Trends in terrestrial andaquatic research
In:
Sutton,S.L. . (Ed .)
Edinburgh
F.B. Golley E.Medina, (Eds.)
et al
&

180
Lopes
Sioli, H. 1971. Resumos de trabalhosrelativos a Amazônia Brasileira.
, 16:1-7.
Walker, I. 1985. On the struture andecology of the macrofauna in theCentral Amazonian forest stream.I g a r a p é d a C a c h o e i r a .
, 122:137-152.Walker, I. 1987. The biology of streams
as part of Amazonian forestecology. , 43: 279-287.
Walker, I. 1990. Ecologia e Biologia dosigapós e igarapés. ,11(64): 44-53.
Walker, I. 1994. The benthic litter-dwelling macrofauna of theAmazonian forest stream Tarumã-Mirim: patterns of colonization andtheir implications for communitystability. ,291(2): 75-92.
Cadernos de Ciências da Terra
Hydrobiology
Experientia
Ciência Hoje
Hydrobiologia
Walker, I. 2003. The benthic insectfauna of the blackwater foreststream Rio Tarumã-Mirim (Manaus,Amazonas): patterns of populationdynamics and their implications forecosystem stability. ,XVII (3/4): 471-480.
Amazoniana
Merritt, R.W.; Cummins, K.W. 1979.. (2ª ed.), Ed.
Kendall/Hunt Publishing Company, Iowa, 441pp.Moulton, T.P. 1998. Saúde e integridade do ecossistema e
o papel dos insetos aquáticos. In: Nessimian, J.L.A.L. Carvalho (Eds.). .Series Oecologia Brasiliensis, vol. V. PPGE-UFRJ. Riode Janeiro, Brasil. p. 281-298.
Nessimian, J.L. 1985.
.Dissertação de Mestrado INPA/UA, Manaus. 108pp. +15 tabelas
Patrick, R.; Aldrick, F.A.; Cairns Jr., J.; Drouet, F.; Hohn,M.H.; Roback, S.S.; Skuja, H.; Spangler, P.J.; Swabey,Y.H.; Whitford, L.A. (1966). The catherwoodfoundat ion peruvian-amazon expedit ion:Limnological and systematic studies.
Philadelphia. n. 14. 375pp.Pes, A.M.O. 2001.
Dissertação deMestrado INPA/UA, Manaus. 166pp.
Reiss, F. 1977. Qualitative and quantitative investigations on the macrobenthic fauna of Central Amazonlakes. 1. Lago Tupé, a black water lake on the lowerRio Negro. 6 (2): 203-235.
Rosenberg, D.M.; Resh, V.H (eds.).1993.
Chapman Hall, Inc. New York and London. 488pp.
An introduction tothe aquatic insects of North America
Ecologia de Insetos Aquáticos
Estudo sobre a biologia e aecologia da fauna invertebrada aquática na liteirasubmersa das margens de dois lagos do arquipélagode Anavilhanas (Rio Negro, Amazonas, Brasil)
,
Monographs ofthe Academy of Natural sciences of Philadelphia.
Taxonomia e estrutura de comunidadede Trichoptera (Insecta) no município de PresidenteFigueiredo, Amazonas, Brasil.
,-
Amazoniana,Freshwater
biomonitoring and benthic macroinvertebrates.
&
&

Diversidade BiológicaCapítulo 13
Diversidade e composição da ictiofaunado Lago Tupé
RESUMO - O sistema hidrológico do rio Negro é caracterizado pela variabilidade debiótopos colonizado por muitos organismos aquáticos, especialmente os peixes com cercade 450 espécies. Considerando que nas Unidades de Conservação implantadas na bacia dorio Negro, as informações sobre a ictiofauna ainda são incipientes, esse trabalho propõeinvestigar a diversidade e composição específica sazonal da ictiofauna no lago que fazparte da Reserva de Desenvolvimento Sustentável do Tupé.Ahipótese a ser testada baseia-se na existência de diferenças sazonais na composição, diversidade e riqueza da ictiofaunaresultante das variações na disponibilidade de habitats durante o ciclo hidrológico. Ascoletas foram realizadas mensalmente de julho de 2002 a dezembro de 2003, com umabateria de malhadeira. A estrutura de associações de peixes foi descrita pela abundância
Maria Gercilia M. SOARES
Kedma Cristine YAMAMOTO
e-mail: [email protected] Nacional de Pesquisas da Amazônia (INPA),
CP. 478, 69011-090, Manaus-AM, Br.
e-mail: [email protected] Universitário Luterano de Manaus (CEULM/ULBRA),
Avenida Solimões 02, Conj. Atílio Andreazza, Bairro Japiim II. 69077-730.Departamento de Ciências Biológicas.
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

relativa, riqueza, diversidade, equitabilidade e número de indivíduos. Foram capturados2.292 exemplares distribuídos em 5 ordens, 22 famílias, 64 gêneros e 84 espécies. Acomposição específica da ictiofauna mostra variações sazonais em nível de família,espécie e abundância de indivíduos. Characidae, Hemiodontidae, Auchenipteridae,Curimatidae, Cichlidae foram às famílias mais abundantes e em nível de espécie
sp., e .Os resultados suportaram a hipótese de que existem diferenças sazonais na diversidade eriqueza da ictiofauna do lago. Essas condições são mantidas por meio de uma área de terrafirme, relativamente intacta e grande área alagada, inclusive na seca que permite aexistência de habitats para a colonização de muitas espécies de peixes.
Amazônia, rio Negro, peixes, igapó.
Acestrorhynchus falcirostris, Moenkhausia intermedia, Hemiodus immaculatus,
Hemiodus Hemiodus gracilis, Hypophthalmus edentatus Cyphocharax abramoides
PALAVRAS-CHAVE:
IntroduçãoOs peixes compõem o grupo de vertebrados mais
diversificado do mundo. Estimativas do total deespécies que possam estar presentes na imensa área dedrenagem da bacia amazônica variam entre 1500 e 5000(Bohlke ., 1978; Goulding ., 1988; Malabarba
., 1998)No sistema hidrológico do rio Negro, caracterizado
pela variedade de biótopos, como: praias arenosas,corredeiras, remansos, ilhas, paranás e lagos, já foramidentificadas cerca de 450 espécies (Goulding .,1988). Os autores consideram que as característicasquímicas das águas pretas, como: alto teor decomponentes húmicos, baixo pH e nutrientes, nãopodem ser considerados fatores limitantes para adiversidade de peixes e apontaram a necessidade deincrementar as amostragens nestes biótopos eestimaram a possibilidade da riqueza para mais de 700espécies.
Apesar da riqueza, a fauna de peixes nos lagos deágua preta tem sido pouco investigada quandocomparada com a de lagos de várzea. Informaçõesimportantes têm sido disponibilizadas sobre a estruturade comunidades de peixes no lago Prato, Arquipélago dasAnavilhanas (Saint-Paul ., 2000); associações entrepeixes e macrófitas aquáticas, Arquipélago dasAnavilhanas (Araújo-Lima ., 1986); migrações dosjaraquis no rio Negro (Ribeiro, 1983; Ribeiro Petrere,1990); riqueza, diversidade e relações tróficas em vários
et al et al etal .
et al
et al
et al&
habitats do rio Negro (Goulding .,1988); diversidade e distribuição nolago do Prato (Garcia, 1995);comportamento alimentar no rioNegro (Ferreira, 1981; CarvalhoGoulding, 1985; Borges, 1986), rioPreto da Eva (Marlier, 1968) e nahidrelétrica de Curuá-Una, (Ferreira,1984). Santos (1983) reveloudiferenças na estrutura de comunida-des de peixes capturados no lago Tupée no rio Negro através de pescaexperimental.
Na Amazônia, a conservação e o usosustentável da biodiversidade emecossistemas aquáticos representamum desafio, especialmente por causada escassez de conhecimento sobre osorganismos, suas relações e seuaproveitamento para o consumohumano. Nas Unidades de Conservaçãoimplantadas na bacia do Rio Negro,ainda são incipientes as informaçõessobre a ictiofauna. Neste contextoesse trabalho propõe investigar adiversidade e composição específicasazonal da ictiofauna no lago que fazparte da RDS Tupé, submetido à fortepressão antrópica pela proximidade deManaus. A hipótese a ser testada
et al
&
182
Soares Yamamoto&

baseia-se na existência de diferençassazonais na composição, diversidade eriqueza da ictiofauna no lago,resultante das variações na disponibili-dade de habitats durante o ciclohidrológico.
O Tu p é ( 0 3 º 0 2 3 5 , 4 S e60º15 17,5 W) é um lago de águapreta, situado na RDS Tupé à margemesquerda do rio Negro, na AmazôniaCentral, cerca de 25 km a oeste deManaus. A flutuação anual média donível da água é de 10 m. A enchente,em geral, começa em novembro econtinua até o final de junho, Fig. 1(Aprile Darwich, 2005, nestevolume), atingindo a cota máxima emjulho; a vazante começa em agosto e aágua flui do lago para o rio, atingindonível mais baixo em outubro.
As coletas dos peixes foramrealizadas mensalmente de julho de2002 a dezembro de 2003, próximo àestação E 10
com bateria demalhadeira, malhas variando de 20 a90mm entre nós, medindo 25m decomprimento e 2,5m de altura. Asmalhadeiras foram armadas na regiãode água aberta do lago, partindo daslaterais para o centro do lago ondepermaneceram durante 24h. Asdespescas ocorreram em intervalos de6h.
Os peixes foram identificados nocampo com o auxílio de chaves deidentificação (Géry, 1977; Malabarba
., 1998; Kullander ., 2003).Porém, várias espécies foram fixadasem solução de formol, 10%, etransportados para o laboratório paraserem identificadas com a colaboração
Material e Métodos
’ ”’ ”
T Fig. 1 (Darwich ,2005, neste volume)
&
et al.
et al et al
de especialistas do INPA.A estrutura de associações de peixes foi investigada
através da abundância relativa (%) em número deindivíduos (N) e biomassa (g), riqueza (R), índice dediversidade de Shannon-Wiener (H') (Krebs, 1989) e índicede equitabilidade (E) (Magurran, 1988). Os dados foramagrupados em quatro períodos do ciclo hidrológicoenchente, cheia, vazante e seca.
Nas coletas efetuadas na água aberta do lago Tupéforam capturados 2292 exemplares de peixes distribuídosem 5 ordens, 22 famílias, 64 gêneros e 84 espécies (Tab.1; Anexo 1). Deste total, a ordem Characiformes (10famílias, 30 gêneros e 48 espécies) predominou commaior número de indivíduos em todos os períodos, 76,3%na enchente, 50,6% na cheia, 49,7% na vazante e 85,8% naseca. Seguida pela ordem Siluriformes (7 famílias, 15gêneros e 20 espécies) capturados na enchente, cheia eseca. Clupeiformes (2 famílias, 3 gêneros e 3 espécies),Perciformes (2 famílias, 13 gêneros e 16 espécies) eGymnotiformes (1 família, 2 gêneros e 2 espécies) são asordens que apresentaram menor número de indivíduos(Fig. 1). A seca (N=1.307) e a enchente (N=579)correspondem aos períodos nos quais foram capturados asmaiores quantidades de exemplares (Fig. 3).
As famílias mais representativas em número deindivíduos, em ordem decrescente de abundância, foramCharacidae, Hemiodontidae, Auchenipteridae,Curimatidae, Cichlidae, Pimelodidae, Pristigasteridae,Ctenoluciidae, Prochilodontidae e Scianidae comdominância variando sazonalmente. Na enchente é maioro número de exemplares pertencentes à famíliaCharacidae (61,3%), Auchenipteridae (13,5%) eCtenoluciidae (5,5%); na cheia, Hemiodontidae (25,3%),Charac idae (12 ,6%) , Pime lod idae (12 ,3%) ,Auchenipteridae (5,7%) e Cichlidae (4,6%); na vazantePimelodidae (26,9%), Hemiodont idae (20%)Auchenipteridae (10,3%) e Curimatidae (8,3%), efinalmente na seca Characidae (49,3%), Hemiodontidae(19,1%), Curimatidae (8,3%) e Cichlidae (7,4%) (Fig. 2).
As espécies com maior abundância no lago foram(27,1%),
(7,29%), (6,68%),
Resultados
Acestrorhynchus falcirostris Moenkhausiaintermedia Hemiodus immaculatus
Diversidade e composição da ictiofauna do Lago Tupé
183

Ordem, FamÍlia, Nome vulgar Enchente Cheia Vazante SecaGênero e Espécie N (%) N (%) N (%) N (%)Clupeiformes
PristigasteridaePellona flavipinnis (Valenciennes, 1836) apapá amarelo, sardinhão 2 0,3 3 1,1 10 6,9 1 0,1IIisha amazonica (Miranda-Ribeiro, 1920) apapá 1 0,2 5 1,9EngraulididaeLycengraulis batesii (Günther, 1868) manjuba 9 1,6 37 14,2Characiformes
ErythrinidaeHoplias malabaricus (Bloch, 1794) traíra 2 0,8 24 1,8CtenoluciidaeBoulengerella lucius (Cuvier, 1816) pirapucu 32 5,5 8 3,1 22 1,7AnostomidaeSchizodon vittatum (Valenciennes, 1850) aracú 9 6,2Schizodon fasciatus, Agassiz, 1829 aracú 1 0,2 1 0,1Anostomus sp. aracú 1 0,2 2 0,8 23 1,8Anostomus taeniatus (Kner, 1858) aracú 2 0,3Rhytiodus argenteofuscus (Kner, 1858) aracú, pau-de-nego 1 0,2Rhytiodus sp. aracú, pau-de-nego 7 0,5Leporinus affinis (Günther, 1864) aracú flamengo 2 0,3 6 0,5Laemolyta sp. aracú caneta 6 0,5HemiodontidaeAnodus elongatus, Agassiz, 1829 orana, charuto 5 0,9 1 0,4 10 6,9Anodus orinocensis (Steindachner, 1887) orana, charuto 1 0,2Hemiodus gracilis Günther, 1864 orana branca 15 2,6 23 8,8 2 1,4 59Hemiodus immaculatus Kner, 1858 orana, cubiu 9 1,6 38 14,6 27 18,6 79 6,0Hemiodus sp. orana 5 1,9 109 8,3Hemiodus goeldi Steindachner, 1908 orana 2 0,3Micromischodus sugilatus (Roberts, 1971) orana colarinho 3 0,2ProchilodontidaeSemaprochilodus insignis (Jardine & Schomburgk, 1841) jaraqui escama grossa 1 0,2 7 2,7 19 1,5Semaprochilodus taeniurus (Valenciennes, 1817) jaraqui escama fina 1 0,2 9 3,4 8 0,6CurimatidaeCurimatella meyeri (Steindachner, 1882) branquinha 5 0,4Curimata vittata (Kner, 1858) branquinha 53 4,1Curimata sp. branquinha 1 0,4Potamorhina altamazonica (Cope, 1878) branquinha cabeça lisa 2 1,4 1 0,1Potamorhina latior (Spix, 1829) branquinha peito-de-aço 2 1,4Psectrogaster rutiloides (Kner, 1858) branquinha cascuda 1 0,2 5 3,4Cyphocharax abramoides (Kner, 1859) branquinha 13 2,2 1 0,4 48 3,7Curimatopsis sp. branquinha 3 2,1CynodontidaeHydrolycus scomberoides (Cuvier, 1816) Peixe-cachorro 1 0,7Cynodon gibbus Spix & Agassiz, 1829 zé-do-ó 1 0,4AcestrorhynchidaeAcestrorhynchus falcirostris (Cuvier, 1819) Peixe-cachorro 41 7,1 1 0,4 1 0,7 578 44,2Acestrorhynchus microlepis (Schomburgk, 1841) Peixe-cachorro, dente de cão 32 5,5 1 0,7CharacidaeCatoprion mento (Cuvier, 1819) piranha 2 0,2Serrasalmus elongatus Kner, 1860 piranha mucura 1 0,4 2 1,4Serrasalmus rhombeus (Linnaeus, 1766) piranha preta 1 0,1Serrasalmus sp. piranha 1 0,7Agoniates anchovia Eigenmann, 1914 cruzador 1 0,7 2 0,2Brycon melanopterus (Cope, 1872) matrinchã 1 0,2 4 1,5 1 0,1Ctenobrycon hauxwellianus (Cope, 1870) piaba 5 3,4Holobrycon sp. piaba 3 1,1Moenkhausia lepidura (Kner, 1858) piaba 23 8,8 9 0,7Moenkhausia intermedia Eigenmann, 1908 piaba 167 28,8Moenkhausia sp. piaba 2 0,8 2 0,2Tetragonopterus sp. piaba 47 3,6Bryconops sp. piaba 5 0,4Lonchogenys sp. 1 piaba 51 8,8Lonchogenys sp.2 piaba 63 10,9
Tabela 1. Lista das espécies capturadas nos períodos de enchente, cheia, vazante e seca no lago Tupé.
continua >
184
Soares Yamamoto&

> continuação
185
Diversidade e composição da ictiofauna do Lago Tupé
Ordem, FamÍlia,Gênero e EspécieGymnotiformes
HypopomidaeBrachyhypopomus sp.RhamphichthyidaeRhamphichthys sp.Siluriformes
PimelodidaeHypophthalmus edentatus Spix & Agassiz, 1829Hypophthalmus fimbriatus Kner, 1858Hypophthalmus marginatus Valenciennes, 1840Pimelodus blochii Valenciennes, 1840Sorubim lima (Bloch & Schneider, 1801)AuchenipteridaeAgeneiosus vitattus Steindachner, 1908Ageneiosus brevifilis Valenciennes, 1840Ageneiosus sp.1Ageneiosus sp.2Auchenipterus sp.Centromochlus heckelii De Filippi, 1853Parauchenipterus galeatus (Linnaeus, 1766)Tatia sp.Auchenipterichthys thoracatus (Kner, 1858)CallichthyidaeHoplosternum littorale (Hancock, 1828)DoradidaeHassar sp.Scorpiodoras sp.Oxydoras niger (Valenciennes, 1821)LoricariidaeAncistrus sp.Rineloricaria sp.Perciformes
CichlidaeAcarichthys heckelii (Müller & Troschel, 1849)Cichla temensis Humboldt, 1821Cichla monoclus Spix & Agassiz, 1831Cichla sp.Geophagus sp.Astronotus ocellatus (Agassiz, 1831)Satanoperca sp.Satanoperca acuticeps (Heckel, 1840)Crenicichla sp.Uaru amphiacanthoides Heckel, 1840Acaronia nassa (Heckel, 1840)Chaetobranchus sp.Heros sp.SciaenidaePlagioscion squamossissimus (Heckel, 1840)Pachyurus sp.Pachypops sp.TOTAL
Nome vulgar Enchente Cheia Vazante SecaN (%) N (%) N (%) N (%)
sarapó 2 0,3
sarapó 1 0,1
mapará 1 0,2 6 2,3 35 24,1 1 0,1mapará 3 0,5 2 1,4mapará 0,0 26 10,0 2 1,4 1 0,1mandi 3 0,5 1 0,4
bico de pato 16 1,2
mandubé 25 4,3 1 0,4mandubé 1 0,4mandubé 1 0,2 1 0,7mandubé 1 0,2
mandi 13 5,0mandi peruano 21 3,6 11 7,6 19 1,5
camgati 4 2,8 4 0,3mandi 31 5,4 2 0,8 9 0,7mandi 26 4,5 19 1,5
tamoatá 2 1,4
botinho 1 0,41 0,2 9 0,7
cuiu-cuiu 1 0,2
bodó 2 0,3 2 0,8cachimbo 1 0,4 5 0,4
acará 4 0,3tucunaré 5 0,9 6 2,3 27 2,1tucunaré 1 0,1tucunaré 2 0,2
acará, papa-terra 6 0,5acara-açú 1 0,4
acará 2 0,8 27 2,1acará 14 1,1
jacundá 2 0,8 1 0,7 1 0,1acará bararuá 1 0,4 2 0,2
acará 1 0,2 10 0,8acará prata 2 0,2acará preto 1 0,1
pescada branca 1 0,2 18 6,9 5 3,4pescada, corvina 2 0,2pescada, corvina 2 0,2
125 84 63 185
Hemiodus Hemiodusgracilis M. intermedia (
H.immaculatusLycengraulis batesii
Hypophthalmus edentatusH. immaculatus
A. falcirostris
sp. (4,97%) e(4,32%). piaba)
é dominante na enchente,(orana, cubio) e
(manjuba) nacheia,(mapará) e na vazantee (peixe-cachorro) naseca (Fig. 3).
Há diferenças na diversidade entre os períodos commaiores valores na cheia e menores na seca. AEquitabilidade não variou muito entre os períodos, masapresentou menor valor na seca (0,614), provavelmentepor causa da dominância de . Na cheia, émaior a uniformidade no número dos exemplares entreas espécies o que aumenta a diversidade (H´= 4,13) (Fig.3).
A. falcirostris

Anexo 1. Fotos das principais espécies de peixes capturadas no lago Tupé.
186
continua >
Ilisha amazonica
Pellona flavipinnis
Lycengraulis batesii
Hoplias malabaricus
Boulengerella lucius
Leporinus affinis
Anostomus .sp
Hemiodus immaculatus
Hemiodus gracilis
Anodus orinocensis
Semaprochilodus insignis
Curimata vittata
Potamorhina latior
Potamorhina altamazonica
Psectrogaster rutiloides
Cyphocharax abramoides
Serrasalmus elongatus
Acestrorhynchus microlepis
Soares Yamamoto&

> continuação
187
Acestrorhynchus falcirostris
Moenkhausia lepidura
Lonchogenys sp.
Ctenobrycon hauxwellianus
Moenkhausia intermedia
Hypophthalmus edentatus
Hypophthalmus fimbriatus
Ageneiosus vittatus
Auchenipterichthys thoracatus
Parauchenipterus galeatus
Pseudepapterus sp.
Centromachlus heckelii
Hoplosternum littorale
Escorpiodoras sp.
Ancistrus sp.
Cichla temensis
Acarichthys heckelii
Plagioscion squamosissimus
Diversidade e composição da ictiofauna do Lago Tupé

Figura 1. Abundância relativa (%) das ordens de peixescapturadas no lago Tupé.
Figura 2. Abundância relativa (%) das famílias de peixes maiscapturadas no lago Tupé.
188
Discussão
A ictiofauna capturada no lago Tupémostra dominância de Otophysi(Characiformes, 77,1%; Siluriformes,13,5%). Os Perciformes (6,3%) e osPristigasteridae (3%) ocorreram compoucos exemplares. As freqüênciasrelativas das ordens no lago estão emconformidade com aquelas indicadaspara os rios (Ferreira, 1993; Ferreira
., 1988) e lagos de várzea daAmazônia Central (Junk ., 1983;Souza-Pereira, 2000; Saint-Paul .,2000; Yamamoto, 2004). A composiçãoespecífica da ictiofauna no lago mostravariações sazonais em nível defamílias, espécie e abundância deindivíduos.
Os maiores números de exemplarescapturados em todos os períodospertencem às famílias Characidae,Hemiodontidae, Auchenipteridae,Curimatidae e Cichlidae. Curimatidaee Hemiodontidae, que são famíliasconstituídas por várias espécies queformam cardumes e realizam migra-ções de longa (rio acima) e curta(dentro da RDS Tupé) distância.Desenvolvem parte do seu ciclo de vidanos lagos, na época de alagação, eparte nos rios, na época de seca. Nosistema lago do Rei foi observadomovimentos migratórios do lago emdireção ao rio Amazonas envolvendoespécies dessas famílias (FernandesMerona, 1988).
Os hemiodontídeos,(cubiu) e (orana branca)
e curimatídeos,e
(branquinhas), foram os peixes maisabundantes na cheia e vazante,respectivamente. Essas espéciesentram no lago com a enchente embusca de abrigo e alimentação, os
etal
et alet al
H. immacula-tus H. gracilis
C. abramoides, P.rutiloides, P. altamazonica P. latior
&
0 20 40 60 100
Characidae
Auchenipteridae
Acestrorhynchidae
Ctenoluciidae
Hemiodontidae
Curimatidae
Pristigasteridae
Anostomidae
Cichlidae
Pimelodidae
Rhamphichthyidae
Doradidae
Loricariidae
Prochilodontidae
Sciaenidae
Erythrinidae
Cynodontidae
Engraulididae
Callichthyidae
Abundância relativa (%)
Enchente
Cheia
Vazante
Seca
0 20 40 60 80 100
Characiformes
Clupeiformes
Siluriformes
Perciformes
Gymnotiformes
Abundância relativa (%)
EnchenteCheia
VazanteSeca
Soares Yamamoto&

189
Diversidade e composição da ictiofauna do Lago Tupé
Enchente
0 20 40 60 80 100
M. intermedia
Lonchogenys sp.1
Lonchogenys sp.2
A. falcirostris
B. lucius
A. vitattus
C. heckelii
C. abramoides
H. immaculatus
C. temensis
H. fimbriatus
A. taeniatus
H. goeldi
Ancistrus sp.
A. microlepis
Tatia sp.
A. thoracatus
H. gracilis
L .batesii
A. elongatus
P. blochii
P. flavipinnis
L. affinis
Brachyhypopomus sp.
Abundância relativa (%)
continua >

0 20 40 60 80 100
H. immaculatus
M. lepidura
L. batesii
P. squamossissimus
S. taeniurus
S. insignis
C. temensis
I.amazonica
P. flavipinnis
Holobrycon sp.
Moenkhausia sp.
Ancistrus sp.
Satanoperca sp.
H..marginatus
H. gracilis
Auchenipterus sp.
B. iucius
H. edentatus
Hemiodus sp.
B. melanopterus
S. taeniurus
H. malabaricus
Tatia sp.
Anostomus sp.
Crenicichla sp.
Abundância relativa (%)
Cheia
> continuação
190
continua >
Soares Yamamoto&

Vazante
0 20 40 60 80 100
H. edentatus
H. immaculatus
C. heckelii
P. flavipinnis
A. elongatus
S. vittatum
C. hauxwellianus
P. rutiloides
P. squamossissimus
P. galeatus
Curimatopsis sp.
H. gracilis
P. altamazonica
P. latior
S. elongatus
H. fimbriatus
H. marginatus
H. littorale
Abundância relativa (%)
191
> continuação
continua >
Diversidade e composição da ictiofauna do Lago Tupé

Seca
0 20 40 60 80 100
A. falcirostris
Hemiodus sp.
H. immaculatus
H. gracilis
C. vittata
C. abramoides
Tetragonopterus sp.
Cichla temensis
Satanoperca sp.
H. malabaricus
Anostomus sp.
B. lucius
S. insignis
C. heckelii
A. thoracatus
C. heckelii
S. lima
A. thoracatus
S. acuticeps
A. nassa
Tatia sp.
Scorpiodoras sp.
M. lepidura
S. taeniurus
Rhytiodus sp.
L. affinis
Laemolyta sp.
Geophagus sp.
Abundância relativa (%)
192
Figura 3. Abundância relativa (%) das principais espécies nos períodos de enchente, cheia,vazante e seca no lago Tupé.
> continuação
Soares Yamamoto&

hemiodontídeos o fitoplâncton (Claro-Jr. 2003); na águaaberta, e os curimatídeos os detritos (Soares .,1986; Ferreira, 1993; Pouilly 2003) na cabeceirado lago. A retração das águas força os peixes a migraremem direção à saída do lago que é próxima ao sítioamostral e, portanto torna passíveis de captura pelasmalhadeiras.
Nas famílias Characidae, Auchenipteridae,Engraulididae, Acestrorhynchidae e Cichlidae a maioriados peixes é residente, desenvolvendo todo o ciclo devida no lago como as espécies de pequeno porte,
(piaba), (piaba),sp.1 (piaba), sp.2 (piaba) e(manjuba); e de médio porte como (peixecachorro), (peixe cachorro), sp.(mandi), (mandi), (mandi),
(mapará), (mapará),(tucunaré), spp. (acará) e
(pirapucu). Durante o ciclo hidrológico, essasespécies estiveram presentes em quase todas aspescarias, embora a abundância tenha apresentadovariações. A maioria é considerada espécie pelágica eexplora os recursos disponíveis na área aberta do lago.Como exemplos podem ser mencionados
e que ingeremzooplâncton (Goulding , 1988; Garcia, 1995),
, que comem insetos (Garcia, 1995)e e
cuja dieta é constituída porpeixes (Goulding 1980; Goulding ., 1988; Garcia,1995; Merona , 2001).
Os caracídeos de pequeno porte (sp.1 e sp.2) dominantes na
enchente e cheia têm as populações reduzidas durante aseca. Isto provavelmente está relacionado à fortepressão de predação exercida por espécies como
, que são vorazes predadores (Goulding., 1988; Claro Jr., 2003; Oliveira Jr., 1998; Soares,, 1986) muito freqüentes e às vezes dominantes na
comunidade íctica do Tupé. Logo após o início daalagação os pequenos caracídeos e ciclídeos járecuperaram as perdas ocorridas na seca, indicado peladominância de sp.1 e
sp.2, na enchente.Mudanças na composição específica causadas pelo
,et al
M.intermedia M. lepidura Lonchogenys
Lonchogenys L. batesiiA. falcirostris -
A. microlepis TatiaC. heckelii A. thoracatus H.
marginatus H. edentatus C.temensis Satanoperca B.lucius
H. marginatus,H. edentatus, L. batesii A. surinamensis
et al. P.galeatus, C. heckelii
A. falcirostris, A. microlepis, B. lucius, S. lima C.temensis, H. malabaricus
et alet al.
M. lepidura,Lonchogenys Lonchogenys
A.falcirostris, B. lucius, P. flavipinnis, C. temensis, H.malabaricus etal - etal.
M. intermedia, LonchogenysLonchogenys
et al.,
os
ciclo hidrológico têm sido amplamenterelatadas em lagos de várzea dosistema Solimões/Amazonas, Inácio,Rei, Catalão (Merona Bittencourt,1993; Saint-Paul ., 2000; Vale,2003), lago do rio Negro,(Garcia, 1995; Saint-Paul ., 2000)e Rios Machado e Madeira (Goulding,1980). Na maioria dos trabalhos acimacitados, é ressaltado que durante operíodo de alagação (enchente) váriasespécies migram da calha do rio paraos lagos e no período da vazante,retornam ao rio. Na seca os lagos sãohabitados principalmente por espéciestípicas de ambientes lênticos,conforme observado no lago Tupé.
Nos períodos de enchente e cheia aabundância de peixes é grande, porémcom o aumento da área alagada e amaior disponibilidade de habitats pararefúgios, a malhadeira por ser umaparelho de pesca passivo, deixapossivelmente, de capturar muitasespécies e exemplares afetando assim,as amostragens durante este período.Na cheia e vazante as amostragenstambém foram prejudicadas pelapresença constante de botos queconsumiam os peixes e destruíam asmalhadeiras, resultando na menorcaptura de indivíduos. Na seca, aretração das águas diminui a áreaalagada e obriga os peixes a sedeslocarem em direção à área centrale mais profunda do lago, ondepermanecem até o próximo período dealagação. Uma vez concentrados naárea aberta do lago os peixes sãofacilmente capturados pelas malha-deiras.
Os valores de diversidade eequitabilidade revelam uma comuni-dade onde existe uma grandediferença na distribuição dos númerosde exemplares entre as espécies. Isso
&et al
et alPrato
193
Diversidade e composição da ictiofauna do Lago Tupé

por causa da dominância de espéciesque formam cardumes tanto asmigradoras ( ) como asresidentes no lago (
e ).A dominância de naenchente e de na seca,favorece a similaridade entre osvalores de diversidade nos períodos,embora na vazante a riqueza sejamuito baixa. Na cheia e vazante adominância de
e , resulta emmenor diferença na distribuição donúmero de exemplares entre asespécies elevando os valores daequitabilidade. Santos (1983) tambémindica essas espécies como abundantesno lago Tupé na década de 80. Apresença de espécies dominantes nacomunidade de peixes no Tupéconcorda com o padrão que tem sidodetectado em comunidades naturaisem outros lagos da Amazônia Centralde águas pretas (Garcia, 1995) e águabranca (Saint-Paul ., 2000;Yamamoto, 2004).
Comparando a riqueza ediversidade desse lago (R=157, H'=3,85) com outros trabalhos realizadosem lagos de várzea da AmazôniaCentral: Comandá (R=59, H'=3,9),Praia (R=76, H'=4,8) e Tracajá (R=52,H'=4,3) (Yamamoto, 2004), a região deágua aberta do Tupé pode serconsiderada com alta diversidade. Asdiferenças estão na biomassa dospeixes, pois em um lago de várzeapode ocorrer uma biomassa cincovezes maior do que em um lago do rioNegro (Saint-Paul ., 2000).
Na comunidade de peixes em umlago nem sempre às espécies maisabundantes correspondem ascomercializadas pela população(Pereira, 1999; Freitas, 1999). Dentre
H. immaculatusM. intermedia, A.
falcirostris, L. batesii H. edentatusM. intermedia
A. falcirostris
H. immaculatus, H.edentatus L. batesii
et al
et al
as 14 espécies de maior importância em termos deabundância capturadas no Tupé (
sp.,sp.1 e
sp.2, sp., C.e sp.) nenhuma
tem alto valor comercial. Somente dois grupos deespécies ( spp., spp.) são, empequena proporção, comercializados nos mercados deManaus (Batista, 1998). Por outro lado, peixes com altasazonalidade e baixa abundância como os
spp. (jaraquis), spp.(tucunarés), (acará-açu) espp.(acará-prata), spp. (pacu), spp.(matrinchã), (traíra) (aracú)e spp. (aracu) estão incluídos na dieta dascomunidades ribeirinhas localizadas na RDS TemaRebelo, 2005, neste volume).
Os resultados acima descritos têm importantesimplicações para o manejo da ictiofauna no lago Tupé,que ainda apresenta grandes áreas com floresta alagada.A variação na estrutura da ictiofauna associada com ociclo hidrológico está fortemente relacionada com adisponibilidade de habitats, que em lagos de água pretaestão principalmente restritos ao igapó e a água aberta.Saint-Paul . (2000) estudando a ictiofauna dasregiões de água aberta e igapó no lago Prato,Arquipélagode Anavilhanas, destaca que mais da metade (74%) dospeixes foi coletado no igapó, sendo superior em númerode espécies a captura na floresta de várzea do lagoInácio, rio Solimões. Portanto, os igapós podem serconsiderados habitats chave, de importância vital para odesenvolvimento de muitas populações de peixes(Goulding ., 1988).
Os resultados suportaram a hipótese de que existemdiferenças sazonais na diversidade e riqueza daictiofauna do lago Tupé por causa da presença de área deterra-firme, relativamente intacta, e a grande áreaalagável, que permitem a existência de habitats para acolonização de muitas espécies de peixes. Esses fatoressão de fundamental importância por possibilitar amanutenção da diversidade e abundância de espécies,haja vista que a proximidade do lago Tupé à cidade deManaus, tem favorecido, nas ultimas décadas, umaintensa pressão antrópica.
A. falcirostris, M.intermedia, H. immaculatus, Hemiodus H. gracilis,B. lucius, C. abramoides, C.vittata, LonchogenysLonchogenys Tetragonopterus heckelii, H.edentatus, L. batesi, A. thoracatus Tatia
Hemiodus Hypophthalmus
Semaprochilodus CichlaA. ocellatus Chaetobranchus
Mylossoma BryconH. malabaricus S. fasciatus
Leporinus(
et al
et al
&
194
Soares Yamamoto&

Agradecimentos
Bibliografia citada
Agradecemos o apoio financeiro recebido através dosprojetos INPA/ Max-Planck e PPI (N I-3540 ) e ao Dr.Jansen Zuanon e Hélio D. B. dos Anjos pela ajuda naidentificação dos peixes.
Aprile F. M.; Darwich A.J. 2005.: Santos-Silva, E. N.; Aprile,
F. M.; Scudeller, V. V..; Melo, S. (Orgs.) Ed. INPA, AM.
Araújo-Lima, C.A.R.M.; Portugal, L.P.S.; Ferreira, E.G.1986. Fish-macrophyte relationship the AnavilhanasArchipelago, a black water system in the CentralAmazon. ., 29: 1-11.
Batista, V.S. 1998.. Tese de
Doutorado, Instituto Nacional de Pesquisas daAmazônia/Fundação Universidade do Amazonas,Manaus,Amazonas. 291pp.
Bohlke, J.E.; Weitzman, S.H.; Menezes, N.A. 1978.Estado atual da sistemática dos peixes de água docedaAmérica do Sul. , 8 (4): 657-677.
Borges, G.A. 1986.
. Dissertação deMestrado, Instituto Nacional de Pesquisas daAmazônia/Fundação Universidade do Amazonas,Manaus,Amazonas. 131pp.
Claro Jr. L. H. 2003.
. Dissertação deMestrado, Instituto Nacional de Pesquisas daAmazônia/Fundação Universidade do Amazonas,Manaus,Amazonas. 61pp.
Carvalho, M. L.; Goulding, M. 1985. On the feedingecology of the catfish inblackwater rio Negro of the Amazon basin.
., 3(1):33-41.Darwich, A. J.; Aprile, F. M., Robertson, B. A.,2005.
Variáveis Limnológicas: contribuição ao estudo
o
Modelos geomorfológi-cos para o lago Tupé. In
J. Fish. BiolDistribuição, dinâmica da frota e dos
recursos pesqueiros da Amazônia Central
ActaAmazonicaEcologia de três espécies do gênero
Brycon no Rio Negro (Amazonas), com ênfase nacaracterização e alimentação
A influência da floresta alagada naestrutura trófica de comunidades de peixes em lagosde várzea da Amazônia Central
Hypophthalmus fimbriatusRevta
Bras. de Zool
Biotupé: meio físico, diversidade biológica esociocultural do baixo Rio Negro,Amazônia Central.
espaço-temporal de águas pretasamazônicas. :
Fernandes, C.C.; Merona, B. 1988.Lateral migration of fishes on afloodplain system in the CentralAmazon (Careiro, Island, lake ofRei) AM, BR.
, Tomo XLVIII(suplemento): 409-433.
Ferreira, E.J.G. 1981
. Dissertação deMestrado, Instituto Nacional dePesquisas da Amazônia/FundaçãoUniversidade do Amazonas,Manaus,Amazonas. 254pp.
Ferreira, E.J.G. 1984. A ictiofauna darepresa hidrelétrica de Curuá-Una,Santarém, Pará. Lista e distribuiçãodas espécies. , 8(3):351-363.
Ferreira, E.J.G. 1993. Composição,distribuição e aspectos ecológicosda ictiofauna de um trecho do rioTrombetas, na área de influência dafutura UHE Cachoeira Porteira,Estado do Pará, Brasil.
, 23 (suplemento): 1-89.Ferreira, E.J.G.; Santos, G.M. dos;
Jégu, M. 1988. Aspectos ecológicosda ictiofauna do rio Mucajaí, naárea da ilha Paredão, Roraima.
, 10(3): 339-352.Ferreira, E.J.G., Zuanon, J.A.S.;
Santos, G.M. 1998.
. IBAMA.Brasília. 211pp.
Freitas, C. E. C.; Batista, V. S. 1999. Apesca e as populações ribeirinhas
In
Sociedad de CienciasNaturales La Salle
Alimentação deadultos de doze espécies decichlídeos (Perciformes, Cichlidae)do rio Negro, Brasil
Amazoniana
ActaAmazonica
Amazoniana
Peixescomerciais do médio Amazonas:Região de Santarém, Pará
Santos-Silva, E. N.;Aprile, F. M.; Scudeller, V. V..;Melo, S. (Orgs.) Ed. INPA, AM.Biotupé: meio físico, diversidadebiológica e sociocultural do baixoRio Negro,Amazônia Central.
195
Diversidade e composição da ictiofauna do Lago Tupé

J.Linn. , 45: 103-144.Magurran, A. E. 1988.
. Chapman e Hall, London. 179pp.Malabarba, L.R.; Reis, R.E.; Vari, R.R.; Lucena, Z.M.S.;
Lucena, C.A. 1998.. EDIPUCRS, PortoAlegre 603pp.
Marlier, G. 1968. Etude sur lês lacs de l'amazonie Central.II. Le placton; Lês poissons du lac Redondo et leurregime alimentaire; le chaines tropiques du lacredondo; lês poissons du rio Preto da Eva.
, 11:1-57.Mérona, B.; Bittencourt, M.M. 1993. Lês peuplements de
poissons du 'lago do Rei', um lac d'inondationd'Amazonie Central: description générale.
, 12: 415-441.Merona, B.; Santos, G.M.; Almeida, R.G. 2001. Short
term effects of Tucurui Dam (Amazônia, Brazil) on thetrophic organization of fish communities.
Oliveira Jr, A. B. 1998. Táticas alimentares ereprodutivas do Tucunaré comum (AGASSIZ, 1813) no reservatório da UHE de BalbinaAM, Brasil. Dissertação de Mestrado, InstitutoNacional de Pesquisas da Amazônia/FundaçãoUniversidade doAmazonas, Manaus,Amazonas. 74pp.
Pereira, H.S. 1999.Ph.D. Thesis. The Pennsylvania
State University. 119pp.Pouilly, M; Lino, F.; Bretenoux, J.G.; Rosales, C. 2003.
Dietary-morphological relationships in a fishassemblage of the Bolivian Amazonian floodplain.
. 62 : 1137-1158.Ribeiro, M.C.L.B. 1983. As migrações dos jaraquis
(Pisces, Prochilodontidae), no rio Negro, Amazonas,Brasil. Dissertação de Mestrado, Instituto Nacional dePesquisas da Amazônia/Fundação Universidade doAmazonas, Manaus,Amazonas. 192pp.
Ribeiro, M.C.L. B.; Petrere, Jr. M. 1990. Fisheries ecologyand management of the jaraqui (
) in Central Amazon., 5: 195-215.
Saint-Paul, U.; Zuanon, J.; Villacorta-Correa, M.A.;Garcia, M. Fabré, N.N.; Berger, U.; Junk, W.J. 2000.Fish communities in Central Amazonia white and theblack waters floodplains. ,57:235-250.
Soc. (Zool)Ecological diversity and its
measurement
Phylogeny and classification ofneotropical fishes .
CadernosdaAmazônia
Amazoniana
Environ.Biol. Fishes, 60: 375-392.
Cichla monoculus
Common-property regimes inAmazonian fisheries.
J.Fish. Biol
Semaprochilodustaeniurus, S. Insignis RegulatedRivers: Research Management
Environ. Biol. Fishes
&
da Amazônia Central., Rio Claro, SP,
Brasil 01: 32-36.Garcia, M. 1995.
.Dissertação de Mestrado, InstitutoNac iona l de Pesqu i sa s daAmazônia/Fundação Universidadedo Amazonas, Manaus, Amazonas.95pp.
Gery, J. 1977. .TFH Publications. Neptune City,672p .
Goulding, M. 1980.
. University ofCalifornia Press, Berkeley. 280pp.
Goulding, M.; Carvalho, M.L.; Ferreira,E.G. 1988. Rio Negro: rich life inpoor water:
SPBAcademic Publishing The Hague.200pp.
Junk, W. J., Soares, M. G. M.Carvalho, F. M. 1983. Distribution offish species in a lake Amazon riverfloodplain near Manaus lagoCamaleão with special reference toextreme oxigen conditions.
, 7(4): 397-431.Krebs, C.J. 1989.
. New York:654pp.
Kullander, S.; Reis, R.; Ferraris, C.2003. Check list of the freshwaterfishes of South and CentralAmerica. EDIPUCRS, Porto Alegre,944p .
Lowe-McConnell, R.H. 1964. The fishesof the Rupunini savanna district ofBritish Guiana, Pt 1. Groupings offish species and effects of theseasonal cycles on the fishes.
BrazilianJournal of Ecology
,Aspectos ecológicos
dos peixes das águas abertas de uml a g o n o A r q u i p é l a g o d a sAnavilhanas, Rio Negro, AM
Characoids of the world
The fishes and theforest. Explorations in Amazoniannatural history
Amazonian diversityand food chain ecology as seenthrough fish communities.
,
AmazonianaEcological
Methodology .
p
Krebs, C.J
p
&
196
Soares Yamamoto&

Biotupé: meio físico, diversidadebiológica e sociocultural do baixoRio Negro,Amazônia Central.
Vale, J.D. 2003.
. Dissertação deMestrado, Instituto Nacional dePesquisas da Amazônia/FundaçãoUniversidade do Amazonas,Manaus,Amazonas. 66pp.
Yamamoto, K.C. 2004.
.Dissertação de Mestrado, InstitutoNac iona l de Pesqu i sa s daAmazônia/Fundação Universidadedo Amazonas, Manaus, Amazonas.71pp.
Composição,diversidade e abundância daictiofauna na área do Catalão,Amazônia Central
A estrutura decomunidades de peixes em lagosmanejados da Amazônia Central
Santos, G.M.; Jegu, M.; Merona, B. 1984.. Projeto
Tucuruí. ELETRONORTE/CNPq/INPA, Manaus 83pp.Santos, G.M. 1983. Aspectos ecológicos da pesca
experimental em sistemas “lacustres” e fluvial nobaixo rio Negro. Anais do
, 367-391.Soares, M.G.M., Almeida, R.G.; Junk, W.J. 1986. The
tropic status of the fish fauna in lago Camaleão emacrophyte dominated floodplain lake in the middleAmazon. , 9(4):511-526.
Souza-Pereira, A. 2000.
.Monografia, Fundação Universidade do Amazonas,Manaus,Amazonas. 31pp.
Terra, A. K. Rebelo,G. H., 2005. O uso da fauna pelosmoradores da comunidade Saõ João e colôniaCentral.
Catálogo depeixes comerciais do baixo rio Tocantins
.
III Congresso Brasileiro deEngenharia de Pesca CONBEP. ManausAM
AmazonianaVariáveis limnológicas e sua
relação com as espécies de peixes onívoras eherbívoras do lago Camaleão, Amazonas, Brasil
&
In: Santos-Silva, E. N.; Aprile, F. M.;Scudeller, V. V..; Melo, S. (Orgs.) Ed. INPA, AM.
197
Diversidade e composição da ictiofauna do Lago Tupé


Diversidade BiológicaCapítulo 14
Bancos de dados sobre biodiversidadena Amazônia: a experiência do ProjetoBiotupé
RESUMO - A criação de bancos de dados por projetos que envolvem a coleta e o estudo deespécimes biológicos, as espécies a que pertencem e os locais em que ocorrem, é um pré-requisito indispensável para a conservação e uso sustentável dos recursos naturais.Contudo, o grande volume de dados oriundos de levantamentos ecológicos de campodemanda a aplicação de novas técnicas e instrumentos para seu armazenamento,recuperação, análise e disseminação. O Projeto Banco de Dados do Biotupé estádirecionado ao manejo e análise dos dados sobre a Reserva de DesenvolvimentoSustentável do Lago Tupé e sua integração em um sistema de informação distribuído sobrebiodiversidade, que possa fornecer apoio para as atividades de pesquisa e extensão emandamento na área. Os recursos computacionais disponíveis ao projeto são utilizados nogerenciamento dos dados sobre espécies e indivíduos coletados em campo, através dautilização de sistemas gerenciadores de banco de dados específicos para levantamentosecológicos de campo e inventários biológicos (ECOLOG e ALINE); na produção de guias decampo e manuais de identificação, por meio de sistemas de identificação auxiliada porcomputador baseados no formato DELTA; e na análise multivariada de dados ecológicosatravés de vários métodos de ecologia numérica. A estratégia do sistema de informação
Mauro José CAVALCANTIe-mail: [email protected]
Departamento de Vertebrados, Museu Nacional/Universidade Federal do Rio de JaneiroQuinta da Boa Vista, São Cristóvão, 20940-040 Rio de Janeiro, RJ Brasil
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

adotada pelo Projeto Biotupé é de baixo risco, usando tecnologia e programas jádisponíveis e testados em outros projetos, e espera-se que possa oferecer um modelooperacional útil para as organizações e indivíduos envolvidos no desenvolvimentosustentável e conservação da biodiversidade naAmazônia.
bancos de dados, bioinformática, levantamentos ecológicos,inventários de biodiversidade, DELTA.PALAVRAS-CHAVE:
Introdução
O Projeto Biotupé é uma proposta de estudomultidisciplinar de longo prazo em execução na Reservade Desenvolvimento Sustentável do Tupé (RDS do Tupé),envolvendo pesquisadores de várias instituições. Oprincipal objetivo deste projeto é inventariar, identificare quantificar a biodiversidade dessa área. Para isso estãosendo desenvolvidos estudos sobre vegetação,hidrologia, hidroquímica, zooplâncton, fitoplâncton,esponjas, peixes, coliformes, insetos aquáticos,crustáceos intersticiais, uso de recursos e estudos sócio-ambientais. Assim, o Projeto Biotupé compreende umacoleção de projetos interrelacionados, cada um delescom seus próprios objetivos e prioridades locais.Todavia, uma necessidade comum a todos estes projetosé o armazenamento, recuperação e disseminação dasinformações sobre a diversidade biológica na áreapesquisada.
A criação de bancos de dados por projetos queenvolvem a coleta e o estudo de espécimes biológicos, asespécies a que pertencem e os locais em que ocorrem,constitui-se num pré-requisito indispensável àconservação da biodiversidade e uso sustentável dosrecursos naturais. Contudo, o manuseio do volumeconsiderável de dados oriundos de levantamentosecológicos de campo e inventários biológicos constitui-seem um grande problema (Yoon, 1993; Janzen, 1994;Lawton ., 1998; Sharkey, 2001), demandando oemprego de novas técnicas e instrumentos para autilização eficaz das informações disponíveis nosprocessos de tomada de decisões referentes àadministração do meio ambiente (Cavalcanti, 1998a).
Neste contexto, a implementação de sistemas debancos de dados apresenta-se como uma forma eficientede organização e armazenamento das informações
et al
obtidas pelos diversos projetosenvolvidos no estudo da RDS do Tupé,possibilitando aos mesmos comparti-lhar e disseminar eficazmente os dadoscoletados e armazenados de formapadronizada por cada um deles,reduzindo consideravelmente o tempode processamento da informação eacelerando a produção de resultadosúteis.
O Projeto Banco de Dados doBiotupé é direcionado ao manejo eanálise dos dados sobre a RDS do LagoTupé e sua integração em um sistemade informação sobre a biodiversidadedesta área, que possa fornecer apoiopara as atividades de pesquisa,conservação e uso sustentável dosrecursos naturais.
O sistema de informação sobrebiodiversidade do Projeto Biotupéfuncionará como um grupo de bancosde dados individuais, ligados atravésde padrões comuns de dados,protocolos de intercâmbio de dados eprogramas aplicativos. O sistema assimestabelecido canalizará dados críticos,acumulados no âmbito de cada projetoindividual ou armazenados nas fontesexistentes, para os projetos de uso derecursos e desenvolvimento sustentá-vel e disseminará a informação dos
O PROJETO BANCO DE DADOS DOBIOTUPÉ
200
Cavalcanti

projetos para um público mais amplo.A flexibilidade oferecida pelos
sistemas gerenciadores de bancos dedados utilizados, dará a cada projetototal autonomia para controlar os tiposde dados que serão compartilhados(espécies, indivíduos, descritoresmorfológicos, variáveis ambientais,etc.). Esta flexibilidade é crítica para oProjeto Biotupé, uma vez quediferentes bancos de dados devem serimplementados por diferentesprojetos, de acordo com as suasnecessidades e prioridades específi-cas. Contudo, para que o sistema deinformação do Projeto Biotupé alcanceseu maior objetivo, a integração,será necessário estabelecer um altograu de padronização na definição decategorias de dados como habitats(comunidades vegetais, por exemplo)e certos descritores (outros,entretanto, poderão ser definidos porcada projeto para atender a suaspróprias necessidades e prioridadesespecíficas) visando assegurar acompatibilidade entre os diferentesbancos de dados (Dalcin, 1998). Oestabelecimento de padrões para osdados e a adoção de estruturas dearmazenamento comuns gerenciadaspor sistemas utilizados por todos osprojetos integrantes do ProjetoBiotupé possibilitará uma troca efetivade informação entre os vários gruposde pesquisa envolvidos. Estaestratégia já foi aplicada com sucessoem outros sistemas de informaçãosobre biodiversidade, envolvendop r o j e t o s d e s e n v o l v i d o s e mcolaboração por várias instituições,como International Legume Databaseand Information Service (Bisby, 1989,1993, 2000; Zarucchi ., 1993),World Biodiversity Database (Schalk,1992) e Plantas do Nordeste (Allkin,
et al
1997, 1998).O núcleo de informação de cada projeto (nomes
científicos, distribuição geográfica, tipos de utilizaçãode plantas e animais, nomes vulgares, etc.) será idênticoe seguirá uma padronização (por exemplo, por tipos deambiente, nomes de localidades e códigos de unidadesde amostragem).Adisponibilidade de uma base de dadostaxonômica única para cada grupo botânico ouzoológico, fornecendo às bases de dados orientadas aosindivíduos (coleções biológicas) um nome único dotáxon, partilhado por todos os projetos, permitirá aintegração consistente e efetiva dos dados, assegurandosua confiabilidade e facilitando o seu intercâmbio(Dalcin, 1998). Outros padrões de dados e terminologias,entretanto, poderão ser estabelecidos individualmentepor cada projeto, segundo suas próprias necessidades.
O Projeto Banco de Dados do Biotupé irá assegurarque os dados, os programas aplicativos e o equipamentocomputacional disponíveis sejam gerenciados de umamaneira efetiva e apropriada através do:
1) Estabelecimento e apoio aos bancos de dadosindependentes desenvolvidos pelos projetos individuaisno âmbito do Projeto Biotupé;
2) Interligação destes bancos de dados a fim de que osprojetos possam compartilhar informações;
3) Desenvolvimento de padrões de dados, formatos emecanismos para assegurar que os dados possam serintercambiados.
Além disso, o Projeto Banco de Dados do Biotupé tempor objetivo criar uma maior auto-suficiência dentro daregião em todos os níveis. Isto será efetuado com o:
1) Fornecimento de treinamento especializado emgerenciamento de dados biológicos, inclusiveoferecendo cursos técnicos de treinamento no uso desistemas de informação e na análise de dados sobrebiodiversidade;
2) Formação de pessoal técnico capacitado na área debioinformática e manejo de bancos de dados biológicos.
O equipamento computacional utilizado no âmbito doProjeto Biotupé foi escolhido com base nos custos,confiabilidade e fácil acesso. São utilizados basicamente
RECURSOS COMPUTACIONAIS
Hardware
201
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé

microcomputadores da linha IBM-PC ou compatíveis. Ointercâmbio eletrônico de dados será efetuado atravésde redes locais (intranet do INPA) e de longa distância(internet).
Sistemas de confiança são um fator crítico para osucesso dos projetos de bancos de dados sobrebiodiversidade (Allkin Winfield, 1993). Dadosbiológicos complexos e variáveis requerem estruturas dedados adequadas para representá-los e programassofisticados para manejá-los, mas que também sejam defácil utilização (Allkin, 1988; Allkin Bisby, 1988; Allkin
., 1992b; White ., 1993). Os custos para odesenvolvimento de tais sistemas são elevados e, emgeral, grosseiramente subestimados. Projetosindividuais de levantamentos ecológicos de campo einventários biológicos raramente dispõem do tempo oudos recursos necessários para criar os próprios sistemasde gerenciamento de bancos de dados de quenecessitam.
Com freqüência, os sistemas de bancos de dados demuitos projetos não funcionam adequadamente quandosão necessários ou são de uso tão complexo que apenasespecialistas ou membros da equipe especialmentetreinados podem acessar o sistema. Os participantes detais projetos dispendem uma considerável quantidadede tempo entendendo sistemas comerciais (comoplanilhas eletrônicas e gerenciadores de bancos dedados), tentando adaptá-los às suas necessidadesespecíficas e muitas vezes duplicando esforços dentro dacomunidade científica. Estes problemas tornam os dadosefetivamente inacessíveis e incertos a longo prazo eimpedem que seus usuários se beneficiem integralmenteda moderna tecnologia disponível de banco de dados(Allkin Winfield, 1993).
A estratégia adotada pelo Projeto Banco de Dados doBiotupé é de baixo risco, usando tecnologia simples eprogramas aplicativos já desenvolvidos e testados. Osistema de informações do Projeto Biotupé utilizarásistemas já disponíveis e facilmente acessíveis,preferencialmente programas de aplicação científicadifundidos, cujo funcionamento é conhecido e que estãoem uso bem sucedido em muitos projetos similares. O
Software
&
&
&
et al et al
sistema irá se beneficiar, assim, dosinvestimentos prévios efetuados nodesenvolvimento desses programas. OProjeto Banco de Dados do Biotupéoferecerá a cada projeto individual umconjunto de ferramentas para om a n e j o d e d a d o s s o b r ebiodiversidade, cada qual projetadapara um propósito diferente (Die-derich Milton, 1993; White .,1993), proporcionando uma grandeflexibilidade na gestão, compartilha-mento e publicação de suas informa-ções. Três sistemas básicos serãoutilizados por todos os projetosintegrantes do Projeto Biotupé: ALINEpara o manejo de dados sobreespécies, ECOLOG para o manejo dedados sobre espécimes e DELTA para omanejo de dados taxonômicos (Fig. 1).
Os recursos computacionaisdisponíveis ao projeto serão utilizadosbasicamente:
1) No gerenciamento dos dadossobre espécies e indivíduos coletadosem campo, através da utilização desistemas gerenciadores de banco dedados com capacidade relacional,específicos para o manejo de dadosobtidos em levantamentos ecológicosde campo e inven tá r i o s debiodiversidade (ECOLOG eALINE);
2) Na produção de guias eletrônicose manuais de identificação de campo,para diferentes usuários, por meio desistemas de identificação auxiliada porcomputador baseados no formatoDELTA(Dallwitz, 1980, 1993).
3) na análise dos dados através demétodos estatísticos e técnicasmultivariadas (Gauch, 1982; DigbyKempton, 1987; Ludwig Reynolds,1988; Legendre Legendre, 1998);Cada uma destas aplicações deferramentas computacionais noProjeto Biotupé será descrita mais
&
&&
&
et al
202
Cavalcanti

detalhadamente a seguir.
O armazenamento, recuperação eanálises básicas dos dados sobreespécies e indivíduos coletados emcampo serão efetuados através dosistema gerenciador de bancos dedados ECOLOG desenvolvido porCavalcanti (1991, 1993), que seráusado por todos os projetos queenvolvem a coleta e o estudo deespécimes biológicos.
O sistema ECOLOG foi especificamente projetado para o manejo dedados obtidos em levantamentosecológicos de campo e inventáriosbiológicos em microcomputadores dalinha PC e conta com mais de 10 anosde investimento em seu desenvolvimento. Sua flexibilidade, que pode sertestemunhada pela natureza diversados projetos que o utilizam (Sá .,1991; Dalcin ., 1997; KurtzAraujo, 2000) e maturidade inspiramconfiança em seus usuários. Estesistema apresenta como principaiscaracterísticas a utilização emequipamentos de baixo custo e fácilacesso (microcomputadores da linhaIBM-PC); a rapidez e facilidade deoperação, dispensando conhecimentosespecializados em computação porparte dos usuários; a validaçãocompleta dos dados de entrada; aflexibilidade na recuperação deregistros para consulta e produção derelatórios (a partir de qualqueratributo ou combinação de atributos);a possibilidade de produzir análisesquantitativas de dados ecológicos(cálculo de índices de diversidade,freqüência e dominância, curvas derarefação, etc.); a emissão delistagens de espécies e de etiquetas de
O SISTEMAECOLOG
-
-
et alet al &
Figura 1. Sistemas do Projeto Biotupé
coleta de campo; a exportação de dados em váriosformatos padronizados para utilização em outrossistemas; e a integração com dispositivos eletrônicos deaquisição de dados, como computadores de mão esistemas de geoposicionamento (Morain, 1993; Herring,1996).
O sistema ECOLOG utiliza uma estrutura relacionalsimples e eficiente, baseada em três tabelas de dadosprincipais (indivíduos, espécies e locais) e duas tabelasauxiliares (descritores morfológicos e variáveisambientais), que se relacionam por camposcompartilhados (Fig. 2). Esta estrutura pode serconsiderada uma versão simplificada do modelo geral deinformação para coleções biológicas proposto porBerendsohn . (1999). As tabelas de dados do sistemaarmazenam dados sobre as características gerais de cadaespécie (como taxonomia e nomenclatura, os dados decoleta de cada espécime (coletor, data, procedência,biometria, morfologia, notas) e a localização espacial(coordenadas geográficas, altitude/profundidade) decada unidade amostral. A partir de cada uma destastabelas de dados, são criados arquivos de índices quepermitem a busca e recuperação de informações atravésde múltiplas chaves. A utilização de uma estruturarelacional permite reduzir a duplicação e a redundâncianos dados e favorece a eficiência no seu armazenamen-to. Assim, por exemplo, uma vez que cada espéciecompreende um registro único na tabela de espécies, énecessário armazenar apenas o número do táxon para osregistros daquela espécie, ao invés de repetir o nome
et al
203
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé
Gerenciamentodos dados sobre
espécimes
Gerenciamentodos dados sobre
espéciesDivulgação
Etiquetasde coleta
Relatórios deManutenção
Lista de espécies,monografias
Chaves deidentificação
gias de campo
ECOLOG ALINE DELTA
ATF DELTA

completo da espécie para cada coleta no banco dedados. Da mesma forma, qualquer alteração em umregistro nas tabelas de espécies ou locais éautomaticamente refletida em todos os registros deindivíduos relacionados àquela espécie ou àquele local.
Como as tabelas de dados do sistema possuem camposde tamanho fixo, procurou-se otimizar ao máximo otamanho de cada campo para assegurar oarmazenamento adequado dos dados, sem um consumoexcessivo de espaço em disco. Esta restrição implica noaumento do espaço de armazenamento necessário aobanco de dados, bem como da complexidade daestrutura lógica do mesmo, porém não impõe limites aotamanho do mesmo.
Todos os projetos no âmbito do Projeto Biotupé irãoadministrar seus bancos de dados sobre espéciesutilizando o sistema ALINE desenvolvido por Cavalcanti
O SISTEMA ALINE
(1998b). O sistema ALINE é umaferramenta genérica, fundamentadaem um modelo de dados relacionalprojetado pelo International LegumeDatabase and Information Service(ILDIS), que tem como objetivo criar emanter um banco de dados global cominformações taxonômicas, ecológicase econômicas sobre as leguminosas,envolvendo grupos de pesquisa emmais de 20 países (Bisby, 1989, 1993,2000; Zarucchi ., 1993). Estaestrutura assegura a qualidade eintegridade dos dados armazenados,permitindo ainda aos usuários definirseus próprios descritores morfológicose configurar seus bancos de dados deacordo com suas necessidadesespecíficas (Fig. 3).
ALINE é um sistema orientado ao
et al
Figura 2. Tabelas relacionadas do Sistema ECOLOG
204
Cavalcanti
Número do IndivíduoNúmero da EspécieLocalColetorData da ColetaHora da ColetaObservações
Número da EspécieGrupoFamíliaGêneroEspécieAutorSubespécie
LocalNome do LocalAltitude/ProfundidadeLatitudeLongitude
Tabela de Indivíduos
Tabela de Locais
Tabela de Espécies
Número do IndivíduoDescritorDescrição
Tabela de Descritores
LocalDataHoraVariávelValor
Tabela de Variáveis Ambientais

táxon com capacidade relacional,disponível para microcomputadores dalinha IBM-PC (Cavalcanti, 1998b). OALINE será usado por todos os projetospara a criação e manutenção de bancosde dados sobre a flora e a fauna da áreade estudo, incluindo informaçõessobre taxonomia e nomenclatura, usoseconômicos, nomes vulgares,ecologia, morfologia, distribuiçãogeográfica e bibliografia relevantepara cada espécie. O sistemaapresenta várias características quegarantem a consistência das informa-ções armazenadas no banco de dados,tais como: completa verificação dosdados assim que introduzidos,ferramentas para a manutenção dosbancos de dados e referências cruza-das, com todos os dados relacionadosàs suas fontes bibliográficas. Departicular importância é a capacidadede exportação de quaisquer subcon-juntos de dados, selecionados pelousuário, em vários formatos padroni-zados, como DELTA (Dallwitz, 1980),NEXUS (Maddison ., 1997) e XDF(White Allkin, 1992). Isto permite aosprojetos individuais utilizarem outrosprogramas quando necessário (porexemplo, para identificação interativae construção de chaves taxonômicasatravés do sistema DELTA) ecompartilhar subconjuntos selecio-nados de dados com outros projetos. Acapacidade de exportar dados emvários formatos oferece segurança eassegura a longevidade das informa-ções armazenadas através do ALINE,permitindo ao projetos integrantes doProjeto Biotupé o intercâmbio deinformações entre si e com outrasorganizações. Entre suas funções,i n c l u e m - s e t a m b é m a q u e l a snecessárias à disseminação dainformação, pois os bancos de dados
et al&
Figura 3. Tabelas relacionadas do Sistema ALINE
criados através do ALINE podem ser publicados comorelatórios impressos, guias eletrônicos em CD's ou comopáginas na Internet, utilizando o formato HTML.
O sistema DELTA (DEscription Language forTaxonomy) consiste de um formato flexível para acodificação da informação taxonômica descritiva e umconjunto de programas específicos para oprocessamento de descrições taxonômicas (Dallwitz,1980, 1993; Askevold O'Brien, 1994). Este sistema foidesenvolvido na Austrália em meados dos anos 70,inicialmente para computadores de grande porte edepois adaptado para microcomputadores da linha IBM-PC, oferecendo uma série de facilidades para o manejo ea organização da informação taxonômica. Especialmen-te úteis são a possibilidade de construção de chaves deidentificação de diversos tipos, a produção de descriçõesem linguagem natural e a preparação de matrizes dedados para análises fenéticas e cladísticas, além daidentificação interativa e recuperação de informações.
O SISTEMADELTA
&
205
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé
espécie
nomes vulgares
usos
dist. geográfica
habitats
sinônimos
notas
descrições
família
dicionáriogeográfico
Referênciasbibliográficas
dicionário dedescritores
imagens

O formato DELTA aceita todos os tipos de caracteres,tanto qualitativos (duplo estado e multiestado,ordenados ou não ordenados) quanto quantitativos(contínuos e descontínuos) e inclui instruções (diretivas)para controlar o processamento dos dados pelos diversosprogramas componentes do sistema. Uma vezcodificados no formato DELTA, os mesmos dados podemser utilizados para produzir descrições e chaves deidentificação com própositos gerais ou específicos,omitindo parte dos caracteres, ou dando ênfase a algunsdeles. Isto torna possível a construção, a partir damesma base de dados, de diferentes tipos de chaves,adequadas ao uso em laboratório ou em campo,específicas para a fauna ou a flora de certas localidadesou determinados estágios do ciclo de vida dos organismos(Stevenson ., 2003).
Em 1988, o formato DELTA foi adotado peloInternational Working Group on Taxonomic Databases forPlant Sciences (TDWG) como um padrão internacionalpara a representação e intercâmbio de dadostaxonômicos descritivos. Desde então, o sistema DELTAjá mostrou-se uma valiosa ferramenta para inventáriossistemáticos de biodiversidade (Allkin ., 1992a;Sharkey, 2001) e levantamentos ecológicos de campopara estudos de impacto ambiental (Ellis, 1988).
O sistema DELTA original é composto de quatroprogramas principais (Dallwitz ., 1993; AskevoldO'Brien, 1994): CONFOR (para conversão de formatos,produção de descrições em linguagem natural e análiseestatística de dados), DELFOR (para ordenação, inclusãoou exclusão de caracteres e seus estados), KEY (paraconstrução de chaves de identificação) e INTKEY (paraidentificação interativa e recuperação de informaçõesdo banco de dados taxonômico). Além destes, existemprogramas complementares, que permitem prepararconstruir matrizes de distância fenética entre táxons(DIST) e realizar análises de agrupamento e similaridade(PCLASS e NSIM). Ambientes interativos para a utilizaçãointegrada dos vários programas que compõem o sistemaDELTA estão disponíveis (Cavalcanti, 1996; Dallwitz
, 1999).O programa INTKEY, para identificação interativa e
recuperação de informações, permite ao usuário acessaro conteúdo de um banco de dados taxonômico a partir dequalquer caráter e na ordem ou combinação em quedesejar, iniciando de novo tantas vezes quantas
et al
et al
et al
etal.
&
necessárias até a determinação de umtáxon ser confirmada (Dallwitz, 1993;Dallwitz ., 1995). O INTKEYpermite também o armazenamento deilustrações digitalizadas associadas àsdescrições dos caracteres para guiar ousuário ao longo da chave. Váriosbancos de dados taxonômicos têm sidocriados com o sistema DELTA,compreendendo desde gêneros degramíneas e leguminosas até espéciesde corais, formigas e besouros(Askevold O'Brien, 1994). Muitosdestes bancos de dados incluemimagens digitalizadas dos táxons ecaracteres, e vários deles estãodisponíveis na Internet (www.delta-inkey.com). No âmbito do ProjetoBiotupé, o sistema DELTA será usadopara produzir guias de identificação(em forma eletrônica ou como manuaisimpressos) a partir de descrições dasespécies. Estas descrições estarãoarmazenadas em bancos de dados dosistema ALINE e, quando necessário,subconjuntos de dados (por exemplo,as plantas de uso medicinal da RDS doTupé) serão exportados doALINE para oformato DELTA, já prontos para aprodução dos guias. Desta forma, oProjeto Banco de Dados do Biotupéespera contribuir para a implementação de uma metodologia prática eeficaz para a elaboração de guiaseletrônicos de biodiversidade (Stevenson ., 2003), para diferentesusuários, que permitam a pessoastecnicamente capacitadas, mas nãoespecializadas em botânica ouzoologia, identificarem as espécies deplantas e animais da região.
O uso de técnicas de análisemultivariada em ecologia de comuni-
et al
-
-et al
&
ANÁLISE DE DADOS
206
Cavalcanti

dades tem se intensificado bastantenos últimos anos, devido, sobretudo, àgrande disponibilidade dos microcom-putadores e pacotes estatísticos. Estastécnicas permitem considerar simulta-neamente os padrões de variação ecovariação de um grande número deatributos do sistema sob estudo,mostrando-se muito úteis na classifica-ção e ordenação de comunidadesecológicas (Green, 1980; Gauch, 1982;Digby Kempton, 1987; LudwigReynolds, 1988; Legendre Legendre,1998) e possibilitando que parâmetrosimportantes subjacentes às relaçõesentre amostras ou espécies possam sermais facilmente detectados e inter-pretados. Em vista de sua utilidade nadescrição e interpretação de dadosecológicos complexos, bem como nageração de hipóteses que podem sertestadas em campo (Green, 1980) einclusive no manejo e conservação decomunidades vegetais (Cooper, 1984;Margules, 1984; DeVelice ., 1988),é de se esperar que a aplicação de taismétodos produza resultados valiososnos estudos ecológicos realizados noâmbito do Projeto Biotupé.
As técnicas de classificaçãopermitem agrupar as amostras("análise normal ou do tipo Q") ou asespécies ("análise inversa ou do tipoR") de acordo com seus atributoscomuns (Sneath Sokal, 1973; Ludwig
Reynolds, 1988; LegendreLegendre, 1998). As técnicas deordenação, por sua vez, podem serusadas tanto para a classificação peloagrupamento de amostras ou espécies,quanto para o arranjo das amostras ouespécies em relação a gradientesambientais, com o objetivo dedetectar a variação conjunta nacomposição da comunidade em funçãode fatores do ambiente (Gauch, 1982).
& &&
&& &
et al
Para a classificação das amostras e das espécies,serão computadas matrizes de similaridade pelocoeficiente de distância de Bray-Curtis (DigbyKempton, 1987; Ludwig Reynolds, 1988), calculadoentre cada par de amostras (análise normal) ou deespécies (análise inversa) da matriz básica de dados.Essas matrizes serão submetidas à análise deagrupamentos pelo método de ligação média não-ponderada (UPGMA; Sneath Sokal, 1973), obtendo-sedendrogramas que representam, em graus desimilaridade, as relações entre o conjunto de amostrasou de espécies consideradas. A distorção produzida pelosdendrogramas será avaliada mediante o cômputo docoeficiente de correlação cofenética (Sneath Sokal,1973). Os valores deste coeficiente variam geralmenteentre 0,60 e 0,90, sendo os valores superiores a 0,80indicadores de baixa distorção em relação à matriz desimilaridade original (Sneath Sokal, 1973). O nível desimilaridade considerado para a delimitação dosagrupamentos será de 50%.
Como técnicas de ordenação serão utilizadas aanálise de componentes principais (Sneath Sokal,1973; Digby Kempton, 1987; Ludwig Reynolds, 1988)e a análise de correspondências pelo método das médiasrecíprocas (Gauch, 1982; Digby Kempton, 1987).
A análise de componentes principais será efetuada apartir da matriz de covariância não centrada (LudwigReynolds, 1988; Legendre Legendre, 1998) entre asespécies. Desta matriz serão extraídos os autovalores eseus autovetores associados, sendo os dados originaiscombinados aos autovetores e projetados sobre os doisprimeiros eixos ortogonais resultantes. Esses eixos,mutuamente perpendiculares e não correlacionadosentre si, explicam sucessivamente a maior parte davariação presente nos dados originais, com um mínimode perda de informação.
A análise de correspondências pelo método dasmédias recíprocas será aplicada a uma matriz deassociação de qui-quadrado, computada simultanea-mente entre as espécies e as amostras, sendo os valoresresultantes ajustados em percentagens. A análise decorrespondências também é uma técnica vetorial;porém difere da análise de componentes principais pelaordenação simultânea das amostras e das espécies,permitindo destacar a correspondência entre ambas, epelo uso de uma matriz de associação de qui-quadrado
&&
&
&
&
&& &
&
&&
207
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé

em lugar de uma matriz de covariância ou correlação(Gauch, 1982).
A eficiência das análises de ordenação efetuadas poresses métodos será julgada de acordo com os seguintescritérios: 1) facilidade de interpretação ecológica; 2)dispersão efetiva dos pontos em contraste com suaaglomeração em um único grupo; 3) anulação do "efeitode arco" uma distorção curvilinear do segundo eixo deordenação devida à dependência quadrática, ou demaior grau, deste eixo com o primeiro, e que pode afetarseriamente o resultado de uma ordenação (Gauch, 1982;Ludwig Reynolds, 1988).
Todas as análises numéricas serão efetuadas atravésdos sistemas de computação científica Octave (Eaton,1997) e R (Ihaka Gentleman, 1996), que combinam acapacidade analítica com a visualização gráfica emambientes interativos versáteis e flexíveis. dotados deuma linguagem de programação extensível.
Os maiores obstáculos ao intercâmbio efetivo deinformação biológica não são técnicos, mas resultam dediferenças entre as opiniões dos pesquisadores e aspráticas de manejo dos dados biológicos.
A adoção de definições comuns para terminologiasbiológicas, ecológicas e geográficas é um pré-requisitopara a comunicação genuína (Blake ., 1994; Karp,1993; Allkin, 1997, 1998). O Projeto Banco de Dados doBiotupé possibilitará uma troca efetiva de informaçãoatravés do estabelecimento e documentação dapadronização de dados e de protocolos para o in-tercâmbio de dados. Padrões internacionalmenteaceitos (como o formato DELTA), quando existirem,serão adotados e seguidos.
Também necessários são protocolos de troca de dadoscomuns a todos os projetos. No âmbito do ProjetoBiotupé, será usada para intercâmbio de dados aLinguagem de Marcação Extensível (XML), entre projetosindividuais e sistemas de software. A Linguagem deMarcação Extensível (XML) é uma linguagem demarcação poderosa e independente de plataforma,crescentemente usada para representação eintercâmbio de informação na Internet (Bosak Bray,1999). Aplicações que consomem e servem seus dadosem XML estão imediatamente aptas a interoperar,mesmo que implementadas em linguagens diferentes e
&
&
&
INTERCÂMBIO DE INFORMAÇÃO
et al
execu t a n do em p l a t a f o rma sdiferentes. Muitos grupos de pesquisajá vem usando XML para troca degrandes volumes de informação entrebancos de dados em instituiçõescientíficas que utilizam plataformascomputacionais, programas eestruturas de dados completamentediferentes (Cavalcanti, 2001).Aplicações específicas de XML têmsurgido em áreas da Biologia como aSistemática Biológica (Gilmour, 2000).Por fim, além de seu potencial para adefinição de formatos padronizadospara o intercâmbio de informaçãocientífica, XML oferece ainda umasolução simples e flexível para oproblema da representação demetadados específicos do domínio deconhecimento, de difícil suporte emmodelos de dados predefinidos(Diederich ., 1989; DiederichMilton, 1991; Michener ., 1997;Senso Rosa Piñero, 2003).
Cada banco de dados criado noâmbito do Projeto Biotupé será usadopara prover uma série de produtos deinformação e publicações específicas,cada qual com um conteúdo e formatodiferenciado, dependendo da audiên-cia identificada.
Um dos modos mais inovadores peloqual o Projeto Biotupé estará tornandodisponível a informação gerada pelosseus projetos integrantes terá lugar naInternet. Esta forma de publicaçãopermitirá ao Projeto Biotupé alcançaruma audiência maior, proverinformação atualizada regularmente emeios para buscar contribuiçõesdiretamente, estabelecendo umcontato com a informação mais direto,atual e interativo entre os potenciaiscolaboradores e usuários dos bancos
et alet al
&
&
DISSEMINAÇÃO DAINFORMAÇÃO
208
Cavalcanti

de dados (Green, 1994; Bisby, 2000;Edwards ., 2000; Stevenson .,2003).
Os créditos serão, em todos oscasos, atribuídos aos autores dasinformações incluídas nos bancos dedados. Todas as informações adiciona-das aos bancos de dados serão,também, associadas à fonte original dainformação. Isto possibilitará a todosos participantes do Projeto Biotupé eaos usuários de seus bancos de dadosconhecer não só quem foi responsávelpor uma determinada informação, mastambém porque esta informação foiincluída.
Será dada prioridade à publicaçãode certos tipos de informação, taiscomo:
1) Publ icação de rev i sõestaxonômicas e monografias de gruposimportantes de plantas e animais;
2) Elaboração de listas de espécies,faunas e floras;
3) Produção de manuais deidentificação sobre grupos animais evegetais importantes, guias de campoilustrados e vários outros tipos depublicação dirigidos ao grandepúblico.
Além disso, espera-se publicarversões eletrônicas (em CD-ROM e naInternet) dos diferentes bancos dedados, regularmente atualizadas,buscando manter um alto grau deconfiabilidade e acessibilidade dainformação.
Atualmente, o Projeto Banco deDados do Biotupé está dando osprimeiros passos para compilar eintegrar a informação obtida pelosvários projetos integrantes do Projeto
et al et al
Considerações Finais
Biotupé durante os últimos dois anos. No decorrer dospróximos anos estes bancos de dados crescerão emtermos do número de espécies e indivíduos, aumentarãoem quantidade de conhecimento para cada espécie emelhorarão em termos de consistência e confiabilidade.
Os projetos de bancos de dados não consistem apenasde dados, programas e computadores, envolvendo aconsideração e discussão de diversos aspectostecnológicos, epistemológicos, cognitivos e sociaisinterrelacionados (Wersig, 1993). Para que estesprojetos sejam bem sucedidos, é necessário que sejamorganizados de maneira a serem sensíveis àsnecessidades, capacidades e interesses dos seusparticipantes.
O Projeto Banco de Dados do Biotupé pretendeestabelecer uma confederação de bancos de dados e nãoum único banco de dados monolítico, ou mesmo imporuma estrutura de dados rígida a todos os participantes(Fig. 4). Este modelo reconhece que cada projeto temseus próprios interesses locais e prioridades quenecessitam ser acomodados, ao mesmo tempo em quetambém reconhece a necessidade de coordenar epadronizar o manejo dos dados tanto quanto possível.Esta forma de organização implica que os programasescolhidos para utilização por todos os projetos deverãoatender a certas demandas específicas.
209
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé
Figura 4. Bancos de dados distribuídos do Projeto Biotupé,organizados em uma federação virtual de SGBDs baseados emesquemas globais.
Rede de
Comunicação
SGBD Distribuído
Rede deComunicação
SGBD Distribuído
INPA UCDB
INPA
Peixes
ULBRA
Vegetação
PlânctonVegetação

Um benefício adicional da distribuição do controle dainformação é que o Projeto Banco de Dados do Biotupénão será dependente do sucesso individual de nenhumdos projetos componentes dentro de um tempodeterminado. Nenhuma estrutura rígida será impostadentro de um esquema ambicioso ou centralizado, e abase de conhecimento do Projeto Biotupé crescerágradativamente, na medida em que forem dandoentrada os dados de alta qualidade provenientes dosprojetos participantes.
Uma ampla gama de usuários será capaz de acessarestes recursos de informação em vários locais, paradiferentes propósitos. Os pesquisadores participantesterão mais facilidades para publicação, e os usuáriosterão pronto acesso a dados de qualidade através dequalquer aspecto dos organismos, incluindo usos, nomesvulgares ou sinônimos.
A abordagem do Projeto Banco de Dados do Biotupé,com respeito ao manejo da informação sobre biodiver-sidade, enfatiza a flexibilidade, o compartilhamento e ocontrole local da informação. O modelo permite integrardados de alta qualidade sobre biodiversidade, cominformações sobre uso e as propriedades relativas aouso, e apresentar esta informação em um formato útilpara as instituições e indivíduos responsáveis pelo desen-volvimento sustentável, conservação da biodiversidadee planejamento ambiental naAmazônia.
Aos Drs. Robert Allkin (Royal Botanic Gardens, Kew),Mike Dallwitz (CSIRO Division of Entomology, Canberra) eRichard Pankhurst (Royal Botanic Gardens, Edinburgh)pelo envio de publicações; ao Prof. Paulo Roberto DuarteLopes (UEFS) pelas valiosas sugestões; e, em especial,aos Drs. Eduardo Couto Dalcin (CNIP/PNE), EdinaldoNelson dos Santos Silva (CPBA/INPA) e Veridiana VizoniScudeller (CEULM/ULBRA), pelo contínuo apoio eincentivo.
Allkin, R. 1988. Taxonomically intelligent database
Agradecimentos
Bibliografia Citada
programs. Hawksworth, D.L.(Ed.). .Oxford University Press, Oxford, p.315-331.
Allkin, R. 1997. Data managementwithin collaborative projects.Dransfield, J.; Coode, M.J.E.Simpson, D. (Eds.).
Royal BotanicGardens, Kew, London p. 5-24.
Allkin, R. 1998. Effective managementand delivery of biodiversityinformation. Bridge, P.; Jeffries,P.; Morse, D.R. Scott, P.R. (Eds.).
CABInternational, Wallingford p. 87-102.
Allkin, R. Bisby, F.A. 1988. Thes t r u c t u r e o f monog r aph i cdatabases. , 37: 756-763.
Allkin, R.; Moreno, N.P.; GamaCampillo, L. Mejia, T. 1992a.Multiple uses for computer-storedtaxonomic descriptions: keys forVeracruz. 41(2): 413-435.
Allkin, R.; White, R.J. Winfield, P.J.1992b. Handling the taxonomicstructure of biological data.
, 16: 1-9.Allkin, R. Winfield, P.J. 1993.
Cataloguing biodiversity: newapproaches to old problems.Biologist, 40(4): 179-183.
Askevold, I.S. O'Brien, C.W. 1994.DELTA, an invaluable computerprogram for generation oftaxonomic monographs.
, 87(1): 1-16.Berendsohn, W.G.; Anagnostopoulos,
A.; Hagedorn, G.; Jakupovic, J.;Nimis, P.L.; Valdés, B.; Güntsch, A.;Pankhurst, R.J. White, R.J. 1999.A comprehensive reference model
In:Prospects in Systematics
.
In:
Plant Diversityin Malesia III.
.
In:
Information Technology, PlantPathology and Biodiversity.
.
Taxon
Taxon,
Mathematical and ComputerModelling
Annals ofthe Entomological Society ofAmerica
&
&
&
&
&
&
&
&
210
Cavalcanti

211
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé
for biological collections and surveys., 48: 511-562.
Bisby, F.A. 1989. Databases, informationsystems, and legume research. In: Stirton,C.H. Zarucchi, J.L. (Eds.).
. Missouri Botanical Garden,St. Louis p. 811-825.
Bisby, F.A. 1993. Species diversity knowledgesystems: the ILDIS prototype for legumes.
, 700: 159-164.Bisby, F.A. 2000. The quiet revolution:
biodiversity informatics and the Internet., 289: 2309-2312.
Blake, J.A.; Bult, C.J.; Donoghue, M.J.;Humphries, J. Fields, C. 1994.Interoperability of biological data bases: ameeting report. , 43(4):585-589.
Bosak, J. Bray, T. 1999. XML and the second-generation Web. ,280(5): 79-83.
Cavalcanti, M.J. 1991. ECOLOG: um sistema debanco de dados para levantamentosecológicos de campo.
, Salvador,557pp.
Cavalcanti, M.J. 1993. ECOLOG: um sistemagerenciador de bancos de dados paralevantamentos ecológicos de campo.R
, Campinas p. 44-45.
Cavalcanti, M.J. 1996. Designing a graphicaluser inter face for DELTA: someconsiderations. , 12: 2-4.
Cavalcanti, M.J. 1998a. Aplicações de SistemasGerenciadores de Bancos de Dados emEcologia e Sistemática.
, 20(1): 5-20.Cavalcanti, M.J. 1998b. A Windows-based data
capture program for ATF., 6: 13.
Cavalcanti, M.J. 2001.
Taxon
Advances inLegume Biology
.
Annals of the New York Academy ofSciences
Science
Systematic Biology
Scientific American
Resumos do XVIIICongresso Brasileiro de Zoologia
esumos do I Simpósio de Aplicações daInformática em Biologia .
DELTANewsletter
Acta BiologicaLeopoldensia
Alice SoftwareNews
Representação eintercâmbio de informação científica naInternet através da Linguagem de Marcação
&
&
&
Extensível (XML)
Plumeria
Serra deMacaé de Cima: Diversidade Florística e Conservaçãoem Mata Atlântica
.
Taxon
Advances in Computer Methods for SystematicBiology: Artificial Intelligence, Databases, ComputerVision
.
Guide to the DELTA System: A General System forProcessing Taxonomic Descriptions
User'sGuide to Intkey: A Program for InteractiveIdentification and Information Retrieval
User'sGuide to the DELTA Editor
Conservation BiologyMultivariate
Analysis of Ecological Communities
In:
. Tese de Mestrado, InstitutoBrasi le iro de Informação em Ciência eTecnologia/Universidade Federal do Rio de Janeiro,Rio de Janeiro, RJ. 52pp.
Cooper, A. 1984. Application of multivariate methods tothe conservation management of hazel scrub inNortheastern Ireland. Biological Conservation, 30:341-357.
Dalcin, E. 1998. Aspectos na implementação de sistemasde registros informatizados em jardins botânicos.
, 6: 34-46.Dalcin, E.; Solano, L. Pizarro, R. 1997. De banco de
dados a centro de informações e serviços: umaexperiência para a Reserva Ecológica Macaé de Cima.In: Lima, H.C. Guedes-Bruni, R.R. (Eds.).
. Instituto de Pesquisas JardimBotânico do Rio de Janeiro, Rio de Janeiro p. 307-314.
Dallwitz, M.J. 1980. A general system for codingtaxonomic descriptions. , 29(1): 41-46.
Dallwitz, M.J. 1993. DELTA and INTKEY. In: Fortuner, R.(Ed.).
. The Johns Hopkins University Press,Baltimore, Maryland p. 287-296.
Dallwitz, M.J.; Paine, T.A. Zurcher, E.J. 1993. User's
. 4th edition.CSIRO Division of Entomology, Canberra. 136pp.
Dallwitz, M.J.; Paine, T.A. Zurcher, E.J. 1995.
. 1st edition.CSIRO Division of Entomology, Canberra. 28pp.
Dallwitz, M.J.; Paine, T.A. Zurcher, E.J. 1999.. CSIRO Division of
Entomology, Canberra. 42pp.DeVelice, R.L.; DeVelice, J.W. Park, G.N. 1988.
Gradient analysis in nature reserve design: A NewZealand example. , 2: 206-217.
Digby, P.G.N. Kempton, R.A. 1987.. Chapman Hall,
London. 206pp.Diederich, J.; Fortuner, R. Milton, J. 1989. Building a
knowledge base for plant-parasitic nematodes:description and specification of metadata.
&
&
&
&
&
&
&&
&

212
Cavalcanti
Fortuner, R. (Ed.).Plenum Publishing, New
York p. 65-76.Diederich, J. Milton, J. 1991. Creating domain specific
metadata for scientific data and knowledge bases.I
, 3(4): 421-434.Diederich, J. Milton, J. 1993. Expert workstations: a
tools-based approach. In: Fortuner, R. (Ed.).
The Johns Hopkins University Press,Baltimore, Maryland p. 103-124.
Eaton, J.W. 1997.
. Department of ChemicalEngineering, University of Wisconsin, Madison.346pp.
Edwards, J.L.; Lane, M.A. Nielsen, E.S. 2000.Interoperability of biodiversity databases:biodiversity information on every desktop. ,289: 2312-2314.
Ellis, D.V. 1988. Quality control of biological surveys., 19(10): 506512.
Gauch, H.G., Jr. 1982.. Cambridge University Press,
Cambridge. 239pp.Gilmour, R. 2000. Taxonomic markup language: applying
XML to systematic data. , 16(4): 406-407.
Green, D.G. 1994. Databasing diversity - a distributed,public-domain approach. , 43(1): 51-62.
Green, R.H. 1980. Multivariate approaches in ecology:The assessment of ecologic similarity.
, 11: 1-14.Herring, T.A. 1996. The Global Positioning
, 274(2): 32-38.Ihaka, R. Gentleman, R. 1996. R: A language for data
analysis and graphics., 5: 299314.
Janzen, D.H. 1994. Priorities in tropical biology., 9(10): 365-367.
Karp, P.D. 1996. Database links are a foundation forinteroperability. , 14: 273-279.
Kurtz, B.C. Araujo, D.S.D. 2000. Composição florística
Nematode Identification andExpert-System Technology.
.
EEE Transactions on Knowledge and DataEngineering
Advances in Computer Methods for SystematicBiology: Artificial Intelligence, Databases, ComputerVision.
.GNU Octave. A High-Level Interactive
Language for Numerical Computations, Edition 3 forOctave version 2.1.x
Science
Marine Pollution BulletinMultivariate Analysis in
Community Ecology
Bioinformatics
Taxon
Annual Reviewof Ecology and Systematics
System.ScientificAmerican
Journal of Computational andGraphical Statistics
Trendsin Ecology and Evolution
Trends in Biotechnology
&
&
&
&
&
e estrutura do componente arbóreode um trecho de Mata Atlântica naEstação Ecológica Estadual doParaíso, Cachoeiras de Macacu, Riode Janeiro, Brasil. , 51:69-112.
Lawton, J.H.; Bignell, D.E.; Bolton, B.;Bloemers, G.F.; Eggleton, P.;Hammond, P.M.; Hodda, M.; Holt,R.D.; Larsen, T.B.; Mawdsley, N.A.;Stork, N.E.; Srivastava, D.S.Watt, A.D. 1998. Biodiversityinventories, indicator taxa andeffects of habitat modification intropical forest. , 391: 72-76.
Legendre, P. Legendre, L. 1998.. 2nd ed.
Elsevier Science, Amsterdam853pp.
Ludwig, J.A. Reynolds, J.F. 1988.
. JohnWiley Sons, New York. 337pp.
Maddison, D.R., Swofford, D.L.Maddison, W.P. 1997. NEXUS: anextensible f i le format fors y s t e m a t i c i n f o r m a t i o n .
, 46(4): 590-621.
Michener, W.K.; Brunt, J.W., Helly,J.L., Kirchner, T.B. Stafford, S.G.1997. Nongeospatial metadata forthe ecological sciences.
, 7: 330-342.Morain, S.A. 1993. Emerging
technology for biological datacollection and analysis.
, 80:309-316.
Pankhurst, R.J. 1991.. Cambridge
Rodriguésia
Nature
Numerical Ecology.
Statistical Ecology: A Primer onMethods and Computing
Systematic Biology
EcologicalApplications
Annals ofthe Missouri Botanical Garden
PracticalTaxonomic Computing
&
&
&
&&
&
Margules, C.R. 1984. Vegetationinventory and conservationevaluation: a case study from theNorth York Moors, England.
, 4: 293-307.Applied
Geography

213
Bancos de dados sobre biodiversidade na Amazônia: a experiência do Projeto Biotupé
University Press, Cambridge. 202pp.Sá, C.F.C.; Araujo, D.S.D.; Lima, H.C.;
Cavalcanti, M.J.; Pereira, M.C.A.Fontoura, T. 1991. Composição florística eestrutura da floresta de cordão arenoso daReserva Ecológica de Jacarepiá, Saquarema(RJ): dados preliminares.
p. 527pp.Senso, J.A. Rosa Piñero, A. 2003. El concepto
de metadato. Algo más que descripción derecursos electrónicos.
, 32(2): 95-106.Schalk, P.H. 1992. Computer-aided taxonomy.
, 4: 124-126.Sharkey, M.J. 2001. The All Taxa Biological
Inventory of the Great Smoky MountainsNational Park. , 84(4):556-564.
Sneath, P.H.A. Sokal, R.R. 1973.
. W.H. Freeman,San Francisco. 573pp.
Stevenson, R.D., Haber, W.A. Morris, R.A.2003. Electronic field guides and user
&
&
&
&
Resumos do XLIICongresso Nacional de Botânica, Goiânia.
Ciência daInformação
Binary
Florida Entomologist
NumericalTaxonomy: The Principles and Practice ofNumerical Classification
communities in the ecoinformatics revolution., 7(1): 3 [online]. URL:
http://www.consecol.org/vol7/iss1/art3.Wersig, G. 1993. Information Science: the study of
postmodern knowledge usage., 29: 229-239.
White, R.J. Allkin, R. 1992. A language for thedefinition and exchange of biological data sets.
, 16(6/7):199-223.
White, R.J.; Allkin, R. Winfield, P.J. 1993. Systematicdatabases: the BAOBAB design and the ALICE system.
Fortuner, R. (Ed.).:
The Johns HopkinsUniversity Press, Baltimore, Maryland p. 297-312.
Yoon, C.K. 1993. Counting creatures great and small., 260: 620-622.
Zarucchi, J.L.; Winfield, P.J.; Polhill, R.M.; Hollis, S.;Bisby, F.A. Allkin, R. 1993. The ILDIS project on theworld's legume species diversity. Bisby, F.A.;Pankhurst, R.J. Russell, G.R. (Eds.).
ClarendonPress, Oxford p. 131-144.
Conservation Ecology
InformationProcessing and Management
Mathematical and Computer Modelling
In: Advances in Computer Methodsfor Systematic Biology Artificial Intelligence,Databases, Computer Vision.
.
Science
In:Designs for a
Global Plant Species Information System..
&
&
&
&


3CAPÍTULO 15 - Histórias: narrativas e depoimentos das comunidades São João do Tupé
e Colônia Centralenrique REBÊLO; Adriana Kulaif TERRA;Viviane Maria Guedes LAYNE Tábita
Maciel AMORIMGeorge H &
CAPÍTULO 16 - A visão vernacular e o desafio cultural para construções em mutirãoLeandro GASPARINI André Munhoz de Argollo FERRÃO&
DiversidadeSociocultural
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),


Diversidade SocioculturalCapítulo 15
Histórias: narrativas e depoimentos dascomunidades São João do Tupé eColônia Central
RESUMO
PALAVRAS-CHAVE:
- Este capítulo retrata a história de duas comunidades da Reserva deDesenvolvimento Sustentável do Tupé (RDS Tupé). As narrativas e depoimentos foramregistrados por quatro pesquisadores. Das vinte entrevistas gravadas com seisinformantes, foram selecionados trechos que: (1) contam as origens dos atuais moradoresda RDS Tupé, e a história das comunidades (2) São João do Tupé e (3) Colônia Central.Ambas as comunidades são exemplos distintos da capacidade de seus moradores emtransformar a paisagem.Apraia do Tupé foi construída por seus moradores para terem umanova fonte de renda. O ramal da Colônia Central foi aberto em mata bruta por gente que foienganada. Este registro deveria ser replicado nas demais comunidades para subsidiar omanejo da RDS Tupé.
história oral, narrativasAmazônicas, história ecológica.
George Henrique REBÊLO
Adriana Kulaif TERRAViviane Maria Guedes LAYME
Tábita Maciel A
1
1
2
& MORIM2
e-mail: [email protected].
INPA, Coordenação de Pesquisas em Ecologia, Caixa Postal 478, 69086-970, Manaus-AM,UNIR, Instituto de Biologia, Porto Velho-RO
1
2
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

Introdução
Material e Métodos
A história oral de duas comunidades foiregistrada por quatro pesquisadores. São João do Tupé éuma comunidade no Lago Tupé de onde se avista o RioNegro. O encantador lago Tupé já produziu lenha,pedras, peixes e agora serve ao turismo de massa. AColônia Central é uma área de mata alta em torno de umramal central ocupado por casas, roças e capoeiras. Oramal rasga a floresta entre as cabeceiras do Tupé, o RioNegro, o igarapé Acácio e o Tarumã-Mirim há apenas dezanos e a mata ainda se conserva bela e imponente.
Ambas as comunidades partilham a área de umamesma unidade de conservação. Criada à revelia de seusmoradores, a hoje Reserva de DesenvolvimentoSustentável do Tupé (RDS Tupé) prevê o uso prudente esustentável dos recursos na área, zona rural de Manaus(AM). Para conhecer seus moradores e sua relação com olugar, gravamos e transcrevemos cerca de vinteentrevistas e depoimentos contados nas comunidades deSão João e Colônia Central por sete pessoas, de váriasidades e funções, entre maio de 2002 e janeiro de 2004.As transcrições registram com maior fidelidade (exceto apronúncia) a fala dos informantes. Selecionamosalgumas narrativas e depoimentos do pequeno númerode informantes independentes: Barú, Domingos,Glicério, Jaumir. As pessoas mais velhas entre os que sedispuseram a separar parte do seu tempo para vasculharsuas lembranças e contar histórias que vão subsidiar omanejo dos recursos da RDS Tupé.
Um capítulo em três atos: no primeiro contam ahistória anterior e as origens, dos atuais moradores. Nosegundo histórias da fundação da comunidade São Joãodo Tupé e os usos atuais do Lago. No terceiro a fundação ehistória recente da Colônia Central. Todos de acordo coma versão transcrita pelos pesquisadores do PROJETOBIOTUPÉ (2004). As convenções se baseiam em trabalhossobre narrativas orais populares da Amazônia (p.ex.Simões Golder 1995), daí a presença de marcas deoralidade, traços regionais, expressões e construçõespeculiares relacionadas as circunstâncias da gravação e à
&
fidelidade de transcrição.Aparecem, os seguintes símbolos:[ ] utilizados para enquadrar
palavras ou expressões que causaramdúvidas ao transcritor ou esclarecem afala. Quando vazios, indicam silêncioprolongado (hesitações, embaraços,timidez).
( ) enquadram a fala dopesquisador, ou sua descrição dosefeitos da narrativa sobre os presentes(riso, raiva, etc.).
ACAR -Associação Comunitária Agrícola Ruralda Comunidade Colônia Central. ACIRU- Associação das ComunidadesIndígenas do Rio Umari. CEM - CentraisElétricas de Manaus. FUNAI - FundaçãoNacional do Índio. IBAMA - InstitutoBrasileiro do Meio Ambiente e dosRecursos Naturais Renováveis. INCRA -Instituto Nacional de Colonização eReforma Agrária. SEDEMA - SecretariaMunicipal de Desenvolvimento e MeioAmbiente. SEMTA Serviço Especial deMobilização de Trabalhadores para aAmazônia (Mart inel lo 1988).SIDERAMA - Companhia Siderúrgica daAmazônia. SUSAM - Secretaria deEstado da Saúde doAmazonas.
O Soldado da BorrachaDepoimento de Glicério Luis
Ferreira, 81 anos, aposentado, nuncaestudou.
-Eu sou nordestino. Eu sou de umlugar por nome Caiçara, mas euquando vim pro Amazonas eu tavanuma cidade do Rio Grande do Nortepor nome Areia Branca, é de lá foi queeu vim.
(e o senhor veio pra cá por quê ?)-Naquela época eu era soldado da
borracha. Tinha aquele alistamento,
Lista de Abreviaturas:
,
Origens dos atuais moradores
218
Rebêlo et al.

219
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central
eu cheguei em Manaus em [1943].(o senhor veio pra cortar seringa?)-Foi pra trabalhar na seringa.Aí eu [
], naquele tempo aí eu [ ], era abaixo,aí abaixo um pouco daquele aeroportode Ponta Pelada, era o nosso barracão,que nós ficava, era ali. Aí eu chegueinum dia e no outro dia eu vim pra[estiva]. Só que quando eu cheguei lá,cheguemo aqui, ainda fizemo,cheguemo aqui no dia oito desetembro. No dia sete de setembro nóstivemo em Santarém, nós passamo emSantarém, aí parou lá, aí o pessoal tavamarchando [...] ficamo pra lá. Aí assimque eu cheguei, um rapaz chegouassim perto de mim, falou comigo,perguntou de onde eu era, eu disse queeu era cearense e ele disse que eraparaibano de Patos [ ]. Aí ficamo logoamigo, ele já tava ali há dias, hátempos, há meses, né? E eu cheguei nooutro dia, aí muito bem, conversamos,quando foi no outro dia de manhã nahora do café, conversamos de novo equando foi na hora do almoço [ ] aí ele:“e aí cumprade tu não quer trabalharna estiva?', “Quê rapaz? Quero, sim...”Era um tenente da polícia que estavaalistando 20 homens. “Vem cá”, aí eusaí da fila, deixei o negócio da comidapra lá, cheguei lá, o tenente [ ], otenente olhou. Aí ele: “Tenente!” queele já tinha trabalhado com o tenentena estiva. Não sei o problema da estivaque eu não sei o que era, quandofaltava gente ele ia lá e listava aquelaquantidade que precisava. Aí ele disse:“eu trouxe aqui este rapaz prosenhor.” “Ah, já tá cheia a lista.”Aí eleolhou: “é esse aí?”, “é”, “ah, eu voutirar um daqui e boto ele.” Aí tirou ooutro que era só [ ], tirou o camaradalá, que eu não sei quem era e colocou omeu nome. Aí ele disse: “olha quandovocês terminarem de almoçarem, cê
acompanhe ele que ele já sabe pra onde é que é.”Justamente era pra nós ir lá na [ ] o nosso lugar era ali naSão Vicente, bem na pra li, quase próximo ao quartel do[Jessias]. Nós trabalhava na estiva e dormia ali, porãoduma casa, mas tinha tudo, tinha banheiro tinha tudo.Quando [ ], quando terminava o serviço, naquele tempoera o tempo da guerra, que tinha chegado um navio aí o [], carregado de mercadoria, nós vimo de noite, viajandode noite [ ]. Quando terminou o serviço nós ia pra PontaPelada, aí a gente ia trabalhar no aeroporto [ ], eranaquele aeroporto que tinha ali , antes ainda trabalheiali, trabalhei ali. Aí era pra ir pro [rio] Purus, [ ] era pra irpro Purus e nunca era chegava pra ir pro Purus. A gentequeria pra trabalhar, aí, aí quando, mas só tinha a gentesó ia pra estiva, quando tinha serviço [ ], assim[ ] demaisné? Quando era mesmo serviço pra estiva, a estivatomava de conta, pra trabalhar. Nóis só ia quando eraassim, muito, aí que ia lá pegar. Quando terminava agente ia lá pra, pro nosso barracão lá. Nisso era que nóstrabalhava, sempre [ ], levava nós pra lá, pra nóstrabalhar, quer dizer que, na estiva nós ganhava dois milréis, naquele tempo. E ganhava três pela companhia.Que de lá que, que pra gente [ ], pros casado ficavaganhando oito. Sempre a família residia lá. E três[ ] elesrecebia todos os meses em viagem. E os solteiros sóganhavam três reais, pela companhia.
(como pela companhia?)-Era uma companhia que trazia os soldados da
borracha de lá, era uma companhia, [ ] SEMTA, umacoisa assim. Era um negócio de, de, naquele tempo daguerra. Eu não sei quem se lembra dessa companhia, eusei que na época foi um aperreio. Nós viemo, de, do, eusaí do Rio Grande do Norte, de Areia Branca pra Mossoróe de Mossoró [ ], até São Luiz do Maranhão, nós em cimade um caminhão de [ ], viemo pegar navio aqui em SãoLuiz do Maranhão e de São Luiz do maranhão pra Belém,quando a gente saía era todo o dia um avião em cima dagente, era muito perigoso, negócio da guerra, onde tinhaaquele trecho ali do São Luiz pra, pra[ ], de São Luiz praBelém, tinha uma passagem lá que era muito perigosa ede noite ia viajar. O avião rodava o dia quase todo.Quando eu [ ], quando nós se alistemo pra, pra vim praestiva, um rapaz que era lá do Rio Grande do Norte, euconheci ele lá, num pegô a vaga, mas tinha outroalistamento [ ] lá pra olaria do Brito, ali do outro lado. NoCacau [Pirera], ali perto do Cacau, pois bem. Eu pra vim

220
Rebêlo et al.
pra cá eu morava lá, pois bem. Ele não pegou pra estivamais pegou pra lá. E ele sabia que nós tava na estiva. Aíele veio [ ], quando foi um dia ele veio bater lá onde nóstava, e contou que lá era barato e o serviço era barato,era 11 mil rés, 11 reais, cruzeiro, naquele tempo nãotinha nada de real, era 11 cruzeiro. E quando faltouserviço aí, aí nós fumo[ ], era dum doutor, que morava aína, no [ ], na rua do[ ], era ali pra dentro que ele morava.Nós fumo lá com ele, se vocês querem lá, pra trabalharpra lá, tem lugar lá, e se vocês quiserem lá pro Curari, eutenho um terreno lá no Curari, que tá precisando degente, mas nós já tinha se conhecido lá na olaria, aí nósfumo trabalhar na olaria. Trabalhei na olaria.Trabalhemo lá quatro anos lá, aí foi tempo que euinventei de pescar, porque o dono, o chefe mesmo queera seu Brito, foi embora pro Portugal, né? Que era [ ] umdos portugues, um dos dono. Afinal de contas eram ostrês portugueses, [ ] era chefe da companhia, o patrão, odono. Aí foi pra Portugal e lá morreu. Aí foi [ ], aímeteram outro sócio não deu certo de jeito nenhum. Aíeu fui pescar, pescar e tal e tal. Passei uns anospescando.
(O que pescava)?-Era [ ] peixe [ ], pescava de rede pra vender em
Manaus. Naquele tempo dava muito Tambaqui.(na malhadeira?)-Não, não. Naquele tempo, quando eu comecei a
pescar ainda não havia malhadeira. Depois de um certotempo, é que começou a malhadeira.
(a rede que o senhor fala é de?)-É de arrastar. Aí eu trabalhei, trabalhei, foi o tempo
que [ ] eu arrumei família, aí eu digo: não, vou mequietar num canto[ ].Aí eu [ ] perguntei da minha mulheraqui como era, que eu não conhecia aqui não. Morava noCacau mas eu só me dei, eu só achei ela porque ela tinhauma irmã pra lá e foi passar uns dia pra lá, lá pro Cacau eeu tava lá né? Por lá nós se ajeitemo né? Aí nós, eu jáconhecia a mãe dela, aí ela contou a situação dela aqui.Será que dá pra nós vive lá? Ela disse que dá [ ], certo.[Deixei] a minha pescaria e vim me embora pra cá [Tupé].
Depoimento de Domingos Velozo Vaz, anos, índioDessana.
-Nós somos naturais do município de São Gabriel da
Aqui é onde cai o céu
Cachoeira, nós somos do Distrito dePari Cachoeira, nas fronteiras daColômbia e Brasil. É [ ] um Distrito, éuma missão dos padres. É uma aldeia,era uma aldeia antigamente e hoje emdia é um distrito [ ] local. Chama-sePari Cachoeira. Onde nós nascemos,onde nos criamos e onde nos formamosa primeira [ ], o nosso estudo daprimeira série à oitava. Bom, lá nóssomos várias etnias. Somos váriasetnias, somos 23 etnias, lá em PariCachoeira. Então [ ] eu, ele, todos nósque estamos aqui: Raimundo VelosoVaz é meu irmão maior da família [ ] é,e a Clemência Vaz que é a nossa irmã, aprimeira filha, da família, segundo é oRaimundo, terceiro é o Henrique que éadministrador da FUNAI lá em SãoGabriel da Cachoeira, e o quarto soueu e a quinta é a Ester, a nossa caçula, [] é da etnia [Dessana]. Agora orestante: essa aqui é a mulher, é amulher dele, a esposa dele, é a[Tukano] Tukana, [ ] é o nome dela éAurora. Ela é da etnia TukanaBatitorobó. Aí parte para a parte da,da, da minha [ ] irmã, ela é viúva einclusive tá aqui com a gente e o filhodela tá aqui, [ ] que é Tukano. Entãoela se casou com um Tukano e hoje emdia [ ], ela é Dessana, casou com umTukano e ele é Tukano, filho de Tukano,aí ele é Tukano ele, nosso sobrinho. Eleé [ ] da etnia Tukana, da classe [ ]. Aídepois parte o filho dele que é oprimeiro filho Zé Maria [ ].
(Esse primeiro aqui ou o outro?)-O outro.[ ] Zé Maria por favor vem
por aqui, tá? Fica feio assim na rede. [ ]Então o filho dele vem como o primeirofilho dele. A esposa dele é Tukana, queé da mesma classe dela, que é [ ]. AMaria da Conceição [ ], é de lá tambémda, todo mundo é de Pari Cachoeira. Aídepois vem ele, que é o filho dele que

é solteiro. Aí depois vem a outra que táem Barcelos, a filha dele e depois vemessa menina que tava ainda agora aqui,que ta pra lá e depois vem essa aqui. Euvenho depois, a minha esposa éTerezinha Prado, ela é Tuyuka. De látambém, das cabeceiras [ ] sabeJacaré, das cabeceiras do Rio [ ]. Ela élegítima e original Tuyuka [ ] índiamesmo (risos). É por isso que ela veiopra cá, pra [ ] cansou de viver no matoe veio pra cá viver num [ ] espaçogrande pra [ ]dizem que, lá nasfronteiras, dizem que aqui é onde cai océu, o limite do céu, [ ] do céu, dizemaqui. Antigamente consideravam comoum bairro, pois é e hoje em dia écapital. Então, ficou assim. Então nósestamos aqui [ ] eu tenho cinco filhos,o meu primeiro filho vai se formar parao ano, terceiro ano do segundo grau,aqui em Manaus mesmo [ ] e a outrasegunda filha tá se formando esse anooitava série [ ]e o outro tá fazendosexta série, a outra tá fazendo terceira[ ] e o pequenininho tá fazendo [ ] é,primeiro ano. Eu tenho cinco filhos.Tão todos em Manaus. [ ] Aí depois vêma nossa caçula [ ], ela só tem um filho,ele tá se formando na oitava sérietambém esse ano [ ], ela só tem umfilho. É a Ester. Ela só tem um filho.Então, a gente se forma uma família.Fora dos que estão aqui, temos aindaoutras famílias que vão vim ainda maiscinco famílias pra cá e vamoscompletar umas dez famílias pra poderlevar, trabalhar à noite e [ ] fazer oserviço que paguem o nosso trabalhotambém, como cidadão.
Depoimento de Álvaro OliveiraBastos, o Barú, 49 anos.
NasciAqui
-Nasci aqui no Tarumã-Mirim, em 1954, 26 de outubrode 1954. Meu pai entrou pra trabalhar aqui nessa área em1949. Então nós passamos 10 anos fora daqui, na cidade,meu pai trabalhou no Tarumã também um tempo, né?Depois voltamos pra cidade de novo e retornamos pra cápro Tarumã em 1962. Então nós tamos morando aídefinitivamente desde 1962.
(O Julião existe desde quando?)-Julião existe há muitos anos, muito antes de 49 já
existia esse nome Julião.(Já era uma Comunidade?)-Não era só um igarapé, as casas eram tudo distante
uma da outra, né? Pra se visitar um amigo tinha que tirarum domingo. Saía de manhã e só retornava à noiteporque era só remando e era dificultoso, os igarapés eramuito cheio de mato, né? Poucas casas mesmo. Bom,essa área aqui nós tamos no Tarumã, área do Julião, erauma área que não se tocava em nada, era uma áreaintocável, né? Porque eles tinham título definitivo e umcidadão por nome Carlos [Stordate]. Eu tenho o mapadado pelo INCRA, né? Eu peguei dia 15 de maio de 2002,eu tenho completo ele lá com nome, metragem da áreatodinha, né?
(Lá no Julião?)-Julião.Ainda hoje eu tava olhando esse mapa, né?(Mas qual que é a área?)-Essa área toda que engloba: Tarumã-Miri, as
cabeceiras do Tarumã-Miri, Acácio, extremando com oRio [Cuieras].
(Era o que, era seringal?)-Não. Antigamente, dizem que existia um seringal,
né? Em 63, nós conversamos com um senhor de idade, elejá tinha uns 85 anos e ele contava que existe um seringalentre a cabeceira doAcácio e a cabeceira doArara. E queeles jogavam a borracha lá no Sucuba. Lá existia umbarracão muito grande e em 45, no tempo do término daguerra, né? Esse barracão ele pegou fogo e desde entãofoi abandonado isso aí. E agora a gente descobriu noINCRA que tinha esse grande título aí, né? [ ] Ninguémpodia tirar uma árvore, alguém ia lá tinha um vigia,comunicava a polícia, a polícia buscava o cidadão nabase da peia. No tempo que ficou assim passou pra mãode um português chamado José Grilo. Dono de umaolaria, né? Aí, José Grilo morreu, ficou na mão da esposadele que era dona Emília, Emília Grilo. Depois ela vendeua posse do finado pro José Sobreira do Nascimento.
221
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

(E aí?Agora tá aonde?)-Agora tá nisso aí, né? Ele morreu e ficou pros
herdeiros, mas conforme o Coordenador da SEDEMA, elesnos orientaram que antes de ele morrer ele passou essaárea do Tupé pra ser uma área de Reserva. Foi Tombadapela SEDEMA.
(Então, pronto. Então é área de RDS)-É, já foi constatado que é.(Aí agora é área do Estado pra ser administrada com
termo de ocupação de posse de todos os moradorescadastrados. Então por isso que as comunidades aqui temque se organizar. Através dessas entidades tem quereivindicar isso. Entendeu? Mesmo por que aqui tem ummonte de outras comunidades além dessa)
Bom, como eu ia contando, né? Quando passou pra oNascimento, aí o povo começaram entrar, tirar umamadeira, cortar madeira, começaram entrar, ele já não[importava] mais. Até que muitos começaram fazer casano terreno, chegou um ponto dele chamar aí em 1972,ele teve uma reunião com o pessoal aí, né? Aí disse queele não ia mexer com ninguém, ia deixar as pessoas quequisessem trabalhar na terra, fazer plantio, né? Mas nãoplantasse [bem de raiz], porque ele não ia brigar com opovo pequeno e ele ia brigar com aquelas pessoasgrandes que ganham dinheiro. [ ] a posse de uma pessoado que brigar com ele. [ ] o Deus dele era o dinheiro. Elenão acreditava em Deus. Então agora ele já morreu, né?Antes dele morrer ele passou essa área pra Reserva, né?Mas ainda tem os herdeiros que parecem que aindafazem alguma questão sobre isso aí. Não tô beminformado, mas existe ainda. Então depois disso criou aprimeira Comunidade: Livramento, depois veio Julião,depois do outro lado também [ ] outro título que é dooutro português chamado Tino.
(Mas isso é do outro lado do rio?)-Do outro lado do rio.
Depoimento de Glicério Luis Ferreira, 81 anos,aposentado, nunca estudou.
(Quando o senhor chegou aqui?)-em [1957]
HISTÓRIADACOMUNIDADE SÃO JOÃO DO TUPÉ
Assim que o povo ficou vigia do lugar
(como era aqui?)-Era um lugar quase sem [ ] o lugar
não tinha nada, aí veio ter nome de umcerto tempo pra cá, foi que veiocomeçar a ter o [ ] porque [ ] quandoesse pessoal conheceu o Tupé.
(mas já chamava Tupé?)-Tupé, agora o pessoal já não
chama mais Tupé, chama: praia doTupé, que não tinha. Essa praia foifeita, todinha ela foi feita manual.
(quem fez essa praia?)-Uma família [ ] a família da
Florência, onde eu tô no meio dafamília. Eu sou casado com a filha dela,que já é falecida. Lá no colégio nóstemos um retrato de uma preta lá,Florência, da Deuza, Deuzuide, que éminha esposa. Ela era merendeira daí.Pois é eu cheguei aqui pra cortarlenha,cortar lenha pro estado e paraser dono de olaria. Quando ela chegoupra cá, minha sogra com ela, esse lugaraqui não era de ninguém [ ], não tinhaninguém,era do estado[ ],era doestado. Agora depois foi que apareceuuma família, um pessoal, de unsportugueses aqui.
(ameaçando?)-Ameaçando. Chegou a minha sogra
[ ], eles[ ], ela chegou aqui em [1945],minha sogra. Aí chegou uns portugues,foi que pediram uma permissão a elapra [ ], que iam cadastrar, pra deixarlegalizar o lugar,[ ]não tinha problema,problema não, com ela não.Justamente eram dois portugueses quetinham ali, na Leonora do[ ], eram doisirmãos, José Brito e Manuel Brito. Eradono de olaria, um do lado e outro dooutro. Pois bem e é assim que o povoficou, vigia do lugar. E foi quando euvim pra cá, eu cheguei, ela chegou em[1950] prá cá, em 50, em 45 e eucheguei em 57.
(veio trabalhar pro estado?)
222
Rebêlo et al.

-Cortava lenha pro estado e prodonos da olaria
(e o estado usava essa lenha emquê?)
-Ah naquele tempo ainda erabombeamento, ainda era [ ] uma parteda, muita parte cortava lenha. [ ] veiopassar, era pra coisa, como nós tinha,era [ ] , depois é que passou pra CEM,foi que diminuiu a lenha. Por que nãose lembra que tinha esse, era Manaus []? Depois que passou pra queimar,depois que passou pra CEM que ficouqueimando diesel, já diminuiu muito ocorte da lenha. Daí foi rebaixando ocorte de lenha, mas era só pra umbombeamento, pra uma padaria etudo.
(e nessa época vocês tiravam lenhade onde?)
-Dessa beira de rio, todo o pessoaltirava e tirava daqui de dentro, só queaí nessa beira de rio não alcancei mais,por causa da tocaria, só dava tocaria,aí eu cortava aqui pra dentro já
(e tinha alguma árvore preferida outirava qualquer uma?)
-Não, não, tirava de qualquer uma(e como é que vocês faziam?)-Eram madeiras quase de igapó que
a gente tirava. Era de igapó mesmo,não tinha muito da terra não, da beirada terra não. Era assim inda não eramadeira de lei era madeira mesmo, erasó mesmo pra fazer as coisas, praqueimar, não tinha madeira de lei não.
(como vocês trabalhavam eram deturma?)
-Era de turma, quer dizer [ ] aequipe aqui era do estado vamos dizerassim né: o senhor que tinha umavontadezinha chegava aí, o senhorquer trabalhar comigo? O que é? Écortar lenha pro estado. Mas comovocê faz? Eu faço a tantos metros ouque fosse de tonelada que o senhor
vendia, eu pago já, tanto! Ah, eu vou! Aí se juntava unsdez ou quinze homens aí entravam pra um lugar assim, aíformava aquela, assim era o dono de um ponto, chegavanuma ponta dessa qualquer, limpava, ajeitava e ficavapra carregar a lenha de canoa naquele ponto, no pontoda embarcação pegada. Quer dizer o senhor era o donode um ponto. Outro chegava e fazia a mesma coisa, 10,15 ou 20 daí por diante.
(e aí tirava no machado?)-É só no machado, naquele tempo ainda não existia
motoserra, se existia, mas não aqui.(e aí arrastavam pra beira?)-É, aí carregava de canoa, colocava no canto e
quando a embarcação, o motor vinha pra carregar etrazer os batelão, aí é que a gente, as vezes era lugar quenão podia nem chegar, tinha que embarcar de canoa denovo pra gente desembarcar nos batelão.
(e tinha uma época certa?)-Tinha, é porque a gente cortava o [ ] hoje, o seu
produto hoje, porque o senhor era o dono. Nós não tinhanada com o estado e nem com qualquer que fosse, onosso negócio era com o senhor, porque o senhor eraquem mandava no seu [ ]. Aí o senhor ia lá e custava prachegar a embarcação pra levar a lenha e o pessoal quequeria o dinheiro já tinha às vezes a carga de um batelãode dois. O senhor ia lá onde o senhor contratou né? Proestado ou quem quer que fosse, aí você guerreava,queria que o batelão viesse logo que é pra levar aquele, efosse em um lugar assim como esse que fecha assim aboca, então[ ] tinha que levar o produto antes de fechar.
(o inverno era a melhor época?)-Era, da enchente cheia.(e quanto eles pagavam?)-Vixe! Era uma mixaria, tinha cada tonelada. Depois
de muito tempo já que passou de 60 cruzeiros. Naqueletempo era cruzeiro ainda.
(uma tonelada?)-É uma tonelada.(o que dava pra fazer com isso?)-Rapaz não dava pra quase nada, mas o jeito que
tinha era dá. Porque não tinha outro meio. Se você faziauma roça, a pessoa fazia uma roça aqui, aí fazia afarinha, aí saia pra vender. Pra vender, naquele tempo afarinha quase toda era empaneirada, não era, agora étoda encharcada, preta, quase toda era empanada. Agente pegava às vezes 10,15,8,6 [ ] paneiros de farinha
223
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

pra levar pra Manaus e passava uma semana andando deSão Raimundo, pu Educando,[ ], levanta pra cuidar dafarinha, ah, não quero ir não, isso e aquilo outro. Eramuito era ruim, era ruim pra ir, só era ruim. Bom pracomer, porque peixe tinha muito, mas pra vender essascoisas assim. Onde pegava um dinheirinho mais fácil erana [lenha] que entregava, o caba recebia logo.
Depoimento de Glicério Luis Ferreira, 81 anos,aposentado, nunca estudou.
(durante quanto tempo o senhor trabalhou na lenhaaqui no Tupé?)
-Eu trabalhei só uns dois anos, não, uns três anos maisou menos na lenha.Aí foi o tempo que veio [fracassiando]também, aí eu, fui trabalhar na roça, fazer roça.
(já era aqui neste sítio mesmo?)-Não, morava mais aqui em baixo um pouco, [...] que
esse igarapé aqui mesmo, que vai aqui, ele tem o nomeda minha sogra, igarapé da Florência e esse outro aqui éo igarapé chefe. Tem muitos braços e todos, todos essesbraços tem nome. Mais os braços chefe mesmo é esse oigarapé da Florência e esse outro é o igarapé Chefe. Poisé, eu fui fazer tupé, trabalhei, trabalhei. Aí, mas antesdisso [ ] começou a chegar aqui um barco, um barcogrande. Isso em [1963], um barco grande. Chegava, []onde tinha esse negócio que é da SEDEMA, [centro devisitantes], naquela parte ali, ele encostava ali, minhasogra morava bem ali perto do barraco. Aí eles falaramcom minha sogra se ela não queria fazer uma limpazinhaali onde tinha um pau, separado de uma [árvore], aindahoje tem uma árvore separada.Aquela árvore tinha que [], era tudo família, era tudo policial, era tudo policial,era o pessoal do exército. Com a família, era tudofamília, era o pessoal do exército, a maioria deles tudosargento reformado, tudo pessoal que fazia serviço prosseus netos, seus [ ], era família né? Não era assim, nãoera nada de bagunça, era só, aí nós fizemo aquela limpa.Encostavam a lancha deles, dia de sábado, todo o sábadoeles vinham.
(então não existia a praia?)-Não. Aí[ ], aí ficou. Ficou ali, ficou ali, não sei o quê
que houve [ ] rodeando, aí entrou a coisa, o [ ] Carlos,
Aconstrução de uma praia:“isso aqui vai ajudar a criar os filhos de vocês”
esposo da que era gerente da VASP,seu Carlos Souza. Ele foi tomar umbanho aí na Ponta Negra e [ ] ele não sedeu lá. Aí um camarada lá informouaqui o Tupé, que tinha uma ponta deterra bonita, justamente a ponta aliera linda, isso aqui, daquele lado dali,só quem ficava daquela ponta alipassava semanas ali, naquela ponta [ ]os que vinham ali passavam semanas,que aqui era muito bonito[ ] caia muito[jacaré], ainda mais pegava [ ] ovos detracajá, rapaz tava aí, passavam erasemana aí. Aí o seu Carlos Souzachegou aí e tal aí falou pra fazer um,limpar ali perto da ponta, a ponta nadarapaz, [ ], [ ], a parte mais pra cima,pra poder fazer uma limpeza ali, mas écoisa pouca. Aí limpamo ali, [ ], aí foise chegando, se chegando mais era sóembarcaçãozinha pequena, não era.
(isso foi já em que ano, que o CarlosSouza chegou?)
-Rapaz não sei em que ano não,mais faz um bocado de ano.
Pois bem, fizemo lá o serviço e ficô.Aí foi quando chegou o pessoal dodistrito. Cristóvão Pinto, ele era nãosei o que da [ ], e ele parece se eu nãotô enganado ele era não sei o que daSanta Casa, ele era agora eu não sei setá ainda. Aí ele chegou aí e pagou pragente fazer aquela limpa daquele ladodo colégio.Aí a gente foi fazendo e tal,aí o pessoal foram chegando, foram seanimando e foram chegando,trabalhando. Aí foi e disseram: “olha,vocês vão limpar isso aqui, pra quevão, vocês limpam, isso vai servir praajuda de vocês. Pra vocês deixarem detrabalhar na roça, serviço de roça nãovai pra frente, vocês tão, táprejudicando o meio [ambiente].Vocês trabalham pra morrer, e ossenhores não vão fazer nada. Vocêslimpam isso aqui, que isso aqui vai
224
Rebêlo et al.

servir pra vocês [ ], ajudar a criar osfilhos de vocês” [ ]. Começemo atrabalhar. Aí quando era dia de sábadoeles traziam uma coisinha, um troço,um quilo de açúcar, um quilo de um,um pacote de café, um quilo de pão,uma coisa assim. Eles passavam,sábado e domingo, saia dava aquelagratificação pra gente.[ ] Sei que nóstrabalhemo aí, um bucado.
(e vocês continuaram trabalhandona roça?)
-Continuemo, trabalhando na, quedizer, que nós gente trabalhava assim,meio dia pra cá e o resto do dia pra lá.Foi, foi, foi, aí chegou o Jalmir que é opresidente o, o presidente dacomunidade. Ele é casado com umafilha minha, aí foi que ajudou. Aíquando nós aprontemo, isso aquiquando nós aprontemo, eles disseram,bom, isso aqui não é pra nós [ ]. Vocêsfizeram, isso é de vocês, nós só quêfica aqui os fim de semana. Vocêsfizeram isso aqui e isso é pra vocêsganharem o dinheirinho de vocês, quevocês não tem da onde tirar. Vocêsdeixarem de trabalhar na roça, nãotrabalhem mais na roça, quando vocês,se vocês chegarem a trabalhar na roça,vocês fazerem só um [ ], meia quadra,uma coisa assim, pra vocês terem umafarinhazinha fresquinha e tal. E nãocorte mais a mata, não corte mais amata. Mas aí deu, não deu praenriquecer, mas dava pro sujeito. Nósmelhoremo muito a situação com aentrada do [ ]. Quando era doença, nósnão tinha preocupação, era só falarcom eles, que se o médico tivesse aí,resolvia as vezes o problema, e se não,a gente já ia encaminhado, chegava lánão ia pra firma não, ia [ ] direto falarcom os médicos, por causa deles. Aí foiindo, foi indo, acho que não passou umano, eles diziam pra nós, vocês gente
cobram, porque vocês foi que trabalharam aqui, vocêscobram de quem encostar vocês cobram, o que deremvocês recebe, um real, ou um quilo disso ou o quilo deuma coisa, sim o que derem vocês recebem. Tá na áreade vocês. Mas tinha muito, tinha muito que não dava eranada, ah, hoje eu não dô deixa da outra vez e tal ecoisa.Tá, tá bom, não tem problema. Depois meu irmão,antes de terminar, aí veio uma bronca trazida do IBAMA.
(quando que foi isso?)-Não tô lembrado.(mas porque que o IBAMAveio pra cima de vocês?)-Porque o IBAMA ele, ele veio pra cá, porque aqui
tinha aqui pra dentro d'água do Tupé tinha um [ ], tem unsigarapé que tem pedra né? Aí quando chegou acomunidade pra cá, aí a comunidade não queria mais queo pessoal tirasse as pedras. Aí [neste espaço que elestiravam não paravam] aí eles foram chamar o IBAMA praver. Daí ele encasquetou, aí fez uma reunião aí do, dessacomunidade Assombrado, o lugar não era tão coisa, masera meio assombrado. Aí o IBAMA veio botaram eles prafora e depois veio fazer o, uma reunião com nós aí napraia. Eles falaram um bocado de coisa, sei que chegou aconclusão de que aqui dentro eles iam colocar umflutuante aqui no meio d'água, armado até os dentes. [ ]eu acho que ele calculou isso, foi, mas pra que se acomunidade até que não tinha muita gente, armado atéos dentes pra quê? Aqui não tinha bandido [ ], nemtraficante, nem essas coisas. Aqui tinha um bocado develhote, pai de família e criança, que não tinha nem aulanaquele tempo ainda.
(mas era o pessoal da comunidade que tirava pedra?)-Não, não. Não era não senhor.(mas então porque a bronca?)-Sabe porque era, eu calculo assim, era porque veio
também outras qualidades de gente. Aí depois, que veio,veio, vinha gaúcho, vinha, tinha colombiano, eu achoque eles tavam cismado de ter tráfico, que colocaramtudo aí pra dentro, eu acho que eles tavam calculandoque tivesse alguém plantando droga, essas coisas assim.Aí mandou o prefeito implantar um monte de [ ]. Eu achoque eles tavam cismado por isso. Aí se alguém plantoueste mal não dava aqui. Pois é [ ], daí [ ], que até uns dianóis tava plantando uma farinha aí, a nossa casa defarinha era mais ali, mais embaixo um pouco, onde temas mangueira, ai [ ] tinha uma roça [ ]. Como noisterminamo de fazer a farinha, era assim uma hora da
225
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

tarde mais ou menos, ai vinhemo pra cá [ ], vinhemo pracá e aí dá uma chuvinha, quando dá uma chuvinha [ ] nóisjá tinha feito muito beiju, o senhor conhece beiju? Pois éeu tinha feito [ ]. Ele ainda olhô e disse “é beiju, né?” Eudigo: “é”. Aí ele queria que eu dissesse onde que era umbarracão que tinha aqui em cima daquela terra. “Poxa, [] conheço não” [ ]. “Mas você não mora aqui?” Eu digo:“moro, mas eu moro aqui. Eu não sou, não, não, não sei oque, não sei o [que] desse barracão, eu, eu sinceridadeeu moro aqui, mas não sei se tem esse barracão, nuncanem ouvi falar. [ ] que colocaram aí pra dentro, que eunuca mais fui ai pra dentro, e é isso mesmo. [ ] É, mais dizque tem um barracão aqui dentro, que tem um pessoaltrabalhando num sei o que [ ]. Se tem, eu não vou dizerque [ ] tem ou não tem? Digo logo assim não, não tem,não sei se tem, é mal. Eu não, eu é que não sei”. [ ]ficamo conversando daí, conversava, aí ele [ ] “e obeiju?” Pra mulher. “Está aí, o senhor come?”, “claro,mas claro!” Aí ela pegou e deu beiju pra eles [ ]. Daíforam nas minha roça ali dentro [ ] tinha dado um bucadode plantação de café. Tava tudo [ ] numa sacolazinha.Arrancaram o café quase todo, jogaram. Acho que éporque [ ] daqui, né? [ ]. Não sei se eles pensavam queera droga, não sei não, não sei que diabo foi que elespensaram, pra ficar [ ].
(esse pessoal era do IBAMA?)-Não, não, era da polícia, era da polícia. Aí eles
ficaram com essa coisa. Depois [ ], depois chegou-se aSEDEMA. A SEDEMA foi quem [ ] botou nóis pro mato denovo. SEDEMAchego [ ], aí o pessoal que era nosso caíramfora [ ]. Caíram fora e aí liberou pro povo lá [ ]. o povo damata [ ].
(fazer roça)-Fazer roça.(Mas o movimento na praia nessa época já era desse
jeito?)-Já era bem desenvolvido, já. Aquela praia foi
crescendo. Mas só que essa praia dali, essa praia dali,aquela praia dali. era lá do [ ], na verdade era com oJaumir [ ], essa praia de lá [ ] pra dentro da vila, aquilo aliera, era nosso. Era nóis que mandava naquilo ali. Aquiloali ficava intrucheirado só de família.
(vocês tinham barraca ali?)-Tinha uma barraca.(e vendia o que lá na barraca?)Não vendia nada. Só fazia ao mandado deles.
(Ah, o pessoal chegava...)Era, tinha uma casa grande, [ ] uma
barraca grande assim e uma mesatambém, mas aquilo ali era, era, eramandado sempre por eles, né. Aquelecara ali [ ] da vida dele e tal e coisa.
(Esse pessoal do [distrito]?)-Esse pessoal de Manaus, esse
pessoal [ ] pois bem. Aí [ ], aí nóistentava fazer esse serviço aí nãoaparecia ninguém pra empatar.Quando o serviço tava quase feito [ ]SEDEMA, o IBAMA. Aí vinha lá umdoutor lá, um doutor outro, jacarémuito bravo. [ ] perguntando quem erao brabo: quem é o brabo daqui?
Depoimento de Jaumir Matias deMagalhães, 44 anos, funcionário daSEDEMA, 1º grau incompleto.
(Por favor, Jaumir conta pra gentecomo é que foi o histórico aqui daComunidade do São João, quando quecomeçou, se já tava aqui, se não tava,como que cê pegou isso aqui?)
-Eu cheguei aqui em 22 de outubrode [1986] e tinha um morador, né? Ládentro do Lago. Tinha um aqui naentrada, logo na praia, que era a sogrado meu sogro, né? Seu Glicério. E foibom. Minha chegada aqui graças àDeus eu tive um, ass im umassentamento bem, eu vim com umplano de trabalho, né? E o Tupé ele eradiferente do que é hoje. O Tupé eletinha uma visão, falando de meioambiente, de quando eu cheguei aqui oTupé tinha uma visão muito melhor. Eletinha um padrão de meio ambientemuito avançado. Só que naquela épocatambém o meio ambiente ele não tinhaexpansão pro interior assim, né?
A Comunidade era o povo todomesmo do interior
226
Rebêlo et al.

Quando se falava em meio ambientenaquela época era só IBAMA, essascoisas, né? Sobre fiscalização de caça epesca, essas coisas. Simplesmentefalavam em flora, né? Preservação dasmatas, essas coisas. E quando foi em[1994] se eu não me engano, foi quesurgiu a Secretaria Municipal de MeioAmbiente, SEDEMA, né? Que elacomeçou um trabalho de conscientiza-ção.
(Isso foi em 88, 89?)-Não a SEDEMAela já é de(Quem criou a SEDEMA foi o Artur
Neto)-Foi, o Artur Neto entrou em
noventa e ....(Na prefeitura ele entrou em 88 eu
acho)-Noventa e dois, ele entrou.(Noventa e dois?)-Foi.(Mas aí ele criou a SEDEMA por aí
logo depois?)-Foi, logo em seguida ele criou. Só
que falta de orientação, nóstrabalhamos muito com agriculturaaqui no Tupé, né? Foi quando começousurgir a Comunidade, em 97, em 87, eucheguei em 86, logo em seguida, em 87a Comunidade começou.
(A Comunidade São João foifundada em 87?)
-Em 87.(Com quantas famílias?)-Foi uma faixa de umas 20 famílias
quando ela começou.(20 famílias)-É, quando a gente começou, né?
Eles chegaram. Era um pessoal que jáera do Baixo Amazonas, já tavam naRedenção há vários anos, né? Edescobriram que essa área tava aquidesocupada e a necessidade daagricultura, né? O povo era todo dointerior, e eles vieram pra cá e a gente
começou a Comunidade. A gente se reuniu aqui ecomeçou. Só que nós não tinha uma formação ambiental.Então nós trabalhamos muito, nós desmatamos muitonesta época, né?Até que a SEDEMA...
(Nessa época o pessoal vivia de que aqui? Elesdesmataram isso aqui pra plantar roça?)
-Da agricultura, era plantio de roça, mandioca, né?(Nessa época a farinha é que era o forte)-Isso, exatamente.(Não tinha essas barracas na praia?)-Não, nessa época eu acho que eu era o único daqui
que não tinha roça. Eu cheguei a colocar na época atéuma roça, né? Eu desmatei uma capoeira na época, já profinal de 87 eu fiz uma rocinha pra mim, né? Porque o riosecou e eu fiquei sem [ ], quando eu cheguei no Tupé omeu trabalho já foi desmatar. Eu comecei limpar a praia,comecei a, o pessoal que vinha pra cá começou atrabalhar comigo e eu fui investindo, fui limpando apraia, fui fazendo. E entãoci tinha pouco tempo decuidar de roça, eu coloquei uma roça só. Depois eucomprei outra roça de outro rapaz. Quando foi notrabalho da SEDEMA, já mesmo severo assim, que ela játinha mesmo assentado, né? Entãoci eu parei de botarroça, mas o pessoal ficaram colocando roça. AComunidade era o povo todo mesmo do interior. Elesmoravam na cidade, mas era um povo que já era dointerior, né? Eles trabalhavam só com agricultura. Nóstivemos também exploração de pedra na época, aquidentro, né? Nós temos três igarapés dentro do Tupé, quetem pedra, de onde eles extraíam pedra. Antigamentetambém antes que eu chegar o pessoal extraiu pedra. Naépoca do seu Glicério, ele chegou mais ou menos em 58,né? O povo tiravam lenha pra chata, né? Chamavam lenhade tonelada. Intãoci eu não alcancei, quando eu chegueiaqui não tinha mais isso aí já, né? Até a pedra quando eucheguei, eles tinham parado de tirar. Agora depois, comdois anos depois da Comunidade fundada, o presidentena época ele liberou que o pessoal tirasse pedra. Só quedava muito turista aqui nessa época, muito turista.
(De quem foi a idéia de criar, de fundar umaComunidade?)
-Essa idéia veio de Manaus, foi desse pessoal quevieram de Manaus, tá? É uma família, que segundo que euconheço a história eles já têm três Comunidades grandesfundadas, né? Que eles fundam essas Comunidades.
(Eles tão aqui ainda?)
227
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

-Não, não. Tem apenas um que ainda tem terrenoaqui dentro, né? Mas ele tá em Manaus.
(Ele não participa?)-É, ele vem pouco, né? Porque ele já tá se colocando
lá pro [Autazes], ele já conseguiu outro terreno pra lá,mas eles moram na Redenção, né? Só que eles fundaramuma Comunidade lá no Mucambo, Baixo Amazonas, eunão conheço, Comunidade São João, eles fundaram aComunidade da Redenção, uma Comunidade São João láque é, né? Que foi fundada por eles também.
(Gostam de São João)-Gostam, eles são devotos a São João. E fundaram
essa aqui também São João, foi começada por eles, né?Seu Manoel Palheta, Seu Antônio Palheta, o seu [Chico]Palheta.
(Quem foi o primeiro presidente aqui daComunidade?)
-O primeiro presidente foi o Carlos Alberto que veiojunto com eles, na época, né? Um rapaz novo na época eele tirou dois mandatos como presidente. Quatro anos,né? Que é o primeiro e segundo mandato.
(CarlosAlberto de quê?)-Carlos Alberto Lobato. Foi um bom camarada, até
hoje um amigo nosso, né? Foi que fundou junto com aesposa dele.
(Ele ainda mora aqui?)-Não ele mora em Manaus, mora na Redenção. Ele é o
presidente da Comunidade Redenção lá, lá emRedenção. Na Comunidade São João de Redenção.
(Ai depois que ele saiu quem que entrou?)-Entrou o Raimundo Silva, que ficou no lugar dele, né?
Foi o segundo presidente. Raimundo Silva tirou mandatode dois anos e depois passou pra dona Erundina. AErundina tirou um ano de mandato, era dois anos omandato, né? Como é até agora, é dois anos de mandato,ela tirou um ano, passou pra o vice, que era o Seu Dedé,que era parente também do Seu Manoel Palheta, que agente chama ele de Teco apelido de casa, e o Seu Dedéterminou o mandato. Aí depois do término do mandatodo Seu Dedé, entrou o Nidoval, né? Que a gente chamaele de Doval também. Intãoce ele fez dois mandato. Eletirou o primeiro mandato de dois anos e o segundomandato ele tirou parece que oito meses, seis meses,oito meses ele tirou mandato, aí deu um pequenoproblema e ele teve que renunciar, né? Foi o quartopresidente. E na renuncia dele eu assumi como interino,
né? Por quatro meses. Aí nós fizemosuma pequena eleição e por motivo dedoença eu renunciei, passei apresidência pra pessoa que eu tinhacolocado como vice, né? E ele tirou omandato, terminou o mandato doDoval, fez o período dum mandato emum ano, da eleição que ele tinha feitona época, né? E com um ano eletambém teve que renunciar, teve quesair, Ele não mora mais aqui, é o Roque.Parece que é Raimundo Roque - Régis onome dele. Foi um bom presidentetambém, ele trabalhou, fez a gentenão fazer certas coisas, porque aComunidade inteira também eladepende, né? Ela tem que ajudar e àsvezes quando a Comunidade não ajudao presidente não pode nada, né? Com asaída, com a renúncia do Roque, aí eutornei a assumir como interino, eu tireium mandato de um ano dele, o restoque faltava o mandato dele, comointerino. Nós fizemos a eleição, aondeeu disputei novamente a presidênciajunto com o Doval e eu ganheinovamente a eleição, né? E completeium ano agora dia 21 [21.04.03].
(Desse mandato agora?)-É, desse mandato que eu tô.
Completei um ano, só tinha um ano.Intonce foi mais ou menos isso.
(E nesse tempo a Comunidadeaumentou de tamanho, diminuiu, oquê você acha?)
-A Comunidade ela diminuiu. Eladiminuiu porque cada uma, a nossaComunidade ela não tinha um local,um centro, um local, como nós temoshoje, né? Hoje nós temos um campo,temos uma escola, temos um posto desaúde, temos a Sede, que é o CentroSocial, nós temos uma faixa de, entretudo, 16 casas, ou é 18 casas.
(De quando que é essa Vila? EssaVila aqui de dentro da Comunidade? De
228
Rebêlo et al.

quando que é? Por que quandofundaram não existia?)
-Não, não. Quando nós fundamos aComunidade, o Lago era, foi até hojeela é assim, ela faz parte, todo Lago[Lago Tupé] faz parte da Comunidade,agora nós colocamos ta com uns 6 anosmais ou menos. Desde então tá commais de sete anos, uns sete anos. Nósindenizamos, que isso aí era o terrenode uma pessoa, né?
(De quem que era esse terreno?)-Esse era do primeiro presidente,
esse terreno. CarlosAlberto.(Vocês compraram dele?)-A Comunidade se reuniu,
conseguiu o dinheiro e indenizou abenfeitoria dele, que ele tinha umacasinha, tinha umas plantas, né? E agente, aí a Comunidade indenizou, foina época do Seu Dedé, na presidênciado Seu Dedé, né?
(E aí distribuíram os lotes?)-Os lotes, exatamente. Pra fazer a
C o m u n i d a d e , o C e n t r o d aComunidade. Agora temos a escola,nós temos o posto de saúde, o postocomunitário de saúde, né? Nós temos ocentro social, então nós faz atéreuniões, faz a festa da Comunidade.
(Que é ali atrás da escola?)-Isso, aquela casa ali.(Quando foi construído o templo da
Assembléia de Deus e a Igreja de SãoJoão? Elas são mais ou menos damesma época, como é que é?)
-Não. A Igreja Católica ela é umpouco mais velha construída aí.
(Ela é desde a época que foi criadaa Vila?)
-Isso, exatamente. Eu acho que foiuma das primeiras casas que foramconstruídas aí, logo depois que a gentefez uma escolinha pequenininha, né?Foi a primeira casa que foi feita ali, foiuma escolinha pequenininha. Logo em
seguida a Comunidade se reuniu, né? Porque de religiõessão duas igrejas, né? Assembléia e a católica. Acomunidade católica, a Igreja católica se reuniu econstruiu a Igreja. A nossa igreja ela era num terreno ládentro no final do Lago, né? Lá na boca do Igarapé daCachoeira. Lá que construíram nossa Igreja. Mas aí agente viu que melhor seria na comunidade, no centro daComunidade, né? Nós reunimos a Igreja Evangélica, né? Enós construímos o templo daAssembléia de Deus.
(Quando que foi isso?)-Olha, eu não tô assim, pra dizer pra ti, foi no dia tal,
no ano tal.(Mais ou menos)-Tá com uns quatro anos agora. Quatro anos que foi
construída essa igreja.(Como que funciona? Tem pastor?)-Tem, nós temos pastor. Só que o pastor ele não fica
aqui exatamente, porque o pastor ele toma conta, muitolonge, dá horas e horas aqui, é o [campo] que a gentechama, né? Da área que estende a Igreja Evangélica,então ele trabalha daqui até Manaus, ele tem uma áreaem Manaus também. Tarumã, tudinho, ele tem a Igrejaonde ele [campia]. Então cada uma Igreja dessa tem umapessoa responsável, que é o dirigente, né? Então nóstemos uma dirigente que mora aqui, aArtemis.
(Artemis que é dirigente?)É, a Artemis que é dirigente, né? E a gente se
congrega. Eu confesso também na Igreja, né. Eu vejo,prego, canto junto, né? E a gente se congrega junto.
(Quem que é o pastor?)-É o pastor Gilberto. Ele mora em Manaus. Eu não
lembro o sobrenome dele, to sem memória.(E o pessoal da Igreja Católica, também é
semelhante, não tem padre aqui, né?)-Não.(Mas tem um padre que visita aí).-É. Quando tem algum movimento, quando a igreja
pede a presença do padre, ele vem, né? Porque o padretambém dessa área ele tem uma área muito grande.
(Quem que é o padre dessa área?)-Padre Geraldo, mas eu também não lembro o
sobrenome dele. Padre Geraldo é uma homem muitolegal, gosto muito dele, né? Dou muito com ele. E sempreeles vêm aqui na Comunidade. Qualquer movimento quetem aqui eles vêm, né?
(Mas quem que fica responsável?)
229
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

-A Dona Jane, a mãe do [John], que é umcompanheiro de trabalho, meu colega de trabalho, né?Ela que é a dirigente aqui, a ministra da Igreja Católica.
(Agora, cê diz que quando começou tinha uma escolapequena.Aí depois fizeram essa escola grande)
-A escola primeira que tinha ela era comunitária, foinós que fizemos a escola pra começar Porque primeironós tinha [paradeiro].
Depoimento de Álvaro Oliveira Bastos, o Barú, 49anos.
-A história desse ramal, nós távamos em milnovecentos e acho que 94, um cidadão por nome Ciro elenos procurou, procurou a direção da Comunidade doJulião, né? E queria uma pessoa que informasse ondetinha uma área que não pertencia, que já tinhaconhecimento que essa área onde nós estamos aqui, né?Pertencia à União. Ele já tinha tido um problema aí pracá. Na margem do Acácio tem um terreno do SeuSebastião [Monterusco]. É um título. Então ele tavatrabalhando dentro dessa área aí e foi embargado, tevebriga com eles lá e ele foi tirado de lá.
(Mas o quê que ele tava fazendo lá?)-Ele tava com uns projetos de uma cooperativa, né?
Então ele foi embargado.(Uma cooperativa pra loteamento?)-Loteamento da forma como tá aqui. Isso aqui tudo
foi loteado, né? E assim ele tava fazendo lá. Mas como elenão teve acesso nenhum lá, né? Ele procurou aComunidade do Julião e nessa época o Delegado Sindicalera outra pessoa, né? Era o próprio presidente dacomunidade lá da Associação. Aí ele informou que sabiaque eu conhecia muito aqui, né?Aí foi que dia 24 de julhode 1994 eles deram entrada nesse ramal lá. Tinha umtopógrafo que pagava a gente pra fazer esse trabalho aí,né?
(Julho de 94 foi época de Copa do Mundo, né?)-Foi nessa época mesmo e o pessoal trabalhava e no
dia do jogo eles corriam pra lá pra comunidade e a genteia apreciar tudo junto.
HISTÓRIADACOLÔNIACENTRAL
Aabertura do Ramal: era mata fechada mesmo, nãotinha trabalho de nada
(Lá no Julião?)-É. Então eu não queria trabalhar
com esse homem, porque eu nuncatinha trabalhado assim. Ele veio umasquatro vezes em casa e disse que mepagava bem. Até que eu conversei como Delegado Sindical e ele disse: rapazvai trabalhar, porque o homem é bom epaga e ele precisa de ti.
(Aí abriram essa picada?)-Aí foi que nós abrimos essa picada
aqui.(De uma ponta a outra?)-Até o fim lá. Mas o ideal era
atravessar oAcácio.(A idéia era atravessar oAcácio?)-Porque teve essa sobra aqui até o
Acácio é aberto, mas aqui pra dentro.Então essa direção aqui vai atravessaraqui (mostrando o mapa).
-(E por que não chegou lá?)-Porque quando chegou bem aqui
denunciaram ele.(Quem que denunciou ele?)-O pessoal mesmo, que tavam com
ele aqui. Tavam sendo enganado. Tavacobrando uma taxa.
(Quanto que era?)-Na época eu não tô lembrado. [ ]
Aí ele abandonou, a Polícia Federalparece que andou atrás dele. Inclusiveele ficou me devendo.
(Ah, ele ainda ficou te devendo? Elesaiu daqui preso?)
-Ele se escondeu, mas não pegaramele não, se escondeu. [ ] Inclusive elefez negócio comigo com uma moto-serra nova, né? Fez umas cinco moto-serra, era pra uma madeireira, eledesviou esses motor pra cá. Pegou umterreno lá onde eu tô agora morando,ele teve serrando lá, ele devastou amadeira quase toda do terreno, aí foi aépoca que denunciaram ele e elesumiu, né? Quando eu tava lá numaboa, né? [ ] derrubar 20 hectares de
230
Rebêlo et al.

mato pra gente fazer plantio de cana.Aí conforme ele não veio mais aí jáquem apareceu lá na minha casa foi umagente da polícia civil, pra pegar omotor, né? Mandou deixar uma ordem,eu não tava em casa nesse dia, né? Tavana cidade. Levou o motor e deixou aordem pra eu comparecer lá nadelegacia em São Lázaro.
(E ainda sobrou pra você?)-Foi. Aí eu fui lá, esclareci que eu
tinha ido trabalhar com ele aí, né? E eupedi uma garantia de ficar no terrenolá enquanto ele não me pagasse. Nóssomamos naquela época o trabalhotodo que eu tinha na mão dele,trabalho aqui, trabalho de lá, dava3.770 reais. Aí fui na delegacia,registrei a queixa, pedi autorizaçãopra mim ficar lá no terreno. Aí odelegado disse que não eraaconselhável, né? Mas que garantissepra ficar até na época que ele chegassepra me dar o pagamento e eu sair dolocal. [ ] delegado é o seguinte: eu nãotô precisando de mais terra não. Eutenho outro terreno que era bemextremando com este. Eu só quero irpra lá por causa do meu trabalho. Eesse homem até hoje não apareceu.
(Nunca mais? Já vai fazer 10 anos?)-Dez anos. Dois anos depois
apareceu a esposa dele lá querendome tapear com 500 reais, né?
(Quem que é a esposa dele?)-Era uma senhora por nome Chagas.
Falei pra ela que eu não tinha negócionão com ela. O delegado me orientouque eu não fizesse negócio nenhumcom ele aqui, se ele aparecesse era pranós ir lá na delegacia pra genteresolver isso por lá. Chegou com umcasal pra ficar no terreno e eu disse praela: “cê paga meu trabalho aítodinho”, depois disso eu derrubeiquatro hectares, quando ela chegou já
tinha plantado tudinho, a minha roça tava dessa altura,por igual. E o rapaz foi lá com meu irmão, “olha lá na casalá. Quem fez esse trabalho aí, meu irmão? Rapaz, opessoal não tem mais direito de sair fora daqui”. Eledisse: “não tem enquanto eles não vierem pagar”,porque eu não fazia questão daquela terra não. Faziaquestão do meu trabalho, né? E até hoje se ele chegar eme pagar, se der pra pagar meu trabalho, as despensaminha lá em cima, né? Eu saio, mas não apareceu. Bom,ela queria me dar 500 reais, eu disse: “não posso ficar. Etem outra coisa: eu não fiz negócio com a senhora. Quemme procurou foi seu marido. E diga pra ele que eu vouesperar ele aqui da mesma forma que ele me procurou,eu recebo ele. Não vou receber ele com briga não”. E eladisse que na outra semana ele vinha falar comigo. Foi em1997, dezembro de 97. Desde então ele não apareceumais. Eu ouvi falar que um dia ele passou lá por SãoSebastião, aqui na Comunidade próxima aoassentamento do [INCRA]. Aí falou com um senhor queele tinha um terreno aí, inclusive ele ia fazer procuração.Tô esperando. Se chegar e me pagar eu saio.
(Baru me diz uma coisa, e quanto tempo vocêslevaram pra fazer esse ramal?)
-Nós passamos quase um mês.(Um mês?)-Trabalhamos desde agosto, o resto de julho e agosto
até o dia 22 de agosto eu trabalhei.(E como é que era aqui essa floresta?)-Era mata fechada mesmo, não tinha trabalho de
nada. Mata bruta mesmo.(Vocês tiraram muita madeira aí legal? O quê que
tinha de madeira?)-Não. Madeira não. Não tem condição de tirar
madeira daqui. Não tem condição de tirar madeira daqui.(Foi só abrir picada?)-Só a picada.(Tinha muita caça?)-Sempre tinha, né? Eu sempre gostava de caçar por
aqui. E ainda tem alguma.(E os primeiros moradores foram chegar quando?)-Quando eles denunciaram ele, já existia alguns
daqui, né? Já tavam derrubando roçado. Logo quecomeçou, né? E eu fiquei morando lá fora e alguém quequeria ver algum terreno eles vinham primeiro comigo.Aícom o tempo eu passei a ser o Delegado Sindical lá doJulião aí eu trabalhei pra esse pessoal tudinho aqui.
231
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

Chegavam perguntavam. Já sabiam que tinha esseloteamento aqui, né? [ ] eu mostrava, mas nunca cobreinada.Aúnica coisa que eles faziam me dar era a diária dotrabalho de ir lá mostrar pra eles, só. Pode perguntar aípra qualquer um morador se algum dia eu cobrei algumacoisa. Até eles se admiravam, porque tem muito lugar aíque os camaradas só faz se for por dinheiro. Taí. Osprimeiros mesmo que vieram, um monte desistiram.
(Quem que foram os mais veteranos aí?)-Os mais veteranos têm o Seu Adison, Seu Raimundo,
DonaAna,Ana Paes Batista.(O Raimundo ta aí ainda, né?)-Tá, o Luiz.(OAdison também?)-Tá, tá o Luiz, seu Paulino, seu José Barbosa, esses
foram os primeiros que ficaram no [retiro]. Tinha o outroo Zé Pascoal, mas o Pascoal ele fez um sítio bonito aí,mas ele não mora mais aí. Tá lá pra Macapá. Depois dissofoi entrando esses outros.
(Nessa época tú que era o Delegado Sindical aqui)-É.(Até lá na ponta?)-Até na ponta. E foi o tempo que houve uma, de uma
gerar a diretoria da Comunidade, né? A Associação aquidentro.
(Isso foi quando?)-Noventa e oito.(Noventa e oito?)-Criada a associação.(Aí os índios já tavam aí?)-Já.(O Domingos, o Raimundo? Foi tu que chamou ele?)-Não, eles vieram até mim, né? E só vim mostrar a
área pra eles aí. Eles vieram através de um rapazchamado Amaral, né? Pra vim pra cá. [ ] Aí eu mostrei aárea aí, né? Era pra trabalhar aí. Inclusive o Domingos elefoi o primeiro presidente da Associação. E agora tá namão do Silvestre. [Que] foi vice do Domingos.
(Mas e o Delegado Sindical agora é o Manoel, né?)-Daqui [Colônia Central] é.(Desde essa época? 98?)-Foi. Nessa época aí.(Aí depois deles é que chegou essa outra turma?
Arlindo, Joel?)Não, o Joel ele entrou aqui em 97. Nessa época o
único trabalho desse que eu peguei aqui foi dois
trabalhos. Só tinha eu que trabalhavacom moto-serra aí. Vinha pra cá, sófazia roçar o mato [ ]. Mas aí já veio unsque pegaram o terreno e não seacostumaram, saíram fora daí. [ ]Porque tem dificuldade em colocarproduto pra beira, né? Pode ver que atéa madeira mesmo ninguém se [atreve]a tirar porque: carregar como? Essaárvore aí isso é Angelim, é bom pramóvel. Se derrubar fazer prancha aídava cinco a seis metros de prancha.Não pode, não tem nem comoaproveitar.
(Aí depois fizeram essa sede?)-Depois fizeram essa sede aqui. [ ]
Foi em [1999]. Então essa é a história,nossa história do Tarumã-Miri, ColôniaCentral, Tupé, Tatu. Isso aí eu conheçode noite e de dia nessa mata aí.
Depoimento de Domingos VelozoVaz, anos, índio Dessano.
(Você foi o fundador aqui daColônia Central, né?)
-Fui.(Como que foi isso aí?)-Eu fui um fundador daqui da
Colônia Central porque muitos deles,muitos daqui do, do, da ColôniaCentral, eles [ ] já estavam antes,como eu estava dizendo ontem, játinham dez famílias aqui, mas só queeles não eram fundadores, [ ] e eleseram simplesmente [ ], eles eraminvasores.
(Não tinha umaAssociação?)-Não tinha nenhuma Associação,
simplesmente eles eram uma família,mas [ ] na prática eles eram unsinvasores. Aí quando eu cheguei, aí vique não dava certo aí eu, fundei.
(Mas já tinha esse ramal?)
Anascente da água
232
Rebêlo et al.

-Já tinha esse ramal, há muitotempo.
(Eles invadiram o ramal, é isso?)-Eles invadiram o ramal. Não, só
que eles vieram através de um [ ], deum político [ ], que era agente dogoverno, ele era agente do governo, aísaiu expulso [ ] dos trabalhos dele [ ] eele visou, porque essa área era daUnião [ ]. É, aí, então, o quê que elefez, ele saindo do governo, dostrabalhos do governo, ele teve quefazer uma Associação aqui chamada,ele fundou também uma AssociaçãoPaz e Bem, chamada Paz e Bem.Histórico dela ela tem também aqui. Éoutra história. É outra Associaçãotambém que ele fundou antes de mim.Ele fundou com uma Associaçãochamada Paz e Bem. Aí o quê que elefez?
(Quem que era esse político aí?)-Ele era um, ele era um, um
[geólogo], ou algo da, do governomesmo, eu não sei quem ele eratambém, mas o pessoal não sabeexplicar também, porque [ ], eu sounovato aqui eu não sei como, eles nãosabem se explicar também. Eu nãocheguei a conhecer. Então eles dizemque esse, esse senhor, ele, ele que feza, os limites tudinho desse ramal.Treze quilômetros medido tudinho [ ],fotografia, tudo, tudo, tudo, tudo eleque fez esse aí. Então, depois que elefez isso [ ], aí o governo botou fora ele,então o quê que ele fez? Praaproveitar, ele fundou uma Associaçãoclandestina chamada Paz e Bem. Aítodo mundo caiu, né? Hoje em dia éassim, né? [risos].Ah, o governo vai darisso, não sei o quê, ta, ta, ta, ta equalquer um pode inganar, né? E assimque ele usou o pessoal, então o pessoalde Manaus, esses recém chegados de,de Maranhão, Pará, entendeu? Vieram
pra cá. Ele cobrou taxa de 50 reais [ ] pra cada lote e olhaque tem 56 lotes aqui, [ ] 50 reais. Aí encheu. Na, narealidade não era pra agricultura, ele estava visando praextrair madeira aqui dentro. Era pra extrair madeira, elejá vinha extraindo lá do Julião pra cá, que aqui tem muitamadeira, de lei. Então, a intenção dele era essa.
(Que madeira que ele tirava daqui?)-Ele tirava de tudo.(Tudo?)-Tudo, tudo, tudo, tudo. Só deixava mesmo a madeira
fina. E vinha acabando. Aí, descobriram que a Associaçãoera clandestina. Aí ele foi varrido pela Polícia Federal,sumiu.Aí todos que tinham ocupado por aqui, no ramal [ ]vendo essa [ ], essa visão aí na frente, né? Aí, seafastaram, foram embora, porque era difícil, era tudomato. Não tinha nada de concretização aqui dentro, elesforam embora. Foram embora e ele já tinha ganho 50reais de cada lote. Fora a [madeira] que ele levou. Aí,então, restaram dez famílias aqui. Ficaram aqui dentro.Restaram dez famílias, ficaram permanentes, calados.Aío quê que eles fizeram? Derrubaram, como a lei [ ]comotodo mundo faz, né?Aí começaram fazer roçado.
(alguma dessas dez famílias ainda ta aí hoje?)-Tão aqui. Ce vai conhecer, quando vocês forem, da
próxima vez vocês vão conhecer eles. E eles, lá docomeço.
(Nesse trecho aqui, não?)-Não, não, daqui não. Aqui é tudo depois já. Eles
entraram depois que eles já tavam com quatro anos.Entraram já quase igual como a gente. Já vão fazerquatro anos eles. Mas só que pra cá funcionou rapidinho[risos]. O pessoal fizeram logo, ta, ta, ta, tá bonito pralá, né? Mas só que, uma hora dessa até que ta bonito, masmeio dia, uma hora é [ ]. Aí, então, o quê que aconteceu?Aí eles fizeram, ficaram lá: dez famílias. Trabalhando,cultivando: maniva, cupuaçu e o que tinha que fazer, né?Aí nessas alturas eu cheguei em 98 [ ], 98 eu cheguei. Evim de [ ] da Associação ACIRU- da Associação dasComunidades Indígenas do Rio Umari, lá dos Distritos dePari Cachoeira, eu era presidente dessa Associação,fundada por mim mesmo, eu vim dá uma volta aqui, eletava já em Cravoeiro, ele tava em Cravoeiro aqui noMunicípio de Barcelos. Eu vim é dá uma volta, dá uma [ ]com ele, conhecer como é que era, aí vim tudinho comtoda minha família. Cheguei em São Gabriel, de lá passeipra Cravoeiro. Pra mim retornar foi uma luta. É um
233
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

dinheiro alto já, né? Tanto pra mim retornar com minhafamília. Aí não consegui mais retornar pra lá. Acabeificando em Barcelos, em Cravoeiro, município deBarcelos. Aí acabei ficando com [ ]um ano, aí matriculei,os meus filhos não prejudicaram nada. Era tempo dematrícula ainda, aí matriculei logo os meus filhos. Só queo primeiro já tava em Manaus, já tava fazendo sexta?Sétima? Sexta [ ], já tava fazendo. Aí os pequeno eumatriculei lá e agüentei um ano, aí não pude maisretornar pra Pari Cachoeira. Aí o quê que acontece? Aínessas alturas, aí a minha irmã, a nossa irmã que tá aqui,a Éster, ela pegou, ela tinha amigos delas, né, queestudavam junto com ela, acadêmico, né, ela estudavaaqui em Manaus e aí [ ], nosso vizinho do outro lado,chamava Amaral. Ele dizia pra ela, poxa Ester, lá tem unsterrenos bonitos, ce não quer pegar, não? Ela dizia não.Dois anos, três anos relevando. Aí certo momento ela viuque [ ], ele já levava já cará [ ], bonito. Ele já levava já daroça dele. A roça dele era bem aqui aonde tá a sede.Então ele plantava cará e ele levou dois cará pra ela [ ],cará bonito. Aí ela se animou, aí ela [ ] veio também comele [ ] sozinha. Veio pra cá, chegou aqui, ela ficouolhando [ ]. Aí o pessoal tinha pulado esse aqui. Daqui doseu Vicente pra lá, os terreno já vão embora, né? Agoraesse aqui eles não pegaram porque os terrenos erampequenos e o pessoal aqui é, quem veio lá de fora eles sópensam em área grande, né? Muita terra. Aí, fecharamessa parte, né? Essa parte era limpa, por que? Só tinhaaquela partezinha na beira do ramal e pra cá é só igapó,né? Ninguém podia trabalhar [ ].Aí o quê que ela fez, ela,como ela era índia [ ] ela visou logo na água, ela não visouno terreno não. Ela visou na água onde tinha nascente daágua, ela visou nesse sentido, ela pegou esse terrenoaqui. Pegou esse terreno, ficou com um ano parado.Pegou esse terreno demarcou a área, né? Aí, com um anoela convidou ele [Raimundo], que ele tava em[Carvoeiro]. Eu já tava aí também. Ela convidou ele pravim derrubar essa área aqui onde nós estamos agora. Praderrubar uma quadra, a roça dela, que ela tinha pego umterreno no município de Manaus, ninguém sabia ondeera. Só ele que veio com o filho dele [ ], né? Com oReginaldo. Ela já tinha brocado aí. Ela já tinha brocado eele veio derrubar [ ]essa área aqui. Ele veio derrubou,voltou [ ] pra Cravoeiro.Aí, quando foi pra queima, eu fuiconvocado. Eu tinha dito pra ele, vá, agora você e [ ] vãopra lá. Ele disse, não, é muito longe pra lá, não dá pra
gente ir lá, muito longe. Ele não quis ir,aí eu digo: tudo bem, eu vou. Aí eu vimcom ele. [Comentário do Raimundo] Aíentão, o quê que eu fiz? Aí eu vim como filho dele, ai com o filho dele eu vim,né? Ai eu vim, não, o filho dele tavaaqui em Manaus. Aí eu vim de lá e eutrouxe ele e ele me trouxe, eu nãoconhecia onde era. Aí nós viemos aquie tocamos fogo no mês de setembro de1998. Nós tocamos fogo aqui.
(Tinham quantas quadras já?)-Uma quadra só [ ]. Essa quadra que
ela tinha derrubado, porque prá lá eratudo igapó, né? E tem lá em cimatambém. Mas o importante é a água,né? A nascente da água. Aí então o queaconteceu? Aí nós queimamos, aí fuiembora, voltei pra, regressei praCravoeiro, aí voltei [ ], aí falei: tudobem, tá. Já que já iniciamos entãovamos lá. Aí eu peguei 12 feixe demaniva de lá, [ ] me mandei de novopra lá. Essa maniva que nós temos aquié de lá, de Cravoeiro, por isso quenossa farinha é bem amarelinha.
(É diferente mesmo)-Diferente. Nós trouxemos 12
feixes, nós carregamos lá do Julião,pra trazer até aqui.
(Vocês vieram por lá?)-É, porque não tinha nem acesso
por aqui ainda. Aqui não tinha acessoainda. Aqui não tinha nem acessoainda, aqui era tudo igapó, aquininguém [ ], ninguém sabia onde era, oacesso era tudo pelo Julião ainda, umramalzinho assim. Tudo bem. Aí nóscarregamos, plantamos, aí ficamos. Aía minha irmã disse pra mim: olha,poxa, já que nós estamos começando,então vamos fazer o seguinte: traga asua família [ ], bota seus filhos praestudar aqui em Manaus e você fica láno sítio. Vamos cuidar do nossotrabalho. Tudo bem. Aí mudei de vez.
234
Rebêlo et al.

Em vez de voltar lá pra [ ] eu vim praManaus. [ ] Bom, aí eu, só eupensando, né? O que será que vai darno final, né? (risos). Aí no sentido depensar eu tava sendo [nômade] jásabe? Eu, degrau, degrau, descendoaté encostar aqui em Manaus.
(Chegando como é o nome?Barreira?)
-Santa Barra. E aconteceu, tudobem, aí eu trouxe a família e [ficamos]lá. Viemos aqui, matriculamos e então[ ] começaram estudar, né? Tudo bem.Aí, nós passamos pra cá [fazê lá] osítio, a roça, né? Um ano que nóspegamos muita malária 1999 e 2000.
(Tinha muita malária aqui?)-Aqui era [ ] infestada de malária. A
gente passava 15 dias, com 15 dias agente já tinha que ir lá. E não tinhanem agente de endemias aqui, nãotinha nada. Nem aqui no Tupé nãotinha. [ ] E passaram, dois anospassamos ida e volta, ida e volta, porisso que nós atrasamos tanto. Nóséramos só nós dois eu e aquele um. Nasférias eles vinham também aqui, elesvinham passavam com a gente 15 dias,com 15 dias todo mundo pegavamalária. Todo mundo voltava demalária, pra lá de novo, lá no HospitalTropical. Chega dava pena. E graças àDeus hoje em dia não tem. Foicombatido [ ] e o posto taí também, aArtemis e aqui nós temos hoje tambémagente de epidemia aqui dentro.
(Aqui, na comunidade aqui comvocês?)
-Uma é essa aí, a Terezinha minhaesposa, o outro éAmaral. Fizemo nesseponto, aí eu fiz tudinho isso aí. Era pramim ter feito mais coisa, mas só que eunão pude fazer, naquele tempo, no queeu disse ontem, aí ficou difícil. Pois é.Aí então [ ], nós ficamos aqui [ ], masporque não deu, não deu pra gente
levar avante, devido que não dava pra gente levar deacordo como a gente queria. Aí ficou difícil. A gentequeria trabalhar [ ], fazer o nosso trabalho, mas só queficava difícil. Aí foi que eu fiz, eu um dia reuni o pessoallá, né? Com esse pessoal lá da frente, ai eu falei comeles, “olha, pôxa, que tal a gente fazer uma Associação,fundar uma Associação tal, são muitos lotes. Quem sabe,de repente a gente se reune umas 300 pessoas aqui, dápra fazer uma comunidade bem grande aqui. Bóra fazerisso aí? Eu tô acostumado, eu sei como fazer, eu conheçoesse ramo aí”. Eles disseram: “pôxa, então bóra fazer”.Todo mundo animado, né?
(Foi quando?)-[1998]. No ano que eu cheguei, 98. Aí o quê que nós
fizemos, tudo bem, então bóra começar. Aí nós tivemosque arcar do nosso bolso com [200] reais pra podercomeçar. Aí o quê que nós fizemos, nós começamos pelaDelegacia Sindical, lá em Julião. Elegemos o presidenteda Delegacia Sindical, o seu [ ] Adson, um dos primeirosque chegou aqui já tão fazendo sete anos, oito anos. Eleficou eleito como delegado sindical da Colônia Central. Eesse nome da Colônia Central já vinha com esse senhorque tinha feito com Paz e Bem, a Associação Paz e Bem.Já vem desse nome, da Colônia Central, entendeu? Aí jáveio. Então foi essa forma que eu tive que, tive queregistrar nesse sentido também, da Colônia Central e nãolargava mais da [pauta], aí ficou desse jeito.
(Como que ficou chamando aAssociação?)-A Associação ficou chamada como: Associação
Comunitária Agrícola [ ] Rural da Comunidade ColôniaCentral, ACAR. Foi registrado dessa forma. Teve quelutar muito, eu lutei muito pra essas coisas, pra poderregistrar e fazer tudo, tudo, tudo, [ ] todas essas coisas.
(Registrar o estatuto, fazerAssembléia)-Assembléia. Isso me custou, é! Deu trabalho, graças
à Deus terminei, tá na mão deles agora, né? Mas porque [], aí depois, logo no início de 2000 [ ], nós fizemos aprimeira Assembléia da Associação e elegemos umaDiretoria e fundamos uma Associação que é, que agoraque é Associação Comunitária Agrícola Rural daComunidade Colônia Central. Fundamos a Associaçãocom estatuto, tudinho [ ] e hoje em dia ela é [ ] tá bemregistrada, graças à Deus, se eles souberem levar, elesvão levar. Depois entrego tudo na mão deles. O[argumento] deles tão tudo na mão deles. Registramos,registrei [ ] e fundei essa Associação e que da qual hoje
235
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

em dia está desse jeito e esperamos, eu só to esperando,olhando pra eles o quê que eles vão fazer.
(Quando foi fundada a Associação tinha quantossócios? Quantas famílias eram?)
-Tinha 25, 25 sócios. Nós éramos 25 sócios, nãofamílias, [ ] era só do lote, um sócio de cada lote, nãofamília. Se fosse família era mais gente ainda.
(E quantos eram residentes aqui?)-Residentes só eram: uma, duas [ ], três, quatro
famílias só. E aqui nós éramos só nós dois: eu e minhaesposa. Minha irmã [Éster] morava lá em Manaus. Entãotá: aí começou, aí tudo bem, aí comecei trabalhardurante três anos: 2000, 2001, [ ] 2002, que acabouagora.
(Quem que foi o primeiro presidente aí daAssociação?)
-Foi eu. [ ] E é duro [risos] a gente conviver com umaspessoas que nunca estiveram, que nunca mais tiveramcontato com uma comunidade. Eu acho, né? Aí ficadifícil. Então três anos nesse sentido aí, durante trêsanos foi isso, não era pra gente tá desse jeito, não. Erapra gente ter nossas casinhas também, na beira doramal, tudo bonitinho, como ali no [ ]. O seu Arlindo elenão tem mais problema, chegou lá, taí, tão trabalhandoo lote. Que não tinha compromisso com nada, né? Agoraeu não, teve três anos.
(O seuArlindo é dessa época também?)-Não, ele vai fazer dois anos agora. Então tá. Aí vimos
que o ano passado, nós vimos que isso não adiantava,tentamos fazer um, tentamos fazer uma sede ali, masmorreu. Não adiantava. Quando era dia de comunidadeaparecia um, dois.
(Aquela sede tem pouco uso, tá abandonada, como éque é?)
-Pelo que eu tô vendo agora tá abandonada. Aí então,visando nesse ponto aí, eu me larguei, larguei o mandatoentreguei os papéis tudinho. Pulei fora. Porque eu nãovou perder tempo, eu não gosto de perder tempo comessas coisas aí. Não adianta a gente quebrar a cabeça,não adianta a gente esquentar a cabeça com aquilo quenão vai, que você ta vendo que não vai certo, você nãovai continuar aquele trabalho. Visando nesse ponto eupulei fora.
(Quando que foi isso?)-Foi dia seis de janeiro 2002. Aí eu entreguei tudinho,
prá eles continuarem no começo logo do ano. Eu
entreguei tudinho e fui embora. E nãodeu certo também porque, comopresidente eu era indígena, entãomuitos diziam: “pôxa um índio nãosabe, nunca trabalhou nesse negócioaí, não sabe, ele não sabe o mundobranco. Ele não sabe convivência domundo dos brancos, ele não sabe, nãosabe nada. Nós brancos, nós sabemos”.Eu via, mas não dizia nada. Então issome, entendeu? Me dizia [ ], então eu saíe [ ], então nesse ponto eu larguei. Vi epercebi que aquilo não dava certo pramim. Então pra eles, era só eles e tudobem. Eu aceito. Então, como eu eraíndio, eu poderia trabalhar só no meurumo também. Tá bom. Visando nesseponto aí largamos, entregamos e saí,do jeito que tá. Do jeito que nósentregamos, entregamos tudodireitinho, bem registradinho comdinheiro ainda. Agora que eles não[amolem] lá em casa, isso é problemadeles.
(Você entregou pra quem? Quem éque assumiu a presidência?)
-Taí, o Augustinho. Só que ele morado lado da Comunidade Julião, ele nãopertence aqui, ele não é nem sócio.
(E como é que fica? Quem não ésócio pode ser presidente?)
-Isso coube do pessoal daqui doramal. Eu não sei porque eleselegeram, eu não estive quando eleselegeram. Só estive no dia da [posse]Eu falei na frente dele, eu falei pra ele:“olha, prá você fazer funcionar essaAssociação, você tem que ser sócio emprimeiro lugar, em segundo lugar vocêtem que mudar sua casa e tem quefazer sua casa aqui dentro da sede.Dessa sede pra você puder funcionaressa Associação, caso contrário daí vaimorrer”. Na frente dele, na frentedele. Só assim que vai funcionar, casocontrário nem pensar. Eu que era,
236
Rebêlo et al.

morava aqui eu fazia meus trabalho,não funcionou. Agora que ele mora láno Julião, quando é que ele vai vimvisitar aqui? Quem é que vai capinar aí?Daqui a pouco taí na capoeira [ ],dentro da capoeira. É que custô caroaquela casa.
(Deve ter dado trabalho pra fazeraquela casa)
-Então, aconteceu isso. Então é,terminando isso, nós fizemos,entregamos, aí o que nós fizemos? Nósvisamos nesse sentido. Bom, já que nóssomos indígenas, pra nós tanto faz. Pranós tanto faz, como a gente está, comoa gente é, o que nós somos. [ ] Aí eufalei com a minha turma, né? Com aminha irmã [ ]: “bóra começar tudo denovo” (risos). Aí nós começamos denovo. Aí fizemos, preparamo estatuto,sô acostumado nisso mesmo, né? Praescrever é comigo mesmo (risos). Aí eupreparei estatuto, tudo que taí.
(Uma novaAssociação)-Nova associação, mas agora
indígena. Mil metros de frente, milmetros de fundo. E onde vai serreconhecido uma área ecológica [ ] eonde nós vamos implantar uma, umprojeto de ecoturismo. E essa área vaiser reconhecida pela FUNAI e outrosórgãos não-governamentais que nosapóiam, muita gente vai apareceraqui. Inclusive [vocês] vão fica aquitambém, o dia que nós terminar essacasa, nós vamos fazer a primeiraAssembléia aqui. Vamos convidar todomundo, vamos querer que todo mundoparticipe dessa primeiraAssembléia.
(E a Associação, então, ela não táfundada ainda, essa novaAssociação?)
-Por enquanto ainda não, está emprocesso. Tá em processo, tá emprocesso, mas estamos no final dejunho, maio, junho, creio que tá tudopronto.
(Como que vai chamar?)-Ela vai se chamar Associação Comunitária Indígena,
é, [Umokodiromatá].(Que significa?)-Que significa Associação Comunitária Indígena
Gente de Deus [ ] Desãno. Vai ser registrado dessaforma.
(Como que vai ser esse projeto do Ecoturismo?)-Olha, esse projeto Ecoturismo, nós estamos [ ], eu
como linha de frente do trabalho, eu estou pensando daseguinte forma: nós vamos trabalhar [ ] dois lados:comunidade indígena e a maloca indígena. Então o quenós vamos fazer? Dentro da maloca nós estamosquerendo montar uma escola de pajelança, onde [ ]porque uma pessoa não índio vão aprender as mitologias[ ] da nossa etnia. Inclusive o professor vai ser eu. Nósvamos fundar uma escola de pajelança, centro cultural ecentro de artesanato.
(Isso na maloca?)-Na maloca. Nesse sentido que nós vamos fazer isso.(E lá vai ter lugar pras pessoas ficarem?)-Tudo, tudo, tudo, [hotéis], nós vamos construir
hotel, pras pessoas que vêm de fora, pra passar um dia,dois dias, [ ] nós vamos construir tudinho. É um projetolongo, mas é, se a gente tiver força de vontade acho quecom certeza a gente consegue.
(Fora isso a comunidade vai continuar tocando seuroçado)
-Roçado, não como a gente tinha aquela visão deagricultor. Nós vamos ter que fazer só pro nossoconsumo, a agricultura vai ser só pro nosso consumo. Daía gente vai fazer um, como é que se diz? Reflorestá,depois da maniva a gente refloresta. E aí a gente vaivivendo assim. Prá nós a área é muito grande, [ ]. E prosturistas, agora eu vou ter que estudar e nós vamos terque estudar muito como que a gente vai fazer.Aí tem queentrar em contato com o Jaumir, com o Marcos Antônio esentar numa mesa, como é que a gente vai funcionar essenegócio. E começar lá, né? Porque tudo começou lá.
(Lá no Tupé)-Lá no Tupé. Nós fizemos uma reunião, que não era
nem pai dele, era o, como era? O [Roque], né? Era oRoque antes dele. Conversei com ele prá poder entraraqui, pra ajeitar o acesso e ficar prá cá.
(Desde quando tem esse acesso?)-Tem desde 2000.
237
Histórias: narrativas e depoimentos das comunidades São João do Tupé e Colônia Central

(Cês que abriram?)-Nós que abrimos.
Antes destes depoimentos a história da regiãoonde hoje é a RDS Tupé era desconhecida, dispersa entrelembranças e fatos ignorados que ocorreram nesta árearural do município de Manaus. O planejamento e omanejo da área dependem também de umacompreensão da sua história e de seus moradores. Osvisitantes da praia do Tupé desconhecem que ela foiconstruída por seus moradores, mas também ospesquisadores e planejadores desconheciam isto. Mesmoas pessoas que fizeram esta história desconheciam etalvez ainda desconhecem o termo “manejo depaisagem”, no entanto hoje o turismo recreativo é umadas principais fontes de renda dos moradores dacomunidade São João do Tupé. O ramal Central foiconstruído como uma aventura para ganhar dinheiro,uma origem que tem pouco a ver com os planos dosmoradores atuais, interessados em ampliar suas fontesde renda, escoar a produção e dotar a área deinfraestrutura de saúde e educação.
Esperamos que o mesmo tipo de registro sejafeito nas demais quatro comunidades que compõem aRDS Tupé (Julião, Agrovila, Sacra-mento e Tatu), parareunir histórias representativas de todas a área. Quemais pesquisadores recolham narrati-vas e maisinformantes se disponham a revirar suas memórias para
Considerações finais
deixar vir à tona as suas lembranças.
To d o o p e s s o a l d a scomunidades São João do Tupé eColônia Central, que nos receberam debraços e corações abertos, em especialos que nos contaram suas histórias.Edinéa Mascarenhas Dias forneceubibliografia histórica.
Martinello, P. 1988. A mobilização damão de obra nordestina para aAmazônia ou a epopéia do arigó.
1: 207-249.PROJETO BIOTUPÉ. 2004. Acesso em 23
d e m a i o d e 2 0 0 4 , d o :url:http://biotupe.inpa.gov.br.
Simões, M.S. C. Golder. 1995.Cejup; Universidade
Federal do Pará, Belém 191 pp.
Agradecimentos
Bibliografia Citada
Cadernos UFAC Estudos ePesquisas,
Belémconta...
.
&
238
Rebêlo et al.

Diversidade SocioculturalCapítulo 16
A visão vernacular e o desafio culturalpara construções em mutirão
RESUMO - Os processos tecno-construtivos baseados em conhecimentos populares podemviabilizar a preservação do patrimônio cultural contribuindo para o resgate da memória eda auto-estima de comunidades localizadas em regiões de desenvolvimento sustentável. Opresente trabalho propõe uma discussão das bases teóricas referentes à emergência epreservação de processos tecno-construtivos locais para a construção de novas áreasedificadas, propiciando, inclusive, trabalhos em mutirão, garantindo a possibilidade de umplanejamento sustentável para a RDS Tupé (Reserva de Desenvolvimento Sustentável doTupé), localizada no município de Manaus (AM), abrangendo uma área de 12.000ha,localizada a 25km do centro da cidade. Essa visão vernacular é considerada de relevanteinteresse à comunidade porque garante postura de equilíbrio entre a população local e omeio ambiente. A prática de reproduzir antigas técnicas construtivas nas atuaiscomunidades fortalece a presença de uma arquitetura em equilíbrio com o meio ambiente,
Leandro GASPARINI
André Munhoz de Argollo FERRÃO
Arquiteto, consultor do INPAPesquisador do Laboratório de Engenharia de Empreendimentos (LaborE / FEC-Unicamp
Professor Livre Docente do Departamento de Arquitetura e Construção da Faculdade de EngenhariaCivil, Arquitetura e Urbanismo da UNICAMP.
Coordenador do Laboratório de Engenharia de Empreendimentos (LaborE / FEC-Unicamp).
Biotupé: Meio Físico,Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central
SANTOS-SILVA, APRILE, SCUDELLER,
Editora INPA, Manaus, 2005
Edinaldo Nelson Fábio Marques Veridiana VizoniSérgio MELO (Orgs.),

pela utilização de recursos naturais disponíveis (como a mão-de-obra) existentes naregião. A padronização nos processos de aprendizado e transferência de tecnologiaregional proporciona a interrelação entre os projetos transdisciplinares apresentados paraa região, tornando mais equilibrada a reflexão cognitiva sobre o patrimônio ambiental esua sustentabilidade.
transdisciplinaridade, desenvolvimento sustentável, culturaconstrutiva, tecnologia apropriada, paisagem cultural.PALAVRAS-CHAVE:
Introdução
A maioria das propostas de interferência espacialdificilmente considera os legados culturais herdados dequalquer população; e quando se discute sobrehabitações célula formadora de uma comunidade talfato é ainda mais evidente.
Nas construções, normalmente se utilizam materiaise técnicas mais convenientes ao mercado do que àpopulação que vivencia sua comunidade, desconsideran-do-se, neste processo, a necessária integração que sedeve promover entre a população e o seu entornonatural, conforme Lorenz (1974, apud ChiapperoSupisiche, 2003):
Ao se definir que, mais importante que o modoconvencional construtivo usado numa construção, ointeresse maior está na relação do ser humano com omeio ambiente, vislumbra-se um restauro culturalnecessário à comunidade, a partir da inserção doconceito de construção que utilize os recursos da regiãopara o desenvolvimento de processos auto-construtivos.Dessa forma pode-se contribuir para a discussão sobre aformatação da “célula formadora”, pois como admite
&
“La humanidad civilizada se encamina por si solahacia su ruína ecológica mientras asuela, conobcecación y vandalismo, la naturaleza que le circunday nutre. Tal vez reconozca sus errores cuando sienta porprimera vez las secuelas económicas de tal actitud,pero entonces, probablemente será demasiado tarde.Sin embargo, lo que menos percibe es el daño causado asu alma en el curso de ese bárbaro proceso. La ruindadestética y ética de la civilización actual es imputableen gran medida, al distanciamento generalizado yacelerado de la naturaleza viva”.
Santos (1997), é pelo resgate histórico,ou ainda pela constatação de que tudose organiza para formar um sistema,um novo momento onde se buscamodos antigos para se produzir novasmaneiras de ações que obtem-se orespeito ao meio-ambiente.
Ao se utilizar desses pensamentos,há que se pensar no espaço como palcopermanente de atividades condiciona-das à valorização da cultura de seususuários, e mostrar-lhes que serealmente querem, e podem, usar daTerra para viver e se desenvolver,necessitam, mais que tudo, sealimentar da sua cultura.
Inicialmente, parece inquestioná-vel que nos dias de hoje qualquerconstrução deva utilizar materiais outécnicas construtivas que estejamestritamente relacionadas ao pro-gresso e desenvolvimento e, inegavel-mente dentro desse contexto,pensarmos em projetos que valorizemmais o meio ambiente que o comércio,o capital, se mostra equivocado, ecomo menciona Ferrara (1999) asociedade a todo o momento cria afacilidade de substituir um produto
A PERSPECTIVA DO PARADIGMAAMBIENTAL
EspaçoAmbiental
240
Gasparini Ferrão&

por outro, dando um tom de abstraçãoà possibilidade da finitude ambiental,já que o capitalismo tem como uma desuas características marcantes afragmentação das atividades econômi-cas movidas pela necessidade decrescente especialização e comoresultado a habitação vem sendocrescentemente esvaziada das inúme-ras funções para qual foi criada, fun-ções que não são só física, como tam-bém social, econômico e cultural, quecondiciona a um comportamentodecorrente de um modo de vida.
Esse modo de vida cria umaperspectiva ambiental calcada nocomportamento de relações técnicas(Foladori, 2001), onde o relaciona-mento com o meio ambiente écrescentemente objetivado em coisaproduzida. E não havendo mais apossibilidade de combater tais influên-cias, principalmente devido ao esque-cimento dos antigos costumes, ohomem amazônida vê na adaptação detécnicas e materiais contemporâneosum modo de assegurar sua dignidade(Fig. 1).
Essas contradições dentro doambiente de estudo mostram queexiste uma realidade cultural marcadapela ambigüidade e pela ignorância dohomem amazônida, primeiro por criarna facilidade de adaptação dos novosmateriais um imaginário de conforto esatisfação que responde ao desejo deadquirir sua habitação (Fig. 2), esegundo por não perceber os danosque esse imaginário pode trazer aomeio ambiente, imaginando que areposição das fontes de produtossejam inesgotáveis. Diante dessaperspectiva, deve-se impor “a tarefade repensar e de reorientar os destinosde sua cultura, de sua história e de suamemória, em direção a uma ecologia
Figura 1. Uso de materiais típicos de construções urbanas,telhas de fibrocimento e blocos de concreto.
Figura 2. Uso do espaço sem cuidados com a margem do lago.
humana, ética e civilizada” (Oliveira, 2002), onde ocomunitário possa se desenvolver de maneirasustentável, entendido aqui como algo que estejabaseado nos costumes locais e que não agrida o meio
ambiente e as relações que nele se estabelecem.Ao considerar tais contradições, surge a necessidade
de alimentar um pensamento holístico que tornecompreensível a realidade através do resgate cultural dopovo amazônida, para que o equilíbrio entre homem,demais seres vivos e o ambiente não permaneçavulnerável, e cegue qualquer visão sistêmica que
A visão vernacular e o desafio cultural para construções em mutirão
241

garanta uma estrutura ambiental equilibrada. Para isso“a noção de tempo é fundamental. A sociedade é atual,mas a paisagem, pelas suas formas, é composta deatualidades de hoje e do passado” (Santos, 1997).
O surgimento desse paradigma holístico (Fig. 3) cria apossibilidade de uma relação transdisciplinar na qual oshomens possam produzir os seus meios de subsistência ,contudo isso depende da natureza dos meios queencontram prontos e que necessitam reproduzir(Candido, 2003). Para que seja viabilizada tal relação, ohomem amazônida deve estar compromissado a repensaruma realidade sistêmica, uma vez que ele não é mais queuma parcela no ambiente, e não estando sozinho, o quedeve prevalecer é: sociedade e demais aspectos quecompreendem o que chamamos de natureza, se fundindonuma totalidade equilibrada.
O homem capacitado de um dos instrumentos maisfantásticos que talvez já tenha aparecido o cérebrotem em si a qualidade de buscar em um passado distantequalquer desejo que o instigue. Essa máquina incrívelprojeta-lhe todas os modos de raciocínio capaz de lhefornecer imagens, medos, ou simples-mente umavontade de andar ou se expressar.
Isso o coloca diretamente ligado à idéia de que tudopode ser (in)formado, e que essa atitude involuntáriacria uma teia processual de co-evolução e inclusão,inter-relações que permitem associar e avaliar práticasadvindas do seu imaginário (Gattaz Sobrinho, 1999).Dentro dessa visão de processo fica impossível dissociar aexistência de um projeto mental pré-estabelecido queinflua no desenvolvimento sustentá-vel. E como esseconceito encerra em si valores culturais acima de tudo,devendo considerar o homem como pilar dessedesenvolvimento, já que a premissa é buscar o usoracional dos recursos naturais e conquistar bem estarambiental. Para que a conquista se concretize, não sepode esquecer o valor de multidisciplinaridade oumulticulturalidade pois conforme (Oliveira 2002) ohomem se transforma a medida que transforma o mundoe confere forma humana ao objeto de sua transformação
(In)formaçãoAmbiental
“De todas as estações do ano, o inverno é a maisvelha. Põe tempo nas lembranças. Remete-nos a umpassado distante” (Bachelard).
e promove assim o reconhecimentoinformativo sobre biodiversidade e suavalorização.
Ao trabalhar a habitação coletiva,há necessidade de considerá-la nãosomente como espaço físico, mas umlocal onde se estabelecem relações.Nesse sentido a arquitetura torna-seum gênero de ordem espacial e ganhareferência própria, tornando-se umaforma silenciosa de ensino, quesuporta símbolos culturais e ideológi-cos, e a análise desses espaços tem deser feita como uma construçãocultural que expressa e refletedeterminados discursos.
Não é possível falar em paisagem,criação de um espaço habitável, sem apercepção e a interpretação culturalque interferem na vida cotidiana dohomem amazônida (Fig. 4). Conhecere respeitar as diferenças de cada serabre a possibilidade de convivênciatolerante entre o que o homem precisae o que a natureza pode dar, pois deacordo com Foladori (2001) os sereshumanos, como organismos biológicose sociedades equipadas com determi-nadas bagagens culturais, possuem umcomportamento e um instrumentalpara transformar o meio ambiente deforma qualitativamente diferente dosoutros seres vivos. Para fazer prevale-cer esse equilíbrio, cabe a tarefa deresgatar um ideário e dados culturaisque instruam o comunitário para quetenha iniciativas de utilização do meioambiente sem acarretar, ou pelomenos minimizar, danos que sejamirreversíveis, tanto para si como para oambiente.
A área demarcada e local doassentamento a ser estudado .Além da questão econômica, odistanciamento verificado em relaçãoaos pontos de abastecimento de
(Fig. 4)
242
Gasparini Ferrão&

Figura 4. A área demarcada e local do assentamento a serestudado.
região próxima de Manaus, já observava que um certo“conjunto arquitetônico se compunha de casa-grande euma dúzia de casinhas, muito semelhantes às casa deadobe e sapé do sul” (Andrade, 1976) deixando claro aimportância da existência de técnicas construtivas emque são utilizados materiais da região onde se vive.Também pelo grau de influência climática, o homemamazônida adapta seu abrigo com espaços abertos comovarandas (Holanda, 1988)). Todos os relatos fortalecem,perante a população da RDS Tupé, a necessidade da
A visão vernacular e o desafio cultural para construções em mutirão
243
Figura 3. Integração de materiais, técnicas construtivas e oentorno.
recursos estratégicos, faz com que asfamílias nesse local extrapolem asrelações congênitas e criem situaçõesde esforços laborais surgindo um grupo“doméstico-familiar”.
Tal proposta requer capacitaçãopara que o comunitário assimile umaidentidade que restaure do seu sub-consciente conhecimentos herdados,e, de modo criativo e compromissado,torne sua consciência sustentável.Tornar consciente toda sua herançasignifica criar expectativas e mostrarcaminhos para romper “com atendência de viver em um mundo decertezas, de solidez perceptiva nãocontestada” (Maturana Varela,2001).
Saliente-se que não se devecapacitar o homem como indivíduo,nem por distinção sexual, já que oentendimento dos papéis sexuais, bemcomo os sociais, são construídosculturalmente e não determinadosbiologicamente (Chaves , 2004).O desafio de aglutinar esforçosindependente-mente de seu papelsexual reforça a idéia de mutirão, oumelhor, de aplicação de esforçoscoletivos para melhoria do espaçosocial, pois o contexto que se querformatar é de desenvolvimento,conservação, proposta apropriada deresgate da tradição herdada. Detecta-da a prevalência do grupo emdetrimento ao indivíduo, vale concluirque a conscientização vernacular nosentido estrito que significa próprioda região em que está é caminhofactível para a busca de sustentabili-dade.
Mario de Andrade, na viagem emque fez ao Amazonas em 1927, em uma
&
OAmbiente Vernacular
et al.

viabilidade das técnicas construtivasexistentes na sua comunidade, combase em materiais da sua própriareserva natural: madeira, pedra eterra, empregados com conhecimentopróprio do seu sub-consciente.
Como se nota na Figura 5, aocontrário do que se imagina comocaracterísticas de construção daprópria região, nem sempre o uso demateriais como a madeira pode trazerbenefícios para se habitar melhor: porisso o primordial para a população daRDS Tupé talvez seja criar formasexplicativas para valorização do
Figura 5. Casa pré-moldada: problemas de conforto ambientale espacial.
manejo da madeira (por exemplo, suascaracterísticas); assim como, da terra(sua facilidade de adaptação àconstrução); ou seja, conscientizá-losde que esses materiais foram osprimeiros a serem utilizados pelohomem para a criação do abrigo“artificial” depois que abandonou acaverna, sua “habitação primitiva”.
O uso da madeira e da terra comomaterial construtivo se encontrapresente em quase todos os lugares domundo e em quase toda as etapasconstrutivas de qualquer edificação.Nesse ponto, o salto qualitativo que sepretende é fazer com que as
244
Gasparini Ferrão&
Figura 6Ae B. Utilização de taipa na casa de farinha.
Figura 7. Casa de morador assentado.
A B

proporcionará ao comunitário da RDS Tupé maiorconhecimento e reconhecimento da concepçãoestrutural da obra para facilitar a conservação e futurasintervenções, sem deixar de lado os conhecimentos dosantepassados, confirmado por Hertz (1998) ao lembrarque toda proposta construtiva deve levar em conta todosos aspectos climáticos, e ninguém reconhece melhoressa interrelação com o meio que deve ser projetado doque o próprio homem amazônida.
A criação de uma Reserva de DesenvolvimentoSustentável, antes de ser uma norma, deve ter comopapel principal desenvolver um planejamento ambientalpara a região que se deseja preservar. Alguns pontostornam-se relevantes para que se consiga converter asRDS de área de preservação biológica em espaços aptospara a vida humana, não só para o presente como para asfuturas gerações, desmistificando a impossibilidade dehabitabilidade dessas Reservas, e nisso reside a decisãode proporcionar a transdiciplinaridade de pensamentosna concepção de projeto e construção das edificaçõescom a utilização prioritária de materiais locais, além dasculturas construtivas apropriadas, que garantam aparticipação e autonomia da comunidade na construçãode suas habitações.
Uma vez destacados os pontos acima como prioridadepara o bom planejamento das intervenções construtivasnas RDS, e principalmente na RDS Tupé, deve-se salientarentão os caminhos para sua efetivação, e dentro dessecontexto alguns mecanismos devem ser estudados:
1.Anecessidade de desenvolvimento de instrumentospara o estudo dos fenômenos de crescimento dademanda por habitações nas comunidades.
2. A reversão do atual processo construtivo negativo(com alto impacto ambiental pela utilização demateriais trazidos de locais com característicasdiferentes da RDS) no sentido de incentivar um processosustentável que contribua ao equilíbrio ambiental etambém à melhoria das condições de vida das pessoasatravés de sua independência técnica e construtiva.
3. Estímulo à consolidação de uma cultura detrabalho participativo entre todos os comunitáriosenvolvidos nas construções através conscientização das
Conclusão
A visão vernacular e o desafio cultural para construções em mutirão
245
construções sejam essencialmentebaseadas em técnicas construtivas quese apropriem desses materiais comoestruturas e coberturas em madeira,fechamentos de adobe ou taipa (Fig.6A e B) etc., visto que é da natureza docomunitário do Tupé, e mesmo deoutros locais da Amazônia, essa visãotecno-construt iva. Pensar naconstrução com essas característicasnada mais é que organizar o processoconstrutivo, otimizando o uso dosmateriais para que sua exploração nãoseja agressiva à natureza, desenvolverambiente de treinamento queproporcione rápida qualificação dacomunidade, inibindo a concentraçãodo conhecimento nas mãos de poucosindivíduos.
Outro aspecto que deve serconsiderado conjuntamente com aqualificação da população diz respeitoao valor que se deve dar aos modosprojetuais empíricos (Fig. 7) utilizadosna área da RDS Tupé: catalogando edocumentando suas formas e seupensar espacial, para que os resultadosadquiridos e estudados auxiliem nacomplementação das propostasconstrutivas, que estariam divididasem:
Padronização das formas deutilização da terra da região, deacordo com períodos do ano,orientando seu manejo para criaçãode adobe ou taipa, para queadquiram durabilidade e resistência;Criação de posturas que respeitem omeio ambiente: planejar etapasconstrutivas de acordo com ascaracterísticas das madeiras queserão utilizadas, para que estassejam retiradas somente na épocacorreta de corte;
Conseguir integrar essas etapas
�
�

Janelas para a biodiversidade noParque Nacional do Jaú: umaestratégia para o estudo dabiodiversidade na Amazônia,Fundação Vitória Amazônica,Manaus, p.63-78.
Chiaperro, R. O. & Supisiche, M. C.2003. Architectura en la Tierracruda, breve consideración sobre laconservaciõn y la restauración.Nobuko, Buenos Ayres, Argentina.79pp.
Ferrara, L A 1999,
. 2a.ed., Editora daUniversidade de São Paulo, SãoPaulo 280pp
2001,
Campinas225pp.
Gattaz S F 1999.
Campinas Mundo em Processo, EmCD.
Hertz, J B. 2003: Como projetar nos
Trópicos úmidos do Brasil PioneiraThompson Learning, São Paulo.125pp.
1988,Olympio, Rio
de Janeiro. 158pp.Maturana, H R. VARELA, F J. 2001,
. Pala Athena, São Paulo, .288 pp.
Oliveira, J A de 2002,. Editora
Valer Governo do Estado doAmazonas Manaus 54pp.
Santos, M. 1997,, 4a.ed., Editora
Hucitec, São Paulo. 66pp
. ., Olhar Periférico:informação, linguagem, percepçãoambiental
.
obrinho, . . A MáquinaCon tex tua l no s Negóc i o s
,
. . Ecotécnicas emArquitetura
,
. , . , AÁrvore do Conhecimento: as basesbiológicas da compreensãohumana
. . , Cultura,História e Memória
/, .
, Pensando o Espaçodo Homem
Foladori, G.
Editora Unicamp, .
Holanda, S B de.. 20a.Ed., José
Limites doDesenvolvimento Sustentável,
. . , Raízes doBrasil
246
Gasparini Ferrão&
reais responsabilidades desses agentes na preservaçãode valores culturais e ecológicos para si e para as futurasgerações.
4. Resgate cultural coletivo, sem distinção de classeou posição dentro da comunidade com a certificaçãopelos estudos históricos, tanto sociais comoarquitetônicos, e por levantamentos da originalidade deocorrência de casos representativos dentro das regiõesde RDS.
5. Avaliação de pós-ocupação das construções paraverificação e consolidação das atitudes transdisciplina-res atuantes no planejamento das edificações, bemcomo para novos direcionamentos corretivos na co-evolução entre os métodos de trabalho empregados.
6. Garantir que a habitação elaborada mediante usosatisfatório dos recursos naturais atinja um mínimo vitalcompatível com as necessidades do comunitário.
O desenvolvimento e consolidação das etapasdescritas leva em conta que, para uma intervenção emuma cultura construtiva regional, é impossível ignorar oslimites físicos do meio ambiente enfocado e pensar que ohomem e meio ambiente devem caminhar solidários;neste sentido o desenvolvimento e qualificaçãoprofissional são necessários para construção de espaçosacessíveis e salubres, que tenham característica dereparabilidade e reutilização dos próprios materiaisempregados; a auto-construção deve ser vista comoaglutinador social e, facilitar a conscientização sobre aimportância do desenvolvimento sustentável, e por fimsalientar que as RDS só se manterão sustentáveis se todasua paisagem cultural for preservada visto que todasociedade não pode ser abstraída do seu contextohistórico-ambiental.
Referências Bibliográficas
Andrade, M 1976 Duas Cidades:Secretaria da Cultura, Ciência e Tecnologia, SãoPaulo. 382pp.
Candido, A. 2003. Os parceiros do rio Bonito. 10 ed.Livraria Duas Cidades Ltda. São Paulo. 376pp.
Chaves, M. Do S. P. R., Abreu, J. P., Bindá, F. 2004, Ascondições de vida e uso dos recursos naturais pelosmoradores do Parque Nacional do Jaú, in: Borges, S.H.; Iwanaga, S.; Durigon, C. C.; Pinheiro, M. R., (Eds.)
., . O Turista Aprendiz,
a
Copyright © 2022 FDOKUMEN




















