
62 nd Ward Republican Executive Committee - Philadelphia ...

Biosedimentology of Microbial Buildups IGCP Project No. 380
Proceedings of 2 nd Meeting, G6ttingen/Germany 1996
edited by
Fritz Neuweiler, Joachim Reitner, G6ttingen and Claude Monty, Nantes
with con t r ibu t ions b y
M a r k F e l d m a n n , Ztirich; Stan A w r a m i k , St. Barbara; Pierre-Andr6 Bourque , Quebec; Simonetta Cir i l l i , Pe ru#a ; Pascale G a u t r e t , Jean M a r c o u x , Jean Claude Plazia t , Paris; Christian D~farge, Jean Tr lche t , Orl6ans; Claude Monty , Nantes; James L. Pinckney, Morehead City; Janine B e r t r a n d - S a r f a t i , Alexis Mouss ine -Pouchk ine , Montpellier; Pamela Reid, Miami; Martina M e r z - P r e i S , Marburg; S6basden Lab iaux , Louvain-la- Neuve; Aymon Baud, Lausanne; Volker Thiel , Walter Michael is , Ursula Gal l ing , Hamburg; Joachim Reitner, Fritz Neuwei ler , Hartmut M~icker, Gernot Arp , G6tdngen; Jenny K r u t s c h i n n a , Frankfurt; Robert Rid ing , Cardiff; Gregory E. W e b b , John S. Je l l , Brisbane; Andr6 F re iwa ld , Bremen; Manmohan Mohanti, Srikanta Das, Bhubaneswar; Matthias Bergbauer, Gabriela Schumann-Kindei, Werner Manz , Ulrich Szewzyk, Berlin
KEYWORDS: BIOSEDIMENTOLOGY -BIOLOGY- PALEONTOLOGY - MICROB IAL COMM UNITIES -CYANOBACTERIA - FUNGI- STROMATOLITES - SPONGES - CORALS - BIOMINERALIZATION - BIOMARKER - BIOFILMS - ORGANOMINERALIZATION - MARINE ENVIRONMENTS - BIOGENIC MOUNDS - MUD MOUNDS - REEFS -COLD-WATER CARBONATES -NON-MARINE ENVIRONMENTS -MASS EXTINCTION/RECOVERY - AFRICA- AMERICA (BAHAMAS; USA) - ASIA (INDIA; INDONESIA; CENTRAL PACIFIC) - AUSTRALIA (BARRIER REEF; SHARK BAY; LAKE THETIS) - EUROPE (GERMANY; IRELAND; ITALY; SPAIN) - PRECAMBRIAN - CAMBRIAN - DEVONIAN - CARBONIFEROUS - PERMIAN - TRIASSIC - JURASSIC - CRETACEOUS - TERTIARY - HOLOCENE
Part VI: * Organic Matter in Great Salt Lake Ooids (Utah, USA) - First Approach to a Formation via Organic Matrices (J. R E r r ~ , G. ARP, V. TBIEL, P. GAU'mET, U. GALLING & W. MICHAnLIS) . . . . . . . . . . . . . . . . . 210
Part VII: Cryptic Microbialite in Subtidal Reef Framework and Intertidal Solution Cavities in Beachrock, Heron Reef, Australia: Preliminary Observations ( G. E. WEBB & J. S. JELL) . . . . . . . . . . . . . . . . . . . . . . 219
Part VIII: MicrobialAherationoftheDeep-waterCoralLophelia pertusa: Early Postmortem Processes (A. FREIWALD, J. REITNEa & J. KRtrrscmN'NA) . . . . . . . . . . . . . . . 223
Ancient Stromatolites and Mierobialites Part: IX: Stromatolite Decline: A brief Reassessment (R. RtD-
~G) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 227 Part: X: Archean and Proterozoic Lacustrine Stromatolites
(S. AWXArmK) . . . . . . . . . . . . . . . . . . . . . . . . . . 230 Part: XI: Vermicular Structures in the Neoproterozoic of the
West African Craton: Microbialites versus 'Molar Tooth' (J. BERTRAND-SARFATI, J.C. PLAZIAT & A. MOUSStNE-PoucnKB~E) . . . . . . . . . . . . . . . . . . . . 231
C O N T E N T S
Introduction Part I: Project 380 of the International Geological Cor-
relation Programme (IGCP): Biosedimentology of Microbial Buildups. (C. MONTY, F. NEtrWEILER, J. Rnrrmm) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 197
Part II: IGCP 380 - 2 M Meeting Report (F. NE .UWEJLE~)..197
Modern benthic microbial communities and their rela- tionships to mineralization events
Part III: Stromatolitic Laminae Formation and Carbonate Pre- cipitation associated with Microbial Mats from Mod- em Bahamian Environments (M. FELDMANN)...200
Part IV: Productivity and Community Composit ion of Stromatolitic Microbial Mats in the Exuma Cays, Bahamas (J.L. PI~CKrrnY & R.P. REID) . . . . . . 204
Part V: High Resolution Cryo-scanning Electron Microscopy of Modem Stromatolites (Lake R2, Rangiroa Atoll, Tuamotu Archipelago, Central Pacific Ocean) (J. TRICtmT & C. D~FAR~E) . . . . . . . . . . . . . . . . . 207
Addresses: Dr. F. Neuweiler, Prof. Dr. J. Reimer, Institut for Geologic and Pal~iontologie, Goldschmidtstr . 3, D-37077 G6ttingen, Germany, Fax:+ 49-551-397996, email: [email protected]; j rei [email protected]; Prof. Dr. C. Monty, Lab. de Biosedimentologie, Univ. de Nantes, 2 rue de la Houssini~re, F-44072 Nantes cedex 3, France, Fax: +33-240-293251

196
Part: XII:
Part: Xl~:
Part: XIV:
Part: XV:
Microbial Signatures in Lacustrine and Fluvial Car- bonates: Gondwana (Permian) and Holocene Exam- pies, India (M. MorlArfrx & S. D/d) . . . . . . . . . 234 Biotic Response to Mass Extinction: the Lowermost Triassic Microbialites (A. B^UD, S. CmmLX & J. M~cotrx) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238 Facies Analysis in Upper Jurassic Stromatolites: Sup- port by Palynological and Stable Isotope Data (M. M~,z-PR~ss) . . . . . . . . . . . . . . . . . . . . . . . . . . 242 Upper Cretaceous and Lower Tertiary Brackish to Freshwater Oncoids and Stromatolites oftheGarumrtian Facies in the/~,ger Basin (Central Southern Pyrenees/ Spain) (H. MhcK~) . . . . . . . . . . . . . . . . . . . . . 246
Carbonate Mud Mounds and Biogenic Mounds Part XVI: PaleozoicFinelyCrystaUineCarbonateMounds:Cryp-
tic Communities, Petrogenesis and Ecological Zona- tion (P.-A. BOURQUE) . . . . . . . . . . . . . . . . . . . . . . . . 250
Part XVII: Sponges in Waulsorfian-type Mudmounds atTralee Bay, Co. Kerry, Southwest Ireland (S. LAnIAUX) . . . . . . 253
Part XVIII:Environmental v e r s u s Organic Controls on Bio- genic Mounds: examples from the Upper Triassic of Northern and Central Apennines (Italy) (S. CIRILL1) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 257
Part XIX: **Mud Mound type Community Replacement on Carbonate Platforms and Coralgal Reefs (F. N~trw~mER) . . . . . . . . . . . . . . . . . . . . . . . . . . . 262
Part XX: Aerobic and Anaerobic Microorganisms in Modern Sponges: a Possible Relationship to Fossilization Processes (G. S CHUMANN-KINDEL, M. BERGBAUF~, W. MASZ, U. SZEWZYK & J. RErrm~) . . . . . . . . . . . 268
Part XXI: Pyrite in Mineralized Sponge Tissue - Product of Sulfate Reducing Sponge-related Bacteria? (J. & G. ScmrMAsN-K~DFI . . . . . . . . . . . . . . . . . . 272
* Contribution 1 and ** Contribution 2 SFB 468: "Wechselwirkungen an geologischen Grenzlq~ichen'.
Addresses: Dr. M. Feldmann, Geological Institute, ETH Zentrum, CH-8092 Ziirich, Switzerland Prof. Dr. S.M. Awramik, Department of Geological Sciences, Preston Cloud Research Laboratory, University of California, Santa Barbara, CA 93106, USA Prof. Dr. P.A. Bourque, Department of Geology, Laval University, Quebec, Canada G1K 7P4, Fax: +418-656-7339 Dr. S. Cirilli, Dipartimento Scienze della Terra, Univ. degli Studi di Perugia, Piazza UniversitY, 1-06100 Perugia, Italy Dr. Pascale Gautret, Universit6 Paris 11-Orsay, Bat. 505, F-91405 Orsay Cedex, France; Fax: +33-1-69156123 Prof. Dr. J. Marcoux. Lab. De Grol., Univ. Paris 7, Paris, France Dr. J.C. Plaziat, Laboratoire de Grologie srdimentaire, Bat 504, Universit6 Paris Sud, F-91405 Orsay, France Dr. C. Dgfarge, Prof. Dr. J. Trichet, Institut des Sciences de la Terre d'Orlrans, C.N.R.S., Univ. d'Orl6ans, Batiment des Grosciences, B.P. 6759, F-45067 Orlgans cedex 2, Fax: +33-2417308 Prof. C. Monty, Lab. de Biosedimentologie, Univ. de Nantes, 2 rue de la Houssini&e, F-44072 Nantes cedex 3, France, Fax: +33-40293251 Dr. J. L. Pinckney, Institute of Marine Sciences, University of North Carolina at Chapel Hill, 3431 Arendell Street, Morehead City, North Carolina 28557, Fax: +1-919-726-2426 Prof. Dr. J. Bertrand-Sarfati, Laboratoire de Palrobotanique, UMR 5554 ISEM, CP 62, UM2, F-34095 Montpellier, France, Fax: +33-67-042032 Dr. A. Moussine-Pouchkine, Grofluides, Bassin Eau, UMR 5569 ISTEM, CP 57, UM2, F-34095 Montpellier, France Dr. R. Pamela Reid, Rosenstiel School of Marine and Atmospheric Science, University of Miami, 4600 Rickenbacker Causeway, Miami, Florida, 33149, USA, Fax: +1-305-361-4632 Dr. M. Merz-PreiB, Institut fur Geologie und Pal~iontologie, Hans-Meerwein-Stral3e, D-35032 Marburg, Germany, Fax: +49-40-41236347 Dr. S. Labiaux, Universit6 Catholique de Louvain-la-Neuve, Place Louis Pasteur 3, B- 1348 Louvain-la-Neuve, Belgium Dr. A. Baud, Musre de Ggologie, BFSH2-UNIL, CH-I015 Lausanne, Switzerland Dipl.-Geol. V. Thiel, Prof. Dr. W. Michaelis, Dipl.-Geol. U. Galling, Institut f~ir Biogeochemie und Meereschemie, Universit~t Hamburg, Bundesstr.55, D-20146 Hamburg, Germany; Fax: +49-40-4 123-6347 Prof. Dr. J. Reitner, Dr. F. Neuweiler, Dipl.-Geol. H. Macker, Dipl.-Geol. G. Arp, Institut und Museum fiir Geologie und Pal~iontologie (IMGP), Goldschmidtstr. 3, D-37077 Gi3ttingen, Germany, Fax:+ 49-551-397996, email: [email protected], [email protected], [email protected], [email protected] Dipl.-Biol. J. Krutschinna, Zoologisches Institut, J. W. Goethe Universit~t, Siesmaierstr. 70, D-60054 Frankfurt a.M., Germany Dr. R. Riding, Department of Earth Sciences, University of Wales, Cardiff CF1 3YE, United Kingdom; Fax: +44-01222- 874326 Dr. G.E. Webb, Dr. J.S. Jell, Department of Earth Sciences, The University of Queensland, Brisbane, QLD, 4072, Australia Dr. A. Freiwald, Universit~t Bremen, Fachbereich Geowissenschaften, Postfach 330 440, D-28334 Bremen, Germany Prof. Dr. M. Mohanti, Dr. S. Das, Department of Geology, Utkal University, Bhubaneswar-751004, Orissa, India Dipl. Biol. G. Schumann-Kindel, Dr. M. Bergbauer, Dr. W. Manz, Prof. Dr. U. Szewzyk, Okologie der Mikro- organismen, Technische Universit~t Berlin, Franklinstr.29, D-10587 Berlin, Germany, Fax: +49-30-31473461

197
- INTRODUCTION - Part I
PROJECT 380 OF THE INTERNATIONAL GEOLOGICAL CORRELATION PROGRAMM
(IGCP): BIOSEDIMENTOLOGY OF MICROBIAL BUILDUPS
C. Monty, F. Neuweiler and J. Reitner
As stated by UNESCO (access via http://unesco.org), the International Geological Correlation Programme (IGCP) is a co-operative enterprise of UNESCO (United Nations Educational, Scientific and Cultural Organization) and IUGS (International Union of Geological Sciences). Launched in 1972, IGCP currently operates world-wide with several thousand scientists in about 150 countries. Focusing on 'Geoscience in the Service of Society' the main objectives of IGCP are i) to increase our understand- ing of the factors controlling the global environment in order that human living conditions may be improved, ii) to develop more effective ways to find and assess natural resources of energy and minerals, iii) to increase knowl- edge of geological processes and geological concepts through correlative studies of many locations around the globe, and iv) to improve standards of research, methods and techniques of carrying out research. Major aim of the programme was to bring together scientists from East and West and to encourage the involvement of developing countries.
IGCP 380: Biosedimentology of Microbial Buildups
IGCP 380: Bioscdimcntology of microbial buildups was launched by Claude Monty, October 1995 (1 st Meet- ing, Paris). The project intends to adopt a widcr perspec- tive than IGCP 261 which was concerned with stromatolites. The field of IGCP 380 covers microbial sedimentation, including stromatolitcs and thrombolitcs, as well as non- laminated microbial buildups. Therefore it is concerned with detailed study of biological mediations within sedi- mentary proccsscs and resulting products. These biomediations comprise biological, biogeochemical, organochemical and/or biophysical interferences which may completely transform sedimentary products. Linked with the activity of living microbial systems, IGCP 380 focuses on life as a geological force, and as life evolves through time, any approach to microbial buildups is framed into an historical and spatial framework. Stromatolitcs appear to have the longer story of all known microbial buildups, i.e. microorganisms have been interacting with sediments for over 3800 million years. But to understand micro- and macromorphologies we have to elucidate the processes acting between living microbial mats/biofilms and the host sediment to find out 'who' is doing 'what', 'where' and 'what for'.
Mud mounds are particular non-laminated microbial buildups which rise in the Lower Phanerozoic (and per- haps in the Late Precambrian). These structures are essen- tially made of carbonate mud which formed locally. As
opposed to stromatolitcs, mud mounds may also incorpo- rate metazoans to a different extent and appear to be strictly limited to marine environments. From the lowermost Cambrian to theLatc Cretaceous mud mounds show evolu- tionary paths. However, there is an urgent need of systematic studies on mud mound constitution through time.
We can summarisc that research within IGCP 380 is concerned with the roles of living microbial communities and non-living organic substratcs to control, mediate and/ or influence mineralization patterns in stromatolites, mud mounds and other related microbial carbonate buildups. This research is based on five main topics i) analysis of modern microbial communities and their
relationships to mineralization events ii) qualification of time/space relationships of bio- and
organomineralization iii) selective characterization of processes and mineralization
products iv) application of tracer techniques to transfer these char-
acteristics into the fossil record v) elucidation of the history and environmental signifi-
cance of microbial sediments such as stromatolitcs, thrombolitcs and mud mounds.
Editorial Note This compilation comprises very different kinds of
papers (conceptual, case studies, short notes and state- ments), representing rather the scientific activity of indi- vidual IGCP 380 members than the outcome of a research group. IGCP 380 is a young project and results of joined research are still exceptional. Bearing this in mind, the main purpose of this compilation is to state that IGCP 380 is a project of active research on global scale providing excellent opportunities for stimulating and co-operative scientific results.
F. Neuweiler wishes to thank all contributors for their help to submit this compilation already 3 months after our 2 nd meeting. While preparing the final manuscript additional support was provided by G. Arp, F. Gunkel and C. Hake (G6ttingen). For additional information on IGCP 380 via Internet see http://www.gwdg.de/~fneuwei/380home.htm
Part II IGCP 380 - 2 nd MEETING REPORT
F. Neuwciler
1 INTRODUCTION The 2 nd Meeting of the IGCP 380 focused on'Microbialites
- Processes and Products' and was held from October 5 ~ to 8 ~h 1996 at the 'Institut and Museum ftir Geologic and Palaontologic' (IMGP), University of GOttingen, Germany. During these days a total of 30 participants from 10 countries (Australia, Belgium, Canada, France, India, Italy, Switzerland, United Kingdom, USA and Germany) met for three field work shops and two scientific sessions on microbialites and mud mounds. The meeting was con- vened by J. Reitner and F. Neuwciler with the collabora- tion of J. Paul (G0ttingen), H. Weller (Greifswald), V. Thicl (Hamburg), M. Merz-Preiss and H. Zankl (Marburg).

198
2 FIELD WORK SHOPS The 18t field work shop entitled 'Kalkowsky's Stromato-
lites revisited (Triassic)' covered 4 major outcrops within the type region for stromatolites and ooids along the northern and southern borders of the Harz Mountains (Scythian, Lower Buntsandstein). We followed a transect fxom marginal, fluviatile deposits consistingofred sandstones with small amounts of ooids and stromatolitic encrusta- tions towards the center of stromatolite occurrence charac- terized by pure oolithic limestones and various forms of stromatolites ranging from small encrustations and domal overgrowths upon ooids to conical shaped, complex forms up to 1.2 m in hight. Discussion focused on the variety of microbial textures, indications of early induration, and specific growth patterns of larger ooids and evident rela- tionships with modem counterparts like those from the Great Salt Lake (USA) (see REn~tER et al. this vol.).
The 2 nd field work shop entitled 'Devonian Microbial Carbonate Muds and associated Zebra-Limestones; Harz Mountains' dealt with a local occurrence of Frasnian mi- crobial and detrital muds in the central part of the Harz Mountains. Discussion on the origin of stromatactoid cavities, the relationship of microbial muds with Epiphyton, hemipelagic muds bearing dacryoconarides, nautiloids and conodonts, and the formation of extended cavities related to sheet cracking and collapse structures were held in the field. There is strong evidence for a deeper-water assemblage including hexactinellid sponges and for pri- mary slopes up to 25 ~ resulting in major gravity mass movement. However, there was no direct evidence that these deposits were originally related to larger mud mound structures.
The 3 rd field work shop entitled 'Non-Cyanobacterial Stromatolites (Thfiste, Late Jurassic)' covered the regres- sive sequence within the Portlandian south of Hannover (see MEI~z-PRmsS this vol.). Marginal marine serpulite and oolite beds grade into intertidal stromatolites, which are covered by ostracod-bearing marls indicating brackish conditions. The stromatolites occur as isolated, oval to cone-like bodies lining a continuous stratigraphic level. Within the core there are remnants of serpulid reefs and related sediment overgrown by thick stromatolitic crusts. The outer part consists of dense stromatolitic laminae with continuous and columnar growth forms. Background data on palynofacies, stable isotopes and biomarkers contrib- uted to a comprehensive discussion resulting in perspec- tives concerning the compilation of different methods towards a better understanding of formation processes. For example, petrographic evidence points to the exist- ence of cyanobacterial mats at the surface, biomarkers provide strong evidence that rock-forming processes were strongly related to anaerobic bacteria, resulting in a spe- cific and well preserved organofacies lacking any traces of cyanobacterial origin.
3 SCIENTIFIC RESULTS Scientific sessions were held at the IMGP at October
7th and 8th covering case studies from the Neoproterozoic and Phanerozoic rock record as well as modem examples
from temperate to tropical regions. Modem occurrences of marine, lagoonary and lacustrine microbial benthic com- munities are studied in tropical/subtropical regions like the Bahamas, the Great Barrier Reef (E Australia), various atolls in French Polynesia, Great Salt Lake (USA), Satonda Crater Lake (Indonesia), and Lake Thetis (W Australia), as well as within temperate regions of the Norwegian Shelf.
3.1 Analysis of modern microbial communities and their relationships to mineralization events
The formation of stromatolitic laminae in modern Bahamian environments (FELDr~a~) can be related to five distinct processes: i) Calcification of mat forming cyano- bacteria (Dichothrix bornetii), ii) precipitation ofmicrospar within a microbial mat environment dominated by Micro- coleus, iii) dissolution and reprecipitation of calcium car- bonate associated with Schizothrix, iv) deposition of detri- tal layers mainly stabilized by the green alga Ostreobium, and v) precipitation of calcium carbonate linked to the degradation of organic matter. Initial studies of Exuma stromatolites (Bahamas; REID) indicate that the microstruc- tures of lithified micritic layers are determined by bio- logically derived gradients with respect to pH, CO 2 and 0 2.
Reef cave microbialites from Lizard Island (Great Barrier Reef; REIT~CZR) are mainly a product of matrix mediated calcification via acidic organic macromolecules. Microbial biofilms with coccoid and rod shaped bacteria cover thrombolitic surfaces of autochthonously formed micrite. They control the release of organic substances as well as the settlement and distribution of benthic organ- isms. Geochemically these microbialites are high Mg- calcites. The stable isotope composition (813C +3.0 to +3.8) indicates that primary micrite formation is close to the expected equilibrium. Micrite production in place also occurs during sponge soft tissue diagenesis and may even result in the preservation of different types of mesohyle structures (bacteriosponges with minipeloidal structures; syncytial tissue of hexactinellids with dense, aphanitic structures).
S ubtidal microbialites in reef frameworks of the Great Barrier Reef (Heron Reef, WEB~ & JELL) share some important features with those from Lizard Island reef caves. They occur as crusts characterised by a high variety of morphologies but exhibit a uniform Mg-calcite miner- alogy and stable isotope composition (813C +3.3 to +3.6). Satonda Crater Lake (Indonesia) is characterised by highly alkaline conditions favouring microbial calcification (Am, REn-~R). Due to specific hydrochemical conditions the biota of the lake are very specific and endemic. Cyanobacteria and heterotrophic microbes exhibit a high diversity, con- trasting a single sponge taxa (Suberites/Polymastia n. sp.), Common taxa of the microbial community arePleurocapsa, Phormidium, Calothrix, Spirulina and Microcystacea. In part, calcification is related to decaying green algal fila- ments. The importance of in situ calcifying cyanobacterial biofilms is not clear yet, because only a few traces of calcifying microbial sheets have been observed.
Domal microbialite bioherms occur along the shore of Lake Thetis (W Australia; REIa~ER, ARV & PAUL). This

199
alkaline, saline lake exhibits subfossil microbialites con- sisting of Scytonema filaments embedded within fibrous aragonite. Actually, growth of bioherms is due to calcify- ing Entophysalis mats forming weakly laminated crusts at the surface. Calcification proceeds from intermediate parts of the mat where heterotrophic bacteria are associated with basophilic organic matter.
3.2 Qualification of time/space relationships of organomineralization
Tropical ponds on reef margins in French Polynesia are covered by abenthic microbial community with filamentous cyanobacteria of the taxon Phormidium (Ttncazr, DI~FARGE). Polymers extend outside the bacterial cell or may even be separated from the living microbial community. The slime is composed of polydispersed polysaccharide fibres, which reorganise into a honeycomb-like chambered structure during early diagenesis. This structure serves as an organic framework for the precipitation of Mg-calcite within or- ganic rich microbial sediments. Advanced techniques (field emission electron microscopy coupled with cryo-tech- niques; TRXCrmT, DEFARGE) illustrate various states of organomineralization with a resolution of 4-5 nm.
Organomineralization is also a crucial process in the formation of the Great Salt Lake ooids (REITNER, ARP, TmEL, GAmX~, MICHAELIS, NEtrW~mER). Ooids exhibit intracrystalline organic matter of up to 500 lag/g carbonate with an amino acid composition dominated by glutamic (30-40 %) and aspartic acid (10-15 %). The ooids actively grow within dysoxic sediment and are covered by high amounts of this Ca2+-binding organic mucus - a character- istic organofilm.
Mineralization upon and/or within non-living organic substrates is also crucial for micrite production in place as observed within Lizard Island reef caves. This occurs at the surface and within semi-closed and closed pore sys- tems (REITNER, GAUTRET, NEUWEILER).
Late preservational stages in modern deep-water coral reefs (azooxanthellate Lophelia pertusa) exhibit large amounts of carbonate mud (FP~IWALD). The conversion of a pure coral framework to a coral rubble facies and a biodetrital mud mound is controlled by sponge excavation and the supply of bioerosionally produced carbonate mud, exemplifying a residual buildup produced by degradational processes rather than an upward accretion.
3.3 Characterization of processes and mineralization products
Mineralization products within microbialites/stromato- lites and the production of carbonate mud - deposited in- place and/or biodetrital - can be characterised by mineral- ogy, stable isotope geochemistry and biogeochemistry. Beyond the scope of comparative petrography (macro- and microfabrics) this multidisciplinary approach helps to cor- relate products and processes in the modern, and provides a powerful tool to trace these processes back into the geological record.
Mineralogy: In general organic macromolecules are the
key compounds in mineralization processes. The orienta- tion and relative distance of Ca-ions as well as the presence of other ions (Sr 2+, Mg 2§ and kinetic factors control the mineralogy (calcite, aragonite, vaterite). Modern exam- pies provide in-situ mineralized calcite and aragonite upon living and non-living organic substrates as well as biodetrital mixtures of various sources, which will help to enlarge our database of natural intracrystalline organic compounds and their function concerning mineralogy, crystal shape and initial seeding (TRIcHE'r, Dt~FARGE, REITNER, GAUTRET, NEUWEILER, TI-ImL, MICHAELIS). Stable isotope geochemistry: Stable isotope geochemistry (813C, 8~sO) of calcium carbonates reflects different modes of calcification (vital effects, kinetic effects), chemical variation of ambient waters (marine, freshwater etc.; e.g. freshwater microbialites; MXcmza, MOVtANTQ, and later stage of diagenetic overprinting. Analysis of 813C and 81sO was applied in most of our case studies and provide additional data of reference for the geological record. Analyses of amino acids: Intracrystalline organic com- pounds can be analysed for amino acids to detect the relative amount of asp and glu - acidic amino acids which are able to bind divalent ions like Ca 2§ and Mg 2+. In addition to this signature of matrix mediated calcification, whole spectra of amino acids provide a powerful tool to correlate processes and products and to elucidate early diagenetic alterations (GAUTRET, GUIF, TRICHET, DEFARGE, REITNER, NEUWEILER) Biomarker analysis: Biomarkers are organic compounds (lipid constituents) derived from living organisms and retain their typical structure in the geological record. They are used to provide information on the source of organic matter, the paleoenvironmental conditions, and postdepos- itional processes like thermal and microbial alteration (THIEL, MICHAELIS, TRICHET, DEFARGE)
3.4 Application of tracer techniques to transfer these characteristics into to fossil record
i) Matrix-mediated calcification is crucial for the de- velopment of Lower Cretaceous mud mounds including an important amount of metazoan-related micrite production in-place (sponge soft tissue diagenesis). Comparative analysis of macro- and microfabrics, mineralogy, stable isotope geochemistry, amino acids and biomarkers point to a common mode of mineralization, i.e. micrite production (organomicrite) upon non-living organic substrates (NEuWEILER, REITNER, GAUTRET, THIEL, MICHAELIS). These processes are presumably signifcant for Paleozoic mud mounds (LABIAUX, BOURQUE, FLAJS) and most of Late Jurassic microbial sponge mounds and mud mounds (SCHVnD, LEINFELDER).
ii) Upper Jurassic stromatolites (Tithonian of Thilste, Germany) were studied using macro- and microfabrics, stable isotope geochemistry and biomarker analysis (ZANKL, ~[VIERZ-PREISS, THIEL, MICHAELIS). There is good evidence that apart from the preservation of filamentous bacteria (cyanobacteria) mineralization processes were confined to anoxic micro-environments and the presence of anaerobic bacteria (methanogene Archaea).

200
iii) Comparative analyses of ooids from the Great Salt Lake, Kalkowsky's type region (Lower Triassic of the marginal Harz Region, Germany) and the Precambrian of Nevada/USA (Beck Spring Formation) indicate an analo- gous organomineralization within modern and ancient salt lakes (REITNER, ARP, GAtn'm~T, TmEL, MICHAEl.IS).
3.5 Elucidation of the history and environmental significance of microbial sediments
So far, case studies involved in IGCP 380 cover a time span from the Proterozoic (e.g. BER~Ar~D-SARFA~) and most parts of the Phanerozoic. The 'Environmental Con- straint Model' (RmiNc) provides a new perspective con- cerning the response of microbes (and metazoans) to environmental factors like e.g. temperature, 02 and CO 2. According to R m ~ the mid-Proterozoic decline of micro- bial carbonates correlates with a decline in temperature and oxygen increase. Analogous shifts are present during mid-Ordovician time. It appears that when microbes de- clined then metazoans were able to expand, but when microbes were abundant then metazoans declined.
The intrinsic biotic response to the Permian-Triassic boundary mass extinction was a bloom of disaster forms like bacteria, cyanobacteria and fungi (BAtrO, CtRtLLI). Eurytopic and/or primitive forms emerge from deep-water and marginal settings to recolonise the relatively vacant marine setting as r-selected generalists.
This sort of environmental perturbations should be recorded in the geological record whenever ecological threshold values of stenotopic organisms (resp. communi- ties dominated by K-strategists) are exceeded. The geo- logical record provides the time span to observe the col- lapse of natural systems, the interval of disaster forms and the successive recovery by adapted stenotopic communi- ties (Rmlr~, BAuo, CmmtJ, NEtrW~LER). Opportunistic systems are part of cryptic communities of modern coralgal reef bodies (Rm-~rER) and their behaviour could be moni- tored in modern, ecologically stressed coralgal reefs.
- MODERN BENTHIC M I C R O B I A L COMMUNITIES AND THEIR RELATIONSHIP
TO M I N E R A L I Z I N G EVENTS - Part III
STROMATOLITIC LAMINAE FORMATION AND CARBONATE PRECIPITATION
ASSOCIATED WITH MICROBIAL MATS FROM MODERN BAHAMIAN ENVIRONMENTS
M. Feldmann
! INTRODUCTION The most conspicuous feature of stromatolites is their
distinct laminated structure, the significance of which has been discussed by numerous authors during this century (see KRUrCmEJN 1983). KAt~OWSKV (1908) defined the term stromatolite on the basis of the presence of laminae, the formation of which he related to an organic origin. Based on Kalkowsky's definition, stromatolitic laminae com-
monly have been regarded to have formed primarily due to two interacting processes, namely sedimentation and mi- crobial growth (PARK 1976). However, preliminary studies of microbial mats from modern environments in the area of Lee Stocking Island, Bahamas (Fig. 1), show that at least five distinct mechanisms of precipitation and sedimenta- tion can be distinguished, which can lead to the formation of stromatolitic laminae.
2 FORMATION OF STROMATOLITIC LAYERS 2.1 Lamina formation by calcification of mat-
forming cyanobacteria An example of recent upper intertidal stromatolites
occurs in a small, very shallow bay at the northwestern end of Lee Stocking Island (Fig. 1). These stromatolites are growing on exposed bedrock, possibly Pleistocene in age, and are surrounded by beach sand. Their surface mats are dominated by the filamentous cyanobacterium Dichothrix bornetii (GoLtn3ir pers. comman.). These cyanobacteria form a 'grassy' surface mat with a thickness of about 1 mm. These organisms appear to live chasmolithic in the pore spaces between adjacent grains at the top of the surface. The individual filaments, which have diameters of up to 30 I.tm and lengths of up to 1 ram, are heavily encrusted by calcium carbonate (PI. 52/1). These precipitates which appear as rhombohedral crystals are probably the result of photosynthetic activity. They are attached to the secreted organic sheath surrounding the trichome (PI. 52/2). With increasing calcification of adjacent cyanobacterial fila- ments a stromatolitic lamina forms (P1. 52/3).
2.2 Lamina formation by precipitation of microspar within a microbial mat environment
A qualitative experiment using pieces of a microbial mat was performed in April and May, 1992 in the Wet Laboratory of the Caribbean Marine Research Center on Lee Stocking Island. Several lime-mud pieces, each with an area of about 10xl0 cm 2, and a thickness of 1 cm, were taken from a desiccated pond (Fig. 1), which is usually only flooded after rainfall. About 1 mm below the surface of the pieces, a very thin, dark green layer occurred which was dominated by the filamentous cyanobacterium Micro- coleus sp. The mud pieces were placed on a wet table with slowly, but continously, flowing normal marine water passing over them. The wet tables were in an open labora- tory covered by a straw roof which protects the tables from direct sunlight.
After the mud pieces had been covered with water on the wet table for four hours, some initial green spots appeared at the edges of the patch surface. After two days the surfaces of the mats were entirely overgrown with a green gelatinous microbial mat having a thickness of about 100 gm. The mat was dominated by the filamentous cyano- bacterium Microcoleus sp. Subsequently, some of the mud pieces were completely covered with an ooidal sand layer about 1-2 mm thick. After approximately four hours, new green cyanobacterial spots appeared on top of the sand layer. This time the spots were not exclusively at the edges but distributed over the entire surface. The cyanobacteria

201
Fig. 1. Location of the subtidal strom- atolites, the Recent intertidal stromatolites, and the island pond in the area of Lee Stocking Island, Bahamas. Inset map of the Bahamas shows the geographic rela- tion to the study area. Modified after DILL (1991).
apparently had migrated through the sand cover. After two days, the mud pieces were intensely overgrown with a new microbial mat which by now was organically binding the underlying grains (PI. 52/4). After about seven days, tiny white spots or precipitates appeared within the gelatinous part of the mats. These spots are composed of aggregates of euhedral to subhedral crystals. The aggregates have grain sizes up to 20 ttm and occur as floats within the mat. After about 4 weeks the precipitated aragonite began to form a crust resulting in the formation of a new stromatolitic lamina (P1.52/5).
2.3 Lamina formation by deposition of a detrital layer
In the area of Lee Stocking Island, the seawater is highly saturated with suspended carbonate grains such as ooids and peloids. These detrital grains are transported by wave action and tidal currents and are deposited on sur- faces during slack time forming stromatolitic sedimentary laminae. On subtidal stromatolite surfaces, such detrital layers are subsequently often stabilized by the alga Ostreobium.
2.4 Lamina formation by dissolution and reprecipitation of previously deposited carbonate
Most of the lower intertidal and upper subtidal stromatolite surfaces in the area of Lee Stocking Island are covered by microbial mats which are dominated by the filamentous
cyanobacterium Schizothrix sp. (BRownE 1993; FELDMANN 1995). The detrital carbonate grains which serve as substrate for the microbes are commonly incorporated in the micro- bial mat (PI. 52/6). Endolithic cyanobacteria, such as Schizothrix, secrete an organic acid which enables them to dissolve calcium carbonate as shown by abundant bore holes and dissolution tracks on carbonate grains (Hor,~ & WicKs 1980). Ca 2§ ions are released and probably metabolically pumped into the non-acidic, extracellular, organic microenvironment, where they are concentrated leading to the precipitation ofCaCO 3 with supersaturation. The ability of Schizothrix to dissolve calcium carbonate weakens the particles leading to their ultimate disintegra- tion. This profess, resulting in the formation ofa stromatolitic micrite lamina, possibly is most important in the formation of modern Bahamian stromatolites (PI. 52/7).
2.5 Lamina formation associated with bacterial degradation of organic matter
After being covered by sediment particles, cyanobacteria are forced to migrate upwards to the surface, whereby the sheaths and organic matter, which was produced to large portions by eukaryotic algae on subtidal stromatolites, are left behind in the subsurface. The biological assemblage in subsurfaces from Lee Stocking stromatolites is dominated by various bacteria (PI. 52/8), although individual cyano- bacteria such as Oscillatoria sp. still occur. The bacterial degradation of the organic matter in the subsurface, asso-

2 0 2
ciated with a specific chemical environment produced by microbes (i.e. sulfate reduction), may lead to the precipi- tation of carbonate and subsequently to the formation of a lithified layer (see Em~iacri 1996).
3 THE SIGNIFICANCE OF LAMINAE FORMATION UNDER VARIOUS
CONDITIONS Although five mechanisms for the formation of
stromatolitic laminae associated with living microbial mats were observed, stromatolites commonly only reveal two to three distinct alternating stromatolitic layers. This may be explained by the nature and composition of the stromatolite forming microbial mat. Lower intertidal and upper subtidal stromatolites from Lee Stocking Island, commonly have alternating detrital and micritic layers. The detrital layer consists of deposited carbonate sand, the micritic layer apparently is precipitated aragonite of previ- ously dissolved carbonate grains (FF0..OMAr~ 1995; FEt.OMAr~ & McKENzm in press). This process is associated with the endolithic activity of the filamentous cyanobacterium Schizothrix. Upper intertidal stromatolites having a Dichothrix mat commonly have alternating layers of detri- tus and microspar, whereby the microspar is the product of filament calcification. Both, Schizothrix and Dichothrix are adapted to a detrital-rich environment and do not form well developed gelatinuous surface mats. In contrast to both, Schizothrix and Dichothrix, Microcoleus forms a relatively thickgelatinuous microbial matcoveringa substrate in a detrital-poor environment. If there is sufficient expo- sure time, carbonate can precipitate and form a layer in association with the photosynthetic activity of the cyanobacteria.
Because lamina formation associated with bacterial degradation of organic matter is a subsurface process, its significance in stromatolite formation remains specula- five. The microfabrics of most ancient stromatolites dif- fers from that of their modem counterparts. Ancient stromatolites often have alternating layers of micros'par and organic-rich micrite, if not altered. This feature is basically absent in modern forms, such as the Bahamian stromatolites (PI. 52/7), and can not be explained with lamina formation on the surface alone. However, it can be explained by the combination of a surface and a subsurface process. Fine sediment can derive from the water column or from photosynthetically induced precipitation within a microbial mat and form a layer which covers large portions of mat forming microbes. Thus, they are forced to migrate upwards to the surface. With the bacterial degradation of the organic material left behind, an alternating layer sub- sequently could form in the subsurface under favoured conditions. Consequently, such combined processes re- quire an environment where microbial mats have suffi- cient time to produce sufficient organic material. A tidal flat area, for example, could provide such an environment. However, if such stromatolite forming processes occur currently, it is difficult to recognize them because they take place in the subsurface.
4 CONCLUSION The examples presented above show that the formation
and the microstructure of stromatolitic laminae depend on the microbial mat and its biologic composition which itself is controlled by environmental conditions. Distinct micro- bial mats from different environments form distinct stromatolitic laminations.
P l a t e 52
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig. 8.
Stromatolitic laminae formation and carbonate precipitation associated with microbial mats from modem Bahamian environments
SEM photomicrograph of Dichothrix filaments heavily encrusted by micritic carbonate. These precipitates form stromatolitic laminae in the presence of Dichothrix dominated mats. Scale bar=50 lain Photomicrograph showing carbonate mineralization in the sheath of Dichothrix. These rhombohedral crystals probably were precipitated as a result of photosynthesis which increased the pH within the sheath. SEM photomicrograph of the two top layers of a freshly formed Dichothrix dominated stromatolitic structure. Because the grain layer is relatively thick, it shows clearly the tendency of cyanobacteria to migrate to the surface, whereby the sheaths are left behind (lower dense layer in the center). Photomicrograph showing a mature microbial mat dominated by the filamentous cyanobacteriaMicrocoleus binding the underlying sediment particles. The tiny white spots in the mat are floating precipitates of carbonate which appeared about seven days after the mat had formed. Photomicrograph showing how carbonate, which was precipitated within the microbial mat, begins to form a crust resulting in the formation of a stromatolitic lamina. SEM photomicrograph of the surface of a stromatolite from the lower intertidal zone. During subaerial exposure, cyanobacteria migrate below the surface crust as seen by the distribution of filaments. Note the abundant boreholes in the grains at the surface which are incorporated in the microbial mat. This endolithic activity could cause dissolution/precipitation processes which may lead to the rapid formation of a surface crust. Thin-section photomicrograph of carbonate grains incorporated in the micritic layer. The carbonate grains are often heavily bored and micritized, whereas the grains of the overlying detrital layer are basically pristine. Crossed nicols. Width of Fig.=l.5 cm SEM photomicrograph showing numerous unidentified microbes in the lower part of a microbial mat from a lower intertidal stromatolite from Lee Stocking Island. The background is organic material which was secreted by the filamentous cyanobacterium Schizothrix. Scale bar=-ll.tm

P l a t e 52 2 0 3

204
Part IV PRODUCTIVITY AND COMMUNITY COMPOSI- TION OF STROMATOLITIC MICROBIAL MATS
IN THE EXUMA CAYS, BAHAMAS
J.L. Pinckney and R.P. Reid
The microalgal community composition, productivity, and oxygen consumption of microbial matsfrom stromatolites in three different habitats in the Exuma Cays, Bahamas were measured to characterize the structure and function of these communities under different environmental con- ditions. Stromatolitic mats were collected from intertidal, subtidal channel, and subtidal bay habitats near Lee Stocking Island, Bahamas. Photopigment composition was determined by high performance liquid chromatography (ttPLC) and productivity was measured using 02 tech- niques. The microalgal community of stromatolitic mats in all three habitats was composed primarily of cyanobacteria with a minor contribution by diatoms. Phototroph biomass (nmol Chl acm "2) in intertidal habitats was nearly double that of subtidal habitats. 02 production was higher in the subtidal bay than intertidal and subtidal channel habitats. Biomass-normalized 02 production (tool 02 tool Chl a -1 h 1) was highest in the subtidal bay, intermediate in the subtidal channel, and lowest in the intertidal. Relatively high ratios of 02 production relative to consumption in all three habitats suggests that fixed carbon, in the form of biomass and extracellular polymeric substances (EPS), accumu- lates within the mats. Heterotrophic consumption of EPS may facilitate CaC O s precipitation and formation of micritic laminae preserved within stromatolites.
1 INTRODUCTION Microbial buildups ranging from stromatolites to
thrombolites are common in the Exuma Cays, Bahamas, occurring in a variety of environments ranging from inter- tidal to subtidal (DRAWS 1983; DILL et al. 1986; REIn et al. 1995). The Exuma stromatolites are characterized by millimeter-scale lamination with a characteristic micro- structure (REtt) et al. 1995; MAcncrva8 et al. 1996). This lamination reflects periodic formation of lithified micritic crusts within cyanobacterial mats on the surfaces of the stromatolites.
Lithification processes forming micritic laminae in Exuma stromatolites appear to be biologically-mediated (MAcn, rrVRE et al. 1996). Previous studies of microbial mats (e.g. PrNcr~EV et al. 1995b; STAL 1995; PAERL P ~ c r ~ v 1996) suggest that the cyanobacteriai mats on the surfaces of the Exuma stromatolites are likely to harbor a physiologically-diverse phototrophic and heterotrophic community that is tightly-coupled with biogeochemical gradients. A thorough understanding of the ecophysiology of modern stromatolitic mat communities will provide a critical link between biological processes and geological structures. In particular, identifying and quantifying the biologically-mediated processes responsible for forming micritic laminae in Exuma stromatolites may elucidate microbial processes responsible for the formation of an- cient (Precambrian) stromatolites. The purpose of this
research was to compare microalgal community composi- tion, productivity, and oxygen consumption of microbial mats from stromatolites in three different habitats in the Exuma Cays, Bahamas.
2 STUDY SITE The Exuma Cays form a NW-SE oriented chain of
islands on the eastern margin of Great Bahama Bank, Bahamas. Stromatolites occur in varying abundance, shapes, and sizes near Lee Stocking Island (23 47' N, 76 06' W), one of the Exuma Cays (DILL 1991; R~rD et al. 1995). Samples of cyanobacterial mats were collected from the surfaces of stromatolites in three habitats: 1. intertidal stromatolites on a sandy beach at the northwestern end of Lee Stocking Island (Site 14 in REID et al. 1995); 2. subtidal stromatolites in a tidal channel (Adderly Cut; 5-10 m depth) between Lee Stocking Island and Norman's Pond Cay (Site 1 in REID et al. 1995); 3. shallow subtidal stromatolites in a calm, sandy embayment (1-2 m depth) at the northern end of Little Darby Island (Site 11 in REID et al. 1995). In this paper, these three sites are denoted as intertidal, subtidal channel, and subtidal bay, respectively. Previous studies have determined that Schbothrix sp. is the dominant filamentous cyanobacterium in mats from all three sites (BRowN 1993; REID et al. 1995).
3 MATERIALS AND METHODS Several (>10) small sections (50 cm 2 x 2 cm) of mat
were collected on 3-6 June 1996 from each of the three habitats. Core samples (0.50 cm2x 0.5 cm) for photopigrnent analysis were obtained from the larger sections, stored in 2 ml microfuge tubes, wrapped in foil, and frozen. Photopigments were extracted in 1.5 ml of solvent (45% methanol, 45% acetone, 10% deionized I-LzO) following sonication (30 s) (BOwLES et al. 1985; PINCr,~rEY et al. 1994). High performance liquid chromatography (HPLC) techniques were used for photo-pigment identification and quantification (MtLHE et al. 1993; VAN I-IEtrr~LEr~ et ai. 1994; PINCKNEV et al. 1996). Pigment analyses allow esti- mation of the relative abundances of major microalgal groups, including cyanobacteria, diatoms, purple photo- synthetic bacteria, chlorophytes, dinoflagellates etc. (ROWAN 1989; P ~ e et al. 1995a,c). In this study,chemosystematic carotenoids and chorlophylls were used as indicators of the relative abundance of cyanobacteria and diatoms.
Photosynthetic measurements consisted of light and dark incubations of mat samples in ambient seawater (37~ Intact mat cores (6.0 cm 2 x 0.5 cm) were placed in clear plastic bags (1 pint Ziploc) and filled with seawater (100 - 300 ml) collected from Exuma Sound. Incubations were conducted outdoors under ambient irradiance in small pools continuously flushed with flowing seawater for temperature control. Neutral density screens (fiberglass mesh) were used to simulate in situ irradiances for the different habitat types. For dark incubations, containers were wrapped in 3 layers of aluminum foil and submerged in the incubation pools. All incubations were conducted between 10:00 am and 2:00 pm. Dissolved 02 was meas- ured at 30 min. intervals during incubations by withdraw-

205
Fig. 2. Microalgal photopigment concentrations and molar ratios for intertidal, subtidal channel, and subtidal bay stromatolitic mats. Values are the mean + 1 standard deviation. Letters (a, b, c) indicate results of means comparisons, with 'a' being the highest value and 'c' the lowest. Common letters indicate that the means were not significantly different (p < 0.05); absence of letters indicate no significant differences in mean values. All oxygenic phototrophs contain chlorophyll a and b-carotene; diatoms contain diatoxanthin and fucoxanthin; cyanobaeteria contain echinenone, myxoxanthophyll and zeaxanthin.
ing 500 I.tl of the incubation water from the sealed plastic bags using a gas-tight syringe. The partial p~essure of dissolved oxygen (pO z) was measured using a small vol- ume sample chamber (200 lal; Cameron Instruments BC202 Gas Cell and BGM 200 Analyzer) equipped with a Clarke- style oxygen electrode. O 2 production was defined as the net increase in [0 2 ] per unit time and equivalent to oxy- genic net primary productivity. 0 2 consumption was the net reduction in [0 2 ] per unit time and represents the sum of aerobic respiration and abiotic oxygen consumption.
Non-parametric statistical procedures (Wilcoxon Rank Sum W test, Kruskal-Wallis one-way ANOVA, =0.05) were used for comparisons of treatment groups. Means comparisons were achieved using a Bonferroni-type non- parametric procedure (=0.05) (NwrER et al. 1985).
4 RESULTS AND DISCUSSION The photopigment composition of microbial mats from
all three habitats was similar but concentrations differed (Fig. 2). Chlorophyll a (Chl R) and b-carotene (common to
all oxygenic microalgae), diatoxanthin and fucoxanthin (diatoms), and the cyanobacterial pigments echinenone, myxoxanthophyll , and zeaxanthin were the major photopigments in all mats. Chl a, b-carotene, echinenone, myxoxanthophyll, and zeaxanthin concentrations (nmol pigment cm -2) were significantly higher in the intertidal than subtidal mats. Photopigment concentrations of stromatolitic mats from the channel and bay subtidai habitats were generally not significantly different. The subtidal bay mats had, however, higher fucoxanthin con- centrations. Pigment molar ratios, which normalize pig- ment concentrations to total microalgal biomass (Chl ~), exhibited trends similar to the non-normalized concentra- tions (Fig. 2). One exception was that echinenone molar ratios did not differ among habitats. Fucoxanthin molar ratios were significantly higher in the subtidal than the intertidal mats. The molar ratio ofzeaxanthin to fucoxanthin (ZF ratio) measures the relative contribution of cyanobacteria and diatoms to the total accessory photopigment pool in these communities. Differences in this ratio indicate the

2 0 6
Fig. 3. Stromatolitic mat O 5 production (photosynthesis) and consumption, Chl_a - specific O 2 production and consumption, and the ratio of 02 production to consumption. Values in the lower two panels are the mean + 1 standard deviation. Values in the upper panel were calculated using pooled data for each habitat type. Letter designations are the same as for Fig. 2.
relative photosynthetic contribution of cyanobacteria and diatoms. The ZF ratio ranged from 4.26 + 1.21 (mean _+ 1 standard deviation) for subtidal bay mats, to 5.90 + 2.86 for subtidal channel mats and 21.42 + 13.50 for intertidal stromatolitic mats. Intertidal mats had a significantly higher ZF ratio than subtidal (channel and bay) mats.
The photopigment data indicate that cyanobacteria and diatoms were the major microalgal groups in stromatolitic mats from all three habitat types. However, the area- specific photopigment concentrations, relative photosyn- thetic contribution, and relative abundances ofcyanobacteria and diatoms differed with habitat type. Intertidal mats had nearly double the total microalgal biomass (Chl ~ com- pared with subtidal channel and subtidal bay stromatolites. b-carotene, a potential photoprotective and antioxidant pigment, was higher in the intertidal mats and may provide a necessary 'sunscreen' for protection from exposure to high irradiances. The cyanobacterial pigments (echinenone.
myxoxanthophyll, zeaxanthin) mirror changes in Chl a concentrations for intertidal mats and provide evidence that the phototroph community is composed primarily of cyanobacteria. The molar ratios of cyanobactefial pig- ments indicate a decrease in the relative contribution of cyanobacteria with increasing water depth. The higher ZF ratio for intertidal mats suggests that diatoms play a greater role in community composition and primary productivity in the subtidal stromatolitic mats than in the intertidal mats. In all three types of mats, however, diatom abun- dance was low compared to microbial mats from other areas. For example, ZF ratios for intertidal microbial mats in North Carolina (USA) range from 0 - 2 ( l~Ncr~u et al. 1995c), considerably lower values than the ratios of 4 - 21 in Ex uma stromatolitic mats. The role of diatoms as major contributors to overall metabolism in Exuma mats seems to be small relative to the m uch more abundant cyanobacteria. Oxygenic phototroph production (net primary productiv-

207
ity) was significantly higher for the subtidal bay mats (327 + 55 nmol 02 cm -2 h "1, mean + 1 SD) compared to intertidal ( 188 + 73) and subtidal channel (168 + 42) habitats (Fig. 3). O 2 consumption averaged 67.3 + 22.5 nmol O z cm 2 h -I for all three habitat types. Biomass (Chl ~ normalized 02 production and consumption showed similar trends, with subtidal bay mats exhibiting significantly higher rates [69.2 + 33.7 moles O 2 (moles Chl a) -1 h -1] than intertidal (18.8 + 8.1) and subtidal channel (38.9+ 14.8) stromatolitic mats. Chl a-specific O 2 consumption was significantly lower in the intertidal mats (7.00 + 1.73) compared to the subtidal channel (18.1 + 11.3) and subtidal bay (14.5 + 3.85) stromatolites. In addition, the ratio of mean O 2 production to consumption ranged from 2.2 to 4.8 and was higher for the subtidal bay mats.
The 02 production and consumption data confirm that all three mat types were actively growing with substantial net primary productivity. The higher microalgal produc- tivity in the subtidal bay mats can be attributed to both a higher biomass and higher photosynthetic efficiency (Chl a-specific production) relative to intertidal and channel mats. Because all mat types had similar 02 consumption rates, net carbon production should be highest in the subtidal bay mats. In all mat types, the high production to consumption ratio (>2 during the daylight period) suggests that fixed carbon, in the form of biomass and extracellular organic substances, accumulates within the mats. Similar results have been reported for intertidal stromatolitic mats on Stocking Island, Bahamas (P~Cr~EY et al 1995b). The metabolic products of excess production in nutrient-lim- ited environments may be secreted as extracellular poly- meric substances (EPS) that effectively bind Ca a§ ions (DEcHO 1990). EPS degradation by bacterial heterotrophs may result in the release of Ca 2§ ions that lead to localized (microscale) high concentrations of Ca 2§ which facilitate CaCO 3 precipitation. This mechanism may possibly ex- plain the microbially-mediated formation of micritic lami- nae in modern-day stromatolites.
Several reports (AwRAremc & RmtNO 1988; RmlNC et al. 1991; Rm~o 1994) have suggested that Exuma stronmtolites are constructed by eukaryotic microalgae (diatoms, chlorophytes, etc.). In contrast, our results indicate that the stromatolites are primarily a product of cyanobacterial (prokaryotic microalgae) growth. In particular, the photopigment data indicate a lack (below detection limits) of microalgal groups other than cyanobacteria and minor diatoms in the surficial mats. In addition, micro- autoradiography of similar mats from Exuma stromatolites in other localities indicate that cyanobacteria are the major COz-fixers in these communities (H. PAERL unpublished).
The photosynthetic component of the stromatolitic mats was the primary target of our investigation. The role of heterotrophic (non-pigmented) bacteria in growth of Exuma stromatolites may, however, be highly relevant, especially with respect to CaCO 3 precipitation and the formation of micritic laminae (e.g. KRU~mEIN et al. 1977; CHAF~rZ & BtJczwsra 1992). A more complete under- standing of the metabolic product exchanges in consortial relationships (PAERL & P t N ~ y 1996) between phototrophs
and heterotrophs may lead to new insights into the ecophysiological factors that regulate stromatolite forma- tion and growth in Bahamian waters. Research into resolv- ing the linkages between heterotrophic and phototrophic mat community components is currently in progress.
Part V HIGH RESOLUTION
CRYO-SCANNING ELECTRON MICROSCOPY OF MODERN STROMATOLITES
(LAKE R2, RANGIROA ATOLL, TUAMOTU ARCHIPELAGO, CENTRAL PACIFIC OCEAN)
J. Trichet and C. Dtfarge
Awlls in the Central Pacific are the seat of accumula- tion of significant amounts of microbially ( cyanobacterially and bacterially) derived organic matter. Such deposits, purely organic during the early steps of their evolution, are progressively mineralized through the authigenic pre- cipitation of carbonate minerals, essentially high-Mg cal- cite. The successive deposition of organic laminae and the preferential development of calcite within certain laminae give a stromatolitic facies to the sediment.
Transmission and conventional Scanning Electron Microscopies revealed that the organic part of the sedi- ment looks like a honeycomb network resulting from the re-organization of the polysaccharide fibers which build the external sheaths of cyanobacterial filaments. After their dispersion (following the death of the microbes), those fibers assemble under such an original organic network delimitatin g alveoli surrounded by polysaccharide walls.
Refined pictures of the microstructure of cyanobacterial sheaths and of the incorporation of their constituting fibers into the sedimentary organic network are given in this paper. Those pictures were obtained by a Field Emis- sion Scanning Electron Microscope equipped with a freeze- drying sample preparation system. This method is well adapted for the study of hydrated,fragile organic samples. The problems addressed by the nucleation and growth of calcite minerals within such a network are evoked at the end of the paper.
1 INTRODUCTION Present atoll-type coral reefs in the Central Pacific
provide environments which are the seat of accumulation of significant deposits of microbial sediments. These sediments proceed from cyanobacterial biomasses and are the seat of authigenic carbonate precipitation, essentially under the form of high-Mg calcite. The sediment is lami- nated, from amm to acm scale, the lamination resulting from the superposition of successive organic red-coloured (due to the presence of carotenoidic pigments) laminae or from the differential deposition of carbonate within certain organic laminae (under the form of more or less continuous whitish layers).
The mean size of the sedimentary basin in which such sediments accumulate varies between some meters or tens

208
of meters (as pools or small lakes on the atoll surface), up to some kilometers as in Niau atoll (in the Tuamotu Archipelago, French Polynesia), where the microbial de- posit occupies the whole lagoon of the atoll (approxi- mately 9 km in diameter). Native people in the Tuamotu Archipelago call these sediments 'kopara'. The geomorpho- logic situation, the global sedimentary and geochemical features of kopara deposits are presented in DI~FARGE et al. (1994).
2 RECENT DATA ON THE MICROSTRUCTURE OF KOPARA DEPOSITS
Early observations on the microstructure of kopara deposits provided evidence that the organic sediment was remarkably structured under the form of a honeycomb-like framework (PI. 53/1) in which the walls of the unit-cells are essentially polysaccharidic and, to a lesser extent, polypeptidic (Dt~ARGE et al. 1994, 1996).
2.1 Materials and methods The aim of this work has been to take benefit of the
coupling of a Field Emission Scanning Electron Micro- scope (FESEM) and of a freeze-drying sample preparation system (cryosystem) in order to have a new and deeper insight in the microstructure of the organic constituents of kopara sediments. FESEM has over conventional SEMs the advantage of far higher resolution at low voltage processing. This allows a better observation of the surface of the samples, under less destructive conditions.
The freeze-drying cryosystems involve the ultra-rapid freezing of the pore water of the sample (at -210~ in nitrogen slush), followed by a sublimation of the superfi- cial ice (by increasing temperature up to -90 ~ to -70~ This rapid quenching results in the solidification of water without significant volume increase (Gut~ 1991), i.e., without a mechanical tear of the fragile organic tissues at the scale of observation.
Field Emission cryo-scanning Electron Microscopy has given the pictures of the kopara organic network presented here. Pictures given in PI. 53 are from metal- coated samples (Au or Au/Pd). The samples studied in this work have been collected in lake R2, on Rangiroa atoll (Tuamotu Archipelago, French Polynesia), in the highly red-coloured top 10 centimeters of the deposit (D~vARGE et al. 1994).
2.2 Results PI. 53 illustrates the microstructure of a bulk piece of
kopara sample (PI. 53/1) and of dead cyanobacterial sheaths (PI. 53/2-4). The honeycomb-like structure of the organic sediment is clearly evidenced. Mean diameter of the al- veoli is ca. 5 I.tm (PI. 53/1). This network encloses remains of cyanobacterial sheaths. The external limit of the emp- tied sheath is either sharp and well delineated (top right part of Pl. 53/1) or less well delineated (as evidenced in the two specimens visible in the center of the same picture). PI. 53/2-4 show the structure of emptied sheaths of cyanobacterial filaments. PI. 53/2,4 show that the sheaths themselves are made of an organic network presenting a high porosity and a fibrous polysaccharide texture. The fibrous polysaccharide structure of the network has been evidenced by MOSr~ER-ARNoU & TRIcrm'r (1979) and I~AR~E et al. (1994) by using the polysaccharide staining tech- nique of THrERRY (1967) on ultrathin sections of samples then observed by Transmission Electron Microscopy. Such observations proved that the bulk honeycomb-like net- work of kopara results from the reorganization of the polysaccharide fibers of cyanobacterial filaments. After the death of the cyanobacteria, their sheath undergoes a physical dispersion, which is followed by the reorganiza- tion of the sheath-constituting fibers (D~ARCE et al. 1994). The progressive passage from well-delineated to poorly- delineated sheaths to the honeycomb structure can be understood as the result of those processes.
3 DISCUSSION AND CONCLUSION The enhanced resolution of the FESEM microscope
helped to depict the microstructure of the organic network which constitutes the kopara microbial sediments. As visible in the top left part in P1.53/2, the walls of the alveoli are made of a porous (at tens of nm scale) framework resulting from the interwoving of polysaccharide fibers. Such a structure is similar to that of the cyanobacterial sheaths themselves, as shown in PI. 53/2,4. PI. 53/2 illus- trates the transition stage between the porous polysaccharide tissue of a sheath and the organic network. The formation of the bulk organic matter is therefore essentially the result of a reorganization of polysaccharide fibers initially asso- ciated within the filament sheaths and finally within al- veoli walls. The fibrous constituent and the porous struc- ture of the sheaths are strikingly similar.
P l a t e 53
Fig. 1.
Figs. 2.-4.
Microbial kopara sediment collected in Lake R2 on Rangiroa Atoll (Tuamotu Archipelago, French Polynesia). These micrographs have been obtained thanks to a Field Emission Scanning Electron Microscope Philips XL 30 FEG equipped with an Oxford CT 1500 HF sample cryopreparation system (courtesy of Philips S.A.).
Microstructural organization of a bulk organic portion of the sediment. The picture shows (top right part) well delineated emptied cyanobacterial sheaths (whose structure is similar to those magnified in Figs.2- 4), and, in its bottom central part, the incorporation of the polysaccharide constituents of the sheaths into the network. Enlarged views of emptied cyanobacterial sheaths enclosed in the bulk organic network.

P l a t e 53 2 0 9

210
Two other data can be added to the previous ones. One refers to the mutual arrangement of individual polysaccharide fibers within the wall of the alveoli of the bulk organic network. The observation of sections of these walls under polarized light shows that the fibers are twisted within liquid-crystalline structures (MArt J F. 1984). The other data deals with the relationship between the nucleation and growth of high-Mg calcite in relation with the organization of the organic network. Calcite crystals form in individual alveoli and nucleate at the surface of the wall of the alveoli. The best pictures of the contact between the organic wall and the first calcite crystals suggest that the latter grow in a direction approximatively perpendicular to the wall's surface (Da-FARO~ et al. 1994).
A relation between the fibrous twisted structure and chemistry of the polysaccharide surface of the wall and the nucleation and growth orientation of carbonate can be anticipated. Further studies are needed to get deeper in- sight into this relation.
Part VI ORGANIC MATTER IN GREAT SALT LAKE
OOIDS (UTAH, USA) -
F I R S T APPROACH T O A FORMATION VIA ORGANIC MATRICES
J. Reitner, G. Aria, V. Thiel, P. Gautret, U. Galling and W. Michaelis
The aragonitic ooids of the Great Salt Lake (Utah, USA) are a product of an organomineralization process. The comparative study of ooids, microbialitic ooid chips, and microbialite crusts demonstrates the crucial role of proteic and glucidic substances in the formation of ooids. The ooids contain strikingly high amounts of intracrystalline organic matter (up to 528 ltg/g carbonate), which consists of about 23% proteic and 77% glucidic material. High values of acidic amino acids (12 mole-% asp and 45 mole- % glu) characterize the proteic phase, whereas mono- saccharids are enriched in fucose, glucose, and mannose. Fluorescence microscopy of histochemically treated ooids reveals distinct dissolution/reprecipitation features prob- ably controlled by the acidity of the organic matter in- volved. Direct influence of the few attached bacteria is negligible, so that our model of ooidformation refers to the free organic substances. We propose, that the free organic substances of ooid surfaces first inhibit carbonate precipi- tation due to their very strong acidity. Increasing neutrali- zation of the free carboxylic and sulfate-groups finally results in a retarded precipitation of aragonite fibres and micrite. Therefore, the ooid fabric is most likely controlled by small-scale biochemical gradients and not by clay minerals, as previously assumed.
GC-MS analyses of the hydrocarbons extracted from the ooids reveal high concentrations of n-alkanes derived from terrigenous sources as well as long-chain mono- methylalkanes. The latter compounds are typical constitu- ents of insect waxes and most likely originate from the salt
fly Ephydra living in the area. In comparison to the hydrocarbon pattern of the microbialitic ooid chips, only small amounts of biomarkers deriving from heterotrophic bacteria and cyanobacteria were observed in the ooids.
1 INTRODUCTION The Great Salt Lake (GSL, Fig. 4) is a relic of the
evaporated Pleistocene Lake Bonneville. Today it is 130 km long and 57 km wide with a maximum water depth less than 15 m. Due to the very gently inclined basin slopes even minor variations in lake level cause considerable shifts of the shoreline.
The lake water is slighdy alkaline, but highly saline with Na+ and CI- as major ions. The salinities vary between 6 and 32%, depending on the influx/evaporation ratio. The North Arm of the lake has an increased salinity in compari- son to the South Arm which receives -90% of the freshwa- ter inflow of the entire lake basin (Fig. 4).
Since the classical scientific work of EARDLEY (1938) and CAROZZl (1962) the lake is regarded as an actualistic scenario for ancient salt lake microbialites ('stromatolites') and large radial ooids resp. oolites. The oolite story was intensely discussed by LOREAU (1969), BArmJRST (1971), KArILE (1974), and SANDBERG (1975), who has established a general model of oolite formation and diagenesis.
The main goal of this paper is to evaluate the role of organic matter during the formation of the GSL ooids in comparison with the microbialites. Our approach is based on the organic matrix theory (DEoENS 1976; WEIrc~n~ et al. 1983 a.o.) which explains the growth of calcium carbonate
Fig. 4. Outline of the Great Salt Lake, Utah, showing the sampling localities. A: Bridger Bay, Antelope Island; B: Lagoon NW of Promontory Point

211
crystals in the presence of free acidic organic macromol- ecules (RmTr~R 1993; RmXr~R et al. 1995; RFaTNER & NEUWEItaR 1995). In marine ooids, remains of these or- ganic matrices were found by TPactmr (1968) and MrrrERER (1968). S~ss & FOrra~R (1972) have shown by experi- ments that organic matter rich in carboxylic groups, e.g. humic acids, may form artificial aragonite ooids.
GSL ooids, microbialitic ooid chips, and microbialites strongly resemble to Lower Triassic (Scythian) 'Rogenstein- Fazies' of central Europe (KALKOWSKY 1908; PAUL & PERVr 1985). Therefore the GSL serves as a modern example for vast ancient closed hypersaline lake systems in add conti- nental settings combined with continous influx of ground- and river waters.
2 M A T E R I A L A N D M E T H O D S The ooid samples were collected during a field cam-
paign in September 1994 (JR) from the northern west coast of Antelope Island (South Arm). In May 1996, ooid and microbialite samples were taken at Bridger Bay, Antelope Island, and from a lagoon NW of Promontory Point (North Arm) by GA & VT.
The samples for histochemical studies were fixed in buffered formol, dehydrated and stored in 70% ethanol. For electron microscopy, 8 samples were fixed in buffered glutardialdehyde (cooled on ice, 24 h) and post-fixed with 2% OsO 4 (for details see RErn~R 1993). SEM studies were carded out with a LEO Gemini field emission SEM, which allows an investigation of uncoated samples at extremely low beam energy (below 1 kV). Some samples were investigated by using an Oxford-Cryosystem in combina- tion with the field emission SEM. This method allows investigations of organic matter without dehydration arti- facts and collapse structures. The TEM studies were car- ried out with a Zeiss EM 10 and a Jeol 100 B.
Cyanobacterial genera were determined by using fixed, sectioned, as well as living specimens following GEITLER (1932), KOMAREK & ANAGNOSTIDIS (1986) and WATERBURY (1992). Microbial populations were taken in culture to investigate the cyanobacteria and bacteria. In situ staining experiments with the Ca2+-chelating fluorescent dyes (calcein, tetracycline) were carded out to localise the growing zones of the ooids and microbialites. Addition- ally fixed specimens were blockstained with fluorescent and non-fluorescent dyes (acridine orange, toluidine blue O, basic fuchsine). Sections were prepared by cutting LR- White-embedded, non-decalcified specimen with a hartpart microtome. Some specimens were decalcified, sectioned, and stained with Alcian VBlue, LFB-PAS and Gram.
The samples for biochemical analyses were sun dried. The superficial organic substance of the samples was removed by Na-hypochlorite, so that macromolecular analy- ses only cover the carbonate phase. For decalcification the ooids were put in pH 4 controlled acetic acid for 24 hours. The insoluble fraction (high contents in clay minerals) was removed by centrifugation. The remaining organic matter was desalted by low-pressure gel filtration chromatogra- phy on PHARMACIA-G25C with UV detection (280 n m). Amino acids: 24 hrs, 110~ 6N HCI hydrolysis; PITC
derivatization; HPLC reverse phase chromatography (BECKMAN system) with Hypersil 100 column (C18 51am 250x4.6mm); UV detection 245 nm. Monosaccharides: 6 hrs 2N TFA hydrolysis; HPLC Ion exchange chromatog- raphy (DIONEX system) with CarboPac PAl(250x4mm); pulsed amperometric detection. Molecular weights: HPLC gel filtration using a TSK G2000SWXL column eluted by sodium citrate adjusted to pH4.5 (0.33M NaCI added); UV scan detection (LDC Analytical-Spectro-Monitor 5000 photodiode array detector); LKB differential refractometer. Amino acid and monosaccharide analyses were carried out on the total Soluble Organic Matrix (SOM), after desalting, ultra filtration against MilliQ water (FILTRON ultrafiltration cells, 3K), and lyophilization. Amino acid composition was tested again on separated fractions collected during HPLC gel filtration used as a preparative fractionating system. Previous to the usual hydrolysis and derivatization procedures, fractions were vacuum concentrated, ultrafiltred against MilliQ water, and lyophilized.
Hydrocarbon investigations were performed on a microbialite, ooid sands (total sediment; surface layer, 0- 1 cm), purified ooids, and a deposit largely consisting of salt fly pupas (Ephydra sp.). The samples were air dried immediately after collection and sealed for transport to- gether with a large amount of drying agent. Prior to solvent extraction, the sediment samples were cleaned by washing with diluted HCI and acetone. Purified ooids were ob- tained by repeated washing of the total sediment and decanting the wash water. Subsequently, the carbonate matrix of each sample was slowly removed by dropwise addition of 6N HCI. The residues were washed with distilled water, centrifuged, and ultrasonically extracted with dichloromethane/methanol (3:1; v:v). Hydrocarbons were separated from the total organic extracts by silica gel column chromatography using n-hexane as elution agent. Further analyses were carried out with gas chromatogra- phy and combined gas chromatography/mass spectrometry as described by HEFTER et al. (1993).
3 RESULTS 3.1 Histochemical characterist ics of ooids
The histochemical studies using Alcian Blue and PAS stainings of the ooids indicate an enrichment of free COO- groups of the acidic glucidic mucus on the surfaces and in small depressions of the ooids. Most of the ooids have a mean size of 300 - 1000 I.tm and they exhibit a radial fabric of large aragonite crystal bundles (palisades) (PI. 54/1-6). This pattern is sometimes interrupted by concentric layers of micrite with an irregular or modified spherulitic ar- rangement of small aragonite crystal needles (1-2 I.tm) (PI. 54/4). Depressions or pockets between the large radial crystals are filled with an aragonitic micrite (cerebroid ooids) (P1.54/4,6). The large aragonite crystals often show dark colored tangential growth-lamellae. Not all ooids exhibit these complex features but may solely consist of large radial crystals or concentric rings ofaragoniticmicrite, respectively. Most ooids have a large nucleus of a quartz grain or other detritus (PI. 54/1-3,6). Of special interest are the depressions between the aragonite palisades, because

212
Fig. 5. Amino acid and monosaccharide compositions of the soluble organic matrix extracted from aragonitic oolites
the acidic organic mucus is enriched within these spaces (PI. 54/1,2). The remaining surface is covered by a thin organic film (P1. 54/4,5) with portions of a true biofilm composed of small (500 nm) red-shaped bacteria. The organic matter of the depressions is characterized by distinct fluorescence properties when using the Ca 2§ -chelating calcein and acridine orange (PI. 54/1,2). Calcein is used as a Ca 2§ detector in acidic organic matter and its fluores- cence intensity reflects the concentration of free Ca 2§ which is in this case dependent on the degree of neutralized carboxyl-and/or sulfate-groups. The upper portions of the organic mucus show a typical orange color (acridine or- ange). The bottoms and the margins of the depressions exhibit a green-yellow fluorescence showing the begin- ning nucleation of smaU CaCO 3 - crystals (PI. 54/2). Under crossed nicols the resulting anhedral seed crystals are easily seen because of a strong birefringance. This calcifi- cation process results in a characteristic aragon itic automicrite (PI. 54/4). It seems that the tips of the large palisade crystals exhibit dissolution fabrics (PI. 54/4,5). This fea- ture may be explained by the highly acidic character of the upper portion of the organic matter which dissolves some parts of the palisade crystal. At the margins of the interme- diate zone of the organic mucus, epitactical reprecipitation of the CaCO 3 starts again on the tips of the palisade crystals (recycling process). In some cases the automicrites are prograding over the palisades forming the concentric lay- ers.
This process is not yet fully understood but it is prob- ably dependent on the biochemical properties of the or- ganic mucus and the saturation with Ca 2§ and HCO3-. SEM studies have shown that small biofilm patches occur inside the depressions together with the acidic organic mucus. However, microbes play only a minor role for the entire organic matter and are difficult to detect. Most of the small bacteria (500 nm-I I.tm) are gram-negative. Only few show a gram-positive staining (perhaps proteolytic or saccharolytic microbes). Cyanobacteria are rarely present on the ooids.
3.2 Macromolecular content in ooids The soluble organic matter (SOM) of the ooids exhibits
some remarkable characteristics. We have measured 528 t~g/g carbonate SOM, which is relatively high. Proteins account for only less than 25% of the soluble matrix which is strongly glucidic, as deteted with histochemical meth- ods. The relative amino acid composition is remarkable because of the very high content in glutamic acid (45%). Other amino acids are the glycine (14.66%), aspartic acid (12.9%), alanine (7.3%), serine (4.8%). Relative mono- saccharide composition is dominated by mannose (26.33 %, incl. xylose), glucose (18.13%), and fucose (9.27%). Other monosaccharides are also well represented with concentrations ranging from 6 to 10% (Fig. 5).
HPLC gel filtration shows no very high molecular weight compounds elufing at the elusion limit of the column (100kD). Heaviest fraction (HMW F, Fig. 6, ap- parent molecular weight 80-90kD) elutes as a composite mixture of badly resolved compounds with a very strong refractive index and also a strong UV absorption at short wave lengths from 225 to 240 nm, but a weak UV absorp- tion at 280 nm. These characteristics are typical for glucid- rich high molecular weight compounds (Cur~ & GAUTRET 1995). The low molecular weight fraction (LMW F) cor- responds to a more or less continuous arrival of small molecules detected by UV absorbance only suggesting that no glucides are covalently bound.
Amino acid compositions of each of these two frac- tions were determined (Fig. 6; graphs on the right). Amino acids represented by black bars on the graphs are those exhibiting clearly distinct concentrations in the two mo- lecular weight fractions. Most of the glutamic acid which is the highly dominant amino acid of the soluble matrix is clearly allocated to the Low-MW fraction as well as the aspartic acid with a lower concentration. This should make these small proteins or polypeptides very acidic. On the opposite the glucide-rich High-MW fraction is dominated by the glycine and it contains high amounts of serine presumably serving as the attachment residue for O- glycosylation.

213
Fig. 6. HPLC gel filtration fractionation of the soluble organic matrix with UV (280rim) detection and refractive index (RI) slzongly indicative for the presence of covalently bound glucides. High molecular weight fraction (HMW F) and low- (LMW F) were collected separately for determination of their anaino acid compositions (histograms on the right : black bars correspond to the main distinctive compounds)
HPLchromatography is in accordance with the histochemical dataand shows clearly that the intracrystalline organic matter (SOM) is a mixture of various organic remains. The sugar-rich HMW fraction of the SOM is related to remains of EPS of biofilms (polysaccharides). The origin of the LMW fraction with the high content of acidic amino acids is still unclear.
3.3 Biofilm composition on microbialites During the sampling campaigns in September 94 and
May 96 the classical microbial bioherms (cf. EARDLEY 1938; CAROZZI 1962) were completely submerged and hardly accessible from the lake shore (lake level 1280 m). Inactive, subfossil bioherms have been visited by the co- authors (GA, VT) NW' of Promontory Point, only. In September 1994 chips of ooid sand ('shingles' of EARDLEY 1938), which were lithified by a thin microbialitic crust, were sampled at the NW coast of Antelope Island. Their surface is covered by a green biofilm of Gloeocapsa, showing a distinct and thin mineralization zone at its base (PI. 54/7). This type of ooid chip is designed in here as 'microbialitic ooid-chip'. Microbialitic carbonate of these chips comprises a thin laminated veneer, which exhibits a fabric strikingly similar to that of ooids, and micritic aragonite precipitates between the ooids within the chip. Apart from that, we recognized in May 1996 mm-thick carbonate crusts on cobbles at Bridger Bay, Antelope Island, in 30-50 cm water depth. Bridger Bay has been intensively monitored for microbialites in September 94
(GA), when the lake level was approximately 50 cm lower. At that time no carbonate crusts were found and the cobbles were free of encrustations. Pm~orcE & FOLK (1996) apparently recognized the same crusts at Stansbury and Antelope Island and argued that they must have been formed since 1983. Our observations indicate an age of less than 2 years for these still growing crusts!
The crusts are up to 9 mm thick, show a knobby surface, and are best developed on lateral and top sides of the cobbles. The lower part of the cobbles lies in ooid sand which is already blackened few mm below the sediment surface by sulfate reduction. The microstructure of the crusts is characterized by a clotted fabric which results from irregularly precipitated anhedral aragonite crystals of micritic size. Aragonite needles are restricted to few intercalated layers, giving the uppermost part a rough stromatolitic appearance. Trapped quartz grains are scat- tered throughout the crusts, whereas ooids are restricted to pockets.
The dark-green to yellow-green biofilm covering the crusts is in average 250-600 ~tm thick. The biofilm is primarily composed of coccoid cyanobacteria of the genus Gloeocapsa (PI. 54/10,11). Pennate diatoms are only fre- quent in depressions of the biofilm. Sections exhibit ir- regular Gloeocapsa-colonies with ceils grouped by subse- quent fissions in three planes (PI. 54/11). Their layered sheaths can be well stained with toluidine blue. Locally the coccoidcyanobacteriumAphanothece (PI. 54/9,11) ispresent between the Gloeocapsa colonies, too. Their red-shaped

214
cells show diffluent sheaths and divide by binary fission in one plane. 'Aphanothece packardii' (syn.: Coccochloris elabens, FELIX & Rusrn~oRava 1980) was repeatedly re- ported from the GSL to form the extended bioherms (EARoLEY 1938; CAROZZI 1962; Gwa'rr & MuRpaV 1980), but at the present conditions (May 1996), Gloeocapsa is obviously the prevailing genus in calcifying biofilms of the aragonite crusts (P1.54/7,11). Both genera are growing at laboratory on agarplates in association with bacteria and filamentous cyanobacteria, but their pure isolation has failed until now for an unknown reason.
Semi-thin sections show that numerous bacteria are associated within the Gloeocapsa-biofilm. The cyano- bacterial sheaths appear to be essentially free of bacteria, but in fissures and pockets between EPS-encapsulated Gloeocapsa-cells, bacterial microcolonies are common (mainly rods). Filamentous bacteria are common on top of the biofilm. The base reveals patches of diverse bacteria as well, thriving on decaying and lysed cyanobacterial cells. The resulting organic mucus forms a narrow zone or irregular patches which display traces of carbonate crys- tals (transition to massive carbonate crust below). Identi-
P l a t e 54
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.-5.
Fig. 6.
Fig. 7.
Fig. 8.
Fig. 9.
Fig. 10.
Fig. 11.
Recent growing ooids, microbialitic ooid chips, and microbialite crusts of the Great Salt Lake, Utah.
Survey of ooid morphologies. Some ooids are entirely radially structured, some show alternations of tangentially and radially structured layers. Quartz grains and faecal pellets serve as nuclei. Note reddish stained superficial organic substance of the still growing ooids (arrows). Loose ooid sand from the NW coast of Antelope Island, South Arm. Sample GSL 94/1, stained with acridine orange, bright field. Scale bar = 400 lain. Growing ooid stained with acridine orange. Retarded calcification within the depressions proceed at the green-yellow seam (arrow), which contains small CaCO3-crystals. Palisade crystals between organic depressions grow epitactically. Sample as Fig. 1., epifluorescence (UV excitation: BP 365/11, LP 397). Scale bar = 50 tun. Radially and tangentially structured ooid showing the strong autofluorescence of depressions (of two different layers). Opaque dots are iron sulfide (arrows), which is a result of sulfate reduction. Cemented ooid chip from a NaCl-saturated lagoon NW of Promontory Point, North Arm. Sample GSL 96/B4, epifluorescence (blue excitation: BP 450-490 nm, LP 520 nm). Scale bar = 100 lxm. Bacterial microcolonies (arrow) attached to depressions of an ooid surface. Dissolution and reprecipitation occur simultaneously adjacent to each other. Note dissolution features of palisade crystals. Reprecipitated aragonitic micrite (a) exhibits a strong autofluorescence (fig. 5). Surface of sample Fig. 3.; Fig. 4: phase contrast, Fig. 5: epifluorescence (blue excitation: BP 450-490 nm, LP 520 nm). Scale bar = 50 lxm. Predominantly radially structured ooid trapped in a halobacterial mat from the North Arm. Several layers of the ooid show fluorescent organic matter within the aragonite of depressions. Wet ooid sandflat NW of Promontory Point, North Arm. Sample GSL 96/B2, epifluorescence (UV excitation: BP 365/11 nm, LP 397 nm). Scale bar = 200 Ixrn. Calcein-stained Gloeocapsa-biofilm (a) of a microbialitic ooid-chip (b) from the NW coast of Antelope Island. The ooid chip is veneered by a fibrous carbonate crust (c), which results from a narrow mineralization zone at the base of the Gloeocapsa-biofilm. Consequently, this type of ooid chip reveals significant amounts of cyanobacterial biomarker. Increased free Ca2§ at the mineralization zone is labeled by calcein-Na 2, indicated by the yellow fluorescing seam (arrow). Sample GSL 94/5, stained with calcein-Na.2, epifluorescence (blue excitation: BP 450-490 rim, LP 520 nm). Scale bar -- 100 ktm. Bisaccate pollen of plants and Artemia eggs are trapped between ooids of a chip. Following degradation the plant material and arthropod waxes are enclosed in the carbonate of the chips, resulting in a corresponding biomarker spectrum. Sample GSL 94/B4, epifluorescence (blue excitation: BP 450-490 nm, LP 520 nm). Scale bar --- 200 lxrn. Aphanothece from Gloeocapsa-dominated biofilms of Bridger Bay, Antelope Island. The rod-shaped cells show binary fission perpendicular to the long axis. The diffluent sheaths are hardly visible. Sample GSL 96/A6, phase contrast. Scale bar = 20 lxm. Gloeocapsa from Gloeocapsa-dominated biofilms of Bridger Bay, Antelope Island. The spherical cells are arranged within a common sheath by subsequent fissions in three planes. Same sample as in Fig.9., Differential interference contrast. Scale bar = 20 pan. Semithin section of a Gloeocapsa-dominated biofilm, which forms the carbonate crusts on cobbles at Bridger Bay, Antelope Island. Rod-shaped and filamentous bacteria (a) are abundant on top and within the Gloeocapsa-biofilm. The sheaths of Gloeocapsa (b) and (to a lesser extent) of diatoms (c) are stained by toluidine blue. Note that the EPS matrix of degraded and lysed cyanobacteria and bacteria at the biofilm base is packed with (carbonate) crystal traces (d). Sample GSL 96/A6, decalcified, semithin section stained with toluidine blue, phase contrast. Scale bar = 20 Ixm.
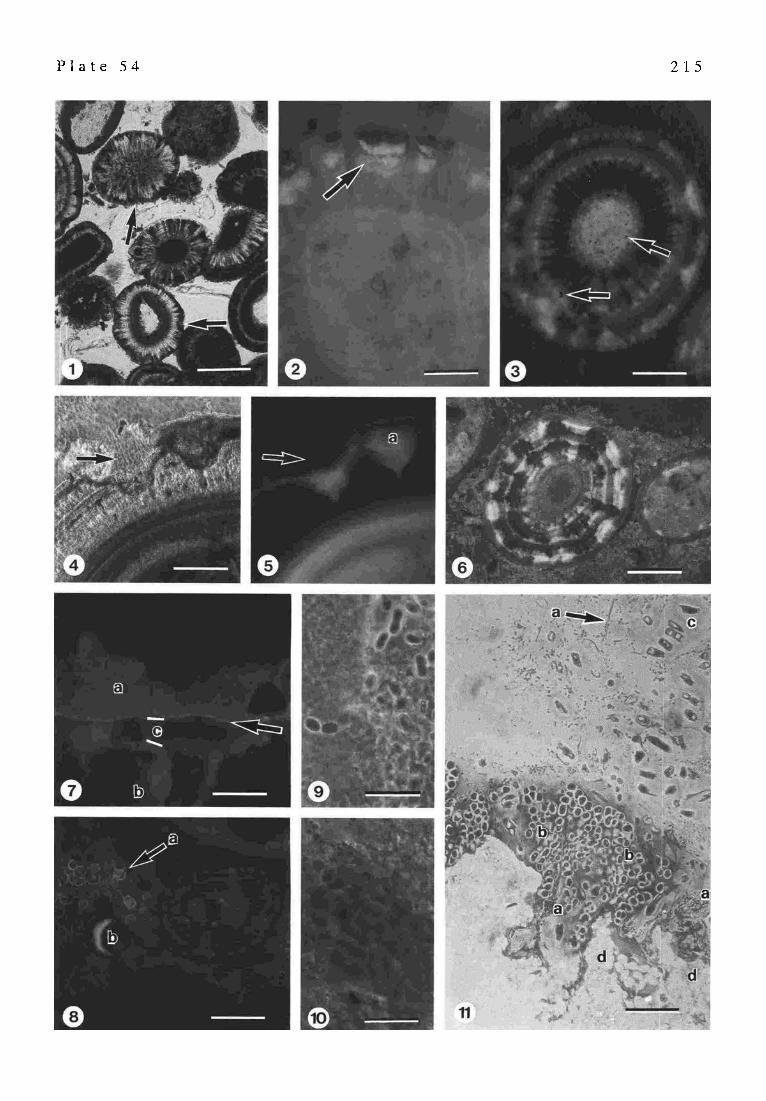
P l a t e 54 2 1 5

216
t-
Ooid sands (total sediment)
Time
29
I
.11 17 �9 I , MMA
l ' " l l J l " " ? __.? ,.tl.#..~-- . . . a l .i__j__ . _ j _ , ] . I d . ,
29
P u r i f i e d o o i d s
17 "~ ~Pr ? = r / � 9 v
_= Time
t--
c-
Ephydra pupas
Time
" , T T I . . [ M M A
1~_ 1!_~�9 ~?,t - . . . . . .
MMA
29
�9 J �9 �9 I . . r . , , I _._T.,
symbols and abbreviations �9 n-alkanes (carbon numbers denoted) �9 mid-chain branched monomethylalkanes i , n-heptadecenes
MMA long-chain monomethylalkanes
Pr pdstane Ph phytane IS internal standard UCM unresolved complex mixture
Fig. 7. Gas chromatograms of the total hydrocarbon fractions obtained from the ooid sands (total sediment, top), the purified ooids, and the deposit enriched in salt fly pupas (Ephydra sp.; bottom).
fication of the bacteria, their physiological activity, and their role in calcification remains to be investigated in detail. Until now we have no indication of abundant bacteria of nm-size ('nanobacteriaL') involved in the for- mation of the crusts.
3.4 Biomarker investigations on ooids and microbialites
Hydrocarbon investigations were performed on (1) a microbialitic ooid chip, (2) ooid sands (total sediment; surface layer, 0-1 cm), (3) purified ooids, and (4) a deposit largely consisting of salt fly pupas (Ephydra sp.) found adjacent to the sediment sampling sites.
Gas chromatograms of the total hydrocarbon fractions
are shown in Figs. 7, 8. Each peak in such a chromatogram represents an individual compound, whereas the peak areas are directly proportional to the concentrations of the respective components. In each case. the main compo- nents are n-alkanes with chain lengths ranging from 15 to 33 carbon atoms. Other individual hydrocarbons are monomethylalkanes, heptadecenes and the isoprenoids pristane and phytane. Variations in the occurrences and the relative abundances of these compounds signify the role of different sources of organic matter contributing to the respective facies types.
The hydrocarbon pattern of the microbialitic ooid chip (Fig. 8) is dominated by short-chain n-alkanes with chain lengths up to 21 carbon atoms. They show a distinct

217
17
, .PI I ! Time
Carbonate crust
I * Is
11 v �9 w � 9 �9 �9
Fig. 8. Gas chromamgram of the total hydrocarbon fraction obtained from the carbonate crust. For sym- bol explanations see Fig. 7.
maximum in n-heptadecane (n-C17). With increasing chain lengths, n-alkane concentrations rapidly decrease, but ex- hibit a minor second maximum at the odd-numbered homo- logues in the region around n-C29. Further significant constituents of the hydrocarbon fraction are short-chain monomethylalkanes and various n-heptadecene isomers. A predominance of n-heptadecane (n-C17) and/or n- heptadecenes is typically observed in sediments receiving a significant contribution of organic matter derived from phototrophic microorganisms. They are particularly prevalent in pure cultured cyanobacteria and in natural cyanobacterial mats (HAN & CALVIN 1969; Boos et al. 1983; see also I-l~vrm~ et al. 1993, and refs. cited therein). A cyanobacterial origin is strongly supported by the co-occurrence of indi- vidual mid-chain branched monomethylalkanes of low molecular weight, which are regarded as characteristic biomarkers for these organisms (SmEA et al. 1990). These molecular signals left by a phototrophic biocommunity are superimposed by a modal distribution of other short-chain n-alkane homologues. Such patterns are generally attrib- uted to autochthonous aquatic sources. They are typically found in sediments in which a pronounced activity of heterotrophic bacteria is observed, e.g. in aphotic micro- bialites (RmTr~R et al. 1996) or the deeper sections of phototrophic microbial mats (e.g. DoBsoN et al. 1988). The imprint of heterotrophic degradation of primary produced organic matter is indicated by the significant hump of an unresolved complex mixture of hydrocarbons (UCM; GRI~LT et al. 1987). The hydrocarbon distribution of the carbonate crust can thus be interpreted as a bacterially induced pattern generated from the interaction/succession of phototrophic primary producers and heterotrophic com- munity members.
In contrast to the microbialite, the hydrocarbon signa- tures of the ooid sands and the purified ooids are domi- nated by high molecular weight compounds. The main components are long-chain n-alkanes showing a clear predominance of the odd carbon numbered homologues. Especially the compounds with 27, 29 and 31 carbon atoms are major constituents of the epicuticular waxes of vascular plants (Eot~rroN & HA~LTON 1967). Their occur- rence in sediments is, therefore, taken as indicative for contributions of terrestrially derived organic matter (PI. 54/8) (T[ssoT & WELTE 1984). A remarkable aspect of the ooid sands and the purified ooids is the presence of high
relative amounts of long-chain monomethylalkanes with carbon chains ranging from C26 to C35. GC-MS analyses revealed that their branching sites are located either at the end (2- and 3-methylalkanes) or close to the centers of the molecules (7-, 9-, 11-. and 13-methylalkanes). Long-chain monomethylalkanes were found as specific constituents of insect waxes (JACKSON & BLOMQUIST 1976; see Ke~io et al. 1995, and refs. cited therein). In the environment studied here, the salt fly Ephydra seasonally occurs in vast amounts and may represent a likely source for these biomarkers. This idea was confirmed by the analysis of a sediment layer enriched with Ephydra pupas which resulted in a hydrocarbon fraction clearly dominated by long-chain monomethylalkanes (Fig. 7, bottom). The presence of these compounds in all samples, therefore, points to a significant organic matter contribution from insects to the GSL sediments.
3.5 Water chemistry During the field campaign in May 1996, water analyses
were carried out at Antelope Island (South Arm) and NW of Promontory Point (North Arm). Due to heavy rainfalls previous to the measurements, the data reflect somewhat uncommon, exceptional conditions. They are not repre- sentative concerning 'normal' conditions, but may be sig- nificant for understandig calcification processes. Samples were taken from open water and from pore water 1-2 cm below the sediment surface. Date measured at Antelope Island and NW' Promontory Point are listed in Tab. 1.
The data reflect a strong and sharp oxycline few mm below the sediment surface. The sediment resp. pore water is heavily influenced by a strong sulfate reduction which increases the alkalinity. During heavy rainfalls anoxic pore water has moved to the surface as seen at the Antelope Island locality. The alkalinity of the open water increases and the Eh values drop significantly.
The situation in the lagoon of Promontory Point is different. The very shallow lagoon is covered by a halite crust which prevents a mixing of anoxic pore water with open water. Important for the growth of the ooids and microbialites is a significant amount of HCO 3- which originates in the sulfate reduction zone. Small pyrite crys- tals are corn mon within the ooids as well as in the microbialites which proves the sulfate reduction process during the ooid growth.

218
T (~ Conductivity (mS/cm) pH Eh (mY) O 2 (ppm) Alk (meq/1)
Bridget Bay, Antelope Island open water pore water
23.9 143.4
8.4 7.95 +85 -335
12.95 0-1.5 8.4
open water Lagoon NW' Promontory Point
pore water
19.7 200
7.67 +144
5.49 5.4
-440 0-1.02
Tab. 1. Data for water samples taken in May 1996 at the Great Salt Lake, Utah, USA.
4 G E N E S I S OF O O I D S A N D M I C R O B I A L I T E S - D I S C U S S I O N
Concerning the growth and fabric ofGSL ooids, previ- ous works focused on physico-chemical mechanisms of carbonate crystal growth and the influence of clay miner- als. EARI~Y (1938) intensively discussed the possible effects of negatively charged clay particles occluded to ooid surfaces and enmeshed within them. He already mentioned the presence of organic matter upon and with in the ooids was already mentioned by him (EARD~Y 1938), later K A ~ (1974) and SA~BERa (1975) agreed with EARDt~Y, but attributed only an insignificant, passive role to organic substances.
SA~BER~S achievement was to demonstrate the pri- mary nature of the radial aragonite needles and palisade crystals by SEM studies of ooid sections. Clay content and microprobe maps of Mg and Si gave rise to the assumption that the modification of normal ooid to cerebroid morphologies is solely a result of adhering clay minerals (sepiolite). KAt~I~ (1974) noted, that the depressions of the (cerebroid) ooids may originate from initial corrosion pits by bacterial activity or other processes, followed by an accumulation of clay at these points.
The precipitation and growth of ooids in general was considered as purely physico-chemical by KAa~ (1974) and SA~BER~ (1975: 507), although the straight-forward paper of Mri-r~RER (1968) had already shown that consid- erable amounts of acidic amino acids capable of Ca 2§ concentration are present within the ooid carbonate. Espe- cially high values of aspartic acids (asp) and, to a lesser extent, of glutamic acid (glu) characterize the GSL ooids (Mrrr~v.Ea 1968, and own data). Our analyses show that beside of proteinaceous matter large amounts of carbohy- drates are present as well. Additionally, labelings with fluorescent and non-fluorescent dyes demonstrate the pres- ence of Ca2+-attracfing (heterogeneous) organic substances upon still growing ooids.
We therefore propose, that growth and fabric of the GSL ooids within the carbonate supersaturated lake brine is mediated and controlled by the superficial organic substances adhering to the ooids. Biomarker analyses support the assumption that bacteria or cyanobacteria are not directly involved in their formation, especially when compared to the data of microbialitic veneered ooid chips and crusts. In contrast, hydrocarbon analyses document
the significance of heterotrophic bacteria and cyanobac- teria in forming the microbialites (e.g. mid-chain branched monomethylalkanes of low molecular weight). Sources of organic matter associated with the forming ooids include detritic wind blown organic matter, remains of insects (Ephydra), and microbial polysaccharides. The biological source of the prominent glu-rich proteins is yet unknown. Abundant nanobacteria were neither found within the biofilms of microbialites, microbialitic ooid chips, nor upon the surface of ooids. PEOONE & FOLK (1996) inter- preted small bead-like carbonate bumps (0,1 gin) within and upon an aragonite cement ofa microbialite ('stromatolite3 as calcified products of nanobacteria, but provided no direct evidence for their presence. So far, our studies by field emission SEM and TEM do not show the presence of nanobacteria associated with carbonate precipitation in the GSL. Instead, small calcareous bodies of comparable size are recognized as a part of the initial calcification of extracellular polymeric substances (EPS). Further investi- gations may clarify this contradiction to the paper of PEDOSrE & FOLK (1996).
5 CONCLUSIONS 1. The aragonitc radial ooids of the GSL contain up to
528~tg/g CaCO 3 intracrystalline soluble organic matter (SOM). 23 % is proteic matter and 77% is glucidic matter.
2. The SOM is characterized by two fractions of mol- ecules. High Molecular Weight Fraction (HMWF) of molecules with 80-90KD and a Low Molecular Weight Fraction (LMWF) with a large range of smaller mol- ecules between 70KD- 13KD. The LMWF molecules are extremely acidic and characterized by more than 40% glu and nearly 20% asp.
3. Calcein and tetracycline stained ooids show distinct fluorescence properties, caused by the Ca 2+ enrichment of the acidic organic matter. The intensity of the fluores- cence is probably dependent on the amount of the Ca 2§ neutralized COO- and SO42-groups. Automicrite is formed within depressions filled up with the organic matter. Remaining intracrystalline organic matter exhibits a strong autofluorescence.
4. Two types of microbialites are recognized: microbialitic told chips and microbial crusts ('stromatolites'). The investigated chips from Antelope Island are covered by

219
thin biofilms of the cyanobacterium Gloeocapsa. Car- bonate precipitation is restricted to a narrow zone at the biofilm base. The microbial crusts are made by complex biofilms with the cyanobacteria Gloecapsa and Aphano- thece and various non-phototrophic bacteria. EPS and biofilm baselayer calcifications form the microbialites.
5. A significant 'background' hydrocarbon contribution to all sediment samples is derived from terrestrial plants, insect waxes and, to a minor degree, autochthonous microorganisms.
6. In the microbialitic ooid chip, the background signal is superimposed by prominent microbial hydrocarbon mark- ers. These findings clearly illustrate that the formation of these deposits is linked to the activity of complex micro- bial communities.
7. A strong similarity between the hydrocarbon fractions of the total sediment and the purified ooids suggests, that the formation of the ooids is not connected to specific biological activity (biofilms). This result is clearly sup- ported by the histological data.
8. The GSL ooids are a product of organomineralization processes in combination with an increased carbonate alkalinity, which is caused by a strong sulfate reduction in the sediment, and a permanent input of divalent cations brought in by rivers.
Part VII CRYPTIC MICROBIALITE IN SUBTIDAL REEF FRAMEWORK AND INTERTIDAL SOLUTION
CAVITIES IN BEACHROCK, HERON REEF, GREAT BARRIER REEF, AUSTRALIA:
PRELIMINARY OBSERVATIONS
G.E. Webb and J.S. Jell
Subtidal, M g-calcite microbialite crusts are pervasive in the shallow, cryptic framework environment on the leeward side of Heron Reef, a small platform reef in the southern Great Barrier Reef. Microbialite-bearing reef framework was recovered from large framework blocks from the sides of Heron boat harbour and from cores drilled beneath Heron Island. The microbialite consists of dense to clotted automicrite typically occurring as microcolumns with diameters between 0.3 and 2.2 ram. Crust surfaces are characteristically 'knobby'. Crusts grew almost exclusively on the floors and sides of cavities with microcolumns aligned vertically or preferen tially inclined to one direction. Crusts postdate scleractinian corals, coralline redalgae, most, but not all, encrustingforaminifers, and encrusting bryozoans, but may overlie or underlie calcareous worm tubes. The crusts grew in a very poorly illuminated, but extensive, open cavity system in the very shallow framework with freely circulating normal marine water. The microbialite does not appear to play a major role in the initial accretion of the skeletal framework, but binds abundant coarse scleractinian, algal and molluscan debris on the cavity floors,secondarily strengthens the
framework, and possibly protects skeletal framework from cryptic bioerosion.
Thin microbialite crusts also occur growing down- ward from the ceilings of solution cavities in the upper intertidal portion of beachrock on the south side of Heron Island. These downward growing crusts have similar micro- columnar to digitate morphology to the subtidal crusts, but generally do not exceed 3 mm in thickness. Both types of microbialite must have accreted very rapidly within the last few thousand years and in the case of the beachrock crusts, almost certainly much more recently.
1 INTRODUCTION Microbialites (e.g., stromatolites, thrombolites) have
long been considered important biologically produced structures in the geological record, but their role in modern normal marine settings has been under-appreciated. Once considered restricted to marine settings with highly abnor- mal water chemistry (e.g., Shark Bay, Western Australia, LOGAN 1961; nearshore Belize, RASMIJSSm~ et al. 1993), agglutinated stromatolites (sensu RIDING 1991) are now known in both subtidal and intertidal settings in the Exuma Cays, Bahamas (D~vis 1983; DmL et al. 1986; DILL 1991 ; Jo~s & HlmaXR 1991; REiD & BROWNE 1991; RIDING et al. 1991a; REID et al. 1995; MAClrCrYRE et al. 1996). Addition- ally, non-agglutinated, thrombolites and stromatolites were described from subtidal reef framework in Tahiti (MONTAGGIONI & CAMOIN 1993; CAMOIN (~ MONTAGGIONI 1994), and from cryptic reef cave settings at Lizard Island, Great Barrier Reef (RErrNER 1993) and at St. Croix in the Caribbean (ZANKL 1993). It is becoming increasingly ap- parent that the high magnesium calcite 'micrite crusts' and 'micrite cements' common in many extant and fossil Qua- ternary reefs (e.g., LAND. & GOREAU 1970; JAr, mS & GrNSBtmG 1979; LAND & MOORE 1980; MARSHALL 1983; MAClNTYmS & MARSHALL 1988; JONES & Htncrm~ 1991) are the products of calcification of microbial or eukaryote biofilms (REIn, mR 1993; Znr~d. 1993, WEBB 1996). Hence, microbialites are important constituents of modern coralgal reefs and ap- pear to have been so throughout the Phanerozoic (PRATt 1982; WEBB 1996).
This paper documents Holocene microbialites from Heron Reef, southern Great Barrier Reef Province (Fig. 9). Subtidal microbialites similar to those described from Tahitian reefs (MorCrAGGIO~a & CAMOn~ 1993; CAMO~ & MorCrAGG~O~ 1994) Occur in Heron reefrock and in inter- tidal solution cavities in Heron Island beachrock. Al- though algae and microbes previously have been impli- cated in the formation of beachrock (e.g., MAXWELL 1962), definitive microbialites have not been described previ- ously from beachrock cavities. The occurrence of microbialite in both subtidal and intertidal settings at Heron Reef suggests that microbial and other biofilm influences on reefal lithification may be more pervasive than generally has been accepted.
2 MATERIALS Samples were obtained from three sources on the
leeward part of Heron Reef. Blocks of reefrock (PI. 55/1)

2 2 0
- ' "
",,, ", [ ~e 0 ,, m . e t e [ s 590
~on Reef. ~ Heron Island N"~, Boat Harbor , \ ) ~ l ~
Beachrock One Tree R f ~ '~f.~.
c
Fig. 9. Location map. (A) Northeastern coast of Queensland, Australia with outline of Great Barrier Reef Marine Park. (B) Reefs in vicinity of Heron Reef. (C) Western end of Heron Reel Open squares: locations of bore holes wherein cores lack framework cavities; half-filled squares: locations of bore holes wherein apparent framework cavities do not contain obvious microbialite; filled squares: locations of bore holes wherein framework cavities contain microbialite crusts; solid triangles: microbialite locations in beachrock; small arrows: area from which dredged blocks were removed from boat harbor walls.
were obtained between 1988 and 1996 from material dredged during enlargement of the Heron Island boat harbor in 1987 (Fig. 9). Reefrock blocks were removed from the edge of the existing harbor by an excavator and were deposited on the beach around the edge of a settling pond/spoil dump on the western edge of the island south of the helipad (Gotau~v & JELL 1993). The exact location of each sampled block in the reef is not known, but all were from the uppermost two meters below the reef flat around the edge of the harbor (Fig. 9). Largest reef blocks ex- ceeded one meter in diameter. Orientation was established in reference to the growth fabric of in situ framework corals (PI. 55/2) and preservation of the reef flat surface. Additional samples were obtained from subtidal reefrock in three of thirteen shallow cores (Fig. 9) drilled beneath the island in 1991 as part of a Heron Island groundwater monitoring program. Additional microbialite samples were recovered from upper intertidal solution cavities in beachrock on the southern shore of Heron Island.
3 SUBTIDAL MICRO BIA LITE Subtidal microbialite crusts occur preferentially on the
floors of reef framework cavities. Most crusts are macro- scopically thrombolitic (clotty),dendrolitic or microcolumnar (PI. 55/3-5). Stromatolitic (laminated) crusts are uncom- mon. Crusts are generally less than 1 cm thick, although thicker vertical accumulations (several cm) occur on the sides of cavities or as shrub-like masses connecting cavity floors and ceilings. Crust thickness is irregular within a given cavity and may vary significantly between adjacent cavities. In some cases, crust thickness increases system- atically towards a particular direction within a cavity and microcolumns may show preferred lateral orientation. Local crust morphology partly reflects cavity size and geometry and the position within the cavity. Five major intergrading crust morphologies occur. 1) Isolated or weakly aggregated clots (diameters 0.3-1.0 mm) occur rarely on the ceilings of cavities and more commonly on the lower surfaces of microbialite that has bridged lateral gaps be- tween coral branches. 2) Clots are commonly stacked into
P l a t e
Fig. 1. Fig. 2. Fig. 3.
Fig. 4.
Fig. 5. Fig. 6.
Fig. 7.
Fig. 8.
55 Holocene microbialite crusts from Heron Reef, Great Barrier Reef, Australia.
Dredged reef framework block. Note microbialite crust coating cavity floors. Scale bar=10 cm. Dredged reef framework block. Note scleractinian corals in growth position. Scale bar=approx. 10 cm. Microbialite crusts extending upward from cavity floors in dredged framework block. Note bivalve detritus (B) cemented into place by microbialite. Scale bar=-I cm. Thrombolitic to dendrolitic crust with calcareous worm tubes in dredged block. Note central, free-standing dendrolite connecting floor to ceiling of cavity. Scale bar=l cm. Coalescing microcolumnar crust from large cavity in dredged block. Scale bar=l cm. Thin section photomicrograph from dredged block showing typical succession from scleractinian coral (S) to coralline red alga (C) to microbialite (M). Scale bar=l ram. Calcareous annelid tube resting on thrombolitic crust on sides and ceiling of framework cavity from core. Scale bar=l cm. Surface of microbialite crust from ceiling of solution cavity in upper intertidal part of beachrock, southern Heron Island. Scale bar=-I cm.

P l a t e 55 2 2 1

222
ORGANISM
scleractinian coral
coralline red algae
encrusting foraminifers
bryozoans
calcareous worm tubes
microbialite
BENTHIC CRYPTIC Fig. 10. Sequence of preserved bi- otic encrustation in Heron Reef framework cavities.
irregular vertical microcolumns (diameters 0.3-2.2 mm), or, less commonly, into digitate shrub-like structures (PI. 55/4) on the sides of cavities and on the floors of small or constricted cavities (i.e., those with initial vertical dimen- sions of less than 2 cm). 3) More regular, relatively smooth microcolumns (diameters typically 0.7-2.2 mm) are abun- dant on the floors of larger cavities, especially in relatively protected low-lying areas, and make up the majority of crusts. 4) In very large, open cavities (vertical dimensions of many centimeters) individual microcolumns coalesce into larger column aggregates (diameters greater than I cm) (PI. 55/5). Individual coalesced columns may expand distally giving rise to a solid, bulbous crust with relatively smooth surfaces or may link to form laterally continuous blade-like projections. 5) Laminated crusts are rare, occur- ring over local highs on the floors of large cavities.
Crusts consist of Mg-calcite and have a peloidal to dense microstructure (PI. 55/6). Fine angular skeletal de- bris occurs in some crusts and many appear brownish in thin section suggesting the occurrence of abundant organic matter. The spaces between columns may contain lithified fine carbonate that is lighter in color in reflected light than the columns. This material is indurated to the same extent as the columns, but stands in lower relief. In other cases, spaces between columns are empty or contain sediment. Microbialite crusts occur on the vast majority of lower and vertical surfaces examined in the leeward Heron reefrock and bind coarse coral, red algal and molluscan debris into the crypts. Crusts invariably postdate in situ scleractinian coral and coralline red algae (P1. 55/6), although rare examples of direct encrustation of scleractinian corals, such as Pocillopora, have been observed. Encrusting foraminifers (e.g., Homotrema) are abundant in the frame- work cavities and mostly predate microbialite crusts, al- though interlayering of microbialite and foraminifers has been observed. Encrusting bryozoans also occur in cavi- ties, postdating some foraminifers, but not microbialite. Various calcareous worm tubes are also abundant in cavi- ties. S mall, coiled worm tubes typically predate microbialite, but small- and large-diameter, straight and irregular tubes may be interlayered with microbialite or postdate it. Post- microbialite worm tubes are most abundant on the walls and ceilings of cavities (P1. 55/7). Coralline sponges and bryozoans, which occur on the surfaces of microbialites at
St. Croix and Lizard Island (RErrnER 1993; ZANKL 1993) have not been observed on the crusts at Heron Island. The sequence of cavity encrustation is shown in Fig. 10.
4 INTERTIDAL MICROBIALITE Intertidal microbialite crusts occur preferentially on
the ceilings of laterally extensive solution cavities, or on the undersides of isolated eroding blocks, high in the upper portion of beachrock. They typically reach thicknesses of less than 4 ram. Crusts consist of irregular anastomosing microcolumns, shrubs, and clots, typically between 0.5 and 1.8 mm in diameter (P1.55/8). Thinner crusts consist of similar textures with smaller dimensions. Crusts of 0.1 mm thicknesses consistofthin'knobby'coatings. Micro- columns are typically somewhat angular in appearance with scalloped or pitted margins. Column extremities appear white when first collected but spaces between columns contain an unidentified, dark green to black algal/ cyanobacterial community.
5 DISCUSSION S ubtidal Heron Reef microbialite crusts are interpreted
as the products of a cryptic community that requires lower light intensity than coralline algae. The nature of the community is unknown owing to the lack of freshly col- lected, 'living' material, but is presumed to consist of microbial and/or eukaryotic biofilm producers, similar to those that produced crusts at St Croix (ZAr~L 1993) and Lizard Island (R~a'NER 1993). The community presumably does not contain abundant cyanobacteria as do the benthic communities that are responsible for the Exuma Cays agglutinated stromatolites 0VlACIrCr~a~E et al. 1996). Heron crusts also do not represent the products of an isolated, deep cryptic or burial diagenesis environment, because annelid worms colonized crust surfaces, suggesting nor- mal marine, if poorly illuminated, waters. Hence, the crusts are synsedimentary products and must have formed within the last several thousand years, since Heron Reef reached its current sea level.
Cryptic, Mg-calcite micrite crusts are known from a variety of Holocene scleractinian reefs and occur in a variety of settings. MAOrCrVgE & MARSHALL (1988) re- viewed the distribution of such crusts and suggested that they are inorganic submarine cements. Since then, the

223
biotic nature of similar fossil and 'living' crusts has been well established (RmrNo et al. 1991b; RmTNER 1993; Zankl 1993; CAMOtN & MorcrAooiorn 1994), and they have been described from high-energy cryptic framework (CAMo~ & MorcrA~OIoM 1994), well-washed, shallow, sub-reef fiat cryptic framework (here described), shallow to deeper reef caves (REIT~mR 1993; ZAr4r, L 1993), deep fore-reef slopes (BRACm~RT & Dt.rU~ 1991), reef lagoons (JONES & Htm'rER 1991), and as replacive fabrics in heavily bioeroded frame- works ( M A ~ & MARSI~LL 1988).
Heron subtidal microbialite crusts are pervasive and bind fine and coarse sediments (PI. 55/3) in the shallow reef framework, but probably are not major contributors to the growing reef edge on the leeward side of the reef. Although the relationship of microbialite crusts to the accreting reef edge has not been observed directly, the breakage of large dredged reefrock blocks appears to have been controlled by original, benthic scleractinian-coralline algal framework, and not by the subsequent microbialite crusts. Therefore, the crusts strengthen and partly lithify the framework, but the coralgal framework would appar- ently be rigid without them. However, the crusts may protect the original skeletal framework from bioerosion, thereby preserving original rigidity. Observed Heron frame- work samples all represent leeward positions in the reef, where submarine cements and biologically-induced car- bonates would not be particularly favored. Similar crusts in the high-energy, windward framework of Tahitian reefs are much thicker and are rarely interlayered with scleractinian corals (CAMOn~ & Mo~rrAooaor,r11994) suggesting that they were important components of the growing framework. Similar crusts are also abundant in most facies of One Tree Reef, also in the southern Great Barrier Reef (MARSHALL 1983). These crusts reach 5 cm in thickness, as in Tahiti, but their distribution in relation to the windward and leeward accreting reef margins is unknown.
Intertidal microbialite in Heron Island beachrock cavi- ties differs from subtidal crusts in growing only downward from the ceiling. Downward-growing crusts occur in subtidal reef caves at Lizard Island (RErrNER 1993) and in the Caribbean (ZAr~L 1993), but not in the shallow Heron Reef framework. The reason for the different morphology is not known, but occurrence of microbialite in the very young and ephemeral beachrock cavities, which are sites of active carbonate dissolution, suggest very rapid accre- tion rates. The beachrock is estimated to be not older than one thousand years and solution cavities are younger still. Additionally, these intertidal microbialites provide an in- dication of the prevalence of microbe- and biofilm-medi- ated calcification in the modern reef environment.
6 CONCLUSIONS AND QUESTIONS Cryptic microbialites occur in shallow subtidal reef
framework and in upper intertidal cavities in beachrock on Heron Reef. Subtidal microbialites from the leeward side of Heron Reef strengthen and lithify existing skeletal framework, but are probably not necessary for short-term reef accretion. They may be more significant in terms of long term framework preservation. Microbialites appear
to be pervasive in modern reef environments and may play a critical role in the creation, and/or preservation, of reef framework rigidity.
Many questions concerning Heron Reef microbialites remain the topics of future research. What type of commu- nity is responsible for the induction of the crusts? What is the nature of framework crusts from the windward side of the reef?. What is the overall role of microbialites in reef accretion?
Part VIII MICROBIAL ALTERATION OF THE DEEP-
WATER CORAL LOPHELIA PERTUSA: EARLY POSTMORTEM PROCESSES
A. Freiwald, J. Reimer and J. Krutschinna
1 INTRODUCTION Azooxanthellate scleractinians show a belt-like distri-
bution along the Northeastern Atlantic continental margin preferentially in water depths between 270 and 450 m (TEIclmRT 1958; WILSON 1979a). In Norwegian waters, however, most deep-water coral occurrences exist on the shelf (FREIWALD 8~ WILSON in press; MORTENSEN et al. 1995), and on deep-seated fjord sills where advection of Atlantic water occurs (DoNs 1944; FREIWALD et al. 1997). A mature reef complex of more than 4 km in lateral extension and up to 45 m in thickness (FREIWALD • WILSON in press) formed byLopheliapertusa has been studied with respect to early postmortem stages of skeletal alteration. Studies on the postmortem alteration of carbonates from deep, aphotic environments are still rare. Best information stems from the Straits of Florida (ZEFF & P~ERKINS 1979; HOOK et al. 1984), the Puerto Rican Shelf (BUDD & PERKINS 1980) and from the Carolina Shelf (PERKINS & HALSEY 1971). These authors noted the importance of endolithic fungal associations in the alteration process of deep-water skeletal carbonates. Fungi often infect other organisms as parasites or infest dead organic matter as saprobes. The ecological impact of fungi on corals has been analyzed for shallow-water environments by RAGttUKUMAR & BALA- SUBRAMANIAN ( 1991), RAGHUKUMAR & RAGHUKUMAR ( 1991) and LE CA~IoN-ALSUMARD et al. (1995).
The preliminary results provide strong evidence for the importance of fungal-bacteria biofilms as inducing agent in early diagenesis above the sediment-water interface of deep and cold-water-agitated shelf environments. In this study, the initial steps of the formation of well-known and globally w idespread occurrences of iron -manganese stained carbonate particles are documented. Moreover, before destructive processes such as bioerosion and biochemical corrosion dominate the long-term preservational pathway, constructive processes such as development of biofilms which, in turn, stimulate sedentary invertebrates to settle on the barren coral skeletons, are of importance. The results presented here and in FREtWALD & W~LSON (in press) form the base of a taphonomic tiering model for deep shelf coral communities which could help to identify ancient counterparts.

224
Fig. 11. Location of Sula Ridge (circle) on the mid-Norwegian shelf (see inlet map).
2 STUDY AREA The coral reef bearing Sula Ridge is located about 80
km offshore at 64~ and 08~ between Fr~yabank and Haltenbank, mid-Norwegian Shelf(Fig. 11). As a morpho- logical elongation of the Frcyabank, this ridge forms a NE- plunging spur at the southwestern end of the NE-SW- striking Haltendjup Basin in 250 to 300 m depth. The Sula- Ridge is bordered by more than 340 m deep depressions. The general current flow pattern offshore Norway is char- acterized by two northward flowing water masses, the Norwegian Coastal Current (NCC) and the Norwegian Current (NC). The less saline NCC is the prominent water mass on the shelf and forms a westward thinning wedge over the shelf margin (EIo~. 1978). The more saline and
more dense NC water passes along the continental slope but intrudes onto the shelf following the morphological depressions such as the Egga Basin (LJgv_N & Nara~N 1969). The temperature of the NC water near the reef site varies slightly, from 6 ~ to 7~ respectively.
3 LOPHELIA PERTUSA: EPITHELIAL TISSUE AND DISTRIBUTION OF MUCUS
Living Lophelia polyps have a yellowish- to orange- coloured soft tissue that rarely exceeds more than 1 cm beneath the edge-zone so that a Lophelia colony consists of numerous functionally and spatially separated polyps. In general, the skeletal area beneath the living polyps is coated by a Iransparent mucus film so that healthy colonies
P l a t e 56
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Skeletal alteration of living and dead Lophelia pertusa: a taphonomic study
Underwater photograph of living and dead Lophelia pertusa. Living corals with extruded tentacles are visible in the lower right. The soft tissue and/or mucus-protected areas beneath the zones of living polyps appear white. The transition from soft tissue and/or mucus-protected skeletal areas is sharp (arrow). The darker coloration derives from iron-manganese-secreting microbes (bacteria and ?fungi; see Figs. 4 and 5). Scale=2.5 cm Response of living Lophelia to attachment of the carnivore polychaete, Eunice norvegicus. The coral polyps produce mucus sheets around the parchment-tube of the polychaete which, in turn, stimulates precipitation of secondarily formed aragonite, the stereome layer. The arrow marks the most recent open aperture of the encrusted Eunice dwelling while former apertures below have been sealed by mucus- controlled calcification. Scale=l cm Cross-sections through Lophelia corallites and extra calcification triggered by the presence of the polychaete (X) (compare with Fig. 2). Scale= lcm Consortium of ring-like iron-manganese precipitates (encircled area for demonstration) which are probably the result of bacterial metabolism and filamentous fungi (arrow). The metallic accumulation accounts for the postmortal coloration of Lophelia skeletons (see Fig. 1). Scale=10 lam Isolated endolithic fungi which typically occur in Lophelia septa. The perpendicular branch-offs (arrow) and the distal swellings are characteristics of this fungal type. Scale=10 I.tm Thin-section from a dead part of a Lophelia skeleton that lacks a soft tissue or mucus protection. This is the zone were another boring fungi, Dodgella, frequently occurs to form a subexpandary layer beneath the exposed surface of the skeleton (black arrows). Moreover, this is the zone of intense colonization of higher invertebrates such as spirorbid polychaetes (S) and octocorals. The ocotocoral spicules (white arrow) are visible encrusting the spirorbid tube. Scale=150 I.tm Dodgella, isolated from a Lophelia skeleton with its flask-like habit and narrow neck. Scale=6 gtm

P l a t e 56 2 2 5

2 2 6
appear white and clean (PI. 56/1 and see Sa~OM~.zN 1971). This slimy mucus is released by the polyps probably in order to prevent the attachment of sessile invertebrates near living polyps thus reducing the spatial competiton between corals and associated invertebrates.
However, there are few attached organisms which are adapted to compete with the mucus film such as the parasitic foraminifer Hyrrokkin sarcophaga (C~Dr~mEN 1994) and the tube-constructing polychaete Eunice norve gic us. Hyrrokkin sar copha ga (CEDrtAOEN 1994) pref- erentially colonizes in the vicinity of living coral polyps and where it etches deeply into the skeleton of the host (FRVaWALD & ScnO~a~ 1996). In places where Eunice norvegicus forms a pergament-like tube between living Lophelia polyps, mucus release occurs not randomly. Specifically, the mucus is present around the parchment tubes of the carnivorous polychaete Eunice norvegicus. Beneath the slimy mucus sheet, stereome aragonite is precipitated around the parchment tube of the polychaete (PI. 56/2-3).
4 EARLY STAGES OF SKELETAL ALTERATION IN LOPHELIA PERTUSA
Skeletal areas which lack a mucus and soft tissue protection get rapidly colonized by other sessile inverte- brates such as spirorbids, other tube-forming polychaetes (Filograna sp.), brachiopods (Terebratulina retusa, Cra- nia anomala), byssate bivalves (Acesta excavata, Delectopecten vitreus) and encrusting bryozoans (F~ErwALo & Wmsor~ in press).
The consumption of mucus sheets as a food resource by bacteria and grazers is well-known in shallow-water coral reefs (Cov~oTH 1990) but has not yet been investigated in deep-water corals, However, this study presents evidence of early stages in epithelial soft tissue degradation by iron- and manganese-secreting bacteria and filamentous fungi. The elemental composition of the bacteria accumulates has been checked using an energy dispersive X-ray microprobe. Iron and manganese is secreted in the form of rings each measuring 3 to 8 gm in diameter (P1. 56/4). Presumably, these accumulations are deposited around single bacteria or bacteria aggregates. After the decompo- sition of the soft tissue, the metalliferous precipitates remain on the barren coral skeleton thus generating a faint reddish brown coloration. During maturation and epitaxial growth of the minerals, the faint iron-manganese aggre- gates can grow to form coherent iron-manganese crusts.
4.1 Infestation of coral septa by an unknown fungus type
Associated to the bacteria are fungi with branched f'damentous hyphae (PI. 56/5). These fungi consist of numerous straight and elongated cylindrical hyphae. Newly formed hyphae branch-off nearly perpendicular from older hyphae (PI. 56/5). Frequently, the distal growth zone of the hyphae shows globular swellings which may represent sites of fructification. This fungal type is endolithic and is most abundant in the septa of Lophelia. Microscopic studies often show a close spatial association with the iron-
manganese mineralizing bacteria (PI. 56/4). Moreover, this fungus frequently has been found at the outer edge zone of the corals and in the septae. BtrDD & Pm~raNs (1980) and Zm~v & PERKINS (1979) reported a microendolithic organism with similar outer morphological attributes from deep-sea carbonate particles. This microborer was intro- duced as 'tubular form I' and is suspected to be of fungal origin.
4.2 Postmortem Dodgella infestation Coral skeletons which lack any soft tissue or mucus
coverage for a longer period show a different style of fungal colonization. This indicates the existence of a taphonomic tiering from living to decaying to longer dead deep-water corals (see PamWALD & WmSON in press). While the iron-manganese precipitates are still present, another fungus gains importance. Flask-like borings of up to 20 I.tm-depth penetration and about 10 lain in diameter are arranged along all exposed surfaces of the coral skel- eton (PI. 56/6). The long axis of these minute excavations are orientated perpendicular to the skeletal surface. The sac-like borings are interconnected with 2 to 5 gin-thick collar-bearing necks to the substrate surface. Such subexpandary borings are characteristic for the the endolithic chemoheterotroph chytridialid fungi Dodgella (P1. 56/7 and ZERBROWSra 1936; ZF.W & P~.xn~s 1979; Giotto 1994). Owing to their subendary arrangement beneath the surface of the coral skeletons, the term 'Dodgella-layer' is appro- priate (PI. 56/6). The presence of Dodgella on Lophelia is restricted to the skeletal areas which are completely free of any epithelial soft tissue or mucus sheets. However, this zone seems to be attractive for larvae of various rapidly growing sessile invertebrates to attach. This is congruent to an observation provided by PERraNS & HALSEY (1971) that Dodgella appeared shortly after death of an infested organism.
5 PERSPECTIVES OF THE LONG-TERM PRESERVATION
The role of endolithic fungi and metal-accumulating bacteria only describe the early postmortem alterations in deep-water corals. The more advanced taphonomic stages in cold-temperate deep-water coral reefs can be character- ized by the following processes: 1) Increase of sponge and octocoral encrustation of the dead coral framework which result in reducing the framework porosity by the macroorganisms and by trapped pelagic particles (foraminiferal tests, pteropods and coccoliths), 2) Increase of sponge-dominated bioerosion which causes in situ collaps- ing of the reef framework (cf. FREIWALD & WILSON ill press).
The potential outcome of these long-term alterations is the conversion of a pure 'coral-dominated framework'into a collapsed 'biodetrital mound filled with a calcareous muddy matrix'. The latter results from trapping pelagic fines and from internally produced bioerosional muds (FARROW & FY~ 1988). Both, the coral-dominated frame- work and the late taphonomic stage of collapsed biodetrital 'mounds' are present on Sula-Ridge, mid-Norwegian Shelf in about 300 m depth.

227
- A N C I E N T S T R O M A T O L I T E S AND M I C R O B I A L I T E S -
P a r t I X
STROMATOLITE DECLINE: A BRIEF REASSESSMENT
R . Riding
1 INTRODUCTION Stromatolites and associated microbial carbonates
(thrombolites, dendrolites) are the most common and con- spicuous fossils in the Precambrian (IVIASLOV 1959), but are comparatively scarce at the present day. This history of long-term decline has been interpreted as a response to the appearance of metazoans (GARRETr 1970a, b), dating from -675-570 Ma (AwRAMn~ 1971). MGNTV (1973) argued against metazoan competition, but believed that algal competition may have been significant. WALTER & HEYS (1985) re-examined stromatolite data and found that de- cline started near 1000 Ma, but they considered that meta- zoans were extant then and criticized Monty's disparage- ment of the metazoan grazing hypothesis. However, GROrZINCER (1990: Fig. 6) presented qualitative abun- dance data indicating that decline commenced as early as 2000 Ma and suggested that declining carbonate precipita- tion may have been responsible.
Thus, research since the metazoan ecological restric- tion hypothesis was first expounded 25 years ago requires re-examination of this concept, and of the reasons for stromatolite decline (GRoTZINGER 1990; SEMIKHATOV 8L RAABEN 1993). Alternative proposals, that competition with algae or reduction in mineralization may have been important factors influencing stromatolite formation, were first made by FISCHER (1965: 1208-1209), who took the view that stromatolites declined in subtidal environments from the Mid-Ordovician onwards.
Microbial growth and interaction with other organisms is only one side of the microbial carbonate equation. The success of stromatolites and other microbial carbonates depends as much upon synsedimentary lithification, with- out which accretion and preservation cannot occur, as upon microbial growth itself (Rran~G 1994a). Recognition of the importance of synsedimentary calcification and lithification directs attention away from emphasis on eco- logical interactions towards environmental constraints. It has important implications for understanding microbial carbonates throughout their long history, including mod- em examples such as those at Shark Bay in Australia and Lee Stocking Island (PI. 57) in The Bahamas.
2 CALCIFICATION AS A KEY FACTOR Synsedimentary lithification is the key to the forma-
tion, maintenance, preservation and abundance of micro- bial carbonates, and is more important than metazoan interference in determining success through time. Supersaturation with respect to carbonate minerals varies spatially at the present day and directly affects microbial carbonate sedimentation, as can be seen by comparing tufa stromatolite formation in freshwater, at one extreme, with
the absence of stromatolites from cold marine waters at the other. Through time in marine environments, microbial calcification has been common during episodes termed Cyanobacterial Calcification Episodes (CCEs) (RIDrs~ 1992), marked by relative abundance not only of microbial carbonates, but also of non-skeletal carbonates. CCEs are believed to represent periods of elevated carbonate satura- tion. They may correspond with one or more of the follow- ing: high global temperature (which enhances precipita- tion rate), low sea-level and low skeletal abundance (which increase availability of calcium and bicarbonate), and development of alkalinity pumps from stratified basins.
Following discontinuous decline commencing about 2000 Ma (GRoTZ~GER 1990: Fig. 6), which can be attrib- uted to environmental change and which continued until the Vendian, stromatolites and other microbial carbonates recovered significantly in the latest Proterozoic and were abundant during the Cambrian and Early Ordovician. This trend appears to mirror temperature ( R m ~ 1994b: 435). Both metazoans and microbes radiated in the Early Cambrian, and reefs were built by both metazoans (archaeocyaths) and microbes, although microbes were generally much more abundant (RIDIN~ & ZHta~AVLEV 1994). Microbial carbonates did not significantly decline until the mid- Ordovician, as FIscrm~ (1965) pointed out, at the close of the first major Phanerozoic CCE. Subsequent well-de- fined CCEs occur in the Upper Devonian and Mid-Upper Triassic. These recoveries and subsequent declines of microbial carbonates support links with physical/chemical factors rather than biological interference. Positive tem- perature excursions during the past 700 Ma particularly seem to correspond with microbial carbonate formation (RIDING 1992). Temperature appears to have been a key long-term factor that directly affects microbial growth and synsedimentary lithification. Marine microbial calcifica- tion has declined since the late Mesozoic, possibly due to a combination of global temperature decline and to in- creased pelagic removal of skeletal CaCO 3 by plankton such as coccolithophores and globigerines (Rttm~ 1993). CCEs may be recognizable in the Proterozoic (RIDING 1994b: Fig. 2).
3 MICROBIAL INTERFERENCE WITH METAZOANS?
It has been suggested (VOLOGDIN 1962; MoNTY 1973: 603) that microbes delayed metazoan radiation in the Neoproterozoic, so that microbial decline may have fa- voured metazoan emergence. This could account for the late timing (see ERWIN 1993) of metazoan radiation. But what happened in the Phanerozoic? Two problems with the Garrett-Awramik model concern not only the timing of Proterozoic stromatolite decline, which significantly pre- dates metazoan evolution, but also the continued success of microbial carbonates in the Cambrian. Stromatolites continue abundantly during the Cambrian at a time when metazoan interference should have been intensifying. In addition, other microbial carbonates such as thrombolites and dendrolites also become abundant in the Early Cambrian. In fact, in Lower Cambrian reefs, dendrolitic fabrics cre-

2 2 8
CASE TIMING INTERPRETATION BY
METAZOAN INTERFERNCE
INTERPRETATION BY
ENVIRONMENTAL FACTORS
stromatolite decline
stromatolites at mass
extinctions
Shark Bay and Lee
Stocking columnar
stromatolites
Proterozoic
end-Permian
Recent
grazing and disturbance limit stromatolite
diversity and abundance
reduction in metazoan interference due to
mass extinction permits stromatolites to
develop as "Disaster Forms"
stromatolites occupy stressed metazoan-
free refuges
one or more of calcification reduction,
temperature reduction, and oxygen
increase, reduce stromatolite accretion and
preservation
factors favouring microbial growth and
early lithification simultaneously inhibit
metazoans
stromatolites occupy sites of
synsedimentary lithification
Tab. 2. Exarnples of stromatolite events and occurrences and their differing interpretations according to the Garrett-Awramik Metazoan Interference model ( G ~ T r 1970a, b; AW~A~K 1971) and the Environmental Constraint model (Rmmo 1995).
ated by calcified micro fossils thatare probably cyanobacteria, are volumetrically often much more abundant than meta- zoan reef builders such as archaeocyath sponges. It is possible that archaeocyaths were actually outcompeted by calcified microbes ( R m ~ & ZmraAVLEV 1994). It remains to be seen whether subsequent Phanerozoic fluctuations in metazoan development were direct interactive responses to microbes, or were due to the environmental factors that affected microbes (Tab.2).
tidal flats are sites of evaporation which has a similar effect. This early lithification is crucial to stromatolite accretion. Thus, if the possibility of metazoan interference is neglected, it is not difficult to see these present day stromatolite habitats, not as refuges from metazoan inter- ference, but as environments of rapid early lithification (Tab .2). The same reasoning applies to the development of heavily calcified stromatolites in freshwater lakes and pools.
4 STROMATOLITES AS DISASTER BIOTAS? A corollary of the metazoan interference hypothesis is
that when metazoans are scarce, stromatolites will become more abundant. SCHUBERT & BOrTJER (1992) used this reasoning toaccount forincreased abundanceofstromatolites immediately following the Permian-Triassic mass extinc- tion. But an alternative view can be proposed, that increase in stromatolite abundance at times of mass extinction is not due to metazoan reduction but because environmental conditions at those times favoured stromatolite growth and calcification. Consequently, at the end of the Permian for example, metazoan decline may be due to microbial inter- ference and/or the factors that encourage stromatolite formation. According to this new view (RmING 1996), environmental factors that reduce metazoan abundance and diversity at mass extinctions could favour stromatolite formation irrespective of competetive interractions.
5 SHARK BAY AND LEE STOCKING ISLAND One of the reasons that the Garrett-Awramik model
became such a widely accepted part of stromatolite lore is that it accords well with the present-day distribution of marine stromatolites in environments hostile to most meta- zoans, including hypersaline lagoons, desiccated tidal flats and areas of highly mobile sediment (AwaA sw~ 1971: 825). But these are also habitats where carbonate precipitatio n is enhanced. Shark Bay and Lee Stocking are wave and current swept environments respectively where water movement will promote marine cementation, and inter-
6 THE PRECAMBRIAN ENIGMA Although it has been acknowledged that a variety of
microbes could have been involved in building Precambrian stromatolites, it has been generally assumed that the prin- cipal ones were cyanobacteria. But if Proterozoic syn- sedimentary carbonate precipitation was as intense as has been suggested, then we should expect calcif ied cyanobacterial microfossils to be far more common in Proterozoic stromatolites than the reported record indi- cates. This scarcity of calcified cyanobacteria in stromatolites that have been assumed to have been built mainly by cyanobacteria at a time when carbonate precipitation was assumed to be prolific has been termed the Precambrian Enigma (Rtomc 1989, 1994b: 432). This enigma raises the suspicion that at least one of these assumptions is incor- rect, and it has been suggested that cyanobacteria may have been less important in Precambrian stromatolite formation than has been thought (RmING 1994b: 432). The paucity of recorded calcified cyanobacteria suggests that smaller microbes, such as eubacteria, may have dominated Archaean and Proterozoic stromatolite biotas. Calcified fossils which resemble cyanobacteria do not become widely abundant until the latest Neoproterozoic. There have been subsequent changes in stromatolite biotas too (RmxNG 1991: Fig. 1), as when diatom algae entered stromatolite building biotas in the Cenozoic and became important in modern stromatolites such as Shark Bay and Lee Stocking Island (AWRAMIK & RmrNG 1988; RIDING et al. 1991).
Microbes are environmentally sensitive, and there is

P l a t e 5 7 2 2 9
P I a t e 57 Columnar stromatolites, The Bahamas.
Columnar stromatolites in the tidal current-swept channel between Lee Stocking Island and Norman's Pond Ca),, Exuma, The Bahamas. According to the Metazoan Interference model, these large stromatolites occupy a site where mobile sediment and strong currents provide a refuge from metazoan interference. According to the Environmental Constraint model, this site is occupied by stromatolites because it enhances the synsedimentary lithification that is essential to their accretion and preservation. At this site it is difficult to know which of these explanations is better, or whether both apply equally. But overall evidence from the geological record suggests that the Environmental Constraint model (RID~6 1995), which proposes that environmental factors have mainly determined the degree of success of microbial carbonates, may be more valid than the Metazoan Interference model which has dominated interpretations of stromatolite distribution in time and space for the past twenty-five years. Photograph by R. Riding, September 1987.
evidence that the global environment was changing sig- nificantly during the Proterozoic in ways that would di- rectly have affected stromatolite formation. In addition to carbon dioxide decrease (KAs'r~c 1987) which could slow carbonate precipitation, and temperature decline (KARaU & EPSa'E~ 1986) which would slow both lithification and microbial growth, increase in atmospheric oxygen also (KAsTINC 1987; KARrrtJ & HOLLAND 1996) occurred. Many bacteria have growth optima well below modern atmos- pheric oxygen levels, but cyanobacteria have adapted to present day conditions. Thus, eubacteria can be expected to have been been more sensitive than cyanobacteria to oxygen in the Proterozoic. Coincidence of onset of stromatolite decline at ~ 2000 Ma (see GROTZtN~R 1990: Fig. 6) and rapid oxygen increase between 2220-2060 Ma (KARmJ & HOLLArr 1996) is consistent with the view that eubacteria had greater involvement than cyanobacteria in stromatolite formation in the Early-Mid Proterozoic. To-
gether with reduction in calcification potential, I suggest that this may have contributed to the Proterozoic decline of stromatolites (Tab. 2).
7 CONCLUSIONS Stromatolite decline cannot be simply accounted for by
metazoan interference. This does not mean that interfer- ence has not occurred, but it does require a review of explanations that have been taken for granted. Environ- mental factors, through their effects on microbial growth and calcification, are probably more important than bio- logical interference in determining overall microbial car- bonate distribution in time and space (Rmn~6 1995). In particular, synsedimentary lithification/calcification is central to microbial carbonate accretion and preservation. Reap- praisal suggests that stromatolites are not disaster biotas, and that Shark Bay and Lee Stocking Island stromatolites do not occupy stressed refuges from metazoan interfer-

2 3 0
ence. Instead, stromatolites develop at times and in loca- tions where environmental factors are favourable. In these conditions, microbial carbonates can co-exist with or even displaced metazoans
Part X ARCHEAN AND PROTEROZOIC LACUSTRINE
STROMATOLITES
S.M. Awramik
The pre-Phanerozoic contains a rich and diverse record of stromatolites. However, the vast majority of these are from or thought to be from marine sedimentary sequences. Few lacustrine examples have been recognized. The pau- city ofpre-Phanerozoic lacustrine stromatolites may result from one or more of the following reasons: (1) lacustrine environments and their contained stromatolites were rare in the pre-Phanerozoic; (2) the stromatolites themselves were rare in pre-Phanerozoic lacustrine environments de- spite an adequate representation of that environment in the stratigraphic record; (3) the number of lacustrine environ- ments preserved has been reduced from their original numbers due to erosion; and (4) regardless if 1-3 are true, a number of pre-Phanerozoic lacustrine environments and their stromatolites have not been recognized because of the difficulty in recognizing such systems in thepre-Phanerozoic. The identification of the lacustrine nature of stromatolitcs in Phanerozoic deposits is considerably easier than with pre-Phanerozoic deposits and is done largely on the basis of associated fauna and flora or the presence of diagnostic fossils to establish a marine origin. Sedimentological fea- tures, diagnostic minerals (e.g., trona), geochemistry, facies relationships, and basin geometry are also used (I:'tCARD & Hlon 1972). Nevertheless, it is usually the fauna and flora, their nonmarine or marine nature, that are used as the principle and most con fident criteria. For the pre-Phanerozoic, non-biotic criteria were the only ones used. Using stromatolites, I have developed biotic-related cri te- ria found in the stromatolites themselves, to provide clues to whether the stromatolites formed in a marine or lacustrine setting. These criteria resulted from an analysis of thin sections and slabs of stromatolites from 37 localities/ formations ranging in age from pre-Phanerozoic through Recent, and from known lacustrine and marine settings. Based on this study, some generalizations have been de- veloped that represent major features of stromatolites that formed in lacustrine environments: 1. significant morphological variability among adjacent
columns or domes 2. columns, if present, are often small (few cm's) 3. macrolamination (light and dark bands composed of
many individual laminae) 4. light laminae often thicker than dark laminae 5. light laminae have a radial fibrous texture 6. sharp boundaries between light and dark laminae 7. laminae are often continuous and smooth within the
stromatoid
8. laminae are easy to trace laterally from one stromatoid to another, often over many stromatoids
9. laminae are composed of fine-grained sediments (micrite)
No single feature provides compelling evidence for a lacustrine origin; however, the combination of two or more features can provide suggestive to presumptive evi- dence fora lacustrine origin. A study ofstromatolites from pre-Phanerozoic formations interpreted to nonmarine on non-biotic grounds (Tumbiana, Reitgat, Wollogorang, Copper Harbor Conglomerate, and upper Loves Creek Member of the Bitter Springs) shows that the stromatolites satisfy at least three of the above criteria. This approach has worked in at least one instance with the study of Collenia undosa WALCOr'r from the Spokane Shale (Belt Supergroup, USA; AWRA~K et al. 1993) whose attributes included eight of the nine criteria (columns are not present). It must be cautioned, however, that complimentary non- biotic criteria for a lacustrine origin must also be sought to increase the confidence in the interpretation. This is par- ticularly important for any ancient large lake, a hundred or more kilometers in diameter, as such large lakes can have non-biotic features that make a lacustrine or marine inter- pretation difficult (e.g., PACKER 1990). The significance of this study is several fold:
1. Stromatolites can be used as paleoenvironmental indicators suggesting a lacustrine setting.
2. Lacustrine and marine stromatolites differ morpho- logically. An analysis of those features that differ and those that are similar, may help to shed light on factors that affect growth and control morphology in stromatolites. Morphologically distinctive and thus biostratigraphically useful lacustrine stromatolites are very rare. This may result from the fact that lake environments are notoriously variable - conditions change with the seasons (very large lakes can dampen this effect). This suggests that relatively stable environmental conditions over years are conducive for stromatolites to exhibit morphological complexity of limited variability.
3. We now have an opportunity to better understand the early history of life in lakes, at least as far back as about 2750 Ma (e.g., Bt~cK 1992).
4. Stromatolites should be searched for on Mars. Mars is likely to have had lakes during its early history although there is considerable debate as to how wet early Mars was (SQUIRES & KASTING 1994). If life appeared and evolved on Mars, it probably adapted early to lakes, the largest-scale aqueous environment available. The lesson from early Earth suggests that life became adapted early in its history to the largest-scale aqueous environment (which was ma- rine) by 3500 Ma and by 2750 Ma, it adapted to lakes. Knowing the preferred setting for stromatolite formation within pre-Phanerozoic lakes on Earth, one can target similar settings in ancient lacustrine deposits on Mars to search for stromatolites. Since stromatolites are usually at the centimeter to decimeter scale, if present on Mars, they should be detectable through video imaging (with a reso- lution at the scale of several centimeters) of outcrops and eroded boulders and other loose rocks.

231
Part XI VERMICULAR STRUCTURES
IN THE NEOPROTEROZOIC OF THE WEST AFRICAN CRATON:
MICROBIALITES VERSUS 'MOLAR TOOTH' J. Bertrand-Sarfati, J.C. Plaziat and
A. Moussine-Pouchkine
1 INTRODUCTION Vermicular structures were firstly described in carbon-
ate beds from the Neoproterozoic sediments of the West African Craton, in the Hank area, Algeria, in the Adrar area, Mauritania, and all along outcrops between the two areas (BER'rRArco-SAm~ATI 1972). These puzzling struc- tures isolated or gathered in clusters (PI. 58/I) were attrib- uted to an organism lncertae-sedis called Vermiculus contortus. Accordingly, it was used for stratigraphic cor- relations through the northern edge of the Taoudenni Basin (BERTRAND-SARFATI & TRO~m'rE 1976). In more recent studies, 'vermiculus' were referred to as 'shrinkage cracks and/or biologic features' (BERTRAND-SARFATI • MOUSSINE- Poucnrm~ 1988). At that time, they have been compared to 'molar tooth' structures, considered as shrinkage cracks. These 'molar tooth' structures, well known from diverse Proterozoic sections (S~a'H 1968; SIEDLECKA 1978; HORODYSKI 1976), have been recently considered as related to seismic activity (PRATr 1992, 1997; FAmCmLD 1997). In the Algerian part of the Taoudenni Basin, the discovery of liquefaction seismites in relation to the extensional event of the opening of a pan-african rift, in the nearby Hoggar, allows us to propose also a relation between these ver- micular structures and seismic activity (BERTRAND-SARFATI & Moussn,~E-Poucmar~ 1993), however their originality as biological strutures differing from 'molar tooth' were already emphasized (Movssn~-Poucma~ & BERTRAND- SAt~A~ 1997).
Even if some of the white vertical fissure-like struc- tures, associated to seismite structures, can be interpreted as similar to the 'molar tooth', we will insist on all the evidences implying a biological origin to the most striking vermicular structures: lenticular stromatolite and microbialite beds, deformed and reworked in relation to seismically liquefied units.
2 LOCALIZATION The West African Craton is covered by a sedimentary
pile that comprises the Neoproterozoic to Paleozoic for- mations of the Taoudenni Basin (Fig. 12). In Algeria, disconformities outline different tectono-sedimentary units (MousstNE-PoucHKINE & BERTRAND-SARFATI 1996). There, numerous seismic events were registrated in carbonate and siliciclastic sediments (slumps, breccias, liquefied layers and hydroplastic deformations such as pseudo-nodules) and vermicular structures. In Adrar area, the Neoproterozoic sediments are piled without disconformities. However, we have found recently seismites features, in several hori- zons, but vermicular structures are only preserved within a single 6m-thick layer of blue-black limestone. These carbonates can be traced along the continuous 1100 km of
outcrop, from Algeria to Mauritania, containing the same vermicular structures.
3 DESCRIPTION OF FACIES IN ADRAR AREA The section of Adrar area comprises blue-black lime-
stone between thick (>20m) biostromes of stromatolites mainly dolomitic (Fig. 13A). The stromatolite biostrome below the limestones is covered by dolomitic sands with storm features (HCS) in which liquefied and brecciated beds are found including pseudo-nodules with fissures similar to 'molar tooth', i.e. fissures filled with micritic material, that are crossing the bedding planes before the folding and showing features of differential compaction with the encasing sediments. The stromatolites above the layer with vermicular structures are also intercalated with coarse grained carbonate sands that bear some vertical fissures of 'molar tooth' type. Four facies of the limestone bed with vermicular structures (Fig. 13B) will be shortly described, the less disturbed facies at the base are overlain by highly deformed dislocation breccias. The fifth facies is not comprised in the blue-black layer, but found beneath the stromatolitic biostrome which underlies itas well as in the Hank-Algeria.
3.1 Undisturbed Sediment Undisturbed sediment consists of a centimetric to
decimetric alternation of fine-grained carbonate mud with few clay particles, faintly laminated and containing by places carbonate grains (silt size) and microbialites. The microbialites, preserved in thin beds, never exceeding 4-5 cm, are composed of vesicular and/or elongated micritic bodies, with numerous sparitic fenestrae (somewhat porostromata-like), showing a discontinuous horizontal layering.
Fig. 12. Neoproterozoic outcrops in the West African Craton, from Adrax-Mauritania to Hank-Algeria: about 1100 kin.

2 3 2
3.2 Stromatolites flakes On the outcrops stromatolites flakes appear as white
vermicular structures, dislocated in all directions but with a faintorientation. The vermicular structures are stromatolites, disrupted in flakes (80-90%), displaced with almost no transportation and resedimented. The flakes, broken or folded, are separated by a thin veneer of recrystallized matrix (iron-rich rhombs).
3.3 Subspherieal Clusters These are subspherical to ovoid complex masses em-
bedded in a blue-black matrix representing only 20% of the rock. The clusters are made of white elements of different origin described below (P1. 58/1-3). In the recrystallized matrix there are elongate clasts of stromatolites and vein- like structures folded and/or broken and filled with micrite of possible biologic origin.
3.4 Coarse Breccias The polygenic breccias in graded-beds are composed
of clasts of layered carbonate of variable size (millimetric flakes to centimetric angular blocks), including vesicular microbialites and transported clasts of laminated micrite with a bilateral symmetry (PI. 58/10). Their shape looks like reworked veins but they are definitely made of micrite of biological origin.
3.5 Fissure-Like Structures In the field they look like seams of vertical and parallel
fissures clearly orientated. In fact they are much more complex, composed of vertical micritic structures broken and displaced (mm scale) in a micritic matrix. The micritic filling of these bodies shows a bilateral symmetry and concentric laminae. Outside the vertical vein-like struc- tures, the diagenetized matrix also contains numerous
clasts of carbonate, micrite lenses and micritic subcircular to elongated elements. All these structures cannot be referred to as 'molar tooth' veins, because they exhibit a much greater complexity without the typical compaction features.
4 DESCRIPTION OF THE CLUSTERS In the field, the horizontal surface of layers with clus-
ters shows juxtaposed but distinct subspherical white 'balls', about 10 cm diameter, in a blue-black muddy carbonate faintly layered. Numerous large stylolites cut accross the beds horizontally, every 20-30 cm but not frankly accross the clusters (PI. 58/3). In vertical section they are either subspherical or elongated. Generally a stylolite bordered the cluster along its lower side, while the upper part presents elements (stromatolite clasts) wedging the cluster and included in the less recrystallised matrix.
Clusters are composed of elements, broken or not, in a black recrystallised matrix. Among the elements the most abundant are clasts of reworked stratiform stromatolites broken in sort of a vertical fold, or arranged in piles (PI. 58/ 4). These are laminated micritic bodies (P1. 58/7), with local enlargement of the light laminae filled with sparite (PI. 58/9). Other stromatolitic elements are clearly bulbous with convex laminae (PI. 58/5). In the centre of the illus- trated cluster is an elongate vertical element with a bilat- eral symmetry. The overall shape is ellipsoidal-flattened, with undulating laminae arranged symmetrically (PI. 58/ 6) around a very regular central sparitic zone (P1. 58/4). Within the cluster, between the elements, a carbonate mud is deposited with a concave upward layering (PI. 58/5). Between the clusters the matrix is diagenetically modified in a coarse microsparite with a faint undulating layering. Rare grains are still recognizable. In this matrix stromatolite clasts and other laminated elements are embedded in
P l a t e 58
Fig. 1. Fig. 2.
Fig. 3. Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7. Fig. 8.
Fig. 9. Fig. 10.
Vermicular structures in the Neoproterozoic of the West African craton
Surface, almost horizontal, of a layer with clusters showing a pattern in staggered row. Scale=20 cm. Detail of the previous photograph, the elements seem to be rolled in the cluster, some of them uncompletely. Many elements show a bilateral symmetry. Scale=8 cm. Vertical section of the bed, cutting a cluster almost vertically in its centre. Scale=3.5 cm Thin section in the cluster of Fig.3. The cluster is asymmetric: the lower part is lined by a stylolite, the upper part free of compaction. On the right stromatolite clasts are broken in a fold-like pattern; on the left they are piled. The centre of the cluster is a microbial element with bilateral symmetry. It is clear that elements in the cluster were deposited together with the inner mud carbonate that settled down on them. Scale= 1.2 cm. Detail of the sediment within the cluster; layering is concave upward. Arrow points to bulbous stromatolite clast. Scale= 0.50 cm Central element of the cluster, clearly laminated-undulated, with a bilateral symmetry around a central sparitic zone of very constant thickness. Scale= 0.20 cm Detail of the broken stromatolite clasts in the cluster, Fig. 4 on the right. Scale=0.25 cm Between the clusters, in the highly recrystallized matrix (probably after liquefaction) with a faint layering, microbial elements are found: oncolites with expansions and plates with bulbous folds. Scale=l.6 cm Detail of a stromatolite clast clearly showing undulating laminae and sparitic laminae. Scale=0.25 cm Breccia, with carbonate clasts of various size and vein-like elements which are in fact microbial (bilateral symmetry of the thinly laminated body). Scale=l cm

P l a t e 58 2 3 3

234
Fig. 13 A. S tratigraphic scheme of the Neoprotero- zoic cover in Adrar. 13B. Schematic section of the blue-black layer with vermieular structures, in Adrar-Mauritania. The figures are related to the succession of facies described in the text.
elongated bodies with bulbous excroissances and oncolite- like elements with elongated expansions (PI. 58/8). All these elements appear as being biological in origin (stromatolite-like).
5 CONCLUSIONS 1) The partial analogy of the Neoproterozoic vermicular structures of the West African Craton with 'molar tooth' structures do not exclude a biogenic origin for most of them. 2) The original sediment in which the vermicular struc- tures are developed was an alternation of microbialite layers and very fine grained carbonate muds. These microbialites are preserved only in centimetric layers of vesicular structures. We suggest that their opposition to the adjacent strongly lithified stromatolitic biostromes may be related to an exceptional change in the water and sediment geochemistry of the West African Shelf. 3) Seismites with true 'molar tooth' are recorded in beds above and beneath the layer with vermicular structures. In all the layers of the black-blue bed, seismic activity is illustrated by dislocations that imply a liquefied stage. Clusters or vein-like structures included in a liquefied matrix as well as breccias are related to a liquefaction probably leading to a reworking. 4) These vermicular structures can be confounded with 'molar tooth' structures, but a careful petrographical study shows that they have an undulating laminated fabric and sometimes a bilateral symmetry that involve a biological origin. 5) Vermicular structures occur in a single limestone hori-
zon. The carbonate muds liquefied very easily, compared to the biogenic layers probably because of the abundance of particular organic matter. The microbialites behave as more rigid elements being more early lithi fled. That differ- ential comportment explains the strangest structures ob- served (clusters). 6) Why do 'molar tooth' structures as well as vermicular structures mainly occur in Proterozoic rocks? The mecha- nism of their early lithification as a micritic inflll remains still unexplained. Could it be biologically induced in both cases?
Part XII MICROBIAL SIGNATURES IN LACUSTRINE AND FLUVIAL CARBONATES: GONDWANA
(PERMIAN) AND H O L O C E N E EXAMPLES, INDIA
M. Mohanti and S. Das
Laminated microbial carbonates occur in the siliciclastic sequence of the Talchir Formation (Lower Permian) of Gondwana basin, Orissa State, India. The microbial mat largely thrived in a lake-deltaic setting in near-shore, very shallow part of the lake and in peri-lacustrine swamps and marginal fluvial regime. Petrographic and stable isotope studies suggest periodic saline (marine?) groundwater input and evaporation under semi-arid condition. A nearby marine setting seems probable. Late Holocene microbial travertines-tufas (dominantly of stromatolitic crusts) occur in parts of Orissa State and appear to be constructed of possible P hormidium, S cytonema,

235
Schizothrix and diatoms. Stable isotopes suggest domi- nantly an inorganic precipitation from freshwater in flu- vial regime of streams and small river.
1 INTRODUCTION PANV,tA (1987) reported some unusual structures in the
'Talchir Group' in the Talchir Gondwana Basin, Orissa and treated them as 'algal nodules'/'oncolites'. This paper docu- ments the microbial signatures in freshwater lacustrine carbonates of the Gondwana Talchir Formation (Lower Permian) and of Late Holocene fluvial travertine-tufa carbonates of Orissa State, India (Fig. 14).
2 GONDWANA (LOWER PERMIAN) LACUSTRINE CARBONATES
2.1 Field Occurrence Laminated crusts/bodies (P1.59/1) with varyingly mixed
argillaceous and carbonate content are encountered in the siliciclastic sequences in different levels towards the mid- dle and higher part of the Gondwana Talchir Formation exposed along the Nandira Jhor stream (within a stretch of ca. 12 kin) between Bedasar Village in the west and Sakasingha Village in the east in close proximity of Angul town (Angul District). These occur mainly as planar/ undulating crust and ovoidal/discoidal bodies (not less than ca. 40 cm in diameter and ca. 15 cm in thickness). Some discoidal bodies show concentric laminations around a central nucleus (PI. 59/2). They are generally observed in siliciclastic siltstone-shale and marlstone-~hale sequences. Locally, they may occur in intervals of darkish shales and as crusts with reddish brown ferruginous coatings and shrinkage/desiccation cracks on siliciclastic sandstone beds. Laminated depositscommonly exhibit fibrous calcite laminae separated by clayey laminae.
2.2 Microfabric Microfabric of the laminated carbonates exhibits lighter
sparry clacite laminae with radial fibrous/bushy' fibrous
aspect alternating with somewhat darker clayey and quartz silt laminae. Little quartz sand may also be seen. The fibrous laminae bear traces resembling poss~le filamentous microbes. The filaments may be entrusted with micrite and microspar and may measure 3-5 lain in diameter. Weakly calcified filaments reveal branching along with mucilagenous drape/exopolymeric substances (EPS) and may locally appear to be dried and contracted. Few illa- ments are 1-2 gm thin and resemble fungal strands. SEM observation reveals bacteria (ca. 1 gm in diameter, P1. 59/ 3) occurring as tiny spherical bodies in clusters and chains and occasionally as isolated forms (nannobacteria?). A spiky fabric is occasionally developed, the spikes measur- ing 2-10 l.tm in length. Occasionally micrite-microspar laminae are separated by darker organic matter. Locally darker cloudy patches, clotted and peloidal matrix occur. While 'bushy' fabric shows a downward conical tip, the larger conical fibrous forms associated in the fibrous crystal-complex exhibit a chevron pattern which may be suggestive of an original selenitic gypsum association (K. I~LTS and C. Motcrv pers. com. to Mol-~ma) which has subsequently been largely replaced by neomorphic calcite pseudospar. Fabric discontinuity is developed due to aggradational growth of fibrous 'bushes' and 'cones' which have pushed/torn the clayey laminae. It is clear that the laminated deposits with varying extent of fibrous calcites have undergone complex diagenetic changes which, we think, have obliterated the microbial structures to a great extent.
2.3 Stable Isotopes and Strontium The results of stable isotope analysis of 20 samples
give 81sO values in the range of-15.38 to -21.54 permil vs PDB and 813C values in the range of-0.88 to -18.23 permil vs PDB. The oxygen isotope data possibly indicate a freshwater origin. However, the highly negative values are much less than the average value for freshwater carbon- ates, suggesting strong diagenetic alteration. Due to bulk
i t 4 ~ o' 66 ~ o'
o , o , o o , .
04"1 o' e6~ o' Fig. 14. Location map.

236
analyses the interpretation of carbon isotopes is very limited. Values in the range of-4.90 to -9.80 permil largely support a freshwater environment in which the carbon supplied to the lake system is largely organic carbon from terrestrial sources and decaying biogenic matter from water column and bottom sediments. However, the values showing relative enrichment in heavy isotopes (i.e. -0.88 to -3.77) may possibly indicate input of heavy carbon isotopes from a saline/marine(?) source. Exceptionally high negative values (i.e. - 16.14 to - 18.23) probably rep- resent strong diagenetic alteration. Average strontium concentration is 168 ppm. Depletion of strontium might have occurred during diagenesis.
2.4 Environment and Depositional Processes The microbialites probably were formed in a freshwa-
ter lake-deltaic setting in very shallow near-shore to slightly deeper water depth (< 1-10 m), in associated peri-lacustrine marshes/swamps and marginal fluvial regime. In general, the microbial mats thrived on flattish substrate in low- energy condition. The fibrous laminae (calcite and prob- ably also gypsum) and clayey laminae indicate seasonal dry and wet periods respectively. The dry, warmer period apparently has favoured growth ot ~ microbial mats when there was very little siliciclastic influx into the lake basin. The mats may as well have trapped and bound some fine siliciclastics. The ridges and furrows and peeling-up of materials in the microbial bodies (P1.59/1) and crusts may be due to the cumulative result of several crustal cycles during which crustal accretion and dissolution might have occurred. The spiky calcite fabric may have developed due to etching of calcite crystals by decomposing cyanobacteria or fungal activity (JoNEs 1987) at a later state. Possible gypsum association may suggest strong seasonality with semi-arid evaporative situation when the microbial mat was periodically impregnated by saline water due to oscil- lation of ground water table. This may perhaps imply the presence of a nearby marine setting (RoucHv & Morcrv 1981). Some cyanobacterial forms may have also trig- gered precipitation of gypsum (THOMPSON & FERRIS 1990). The implication of a nearby marine setting bears signifi- cance in the context of other different marine signatures recently recorded in the Lower Permian Talchir Formation in other parts of India (CrIAm~BAR~ 1993; CHANt)RA 1996; Guru~ ~r AL. 1996; MurmOVADrrtAV 1996).
3 LATE HOLOCENE FLUVIAL (TRAVERTINE-TUFA) CARBONATES
3.1 Travertine-Tufa Complex Fluvial carbonate deposits ('travertine-tufa'; Late
Holocene and Modem) commonly occur as stromatolitic crusts which may form planar and stratified crusts in diverse dispositions, coatings around tree roots and may be associated with phytohermal (moss-rich) deposits at streams and waterfalls. Pisoids (0.4 to 3 cm in diameter) are enclosed in small pockets and lenses between disrupted crusts. At river channel margins coatings on pebbles/ gravels and pillar-like/columnar stromatolitic growths (ca. 5 mm in height) with transverse ridges and troughs occur (P1. 59/4). Modem crusts show laminae crenulation indi- caring a current direction.
3.2 Microfabrie S tromatolitic travertines are constructed of alternating
lighter sparry laminae and relatively darker micritic lami- nae. While sparry laminae in some crusts contain unbranched filamentous forms of Phormidium, others show branching filaments (Scytonema?; PI. 59/5). Phormidium is consid- ered as a predominating cyanobacterium in travertine-tufa (e.g. FREYTET & tSJ~T 1996). The micritic laminae possibly contain Schizothrix filaments. Few sections show cyano- bacteria of a single genus forming erect growth in sparry laminae and prostrat growth in micritic laminae. Thin sections show small, roundish to diffuse clumps (10-150 lain in diameter) in microbial micrite and spar association. Laminated crusts composed of fibrous aragonite show radial, bushy growths and spherulites (possibly microbially controlled; VERRECCrnA et al. 1995). Columnar growths show light and dark microcrenate laminae with filaments. Porous crusts under SEM reveal 'microphyte biofilms' (PEDLEV 1994) constituted of mucopolysaccharides/EPS spreading over filaments. Both calcified and uncalcified filaments are seen, the calcified filaments are generally tubes of calcite.
Apart from cyanobacteria profuse amounts of diatoms are observed. Other microorganisms noted are coccoid cyanobacteria and fungi. Epiphytic cyanobacteria are also seen in moss-rich deposits. SEM exhibits rhombic, spiky and needle cement crystals. In some crusts, neomorphism has led to inversion of aragonite to calcite forming colum- nar and bladed/fibrous crystals.
P l a t e 59
Fig. 1.
Fig. 2.
Fig. 3. Fig. 4.
Fig. 5.
Microbial signatures in Lacustrine and Fluvial Carbonates: Gondwana (Permian) and Holocene Examples, India
Discoidal microbial carbonate body with crude laminations, central part showing ridges and furrows. Permian, note hammer for scale. Part of a discoidal microbial carbonate body showing concentric laminations around a central nucleus (arrow). Bacterial bodies of Permian age (arrow) in cluster (lower left) and chain (central part). Stromatolitic travertine (Holocene) showing crowded pillar-like/columnar growths. The columns taper upwardly and show transverse ridges. Micritic to microsparry crusts on cyanobacterial template of filaments (Scytonema?) and mucilage (arrow). The mat appears dried and contracted. Holocene

P l a t e 59 2 3 7

238
3.3 Stable Isotopes and Strontium The results of stable isotopic analysis show 813C values
varying from -12.56 to -2.93 permil vs PDB and 81sO varying from -7.16 to -2.00 permil. Sr concentrations vary from 80 to 3100 ppm, the high values representing domi- nantly aragonitic crusts. The range of 813C composition suggests varying degrees of biogenic CO 2 input (ANDREWS et al. 1993) from the soil zone to the bicarbonate precur- sors. Therelatively highervalues may indicate disequilibrium isotopic effects or some (unknown) heavier carbon isotope source. Photosynthesis might have partly enriched travertines in heavy isotopes. The ~i lsO composition of travertines is consistent with precipitation from freshwater. Diagenetic inversion of aragonite to calcite might have resulted in some depletion of heavy isotopes and strontium.
3.4 Depositional Conditions The outgassing of CO 2 from stream water leads to
supersaturation with respect to calcite (or aragonite) and consequent precipitation. The isotope data suggest domi- nantly an inorganic mode of precipitation from the solu- tion. The mucilagenous sheath of cyanobacteria and dia- toms provide a template for surficial mineralization. Bushy and spherulitic fabrics suggest that cyanobacteria and/or bacteria initiate precipitation through photosynthesis and later growth may be of inorganic origin.
The precipitation in response to seasonal changes may correspond to concomitant growth of two genera or single genus with two styles of growth, which form seasonal laminae. The morphology of microbial crusts seem to depend on substrate slope, microbial growth and water current.
PART XIII BIOTIC RESPONSE TO MASS EXTINCTION:
THE LOWERMOST TRIASSIC MICROBIALITES
A. Baud, S. Cirilli and J. Marcoux
1 INTRODUCTION A report on lower Triassic stromatolites as post-mass
extinction disaster forms have been presented by SCmmF.RT & BOI"I'JER (1992). But the main description concerns late Olenekian (Spathian) stromatolites of the Great Basin (NW USA) and not the lowermost Triassic microbialites. 'Tethys just after the end of Permian mass extinction' was the subject of a presentation at the Pangea Conference in Calgary (BaUD 1993). Working on upper Permian paleomap (BAtrD et al. 1993a,b), we can show that one of the main characteristic of the upper Permian paleoenvironment of the Tethys is the growth of giant shallow carbonate plat- forms. High diversity marine fauna and flora with prolif- eration of calcareous algae, foraminifera, echinoderms, bryozoan, brachiopods, and bivalves characterize the car- bonate factory. The latest Permian second order T-R cycle is well recorded on the NW Cimmerian margin (BAUD 1993) in S Armenia (Vedi, Sovetachen), adjacent NW Iran (Dorasham, Dzhulfa, Ali Bashi, ROSTOV'rSEV & AZARIAN
1973, KOTL~'AR et al. 1983)), Elburz (ALrtr~a et al. 1980) and in central Iran (Abadeh, IRANIAN-JAPANESE R.G. 1981).
The natural boundary between Permian and Triassic corresponds to the maximum regression, that is the maxi- mum basinward shifts of coastal onlap. Exposure of large shallow shelves which dramatically reduced the habitat area, oxidation of a great amount of organic carbon, and release of gas hydrates give way to a hyper-weakening of the ecosystem and a devastating extinction. These events and global change, recorded in the large ~13C shift (BAUD et al. 1989), strongly affected the carbonate productivity, stratal patterns and biofacies. A bloom of disaster forms: -fungi (EsrmT et al. 1995), -cyanobacteria, is the intrinsic biotic response to the mass extinction.
During the first step of the very rapid and large scale lowest Triassic transgression, we note in different areas (S Alps, Taurus, Turkey, S Armenia, E Elburz, Iran, Central Iran and Central Afghanistan) the growth of domal stromatolites, thrombolites and other microbial structures. At the dawn of Triassic time, the carbonate factory was dominated by non-skeletal species and by microrganisms able to precipitate carbonate. True reef communities were absent in the earliest Triassic strata and they were locally substituted by microbialite mounds. In fact since reefs are composed of stenotopic organisms they are very vulner- able to changing environmental conditions, but not when they are formed by eurytopic organisms (HALLA~,11994). In the following chapter, we will examine three cases show- ing different macro- and microfabrics according to paleo- environmental setting: the Curuk Dagh profile, a shallow carbonate platform on the Gondwanian margin, and the Sovetachen and Vedi profiles, a pelagic carbonate ramp on the NW Cimmerian margin. We will give a comparison with the Abadeh area (Central Iran).
2 MICROBIALITES ON A SHALLOW WATER CARBONATE PLATFORM
2.1 Setting The Curuk Dagh section (Fig. 15) is situated about
15 km Northwest of Kemer (Antalya nappe, SW Turkey). A general description is given in MARCOUX & BAUD (1986) and/VIARCOUX et al. (1986), for stable isotope data see BAtro et al. (1989).
The upper Permian Pamucak Formation consists of dark, partly cherty, nodular lime wackestone (1 in Pl. 60/ l) with three levels in the upper part rich in brachiopods. Based on fusulinids, small forams and brachiopods, the age is late Dzhulfian - early Changhsingian. This Forma- tion is capped by 30-60 cm of oolitical grainstone (2 in Pl. 60/1) affected by vadose diagenesis. Directly above begins the Kokarkuyu Formation with the first level showing typical domal stromatolitic structures about 40-60 cm in thickness (3 in Pl. 60/1). The following beds are thicker (up to 1 m) and show thrombolite textures. Abundant lower Induan (lower Triassic) microforaminifera of the genera Rectocornuspira, Earlandia, Cyclogyra and Aramodiscus characterise these basal Kokarkuyu deposits.
The stromatolitic-microbialites deposits are concen- trated within the first 12 m of the Kokarkuyu Formation.

239
slaCpos Age Lithologyand i , ~ , ~ , ; , ~s Paleoenvironments groups
)olites t
shallow shelf colonized (9 _<~
nar ~ ""i i |tolites < by microblalltes --~
d micrites ~o_ ~ "~ co _~ : ~ o ~ . ~ ~ ~
natolites i . , Transgressive systems tract ~ ~ ~ ~ ~. grainstone: ~ sBhigh energy ~ nodular ! low energy 2R I
limestone:' shallow shelf i _ _ s I extinction
high diversity I skeletal " skeletal community ackestone 1 I highstand
813CPDB isotope c u r v e (after Baud & el. 1989) " I
R a n g e c h a r t o f m a i n f o s s i l
Fig. 15. Detailed stratigraphy, 813C isotope curve, facies and paleoenvironments of the Permian-Triassic boundary section at Curuk Dagh (Antalya, Turkey)
The main observed textures are described in the following chapters.
2.2 Macro- And Microfabrics In this section, microbialites form mounds up to 40 cm
thick. This type of microbialite can have a planar or stromatolitic (LLH-type like) growth formed by microbial laminae with fenestral fabric; it consists of regular alterna- tions of layers of cements with dark micrite (PI. 60/3). The intercalated layers are composed of dark fine grained bioclastic micrite containing peloids, ostracods, small forams, and/or clotted micrite with remains of twisted filaments and with pseudo-peloids, cemented by microsparite and sparite. The larger cavities are bordered by micro- spherulitic cements (Rivularia-like microbial colony) and completely infilled by micrite or peloidal micrite and/or fine grained bioclastic micrite (PI. 60/4). Fringe cements (2 or more generations) and/or drusy cements infiUed the remaining fenestral porosity. Small forams (Cornuspira mahajeri-Rectocornuspira kalhori assemblage) and Earlandia spp. are abundant and sometimes trapped within crystals, or accumulated parallel to the laminations (PI. 60/ 5). Ostracods and gastropods are also present; the latter are often coated by several generations of fringe cements. Synsedimentary microfractures are present, infilled by the intercalated fine sediment and by fragments of the layers
of cements. They testify to early cementation due to biochemical processes. The associated thrombolite type is composed of clotted micrite which shows traces of straight to undulose micritic filaments, sometimes forming oncoidal- type fabric (PI. 60/6).
3 MICROBIALITES ON A PELAGIC CARBONATE RAMP
3.1 Setting The Sovetachen and the Vedi profiles (Transcaucasia,
S Armenia) are situated in Armenia, about 45 and 60 km SE of Yerevan. They have been descibed by KO~YAR et el. (1983) and by ASt.A~_AN (1984) and work on stable iso- topes is found in BAUD et el. (1989).
In the Sovetachen section, red marls corresponding to the latest Pseudotirolites zone of the Permian overly the Paratirolites red nodular limestones. The following basal Kara Baglyar Formation (Induan, lower Triassic) consists of bedded platy lime mudstone in alternation with microbialites, and its base is dated as the lowermost Triassic H. parvus zone.
In the Vedi profile, overlying the Paratirolites lime- stones (1 in PI. 60/2) locally there are about 5 cm of red marls, corresponding to the latest Pseudotirolites zone of the Permian. The following basal Kara Baglyar Formation (Induan, lower Triassic) consists of 2 m of domal micro-

2 4 0
bial biolithite (2 in PI. 60/2), overlain by bedded platy lime mudstones and microbial limestones. H. parvus also oc- curs in this section.
3.2 Macro- and Microfabrics In these two sections microbialite carbonates are present
as isolated small patches associated with the more com- mon thrombolite types. Locally, microbialites form mas- sive mud mounds up to 2 m high (Vedi section). Laterally, lime mudstones frequently contain floatstones of fine and medium coarse debris sourced from the microbialite mounds. In Sovetachen section they occur as nodules within reddish and greyish wackestone/packstone. In some cases they represent reworked material coming from lateral in situ mounds: crystals are often broken, folded, and disoriented (PI. 60/8); in other cases, the gradual passage between nodules and matrix and the crystal orientation suggest an in situ growing of the elongated crystals in small fan arrays within the muddy matrix.
In Vedi section, microbial textures exhibit the presence of elongated crystals in fan arrays; they form around single filaments or as bundles of parallel filaments (Pl. 60/7). Thicker, bedded platy lime mudstones show clotted micrite with recognizable cyanobacterial filament remains, some- times arranged to form oncoidal type-like structures. Fenestral cavities are filled by sparitic cements. This type of microbialite usually encrust muddy sediments (wackestone and mudstone, rarely packstone) containing ammonoids, thin shelled bivalves, ostracods, small gastropods and rare foraminifers.
4 COMPARISON WITH ABADEH (CENTRAL IRAN)
A detailed survey of the Permian-Triassic boundary beds in the Abadeh area (Central Iran) has been published by IP~aA~-JAPA~SE R.G. (1981). As in Transcaucasia, there is a plurimetric thick zone of'algal biolithite' with H. parvus just above the latest Permian Paratirolites lime- stones. Profiles through the 'stromatolites-thrombolites' unit and microfacies ilustrations are given in I ~ A N - JAPAtZZSE R.G. (1981: Fig. 11, PI.5). We can emphasize the great similarity with the Sovetachen section which is located about 1500 km to the NW. B^unmAr,a (1993) named this basal Triassic unit the 'Thrombolite zone'. According to this author, this "rhrombolite zone' occurs also in the basal Dalan formation of the Zagros Range. This indicates the widespread occurrence of microbialites within the basal Triassic sediments.
5 MASS EXTINCTION AND THE CONQUEST BY DISASTER FORMS
Post-mass extinction aftermath and recovery periods may be times when metazoan-imposed barriers to the nearshore normal marine environments were removed, so that the opportunities for microbial communities to thrive in such settings might increase. Their presence indicates for the Early Triassic a period characterized by drastic diversity reduction and elimination of major taxa from shallow water carbonate environments. As eurytrophic and/or primitive groups, microbial communities emerged
P l a t e 60
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig. 8.
Biotic response to mass extinction: the Lowermost Triassic microbialites
Permian-Triassic boundary (top of GSA scale) at the Curuck Dagh section, l=nodular limestone of the Pamucak Formation. 2='Tesero type' oolitical grainstone. 3=domal stromatolites Permian-Triassic boundary (top of the hammer) at the Vedi 2 section, l=Paratirolites grey nodular limestones. 2=base of the of 2 m thick domal microbial biolithite Curuk Dag section: microbial laminae showing fenestml fabric composed by regular alternation of layers of cements with dark micrite. Cements are formed by elongated crystals growing in fringes and/or in fan arrays. The intercalated layers are composed of dark fine grained bioclastic micrite containing peloids, ostracods, small forams, and/or clotted micrite and pseudo-peloids. Synsedimentary microfractures are present, infilling by the intercalated fine sediment and by fragments of the layers of cements, x 6 Curuk Dag section: the larger cavities are bordered by microspherulitic cements (Rivularia-like microbial colony) and complete infilling by micrite or peloidal micrite and/or fine grained bioclastic micrite, x18 Curuk Dag section: small forams (Cornuspira mahajeri-Rectocornuspira kalhori assemblage) and Earlandia spp. are abundant and sometimes enclosed within crystals or accumulated parallel to the lamination, x 85 Curuk Dag section: oncoid-type masses formed by clotted micrite which shows traces ofmicritic filaments straight or undulating. The sediment is cemented by microsparite to sparite, x 10 Sovetachen section: small digitate microbial stromatolite, composed of alternating lamellae. They exhibit a primary or early diagenetic radial fibrous fabric, which presumably formed around single filaments or a bundle of sub-parallel filaments, x 10 Sovetachen section: small microbial patches as nodules within the reddish and greyish wackstone/ packstone. They represent reworked material coming from lateral in situ mounds: crystals are often broken, folded, and disoriented, x 6
P l a t e 6 0 2 4 I

242
from stressed palaeoenvironments to recolonize the rela- tively vacant normal marine settings (r-selected generalists and opportunists). The main causes of their rapid and wide colonization by microbial communities of marine enviroments could be summarized as follows:
- The absence or great reduction in diversification of metazoans; it resulted in an ecologic relaxation caused by the diminuition of natural selective pressures (predation and competion, VERMmJ 1987). In these palaeoecologic conditions biofilms and microbial mats could spread in environments where extreme conditions exclude most of the most evolved species. The absence of predation and of intense bioturbation allowed the preservation of microbial laminated structures (AWRAMm 1990).
- Decreased space competion for substrate coloniza- tion. Following the Permian-Triassic mass extinction, the proliferation of disaster forms should occur in the habitat where extinctions were most pronounced. Microbial com- munities were facilitated to spread in marine environments depauperated by invertebrate paleocommunities (ScmraERT
BoTrJER 1992). - Low sedimentation rate, due to reduced amount of
skeletal debris and biogenic carbonate sediments linked to the severe biotic crisis; it prevented the burial of microbial mats and it resulted in an increase of the organic matter concentration. The higher concentration of organic com- pounds on marine bottoms provides a highly satisfactory nutrient source for the growth of microbial communities (ScHULTZE-LAM et al. 1993).
- Major changes in atmospheric and water column oxygen content and/or sea water carbonate concentration; biofilms and microbial mats occur in environments where extreme physical and chemical conditions exclude most of the most evolved species. This is because microbial eco- systems form complex communities which have devel- oped a wide spectrum of metabolic functions: autotrophic or heterotrophic species, aerobic or anaerobic species. These characteristics confer on the microbial world an extraordinary capacity to proliferate quickly in media- fluctuating physico-chemical conditions and to adapt them- selves to a various kinds of environments.
The microbial conquest of the earliest Triassic marine bottoms had three main effects as immediate consequences.
1 - Producing copious mucus and including filamentous microbes, microbial communities trapped and bound de- trital grains, strengthen the cohesion of sediments and build biolaminated deposits and microbial mounds. The covering and the mucilage of the microbial mats stabilized sediments and favoured preservation of non-skeletal struc- tures.
2- The microbial proliferation gave way to a oxygen- deficient environment. On the bottom of stagnant water masses, the microbial communities brought anoxic condi- tions which led, in extreme conditions, to the sedimenta- tion of bituminous shales. In this sense, the early Triassic anoxic event (HALLAM 1994) is a consequence of the mass extinction, not the cause. The earliest Triassic rocks are either completly devoid of benthos or have thin horizons
containing fossils low in diversity and high in density. Reef communities and other benthic communities of high diversity were absent in these deposits (Reef gap, FLt3CEL 1994).
3- Inducing mineralizations the microbial community plays a leading part during the fist steps of diagenesis and during fossilization, particularly by promoting the preser- vation of soft-bodied organisms. Precipitation of calcium carbonate is first induced actively in the bacterial mem- branes providing the early cementation of the entire struc- tures (LEADBEA~R & R i o ~ 1986; SCnrCetDER~WrN & HARRIs 1985; REIa'r~R 1993; GALL 1995).
The biotic recovery after the Permian-Triassic mass extinction was very slow and the full recovery of the skeletal carbonate factory did not occur until the middle Triassic. The physico-chemical parameters influenced the nature and the availability of refuges and the composition of the taxa available for the new radiation. The entire early Triassic was characterized by simple, cosmopolitan, low diversity, low complexity paleocommunities (ScriLram~r & BOTUER 1995). Diversification and regrouping of stable, ecologically complex associations does not really happen until stenotopic organisms emerge from refuge (k-selected organisms).
Part XIV FACIES ANALYSIS
IN UPPER JURASSIC STROMATOLITES: SUPPORT BY PALYNOLOGICAL AND STABLE
ISOTOPE DATA
M. Merz-PreiB
Growth and surrounding environment of Upper Jurassic stromatolitesfromNorth Germany have been reconstructed by isotope and palynological data. The stromatolites are embedded in thick marls. This circumstance lead to the continuation of anoxic conditions in the sediment through- out diagenesis. Trapped palynomorphs and plant detritus show an exceptionally good preservation. The documented plant communities allow a reconstruction of the surround- ing environment. The stromatolites seem to have grown in a restricted, marginal marine environment, probably in the intertidal or shallow subtidal zone. Light 813C-values support the interpretation of growth within a restricted lagoon.
1 INTRODUCTION Although a somehow restricted environment is often
assumed for the growth of stromatolites, a detailed recon- struction is often difficult. Various methods such as facies analysis or stable isotope measurements might contribute to a reconstruction. In Upper Jurassic stromatolites from northern Germany palynomorphs proved to be a suitable indicator for the paleoenvironment. This should be a more general feature in stromatolites, because they frequently trap fine grained sediment. Due to the typical stratification of the microbial mats, internal anoxic conditions support-

243
ing the preservation of organic material should have been a common feature of many stromatolites. Where anoxic conditions prevailed throughout diagenesis, palynology might be a valuable tool to support facies analysis.
2 METHODS Samples of the stromatolites have been dissolved in
weak acetic acid. The insoluble residue has been examined for plant debris, crustacean remains and crystal aggre- gates.
Samples for pollen analysis were prepared by dissolv- ing 10 g sample over night in 100 ml HCI (37%), then in 45 ml concentrated HF for 12 hours and again 50 ml HC1 (37%) for 1 hour. Between the treatments the samples were centrifuged (10 min, 4000-5000 rpm) and the acid de- canted. The residue was washed with water, and sieved after a short ultrasonic treatment.
The isotopes were analysed in Erlangen, Germany, Institute for Geology and Mineralogy, in the mass spectrometer laboratory of Prof. Dr. W. Buggisch by Dr. M. Joachimski. Results are given as permil vs PDB.
3 RESULTS 3.1 Setting ofstromatolites
The stromatolites occur in a single horizon within the Upper Jurassic (Portlandian) Mtinder marls located in an area close to the village of Thtiste, Lower Saxony, Ger- many. The sequence has been described in detail by JAHNKE & Rrrzxowsra (1980). A massive, about 13 m thick bed of serpulite is followed by an interbedding of marls with micritic and oolitic limestones. The stromatolites occur in the top part of the sequence, followed by a final marl bed. The sequence is interpreted by JArINKE & Rrrzgowsra (1980) as an increasingly restricted, marginal marine envi- ronment. The serpulite and oolite beds are meant to repre- sent shoals in a shallow marine environment, which pro- tected a landward lagoon. In strata below the stromatolites indicators for an evaporitic environment can be found. The stromatolites grew in the intertidal zone rimming the lagoon. The ostracod fauna in the overlying marls indi- cates brackish conditions.
The stromatolites vary in diameter from 10 to 100 cm. Their surface is brain-like undulated, the sides are smooth (PI. 61/4, 5). In cross section, two growth stages can be seen. The cores are often made up of small serpulid col- onies, overgrown by thick stromatolitic crusts (PI. 61/4).
3.2 Stable Isotopes Along a transect through the laminated facies of a
stromatolite 14 analyses of stable carbon and oxygen isotopes were made. While the oxygen isotopes, with 5180 falling in a range between -2.72 to -4.68 permil show about marine values, the 813C-values (-8.25 to -16.26 permil) of the stromatolites are lighter than should be expected for normal marine equilibrium precipitation. The dissolved inorganic carbon (DIC) of the water must have been considerably enriched in 12C. Various processes, related to microbial activity, could have been responsible for this enrichment.
One possible process is sulfate reduction in the anoxic layer of the microbial mats, generating C-isotopically light HCO 3" out of the land derived plant material (JORGENSEN 1983; LAZAR et al. 1989). The high rate of sulfate reduction in the anaerobic layers of the microbial mats is shown by the abundance of sulfides. This process alone, however, might not be sufficient to account for the light 813C values measured (LAZAR et al. 1989).
Another possible explanation for the light carbon iso- topes might be found in high rates of photosynthesis in the upper, probably cyanobacteria-dominated parts of the mats, which might cause the carbon isotope fractionation be- tween the atmosphere and the water to be similar to the fractionation between air and a hydroxide solution (BA~a~TSC~ 1952; HERCZ~G & FAIP.UANXS 1987; LAZAR & E~EZ 1990), resulting in a DIC-POol as light as -15 permil (BAm~TSCm 1952). Although the reaction of CO 2 with OH should be of no importance in normal marine environments (I-Im~czEG & FAmBANKS 1987) it could well be of importance in a restricted, possibly hypersaline or brackish lagoon (LAZAr & E~Z 1990).
Still, the lowest measured ~13C value of -16.25 permil is very light. For a theoretical DIC of -15 permil the carbonate should show values around -13 permil. A com- bination of the two above mentioned processes, with a possible addition of 12C from oxidized organic material (BEIN 1986) could explain the overall light 813C values of the stromatolites.
The oxygen and carbon isotopes in the samples corre- late well (r=0.93, n= 14). For lacustrine carbonates, TALBOT (1990) interprets a good correlation between 813C and 8180 (r >0.7) as a sign for deposition within a closed lake system. In the Thtiste stromatolites the correlation might therefore reflect the environmental conditions of a re- stricted lagoon with extended residence times. With in- creasing residence time due to restricted exchange with normal seawater, there is time for photosynthesis to enrich 13C in an overall very light DIC-pool or time for the water to approach air equilibrium during slower rates of micro- bial activity (BEIN 1986). AS evaporation of sea water in a restricted lagoon would have resulted in heavier oxygen isotopes, the measured oxygen isotope values might indi- cate influx of evaporated fresh water, which can result in 8180 values typical for marine carbonates (MERZ 1992). This fresh water influx is also shown by an association of spores which is typical for aquatic transport.
3.3 Organic material The stromatolites contain a high amount of well pre-
served organic material. A piece of wood, about 7 cm long, has been found on the surface of one of the stromatolites. Much more abundant, however, is microscopic plant de- bris and palynomorphs trapped by the microbial mats. PI. 61/6-9 give an impression of the excellent preservation of the material.
The dinoflagellates document sediment influx from the normal marine realm into the lagoon. On the other hand there has been considerable influx from the land, shown

2 4 4
not only by the wood debris, but also by spores and pollen. Under the SEM the wood cells can still be seen and even the pit connections between cells are preserved (PI. 61/9). The wood debris has been brought into the lagoon by fresh water runoff, either seasonally or by a permanent river. In probably the same way the trilete spores of the ferns and mosses have been transported. Windblown pollen grains from Gymnosperms and Araucariaceans, prefering well drained, slightly elevated areas, also occur abundantly in the stromatolites. The flora along the shore line is repre- sented by pollen of Cheirolepidaceans, possible swamp vegetation by Taxodiacean-pollen. Inbetween, various ferns, Lycopodiaceans and Sellaginellaceans grew.
The palynomorphs found in the stromatolites fit well with the environment of a restricted lagoon, with occa- sional fresh water influence, surrounded by brackish marsh lands with elevated areas in the proximity.
The only remnants of a macrofauna besides serpulids in the stromatolites are the abundant small pieces of crusta- cean exoskeletons found in the insoluble residue. They are, however, generally to small for an exact determination.
4 DISCUSSION The stromatolites from Thiiste Seem to have grown in
a restricted, marginal marine environment, probably in the intertidal or shallow subtidal zone. The association of stromatolites and serpulids has been described several times and been interpreted to be a marginal marine or brackish environment (LEEDER 1973, 1975 (but compare BuRcHe-l-llz & RmINO 1977); P~RVT 1974; TOOMEy & CYS 1977; WRlGrrr & MAVALL 1981; DAVALrD et al. 1994). The serpulids were taken as the main indicators for a restricted environment, but the very limited associated fauna also indicates restricted conditions.
The most similar occurrence seem to be subrecent (5500 y.b.p.) stromatolites described by DAVAtrO et al. (1994) from a lagoon in Tunisia. The authors notice a distinct facies change from a core facies formed by serpulid
reefs to an outer facies of stromatolitic crusts. They ascribe this facies change to the change from euryhaline to hypersaline conditions in the lagoon with dropping sealevel. There are no indications for the type of environmental changes responsible for the overgrowth of the serpulids with stromatolitic crusts in the Thfiste stromatolites, but changes in salinity could be a plausible reason. Because of the brackish ostracod fauna described from the overlying marls (JArn, a~ & Rrr2xowsr~ 1980), however, a change to hypersaline conditions seems not very likely.
The lithologies described by DAVAUD et al. (1994) are also very similar to the facies found in Thtiste. They found oolites and bioclastic sands, forming beach ridges, protect- ing the lagoon. Within the lagoon, black laminated shales were deposited, while the stromatolites grew along the coastline. Overall, the facies they describe from the Holocene might have been very similar to the Late Jurassic facies of ThiJste.
5 CONCLUSIONS 1. The isotopic composition falls into ranges between
513C: -16.26 to -8.25 and ~5180:-2.72 to -4.68. The ~80 values indicate about equilibrium conditions for pre- cipitation. The 613C values are lighter than equilibrium precipitation from normal seawater. This could be due to strong photosynthesis by the cyanobacteria and bac- terial sulfate reduction. There is a remarkable good correlation between the carbon and the oxygen isotope values (r = 0.92).
2. In stromatolites containing well preserved organic ma- terial palynology can be a valuable tool for reconstruct- ing environmental conditions. In the stromatolites from Thfiste the palynomorphs record marine influence as well as a nearby coastal vegetation and elevated coastal areas.
3. Data presented here support the interpretation that the Th~iste stromatolites developed in a restricted, marginal marine, intertidal to shallow subtidal environment.
P l a t e 61
Figs. 1-3.
Fig. 4.
Fig. 5. Fig. 6.
Fig. 7.
Fig. 8. Fig. 9.
Upper Jurassic stromatolites of Thtiste, Lower Saxony, Germany.
Thin sections through the finely laminated outer parts of a stromatolite. This rim facies seems to contain calcified filaments (arrows in Figs. 1 and 2), which either grew in little bushes, preserved as micritic tubes in cemented holes, or they grew errect on the surface of the stromatolite (Fig. 3). Fig. 1 is cut tangential to the stromatolite surface, Figs. 2,3 are cut vertical to the surface. (Width of Fig. 1 is 1 mm, of Figs. 2, 3 it is 3.5 mm). Cross-section of a stromatolite, showing the to different growth stages. An inner stage with abundant serpulites and micritic clasts is followed by a stage of micrite accretion, probably by cyanobacteria- dominated microbial mats. The diameter of the stromatolite is 22 cm. Brain-like surface of a stromatolite. The shown section of the stromatolite is about 12 cm wide. Dinoflagellate (Rigaudella aemula (DF.~_.~Nt)RE) BELOW 1982), indicating marine influence in the environ- ment. x 710 Callialasporites turbatus (BAL~) ScHtrt.Z 1967 is a Sellaginella spore. Recent Sellaginellaceans grow in humid tropical forests, x 710 Pollen of a gymnosperm, transported by wind, possibly over a large distance, x 710 Wood debris, probably of a gymnosperm. The pit-connections between the cells are still clearly visible.
P l a t e 61 2 4 5

2 4 6
Part XV UPPER CRETACEOUS AND LOWER TERTIARY BRACKISH TO FRESHWATER ONCOIDS AND
STROMATOLITES OF THE GARUMNIAN FACIES IN THE/kGER BASIN (CENTRAL SOUTHERN PYRENEES/SPAIN)
H. M~tcker
Some new outcrops in the terrigenous sequences of the Garumnian facies of La Vail d'Ariet in the southeastern part of the Ager Basin (Central Southern Pyrenees~Spain) show many different growth types of oncoids and stromatolites. They can be used together with the host rock litholo gy and the micro fossil record as good indicators for the depositional history of alluvial, fluvial and deltaic deposits of the Garumnian facies. The different forms of stromatolitic structures are suitable to distinguish several depositional zones of a fluvial system and to sketch recon- struction models of the alluvial parts of the Garumnian: i) oval round and "ear-shaped' concavo-convex oncoids de- posited at the channel base of a river, ii) linked hemispheroidal domes grew near the mouth of a river, iii ) oncoidal fragments are bound to the conglomeratic base of a channel and to channel levee deposits, iv) cauliflower- like oncoids,flat concavo-convex and small elongate struc- tures are typical for abandoned channels or shallow mar- gins of meandering rivers, and v) discrete vertical stacked hemispheroids developed in alluvial ponds under reduced terrigenous input.
1 INTRODUCTION The Ager Basin is an asymmetrical syncline in the
southern part of the Central Southern Pyrenees (Fig. 16). The northern flank has been implicated in the overthrusting of the Montsec. The basin is filled with Mesozoic (pre- dominantly Cretaceous) and Tertiary sediments.
The Garumnnian ranges from the Maastrichtian to the lowermost Eocene. The term has been introduced by LEYraERm (1862) for 'epicretaceous' sediments in the Petit Pyr6n6es (southern France). The lithostratigraphic limits in the study area correspond to the top of rudist bearing limestones of the Bona Formation (sensu Mzv et al. 1968) and at the top to Alveolina limestones of the Ager Forma- tion (sensu Ltrrm~AcrmR 1969). The Garumnian facies in La Vail d'Ariet is characterised by several changes from alluvial to lacustrine and paludine regime. Stromatolites and oncoids are restricted to the alluvial deposits which mainly consist of fluvial to deltaic sandstones and con- glomerates and partly tidally influenced alluvial red beds. Former investigations ofoncoid-bearing sediments in north- eastern Spain and southern France have been made e.g. by ANADON • ZAMARREIq'O (1981), FRE'tTEr & PLAZIAT (1982), LLOMPART & KaAUSS (1982), NICKEL (1983 ), and BERaXAm)- SARFAT~ et al. (1994).
There are two terrigeneous sequences in the working area: one in the Maastrichtian (Gansserina gansseri zone and Abathomphalus mayorensis zone) and another one during the Upper Paleocene (Thanetian).
2 LOWER TERRIGENEOUS SEQUENCE The Lower Terrigeneous Sequence is limited at its base
by lacustrine charophyte limestones which form the first Garumnian deposits in the study area and at its top by paludine limestones of Danian age.
This sequence is dominated by coarse grained cross- stratified and partly graded fluvial to deltaic limy quartz sandstones and conglomerates which indicate a high en- ergy deposition. They contain freshwater oncoids which form concentrically stacked spheroids. Space-linked hemispheroidaI stromatolites occur near the top. Dinosaur bones and lignitic rests of plants are bound to the conglom- eratic base of channel fills. Gastropods are very rare.
P l a t e 62
Fig. la/b.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Stromatolitic structures of the Garumnian. Upper Maastrichtian to Upper Paleocene. All samples from La Vail d'Ariet, Ager Basin, Central Southern Pyrenees, Spain.
~Ear-shaped' concavo-convex oncoids of the Lower Terrigeneous Sequence. The nucleus consists of a disarticulated and broken shell of Unio. Note the alternation of light coloured carbonate-rich and dark coloured organic-rich laminae. (xl resp. x 1.5) Coated pebble in a basal conglomeratic layer. The core consists of a marine limestone of probably Cretaceous age with foraminifera. The pebble is surrounded by fragments of oncoids, different limestone clasts and Microcodium. Upper Terrigeneous Sequence, Interval A. Cauliflower-like oncoid which shows in one part directed growth between two oncoidal phases caused by decreased water current. Upper Terrigeneous Sequence, Interval B. Scale bar = 1 cm Small elongate structures which are interpreted as algal incrustations around roots or little branches. Upper Terrigeneous Sequence, Interval A. Scale bar = 1 cm Loaf-shaped stromatolite which form discrete vertical stacked hemispheroids. They developed in colour- mottled reddish marls which are interpreted as the top layer of an alluvial pond. Upper Terrigeneous Sequence, Interval B.

P l a t e 62 2 4 7

248
Fig. 16. The Garurnnian facies in the Central Southern Pyrenees (modified after G,~cI^-V~a.t~s et al. 1993)
Silty bioturbated red beds are rather rare and contain foraminifera (e.g. Globotruncanita conica,Abathomphalus intermedius and Abathomphalus mayorensis), ostracods (e.g. Frambocythere tumiensis), charophytes and gyro- gonites, Microcodium and corroded quartz grains.
Most of the oncoids are oval and concavo-convex ('ear- shaped') because the cores consist of disarticulated and fragmented shells of Unio (PI. 62/1a,b). The autochtonous malaco-fauna demonstrates the freshwater conditions (FRLa'~T & ~.~zaAr 1982). The oncoids have an undulat- ing surface. Other core materials (e.g. dinosaur bones, gastropods like Melanopsis or limestone clasts) have been described by other authors for the Pyrenees (LLo~,u'AR7 & KRAUSS 1982; F~CrET & Pt.~ZIAT 1982) but were not found in the study area. Sometimes the original nucleus is re- placed by white chalky micrite so that the broken oncoid shows the mould of the mussel. All oncoids show an alternation of light coloured carbonate-rich and dark col- oured organic-rich laminae. Vertical holes are filled with detritus (quartz grains and clay minerals). The sorting of the oncoids is very good. The oncoids of one single bed have almost the same diameter. The size ranges from 1.5 to 10 cm. On some plains the oncoids are enriched by weathering and cover almost the whole surface of an outcrop.
The stromatolites are restricted to one small outcrop near the top of the Lower Terrigeneous Sequence and form linked hemispheroidal domes with a maximum height of 50 cm. They probably developed near the mouth of a river and are together with the foraminifera of the alluvial red beds evidence for marine influence. The top of the Lower Terrigeneous Sequence is formed by a thin caliche soil which passes into rhizolitic limestones of the Lower Paludine Limestones.
The depositional environment of the Lower Terrigeneous Sequence was a fluvial to tidally influenced deltaic com- plex which was connected with a marine influenced allu- vial plain will] paleosoils and ephemeral freshwater lakes.
3 UPPER TERRIGENEOUS SEQUENCE The Upper Terrigeneous Sequence is limited at its base
by the Lower Paludine Limestones and at its top by the Upper Paludine Limestones which are the youngest Garumnian beds in the working area.
The facies of the Upper Terrigeneous Sequence is more variable than the deposits of the Lower Terrigeneous Sequence. The oncoid-bearing fluvial limy sandstones and conglomerates are of minor spreading than in the Lower Terrigeneous Sequence but they are rather impor- tant to interpret the environmental settings of the Upper Paleocene. The Upper Terrigeneous Sequence is com- posed of two intervals which are separated by a sequence boundary.
3.1 Interval A Interval A is predominantly composed of alluvial
sandstones, siltstones and silty to marly paleosoils with pedogenetic features like colour mottling, root casts, root moulds and Microcodium. Lacustrine limestones are very rare and restricted to ephemeral lakes of some tens to 100 m in diameter. The oncoids developed in single-channeled meandering rivers.
A correlation exists between the host rock lithology and the degree of preservation of oncoidal structures. Thin-sections ofpolymict conglomerates which often form the base of a fluvial section only show small fragmented oncoids and ooid-like oncoids (sensu FREV'I~T & PLAZIAT 1982, coated pebbles sensu NICKEL 1983) (PI. 62/2). The

249
Fig. 17. Schematic diagram of the Thanetian environmental settings and the distribution of oncoids and stromatolites. Facies which is not exposed in the study area but described by other authors in the Pyrenees is marked with an asterisc.
cores of these coated pebbles consist either of extraclasts or of small oncoidal fragments (even the alluvial red beds contain small fragmented oncoids, caused by levees which transported channel sediments onto the flood plain). Flu- vial limy sandstones deposited in meandering single-chan- nelled rivers of lower current contain oval to spherical oncoids with a maximum diameter of 10 cm. The nucleus mostly consists of Garumnian limestones. The internal structures are very similar to the oncoids of the Lower Terrigenous Sequence. The oncoids incorporate in vertical holes detrital grains (quartz and clay minerals) and frag- ments of Microcodium.
Interval A is capped by a caliche soil - the so-called Colmenar-Tremp- Horizon-which is a widespread paleosoil in the whole Central Southern Pyrenees and marks an important regional sequence boundary.
3.2 Interval B Interval B is more influenced by transgressive proc-
esses. There are two intercalations with brackish, marine lagoonal and hypersaline facies associations, e.g. packstones to grainstones with miliolids, rotaliids, ostracods, red algae of the genus Distichoplax biserialis and echinoid debris. Oncoids respectively stromatolites are restricted to two horizons.
The first outcrop contains cauliflower-like oncoids an d oncoidal stromatolites with a maximum diameter of 30 cm and flat concavo-convex structures with a maximum diam- eter of 10 cm which may be coatings around bark or leaves (BERTRAND-SARFAaa 1994). Some oncoids show directed growth caused by decreased water current (PI. 62/3). The oncoids have a wavy surface and show lighter brownish and darker greyish laminae and vertical holes which are
filled with detritus, fragments of Microcodium and ostracods. Small elongate structures are interpreted as incrustations around roots or little branches (BERTaAr~D-SARFAa~ 1994) (P1. 62/4). They are enriched by weathering and cover almost the whole top of the outcrop. Such an association of different stromatolitic structures is typical for shallow or abandoned channels of meandering rivers (NIcI~EL 1983). The second outcrop shows loaf-shaped stromatolites which form discrete vertical stacked hemispheroids (PI. 62/5). They occur in colour-mottled reddish marls which are interpreted as the top layer of an alluvial pond (ANADON & ZAraARRE~O 1981) and suggest a low terrigeneous sedi- ment supply.
The depositional environment of the Upper Paleocene (Fig. 17) was an alluvial flat with meandering rivers with concentrically stacked oncoids. Stromatolitic oncoids and algal incrustations developed at the margins of fluvial channels or in abandoned channels. There are paleosoils, ephemeral lakes and alluvial ponds with loaf-shaped stromatolites. Two transgressions are reflectedby hypersaline lakes, marine lagoons and tidal flats with evaporites.
4 CONCLUSIONS (1)The different growth types of stromatolitic structures
are very suitable for reconstructing the depositional history of terrigeneous deposits, especially fluvial and deltaic sediments.
(2) The form is evident for freshwater or marine influence. (3) It is possible to distinguish several deposition zones in
a fluvial system (channel base, abandoned channels, shallow margins, mouth of a river) by the form, size and degree of preservation of the stromatolites and/or oncoidso

2 5 0
- C A R B O N A T E MUD M O U N D S AND B I O G E N I C M O U N D S -
Part XVI PALEOZOIC
FINELY CRYSTALLINE CARBONATE MOUNDS: CRYPTIC COMMUNITIES, PETROGENESIS
AND ECOLOGICAL ZONATION
P.-A. Bourque
1 INTRODUCTION Mounds are structures whose accretion was controlled
by small, commonly delicate and/or solitary elements (biogenic mounds) or by inorganic accumulation of mud with variable amount of fossils (mud mounds), in contrast with reefs that are structures constructed by large, usually clonal elements, and capable of thriving in energetic envi- ronments (JAMES & BOURQUE 1992). This extended abstract of an oral presentation given at the IGCP-380 meeting held in GOttingen, October 1996, is concerned with geological structures that are mound-shaped bodies composed mainly of finely crystalline carbonates. These structures were primarily mounds, but are often collectively, and improp- erly, called mudmounds (or mud-mounds, or mud mounds) since the accretion of most of them was not controlled by accumulation of loose lime mud alone but by organisms, although they may be mud-rich. Despite they have been the subject of numerous studies, these mounds remain enigmatic. For those workers concerned with the under- standing of life evolution and biological community struc- ture in the past, their study is as much rewarding as is the study of reefs for instance, since they often comprise an important cryptic fauna and flora.
This note deals with the nature and composition of the primary organic communities of the Paleozoic finely crys- talline carbonate mounds, the petrogenesis of the mound facies, and their environmental significance. It insists on the importance to recognize the primary communities responsible for mound accretion prior to any attempt to propose community evolution or ecological zonation, and, although not minimalizing the overestimated, in my opin- ion, role of microbes, reasserts the multispectrum origin of these carbonate mounds. Selected examples from three basic types of Paleozoic mounds are used here: the Cambrian- Devonian 'rrcifs rouges'-type mounds; the Carboniferous Waulsortian-type mounds; the peculiar Devonian 'kess- kess' mounds of Morocco.
2 PETROGENESIS AND COMMUNITY SPECTRUM
Three time-related aspects of mounds should be con- sidered when dealing with petrogenesis: the nature of the primary organic community (if present) and its proportion with respect to loose lime mud (autochthonous and allochthonous); the early transformations (decay, lithi- fication) of the primary organic material, mainly through bacterial activity (biodiagenesis); the later diagenetic im- print that may have changed significantly petrographical
characteristics and some geochemical parameters (e.g., stable isotope signature).
At the outcrop level, most mounds exhibit a finely crystalline carbonate commonly with very few fossils, suggesting at first glance that the carbonate corresponds to primary loose lime mud. However, during the last decades, most studies have highlighted the necessary role of organ- isms in finely crystalline carbonate mound accretion or early diagenesis, in particular that of the microbes, to the point that several workers currently refer to mound facies collectively as microbialites (or microbolites), thus under- estimating and marginalizing the significant role of other organisms, such as the sponges.
2.1 Ro le o f Sponges Importantly, most Paleozoic mounds are not single
component as often presented in many discussions on 'mud mounds', but rather are made up of two or more facies. A first example is the so-called 'rrcifs rouges' of the Belgian Ardennes (which are not reefs and not entirely red) that constitute, to some extent, the type of the stromatactis-rich reddish mounds known from the Cambrian to the Devonian throughout the world. These mounds are composed of three distinct facies, each with a number of sub-facies: a red stromatactis-rich facies, a pink to pinkish grey coral- crinoid-bryozoan facies, and a grey coral-stromatoporoid- microbial facies (BotJRQtrE 1983; BOLrLVA~ 1990). Other examples are the Silurian mounds of the Qurbec Appala- chian s which are made up of the same red stromatactis-rich facies as in Belgium, and of a grey bryozoan-stromatoporoid- microbial facies (BOURQUE & GIGNAC 1983, 1986; BOURQUE & RAYMOND 1988), and the Devonian mounds of the Montagne Noire in France composed of the red stromatactis- rich facies, and of a pink fenestellid bryozoan-sponge facies (BotJRROtaLH & BOURQUE 1995).
For one, the red stromatactis-rich facies has shown to be very rich in sponge spicules, and, in some cases in sponge bodies, everywhere it has been studied (e.g., BOURQUE & GIONAC 1983; KERANS 1985; WALLAC~ 1987; BOtrLVArN 1990; JAMES & GRAWSTOCZ 1990; BOtrRROtaLH & BOURQUE 1995). This facies has been proposed to have originated from the transformation and cementation of a primary sponge network through microbial decay during very early diagenesis, destroying the original shape of the sponges and much of the spicules, and leaving a finely crystalline limestone often seen as a pelletoidal microspar under the light microscope, in contrast with uniform microspar thought to represent primary loose lime mud often forming geopetal infillings (BotJRQtrZ & BOLrLVA~N 1993). It ensues that the now observed finely crystalline limestone corresponds to both the early lithified sponge network and a certain amount of loose lime mud, an amount which obviously was lesser than suggested at the first glance by the bulk of the finely crystalline limestone. We have few data as to the proportion of mud versus the sponge network. In the Silurian mounds of Qurbec for instance, the pelletoidal sponge spicule-rich microspar/uniform microspar ratio has been evaluated to be 2:3 (BoURQUE & GIGNAC 1983).
The occurrence and significant role of sponges can also

251
MUD MOUNDS
/ / / ~ Small delicate Calcimicrobes f MiJd N skeletons o:nt
BIOGENIC,~/f MOUNDS/ SKELE'I'AL ~
Stromatolites Thrombolites Sponges Shells
Fig. 18. Facies spectrum of mounds. Distinction between biogenic mounds and mud mounds is based on accretionary controls (biogenic vs inorganic mud accumulation), not on relative per- centages of components. Slightly modified from JAMES & BOIJRQtJE (1992).
be asserted in at least some Waulsortian-type mounds. As here understood, Waulsortian-type mounds are those mounds which in the common understanding are Carboniferous and characterized by a mud-fenestrate bryozoan-crinoid association. They include the classical Waulsortian mounds (those of Tournaisian-early Vistan age; LEES 1988) which are characterized by a distinct skeletal grain association set (LEES 1988; LEES et al. 1985; LEES & MILLER 1985), and all those other Carboniferous mud-rich mounds that are slightly younger (= non-Waulsortian bioherms ofWE~B 1994). It is a widespread practice in the studies of the classical Waulsortian mounds to discriminate between mound facies according to grain-type associations using a ranking pro- cedure based on pointcounting or visual estimates of fossils in thin-sections (e.g., I-IENr~BERT & LEES 1991; LEES
~LLER 1985; AnR & STArcroN 1996; DEHANTSCHIJ I-I-ER & LEES 1996), and to assign them to the four Waulsortian Phases of LEES & MILLER (1985). In most studies, sponge spicules are found to be ubiquitous and often abundant in LEES & MrL~R'S (1985) phases B and C, although sponge bodies are commonly absent. However, the presently ob- served amount of sponge spicules more likely represents a small proportion of the initial abundance. Indeed, siliceous sponge spicules are very early dissolved: the microscleres are usually destroyed, and only a small proportion of the macroscleres is preserved (BOURQUE & GIGNAC 1983; MADi et al. 1996). Despite this, most workers deny a significant role of sponges in the mound accretion.
In Upper Vistan Waulsortian-type mounds of north- western Algeria, two of the mound facies contain both ubiquitous sponge spicules and sponge bodies (BOURQUE et al. 1995; MADI et al. 1996): a lower sponge boundstone and wackestone, and an overlying sponge/fenestellid bryozoan boundstone and wackestone. In both facies, sponges were concluded to have play a significant role in mound accre- tion. The texture of the spicule-rich microspar in the Algerian mounds is often pelletoidal, like that of the red stromatactis-rich facies of the 'rtcifs rouges', contrary to that of the classical Waulsortian mounds which is more commonly uniform. As in the Cambrian-Devonian 'r6cifs
rouges'-type mounds, the spicule-bearing pelletoidal microspar of the Carboniferous Algerian mounds is inter- preted as the end product of the biodiagenesis of a primary sponge network by microbial community. It may not be the case for the spicule-bearing uniform microspar of the classical Waulsortian mounds. An alternative hypothesis to the role of builders by sponges at mound surface is that the sponges were boring sponges rather. In this view, recent findings by I~EIWALD & WmSON (in press) of de- struction of the Lophelia frame of deep water mounds by boring sponges in the NE Atlantic, and massive mud production derived from this boring offers an attractive hypothesis to explain mud accumulation without apparent organic frame.
2.2 Role of Microbes The finely crystalline limestones are relatively easy to
relate to a primary microbial accretion processes when they have stromatolitic or thrombolitic fabrics. However this is not the usual case in mounds where more commonly a microbial origin is diagnosed based on the pelletoidal texture of the microspar (review and discussion by P1cv~Pa~ 1996). Microbes may play different roles in petrogenesis of mound facies, among which: trappers and binders of allochthonous mud; in situ loose mud producers; and micrite-microspar cement producers via vital activity of microbes (e.g., by photosynthesis), or via organic matri- ces. In the last case for instance, decaying sponges (BotmQtrE & BOtaWAtN 1993; RErrY~R et al. 1995; MAOI et al. 1996) or decaying former microbial community (TRICnEW & D~ARt"E this report; D~'FAROE et al. 1996) may be used as substrate for calcite nucleation. In both cases, the end product is a finely crystalline limestone with a pelletoidal micrite or microspar texture, a situation that renders difficult the deciphering of the primary community. When sponge bodies, or pieces of sponge bodies, or even only sponge spicules occur ubiquitously within thepelletoidal microspar, a sponge precursor may be concluded with a certain level of confidence, or at least strongly suspected. But when the pelletoidal microspar is devoid of these, although primary accretion by microbial community is often concluded, we are left with uncertainty.
No doubt that the microbial community can act as primary builders and play a significant role in mound accretion. For instance, extensive microbial encrustation observed as vaguely laminated to unlaminated pelletoidal microspar on skeletal material is a common facies in mounds. Examples are: the grey coral-stromatoporoid- microbial facies of the Devonian mounds of Belgium (BotrLVA[N 1990), the grey bryozoan-stromatoporoid-mi- crobial facies of the Silurian mounds ofQutbec (BotmQtrE et al. 1986; BOURQUE & RAYMOND 1988), and the coral- microbial framestone of Algerian Carboniferous mounds (MADI et al. 1996).
2.3 Biofacies Spectrum v e r s u s Microbialites The facies spectrum of mounds is illustrated by the
conceptual classification of reefs and mounds of JAMZS & BOURQUE (1992) (Fig. 18). A biogenic mound is one that

252
CARBONIFEROUS CAMBRIAN-DEVONIAN RF.CIFS ROUGES-TYPE MOUNDS WAULSORTIAN-TYPE MOUNDS
SILURIAN - QUEBEC DEVONIAN - FRANCE DEVONIAN - BELGIUM WAULSORTIAN-TYPE CLASSICAL DEPTH-RELATED Bourque & Gignac, Bourrouilh & Boulvain, 1990; - NW ALGERIA WAULSORTIANS COMMUNITIES 1983; Bourque & Bourque, 1995 Bourque & Boulvain, Bourque et al., 1995; Lees & Mi/ler, 1985; Raymond, 1989 1993 Madi et al., 1996 Lees, 1988
Fenestelnds Corals Rugose corals Phase D: DELICATE SKELETONS & ,,3 Stromatoporolds Stromatoporoids Microbial encrusters C + grain coating, MICROBIAL ENCRUSTERS i - Calcareous algae Microbial encrusters Calcareous algae Calcareous algae O Microbial encrusters 1-
,.1
Feoestellids Sponge (spicules) Crinoids + mud
Corals Crinoids ~'yozoBRs + mud
Crinoids Fenestellids Sponge (spicules) Red algae + mud
Sponges Fenestellids
Phase C: B + Plurilocular foraminifers
Phase B: A + Sponge spicules
Phase A: Fenestellids Crinoids
DELICATE SKELETONS & SPONGES § MUD
FENESTELLIDS & SPONGES
Sponges Sponges Sponges (red stromataCtis (red stromatactis (red stromatac~is Sponges SPONGES limestone) limestone) limestone)
Fig. 19. Facies spectrum of selected Paleozoic mound examples and bathymeu'ic frame.
accretion is controlled by organic community, regardless of the percentage of organisms. An individual biogenic mound may be composed of an array of biofacies, and may contain variable amount of loose mud. Obviously, to be meaningful this classification must use the primary com- ponents of a mound. Recently, the term microbialite has come widely used to name mound lithology. As originally defined, microbialite is 'organosedimentary deposits that have accreted as a result of a benthic microbial community trapping and binding detrital sediment and/or forming the locus of mineral precipitation' (Btrar~ & MOORE 1987). Although this definition may be very broadly interpreted, I think it would be wiser to restrict the term microbialite to facies primarily built by benthic microbial community, excluding those resulting from early lithification of a non- microbial community (like sponges) through microbial mediation, otherwise the term becomes meaningless.
2.4 Mud Mounds Mud mounds (sensu JAMES & BOURQUE 1992) are cer-
tainly not structures as common as the use of the term suggests. Some Paleozoic mounds however can be attrib- uted to true mud mounds, that is they were built by mud accretion without the help of organic communities. The peculiar Devonian conical kess-kess mounds of Morocco are good examples. Although they have been often assimi- lated to the 'rtcifs rouges'-type mounds (e.g. Ft~JS & HussNr~ 1993) and although they share with them several attributes like the finely crystalline nature of the lime- stone, the presence of stromatactis-like spar bodies, and the deep water setting, they fundamentally differ in the following: a) spectacular conical shape, with steep slopes (up to 60 ~ and a near circular outline in plan view; b) internal architecture with accretion surfaces parallel to mound surface; c) scarce delicate skeletal fauna, indicat-
ing no skeletal implication during mound accretion; d) no evidence of cryptic binders or builders such as microbes or sponges; e) embedding in finely crystalline limestone identical to the mound limestone; and f) mound location restricted to strata above a volcanic high (Hamar Laghdad massif) (Moth, J1 et ai. 1996), For one, the unique conical geometry of these mounds is difficult to conciliate with an organic control on their accretion. Integrating information from mound architecture, isotope signatures and overall geological setting, it has been concluded that the mounds were built by precipitation of finely crystalline carbonate mud from venting hydrothermal water (MouNJI et al. 1996). This water infiltrated into the volcanic and sedimentary rock pile was driven to sea floor by thermal flux above the Hamar Laghdad magmatic massif, forming volcano-like mud cones. The mounds were progressively cemented in the deep water marine environment during accretion, per- mitring encrustation by marine fauna.
3 ECOLOGICAL ZONATION There seems to have no significant changes in the commu- nity structure between the S ilurian-Devonian 'rtcifs rouges'- type and Waulsortian-type mounds during the Paleozoic time, a community structure which I believe was related to environmental factors, largely the water depth. Fig. 19 is an attempt to place the facies mosaic of selected Paleozoic examples in a bathymetric frame. Four bathymetric zones based on benthic communities emerge, from the deepest to the shallowest: 1) the sponge zone, typified by the red stromatactis lime-
stoneofCambrian to Devonian'rtcifs rouges'-typemounds, but also present in the Waulsortian-type mounds of Algeria; the sponge-dominated community is almost devoid of other fossils;
2) the fenestellid-sponge zone, best known from the

2 5 3
Waulsortian-type mounds of Algeria; 3) the delicate skeleton-sponge zone with significant amount
of loose lime mud; sponges are known mainly on more or less abundant scattered spicules;
4) the delicate skeleton-microbial encruster zone.
Phase A to D scheme of LEES & MmLm~ (1985) is incorporated here with much uncertainty, mainly because these phases are defined on grain-type associations based on a ranking procedure, not taking into account taphonomic processes that may have favoured one type of skeletons to the detriment of an other type (e.g., crinoid ossicles vs sponge spicules), nor texture of the micrite/microspar. For instance, more or less vague, clotted or peloidal fabrics in the primary muds in the lowest parts of the Waulsortian buildups in Waulsort are logged as matrix peloids (DEHANTSCmrrrER & LEES 1996). The texture of phase B limestones at Knockastia Quarry in Ireland is very similar to the spicular pelletoidal microspar of the fenestellid- sponge facies of the Waulsortian-type mounds of Algeria and the red stromatactis limestone of 'r6cifs rouges'-type mounds (my personal observations). Moreover, even if sponge spicules are not listed in phase A by LEES & MILLER (1985), it appears that they occur in at least some exam- pies. For instances, sponge spicules are listed in phases A and B of Waulsortian buildups atWaulsort (DEHArcrscuu-r~ ~ & LEES 1996); spicules are more common in the deeper mound growth phases in the Waulsortian mounds of the Sacramento Mountains in New Mexico (AnR & STArCrON 1994). It is therefore probable that phase A and B corre- spond to the fenestellid-sponge community, perhaps going up into the shallower delicate skeleton organism-sponge community.
4 CONCLUSIONS 1. Although single component mounds exist, Paleozoic
finely crystalline mounds are often composed of two or more facies. The mound classification of JAMES & BOURQUE (1992) illustrates the facies spectrum.
2. Microbes often use organic matter as matrix for carbon- ate precipitation. Deciphering between the original com- munity and the secondary microbial community that used the organic matter of the first is not easily achieved in ancient rocks, an achievement however necessary otherwise any conclusion on the community structure and ecological zonation of mounds is meaningless.
3. The mound community structure is concluded to have encountered no significant changes during the Paleozoic and their distribution to have been controlled by envi- ronmental factors, largely by water depth. Four bathymetric zones based on benthic communities are recognized.
4. It is much probable that microbial communites have played a role in most facies petrogenesis, but their role as primary builders is recognized only in the shallowest bathymetric zone.
5. To be meaningful, the term microbialite should be applied only to those facies that were primarily built by a benthic microbial community.
Part XVII SPONGES IN WAULSORTIAN-TYPE
MUDMOUNDS AT TRALEE BAY, CO. KERRY, SOUTHWEST IRELAND
S. Labiaux
Petrographic study of enigmatic organic structures resembling sponges collected from Waulsortian buildups (and not in the lateral equivalent or cover facies) on wave- polished surfaces on the coast of Co. Kerry, southwest Ireland, shows that they are indeed sponges or sponge- derived. This is the first record of sponge body fossils from undoubted Waulsortian facies where sponge remains in the form of scattered spicules or spicule bundles are known to be sometimes common. The sponges are varied in shape and size and also in their state of fragmentation and preservation. The present study reviews their macroscopic and microscopic features. Some of the primary and early- diagenetic textures that can readily be attributed to sponges are presented. The distribution of those fossils within the Waulsortian in the Co. Kerry outcrops is discussed as is the reason for their apparent concentration in this area as compared with Waulsortian buildups elsewhere.
1 INTRODUCTION Recently, there has been a growing interest in the role
of sponges in the genesis ofPaleozoic mudmounds (BoURQUE GIGNAC 1983; BOURQUE & BOULVAIN 1993; WARNKE
1995). This has received added stimulus from the analysis by REITr<ER (1993) and REITNER et al. (1995) of the role of sponges and their related microbial communities in car- bonate production within modem reef environments in Australia. However, none of the studies of the role of sponges in Paleozoic mudmounds has been concerned with the widespread Waulsortian facies of the Dinantian. Sponges certainly grew on many Waulsortian mounds and played a significant role in the early diagenesis of the sediments, but their remains have only been known in the form of spicules or spicule bundles (LEEs & ]VIILLER 1995: 206), never as body fossils. The discovery of large sponge- like fossils and their subsequent identification as true sponges or sponge-derived structures, was thus unex- pected. The sponges occur in well-exposed wave-polished outcrops of Waulsortian facies on the coast of Tralee Bay, Co. Kerry, south west Ireland (Fig. 20). They have not been observed in off-bank facies.
Micropaleontological dating by foraminifera and conodonts shows that the Waulsortian of this area extends to a higher stratigraphic level than most of the Waulsortian Complex in Ireland. A continuous outcrop shows that it can be dated from the late Tournaisian well into the Arundian part of the Lower Vis6an (Fig. 21).
Around Tralee, the Waulsortian facies occurs as a laterally continuous aggregate of overlapping and/or coa- lescent banks in the stratigraphically lower part of the sequence. At the top, growth was reduced to smaller scattered banks or bank aggregates (Fig. 21). The classic bryozoan-rich Waulsortian lithologies with a wide range of sparry fabrics (stromatactoid and others) occur in the

254
Fig. 20. Map of study area showing localities mentioned in the text
lower, continuous bank development. The scattered banks at the top of the sequence are of different character: they usually bear higher proportions of mud and sometimes exhibit abundant corals, suggesting that these accumula- tions are not of normal Waulsortian type. Sponges occur widely in normal Waulsortian lithologies, but rarer exam- ples have also been found in the atypical bank lithologies towards the top of the sequence.
2 OBSERVATIONS 2.1 Macroscopic features
The sponges (PI. 63/1,2) correspond to lithistid demosponges and hexactinellids. No thorough taxonomic investigation was possible because most of the spicule frameworks, on which classification is based, are too badly preserved.
All the large specimens (15 to 30 cm in length) are lithistids and usually marrow-shaped with a radial struc-
ture (channel system) and an obvious central cavity. Smaller lithistids, probably in an earlier state of maturity, have the same characteristics and occur in circular sections. Lithistids seem to be the dominant type in the rocks observed. Their distinct large central cavity is either filled with blocky spar or geopetal muds.
Hexactineltids are much rarer. Although they do not show a large central cavity, they have an internal or- ganization that reflects the initial organization of spicule networks (Pi. 63/3). Their size does not exceed 5 cm in diameter. This statement takes no account of Hyalostelia, whose shape and size are unknown (HINDE 1886), but which has been here identified by its large monoaxone root-tuft spicule bundles recovered in the sediment.
Large sponges tend to occur as isolated individuals in the buildup material whereas small ones occur together in clusters. All types are associated with fenestrate bryozoan fronds, and usually occur in spar-rich layers (PI. 63/2).
P l a t e
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
63 Sponges in Waulsortian-Type Mudmounds, Southwest Ireland
Flat-lying, marrow-shaped lithistid sponge at outcrop exhibiting radial structure and large spar-filled central cavity with thin geopetal sediment. Waulsortian Complex, Maharees, Co. Kerry Lithistid sponge (arrow) in spar-rich band of Waulsortian lithology with its central cavity incompletely filled with bioclastic mud. Waulsortian Complex, Barrow Harbour, Co. Kerry Negative print of thin-section showing a hexactinellid sponge with its spicular network. Moulds of megascleres (arrow) are obvious and have locally been enlarged to form stromatactoid cavities. Width of Fig.=5 cm. Upper bank, Maharees, Co. Kerry Degraded sponge material (arrows showing two individuals) in a Waulsortian facies rich in stromatactoid sparry masses. Camp, Co. Kerry Polished slab of Waulsortian bank facies. Large, arcuate and laterally extended cavities parallel to bedding are interpreted as occupying the positions of dissolved or collapsed sponge-walls (arrows). Picture height is 6 cm. Maharees, Co. Kerry Peloidal microspar. Scale bar=500 ~tm. Maharees, Co. Kerry (same sample as Fig.5.)

P l a t e 63 2 5 5
256
Fig. 21. Stratigraphy and regional pattern of the Waulsortian Complex in the Tralee Bay area.
Most of them are lying parallel to bedding suggesting that they simply toppled over. There is only one record of a sponge found upright in life position. Lobate sparry masses within spar-rich and bryozoan-rich bands are interpreted as representing fragments of sponges in a more degraded state (P1.63/4) because they systematically appear in the neighbourhood of, and have the same radius as well- preserved sponges. Altogether, 10 large specimens similar to those of PI. 63/1,2 were found. Smaller individuals in clusters have not been counted.
Because the state of preservation varies so widely, it is difficult to determine how much the sponges originally contributed to the total rock volume. However, they were probably important in the bank genesis, at least locally where they may well have been the main component of the epibenthic community living on the bank.
2.2 Microscopic features directly attributed to sponge relics
Although sponges are quite common in the field, they are difficult to sample, especially when the relationship with the surrounding sediment has to be investigated. The best example obtained shows two sponge bodies on a polished slab (PI. 63/5) which was subsequently cut into two large thin-sections (7xl lcm) so that the fabrics could be examined in detail. Several microscopic fabrics have been identified as being associated with sponge relics:
-Peloidal microspar consists of irregular peloids (50 to i00 microns in diameter) more or less scattered in a microsparitic matrix. This fabric, devoid of bioclasts, never occurs in a geopetal situation, seems to have formed firm masses (PI. 63/6), and is here attributed to in situ calcification of sponge tissues. Such fabrics would have been included in the category of'matrix peloid' in previous Waulsortian studies (e.g. LE~s & MILLER 1995: 208-9). -Geopetal bioclastic muds occur sometimes in large quantities within the central cavity of the sponge body.
These muds must have originated outside the sponges and form part of the sequence of polymuds sensu LEES & M[LtZR (1995: 209-210). Their accumulation in this situ- ation indicates that the sponges must have been suffi- ciently rigid to support open cavities. -Stromatactoid cavities are formed by the collapse of incompletely lithified sponges. Commonly, sponge frag- ments lay randomly on top of each other to form mechani- cal 'bridges' producing shelter cavities beneath (cf. the mechanism suggested by LEEs (1964: 521-522) to explain stromatactoid cavities in the Waulsortian of Ireland). Geopetal sediments within the cavities helped to produce the stromatac" id shapes. Other workers (WARNK~ 1995; BOURQUE & BOULVAIN 1993) have invoked this mechanism for the formation of stromatactis. Large, arcuate and later- ally extended cavities parallel to bedding are also inter- preted as occupying the positions of dissolved or collapsed sponge-walls (PI. 63/5). However, that is not meant to imply that all stromatactoid cavities in the Waulsortian are sponge-related.
Spicules, which would be expected to be the main remnants of sponges, are rarely recovered from the sponge bodies in the material under study. Only in the case of hexactinellids, which possess rigid spicule networks, can the relationship between the fossilized object and the original skeleton be established (PI. 63/3). In most in- stances in this study, 'objects' that can be readily identified as the remains of sponge walls are systematically devoid of spicules even though spicules are found dispersed within the nearby muds, thus suggesting that they are not hexactinellids. These latter spicule-rich muds are thought to be identical to the Mla primary muds of LEEs & Mmt.ER (1995: 209).
It is interesting to observe that a single sponge body represents, in itself, a mini-Waulsortian system where all the components of the Waulsortian are present: multi- component muds, stromatactoid sparry fabrics and bioclasts. This mini-system may have been reproduced at various

257
scales from millimetric to decimetric - a factor that must be taken into account when quantifying the sponges in terms of rock volume.
2.3 Cementation Cathodoluminesence of the calcite cements fringing
sponge-walls has revealed the existence of a non-fibrous, "black" (i.e.non-luminescent) cementation phase predat- ing the crypto-fibrous cement and other, blocky cements. This is most unusual as the fibrous cement is generally the earliest one. As this 'black' cement phase seems to be restricted to the immediate neighbourhood of sponges in the material studied, it is therefore provisionally attributed to the presence of the sponges and their associated micro- bial communities which might have had a strong influence on the chemistry of the surrounding waters. This assess- ment needs to be confirmed by additional C.L. studies of more samples.
3 DISTRIBUTION OF SPONGES IN THE WAULSORTIAN
3.1 In the study area In the studied area of Co. Kerry, the sponge body
fossils do not seem to be restricted to any particular stratigraphic interval. However, here within the Waulsortian facies only lithistids are present and they only occur in the banded, fenestrate bryozoan-rich facies. In terms of LEES & M~Lt~RS'S Waulsortian Phase model (1985, 1995), they extend from phase B (subphotic) to phase D (shallow photic zone indicated by the presence of dasyclad algae). The large spicules of Hyalostelia are ubiquitous.
Sponge body fossils are usually absent from the smaller lens-shaped muddy banks in the beds overlying the con- tinuous Waulsortian Complex, although those facies con- tain numerous spicules, often of hexactinellid type (among them the large spicules of Hyalostelia). The rare examples of undoubted hexactinellid body fossils (PI. 63/3) have been recovered from these upper levels.
There is thus an apparent zonation in the distribution of the main types of sponge body-fossils: lithistids are domi- nant in the lower, continuous part of the sequence of typical Waulsortian lithologies, whereas only a few hexactinellids dominate at the top in bank lithologies devoid of lithistids. This zonation may be related to selec- tive preservation rather than original distribution: lithistids may be prone to preservation in common Waulsortian lithologies but not in the lithologies of the upper banks. Preservation potential could well be a key factor control- ling the apparent abundance of sponges in Waulsortian rocks in general.
3.2 Elsewhere in Ireland Sponge body fossils have never been recorded from the
many exposures of the Waulsortian Complex elsewhere in Ireland and none have yet been found in the few, large samples examined in the present study. Petrographic fab- rics similar to those described above have been found in samples from Mullawornia and Feltrim (see appendix for locality details) but no clear relationship between these
fabrics and sponge body-fossils can be established. For reasons not yet understood, preservation processes did not lead to the fossilization of entire organisms.
4 DISCUSSION On present knowledge, the sponge body fossils found
in Co. Kerry thus represent a unique occurrence. The characters of the rocks themselves offer no explanation for this - they seem to be normal Waulsortian of Phases B - D. Presence of patches of characteristic fabrics in Waulsortian limestones from elsewhere in Ireland suggests that sponges similar to those found in Co. Kerry were originally present (note that the presence of sponges was not in doubt as the rocks often contain spicules of hyalosteliid and probably other siliceous sponges).
The apparent restriction of the sponge body-fossils to the Waulsortian of Co. Kerry may be due less to unusual conditions of preservation there than to inadequate expo- sure quality elsewhere. The presence of wave-polished coastal exposures in Kerry is unusually favourable for observation. The absence of observations of sponge body- fossils from quarries in the same Waulsortian limestones in inland quarries nearby corroborates this argument.
The fact that such fossils have not been recorded during the detailed petrographic study of Waulsortian rocks which have been undertaken in Europe and North America dur- ing the last 20 years is not surprising - standard-size thin- sections are inadequate tools for detecting recognizable body-fossils of this type.
Appendices Feltrim: quarry, 2.5 km SE of Swords, Co. Dublin. INGR: O 193 447 (PHILCOX 1963). Mullawornia: disused quarry, 3 km NW of Ballymahon, Co. Longford; 16.5 km S of Longford. INGR N 130590
Part XVIII ENVIRONMENTAL VERSUS ORGANIC CON- TROLS ON BIOGENIC MOUNDS: EXAMPLES FROM THE UPPER TRIASSIC OF NORTHERN
AND CENTRAL APENNINES (ITALY)
S. Cirilli
1 INTRODUCTION Different types of biogenic mounds have been investi-
gated, in order to define general criteria to decribe the main characterising textures and their environmental signifi- cance. The biogenic mounds come from two different stratigraphic and palaeogeographic settings of the Upper Triassic Northern and Central Apennines: the Monte Cetona Formation (Rhaetian, Northern Appennines) and the Norian- Rhaetian succession of the Monti Simbruini area.
2 GENERAL SETTINGS 2.1 Northern Apennines (southern Tuscany-western
Umbria) The Monte Cetona Formation (about 150 m thick)
consists of a lower carbonate-argillaceous member, made

258
of thin to medium bedded limestones (pack-/wackestone) intercalated with thinly bedded marls and clays; and an upper carbonate member made of thinly bedded dark limestones (pack-/wackestone), interbedded with carbon- ate packstones, seldom grainstones (CtAaAPmA et al. 1987). The depositional environment was referred to a mixed carbonate-siliciclastic homoclinal ramp characterised by dysoxic-anoxic conditions (S~FAr~ & Tao~ea3m-rA 1989; Cmmu et al. 1994). The terrigenous input was more abun- dant during the deposition of the lower member.
Biogenic mounds are present throughout the forma- tion, but with different morphologies and textures. Mounds from the lower member are intercalated with detritic- bioclastic wacke-/packstones, frequently burrowed. They show a flat lenticular shape and their thickness ranges from about 50 cm to 2 m. The main texture is represented by microbial bindstone. Biogenic mounds from the upper member display a lenticular shape and they are about 10- 30 m in length and 5-15 m in height. They are mostly composed of serpulid framestone and microbial bindstone. The mounds are situated upon wacke-/packstone contain- ing oncolites, foraminifers and calcareous algae. The lat- eral sediments are mainly bioclastic packstones which locally onlapped the mounds. On a larger scale they are lateral to thinly bedded limestones, which are definitely younger than the mound complex. Immediately above the mounds the sediments are dark mudstone intercalated with wackestones.
2.2 Central Apennines (Monti Simbruini) The Norian-Rhaetian succession consists of about 500 m
of carbonate (limestones and dolostones) whose lithofacies were referred to shallow water carbonate platform depos- its lateral to anoxic sediments deposited within an intraplatform basin (Ctan~ 1992,1993). The studied biogenic mounds come from the carbonate platform complex. The interior platform is represented by a sequence of shallowing upward cycles. The marginal facies consists of a thick succession of massive dolostones in the lower part which grades upward into cyclothemic dolostones which repre- sent the back margin area. Biogenic mounds are present in
the lower massive portion and in the subtidal horizons of the cyclothemic dolostones. In the first case they show lenticular shape and are about 30 m thick. They are mostly composed of serpulid framestones. They occur beneath and upon detrital-bioclastic pack- and grainstones and are associated with float- and rudstones. In the second case biogenic mounds have a flat lenticular morphology and are less than 1 m thick. They occur upon and lateral to bioclastic pack-/grainstones in the back margin and wackestones in the platform interior.
3 MACRO- AND MICROFABRICS OF BIOGENIC MOUNDS
Two main types of biogenic mounds have been distin- guished (Cmmu 1996), i) microbial mounds, including stromatolites, thrombolites, and microbial crusts, and ii) serpulid mounds, present with two main different tex- tures: serpulids with micrite and serpulids with cement.
Stromatolites occur as planar or domal stromatolites (LLH-type stromatolite) of generally low relief rarely exceeding 20-30 cm in height. The undulated laminar stromatolites are sometimes formed by Girvanella-like microtubes in fenestral pelmicrite and bioclastic wacke-/ packstone (PI. 64/1). Bioclasts and other detrital sediment was trapped within the mats and usually concentrated to form individual laminae. Fenestral and other micro- and macrocavities are bordered by several generations of fi- brous isopachous cement, marine in origin. Blocky ce- ments or internal silty sediment occur in the innermost part of cavities.
Thrombolites consist of millimeter and centimeter- sized dark micritic clots, surrounded by sandy sediment or sparry cement (P1.64/2). Terrigenous silt, peloids, ooids, and bioclasts are scattered within and between mesoclots. The interstitial sediment is frequently burrowed and/or bored.
Microbial crusts are formed by alternations of thin layers of brownish fringe cements with dark and dense micritic microbial coatings. Their origin is linked to bio- chemical processes activated by microbial communities
P l a t e 64
~g.1.
Fig.2.
Fig.3.
Fig.4.
Fig.5.
Upper Triassic Biogenic Mounds of Northern and Central Apennines (Italy)
Stromatolite type: Girvanella-like microtubes in fenestral pelmicrite. Fenestral fabric is commonly enlarged by dissolution processes. Micro- and macrocavities are bordered by several generations of fibrous isopachous cement. Blocky cements in the innermost part of cavities, x 35 Thrombolite type: centimeter-sized dark micritic clots surrounded by sandy sediment or sparry cement.
Note trapping of bioclasts, peloids and other grains, x 15 Serpulids with cements: serpulid framestone; the framework porosity is infilled by several generations of fibrous isopachous cement. The final voids are closed by equigranular, blocky cement. (Negative print, x 5) Isolated microbial crusts: alternations of thin layers of brownish fringe cements with layers of dark and dense microbial micrite. (Negative print), x 8 Serpulids with micrite: small cluster of serpulid tubes whithin a micritic matrix. Aggregates are surrounded
by microbial crusts, final voids are in filled by blocky cements, x 6
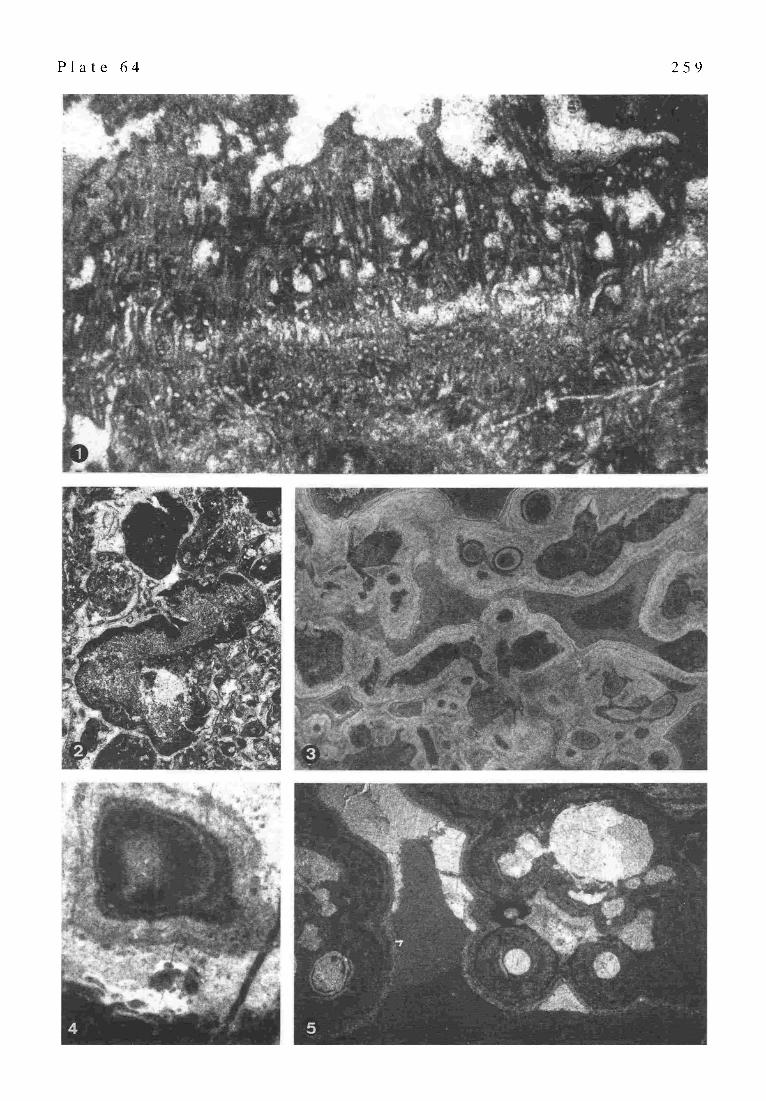
P l a t e 6 4 2 5 9

260
microbial.
Tr
St
environmental parameters
water energy
surface
patchy colonization by microbs
sediment supply
episodic
wide and colonizable regular by mlcrobs
high
turbulence
low to moderate
encrusters & bioeroders
present
absent
biologic parameters
burrowers
present
absent
other organisms
present
algae enhancing trapping ability
mma~s
aboundant and diverse metazoans
metazoans devoid
IA
increase
light ~ deere~
shallowwater high energy, patchy colonization episodic sediment supply presence of encrusters, bioeroders, burrowers
pollution, suspended sediments independently from depth
deep water low sedimentation rate high organic matter concentration
thrombolites
thrombolites
stromatolites
Fig. 22. A: the main environmental and biogenic parameters which control the growth of microbialites; Tr: thrombolites; Sr: stromatolites. B: the control of l.;ght variations in respect to water depth and other parameters on the development of stromatolites or thrombolites.
(Kau~,mEIN 1976; MONTe 1995). The crusts may occur isolated (PI. 64/4) but more frequently as coatings of serpulid boundstones and/or detrital sediments (PI. 64/5).
Serpulids with micrite correspond to serpulid boundstones in which the aggregates are mostly disoriented or occur as small clusters with a maximum diameter of about 3 cm embedded within a micritic matrix. This matrix contains small peloids, encrusting foraminifers, and small ostracods. This type of mound is frequently characterised by the presence of thick microbial crusts coating single serpulid tubes and entire clusters (PI. 64/5).
Serpulids with cements correspond to serpulid framestone with aggregates sometimes encrusting large bioclasts (e.g. gastropods, brachiopods) or forming a continuous frame- work (PI. 64/3). The serpulids may be the only organisms or are associated with corals, calcareous sponges, soleno- poraceae, Girvanella sp., Thaumatoporellaparvovesiculifera (RA~Rz), and Muranella sp.. Serpulid tubes and, occa- sionally the associated fauna, are encrusted by fibrous
cement. The remaining voids are filled with sandy carbon- ate sediment composed ofpeloids and bioclasts, mainly of foraminifers and ostracod shells. The final voids are closed by equigranular, mosaic cement.
4 ENVIRONMENTAL AND ORGANIC CONTROLS ON MACRO- AND MICROFABRICS
Microbial mounds including thrombolites are common in the deeper portion of the homoclinal carbonate-siliciclastic ramp which is characterised by relatively high sedimenta- tion rates associated with abundant terrigenous influx. Light was drastically reduced by particles in suspension. They are also frequent in the back margin area, within shallow waters affected by high water energy and high illumination. Microbial mounds with stromatolites are common in the tidal flat of the inner carbonate platform and in the more protected areas of the back margin. Serpulid mounds with cement are common at the margin area of the carbonate platform. Serpulid mounds with

261
micrite, frequently associated with microbial crusts, grew in the upper portion of the carbonate ramp under stressed conditions: reduced light, carbonate mud and fine terrigenous particles in suspension, anomalous salinity, and dysoxic to anoxic conditions.
The main factor discriminating serpulid and microbial mounds is the nature of substrate: hard substrate favoured the settlement of serpulid colonies, while the development of microbial mounds is independent from this. It is strongly controlled by other factors such as sediment supply, water energy, light and depth (Fig. 22A).
The growth of stromatolites seems to be favoured by wide surfaces under regular sediment supply and a rela- tively low to moderate water energy in absence of water turbulence, skeletal encrusters, burrowers, micro- and macroborers. It results in well-developed and continuous lamination. For stromatolites growing in shallow water, the presence of algae living within the photic zone, may enhance the trapping ability of microbial communities taking part in the accretion of microbialites.
Thrombolites can originate in shallow water environ- ments where water turbulence caused an irregular distribu- tion of poorly sorted detrital-bioclastic sediment. They can develop also in deeper environments, where the episodic terrigenous influx results in a discontinuous sediment supply (i.e. the mixed terrigenous-carbonate deep ramp where the lower member of the Monte Cetona Formation was deposited). However, in each case thrombolite-form- ing microbial communities are commonly associated with abundant and diverse metazoans, including encrusters and bioeroders. Under their presence it is also possible that syndepositional disturbance such as physical damage, bioturbation and local bioerosion could convert stromatolitic to thrombolitic fabrics (BRAGA et al. 1995).
Variation of illumination can play a different role in the accretion pattern of microbialites (Fig. 22B). In shallow water environments, the light increase and the high water energy, together with patchy colonization of the surface by micro- and macroorganisms result in thrombolitic or ir- regular stromatolitic macrofabrics. The presence of sus- pended particles within the water column (light decrease) provides nutrients for the epi- and infauna resulting in a wide-spread bioturbation. Under such conditions both irregular sediment supply and syndepositional disturbance can be the main causes for the formation of thrombolites.
A light decrease towards deeper quiet water associated with low sedimentation rates (i.e. during transgressive phases) can favour the development of stromatolites. The low sedimentation rate may result in an increase of the organic matter concentration which provides a sufficient nutrient source for the growth of microbial communities (ScHtr~TZE-LAM et al. 1993). A regular microbial growth and supply of well-sorted, fine grained sediment in deep water resulted in well-developed stromatolitic layering. An example of this has been reported from the Lower Jurassic of Umbria-Marche Appennines, Central Italy (BA~a'ou~ & Cmmu 1994). Microbialites formed by thinly laminated crusts mainly of LLH-stromatolite type, have been found within pelagic sediments from condensed
sequences referred to the Late Carixian (Pliensbachian).
5 SERPULIDS AND MICROBIALCOMMUNITIES:
IS IT A SYMBIONTIC RELATIONSHIP? The presence of serpulid mounds depends on the origi-
nal substrate, because the serpulid larvae require a hard substrate for settlement. The shape of aggregations is the result of upward and outward growth to reach suitable suspension feeding sites and to prevent the burial by sediment. Light and nutrients are important factors for the settlement of serpulids and the growth of these buildups. The more important factors leading to the death of modern serpulid colonies are represented by physical and chemical factors. Large amount of suspended mud, low oxygen levels, presence of hydrogen sulphide represent lethal conditions for serpulid reefs (BosENCE 1979). But these data do not match with several records in ancient environ- ments of serpulid mounds in both shallow and deep water, characterised by high rates of muddy sedimentation and dysoxic to anoxic conditions. In the Monte Cetona Fro, serpulid boundstones in micritic matrix occur in the upper portion of a carbonate ramp, in a dysoxic to anoxic envi- ronment, characterised by high rates of muddy sedimenta- tion as suggested by the lateral dark mudstones and by the type of the organic content (CmILU et al. 1994). This type of mound is usually associated with thick microbial crusts suggesting a cyanobacterial symbiosis as a main process to assure survival to the serpulid colonies.
In modern environments symbionts include a variety of very different organisms including cyanobacteria (TgENCH 1987; COWEN 1988). In some modern sponge reefs the oxygen produced by cyanobacterial symbiontic relation- ships exceeds almost by the factor of 1.5-2 the required oxygen for respiration (WILKINSON 1987). The excess oxy- gen is dissolved in the water column and avaible to be used by other organisms. The same process could occur within the serpulid-microbial ecosystem, where oxygen surplus added to the water column improved the local conditions of oxygenation.
The symbiontic relationships between serpulids and microbial communities could play an important role in the early processes of cementation. Usually the amount of organic carbon fixed by symbionts (zooxanthellae) in the coral ecosystems exceeds the requirement rate for their physiologic processes (Musct, a'INE et al. 1981). SPENCER DAV1ES (1984) estimated that in a zooxanthellate coral, about 51% of the energy fixed by the zooxanthellae is used in respiraton; 48% is lost from the system and only 1% is used for growth. KINSEY (1985) suggested that almost 10- 30% of the organic carbon produced is used in the forma- tion of skeletal carbonate by the corals. In some modern reefs of the Red Sea more than 95% of the carbon fixed by the symbionts of the coral Stylofora is released to the host for respiration, reproduction and growth (MuSCATtNE et al. 1985). The surplus is released as mucus and dissolved organic matter. But the coral reefs are ecosystems with high primary productivity rates relative to the serpulid reefs. This means that there is a great amount of carbon

262
flux escaping from the serpulid ecosystems: the budget of carbon fixed by the symbionts should be lower, while the surplus of carbon higher and the mucus produced more abundant. The availability of the carbon surplus could be involved in the early cementation processes. As empha- sized by several authors (KRuM]]EIN 1976; SCrm'mDERMA~rN & HARRIS 1985; Btmr,rE & MOORE 1987; Rmn~G 1991; REtTr~R 1993) these processes are enhanced by the pres- ence of microbial communities that actively participate in the processes of carbonate precipitation. Evasion of CO 2,
�9 mainly close to the metabolism of the microbial communi- ties, speeds up the nucleation of the calcite crystals, and therefore early cementation of the microbial crusts and associated sediments.
Part X1X MUD MOUND TYPE COMMUNITY
REPLACEMENT ON CARBONATE PLATFORMS AND CORALGAL
REEFS
F. Neuweiler
1 INTRODUCTION The primary aim of studies concerned with mud mounds
is the documentation of the compositional and petro- graphic inventar of these prominent structures (palaeon- tology, sedimentology, diagenesis, carbonate mud budget). Despite detailed descriptions of mound composition little is known about the compositional variation along lateral stratigraphic equivalents, especially at the level of the initial mound growth. Beyond of that, generally little attention is given to occurrences of analogous facies (i.e. to those found within mud mounds) but lacking any mound morphology. This is a short note which provides examples for both observations lateral to the initial growth of a mud mound and morphologically indistinct occurrences of'mud mound facies'. Examples are from the Lower Cretaceous (Albian) of the Soba Reef area, N Spain (Fig. 23). Discus- sion will focus on the controls of community replace-
merits, associated changes in the trophic structure and resulting facies geometries.
The Soba Reef area (Fig. 23) is part of the western margin of the Vasco-Cantabrian Basin. From W to E and from N to S the Aptian/Albian carbonate and mixed facies platform deposits ('Urgonian', mainly rudist-bearing lime- stones) grade into basin deposits bearing marls, marly limestones, silici- and calciturbidites. Along the slope large-scale mud mounds occur, exhibiting a highly vari- able, depth related megabenthos community and 50-80 vol.-% of autochthonously formed micrite (NEuwEIt.eR 1995). This automicrite may form thrombolitic and stromatolitic structures and therefore was attributed to different types of microbialites by NEUWmt~R (1993). On the base of an integrated data set (comparative petrogra- phy, stable isotopes, intracrystalline organic compounds, and biomarker) an organic i.e. non-living control was suggested for these types of automicrites (organic matrix mediated organomicrites sensu RmT~rER et al. 1995; NEUWEZU~R 1995; NEUWEU.ER et al. 1996). For details on the principles of organomineralization see MITTERER & CUr,rNtNGHAM (1985); RErr~R et al. (1995), and TR~CHET & D~FARGE (1995).
2 COMMUNITY REPLACEMENT LATERAL TO INITIAL MOUND GROWTH The section illustrated in Fig. 24 is part of the Top Val
de Ason/Gandara section (Upper Aptian to Middle Albian) which comprises a 260 m thick succession of mostly lagoonary deposits charcterized by thickly bedded rudist- bearing limestones (mostly requienid rudists like Toucasia and Pseudotoucasia) and episodic intercalations of deltaic sandstones (N~uwEILER 1995: 35-58). According to data of PASCAL (1985) and NEtrWmLER (1995) this interval (Fig. 24) represents the Lower/Middle Albian boundary inter- val. For the purpose of detailed description, the section is subdivided into a lower carbonate unit, a siliciclastic unit, and an upper carbonate unit. Individual subunits are con- secutively numbered (subunits A-H). Stratigraphic corre- lation with the development of the mud mound 'La Gandara'
Santander ~ N
r-h~-~J ~Ramalesl ~ ASTRANA ao(~ Santander I ~ _J I ~(~ ~"
I L , ~ " I c ~,t CANEDO '
Fig. 23. Locality map with names mentioned in the text. l=replacement interval lat- eral to the mud mound 'La Gandara', 2=replacement interval upon coralgal reef close to the village Aja.

2 6 3
Fig. 24. Log of replacement interval lateral to the mud mound 'La Gandara'. l=requienid rudists, 2=chondrodont bivalves, 3=miliolid forams, 4=ataxophragmid forams, 5=orbitolinid forams, 6=Lithocodium, 7=Bacinella, 8=Scleraetinia, 9=Pienina oblonga (octocoral sclerites), lO=Euzadiella, 1 l=Murania, 12=cryptic Acanthochaetetes, 13=Porostromata, 14=encrusting red algae. 15=dendroid red algae, 16=Marinella, 17=dasycladaeean algae, 18=codiacean algae, 19=organomicrites, 20=lithistid sponges, 21=encrusting forams, 22=polychaetes, 23=thecidean brachiopods, 24=other brachiopods, 25=encrusfing bryozoans, 26=pharetronid sponges, 27=record of microborings
is based on mapping (Pl. 65/1) and sedimentological cri te- ria (supply of mud mound intraclasts).
2.1 Description of compositional variation The lower carbonate unit (with subunits A-C, Fig. 24)
comprises 'typical Urgonian' rudistidlimestones and mixed facies bioclastic rocks rich in orbitolinids and oncoids. The rudist-bearing limestones (subunit A) exhibit an
autochthonous association of requienid rudists (Toucasia, Pseudotoucasia) together with Lithocodium, Bacinella, miliolid, ataxophragmid and orbitolinid foraminifera, the coralline sponge Murania and various Porostromata (e.g. Cayeuxia). The density of microborings within rudists is very high resulting in a quasi autochthonous packstone texture. The depositional environment is that of a re- stricted lagoon.

2 6 4
These rudist-bearing limestones are succeeded by mixed siliciclasric/carbonate deposits (subunits B, C) which in- tercede between the lower carbonate unit and the siliciclastic unit above. Despite minor variation relative to subunit A (oncoids, calcareous algae, Fig. 24) these deposits still represent a common facies of Urgonian platforms, spa- tially related to siliciclastic input of adjacent deltaic sys- tems. There is an erosional boundary between subunits B and C.
Siliciclastic unit This is a ca. 12 m thick succession of fine- to medium
grained quartz sandstones with minor amounts of clay and bioclasts (exclusively orbitolinid foraminifera). Sedimen- tary structures comprise asymmetrical ripples, isolated ripples, trough cross-bedding, and planar cross-bedding. There are minor indications ofbioturbation. The depesitional environment is that of a very shallow subtidal/intertidal shelf (in part mixed- and sandflat). The siliciclastic unit is part of a widespread shallowing-upward cycle covering former carbonate platform areas and most parts of the adjacent basin (Nztm'mLER 1995).
Upper carbonate unit In relation to the development of the mud mound 'La
Gandara' the upper carbonate unit can be subdivided into
a pre-mound interval (subunit E), an initial mound interval (subunit F), and a syn-mound interval with subunits G and H.
By composition and texture the pre-mound interval is similar to the mixed facies deposits of subunit C, variation concerns a compositional trend towards a detrital coral facies with various amounts of lagoonary elements (e.g. the coralline sponge Euzkadiella). Towards the top of subunit E the detrital coral facies converts to an autochthonous assemblage of platy, mainly microsolenid corals and litho- phagous bivalves.
The initial mound interval (subunit F) is characterized by a repetitive replacement ofa scleractinian coral/calcar- eous algae community by a composite organomicritic facies (F1, F2; PI. 65/2-4) and a successive recovery of coralgal facies towards the top (F3; PI. 65/5). The coralgal community forms a biostrome and is composed of scler- actinian corals such as Placophyllia, Calamophylliopsis, an undetermined species with meandro-flabellate growth form, encrusting red algae (mostly the squamariacean Polystrata alba (= Ethelia, = Pseudolithothamnium), less frequent the corallinacean Lithophyllum), and encrusting foraminifera such as Placopsilina, Tolypammina, and Coscinophragma.
At the level of initial mound growth (F1) the outer surfaces of the coralgal community show thin coatings of
P l a t e 65
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig. 8.
Fig. 9.
Fig. 10.
Fig. 11.
Mud mound type community replacement on carbonate platforms and coralgai reefs (Lower Creta- ceous, Lower Albian, N Spain)
The mud mound 'la Gandara' is situated at the upper platform margin seperating basin marls (B) and lagoonary sequences. The correlative interval of initial mound growth (arrow) exhibits repetetive replacements of the benthic community. Replacement interval (interval F in previous text) from coralgal to organomicritic coatings, l=Polystrata alba, 2=Acanthochaetetes in cryptic position. Center=terebratulid brachiopod. Hight of Fig.=21 mm Replacement interval: meandroid and dendroid corals (Calamophylliopsis) overgrown by organomicritic coatings and thrombolites. Note associated polychaetes worm tube and encrusting foraminifera (arrows). Hight of Fig.=21 mm Organomicritic facies (coatings, thrombolites, lithistid sponge) in part covering debris of corals, codiacean algae and gastropods. Hight of Fig.=21 mm Partial recovery of coralgal community (interval F3 in previous text). Encrustations by red algae dominate again. Width of Fig.=21 mm Post-mound interval (interval G in previous text): autochthonous coralline sponge Euzkadiella (1) with Acanthochaetetes in cryptic position. Note frequent fragments of ancestral dendroid red algae (2, arrows). Hight of Fig.=21 mm Top of coralagal reef body in the vicinity of the village Aja. Arrows point to the retrogradational shift of facies along the transitional zone from marly basin rhythmites to massiv organomicritic/siliceous sponge limestones. Detrital reef core sediments with scleractinian corals, encrusting red algae and biodetritus of rudists and chondrodont bivalves. Width of Fig.=13 mm Establishment of an organomicritic facies (arrow) within pore space of the growth framework. Width of Fig.=13 mm Replacement interval: association of organomicrites, lithistid, hexactinellid sponges and terebellid
polychaetes. Note Acanthochaetetes (arrow) in cryptic space. Width of. Fig.=22 mm Replacement interval: stratiform occurrence of 'mud mound facies' with fine-grained internal sediments and stromatactoid cavities. Width of Fig.=22 mm

P l a t e 65 2 6 5

266
organomicrites. Some larger fragments of scleractinian corals already exhibit a constructive outer rim of irregular organomicrites. Internally, within cryptic spaces there is a succession from crusts of Murania to thrombolitic organomicrites associated with encrusting foraminifera, siliceous sponges and calacareous polychaete tubes. Occassionaly Acanthochaetetes seunesi is found but al- ways as small cm-sized specimens in cryptic position (PI. 65/2). Minor elements are encrusting bryozoans and pharetronid sponges. Despite the initial phase with Murania, the benthic community related to organomicrites (thrombolites) is one of those found within mud mounds. Finally the biostrome was buried by a marly, bioclastic packstone.
Above the level of initial mound growth (F2) there is a general repetition with a coralgal facies at the base and an organomicritic facies towards the top. The most important difference relative to level F 1 below is that the organomicritic facies is not only well developed within cryptic space, but also covers outer spaces of the coralgal community. The migration of the composite organomicritic facies from cryptic to non-cryptic space is clearly evident from the fact that coarse allochthonous debris (e.g. codiacean algae, gastropods, fragments of scleractinian corals) exhibit thick crusts of organomicrite (PI. 65/3-4). Acanthochaetetes is still in cryptic position and all contacts of encrusting fabrics with the final marly packstone cover correspond to organomicrites.
The F3-1evel shows drastically reduced organomicritic structures at outer rims as well as within cryptic space (PI. 65/5). Instead there is a partial recovery of the coralgal community composed ofPlacophyllia, Lithophyllum, and entrusting foraminifera ( P lacospilina, Coscinophragma).
The lower part of the syn-mound interval (subunits G- H) episodically exhibits an autochthonous fauna of pre- dominantly microsolenid corals and thick encrustations by the coralline sponge Euzkadiella. The cryptic space below Euzkadiella is frequently occupied by Acanthochaetetes seunesi (PI. 65/6), i.e. in identical position as within the replacement in terval below. Most parts of the interval bear biodetrital rocks such as tempestite beds dominated by requienid rudists and chondrodont bivalves as well as oncoid grain-/rudstones (subunit H). On regional scale the base of subunit Grepresents the first occurrence of dendroid red algae with Agardhiellopsis cretacea and Paraphyllum primaevum (PI. 65/6). This peculiar level is of high strati- graphic significance and allows the correlation with del- taic deposits and mixed siliciclastic/calcareous turbidite beds within the basin (NEtrW~t~R 1995). Despite this and the fact of incorporated organomicrite clasts and larger fragments of lithistid sponges (which indicate the syn- mound character), by general composition and texture this interval again represents a common deposit of adjacent Upper Aptian and Lower/Middle Albian platforms.
2.2 Community replacement, throphic structure and facies geometry
The compositional variation of the benthic community in lateral beds of initial mound growth provides direct
evidence that mound growth is not only a local phenom- enon but also exhibits a regional effect documented as a biofacies discontinuity. This discontinuity can be described as a replacement of a highly skeletonized association, mainly calcareous, i.e. the coralgal community, by an association of low skeletal production, calcareous, siliceous and agglutinated, i.e. the composite organomicritic facies. Along this replacement the trophic structure seems to change from an assemblage dominated by tentacle feeding microcarnivores and photoautotrophic primary producers to an assemblage dominated by decomposers and actively filter feeding organisms. Organomicritic facies exhibit a strong evidence for high rates of biodegradation as con- cluded from biomarker analyses (Trrt~ pers. comm.; cf. NEtrWmLE~ 1995: 219-220). Outocrop conditions are not good enough to give accurate information on the geometry of this replacement level. About 400 m to the W the marly lithofacies grades into limestones dominated by scleractinian corals, about 300 m to the E it should grade into the lower parts of the mud mound 'La Gandara' (cf. Pl. 65/1). The marly lithofacies and the associated benthic community indicate episodes of decreasing carbonate productivity suggesting stratigraphic condensation. Composition of beds immediatly above the replacement level suggests that there was already a remarkable mound relief by time these were deposited ('empty bucket' geometry, base ofsubunit G).
3 COMMUNITY REPLACEMENT AT CORALGAL REEF BODIES
From the mud mound of'La Gandara' about 6 km to the E there is an occurrence of an isolated reef platform in vicinity of the village Aja (Fig.23). The upper parts of these deposits interfinger with a sequence of rhythmically bedded limestone/marl alternations (PI. 65/7). This se- quence bears belemnites ofNeohibolites cf. minimus thus indicating the higher parts of the Lower Albian (Douvilleicerasmammillatum zone). At this level a transect of about 100 m with a thickness of 5 m was suitable for sampling (cf. Fig. 25).
3.1 Description of compositional variation (Fig. 25) Reef core sediments are poorly washed pack- and
rudstones (PI. 65/8). The detrital reef rock bears meandroid and dendroid scleractinian corals, radiolitid and requienid rudists, codiacean algae (Arabicodium) and encrusters such as Polystrata, Porostromata, Lithocodium, and bryozoans. Endolithic traces of microborers are very fre- quent. At the reef margin there is an autochthonous docu- mentation of scleractinian corals (dendroid, meandroid, massiv) encrusted by Polystrata. In cryptic spaces small siliceous sponges (mainly lithistid demosponges) occur. Minor elements are encrusting foraminifera, brachiopods and stromatoporoid sponges. In course ofa retrogradational trend of facies this coralgal facies was covered by an organomicritic facies dominated by lithistid demosponges and hexactinellids, a community weU-known from adja- cent mud mounds (cf. NEtr~mmE~ 1995).
The establishment of the organomicritic facies resulted in a secondary frame, filling void space which otherwise

267
Fig. 25. Sketch of facies geometry along the re- placement of a eoralgal reef community by an organomicritic/silieeous sponge cover (vicinity of the village Aja, ef. Fig. 23)
would be simply occupied by soft detrital reef sediment (PI. 65/9). Growth forms of organomicrite comprise thin coatings upon scleractinian corals and stromatolitic fab- rics. At this early stage of space filling the community consists of encrusting foraminifera (Placopsilina), monocrepid and tetracrepid lithistid sponges, non-lithistid demosponges, Acanthochaetetes seunesi, and thecidean brachiopods. The general decrease of coral fragments and the close vicinity to the final surface of the whole reef body with a massive organomicritic facies rather indicate a descendent entering of the organomicritic facies from the top of the carbonate body!
This uppermost level of autochthonous carbonate pro- duction consists of organomicrites (thrombolites, stromatolites, massiv fabrics) and a diverse association of lithistid demosponges, non-lithistid demosponges, Hexactinosa and terebellid polychaetes (PI. 65/10-11). At this levelAcanthochaetetes seunesi is still found in cryptic position (PI. 65/10)! Residual pore space is partly filled with a microbioclastic wackestone rich in small intraclasts and scattered sponge spicules (PI. 65/11), a facies identical to that of the adjacent, rhythmically bedded limestone marl alternation.
3.2 Community replacement, throphic structure and facies geometry
In this case, in cause of the lack o f accurate stratigraphic correlation, observations on the compositional variation of the benthic community are restricted to the local scale. Again the biofacies discontinuity represents a replacement of a highly skeletonized association by one of low skeletal production and the trophic structure changes again from tentacle feeding microcarnivores and photoautotrophic primary producers to active filter feeding organisms and decomposers. Relatively low rate of carbonate production is not expressed by lithology (massiv limestones), as it was the case along the attached carbonate platform of the previous example. But lower rates are well-expressed along the replacement level by a spatial limitation of massive carbonate lithology to 1/3 of the precursory coralgal reef body (cf. Fig. 25). The retrogradational geometry proceeds and lead to a final burial of the carbonate body by prodelta-deposits.
4 CAUSES OF COMMUNITY REPLACEMENT- A DISCUSSION
Replacements of the benthic community and associ- ated shifts of facies described above can be referred to the initial definition of 'platform drowning' or reef drowing (ScnLAGER 1981: 197). With time the concept of platform drowning evolved far beyond the causal mechanism of a relative rise of sea-level (e.g. SCHLAGER 1989; HALLOCK & SCHLAGZR 1986; MULUNS & H~E 1989). E.g. ERUCH et al. (1990) summarized 20 possible causes of drowning events. Within this context three possible causes should be dis- cussed in more detail: i) a rise of relative sea-level, ii) an oceanic anoxic event and possibly related nutrient floods, and iii) other modes of increased nutrient supply.
In both cases a relative rise of sea-level can be deduced from large-scale backstepping, sequential patterns, and distinct decreases ofbioerosion by microborers (NEUWEmER 1995). However, in the case of lateral beds adjacent to the mound 'La Gandara' the fact that Acanthochaetetes re- mains within its cryptic position throughout the sequence demonstrates that relative rise of sea-level could not sub- merge this platform area below the photic zone! Aphotic environments reveal open water occurrences of Acantho- chaetetes two to fivefold in size (REITNER et al. 1995; NEVW~LER 1995). In the case of community replacement upon a coralgal reef, A canthochaetetes is not as frequent to provide a clear picture throughout the sequence. Despite that I stress the point that Acanthochaetetes is still found within its cryptic position at the uppermost level of the organomicritic/siliceous sponge cover. Therefore, there is no direct evidence that at the levels of community replace- ment any of the platform areas described above was sub- merged below the photic zone during the relative rise of sea-level.
Mid-Cretaceous oceanic anoxic events (OAE) cover the Aptian/Albian episode with possible subevents in the Lower Albian (ARTHUR et al. 1990). OAEs may effect the growth potential of carbonate platforms via nutrient floods resulting from mid-ocean overturn (HALLOCK & SCHLA~Fa 1986; SCHLAOER & PmLW 1990). In the regional context of the Soba Reef area there is no evidence of black shales within the adjacent basin areas. TOC values of dark basinal

268
sediments aregenerallybelow 1% (PAscAL 1985; NEtrw~u~R 1995), and no redox-related benthic events (sensu SAWDA et ai. 1991) have been recognized so far. However, selec- tive measurements of ~513C (NEuwEIt~R 1995: Fig. 89) fit well with the elevated values of the average carbon iso- topic composition to be expected for the Aptian/Albian OAE (cf. AR~trR et al. 1990: Fig. 2). In conclusion, there is no evidence for major effects of anoxic events and related nutrient floods via oceanic overturns.
Other modes of increased nutrient supply comprise episodic upwelling of relatively cold, nutrient rich bottom waters and, from opposite direction, terrestial derived nutrients brought in by rivers and related deltaic systems. Both cases fit well with the general depositional condi- tions found within the Soba Reef area. The siliciclastic unit below the establishment of the mud mound 'La Gandara' represents an extensive period of continental influence, prograding into larger parts of the basin (NEtrw~.it~R 1995: 189-192). In the case of the coralgal reef, prodel ta deposits occur in the course of the proceeding deepening-upward cycle. But in any case there is a clear time-offset between siliciclastic influences as confined to the deposition of siliciclastics and community replacements. It is not clear whether the associated water masses could influence the benthic community 'out of phase' with siliciclastic sedi- ment accumulation.
Episodic influences of nutrient-rich bottom waters migrating along, and episodically across, contour lines could explain the spatial distribution of mud mounds along the platform to basin transitional zone, ranging from deeper aphotic environments up to the photic zone (cf. NEVWEmER 1995 megabenthos variation within those mounds). These influences may be facilitated during times of relatively rising sea-level. The combined variations in nutrients, water temperature and (slight) water depth might be strong enough to compete the stenotopic coraigal community out. In order to elucidate such processes we need to combine palaeontological data with fabric-selective data from biogeochemistry and geochemistry on the base of an enhanced stratigraphic resolution.
Part XX AEROBIC AND ANAEROBIC
MICROORGANISMS IN MODERN SPONGES: A POSSIBLE RELATIONSHIP TO
FOSSILIZATION-PROCESSES
G. Schumann-Kindel, M. Bergbauer, W. Manz, U. Szewzyk and J. Reimer
In this study bacteria present in the tissue of the Mediterranean sponges Chondrosia reniformis andPetrosia ficiformis were examined by cultivation and culture-inde- pendent methods. In this approach, culture dependent techniques have been combined with in situ hybridization usingfluorescently labeled rRNA-targeted oli gonucleotide probesspecificforBacteriaandsubclassesofProteobacteria.
Hybridization of the associated microbial populations in both sponges showed that the majority of bacteria are members of the gamma-subclass of Proteobacteria. Using highly specific oligonucleotide probesfor detecting sulfate- reducing bacteria, distinct signals could be found scat- tered in native sponge tissue of both sponges. In all enrichment cultures with lactate as sole carbon source which were inoculated with tissue from the two sponges C. reniformis and P.ficiformis, bacteria could be detected by in situ hybridization using a highly specific probe for sulfate-reducing bacteria of the Desulfobacter-cluster. Not only sulfate-reducing bacteria were enriched but also other fermentative bacteria who are involved in the degra- dation of sponge tissue. Sulfate reducing bacteria may control the calcification of the sponge tissue during degra- dation because increasing the carbonate alkalinity.
1 INTRODUCTION The presence of bacteria in living sponges is well
documented, mostly by microscopic (VAcELET & DONAOEY 1977; WmraNsoN 1978a,b; R~la'r~a 1993) and culture tech- niques (SAr,rrAwv et al. 1990; WILraNSON 1978a). Since only a small portion of the bacteria could be selected and partly characterized from few sponges, the function of the majority of the sponge-associated microorganisms is still unknown. Especially, it is not known if and how the sponge and its microbial residents interact and if some of the microorganisms might be obligatory for the survival of the sponge. Possible interactions might range from nutri- tional aspects, similar to the action of bacteria in the intestinal tract of higher animals, to processes like the formation of an inorganic skeleton. Such organo- and/or biomineralization processes might even have occurred during the fossilization of sponges, as indicated by the presence of pyrite, on various sponge-fossils, but also fossils of other animals (e.g. ammonites, echinoderms etc.). Pyrite is one of the most common type of iron/ sulphur minerals in nature. For its formation, anaerobic conditions are required, as they establish during degrada- tion of organic matter. Since the isolation and cultural recovery do not correlate with the real existing microbial populations, and therefore their function, ecology "and survival strategies of the aerobic and anaerobic bacteria in sponges are poorly understood. The aim of this study is the identification and characterization of all inhabitants, espe- cially of the sulfate-reducing microorganisms (SRB), within the selected sponges. Furthermore we plan to evaluate the relevance and function of the bacteria for the sponge. Additionally, the involvement of these sponge-associated bacteria in biomineralization processes, e.g. the formation of pyrite during fossilization, shall be investigated.
2 EXPERIMENTAL DESIGN Aerobic and anaerobic cultures and isolation of pure
cultures specimen of C. reniformis and P. ficiformis were taken in rocky habitates at 5-10 m water depth by SCUBA- diving. Sampling sites were located at the western shore of Corfu, Greece. Samples for the molecular analysis as well as inocula for cultivation studies were prepared under

269
aseptic conditions from the inner part of a specimen. Cultivation of aerobic sponge-associated bacteria was performed with marine media: commercial (Marine Agar/ Broth 2216, Difco) and modified V-medium and Lewins marine medium (LEwIN & LOtrNSBERV 1969; V~,AT~,NEN 1976) differing in their nutrient conditions, each prepared as agar plates as well as liquid assays.
Anaerobic bacteria were cultivated in an anoxic, bicar- bonate-buffered, sulfide-, respectively titaniumcitrate-re- duced mineral salt medium; chemical stock solutions were prepared according to WrODEL & P ~ o (1981) and asepti- cally added to the autoclaved medium. For enrichment of sulfate-reducing bacteria, basal media were supplemented with lactate (20 raM), propionate (10 mM), benzoate (5 raM), acetate (20 mM) or yeast-extract (0.1%), respec- tively. Na2SO 4 was supplemented to all enrichments at a final concentration of 20 mM.
2.1 in situ hybridization Sponge tissue and cultured bacteria were fixed in 3.7%
formaldehyde, in situ hybridization was done with fluores- cent-labeled oligonucleotide probes complementary to regions of the 16S and 23S rRNA characteristic for the domains Archaea (probe ARCH915, S'rAm. & AMANN 1991) and Bacteria (probe EUB338, AMANN etal. 1990) as well as the alpha-, beta-, gamma-, and delta-subclasses of Proteobacteria (probes ALFlb, BET42a, and GAM42a, respectively; MArCzetal. 1992),Flavobacteria-Cytophaga- group (probe CF319, MArCZ et al. 1996) and species- specific probes for sulfate-reducing bacteria (developed by Manz et al. pers. comm.). Probe-conferred fluorescence was detected with aZeiss Axioskop fitted for epifluorescence microscopy with corresponding filter sets.
2.2 Sequencing and phylogenetical characterization The 16S rRNA-genes from single bacterial species
were amplified by PCR (polymerase-chain reaction) using universal primers (27F, 1492R, M~t~LIN et al. 1988). The retrieved amplificates were sequenced non-radioactively after purification with a direct-blotting electrophorese system (HoEFER). The received sequences were analyzed phylogenetically by computer-aided alignment-analysis (Software: ARB-program, developed by Sa~u~a< et al., TU M~inchen).
3 RESULTS In this study bacteria present in the tissue of the Medi-
terranean sponges Chondrosia reniformis and Petrosia ficiformis were examined by cultivation and culture-inde- pendent methods. Thereby, culture dependent techniques (aerobic and anaerobic) have been combined with in situ hybridization using fluorescently labeled rRNA-targeted oligon ucleotide probes specific for Archaea, Bacteria and subclasses of Proteobacteria.
3.1 Hybridization of sponge tissue The sponge material prepared for the inoculation of the enrichment cultures was used in parallel assays for in situ determination of bacteria without prior cultivation. In situ
hybridization of native sponge tissue with the Bacteria- specific probe EUB 338 gave strong fluorescent signals. Hybridized bacteria of various morphological types ob- tained from P. ficiformis are shown in PI. 66/1. Bacteria within the sponge tissue of C. reniformis, hybridized with the same probe (EUB338), were dominated by rod-shaped cells (PI. 66/2). Hybridization of the associated microbial populations in both sponges revealed that the majority of bacteria are members of the gamma-subclass of Proteo- bacteria. Using newly developed, highly specific oligonucleotide probes for characterization of sulfate- reducing bacteria, distinct signals could be found scattered in native sponge tissue of both sponges.
3.2 Aerobic cultivation of sponge material Sponge-associated bacteria could be successfully cul-
tivated from both sponges. The obtained bacteria revealed a broad range of different morphotypes within the culturable population. These organisms differed in shape, size, and morphology of agar-colonies. In situ hybridization with domain- and subclass-specific probes revealed that all detectable bacteria in the enrichment cultures inoculated from C. reniformis and P. ficiformis could be phylogeneti- cally characterized as members of the alpha- and gamma- subclass of Proteobacteria. Bacteria forming colonies on agarplates were selected for isolation according to colony and cell morphology. From the pure cultures, obtained from the sponges, DNA was isolated and subsequently sequenced in parts. Most of the isolated bacteria are facultative anaerobes. First results of computer-aided se- quence analysis of these isolates indicated that they are new species, not described till now, clustering in the Vibrio-group of gamma-Proteobacteria. Bacteria of the alpha-subclass could not be isolated so far.
3.3 Anaerobic enrichment cultures In all enrichment cultures inoculated with tissue from
the two sponges C. reniformis and P. ficiformis and sup- plemented with lactate as sole carbon source, bacteria could be detected by in situ hybridization using a highly specific probe for sulfate-reducing bacteria of the Desulfobacteriaceae-cluster. While the subpopulation of sulfate-reducers of C. reniformis was dominated by small vibrios, the respective population of P. ficiformis con- sisted mostly of vibrios of different size. This is clearly demonstrated in P1.66/3 and PI. 66/4a, where both cultures were simultaneously hybridized with the same probe.
Additionally, it is shown in PI. 66/4b that not only sulfate-reducing bacteria were enriched but also other fermentative bacteria who are involved in the degradation of sponge tissue. In the enrichment cultures obtained from these sponges the subpopulation of sulfate reducers was never dominated by one species, but always consisted of at least three morphotypes species (PI. 66/5). These vibrios isolated from lactate-enrichment, hybridized with a probe for the Desulfovibrionaceae-cluster, differed evidently in morphology from those vibrios detected with a probe for the Desulfobacteriaceae-cluster.

2 7 0
Sea Salt e.g. S042
H2S '
H2S Fe 2+
' I Sulfate-reducti~
, Fe + , T [f i j ] ,
FeS2 + H2
l M~icrobial J degradation: fermentation
4 DISCUSSION There is a distinctive population of facultative and
strict anaerobic bacteria within the selected sponges. Due to the different enrichment assays of the sponge-associ- ated anaerobic bacteria it is possible to characterize the involved organisms in regard to their physiological prop- erties and phylogenetical affiliation. Examinations with SRB-specific probes revealed that there is no dominance of a certain sulfate-reducing organism. The cultivated sulfate reducing organisms cluster in the Desulfobacter- and Desulfovibrio-gronp of the delt,!-Proteobacteria. Apart from the sulfate-reducing bacteria a lot of other anaerobic organisms with different morphotypes could also be en- riched. Interestingly, they gave a strong hybridization signal with the universal Bacteria-probe EUB338, but these organisms emitted no signal after hybridization with commonly used group specific probes. This indicates, that a greater portion of the sponge associated population are unknown organisms. Their relationship to the sponge will be elucidated by isolation and subsequent physiological and phylogenetical characterization.
The design of individual probes, which is currently in progress, will allow to clarify the relevance and specificity of these putative fermentative organisms within the sponge. Especially these organisms play a significant role in the degradation of organic material. During sponge-degrada- tion, it is obvious that the inner part of the sponge will become anoxic as soon as the inflow of fresh seawater
Sponge (source of C,S,N,P,Fe)
NH3 + C02 + Ca 2+
GAG03
Fig. 26. Summary dia- gram illustrating the re- lationship between meta- zoa degradation and mi- crobial influence onpyrit- ization and calcification
stops. Due to the combined action of the mixed populations within the sponge, the organic carbon of the tissue will be oxidized to CO 2 and sulfide will be formed from the degradation of organic S-compounds and from the reduc- tion of sulfate. The ongoing degradation process will result in a further decrease of the redox potential in the remnants of the sponge. In this specific environment, it is possible that sulfide is immediately precipitated as FeS and, pro- vided that high concentrations of sulfide and iron are present, as FeS 2 (pyrite). The formation of pyrite during growth of anaerobic, sulfide producing bacteria is well documented, even for pure cultures (S~TrER et al. 1983). Additionally, it was shown that the attachment o f anaerobic bacteria to amorphous ferrous sulfide increases their activ- ity, so that the preliminary stage of pyrite supports the formation of an anaerobic biofilm (SzzwzvK & ScmNr 1991).
Organomineralization processes are very complex and based on various mechanisms. For sulfide-based bit- mineralization processes, especially for the pyritization of marine sponges, it can be assumed that the main source of sulfide-production is the dissimilatory sulfate-reduction, carried out by a variety of bacteria, referred to sulfate- reducing bacteria Another source of sulfide formation is the degradation of organic sulfur compounds, e.g. sulfur containing amino acids. With this hydrogen sulfide and chemically or bacterially catalyzed reduction of ferric iron to ferrous sulfide (FeS), pyrite formation takes place
P l a t e 66
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Aerobic and anaerobic microorganisms in modem sponges: a possible relationship to fossilization- processes (Scale bar of all Figs.= 10 I-un)
Epifluorescence micrographs of in situ hybridization of Petrosia ficiformis tissue with the Bacteria- directed probe EUB338. Epifluorescence micrographs of in situ hybridization of Chondrosia reniformis tissue with FLUOS- labeled Bacteria- probe EUB338. Epifluorescence micrographs of in situ hybridization of anaerobic enrichment culture with lactate from Chondrosia reniformis tissue with CY3-1abeled highly specific probes for sulfate-reducing bacteria of the Desulfobacteriaceae-cluster. Epifluorescence (a) and phase-contrast (b) micrographs (identical fields) of in situ hybridization of anaerobic enrichment culture with lactate from Petrosia ficiformis tissue with a highly specific CY3- labeled probe for sulfate-reducing bacteria of the Desulfobacteriaceae-cluster. Epifluorescence micrographs of in situ hybridization of anaerobic enrichment culture with lactate from Petrosia ]iciformis tissue with highly specific CY3-1abeled probes for sulfate-reducing bacteria of the Desulfovibrionaceae-cluster.

P l a t e 66 2 7 1

272
accompanied by a release of hydrogen (DRoBNER et al. 1990).
2 FeS + 2 H2S --> 2 Fe(SH)2 2 Fe(SH) 2 --> 2 FeS 2 + 2 H 2
Under anaerobic conditions, reduction of ferric iron to the ferrous state can occur if appropriate electron-donors are present, such as organic compounds or H 2. The hydro- gen then can also be used for CO2-fixation by hydrogen- consuming bacteria or it can diffuse out of the site of pyritization. The pyritization can go hand in hand with the calcification. The degradation of organic matter leads to the formation of NH 3 and CO 2 as a result of the total oxidation of organic matter. During this oxidation process, the received organic acids temporarily compensate the alkalinity and so does the increased CO2-concentration. The organic acids are oxidized to CO 2 which is partly precipitated as calcium carbonate, provided sufficient concentrations of Ca-ions. Therefore, the pH may slowly increase.
These processes are summarized in Fig. 26. This scheme points out that central compounds for pyritization and calcification, such as H2S, CO 2, H 2, and NH 3, occur from microorganisms. In living sponges, sulfate reducing bac- teria are distributed rarely within the tissue. This indicates the presence of anoxic microniches, which allow survival and subsequent growth of anaerobic bacteria. For sulfate- reducing bacteria, it was even shown that some species are somewhat tolerant towards oxygen or even use it as elec- tron-acceptor for the generation ofATP (DmLtNG & CYPIONKA 1990).
Further investigations of culturable sponge-microor- ganisms are currently carried out to investigate the ecol- ogy of sponge-associated microorganisms.
This study will elucidate the physiological properties and, by molecular analysis, the phylogenetical affiliation of these organisms. Occurrence and spatial distribution of even uncultured bacteria in sponge tissue will be analyzed by in situ hybridization with specifically designed rRNA- targeted oligonucleotides.
Part XXI PYRITE IN MINERALIZED SPONGE TISSUE - PRODUCT OF SULFATE REDUCING SPONGE
RELATED BACTERIA?
J. Reitner and G. Schumann-Kindel
Isolated pyrite crystals are common in taphonomically mineralized sponges tissues (automicrites). In the sur- rounding sediment, pyrite is absent or rare. The sponge tissue automicrites are often dark coloured due to statisti- cally distributed very fine pyrite crystals (approximately I pm or smaller). Besides the smallpyrite, larger crystals often exhibit patchy concentrations or they are arranged in rows. Pyrite formation is probably linked with sulfate reducing symbiontic bacteria in the sponge mesohyle. Modern sponges possess mesohyle symbiontic bacteria
including sulfate reducing and further anaerobic bacteria. During early decaying processes of the sponge tissue the internal sponge space is becoming entirely anaerobic which favours the growth of the sulfate reducing bacteria. The sulfate reducing process significantly increases the carbonate alkalinity which is necessary to form automicrite. This process may explain the rapkt calcification of sponge tissue in modern marine microbialites and ancient sponge mud mounds.
1 INTRODUCTION Sponges are a main faunistic element in many ancient
micritic buildups (mud mounds) and modern marine microbialites (REITNER 1993; REITNER & NEUWEILER 1995; WARNKE 1994; FLAJS 8Z Ht~SSNER 1993; NEUWEILER 1993, 1995; BOURQUE & GIGNAC 1993; KEUPP et al. 1990; a.m.o.). Most of the sponges are nearly entirely preserved due to a special taphonomic calcification (automicrite formation). From biomarker analyses we know that sponge related microbes play a major role during the automicrite forma- tion (THIEL et al. 1996). Investigations on symbiontic bacteria in sponges by SCHUMANN-KINDEL et al. (this vol- ume) have shown that sulfate reducing bacteria are an integrated part of the sponge related bacterial community. In decaying tissues of sponges from a spiculitic biofacies of the Arctic Seamount Vesterisbanken (HENRtCH et al. 1992), pyrite was commonly observed within the degraded tissue. Taphonomic studies on open water sponges was carried out by LINDENBZRGER (1993). She has demostrated that under such conditions a taphonomic calcification does not happens. Purpose of the presented paper is to demon- swate the pyrite distribution in some taphonomically lithified sponges from marine microbialites, from Cretaceous sponge dominated basin marls, and from Carboniferous and Cre- taceous mud mounds. Biogeochemical and geochemical investigations are still in progress and, therefore, the presented paper is only a status report and should be understood as a concept study.
2 MATERIAL AND METHODS The subfossil sponge specimens were collected in reef
caves of Lizard Island and the nearby islands (Great Barrier Reef, Australia). In these caves, the sponges are integrated in thrombolitic microbialims. Abundant mate- rial from spiculithic mats was collected during the cruise ARK VII/4 oftheRV Polarstern 1990 to the Vesterisbanken Seamount in the central Greenland Sea (HENRICH et al. 1992). The fossil material for this study was collected by Dr. WARNKE in the Sligo mud mound (North-Western Ireland) (WARNKE & MEISCHNER 1995; WARNKE 1994), by JR in Lower Cretaceous mud mounds of northern Spain and from the Campanian basin marls of the HOver quarry (near Hannover, northern Germany).
The recent/subfossil specimens were biologically fixed in buffered formol or buffered glutardialdehyde. The speci- mens were embedded in LRWight and sectioned with a hardpart microtome and then stained with various histo- logical staining dyes (detailed procedure in REITNER 1993). Pyrite was decteted with X-Ray diffraction, SEM-EDX,

273
and microscopically. S-isotope analyses are still in progress. For cultivation procedure and determinations of recent sponge bacteria see SCmrMANN-KItcOEL et al. (this volume).
3 RESULTS Mineralizing events of sponge tissues are restricted to
sponges which are located in small caverns like boring cavities of excavating sponges (e.g.Aka, PI. 67/4), semiclosed sedimentary pockets, specimens within thick spiculitic mats or sponges with a rigid skeleton combined with thick organic tissues (rigid hexactinellids, lithistids, sponges with a basal skeleton). We never observed any taphonomic calcification in sponges without these protective features. During decay, the tissue is becoming disarticulated and is normally fed or oxidized by various decomposers (e.g. proteolitic and saccharolytic bacteria like gram positve taxa of the Clostridium-group). Recent investigations on microbiological enrichments cultures of some sponge re- lated bacteria have demonstrated that besides aerobic bacteria, anaerobic ones are members of the sponge symbiontic microbial pobulation.
In situ hybridization of bacteria under aerobic condi- tions has shown that culturable bacteria from the hadromerid demosponge Chondrosia reniformis and the haplosclerid demosponge Petrosiaficiformis are phylogenetically char- acterized as members of the alpha- and gamma-subclass of Proteobacteria. First results have demostrated that these forms are linked with the Vibrio-group of gamma- Proteobacteria (PI. 67/5).
In anaerobic enrichment cultures, the subpopulation of sulfate reducers was never dominated by one species, but always consisted of at least three species. The cultivated sulfate reducing organisms are related in theDesulfobacter- and Desulfovibrio-group of the delta-Proteobacteria (ScHuMaNN-KJr~EL et al. this volume). These organisms also play an important role in the degradation of organic material. Currents, wave action, and biological activity disarticulate and transport the remains of the tissue. Only spicules or parts of connective tissue with spicules are at least found in the sediments (LINDENSERCER 1993). Taphonomic calcification of sponge remains were never observed under such conditions.
3.1 Lithified Sponges in Recent Marine Microbialites
Within marine thrombolitic microbialites of reef caves of Lizard Island and North Direction Island (northern Great Barrier Reef), sponges are important autochtonous metazoan fossils. The microbialitic crusts are ca. 3500 y old and mainly constructed of various types ofautomicrites (REn'N~R 1993; REITNER et al. 1995a,b). Biomarkers of sponges, e.g. demosponge fatty acids, are very common (TrnEL et al. 1996). Beside sponge related fossil molecules biomarkers of heterotrophic bacteria are present including terminally branched fatty acids characteristic for anaerobic bacteria. This type of fatty acids was also detected in various taxa of sponges growing in the reef caves with microbialitic crusts. Spicule skeletons are abundant within the microbialite often in inactive Aka boring cavities. The
spicule arrangements are surrounded by sponge character- istic taphonomic minipeloids or dark aphanitic automicrites. In the example presented here, the mesohyle tissue of the haplosclerid boring spongeAka is preserved in an aphanitic automicrite. Isolated pyrite crystals are common, some- times concentrated in small clusters of 50 gm size (PI. 67/ 4). The pyrite crystals are mostly anhedral orpolycrystalline and they have a size of ca. 5-10 gm. Framboid pyrite was never observed. Within the very fine crystalline automicrite the pyrite is statistically distributed. The pyrite crystals have a size of 1-2 Iam. Not in all taphonomically calcified sponge skeletons pyrite was observed, but sulfur and Fe was detected with EDX-analyses.
3.2 Pyrite in Decaying Sponges In histological sections of decaying sponges from auto-
chthonous spiculite mat of the top of the Artic Seamount Vesterisbanken, taphonomic minipeloid-automicrite for- mation was observed. The peloid formation starts within the bacteria loaded mesohyle, but the choanocytes resp. choanocyte-chambers are still preserved (PI. 67/1-2). The peloids are strictly related to the mesohyle bacteria (PI. 67/ 3). One type of the calcified bacteria is preserved as spindle shaped calcitic bodies with spherulitic crystal arrangements, a second type are clusters of automicritic small ovoid bodies (minipeloids with a size of 10-30 ~tm, PI. 67/3 and REITNER et al. 1995a). Pyrite crystals are present, but rare (P1. 67/2). They are randomly distributed and restricted to the mesohyle area. The crystals are small (2-3 ~m) and anhedral. In few cases the peloids have a dark core which is probably made by very fine distributed pyrite (PI. 67/3).
3.3 Pyrite in Sponges from Lower Campanian Basin Marls
A characteristic feature of all sponges from the Lower Campanian basin marls of the HOver quarry (Hannover, northern Germany) is a strong mineralization with pyrite. Pyrite occurs in all taxa, within non-rigid demosponges, lithisitid demosponges, and all types of hexactineilids. Distribution and type of pyritization is dependent on the taxon. The mesohyle structures of the various sponges are quite different and the amount of bacteria is variable between 10-60% of the entire biomass of the sponge (REITNER 1993). AS a rule hexactinellids bear fewer amounts of sponge related bacteria then demosponges. In lithistid demosponges up to 60% of the sponge biomass is related to the bacteria. Beside pyrite enrichments in the former mesohyle areas of the sponges, pyrite is often concentrated within the axial canals of the spicules. These axial canals are perfect anaerobic spaces and sulfate reducers degrade the proteic axial filament. Within the axial canals, framboid pyrite is common. Pyrite within the sponge related automicrites exhibits two morphological types. In hexactinellids, dense rows of small pyrite crystals are common (PI. 67/6), within lithistids, framboid pyrite are concentrated in heaps (PI. 67/7). Non-rigid demosponges exhibit, beside mesohyle pyrite often entirely pyritized spicules.

2 7 4
3.4 Pyrite Occurrences in Sponges of Fossil Micritic Buildups
In many late Paleozoic and Mesozoic sponge mud mounds, pyrite is enriched within the taphonomically lithified sponge bodies. Not all sponge mud mounds dem- onstrate this feature which is up to now not yet fully understood. It is probably related with the type of sponges (not all have significant amounts of bacteria) and the presence and amounts of Fe z+ . The distribution of pyrite within the sponge automicrites is comparable with those ones seen in the marine microbialites. Common are dark grey aphanitic automicrites and minipeloidal fabrics (PI. 67/8-9). The dark color is related to fine distributed pyrite with grain sizes below one micron (PI. 67]8). Framboid pyrite bodies are rarely observed and probably a diagenetic feature (FARaANO 1970). Within certain places of the taphonomically formed automicritepyrite exhibits a patchy concentration (P1. 67/9), aggregates in rows are not very common. Pyritization is in all observed cases strictly related to the mineralized sponge body.
4 DISCUSSION Within many modern sponges, beside aerobic bacterial
communities distinctive populations of facultative and strict anaerobic bacteria are present and related to the Desulfobacter- and Desulfovibrio-group of the delta- Proteobacteria. Apart from the sulfate-reducing bacteria other anaerobic microorganisms are also be enriched. In few cases, gram positive Clostridium bacteria were ob- served in histological sec tions which are typcial for anaerobic
proteolytic and/or saccharolytic bacteria. All these organ- isms generally play a significant role in the degradation of organic material. After the dead of a sponge, the inner part of the sponge tissue will become anoxic, if the specimen is located in a protected container situation. Only this taphonomic supposition allows at least an entire preserva- tion of sponge skeletons via taphonomic calcification. It is supposed that after the death of the sponge, the sponge related bacteria get out of control of the sponge bioactive substances like certain antibiotics. It starts a very rapid growth rate of some of the sponge related bacteria as observed in artificial decaying experiments carried out in small seawater aquaria during field work on the Lizard Island (Australia) and Dravuni (Fiji) research stations. The degradation process led to decreasing redox potential (- 400 mV), zero oxygene and an decrease of the pH from slightly alkalic values (8,2 normal seawater) to slightly acidic conditions (6,8) in the remnants of the sponge. The microbial populations oxidize the organic carbon of the tissue to CO 2 and sulfide will be formed by the degradation of organic S-compounds and from the reduction of sulfate. Under these conditions, it is possible that sulfide is rapidly precipitated as FeS. If higher concentrations of sulfide and iron are present, FeS z could be precipitated. The bacterial sulfate reduction increases significantly the carbonate alkalinity which controls the taphonomic calcification events. We have measured in the sponge decaying experi- ments high values of total alkalinity (10-40 meq/l). This process is completed by ammonification processes which additional increase the alkalinity.
P l a t e 67
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig.8
Fig. 9.
Pyrite in mineralized sponge tissue -Product of sulfate reducing sponge related bacteria?
Decaying haplosclerid demosponge with initially formed peloids and calcified mesohyle bacteria. Spongin fibres are colored in red, stained with basic fuchsin. Vesterisbanken Seamount, Central Greenland Sea. x Nicols + gypsum. Scale=500 IJ.m. Detail of Fig. 1 with still preserved ball-shaped choanocyte chambers and small pyrite crystals within the mesohyle (arrow). Scale=100 I.tm. Detail of Fig.2 with a cluster of minipeloids and small spherical bodies of entirely calcified bacteria. The dark color in the center of the peloids is fine pyrite. Scale=50 p.m. Entirely calcified (high Mg-calcite) boring haplosclerid demosponge Aka in life postion. Pyrite is arranged (dark spots) are arranged in rows (arrow); Lizard Island Bommie Bay reef cave, Great Barrier Reef, northern Australia. Scale--200t.tm. TEM section of the mesohyle of a haplosclerid sponge (Petrosia sp.) of Lizard Island with a large archaeocyte and numerous Vibrio-type symbiontic bacteria. Scale=2 Iaxn. Lyssacinose hexactinellid from the Lower Campanian of HOver (Hannover, northern Germany). Pyrite crystals are arranged in rows forming a dense network within the sponge automicrite. Scale=100 tJ.m. Patchy clusters of framboid pyrite within the lithified mesohyle of a lithistid demosponge (Megamorina) from the Lower Campanian of HOver. x Nicols; Scale=100 I.tm. Nearly complete preserved bail-shaped lithistid demosponge (Tetracrepida) of a late Albian mud mound (Albeniz, Northern Spain). The sponge mesohyle demonstrates a dark peloidal automicrite. The dark colour is related to very fine distributed anhedral pyrite. Larger pyrite crystals are randomly distributed within the automicrite. The dermal layer of sponge exhibits an aphanitic automicrite. Scale=100 ~m. Dark sponge automicrite from a lyssacine hexactinellid of the Lower Carboniferous Sligo mud mound (Northwestern Ireland) (leg. Dr. WARN~). Pyrite occurs as clusters and is concentrated within the mesohyle or former syncytial tissue. Lyssacine hexactinellids are the most important mud mound builders in the Paleozoic. Scale=50 ILtm
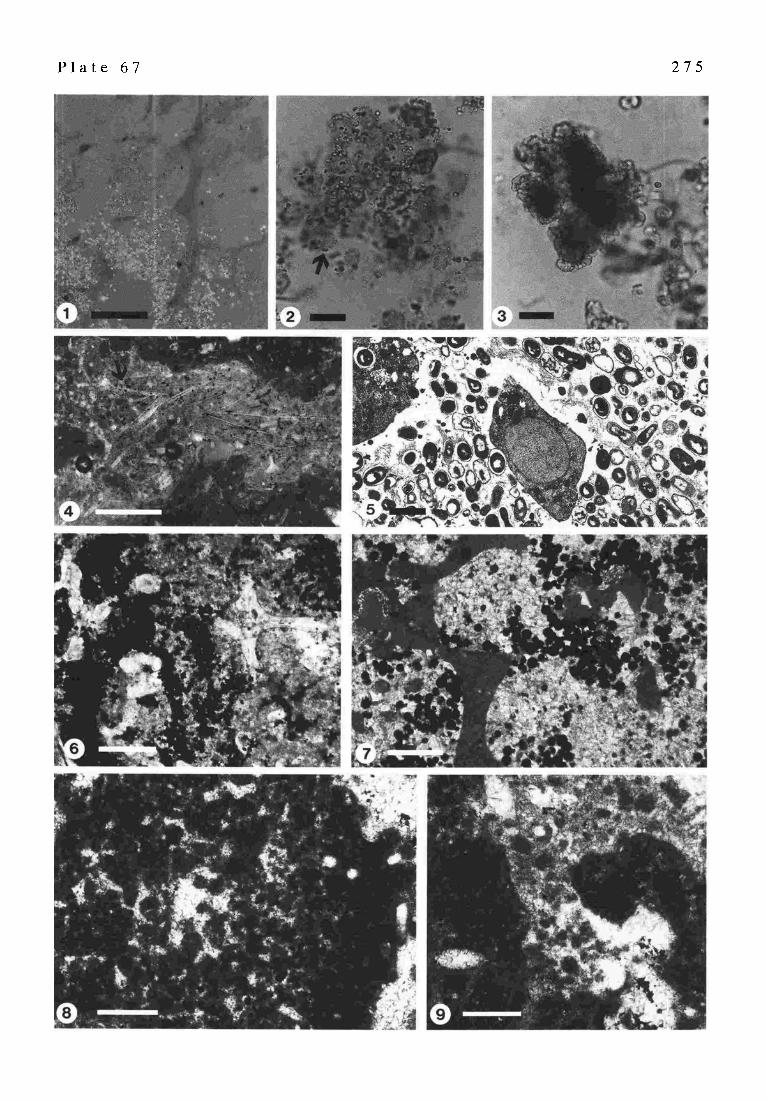
P l a t e 67 2 7 5

2 7 6
Sulfate reduction and ammonification increase alka- linity and favour the special taphonomic calcification process which is characteristic for many fossil sponge occurrences mainly in autochthonous spiculites and mud mounds,
ACKNOWLEDGEMENTS Mark Feldmann gratefully acknowledges the financial
support at the inception of his study from the SANW (Swiss Academy of Natural Sciences). The logistical sup- port was provided by the Caribbean Marine Research Center, Lee Stocking Island, Bahamas. M. Feldmann thanks Iwan St0ssel for his critical review which significantly improved earlier versions of his manuscript.
Stan Awramik's research is supported by a grant from NASA's Exobiology Program.
Pierre-Andr6 Bourque's research on bioconstruction through geological time, including the mound facies, is supported by an individual research grant from Natural Sciences and Engineering Research Council of Canada. He acknowledges a travel grant from the local German IGCP Committee to participate in the G0ttingen meeting.
Simonetta Cirilli wishes to thank Dr. R. Rettori (Perugia Univ., Italy) for his support in the determination of the palaeontological content. Her gratitude also to Dr. R. Riding (Cardiff Univ., UK), to Dr. J. Trichet (Univ. d'Orltans, France) and to Dr. H. Zankl (Marburg Univ., Germany) for the interesting and stimulating suggestions for continuing studies on this topic. Her research was supported by MURST 60 % Cirilli S. and CNR funds (PALAEO- PELAGOS-project).
James L. Pinckney & R.Pamela Reid are grateful for laboratory facilities and logistical support for sample col- lections and incubations provided by the Caribbean Ma- rine Research Center on Lee Stocking Island. They thank H. Paerl, M. Feldmann, P. Visscher, and K. Browne for comments, inspiring discussions, and assistance in the field. Their research was supported by National Science Foundation Grants OCE-9116296 and OCE-9415985.
Martina Merz-Preil3 is very grateful to E. Engel, Chemnitz, and L. Aghai-Soltani, Marburg, for the determination of the palynomorphs, to M. Joachimski, Erlangen, who meas- ured the stable isotopes and A. Schaper, Marburg, for the SEM work. Many discussions with H. Zankl, Marburg, increased her interest in the Thiiste stromatolites.
S6bastien Labiaux extends grateful thanks to Prof. Alan Lees for supervision (during studies for a doctorate) and useful comments on a draft manuscript, to Prof. G.D Sevastopulo, Dr. M. Philcox, and to Dr. J. Dehantschutter for their help in the field.
Aymon Baud and co-workers' research was supported in part by Swisss National fund (A. Baud, grant 20-45455- 95) by CNR and MPI 60% for Simonetta Cirilli and by CNRS for Jean Marcoux. Field works in Turkey receive a strong support from Prof. Celal Seng0r (Istanbul) who is gratefully acknowledged. Thanks to Glynn Ellis for im- proving the English text.
Gregory E. Webb and John S. Jell wish to thank P.
Coles of the Department of Earth Science, The University of Queensland for providing thin-sections of Heron micro- bialites.
Andr6 Freiwald et al. wish to thank the captain and his crew from R/V VICrOR H~NSEN for skillful operation on Sula Ridge, Norwegian Shelf. Their study is funded by the Deutsche Forschungsgemeinschaft and is part of the Prior- ity Programm 'Biogene Sedimentation' (Grant He-1671/ 1).
Manmohan Mohanti & Srikanta Das are grateful to Prof. E. Usdowski (Gtttingen, Germany), Dr. H. Edenkeuser (Kiel, Germany), and Dr. Ian M. Billing (Rijswijk, Neth- erlands) for isotope data of travertine-tufas and Prof. Howard J. Spero (Davis, U.S.A.) for isotope data of Gondwana carbonates. They thank Dr. W. de Priester and Dr. C.F. Winkler Prins (Leiden, Netherlands) for kind assistance with SEM micrographs.
Joachim Reitner and co-authors wish to thank D. KOrr~o, Max-Planck-lnstitute of Biophysical Chemistry, for teaching in microtomy and introduction to the TEM, Dr.WAONER from LEO Elektronenmikrokopie GmbH for the access to the LEO field emisson SEM Gemini with a cryostation at the GBF (Braunschweig), and Gabriela Schumann-Kindel (Microbiology, TU Berlin) for assisting cultivation of microorganisms. The'Studienstiftung des deutschen Volkes' is greatly acknowledged for travel expenses (Gernot Aria) The Deutsche Forschungsgemeinschaft has partly fi- nanced their study (project: 'Evolution of MulticeHular Systems and Organomineralization (EMSO), Leibniz Pro- grammeRe 665/12-1 and SFB 468/A 1). His work on ooids is a contribution to 'SFB 468 - Geologische Grenzflachen', Project A1; publ.-no. 1
Joachim Reitner and Gabriela Schumann-Kindel wish to thank Dr. K. Warnke (GOttingen) for the possibility to study thin-sections of the Sligo mud mound of Ireland. The Deutsche Forschungsgemeinschaft is greatly acknowl- edged for financing this study (Re 665/5, 7, 8, 12-1).
Fritz Neuweiler acknowledges the support of the Deutsche Forschungsgemeinschaft (German Research Foundation, Re 665/7, 8): His paper contributes to 'SFB 468: Geologische Grenzfl~ichen' (Project A 1; publ.-no.2) and EMSO: Evolu- tion of Multicellular Systems and Organomineralization.
REFERENCES
Aria, W.M. & S TArCroN, R.J. Jr. (1994): Comparative sedimentology and paleontology of Waulsortian mounds and coeval level- bottom sediments, lower Lake Valley Formation (lower Mississippian), Sacramento Mountains, New Mexico.-Abh. Geol. Bundesanst., 50, 11-24, Wien
-- & -- (1996): Constituent composition of Early Mississippian carbonate buidups and their level-bottom equivalents, Sacramento Mountains, New Mexico. In: S~OOEN, P., SOr.~aERVa.La, I.D. & JONES, G.L. (eds.): Recent Advances in Lower Carboniferous Geology. - Geol. Soc. Spec. Publ., 107, 83-95, London
ALVr~V.R, D., BAUD, A., G~x, J. & ST~reRZ, G. (1980): La limite Permien-Trias dans quelques loealitts du Moyen-Orient): recherches stratigraphiques et rnicropal~ontologiques. - Riv. hal. Paleont. Strat. 85, 683-714, Milano
AMAr~, R.I., Bm-o~, B.J., OLSON, R .J., Cmsnot~, S.W., DEvmEtm,

277
R. & S'rArm, D.A. (1990): Combination of 16S rRNA-tar- geted oligonucleotide probes with flowcytometry for analyzing mixed microbial populations. - Appl. Environ. Microbiol., 56,1919-1925
ANADON, P. & Z~ARR~O, I. (1981): Paleogene Nortrnarine Algal Deposits of the Ebro Basin, Northeastern Spain. In: MONTY, C. (ed.): Phanerozoic Stromatolites, 140-154, Berlin (Springer).
ANDREWS, J.E., RmmG, R. & DEm, as, P.F. (1993) : Stable isotopic compositions of Recent freshwater cyanobacterial carbon- ates from the British Isles: Local and regional environmental controls .- Sedimentology, 40, 303-314, Oxford
ARTllUR, M.A., JENKYNS, H.C., BRUMSACK, H.J. & SCHLANGER, S.O. (1990): Stratigraphy, geochemistry, and paleoceanography of organic carbon-rich Cretaceous sequences. In: G~sauRo, R.N. & BEAUDOIN, B. (eds.): Cretaceous Resources, Events and Rhythms. - NATO ASI Series, Series C, 304, 75-I19, Dordrecht (Kluwer Acad. Pub1)
ASLANIAN, A.T. (1984): Permian and Triassic deposits of the Transcaucasus. In: "The Armenian Soviet Socialist Republic Excursion t02, 27th Intemat. Geol. Congress, Guide book', 86-98, Moscow.
AWRAMIK, S.M. (1971): Precambrian columnar stromatolite di- versity: reflection of metazoan appearance. - Science, 174, 825-827, Washington
-- (1990): Stromatolites.- In: BRINGS, D.E.G. & CROW'nmR, P.R. (eds): Paleobiology: - A synthesis, 336-341, London (Blackwell Scient. Publ.)
AWRA~K. S.M., KIDDER, D.L., AND ZING, J. (1993): Collenia undosa, Charles W~COT'r, and lacustrine stromatolites. - Geol. Soc. Amer., Abstr. Progr., 25/6, A357.
AWRA~K, S.M. & RID~G, R. (1988): Role of algal eukaryotes in subtidal columnar stromatolite formation. - Proc. Nat. Acad. Sci. USA, 85, 1327-1329, Washington
BAEWI~CHI, P. (1952): Die Fraktionierung der Kohlensto ffisotopen bei der Absorption von Kohlendioxid. - Helv. Chim. Acta, 35, 1030-1036, Basel.
BAGHt~aN~, D. (1993): The Permian Sequence in the Abadeh Region, Central Iran. Occasional Publications ESRI, New Series 9, 7-22, Columbia (South Carolina Univ.)
B,xa't'ot.IN~ A. & Cmn.ta S.(1994): Early lithified microbialite on Jurassic palaeohigh of Sasso di Pale (Central Appennines, I ta ly) . - 15th I.A.S. Meeting, Ischia, Abstract book, 46.
B aT,URST, R.G.C. ( 1971): Carbon at sediments and their diagenes is. - Dev. Sedimentol., 12, 620 pp., Amsterdam (Elsevier)
BAUD, A. (1993a): From Murgabian (late Permian) to Olenekian (early Triassic) sequence stratigraphy: the Tethyan margins record. - Carboniferous to Jurassic Pangea Conference, Pro- gram and Abstract, 14, Calgary
-- (1993b):Tethysjustafter theendofPermianmassextinction. Carboniferous to Jurassic Pangea Conference, Program and Abstract, 15, Calgary
BAUD, A., MAGARITZ, M. &HoLsER, W.T. (1989): Permian- Triassic oflhe Tethys: Carbon isotope sudies.-Geol. Rundschau 78, 649-677, Stuttgart
BAUD, A., MARCOUX, J., GUIRAUD, R., Rtcou, L.E. & GAETaNa, M. (1993a): Late Murgabian (266-264 Ma). In: DERCOURT, J., R~cou, L.E. & VRIELYNCK, B. (eds.): Atlas Tethys, Palaeenvironmental maps, explanatory notes, 9-21, Paris (Gauthier-Villars)
BAUD, A., MARCOUX, J., GUIRAUO, R., Rtcou, L.E. & GAETANI, M. (1993b): Late Murgabian P alaeoenvironments (266-264 Ma). - BECIP-FRANLAB. CCGM, Reuil-Malmaison.
BE~, A. (1986): Stable isotopes, iron and phosphorus in a sequence o flacustrine carbonates - paleolimnic implications. - Chem. Geol., 59, 305-313, Amsterdam.
BE|X~W, R. (1982): Rigaudella, einneues Genus von Dino flagellaten- Zysten. -N.Jb.Geol. Pal., Mh., 1982, 137-150, Stuttgart.
BER'mAND-SARFA~, J. (1972): Les stromatolites du Prrcambrien Supdrieur du Sahara Nord-Occidental; inventaire, morphologie
et microstructure des laminations, corrrlations stratigraphiques. - Publ., Centre de Recherches sur les zones arides, Geol., 14, 240 p., Paris (C.N.R.S)
BERTRAND-SARFATI, J. & MouSSmE-PoucHKm'E, A. (1988): Is cratonic sedimentation consistent with available models? an example from the Upper Proterozoic of the West African Craton. In: JACKSON, M.J. (ed.): Aspects of Proterozoic sedi- mentary Geology. - S e d . Geol., 58, 255-276, Amsterdam
-- & -- (1993): Structures srdimentaires (seismites) dans des carbonates Nroprotrrozoiques du craton Ouest Africain (Algrrie). - 4 ~ Congr~s fran~ais de Srdimentologie, 45-46, Lille
BERTRAND-S ARFATI, J. & TROMPETrE, R. (1976): Useofstromatolites for intrabasinal correlation: example from the late Proterozoic of the northwestern margin of the Taoudenni basin. In: WALTER, M.R. (ed): Stromatolites, 518-522, Amsterdam (Elsevier)
BEarR~D-S~a~FA~, J., FREV~T, P. & I~.m~AT, J.-C. (1994): Micro- structures in Tertiary Nonmarine Stromatolites (France). Comparison with Proterozoic. - In: BER~o-SARFArI, J. & MONa'V, C. (eds.): Phanerozoic Stromatolites II. - 155-191, Berlin (Springer)
BOON, J.J., Hn~s, H., BURLING/~ME, A. L., RDPSTRA, W.I.C., DE L~UW, J.W., EDMONDS, K. E. & EGL~CrON, G. (1983): Organic geochemical studies of Solar Lake laminated cyanobacterial mats. In: BJOROY, M. et al. (eds.): Advances in Organic Geochemistry 1981,207-227, New York (Wiley)
BOSENCE D.W.J. (1979): The factors leading to aggregation and reef formation inSerpula vermicularis L.. In: L.~wooD G. & ROSEN B.R. (eds): Biology and systematics of colonial organ- i sms . - Syst. Assoc. Spec. Publ., 11,299-318, London (Aca- demic Press)
BOULVAn~, F. (1990): Srdimentologie et diagen~se des monticules micritiques frasniens 'F2j' de Betgique. - unpubl, doctoral thesis, Universit6 Libre de Bruxelles, 535 p., Bruxelles
BOUROUE, P.-A. (1983): Determinant role of sponges in the genesis of stromatactis facies in the Upper Frasnian red bioherms of Belgium. - I.A.S., 4th European Meeting, Split, Yougoslavia, Abstracts, 31-32, Split
BOURQUE, P.-A. & BOULW~q, F. (1993): A model for the origin and petrogenesis o f the red stromatactis limes tone of Paleozoic carbonate mounds.- J. Sediment. Petrol., 63, 607-619, Tulsa
Bo~OUE, P.-A. & GIGNAC, H. (1983): Sponge constructed stromatactis mud mounds, Silurian of Gaspr, Quebec. - J..Sediment.Petrot., 53,521-532, Tulsa
BOtrROUE, P.-A., MADI, A. & M~ET, B.L. (1995): Waulsortian- type bioherm development and response to sea-level fluctua- tions: Upper Vis~an of Brchar Basin, Western Algeria. - J. Sediment. Res., B65, 80-95, Tulsa
BOURQUE, P.-A. & RAYMOND, L. (1988): Non-skeletal bioherms of the lower reef complex of the West Point Formation, Late Silurian, Gasp~ Peninsula, Qurbec. - In: GELDSE'r~R, N.P., JAMES, N.P. & TEaBUYr, G.E. (eds.): Reefs, Canada and Adjacent Area. - Canad. Soc. Petrol. Geol., Mem. 13, 258- 262, Calgary
BotmQUE, P.-A., AMYOT, G., DESROCHERS, A.,GmNAC, H.,GossEL~, C., LACHAMBRE, G. & LPa.mER~, J.-Y. (1986): Silurian and Lower Devonian reef and carbonate complexes of the Gasp6 Basin, Qurbec - a summary. - Bull. Canad. Petrol. Geol., 34, 452-489, Calgary
BOURROtaLH, R. & BOURQUE, P.-A. (1995): Marqueurs d'rvolution de marges continentales palrozoYques: les monticules carbonatLs ~ stromatactis.-Bull. Soc. Grol. France, 166, 711-724, Paris
BOWLES, H., PaERL, H. & TUCKER, J. (1985): Effective solvents and extraction periods employed in phytoplankton carotenoid and chlorophyll determination. - Can. J. Fish. Aquat. Sci., 42, 1127-1131
BRAClIERT, T.C. & D~LO, W.-C. (1991): Laminar micite crusts and associated foreslope processes, Red Sea. - J. Sediment. Petrol., 61,354-363, Tulsa

278
BRAGA J.C., MAaTm J.M & Rmmo R. (1995): Controls on micro- bial dome fabric development along a carbonate-siliciclastic shelf basin transect, Miocene, SE Spain. - Palaios, I0, 347- 361, Tulsa.
BRow~, K. (1993): Lamination in recent Bahamian subtidal stromatolites: origin and lithification. -unpubl. Ph. D. thesis, University of Miami, 296 p., Coral Gables, FL,
BtmD, D.A. & PERg~s, R.D. (1980): Bathymetric zonation and paleoecological significance ofmicroborings in Puerto Rican shelf and slope sediments. - J. Sediment. Petrol., 50, 881- 904, Tulsa
BtncK, R. (1992): The antiquity of oxygenic photosynthesis - evidence from stromatolites in sulphate-deficient Archean lakes. - Science, 255,74-77, Washington
Btraca~-m, T.P. & Rmn~o, R. (1977): Attached vermiform gas- tropods in Carboniferous marginal marine stromatolites and biostromes. - Lethala, 10, 17-28, Oslo.
Btah'qE, R.V. & Moose L.S. (1987): Microbialites: organosedi- mentary deposits of benthic microbial communities. - Palaios, 2, 241-254, Tulsa.
C~om, G. F. & MoYrA~ot, v., L. F. (1994): High energy coralgal- stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia). - Sedimentology, 41, 655-676, Oxford
Cm~ozzl, A.V. (1962): Observations on algal biostromes in the Great Salt Lake, Utah. - J. Geol., 70, 246-252, Chicago
Ca~':ET'Z, H.S. (1986): Marine peloids: a product of bacterially induced precipitate of calcite. - J. Sediment. Petrol., 56, 812- 817, Tulsa
CHAF~TZ, H.& BUCZ'CNSKI, C. (1992): Bacterially-induced lithification of microbial mats. - Palaios, 7, 277-293, Tulsa
CaArd~Aa~an% A. (1993): Marine influence during Talchir depo- sition, Girdih basin, Bihar. - Ind. J. Geol., 65/4, 290-292, Calcutta.
C-'n~aqDaA, S.K. (1996): Marine signatures in the Gondwanas of Peninsular India and Permian palaeogeography.- Gondwana Nine, 1, 529-538, Geol. Surv. India, Calcutta.
ChemIC^ G., CmILU S., PASSERI L., TRINCIANTI E. & Z ~ l L. (1987): 'Anidriti di Burano' et 'Formation du Monte Cetona' (nouvelle Formation), biostratigraphie de deux series-types du Trias superieur darts l'Apermin septentrional. - Rev. Paleobiol., 6/2, 341-409, Genrve.
CnuLta S. (1992): Facies euxiniche nel Trias superiore dei Monti Simbruini (Appennino centrale).- Studi Geol. Camerti, Vol. Spec.1991/2, CROP 11,157-159, Camerino.
-- (1993) : I1 Trias di Filettino-Vallepietra (Monti Simbruini, Appennino Centrale). - Boll. Soe. Geol. Ital., 112, 371-394, Roma.
-- (1996): UpperTriassic biogenic mounds in the Northern and Central Apennines (Italy): palaeoecology and palaeobio- geographic implications. - Paleopelagos, 6, Roma (in press)
Cma.ta S., BUCra,ALO R. & Por,"rlrn R. (1994): Palynostratigraphy and palynofacies of the Late Triassic R. contorta facies in the Northern Apennines: II) The Monte Cetona Formation. - Rev. de Palrob., 13 (2), 319-339, Genrve.
COF~Ora, M. A. (1990): Mucous sheet formation on poritid corals: An evaluation of coral mucus as a nutrient source on reefs. - Marine Biology, 105, 39-50
C o w ~ R. (1988) : The role of algal symbiosis in reefs through time. - Palaios, 3, 221-227,Tulsa.
Cuw, J.P. & GAUTSET, P. (1995): Glucides et prot~ines de la matrice soluble des biocristaux de scleractiniaires acroporides. - C.R. Acad. Sei. Paris, 320, ser.IIa : 273-278, Paris.
DAV^UD, E., STgASSm~, A., JEDOUt, Y. (1994): Stromatelite and serpulid bioherms in a Holocene restricted lagoon (Sabkha El Melah, Southeastern Tunisia). In: BERTRAND-SARFATI, J. & MON~ C., (eds.): Phanerozoic stromatolites II, 131-151, Amsterdam (Kluwer)
DEc'no, A. (1990): Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine proc- esses. - Oceanogr. Mar. Biol. Ann. Rev., 28, 73-154
DI~FAROE, C., TRICHET, J., JAUNET, A.-M., ROBERT, M., TRIABLE, J. & Spdqso~m, F.J. (1996): Texture of microbial sediments revealed by cryo-scarming electron microscopy. - J. Sedi- ment. Res., 66, 935-948, Tulsa
I~9.FAROE, C., TRICHET, J., MAURIN, A. & HUCH~, M. (1994): Kopara in Polynesian atons: early stages of formation of calcareous stromatolites. - Sed. Geol., 89, 9-23, Amsterdam
Deom~s, E.T. (1976): Molecular mechanisms on carbonate, phos- phate and silica depositions in the living cell. - Topics on curt. Chem., 64, 1-12, Berlin
DEH,~acrscHtrrr~, J.A.E. & I.,8~, A. (1996): Wanlsortian buildups of Waulsort, Belgium. -Geo l . J., 31, 123-142, Chiehester
DILL, R. (1991): Subtidal stromatolites, ooids, and crusted-lime muds at the Great Bahama Bank margin. In: Osaoar~ R.H. (ed.): From Shoreline to Abyss. - Soc. Econ. Paleont. Min., Spec. Publ., 46, 147-171, Tulsa
Da.L, R.F., SHrm~, E. A., Jo~.s, A. T., KELLY, K. & ST~I'~, R. P. (1986): Giant subtidal stromatolites forming in normal salin- ity water. - Nature, 324, 55-58, London
Da .L~ , W. & CYPmNK^, H. (1990): Aerobic respiration in sulfate-reducing bacteria. - FEMS Microbiol. Lett., 71,123- 128
DONs, C. (1944) Norges korallrev. - Det Kongelige Norske Videnskabers Selskabs Forhandlinger, 16, 37-82
DR^vis, J. (1983): Hardened subtidal stromatolites, Bahamas. - Science, 219, 385-386, Washington
DRosr~, E., HusEa, H., W~,CHTERSH~,USSR, G., ROSE, D., STETr~g, K.O. (1990): Pyrite formation linked with hydrogen evolu- tion under anaerobic conditions. - Nature, 346, 742-744, London
EARDLEY, AJ. (1938): Sediments of Great Salt Lake, Utah. - Amer. Ass. Petrol. Geol. Bull., 22, 1305-1411, Tulsa.
EGLINTON, G. & Ha.~IILTON, R.J. (1967): Leaf epicuticular waxes. - Science, 156, 1322-1335, Washington
ErmLICH, H. L (1996): Geomicrobiology, pp719, New York (Marcel Dekker, Inc.)
EI~, L.I. (1978): Ocean currents on the Halten and Malangsgrunnen banks. - IKU-Report, 97, 1-302
ERt~CH, R.N., BAgREa'r, S.F. & GUo B ~ Ju (1990): Seismic and geological characteristics of drowning events on carbonate platforms.- Amer. Ass. Petrol. Geol. Bull., 74, 1523-1537, Tulsa
ERW~, D.H. (1993): The origin of metazoan development: a palaeobiological perspective. - Biological Journal of the Lirmean Society, 50, 255-274
ESaET, Y., R~,lpINO M., R. & VIsscHm, H. (1995): Fungal event and palynological record of ecological crisis and recovery across the Permian-Triassic boundary. - Geology, 23, 967- 970, Boulder
FtaRCmLD, I. J., EINSZLE, G. & SONG, T. (1997): Possible seismic origin of molar tooth structures in Neoproterozoic carbonate ramp deposits, North China. - Sedimentology (in press)
Ft, RR~aqD, M. (1970): Framboidal sulphides precipitated syntheti- cally. - Min. Deposits, 5, 237-247, Berlin
FARROW, G.E. & FYFS, J.A. (1988): Bioerosion and carbonate mud production on high-latitude shelves. - Sed. Geol., 60, 281-297, Amsterdam
F~LDM~r~, M. (1995): Controls on stromatolite formation: A comparative study of modern stromatolites from the Baha- mas with Messinian examples from southeast Spain - PhD Thesis, ETH Zilrich, 128 p.
F~LDr, t ~ , M. & McKmcz~ J.A. (in press): S tromatolite-thrombolite associations in a modern environment, Lee Stocking Island, Bahamas - Palaios, in press.
F1scam~, A,G. (1965): Fossils, early life, and atmospheric history. - Proc. Nat. Acad. Sci. USA, 53, 1205-1215, Washington
FLus, G. & Huss~g, H. (1993): A Microbial Model for the Lower Devonian Stromatactis Mud Mounds of the Montagne Noire (France). - Facies, 29, 179-194, Erlangen
FL0on. (1994): Pangean shelf carbonates: Controls andpaleoclimatic

279
signifiance of Permian andTriassic reefs . - Geol. Soc. Amer. Spec. Paper, 288, 247-266, Boulder
FREaWALD, A., HE~CH, R. & PXTZOLD, J. (1997) Anatomy of a deep-water coral reef mound from S tjemsund, West-Finnmark, Northern Norway. In: JAMES, N.P. & CLARKE, J. (eds.): Cool- water carbonates.- Soc. Econ. Paleont. Min., Spec. Publ, 56, 141-162
FRFaWALD, A. & SCH6NFELD, J. (1996): Substrate pitting and boring pattern of Hyrrokkin sarcophaga CEDHAGEN 1994 (Foraminiferida) in a modern deep-water coral reef mound. - Marine Micropaleontology, 28, 199-207
FR~WALD, A. & WmSON, J. B. (in press): Taphonomy of modern deep, cold-temperate-water coral reefs. - Historical Biology
FREYTET, P . & PLAZtAT, J.-C. (1982): Continental Carbonate Sedimentation and Pedogenesis - Late Cretaceous to Early Tertiary of Southern France. - Contr. Sediment., 12:pp.213 pp., Stuttgart (E. Schweizerbarth).
FREYTET, P. & PLET, A. (1996): Modern Freshwater Microbial Carbonates : ThePhormidium Stromatolites (Tufa-Travertine) of Southeastern Burgundy (Paris Basin, France). - Facies, 34, 219-238, Erlangen.
FRITZ, G.K. (1958): Schwammstotzen, Tuberoide und Schutt- breecien im WeiBen Jura der Schwtibischen Alb.-Arb. Geol. Paltiont. Inst. TH Stuttgart, NF, 13, 1-118, Stuttgart.
GALL, J.C. (1995): B iofilms et mattes microbiennes: leur contri- bution h la s~dimentogen~se. - C.R. Acad. Sci. Paris, 321, s6rie Ha, 823-835, Paris
GARC~A-VALL~.S, M., GONZALEz-PRADO, J. & VENDRELL SAZ, M. (1993): Caracterizaci6n de los lignitos garumnienses del Pireneao Central catahln. -Cuad. Geol. Ib&ica, 17, 85-107, Madrid
GARRET"r, P. (1970a): Deposit feeders limit development of stromatolites. - Amer. Ass. Petrol. Geol. Annual Conven- tion, Calgary, June 22-24, 1970, Abstracts p. 848.
- - (1970b): Phanerozoic stromatolites: noncompeteive ecologic restriction by grazing and burrowing animals.- Science 169, 171-173, Washington
GErrLER, L. (1932): Cyanophyceae. In: Rabenhorst's Kryptogamen- Flora yon Deutschland, Osterreich und der Schweiz, 2 nd ed., 14, 1196 pp., Leipzig.
GLAUB, I. (1994): Mikrobohrspuren in ausgewtihlten Ablagerungs- raumen des europ~iischen Jura und der Unterkreide. - Courier Forschungsinstitut Senckenberg, 174, 1-289, Frankfurt a.M.
GOURLAY, M.R. & JELL, J.S. (1993): Heron Island spoil dump. - Res. Publ. Great Barrier Reef Marine Park Authority, 28,221 pp., Townsville
GRIMALT,J.O., TORRAS, E. & ALBAIGflS, J. (1987): Bacterial re- working of sedimentary lipids during sampling storage. In: MATTAVELU, L. & NOVELLI, L. (eds.): Advances in Organic Geochemistry 1987. - Org. Geochem., 13,741-746, Oxford
GROTZaNGER, J.P. (1990): Geochemical model for Proterozoic stromatolite decline. - Amer. J. Science, 290-A, 80-103, Washington
GUHA, P.K.S., MUKHOPADHYAY, G. & DAS, R.N. (1996) : Trace fossils as indicator of palaeoenvironment of Talchir Forma- tion in Raniganj and Deogarh Group of coalfields, India. - Gondwana Nine, 1,491-504, Geol. Surv. India, Calcutta.
GULIK, T. (1991): CoAgUlation des ~chantillons biologiques. In: MOREl., G. (ed.): Microscopic ~lectronique. Cryom~thodes, immunocytologie, autoradiographie, hybridation in situ. - Editions INSERM, 5-20, Paris.
GWYNN, J.W. & MURPHY, P.J. (1980): Recent sediments of the Great Salt Lake basin. In: GWYNN, J.W. (ed.): Great Salt Lake. A scientific, historical and economic overview. - Utah Geol. Min. Surv., Bull., 116, 83-96, Salt Lake City.
HALLAM, A. (1994): The earliest Triassic as an anoxic event, and its relationship to the end-Palaeozoic mass extinction.- Can. Soc. Petrol. Geol. Mem, 17, 797-804, Calgary
HALLOCK, P. & SCItLAGER, W. (1986): Nutrient excess and the demise of coral reefs and carbonate platform. - Palaios, 1,
389-398, Tulsa HAN, J. & CALVIN, M. (1969): Hydrocarbon distribution of algae
and bacteria and microbial activity in sediments. - Proc. Nat. Acad. Set. USA, 64, 436-443, Washington
HEF'IER, J., TmEL, V., JEN'ISCH, A., GALLING, U., KEMPE, S. & MICHAELIS, W. (1993): Biomarker indications for microbial contribution to recent and late Jurassic carbonate deposits. - Facies, 29, 93-106, Erlangen.
HENNEBERT, M. & LEES, A. (1991): Environmental gradients in carbonate sediments and rock detected by correspondence analysis: examples fron theRecent of Norway and the Dinantian of southwest England. - Sedimentology, 38, 623-642, Ox- ford
HENRICH, R., HARTMANN, M., REITNER, J., SCH~,FER, P., FREIWALD, A., STEINMETZ, S., DIETRICH, P. & THmDE, J. (1992): Facies belts and communities of the Arctic Vesterisbanken Seamount (Central Greenland Sea) . - Facies, 27, 71-104, Erlangen
HERCZEG, A.L. & FAmBANKS, R.G. (1987): Anomalous carbon isotope fractionation between atmospheric CO 2 and dis- solved inorganic carbon induced by intense photosynthesis. - Geochim. Cosmochim. Acta, 51,895-899, Amsterdam.
HooK, J.E., GOLUBIC, S. & MILLIMAN, J.D. (1984): Micritic ce- ment in microborings is not necessarily a shallow-water indicator. - J. Sediment. Petrol., 54, 425-431, Tulsa
HORODYSKL R.J. (1976): S tromatolites in the Upper Siyeh Lime- stone (Middle Proterozoic), Belt Supergroup, Glacier Na- tional Park, Montana. - Precambrian Res. 3, 517-536, Am- sterdam
HUMM, H. J. & WICKS, S. R. (1980): Introduction and guide to the marine Bluegreen Algae. - p.194, New York (Wiley)
IRANIAN-JAPANESE RESEAROt GROUP (1981): The Permian and the Lower Triassic Systems in Abadeh Region, Central Iran. - Memoirs of the Faculty of Science, Kyoto University, Series of Geology & Mineralogy, XLVII, 61-133, Kyoto
JACKSON, L.L. & BLOMQUIST, G.J. (1976): Insect waxes. In: KOLA~I"UKUDY, P.E. (ed.): Chemistry and biochemistry of natural waxes, 201-229, Amsterdam (Elsevier).
JAHNKE, H. & RrrzxowsKr, S. (1980): Die Fazies-Abfolge im Miinder Mergel der Steinbr(iche bet Thi.iste (Ober-Jura, Hilsmulde). - Ber. naturhist. Ges. Hannover, 123, 45-62, Harmover
JAMES, N.P. & BOURQUE, P.-A. (1992): Reefs and Mounds. In WALKER, R.G. & JAMES, N.P. (eds.): Facies Models - Re- sponse to Sea-Level Change. - Geol. Assoc. Canada, 323- 347, St. John's
JAMEs, N. P. & G~SBURG, R. N. (1979): The seaward margin of Belize Barrier and Atoll reefs: morphology sedimentology, organisms distribution and late Quaternary history. - I.A.S., Spec. Publ., 3, pp.191, Oxford
JAMES, N.P. & GRAVESTOCK, D.I. (1990): Lower Cambrian shelf and shelf margin buildups, Flinders Range, South Australia. - Sedimentology, 37, 455-480, Oxford
JONES, B. (1987): The alteration of sparry calcite crystals in a vadose setting, Grand Cayman Island. - Can. J. Earth Sci., 24, 2292-2304
JONES, B. & HUNTER, I. G. (1991 ): Corals torhodoliths to microbialites - a community replacement sequence indicative of regressive conditions. - Palaios, 6, 54-66, Tulsa
J~ROENSEN, B.B. (1983): Themicrobial sulphur cycle. In: KRUMBEXN, W.E. (ed.): Microbial Geochemistry, 329, 91-124, Oxford.
KAItLE, C.F. (1974): Ooids from Great Salt Lake, Utah, as an analogue for the genesis and diagenesis of ooids in marine limestones. - J. Sediment. Petrol., 44, 30-39, Tulsa.
KALKOWSKY, E. (1908): Oolith und Stromatolith irn norddeutschen Buntsandstein. - Z . deutsch, geol. Ges., 60, 68-125, Berlin
KARHU, J.A. &EPSTEtN, S. (1986): The implication of the oxygen isotope records in coexisting cherts and phosphates.-Geochim. Cosmochim. Acta, 50, 1745-1756, Amsterdam
KARttU, J.A. & HOLLAND, H.D. (1996): Carbon isotopes and the rise of atmospheric oxygen. - Geology, 24, 867-870, Boulder

280
K^STINO, J.F. (1987): Theoretical consta-aints on oxygen and carbon dioxide concentrations in the Precambrian atmos- phere. - Precambrian Res., 34: 205-229, Amsterdam
IO.~o, F., SINNINOHE-DAMsTE, J.S., KOC~-W~N D~,~,A.C., Rm~sw~ W.I.C., Huc, A.Y. & DE l.~EtrW, J.W. (1995): Occurrence and origin of mono-, di-, and trimethylalkanes in modern and Holocene cyanobacterial mats from Abu Dhabi, United Arab Emirates. - Geochim. Cosmochim Acta, ~9, 2999-3015, Oxford
K~RANS, C. (1985): Petrology of Devonian and Carboniferous carbonates of the Canning and Bonaparte Basins. - Western Austral. Min. Petrol. Res. Inst., Report 12, p.203, Perth
K~uPe, H., KocH, R. & ~ D E ~ , R.R. (1990): Steuerungsprozesse der Entwicklung yon Oberjura-spongiolitben S~ddeutschlands: Kermmisstand, Probleme und Perspektiven. - Facies, 23, 141-174, Erlangen.
Kn~sE't D.W. (1985): Metabolisms, calcification and carbon production. I. Systems level studies: Proceeding of the 5th International Coral Reef Congress, Tahiti, 1985, 4, 505-526.
KOM/~RZK, J. & ANAONOST~D~S, K. (1986): Modem approach to the classification system of eyanophytes. 2 - Chroococcales. - Arch. Hydrobiol. Suppl., 73 (Algological Studies 43), 157- 226, Stuttgart.
KOTLY~'a~, G.V., ZH~a~OV, Y.D., Kocz~w~cT_, G .S., KROPACrmV~, G.S., ROSTOVCEV, K.O., CHEDJ~, L., VUKS, G.P. & GUs~v^, E.A. (1983): Evolution of the latest Permian biota, Djulfian and Dorashamian regional stages in the URSS. - Leningrad Dep. Pub. House, Nauka, pp.200, Leningrad
KRU~UE~,~, W. E. (1983): Stromatolites - The challenge of a term in space and time. - Precambrian Res., 20,493-531, Amster- dam
-- (1986): Biotranfer of minerals by microbs and microbial mats. - I n : LEAD~E^TER, B.S.C. & R~DIN~, R. (eds.): Biomineralization in Lower Plants and animals. - T h e Syst. Assoc. Spec., 30, 55-72, Oxford (Clarendon Press)
KRUMB~.n~, W. E., COHF~,Y. & SH;LO, M. (1977): Solar Lake (Sinai). 4. Stromatolitic cyanobacterial mats. - Limnol. Oceanogr., 22, 635-656.
LAND, L.S. & GOREAU, T.F. (1970): Submarine lithification of Jamaican reefs. - J. Sediment. Petrol., 40, 457-462, Tulsa
LAND, L.S. & MOORE, C.H., Jr. (1980): Lithification, mieritization and syndepositional diagenesis o fbiolithites on the Jamaican island slope. - J. Sediment. Petrol., $0, 357-370, Tulsa
LAZAR, B. & ER~Z, J. (1990): Extreme x3C depletions in seawater- derived brines and their implications for the past geochemical carbon cyc l e . - Geology, 18, 1191-1194, Boulder
LAZAR, B., JAVOR, B., ER~z, J. (1989): Total alkalinity in marine- derived brinesand pore waters associated with microbial mats. In: COHE~ Y. & ROSEN~E~O, E. (eds.): Microbial Mats, physiological ecology of benthic microbial communities, 84- 93, Washington
L~ C,~'~v~oN-ALst~O, T., GOLUB~C, S. & P~mss, K. (1995): Fungi in corals: symbiosis or disease? Interaction between polyps and fungi causes pearl-like skeleton biomineralization. - Marine Ecology Progress Series, 117, 137-147
LEEDE~, M. (1973): Lower Carboniferous serpulid patch reefs, bioherms and biostromes. - Nature, 242, 41-42, London.
-- (1975): Lower Border Group (Toumaisian) stromatolites from the Northumberland basin. - Scott. J. Geol., II, 207- 226, Glasgow
LEES, A. (1964): The Structure and Origin of the Waulsortian (Lower Carboniferous ) 'reefs' of West-Central E i r e . - Phil. Trans. Roy. Soe.London, ser. B, 247, 483-531, London
-- (1988): Waulsortian 'reefs': the history of a concept. - Inst. G6ol. Univ. Louvain Memoir, 34, 43-55, Louvain-la-Neuve
l_s A., HALLET, V. & Hmo, D. (1985): Facies variation in Waulsortian buildups, Part 1; a model from Belgium.- Geol. J., 20, 133-158, Chichester
LEES, A. & MmLER, J. (1985): Facies variation in Waulsortian buildups, Part 2; Mid-Dinantian buildups from Europe and
North America. - Geol. J., 20, 159-180, Chichester -- & -- (1995): Waulsortian Banks. - In : MONTY, C., BOSEr~CE, D.,
BPaDOES, P. & I~^TT, B. (eds): Mudmotmds: their Origin and Evolution.- I.A.S. Spec. Publ., 23,191-271, Oxford (Blackwell)
l.s R.A, & LotmsB~Ry, D.M. (1969): Isolation, cultivation and characteristics of flexibacteria. - J. Gen. Microbiol., 58,145-170
I.,EYM~m, A. (1862): Compte-rendu de la r~union de la Soc. g~ol. de France h Saint-Gaudens. - Bull. Soe. g~ol. France, 2/19, 1089-I 163, Paris
l_aNom'~rB~o~, M. (1993): Poriferenskleren (Demospongea und Calcarea) in subrezenten S edimenten aus dem Ktlstenbereich von Banyuls-sur-Mer (Mittelmeer): Bestandsaufnahme und Vergleich mit der rezenten Poriferenfauna. - Unpubl. Di- ploma Thesis, Geowiss. Fac., 58 p., FU-Berlin
La~Er~, R. & N~aCK~N, O. (1969): On the hydrography of the shelf waters offM~re and Helgeland.- Fiskeri Direktoratets Skrifter Serie Hay Unders/,gelse, 15, 285-294
I.,LO~',~a~T,C, & KO.AOss, S. (1982): Restos demoluscosy dinosaurios en formaciones estromatolfticas garunmienses al S del Montsec (Prov. de l.Arida). - Bol. Geol. Min., XCIII/V, 371-378, Madrid.
Loo~a~, B.W. (1961): Cryptozoan and associated stromatolites from the Recent, Shark Bay, Western Australia. - J. Geol., 69, 517-533, Chicago
LOREAtr, J.-P. (1969): Ultrastructures et diageneses des oolithes marines anciermes (jurasiques).- C.R. Acad. Sci. Paris., 269, 819-822, Paris
LUTERaACHER, H.-P. (1969): Remarques sur la position strati- graphique de la Formation d'Ager (Pyrenees M~ridionales). Coll. Sur l'Eocene, Paris. -Bur. Reeherches G~ol. et Mini&es, 69, 225-232; Paris
M A ~ R E , I.G. & M ARSH~.L, J.F. (1988): Submarine lithification in coral reefs: some facts and misconceptions. - Proc. 6th Intemat. Coral Reef Symp., Australia, 1988, 1, 263-272, Townsville
M A ~ E , I., REro, R.P. & STEr~CK, R. (1996): Growth history of stromatolites in a holocene fringing reef, Stocking Island, Bahamas. - J. Sediment. Res., 66, 231-242, Tulsa
M~,cI~R, H. (I 996): S tratigraphie und Mikrofazies des [3berg angs Kreide- Terti~r (Garumnium) im Tal yon Ariet am st~d6sflichen Rand des ~kger-Beckens unter besonderer Berficksichtigung der Oberen Terrigenen Sequenz. - Diploma Thesis Univ. Frankfurt, 119 pp. (unpublished)
MAre, A., BOURQUE, P.-A., & M^MET, B.L. (1996): Depth-related Ecological Zonation of a Carboniferous Carbonate Ramp: Upper Vis~an of B6char Basin, Western Algeria. - Facies, 35, 59-80, Erlangen
M ~aLLe, F. (1984): C ontribu tion ~ la connalssance des m~canismes biog~ochimiques darts deux milieux de s&timentafion r~cifaux. - Rapport de Dipl6me d'Etudes Approfondies, Univ. Orleans, 60 pp. (unpublished)
M~rz, W., AMJ~m, R., LoDwto, W., VANCANNEYT, M. & Scrn~F~, K.-H. (1996): Application of a suite of 165 rRNA-specific oligonucleotide probes designed to investigate bacteria of the phylum cytophaga-flavobacter-bacteroides in the natural en- vironment. - Microbiology, 142, 1097-1106
M ~ z , W., AM^NN, R.I., LUDWIG, W., W^GNER, M. & SCnL~'F~, K.-H. (1992): Pbylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: problems and solu- tions. - Syst. Appl. Microbiol., 15, 593-600
M~cotm, J. & BAUD, A. (1986): The Permo-Triassic boundary in the Antalya nappes (Western Taurides, Turkey). Mem. Soc. Geologica It.al, 34, 243-252, Roma
M~a~cou'x, J., B^UD, A., KReSTYN, L. & MONOD, O. (1986): Field Workshop 'Late Permian and Triassic in Western Turkey'. - Guide Book part 2, Western Tauride (Antalya-Seydisehir- Isparta-Antalya). 65 pp., Istanbul Technical University and Subeommission on Triassic Stratigraphy
M~sr~LL, J. F. (1983): Submarine cementation in a higher-

281
energy platform reef: One Tree Reef, southern Great Barrier Reef. - J. Sediment. Petrol., 53, 1133-1149, Tulsa
MasLov, V.P. (1959): Stromatolites and facies. Dokl ady Akademia Nauk SSSR 125, 1085-1088, Moskva (in Russian)
MAXWELL, W.G.H. (1962): Lithification of carbonate sediments in the Heron Island Reef, Great Barrier Reef. - J. Geol. Soc. Austral., 8, 217-238, Adelaide
MEDLIN, L., ELWOOD, H.J., STICrmL, S. & SoGm, M.L. (1988): The characterization of enzymatically amplified eukaryotic 16S- like RNA-coding regions. - Gene, 71,491-499
MERZ, M. (1992): The biology of carbonate precipitation by cyanobacteria. - Facies, 26, 81-102, Erlangen.
MEY, P.H.W., NAGTEGAAL, P.J.C., ROBERTI, K.J. & HARTEVELT, J.J.A. (1968): Lithostratigraphic Subdivision of Post-Herzynian Deposits in the South-Central Pyrenees, Spain. - Leidse Geol. Meded., 41,221-228, Leiden
Mn.Ln~., D., PmRL, H. &. HURLEY, J. (1993): Microalgal pigment assessments using high performance liquid chromatography: a synopsis of organismal and ecological applications. -Can. J. Fish. Aquat. Sci., 50, 2513-2527
MIT~RER, R.M. & CtmmNOtAra, R. (1985): The interaction of natural organic matter with grain surfaces: implications for calcium carbonate precipitation. In: Schneidermann, N. & Harris, P.M. (eds.): Carbonate Cements. - Soc. Econ. Paleont. Miner., Spec. Publ, 36:117-31, Tulsa
MOr~'rAGGtONt, L.F. & C ~ o m , G.F. (1993): Stromatolites associ- ated with coralgal communities in Holocene high-energy reefs. -Geology, 21, 149-152, Boulder
MONTY, C.L.V. (1973): Precambrian background and Phanerozoic history of stromatolitic communities, an overview.- Annales Soc. Gdol. Belgique 96, 585-624, Bruxelles
-- (1995): The rise and nature of carbonate mud-mounds: an introductory actualistic approach.- In: MONTY, C.; BOSENCE, D., BmD~ES, P. & PRATT, B. (eds): Mudmounds: their Origin and Evolution. - I.A.S. Spec. Publ., 23, 11-48, Oxford (Blackwell)
MORTENSEN, P.B., HOVLAND, M., BRATTEOARD, T. & FARESTVEIT, R. (1995): Deep water bioherms of the scleractinian coral Lophelia pertusa (L.) at 64~ on the Norwegian shelf: structure and associated megafauna. - Sarsia, 80, 145-158
MOSNIER-ARNOU, M. & TRIO-lET, J. (1979): Premiers r6sultats d'observ ations sur la structure de s6diments algaires richement organiques darts lesquels pr6cipitent des carbonates de cal- cium. - Rapport interne Compagnie Franqaise des P&roles- Total, 7 p., (unpublished)
MOt/N/I, D, BOURQUE, P.-A. & SAVED, M. (1996): Architecture and isotopic constraints on origin of Lower Devonian conical mounds (Kess-Kess) of Tafilalt, Anti-Atlas, Morocco. - I.A.S., 17tb African-European Meeting, Sfax, Tunisia, Ab- stracts, 192-193
MOUSSL~E-PoucHrarm, A. & BER'mAND-S,~,FA'n, J. (1997):Tectono- sedimentary subdivisions in the Neoproterozoic Cover of the Taoudenni Basin (Algeria-Mauritania-Mali). - J. African Earth Sci., (in press).
MUK~OPAm~YAY, G. (1996) : Facies analysis o fTalchir Sediments (Permo-Carboniferous), Dudhi Nala, Bihar, India - A Glaciomarine model. - Gondwana Nine, 2, 737-753, Geol. Surv. India, Calcutta.
Mt.rt.LtNS, H.T. & H~E, A.C. (1989): Scalloped bank margins: beginning of the end for a carbonate platform?- Geology, 17, 30-33, Boulder
MUSCA'r~NE et al. (1981): Estimating the daily contribution of carbon from zooxanthellae to coral animal respiration. - Limonl. Oceanogr., 26, 601-611
MUSCATINE, L., McCLOsKEY, L.R., & Low,, Y. (1985): A com- parison of the growth rates of zooaxanthellae and animal tissue in the Red Sea coral Stylophora pistillata: Proceeding of the 5th International Coral Reef Congress, Tahiti, 1985, 6, 119-123
NETER, J., WASSERMAN, W. & KUTNER, M. (1985): Applied linear
statistical models. -832 pp., Homewood, Illinois (Irwin) N E A R , F. (1993): Development of Albian mierobialites and
microbialite reefs at marginal platform areas of the Vasco- Cantabrian Basin (Soba Reef Area, Cantabria, N.Spain.). - Facies, 29, 231-250, Erlangen
-- (1995): Dynamische Sedimentationsvorg~ge, Diagenese und Biofazies unterkretazischer Plattformriinder (Apt/Alb; Soba- Region, Prov. Cantabria, N-Spanien). - Berliner geowiss. Abh., E, 17, 235 pp., Berlin
NEUWE~R F., RErm~, J. & ARP, G. (1996): Controlling Factors and Environmental significance of Organomicrite Produc- tion and Buildup Development. In: RErrN~, J., NEUWEUJ~R, F. & Gtnqr, EL, F. (eds.): Global and Regional Controls on Biogenic Sedimentation, I. Reef Evolution. Research Reports.- G6ttinger Arb.Geol.Pal~iont., SB2, 185-192, G6ttingen
NICKEL, E. (1983): Environmental Significance of Freshwater Oncoids, Eocene Guarga Formation, Southern Pyrenees, Spain. In: PERY'r, T.M. (ed.): Coated Grains, 308-329, Berlin (Springer).
PACKER, B.M. (1990): Sedimentology, paleontology, and stable- isotope geochemistry of selected formations in the 2.7- billion-year-old Fortescue Group, Western Australia. - Un- published Ph.D. Thesis, UCLA, 187 pp.
P~RL, H.& P~cr,.r~v, J. (1996): A mini-review of micmbiai consortia: their roles in aquatic production and biogeochemical cycling. - Microb. Ecol., 31,225-247
P~a~OYA, K.L. (1987): Some unusual structures in the Talchir Group, Talchir Gondwana Basin, Orissa. - J. Geol. Soc. India, 30, 305-308, Bangalore.
PARK, R. (1976): A note on the significance of laminations in stromatolites- Sedimentology, 23, 379-393, Oxford
PASCAL, A. (1985): Les syst~mes bios~dimentaires urgoniens (Aptien-Albien) stir la marge Nord lb&ique. - M~m. G~ol. Univ. Dijon, 10, 569 pp., Dijon
PAUL, J. & PERYT, T.M. (1985): Oolite und Stromatolithen im Unteren Buntsandstein des Heeseberges bei Jerxheim, Kreis Wolfenbtittet. - Ber. naturhist.Ges. Harmover, 126, 175-186. Hannover
PEDLEY, H.M. (1994): Prokaryote - Microphyte Biofilms and Tufas : A Sedimentological Perspective. - Kaupia, 4, 45-60, Darmstadt
PEDONE, V. & FOLK, R.L. (1996): Formation of aragonite cement by nannobacteria in the Great Salt Lake, Utah. - Geology, 24, 763-765, Boulder
PERK,S, R.D. & HALSEY, S.D. (1971): Geologic significance of microboring fungi and algae in Carolina shelf sediments. - J. Sediment. Petrol., 41,843-853, Tulsa
PERYT, T.M. (1974): Spirorbid-algal stromatolites.- Nature, 249, 239-240, London.
PICARO, M.D. & HmH, L.R., Jr. (1972): Criteria for recognizing lacustrine rocks. In: RmBY J.K.& H~a~Bt.m W.K. (eds.): Recognition of Ancient Sedimentary Environments. Soc. Econ. Paleont. Min., Spec. Publ, 16, 108-145, Tulsa
PICK~a~D, N.A.H. (1996): Evidence for microbial influence on the development of Lower Carboniferous buildups. In: Sa~OOEN, P., SOMMERVn.LE, I.D. & JoinT, S, G.L. (eds.): Recent Advances in Lower Carboniferous Geology. - Geol. Soc. Spec. Publ, 107, 65-82
PtYC~rEY, J., Mint.m, D., HowE, K., PAERL, H. & HURLEY, J. (1996): Flow scintillation counting of 14C-labeled microalgal photosynthetic pigments. - J. Plank. Res., 18, (in press)
P m c ~ Y , J., P~a~RL,H. & BF.Botrr, B. (1995a): Salinity control of benthic microbial mat community production in a B ahamian hypersaline lagoon. - J. Exp. Mar. Biol. Ecol., 187, 223-237
P~cr, r~v, J., P~RL, H. & Fla-Ta, a'mmK, M. (1995c): Impacts of seasonality and nutrients on microbial mat community struc- ture and function. - Mar. Ecol. Prog. Ser., 123, 207-216.
Pmcm',mv, J., PmRL, H., REIn, R.P. & BnBotrr, B. (1995b): Ecophysiology of stromatolitic microbial mats, Stocking Island, Exuma Cays, Bahamas. - Microb. Ecol., 29, 19-37.

282
PINCKNEY, J., PAPA, R. & Z~GMARK, R. (1994): Comparison of high performance liquid chromatographic, spectrophotometric, and fluorometric methods for determining chlorophyll a concentrations in estuarine sediments. - J. Microbiol. Meth- ods, 19, 59-66
PRA1"r, B.R. (1982): Stromatolite decline - a reconsideration. - Geology, 10, 512-515, Boulder
-- (1992): Shrinkage features ('molar tooth' structures) in Proterozoic limestones- New model for their origin through synsedimentary earth-quake induced dewatering. - Geol. Soc. Amer., Abstr., 1490
-- (1997): The origin of 'molar tooth' structures in Proterozoic carbonate rocks by synsedimentary earthquake induced shrink- age and fluidization, and implications for the nature and evolution of basin and marine sediments. -Geol . Soc. Amer., (in press)
RAGHUKUMAR, C. & RAGHUKUMAR, S. (1991): Fungal invasion of massive cora ls . - P.S.Z.N.I.: Marine Ecology, 12, 251-260
RAGIIUKUMAR, S. & BALSUBRAMANIAN, R. (1991): Occurrence of thraustochytrid fungi in corals and coral mucus. - Indian J.Marine Sciences, 20, 176-181
RASMUSSEN, K.A., MACINTYRE, I.G. & PRUFERT, L. (1993): Mod- em stromatolite reefs fringing a brackish coastline, Chetumal Bay, Belize. - Geology, 21, 199-202, Boulder
REIn, R.P. & BROWNE, K.M. (1991): Intertidal stromatolites in a fringing Holocene reef complex in the Bahamas. -Geology, 19, 15-18, Boulder
REID, R.P., MACINTYRE, I., BROWNE, K., S ~TENECK, R., & MILLER, T. (1995): Stromatolites in the Exuma Cays, Bahamas: Uncom- monly common. - Facies, 33, 1-18; Erlangen
RErrNr~ J. (1993): Modern cryptic microbialite/metazoan facies from Lizard Island (Great Barrier Reef, Australia) - Forma- tion and concepts. - Facies, 29, 3-40, Erlangen
RErrN~R, J., GAU~ET, P., MARtN, F. & NEtrW~a.ER, F. (1995): Automicrites in a modern marine microbialite. Formation model via organic matrices (Lizard Island, Great Barrier reef, Australia). - Bull. Inst. oc6anograph. Monaco, spec. 14/2, 237-263, Monaco
REITNER, J & NEt/WEILER, F. (coord.) (1995): Mud Mounds: A polygenetic spectrum of fine-grained carbonate buildups. - Facies, 32, 1-70, Erlangen
RErrN~, J., NEUWEn~R, F. & GAtrmET, P. (1995): Modern and fossil automicrites: implications for mud mound genesis. - In: RErm~, J. & NEUWEa.ER, F. (coord.): Mud Mounds: A Polygenetic Spectrum of Fine-grained Carbonate Buildups.
- Facies, 32, 4-17, Erlangen RErmF.R, J., WU.MS~, M. & NEVWEn~R, F. (1995): Cenomaniard
Turonian sponge microbialite deep-water hardground com- munity (Liencres, Northern Spa in) . - Facies, 32, 203-212, Erlangen
R~TN~, J., WORHmOE, G., Tram., V. & GAIJTRET, P. (1996): Reef Caves and cryptic habitats o f Indo-Pacific reefs- Distribution patterns of coralline sponges and microbialites. In: REIa~mR J., NEtrW~V,.ER, F. & G t r ~ L , F. (eds.): Global and Regional Controls on Biogenic Sedimentation, I. Reef Evolution. Research Reports. - G6ttinger Arb.Geol.Palaont., SB2, 91- 100, GOttingen
RIDING, R. (1989): Calcified cyanobacteria in Phanerozoic reefs. - Algae in Reefs Symposium, Granada 1989, Abstracts: 3-4
RIDING R. (1991): Classification of microbial carbonates.- In: RIDn~G, R. (ed.): Calcareous Algae and stromatolites, 2 . - 1- 51, Berlin (Springer)
-- (1992): Temporal variation in calcification in marine cyanobacteria. - J. Geol. Soc., London 149, 979-989.
-- (1993): Phanerozoic patterns of marine CaCO 3 precipitation. - Naturwissenschaften, g0, 513-516.
-- (1994): Strornatolite survival and change: the significance of Shark Bay and Lee Stocking Island subtidal columns. In: KR UMBEIN, W., PAIERSON, D. & STAL, L. (eds.): Biostabilization of Sediments, 183 -202, Oldenburg, Germany (Univ. Oldenburg)
-- (1994a): Microbial calcification in time and space. - Death Valley Internat. Stromatolite Symposium, Laughlin, Ne- vada, 15-17 October 1994, Abstracts, 65-66.
-- (1994b): Evolution of algal and cyanobacterial calcification. In: BENGTSON, S. (ed.): Early life on Earth. -Nobe l Sympo- sium 84, 426-438 (Columbia University Press, New York)
-- (1995): Microbes and the rise of Metazoa: a reassessment. - Sardinia 95, 6th Paleobenthos Internat. Symposium, Sar- dinia, Italy, 25-31 October 1995, Abstract-book, 46-47.
-- (1996): Stromatolite decline: reassessment of the concept of metazoan competition.-Palaeontological Association, 40th Armual Meeting, Birmingham, England, 16-19 December 1996, Abstracts, xxv-xxvi.
RIDmG, R., AWRAMIK, S.M., W~SBOROUGH, B.M., G R I N , K.M. & DmL, R. F. (1991a): B ahamian giant stromatolites: micro- bial composition of surface mats. - Geol. Mag., 128, 227- 234, Cambridge
RIDING, R.,MARTty, J.M. & BRAGA, J.C. (199 lb): Coral-stromatolite reef framework, Upper Miocene, Almeria, Spain. - Sedimentology, 38, 799-818, Oxford
RID~G, R. & ZHURAVLEV, A.Yu. (1994): Cambrian reef builders: calcimicrobes and archaeocyaths. - Ecological Aspects of the Cambrian radiation. IGCP Project 366, Cambridge, Eng- land, 12-15 December 1994. - Terra Nova, Abstract Supple- ment 3, p. 7
ROSTOVTSEV, K.O. & AZARIA~, N.R. (1973): The Permian-Triassic Boundary in Transcaucasia. In: The Permian and Triassic Systems and their mutual Boundary. Mem. 2, 89-99, Calgary
RoucHY, J.M. & MoN'rr C. (1981): SLromatolites and cryptalgal laminites associated with Messinian gypsum of Cyprus. - In: MONTY, C. (ed.): Phanerozoic Stromatol i tes . - 155-180, Heidelberg (Springer)
RowAN, K. (1989): Photosynthetic Pigments of Algae. 382 pp., New York (Cambridge Univ. Press)
SJ~'~DBERG, P.A. (1975): New interpretations of Great Salt Lake ooids and of ancient non-skeletal carbonate mineralogy. - Sedimentology, 22, 497-537, London
SASTAVY, D.L., WILLErCZ, P.& COLWELL, R.R. (1990): Phenotypic study of bacteria associated with the carribean sclerosponge, Ceratoporella nicholsoni.-Appl. Environ. Microbiol., $6,1750- 1762
SCHLAGER, W. (1981 ): The paradox of drowned reefs and carbon- ate platforms. -Geol . Soc. Amer. Bull., 92, 197-211, Boulder
-- (1989): Drowning unconformities on carbonate platforms.- In: CREVELLO, P.D., WmSON, J.L., SARO, J.F. & READ, J.F. (eds.): Controls on carbonate platform and basin develop- ment. - Soc. Econ. Paleont. Miner., Spec. Publ, 44, 15-25, Tulsa
SCHLAGER, W & PmLW, J. (1990): Cretaceous carbonate plat- forms. - In: GINSBLrRG, R.N. & BEAtmO~, B. (eds.): Creta- ceous Resources, Events and Rhythms. - NATO ASI Series, Series C, 304, 173-195, Dordrecht (Kluwer Acad. Publ)
SCHNEIDERMANN, N.& HARRIS, P.M. (1985, eds.): Carbonate Ce- ments. - Soc. Econ. Paleont. Miner., Spec. Publ., 36; Tulsa
SCHUa~T, J.K. & Bo'rrmR, D.J. (1992): Early Triassic stromatolites as post-mass extinction disaster forms. - Geology, 20, 883- 886, Boulder
-- & -- (1995): Aftermath of the Permian-Triassic mass extinction event: Paleoecology of the Lower Triassic carbonates in the western USA. - Palaeogeogr., Palaeoclimat.,Palaeoecol., 116, 1-39, Amsterdam
SCHULXT~-LAM, S. THOMSON, J.B. & BEVERttmE, T.J. (1993): Metal ion immobilization by bacterial surfaces in freshwater environments.- Water Poll. Res. J. Canada, 28/1, 51-81
SCHULZ, E. (1967): Sporenpal~iontologische Untersuchungen rritoliassischer Schichten im Zentralteil des Germanischen Beckens. - Pal~iont. Abh., B, 2, 541-633, Berlin
ScorN, T.P., ALEXANDERSSON, E.T., BOWF.S, G.E., CLO~E, J.J., FARROW, G.E. & MILL/MAN, J.D. (1980): Recent, temperate, sub-photic, carbonate sedimentation: Rockall Bank, north-

283
east Atlantic. - J. Sediment. Petrol., 50, 331-356, Tulsa SE~KHATOV, M.A. & RAABE~, M.E. (1993): Dynamics of the
taxonomic diversity of Riphean and Vendian stromatolites in northern Eurasia. - Stratigraphy and Geological Correlation 1 (2): 133-141
Sm~, J., BRASSELL, S. C. & WARD, D. M. (1990): Mid-chain branched mono- and dimethyl alkane in hot spring cyanobacterial mats: A direct biogenic source for branched alkanes in ancient sediments?-Org. Geochem., 15, 223-231, Oxford
SEDLECKA, A. (1978): Late Precambrian Tidal-fiat deposits and algal stromatolites in the Batsfjord Formation, East Finnmark, North Norway. - Precambrian. Res., 21, 277-310, Amster- dam
SMITH, A.G. (1968): The origin and deformation of some 'molar tooth' structures in the Precambrian Belt Supergroup. - J. Geol., 16, 426-443, Chicago
SPENCF~ DAVIES, P. (1984): The role of zooxanthellae in the nutritional energy requirements of Porcillopora eydouxi - Coral Reefs, 2, 181-186, Berlin
SOUmEs, S.W. & KAS'r~G, J.F. (1994): Early Mars: How warm and how wet? - Science, 265,744-749, Washington
STAHL, D.A. & AMm~rN, R.L (1991): Development and application of nucleic acid probes. In: STACKEBRANDT, E. & GOODFELLOW, M. (eds.): Nucleic acid techniques in bacterial systematics, 205-248, Chichester (Wiley)
STAL, L. (1995): Physiological ecology of cyanobacteria in mi- crobial mats and other communities. - New Phytol., 131, 1- 32
ST~ANI M. & TaO~iBETrA, P. (1989): Le successioni retiche della Toscana orientale e dell'Umbria: cicli sedimentari asimmetrici in un bacino marnoso-calcareo dominato dalle tempeste. - Boll. Soc. Geol. It., 108, 591-606, Roma
S'm'rrza, K.O., K~t~, H. & STACV~Ba,~T, E. (1983): Pyrodictium gen. nov., a new genus of submarine disc-shaped sulphur reducing archaebacteria growing optimally at 105~ - Sys- tem. Appl. Microbiol., 4, 535-551
SrRO~aGREN, T. (1971): Vertical and horizontal distribution of Lophelia pertusa (Linn6) in Trondheimsfjorden on the west coast of Norway. -Kongel. norske Videnskaber Selsk. Skrifler, 6, 1-9, Oslo
StrEss, E. & Ft,zr'mRr~, D. (1972): Aragonitic ooids: experimental precipitation from sea water in the presence of humic acid. - Sedimentology, 19, 129-139, Oxford
SZEW'ZW:, U. & SCmNK, B. (1991): Attachment to amorpho us iron sulfide increases the activity of strictly anaerobic, gallic acid- degrading bacteria. - FEMS Microbiology Letters, 78, 115- 120
TALBOT, M.R. (1990): A review of the palaeohydrological inter- pretation of carbon and oxygen isotopic ratios in primary lacustrine carbonates. - Chem. Geol., 80, 261-279, Amster- dam
TEXC:-IERT, C. (1958) Cold- and deep-water coral banks. - Amer. Ass. Petrol. Geol. Bull., 42, 1064-1082, Tulsa
THmL, V., REfinER, J, & MIca~LtS, W. (1996): Biogeochemistry of Modern Porifera and Microbialites from Lizard Island (Great Barrier Reef, Australia) and fossil analogous. In: RErrNER, J., NmrwEmEa, F. & GtrNr~L, F. (eds.): Global and Regional Controls on Biogenic Sedimentation, I. Reef Evo- lution. Research Reports. - GOttinger Arb.Geol.Pal~ont., SB2, 129-132, G6ttingen
TrlmRV, J.-P. (1967): Mise en 6vidence des polysaccharides sur coupes fines en microscopic 61ectronique. - J. Microscopic, 6, 987-1018
THOMPSON, J.B. & FERPaS, F.G. (1990): Cyanobacterial precipita- tion of gypsum, calcite and magnesite from natural alkaline lake water. - Geology, 8, 995-998, Boulder
TISSOT, B.P. & W~-LTE, D.H. (1984): Petroleum formation and occurrence, pp. 699, New York. (Springer)
TOOMEV, D.F. & CYS, J.M. (1977): Spirorbid/algal stromatolites,
a probable marginal marine occurrence from the Lower Permian of New Mexico, USA. - N, Jb. Geol. Pal. Mh., 6, 331-342, Stuttgart
TRF.NCn R.K. (1987): Non-parasitic symbioses. In: Taylor F.R. (ed.): The biology of Dinoflagellates, 348-355, Oxford (Blackwe11)
TRtCnET, J. & DtFARGE, C. (1995): Non-biologically supported organomineralization. - Bull. Inst. Oc~anograph., Monaco, num. Sp~c. 14 (2), 203-236, Monaco
TRICHET, J. (1968): ~tudede la composition de la fraction organique des oolites. Comparison avec celle des membranes des bactfiries et des cyanophyc~es. - C.R.Aead. Sci.Paris, 267, 1492-1494, Paris.
TUDHOPE, A.W. & SCOFFr~, T.P. (1995): Processes of sedimenta- tion in Gollum Channel, Porcupine Seabight: submersible observations and sediment analysis. - Transactions Royal Soc. Edinburgh, Earth Sciences, 86, 49-55, Edinburgh
V~,XT~NEN, P. (1976): Microbiological studies in coastal waters of the Northern Baltic sea. I. Distribution and abundance of bacteria and yeasts in the Tvtirminne area. - Walter and Andre de Nottbeck Found, Sci. Rep., 1, 1-58
VACELET, Jr. & DONADEY, C. (1977): Electron microscope study of the association between some sponges and bacteria. - J. Exp. Mar. Biol. Ecol., 30, 301-314
VAN HEUKELEM, L., LEwrrus, A., KANA,T. & CRAFT, N. (1994): Improved separations of phytoplan.kton pigments using tem- perature-controlled high performance liquid chromatogra- phy. - Mar. Ecol. Prog. Set., 114, 303-313
VERMEU, G.J. (1987): Evolution and escalation: an ecological history of life, 527 pp. Princeton, New Jersey (Princeton Univ. Press).
VERRECCHIA, E.P., FREY1ET, P., VERRECCIIIA, K.E. & DtrMOm', J.L. (1995): Sphcrulites in calcrete laminar crusts : Biogenic CaCO 3 precipitation as a major contributor to crust forma- tion. - J. Sediment. Res., A 65, 690-700, Tulsa
VOLOGD~, A.G. (1962): Oldest algae of the USSR. - Publishing house of the USSR Academy of Sciences, Moscow, 656 pp. (in Russian).
WM.LACE, M.W. (1987): The role of internal erosion and sedi- mentation in the formation of stromatactis mudstones and associated lithologies. - J. Sediment. Petrol., 57, 695-700, Tulsa
WALTER, M.R. & HEYS, G.R. (1985): Links between therise of the Metazoa and the decline of stromatolites. - Precambrian Res., 29, 149-174, Amsterdam
WARNKE, K. (1994): Die unter-karbonischen Mud Mounds in den Counties Sligo und Leitrim, Nordwest-lrland. Fazies, Bildungsbedingungen und Diagense. - Documenta naturae, 88, 72 pp., Mtinchen
-- (1995): Calcification processes o f siliceous sponges in Vis6an limestones (Counties Sligo and Leitrim, NW Ireland.- Facies, 33, 215-228, Erlangen.
WARNKE, K. & MEISCItNF.R, D. (1995): Origin and depositional environment of Lower Carboniferous mud mounds of North- western Ireland.- In: REITNER, J. & NELIWE1LER. F. (coord.): Mud Mounds: A polygenetic Spectrum of fine grained car- bonate Buildups. - Facies, 32, 362,2, Erlangen
WATERBURY, J.B. (1992): The cyanobacteria - isolation, purifica- tion, and identification. In: B ALOWS, A.,TROPER, H.G., DWORKIN, M., HARDER, W., SC|ILEIIqER, K.-H. (eds.): The prokaryotes, 2 ad ed., 2058-2078, Berlin (Springer)
WEBB, G.E. (1994): Non-Waulsortian Mississippian bioherms: a comparative analysis. - In: BEAUCHAMP, B., EMBRY, A.F. & GLASS, D.J. (cds.): Pangea: Global Environments and Re- sources. - Canad. Soc. Petrol. Geol., Mem., 17, 701-702, Calgary
-- (1996): Was Phanerozoic reef history controlled by the distribution of non-enzymatically-secreted carbonates (mi- crobial carbonate and biologically-induced cement)? - Sedimentology, 43,947-971, Oxford

284
WF.~R, S., TRAtrB, W. & LOW~S'rAM, H.A. (1983): Orgaruc matrix in calcified exoskeletons. In: WESa'aROEK, P. & rJE JoNc, E.W. (eds.): Biomineralization and biological Mee-z! accumulation, 205-224, Amsterdam
WtDDEL, F. & P~r, rmo, N. (1981): Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids, I. Iso- lation of new sulfate-reducing bacteria enriched with acetate from salineenvironment. DescripfionofDesulfobacterpostga~e~ gen. nov. sp. nov.. - Archives MicrobioL, 129, 295-4/'09
Wmra-~soN, C.R. (1978a): Microbial association~ ;.n sponges. I. Ecology, physiology and microbial populations of coral reef sponges. - Mar. Biol., 49, 161-167, Berlin
-- (1978b): Microbial associations in sponges. III. Ultrastructure of the in situ associations in coral reef sponges. - Mar. Biol., 49, 177-185, Berlin
-- (1987): Productivity and abundance of large spongepopulafions on Flinders Reef flats, Coral Sea.- Coral Reefs, 5, 183-188, Berlin
Wn.soN, J.B. (1979a): The distribution of the coral Lophelia pertusa (L) [L. prolifera (Pallas)] in the north-east Atlantic. - J . Mar. Biol. Assoc. United Kingdom, 59, 149-164
-- (1979b): 'Patch' developmentofthedeep-watercoralLophelia pertusa (L.) on Rockall Bank. - J. Mar. Biol. Assoc. United Kingdom, 59, 165-177
WRIcH'r, V.P. & MAY,~a~L~ M. (1981): Organism-Sediment inter- actions in stromatolites: an example from the Upper Triassic of South West Britain. In: Mor~TY, C.L.V. (ed.): Phanerozoic Sromatolites, 75-84, Berlin (Springer)
ZtaqKa~, H. (1993): The origin of high-Mg-calcite microbialites in cryptic habitats of Caribbean coral reefs - their dependence on ligh.' and turbulence. - Facies, 29, 55-59, Erlangen
ZF.~, M~L. & PERK,S, R.D. (1979): Microbial alteration of Bahamian deep-sea carbonates. - Sedimentology, 26, 175- 201, Oxford
Z~RBaOWSKI, G. (1936): New genera of Cladochytriaceae. - Annales Missouri Botanical Garden, 23, 553-'564
Manuscript received January 23, 1997
Copyright © 2022 FDOKUMEN




















