
Biogenic amines formation in Streptococcus thermophilus isolated from home-made natural yogurt
8
Biogenic amines formation in Streptococcus thermophilus isolated from home-made natural yogurt Yekta Gezginc a , Ismail Akyol b,c , Esmeray Kuley d,⇑ , Fatih Özogul d a Kahramanmaras Sutcu Imam University, Faculty of Agriculture, Department of Food Engineering, 46100 Kahramanmaras, Turkey b Kahramanmaras Sutcu Imam University, Faculty of Agriculture, Department of Agricultural Biotechnology, 46100 Kahramanmaras, Turkey c Kahramanmaras Sutcu Imam University, Microbiology and Microbial Genetics Laboratory, USKIM 46100, Kahramanmaras, Turkey d Cukurova University, Faculty of Fisheries, Department of Fish Processing Technology, 01330 Balcali, Adana, Turkey article info Article history: Received 28 August 2012 Received in revised form 9 October 2012 Accepted 25 October 2012 Available online 10 November 2012 Keywords: Biogenic amines Streptococcus thermophilus PCR hdcA gene Yogurt abstract Twelve different biogenic amines formation in 58 isolates of Streptococcus thermophilus from home-made natural yogurt were investigated in histidine (HDB) and lysine decarboxylase broth (LDB). All S. thermophilus isolates had an ability to produce twelve different biogenic amines in HDB and LDB. Most of the S. thermophilus isolates formed low amounts of histamine (1–50 mg/L) from histidine. Apart from one isolate, S. thermophilus produced tyramine at low (47 isolates) and medium (10 isolates) levels. The amount of each specific biogenic amine produced by S. thermophilus was generally lower than 100 mg L 1 . Also, the presence of hdcA gene was investigated using PCR technique and relation between gene and histamine production was conducted in S. thermophilus isolates. This study showed that most of the S. thermophilus isolates have the ability to form biogenic amines, especially histamine, and tyramine, which is an important consideration when selecting strains as starter cultures. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction Streptococcus thermophilus is a thermophilic Gram-positive microorganism which belongs to the lactic acid bacteria group (LAB). It is used for production of fermented milks such as yogurt and for the production of some Italian and Swiss type cheeses, such as Mozzarella and Emmental (Iyer, Tomar, & Singh, 2010). The main roles of S. thermophilus in milk fermentation are to utilise gal- actose and provide rapid acidification with lactic acid production. Several other important technological properties, such as sugar metabolism, proteolytic activity, and urease activity are also known, and represent the degree of phenotypic diversity existing within the species. At the end of metabolic activities, some LAB strains can produce toxic compounds such as biogenic amines (BA) (Ladero et al., 2012). BA are organic, basic nitrogenous compounds of low molecular weight (Bodmer, Imark, & Kneubuhl, 1999). Removal of the a- carboxyl group from an amino acid leads to the corresponding biogenic amine. The most important BA in food are tyramine, histamine, cadaverine, putrescine, 2-phenylethylamine and agma- tine are produced by decarboxylation of the amino acids tyrosine, histidine, lysine, ornithine, phenylalanine and arginine, respec- tively (Linares, Martin, Ladero, Alvarez, & Fernandez, 2011). The toxicological level of BA is very difficult to establish because it de- pends on individual characteristics and the presence of other amines in food. However, a maximum total BA (the sum of tyra- mine, histamine, putrescine, and cadaverine) level of 750– 900 mg/kg has been proposed as a safety limit (Ladero, Fernandez, & Alvarez, 2009). The presence of BA depends on decarboxylase, the enzyme that catalyses the decarboxylation of amino acids and a transporter responsible for amino acid–BA interchange (Linares et al., 2011). Decarboxylation increases survival under stress conditions (Rhee, Kim, & Choi, 2005) via the consumption of protons and the excretion of amines and CO 2 , helping to restore the external pH. BA production may also offer a way of obtaining energy. This function is important for microorganisms, such as lactic acid bacte- ria (LAB), which lacking a respiratory chain for generating high yields of ATP (Vido et al., 2004). A relation was observed between an acidic pH and increase in BA synthesis (Fernandez, Linares, Rodriguez, & Alvarez, 2007). Decarboxylase enzymes have an opti- mum pH of around 5.0 (Moreno-Arribas & Polo, 2008). High temperatures can accelerate amino acid accumulation and, hence, stimulate amine formation (Joosten and van Boeckel, 1988). Addition of NaCl to milk can also affect BA formation. The addition of NaCl to milk with Lactococcus lactis ssp. cremoris and Lactococcus lactis spp. lactis slowed down the formation of agmatine, spermidine, spermine, putrescine, histamine, tyramine 0308-8146/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.foodchem.2012.10.138 ⇑ Corresponding author. Tel.: +90 322 3386084x2961; fax: +90 322 3386439. E-mail address: [email protected] (E. Kuley). Food Chemistry 138 (2013) 655–662 Contents lists available at SciVerse ScienceDirect Food Chemistry journal homepage: www.elsevier.com/locate/foodchem
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Biogenic amines formation in Streptococcus thermophilus isolated from home-made natural yogurt

Food Chemistry 138 (2013) 655–662
Contents lists available at SciVerse ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Biogenic amines formation in Streptococcus thermophilus isolated fromhome-made natural yogurt
Yekta Gezginc a, Ismail Akyol b,c, Esmeray Kuley d,⇑, Fatih Özogul d
a Kahramanmaras Sutcu Imam University, Faculty of Agriculture, Department of Food Engineering, 46100 Kahramanmaras, Turkeyb Kahramanmaras Sutcu Imam University, Faculty of Agriculture, Department of Agricultural Biotechnology, 46100 Kahramanmaras, Turkeyc Kahramanmaras Sutcu Imam University, Microbiology and Microbial Genetics Laboratory, USKIM 46100, Kahramanmaras, Turkeyd Cukurova University, Faculty of Fisheries, Department of Fish Processing Technology, 01330 Balcali, Adana, Turkey
a r t i c l e i n f o a b s t r a c t
Article history:Received 28 August 2012Received in revised form 9 October 2012Accepted 25 October 2012Available online 10 November 2012
Keywords:Biogenic aminesStreptococcus thermophilusPCRhdcA geneYogurt
0308-8146/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.foodchem.2012.10.138
⇑ Corresponding author. Tel.: +90 322 3386084x29E-mail address: [email protected] (E. Kuley).
Twelve different biogenic amines formation in 58 isolates of Streptococcus thermophilus from home-madenatural yogurt were investigated in histidine (HDB) and lysine decarboxylase broth (LDB). AllS. thermophilus isolates had an ability to produce twelve different biogenic amines in HDB and LDB. Mostof the S. thermophilus isolates formed low amounts of histamine (1–50 mg/L) from histidine. Apart fromone isolate, S. thermophilus produced tyramine at low (47 isolates) and medium (10 isolates) levels. Theamount of each specific biogenic amine produced by S. thermophilus was generally lower than100 mg L�1. Also, the presence of hdcA gene was investigated using PCR technique and relation betweengene and histamine production was conducted in S. thermophilus isolates. This study showed that most ofthe S. thermophilus isolates have the ability to form biogenic amines, especially histamine, and tyramine,which is an important consideration when selecting strains as starter cultures.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Streptococcus thermophilus is a thermophilic Gram-positivemicroorganism which belongs to the lactic acid bacteria group(LAB). It is used for production of fermented milks such as yogurtand for the production of some Italian and Swiss type cheeses, suchas Mozzarella and Emmental (Iyer, Tomar, & Singh, 2010). Themain roles of S. thermophilus in milk fermentation are to utilise gal-actose and provide rapid acidification with lactic acid production.Several other important technological properties, such as sugarmetabolism, proteolytic activity, and urease activity are alsoknown, and represent the degree of phenotypic diversity existingwithin the species. At the end of metabolic activities, some LABstrains can produce toxic compounds such as biogenic amines(BA) (Ladero et al., 2012).
BA are organic, basic nitrogenous compounds of low molecularweight (Bodmer, Imark, & Kneubuhl, 1999). Removal of the a-carboxyl group from an amino acid leads to the correspondingbiogenic amine. The most important BA in food are tyramine,histamine, cadaverine, putrescine, 2-phenylethylamine and agma-tine are produced by decarboxylation of the amino acids tyrosine,histidine, lysine, ornithine, phenylalanine and arginine, respec-
ll rights reserved.
61; fax: +90 322 3386439.
tively (Linares, Martin, Ladero, Alvarez, & Fernandez, 2011). Thetoxicological level of BA is very difficult to establish because it de-pends on individual characteristics and the presence of otheramines in food. However, a maximum total BA (the sum of tyra-mine, histamine, putrescine, and cadaverine) level of 750–900 mg/kg has been proposed as a safety limit (Ladero, Fernandez,& Alvarez, 2009). The presence of BA depends on decarboxylase,the enzyme that catalyses the decarboxylation of amino acidsand a transporter responsible for amino acid–BA interchange(Linares et al., 2011).
Decarboxylation increases survival under stress conditions(Rhee, Kim, & Choi, 2005) via the consumption of protons andthe excretion of amines and CO2, helping to restore the externalpH. BA production may also offer a way of obtaining energy. Thisfunction is important for microorganisms, such as lactic acid bacte-ria (LAB), which lacking a respiratory chain for generating highyields of ATP (Vido et al., 2004). A relation was observed betweenan acidic pH and increase in BA synthesis (Fernandez, Linares,Rodriguez, & Alvarez, 2007). Decarboxylase enzymes have an opti-mum pH of around 5.0 (Moreno-Arribas & Polo, 2008).
High temperatures can accelerate amino acid accumulationand, hence, stimulate amine formation (Joosten and van Boeckel,1988). Addition of NaCl to milk can also affect BA formation.The addition of NaCl to milk with Lactococcus lactis ssp. cremorisand Lactococcus lactis spp. lactis slowed down the formation ofagmatine, spermidine, spermine, putrescine, histamine, tyramine

656 Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662
and tryptamine (Santos, Souza, Cerqueira, & Gloria, 2003). Simi-larly, the addition of high concentrations of sodium chloride(5%) to milk inoculated with Enterococcus faecalis, reduced to aminimum the amount of 2-phenylethylamine and tyramine pro-duction (Gardini, Martuscelli, Caruso, & Galgano, 2001). However,addition of 2% NaCl in a medium inoculated with Enterobacter aer-ogenes at 37 �C caused a slight increase in amine formation, andadditions of 3% or more lacked any stimulatory effect on amineformation (Greif, Greifova, & Karovicova, 2006). The highest pro-duction of tyramine by Lactococcus lactis subsp. lactis and L. lactissubsp. cremoris was obtained within the culture with the highest(2% w/v) salt concentration applied (Bunkova, Bunka, Pollakova,Podesvova, & Drab, 2011). Many factors such as bacterial density,synergistic effects between microorganisms, level of proteolysis(availability of substrate), salt-in-moisture level and ripeningand storage temperatures are found to have limiting effects onthe build-up of amines (Valsamaki, Michaelidou, & Polychroniadou,2000).
An ingestion of BA can cause headache, hypertension, pyrexia,or heart disease. Histamine is the most toxic biogenic amine de-tected in foods. It can result in unpleasant reactions by stimulationof the nervous system and blood vessels in the human body (Gao,Guo, Wang, & Zhang, 2011). On the other hand, BA biosynthesis isessential since these compounds act as precursor of the synthesisof hormones, alkaloids, nucleic acids, and proteins in eukaryoticcells (Premont, Gainetdinov, & Caron, 2001).
Yogurt is milk fermented by the lactic acid bacteria S. thermo-philus and Lactobacillus bulgaricus (Sieuwerts et al., 2010). The tech-nological conditions used in the production of dairy products mayfavour the synthesis and accumulation of BAs (Calles-Enriquezet al., 2010). Several strains of S. thermophilus (Calles-Enriquezet al., 2010; La Gioia et al., 2011), Lactobacillus brevis, Lactococcuslactis (Ladero et al., 2011) and Lactobacillus curvatus (Ladero,Sanchez-Llana, Fernandez, & Alvarez, 2011), obtained from dairysources, have been identified as histamine, tyramine, or putrescineproducers (Linares et al., 2012).
The major biogenic amine of stirred yogurt samples is hista-mine. In unripened cheeses, milks, and yogurts, spermidine andspermine are the prevailing amines (Novella-Rodriguez, Veciana-Nogues, & Vidal-Carou, 2000); however, it remains unclearwhether they are synthesised by microorganisms or have anendogenous origin (Linares et al. 2012). S. thermophilus strains pos-sessing a histidine decarboxylase gene represented 6% of dairy iso-lates screened (Rossi et al., 2011). Some strains of lactic acidbacteria synthesise histamine, their ability strictly related to thepossession of the histidine decarboxylase gene (Landete, Ferrer, &Pardo, 2005). Two S. thermophilus strains, from a total of 69 S. ther-mophilus strains screened showed a capacity to produce histamine(Calles-Enriquez et al., 2010).
The production of BAs by lactic acid bacteria is not a desirableproperty (Linares et al., 2011). The presence of BA in fermentedfood has traditionally been used as an indicator of starter quality.Biogenic amine production by bacteria was shown to be strain-dependent rather than being related to species specificity (Garai,Duenas, Irastorza, & Moreno-Arribas, 2007). It is important to notethat it is not always possible to make such modifications to the fer-menting process without causing changes in texture flavour orquality.
The most important biogenic amines in dairy foods are hista-mine, tyramine and cadaverine (from histidine, tyrosine and lysinedecarboxylation, respectively). The occurrence of these amines ismainly due to microbial groups that participate in the fermenta-tion process. Therefore, the aims of the study were to investigatebiogenic amines formation by S. thermophilus isolated fromhome-made natural yogurt, and in lysine and histidine decarboxyl-ase broth.
2. Material and methods
2.1. Bacterial strains and growth conditions
A total of 67 LAB, including S. thermophilus, were isolated fromtraditional home-made yogurt samples randomly collectedfrom different region of Turkey in a previous study (Gezginc,2010). Isolated LAB were identified using biochemical tests andsequencing. The 16S rRNA region PCR amplification and sequenceanalysis of strains were performed according to the methods ofTabasco, Paarup, Janer, Pelaez, and Requena (2007). A total of 58S. thermophilus strains were examined. S. thermophilus strains wereroutinely grown at 42 �C in SM17 medium (Merck, Germany).
2.2. Culture media and bacterial extraction
The production of ammonia and biogenic amines from all S.thermophilus strains in this work was monitored using histidinedecarboxylase broth (HDB) and lysine decarboxylase broth (LDB),proposed by Klausen and Huss (1987). HDB and LDB contained1 g peptone, 0.5 g Lab-Lemco powder (Oxoid, Basingstoke, UK),2.5 mg NaCl (Merck, Darmstadt, Germany), 4.01 g L-histidinemonohydrochloride monohydrate or L-lysine monohydrochloride,and 2.5 mg pyridoxal HCl (all Sigma, Steinheim, Germany) in500 mL distilled water; the pH was adjusted to 5.5 with 1 M KOHor 6% trichloroacetic acid (both Riedel-de-Haën, Seelze, Germany).After preparation HDB and LDB were pipetted into 10-ml bottlesand then autoclaved at 121 �C for 15 min. prior to use.
SM17 broth (Merck, Germany) was used for propagation of S.thermophilus cultures, S. thermophilus strains were incubated at37 �C for 72 h, after which 0.5 ml of these bacterial cultures(1011 cfu/ml) were removed and put into the LDB and HDB, to al-low them to decarboxylate the lysine and histidine.
In order to identify the S. thermophilus strains showing the abil-ity to decarboxylate, 5 mL of the LDB and HDB containing S. ther-mophilus strains were removed to separate bottles and then 2 mltrichloroacetic acid (6%) were added. They were then centrifugedat 3000 g for 10 min and then filtered through a filter paper(589/2, Ø125 mm, Schleicher & Schuell, Dassel, Germany). Afterthat, 4 mL of bacterial supernatant were taken for derivatisationin order to be analysed by HPLC.
2.2.1. Chemical reagentsL-Lysine, L-histidine and all BA standards were purchased from
Sigma–Aldrich (Munich, Germany). The used BA standards werehistamine dihydrochloride, tyramine hydrochloride, tryptaminehydrochloride, putrescine dihydrochloride, 2-phenylethylaminehydrochloride, cadaverine dihydrochloride, spermidine trihydro-chloride, spermine tetrahydrochloride, 5-hydroxytryptamine(serotonin), 3-hydroxytyramine hydrochloride (dopamine), agma-tine sulphate, trimethylamine hydrochloride and ammonium chlo-ride. Each standard was dissolved in 10 mL HPLC-grade water. Thefinal concentration of the free base for each amine was 10 mg/mLsolution.
2.2.2. Derivatisation of bacterial extractionA stock solution was prepared by dissolving 2% benzoyl chloride
in acetonitrile to enhance the reaction with amines. For derivatisa-tion of standard amine solutions, 100 lL were taken (4 mL for ex-tracted bacterial cultures) from each free base standard solution(10 mg/mL). Sodium hydroxide (1 mL of 2 M) was added, followedby 40 lL of benzoyl chloride and the solution mixed on a vortexmixer for 1 min. The reaction mixture was left at room tempera-ture for 20 min. After that, the benzoylation was stopped by adding2 mL of saturated sodium chloride solution and the solution

Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662 657
extracted twice with 2 mL of diethyl ether. The upper organic layer(4 mL) was transferred into a clean tube after mixing. Afterwards,the organic layer was evaporated to dryness in a stream of nitro-gen. The residue was dissolved in 1 mL of acetonitrile and 10-lLaliquots were injected into the HPLC (Ozogul, 2004).
2.3. Analytical method
For the biogenic amines analyses, a Shimadzu Prominence HPLCapparatus (Shimadzu, Kyoto, Japan) equipped with a SPD-M20Adiode array detector and two pumps (Shimadzu LC-10AT), auto-sampler (SIL 20AC), column oven (CTO-20AC), and a communica-tion bus module (CBM-20A) with valve unit FCV-11AL was used.The column was ODS Hypersil, 5 lm, 250 � 4.6 mm (Phenomenex,Macclesfield, UK).
Biogenic amines analysis was done using the method of Ozogul(2004) and measured in mg amines per litre broth. The confirma-tion of biogenic amines production was accomplished using a rapidHPLC method with a reversed phase column by using a gradientelution program. The same analytical method was used for ammo-nia and trimethylamine separation. Chromatographic separationwas carried out using continuous gradient elution with acetonitrile(eluent A) and HPLC grade water (eluent B). The total separationtime was less than 20 min. and the injection volume was 10 lL.Detection was monitored at 254 nm. A standard curve forammonia and each of the twelve amines in the range of 0–50 mg/mL was prepared. Correlation coefficient of peak areaagainst amine standard concentrations for each compound wascalculated after injecting five replicates of each standard solutionof amine. The correlation coefficient (r2) in the curve was >0.99for each benzoylated amine and ammonia, which showed a linearrelationship between amine concentration and detector response;therefore the gradient elution program used in this work wassatisfactory.
2.4. Molecular studies
Chromosomal DNA extraction was carried out using an isolationkit (Fermentas, Thermo Fisher Scientific, Waltham, MA). PrimersHdc3F (50AGATGGTATTGTTTCTTATGA30) and Hdc4R (50CAAACACCAGCATCTTC30) were designed for amplification of a 435 bp lengthpart of hdc gene (Coton & Coton, 2005). Oligonucleotides were ob-tained from Iontek (Istanbul, Turkey). PCR was done in 40 lL of mix-ture by using Fermentas DNA polymerase, 5 U/lL, and 10� PCRbuffer (Amersham, Little Chalfont, UK). Twenty picomoles of eachprimer were used, and deoxynucleoside triphosphates (dNTPs) (Fer-mentas) were used at a concentration of 250 lM each. The genomicDNA of S. thermophilus isolates was used as a template. PCR ampli-fications were performed in a DNA thermal cycler with the follow-ing thermal cycling program: initial cycle, 2 min at 95 �C; next 30cycles, 1 min at 94 �C; 1 min at 65 �C, and 2 min at 72 �C. AmplifiedPCR products were run on 1.5% electrophoresis gel (Bio-Rad, Hercu-les, CA) and analysed under UV light.
2.5. Statistical analysis
The mean values and standard deviations were calculated basedon data taken from at least three independent experiments con-ducted on separate days using freshly made reagents. One-wayANOVA was used to determine the significance of differences atp < 0.05. All statistics were performed using SPSS 15.0 for Windows(SPSS Inc., Chicago, IL).
3. Result and discussion
3.1. Biogenic amine production by isolates in histidine enrichmentdecarboxylase broth
HPLC shows that a number of S. thermophilus isolates producedbiogenic amines in histidine decarboxylase (HDB) (Table 1). All S.thermophilus isolates had the ability to produce ammonia. Thirty-nine strains of S. thermophilus produced below 100 mg/L of ammo-nia, while the other strains produced more than 100 mg/L.
S. thermophilus isolates produced not only histamine or cadav-erine but also other amines (putrescine, tyramine, etc.) in specificdecarboxylase broths, although the other amino acids (ornithine,tyrosine, etc.) were not added into HDB and LDB. In the presentstudy, the broth used does contain peptone and beef extract, whichcontain the other amino acids. The amounts of glutamic acid, argi-nine, lysine, tyrosine and tryptophan in peptone and beef extractare calculated as 304, 170, 132, 14, and 16 mg/L, respectively (Ox-oid L37; Oxoid L29). The observed other biogenic amines are thuslikely to have been produced from the amino acids in peptone andbeef extract. Most of the isolates produced twelve BA in HDB,although dopamine and agmatine were formed by most isolatesat low levels (<100 mg/L). Biogenic amine production by isolateswas generally below 100 mg/L. The lactic acid bacteria strains (E.faecalis, Leuconostoc mesenteroides, Lactobacillus fermentum, Lacto-bacillus acidophilus, Lactobacillus casei) isolated from yoghurt pro-duced BA in amino acid decarboxylase broth, although S.thermophilus did not accumulate BA (Omafuvbe & Enyioha, 2011).
Bunkova et al. (2009) reported that L. lactis subsp. cremoris, S.thermophilus and Lactobacillus delbrueckii subsp. bulgaricus producetyramine, whereas the other tested BA, such as histamine, putres-cine and cadaverine were not detected. Omafuvbe and Enyioha(2011) found that some of the LAB strains belonging to Lactobacil-lus, Lactococcus, Leuconostoc, Streptococcus and Enterococcus generacan decarboxylate tyrosine and lysine. Lactic acid bacteria are themain bacteria responsible for putrescine production in wine(Moreno-Arribas & Polo, 2008). Several lactic acid bacteria canuse agmatine via the agmatine deaminase pathway to produceputrescine. Putrescine, one of the main biogenic amines associatedwith microbial food spoilage, can be formed by bacteria from argi-nine via ornithine decarboxylase (ODC), or from agmatine via agm-atine deiminase (AgDI). Putrescine production from agmatinecould be linked to the aguA and ptcA genes in Lactobacillus hilgardiiX1B and E. faecalis ATCC 11700 (Landete, Arena, Pardo, Manca deNadra, & Ferrer, 2010). In the current study, 11 isolates of S. ther-mophilus did not produce putrescine, whilst two isolates werecharacterised as medium putrescine producers (<500 mg/L). Onestrain of S. thermophilus did not produce cadaverine, although mostof the isolates had an ability to form cadaverine in the range of 1–100 mg/L. Low concentrations of cadaverine production (below10 mg/L) were reported for 17 strains of lactobacilli, two strainsof enterococci and one strain of Pediococcus isolated from dairyproducts (Lorencova et al., 2012).
Spermidine production by most S. thermophilus isolates (40 iso-lates) was below 100 mg/L, but two isolates were good spermidineproducers. Although ten isolates of S. thermophilus did not inducetryptamine and spermine production, a medium amount of amineproduction was observed by one and three isolates of S. thermophi-lus, respectively.
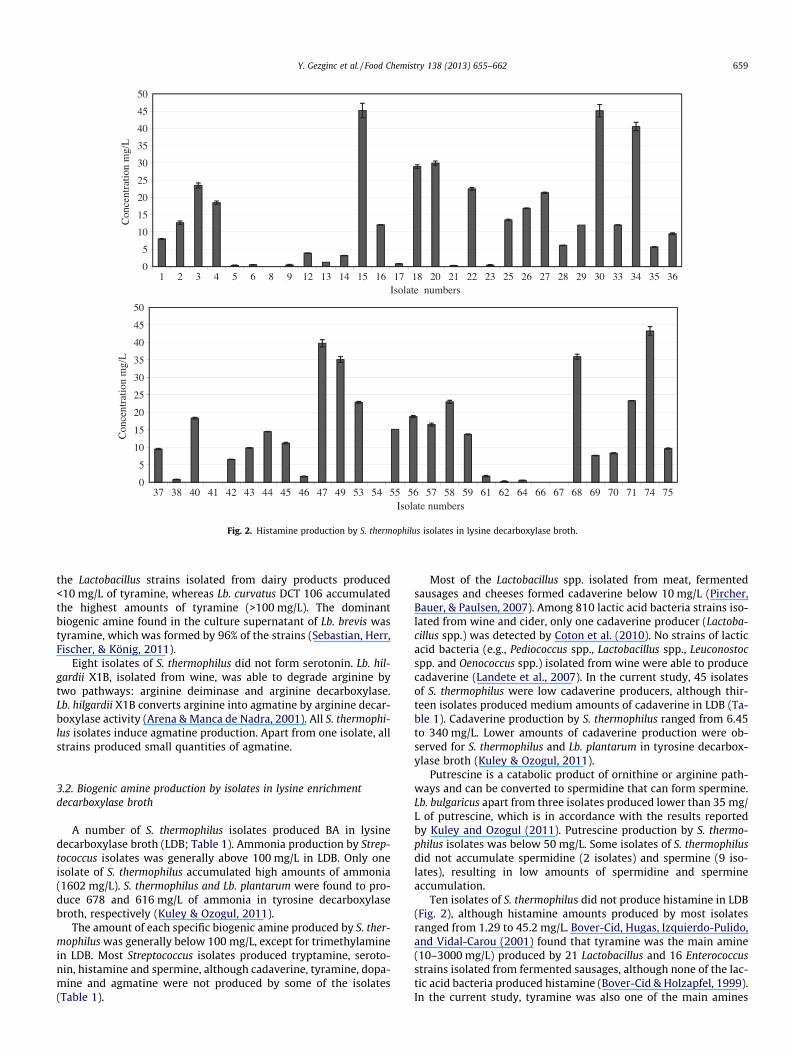
Sixteen isolates of S. thermophilus did not produce histamine inHDB. Most of the S. thermophilus isolates formed low amounts ofhistamine (1–50 mg/L) from histidine whilst one isolate was a pro-lific amine producer, with amine production above 36.33 mg/L(Fig. 1). Landete, de Las Rivas, Marcobal, and Munoz (2007)found that Oenococcus oeni, Lb. hilgardii, Lactobacillus mali, Le.
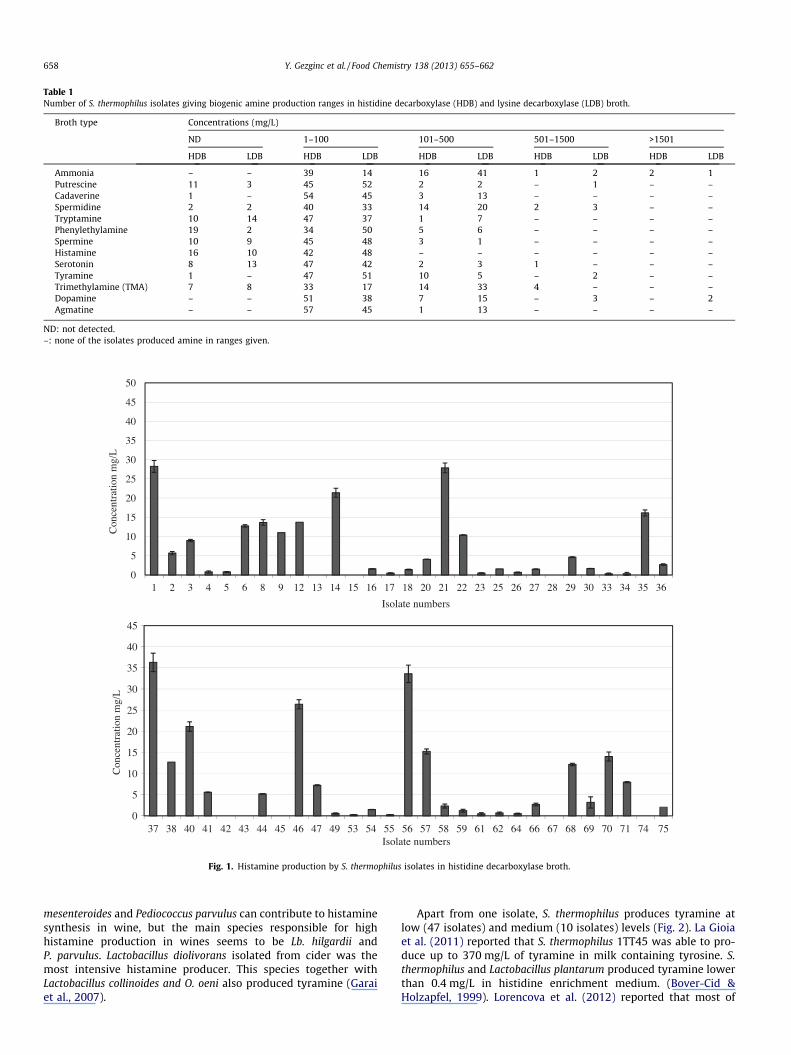
Table 1Number of S. thermophilus isolates giving biogenic amine production ranges in histidine decarboxylase (HDB) and lysine decarboxylase (LDB) broth.
Broth type Concentrations (mg/L)
ND 1–100 101–500 501–1500 >1501
HDB LDB HDB LDB HDB LDB HDB LDB HDB LDB
Ammonia – – 39 14 16 41 1 2 2 1Putrescine 11 3 45 52 2 2 – 1 – –Cadaverine 1 – 54 45 3 13 – – – –Spermidine 2 2 40 33 14 20 2 3 – –Tryptamine 10 14 47 37 1 7 – – – –Phenylethylamine 19 2 34 50 5 6 – – – –Spermine 10 9 45 48 3 1 – – – –Histamine 16 10 42 48 – – – – – –Serotonin 8 13 47 42 2 3 1 – – –Tyramine 1 – 47 51 10 5 – 2 – –Trimethylamine (TMA) 7 8 33 17 14 33 4 – – –Dopamine – – 51 38 7 15 – 3 – 2Agmatine – – 57 45 1 13 – – – –
ND: not detected.–: none of the isolates produced amine in ranges given.
0
5
10
15
20
25
30
35
40
45
50
1 2 3 4 5 6 8 9 12 13 14 15 16 17 18 20 21 22 23 25 26 27 28 29 30 33 34 35 36
Con
cent
ratio
n m
g/L
Isolate numbers
0
5
10
15
20
25
30
35
40
45
37 38 40 41 42 43 44 45 46 47 49 53 54 55 56 57 58 59 61 62 64 66 67 68 69 70 71 74 75
Con
cent
ratio
n m
g/L
Isolate numbers
Fig. 1. Histamine production by S. thermophilus isolates in histidine decarboxylase broth.
658 Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662
mesenteroides and Pediococcus parvulus can contribute to histaminesynthesis in wine, but the main species responsible for highhistamine production in wines seems to be Lb. hilgardii andP. parvulus. Lactobacillus diolivorans isolated from cider was themost intensive histamine producer. This species together withLactobacillus collinoides and O. oeni also produced tyramine (Garaiet al., 2007).
Apart from one isolate, S. thermophilus produces tyramine atlow (47 isolates) and medium (10 isolates) levels (Fig. 2). La Gioiaet al. (2011) reported that S. thermophilus 1TT45 was able to pro-duce up to 370 mg/L of tyramine in milk containing tyrosine. S.thermophilus and Lactobacillus plantarum produced tyramine lowerthan 0.4 mg/L in histidine enrichment medium. (Bover-Cid &Holzapfel, 1999). Lorencova et al. (2012) reported that most of
0
5
10
15
20
25
30
35
40
45
50
1 2 3 4 5 6 8 9 12 13 14 15 16 17 18 20 21 22 23 25 26 27 28 29 30 33 34 35 36
Con
cent
ratio
n m
g/L
Isolate numbers
0
5
10
15
20
25
30
35
40
45
50
37 38 40 41 42 43 44 45 46 47 49 53 54 55 56 57 58 59 61 62 64 66 67 68 69 70 71 74 75
Con
cent
ratio
n m
g/L
Isolate numbers
Fig. 2. Histamine production by S. thermophilus isolates in lysine decarboxylase broth.
Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662 659
the Lactobacillus strains isolated from dairy products produced<10 mg/L of tyramine, whereas Lb. curvatus DCT 106 accumulatedthe highest amounts of tyramine (>100 mg/L). The dominantbiogenic amine found in the culture supernatant of Lb. brevis wastyramine, which was formed by 96% of the strains (Sebastian, Herr,Fischer, & König, 2011).
Eight isolates of S. thermophilus did not form serotonin. Lb. hil-gardii X1B, isolated from wine, was able to degrade arginine bytwo pathways: arginine deiminase and arginine decarboxylase.Lb. hilgardii X1B converts arginine into agmatine by arginine decar-boxylase activity (Arena & Manca de Nadra, 2001). All S. thermophi-lus isolates induce agmatine production. Apart from one isolate, allstrains produced small quantities of agmatine.
3.2. Biogenic amine production by isolates in lysine enrichmentdecarboxylase broth
A number of S. thermophilus isolates produced BA in lysinedecarboxylase broth (LDB; Table 1). Ammonia production by Strep-tococcus isolates was generally above 100 mg/L in LDB. Only oneisolate of S. thermophilus accumulated high amounts of ammonia(1602 mg/L). S. thermophilus and Lb. plantarum were found to pro-duce 678 and 616 mg/L of ammonia in tyrosine decarboxylasebroth, respectively (Kuley & Ozogul, 2011).
The amount of each specific biogenic amine produced by S. ther-mophilus was generally below 100 mg/L, except for trimethylaminein LDB. Most Streptococcus isolates produced tryptamine, seroto-nin, histamine and spermine, although cadaverine, tyramine, dopa-mine and agmatine were not produced by some of the isolates(Table 1).
Most of the Lactobacillus spp. isolated from meat, fermentedsausages and cheeses formed cadaverine below 10 mg/L (Pircher,Bauer, & Paulsen, 2007). Among 810 lactic acid bacteria strains iso-lated from wine and cider, only one cadaverine producer (Lactoba-cillus spp.) was detected by Coton et al. (2010). No strains of lacticacid bacteria (e.g., Pediococcus spp., Lactobacillus spp., Leuconostocspp. and Oenococcus spp.) isolated from wine were able to producecadaverine (Landete et al., 2007). In the current study, 45 isolatesof S. thermophilus were low cadaverine producers, although thir-teen isolates produced medium amounts of cadaverine in LDB (Ta-ble 1). Cadaverine production by S. thermophilus ranged from 6.45to 340 mg/L. Lower amounts of cadaverine production were ob-served for S. thermophilus and Lb. plantarum in tyrosine decarbox-ylase broth (Kuley & Ozogul, 2011).
Putrescine is a catabolic product of ornithine or arginine path-ways and can be converted to spermidine that can form spermine.Lb. bulgaricus apart from three isolates produced lower than 35 mg/L of putrescine, which is in accordance with the results reportedby Kuley and Ozogul (2011). Putrescine production by S. thermo-philus isolates was below 50 mg/L. Some isolates of S. thermophilusdid not accumulate spermidine (2 isolates) and spermine (9 iso-lates), resulting in low amounts of spermidine and spermineaccumulation.
Ten isolates of S. thermophilus did not produce histamine in LDB(Fig. 2), although histamine amounts produced by most isolatesranged from 1.29 to 45.2 mg/L. Bover-Cid, Hugas, Izquierdo-Pulido,and Vidal-Carou (2001) found that tyramine was the main amine(10–3000 mg/L) produced by 21 Lactobacillus and 16 Enterococcusstrains isolated from fermented sausages, although none of the lac-tic acid bacteria produced histamine (Bover-Cid & Holzapfel, 1999).In the current study, tyramine was also one of the main amines
660 Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662
produced by S. thermophilus isolates. Two isolates of S. thermophilusproduced significant amounts of tyramine, 504 and 539 mg/L. Theformation of dopamine by S. thermophilus ranged from 1.17 to2160 mg/L, while agmatine was produced by 13 isolates of S. ther-mophilus at levels above 100 mg/L.
3.3. Amplification of the hdcA gene in isolates
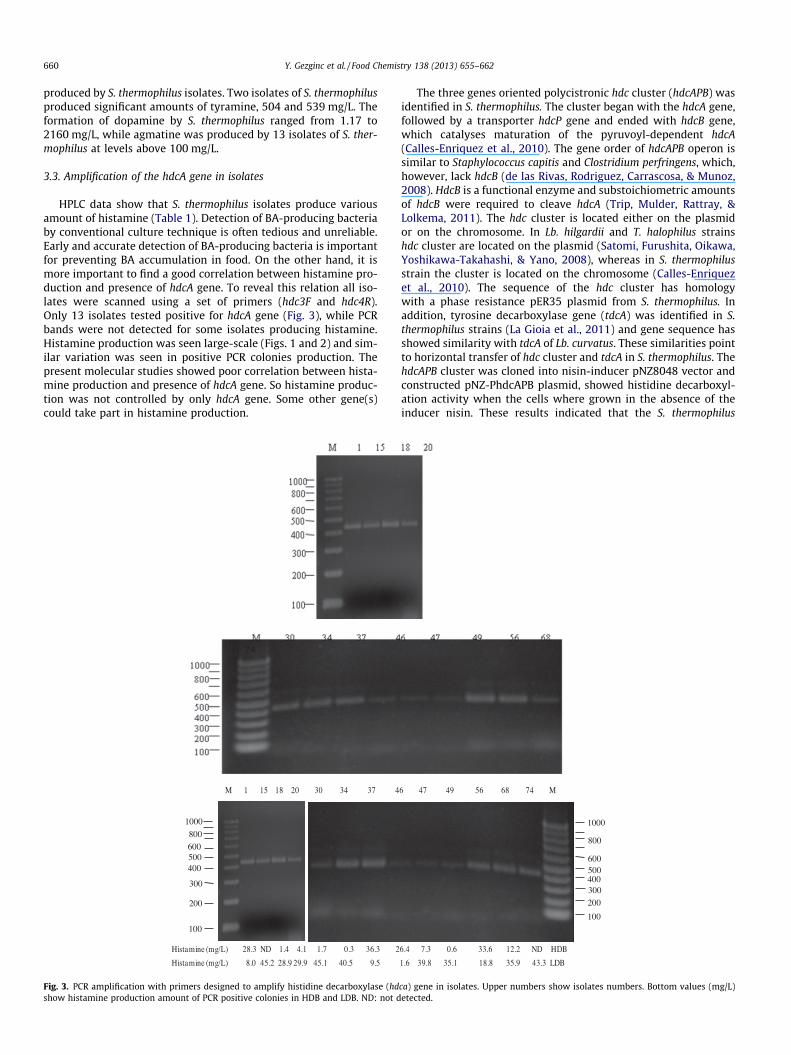
HPLC data show that S. thermophilus isolates produce variousamount of histamine (Table 1). Detection of BA-producing bacteriaby conventional culture technique is often tedious and unreliable.Early and accurate detection of BA-producing bacteria is importantfor preventing BA accumulation in food. On the other hand, it ismore important to find a good correlation between histamine pro-duction and presence of hdcA gene. To reveal this relation all iso-lates were scanned using a set of primers (hdc3F and hdc4R).Only 13 isolates tested positive for hdcA gene (Fig. 3), while PCRbands were not detected for some isolates producing histamine.Histamine production was seen large-scale (Figs. 1 and 2) and sim-ilar variation was seen in positive PCR colonies production. Thepresent molecular studies showed poor correlation between hista-mine production and presence of hdcA gene. So histamine produc-tion was not controlled by only hdcA gene. Some other gene(s)could take part in histamine production.
100
1000
800
600500400
300
200
M 1 15 18 20 30 34 37 4
Histamine (mg/L) 28.3 ND 1.4 4.1 1.7 0.3 36.3 2
Histamine (mg/L) 8.0 45.2 28.9 29.9 45.1 40.5 9.5
Fig. 3. PCR amplification with primers designed to amplify histidine decarboxylase (hdshow histamine production amount of PCR positive colonies in HDB and LDB. ND: not d
The three genes oriented polycistronic hdc cluster (hdcAPB) wasidentified in S. thermophilus. The cluster began with the hdcA gene,followed by a transporter hdcP gene and ended with hdcB gene,which catalyses maturation of the pyruvoyl-dependent hdcA(Calles-Enriquez et al., 2010). The gene order of hdcAPB operon issimilar to Staphylococcus capitis and Clostridium perfringens, which,however, lack hdcB (de las Rivas, Rodriguez, Carrascosa, & Munoz,2008). HdcB is a functional enzyme and substoichiometric amountsof hdcB were required to cleave hdcA (Trip, Mulder, Rattray, &Lolkema, 2011). The hdc cluster is located either on the plasmidor on the chromosome. In Lb. hilgardii and T. halophilus strainshdc cluster are located on the plasmid (Satomi, Furushita, Oikawa,Yoshikawa-Takahashi, & Yano, 2008), whereas in S. thermophilusstrain the cluster is located on the chromosome (Calles-Enriquezet al., 2010). The sequence of the hdc cluster has homologywith a phase resistance pER35 plasmid from S. thermophilus. Inaddition, tyrosine decarboxylase gene (tdcA) was identified in S.thermophilus strains (La Gioia et al., 2011) and gene sequence hasshowed similarity with tdcA of Lb. curvatus. These similarities pointto horizontal transfer of hdc cluster and tdcA in S. thermophilus. ThehdcAPB cluster was cloned into nisin-inducer pNZ8048 vector andconstructed pNZ-PhdcAPB plasmid, showed histidine decarboxyl-ation activity when the cells where grown in the absence of theinducer nisin. These results indicated that the S. thermophilus
1000
100
800
600500400300200
6 47 49 56 68 74 M
6.4 7.3 0.6 33.6 12.2 ND HDB
1.6 39.8 35.1 18.8 35.9 43.3 LDB
ca) gene in isolates. Upper numbers show isolates numbers. Bottom values (mg/L)etected.

Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662 661
promoter was active in L. lactis and horizontal transfers werepossible (Trip, Mulder, & Lolkema, 2012; Trip et al., 2011).
The expression of hdcA was increased only in stationary phasein the presence of histidine. hdcA gene expression was comparedwith histamine production under different durations and temper-atures. Compared to 4 �C, 10 times higher histamine concentrationwas observed at 42 �C after 24 h (Calles-Enriquez et al., 2010). Nomajor differences were found in hdcA expression levels under thesetemperature conditions, thus indicating that different factors couldbe implicated in the induction of the hdc cluster and histamineproduction (Trip et al., 2011). The hdcA enzyme was synthesisedin milk even in the absence of histidine. The enzyme activity incell-free extract obtained from cultures grown with or without his-tidine was not significantly different (Rossi et al., 2011).
Tabanelli, Torriani, Rossi, Rizzotti, and Gardini (2012) evaluatedfactors (pH, NaCl concentration, temperature, and lactose availabil-ity) influencing HdcA activity. The hdcA was mostly active at pH 4.5but reduced activities were observed at higher pH (pH 8.0). In thepresence of salt concentration up to 5% (w/v) no significant differ-ences were observed in comparison with the control HdcA group.For higher NaCl concentrations, the histamine amount decreased.The optimum thermal stability of hdcA of S. thermophilus was50 �C but increasing temperate reduces activity. The addition oflactose as carbon source caused only slight induction in hdcAexpression (Calles-Enriquez et al., 2010). The present molecularstudies indicated that the presence of hdcA gene was not correlatedwith histamine production. Occurrence of BA production should betaken into consideration rather than presence of genes.
4. Conclusion
The use of amine-negative starter cultures for fermented foodhas been suggested to control biogenic amines production. Bio-genic amine production capability of 58 S. thermophilus isolatesin yogurt was investigated. The results of the study showed thatmost of the S. thermophilus isolates have an ability to form biogenicamines, especially histamine and tyramine, which are the mosttoxic amines for human health. It is important to select suitablestrains as starter cultures.
References
Arena, M. E., & Manca de Nadra, M. C. (2001). Biogenic amine production byLactobacillus. Journal of Applied Microbiology, 90, 158–162.
Bodmer, S., Imark, C., & Kneubuhl, M. (1999). Biogenic amines in foods: Histamineand food processing. Inflammation Research, 48, 296–300.
Bover-Cid, S., & Holzapfel, W. H. (1999). Improved screening procedure for biogenicamine production by lactic acid bacteria. International Journal of FoodMicrobiology, 53, 33–41.
Bover-Cid, S., Hugas, M., Izquierdo-Pulido, M., & Vidal-Carou, M. C. (2001). Aminoacid-decarboxylase activity of bacteria isolated from fermented pork sausages.International Journal of Food Microbiology, 66, 185–189.
Bunkova, L., Bunka, F., Hlobilova, M., Vanatkova, Z., Novakova, D., & Drab, V. (2009).Tyramine production of technological important strains of Lactobacillus,Lactococcus and Streptococcus. European Food Research Technology, 229,533–538.
Bunkova, L., Bunka, F., Pollakova, E., Podesvova, T., & Drab, V. (2011). The effect oflactose, NaCl and an aero/anaerobic environment on the tyrosine decarboxylaseactivity of Lactococcus lactis subsp. cremoris and Lactococcus lactis subsp. lactis.International Journal of Food Microbiology, 147, 112–119.
Calles-Enriquez, M., Eriksen, B. H., Andersen, P. S., Rattray, F. P., Johansen, A. H.,Fernandez, M., et al. (2010). Sequencing and transcriptional analysis of theStreptococcus thermophilus histamine biosynthesis gene cluster: Factors thataffect differential hdcA expression. Applied and Environmental Microbiology, 76,6231–6238.
Coton, E., & Coton, M. (2005). Multiplex PCR for colony direct detection of gram-positive histamine- and tyramine-producing bacteria. Journal of MicrobiologicalMethods, 63, 296–304.
Coton, M., Romano, A., Spano, G., Ziegler, K., Vetrana, C., Desmarais, C., et al. (2010).Occurrence of biogenic amine-forming lactic acid bacteria in wine and cider.Food Microbiology, 27, 1078–1085.
de las Rivas, B., Rodriguez, H., Carrascosa, A. V., & Munoz, R. (2008). Molecularcloning and functional characterization of a histidine decarboxylase fromStaphylococcus capitis. Journal of Applied Microbiology, 104, 194–203.
Fernandez, M., Linares, D. M., Rodriguez, A., & Alvarez, M. A. (2007). Factors affectingtyramine production in Enterococcus durans IPLA 655. Applied Microbiology andBiotechnology, 73, 1400–1406.
Gao, P. F., Guo, X. F., Wang, H., & Zhang, H. S. (2011). Determination of trace biogenicamines with 1,3,5,7-tetramethyl-8-(N-hydroxysuccinimidyl butyric ester)-difluoroboradiaza-s-indacene derivatization using high-performance liquidchromatography and fluorescence detection. Journal of Separation Science, 34,1383–1390.
Garai, G., Duenas, M. T., Irastorza, A., & Moreno-Arribas, M. V. (2007). Biogenicamine production by lactic acid bacteria isolated from cider. Letters in AppliedMicrobiology, 45, 473–478.
Gardini, F., Martuscelli, M., Caruso, M. C., & Galgano, F. (2001). Effects of pH,temperature and NaCl concentration on the growth kinetics, proteolytic activityand biogenic amine production of Enterococcus faecalis. International Journal ofFood Microbiology, 64, 105–117.
Gezginc, Y. (2010). The investigation of potential usage in food industry in terms ofplasmid content and biogenic amine production of lactic acid bacteria isolated fromtraditional yoghurts. PhD Thesis. University of Kahramanmaras Sutcu Imam.
Greif, G., Greifova, M., & Karovicova, J. (2006). Effects of NaCl concentration andinitial pH value on biogenic amine formation dynamics by Enterobacter spp.bacteria in model conditions. Journal of Food and Nutrition Research, 45, 21–29.
Iyer, R., Tomar, S. K., & Singh, A. K. (2010). Response surface optimization of thecultivation conditions and medium components for the production of folate byStreptococcus thermophilus. Journal of Dairy Research, 77, 350–356.
Joosten, H. M. L. J., & van Boeckel, M. A. J. S. (1988). Conditions allowing theformation of biogenic amines in cheese: A study of the kinetics of histamineformation in an infected Gouda cheese. Netherlands Milk and Dairy Journal, 41,3–24.
Klausen, N. K., & Huss, H. H. (1987). Growth and histamine production byMorganella morganii under various temperature conditions. InternationalJournal of Food Microbiology, 5, 147–156.
Kuley, E., & Ozogul, F. (2011). Synergistic and antagonistic effect of lactic acidbacteria on tyramine production by food-borne pathogenic bacteria in tyrosinedecarboxylase broth. Food Chemistry, 127, 1163–1168.
La Gioia, F., Rizzotti, L., Rossi, F., Gardini, F., Tabanelli, G., & Torriani, S. (2011).Identification of a tyrosine decarboxylase gene (tdcA) in Streptococcusthermophilus 1TT45 and analysis of its expression and tyramine production inmilk. Applied and Environmental Microbiology, 77, 1140–1144.
Ladero, V., Fernandez, M., & Alvarez, M. A. (2009). Effect of post-ripening processingon the histamine and histamine-producing bacteria contents of differentcheeses. International Dairy Journal, 19, 759–762.
Ladero, V., Fernandez, M., Calles-Enriquez, M., Sanchez-Llana, E., Canedo, E., Martin,M. C., et al. (2012). Is the production of the biogenic amines tyramine andputrescine a species-level trait in enterococci? Food Microbiology, 30(1),132–138.
Ladero, V., Rattray, F. P., Mayo, B., Martin, M. C., Fernandez, M., & Alvarez, M. A.(2011). Putrescine producing Lactococcus lactis: sequencing and transcriptionalanalysis of the biosynthesis gene cluster. Applied Environmental Microbiology, 77,5507–5511.
Ladero, V., Sanchez-Llana, E., Fernandez, M., & Alvarez, M. A. (2011). Survival ofbiogenic amine-producing dairy LAB strains at pasteurization conditions.International Journal of Food Science and Technology, 46, 516–521.
Landete, J. M., Arena, M. E., Pardo, I., Manca de Nadra, M. C., & Ferrer, S. (2010). Therole of two families of bacterial enzymes in putrescine synthesis from agmatinevia agmatine deiminase. International Microbiology, 13, 169–177.
Landete, J. M., de Las Rivas, B., Marcobal, A., & Munoz, R. (2007). Molecular methodsfor the detection of biogenic amine-producing bacteria on foods. InternationalJournal of Food Microbiology, 117, 258–269.
Landete, J. M., Ferrer, S., & Pardo, I. (2005). Which lactic acid bacteria are responsiblefor histamine production in wine? Journal of Applied Microbiology, 99, 580–586.
Linares, D. M., del Río, B., Ladero, V., Martínez, N., Fernández, M., Martín, M. C., et al.(2012). Factors influencing biogenic amines accumulation in dairy products.Frontiers in Microbiology, 3, 180.
Linares, D. M., Martin, M. C., Ladero, V., Alvarez, M. A., & Fernandez, M. (2011).Biogenic amines in dairy products. Critical Review of Food Science and Nutrition,51, 691–703.
Lorencova, E., Bunkova, L., Matoulkova, D., Drab, V., Pleva, P., Kuban, V., et al. (2012).Production of biogenic amines by lactic acid bacteria and bifidobacteria isolatedfrom dairy products and beer. International Journal of Food Science andTechnology, 47, 2086–2091.
Moreno-Arribas, M. V., & Polo, M. C. (2008). Occurrence of lactic acid bacteriaand biogenic amines in biologically aged wines. Food Microbiology, 25,875–881.
Novella-Rodríguez, S., Veciana-Nogués, M. T., & Vidal-Carou, M. C. (2000). Biogenicamines and polyamines in milks and cheeses by ion-pair high performanceliquid chromatography. Journal of Agriculture and Food Chemistry, 48,5117–5123.
Omafuvbe, B. O., & Enyioha, L. C. (2011). Phenotypic identification and technologicalproperties of lactic acid bacteria isolated from selected commercial Nigerianbottled yoghurt. African Journal of Food Science, 5, 340–348.
Ozogul, F. (2004). Production of biogenic amines by Morganella morganii, Klebsiellapneumoniae and Hafnia alvei using a rapid HPLC method. European Food Researchand Technology, 219, 465–469.

662 Y. Gezginc et al. / Food Chemistry 138 (2013) 655–662
Pircher, A., Bauer, F., & Paulsen, P. (2007). Formation of cadaverine, histamine,putrescine and tyramine by bacteria isolated from meat, fermented sausagesand cheeses. European Food Research and Technology, 226, 225–231.
Premont, R. T., Gainetdinov, R. R., & Caron, M. G. (2001). Following the trace 2 ofelusive amines. Proceedings of National Academic Sciences of the United States ofAmerica, 98, 9474–9475.
Rhee, J. E., Kim, K. S., & Choi, S. H. (2005). CadC activates pH-dependent expressionof the Vibrio vulnificus cadBA operon at a distance through direct binding to anupstream region. Journal of Bacteriology, 187, 7870–7875.
Rossi, F., Gardini, F., Rizzotti, L., La Gioia, F., Tabanelli, G., & Torriani, S. (2011).Quantitative analysis of histidine decarboxylase gene (hdca) transcription andhistamine production by Streptococcus thermophilus PRI60 under conditionsrelevant to cheese making. Applied and Environmental Microbiology, 77,2817–2822.
Santos, W. C., Souza, M. R., Cerqueira, M. M. O. P., & Gloria, M. B. A. (2003). Bioactiveamines formation in milk by Lactococcus in the presence or not of rennet andNaCl at 20 and 32 �C. Food Chemistry, 81, 595–606.
Satomi, M., Furushita, M., Oikawa, H., Yoshikawa-Takahashi, M., & Yano, Y. (2008).Analysis of a 30 kbp plasmid encoding histidine decarboxylase gene inTetragenococcus halophilus isolated from fish sauce. International Journal ofFood Microbiology, 126, 202–209.
Sebastian, P., Herr, P., Fischer, U., & König, H. (2011). Molecular identification oflactic acid bacteria occurring in must and wine. South African Journal for Enologyand Viticulture, 32, 300–309.
Sieuwerts, S., Molenaar, D., van Hijum, S. A. F. T., Beerthuyzen, M., Stevens, M. J. A., &Janssen, P. W. M. (2010). Mixed-culture transcriptome analysis reveals themolecular basis of mixed-culture growth in Streptococcus thermophilus andLactobacillus bulgaricus. Applied and Environmental Microbiology, 76, 7775–7784.
Tabanelli, G., Torriani, S., Rossi, F., Rizzotti, L., & Gardini, F. (2012). Effect of chemico-physical parameters on the histidine decarboxylase (HdcA) enzymatic activityin Streptococcus thermophilus PRI60. Journal of Food Science, 77, 231–237.
Tabasco, R., Paarup, T., Janer, C., Pelaez, C., & Requena, T. (2007). Selectiveenumeration and identification of mixed cultures of Streptococcusthermophilus, Lactobacillus delbrueckii subsp. bulgaricus, L. acidophilus, L.paracasei subsp paracasei and Bifidobacterium lactis in fermented milk.International Dairy Journal, 17, 1107–1114.
Trip, H., Mulder, N. L., & Lolkema, J. S. (2012). Improved acid stress survival ofLactococcus lactis expressing the histidine decarboxylation pathway ofStreptococcus thermophilus CHCC1524. Journal of Biological Chemistry, 287,11195–11204.
Trip, H., Mulder, N. L., Rattray, F. P., & Lolkema, J. S. (2011). HdcB, a novel enzymecatalysing maturation of pyruvoyl-dependent histidine decarboxylase.Molecular Microbiology, 79, 861–871.
Valsamaki, K., Michaelidou, A., & Polychroniadou, A. (2000). Biogenic amineproduction in Feta cheese. Food Chemistry, 71, 259–266.
Vido, K., le Bars, D., Mistou, M. Y., Anglade, P., Gruss, A., & Gaudu, P. (2004).Proteome analyses of heme-dependent respiration in Lactococcus lactis:Involvement of the proteolytic system. Journal of Bacteriology, 186, 1648–1657.




















