
Control of grape powdery mildew with synthetic, biological ...

Biochemical alterations in grape infected with three
phytonematode species with emphasis on root-knot
nematode control.
Hosny H. KesbaZoology and Agricultural Nematology Department, Faculty of
Agriculture, Cairo University, Giza 12613, Egypt.
Abstract
Grape rootstocks, Flame seedless, Flame
seedless/Freedom and Freedom were reacted differently
to Meloidogyne incognita, Rotylenchylus reniformis and Tylenchulus
semipenetrans according to rootstock progenitor. Freedom
reduced significantly the nematode criteria and build
up. Humic and fulvic acids were tested at the rate of
2 and 4 ml/pot against the root-knot nematode. On
Freedom, all treatments reduced significantly
nematode build up and the higher dose was more
effective than the lower one. As a result of humic
and fulvic applications, the lipid peroxidation (MDA)
and H2O2 contents were significantly reduced after
treatments while the antioxidant compounds
glutathione (GSH) and ascorbic acid (AsA) contents
significantly increased when compared with check.
Antioxidant defense enzymes (ascorbate peroxidase
(APX), superoxide dismutase (SOD) and catalase (CAT))
showed significant increase in their activities.

Total phenol content and polyphenol oxidase (PPO)
were improved significantly in treated plants
compared to inoculated untreated check.
Keywords- Meloidogyne incognita, Rotylenchulus reniformis,
Tylenchulus semipenetrans, grape, humic acid, fulvic
acid, non enzymatic antioxidants, Antioxidant
enzymes.
Introduction
Plant resistance to nematode infection is one of
the most important agricultural measures in nematode
management strategies especially in perennial crops.
Under such circumstances, perennials are subjected to
invasion with progressive numbers of nematodes due to
the unbroken build up notably when environmental
conditions are in favor to nematode activities.
Accordingly, cultivation of resistant rootstocks or
inducing resistance in growing plants is a smart
choice to control nematode populations. On grapes,
the root-knot and the reniformis nematodes are highly

distructive to most grape varieties, however, they
are found to be poor hosts for the citrus nematode
(Kesba, 2003).
Resistant grape varieties or rootstocks to
different plant parasitic species had been studied by
many researchers (Walker et al., 1994; Walker, 1997;
Al-Sayed et al., 1999 & 2005; Anwar & McKenry, 2000 &
2001; McKenry et al., 2001).
Humic and fulvic acids are among the organic
acids that induce plant resistance to nematode
infection (Al-Sayed et al., 1988; Nandi et al., 2000;
Daneel et al., 2000; Zhang & Schmidt, 2000; Zaki et al.,
2004; Kesba & Al-Shalaby, 2008). They enable plants
to produce different compounds that improve the
physiological defense mechanism in infected plants e.g.
enzymatic and non-enzymatic compounds against
parasitic nematodes (Kesba & El-Beltagi, 2008), H2O2
(Waetzig et al., 1999; Peltzer et al., 2002), jasmonic
acids (Reymond & Farmer, 1998), reactive oxygen
(Neill et al., 2002; Montes et al., 2004; Murgia et al.,
2004), enzymatic and non-enzymatic ascorbate (Asada &
Takahashi, 1987; Asada, 1992; Jespersen et al., 1997;
Yoshimura et al., 2004), peroxidase and polyphenol
oxidase (Vaughn & Duke, 1984; Sarowar et al., 2005),
antioxidant (Halliwell, 1996; Reed, 1995; Decker,

1997), polyphenols (Buffle, 1988; Stackhouse &
Benson, 1988; Laughton et al., 1989; Metodiewa et al.,
1999).
Therefore, the objectives of this study were to:
(1) study the relationship between rootstock and
scion against three phytonematode genera and its
biochemical alterations. (2) evaluate the effect of
humic and fulvic acids on root-knot nematode, M.
incognita control and there role in enhancing plant
resistance against nematode infection.
Materials and Methods
Nematode species stock cultures
Pure cultures of the root-knot nematode, M.
incognita, the reniform nematode, R. reniformis and the
citrus nematode, T. semipenetrans were obtained from
isolates belonging to the Nematology Research Center,
Faculty of Agriculture, Cairo University. Nematode
species have been propagated separately, (M. incognita on
eggplant cv. Classic), (R. reniformis on pigeon pea) and
(T. semipenetrans on sour orange) grown in 20 cm diameter
clay pots filled with sterilized loamy soil. To avoid
contamination, cultures of each species were arranged
separately, examined and periodically renewed in

order to ensure continuous supplies of inocula for
the experimental work.
Glasshouse experiments
One year old seedlings of grape rootstocks, Flame
seedless, Flame seedless/Freedom and Freedom with
uniform size were obtained from Grape Department,
Horticulture Research Institute, Agriculture Research
Center and cultivated singly in 20 cm diameter clay
pots filled with steam sterilized sandy loam soil
(1:1, v/v). One month after, 5 seedlings of each
rootstock were inoculated separately with 5000
infective stages of M. incognita, R. reniformis or T.
semipenetrans by pipetting the nematode water suspension
into 4 holes around the root system and immediately
covered with soil. Pots were labeled and arranged
randomly on a glasshouse clean bench, receiving
similar horticulture treatments. Seedlings were left
out for 4 months after inoculation. Soil populations
were extracted by means of (Hooper et al., 2005) and
counted. The nematode embedded stages were also
counted.
For testing the effect of humic and fulvic acids
(extracting and preparation at By-product Research
Department, Agricultural Research Center, Ministry of
Agriculture) on the root-knot nematode development

and reproduction, another 5 seedlings of each
rootstock were inoculated with 5000 J2 of M.
incognita/pot. Two weeks after inoculation, two
commercial products of humic and fulvic acids were
applied at the rate of (2, 4 ml/plant) as soil
drench. All treatments were arranged in a fully
randomized design on a clean bench in the glasshouse
at 32 ±5 oC receiving similar horticultural
treatments. After 4 months, nematode soil populations
were extracted and counted using a Hawksley counting
slide, under a binocular microscope. A subsample (5
g) of roots from each plant was stained and gall
numbers, embedded stages (developmental stages +
eggmasses) per root were calculated, final population
(embedded stages + nematodes in soil), nematode build
up (Pf/Pi), average of eggs/eggmass were estimated.
Plant chemical analysis
Subsamples of fresh root of each treatment were
chemically analyzed at Central Chemistry Lab.,
Faculty of Agriculture Research Park (FARP), Faculty
of Agriculture, Cairo University as follows:
Determination of non-enzematic antioxidants
(oxidative burst)
Assay of lipid peroxidation (MDA) according to
Hodges et al., (1999), hydrogen peroxide (H2O2) by the

method described by Capaldi and Taylor (1983), total
glutathione (GSH) according to De Vos et al., (1992),
total ascorbic acid (TAA) according to Abdulnabi et
al., (1997), total phenols by the method of Zieslin
and Ben-Zaken (1993).
Determination of antioxidant defense enzymes activity
Assay of superoxide dismutase (SOD) using the
method of Beauchamp and Fridovich (1971), ascorbate
peroxidase (APX) using the method of (Nakano & Asada,
1981), catalase (CAT) using the method of Dhindsa et
al. (1981), polyphenol oxidase (PPO) according to
Coseteng and Lee (1987).
Statistical analysis
Data were compared by Duncan's Multiple Range Test
(DMRT) at the 5% level of probability using MSTAT
version 4 (1987).
Results
Development and reproduction of the root-knot, the
reniform and the citrus nematodes on grape rootstocks
Data in (Table 1) revealed that both the root-
knot nematode, M. incognita and the reniform nematode, R.
reniformis reproduced normally on the three rootstocks
with significant differences in their susceptibility.

Flame seedless was the most susceptible, followed by
Flame seedless/Freedom but Freedom showed some
resistance to both nematodes as measured by numbers
of galls, embedded stages, final population and the
rates of build up and egg deposition. The three
rootstocks are not good hosts to the citrus nematode,
T. semipenetrans on which the nematode folded hardly. It
is noticeable that nematode population declined on
Freedom. In all cases, grafting Flame seedless on
Freedom rootstock resulted in reducing the final
population of the root-knot nematode by 20%, the
reniform nematode by 57% and the citrus nematode by
31% as compared to Flame seedless.
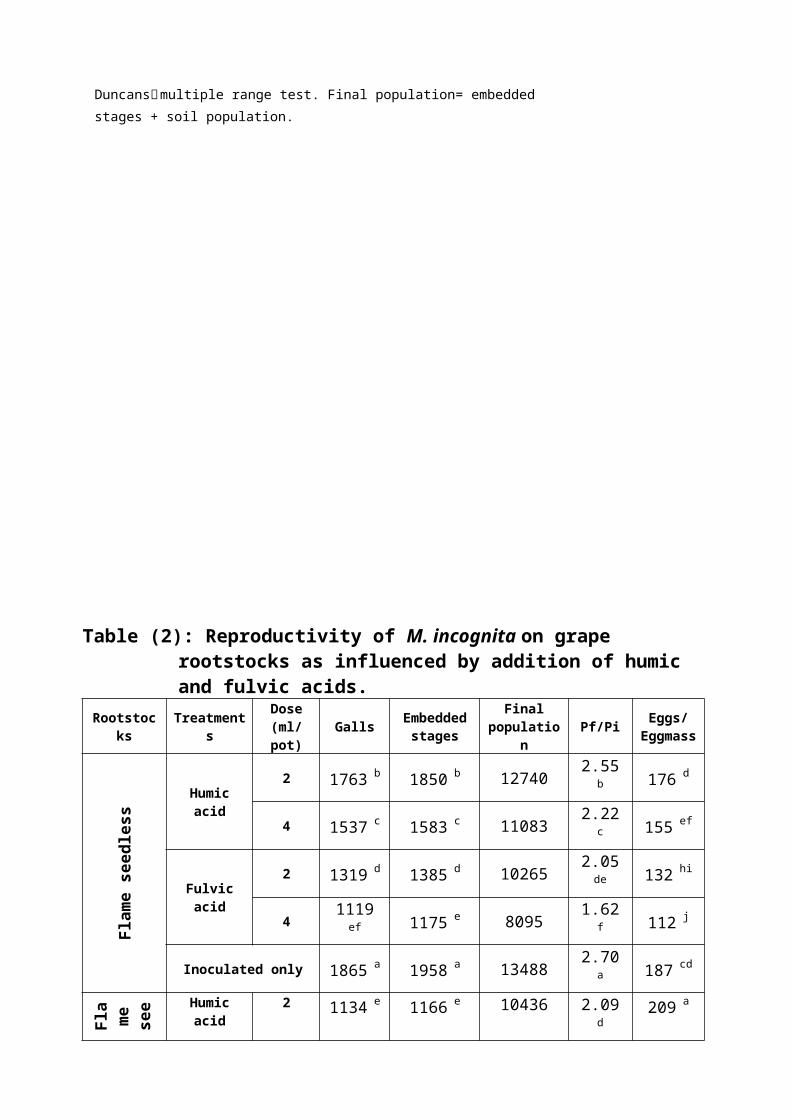
Effect of humic and fulvic acids on the development
and reproduction of M. incognita on grape
On Flame seedless, the susceptible rootstock,
both acids at the two concentrations significantly
reduced the number of galls and the embedded stages
on the infected roots as compared with the check. The
higher dose (4ml/pot) was significantly efficient
than the lower one. Similar results were observed
with the final population and the rate of nematode
build up as well as the number of eggs/eggmass.
Fulvic acid was significantly better than humic acid

in suppressing the root-knot nematode reproduction on
the susceptible rootstock (Table 2).
On Freedom, the less susceptible rootstock, an
opposite results were obtained whereas the
application of organic acids significantly increased
root galling and the number of embedded stages.
However, they reduced the final population, the rates
of build up and egg deposition. Also, significant
differences were observed between lower and higher
doses.
On Flame seedless/Freedom rootstock, the
increases in numbers of galls and the embedded stages
varied significantly but not drastically with the
check. Also, the reductions in the final population
and the rates of build up and egg deposition were not
satisfactory. No significant variations could be
observed between concentrations in the majority of
cases.
Results in (Table 3) manifested that single
infection of grape rootstocks with the root-knot, the
reniform and citrus nematodes resulted in increasing
root contents of both lipid peroxidation (MDA) and
H2O2 with significant differences within nematode
genera and between them and the uninoculated healthy
roots. The highest rates of increment were

implemented by M. incognita in all rootstocks and by all
nematode genera on Freedom.
Nematode infection also boosted the contents of
enzymatic antioxidant, total glutathione (GSH), total
ascorbic acid (TAA) and total phenols. It is clear
that the highest rates of enhancement of enzymatic
antioxidant were observed in roots infected with the
reniform nematode followed by those infected with the
citrus nematode, however, the rates of increase in
the roots infected with the root-knot nematode ranked
statistically in the third group. The utmost increase
rates were observed in roots of Freedom.
Concerning the influence of organic acids on the
contents of roots of lipid peroxidation, results
showed that both humic and fulvic acids significantly
reduced MDA and H2O2 in grape roots infected with M.
incognita as compared to non treated infected roots
(Table 4). With higher concentration, the greater
reduction in both MDA and H2O2 were observed with
humic acid which is efficient than fulvic acid with
Flame seedless and Flame seedless/Freedom rootstocks
and vice versa with Freedom. Both acids succeeded to
reduce the contents of lipid peroxidation on Freedom
to be almost near that in healthy plants.

All treatments significantly improved the levels
of non-enzymatic antioxidant molecules in the roots
of infected rootstocks especially at the higher
concentrations. Humic acid also achieved the best
results in the majority of cases. Also, all
treatments increased the levels of antioxidant
enzymes. Higher levels of the antioxidant enzymes
were recorded in roots of plants treated with the
higher dose. The highest significant increase in APX
was observed in roots of Flame seedless/Freedom
treated with humic acid at the high dose. With SOD
the highest significant increase was achieved by the
high dose of humic acid in roots of Freedom. However,
the highest significant increase in CAT and PPO was
accomplished by fulvic acid (the high dose) in roots
of Flame seedless and Flame seedless/Freedom
rootstocks, respectively.
Discussion
The present results proved that the nematode
species M. incognita, R. reniformis and T. semipenetrans
developed and reproduced variably on the tested
rootstocks. Flame seedless was the most susceptible
followed by Flame seedless/Freedom but Freedom showed
some resistance. Variability in the development and

reproduction of different nematode species on
different grape rootstocks had been documented by
many researcher workers (Saur, 1977; Walker et al.,
1994; Mckenry et al., 2001; Al-Sayed et al., 2005).
Both humic and fulvic acids significantly reduced
the numbers of the root-knot nematode in soil and on
roots of the susceptible rootstock, Flame seedless
and fulvic acid was more suppressive than humic. The
role of organic acids in suppressing the plant
parasitic nematodes populations was reported by Al-
Sayed et al., 1988; Al-Sayed et al., 2007 and Kesba and
El-Beltagi, 2008.
An opposite results were obtained in case of the
less susceptible rootstock, Freedom whereas treatment
with organic acids increased root galling and root
population, but suppressed soil and final population.
On Flame seedless/Freedom rootstock both acids
resulted in moderate increase in root populations,
and unsatisfactory reduction in the final population.
It seems that humic and fulvic acids somehow affect
the nematode fecundity and showed potential toxic
action against the free living stages in the soil.
The increase of proteins and fatty acids in root
tissues as a result of treating plants with organic
acids may enhance some biochemical compounds able to

retard nematode reproduction (Al-Sayed et al., 2005;
Kesba & Al-Shalaby, 2008).
Incompatible resistant interactions between
plant and pathogen are often determined by the
formation of reactive oxygen species (ROS) by the
pathogen (Baker & Orlandi, 1995; Montes et al., 2004).
Reactive oxygen species (ROS), such as hydrogen
peroxide (H2O2), is some of the most damaging
stressors in plants. Thus, H2O2 from the oxidative
stress plays a key role in the orchestration of a
localized hypersensitive response during the
expression of plant disease resistance (Levine et al.,
1994). ROS induced lipid peroxidation may be one of
the mechanisms accounting for cell death (Jabs,
1999). The results agreed with those facts and showed
that, with comparison to healthy plants of grape
rootstocks (Flame seedless, Flame seedless/Freedom
and Freedom) infection with (M. incognita or R. reniforms)
enhanced the content of MDA and H2O2. The results also
indicated that plants possess both enzymatic and non-
enzymatic antioxidant defense systems to counteract
ROS under nematode infections. The significant
increase of non-enzymatic antioxidant such as (GSH,
TAA and TPH) contents may be driven by enhancement of
MDA and H2O2 formation in the nematode infections.

However, GSH may play a protective role in scavenging
of singlet oxygen, peroxides and hydroxyl radicals
and is involved in recycling reduction of ascorbic
acid (AsA) in the ascorbate-glutathione cycle (Foyer,
1993). On the other hand, the significant increase in
the total soluble phenols (TPH) contents could be
affected as strong antioxidant natural products
induced under oxidative stress condition controlling
the oxidative damage. These data was in accordance
with (Goodman et al., 1967), who found that, multifold
increase of phenols after challenging with
elicitation may be due to the excess production of
H2O2 in elicited plant cells through increased
respiration (Farkas & Kiraly, 1962) or due to the
activation of hexose-monophosphate pathway, acetate
pathway and release of bound phenols by hydrolytic
enzymes. In addition, the nematode infection caused
marked increase in the activity of antioxidant
enzymes (APX, SOD, and CAT) which are involved in
scavenging excess ROS in plant cells (Yoshimura et al.,
2004). Catalase and Ascorbate peroxidase play an
essential role in scavenging from the H2O2 toxicity.
The combined action of CAT and SOD converts the toxic
superoxide radical (O2.−) and hydrogen peroxide (H2O2)
to water and molecular oxygen (O2), thus averting the

cellular damage under unfavorable conditions like
infection by nematodes (Asada, 1992; Zacheo & Bleve-
Zacheo, 1988). While the increase in polyphenol
oxidase (PPO) activity after nematodes infection may
be due to autooxidation of the total phenol substrate
which interact with H2O2 and may prevent nematode
spread to healthy tissue (Trudgill, 1991).
Treatments infected with M. incognita reduced the
contents of MDA and H2O2 in all rootstocks by
improving the contents of non-enzymatic antioxidants
(GSH, TAA and TPH) and increase the activities of PPO
and antioxidants enzymes (APX, SOD and CAT) and
reached its maximum induction at the higher dose (4
ml). In addition, our results indicated that the high
dose (4 ml) at Flame seedless/Freedom rootstock was
the best treatment that suitable plant treatments to
reduce the ROS, lipid peroxidation formation and
improve the induction of antioxidant defense system.
It could be conclude that humic and fulvic acids
treatments may play an important role in antioxidant
defense system of plants, it is supposed that low
concentration might be a signal to induce the
expression of many antioxidative molecules and
enzymes and reduce the ROS in plant cells (Karasyova
et al., 2007).

ReferencesAbdulnabi, A.A., Emhemed, A.H., Hussein, G.D. and Biacs, P.A.
(1997). Determination of antioxidant vitamins in tomato.Food Chemistry, 60: 207-212.
Al-Sayed, A.A., Ahmed, S.S. and Montasser, S.A. (1988). Effects of some organic acids on egg hatchability of Meloidogyne incognita and Rotylenchulus reniformis. Annals of Agricultural Science, Moshtohor, Benha University, 26: 1325-1332.
Al-Sayed, A.A., Kheir, A.M., El-Naggar, H.I. and Kesba, H.H.(1999). Varietal reaction of some grape Rootstocks tothe infection with three plant parasitic nematodespecies. Journal of Agricultural Sciences, MansouraUniversity, 24: 7707-7718.
Al-Sayed, A.A., Kheir, A.M., El-Naggar, H.I. and Kesba, H.H. (2007). Organic management of Meloidogyne incognita on grapesin relation to host biochemistry. International Journal of Agricultural Research, 2: 776-785.
Al-Sayed, A.A., Kheir, A.M., El-Naggar, H.I. and Kesba, H.H. (2005). Could other Vitis species be helpful in nematode management in Egypt's sand soil viticultures? Bulletin of Faculty of Agriculture, Cairo University, 56: 393-406.
Anwar, S.A. and Mckenry, M.V. (2000). Penetration, developmentand reproduction of Meloidogyne arenaria on two new resistantVitis spp. Nematropica, 30: 9-17.
Anwar, S.A. and Mckenry, M.V. (2001). Susceptible ofgenetically transformed Freedom grape rootstocks toMeloidogyne arenaria Pt. Freedom. International Journal ofNematology, 11: 1-7.
Asada, K. (1992). Ascorbate peroxidase: a hydrogen peroxide-scavenging enzyme in plants. Physiologia Plantarum, 85: 235–241.
Asada, K. and Takahashi, M. (1987) Production and scavenging of active oxygen in photosynthesis. In: D. J. Kyle, C. B. Osmond and C. J. Arntzen (Eds.). Photoinhibition. Elsevier Science Publishers, Amsterdam, pp. 227-287.
Baker, C.J. and Orlandi, E.W. (1995). Active oxygen in plant pathogenesis. Annual Review of Phytopatholology, 33: 299–321.

Beauchamp, C. and Fridovich, I. (1971). Superoxide dismutase: improved assays and assay applicable to acrylamide gels.Analytcial Biochemistry, 44: 276–287.
Buffle, J. (1988). Complexation reactions in aquatic systems. Ellis Horwood Series, In Analytical Chemistry. Ellis Horwood Limited, Chichester, England.
Capaldi, D.J. and Taylor, K.E. (1983). A new peroxidase color reaction: oxidative coupling of 3-methyl-2-benzothiazolinone hydrazone (MBTH) with its formaldehydeazine application to glucose and choline oxidases. Analytcial Biochemistry, 129: 329–336.
Coseteng, M.Y. and Lee, C.Y. (1987). Change in apple polyphenol oxidase and polyphenol concentrations in relation to degree of browning. Journal of Food Science,52: 985-989.
Daneel, M.S., De Jager, K., Dreyer, S., Dekker, J. and Joubert, J.P. (2000). The influence of oxihumate on nematode control and on yield (Musa AAA, Cavendish subgroup). Acta Horticulture, 540: 441-452.
De Vos, C.H., Vonk, M.J., Vooijs, R. and Henk, S. (1992). Glutathione depletion due to copper-induced phytochelatin synthesis causes oxidative stress in Silene cucbalus. Plant Physiology, 98: 859-858.
Decker, E.A. (1997). Phenolics: prooxidants or antioxidants? Nutrition Review, 55: 396–398.
Dhindsa, R.S., Plumb-Dhindsa, P. and Thorpe, T.A. (1981). Leafsenescence: correlated with increased levels of membranepermeability and lipid peroxidation, and decrease levelsof superoxide dismutase and catalase. Journal of Experimental Botany, 32: 93–101.
Farkas, G. L. and Kiraly, Z. (1962). Role of phenolic compounds in the physiology of plant disease resistance.Phytopathology, 44: 105–150.
Foyer, C.H. (1993). Ascorbic acid. In: Alscher, R.G. and Hess, J.L. (Eds.). Antioxidants in higher plants. CRC press, Florida, USA. pp: 31-58.
Goodman, R. N., Kiraly, E. and Zaitlin, M. (1967). The biochemistry and physiology of infections in plant diseases. D.Van Nostri and Co., Princeton, N.J. , Inc. p. 534
Halliwell, B. (1996). Antioxidants in human health and disease. Annual Review of Nutrition, 16: 33–50.
Hodges, D.M., DeLong, J.M., Forney, C.F. and Prange, R.K. (1999). Improving the thiobarbituric acid-reactive-

substances assay for estimating lipid per-oxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 207: 604–611.
Hooper, D.J., Hallmann, J. and Subbotin, S.A. (2005). Methods for extraction, processing and detection of plant and soil nematodes. In: Luc, M., Sikora, R.A. and Bridge, J.(Eds). Plant parasitic nematodes in subtropical and tropical agriculture. Wallingford, UK, CABI Publishing, pp. 53-86.
Jabs, T. (1999). Reactive oxygen intermediates as mediators ofprogrammed cell death in plants and animals. BiochemicalPharmacology, 57: 231-245.
Jespersen, H.M., Kjaersgard, I.V.H., Ostergaard, L. and Welinder, K.G. (1997). From sequence analysis of three novel ascorbate peroxidases from Arabidopsis thaliana to structure, function and evolution of seven types of ascorbate peroxidase. Biochemical Journal, 326: 305–310.
Karasyova, T.A., Klose, E.O., Menzel, R. and Steinberg, C.E. (2007). Natural organic matter differently modulates growth of two closely related coccal green algal species. Environmental Science and Pollution Research – International, 14: 88-93.
Kesba, H.H. (2003). Integrated nematode management on grapes grown in sandiness soil. PhD Thesis, Faculty of Agriculture, Cairo University, 189 pp.
Kesba, H.H. and Al-Shalaby, Mona E.M. (2008). Survival and reproduction of Meloidogyne incognita on tomato as affected by humic acid. Nematology, 10: 243 – 249.
Kesba, H.H. and El-Beltagi, H.E.S. (2008). Biochemical changesin grape rootstocks resulted from humic acid treatments in relation to nematode infection. Egyptian Journal of Applied Sciences, Zagazig University, 23: 277-292.
Laughton, M.J., Halliwell, B., Evans, P.J. and Hoult, J.R.S. (1989). Antioxidant and prooxidant actions of the plant phenolics quercetin, gossypol and myricetin. BiochemicalPharmacology, 38: 2859–2865.
Levine, A., Tenhaken, R., Dixon, R. and Lamb, C. (1994). H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell, 79: 583-593.
Mckenry, M.V., Kretsch, J.O. and Anwar, S.A. (2001).Interactions of selected Vitis Rootstocks with endo-parasitic nematodes. American Journal of Enology andViticulture, 52: 310-316.

Metodiewa, D., Jaiswal, A.K., Cenas, N., Dickancait´e, E. and Segura-Aguilar, J. (1999). Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radical Biology and Medicine, 26: 107–116.
Montes, M.J., López-Braña, I. and Delibes, A. (2004). Root enzyme activities associated with resistance to Heteroderaavenae conferred by gene Cre7 in a wheat/ Aegilops triuncialis introgression line. Journal of Plant Physiolology, 161: 493–495.
MSTAT Version 4 (1987). Software program for the design and analysis of agronomic research experiments. Michigan, USA, Michigan State University.
Murgia, I., Tarantino, D., Vannini, C., Bracale, M., Carravieri, S. and Soave, C. (2004). Arabidopsis thaliana plants overexpressing thylakoidal ascorbate peroxidase show increased resistance to paraquat-induced photooxidative stress and to nitric oxide-induced cell death. The Plant Journal, 38: 940–953.
Nakano, Y. and Asada, K. (1981). Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant and Cell Physiology, 22: 867–880.
Nandi, B., Sukul, N.C. & Babu, S.P.S. (2000). Exogenous salicylic acid reduces Meloidogyne incognita infestation of tomato. Allelopathy Journal, 7: 285-288.
Neill, S., Desikan, R. and Hancock, J. (2002). Hydrogenperoxide signaling. Current Opinion in Plant Biology, 5:388–395.
Peltzer, D., Dreyer, E. and Polle, A. (2002). Differential temperature dependencies of antioxidative enzymes in twocontrasting species: Fagus sylvatica and Coleus blumei. Plant Physiology and Biochemistry, 40: 141–150.
Reed, J.D. (1995). Nutritional toxicology of tannins and related polyphenols in forage legumes. Journal of AnimalScience, 73: 1516–1528.
Reymond, P. and Farmer, E.E. (1998). Jasmonate and salicyclateas global signals for defense gene expression. Current Opinion in Plant Biology, 1: 404–411.
Sarowar S., Kim E.N., Young, J.K., Sung, H.O., Ki, D.K., Byung, K.H. and Jeong, S. (2005). Overexpression of a pepper ascorbate peroxidase-like1 gene in tobacco plantsenhances tolerance to oxidative stress and pathogens. Plant Science, 169: 55–63.

Saur, M.R. (1977). Nematode resistant grape rootstocks.Australian Dried Fruits News, 5: 10-14.
Stackhouse, R.A. and Benson, W.H. (1988). The influence of humic acid on the toxicity and bioavailability of selected trace metals. Aquatic Toxicology, 13: 99–108.
Trudgill, D.L. (1991). Resistance to and tolerance of plant parasitic nematodes in plants. Annual Review of Phytopathology, 29: 167–192.
Vaughn, K.C. and Duke, S.O. (1984). Function of polyphenol oxidase in higher plants. Physiologia Plantarum, 60: 106–112.
Waetzig, G.H., Sobczak, M. and Grundler, F.M.W. (1999). Localization of hydrogen peroxide during the defence response of Arabidopsis thaliana against the plant parasitic nematode Heterodera glycines. Nematology, 1: 681– 686.
Walker, G.E. (1997). Effects of Meloidogyne spp and Rhizoctoniasolani on the growth of grapevine rootlings. Journal ofNematology, 29: 1-9.
Walker, M.A., Ferris, H. and Eyre, M. (1994). Resistance inVitis and Muscadinia species to Meloidogyne incognita. PlantDisease, 78: 1055-1058.
Yoshimura, K., Miyao, K., Gaber, A., Takeda, T., Kanaboshi, H., Miyasaka, H. and Shigeoka, S. (2004). Enhancement of stress tolerance in transgenic tobacco plants overexpressing Chlamydomonas glutathione peroxidase in chloroplasts or cytosol. Plant Journal, 37: 21–33.
Zacheo, G. and Bleve-Zacheo, T. (1988). Involvement of superoxide dismutases and superoxide radicals in the susceptibility and resistance of tomato plants to Meloidogyne incognita attack. Physiological and Molecular Plant Pathology, 32: 313–322.
Zaki, M.J., Javad, S., Abid, M., Khan, H. and Moinuddin, M. (2004). Evaluation of some chemicals against root-knot nematode, Meloidogyne incognita. International Journal of Biology and Biotechnology, 1: 613-618.
Zhang, X. and Schmidt, R.E. (2000). Hormone-containing products impact on antioxidant status of tall fescue and creeping bentgrass subjected to drought. Crop Science, 40: 1344-1349.
Zieslin, N. and Ben-Zaken, R. (1993). Peroxidase activity and presence of phenolic substances in peduncles of rose flowers. Plant Physiology and Biochemistry, 31: 333–340.
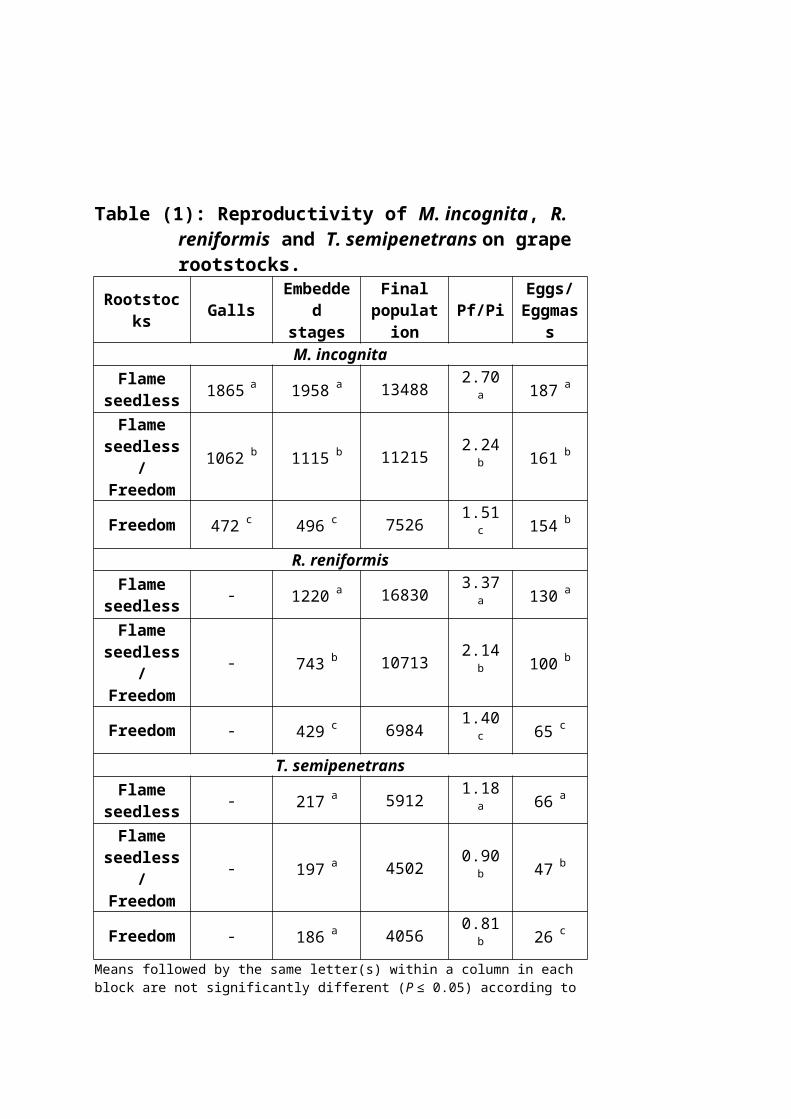
Table (1): Reproductivity of M. incognita, R. reniformis and T. semipenetrans on grape rootstocks.
Rootstocks Galls
Embedded
stages
Finalpopulation
Pf/PiEggs/Eggmas
sM. incognita
Flameseedless 1865 a 1958 a 13488 2.70
a 187 a
Flameseedless
/Freedom
1062 b 1115 b 11215 2.24b 161 b
Freedom 472 c 496 c 7526 1.51c 154 b
R. reniformisFlame
seedless - 1220 a 16830 3.37a 130 a
Flameseedless
/Freedom
- 743 b 10713 2.14b 100 b
Freedom - 429 c 6984 1.40c 65 c
T. semipenetransFlame
seedless - 217 a 5912 1.18a 66 a
Flameseedless
/Freedom
- 197 a 4502 0.90b 47 b
Freedom - 186 a 4056 0.81b 26 c
Means followed by the same letter(s) within a column in each block are not significantly different (P ≤ 0.05) according to
Duncans׳ multiple range test. Final population= embedded stages + soil population.
Table (2): Reproductivity of M. incognita on grape rootstocks as influenced by addition of humic and fulvic acids.
Rootstocks
Treatments
Dose(ml/pot)
Galls Embeddedstages
Finalpopulatio
nPf/Pi Eggs/
Eggmass
Flame
seedle
ss
Humicacid
2 1763 b 1850 b 12740 2.55b 176 d
4 1537 c 1583 c 11083 2.22c 155 ef
Fulvicacid
2 1319 d 1385 d 10265 2.05de 132 hi
41119ef 1175 e 8095 1.62
f 112 j
Inoculated only 1865 a 1958 a 13488 2.70a 187 cd
Fla
me see Humic
acid2 1134 e 1166 e 10436 2.09
d209 a
dles
s /
Freedo
m 41113efg 1149 ef 9969 1.99
de 201 ab
Fulvicacid
21092efg 1171 e 9771 1.95
e 193 bc
41071fg 1193 e 9573 1.92
e 185 cd
Inoculated only 1062 g 1115 f 11215 2.24c 161 e
Freedom
Humicacid
2 684 h 958 g 7288 1.46g 139 gh
4 590 i 767 h 6387 1.28h 123 ij
Fulvicacid
2 660 h 792 h 5712 1.14i 146 fg
4 566 i 623 i 5193 1.04i 116 j
Inoculated only 472 j 496 j 7526 1.51f 154 ef
Means followed by the same letter(s) within a column in each block are not
significantly different (P ≤ 0.05) according to Duncans׳ multiple range test. Final population= embedded stages + soil population.
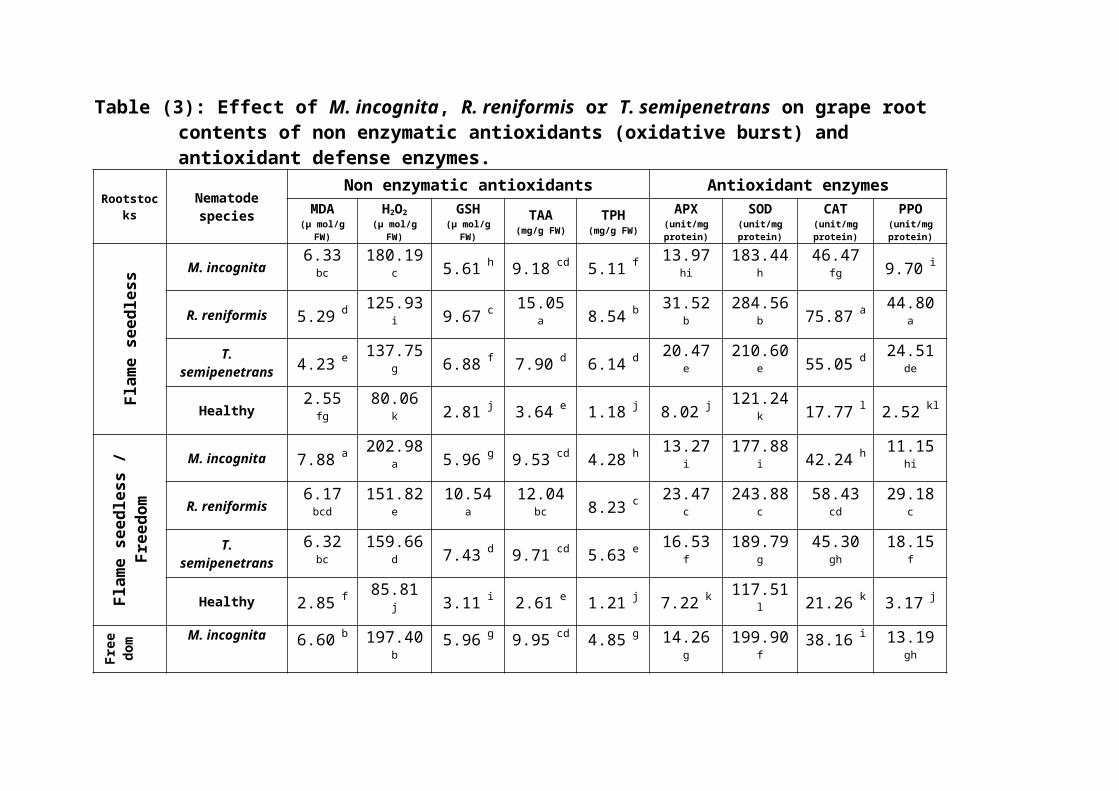
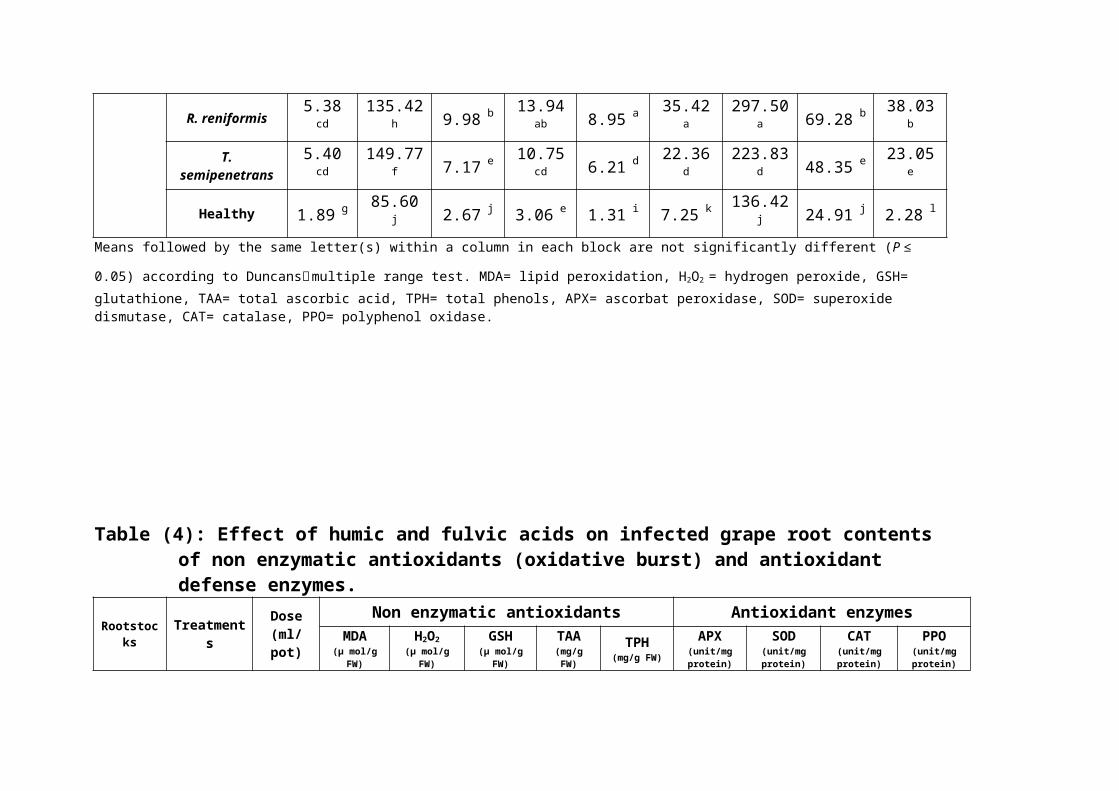
Table (3): Effect of M. incognita, R. reniformis or T. semipenetrans on grape rootcontents of non enzymatic antioxidants (oxidative burst) and antioxidant defense enzymes.
Rootstocks
Nematodespecies
Non enzymatic antioxidants Antioxidant enzymesMDA
(μ mol/gFW)
H2O2(μ mol/g
FW)
GSH(μ mol/g
FW)
TAA(mg/g FW)
TPH(mg/g FW)
APX(unit/mgprotein)
SOD(unit/mgprotein)
CAT(unit/mgprotein)
PPO(unit/mgprotein)
Flame se
edless
M. incognita6.33bc
180.19c 5.61 h 9.18 cd 5.11 f
13.97hi
183.44h
46.47fg 9.70 i
R. reniformis 5.29 d125.93
i 9.67 c15.05
a 8.54 b31.52
b284.56
b 75.87 a44.80
a
T.semipenetrans 4.23 e
137.75g 6.88 f 7.90 d 6.14 d
20.47e
210.60e 55.05 d
24.51de
Healthy2.55fg
80.06k 2.81 j 3.64 e 1.18 j 8.02 j
121.24k 17.77 l 2.52 kl
Flame
seed
less /
Free
dom
M. incognita 7.88 a202.98
a 5.96 g 9.53 cd 4.28 h13.27
i177.88
i 42.24 h11.15
hi
R. reniformis6.17bcd
151.82e
10.54a
12.04bc 8.23 c
23.47c
243.88c
58.43cd
29.18c
T.semipenetrans
6.32bc
159.66d 7.43 d 9.71 cd 5.63 e
16.53f
189.79g
45.30gh
18.15f
Healthy 2.85 f85.81
j 3.11 i 2.61 e 1.21 j 7.22 k117.51
l 21.26 k 3.17 j
Free
dom M. incognita 6.60 b 197.40
b5.96 g 9.95 cd 4.85 g 14.26
g199.90
f38.16 i 13.19
gh
R. reniformis5.38cd
135.42h 9.98 b
13.94ab 8.95 a
35.42a
297.50a 69.28 b
38.03b
T.semipenetrans
5.40cd
149.77f 7.17 e
10.75cd 6.21 d
22.36d
223.83d 48.35 e
23.05e
Healthy 1.89 g85.60
j 2.67 j 3.06 e 1.31 i 7.25 k136.42
j 24.91 j 2.28 l
Means followed by the same letter(s) within a column in each block are not significantly different (P ≤
0.05) according to Duncans׳ multiple range test. MDA= lipid peroxidation, H2O2 = hydrogen peroxide, GSH= glutathione, TAA= total ascorbic acid, TPH= total phenols, APX= ascorbat peroxidase, SOD= superoxide dismutase, CAT= catalase, PPO= polyphenol oxidase.
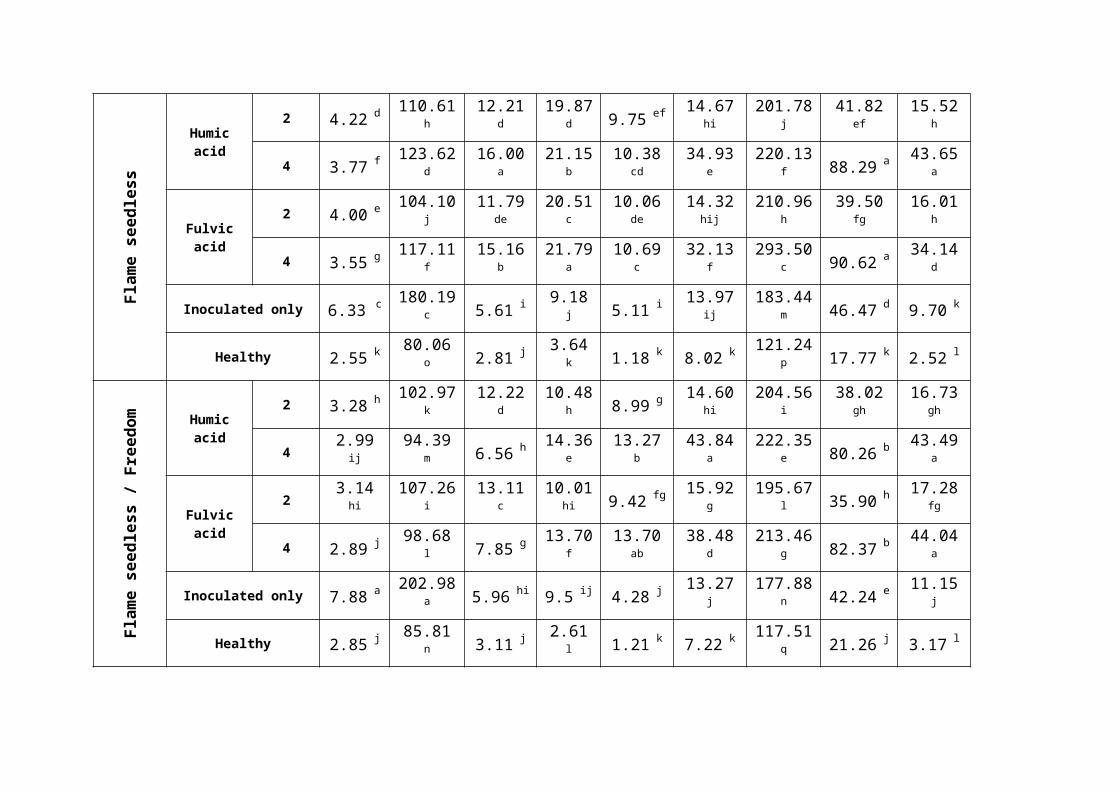
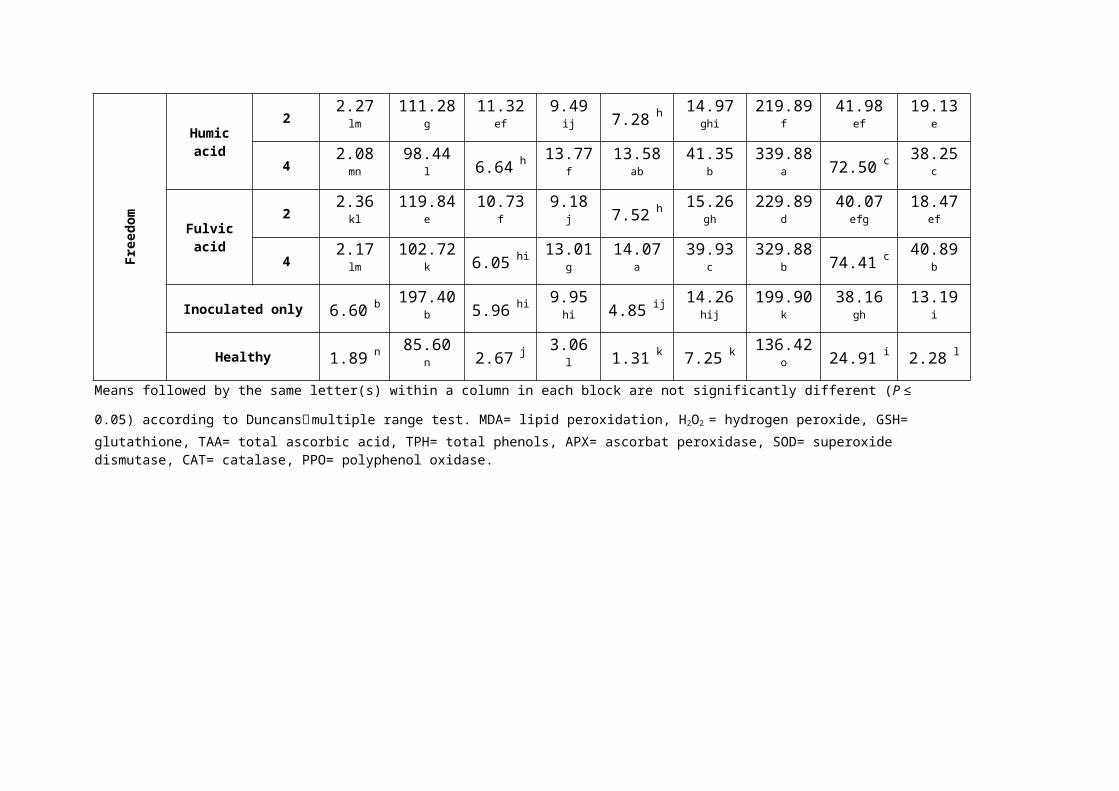
Table (4): Effect of humic and fulvic acids on infected grape root contentsof non enzymatic antioxidants (oxidative burst) and antioxidant defense enzymes.
Rootstocks
Treatments
Dose(ml/pot)
Non enzymatic antioxidants Antioxidant enzymesMDA
(μ mol/gFW)
H2O2(μ mol/g
FW)
GSH(μ mol/g
FW)
TAA(mg/gFW)
TPH(mg/g FW)
APX(unit/mgprotein)
SOD(unit/mgprotein)
CAT(unit/mgprotein)
PPO(unit/mgprotein)
Flam
e seedless
Humicacid
2 4.22 d110.61
h12.21
d19.87
d 9.75 ef14.67
hi201.78
j41.82
ef15.52
h
4 3.77 f123.62
d16.00
a21.15
b10.38
cd34.93
e220.13
f 88.29 a43.65
a
Fulvicacid
2 4.00 e104.10
j11.79
de20.51
c10.06
de14.32
hij210.96
h39.50
fg16.01
h
4 3.55 g117.11
f15.16
b21.79
a10.69
c32.13
f293.50
c 90.62 a34.14
d
Inoculated only 6.33 c180.19
c 5.61 i9.18
j 5.11 i13.97
ij183.44
m 46.47 d 9.70 k
Healthy 2.55 k80.06
o 2.81 j3.64
k 1.18 k 8.02 k121.24
p 17.77 k 2.52 l
Flam
e seed
less
/ Fre
edom Humic
acid
2 3.28 h102.97
k12.22
d10.48
h 8.99 g14.60
hi204.56
i38.02
gh16.73
gh
42.99ij
94.39m 6.56 h
14.36e
13.27b
43.84a
222.35e 80.26 b
43.49a
Fulvicacid
23.14hi
107.26i
13.11c
10.01hi 9.42 fg
15.92g
195.67l 35.90 h
17.28fg
4 2.89 j98.68
l 7.85 g13.70
f13.70
ab38.48
d213.46
g 82.37 b44.04
a
Inoculated only 7.88 a202.98
a 5.96 hi 9.5 ij 4.28 j13.27
j177.88
n 42.24 e11.15
j
Healthy 2.85 j85.81
n 3.11 j2.61
l 1.21 k 7.22 k117.51
q 21.26 j 3.17 l
Freedom
Humicacid
22.27lm
111.28g
11.32ef
9.49ij 7.28 h
14.97ghi
219.89f
41.98ef
19.13e
42.08mn
98.44l 6.64 h
13.77f
13.58ab
41.35b
339.88a 72.50 c
38.25c
Fulvicacid
22.36kl
119.84e
10.73f
9.18j 7.52 h
15.26gh
229.89d
40.07efg
18.47ef
42.17lm
102.72k 6.05 hi
13.01g
14.07a
39.93c
329.88b 74.41 c
40.89b
Inoculated only 6.60 b197.40
b 5.96 hi9.95hi 4.85 ij
14.26hij
199.90k
38.16gh
13.19i
Healthy 1.89 n85.60
n 2.67 j3.06
l 1.31 k 7.25 k136.42
o 24.91 i 2.28 l
Means followed by the same letter(s) within a column in each block are not significantly different (P ≤
0.05) according to Duncans׳ multiple range test. MDA= lipid peroxidation, H2O2 = hydrogen peroxide, GSH= glutathione, TAA= total ascorbic acid, TPH= total phenols, APX= ascorbat peroxidase, SOD= superoxide dismutase, CAT= catalase, PPO= polyphenol oxidase.

ودا مات�� ي حة� ن�� مكاف�� ات� م�ع الاه�ي�مام ب�� ب� ودا ال�ن� مات�� ي واع م�ن� ن�� ت#� ة� ا$ لاث&% ب% ة� ن�� ص�اث�� ة� الا* ح� ي ن, ي ال�عن�ب� ن�- ة�5 ف� ي ن&$ ا وك�مب ي يرات� ال�ن; غ� ال�ت�ور ذ� ذ ال�ج� عق� ت��
ة ي ح�ام�ذ ك�سي� ح�سن�ة� راع�ي ا ال�ز� ب ول�وح�� مات�� ي وان� وال�ن� سم ال�حي راعة� –ق�� ة� ال�ز� اه�رة�– ك�لي ام�غة� ال�ق� ح��
ذم �زي& ذلس م�طعوم ع�لي ف� م س�ب لي ذم وف�� �زي& ص�ل ف� ذلس والا$ م س�ب لي ف� ف�� ة� ك�ل م�ن� ال�صت� ار م�ذى ح�ساس�ي ب� ت� م اح�� ت��ودا ال�موال�ح ) مات�� ي ة� ن�� ودا ال�كلوث� مات�� ي ور وال�ن� ذ� ذ ال�ج� عق� ودا ت�� مات�� ي كل م�ن� ن�� ة� ب�� ص�اث�� عل5000ل�لا* ( ورد ف�� ات� ب� /ن�� ط�ور م�عذى
معذل ك� )ب�� ب ول�ف� وال�ف� وم�ك� ا$ ة� ك�ل م�ن� ح�ام�ض� ال�هي اف� ص�� ر ا* ي ث&% أ$ ل�ك� ي�� ، وك�ذ� ة� ص�اث�� أت� ل�لا* اي&� ب� ي ص�ورة�4 و 2ال�ن� ( ف� ات� ب� م�ل/ن��ور. ذ� ذ ال�ج� عق� ودا ت�� مات�� ي أت� ل�ن� اي&� ب� اومة� ال�ن� ن� م�ق� حسي ة ع�لي ت�� ذرث�� ة� م�ذى ف�� لة� ل�معرف� س�اي�$
ودا مات�� ي ة� ل�ن� كوث�� ذ ال�مب� ع�ذاد ال�عق� ل ا$ لب ق� ذم ت�� �زي& ص�ل ف� ًأ الا$ وي� طاع م�عي� : اس�ت� ها هي حصل ع�لي ح� ال�مي, ات&$ ب� ه�م ال�ن- وك�ان� م�ن� ا$لة� ل ك�ب� ض� داح�� ت وس�ط ع�ذد ال�ن; ل�ك� م�ي� ر وك�ذ� كاث�% ة� وم�عذل ال�ت� ع�ذاد ال�كلي ة� والا$ لف� ت� ط�وار ال�مح� ع�ذاد الا$ ل ا$ لب ق� ور وت�� ذ� ذ ال�ج� عق� ت��
ذم. �زي& ذلس م�طعوم ع�لي ف� م س�ب لي وال�ق� ذلس ا$ م س�ب لي ف� ف�� كل م�ن� ال�صت� ار ب�� ب� ت� ودا م�جل الاح�� مات�� ي واع ال�ن� ت#� ع ا$ مت ض� ل�ج� ت ال�ن;ور م�ن� ك�ل م�ن� ذ� وى ال�ج� أدة� م�حي, ي& ة� ل�ز� ص�اث�� دت� الا* ذ ا$ دت�H2O2 و MDAوف�� . ك�ما ا$ مة� أت� ال�سلي اي&� ب� أل�ن� ة� ي�� ارث�� ة� م�ق� وث� أدة� م�عي� ي& ر�
ور م�ن� ك�ل م�ن� ذ� وى ال�ج� أدة� م�حي, ي& ة� ل�ز� ص�اث�� اط TPH و TAA وGSHالا* س% أدة� ن�� ي& antioxidant enzymes ور�
.PPOو CAT وSOD و APXل�كل م�ن� ب% أت� ح�ن اي&� ب� اومة� ال�ن� ن� م�ق� حسي ي ت�� ك�5 دورًا ف� ب ول�ف� و ح�ام�ض� ال�ف� وم�ك� ا$ كل م�ن� ح�ام�ض� ال�هي وك�ان��ت� ل�لمعام�لة� ب��
ع�لي ) ر� الا$ اصة� الي�رك�ي دت� ال�معام�لات� وح�� اصة� ع�لي4ا$ ة� ح�� وث� ة� م�عي� ذرج� ور ي�� ذ� ذ ال�ج� عق� ودا ت�� مات�� ي ع�ذاد ن�� ل ا$ لب ق� ي ت�� ( ف� ات� ب� م�ل/ن��ور م�ن� ذ� وى ال�ج� ض� م�حي, ف� لى خ�� ة� ع�امة� ا* صف� دت� ال�معام�لات� ب�� ل�ك� ا$ ي�رول. ك�ذ� أل�معام�لة� ال�كن- ها ي�� ي� ارن�- ذ م�ق� ذم ع�ب� �زي& ص�ل ف� الا$
MDA و H2O2ودا مات�� ي ادات� ال�ن� وى م�ن� م�ص� أدة� ال�محي, ي& ط(. ور� ف� ة� ف�� أت� ال�مصاث�� اي&� ب� ي�رول )ال�ن� أل�كن- ة� ي�� ارث�� ة� م�ق� وث� ة� م�عي� ذرج� ي��

GSHو TAAو TPH اط س% أدة� ن�� ي& PPOوCAT و SOD و APX ل�كل م�ن� antioxidant enzymes ور�
.) ر م�عام�لة� ي ة� وال�غ� أت� ال�مصاث�� اي&� ب� ر م�عام�لة� وال�ن� ي ة� وال�غ� ير م�صاث�� أت� ال�غ� اي&� ب� ي�رول )ال�ن� أل�كن- ة� ي�� ارث�� ة� م�ق� وث� ة� م�عي� ذرج� ي��
29
Copyright © 2022 FDOKUMEN




















