
Biobutanol production from rice bran and de-oiled rice bran by Clostridium...
13
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/51857884 Biobutanol production from rice bran and de- oiled rice bran by Clostridium saccharoperbutylacetonicum N1-4 Article in Bioprocess and Biosystems Engineering · December 2011 DOI: 10.1007/s00449-011-0664-2 · Source: PubMed CITATIONS 26 READS 89 3 authors: Some of the authors of this publication are also working on these related projects: Knowledge Transfer and Implementation of System of Rice Intensification And Associated Sustainable Agricultural Innovations In Tumpat, Kelantan View project Production of Biocontrol & Biofertilizers Using Effective Microorganism Technology for System of Rice Intensification (SRI) View project Najeeb Al-Shorgani National University of Malaysia 40 PUBLICATIONS 209 CITATIONS SEE PROFILE Mohd Sahaid Kalil National University of Malaysia 100 PUBLICATIONS 522 CITATIONS SEE PROFILE Wan Mohtar Wan Yusoff National University of Malaysia 130 PUBLICATIONS 591 CITATIONS SEE PROFILE All content following this page was uploaded by Najeeb Al-Shorgani on 25 January 2014. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.
Transcript of Biobutanol production from rice bran and de-oiled rice bran by Clostridium...

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/51857884
Biobutanolproductionfromricebranandde-oiledricebranbyClostridiumsaccharoperbutylacetonicumN1-4
ArticleinBioprocessandBiosystemsEngineering·December2011
DOI:10.1007/s00449-011-0664-2·Source:PubMed
CITATIONS
26
READS
89
3authors:
Someoftheauthorsofthispublicationarealsoworkingontheserelatedprojects:
KnowledgeTransferandImplementationofSystemofRiceIntensificationAndAssociated
SustainableAgriculturalInnovationsInTumpat,KelantanViewproject
ProductionofBiocontrol&BiofertilizersUsingEffectiveMicroorganismTechnologyforSystemofRice
Intensification(SRI)Viewproject
NajeebAl-Shorgani
NationalUniversityofMalaysia
40PUBLICATIONS209CITATIONS
SEEPROFILE
MohdSahaidKalil
NationalUniversityofMalaysia
100PUBLICATIONS522CITATIONS
SEEPROFILE
WanMohtarWanYusoff
NationalUniversityofMalaysia
130PUBLICATIONS591CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyNajeebAl-Shorganion25January2014.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinblue
arelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

1 23
Bioprocess and BiosystemsEngineering ISSN 1615-7591 Bioprocess Biosyst EngDOI 10.1007/s00449-011-0664-2
Biobutanol production from rice branand de-oiled rice bran by Clostridiumsaccharoperbutylacetonicum N1-4
Najeeb Kaid Nasser Al-Shorgani, MohdSahaid Kalil & Wan Mohtar Wan Yusoff

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL PAPER
Biobutanol production from rice bran and de-oiled rice branby Clostridium saccharoperbutylacetonicum N1-4
Najeeb Kaid Nasser Al-Shorgani • Mohd Sahaid Kalil •
Wan Mohtar Wan Yusoff
Received: 22 August 2011 / Accepted: 28 November 2011
� Springer-Verlag 2011
Abstract Rice bran (RB) and de-oiled rice bran (DRB)
have been treated and used as the carbon source in ace-
tone–butanol–ethanol (ABE) production using Clostridium
saccharoperbutylacetonicum N1-4. The results showed that
pretreated DRB produced more ABE than pretreated RB.
Dilute sulfuric acid was the most suitable treatment method
among the various pretreatment methods that were applied.
The highest ABE obtained was 12.13 g/L, including
7.72 g/L of biobutanol, from sulfuric acid. The enzymatic
hydrolysate of DRB (ESADRB), when treated with XAD-4
resin, resulted in an ABE productivity and yield of 0.1 g/
L h and 0.44 g/g, respectively. The results also showed that
the choice of pretreatment method for RB and DRB is an
important factor in butanol production.
Keywords Biobutanol � Clostridium
saccharoperbutylacetonicum N1-4 � Rice bran �De-oiled rice bran � Pretreatment
Introduction
The growing demand of biobutanol as an alternative form
of fuel for transportation has created considerable interest
due to the occurrence of global warming caused by the
increase of carbon dioxide emissions into the atmosphere
from the combustion of fossil fuels.
As a biofuel, butanol has many advantages over ethanol.
The vapor pressure of butanol is lower than that of ethanol,
while its energy content is higher; these attributes make
butanol to be safer for use in blending gasoline for sub-
sequent use as fuel, because the blend has a better fuel
economy as a fuel than blends obtained from the mixture of
ethanol and gasoline. The ability of butanol to tolerate
contamination from water makes its blends with gasoline
averagely stable and this greatly increases the efficiency of
the existing gasoline supply and distribution channels. In
addition, butanol can be used in conventional engines
without modification [1].
The fermentation process through the use of solvent-
producing Clostridium for the production of butanol has
some problems ranging from toxicity and low productivity
to high recovery cost. However, the results of various
researches have yielded new methods for reducing the
difficulty encountered in the process previously. These new
techniques include the procurement of higher butanol-tol-
erant strains and enhancement of yield through mutagen-
esis or the periodic collection of solvent to prevent their
toxicity [2].
To reduce the cost associated with recovery, many
efforts have been made to develop in situ separation pro-
cesses. These include pervaporation, adsorption, liquid–
liquid extraction, gas stripping and reverse osmosis [2].
Also, to increase the butanol concentration and butanol
selectivity as the major product in the total solvent, com-
plementation of the adhE1 and ctfAB genes in a mega-
plasmid-less C. acetobutylicum M5 strain has been
proposed [3]. The new approach is focused on developing a
more efficient process in the production of butanol through
the use of system-level techniques. Some of these
N. K. N. Al-Shorgani (&) � W. M. W. Yusoff
School of Bioscience and Biotechnology, Faculty of Science
and Technology, Universiti Kebangsaan Malaysia,
43600 UKM Bangi, Selangor, Malaysia
e-mail: [email protected]
M. S. Kalil
Department of Chemical and Process Engineering,
Faculty of Engineering, Universiti Kebangsaan Malaysia,
43600 UKM Bangi, Selangor, Malaysia
123
Bioprocess Biosyst Eng
DOI 10.1007/s00449-011-0664-2
Author's personal copy

techniques, for instance genome sequencing and engi-
neering, construction of genome-scale metabolic net-
works of clostridia, omics studies, transcription machinery
engineering and construction of synthetic pathways, have
been utilized in developing more efficient butanol pro-
ducers [4]. To overcome poor solvent resistance and
low productivity in butanol production, a new strategy
involving the use of in situ recovery techniques alongside
strain design has been proposed [5]. One of the most
important factors in butanol fermentation is the avail-
ability and price of the substrate. Butanol can be produced
from various raw materials or renewable agricultural crops
including sago starch [6, 7], corn [8], molasses [9] and
whey permeate [10]. Rice is the staple food for a large
portion of the world’s population and the annual world
rice production is more than 500 million metric tons. The
primary waste product of the rice industry is rice bran,
of which approximately 40 million metric tons are pro-
duced every year and are discarded as unfit for human
consumption.
Rice bran (RB), the residue of brown rice that is
created during the production of white rice, is abun-
dantly available in Southeast Asia. This agricultural by-
product contains a number of carbohydrates and other
nutrients such as proteins, lipids, fiber, Ca2?, Mg2?,
phosphate, silica, Zn2?, thiamin and niacin [11]; but its
industrial applications are limited to being an animal
feed additive or for the production of rice bran oil. This
residue, after extracting rice bran oil, is called de-oiled
rice bran (DRB). Therefore, there are two different
kinds of rice bran available from the rice-processing
industry: RB and DRB. DRB still contains many car-
bohydrates and cellulosic polysaccharides [12], and the
presence of a high amount of carbohydrate (39% cel-
lulose and 31% hemicellulose) and less lignin (4%)
make it an attractive feedstock for conversion into a
variety of value-added products, such as single cell
proteins and ethanol [13].
Dilute acid hydrolysis is a simple and easy method to
perform and is regularly used for the depolymerization of
biomass into fermentable sugars [13]. However, it has
some limitations. For example, if higher temperatures (or
longer residence time) are applied, the hemicellulosic-
derived monosaccharides will degrade and give rise to
fermentation inhibitors such as furan compounds, weak
carboxylic acids and phenolic compounds. These fermen-
tation inhibitors are known to affect the performance of
bioethanol and biobutanol production in fermenting
microorganisms [14–16].
In the present study, Clostridium saccharoperbutyl-
acetonicum N1-4 was used to produce butanol from RB
and DRB hydrolysates, because this bacterium is known to
be a hyper-butanol-producing strain [17, 18].
Materials and methods
Microorganism and inoculum preparation
Clostridium saccharoperbutylacetonicum N1-4 was pro-
vided by the Biotechnology Laboratory, Chemical and
Process Engineering Department of UKM. It was kept at
4 �C as a suspension of spores in potato glucose medium
(PG medium) as a stock culture. The inoculum was pre-
pared by transferring the suspension of spores (1 mL) into
10 mL of PG medium with subsequent heat shock for
1 min in boiling water, cooling in ice water and incubating
for 1–2 days at 30 �C under anaerobic conditions. The
colony morphology and Gram-staining behavior of the
inoculum were verified to ensure that the culture was pure
[19]. The culture was then transferred to tryptone–yeast
extract–acetate medium (TYA), incubated for 18 h and
used as the inoculum.
RB and DRB pretreatment
RB was collected from Kilang Beras BERNAS, Tanjung
Karang, Selangor, Malaysia; and DRB was obtained by
extracting the oil using hexane. The oil was extracted by
solvent extraction using a Soxhlet apparatus according to
the method of Aryee and Simpson [20]. Briefly, one part of
the rice bran sample of a known weight was injected into a
33 9 94 mm cellulose extraction thimble, covered with
silanized glass wool and extracted with 20 parts of hexane
(1:20 w/v) for 10 h at the boiling point of hexane (70 �C)
[20].
The RB and DRB were treated by acidic and enzymatic
methods prior to fermentation. For acid pretreatment,
100 g of RB or DRB was soaked in 1 L of 1% (v/v) HCl or
1% (v/v) H2SO4 and autoclaved at 121 �C for 1 h (dilute
sulfuric acid pretreatment) or at 80 �C for 3 h (dilute
hydrochloric pretreatment), to generate hydrochloric acid
rice bran hydrolysate (HARB), hydrochloric acid de-oiled
rice bran hydrolysate (HADRB), sulfuric acid rice bran
hydrolysate (SARB) and sulfuric acid de-oiled rice bran
hydrolysate (SADRB), respectively. A combined enzy-
matic and acid treatment was carried out after the acid
treatment to generate EHARB, EHADRB, ESARB and
ESADRB. The pH was then adjusted to 4.5 with 10 M
NaOH, and a mixture of two enzyme solutions (6 mL/
100 g RB or DRB of each enzyme) [Celluclast 1.5 L
(cellulase; Novo, Malaysia) and Novozyme 188 (b-gluco-
sidase; Novo, Malaysia)] was added and the solution was
incubated at 45 �C for 72 h with agitation at 80 rpm. After
72 h, the hydrolysate was centrifuged at 7,0009g for
10 min to remove the sediments, and the supernatant was
supplemented with P2 solution and autoclaved at 121 �C
for 15 min and used as a fermentation medium.
Bioprocess Biosyst Eng
123
Author's personal copy

The DRB was also treated enzymatically using amylase
enzymes. For this, the pH of the DRB solution was
adjusted to 6.2 by 10 M NaOH and calcium was added,
using calcium chloride (Ca?2 C40 ppm), to stabilize the
enzyme. A heat-stable a-amylase (Termamyl 120 L; Novo,
Malaysia) (0.05 mL/100 g DRB) was added to the DRB
mixture, heated to 100 �C and held at this temperature for
10 min. The mixture was then cooled to 90 �C and held at
this temperature for an additional 2 h to increase hydro-
lysis. After liquefaction, the pH was reduced to 4.5 by 5 M
HCl, and the solution was cooled to 60 �C. Saccharification
was carried out by adding glucoamylase enzyme (AMG
300 L; Novo, Malaysia) (0.1 mL/100 g DRB) at pH 4.5
and incubating at 60 �C for 24 h.
Inhibitor removal by overliming and XAD-4 resin
Chemical treatment methods result in the production of
undesirable secondary products that can inhibit microbial
growth. One of the ways to reduce or remove these
inhibitors is through the overliming method, where calcium
hydroxide is added under fast stirring at 80 �C until the pH
rises to 10, followed by the addition of 1 g/L Na2SO3
(overliming). Sedimentation of the inhibitors was allowed
for 1 h with intermittent mixing followed by cooling to
room temperature. The mixture was then filtered using
vacuum filtration to remove precipitates. The pH of the
clear filtrate was adjusted to 7 using concentrated H2SO4
and was again vacuum filtered to remove any remaining
traces of salt residue.
The hydrolysate was centrifuged again at 5,0009g for
10 min, and the fermentation inhibitors were removed by
passing the clear supernatant through a nonionic polymeric
adsorbent resin [Amberlite XAD-4 (Sigma Chemicals)]
column (510 9 15 mm) at a flow rate of 8 mL/min.
Approximately, 500 mL of hydrolysate was passed through
the column, which was then packed using 75 g of XAD-4
resin. Previous studies have shown that 60–80% of furfural,
hydroxymethyl furfural (HMF) and ferulic compounds can
be removed using XAD-4 resin [21].
Analytical methods
Samples were taken periodically for the analysis of sol-
vents, acids, pH and sugars. After centrifugation at
7,0009g for 10 min, the supernatants were filtered through
a 0.2-lm cellulose acetate filter. The solvent (acetone,
butanol and ethanol) concentrations were analyzed by gas
chromatography (7890A GC-System, Agilent Technolo-
gies, Palo Alto, CA, USA) equipped with a flame ioniza-
tion detector and a 30-m capillary column (Equity1;
30 m 9 0.32 mm 9 1.0-lm film thickness; Supelco Co,
Bellefonate, PA, USA). The oven temperature was
programmed to increase from 40 to 130 �C at a rate of
8 �C/min. The injector and detector temperatures were set
at 250 and 280 �C, respectively. Helium, as the carrier gas,
was set at a flow rate of 1.5 mL/min.
Sugars and organic acids (acetic and butyric acid) were
measured by high-performance liquid chromatography
(HPLC 12000 Series, Agilent technologies, Palo Alto, CA,
USA). Acids were analyzed using a Genesis C18 120A
column (25 cm 9 4.6 mm; Jones Chromatography,
Tempe, AZ, USA) at a column temperature of 40 �C, with
20 mM H2SO4 as a mobile phase, at a flow rate of 0.6 mL/
min. Detection was accomplished with a UV detector at a
wavelength of 220 nm. Sugars were analyzed using a
Shodex Asahipak NH2P-50 4E column (4.6 mm
ID 9 250 mm; Shodex, Japan), and the concentrations
were detected with a refractive index detector (RID 1200,
Agilent Technologies, Palo Alto, CA, USA) at 30 �C and a
flow rate 1 mL/min, using a mixture of acetonitrile and
water (H2O/CH3CN = 40/60) as a mobile phase.
The glucose concentration was detected with a Bio-
chemistry Analyzer (YSI 2700D, YSI Inc., Life Sciences,
Yellow Springs, OH, USA). Cell density was measured
using a Genesys-10 Scanning UV–VIS Spectrophotometer
(Thermo Spectronic, Rochester, USA) at 660 nm. ABE
yield was defined as the amount of ABE produced divided
by the total amount of sugars, and productivity was defined
as the ABE concentration divided by the fermentation time.
Media preparation
PG medium was used to prepare the inoculum and consisted of
150 g/L potato, 10 g/L glucose, 0.5 g/L (NH4)2SO4 and 3 g/L
CaCO3.
TYA medium was also used to prepare the inoculum and
consisted of the following components: 20 g/L glucose,
6 g/L tryptone, 2 g/L yeast extract, 3 g/L ammonium
acetate, 0.5 g/L KH2PO4, 0.3 g/L MgSO4�7H2O, and
0.01 g/L FeSO4�7H2O.
P2 medium was used as a fermentation medium and
consisted of the following: 0.5 g/L KH2PO4, 0.5 g/L
K2HPO4, 0.4 g/L MgSO4�7H2O, 0.01 g/L MnSO4�4H2O,
0.01 g/L FeSO4�5H2O, 1.0 g/L yeast extract and 0.5 g/L
cysteine. A final concentration of 80 lg/L biotin and 1 mL
of a solution containing 1 mg/L 4-aminobenzoic acid were
added to 1 L of P2 medium.
Batch fermentation
The anaerobic batch fermentation was conducted in
250-mL Schott (Duran) bottles with a working volume of
150 mL. P2 medium was used as a fermentation medium,
and 10% (w/v) of either RB or DRB hydrolysates were
used as the sole carbon.
Bioprocess Biosyst Eng
123
Author's personal copy
The initial pH of the medium was adjusted to 6.5. The
medium was then sterilized by autoclaving at 121 �C for
15 min. To generate an anaerobic condition, the medium
was sparged with oxygen-free nitrogen, and the vitamin
solution was filter-sterilized and added aseptically into the
sterilized medium. The batch culture was initiated by
inoculation of the medium with 10% fresh inoculum of
C. saccharoperbutylacetonicum N1-4 that had been previ-
ously grown in TYA medium for 18 h. The Schott bottles
were incubated at 30 �C without shaking under anaerobic
conditions. All fermentations were performed in duplicate,
and measurements are mean values.
Results and discussion
In this study, RB and DRB were pretreated prior to fer-
mentation by various methods. This pretreatment included
using diluted sulfuric and hydrochloric acid (1% v/v) fol-
lowed by a combination of acidic and enzymatic hydroly-
sis. Treatment with H2SO4 and HCl generates SARB,
HARB, SADRB and HADRB. Subsequent treatments of
RB and DRB using the combination of dilute acid followed
by enzymatic hydrolysis generated ESARB, EHARB, ES-
ADRB and EHADRB, respectively.
RB as a by-product of rice production and DRB as a by-
product of rice oil production are cheap and renewable
agricultural products available around the world. Hydro-
lysis of 100 g/L RB and DRB using dilute sulfuric acid
produced sufficient amounts of reducing sugars for butanol
fermentation by C. saccharoperbutylacetonicum N1-4 at
31.43 and 33.35 g/L, respectively. The solvent produced
from the DRB hydrolysate, after the XAD-4 resin treat-
ment, showed concentrations as high as 12.13 g/L of ABE.
ABE fermentation using untreated RB and DRB
as media
RB and DRB without hydrolysis/treatment were used as a
control and supplemented in P2 medium. The reduced
sugar content in untreated rice bran was found to be 6.74 g/
L. The presence of this small amount of sugar may result
from the sterilization effect, which causes the thermal
degradation of polysaccharides in RB. DRB showed lesser
amounts of reducing sugars (2.7 g/L), possibly due to the
oil extraction process.
During the fermentation of untreated RB using C. sac-
charoperbutylacetonicum N1-4, the sugar concentrations of
8.43 and 9.88 g/L were detected after 12 and 24 h,
respectively, demonstrating the ability of C. sacchar-
operbutylacetonicum N1-4 to secrete the respective
enzymes for the hydrolysis of different RB constituents
(Fig. 1). A similar phenomenon was observed with the
fermentation of the untreated DRB (Fig. 2).
ABE production by C. saccharoperbutylacetonicum
N1-4 was carried out using untreated RB and DRB as a
carbon source in P2 medium. During the 96 h of RB fer-
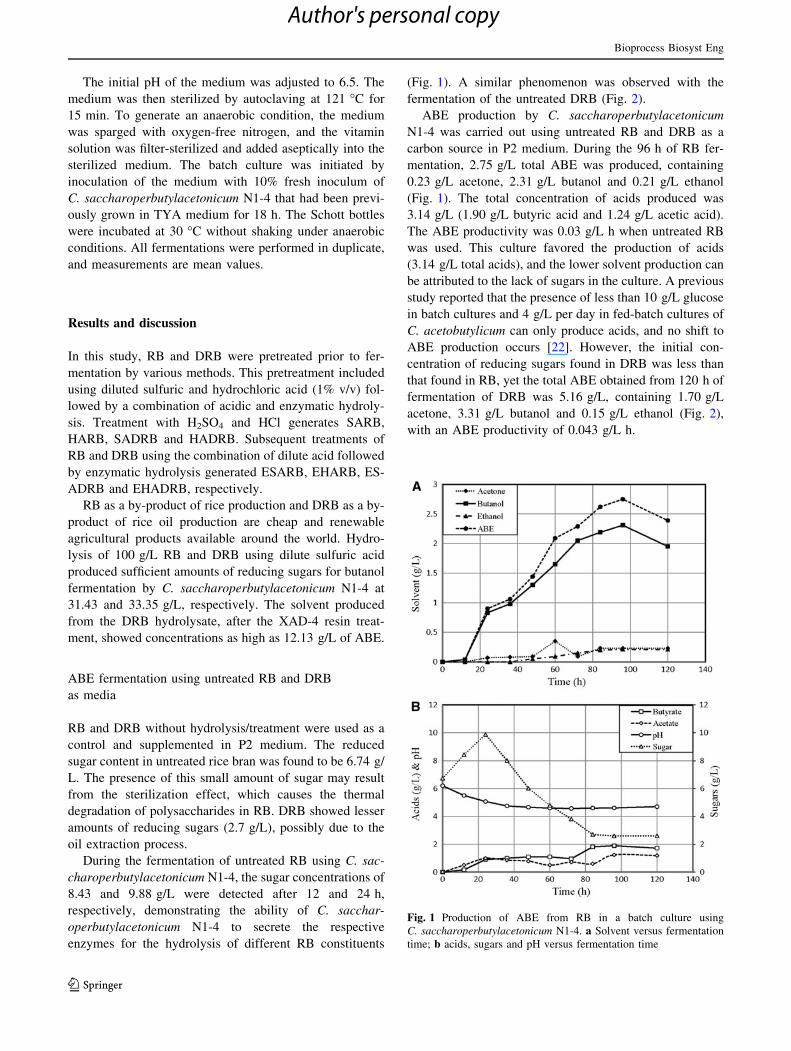
mentation, 2.75 g/L total ABE was produced, containing
0.23 g/L acetone, 2.31 g/L butanol and 0.21 g/L ethanol
(Fig. 1). The total concentration of acids produced was
3.14 g/L (1.90 g/L butyric acid and 1.24 g/L acetic acid).
The ABE productivity was 0.03 g/L h when untreated RB
was used. This culture favored the production of acids
(3.14 g/L total acids), and the lower solvent production can
be attributed to the lack of sugars in the culture. A previous
study reported that the presence of less than 10 g/L glucose
in batch cultures and 4 g/L per day in fed-batch cultures of
C. acetobutylicum can only produce acids, and no shift to
ABE production occurs [22]. However, the initial con-
centration of reducing sugars found in DRB was less than
that found in RB, yet the total ABE obtained from 120 h of
fermentation of DRB was 5.16 g/L, containing 1.70 g/L
acetone, 3.31 g/L butanol and 0.15 g/L ethanol (Fig. 2),
with an ABE productivity of 0.043 g/L h.
Fig. 1 Production of ABE from RB in a batch culture using
C. saccharoperbutylacetonicum N1-4. a Solvent versus fermentation
time; b acids, sugars and pH versus fermentation time
Bioprocess Biosyst Eng
123
Author's personal copy
From these results, it can be suggested that the treatment
of RB and DRB or other agricultural residues prior to
fermentation is an important factor in improving the ABE
yield and productivity. Similar results were obtained from a
study conducted by Lee et al. [11], where only 2.22 g/L
butanol was produced from untreated RB; however, when
it was supplemented with P2 medium and fermented using
C. beijerinckii NCIMB 8052 in a batch reactor, 5.03 g/L of
ABE was obtained versus 5.16 g/L of ABE obtained from
DRB in our study. Our results are in agreement with those
reported by Lee et al. [11].
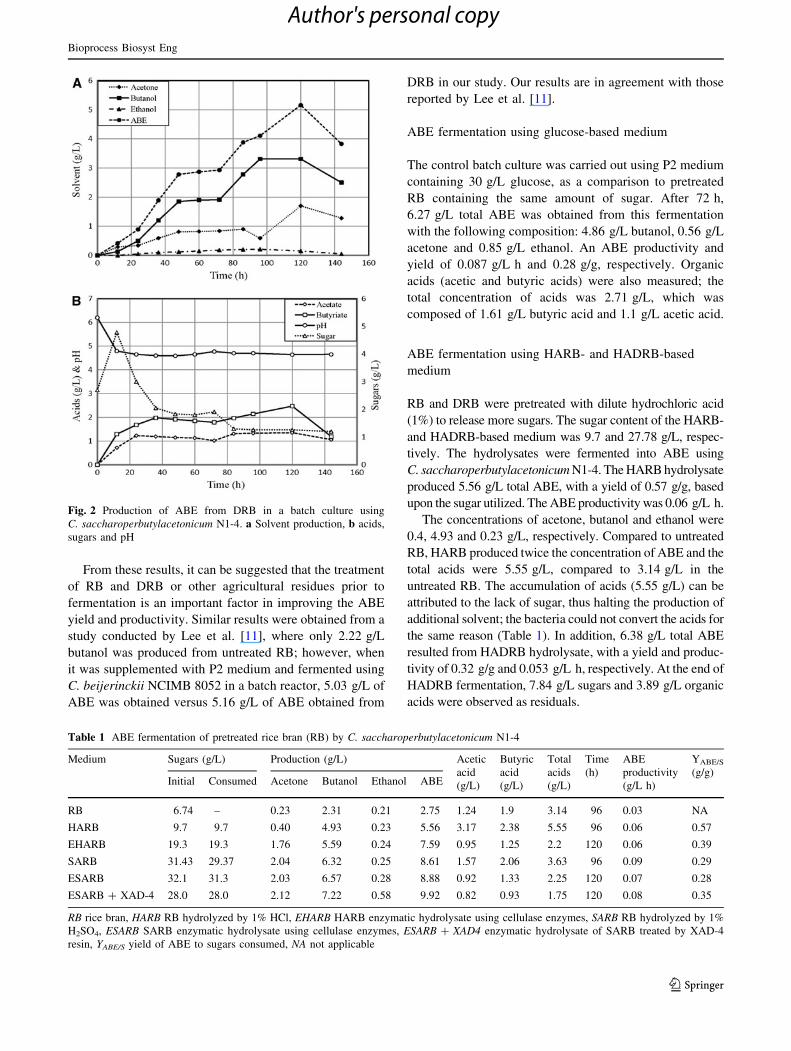
ABE fermentation using glucose-based medium
The control batch culture was carried out using P2 medium
containing 30 g/L glucose, as a comparison to pretreated
RB containing the same amount of sugar. After 72 h,
6.27 g/L total ABE was obtained from this fermentation
with the following composition: 4.86 g/L butanol, 0.56 g/L
acetone and 0.85 g/L ethanol. An ABE productivity and
yield of 0.087 g/L h and 0.28 g/g, respectively. Organic
acids (acetic and butyric acids) were also measured; the
total concentration of acids was 2.71 g/L, which was
composed of 1.61 g/L butyric acid and 1.1 g/L acetic acid.
ABE fermentation using HARB- and HADRB-based
medium
RB and DRB were pretreated with dilute hydrochloric acid
(1%) to release more sugars. The sugar content of the HARB-
and HADRB-based medium was 9.7 and 27.78 g/L, respec-
tively. The hydrolysates were fermented into ABE using
C. saccharoperbutylacetonicum N1-4. The HARB hydrolysate
produced 5.56 g/L total ABE, with a yield of 0.57 g/g, based
upon the sugar utilized. The ABE productivity was 0.06 g/L h.
The concentrations of acetone, butanol and ethanol were
0.4, 4.93 and 0.23 g/L, respectively. Compared to untreated
RB, HARB produced twice the concentration of ABE and the
total acids were 5.55 g/L, compared to 3.14 g/L in the
untreated RB. The accumulation of acids (5.55 g/L) can be
attributed to the lack of sugar, thus halting the production of
additional solvent; the bacteria could not convert the acids for
the same reason (Table 1). In addition, 6.38 g/L total ABE
resulted from HADRB hydrolysate, with a yield and produc-
tivity of 0.32 g/g and 0.053 g/L h, respectively. At the end of
HADRB fermentation, 7.84 g/L sugars and 3.89 g/L organic
acids were observed as residuals.
Fig. 2 Production of ABE from DRB in a batch culture using
C. saccharoperbutylacetonicum N1-4. a Solvent production, b acids,
sugars and pH
Table 1 ABE fermentation of pretreated rice bran (RB) by C. saccharoperbutylacetonicum N1-4
Medium Sugars (g/L) Production (g/L) Acetic
acid
(g/L)
Butyric
acid
(g/L)
Total
acids
(g/L)
Time
(h)
ABE
productivity
(g/L h)
YABE/S
(g/g)Initial Consumed Acetone Butanol Ethanol ABE
RB 6.74 – 0.23 2.31 0.21 2.75 1.24 1.9 3.14 96 0.03 NA
HARB 9.7 9.7 0.40 4.93 0.23 5.56 3.17 2.38 5.55 96 0.06 0.57
EHARB 19.3 19.3 1.76 5.59 0.24 7.59 0.95 1.25 2.2 120 0.06 0.39
SARB 31.43 29.37 2.04 6.32 0.25 8.61 1.57 2.06 3.63 96 0.09 0.29
ESARB 32.1 31.3 2.03 6.57 0.28 8.88 0.92 1.33 2.25 120 0.07 0.28
ESARB ? XAD-4 28.0 28.0 2.12 7.22 0.58 9.92 0.82 0.93 1.75 120 0.08 0.35
RB rice bran, HARB RB hydrolyzed by 1% HCl, EHARB HARB enzymatic hydrolysate using cellulase enzymes, SARB RB hydrolyzed by 1%
H2SO4, ESARB SARB enzymatic hydrolysate using cellulase enzymes, ESARB ? XAD4 enzymatic hydrolysate of SARB treated by XAD-4
resin, YABE/S yield of ABE to sugars consumed, NA not applicable
Bioprocess Biosyst Eng
123
Author's personal copy
ABE fermentation using EHARB- and EHADRB-based
medium
The combined pretreatment using dilute hydrochloric acid
(1%) followed by cellulolytic enzymes was performed on
RB and DRB to generate more sugars. This treatment
generates EHARB and EHADRB hydrolysates, and it
resulted in an increased production of up to 19.3 g/L
reducing sugars in EHARB, having 18 g/L glucose and
1.3 g/L xylose. The fermentation of EHARB resulted in
7.59 g/L ABE, which contained 1.76 g/L acetone, 5.56 g/L
butanol and 0.24 g/L ethanol. ABE productivity and yield
were 0.06 g/L h and 0.39 g/g, respectively. Although the
ABE production from EHARB improved compared to
HARB, the productivity remained unchanged (0.06 g/L h);
this is likely due to the fermentation time. The prolonged
fermentation time may be due to the existence of microbial
inhibitors resulting from the hydrolysis of lignocellulosic
materials, although we avoided the use of high tempera-
tures in this treatment (only heating at 80 �C for 3 h).
The total sugar concentration resulting from the pretreat-
ment of HADRB with cellulolytic enzymes was 29.51 g/L,
and the fermentation of this hydrolysate produced 6.56 g/L
total ABE, containing 1 g/L acetone, 4.71 g/L butanol and
0.84 g/L ethanol. In this study, EHARB produced more ABE
and butanol compared to EHADRB; the productivity was also
increased, although the reducing sugar concentration was
higher in EHADRB.
ABE fermentation using SARB- and SADRB-based
medium
The SARB- and SADRB-based medium contained 30.79
and 33.35 g/L reducing sugars, respectively. The concen-
trations of glucose, xylose and cellobiose in SARB were
23.46, 5.64 and 1.69 g/L, respectively; and in SADRB, the
concentrations were 19.96, 8.21 and 5.18 g/L, respectively.
The glucose percentage was 76.2% in SARB and 60% in
SADRB, indicating the efficiency of using dilute sulfuric
acid as a pretreatment method to generate simple sugars
from agricultural residues.
The fermentation profiles of the SARB and SADRB
hydrolysates by C. saccharoperbutylacetonicum N1-4 in
batch culture utilized 29.37 and 28 g/L of reducing sugar,
respectively. During the first 96 h of SARB hydrolysate
fermentation, 93% of the reducing sugar was consumed,
resulting in a total ABE production of 8.61 g/L, containing
6.23 g/L butanol, 2.04 g/L acetone and 0.25 g/L ethanol.
An ABE yield of 0.29 g/g of sugar was consumed, and a
productivity of 0.09 g/L h was achieved. A regular
increase in solvent production was observed until 96 h of
incubation and became constant thereafter. The maximum
solvent production (9.66 g/L) was measured after 120 h
from SADRB medium, with an ABE yield of 0.35 g/g and
a productivity of 0.081 g/L h (Table 2).
A maximum value of 8.61 g/L ABE resulting from the
SARB fermentation was lower than that reported by Lee
et al. [11]. This may be due to the ability of the C. beijerinckii
NCIMB 8052 strain to more efficiently convert the SARB to
butanol compared to C. saccharoperbutylacetonicum N1-4.
It may also be due to the presence of inhibitors in the SARB.
The sulfuric acid showed a superior effect for the hydrolysis
of hemicelluloses, but generated inhibitors for microbial
fermentation.
ABE fermentation using ESARB- and ESADRB-based
medium
SARB and SADRB were treated with cellulase enzymes
(cellulase and cellobiose) as mentioned in ‘‘Materials and
methods’’. The hydrolysates of ESARB ESADRB were
detoxified by overliming and used for solvent production.
Table 2 ABE fermentation of pretreated de-oiled rice bran (DRB) by C. saccharoperbutylacetonicum N1-4
Medium Sugars (g/L) Production (g/L) Acetic
acid
(g/L)
Butyric
acid
(g/L)
Total
acids
(g/L)
Time
(h)
ABE
productivity
(g/L h)
YABE/S
(g/g)Initial Consumed Acetone Butanol Ethanol ABE
DRB 2.70 – 1.70 3.31 0.15 5.16 1.35 2.47 3.83 120 0.043 NA
HADRB 27.78 19.94 1.83 4.39 0.16 6.38 2.00 1.89 3.89 120 0.053 0.32
EHADRB 29.51 24.30 1.00 4.71 0.84 6.56 0.78 1.38 2.16 120 0.055 0.27
SADRB 33.35 28.00 2.55 6.75 0.36 9.66 2.16 0.74 2.90 120 0.081 0.35
ESADRB 32.65 26.28 2.12 7.10 1.29 10.51 0.87 1.04 1.91 120 0.088 0.40
ESDRB ? XAD-4 30.00 28.57 3.18 7.72 1.23 12.13 1.63 0.50 2.13 120 0.101 0.44
EDRB 29.00 24.54 1.22 4.96 0.19 6.38 2.73 1.21 3.94 120 0.053 0.26
DRB de-oiled rice bran, HADRB DRB hydrolyzed by 1% HCl, EHADRB HARB enzymatic hydrolysate using cellulase enzymes, SADRB DRB
hydrolyzed by 1% H2SO4, ESADRB SADRB enzymatic hydrolysate using cellulase enzymes, ESDRB ? XAD4 enzymatic hydrolysate of DRB
treated by XAD-4 resin, EDRB DRB treated by amylase enzymes, YABE/S yield of ABE to sugars, NA not applicable
Bioprocess Biosyst Eng
123
Author's personal copy

ESARB did not enhance the production of reducing
sugars and the sugar content in ESARB was 32.1 g/L
compared to the 31.43 g/L found in SARB; a similar effect
was observed with ESADRB (Tables 1, 2). These results
demonstrate that a dilute sulfuric acid treatment has the
ability to convert the lignocellulosic materials to ferment-
able sugars while saving the cost associated with enzymatic
hydrolysis. During the fermentation of the ESARB
hydrolysate, only 8.88 g/L total ABE was produced,
resulting in a productivity of 0.07 g/L h and a yield of
0.28 g/g, while ESADRB resulted in 10.51 g/L total ABE,
with a productivity of 0.088 g/L h and an ABE yield of
0.4 g/g. Moreover, enzymatic hydrolysis of DRB using
amylolytic enzymes resulted in 29 g/L reducing sugars and
only 6.38 g/L of ABE with a productivity of 0.053 g/L h,
which is similar to that seen in HADRB.
Previously, it was reported that butanol fermentation
from renewable agricultural residues requires pretreatment
to increase the amounts of fermentable sugars [15, 23, 24].
The most popular and effective hydrolysis method for these
renewable lignocellulosic materials is acid treatment using
dilute sulfuric acid; yet, this type of treatment suffers from
microbial inhibitor generation, such as furans, aliphatic
acids and phenolic compounds [25, 26], thus reducing the
yield and productivity of butanol. In addition, the inhibitors
that affect microbial growth and ABE fermentation are
required to be reduced or removed. However, this acid
treatment has been confirmed to be a cost-effective pre-
treatment method [11]. In this study, the presence of more
than 30 g/L reducing sugars was enough for solvent pro-
duction by C. saccharoperbutylacetonicum N1-4.
In an attempt to resolve the inhibition problem, XAD-4
resin was used to remove the fermentation inhibitors from
the ESARB and ESADRB hydrolysates prior to fermen-
tation. It has been reported that XAD-4 resin has the ability
to reduce furfural and hydroxymethyl furfural (HMF),
which are the common inhibitors resulting from biomass
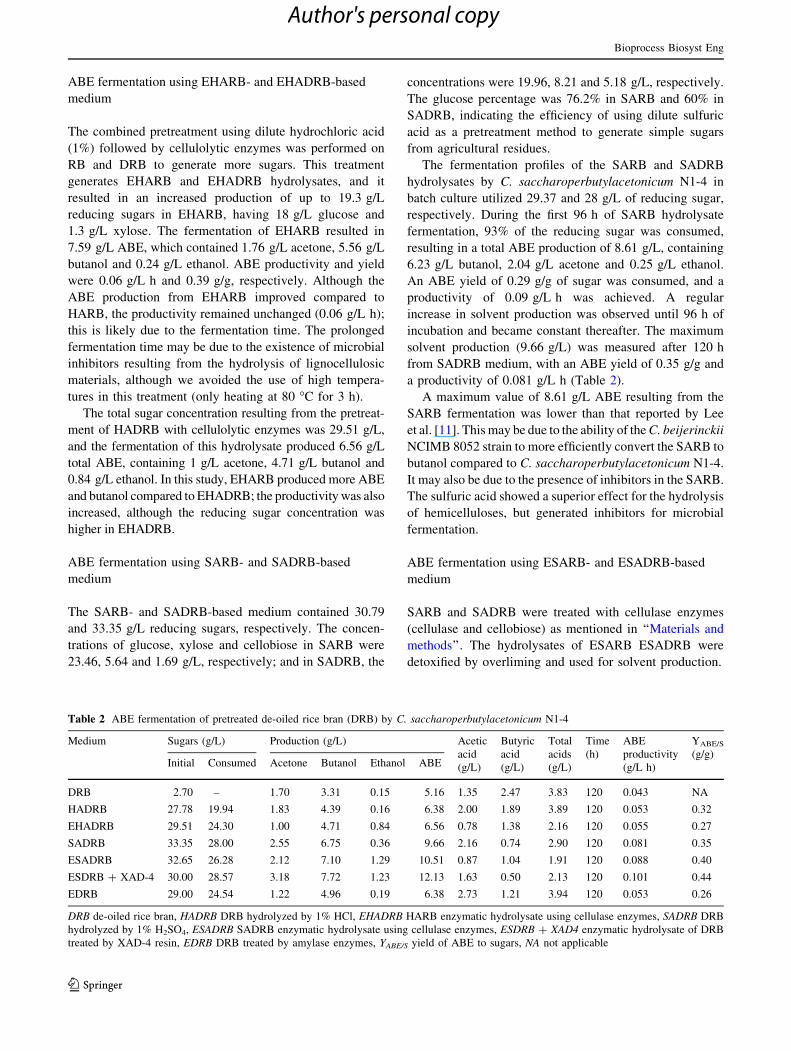
treatment [21]. Fermentation of XAD-4 treated ESARB
produced 9.92 g/L total ABE, 2.12 g/L acetone, 7.22 g/L
butanol and 0.58 g/L ethanol (Fig. 3). The ABE produc-
tivity and yield were 0.08 g/L h and 0.35, respectively.
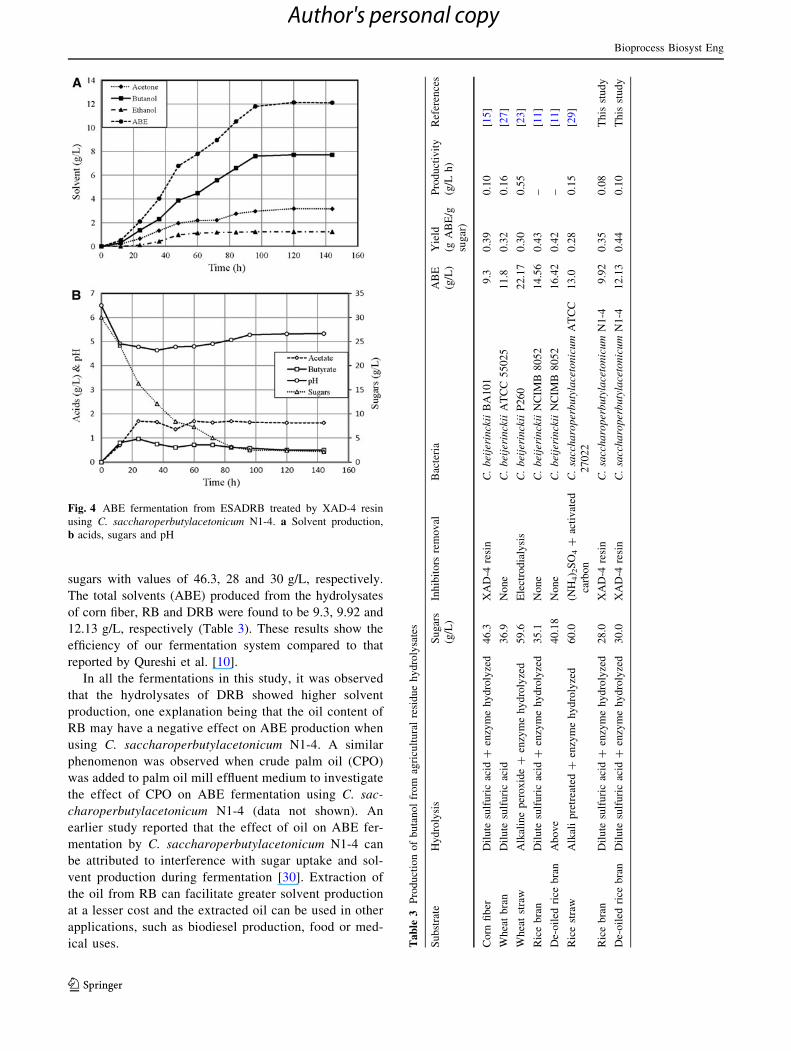
Furthermore, XAD-4 treatment of ESADRB improved
solvent production, productivity and yield. In this study, a
maximum ABE production of 12.13 g/L was obtained from
ESADRB treated with XAD-4 resin, which gave a pro-
ductivity of 0.101 g/L h (Fig. 4). These results showed that
XAD-4 resin markedly enhanced the ABE concentration,
yield and productivity compared to that obtained from non-
treated ESARB and ESADRB (Tables 1, 2).
In a similar study, Qureshi et al. [15] enhanced the
performance of corn fiber hydrolysate significantly when
the hydrolysate was treated with XAD-4 resin. In their
study, only 1.7 g/L ABE was produced from untreated corn
fiber hydrolysate, but the treatment with XAD-4 improved
the production to 9.3 g/L ABE.
Various agricultural residues, including corn fiber [15],
wheat bran [27], wheat straw [28], rice bran [11] and rice
straw [29], have been reported to have ABE production
when using different species of solvent-producing clos-
tridia (Table 3). In our study, a maximum ABE concen-
tration of 12.13 g/L was identified in the XAD-4-treated
ESADRB fermentation, which is higher than that reported
by Qureshi et al. [15] and Liu et al. [27]. However, it is
lower than that reported by Qureshi et al. [23], Lee et al.
[11] and Soni et al. [29], as summarized in Table 3. The
differences in ABE production may be due to the differing
nature of the agricultural substrates, the sugar content, the
different Clostridium strains and the existence of the fer-
mentation inhibitors still present in the hydrolysate even
after XAD-4 treatment. Both may have contributed to the
inefficient utilization of the sugars present in the ESADRB
treated with XAD-4 resin for solvent production.
Comparison of corn fiber [15], RB and DRB (our study)
using the same pretreatment method (enzymatic hydrolysis
followed by XAD-4 resin to remove fermentation inhibi-
tors) was found to produce different amounts of reducing
Fig. 3 ABE fermentation from ESARB treated by XAD-4 using C.saccharoperbutylacetonicum N1-4. a Solvent production, b acids,
sugars and pH
Bioprocess Biosyst Eng
123
Author's personal copy
sugars with values of 46.3, 28 and 30 g/L, respectively.
The total solvents (ABE) produced from the hydrolysates
of corn fiber, RB and DRB were found to be 9.3, 9.92 and
12.13 g/L, respectively (Table 3). These results show the
efficiency of our fermentation system compared to that
reported by Qureshi et al. [10].
In all the fermentations in this study, it was observed
that the hydrolysates of DRB showed higher solvent
production, one explanation being that the oil content of
RB may have a negative effect on ABE production when
using C. saccharoperbutylacetonicum N1-4. A similar
phenomenon was observed when crude palm oil (CPO)
was added to palm oil mill effluent medium to investigate
the effect of CPO on ABE fermentation using C. sac-
charoperbutylacetonicum N1-4 (data not shown). An
earlier study reported that the effect of oil on ABE fer-
mentation by C. saccharoperbutylacetonicum N1-4 can
be attributed to interference with sugar uptake and sol-
vent production during fermentation [30]. Extraction of
the oil from RB can facilitate greater solvent production
at a lesser cost and the extracted oil can be used in other
applications, such as biodiesel production, food or med-
ical uses.
Fig. 4 ABE fermentation from ESADRB treated by XAD-4 resin
using C. saccharoperbutylacetonicum N1-4. a Solvent production,
b acids, sugars and pH
Ta
ble
3P
rod
uct
ion
of
bu
tan
ol
fro
mag
ricu
ltu
ral
resi
du
eh
yd
roly
sate
s
Su
bst
rate
Hy
dro
lysi
sS
ug
ars
(g/L
)
Inh
ibit
ors
rem
ov
alB
acte
ria
AB
E
(g/L
)
Yie
ld
(gA
BE
/g
sug
ar)
Pro
du
ctiv
ity
(g/L
h)
Ref
eren
ces
Co
rnfi
ber
Dil
ute
sulf
uri
cac
id?
enzy
me
hy
dro
lyze
d4
6.3
XA
D-4
resi
nC
.b
eije
rin
ckii
BA
10
19
.30
.39
0.1
0[1
5]
Wh
eat
bra
nD
ilu
tesu
lfu
ric
acid
36
.9N
on
eC
.b
eije
rin
ckii
AT
CC
55
02
51
1.8
0.3
20
.16
[27]
Wh
eat
stra
wA
lkal
ine
per
ox
ide
?en
zym
eh
yd
roly
zed
59
.6E
lect
rod
ialy
sis
C.
bei
jeri
nck
iiP
26
02
2.1
70
.30
0.5
5[2
3]
Ric
eb
ran
Dil
ute
sulf
uri
cac
id?
enzy
me
hy
dro
lyze
d3
5.1
No
ne
C.
bei
jeri
nck
iiN
CIM
B8
05
21
4.5
60
.43
–[1
1]
De-
oil
edri
ceb
ran
Ab
ov
e4
0.1
8N
on
eC
.b
eije
rin
ckii
NC
IMB
80
52
16
.42
0.4
2–
[11]
Ric
est
raw
Alk
ali
pre
trea
ted
?en
zym
eh
yd
roly
zed
60
.0(N
H4) 2
SO
4?
acti
vat
ed
carb
on
C.
sacc
ha
rop
erb
uty
lace
ton
icu
mA
TC
C
27
02
2
13
.00
.28
0.1
5[2
9]
Ric
eb
ran
Dil
ute
sulf
uri
cac
id?
enzy
me
hy
dro
lyze
d2
8.0
XA
D-4
resi
nC
.sa
cch
aro
per
bu
tyla
ceto
nic
um
N1
-49
.92
0.3
50
.08
Th
isst
ud
y
De-
oil
edri
ceb
ran
Dil
ute
sulf
uri
cac
id?
enzy
me
hy
dro
lyze
d3
0.0
XA
D-4
resi
nC
.sa
cch
aro
per
bu
tyla
ceto
nic
um
N1
-41
2.1
30
.44
0.1
0T
his
stu
dy
Bioprocess Biosyst Eng
123
Author's personal copy

Conclusion
This study suggested that rice bran (RB) and de-oiled rice
bran (DRB) should be hydrolyzed to increase the fer-
mentable sugars and consequent butanol productivity. The
use of dilute sulfuric acid and enzymatic hydrolysis by
cellulolytic enzymes as pretreatment methods for RB and
DRB demonstrated similar amounts of reducing sugars,
whereas hydrolysis of DRB by amylase enzymes produced
comparable concentrations of total ABE and reducing
sugars when compared with dilute hydrochloric acid
hydrolysis of DRB. Among all the hydrolysates, the highest
total solvent and butanol production (12.13 and 7.72 g/L,
respectively) was obtained when ESADRB was treated
with XAD-4 resin, with a productivity of 0.1 g/L h and a
yield of 0.44 g/g.
It was found that the poor butanol fermentation perfor-
mance in RB hydrolysates compared to DRB hydrolysates
was likely due to the presence of oil in the rice bran. This
study showed that the extraction of oil from rice bran could
improve the butanol production by 88%. The oil produced
can then be used in other applications such as with bio-
diesel. An enhancement of butanol production was found
when the hydrolysates of RB or DRB were treated with
XAD-4 resin, thus demonstrating the presence of fermen-
tation inhibitors accompanying the hydrolysis of cellulosic
biomass and the ability of XAD-4 resin to remove some
inhibitors.
Acknowledgments We would like to thank Prof. Dr Yoshino
Sadazo, Kyushu University, Japan, who provided us with C. sacchar-operbutylacetonicum N1-4. This research was supported by the
UKM-GUP-KPB -08-32-128 grant.
References
1. Durre P (2007) Biobutanol: an attractive biofuel. Biotechnol J
2(12):1525–1534. doi:10.1002/biot.200700168
2. Lee SY, Park JH, Jang SH, Nielsen LK, Kim J, Jung KS (2008)
Fermentative butanol production by clostridia. Biotechnol Bioeng
101(2):209–228. doi:10.1002/bit.22003
3. Lee JY, Jang Y-S, Lee J, Papoutsakis ET, Lee SY (2009) Met-
abolic engineering of Clostridium acetobutylicum M5 for highly
selective butanol production. Biotechnol J 4(10):1432–1440. doi:
10.1002/biot.200900142
4. Jang Y-S, Lee J, Malaviya A, Seung DY, Cho JH, Lee SY (2011)
Butanol production from renewable biomass: rediscovery of
metabolic pathways and metabolic engineering. Biotechnol J.
doi:10.1002/biot.201100059
5. Ezeji T, Milne C, Price ND, Blaschek HP (2010) Achievements
and perspectives to overcome the poor solvent resistance in
acetone and butanol-producing microorganisms. Appl Microbiol
Biotechnol 85(6):1697–1712. doi:10.1007/s00253-009-2390-0
6. Al-Shorgani N, Kalil M, Yusoff W (2011) Fermentation of sago
starch to biobutanol in a batch culture using Clostridium sac-charoperbutylacetonicum N1-4 (ATCC 13564). Ann Microbiol
1–12. doi:10.1007/s13213-011-0347-x
7. Madihah MS, Ariff AB, Sahaid KM, Suraini AA, Karim MIA
(2001) Direct fermentation of gelatinized sago starch to acetone–
butanol–ethanol by Clostridium acetobutylicum. World J Micro-
biol Biotechnol 17(6):567–576. doi:10.1023/a:1012351112351
8. Qureshi N, Blaschek H (2001) ABE production from corn:
a recent economic evaluation. J Ind Microbiol Biotechnol
27:292–297
9. Qureshi N, Lolas A, Blaschek HP (2001) Soy molasses as fer-
mentation substrate for production of butanol using Clostridiumbeijerinckii BA101. J Ind Microbiol Biotechnol 26(5):290–295.
doi:10.1038/sj.jim.7000131
10. Ennis BM, Maddox IS (1985) Use of Clostridium acetobutylicumP262 for production of solvents from whey permeate. Biotechnol
Lett 7(8):601–606. doi:10.1007/bf01026457
11. Lee J, Seo E, Kweon D-H, Park K, Jin Y-S (2009) Fermentation
of rice bran and defatted rice bran for butanol production using
Clostridium beijerinckii NCIMB 8052. J Microbiol Biotechnol
19(5):482–490. doi:10.4014/jmb.0804.275
12. Tanaka T, Hoshina M, Tanabe S, Sakai K, Ohtsubo S, Taniguchi
M (2006) Production of D-lactic acid from defatted rice bran by
simultaneous saccharification and fermentation. Bioresour Tech-
nol 97(2):211–217
13. Chandel AK, Narasu ML, Rudravaram R, Pogaku R, Rao LV
(2009) Bioconversion of de-oiled rice bran (DORB) hemicellu-
losic hydrolysate into ethanol by Pichia stipitis NCM3499 under
optimized conditions. Int J Food Eng 5(1)
14. Palmqvist E, Hahn-Hagerdal B (2000) Fermentation of ligno-
cellulosic hydrolysates. II: inhibitors and mechanisms of inhibi-
tion. Bioresour Technol 74:25–33
15. Qureshi N, Ezeji T, Ebener J, Dien B, Cotta M, Blaschek H
(2008) Butanol production by Clostridium beijerinckii. Part I: use
of acid and enzyme hydrolyzed corn fiber. Bioresour Technol
99(13):5915–5922
16. Chandel AK, Kumar KR, Singh AK, Kuhad RC (2007) Detoxi-
fication of sugarcane bagasse hydrolysate improves ethanol pro-
duction by Candida shehatae NCIM 3501. Bioresour Technol
98:1947–1950
17. Kosaka T, Nakayama S, Nakaya K, Yoshino S, Furukawa K
(2007) Characterization of the sol operon in butanol-hyperpro-
ducing Clostridium saccharoperbutylacetonicum strain N1-4 and
its degeneration mechanism. Biosc Biotechnol Biochem 71(1):
58–68
18. Tashiro Y, Takeda K, Kobayashi G, Sonomoto K (2005) High
production of acetone–butanol–ethanol with high cell density
culture by cell-recycling and bleeding. J Biotechnol 120(2):
197–206
19. Al-Shorgani N, Ali E, Kalil M, Yusoff W (2011) Bioconversion
of butyric acid to butanol by Clostridium sacchar-operbutylacetonicum N1-4 (ATCC 13564) in a limited nutrient
medium. BioEnergy Res 1–7. doi:10.1007/s12155-011-9126-6
20. Aryee ANA, Simpson BK (2009) Comparative studies on the
yield and quality of solvent-extracted oil from salmon skin.
J Food Eng 92(3):353–358
21. Weil JR, Dien B, Bothast R, Hendrickson R, Mosier NS, Ladisch
MR (2002) Removal of fermentation inhibitors formed during
pretreatment of biomass by polymeric adsorbents. Ind Eng Chem
Res 41(24):6132–6138. doi:10.1021/ie0201056
22. Fond O, Matta-Ammouri G, Petitdemange H, Engasser JM
(1985) The role of acids on the production of acetone and butanol
by Clostridium acetobutylicum. Appl Microbiol Biotechnol
22(3):195–200. doi:10.1007/bf00253609
23. Qureshi N, Saha BC, Hector RE, Cotta MA (2008) Removal of
fermentation inhibitors from alkaline peroxide pretreated and
enzymatically hydrolyzed wheat straw: production of butanol
from hydrolysate using Clostridium beijerinckii in batch reactors.
Biomass Bioenergy 32(12):1353–1358
Bioprocess Biosyst Eng
123
Author's personal copy

24. Ezeji TC, Qureshi N, Blaschek HP (2007) Bioproduction of
butanol from biomass: from genes to bioreactors. Curr Opin
Biotechnol 18(3):220–227
25. Purwadi R, Niklasson C, Taherzadeh MJ (2004) Kinetic study of
detoxification of dilute-acid hydrolyzates by Ca(OH)2. J Bio-
technol 114(1–2):187–198
26. Larsson S, Palmqvist E, Hahn-Hagerdal B, Tengborg C, Stenberg
K, Zacchi G, Nilvebrant N-O (1999) The generation of fermen-
tation inhibitors during dilute acid hydrolysis of softwood.
Enzyme Microb Technol 24(3–4):151–159
27. Liu Z, Ying Y, Li F, Ma C, Xu P (2010) Butanol production by
Clostridium beijerinckii ATCC 55025 from wheat bran. J Ind
Microbiol Biotechnol 37(5):495–501. doi:10.1007/s10295-010-
0695-8
28. Marchal R, Rebeller M, Vandecasteele JP (1984) Direct bio-
conversion of alkali-pretreated straw using simultaneous enzy-
matic hydrolysis and acetone–butanol fermentation. Biotechnol
Lett 6(8):523–528. doi:10.1007/bf00139996
29. Soni B, Das K, Ghose T (1982) Bioconversion of agro-wastes
into acetone butanol. Biotechnol Lett 4:19–22
30. Hipolito CN, Crabbe E, Badillo CM, Zarrabal OC, Morales Mora
MA, Flores GP, MdA HC, Ishizaki A (2008) Bioconversion of
industrial wastewater from palm oil processing to butanol by
Clostridium saccharoperbutylacetonicum N1-4 (ATCC 13564).
J Clean Prod 16(5):632–638
Bioprocess Biosyst Eng
123
Author's personal copy




















