
Bell , A. and Everhart, M.J. 2011. Remains of small ornithurine birds from a Late Cretaceous...
9
Remains of small ornithurine birds from a Late Cretaceous (Cenomanian) microsite in Russell County, north-central Kansas AlyssA Bell 1 And MichAel J. everhArt 2 1. Department of Earth Sciences, University of Southern California, Los Angeles, California 90007 Corresponding author: [email protected] 2. Sternberg Museum of Natural History, Fort Hays State University, Hays, Kansas 67601 [email protected] Our analysis of vertebrate remains collected from a Cenomanian microsite in the Lincoln Limestone, the basal-most member of the Greenhorn Formation in Russell County, Kan- sas, identifies ornithurine (sensu Chiappe, 1996) avian fossils. The specimens presented here are slightly younger than the oldest avian remains known from North America (from the Woodbine Formation, Texas) and of a similar age as the avians from the Asheville Formation in Saskatchewan, Canada. This find thus connects the extensive geographic range of the earliest North American birds from Saskatchewan south through central Kansas and on to Texas. The specimens discussed here are fragmentary, yet show features definitive of ornithurine birds. One specimen is attributable to Ichthyornis, whereas another preserves teeth characteristic of ornithurine birds. In addition to bird bones, the microsite yielded numerous bony fish remains, shark teeth, coniasaur vertebrae, and other lizard bones. Key words: Avian, Ichthyornis, ornithurine, Lincoln Limestone, Greenhorn Formation, teeth t rAnsActions of the KAnsAs AcAdeMy of science Vol. 114, no. 1-2 p. 115-123 (2011) introduction In terms of actual specimens, sediments deposited by the Western Interior Seaway in Kansas have produced one of the most abundant avifaunas known from the Late Cretaceous. This fauna is predominantly composed of the gull-like Ichthyornithiformes and the flightless Hesperornithiformes. The vast majority of these fossils have come from the Smoky Hill Chalk (Coniacian - Campanian) of the Niobrara Formation (Marsh 1880; Martin and Lim 2002). Until recently (Everhart and Bell 2009), the earliest known bird remains in Kansas consisted of the proximal end of a right carpometacarpal of an Ichthyornis from the middle Turonian of Ellis County (Walker 1967; Clarke 2004; Shimada and Fernandes 2006). Mudge (1866) and Snow (1887) reported “bird tracks” from the Dakota Formation (middle Cenomanian) of Kansas in Clay and Ellsworth counties, respectively. Both specimens are now missing, but have since been reinterpreted as the tracks of small dinosaurs (Liggett, 2005). Valid bird trackways have been reported from the Cenomanian of Colorado by Mehl (1931) and from the Early Cretaceous (Aptian) of British Columbia, Canada by Currie (1981). Much less is known of avian evolution prior to the Coniacian-Campanian in North America. The best preserved avifauna older than that in the Niobrara Formation was collected from the Belle Fourche member of the Asheville Formation (middle Cenomanian) of Saskatchewan, Canada (Tokaryk et al. 1997). These bone beds preserve numerous remains of the hesperornithiforms Pasquiaornis hardei and P. tankei, three unidentified species of Ichthyornis, and one unidentified enantiornithine, as well as numerous non-avian remains including plesiosaur, dolichosaur, turtle, and fish (Cumbaa et al. 2006). Outside of these Saskatchewan sites avian remains are limited to single occurrences (e.g. Tykoski and Fiorillo 2010; Everhart and Bell 2009).
Transcript of Bell , A. and Everhart, M.J. 2011. Remains of small ornithurine birds from a Late Cretaceous...

Remains of small ornithurine birds from a Late Cretaceous (Cenomanian) microsite in Russell County, north-central KansasAlyssA Bell1 And MichAel J. everhArt2
1. Department of Earth Sciences, University of Southern California, Los Angeles, California 90007 Corresponding author: [email protected] 2. Sternberg Museum of Natural History, Fort Hays State University, Hays, Kansas 67601 [email protected]
Our analysis of vertebrate remains collected from a Cenomanian microsite in the Lincoln Limestone, the basal-most member of the Greenhorn Formation in Russell County, Kan-sas, identifies ornithurine (sensu Chiappe, 1996) avian fossils. The specimens presented here are slightly younger than the oldest avian remains known from North America (from the Woodbine Formation, Texas) and of a similar age as the avians from the Asheville Formation in Saskatchewan, Canada. This find thus connects the extensive geographic range of the earliest North American birds from Saskatchewan south through central Kansas and on to Texas. The specimens discussed here are fragmentary, yet show features definitive of ornithurine birds. One specimen is attributable to Ichthyornis, whereas another preserves teeth characteristic of ornithurine birds. In addition to bird bones, the microsite yielded numerous bony fish remains, shark teeth, coniasaur vertebrae, and other lizard bones.
Key words: Avian, Ichthyornis, ornithurine, Lincoln Limestone, Greenhorn Formation, teeth
trAnsActions of the KAnsAs AcAdeMy of science
Vol. 114, no. 1-2 p. 115-123 (2011)
introduction
In terms of actual specimens, sediments deposited by the Western Interior Seaway in Kansas have produced one of the most abundant avifaunas known from the Late Cretaceous. This fauna is predominantly composed of the gull-like Ichthyornithiformes and the flightless Hesperornithiformes. The vast majority of these fossils have come from the Smoky Hill Chalk (Coniacian - Campanian) of the Niobrara Formation (Marsh 1880; Martin and Lim 2002). Until recently (Everhart and Bell 2009), the earliest known bird remains in Kansas consisted of the proximal end of a right carpometacarpal of an Ichthyornis from the middle Turonian of Ellis County (Walker 1967; Clarke 2004; Shimada and Fernandes 2006).
Mudge (1866) and Snow (1887) reported “bird tracks” from the Dakota Formation (middle Cenomanian) of Kansas in Clay and Ellsworth counties, respectively. Both specimens are now
missing, but have since been reinterpreted as the tracks of small dinosaurs (Liggett, 2005). Valid bird trackways have been reported from the Cenomanian of Colorado by Mehl (1931) and from the Early Cretaceous (Aptian) of British Columbia, Canada by Currie (1981).
Much less is known of avian evolution prior to the Coniacian-Campanian in North America. The best preserved avifauna older than that in the Niobrara Formation was collected from the Belle Fourche member of the Asheville Formation (middle Cenomanian) of Saskatchewan, Canada (Tokaryk et al. 1997). These bone beds preserve numerous remains of the hesperornithiforms Pasquiaornis hardei and P. tankei, three unidentified species of Ichthyornis, and one unidentified enantiornithine, as well as numerous non-avian remains including plesiosaur, dolichosaur, turtle, and fish (Cumbaa et al. 2006). Outside of these Saskatchewan sites avian remains are limited to single occurrences (e.g. Tykoski and Fiorillo 2010; Everhart and Bell 2009).
Mike
Sticky Note
Bell , A. and Everhart, M.J. 2011. Remains of small ornithurine birds from a Late Cretaceous (Cenomanian) microsite in Russell County, north-central Kansas. Kansas Academy of Science, Transactions 114(1-2):115-123.

116 Bell and Everhart
Here we present an analysis of a new microsite stratigraphically located at the base of the Greenhorn Formation (upper middle Cenomanian) in Russell County, Kansas (Fig. 1). Although the majority of the bones from the microsite are too fragmentary for identification, three elements were identified as avian: the proximal end of a coracoid and femur and two jaw fragments containing teeth. Differences in size suggest that the remains are from more than one individual. The significance of the geologic occurrence and descriptions of these elements is discussed below.
Abbreviations: FHSM – Fort Hays University, Sternberg Museum of Natural History, Hays, Kansas; YPM – Yale Peabody Museum, New Haven, Connecticut
GeoloGicAl occurrence
The microsite is located in a lag deposit in the basal Lincoln Member of the Greenhorn Formation, in direct contact with the underlying Graneros Shale. The layer in which the microsite is located varies from 1 to 4 cm in thickness and appears to consist of material that had filled in shallow erosional depressions in the top of the Graneros Shale. The site is located in a road cut just south of Wilson Lake in Russell County, Kansas. The bird remains were collected in an area of less than 1 m2.
The avian jaw fragments were picked from a sieved portion of the matrix examined under a binocular microscope by the second author (MJE). In addition to the avian remains, the locality also yielded shark, teleost, and pliosaur teeth as well as a coniasaur jaw, vertebrae, and other bones. Other localities in the basal Lincoln Limestone in Kansas have produced large numbers of vertebrate remains (Liggett et al. 2005; Martin and Shimada 2008; Shimada and Martin 2008).
The microsite is located stratigraphically just above two markers used in North American stratigraphic correlations, a thick bentonite seam referred to as the “X-bentonite” (Hattin 1965; Miggins 2004) and the underlying Ostrea beloiti bed (McNeil and Caldwell 1981; Collom 2000). Using 40 Ar/39 Ar dating, the X-bentonite has been variably dated at 94.93 ± 0.53 Ma in the Frontier Formation in Wyoming (Obradovich, 1993), 95.11 ± 0.16 Ma in the Greenhorn Limestone in Colorado (Shimada et al. 2006), and 95.53 ± 0.159 Ma below the Lincoln Limestone Member in Kansas (Miggins 2004).The Russell County microsite is considered to be of a similar age as the Canadian localities (Cumbaa et al. 2006), as definitive stratigraphic analyses have not been conducted in order to precisely determine the relative ages of these two localities.
Figure 1. Locality map showing Russell County, Kansas (highlighted in gray) with the generalized stratigraphy. The arrow indicates the approximate stratigraphic location of the microsite presented here.

Transactions of the Kansas Academy of Science 114(1-2), 2011 117
The oldest definitive avian remains in North America were collected from the lower middle Cenomanian Lewisville Member of the Woodbine Formation in Tarrant County, Texas. This specimen was described by Tykoski and Fiorillo (2010) as an enantiornithine, Flexomornis howei, from the fragmentary, disarticulated remains of a single individual. The minimum age of the overlying Arlington member of the Woodbine Formation has been inferred to be either 96.01 Ma (Gradstein et al. 2005) or 95.73 ± 0.61 Ma (Cobban et al. 2006). Although the error ranges on these dates overlap slightly with those presented for southern portions of the X-bentonite (see above), as Flexomornis was found in the underlying Lewisville Member it is reasonable to treat it as the oldest North American bird fossil, closely followed by the Bainbridge and Carrot river material in Canada and the specimens presented here, however specimens from all three locations could, in fact, be of a similar age.
PAleoenvironMent
During Middle Cenomanian time, Russell County was at the eastern edge of the transgressing Western Interior Sea and under the delta of a large, southwesterly flowing river that deposited the Dakota Formation (Hattin 1965; Hattin and Siemers 1978, fig. 3). As the Greenhorn Transgression progressed, the environment transitioned from marginal marine to near-shore open marine conditions as evidenced by the deposition of the Graneros Shale (Hattin and Siemers, 1978). The transitional beds between Dakota and the Graneros contain abundant teeth of sharks and bony fishes (Everhart et al. 2004; Everhart 2011) as well as teeth, bones and osteoderms of crocodiles. As the water deepened, the Graneros Shale was overlain, disconformably, by the basal Lincoln Limestone Member of the Greenhorn Formation. Lag deposits of calcarenites containing abundant shark teeth and other vertebrate remains sit directly on the scoured upper surface of the Graneros (Shimada and Martin 2008). Liggett et al.
(2005) reported on vertebrate remains from another basal Lincoln deposit, including two teeth and a portion of a pliosaur paddle (cf. Brachauchenius lucasi), coniasaur vertebrae, teeth and a jaw fragment (FHSM VP-13999), and the earliest known remains of a pterosaur (FHSM VP-13998) from Kansas, as well as abundant teeth of sharks and bony fish.
In addition to the bird bones, the basal Lincoln locality reported here produced vertebrae (FHSM VP-17569) and a right dentary of a coniasaur (FHSM VP-15825), a turtle vertebra, and two pliosaur teeth (FHSM VP-14782 and VP-17567), as well as numerous teeth and vertebrae from bony fishes and sharks (MJE, pers. obs. 2002-2009). The bones and teeth are jumbled together in the deposit, often lying in contact with one another. The remains of pterosaurs, lizards, and birds in the predominantly marine basal Lincoln Limestone are indicative of a nearby terrestrial environment.
systeMAtic PAleontoloGy
Aves Linnaeus 1758 Ornithuromorpha Chiappe 1996 Ornithurae Haeckel 1866 Ichthyornithes Marsh 1873 unidentified Ichthyornithes
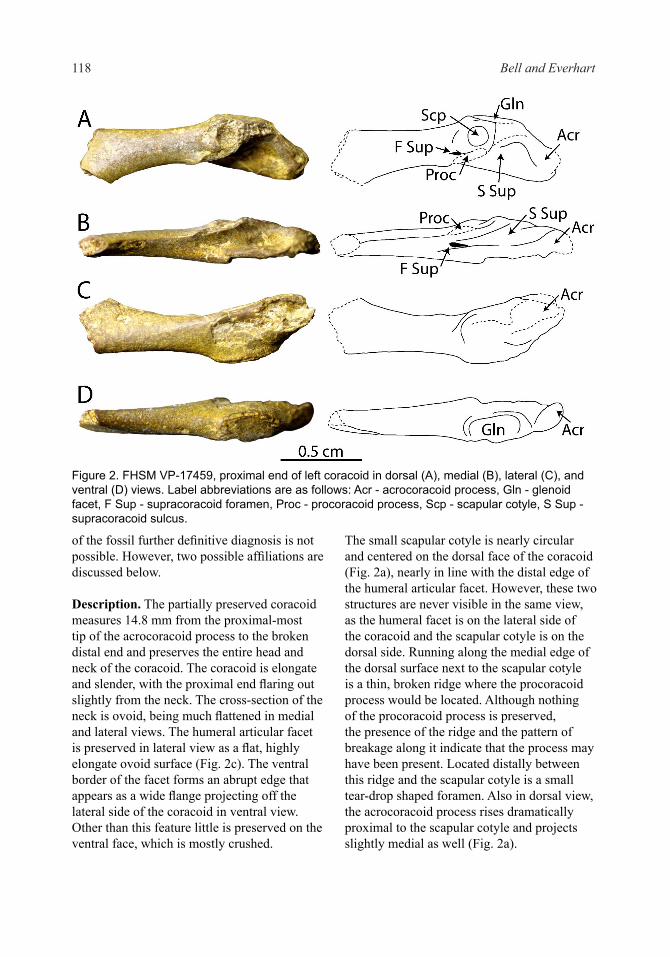
Material - FHSM VP-17459, proximal end of a left coracoid.
Diagnosis - The proximal end of a left coracoid, FHSM VP-17459 (Fig. 2), is identified as avian by its strut-like shape and localized facets for the articulation of the scapula and humerus. It is further assigned to the clade Ornithuromorpha (sensu Chiappe 2002) by the placement of the humeral articular facet well ventral to the acrocoracoid process. The presence of a prominent, medially projecting acrocoracoid process identifies the coracoid as belonging to the Ichthyornithes (Marsh 1873). Due to the fragmentary nature
118 Bell and Everhart
of the fossil further definitive diagnosis is not possible. However, two possible affiliations are discussed below.
Description. The partially preserved coracoid measures 14.8 mm from the proximal-most tip of the acrocoracoid process to the broken distal end and preserves the entire head and neck of the coracoid. The coracoid is elongate and slender, with the proximal end flaring out slightly from the neck. The cross-section of the neck is ovoid, being much flattened in medial and lateral views. The humeral articular facet is preserved in lateral view as a flat, highly elongate ovoid surface (Fig. 2c). The ventral border of the facet forms an abrupt edge that appears as a wide flange projecting off the lateral side of the coracoid in ventral view. Other than this feature little is preserved on the ventral face, which is mostly crushed.
The small scapular cotyle is nearly circular and centered on the dorsal face of the coracoid (Fig. 2a), nearly in line with the distal edge of the humeral articular facet. However, these two structures are never visible in the same view, as the humeral facet is on the lateral side of the coracoid and the scapular cotyle is on the dorsal side. Running along the medial edge of the dorsal surface next to the scapular cotyle is a thin, broken ridge where the procoracoid process would be located. Although nothing of the procoracoid process is preserved, the presence of the ridge and the pattern of breakage along it indicate that the process may have been present. Located distally between this ridge and the scapular cotyle is a small tear-drop shaped foramen. Also in dorsal view, the acrocoracoid process rises dramatically proximal to the scapular cotyle and projects slightly medial as well (Fig. 2a).
Figure 2. FHSM VP-17459, proximal end of left coracoid in dorsal (A), medial (B), lateral (C), and ventral (D) views. Label abbreviations are as follows: Acr - acrocoracoid process, Gln - glenoid facet, F Sup - supracoracoid foramen, Proc - procoracoid process, Scp - scapular cotyle, S Sup - supracoracoid sulcus.

Transactions of the Kansas Academy of Science 114(1-2), 2011 119
In medial view a wide supracoracoid sulcus is present (Fig. 2b), bordered ventrally by the acrocoracoid process and dorsally by the broken ridge discussed above. At the distal-most extent of this sulcus a distinct ovoid foramen is located. This foramen appears to connect to the smaller foramen seen in dorsal view medio-distal to the scapular cotyle and is probably for the passage of the supracoracoideus nerve.
Remarks - This coracoid exhibits a number of similarities to the ichthyornithiform Iaceornis marshi (Clarke 2004), despite being about 75% as large. FHSM VP-17459 shares with the holotype of I. marshi (YPM 1734) a robust, medially-expanded acrocoracoid process. In I. marshi this process bears a slightly bulbous, hooked projection on the proximal-most end, whereas this region of FHSM VP-17459 is heavily eroded, preventing a more exact comparison. Furthermore, the progressively more medio-distal arrangement of the glenoid facet, the scapular cotyles, and the foramen for the supracoracoideus nerve is similar between these specimens. The procoracoid process in I. marshi is small and runs along the medial edge of the supracoracoideus nerve foramen. Although a procoracoid process itself is not preserved in FHSM VP-17459, an area of breakage similar in proportion and location to the procoracoid process of I. marshi is present. FHSM VP-17459 differs from I. marshi in the placement of the glenoid facet, which is rotated further away laterally from the scapular cotyles such that it faces completely laterally. Also, the sulcus for the supracoracoideus muscle is a deeply excavated groove in the medial surface formed by the medial inflection of the acrocoracoid process and the flat dorsal surface of the scapular cotyles and inferred procoracoid process.
Marsh (1872) suggested that Ichthyornis was roughly the size of a pigeon, without providing further explanation. In terms of size, the coracoids of Ichthyornis specimens range from 125% (Iaceornis marshii YPM 1734) to
300% (Ichthyornis validus, YPM 1446) larger than FHSM VP-17459 in measurements of the neck width, diameter of the glenoid and scapular facets, and width of the acrocoracoid process. Based on the size of the coracoid, FHSM VP-17459 would be similar in mass to modern birds such as the pigeons Ptilinopus melanospila (94 g; Schleucher 2002) and P. iozonus (108 g; Klenova et al. 2008), the buttonquail Turnix suscitator (97g; Kendeigh et al. 1977 ), and the lark Melanocorypha yeltoniensis (60 g; Dunning 1993).
Aves Linnaeus 1758 unidentified avian
Material - FHSM VP-17461, one maxillary and one mandibular fragment, each containing one intact tooth.
Diagnosis - FHSM VP-17461 is identified as avian by the recurved, unserrated teeth in combination with numerous foramina along the jaws. Due to the fragmentary nature of the specimen further definitive diagnosis is not possible.
Description - Both fragments of FHSM VP-17461 contain one tooth (Fig. 3). Both teeth are conical and non-serrated. FHSM VP-17461a (Fig. 3a), a fragment of maxilla with a single tooth, measures 1.15 mm from the tip of the crown to the medial jaw line and is only very slightly recurved. FHSM VP-17461b (Fig. 3b), another fragment of mandible with a single tooth, measures 1.22 mm from the tip of the crown to the medial jaw line and is fully recurved. Both jaw fragments are similarly shaped. On the lateral surface the upper edge of the jaw is curved convexly, whereas the lower is concave. On FHSM VP-17461a, a series of foramina are visible in the concave portion of the jaw, whereas one is visible in a similar location on FHSM VP-17461b. The medial surface of the jaw fragments bear a longitudinal groove along the bottom edge and a noticeable bulge around the insertion of each tooth. The top margin of FHSM VP-17461b

120 Bell and Everhart
appears to contain a narrow groove into which the tooth is set, however this may be only due to crushing, as this area is better preserved in FHSM VP-17461a and shows a smooth, rounded top of the jaw on either side of the tooth.
Remarks - Other avian teeth from the Late Cretaceous of Kansas are very similar in form to the teeth described here, however they vary widely in size. In the jaw fragments presented here, the alveoli measure 0.87 mm (FHSM VP-17461a) and 0.99 mm (FHSM VP-17461b)
at the widest point, which is smaller than the range of 1.2 mm to 2.1 mm listed by Clarke (2004) for Ichthyornis dispar (YPM 1470 and FHSM (SMNH) VP-2503, and the range of 1.95 mm to 2.54 mm measured for Hesperornis regalis (YPM 1206). Because neither the teeth nor jaws bear any unique or distinguishing features and the small variation in tooth size may be the result of ontogeny or individual variation, it is not possible to make a more definitive identification of FHSM VP-17461.
Aves Linnaeus 1758 unidentified avian
Material - FHSM VP-17458, proximal end of a right femur.
Diagnosis - FHSM VP-17458 is identified as avian by the preservation of a distinct femoral neck. Due to the fragmentary nature of the fossil further definitive diagnosis is not possible.
Description - The proximal end of a right femur, FHSM VP-17458, has been extensively crushed and preserves only the caudal face of the bone (Fig. 4). The head of the femur is separated from the shaft by a distinct neck. The medially-facing surface of the head possesses a depression for the capital ligament proximally offset on the face. Although the trochanter of the femur is not preserved due to crushing, it appears to have risen to a similar proximal height as the head of the femur. A shallow depression separates the head from the trochanter. A faint groove runs longitudinally down the center of the shaft, the proximal extent of which is obscured by breakage. Due to the extensive crushing little else can be observed, however the shaft of the femur does not appear to taper from the proximal end to the distal-most end.
Remarks - Despite its poor state of preservation, FHSM VP-17458 bears similarities to both Ichthyornis dispar (Marsh, 1873) and Iaceornis marshi (Clarke 2004).
Figure 3. FHSM VP-17461a, maxilla fragment in medial (A) and lateral (C) view. FHSM VP-17461b, mandible fragment in medial (B) and lateral (D) views. Abbreviation: f - foramen.

Transactions of the Kansas Academy of Science 114(1-2), 2011 121
The faint longitudinal groove in FHSM VP-17458 is similar to that seen in better-preserved specimens of both I. dispar and I. marshi (as figured in Clarke 2004). This groove may be the result of similar taphonomic processes; however, it is not observed in other ornithuromorph birds and may, therefore, be a unique feature. Unlike I. dispar, the trochanter of FHSM VP-17458 does not extend proximally beyond the head of the femur. Rather, it more closely resembles the state in I. marshi, where the majority of the trochanter extends to a similar level as the head. The orientation of the head of the femur of FHSM VP-17458 closely resembles that of I. dispar, both of which differ from I. marshi in having a medially-facing surface of the head with a proximally offset fovea for the capital ligament. In I. marshi the face of the head is tilted proximally such that it is almost in alignment with the proximal margin of the femur. FHSM VP-17458 is virtually identical in size to the holotype of Ichthyornis dispar (YPM 1450) and smaller than Iaceornis marshii (YPM 1734), as seen in measurements of the diameter of the femoral head, medio-lateral
width of the proximal end, and proximal width of the shaft (taken just below the head and trochanter). However, Clarke (2004, pg 27) noted that over 90% of Ichthyornis specimens are larger than YPM 1450, making the holotype one of the smallest known specimens. Modern birds that present similar femoral measurements to FHSM VP-17458 include the pigeon Caloenas nicobarica (620 g; Sonnenfield et al. 2002), as well as the rails Aramides cajanea (400 g; McNab 2002) and Rallus longirostris (261 g; Hammons et al. 1968), and the tinamou Crypturellus noctivagus (800 g; Dunning 1993).
conclusions
The identification of numerous avian fossils from deposits predating other avian sites in North America helps push our knowledge of avian evolution in North America back to the earliest Late Cretaceous. Although highly fragmentary, these remains do represent examples of ornithurine birds, some of which may likely belong to the ichthyornithiforms, which are better known from the younger Niobrara Formation. Future work on other sites from this age may yield additional specimens from which a better picture of early avian evolution in North America can be determined.
AcKnowledGMents
We would like to thank Keith Ewell for the discovery of the basal Lincoln Limestone microsite and the donation of the FHSM VP-17458 and VP-17459 specimens, as well as Dr. Luis Chiappe and two anonymous reviewers for comments on the manuscript.
literAture cited
Chiappe, L.M. 1996. The phylogenetic position of the Cretaceous birds of Argentina: Enantiornithes and Patagopteryx defarrariisi. Courier Forschungsintitut Senckenberg 181: 55-63.
Figure 4. FHSM VP-17458, proximal end of right femur in cranial view. Abbreviations are as follows: CL - fovea for the capital ligament, H - femoral head, Tr - trochanter. Only the cranial view is shown due to the heavy crushing and distortion of the femur in other views.

122 Bell and Everhart
Chiappe, L.M. 2002. Basal bird phylogeny: problems and solutions. pp. 448-472 in L. M. Chiappe and L. M. Witmer (eds.), Mesozoic Birds: above the heads of dinosaurs. University of California Press, Berkeley.
Clarke, J.A. 2004. Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bulletin of the American Museum of Natural History 286:1-179.
Cobban, W.A., Walaszczyk, I., Obradovich, J.D. and McKinney, K.C. 2006. A USGS zonal table for the Upper Cretaceous middle Cenomanian-Maastrichtian of the Western Interior of the United States based on ammonites, inoceramids, and radiometric ages. U.S. Geological Survey, Open File Report 2006-1250.
Collom, C.J. 2000. High-resolution stratigraphy, regional correlation and report of molluscan faunas: Colorado group (Cenomanian - Coniacian interval), Late Cretaceous, east-central Saskatchewan. pp. 82-97 in Summary of Investigations 2000, Volume 1, Saskatchewan Geological Survey, Saskatchewan Energy and Mines, Miscellaneous Reports 2000-4.1.
Currie, P.J. 1981. Bird footprints from the Gething Formation (Aptian, Lower Cretaceous) of northeastern British Columbia, Canada. Journal of Vertebrate Paleontology 1(3-4):257-264.
Cumbaa, S.L., Schröder-Adams, C., Day, R.G. and Phillips, A. 2006. Cenomanian bone bed faunas from the northeastern margin, Western Interior Seaway, Canada. pp. 139-155 in S. G. Lucas and R. M. Sullivan (eds.), Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35.
Dunning, J.B., Jr. 1993. CRC handbook of avian body masses. CRC Press, Boca Raton, Florida.
Everhart, M.J. Occurrence of the hybodont shark genus Meristodonoides (Chondrich-thyes; Hybodontiformes) in the Cretaceous of Kansas. Kansas Academy of Science, Transactions 114(1-2):33-46.
Everhart, M.J. and Bell, A. 2009. A hesperornithiform limb bone from the basal Greenhorn Formation (Late Cretaceous; middle Cenomanian) of north central Kansas. Journal of Vertebrate Paleontology 29:952-956.
Everhart, M.J., Everhart, P.A. and Ewell, K. 2004. A marine ichthyofauna from the Upper Dakota Sandstone (Late Cretaceous). Joint Annual Meeting of the Kansas and Missouri Academies of Science, p. 48 (abstract).
Gradstein, F., Ogg, J. and Smith, A. 2005. A geologic time scale 2004. Cambridge University Press, Cambridge, UK.
Hammons, R.L., Hughes, M.R. and Moldenhauer, R.R. 1988. Body water and water flux in fresh water and sea-water acclimated clapper rails, Rallus longirostris. Comparative Biochemistry and Physiology A, Comparative Physiology 91:539-541.
Hattin, D.E. 1965. Stratigraphy of the Graneros Shale (Upper Cretaceous) in central Kansas. Bulletin of the Kansas University Geological Survey 178:1-83.
Hattin, D.E. and Siemers, C.T. 1978. Upper Cretaceous stratigraphy and depositional environments of western Kansas. Guidebook Series 3, AAPG/SEPM Annual Meeting, Kansas Geological Survey, 55 pp.
Kendeigh S.C., Dol’nik, V.R. and Gavrilov, V.M. 1977. Avian energetics. pp. 129–204 in Pinowski J. and Kendeigh, S.C. (eds.), Granivorous Birds in Ecosystems. Cambridge University Press, Cambridge.
Klenova, A.V., Volodin, I.A. and Volodina, E.V. 2008. Duet structure provides information about pair identity in the red-crowned crane (Grus japonensis). Journal of Ethology 26:317-325.
Liggett, G.A. 2005. A review of the dinosaurs from Kansas. Kansas Academy of Science. Transactions 108(1-2):1-14.
Liggett, G.A., Shimada, K., Bennett, S.C. and Schumacher, B. 2005. Cenomanian (Late Cretaceous) reptiles from northwestern Russell County, Kansas. PaleoBios 25(2):9-17.

Transactions of the Kansas Academy of Science 114(1-2), 2011 123
Marsh, O.C. 1872. Notice of a new and remarkable fossil bird. American Journal of Science 4(22):344.
Marsh, O.C. 1873. On a new sub-class of fossil birds (Odontornithes). American Journal of Science 5:161-162.
Marsh, O.C. 1880. Odontornithes: A monograph on the extinct toothed birds of North America. U.S. Geological Exploration of the 40th Parallel (King), Volume 7, 201 pp., 34 pl.
Martin, D.J. and Shimada, K. 2008. Lithostratigraphy and depositional environment of the Lincoln Limestone Member of the Greenhorn Limestone (Upper Cretaceous) in Russell County, Kansas, with special reference to the basal beds. Kansas Academy of Science, Transactions 111(1-2):79-92.
Martin, L.D. and Lim, J.-D. 2002. New information on the hesperornithiform radiation. pp. 165-169 in Zhou, Z., and Zhang, F. (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution.
McNab, B.K., 2002. Short-term energy conservation in endotherms in relation to body mass, habits, and environment. Journal of Thermal Biology 27:459-466.
McNeil, D.H. and Caldwell, W.G.E. 1981. Cretaceous rocks and their foraminifera in the Manitoba Escarpment. Geological Association of Canada Special Paper 21:1-439.
Mehl, M.G. 1931. Additions to the vertebrate record of the Dakota Sandstone. American Journal of Science 21:441-452.
Miggins, D. 2004. 40 Ar/39 Ar results for Cretaceous bentonite beds of Colorado and Kansas. Report provided to USDA Forest Service under intra-agency agreement 04-IA-1102, 11 pp.
Mudge, B.F. 1866. Discovery of fossil footmarks in the Liassic (?) Formation in Kansas. American Journal of Science 41(122):174-176.
Obradovich, J.D. 1993. A Cretaceous time scale. pp. 379-396 in W.G.E. Caldwell and E.G. Kauffman(eds.), Evolution of the Western Interior Basin, Geologic Association of Canada, Special Paper 39.
Schleucher, E. 2002. Metabolism, body temperature, and thermal conductance of fruit-doves (Aves: Columbidae, Treroninae). Comparative Biochemistry and Physiology A, Molecular and Integrative Physiology 131:417-428.
Shimada, K. and Fernandes. M.V. 2006. Ichthyornis sp. (Aves: Ichthyornithiformes) from the lower Turonian (Upper Cretaceous) of western Kansas. Kansas Academy of Science, Transactions 109(1-2):21-26.
Shimada, K. and Martin, D.J. 2008. Fossil fishes from the basal Greenhorn Limestone (Upper Cretaceous, Late Cenomanian) in Russell County, Kansas. pp. 89-103 in G.H. Farleyand J.R. Choate (eds.), Unlocking the Unknown; Papers Honoring Dr. Richard Zakrzewski, Fort Hays Studies, Special Issue No. 2, 153 pp., Fort Hays State University, Hays, Kansas.
Shimada, K., Schumacher, B.A., Parkin, J.A. and Palermo, J.M. 2006. Fossil marine vertebrates from the lowermost Greenhorn Limestone (Upper Cretaceous: middle Cenomanian) in southeastern Colorado. Journal of Paleontology 80:1-45.
Snow, F.H. 1887. On the discovery of a fossil bird track in the Dakota Sandstone. Kansas Academy of Science, Transactions 10:3-6.
Sonnenfield, J.M., Carpenter, J.W., Garner, M.M., Collins, D. and Joslin, J. 2002. Pheochromocytoma in a Nicobar pigeon. Journal of Avian Medicine and Surgery 16:306-308.
Tokaryk, T.T., Cumbaa, S.L. and Storer, J.E. 1997. Early Late Cretaceous birds from Saskatchewan, Canada: the oldest diverse avifauna known from North America. Journal of Vertebrate Paleontology 17:172-176.
Tykoski, R.S. and Fiorillo, A.R. 2010. An enantiornithine bird from the lower Middle Cenomanian of Texas. Journal of Vertebrate Paleontology 30:288-292.
Walker, M.V. 1967. Revival of interest in the toothed birds of Kansas. Kansas Academy of Science, Transactions 70(1):60-66.




















