
Bacteria in Weathered Basaltic Glass, Iceland
18
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Open University Library] On: 20 January 2011 Access details: Access Details: [subscription number 907468021] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Geomicrobiology Journal Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713722957 Bacteria in Weathered Basaltic Glass, Iceland Charles S. Cockell a ; Karen Olsson a ; Felicity Knowles a ; Laura Kelly a ; Aude Herrera a ; Thorsteinn Thorsteinsson b ; Viggo Marteinsson c a Geomicrobiology Research Group, CEPSAR, Open University, Milton Keynes, UK b Hydrology Division, National Energy Authority, Reykjavik, Iceland c Matis/Prokaria - Food Research, Innovation and Safety, Reykjavik, Iceland To cite this Article Cockell, Charles S. , Olsson, Karen , Knowles, Felicity , Kelly, Laura , Herrera, Aude , Thorsteinsson, Thorsteinn and Marteinsson, Viggo(2009) 'Bacteria in Weathered Basaltic Glass, Iceland', Geomicrobiology Journal, 26: 7, 491 — 507 To link to this Article: DOI: 10.1080/01490450903061101 URL: http://dx.doi.org/10.1080/01490450903061101 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Bacteria in Weathered Basaltic Glass, Iceland

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Open University Library]On: 20 January 2011Access details: Access Details: [subscription number 907468021]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Geomicrobiology JournalPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713722957
Bacteria in Weathered Basaltic Glass, IcelandCharles S. Cockella; Karen Olssona; Felicity Knowlesa; Laura Kellya; Aude Herreraa; ThorsteinnThorsteinssonb; Viggo Marteinssonc
a Geomicrobiology Research Group, CEPSAR, Open University, Milton Keynes, UK b HydrologyDivision, National Energy Authority, Reykjavik, Iceland c Matis/Prokaria - Food Research, Innovationand Safety, Reykjavik, Iceland
To cite this Article Cockell, Charles S. , Olsson, Karen , Knowles, Felicity , Kelly, Laura , Herrera, Aude , Thorsteinsson,Thorsteinn and Marteinsson, Viggo(2009) 'Bacteria in Weathered Basaltic Glass, Iceland', Geomicrobiology Journal, 26: 7,491 — 507To link to this Article: DOI: 10.1080/01490450903061101URL: http://dx.doi.org/10.1080/01490450903061101
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Geomicrobiology Journal, 26:491–507, 2009Copyright © Taylor & Francis Group, LLCISSN: 0149-0451 print / 1521-0529 onlineDOI: 10.1080/01490450903061101
Bacteria in Weathered Basaltic Glass, Iceland
Charles S. Cockell,1 Karen Olsson,1 Felicity Knowles,1 Laura Kelly,1
Aude Herrera,1 Thorsteinn Thorsteinsson,2 and Viggo Marteinsson3
1Geomicrobiology Research Group, CEPSAR, Open University, Milton Keynes, MK7 6AA, UK2Hydrology Division, National Energy Authority, Reykjavik, Iceland3Matis/Prokaria – Food Research, Innovation and Safety, Skulagata 4, 101 Reykjavik, Iceland
Bacteria play an important role in rock weathering and yettheir diversity and potential activity in the terrestrial rock weath-ering environment is poorly understood. Culture and culture-independent methods (16S rDNA) were used to investigate thepopulations of bacteria inhabiting a basaltic glass/palagonite sub-glacial (hyaloclastite) deposit subject to weathering in Iceland.The rock hosts a diverse microbial community. The 16S rDNAclones were dominated by Actinobacteria, Proteobacteria, Bac-teroidetes and Acidobacteria. Representatives of Gemmatimon-adetes and Verrucomicrobia were present. Isolation of organismson basalt/palagonite yielded only two isolates, an actinobacteriumand a Bacteroidetes, showing that the active species, at least inthe time scale of laboratory cultivation, are a small proportionof the total diversity. Firmicutes and Actinobacteria were isolatedwhen basalt/palagonite was supplemented with an organic source.Many of the isolates demonstrated tolerance to transition metals(Cr, Cu, Zn, Ni, Co) naturally present in the rock. The growthof the isolates was inhibited at typical pH values for Icelandicrain, which suggests that the increase in pH caused by the con-sumption of protons in rock weathering, for example by palagoniteformation, may play a role in defining which organisms are ac-tive. Colonization experiments show that the filamentous growthhabit of the actinobacterium isolated on basalt/palagonite allows itto actively invade and colonise the basaltic glass. The filamentousgrowth of some actinobacteria may be an important contributorto their role in systemic interstitial rock weathering in the naturalenvironment.
Keywords Actinobacteria, basaltic glass, Iceland, volcanic,weathering
Received 5 February 2009; accepted 20 April 2009.This work was made possible and supported by the Royal Society
and the Leverhulme Trust (Project No. F/00 269/N) to Charles Cockell.Felicity Knowles was supported by a Society for General Microbiology(SGM) Vacation Studentship. We thank Heather Davies for work onthe TEM sections.
Address correspondence to Charles S. Cockell, GeomicrobiologyResearch Group, CEPSAR, Open University, Milton Keynes, MK76AA, UK. E-mail: [email protected]
INTRODUCTIONThe weathering of volcanic minerals is recognized to make a
significant contribution to the global silicate weathering budget(Louvat and Allegre 1998; Dessert et al. 2001; Kisakurek et al.2004), thus influencing carbon dioxide drawdown and climatecontrol, since carbon dioxide is consumed in mineral weatheringreactions. For example, on land the Deccan Traps in India, withan estimated area of 106 km2, are thought to account for 5%of the global silicate weathering flux (Gaillardet et al. 1999).In total, basalt rocks may account for over 30% of the globalcarbon dioxide drawdown in silicate weathering (Dessert et al.2001, 2003).
Microorganisms are thought to play an important part in rockweathering. The role of lichens in volcanic rock weathering hasreceived a great deal of attention (Jackson and Keller 1970;Adamo et al. 1993) and recent laboratory work has begun toelucidate the role of bacteria (e.g., Wu et al. 2007a). A prokary-otic involvement in volcanic rock weathering has been inferredin deep-ocean basaltic glass in which a diversity of microbialalteration textures has been reported (e.g., Thorseth et al. 1992;Fisk et al. 1998; Torsvik et al. 1998; Furnes and Staudigel 1999;Thorseth et al. 2001; Etienne and Dupont 2002; Thorseth et al.2003).
The prokaryotic diversity of volcanic environments has onlyrecently received detailed attention. A study of seafloor basaltfrom arctic spreading ridges showed a diverse bacterial com-munity, whose composition varied with age. Firmicutes wasthe only phylum detected in basalts of all three ages studied(20 yr, 0.1 Ma and 1 Ma) (Lysnes et al. 2004). Bacteroidetesand α-proteobacteria were only found in the 20 yr and 0.1 Masamples, but not in the oldest basalts. Deep-ocean crust environ-ments harbour a diversity of bacterial phyla, some of which haveclades only associated with the ocean crust environment (rootingin the α- and γ -proteobacteria, Acidobacteria, Actinobacteria,Verrucomicrobia), with one archeal clade apparently endemicto basalts (Mason et al. 2007). Proteobacteria were found todominate seafloor basalts from Hawaii and the East Pacific Rise(Santelli et al. 2008) and the basalts had higher diversity thanother deep-sea environments.
491
Downloaded By: [Open University Library] At: 14:43 20 January 2011

492 C. S. COCKELL ET AL.
In terrestrial volcanic environments, bacterial communitycomposition is known to vary with the age of lava flow. In Hawai-ian basalts only Actinobacteria and Acidobacteria were foundin rocks of all ages in lava flows dating back to 1700 (Gomez-Alvarez et al. 2006). Clones from forested lava were dominatedby Proteobacteria. In all other sites clones were dominated byunclassified sequences, including sequences belonging to theChloroflexi, an observation similar to studies of obsidian fromIceland in which a high proportion of unclassified Chloroflexi-related sequences were reported (Herrera et al. 2008, 2009).
In terrestrial environments, it is not clear to what extent bacte-ria are endemic to the endolithic environment. In so far as rockseventually weather to soils, the populations of organisms asso-ciated with the rock-microbe interface might be similar to thoseoccurring in soils, particularly in extensively weathered rocks orthose that host soil and clay-like weathered mineral phases (e.g.,palagonite) in the pore spaces (Thorseth et al. 1991; Stroncikand Schminke, 2002).
A further challenge concerning the examination of the bac-terial involvement in volcanic rock weathering is to estab-lish which organisms might be active. Although a culture-independent (e.g., 16S rDNA) approach can yield an insightinto which organisms are in the rock, it does not reveal whichorganisms are metabolically active. The extraction of RNA frommetabolically active cells might yield evidence of activity, butRNA can still reside in inactive organisms.
In this paper, our investigations were focused on hyalo-clastite. Hyaloclastite is subglacial basaltic glass formed whenlava erupts and comes into contact with ice and/or water andquenches rapidly to form volcanic glass. It is widespread in vol-canic regions associated with icy or water-rich environments.In this study we isolated heterotrophs from weathered basalticglass and sought to understand their potential activity in theweathered rock environment.
MATERIALS AND METHODS
Sampling LocationSamples of weathered hyaloclastite were collected from
Valafell, near Hekla volcano, at location 64◦4.83′ N, 19◦32.53′
W in the south of Iceland (Figure 1a). The material is a mixtureof basaltic glass and its weathering product, palagonite (Cockellet al. 2009). The material is of Pleistocene age (<0.8 Ma old)and forms exposed outcrops throughout the study area (Figure1b) (Jacobsson and Gudmundsson 2008). Some of the materialcontains clasts of crystalline basalt, but these were not exam-ined in this study. Samples of between 1 and 10 cm diameterwere collected from exposed outcrops into sterile plastic bagsto avoid contamination.
Porosity and Surface Area of MaterialTo investigate the physical space available for microorgan-
isms within the rock, two parameters were measured. The sur-
face area of the material was determined by BET (Brunauer–Emmett–Teller) analysis (5-point) (Meritics Ltd., Dunstable,UK) (Brunauer et al. 1938). The porosity of the material wasdetermined by mercury intrusion porosimetry (MCA Services,Meldreth, Cambridge, UK). For both BET and mercury intru-sion porosimetry the mean of four bulk samples of approximatemass 6 grams each was determined.
Scanning Electron Microscopy (SEM) and ElectronMicroprobe Analysis (EMPA)
Blocks of hyaloclastite were examined by Scanning Elec-tron Microscopy (SEM). Samples of rock were carbon coated(15–20 nm thickness) on aluminium stubs and examined in sec-ondary electron mode at a 20 kV accelerating voltage and 7–15mm working distance. The SEM was a Quanta 3D dual beamFIBSEM (FEI, Oregon, USA).
Electron microprobe analysis was used to examine mineralcomposition. EMPA was carried out on polished samples ofmaterial embedded within Struers Epofix resin using a CamecaSX100 electron microprobe (EMP) operating in wavelength-dispersion mode. The following standards and X-ray lines wereused: jadeite (Na Kα), forsterite (Mg Kα), feldspar (Al, Si andK Kα), fluorapatite (P Kα), synthetic KCl (Cl Kα), crocoite (CrKα), rutile (Ti Kα), bustamite (Mn and Ca Kα), hematite (FeKα), Ni metal (Ni Kα). An operating voltage of 20 kV and probecurrent of 20 nA (measured on a Faraday cage) were used. Abeam of 10 µm in diameter was used to minimize volatilizationeffects. Count times varied from 20 to 80 seconds per element.
Microbial EnumerationsTo determine the number of cells within the material two
representative rock samples (each approximately 5 cm in diam-eter) were broken in half in a laminar flow hood and materialwas obtained from the center with a sterilised small chisel. Onegram of rock was obtained from the interior of each sample andpooled. The material was crushed into small pieces (<0.25 mmdiameter) using a sterile mortar and pestle and added to 10 mLof a 1% solution of 0.01% tween-80 and 0.05% peptone in ddH2O as described by Weirich and Schweisfurth (1985). Peptonewas added to encourage amphoteric detachment. The solutionwith rock was sonicated briefly for 10 s at 4◦C to assist cell de-tachment from rock particles. Aliquots were stained with SybrGreen I DNA staining dye (Invitrogen, Paisley, UK) accordingto the manufacturer’s instructions. Controls were also preparedusing autoclaved basalt glass/palagonite from the same samplesto check for non-specific binding of the dye to the palagonite.
Organisms were enumerated using a Leica DMRP micro-scope equipped with epifluorescence (Leica Microsystems,Bensheim, Germany). Stained cells were observed fluorescinggreen using an excitation waveband of 450–490 nm (Leica filtercube I3) and an emission long band cutoff filter of >515 nm.Enumerations were corrected to give the numbers of cells pergram (dried) of material. The procedure was repeated using
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 493
FIG. 1. A. Location of the hyaloclastite examined in this study (Valafell, 64◦4.83′ N, 19◦32.53′ W) in relation to major geological outcrops. B. Image of fieldsite showing outcrops of weathered basaltic glass. Grey clasts evident within the material are crystalline basalt. The rock in the foreground containing clasts isapproximately 50 cm across.
material obtained from the top 5 mm of the 5 cm diametersample to investigate the differences between the interior andnear-surface environment of rocks. Three independent sampleswere examined (expressed as mean and standard deviation).
Microbial Isolation and Culturing of OrganismsTo isolate organisms potentially active within the rock, plates
were prepared using the hyaloclastite with no amendments.Samples of weathered basalt glass were crushed in a mortarand pestle until they were a fine powder. Of this powder, 0.5 gwas added to 20 mL ddH2O and 2% agar (Oxoid, Basingstoke,UK; Bacteriological Agar No. 1) and autoclaved at 121◦C for 15min. The agar was previously washed according to the methodof Ryan (1950) to minimise organic sources from the agar. Theseplates are hereafter referred to as BP (Basalt/Palagonite) plates.These unamended plates were used to culture organisms that
live only on the available minerals and associated organics fromthe rock (however, autoclaved microorganisms within the rockare likely to be lysed, resulting in a potentially higher organicavailability than the natural unautoclaved material).
‘Amended BP plates’ were also prepared with the additionof 0.01 g NH4SO4, 0.005 g NaH2PO4, 0.01 g of yeast extract(0.05% wt%) and 0.01g glucose (0.05% wt%) per 20 mL ddH2Oof to provide organisms with nitrogen, sulfur, phosphorus andan additional carbon source to favour growth. The yeast andglucose concentrations were provided at low concentrations tofavour oligotrophic organisms.
The rock fragments used to inoculate BP and amended BPplates were obtained from a fresh sample of hyaloclastite. Anaseptically collected sample of rock was broken in half witha sterile hammer in a laminar flow hood. A sterile coring de-vice was used to grind ∼0.25 g of material from the centreof the rock. The material was collected in a sterile Petri dish.
Downloaded By: [Open University Library] At: 14:43 20 January 2011

494 C. S. COCKELL ET AL.
Approximately 0.05 g of this material was scattered onto theBP and amended BP plates, which were incubated at 21◦C for4 months. Colonies were removed and repeatedly subcultureduntil single isolates were obtained. Colonies from unamendedBP plates were subcultured further on unamended BP plates.
Colonies from amended BP plates were isolated further on1/10 TSA (Tryptone Soya Agar) (0.3 g/L) (Oxoid, Basingstoke,UK) plates as the addition of basalt/palagonite was not found tobe necessary for their further growth.
To investigate the potential for the isolates to grow in the nat-ural environment all isolates were inoculated onto two typesof agar plates: (1) BP plates in which the basalt/palagonitewas added after autoclaving the agar to prevent alteration toindigenous rock organics caused by autoclaving. In practice,so few organisms grew on just basalt and palagonite in thetime frame of the experiment that contamination from unau-toclaved basalt/palagonite did not interfere with the assess-ment of the growth of the isolate, (2) BP plates in which thebasalt/palagonite/water mixture was sonicated for 20 min at4◦C. This procedure was used to increase the availability of nat-ural rock organics (simulating cell death and lysis), but withoutautoclaving.
Isolates obtained from unamended BP plates were also grownin solution to eliminate the possibility that the agar was provid-ing a carbon source for growth. A colony of each isolate wasadded to 5 g of autoclaved basalt/palagonite in 10 mL ddH2O.After 2 months, suspended material was removed, stained withSYBR Green I (as described above), and examined under epi-fluoresence microscopy to determine if growth had occurred bycomparing cell density to the initial inoculated solution.
The Bacteroidetes (BP16) obtained from unamended BPplates would not grow on a variety of defined minimal saltsmedia including TSA plates, but could be grown on 2 types ofmedia free of basaltic rock: (1) ACM (Autoclaved Cyanobac-terium Medium) agar plates. ACM plates were made from Trisminimal medium. Tris minimal medium consisted of 6.06 g ofTris-HCl, 4.68 g of NaCl, 1.49 g of KCl, 1.07 g of NH4Cl,0.43 g of Na2SO4, 0.2 g of MgCl2· 6H2O, 0.03 g of CaCl2·2H2O, 0.23 g of Na2HPO4, 0.005 g of iron ammonium citrateand 1 mL of trace element solution SL7 (10 mL 25% HCl, 1.5g FeCl2· 4H2O, 190 mg CoCl2· 6H2O, 100 mg MnCl2· 4H2O,70 mg ZnCl2, 62 mg H3BO3, 36 mg Na2MoO4.2H2O, 24 mgNiCl2· 6H2O, 17 mg CuCl2· 2H2O in 1L ddH2O) as describedby Mergeay et al. (1985) in 2% agar (Bacteriological Agar No.1; Oxoid, Basingstoke, UK).
The cyanobacterium Chroococcidiopsis 029 (Culture Collec-tion of Microorganisms from Extreme Environments, Universityof Rome Tor Vergata) was used as a carbon source. This unicel-lular, non-motile cyanobacterium is found growing in endolithichabitats in hot deserts such as the Negev in Israel and cold desertssuch as the Ross Desert of the Antarctic (e.g., Friedmann andOcampo-Friedmann 1985; Billi and Grilli Caiola 1996). Thecultured organism was added to the medium at a concentra-tion of ∼107 cells/mL after culturing as described previously
(Cockell et al. 2005) and the medium was autoclaved at 121◦Cfor 15 min, (2) BG-11 medium (Rippka et al. 1979) with addedyeast extract at a concentration of 0.05% wt%.
The actinobacterial isolate (BP17) obtained from unamendedBP plates was able to grow on 1/10 TSA plates, 1/10 nutrientbroth plates and Tris minimal medium plates with 0.2% glucose.The latter were used for subsequent physiological experiments.
pH Tolerance of IsolatesThe pH range of growth of the isolates was determined by
growing them on 1/10 nutrient broth agar plates adjusted to pH5.6, 7.1 and 8.8 with HCl or NaOH. After one and three weeks,the plates were scored for the presence or absence of growth. Inthe case of isolate BP16, BG-11/yeast extract agar plates wereused.
Heavy Metal Resistance of IsolatesIsolates were plated on a transition metal-amended (TMA)
Tris minimal medium agar plate (Tris minimal medium com-position as described here). The carbon source was supplied as0.2% glucose. In the case of BP16, transition metal-amendedBG-11/yeast extract plates were used. In this study transitionmetals were added to correspond to the highest concentrationsdetermined in rocks of tholeiitic to olivine-tholeiitic concentra-tion by Walsh and Clarke (1982) as follows: metal ion in ppm(and molarity in an equivalent mass of water) (Cr [K2CrO4], 120(2.3 mM); Cu [CuSO4], 160 (2.5 mM); Zn [ZnCl2], 150 (2.3mM); Ni [NiCl2· 6H2O], 140 (2.4 mM); Co [CoSO4· 7H2O],40 (0.68 mM); V [VOSO4], 400 (78 mM)). The compound inbrackets is the form in which the compound was incorporatedinto the medium. These values are similar to a range of basaltsexamined in the Western Volcanic Zone, Iceland (Sinton et al.2005), although the chromium concentration we used is lower(about half) than most values reported by Sinton et al.
Tolerance to individual metals was determined for each iso-late. Metals were added at a concentration of 4 mM (the con-centration of the metal ion itself) in separate agars and theywere serially diluted by 2-fold to a concentration of 0.125 mMand poured into plates. Plates were marked into 15 differentsections and isolates were streaked onto the plates. The follow-ing metals (with the corresponding compounds in which theywere delivered as above) were examined: Cr, Cu, Zn, Ni, Co.We also tested the isolates for resistance to cadmium (in theform of CdSO4· 8H2O). The minimum concentration at whichgrowth was no longer observed was recorded as the MinimumInhibitory Concentration (MIC).
Additional Physiology of Isolates That Grew on BasalticGlass/Palagonite
Additional physiological data were gathered on the two iso-lates obtained from unamended BP plates. Freeze-thaw toler-ance was tested by placing ∼0.02 g of cell material obtained
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 495
from agar plates into 3 separate Eppendorf tubes. The tubeswere frozen at -20◦C. They were removed the following day,the contents of one of the tubes was spread onto BP plates andthe tubes re-frozen. The process was repeated for the remainingtubes. Plates were examined for growth after 3 weeks.
Desiccation tolerance of the isolates was tested by placing∼0.02 g of cell material directly from plates into 2 separateEppendorf tubes with a hole in the lid. The tubes were placedinto a 1.5 L desiccator containing 200 g silica gel. After 3 daysand 1 week the contents of the tubes was plated onto BP plates.Plates were examined for growth after 3 weeks.
The two isolates were tested for psychro- and thermal toler-ance by incubating inoculated BP plates at 4◦, 29◦ and 40◦C.Plates were examined for growth after three weeks.
The range of pH tolerance of growth was tested by growingthe Bacteroidetes and actinobacterium in BG-11/yeast extractand Tris minimal medium (0.2% glucose), respectively, modi-fied with HCl and NaOH to produce a series of media from pH3.0 to 11. Growth was examined after 4 weeks.
Salt tolerance was tested by growing the Bacteroidetesand actinobacterium on BG-11/yeast extract and Tris minimalmedium (0.2% glucose) plates respectively, amended with NaClto the following percentages: 0.1, 0.2, 0.5, 1 and 5%.
The isolates were examined for their tolerance to selectedantibiotics. Antibiotics were selected that have previously beenshown to be produced by soil actinobacteria (Nwosu 2001):Lincomycin, Bacitracin, Penicillin, Nystatin, Streptomycin,Cephalosporin, Kanamycin, Cycloserine, Polymyxin, Grise-ofulvin, Tetracycline, Chlortetracycline. Antibiotic resistancewas tested by growing the Bacteroidetes and actinobacteriumon BG-11/yeast and Tris minimal medium plates, respectively.Antibiotics were dissolved to a final concentration of 0, 5, 15,30, 40 µg/mL or 0, 8U, 40, 60, 80 Units/mL as appropriate.Growth was examined after 4 weeks. All reagents for cultur-ing and antibiotics were obtained from Sigma Aldrich (SigmaAldrich, Gillingham, UK).
Transmission Electron Microscopy (TEM) of IsolatesThe isolates were scraped off agar plates with a sterile scalpel
blade and dropped into 2.5% glutaraldehyde in 0.1M sodium ca-codylate pH 7.4 containing 0.1% ruthenium red (cacod/RR). Bymicrocentrifugation and then aspiration at each step, the clumpsof cells were post-fixed in 2% osmium tetroxide in cacod/RR,dehydrated through an ascending series of acetone, infiltratedin Epon (Shell Chemical Company, San Francisco, CA, USA)minus accelerator at 60◦C with 6 changes. The samples werefinally embedded in complete Epon and polymerised at 60◦Cfor 48 h.
Ultrathin sections of approximately 70 nm were cut using adiamond knife and collected on copper slot grids with a formvarfilm. The sections were counter-stained with uranyl acetate andReynolds lead citrate before examination in a JEM 1400 trans-mission electron microscope (JEOL, Tokyo, Japan) operating
at 80 kV. Digital images were acquired using an AMT XR60digital camera (AMT Imaging, Danvers, MA, USA).
Colonisation of Basaltic Glass by Isolates fromBasalt/Palagonite Plates
Samples of hyaloclastite were embedded within StruersEpofix resin (Struers, Solihull, UK). The blocks were cutthrough the hyaloclastite and the surfaces exposing the rockwere polished. The resin did not penetrate into the vesicles ofthe rock so that they remained unobstructed. These blocks werecut in half perpendicular to the polished surface so that thehyaloclastite was exposed on one side of the block. The blockswere placed into Petri dishes with the polished surface facingupwards and basalt/palagonite agar was poured into the dish un-til it was at the same level as the surface of the block. The contactbetween the agar and the hyaloclastite encouraged moisture up-take by the rock by capillary action from the agar, maintainingmoist conditions, and it allowed bacterial colonisation from theBP agar onto and into the rock. Plates were inoculated eitherwith the Bacteroidetes (BP16) or the actinobacterial (BP17) iso-late obtained from unamended BP plates by adding an inoculumto the agar surface next to the cut side of the block so that colo-nization would occur onto the surface of the polished glass andpotentially into it. The Petri dishes were sealed and incubated at21◦C. After one month the blocks were removed from the agarand the growth of the organisms on the rock surface and withinthe vesicles was observed under SEM as described above.
16S rDNA Analysis of Whole Community Genomic DNAFor community analysis DNA was extracted from whole
basalt/palagonite rocks that were stored aseptically at −80◦Cin sterile bags. Four separate rocks collected from the field site(each separated by approximately three meters) were crushedin a sterilized hollow metal cylinder using a sterile heavy metalplunger (Herrera and Cockell 2007; Herrera et al. 2008) andfive grams of the pooled rock powder was used for subsequentanalysis. All manipulations were carried out under aseptic con-ditions to avoid contamination. DNA was extracted by usingthe PowerMax Soil DNA Isolation Kit (MoBio Laboratories,Cambridge, UK) according to the manufacturer’s instructions.
Ribosomal DNA (rDNA) was PCR amplified directly fromthe DNA extracted from the rock using the primers, Com1 (5′-CAG CAG CCG CGG TAA TAC-3′) (Schwieger and Tebbe1998) and pH (5′-AAG GAG GTG ATC CAG CCG CA-3′)(Bruce et al. 1992). The total volume of the PCR reaction was50 µl and contained: 5 µl of template DNA; 200 µM dNTP;1 µM primers; 2.5U of Taq DNA polymerase; 1X Buffer (50mM KCl; 10 mM Tris-HCl pH8.3 at 25◦C, 1.5 mM Mg(OAC)2)and 1X TaqMaster solution (Eppendorf, Flowgen Bioscience,Nottingham, UK). Amplification consisted of incubation at94◦C for 5 min: this was followed by 35 cycles of 1 min at94◦C, 1 min at 55◦C and a 1 min extension at 72◦C, with a finalextension of 5 min at 72◦C. Negative controls without DNAtemplate were run for all PCR reactions.
Downloaded By: [Open University Library] At: 14:43 20 January 2011

496 C. S. COCKELL ET AL.
The 16S rDNA PCR product was extracted and purified froma 0.8% (w/v) agarose gel (Invitrogen, Paisley, UK) by means ofthe GenElute Gel Extraction kit (Sigma-Aldrich, Poole, UK) ac-cording to the manufacturer’s instructions. The purified productwas ligated overnight at 4◦C, with the pCR
©R 4-TOPO©R vec-
tor. Chemical transformation was conducted with the OneShotTOP10 chemically competent Escherichia coli from the TOPO-TA cloning kit (Invitrogen, Paisley, UK) as per manufacturer’sinstructions. Clones containing inserts were stored in 25% (v/v)glycerol at −20◦C.
The 16S rDNA inserts were sequenced using the T7 andT3 universal primers which are within the cloning vector andcover the complete insert (School of Biological Sciences, Uni-versity of Edinburgh, UK). Each sequence was submitted tothe CHECK CHIMERA program of the Ribosomal DatabaseProject (RDP) (Cole et al. 2003). Potential chimeras were elim-inated. The phylum of each sequence was determined using theClassifier tool of the RDP. Rarefaction analysis was carried outusing FastGroupII (Yu et al. 2006).
For phylogenetic tree construction we used the hypervari-able regions V5 and V6 of the 16S rDNA gene (Chakravortyet al. 2007) corresponding to E.coli nucleotide positions 718-1190. The sequences were aligned with representative bacte-rial sequences from the Ribosomal Database Project (RDP)obtained with the Sequence Match and Classifier tools. Se-quences were aligned using ClustalX (Thompson et al. 1997).The phylogenetic trees were constructed using the neighbour-joining method (Saitou and Nei 1987) and Kimura 2-parameterfor distance correction (Kimura 1980) using the Phylip v. 3.67inference package (DNADIST and NEIGHBOR). SEQBOOTand CONSENSE were used to provide confidence estimatesfor tree topologies for a consensus tree. These trees were thenused as user trees to construct a maximum likelihood tree us-ing DNAML. Trees were visualized with NjPLOT (Perriere andGouy 1996) and TreeView.
16S rDNA Identification of IsolatesMicrobial isolates were identified by direct PCR from
colonies. DNA template was prepared by dispersing a bacte-rial colony in 50 µl of 10 mM TE buffer, pH 7.5, and freeze-thawing at −20◦C. PCRs were performed with the primers pA(5′-AGA GTT TGA TCC TGG CTC AG-3′) and pH (5′-AAGGAG GTG ATC CAG CCG CA-3′) (Bruce et al. 1992). Thereaction mixture contained: 5 µl of template DNA; 200 µMdNTP; 1 µM primers; 1.25U of Taq DNA polymerase; 1X PCRBuffer (20 mM Tris-HCl (pH 8.4), 50 mM KCl) and 2.5 mMMgCl (Invitrogen, Paisley, UK).
Amplification consisted of incubation at 94◦C for 15 min(which was used to assist cell disruption): this was followed by35 cycles of 1 min at 94◦C, 1 min at 55◦C and a 1 min extension at72◦C, with a final extension of 5 min at 72◦C. The PCR productswere run on a 0.8% (w/v) agar gel and extracted as previouslydescribed. DNA sequencing of the PCR products was carried
out directly by Mclab (San Francisco, USA). Sequences werephylogenetically classified and their nearest putative phyloge-netic relatives identified in the GenBank database. For BP16 and17, in addition to sequencing using primers pA and pH, primersCom1 (as above) and Com2 (5′-CCG TCA ATT CCT TTG AGTTT-3′) were used to obtain greater coverage of the 16S rDNA.The sequences were used to obtain 16S rDNA sequences of1463 and 1429 bp, respectively.
Nucleotide Sequence Accession NumbersThe partial sequences of the bacterial clones were deposited
in GenBank (accession numbers EU621919 to EU621981 andFJ360637 to FJ360683). The partial sequences of the isolateswere deposited in GenBank (accession numbers EU295462 toEU295477 and accession number FJ379585).
RESULTS
Surface Area and PorosityThe surface area (BET) of the hyaloclastite was 90 m2/g.
The porosity of the rock was 26.1%. The total porosity resultingfrom pores >2 µm diameter was 19.5%.
Scanning Electron MicroscopyThe material was highly vesiculated (Figure 2A). The re-
sults we observed were qualitatively similar to those reportedby Thorseth et al. (1991, 1992). Palagonite layers were devel-oped on the surface of the glass as the first stable product ofaqueous mafic glass weathering, with alteration textures occur-ring at the glass-palagonite interface. The palagonite (Pal) rindshad a maximum thickness of ∼10 ± 2 (s.d.) µm (n = 30 mea-surements) and exfoliated from the surface of the glass (Glass)(Figure 2B).
EMPA analysis showed the composition (mean of 10 sam-ples) of the glass and its weathered palagonite product for majorcations (expressed as percent oxides) to be: Na2O (glass- 2.57,palagonite - 0.07), K2O (0.39, 0.14), MgO (7.09, 4.13), CaO(10.71, 3.35), MnO (0.20, 0.09), FeO (13.20, 13.90), Al2O3
(15.31, 15.68), SiO2 (46.10, 40.65), TiO2 (2.63, 2.51), Total(98.23, 80.63). The total percentages are less than 100 on ac-count of the presence of bound water within the material, par-ticularly within the claylike palagonite.
Microbial EnumerationsThe total numbers of cells in the rock interior was estimated
as 1.28 ± 0.l2 × 107 cells/g. In the upper 5 mm of rock thenumber of cells was 3.19 ± 0.67 × 107 cells/g.
Isolation and Physiology of OrganismsOn the unamended BP plates 2 organisms were isolated.
One isolate formed red colonies around basalt/palagonite grains
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 497
FIG. 2. Secondary scanning electron microscopic images of the basaltic glass. A. Vesicular nature of microbial habitat (scale bar 500 µm) in which bacterialive, B. Image showing palagonite rind (Pal) (∼10 µm thick) on basaltic glass (Glass). In the center, the palagonite has become detached showing altered glassbeneath (scale bar 30 µm).
after 4 months (BP16) and it corresponded to Bacteroidetes(Table 1). Only 2 grains exhibited growth of this organism outof approximately 200 grains.
The second organism isolated formed salmon-colouredwaxy colonies around grains (BP17) and it correspondedto Actinobacteria (Streptomyces). The organism was abun-dant and formed colonies around approximately half of thebasalt/palagonite grains on the unamended BP plates.
Both isolates obtained on unamended BP plates grew in so-lution made from dd H2O mixed with ground basalt/palagonitewith no other amendments. They grew on BP plates made withunautoclaved basalt/palagonite and sonicated basalt/palagonite.
After 2 months, bacterial colonies developed around the pe-riphery of rock fragments on the amended BP plates. A total of15 isolates were obtained for subsequent analysis.
The 16S rDNA characteristics of all 17 isolates obtainedon the BP plates are shown in Table 1. Eleven of the isolatesobtained on amended BP plates were Firmicutes and 4 wereActinobacteria.
Isolates from the amended BP plates grew on unamended BPplates (with sonicated and autoclaved hyaloclastite), but slowly,with colonies visible after 2 months.
pH Tolerance of the IsolatesTable 2 shows that all of the isolates grew at pH 7.1 and
8.8. However, only nine of the isolates could grow at pH 5.6after 1 week. After 3 weeks all isolates showed growth at pH5.6, suggesting that growth was retarded at this pH, but was notprevented.
Metal Tolerance of the IsolatesNone of the isolates could grow on transition metal-amended
Tris minimal medium agar with combined metals at the concen-trations reported by Walsh and Clarke (1982). The MIC (min-imal inhibitory concentration) of the metals for some of theisolates was <0.125 mM. For each metal there was at least oneisolate that had an MIC >0.125 mM (Table 2).
Additional Physiology of Isolates That Grew on BasalticGlass/Palagonite
Both isolates obtained from unamended BP plates showedviability after 3 freeze-thaw cycles and 1 week of desiccation.The Bacteroidetes isolate showed no growth at 4◦, 29◦ or 40◦C(but it grew at 21◦C), the actinobacterium grew at 29◦C, butnot at the other temperatures. The pH range of growth of theBacteroidetes isolate was pH 6–9, that of the actinobacteriumwas pH 6–10. The Bacteroidetes grew at 0.2% NaCl, but not0.5%, the actinobacterial isolate grew at 1% NaCl, but not 5%.
The antibiotic resistance of the 2 isolates (highest concen-tration at which growth was observed for the Bacteroidetes andactinobacterium; X denotes no resistance) was: Lincomycin (X,X), Bacitracin (X, X), Penicillin (8U, 8U), Nystatin (80U, 80U),Streptomycin (5 µg, X), Cephalosporin (X, 40 µg), Kanamycin(15 µg, X), Cycloserine (X, 40 µg), Polymyxin (80U, 80U),Griseofulvin (40 µg/mL, 40 µg), Tetracycline (X, 40 µg),Chlortetracycline (X, 40 µg).
Transmission Electron Microscopy (TEM) of IsolatesTransmission electron micrographs of each isolate are shown
in Figure 3. Sectioned colonies of the Bacteroidetes isolate
Downloaded By: [Open University Library] At: 14:43 20 January 2011

TAB
LE
116
SrD
NA
info
rmat
ion
for
17is
olat
es(i
sola
tes
BP1
toB
P15
from
amen
ded
BP
plat
esan
dis
olat
esB
P16
and
BP1
7fr
omun
amen
ded
BP
plat
es)
Isol
ate
and
Gen
Ban
kA
cces
sion
No.
Nea
rest
mat
ch(c
ultu
red
orun
cultu
red)
Acc
essi
onN
o.of
near
est
mat
ch
Perc
enta
gene
ares
tm
atch
Com
men
ts
BP1
EU
2954
62U
ncul
ture
dac
tinob
acte
rium
DQ
8281
31.1
99%
Isol
ate
shar
es99
%si
mila
rity
with
othe
run
cultu
red
actin
obac
teri
a.C
lose
stcu
lture
dba
cter
ium
isa
soil
isol
ate
(AF1
8315
1)B
P2E
U29
5463
Paen
ibac
illu
ssp
.A
Y28
9507
99%
Isol
ate
shar
es99
%si
mila
rity
with
othe
rcu
lture
dPa
enib
acil
lus
spp.
BP3
EU
2954
64F
irm
icut
esba
cter
ium
EF6
8291
793
%U
ncul
ture
dF
irm
icut
es(E
F682
917.
1)
BP4
EU
2954
65B
acil
lus
sp.
DQ
1904
2810
0%Sh
ares
100%
sim
ilari
tyw
ithB
acil
lus
sp.f
rom
Moj
ave
dese
rtpl
aya
BP5
EU
2954
66B
acil
lus
sp.X
JU-1
DQ
4854
1410
0%Sh
ows
100%
sim
ilari
tyto
Bac
illu
ssi
mpl
ex.I
sola
tes
from
lake
san
dso
ils.
BP6
EU
2954
67B
acil
lus
sim
plex
AB
3637
3810
0%Sh
ares
100%
sim
ilari
tyto
othe
rB
acil
lus
sim
plex
stra
ins
BP7
EU
2954
68B
acil
lus
sp.
DQ
1904
2810
0%Sh
ares
100%
sim
ilari
tyw
ithB
acil
lus
sp.f
rom
Moj
ave
dese
rtpl
aya
BP8
EU
2954
69U
ncul
ture
dF
irm
icut
esA
Y91
7512
98%
AY
9175
12is
olat
edfr
omre
cent
Haw
aiia
nvo
lcan
icde
posi
ts.C
lose
stcu
lture
dm
atch
(97%
),D
Q15
4300
soil
bact
eriu
m.
BP9
EU
2954
70B
acil
lus
sp.
DQ
1904
2810
0%Sh
ares
100%
sim
ilari
tyw
ithB
acil
lus
sp.f
rom
Moj
ave
dese
rtpl
aya
BP1
0E
U29
5471
Bac
illu
ssp
.D
Q19
0428
100%
Shar
es10
0%si
mila
rity
with
Bac
illu
ssp
.fro
mM
ojav
ede
sert
play
a
BP1
1E
U29
5472
Stre
ptom
yces
sp.
EF6
7264
810
0%O
rgan
ism
isol
ated
from
the
rhiz
osph
ere
BP1
2E
U29
5473
Bac
teri
umSL
5.44
DQ
5171
8299
%Sh
ows
99%
sim
ilari
tyto
DQ
1904
28
BP1
3E
U29
5474
Bac
illu
sci
rcul
ans
EF1
0098
99%
Sim
ilar
sequ
ence
sim
ilari
tyw
ithot
her
Bac
illu
sci
rcul
ans
stra
ins
BP1
4E
U29
5475
Stre
ptom
yces
sp.
EF0
9311
210
0%O
rgan
ism
isol
ated
from
surf
ace
ofSc
ottis
hm
onum
ents
.Nex
tclo
sest
mat
ch(9
9%,E
F010
652)
isol
ated
from
pale
olith
icca
ves
BP1
5E
U29
5476
Unc
ultu
red
actin
obac
teri
umA
Y13
3106
100%
Clo
sest
cultu
red
isol
ate
from
anea
rth-
wor
mbu
rrow
(AY
0394
52)
BP1
6E
U29
5477
Unc
ultu
red
orga
nism
clon
eD
LE
129
EF1
2761
494
%C
lose
stm
atch
foun
din
olde
stst
udie
dic
eon
eart
h.O
rgan
ism
also
show
s94
%si
mila
rity
with
uncu
lture
dse
quen
cefr
omde
epgl
acie
ric
ein
Tib
et.N
eare
stcu
lture
dis
olat
e(9
4%)
isa
Bac
tero
idet
esba
cter
ium
(aer
obic
hete
rotr
ophi
cba
cter
ium
from
soil
ofL
aG
orce
Mou
ntai
ns(8
6◦ 30′ S
,147
◦ W),
Ant
arct
ica.
(DQ
3517
28))
BP1
7FJ
3795
85C
ultu
red
Stre
ptom
ycin
aea
bact
eriu
mC
L4.
126
FM17
3947
.110
0%A
ctin
obac
teri
umw
ith10
0%se
quen
cesi
mila
rity
isol
ated
from
tufa
core
,H
artz
Mou
ntai
n,G
erm
any.
498
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 499
TABLE 2Physiological characteristics of 17 bacterial isolates
Isolate No. Spore formation1,2 pH tolerance3 Metal resistance4
5.6 7.1 8.8 Cd Ni Co Zn Cu CrBP1 + − + + a a a a a aBP2 + + + + 0.25 a a 2.0 a aBP3 + − + + a a a a 0.25 aBP4 + − + + 0.25 a a a 0.25 aBP5 + + + + 0.5 a 0.25 2.0 a 0.25BP6 + + + + 0.5 a 0.25 2.0 a 0.25BP7 + − + + 0.25 a a 0.25 a aBP8 + + + + a a a a a aBP9 + − + + a a a a a aBP10 + − + + a a a a a aBP11 − + + + a a a 2.0 a aBP12 + + + + a 0.25 a 0.5 a aBP13 + + + + a 0.25 a 2.0 a 0.25BP14 − + + + a a a 0.5 a aBP15 − + + + a 0.5 0.25 2.0 a aBP16 − + + + a a a a a aBP17 − − + + a a a 0.25 a a
1Spore formation was tested by growing isolates for 1 week in a modified Schaeffer sporulation medium (Schaeffer et al. 1965) consisting of1.6 g of nutrient broth (Oxoid), 0.2 g of KCl, 0.5 g MgSO4·7H2O in 1L ddH2O with the following ingredients added to the cooled medium (per100 mL): 2 mL 1M Ca(NO3)2, 2 mL 0.1M MnCl2·4H2O, 2 mL FeSO4·7H2O, 2 mL 50% D-glucose monohydrate.
2We cannot rule out spore formation in other media for those that did not display spore formation in modified Schaeffer’s medium.3Growth response measured after 1 week. After 3 weeks isolates showed growth on pH 5.6 plates, suggesting delayed, but not arrested growth.4Metal resistance measured as Minimum Inhibitory Concentration (MIC) in mM. ‘a’ means a MIC of <0.125 mM.
(BP16) show extensive polysaccharide formation (Figures 3A,B), which show as stained strand-like material. Small membranevesicles were also visualised to have been produced by cells(Figure 3B). Cells of the Actinobacteria isolate (BP17) grewas filamentous colonies into the agar (Figure 3C, left of dottedline). On the surface of the agar plate the cell exhibited electrondense characteristics corresponding to spore structures (Figure3D, right of dotted line) consistent with a Streptomyces.
Colonisation of Basaltic Glass by Isolates fromBasalt/Palagonite Plates
Colonization of the basaltic glass blocks occurred with bothisolates obtained from unamended BP plates. The Bacteroidetescolonized the surface of the glass. Its growth was limited to theedge of the block where division of the non-motile coccoid cellsfrom the adjoining agar resulted in colonization. The organismformed a biofilm across areas of exposed glass (Figure 4A).Colonization of vesicles occurred at discrete points where cellsgrew over the edge of a vesicle (Figure 4B; arrow) with thesubsequent formation of a biofilm across the vesicle (Figure4C). The actinobacterial filaments were observed to grow acrossthe surface of the glass from the agar so that after 1 month of
incubation most of the surface of the blocks was covered in amat of filaments (Figure 4D). Filaments were observed to havepenetrated into vesicles within the glass (Figure 4E). Often thishad occurred by the formation of a filament network acrossvesicles with subsequent random downward growth of somefilament tips into the vesicles (Figure 4F).
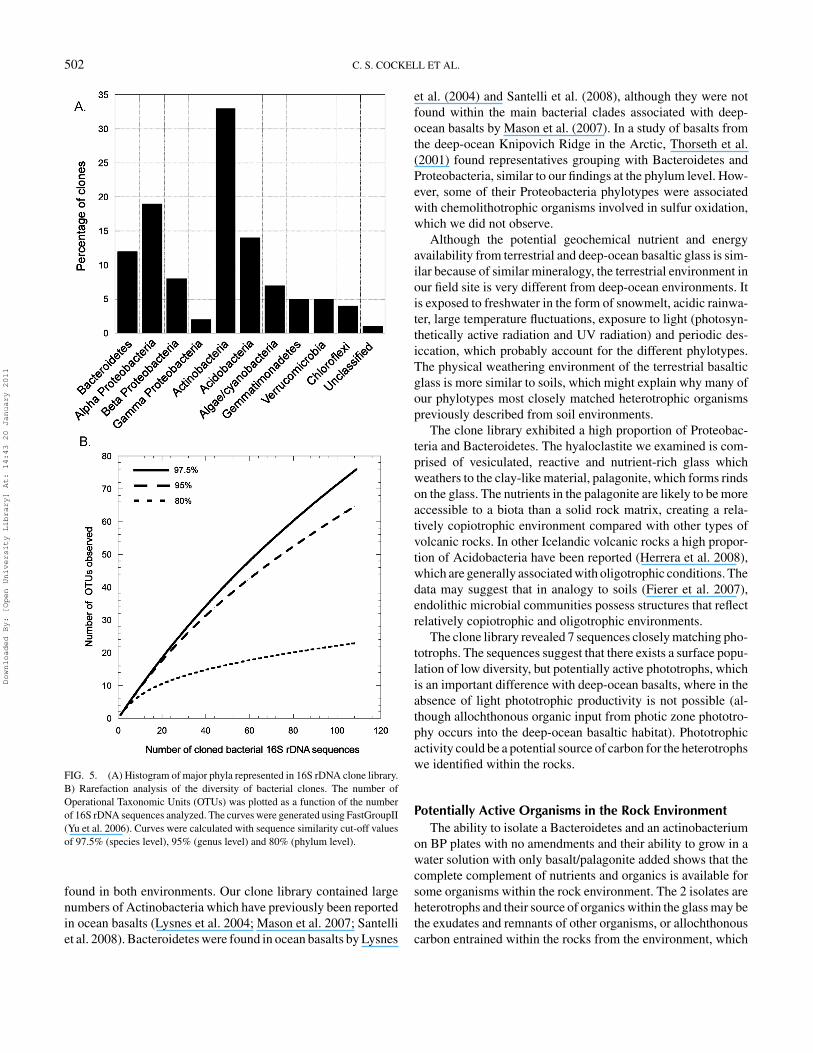
16S rDNA Analysis of Community Genomic DNAThe phylotypes of the bacterial 16S rDNA clone library are
shown in Figure 5A. One hundred and ten clones were examined.Rarefaction analysis showed a high diversity of microbial phylo-types associated with the rocks (Figure 5B). Seventy-six OTU’swere obtained at 97.5% similarity. None of the rocks in our fieldlocation displayed macroscopic signs of phototrophic growth(chlorophyll pigmentation), although 7 of our clones had a clos-est match to phototrophs (3 of them matching cyanobacteria andthe others corresponding to Stichococcus sp. and Klebsormid-ium spp.). Thirty-one of our isolates had a closest sequencematch to organisms previously described in cold soil and/or en-dolithic environments (12 from Antarctic soils [represented bythree Bacteroidetes, one Acidobacteria, 5 Actinobacteria, 1 α-and 1 β-proteobacteria, and 1 unclassified], 7 from Antarctic
Downloaded By: [Open University Library] At: 14:43 20 January 2011

500 C. S. COCKELL ET AL.
FIG. 3. Transmission Electron Micrographs (TEM) of the Bacteroidetes (BP16) and Actinobacteria (BP17) isolates obtained on unamended BP plates.(A) Sectioned colony of Bacteroidetes isolate (scale bar 500 nm), (B) Magnified sectioned Bacteroidetes cells (scale bar 100 nm), (C) Vertical section through agarplate of Actinobacteria isolate (surface of agar is white dotted line; scale bar 2 µm), (D) Magnified sectioned structures from the surface of the agar (scale bar 100 nm).
endoliths [all α-proteobacteria], 5 from glaciers and deglaciatedsoils [3 Bacteroidetes, 1 α- and 1 β-proteobacteria], 3 from theAtacama desert [all Actinobacteria] and 4 from other endolithicenvironments [2 actinobacteria and 1 α- and 1 β-proteobacteria).
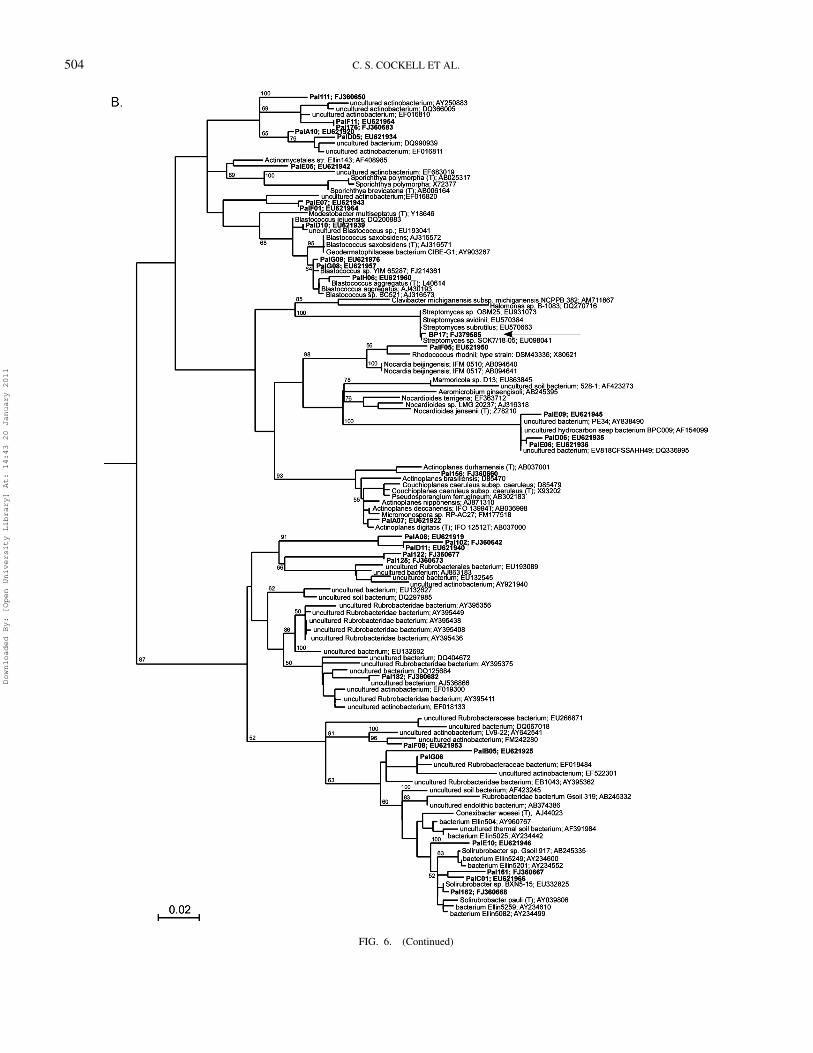
The phylogenetic trees for the Actinobacteria and Bac-teroidetes are shown in Figure 6. The phylogenetic affiliation ofthe 2 isolates obtained on unamended BP plates is also shown.
DISCUSSIONThe weathering of volcanic rocks plays an important role in
the drawdown of carbon dioxide from the atmosphere (Gail-lardet et al. 1999; Dessert et al. 2001) and thus the long-termcarbonate-silicate cycle. A crucial part of understanding thebiological involvement in this process is to identify and charac-terise potentially active bacteria. In this study, we investigatedsubglacial basaltic glass (hyaloclastite). We determined the di-versity of phylotypes within the rock and the characteristics ofculturable heterotrophs.
Bacterial Phylotypes in the Rock EnvironmentThe community within the material was dominated by phy-
lotypes belonging to Actinobacteria, Proteobacteria and Bac-
teroidetes. Actinobacteria and Proteobacteria have previouslybeen shown to be abundant in endolithic habitats (de la Torreet al. 2003; Walker and Pace 2007), and they are importantgroups in soils. Bacteroidetes (formally Cytophaga-Flexibacter-Bacteroidetes) have previously been associated with soil crustsand soils (Shivaji et al. 2004; Nagy et al. 2005; Gundlapallyet al. 2006), although they are also found in freshwater (Wu et al.2007b), marine (O’Sullivan et al. 2004; Murray and Grzymski2007) and microbial mat (Abed et al. 2007) environments andthey may be associated with the degradation of organic mate-rial, a contention supported by our observation of the growth ofthe Bacteroidetes isolate (BP16) on plates containing degradedcyanobacteria. The clone library shows the presence of Gem-matimonadetes, a recently described phylum (e.g., Zhang et al.2003), which is present in diverse environments, including arc-tic permafrost (Steven et al. 2007), cave soils (Zhou et al. 2007),water treatment plants (Qin et al. 2007), and the hyper-arid coreof the Atacama desert (Drees et al. 2006).
The data highlight the difference between terrestrial basalticglass and deep-ocean basaltic glass. None of our sequencesmatched the sequences in the ocean crust clades described byMason et al. (2007) or other previously reported ocean phylo-types. However, representatives of some of the major phyla are
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 501
FIG. 4. Scanning Electron Micrographs (SEM) in secondary electron mode showing colonization of basaltic glass blocks by isolates obtained on unamended BPplates. A-C) Bacteroidetes isolate, (A) Colonization of the surface of the rock (scale bar 5 µm), (B) Colonial growth from surface into vesicle (scale bar 10µm),followed by (C) Formation of biofilm within vesicle (scale bar 30 µm). D–F. Actinobacteria isolate. (D) Formation of network across basalt glass surface (scalebar 50 µm), (E) Invasion of vesicle by filaments (scale bar 25 µm), (F) Example of initial network formation over vesicle with some filaments moving into vesiclespace (scale bar 10 µm).
Downloaded By: [Open University Library] At: 14:43 20 January 2011
502 C. S. COCKELL ET AL.
FIG. 5. (A) Histogram of major phyla represented in 16S rDNA clone library.B) Rarefaction analysis of the diversity of bacterial clones. The number ofOperational Taxonomic Units (OTUs) was plotted as a function of the numberof 16S rDNA sequences analyzed. The curves were generated using FastGroupII(Yu et al. 2006). Curves were calculated with sequence similarity cut-off valuesof 97.5% (species level), 95% (genus level) and 80% (phylum level).
found in both environments. Our clone library contained largenumbers of Actinobacteria which have previously been reportedin ocean basalts (Lysnes et al. 2004; Mason et al. 2007; Santelliet al. 2008). Bacteroidetes were found in ocean basalts by Lysnes
et al. (2004) and Santelli et al. (2008), although they were notfound within the main bacterial clades associated with deep-ocean basalts by Mason et al. (2007). In a study of basalts fromthe deep-ocean Knipovich Ridge in the Arctic, Thorseth et al.(2001) found representatives grouping with Bacteroidetes andProteobacteria, similar to our findings at the phylum level. How-ever, some of their Proteobacteria phylotypes were associatedwith chemolithotrophic organisms involved in sulfur oxidation,which we did not observe.
Although the potential geochemical nutrient and energyavailability from terrestrial and deep-ocean basaltic glass is sim-ilar because of similar mineralogy, the terrestrial environment inour field site is very different from deep-ocean environments. Itis exposed to freshwater in the form of snowmelt, acidic rainwa-ter, large temperature fluctuations, exposure to light (photosyn-thetically active radiation and UV radiation) and periodic des-iccation, which probably account for the different phylotypes.The physical weathering environment of the terrestrial basalticglass is more similar to soils, which might explain why many ofour phylotypes most closely matched heterotrophic organismspreviously described from soil environments.
The clone library exhibited a high proportion of Proteobac-teria and Bacteroidetes. The hyaloclastite we examined is com-prised of vesiculated, reactive and nutrient-rich glass whichweathers to the clay-like material, palagonite, which forms rindson the glass. The nutrients in the palagonite are likely to be moreaccessible to a biota than a solid rock matrix, creating a rela-tively copiotrophic environment compared with other types ofvolcanic rocks. In other Icelandic volcanic rocks a high propor-tion of Acidobacteria have been reported (Herrera et al. 2008),which are generally associated with oligotrophic conditions. Thedata may suggest that in analogy to soils (Fierer et al. 2007),endolithic microbial communities possess structures that reflectrelatively copiotrophic and oligotrophic environments.
The clone library revealed 7 sequences closely matching pho-totrophs. The sequences suggest that there exists a surface popu-lation of low diversity, but potentially active phototrophs, whichis an important difference with deep-ocean basalts, where in theabsence of light phototrophic productivity is not possible (al-though allochthonous organic input from photic zone phototro-phy occurs into the deep-ocean basaltic habitat). Phototrophicactivity could be a potential source of carbon for the heterotrophswe identified within the rocks.
Potentially Active Organisms in the Rock EnvironmentThe ability to isolate a Bacteroidetes and an actinobacterium
on BP plates with no amendments and their ability to grow in awater solution with only basalt/palagonite added shows that thecomplete complement of nutrients and organics is available forsome organisms within the rock environment. The 2 isolates areheterotrophs and their source of organics within the glass may bethe exudates and remnants of other organisms, or allochthonouscarbon entrained within the rocks from the environment, which
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 503
FIG. 6. 16S rDNA phylogenetic trees of (A) Bacteroidetes and (B) Actinobacteria showing positions of 2 isolates obtained on unamended BP plates (arrows)and clones obtained from culture independent methods (labelled ‘Pal’ in bold). A maximum likelihood tree was generated using a neighbour-joining tree (1000bootstraps) as the input tree and using a mask of 472 nucleotide positions (E.coli numbering 718 to 1190 covering V5 and V6). Aquifex sp. (accession numberAB304892) was used as an outgroup. The scale bar represents the number of changes per nucleotide position. Bootstrap values of 50% and greater are shown.(Continued)
Downloaded By: [Open University Library] At: 14:43 20 January 2011
504 C. S. COCKELL ET AL.
FIG. 6. (Continued)
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 505
could potentially come from rain or snow melt. The organismspossessed tolerance to desiccation and freeze-thaw and had onlyslight halotolerance, consistent with the environment within Ice-landic volcanic rocks.
After 4 months of incubation we were unable to isolate otherorganisms. This result suggests either that a very small percent-age of species are active within the rocks or that the metabolicactivity of many of the organisms is low so that they cannotbe cultured on basalt/palagonite material in the time scale ofseveral months. Both explanations are likely to be important.Microorganisms will be entrained within the rocks by numer-ous processes including snowmelt, rainfall and aeolian transport.Many of these organisms may not have the metabolic capabilityto grow within the rock, but would be picked up by culture-independent methods. Despite the relatively nutrient-rich en-vironment in the basaltic glass and palagonite compared withother volcanic rocks, as discussed above, the rates of weatheringof basaltic glass are still low. A 1-mm radius sphere of basalticglass is estimated to have a lifetime of ∼500 years at pH 4.0and a temperature of 25◦C (Wolff-Boenisch et al. 2004). Therelease of nutrients over these time scales, even in palagonite,would suggest that active organisms will grow slowly.
In this study we isolated Firmicutes species on carbon-amended BP plates. Fajardo-Cavazos and Nicholson (2006) iso-lated Bacillus spp. from granite and showed that some isolateswere related, suggesting that the cells might be reproducing in-side the rocks. This is also consistent with the observation thatsome of our isolates are similar or may be identical (Table 1).We did not isolate Firmicutes on unamended BP plates, onlyon plates augmented with organics. However, after transfer ontounamended BP plates the isolates obtained on amended platesdid grow very slowly, suggesting that they can be active in theenvironment, which would be consistent with the data obtainedby Fajardo-Cavazos and Nicholson (2006). We did not detectFirmicutes in our clone library. Although one possible explana-tion is that many of the Firmicutes are present as spores with asmall proportion as vegetative cells, and they were not picked upby our DNA extraction method, preliminary data using an envi-ronmental Phylochip (unpublished data) show they are present.The lack of representation in our clone library suggests incom-plete sampling of the microbial population, consistent with therarefaction curve.
Adaptation to the Rock MicroenvironmentDuring the weathering of rock, protons are consumed, in-
creasing the pH in the local micro-environment (Bland andRolls 2005). If the rock is open to the atmosphere, the pHstabilises to around neutral conditions, but can become alkalineif the interior of the rock is essentially closed to the atmosphere(CO2-diffusion limited). The pH of Icelandic rain water is acidicand has been measured previously as 5.1–5.6 (Gislason et al.1987). The physiological data we present show that the growthof many organisms is retarded at the pH of Icelandic rain watercompared with neutral and alkaline pH, raising two important
possibilities: (1) pH changes caused by rock weathering mightchange the community of active organisms, (2) the activity oforganisms themselves, through their metabolic activity, mightchange the rate of weathering and thus the rate of pH change,acting as a feedback to determine which organisms are metabol-ically active.
In hyaloclastite, pH changes accompany palagonite forma-tion (Thorseth et al. 1991). In some palagonite formation mech-anisms pH may drop to as low as 3, forming ‘white palagonite’.Brown palagonite, such as the material that occurs extensivelyin our rocks, and is accompanied by the deposition of Fe, Tiand Al in the pore spaces, suggests weathering phases requiringhigh pH (> 9) (Thorseth et al. 1991). Thus, the activity of themembers of the microbial community is likely to be sensitiveto the specific phases and types of palagonite formed in basalticglass environments, which will vary temporally and spatially onmicron scales.
Volcanic rocks contain numerous biologically detrimen-tal transition metals at low concentrations, which organismsmust be able to tolerate if they are to be active in the rockweathering environment. They include Cr, Ni, Cu, V and Co(Carmichael 1964; Walsh and Clarke 1982). The transitionmetal-amended agar experiment showed that the highest con-centrations recorded for these elements prevented the growth ofour isolates if they are all present simultaneously. The exper-iment suffers from some simplifications. In the rock, many ofthese elements are strongly bound into the silica matrix. In agarthey will enter more readily into solution and thus come intocontact with the organisms at higher concentrations. This prob-ably explains the inability of the isolates to tolerate the metalswhen they were all combined.
The Minimum Inhibitory Concentration (MIC) of the iso-lates showed that some of them have heavy metal resistance,suggesting either that: (1) it may have been genetically acquired(mutation or gene transfer) by the organism elsewhere, or (2)it may have been acquired by mutation or gene transfer in theparticular habitat under study. The MIC of our isolates is stillat least ten times lower than that recorded for some microor-ganisms isolated from metal-contaminated industrial soils (e.g.,Bopp et al. 1983; Malik and Jaiwal 2000). However, the datashow that the interior of the rocks harbours metal-resistancegenes.
Both of the isolates obtained on unamended BP plates ex-hibited antibiotic resistance. Antibiotic resistance is known insoil microorganisms (Nwosu 2001) and they are produced in aparticularly large variety by the Actinobacteria (Riesenfeld et al.2004). The abundance of Actinobacteria within the rocks andthe presence of antibiotic resistance in the isolates obtained onunamended BP plates suggest that endolithic environments maybe an important source of novel antibiotic resistance genes.
Rock Colonization by ActinobacteriaThe prevalence of Actinobacteria in our clone library,
their representation among the isolates cultured on BP plates
Downloaded By: [Open University Library] At: 14:43 20 January 2011

506 C. S. COCKELL ET AL.
amended with carbon and our ability to culture an isolate onbasalt/palagonite agar that was abundant in glass samples maybe explained by their physiological tolerances, such as the re-sistance to desiccation we observed. However, we also hypoth-esized that a contributing factor may be the filamentous growthhabit of some Actinobacteria, which allows them to effectivelycolonise the vesiculated, porous material. We tested this hy-pothesis with a simple colonization experiment using polishedblocks of basaltic glass. The experiment showed that non-motilecoccoid organisms, such as the Bacteroidetes we isolated, aregenerally limited to moving through the rock by cell division onthe rock surface. In the natural environment they might also betransported through the rock by water flow. In contrast, filamentsof the actinobacterial isolate were capable of directionally grow-ing across the rock surface and branching into the vesicular porespaces. This process is analogous to the invasion of rocks by fun-gal hyphae (Burford et al. 2003). This would give filamentousActinobacteria an ability to systematically colonize permeablerocks by moving in many directions and might partly explainthe abundance of the actinobacterial isolate we obtained com-pared to the Bacteroidetes. A high abundance of Actinobacteriahas similarly been reported in the volcanic pyroclastic depositsformed during the eruption of Mount St. Helens, USA in 1980(Ibekwe et al. 2007). In addition to Acidobacteria, Actinobac-teria were the only phylum found in Hawaiian lava flows of allages studied by Gomez-Alvarez et al. (2007). These observa-tions show that Actinobacteria can play an important role in thecolonization of vesiculated terrestrial volcanic rocks and ash,and potentially their subsequent weathering.
REFERENCESAbed RMM, Zein B, Al-Thukair A, de Beer D. 2007. Phylogenetic diversity
and activity of aerobic heterotrophic bacteria from a hypersaline oil-pollutedmicrobial mat. Syst Appl Microbiol 30:319–330.
Adamo P, Marchetiello A, Violante P. 1993. The weathering of mafic rocks bylichens. Lichenologist 25:285–297.
Billi D, Grilli Caiola M. 1996. Effects of nitrogen and phosphorus deprivationon Chroococcidiopsis sp. (Chroococcales). Algal Stud 83:93–105.
Bland W, Rolls D. 2005. Weathering: An Introduction to the Scientific Princi-ples. Arnold, London.
Bopp HL, Chakrabarty AM, Ehrlich HL. 1983. Chromate resistance plasmid inPseudomonas fluorescens. J Bacteriol 155:1105–1109.
Bruce KD, Hiorns WD, Hobman JL, Osborn AM, Strike P, Ritchie DA. 1992.Amplification of DNA from native populations of soil bacteria by using thepolymerase chain reaction. Appl Environ Microbiol 58:3413–3416.
Brunauer S, Emmett PH, Teller E. 1938. Adsorption of gases in multimolecularlayers. J Am Chem Soc 60:309–319.
Burford EP, Kierans M, Gadd GM. 2003. Geomycology: Fungi in mineralsubstrata. Mycologist 17:98–107.
Carmichael ISE. 1964. The petrology of Thingmuli, a Tertiary volcano in easternIceland. J Petrol 5:425–460.
Chakravorty S, Helb D, Burday H, Connell N, Alland D. 2007. A detailedanalysis of 16S ribosomal RNA gene segments for the diagnosis of pathogenicbacteria. J Microbiol Meth 69:330–339.
Cockell CS, Schuerger AC, Billi D, Friedmann EI, Panitz C. 2005. Effects ofa simulated Martian UV flux on the cyanobacterium, Chroococcidiopsis sp029. Astrobiology 5:127–140.
Cockell CS, Olsson K, Herrera A, Meunier A. 2009. Alteration textures interrestrial volcanic glass and the associated bacterial community. Geobiology7:50–65.
Cole JR, Chai B, Marsh TL, Farris RJ, Wang Q, Kulam SA, Chandra S, Mc-Garrell DM, Schmidt TM, Garrity GM, Tiedje JM. 2003. The RibosomalDatabase Project (RDP-II): previewing a new autoaligner that allows regu-lar updates and the new prokaryotic taxonomy. Nucleic Acids Res 31:442–443.
de la Torre JR, Goebel BM, Friedmann EI, Pace NR. 2003. Microbial diversityof cryptoendolithic communities from the McMurdo Dry Valleys, Antarctica.Appl Environ Microbiol 69:3858–3867.
Dessert C, Dupre B, Francois LM. 2001. Erosion of Deccan Traps determinedby river geochemistry: impact on the global climate and the 87Sr/86Sr ratioof seawater. Earth Planet Sci Lett 188:459–474.
Dessert C, Dupre B, Gaillardet J, Francois LM, Allegre CJ. 2003. Basalt weath-ering laws and the impact of basalt weathering on the global carbon cycle.Chem Geol 202:257–273.
Drees KP, Neilson JW, Betancourt JL, Quade J, Henderson DA, Pryor BM,Maier RM. 2006. Bacterial community structure in the hyperarid core of theAtacama Desert, Chile. Appl Environ Microbiol 72:7902–7908.
Etienne S, Dupont J. 2002. Fungal weathering of basaltic rocks in a cold oceanicenvironment (Iceland): comparison between experimental and field observa-tions. Earth Surf Proc Land 27:737–748.
Fajardo-Cavazos P, Nicholson W. 2006. Bacillus endospores isolated from gran-ite: close molecular relationships to globally distributed Bacillus spp. fromendolithic and extreme environments. Appl Environ Microbiol 72:2856–2863.
Fierer N, Bradford MA, Jackson RB. 2007. Toward an ecological classificationof soil bacteria. Ecology 88:1354–1364.
Fisk MR, Giovannoni SJ, Thorseth IH. 1998. Alteration of oceanic volcanicglass: textural evidence of microbial activity. Science 281:978–980.
Friedmann EI, Ocampo-Friedmann R. 1985. Blue-green algae in arid cryptoen-dolithic habitats. Archiv fur Hydrobiologie 38/39:349–350.
Furnes H, Staudigel H. 1999. Biological mediation in ocean crust alteration:How deep is the deep biosphere? Earth Planet Sci Lett 166:97–103.
Gaillardet J, Dupre P, Louvat CJ, Allegre CJ. 1999. Global silicate weatheringand CO2 consumption rates deduced from the chemistry of the large rivers.Chem Geol 159:3–30.
Gislason SR, Eugster HP. 1987. Meteoric water-basalt interactions. II: A fieldstudy in N.E. Iceland. Geochim Cosmochim Acta 51:2841–2855.
Gomez-Alvarez V, King GM, Nusslein K. 2006. Comparative bacterial in recentHawaiian volcanic deposits of different ages. FEMS Microbiol Ecol 60:60–73.
Gundlapally SR, Garcia-Pichel F. 2006. The community and phylogenetic di-versity of biological soil crusts in the Colorado Plateau studied by molecularfingerprinting and intensive cultivation. Microb Ecol 52:345–357.
Herrera A, Cockell CS. 2007. Exploring microbial diversity in volcanic environ-ments: A review of methods in DNA extraction. J Microbiol Meth 70:1–12.
Herrera A, Cockell CS, Self S, Blaxter M, Reitner J, Arp G, Drose W, Thorsteins-son T, Tindle AG. 2008. Bacterial colonization and weathering of terrestrialobsidian in Iceland. Geomicrobiol J 25:25–37.
Herrera A, Cockell CS, Self S, Blaxter M, Reitner J, Thorsteinsson T, Arp G,Drose W, Tindle AG. 2009. A cryptoendolithic community in volcanic glass.Astrobiology 9:369–381.
Ibekwe AM, Kennedy AC, Halvorson JJ, Yang CH. 2007. Characterization ofdeveloping microbial communities in Mount St. Helens pyroclastic substrate.Soil Biol Biochem 39:2496–2507.
Jackson TA, Keller WD. 1970. A comparative study of the role of lichens and“inorganic” processes in the chemical weathering of recent Hawaiian lavaflows. Am J Sci 269:446–466.
Jacobsson SP, Gudmundsson MT. 2008. Subglacial and intraglacial volcanicformations in Iceland. Jokull 58:179–196.
Kimura M. 1980. A simple method for estimating evolutionary rates of basesubstitutions through comparative studies of nucleotide sequences. J MolEvol 16:111–120.
Downloaded By: [Open University Library] At: 14:43 20 January 2011

BACTERIA IN BASALTIC GLASS 507
Kisakurek B, Widdowson M, James RH. 2004. Behaviour of Li isotopes duringcontinental weathering: the Bidar laterite profile, India. Chem Geol 212:27–44.
Louvat P, Allegre CJ. 1998. Riverine erosion rates on Sao Migual volcanicisland, Azores archipelago. Chem Geol 148:177–200.
Lysnes K, Thorseth IH, Steinbu BO, Øvreas L, Torsvik T, Pedersen RB. 2004.Microbial community diversity in seafloor basalt from the Arctic spreadingridges. FEMS Microb Ecol 50:213–230.
Malik A, Jaiswal R. 2000. Metal resistance in Pseudomonas strains isolated fromsoil treated with industrial wastewater. World J Microbiol Biotech 16:177–182.
Mason OU, Stingl U, Wilhelm LJ, Moeseneder MM, Di Meo-Savoie CA, FiskMR, Giovannoni SJ. 2007. The phylogeny of endolithic microbes associatedwith marine basalts. Environ Microbiol 9:2539–2550.
Mergeay M, Nies S, Schlegel HG, Gerits J, Charles P, Van Gijsegem F. 1985.Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid-bound resistance to heavy metals. J Bacteriol 162:328–334.
Murray AE, Grzymski JJ. 2007. Diversity and genomics of Antarctic marinemicro-organisms. Phil Trans Royal Soc 362:2259–2271.
Nagy ML, Perez A, Garcia-Pichel F. 2005. The prokaryotic diversity of bio-logical crusts in the Sonoran desert (Organ Pipe Cactus National Monument,AZ). FEMS Microb Ecol 54:233–245.
Nwosu VC. 2001. Antibiotic resistance with particular reference to soil mi-croorganisms. Res Microbiol 152:421–430.
O’Sullivan LA, Fuller KE, Thomas EM, Turley CM, Fry JC, Weightman AJ.2004. Distribution and culturability of the uncultivated ‘AGG58 cluster’ of theBacteroidetes phylum in aquatic environments. FEMS Microb Ecol 47:359–370.
Perriere G, Gouy M. 1996. WWW-query: an on-line retrieval system for bio-logical sequence banks. Biochimie 78:364–369.
Qin YY, Liu DT, Yang H. 2007. Investigation of total bacterial and ammonia-oxidizing bacterial community composition in a full-scale aerated submergedbiofilm reactor for drinking water pretreatment in China. FEMS MicrobiolLett 268:126–134.
Riesenfeld CS, Goodman RM, Handelsman J. 2004. Uncultured soil bacteria area reservoir of new antibiotic resistance genes. Environ Microbiol 6:981–989.
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY. 1979. Genericassignments, strain histories and properties of pure cultures of cyanobacteria.J Gen Microbiol 111:1–61.
Ryan FJ. 1950. Selected methods of Neurospora genetics. Meth Med Res 3:51–75.
Saitou N, Nei M. 1987. The neighbor-joining method: a new method for recon-structing phylogenetic trees. Mol Biol Evol 4:406–425.
Santelli CM, Orcutt BN, Banning E, Bach W, Moyer CL, Sogin ML, StaudigelH, Edwards KJ. 2008. Abundance and diversity of microbial life in oceancrust. Nature 452:653–657.
Schaeffer P, Millet J, Aubert JP. 1965. Catabolic repression of bacterial sporu-lation. Proc Natl Acad Sci USA 54:704–711.
Schwieger F, Tebbe CC. 1998. A new approach to utilize PCR-single-strand-conformation polymorphism for 16S rRNA gene-based microbial communityanalysis. Appl Environ Microbiol 64:4870–4876.
Shivaji S, Reddy GSN, Aduri RP, Kutty R, Ravenschlag K. 2004. Bacterialdiversity of a soil sample from Schirmacher Oasis, Antarctica. Cell Mol Biol50:525–536.
Sinton J, Gronvold K, Saemundsson K. 2005. Postglacial eruptive history ofthe Western Volcanic Zone, Iceland. Geochem Geophys Geosys 6: Art. No.Q12009.
Steven B, Briggs G, McKay CP, Pollard WH, Greer CW, Whyte LG. 2007. Char-acterization of the microbial diversity in a permafrost sample from the Cana-dian High Arctic using culture-dependent and culture-independent methods.FEMS Microbiol Ecol 59:513–523.
Stroncik NA, Schminke H. 2002. Palagonite – A review. Intern J Earth Sci91:680–697.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. TheCLUSTAL X windows interface: flexible strategies for multiple sequencealignment aided by quality analysis tools. Nucl Acids Res 25:4876–4882.
Thorseth IH, Furnes H, Tumyr O. 1991. A textural and chemical study ofIcelandic palagonite of varied composition and its bearing on the mechanismof the glass-palagonite transformation. Geochim Cosmochim Acta 55:731–749.
Thorseth IH, Furnes H, Heldal M. 1992. The importance of microbiologicalactivity in the alteration of natural basaltic glass. Geochim Cosmochim Acta56:845–850.
Thorseth IH, Torsvik T, Torsvik V, Daae FL, Pedersen RB. 2001. Diversity oflife in ocean floor basalt. Earth Planet Sci Lett 194:31–37.
Thorseth IH, Pedersen RB, Christie DM. 2003. Microbial alteration of 0-30-Ma seafloor and sub-seafloor basaltic glasses from the Australian AntarcticDiscordance. Earth Planet Sci Lett 215:237–247.
Torsvik T, Furnes H, Muehlenbachs K, Thorseth IH, Tumyr O. 1998. Evidencefor microbial activity at the glass-alteration interface in oceanic basalts. EarthPlanet Sci Lett 162:165–176.
Walker JJ, Pace NR 2007. Phylogenetic composition of rocky mountain en-dolithic microbial ecosystems. Appl Environ Microbiol 73:3497–3504.
Walsh JN, Clarke E. 1982. The role of fractional crystallisation in the formationof granitic and intermediate rocks of the Beinn Chaisgidle Centre, Mull,Scotland. Mineral Mag 45:247–255.
Weirich G, Schweisfurth R. 1985. Extraction and culture of microorganismsfrom rock. Geomicrobiol J 4:1–20.
Wolff-Boenisch D, Gıslason SR, Oelkers EH, Putnis CV. 2004. The dissolutionrates of natural glasses as a function of their composition at pH 4 and 10,and temperatures from 25 to 74◦C. Geochim Cosmochim Acta 68:4843–4858.
Wu L, Jacobson AD, Chen H, Hausner M. 2007a. Characterisation of elementalrelease during microbe-basalt interactions at T=28◦C. Geochim CosmochimActa 71:2224–2239.
Wu X, Xi WY, Ye WJ, Yang H. 2007b. Bacterial community composition of ashallow hypertrophic freshwater lake in China. FEMS Microb Ecol 61:85–96.
Yu Y, Breitbart M, McNairnie, Rohwer F. 2006. FastGroupII: A web-basedbioinformatics platform for analyses of large 16S rDNA libraries. BMCBioinformatics 7:57.
Zhang H, Sekiguchi Y, Hanada S, Hugenholtz P, Kim H, Kamagata Y, NakamuraK. 2003. Gemmatimonas aurantiaca gen. nov., sp nov., a Gram-negative,aerobic, polyphosphate-accumulating micro-organism, the first cultured rep-resentative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int JSyst Evol Microbiol 53:1155–1163.
Zhou JP, Gu YQ, Zou CS, Mo MH. 2007. Phylogenetic diversity of bacteria inan earth-cave in Guizhou Province, Southwest of China. J Microbiol 45:105–112.
Downloaded By: [Open University Library] At: 14:43 20 January 2011




















