
Differential Logging: A Commutative and Associative Logging ...

Experimental and Clinical Psvchopliarniat-ology1998, Vol. 6, No. 3, 248-254"
Copyright 1998 by the Am
Associative Tolerance to Nicotine Analgesia in the Rat:Tail-Flick and Hot-Plate Tests
Antonio Cepeda-Benito, Jose Reynoso, and Eric H. McDanielTexas A&M University
Previous assessments of associative nicotine tolerance may have confounded associativeeffects with novelty-induced stress effects, instrumental learning effects, or both. That is,subjects were tested in novel environments, allowed to practice the test response, or bothduring the tolerance development phase. In the first study, 32 male Sprague-Dawley rats wereinjected with various doses of nicotine and tested for nociception in the tail-flick and hot-platetests to assess nicotine's analgesic effects. In the second study, 35 rats received nicotineexplicitly paired or unpaired with a distinctive test context. All animals were equallypreexposed to the test environment, and none had the opportunity to practice the test response.Paired rats developed greater nicotine tolerance than unpaired rats. This context-dependent(associative) tolerance effect was found with both tail-flick and hot-plate tests.
Drug tolerance is a decrease in the effects of a drug dose
following repeated drug administrations. Drug tolerance is
central to the definition of drug dependence (American
Psychiatric Association, 1994), and several theorists assign
tolerance, or the mechanisms that produce tolerance, an
important role in the genesis and maintenance of addictive
behaviors (see Ramsay & Woods, 1997; Trujillo & Akil.
1995). For example, smokers may smoke more to compen-
sate for the development of tolerance of the reinforcing
effects of nicotine. Likewise, tolerance of the ill effects of
nicotine may increase tobacco consumption by allowing
smokers to smoke more without feeling sick (Alexander &
Hadaway, 1982; Pomerleau, 1995; Trujillo & Akil). Thus,
nicotine tolerance can be a symptom of physical dependence
on nicotine (American Psychiatric Association, 1994) and a
contributor to the severity of the smoker's physical and
psychological dependence (Pomerleau).
There is considerable evidence that many examples of
drug tolerance represent the operation of classical condition-
ing (Young & Goudie, 1994), and some theorists have
proposed that "learning is the primary underlying cause"
(Ramsay & Woods, 1997, p. 170) of all drug tolerance
phenomena. Most of the support for the role of classical
conditioning in the development of drug tolerance comes
from investigations of tolerance to the analgesic effects of
morphine. In associative tolerance studies, animals receiv-
ing morphine explicitly paired with a distinctive test context
Antonio Cepeda-Benito, Jose Reynoso, and Eric H. McDaniel,Department of Psychology, Texas A&M University.
This research was supported by Grant DA10576-01 from theNational Institute on Drug Abuse. We thank Anissa Norrell,Thomas Cross, Andrea Szabo, Michael Drake, Eddy Badrina, PaigeBonner, Amber Shipp, Enrique Pizana, Sarah Manning, and JasonMurphy for help in conducting Ihe experiment.
Correspondence concerning this article should be addressed toAntonio Cepeda-Benito, Department of Psychology, Texas A&MUniversity, College Station, Texas 77843-4235. Electronic mailmay be sent to [email protected].
are more tolerant to morphine's analgesic effects than
animals receiving the same drug exposure regimen in an
environment other than the distinctive context (e.g., Cepeda-
Benito & Tiffany, 1996a). The conditioning explanation for
these findings is that the distinctive context has become a
conditioned stimulus (CS) that elicits associative tolerance
effects (Baker & Tiffany, 1985; Poulos & Cappell, 1991;
Siegel, 1975). Some authors have also proposed that the
interoceptive effects of drugs may also function as CSs
capable of producing associative tolerance effects (e.g.,
Cepeda-Benito & Short, 1997; Grisel, Wiertelak, Watkins, &
Maier, 1994).
Despite the prominence of learning accounts of drug
tolerance, we know of only five studies that investigated the
impact of cue associations on the development of nicotine
tolerance (Caggiula et al, 1991, 1993; Caggiula, Epstein, &
Stiller, 1989; Epstein, Caggiula, Perkins, McKenzie, &
Smith, 1991; Epstein, Caggiula, & Stiller, 1989). These
studies investigated tolerance to nicotine's (a) anorectic
effects (Caggiula et al., 1989, 1991), (b) corticosterone-
elevating effects (Caggiula et al., 1991), (c) tachycardiac
effects (Epstein et al., 1991), and (d) analgesic effects(Caggiula et al., 1993; Epstein et al., 1989). These research-
ers found evidence of nicotine tolerance in subjects tested in
the same context in which they had received repeated
administrations of nicotine. However, these subjects failed
to display tolerance effects when they were tested in the
presence of a novel set of contextual stimuli (challenge test).
Thus, the researchers concluded that these subjects had
developed a context-dependent, or associative, form of
nicotine tolerance.
Alternatively, when animals are tested in a novel environ-
ment, an apparent loss of tolerance also may be accounted
for by the presence of novelty-induced stress in the test
situation. That is, environmental stress may interact with a
drug dose to augment the drug effects being measured. For
example, exposure to a novel environment can elevate
corticosterone levels (e.g., Caggiula et al., 1993) or induce
analgesia (e.g., Netto, Siegfried, & Izquierdo, 1987; Sher-
248

TOLERANCE TO NICOTINE ANALGESIA IN THE RAT 249
man, 1979). Various investigations have demonstrated thatsome examples of context-specific drug tolerance mayreflect the presence of stress effects rather than the presenceof absence of associative tolerance effects (e.g., Bardo &Hughes, 1979; Sherman, 1979; Sherman, Proctor, & Strub,1982).
In the two studies that investigated the development oftolerance to the analgesic effects of nicotine (Caggiula et al.,1993; Epstein et al., 1989), the subjects were allowed topractice the test response during the tolerance developmentphase. This method could be problematic because animalspermitted to practice a task while drugged can develop moretolerance to the disruptive effects of the drug on perfor-mance than animals given as much drug but without theopportunity for drugged practice (sec reviews by Goudie &Demellweek, 1986; Wolgin, 1989; Young & Goudie, 1994).Some investigators have argued that this intoxicated-practice effect represents the operation of instrumentalconditioning resulting in the learning of specific compensa-tions for drug-induced disruptions in performance, that is,behavioral contingent tolerance (e.g., LeBlanc, Gibbins, &Kalant, 1973). In all fairness, some theorists do not concep-tually differentiate between instrumental and associativeforms of drug tolerance (see Poulos & Cappell, 1991).
Our main objective was to obtain contextual nicotinetolerance effects by using procedures that minimize theinfluence of novelty and instrumental confounding factorsduring testing. Given that nicotine has analgesic properties(e.g., Rogers & Iwamoto, 1993), we modeled our proceduresafter those of studies that have produced robust associativetolerance to morphine's analgesic effects (e.g., Carter &Tiffany, 1996; Cepeda-Benito & Tiffany, 1992, 1996a,1996b). In these associative tolerance studies, the influenceof novelty-induced stress was controlled or minimijed byextensively prchabituating the animals to the experimentaland tolerance assessment procedures (sec also Ramsay &Woods, 1997). These associative tolerance investigationsalso showed that intoxicated practice is not necessary for theproduction of robust associative tolerance (e.g., Cepeda-Benito & Tiffany, 1992).
Study 1
We designed Study 1 to verify that we could assessnicotine's antinociceptive effects in a dose-dependent man-ner with tail-flick and hot-plate devices. Although it isgenerally accepted that nicotine reliably produces analgesiceffects in rats, in some cases the antinociceptive effects ofnicotine in the tail-flick test have not been observed (e.g.,Yang, Wu, & Zbuzek, 1992). Unlike previous dose-relatedanalyses (Caggiula, Epstein, Perkins, & Saylor, 1995; Rog-ers & Iwamoto, 1993), our analyses attempted to minimizestress factors during the test session by extensively prehabitu-ating the rats to the experimental procedures and the testenvironment.
Epstein et al. (1989) demonstrated the presence of toler-ance of nicotine's analgesic effects by snowing that withrepealed administrations, nicotine lost its ability to increasetail-withdrawal latencies. In order to establish the specificity
or generality of this reported nicotine tolerance effect intail-flick reflexes, we measured responses mediated bothspinally (tail flick) and supraspinally (hot plate).
Method
Subjects. The subjects, 32 experimentally naive, male Sprague-
Dawley rats, weighed between 300 and 350 g at the beginning of
the experiment. They were housed individually in plastic cages
with a bed of wood shavings. The colony was constantly illumi-
nated, and the rats had continuous access to food and water in their
home cages.
Drugs. Nicotine bitartrate was dissolved in physiological sa-
line to produce the following nicotine concentrations: 0.125, 0.25,0.375, 0.50, 0.60, and 0.75, mg/ml. All solutions were subcutane-
ously injected in the scruff of the neck (1 ml/kg of body weight).Tail-jllck analgesia assessment. The tail-nick test measures the
latency for the rat to move its tail away from a hot beam of light
(e.g., Cepeda-Benito & Tiffany, 1996a). A rat was restrained in an
opaque cylinder (6.8 x 22 cm) that had a Plexiglas base (5.5 X 22
cm). Ventilation holes were made on top of and in front of the tube.
The rat's tail protruded from the back of the tube and was placed in
a grooved plate such that the tail was directly under the light source
(HTC tail flick, model 33B). When the rat moved its tail away from
the light beam, a photosensitive cell tripped a timer and the
tail-flick latency was automatically recorded. To avoid interactions
between tail area stimulated and degree of analgesia (Yoburn,
Morales, Kelly, & Inturrisi, 1984), each assessment was the mean
of three consecutive trials with the location of the beam beingvaried among the proximal, middle, and distal thirds of the rat's
tail. The beam intensity was adjusted such that undrugged animals
nicked their tails at about 4 s. A 15-s limit was used to prevent
damage to the tail. The tester was blind to the rat's group condition.Hot-plate analgesia assessment. The hot-plate test measures a
rat's latency to lick a paw or jump (e.g., Caggiula el al., 1995;
Cepeda-Benito & Tiffany, 1996b; Krank, 1987). A rat was confined
to the hot-plate's surface in a chamber (30 X 30 X30 cm) with a
clear Plexiglass lid. The hot plate consisted of a metal surface
thermostatically controlled to a constant temperature of 50° C
(IITT hot plate, model 35D). Two independent observers, blind tothe rat's group condition, timed to the nearest hundredth of a
second each rat's latency to either lick a paw or jump, whichever
came first (e.g., Krank). The response latency was the mean of the
two observations. The median difference between observers was 1.47 s.
Rats were removed from the hot plate as soon as they either licked a paw
or jumped. Animals that neither licked a paw nor jumped after 60 s were
removed from the apparatus to prevent tissue damage.
Tail-flick prehahituation and testing phase. After 1 week of
accommodation to their new home environment, the rats were
weighed once daily for 3 days and weighed twice daily for an
additional 3 days. Rats were then habituated to injection and
mock-testing procedures. Rats received five saline injections paired
with a distinctive context and five saline injections paired with their
home cage environment. The interval between distinctive context
injections was 48 hr. Each home cage injection was given 24 hr
after each distinctive context exposure. For distinctive context
exposures, each rat was weighed, individually carried in a small
plastic container to the distinctive context room, injected with
saline, placed inside a tube, placed in a dark cabinet, and mocktested in the tail-flick device 4, 8, and 13 min after the injection. In
the distinctive context room, the room's light was dimmed,
apple-cinnamon air fresheners scented the holding cabinet, and
white noise was continuously played. Rats were not removed from
their holding tubes. Each mock tail-flick test consisted of placing
250 CEPEDA BENITO, REYNOSO, AND McDANIEL
the rat in the tail-flick apparatus and going through the motions ofconducting three tail-flick tests (i.e., without aiming the lighl beamat the tail). After each mock test, the rats were returned to thescented cabinets. Rats were returned to their home cage environ-ments 33 min after the last mock lesl. For home cage injections, therats were individually weighed and placed back in their homecages. Each rat was then removed from its home cage, injected withsaline, and returned to its cage.
Rats were tested in the distinctive context 48 hr after the fifthdistinctive context exposure session (or 24 hr after the last homecage injection). Rats were randomly divided into five groups, withone group receiving saline and each remaining group receiving adifferent test dose of nicotine (0.25, 0.5, 0.6, and 0.75 mg/kg). Ratswere tested in actual tail-flick trials 4, 8, and 13 min after beinginjected with nicotine.
Hot-plale prehabituation and testing phase. Rats began fiveadditional distinctive context exposure sessions 48 hr after theywere tested with the tail-flick device. Except for mock tests, theseexposure sessions were identical to the tail-flick exposure sessions.For each mock test, the rat was removed from its tube and placed ona nonfunctional hot plate for 60 s. The rat was then placed backinside its tube and returned to the scented cabinet. Rats receivedsaline injections in their home cage environment 24 hr after eachdistinctive context exposure.
Rats were tested in the distinctive context 48 hr after the lastdistinctive context exposure session (or 24 hr after the last homecage injection). Rats were randomly divided into five groups, withone group receiving saline and each remaining group receiving adifferent dose of nicotine (0.125, 0.25, 0.375, and 0.50 mg/kg).Rats were tested in actual hot-plate trials 4, 8, and 13 min afterbeing injected with nicotine.
Results
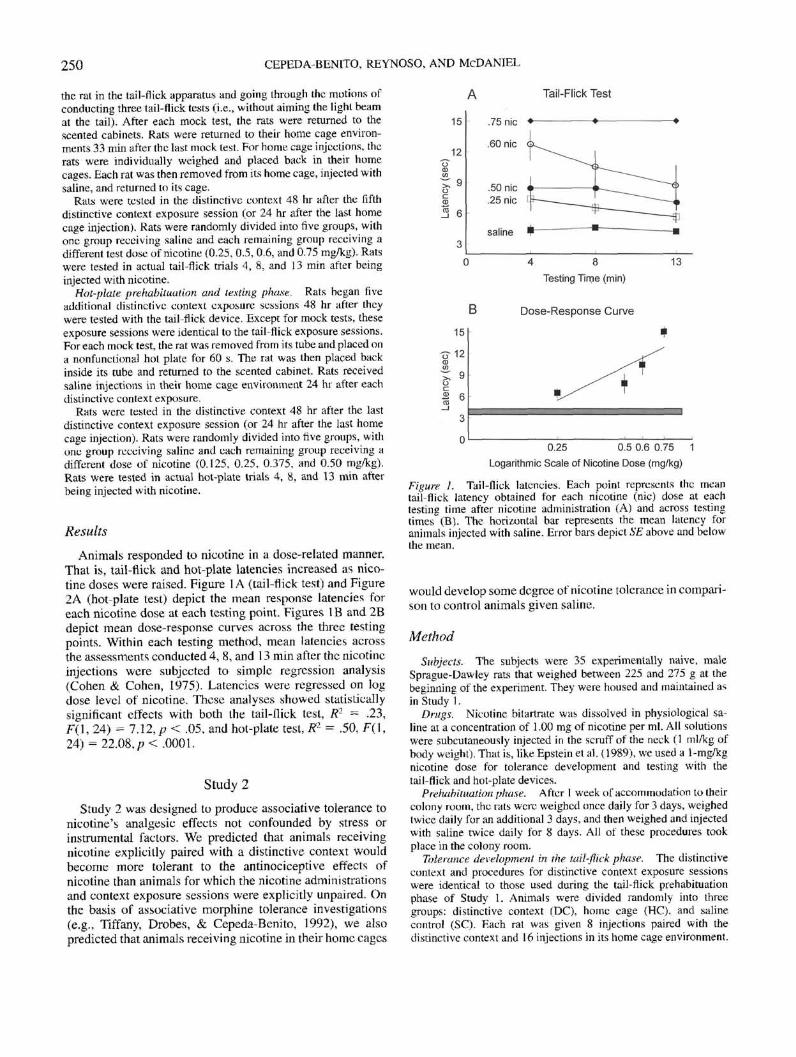
Animals responded to nicotine in a dose-related manner.Thai is, tail-flick and hot-plate latencies increased as nico-tine doses were raised. Figure 1A (tail-flick test) and Figure2A (hot plate test) depict the mean response latencies foreach nicotine dose at each testing point. Figures 1B and 2Bdepict mean dose-response curves across the three testingpoints. Within each testing method, mean latencies acrossthe assessments conducted 4, 8, and 13 min after the nicotineinjections were subjected to simple regression analysis(Cohen & Cohen, 1975). Latencies were regressed on logdose level of nicotine. These analyses showed statisticallysignificant effects with both the tail-flick test, tf2 = .23,F(\, 24) = 1.12,p < .05, and hot-plate test, R1 = .50, F(\,24) = 22.08,p< .0001.
Study 2
Study 2 was designed to produce associative tolerance tonicotine's analgesic effects not confounded by stress orinstrumental factors. We predicted that animals receivingnicotine explicitly paired with a distinctive context wouldbecome more tolerant to the antinociceptive effects ofnicotine than animals for which the nicotine administrationsand context exposure sessions were explicitly unpaired. Onthe basis of associative morphine tolerance investigations(e.g., Tiffany, Drobes, & Cepeda-Benito, 1992), we alsopredicted that animals receiving nicotine in their home cages
Tail-Flick Test
15
12
.50 nic
.25 nic
saline
B
4 8
Testing Time (min)
Dose-Response Curve
13
15
IS 6
0.25 0.5 0.6 0.75 1
Logarithmic Scale of Nicotine Dose (mg/kg)
figure I. Tail-flick latencies. Each point represents the meantail-flick latency obtained for each nicotine (nic) dose at eachtesting time after nicotine administration (A) and across testingtimes (B). The horizontal bar represents the mean latency foranimals injected with saline. Error bars depict 5'£ above and belowIhe mean.
would develop some degree of nicotine tolerance in compari-son to control animals given saline.
Method
Subjects. The subjects were 35 experimentally naive, maleSprague-Dawley rats that weighed between 225 and 275 g at thebeginning of the experiment. They were housed and maintained asin Study 1.
Drugs. Nicotine bitartrate was dissolved in physiological sa-line at a concentration of 1.00 mg of nicotine per ml. All solutionswere subcutaneously injected in the scruff of the neck (1 ml/kg ofbody weight). That is, like Epstein et al. (1989), we used a 1-mg/kgnicotine dose for tolerance development and testing with thetail-flick and hot-plate devices.
Prel-tahituation phase. After 1 week of accommodation to theircolony room, the rals were weighed once daily for 3 days, weighedtwice daily for an additional 3 days, and then weighed and injectedwith saline twice daily for 8 days. All of these procedures tookplace in the colony room.
Tolerance development in the tail-flick phase. The distinctivecontext and procedures for distinctive context exposure sessionswere identical to those used during the tail-flick prehabituationphase of Study 1. Animals were divided randomly into threegroups: distinctive context (DC), home cage (HC), and salinecontrol (SC). Each rat was given 8 injections paired with thedistinctive context and 16 injections in its home cage environment.
TOLERANCE TO NICOTINE ANALGESIA IN THE RAT 251
A Hot-Plate Test
60
50
oj«>, 40
30
20
10
.25 nic
.125 nicsaline
B
Testing Time (min)
Dose-Response Curve
13
60
„ 50o1 40
£ 30CDra 20
10
0.125 0.25 0.375 0.5
Logarithmic Scale of Nicotine Dose (mg/kg)
Figure 2. Hot-plate latencies. Each point represents the meanhot-plate latency obtained for each nicotine (nic) dose at eachtesting time after nicotine administration (A) and across testingtimes (B). The horizontal bar represents the mean latency foranimals injected with saline. Brror bars depict SE above and belowthe mean.
The interval between context exposures was 72 hr. During the 2days that followed each context exposure session, rats received adaily injection in their home cage environment. DC rats receivednicotine in the distinctive context and saline in the home cageenvironment. HC rats received saline in the distinctive context andnicotine and saline for the first and second home cage injections,respectively. SC rats received saline in both environments.
Tolerance development in the hot-plate phase. Following thetail-flick test sessions, animals received two daily saline injectionsin their home cages for 2 days. Then, each rat was given 5injections paired with the distinctive context and 20 injections in itshome cage environment. That is, the ratio of saline to nicotineinjections was greater in the hot-plate phase than in the tail-flickphase. Previous research has shown that these cues can supportassociative tolerance effects in HC animals (e.g., Cepeda-Benito &Tiffany, 1995). After observing high levels of tolerance in HC ratsin the tail-flick assay, we were concerned that HC rats were usinginjection cues to support associative tolerance effects. Therefore.we reduced the likelihood of rats receiving nicotine in the presenceof injection cues.
The interval between context exposures continued to be 72 hr,but home cage injections were administered twice daily after eachdistinctive context exposure. The first of the home cage injectionsconsisted of nicotine for the HC animals. All other home cageinjections consisted of saline for these animals. The interinjectioninterval for each daily pair of home cage injections was 4 hr. DCanimals continued to receive nicotine in the distinctive context and
saline for their home cage injections. SC animals received saline inboth environments. All other experimental procedures were thesame as those used in the hot-plate prehabituation phase of Study 1.
Tail-flick and hot-plate analgesia assessments. Analgesia as-sessments were identical to those conducted in Study 1. Each ratreceived a 1-mg/kg dose of nicotine and was tested 4, 8, and 13 minafter the injection. Tail-flick and hot-plate test sessions wereconducted 72 hr after the eighth and last nicotine context pairings,respectively.
Data analysis. Within each testing method, the data werestudied with a one-between, one-within repeated measures analy-ses of variance. Mean response latency was the dependent variable,group Condition was the between factor, and testing time was thewithin factor. To control for heterogeneity of covariance acrossrepeated measures, we used the Greenhouse-Geisser (1959) tech-nique to adjust the degrees of freedom for the error terms.Following a priori directional hypothesis, we compared meanresponse latencies between SC and HC rats and between HC andDC rats.
Results
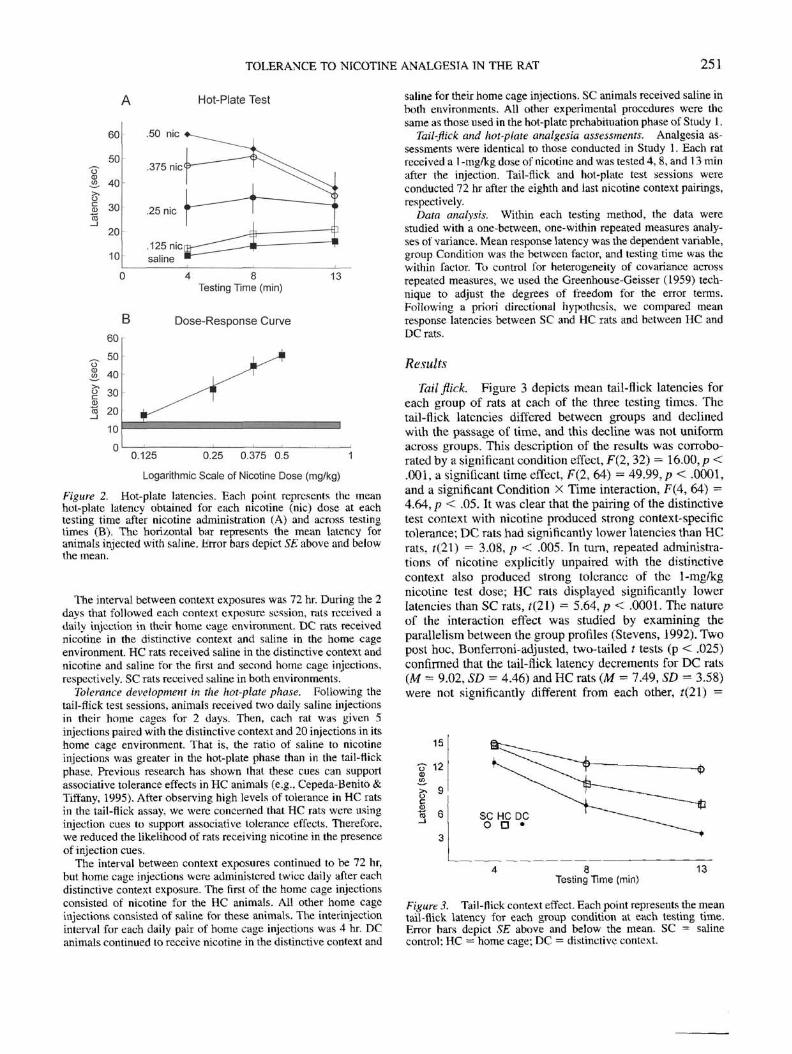
Tail flick. Figure 3 depicts mean tail-flick latencies foreach group of rats at each of the three testing times. Thetail-flick latencies differed between groups and declinedwith the passage of time, and this decline was not uniformacross groups. This description of the results was corrobo-rated by a significant condition effect, F(2, 32) — 16.00, p <.001, a significant time effect, F(2, 64) = 49.99; p < .0001,and a significant Condition X Time interaction, F(4, 64) =4.64, p < .05. It was clear that the pairing of the distinctivetest context with nicotine produced strong context-specifictolerance; DC rats had significantly lower latencies than HCrats, f(21) = 3.08, p < .005. Tn turn, repeated administra-tions of nicotine explicitly unpaired with the distinctivecontext also produced strong tolerance of the 1-mg/kgnicotine test dose; HC rats displayed significantly lowerLatencies than SC rats, t(2l) = 5.64, p < .0001. The natureof the interaction effect was studied by examining theparallelism between the group profiles (Stevens, 1992). Twopost hoc, Bonferroni-adjusted, two-tailed f tests (p < .025)confirmed that the tail-flick latency decrements for DC rats(M = 9.02, SD = 4.46) and HC rats (M = 7.49, SD = 3.58)were not significantly different from each other, £(21) =
15
Testing Time (min)13
Figure 3. Tail-flick context effect. Each point represents the meantail-flick latency for each group condition at each testing time.Error bars depict SE above and below the mean. SC = salinecontrol; HC = home cage; DC = distinctive context.

252 CEPEDA-BENITO, REYNOSO, AND McDANIEL
0.95. p > .05. However, tail-flick latency decrements were
significantly higher for HC rats than for SC rats (M ~ 3.25,
SD = 3.47), f(21) = 2.88, p < .01.
Hot plate. Figure 4 depicts mean hot-plate latencies for
each group of rats at each of the three testing times. The
hot-plate latencies differed between groups and declined
similarly across groups with the passage of lime. This
description of the results was corroborated by a significant
condition effect, F(2, 32) = 12.18, p < .001, a significant
time effect, F(2, 64) = 9.25, p < .001, and no significant
Condition X Time interaction, F< 1. As in the tail-flick test,
it was clear that the pairing of the distinctive test context
with nicotine produced strong context-specific tolerance;
DC rats responded faster than HC rats, r(21) = 3.52, p <
.005. In turn, repeated administrations of nicotine in the
home cage also produced tolerance of nicotine; HC rats
displayed significantly lower latencies than SC rats, r(21) =
4.73, p< .001.
General Discussion
Study 1 replicated the finding that nicotine can produce
analgesic effects in a dose-related manner (Caggiula et al.,
1995; Rogers & Iwamoto, 1993). This effect was found with
both the tail-flick and the hot-plate devices and was indi-
cated by log dose being a significant predictor of tail-flick
and paw-lick latencies. These results were obtained with
procedures that minimized the presence of novelty-induced
stress during testing (see also Ramsay & Woods, 1997; cf.
Caggiula et al., 1995; Rogers & Iwamolo, 1993). That is, rats
were extensively prehabituated to the testing procedures.
The data from Study 2 showed that animals that were
injected with nicotine during the tolerance development
phase were less responsive to nicotine's analgesic effects
than animals that received nicotine only during testing. This
effect was indicated by the lower tail-flick and paw-lick
latencies observed in HC and DC rats than in SC rats. The
data also revealed that animals receiving nicotine administra-
tions explicitly paired with a distinctive environment (DC
rats) were more tolerant than animals receiving as many
nicotine administrations but explicitly unpaired with a
distinctive context (HC rats). The results appear congruent
with the hypothesis that a distinctive context may function
as a CS capable of eliciting associative tolerance effects in
anticipation of nicotine's antinociceptive effects (e.g., Sie-
gel, 1975).
The results of these studies appear to be reliable and
nonspecific, as they were obtained across two functionally
different responses, the spinal tail-flick reflex and the
supraspinal paw-lick response. It is unlikely that the context
effect was caused by the presence of novelty-induced stress
during the tolerance assessment session because all subjects
were extensively familiarized with the experimental and
testing procedures (cf. Caggiula et al., 1989, 1991, 1993;
Epstein et al., 1989; 1991). That is, prehabituation to theexperimental and testing procedures is often regarded as an
effective process for minimizing confounding stress effects
in drug tolerance experiments (e.g., Ramsay & Woods,
1997).
60
tr 50&
£• 40
I-3 30
20
Testing Time (min)13
Figure 4. Hot-plate context effect. Each point represents themean hot-plate latency for each group condition at each testingtime. Error bars depict SE above and below the mean. SC = salinecontrol; HC = home cage; DC = distinctive context.
Our finding that a distinctive context functioned as a CS
for the development of associative drug tolerance replicates
the results of the many studies that have investigated the
development of tolerance to morphine's analgesic effects(e.g., Cepeda-Benito & Tiffany, 1992; Tiffany & Maude-
Griffin, 1988). Therefore, our results are important theoreti-
cally because they help support the generalizability of
associative morphine tolerance phenomena to other drugs.
This assertion is congruent with the statement by Poulos and
Cappell (1991, p. 402) "to go beyond morphine" when
assessing the adequacy of general models of drug tolerance.That is, these authors defended the idea that a comprehen-
sive theory of drug tolerance should establish a generality of
tolerance findings across different drugs (Poulos & Cappell).
We emphasize that instrumental learning could not have
played a role in the context effect because the subjects did
not practice the test response during the tolerance develop-
ment phase (cf. Caggiula et al., 1993; Epstein et al., 1989).
However, this observation merits attention only if instrumen-
tal and associative drug tolerances are defined as being
conceptually different (e.g., Cepeda-Benito & Tiffany, 1995).
Conversely, Poulos and Cappell (1991) defend the idea that
all tolerance that develops to specific drug effects is,
explicitly or implicitly, behaviorally contingent. These au-
thors theorize that, regardless of the experimenter's manipu-
lation, drug tolerance will develop only if drug effects place
behavioral demands on specific physiological systems. That
is, in the absence of behavioral or functional disturbances,
organisms would not have a need to initiate homeostatic
restoration processes, or tolerance.
Nevertheless, from a merely methodological viewpoint,our results at least corroborate the idea that contextual
tolerance can be obtained without assessment of a drug's
effects on behavioral responding after each drug administra-
tion during the tolerance development phase (see also
Cepeda-Benito & Tiffany, 1992; Tiffany et al., 1992).
Therefore, by systematically manipulating various param-
eters within the present design, future research on nicotine
tolerance could further examine the conditions under which
associative nicotine tolerance develops (see Baker & Tif-
fany, 1985; Poulos & Cappell, 1991).
There are two potential problems with our experimental

TOLERANCE TO NICOTINE ANALGESIA IN THE RAT 253
design. First, testing of the same rats first in the tail-flick test
and then in the hot-plate test did not allow us to compare
tolerance effect sizes across both testing methods. That is,
the tail-flick test was given after 8 conditioning sessions,
whereas the hot-plate test was given after 14 nicotine
context pairings. Therefore, any effect size differencesbetween the two methods could not be interpreted. More-
over, we did not control for the possibility that testing of the
rats in the tail-flick test first could have influenced the
hot-plate test results. These problems could have been
avoided by counterbalancing the order of testing across
measurements or by testing independent groups of rats in
each testing device. Nevertheless, we believe that it is safe to
affirm that the order of testing docs not compromise our
interpretation of the results. That is, compared to DC rats,
HC rats had a lower probability of receiving nicotine inside
the distinctive context and a higher probability of receiving
nicotine outside the distinctive context in both test sessions.
Both sets of probabilities are congruent with the prediction
that DC animals would develop a stronger context-drug
association than HC animals (see Rescorla & Wagner,
1972).
The experimental design also could have been improved
by including a placebo test control group for each of the
three conditions, SC, HC, and DC. Given that we repeatedly
paired the testing environment with nicotine for DC but not
SC and HC rats, undrugged control latencies could have
helped to clarify whether the context effects were attribut-
able to differential changes in distinctive context baseline
latencies across groups. For example, the development of
contextual tolerance could have been mediated by a condi-
tioned hyperalgesic response (e.g., Siegel, 1975).
References
Alexander, B. K., & Hadaway, P. K (1982). Opiate addiction: Thecase for an adaptive orientation. Psychological Bulletin, 92,367-381.
American Psychiatric Association. (1994). Diagnostic and statisti-cal manual of mental disorders (4th ed.). Washington, DC:Author.
Baker, T. B., & Tiffany, S. T. (1985). Morphine tolerance ashabituation. Psychological Review, 92, 78-108.
Bardo, M T., & Hughes, R. W. (1979). Exposure to a nonfunctionalhotplate as a factor in the assessment of morphine-inducedanalgesia and analgesic tolerance in rats. Pharmacology Biochem-istry and Behavior, 70,481^185.
Caggiula, A. R., Epstein, L. H., Antelman, S. M., Saylor, S., Knopf,S., Perkins, K. A., & Stiller, R. L. (1993). Acute stress orcorticosterone administration reduces responsiveness to nico-tine: Implications for a mechanism of conditioned tolerance.Psychopharmacology, I I I , 499-507.
Caggiula, A. R., Epstein, L. H., Antelman, S. M., Seymour, M.,Saylor, S., Perkins, K. A., Knopf, S., & Stiller, R. L. (1991).Conditioned tolerance to the anorectic and corticosterone-clevating effects of nicotine. Pharmacology Biochemistry andBehavior, 40, 53-59.
Caggiula, A. R., Epstein, L. H., Perkins, K. A., & Saylor, S. (1995).Different methods of assessing nicotine-induced antinociceptionmay engage different neural mechanisms. Psychopharmacology,122, 301-306.
Caggiula, A. R., Epstein, L. H., & Stiller, R. L. (1989). Changingenvironmental cues reduces tolerance to nicotine-induced an-orexia. Psychopharmacology, 99, 389-392.
Carter, B. L., & Tiffany. S. T. (1996). Cross-tolerance of associativeand nonassociative morphine tolerance in the rat with mu- andkappa-specific opioids. Psychopharmacology, 123, 289-296.
Cepeda-flenito. A., & Short, P. (1997). Morphine's interoceptivestimuli as cues for the development of associative morphinetolerance in the rat. Psychobiology, 25, 236-240.
Cepeda-flenito. A., & Tiffany, S, T. (1992). Effect of number ofconditioning trials on the development of associative toleranceto morphine. Psychopharmacology, 109, 172-176.
Cepeda-Benito, A., & Tiffany, S. T. (1995). Role of drug-administration cues in the associative control or morphinetolerance in the rat. Psychopharmacology, 122, 312-316.
Cepeda-Benito, A., & Tiffany, S. T. (1996a). The failure ofnoncontingent morphine exposure to disrupt context-specificmorphine tolerance. Pharmacology Biochemistry and Behavior,54, 575-580.
Cepeda-Benito, A., & Tiffany. S. T. (1996b). Test-specific manifes-tations of associative tolerance to the analgesic effects ofmorphine in the rat. Psychobiology, 24, 327-332.
Cohen, J.. & Cohen, P. (1975). Applied multiple regression/correlational analysis for the behavioral sciences. Hillsdale, NJ:Erlbaum.
Epstein, L. H.. Caggiula, A. R., Perkins, K. A., McKenzic, S. J., &Smith, J. A. (1991). Conditioned tolerance to the heart rate ofsmoking. Pharmacology Biochemistry and Behavior, 39, 15-19.
Epstein, L. H., Caggiula, A. R., & Stiller, R. L. (1989). Environment-specific tolerance to nicotine. Psychopharmacology, 97, 235-237.
Goudie, A. J., & Demellweek, C. (1986). Conditioning factors indrug tolerance. In S. R. Goldberg & I. P. Stolerman (Eds.),Behavioral analysis of drug dependence (pp. 225-285). NewYork: Academic Press.
Greenhouse, S. W., & Geisser, S. (1959). On methods in theanalysis of profile data. Psychometrika, 24, 95—112.
Grisel, J. E., Wiertelak, E. R.'Watkins, L. R., & Maicr, S. F. (1994),Route of morphine administration modulates conditioned analge-sic tolerance and hyperalgesia. Pharmacology Biochemistry andBehavior, 49, 1029-1035.
Krank, M. D. (1987). Conditioned hyperalgesia depends on thepain sensitivity measure. Behavioral Neuroscience, 101, 854—857.
LeBlanc, A. E.. Gihhins, R. J., & Kalant, H. (1973). Behavioralaugmentation of tolerance to ethanol in the rat. Psychopharmaco-logia. JO, 117-122.
Netto, C. A., Siegfried, B., & Izquierdo, I. (1987). Analgesiainduced by exposure to a novel environment in rats: Effect ofconcurrent and post-training stressful stimulation. Behavioraland Neural Biology, 48, 304-309.
Pomerleau, O. F. (1995). Individual differences in sensitivity tonicotine: Implications for genetic research on nicotine depen-dence. Behavior Genetics, 25, 161-177.
Poulos, C. X., & Cappell, H. (1991). Homeostatic theory of drugtolerance: A general model of physiological adaptation. Psycho-logical Review, 98, 390-408.
Ramsay, D. S., & Woods, S. C. (1997). Biological consequences ofdrug administration: Implications for acute and chronic toler-ance. Psychological Review, 104, 170-193.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovianconditioning: Variations in the effectiveness of reinforcementand nonreinforcement. In A. H. Black & W. F. Prokassy (Eds.),Classical conditioning 11: Current research and theory (pp.64-99). New York: Appleton-Century-Crofts.

254 CEPEDA-BENITO, REYNOSO, AND McDANIEL
Rogers, D. T., & Iwamoto, E. T. (1993). Multiple spinal mediatorsin parenteral nicotine-induced antinociception. Journal of Phar-
macology and Experimental Tlicrapeutics, 267, 341-349.Sherman, J. E. (1979). The effects of conditioning and novelty on
the rat's analgesic and pyretic responses to morphine. Learning
and Motivation, 10, 381-418.Sherman, J. E., Proctor, C., & Strub, H. (1982). Prior hotplate
exposure enhances morphine analgesia in tolerant and drug-naive rats. Pharmacology Biochemistry and Reluivior, 17, 229-
232.
Siegel, S. (1975). Evidence from rats that morphine tolerance is alearned response. Journal of Comparative and Physiological
Psychology, 89, 498-506.Stevens, i. (1992). Applied multivariate statistics for the social
sciences (2nd ed.). Hillsdale, NJ: Erlbaum.Tiffany, S. T., Drobes, D. J., & Cepeda-Benito, A. (1992).
Contribution of associative and nonassocialive processes to the
development of morphine tolerance. Psvchopharmacology, 109,
185-190.Tiffany, S. T., & Maude-Griffin, P. M. (1988). Tolerance to
morphine in the rat: Associative and nonassociative effects.
Behavioral Neuroscience, 102, 534-543.Trujillo, K. A., & Akil, H. (1995). Excitatory ainino acids and drugs
of abuse: A role for W-methyl-D-asparate receptors in drug
tolerance, sensitization and physical dependence. Drug and
Alcohol Dependence, 38, 139-154.Wolgin, D. L. (1989). The role of instrumental learning in
behavioral tolerance to drugs. In A. J. Goudie and M. W.Emmit-Ogiesby (Eds.), Pxvchoactive drugs: Tolerance andsensitization (pp. 17-114). Clifton, NJ: Humana Press.
Yang, C., Wu, W., & Zbuzek, V. K. (1992). Antinociceptive effectof chronic nicotine and nociceptive effect of its withdrawalmeasured by hot-plate and tail-flick in rats. Psvchopharmacol-
ogy, 106, 417-420.Yobiirn, B. C., Morales, R.. Kelly, D. D., & mturrisi, E. C. (1984).
Constraints on the tail-flick assay: Morphine analgesia andtolerance are dependent upon locus of tail stimulation. Life
Sciences, 34, 1755-1762.Young, A. M., & Goudie, A. J. (1994). Adaptive processes
regulating tolerance to behavioral effects of drugs. In F. E.Bloom & D. J. Kupfer (Eds.), Pxychopharmacology: The fourth
generation of progress (pp. 657-811). New York: Raven Press.
Received November 18, 1997
Revision received March 24, 1998
Accepted March 25, 1998
Low Publication Prices for APA Members and Affiliates
Keeping you up-to-date. All APA Fellows, Members, Associates, and Student Affiliates
receive—as part of their annual dues—subscriptions to the American Psychologist and
APA Monitor. High School Teacher and International Affiliates receive subscriptions to
the APA Monitor, and they may subscribe to the American Psychologist at a significantly
reduced rate. In addition, all Members and Student Affiliates are eligible for savings of up
to 60% (plus a journal credit) on all other APA journals, as well as significant discounts on
subscriptions from cooperating societies and publishers (e.g., the American Association for
Counseling and Development, Academic Press, and Human Sciences Press).
Essential resources. APA members and affiliates receive special rates for purchases of
APA books, including the Publication Manual of the American Psychological Association,
and on dozens of new topical books each year.
Other benefits Of membership. Membership in APA also provides eligibility for
competitive insurance plans, continuing education programs, reduced APA convention fees,
and specialty divisions.
More information. Write to American Psychological Association, Membership Services,
750 First Street, NE, Washington, DC 20002-4242.
Copyright © 2022 FDOKUMEN




















