
Associative interference between cues and between outcomes presented together and presented apart:...
23
Behavioural Processes 57 (2002) 163 – 185 Associative interference between cues and between outcomes presented together and presented apart: an integration Ralph R. Miller *, Martha Escobar Department of Psychology, State Uniersity of New York – Binghamton, Binghamton, NY 13902 -6000, USA Accepted 16 November 2001 Abstract In recent years, ‘stimulus competition’ in the study of acquired behavior has referred exclusively to (a) associative competition between cues trained in compound (e.g. overshadowing and blocking). Rarely cited are older experiments cast in the verbal learning tradition, now complemented with data from humans and rats in Pavlovian preparations, that demonstrate (b) competition between cues separately trained with a common outcome (i.e. proactive and retroactive interference). Similarly neglected are numerous examples of (c) competition between outcomes separately trained with a common cue within the verbal learning literature (also proactive and retroactive interference) as well as within the Pavlovian literature (i.e. counterconditioning). Recent data demonstrate (d) competition between outcomes trained in compound, thereby completing the four cells of a 2 ×2 matrix (competing stimuli trained together vs. trained apart and the competing stimuli being cues or outcomes) which highlights the ubiquitous nature of associative stimulus interference/competition. Most contemporary theories of acquired behavior can account for the phenomena in one or at most two cells of this matrix. Whether a common mechanism underlies the phenomena in all four cells of the matrix is currently unclear, but until such time as data preclude a common mechanism, parsimony encourages efforts to develop a model that encompasses all four cells. Here we offer a tentative model that addresses all four cells, albeit with two processes. © 2002 Elsevier Science B.V. All rights reserved. Keywords: Associative interference; Learning theories; Stimulus competition www.elsevier.com/locate/behavproc 1. Introduction The capacity to account for cue competition is a benchmark that currently must be met by any theory of acquired behavior (i.e. learning). Cue competition has held this central position ever since Kamin (e.g. 1968) argued that acquisition of an association between a cue and an outcome requires pairing the cue with a ‘surprising’ out- come, (i.e. the occurrence of the outcome must constitute new information about the environ- ment). For example, if a conditioned stimulus (CS) A is paired with (immediately followed by) an unconditioned stimulus (US) prior to a com- pound of CSs X and A being paired with the same US (i.e. A–US followed by AX–US), X will accrue less control of behavior than if the A–US pairings had not occurred. This deficit in respond- * Corresponding author. Tel.: +1-607-777-2291; fax: +1- 607-777-4890. E-mail address: [email protected] (R.R. Miller). 0376-6357/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved. PII:S0376-6357(02)00012-8
Transcript of Associative interference between cues and between outcomes presented together and presented apart:...

Behavioural Processes 57 (2002) 163–185
Associative interference between cues and between outcomespresented together and presented apart: an integration
Ralph R. Miller *, Martha EscobarDepartment of Psychology, State Uni�ersity of New York–Binghamton, Binghamton, NY 13902-6000, USA
Accepted 16 November 2001
Abstract
In recent years, ‘stimulus competition’ in the study of acquired behavior has referred exclusively to (a) associativecompetition between cues trained in compound (e.g. overshadowing and blocking). Rarely cited are older experimentscast in the verbal learning tradition, now complemented with data from humans and rats in Pavlovian preparations,that demonstrate (b) competition between cues separately trained with a common outcome (i.e. proactive andretroactive interference). Similarly neglected are numerous examples of (c) competition between outcomes separatelytrained with a common cue within the verbal learning literature (also proactive and retroactive interference) as wellas within the Pavlovian literature (i.e. counterconditioning). Recent data demonstrate (d) competition betweenoutcomes trained in compound, thereby completing the four cells of a 2×2 matrix (competing stimuli trainedtogether vs. trained apart and the competing stimuli being cues or outcomes) which highlights the ubiquitous natureof associative stimulus interference/competition. Most contemporary theories of acquired behavior can account forthe phenomena in one or at most two cells of this matrix. Whether a common mechanism underlies the phenomenain all four cells of the matrix is currently unclear, but until such time as data preclude a common mechanism,parsimony encourages efforts to develop a model that encompasses all four cells. Here we offer a tentative model thataddresses all four cells, albeit with two processes. © 2002 Elsevier Science B.V. All rights reserved.
Keywords: Associative interference; Learning theories; Stimulus competition
www.elsevier.com/locate/behavproc
1. Introduction
The capacity to account for cue competition isa benchmark that currently must be met by anytheory of acquired behavior (i.e. learning). Cuecompetition has held this central position eversince Kamin (e.g. 1968) argued that acquisition of
an association between a cue and an outcomerequires pairing the cue with a ‘surprising’ out-come, (i.e. the occurrence of the outcome mustconstitute new information about the environ-ment). For example, if a conditioned stimulus(CS) A is paired with (immediately followed by)an unconditioned stimulus (US) prior to a com-pound of CSs X and A being paired with thesame US (i.e. A–US followed by AX–US), X willaccrue less control of behavior than if the A–USpairings had not occurred. This deficit in respond-
* Corresponding author. Tel.: +1-607-777-2291; fax: +1-607-777-4890.
E-mail address: [email protected] (R.R. Miller).
0376-6357/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved.
PII: S0376 -6357 (02 )00012 -8

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185164
Fig. 1. A 2×2 matrix of different types of associative interference effects. In each cell, some representative examples of interferenceprocedurally appropriate for that cell are listed. ‘X’ represents the target conditioned stimulus, ‘A’ represents the interfering cue,‘US’ represents an unconditioned stimulus, ‘O’ represents an outcome, which might be a biologically significant US or an innocuousstimulus that is later paired with a US, and O1 and O2 (US1 and US2 if they are biologically significant) represent two distinctlydifferent outcomes. The larger font of the overshadowing cue in cell 1 (denoted here as ‘A’) reflects the finding that overshadowingof a target cue is greatest when the overshadowing cue is of considerably higher salience than the target cue. Contemporary modelsof acquired behavior have focused almost exclusively on phenomena of cell 1 to the exclusion of phenomena in cells 2, 3 and 4.
ing to CS X is called ‘blocking’ (Kamin, 1968;Lashley, 1942). In Kamin’s framework, blockingoccurs because the US is not surprising when it ispaired with CS X due to CS A’s already providinginformation that the US is about to occur. SinceKamin’s influential writings, this principle hasbeen incorporated into almost every new model oflearning in one or another form (e.g. McLarenand Mackintosh, 2000; Pearce, 1987; Pearce andHall, 1980; Rescorla and Wagner, 1972). Onereason that this view has been so widely acceptedis that it is convergent with the most apparentfunction of learning, which is to anticipate eventsin the environment so that an organism can pre-pare for and perhaps even modify them. If onecue already predicts an outcome, there is seem-
ingly little utility in learning that a second cuethat accompanies the first cue also predicts thesame outcome. This ‘sensible’ view of learning atthe functional level nicely explains other examplesof cue competition, such as overshadowing(Pavlov, 1927), the relative stimulus validity effect(Wagner et al., 1968), and the overexpectationeffect (Rescorla, 1970). As a result of this accep-tance of Kamin’s view, researchers in recent yearshave focused on phenomena consistent with thisprinciple (i.e. competition between cues trainedtogether [in compound]), and have tended to ig-nore other types of stimulus competition. How-ever, other forms of stimulus competition havebeen reported in the literature on learning overmuch of the last century.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 165
Fig. 1 depicts four different types of potentialstimulus competition, distinguished from one an-other by (1) whether the competing stimuli arecues (i.e. antecedent events, which might serve assignals for events which immediately follow them)or outcomes (subsequent events), and (2) whetherthe competing elements are trained in compound(simultaneous or serial) or in widely separatedtrials. Competition between cues presented to-gether during training, which has received somuch attention since Kamin’s papers were pub-lished in the late 1960s, constitutes what is iden-tified here as cell 1 of the matrix in Fig. 1. Thepurpose of this paper is to summarize old andnew data documenting the occurrence of types ofstimulus interference (i.e. competition) that consti-tute examples of cells 2, 3, and 4. Recognitionthat these other types of interference occur wouldsuggest reassessment of models of learning and/oracquired responding that address only phenomenain cell 1 and fail to speak to phenomena in cells 2,3, and 4.
2. Cell 2—interference between outcomes trainedtogether
Cell 2 refers to competition between outcomesthat are trained together (i.e. a common cue leadsto a compound of two [or more] outcomes). Assuch, it is in some sense the mirror image of cell 1,which depicts competition between cues (i.e. an-tecedent events in the training trial dyad) trainedtogether. The fundamental problem in assessingthe occurrence of this sort of interference (at leastwith nonhuman animals) is that the most straight-forward experimental designs call for presentingtwo unconditioned stimuli (USs) at the same time(i.e. CS�US1+US2). If the two different USssupport the same response (e.g. approach), com-petition is difficult to assess because there wouldbe ambiguity concerning whether CS–US1 orCS–US2 associations underlie the observed re-sponse. And if the two USs support differentresponses (e.g. approach and withdrawal),diminution of the (target) conditioned responseappropriate for one US due to the presence dur-ing conditioning of the other US could be due to
response competition, which is interesting in itsown right, but is likely quite different from theassociative interference in which we here are cen-trally interested. Associative interference is as-sumed to be less peripheral than responsecompetition, and to reflect competition betweenassociations (in this case, CS–US1 and CS–US2)either for retrieval (i.e. performance deficit) or forstorage in memory (i.e. acquisition deficit or lossof information).
One way to circumvent the response competi-tion problem is to examine situations in whichthere are two outcomes, but neither is biologicallysignificant (i.e. at the presumed time of interfer-ence, they do not elicit high levels of responding)during training (when they are presented in com-pound). If neither outcome supports responding,response competition would be an implausiblesource of interference between X–O1 and X–O2,where CS X is the target cue and O1 and O2 arebiologically innocuous outcomes. Of course, toassess whether the presence of O2 during trainingof X–O1 degraded the X–O1 association instored memory strength or in retrievability, O1must be made biologically significant prior totesting. Hence, we performed several studies thatembedded outcome competition in a sensory pre-conditioning procedure. In sensory precondition-ing (Brogden, 1939), two innocuous stimuli withno consequence for behavior are paired. Then oneof these stimuli is paired with a biologically sig-nificant stimulus (US), and responding to theother stimulus is typically observed (see the tophalf of Fig. 2).
2.1. Standard preparation
Before we turn to data, we here describe ourstandard preparation which was used in all of theexperiments that we describe below. Our subjectswere naive, adult, water-deprived rats. The cuesand innocuous outcomes were moderate intensitylights, tones, clicks, and white noise. The US wasa brief, mild, footshock. Subjects were first al-lowed to locate a water tube in the experimentalapparatus. Then they were exposed to the experi-mental treatments. Prior to testing, they wereagain acclimated to the experimental context in
R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185166
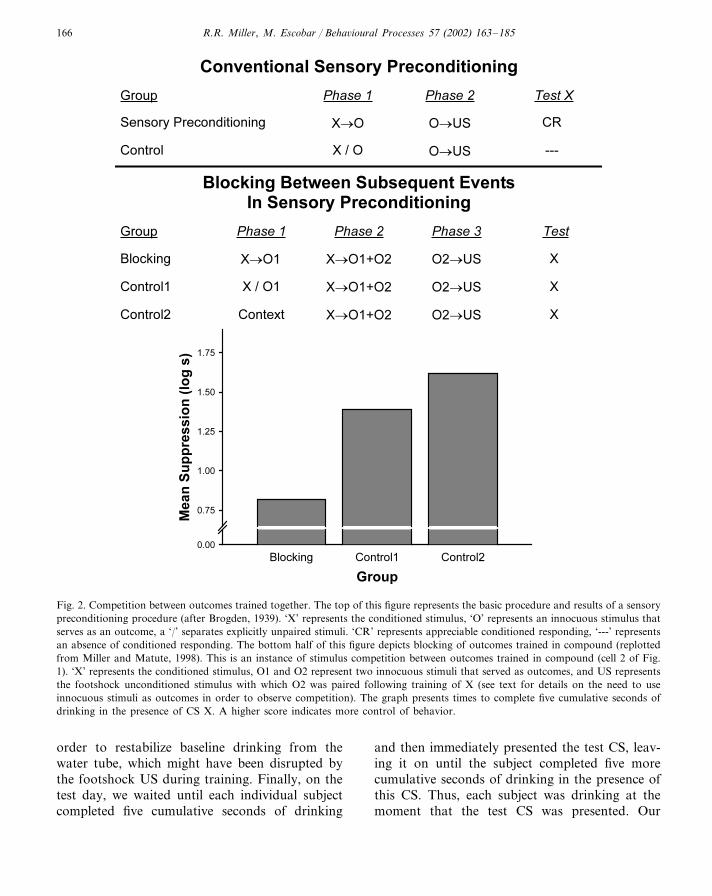
Fig. 2. Competition between outcomes trained together. The top of this figure represents the basic procedure and results of a sensorypreconditioning procedure (after Brogden, 1939). ‘X’ represents the conditioned stimulus, ‘O’ represents an innocuous stimulus thatserves as an outcome, a ‘/’ separates explicitly unpaired stimuli. ‘CR’ represents appreciable conditioned responding, ‘---’ representsan absence of conditioned responding. The bottom half of this figure depicts blocking of outcomes trained in compound (replottedfrom Miller and Matute, 1998). This is an instance of stimulus competition between outcomes trained in compound (cell 2 of Fig.1). ‘X’ represents the conditioned stimulus, O1 and O2 represent two innocuous stimuli that served as outcomes, and US representsthe footshock unconditioned stimulus with which O2 was paired following training of X (see text for details on the need to useinnocuous stimuli as outcomes in order to observe competition). The graph presents times to complete five cumulative seconds ofdrinking in the presence of CS X. A higher score indicates more control of behavior.
order to restabilize baseline drinking from thewater tube, which might have been disrupted bythe footshock US during training. Finally, on thetest day, we waited until each individual subjectcompleted five cumulative seconds of drinking
and then immediately presented the test CS, leav-ing it on until the subject completed five morecumulative seconds of drinking in the presence ofthis CS. Thus, each subject was drinking at themoment that the test CS was presented. Our

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 167
dependent variable was the time that it tooksubjects to complete five cumulative seconds ofdrinking in the presence of the test CS. Scoreswere converted to log seconds in order to betternormalize the distribution of scores withingroups. Longer times to complete five cumula-tive seconds of drinking presumably representgreater fear of the test CS due to its ability tobetter activate the expectation of the footshockUS. All of our CSs were auditory or visualcues, which were appropriately counterbalancedin each experimental design.
2.2. E�idence of interference between outcomestrained together
We first used sensory preconditioning to ex-amine competition between outcomes inEsmoriz-Arranz et al. (1997) and followed thispaper with Miller and Matute (1998), whichmade the same point using somewhat differentcontrol groups. The treatment and mean testscores of some of the experimental groups fromthe Miller and Matute study are depicted in thelower half of Fig. 2. In phase 1, we first estab-lished an association between CS X and out-come 1. Then in phase 2 we paired CS X with asimultaneous compound of outcome 1 and out-come 2 (outcomes 1 and 2 were innocuous stim-uli). Next, we made outcome 2 biologicallysignificant by pairing it with footshock. Finally,we measured fear induced by CS X. Pre-sumably, suppression of drinking by CS X wasa direct function of the strength of the X–O2association because outcome 1 had no biologicalsignificance either during training or testing. Ascan be seen in Fig. 2, prior training of the CSX–O1 association decreased behavioral controlby CS X (group blocking), with such controlnecessarily being mediated by the X–O2 associ-ation as only O2 was paired with the footshock.These studies provide clear demonstrations ofblocking of outcomes presented in compoundduring training. Competition between outcomessuch as this is beyond the domain of Kamin’s(1968) view that the absence of surprise was thebasis of stimulus competition, as well as formost of the more formal models that are de-
scended from it (e.g. Rescorla and Wagner,1972)
Interestingly, Rescorla (1980) (pp. 90–97) ob-served similar results in a second-order condi-tioning design. The fact that he usedsecond-order conditioning allowed responsecompetition as a potential interpretation of hisobservations. However, his central interest inthese experiments was second-order condition-ing, not competition between outcomes. Al-though he noted that his data suggestedcompetition between outcomes trained in com-pound, he never pursued nor publicized thefinding, probably because it did not conform tothe informational hypothesis that was and still iscommonly used to account for stimulus compe-tition (prior knowledge based on a CS X–O1association does not make the occurrence of O2after CS X unsurprising or redundant) (but seeRescorla, 1991).
Other studies have been published demon-strating that subsequent stimuli paired with acommon antecedent stimulus on the same train-ing trial(s) yield the greatest competition whenthe subsequent stimuli have the same temporalrelationship to the antecedent stimulus. The roleof this variable can be observed in at least twodifferent situations. First, the competing subse-quent stimuli can be presented in serial com-pound (e.g. X�O1�O2) as opposed tosimultaneous compound (e.g. X�O1+O2).Second, the subsequent stimuli (O1 and O2) canbe presented simultaneously during target train-ing but on later trials the temporal relationshipbetween them and the common antecedent stim-ulus (X) is shifted so that O1 and O2 come tohave a different temporal relationship to X. Forexample, we could train X�O1+O2 with aPavlovian delay procedure and then give furtherX�O2 training with a trace procedure (i.e. withan interval between X and O2). The usual out-come of this procedure is that interference withthe X–O1 association is reduced, as comparedwith a group that lacks the further X�O2training (Blaisdell et al., 1997; Burger et al.,2000). We return to the implications of this ob-servation later.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185168
3. Cell 3— interference between cues trainedapart
Cell 3 of Fig. 1 refers to competition betweencues that are trained apart, but have a commonoutcome. The pairings of the interfering cue withthe common outcome can come before the targetpairings (ordinarily called proactive interference),after the target pairings (ordinarily called retroac-tive interference), or interspersed with the targetpairings. Although the interspersed situation hasbeen little examined,1 there is a considerable liter-ature concerning proactive and retroactive inter-ference, most of it dating back to the vast verballearning literature published during the middle ofthe 20th century (for a review see Slamecka andCeraso, 1960). Typically, the basic preparationconsisted of human participants being presentedwith a list of paired associates (words, nonsensesyllables, or trigrams) of the form A�O1 (e.g.‘dog-chair’), then a second pair of the form B�O2 (e.g. ‘cat-couch’). At test, participants werepresented with a cue (A if the concern wasretroactive interference by B, or B if the concernwas proactive interference by A) and asked whatoutcome had been paired with it. Although peoplewere clearly able to learn these paired associatesover repeated trials, the likelihood of them re-calling that A had been paired with O1 sometimesdeteriorated if they had also been exposed toB�O1 (i.e. if a common outcome served as O1and O2). These results appear to be clear exam-ples of cell-3 type interference. However, this pro-cedure was used exclusively with humanparticipants, it was usually embedded in a longlist of paired associates, and it was basically a
verbal task which possibly depended upon cogni-tive faculties that did not engage underlying basiclearning processes (e.g. Pavlovian conditioning).For these reasons, some people have wonderedwhether these observations should be used toevaluate modern learning theories, in the sameway that overshadowing and blocking (i.e. compe-tition between cues presented together) are usedbecause verbal learning effects could reflect pro-cesses other than associative learning. Indeed, ver-bal learning studies often lacked control groups toallow assessment of underlying processes, such asthe influence of degrading of the A–O1 correla-tion by presenting O1 in the absence of A (duringthe B�O1 pairings) as opposed to learning asecond association to the same outcome. More-over, the verbal learning literature concerning thissort of interference is ambiguous. Although therewere many reports of such interference (e.g. Post-man, 1962), there were also a number of failuresto obtain it (e.g. Jung, 1963). Unfortunately, in-terest in such interference effects decreased beforethe variables determining this ambiguity werethoroughly studied.
3.1. Interference between cues trained apart innon�erbal preparations
Recently, Matute and Pineno (1998) demon-strated interference between cues trained apart,specifically retroactive interference. Their studiesused human participants playing a video game inwhich they had to anticipate a specific outcome.Verbal instructions were used in their task, butthe actual training experiences (i.e. Pavlovian con-tingencies between colors presented on a com-
Fig. 3. Competition between conditioned stimuli trained apart with a common outcome in first-order conditioning and in sensorypreconditioning. The top panel summarizes the treatments of some groups from a first-order conditioning experiment that failed toobtain retroactive interference, whereas the bottom panel summarizes an experiment that did yield retroactive interference when thetarget and competing training occurred embedded within the first part of a sensory preconditioning procedure (both replotted fromEscobar et al., 2001). The target conditioned stimulus (CS) is represented by ‘X,’ the interfering CS is represented by ‘A,’ and theinitially innocuous stimulus outcome is represented by ‘O.’ After the target training (phase 1) and the interfering training (phase 2)the outcome was paired with a footshock unconditioned stimulus (US). The graphs present times to complete five cumulativeseconds of drinking in the presence of CSs X and A. A higher score indicates more control of behavior. For details, see text andEscobar et al. (2001).
1 If anything, intermixed training should encourage stimulusgeneralization and facilitate learning.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 169
Fig. 3.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185170
puter monitor) and responses used to assess[retroactive] inference (i.e. rate of button pressing)were nonverbal. Our laboratory has recently per-formed analogous studies using rats as subjects,first replicating their basic findings and thenadding studies to further illuminate the underlyingprocesses. Because Matute and Pineno reportedretroactive interference, and the verbal learningexperiments often found retroactive interferenceeasier to obtain than proactive interference, weinitially focused on retroactive interference.
In phase 1, rats were exposed to pairings of thetarget cue and outcome in which the outcome wasa biologically significant unconditioned stimulus(footshock; X�US). Then, in phase 2, groupcompetition was exposed to A�US pairings. Ascan be seen in the top of Fig. 3, subsequentresponding to cue X was not attenuated relative toa control group that received A-alone presenta-tions in Phase 2 (see top panel of Fig. 3). That is,no retroactive interference of X�US training byA�US training was observed. However, onemust note that in this experiment we first endowedcue X with a response potential (in phase 1) andthen in phase 2 we tried to attenuate (interferewith) that response potential. Considerable priorwork in our laboratory has determined that, allother factors being equal, it is distinctly moredifficult to attenuate a response potential that isalready established than to prevent the initialestablishment of that response potential (Dennis-ton et al., 1996; Miller and Matute, 1998; Oberlinget al., 2000). Thus, by using first-order condition-ing in phase 1, we may have been using a situationin which retroactive interference is particularlydifficult to obtain.
In contrast to our study with a footshock out-come, in Matute and Pineno’s (1998) studies, forethical and practical reasons having to do with theuse of human subjects, the outcome was notbiologically significant. To better approximatethat condition in our laboratory with rats assubjects, we recast the initial experiment within thefirst half of a sensory preconditioning procedure.By our avoiding first-order conditioning, in phase2 we were effectively trying to induce retroactiveinterference with an association that did not sup-port responding at the time of phase 2. The
essential features of this study are depicted in thebottom panel of Fig. 3. As in the precedingexperiment, in phase 1 rats were given targetpairings, but now the US was replaced by aninnocuous outcome (i.e. X�O). In phase 2, differ-ent treatments were given to the various groupswith the critical group (competition) receiving apotentially interfering treatment that consisted ofpairing cue A with the same innocuous outcomethat had been used in phase 1 (i.e. A�O). Onlyafter completion of the interference treatment(phase 2) was the Outcome made biologicallysignificant through O�US (footshock) pairingsso that there would be some motivation for behav-ior with which to assess the current efficacy of theX�O association. As can be seen in the lowerpart of Fig. 3, group competition showed substan-tially less responding to cue X than did any of thecontrol groups, which were exposed to either A-alone (group Cx–A), O-alone (group Cx–O), orno event (group Cx-alone) during phase 2. That is,retroactive interference as a result of A�O train-ing was evident. Moreover, the lack of retroactiveinterference in group Cx–O relative to groupCx–alone indicates that the source of the interfer-ence in group comp was not due to degradedcontingency as a result of phase 2 exposure to theoutcome. (Possibly, more phase 2 exposure to Owould have reduced behavioral control by X as aresult of reducing the contingency between X andO.) The details of this study are described inEscobar et al. (2001).
3.2. Context dependency
Toward trying to better understand the basis ofinterference in cell 3 of Fig. 1, we sought toidentify factors that influence when one will ob-serve proactive as opposed to retroactive interfer-ence. The two phases of interference training insome sense create ambiguity with respect to thecommon element, which is the outcome for cell 3.In the basic learning literature, there is strongevidence that simple associations are relativelyindependent of the contexts in which they weretrained. That is, transfer from a training context toa different test context usually results in little lossof behavioral control. However, if there is am-biguity, such as two conflicting associations,

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 171
Fig. 4. Competition between cues trained apart in different contexts. Conditioned stimuli X and A were paired with innocuousstimulus outcome O. Subsequently, O was paired with a footshock US. Subscripts indicate whether the treatment took place incontext 1 or 2. The group names refer first to the phase 2 treatment and then, after the hyphen, to the context in which the twotests occurred. The graph presents times to complete five cumulative seconds of drinking in the presence of CSs X and A. A higherscore indicates more control of behavior. Test X assessed retroactive interference and test A assessed proactive interference. Fordetails, see text and Escobar et al. (2001).
then organisms appear to use the context to dis-ambiguate the situation, that is, determine whichof two conflicting associations immediately ob-tains (e.g. Bouton, 1993). Here, we sought to seeif the inherent ambiguity created by the twophases of interference training (see phases 1 and 2for group comp in the lower half of Fig. 3) alsocreated a context dependency. This was done intwo experiments, one in which we manipulated
the background cues as the contexts of learningand testing, and the other in which we used apunctate (i.e. brief) stimulus to define the immedi-ate context.
In the first experiment, all rats were given X�O pairings in context 1 followed by various treat-ments, most notably A�O pairings (see Fig. 4for the specific treatments received by each group)in context 2, which was distinctly different from

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185172
context 1. Then, the initially innocuous outcomeO was paired with a footshock US in both con-texts 1 and 2. Finally subjects in group comp-1were tested in context 1 with CS X and CS A,whereas all other subjects were similarly tested incontext 2. In group comp-1, behavioral controlwas strong for CS X and weak for CS A (i.e.proactive interference with the A�O associationby the X�O association), whereas in groupcomp-2, the results were reversed (i.e. retroactiveinterference with the X�O association by theA�O association). The remaining three groupsexhibited strong responding to X and weak re-sponding to A (which had not been paired with Ofor these groups). Thus, the specific context usedfor testing appears to have influenced (i.e. primed)which association was most readily retrieved whena test cue (X or A) was presented. Note that, as afunction of test conditions, this experiment ob-tained evidence of both retroactive and proactiveinterference; specifically, the type of interferenceobserved was influenced by which training contextwas used for testing. Importantly, the potential ofthe specific test context background cues to mod-ulate which association was behaviorally evi-denced indicates that at the time of testing bothassociations were still at some level encoded in thecomp subjects; phase 2 training did not produce aloss of the association formed during phase 1.
In the second experiment, we used a singlecontext for all phases of the experiment and at-tempted to prime either the phase 1 or phase 2association by presenting at the start of the testingsessions a stimulus that was intermittently pre-sented either during phase 1 (B) or during phase 2(C) training (see Fig. 5). During training, both Band C were presented sufficiently far away in timefrom the X�O and A�O training trials that Band C acquired little behavioral control. As canbe seen in Fig. 5, group none, which had nopriming stimulus presented during the test trialsexhibited stronger responding to A than X, indi-cating retroactive interference. In contrast, prim-ing the test trials with stimulus B enhancedbehavioral control by X and attenuated it by A.Priming the test trials with stimulus C was seen tohave the opposite effect, although response levelsdid not differ appreciably from those of group
none, consistent with the fact that, without prim-ing, this situation yielded retroactive interference.The notable finding is that a punctate primingstimulus transformed a situation in which retroac-tive interference was evident (group none) into asituation in which proactive interference was evi-dent (group B). As in the preceding experiment,the effectiveness of the priming cues to modulatewhich association was behaviorally evidenceddemonstrates that the observed deficits in re-sponding to X and A were retrieval failures ratherthan a consequence of irrevocably lost memories.(Although we emphasize priming here, surely re-tention intervals and amount of training are alsopotent variables.) These studies in conjunctionwith the several examples of this sort of interfer-ence within the verbal learning literature givecredence to the occurrence of competition be-tween cues trained apart with a common out-come, and begin to suggest some of the factorsthat influence such competition. (Note that we areassuming that there are strong parallels betweenpaired-associate learning and interference phe-nomena within the Pavlovian tradition. See Dona-hoe and Palmer (1994) for a review of thesimilarities and differences between paired-associ-ate and Pavlovian learning.)
4. Cell 4— interference between outcomes trainedapart
Interference between outcomes trained apartwith a common cue is evident in a number ofdifferent situations. Among these are the vastnumber of demonstrations of A�B, A�C inter-ference in the old verbal learning literature (e.g.Postman, 1962). However, as discussed before,verbal learning experiments have sometimes beendiscounted for possibly engaging higher-ordercognitive processes unique to humans processingverbal information. Additional examples of cell4-type interference include counterconditioningand extinction. In counterconditioning (e.g. Dear-ing and Dickinson, 1979; Pavlov, 1927), a CS isfirst paired with one outcome (US1) and thenwith a second outcome (US2). The more obviouseffect is that the X�US2 pairings interfere with

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 173
Fig. 5. Competition between cues trained apart with different punctate cues being presented during phase 1 and phase 2. For eachanimal, training and testing occurred in the same context. Conditioned stimuli X and A were paired with innocuous stimulusoutcome O. Interspersed with the X�O trials of phase 1 were presentations of stimulus B and interspersed with the A�O trialsof phase 2 were presentations of stimulus C. Subsequently O was paired with a footshock US. During test sessions, B or C waspresented some minutes before X or A was presented. The group names refer to the priming cue presented at the beginning of eachtest trial except for group unpaired, the name of which reflects its phase 2 treatment. The graph presents times to complete fivecumulative seconds of drinking in the presence of CSs X and A. A higher score indicates more control of behavior. Test X assessedretroactive interference and test A assessed proactive interference. For details, see text and Escobar et al. (2001).
behavior based on the X�US1 association, andless obviously, but clearly occurring, behaviorbased on the X�US2 association is subject tointerference by the X�US1 experience. In extinc-tion, a CS is first paired with an outcome (X�US) and then the CS is presented alone (X�noUS). That the extinction treatment decreases con-ditioned responding to the CS can also be viewedas a form of interference. Moreover, whether thisinterference is retroactive or proactive can bemodulated by contextual or punctate stimuli. For
example, Bouton and his associates have exten-sively examined the consequences of acquisitiontraining in one context and extinction treatmentin a second context. Although the extinction treat-ment eliminated conditioned responding to X inthe extinction context, vigorous responding wasobserved in both the training context and in athird neutral context (e.g. Bouton and Bolles,1979; for a punctate priming stimulus, see Brooksand Bouton, 1993). This stimulus-induced recov-ery from extinction is called renewal and it has

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185174
now been observed in many different species andpreparations.
4.1. Response competition and temporal factors
As mentioned above, counterconditioning issubject to interpretation in terms of responsecompetition, which is potentially important but isnot to be confused with associative interference,the topic at issue here. Moreover, acquisitionfollowed by extinction uses a US and the absenceof the US as the two outcomes, which may intro-duce some factors not inherent to competitionbetween two different outcomes, each of whichconsists of a stimulus presentation. Hence, we(Escobar et al., 2002) conducted some experi-ments to see if we could obtain competition,specifically retroactive interference, between twodistinctly different outcomes trained apart with acommon cue. In addition to this demonstration,these studies had two additional goals. One wasto compare the magnitude of competition betweenoutcomes trained apart with the magnitude ofcompetition between cues trained apart; and theother was to examine if the temporal relationshipbetween the cue and the two outcomes influencedthe degree of interference. To obviate a role ofresponse competition, as we did in our examina-tion of cell 2, the two phases of interferencetraining were embedded in the first part of asensory preconditioning procedure. In the firstexperiment, during phase 1 all rats were exposedto A�B pairings. In phase 2, group A-samereceived A�C pairings (potentially creating com-petition between outcomes B and C); group A-diffreceived C�A pairings which should have pro-duced associative interference similar to groupA-same only if the temporal order of A and Cmade no difference; and group A-unpaired re-ceived equivalent exposure to A and C unpaired,thereby serving as a control for responding basedon the A�B pairings being attenuated by nonas-sociative exposure to A or C during phase 2 (seethe top of Fig. 6). Groups B-same, B-diff andB-unpaired were treated the same as their A coun-terparts, except that in phase 2 A was replaced byB; thus, these three groups assessed competitionbetween cues A and C rather than outcomes B
and C. In phase 3, all subjects were exposed toB-footshock US pairings and finally subjects weretested for lick suppression in the presence of cueA.
Inspection of the mean suppression scores ofexperiment 1 in Fig. 6 reveals that behavioralcontrol by cue A was lower in group A-same thanin group A-unpaired. This difference documentscompetition between outcomes trained apart andfurther that the observed interference was not dueto degraded contingency as a result of the phase 2exposures to A (i.e. A-noB trials) that were re-ceived by group A-same. Additionally, the lowerscore of group A-same relative to group A-diffindicates that the A–C pairings of phase 2 yieldedmore retroactive interference when stimulus Awas maintained in the same temporal position inphase 2 that it had in phase 1 (i.e. changing A toan outcome in phase 2 reduced its potential tointerfere with the phase 1 memory of it serving asa cue). Parallel comparisons of the three B groupsin experiment 1 encourage the same conclusionsabout competition between cues. Moreover, thedifference observed between groups A-same andA-unpaired seems quite similar to that seen be-tween groups B-same and B-unpaired, indicatingthat, at least in this situation, competition be-tween outcomes was comparable in magnitude tocompetition between cues. Similar conclusionsconcerning the comparable magnitude of interfer-ence between cues trained apart, provided theyhave a common outcome, and between outcomestrained apart, provided they have a common cue,were reached by Pineno and Matute (2000) basedon their study of human participants in a behav-ioral (nonverbal) task.
The above comparison of competition betweenoutcomes and between cues might be faulted be-cause B was consistently paired with the US andtesting was consistently performed with A. Thus,the B groups in experiment 1 were exposed to B inall three phases of treatment, whereas the Agroups were not. Additionally, the A groups weretested on a stimulus that was presented in phase2, whereas the B groups were not. To determine ifeither of these confounds might have contributedto our observations, we replicated the study, butthis time in Phase 3 paired A with the footshock

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 175
Fig. 6. Competition between outcomes trained apart and between cues trained apart. For each subject, all training and testingoccurred in the same context. Stimuli A, B and C were all innocuous during phases 1 and 2. ‘/’ separates stimuli that were presentedexplicitly unpaired, and US represents the footshock unconditioned stimulus. The graph presents times to complete five cumulativeseconds of drinking in the presence of CS A (experiment 1) and B (experiment 2). A higher score indicates more control of behavior.For details, see text and Escobar et al. (2002).
US and then we tested on stimulus B. Assessingthe A�B association by reinforcing A and thentesting on B may seem like grounds for pausebecause it constitutes backward conditioning;however, there is a large and growing literature
that under some conditions backward pairingssupport excitatory responding (e.g. Arcediano etal., 2002; Matzel et al., 1988; Pavlov, 1927; Savas-tano and Miller, 1998). Thus, experiment 2 in-verted the confounds of experiment 1. If the

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185176
results of experiment 1 depended on either ofthese confounds, different results should havecome from experiment 2. However, inspection ofthe group mean scores of experiment 2 in Fig. 6indicates that these changes had no appreciableeffect. Consequently, we may conclude that thesepotential confounds did not influence our originalobservations, and competition between outcomesoccurs and is similar in magnitude to competitionbetween cues.
5. Why have modern models of learning ignoredcells 2, 3 and 4?
Collectively, the experiments reviewed abovedemonstrate competition between cues presentedtogether, cues presented apart, outcomes pre-sented together and outcomes presented apart.Moreover, much of these data are not new, butdate back the better part of 40 years. How andwhy did students of learning forget such a largeand relevant literature? Here we can only specu-late. But we would suggest that there were at leastfive reasons beyond the recurring human problemof failing to remember history.
One reason was that the controls used in theolder studies were not always up to modern stan-dards. As we have demonstrated, these interfer-ence effects can be obtained even with appropriatecontrol groups. A second reason is that the mate-rials were verbal items with human subjects, andconsequently might be explained with higher-or-der mental processes rather than basic associativeprocesses that transcend species. However, thepresent studies suggest that these effects can alsobe obtained in Pavlovian situations with nonhu-man subjects. A third reason for this neglect,applicable to at least cells 3 and 4, was a sensethat different mechanisms accounted for thesetypes of interference, relative to cell 1 (cell 2 wasnot well recognized). But recent observations dis-cussed below (Section 6) challenge the notion ofdifferent mechanisms. A fourth reason that cell 3has been neglected may be based on the view thatcue competition is of greater magnitude when thecompeting cues are trained together than whenthey are trained apart. However, rarely do re-
searchers include appropriate controls to properlycompare cell 1-type interference with cell 3-typeinterference. Moreover, researchers ordinarily re-port only studies with tuned parameters that max-imize the type of interference in which they areimmediately interested; there is no reason to thinkthat the conditions that maximize one type ofinterference will also maximize the other type ofinterference. Because of the focus in recentdecades on cell 1-type interference, we better un-derstand the optimal parameters to obtain thistype of interference in the animal laboratory thanwe do to obtain the other types of interference.
A fifth reason that cells 3 and 4 were ignoredlikely had to do with the functionally compellingnature of the informational account provided byKamin (1968) and Rescorla and Wagner (1972).That is, the central value of the ability to learnenvironmental relationships seemingly is to antici-pate impending events so one might either preparefor or even try to alter the nature of these events.In this framework, redundant information aboutimpending events is viewed as being of little func-tional value and deleterious with respect to theputative limited capacity of memory. Hence, it isappealing to hypothesize a learning mechanismthat is sensitive to redundancy and precludeslearning associations about events that, duringtraining, were expected based on simultaneouslypresented cues. Cell 1-type stimulus competition isentirely consistent with this view, whereas thestimulus competition depicted in the other threecells of the matrix in Fig. 1 are not. As a conse-quence of the appeal of the functional viewpoint,cells 3 and 4 came to be ignored and were eventu-ally forgotten by many researchers. However, it isimportant to note that, as its name indicates, thefunctional viewpoint speaks to the function oflearning, not necessarily the underlying process.As biologists have repeatedly found, process doesnot always reflect function. Process is severelylimited by a species’ evolutionary history andrandom factors in sexual recombination and mu-tation. Several phenomena (e.g. priming effects)suggest that subjects encode great amounts of,mostly redundant, information (process); cuecompetition effects reflect the tendency of organ-isms to use only part of this information (func-

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 177
tion). In summary, learning theory appears tohave allowed an implicit concern for the functionof learning to overly influence the empirical exam-ination of basic phenomena. We have erroneouslyassumed that function dictates process.
6. Common or different mechanisms?
Traditionally, cell 1-type stimulus competitionwas regarded as a learning deficit. That is, sub-jects were assumed unable to exhibit the deficientbehavior without new training trials with thetarget cue (i.e. further training; Rescorla andWagner, 1972). In contrast, cell 3- and 4-typecompetition has been viewed largely as a perfor-mance deficit that could be reversed without fur-ther training with the target cue. For example,retroactive interference was known to wane overincreasing retention intervals (e.g. Postman et al.,1968). The corresponding return of behaviorbased on phase 1 training without further trainingsuggested that the original deficit (i.e. retroactiveinterference) was due to a failure to access anassociation that was still encoded within the sub-ject, rather than an irrevocable loss of the phase 1association. The previously described studies byEscobar et al. (2001) demonstrating priming byeither a specific stimulus or a training context alsospeak to this sort of competition being at least inpart a performance deficit. However, this distinc-tion between the mechanisms underlying cell 1-type interference and mechanisms underlying cells3 and 4-type interference does not stand up tocontemporary scrutiny.
Cell 1-type interference (e.g. overshadowingand blocking) in recent years has been found tobe reversible without further training. Manipula-tions that are often found to undo this type ofstimulus competition include, (1) lengthening theretention interval (i.e. spontaneous recovery, e.g.Kraemer et al., 1988), (2) administration of so-called ‘reminder treatments’ which consist of pre-sentation of either the outcome alone, the cuealone, or the training context (e.g. Balaz et al.,1982), and (3) posttraining massive extinction ofthe overshadowing or blocking stimulus (e.g.Blaisdell et al., 1999b; Kaufman and Bolles, 1981;
Matzel et al., 1985). Although these demonstra-tions of recovery from competition between cuestrained together (particularly, extinction of thecompeting cue) were initially viewed as cleardemonstrations of the original response deficitbeing a performance failure (e.g. Miller andMatzel, 1988, Miller and Schachtman, 1985),newer learning models have been formulated thatview recovery from cell 1-type stimulus competi-tion resulting from reminder treatments or extinc-tion of the competing cue as new learning thatoccurs in the absence of the target cue (Dickinsonand Burke, 1996; Van Hamme and Wasserman,1994); however, these models are less able toaccount for spontaneous recovery in terms of newlearning. But how one accounts for recovery fromcell 1-type cue competition without further train-ing with the target cue is a digression from themain point here, which is that the responsedeficits constituting cells 1–4 in Fig. 1 have nowall been seen to be reversible without furthertraining with the target stimulus (see Miller et al.,1986; Spear, 1976). Although researchers may stillbe arguing over theories, the basic recovery frominterference phenomenon appears to be obtain-able in each cell of the matrix. Of course, thereare many other phenomena besides recovery frominterference on which the behavioral deficits rep-resented in each cell could and should be com-pared. A challenge for the future is thatresearchers make further comparisons betweenthese different types of stimulus competition tosee if the same phenomena can be demonstratedfor each cell. If they can, this would argue for acommon account of interference in all cells; ifthey cannot, this would encourage the develop-ment of different models for different cells of thematrix. But until such time as clear differences inthe phenomena obtainable for each cell of thematrix is documented, the rule of parsimony callsfor learning models to attempt to explain all fourtypes of competition with a single mechanism.
7. A larger perspective
Above we have focused on stimulus interactionto the extent that it interferes with behavioral

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185178
control by a target stimulus (i.e. an inverse rela-tionship exists between the behavioral control ofthe target stimulus and the interacting [compet-ing] stimulus). However, stimulus interaction alsoincludes the potential for facilitation of behav-ioral control by the interacting stimulus (i.e. adirect relationship exists between behavioral con-trol of the target stimulus and the interacting[facilitating] stimulus). Examples of stimulus fa-cilitation between cues trained in compound areprovided by potentiation (Rusiniak et al., 1979)and augmentation (Batson and Batsell, 2000),which are [arguably] the same treatments asovershadowing and blocking, respectively, but re-sult in enhanced rather than attenuated respond-ing to the target cue. Second-order conditioning(Pavlov, 1927), sensory preconditioning (Brog-den, 1939), and mediated extinction (Hollandand Forbes, 1982) provide examples of stimulusfacilitation between stimuli trained apart. Wesuggest that interference and facilitation togethermight best be called stimulus interaction, whichwould refer collectively to all situations in whichthe associative status of one stimulus, C, influ-ences responding based upon an association be-tween two other stimuli (A–B).
Stimulus facilitation has received much lessattention in recent years than has stimulusinterference (including competition), and corre-spondingly far fewer models have been proposedto account for stimulus facilitation (but see Hol-land, 1990). Surely, there must be principledways of determining whether interference or fa-cilitation will be observed, but currently fewmodels make clear predictions concerning theconditions under which one or the other will beobserved (but see McLaren and Mackintosh,2000). Suggested critical variables include stimu-lus similarity (Osgood, 1949), number of trainingtrials (e.g. Yin et al., 1994), whether the elementsin training are presented simultaneously or seri-ally (e.g. Dwyer, 2001), and whether the trainingsituation favors within-compound associationsbeing formed between the interacting stimuli (e.g.Aitken et al., 2001). However, there is muchresearch left to be done before we fully under-stand this issue. A detailed discussion of facilita-tion is beyond the domain of this article. We
mention facilitation here only because it wouldbe misleading to imply that interference is theonly type of stimulus interaction. Facilitation isan important facet of stimulus interaction thatideally some day will be explained by a modelcapable of accounting for both interference andfacilitation including the conditions under whicheach is observed.
To obtain any of these stimulus interactioneffects, interference or facilitation, one needs ap-propriate parameters. These interaction effectsare not as ubiquitous as many textbooks imply.For example, in our initial attempts to obtainblocking, we had to run many pilot studies untilwe identified parameters that consistently re-sulted in blocking. Research toward better identi-fying these parameters surely would help usunderstand the mechanisms underlying stimulusinteraction. For example, [forward] blocking hasbeen found to wane with large numbers of phase2 trials (i.e. AX�US; Azorlosa and Cicala,1988). This rarely cited observation is damagingto models of learning that account for learningin terms of the cue added in phase 2 (theblocked CS) being redundant with the one origi-nally trained during phase 1 (the blocking CS);that is, in an informational framework there isno basis for expecting the added cue to gainbehavioral control if phase 1 training wasasymptotic.
8. A tentative model
First, let us summarize the empirical phe-nomenon that we wish to explain. Organismslearn many relationships between stimuli duringtheir lives which seemingly do not interfere witheach other. Thus, it is not surprising that asso-ciative interference, to the extent that it occurs atall, appears to be limited largely to situations inwhich the two competing associations have acommon element, specifically a common outcomein the cases of cells 1 and 3 and a common cuein the cases of cells 2 and 4 (see Fig. 1). In cells1 and 2 (i.e. competition between stimuli trainedtogether), the requirement of a common elementis largely met automatically by the competing

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 179
elements being trained together.2 This is not thecase with cells 3 and 4 (i.e. competition betweenstimuli presented apart), in which there is agreater probability of differences between in-stances of the potentially common element thatare paired with each of the two competing stimuli.Here, the necessity of an element common to thetwo associations leads to the expectation that anychange in the ‘common’ element between the twophases of training in cells 3 and 4 will attenuateinterference. Consistent with this, stimulus inter-ference appears to be greatest when two associa-tions have one element in common (not twowhich makes phase 2 equivalent to more trainingtrials with the same elements), with ‘common’here including the temporal (and likely spatial)positions of the competing stimuli relative to thecommon element (Escobar et al., 2002; but seeKeppel et al., 1971).
Contemporary models (e.g. Gallistel and Gib-bon, 2000; Mackintosh, 1975; McLaren andMackintosh, 2000; Miller and Matzel, 1988;Pearce, 1987; Pearce and Hall, 1980; Rescorla andWagner, 1972) were designed largely to accountfor competition between cues predicting a com-mon occurrence of a single outcome (cell 1). Mostof these models view competition between cuestrained together as a failure to acquire an associa-tion between the target cue and the outcome (e.g.Rescorla and Wagner, 1972), but some of themregard such competition as a failure to retrieve thetarget cue–outcome association (e.g. Denniston etal., 2001). Regardless of their orientation, thesemodels fail to account for other types of associa-tive competition, such as situations in which there
are competing outcomes or the competing stimuliare not presented in compound. Addressing cell 4alone, Bouton (e.g. 1993) proposed a retrieval-fo-cused model which posits that, in situations inwhich a cue is associated to multiple outcomes,contextual or punctate stimuli present at the timeof testing permit subjects to select which one ofthe competing outcomes will be retrieved to im-mediately influence behavior. In Bouton’s frame-work, retrieval of a specific association isfacilitated by those cues that were present at thetime in which that association was trained. Thismodel has proven efficient in accounting for manyforms of cell 4 interference, but it is unable toaccount for competition between outcomes thatare trained in compound (cell 2), much less anysort of competition between cues (cells 1 and 3).For example, competition between cues trainedapart (cell 3-type interference) is problematic tothis model because the competing stimuli in thiscase are cues, each of which should be able toretrieve representations of the common outcomewithout interacting with one another because eachcue is part of but one association. More generally,exclusive of contingency effects (which have beendemonstrated to be insufficient to account formany apparent interference effects), the existingmodels are all fundamentally unable to explainassociative interference represented by more thanone cell of the matrix in Fig. 1.
The failure of contemporary models of learningto explain more than one cell of the interferencematrix depicted in Fig. 1 suggests that new modelsrequire either different or additional mecha-nism(s) to address all four cells of the matrix. Inseeking such a model, we were greatly influencedby the apparent reversibility of the interferencedeficits in each of the four cells of the matrix (seethe preceding discussion within Section 6), whichsuggests that interference effects have an impactmostly at the retrieval level. We propose thatinterference between cues and outcomes trainedtogether and apart could be accounted for by adual-process ‘retrieval’ model that combines avariation of Denniston et al.’s (2001) comparatorhypothesis (also see Miller and Matzel, 1988),with an extrapolation of Bouton’s (1993) retrievalmodel by Escobar et al. (2001, 2002) and Pineno
2 Even in the case of stimuli trained together, however, thecommon element may have a different relationship with thecompeting stimuli. One such case are situations in which thereare discrepancies in the temporal relationship of the commonelement to the two competing elements. For example, Barnetet al. (1993) and Blaisdell et al. (1997) observed that blockingand overshadowing, respectively, decreased when the temporalrelationships of the two competing cues with respect to theoutcome differed. Barnet et al. changed the temporal relation-ship of the blocking CS with respect to the outcome betweenphases 1 and 2 of the blocking procedure, whereas Blaisdell etal. changed the temporal relationship of the overshadowing CSto the outcome between overshadowing treatment and testing.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185180
and Matute (2000). Importantly, the mechanismsof each of these two processes apply to all fourcells of the matrix but, because of the proce-dures defining each cell, one of the two pro-cesses is moot under the procedures in whichthe other is engaged: the particular associativestructures make one of the two processes ineffi-cient in each cell.
8.1. Process 1: comparator process
We assume that this process, an extension ofthe comparator hypothesis (Denniston et al.,2001; Miller and Matzel, 1988), prevails in cell1- and cell 2-type interference. The comparatorhypothesis’ account of cell 1-type interference(i.e. ‘cue competition’) has been described manytimes previously. To summarize, it asserts thatretrieval of the target CS–outcome associationis a positive function of the strength of thatassociation and, importantly, also a negativefunction of the product of (1) the strength ofthe association(s) between the target cue andother cues present during training (i.e. compara-tor [competing] stimuli) and (2) the strength ofthe association(s) between each of these ‘com-parator’ stimuli and the outcome. Consistentwith previously mentioned observations, this lat-ter (indirectly) accessed representation of theoutcome is assumed to attenuate retrieval of thetarget association most effectively when the tworepresentations of the common outcome (one di-rectly addressed by the target CS and the otherindirectly addressed by the target CS throughcomparator stimuli) are identical, and less effec-tively as they are made more dissimilar either inconventional physical attributes (e.g. Blaisdell etal., 1997; but see e.g. Ganesan and Pearce,1988) or the temporal relationship of the com-mon outcome to the competing cues (e.g. Blais-dell et al., 1999a). Hence, the effectiveness ofthe target cue in retrieving a representation ofthe outcome is relative to that of other cues thatwere present during training of the target cue-outcome association.
We now assert that this account can explaincell 2-type interference (i.e. competition betweenoutcomes trained together), as well as cell 1-type
interference, if we treat the nontarget outcomeas a potential comparator stimulus for the targetcue. Note that the distinction between cues andoutcomes is more likely in the mind of the ex-perimenter than the subject. As Goddard (1999),among others, has demonstrated, outcomes caneffectively signal impending events (i.e. otheroutcomes). The comparator hypothesis assumesthat effective comparator stimuli are all salientstimuli, other than the target CS and the US,that are present during training of the targetCS. During cell 2 treatment, the target cue isnot only being paired with the target outcome,it is being paired with the nontarget outcome,which in turn is also being paired with thetarget outcome. Thus, at test, the cue not onlyhas access to its direct association to the targetoutcome, but also the cue-nontarget outcome as-sociation and hence indirectly to the nontargetoutcome-target outcome association. This latter(indirect) associative pathway presumably atten-uates activation based on the [direct] target as-sociation. To the subject, the nontarget outcomeof cell 2 plays a role almost identical to thenontarget cue of cell 1. However, the conven-tional temporal relationships are slightly differ-ent in that in cell 2 there is typically a serialrelationship between the cue and the competingelement (i.e. nontarget outcome), whereas in cell1 there is ordinarily a simultaneous relationshipbetween the target cue and the competing ele-ment (i.e. nontarget cue). Notably, for each cell,when these two temporal relationships are inte-grated to yield a temporal relationship for theindirect (mediated) association between thetarget cue and the target outcome, the targetcue has the same net temporal relationship tothe outcome as obtains for the direct associationbetween the target cue and target outcome. Theproposed comparator process in principle appliesto situations depicted by all four cells of thematrix, but its action is minimal with respect tocells 3 and 4 because the association betweenthe target cue and interfering cue in cell 3, andthe association between the target outcome andthe nontarget outcome in cell 4 should be weakor nonexistent.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 181
8.2. Process 2: priming process
We assume that this process prevails in cell 3-and cell 4-type interference. We propose that be-havior indicative of interference will be observedwhen two associations have a common element(cue or outcome) and the nontarget (i.e. interfer-ing) association is better primed for retrieval thanis the target association. In this framework, apriming stimulus does not itself activate the repre-sentation of a stimulus stored in memory (unless itindependently has excitatory potential, which isoften the case); rather it increases the ease withwhich one physical stimulus (a cue) can activatethe representation of another stimulus (an out-come), after pairings of the cue and outcome in thepresence of the priming stimulus (i.e. positivepriming; for a similar view, see Neely, 1977). Anexample of this priming function is contextualmodulation of conditioned responding. Bringing asubject back to the context in which a cue–out-come association was previously learned does notresult in the immediate elicitation of a conditionedresponse. However, if the cue is presented in thatcontext, conditioned responding often will morelikely be observed than if the cue was presented inanother context. Importantly, in our view, a prim-ing stimulus not only enhances the retrievability ofrepresentations of stimuli trained in its presencebut, at the same time, it decreases the retrievabilityof (i.e. it negatively primes) any other stimulusrepresentation common to either of the two stim-uli constituting the positively primed association.A typical example of this sort of priming is theso-called renewal effect (Bouton and Bolles, 1979),in which testing an extinguished cue in the contextof acquisition results in renewed responding (i.e. itprimes the cue-outcome experience) and decreasesthe likelihood of retrieval of the cue-no outcome
experience.3 It is the addition of this primingmechanism that accounts for the stimulus modula-tion of interference (proactive vs. retroactive) incells 3 and 4 of the interference matrix. Thismechanism has much in common with the cogni-tive construct of retrieval inhibition (e.g. Bjork,1989), which refers to decreased accessibility ofcertain items that are in fact stored in memory.(The concept of retrieval inhibition is not to beconfused with the concept of conditioned inhibi-tion, which refers to the presumed associativevalue of a stimulus that signals the omission of anexpected outcome.) The role of priming cues forinterference within cells 1 and 2 is minimizedbecause the two competing associations in thesecases are trained together (i.e. simultaneously) andconsequently are ordinarily subject to the samepriming cues. Thus, although priming cues mayinfluence retrieval of associations, they should nottend to differentially favor one association overthe other when the two associations were trainedtogether as is the case in cells 1 and 2.
As previously discussed, Bouton (1993) insight-fully used the priming construct, albeit withoutemphasizing that name, to account for the contex-tual dependency of extinction (i.e. renewal), coun-terconditioning, and the CS-preexposure effect(i.e. cell 4). In addition to Bouton’s work withpriming by contexts, Holland (1992; also seeMiller and Oberling, 1998) has demonstrated ingreat detail how discrete cues, which he calls‘occasion setters,’ can play a similar role as prim-ing stimuli. However, Bouton’s and Holland’saccounts are limited to cell 4 of the Fig. 1 matrix(i.e. priming stimuli would disambiguate the valueof a cue trained with different outcomes). Here wemake that account more general by proposing thatin any situation in which a retrieval cue (be itdiscrete or contextual) is presented, the degree towhich the primed association will control behaviorwill depend on the degree to which that retrievalcue primes any other association that has a com-mon element with the target association. Alterna-tively stated, a stimulus (discrete or contextual)that [positively] primes one association automati-cally negatively primes all other associations thatshare a stimulus representation (i.e. cue or out-come) with the target association.
3 These assumptions give rise to the Jabberwockian state-ment that, in addition to the need for a retrieval cue for thetarget association, retrieval is a function of the strength of thetarget association modulated by its prime (if present), minusthe strength of any competing association modulated by itsprime (if present), minus the degree to which the target primefacilitates retrieval of any competing association, plus thedegree to which any competing prime facilitates retrieval of thetarget association.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185182
In order to explain the effects upon cells 3 and4-type interference of switching the temporal or-der of stimuli between training phases (Escobar etal., 2002), as previously proposed for the com-parator mechanism, we continue here to assumethat a stimulus representation encodes not onlythe traditional physical attributes of the stimulus,but also the spatiotemporal location of the stimu-lus with respect to its associate (see Savastano andMiller, 1998, for the rationale of this assumption).Thus, we consider that the reduced interferenceobserved when the position of the common ele-ment between is switched between the two phasesof training is a consequence of a decrease in thesimilarity of the so-called common element be-tween phases, that is, the common element is lesscommon. In our view, positively priming oneassociation negatively primes a second associationonly if the two associations share a common cueor outcome. Temporal shifting of the order ofelements decreases the commonality of the com-mon element, thereby reducing interference.
Toward placing this priming mechanism in his-torical context, we note that McGeoch (1932)suggested that the most strongly activated associ-ation at the moment of retrieval would be ex-pressed at the expense of other associationssharing some degree of similarity with the acti-vated association. Moreover, he suggested thatbackground (contextual) stimuli are responsiblefor the activation of one or another association(i.e. priming). Congruent with this view, Slameckaand Ceraso (1960) reviewed the literature as of1960 and concluded that greater training on oneassociation increased the likelihood that it wouldinterfere with other associations and would not besubject to interference itself. We (Escobar et al.,2001) have suggested that this observation canaccount for failures to observe interference withstimuli that have inherent or acquired biologicalsignificance, provided one assumes that degree oftraining and biological significance increase thestrength of the association. We can incorporatethese observations with the highly plausible as-sumption that, orthogonal to priming and com-parator processes, strength of an associationincreases its retrievability.
The model presented here lacks parsimony inthat it involves separate mechanisms to explaininterference between stimuli trained together andinterference between stimuli trained apart. How-ever, this is presumably just a first step toward anintegration of the different types of interferencewithin a common framework. At least, here wehave provided an account of all four types ofinterference with only two mechanisms. More-over, these two mechanisms both apply to all fourtypes of interference; only the procedures in twoof the four situations render one process ineffec-tive and the procedures in the other two situationsrender the other process ineffective.
9. Conclusions
Associative competition between events is notlimited to antecedent events trained together. Italso occurs between antecedent events trained sep-arately, as well as between subsequent eventstrained together and apart. Competition betweensubsequent events challenges associative modelsthat emphasize predictive value. Associative com-petition between events trained apart challengesassociative models that account for associativecompetition in a framework that requires simulta-neous activation of the representations of thecompeting stimuli. All four types of associativeinterference (cells 1–4 of the matrix in Fig. 1) areparameter dependent, and all are likely mirroredby associative facilitation under appropriate con-ditions. Stimulus competition/interference appearsto be a much more general phenomenon than iscommonly recognized. Whether the differentforms of stimulus interference arise from a com-mon underlying mechanism is not yet clear. Butherein we propose the outline of a model thataddresses all four types of interference. Thismodel attributes interference to a combination of(1) a comparator mechanism in which the re-trieval of associations is attenuated by the exis-tence of other associations to the elements of thetarget association (an extension of Miller andMatzel, 1988; also see Denniston et al., 2001), and(2) a priming mechanism in which priming stimulinot only positively prime associations acquired in

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 183
their presence, but negatively prime all other asso-ciations (an extension of Bouton, 1993 and Mc-Geoch, 1932). Due to procedural differencesbetween the different cells of the interference ma-trix, the comparator mechanism has its greatestinfluence on competition between stimuli (cues oroutcomes) trained together (i.e. cells 1 and 2), andthe priming mechanism has its greatest influenceon competition between elements (cues or out-comes) that are trained apart (i.e. cells 3 and 4).
Acknowledgements
Support for the preparation of this manuscriptwas provided by NIMH grant 33881. We thankJeffrey Amundson, Francisco Arcediano, Ray-mond Chang, Peter Killeen and Steven Stout fortheir comments on an earlier version of themanuscript.
References
Aitken, M.R.F., Larkin, M.J.W., Dickinson, A., 2001. Re-ex-amination of the role of within-compound associations inthe retrospective revaluation of causal judgments. Q. J.Exp. Psychol. 54B, 27–51.
Arcediano, F., Escobar, M., Miller, R.R., 2002. Temporalintegration and temporal backward associations in humanand nonhuman subjects, submitted for publication.
Azorlosa, J.L., Cicala, G.A., 1988. Increased conditioning inrats to a blocked CS after the first compound trial. Bull.Psychon. Soc. 26, 254–257.
Balaz, M.A., Gutsin, P., Cacheiro, H., Miller, R.R., 1982.Blocking as a retrieval failure: reactivation of associationsto a blocked stimulus. Q. J. Exp. Psychol. 34B, 99–113.
Barnet, R.C., Grahame, N.J., Miller, R.R., 1993. Temporalencoding as a determinant of blocking. J. Exp. Psychol.Anim. Behav. Process. 19, 327–341.
Batson, J.D., Batsell, W.R. Jr, 2000. Augmentation, not block-ing, in an A+/AX+ flavor-conditioning procedure. Psy-chon. Bull. Rev. 7, 466–471.
Bjork, R.A., 1989. Retrieval inhibition as an adaptive mecha-nism in human memory. In: Roediger, H.L. III, Craik,F.I.M. (Eds.), Varieties of Memory and Consciousness:Essays in Honour of Endel Tulving. Lawrence Erlbaum,Hillsdale, NJ, pp. 309–330.
Blaisdell, A.P., Denniston, J.C., Miller, R.R., 1997. Unblock-ing with qualitative change of unconditioned stimulus.Learn. Motiv. 28, 268–279.
Blaisdell, A.P., Denniston, J.C., Miller, R.R., 1999a. Post-training shifting of the overshadowing stimulus-US interval
alleviates the overshadowing deficit. J. Exp. Psychol.Anim. Behav. Process. 25, 18–27.
Blaisdell, A.P., Gunther, L.M., Miller, R.R., 1999b. Recoveryfrom blocking achieved by extinguishing the blocking CS.Anim. Learn. Behav. 27, 63–76.
Bouton, M.E., 1993. Context, time, and memory retrieval inthe interference paradigms of Pavlovian learning. Psychol.Bull. 114, 80–99.
Bouton, M.E., Bolles, R.C., 1979. Contextual control of theextinction of conditioned fear. Learn. Motiv. 10, 445–466.
Brogden, W.J., 1939. Sensory pre-conditioning. J. Exp. Psy-chol. 25, 323–332.
Brooks, D.C., Bouton, M.E., 1993. A retrieval cue for extinc-tion attenuates spontaneous recovery. J. Exp. Psychol.Anim. Behav. Process. 19, 77–89.
Burger, D.C., Mallemat, H., Miller, R.R., 2000. Overshadow-ing of subsequent events and recovery thereafter. Q. J.Exp. Psychol. 53B, 149–171.
Dearing, M.F., Dickinson, A., 1979. Counterconditioning ofshock by a water reinforcer in rabbits. Anim. Learn.Behav. 7, 360–366.
Denniston, J.C., Miller, R.R., Matute, H., 1996. Biologicalsignificance as a determinant of cue competition. Psychol.Sci. 7, 325–331.
Denniston, J.C., Savastano, H.I., Miller, R.R., 2001. Theextended comparator hypothesis: learning by contiguity,responding by relative strength. In: Mowrer, R.R., Klein,S.B. (Eds.), Handbook of Contemporary Learning Theo-ries. Erlbaum, Hillsdale, NJ, pp. 65–117.
Dickinson, A., Burke, J., 1996. Within-compound associationsmediate the retrospective revaluation of causality judg-ments. Q. J. Exp. Psychol. 49B, 60–80.
Donahoe, J.W., Palmer, D.C., 1994. Learning and ComplexBehavior. Allyn and Bacon, Needham Heights, MA.
Dwyer, D.M., 2001. Mediated conditioning and retrospectiverevaluation with LiCl then flavour pairings. Q. J. Exp.Psychol. 54B, 145–165.
Escobar, M., Arcediano, F., Miller, R.R., 2002. Conditionsfavoring retroactive interference between antecedent eventsand between subsequent events. Psychon. Bull. Rev., 8,691–697.
Escobar, M., Matute, H., Miller, R.R., 2001. Cues trainedapart compete for behavioral control in rats: convergencewith the associative interference literature. J. Exp. Psychol.Gen. 130, 97–115.
Esmoriz-Arranz, F.J., Miller, R.R., Matute, H., 1997. Block-ing of subsequent and antecedent events. J. Exp. Psychol.Anim. Behav. Process. 23, 145–156.
Gallistel, C.R., Gibbon, J., 2000. Time, rate and conditioning.Psychol. Rev. 107, 219–275.
Ganesan, R., Pearce, J.M., 1988. Effect of changing the un-conditioned stimulus on appetitive blocking. J. Exp. Psy-chol. Anim. Behav. Process. 14, 280–291.
Goddard, M.J., 1999. The role of US signal value in contin-gency, drug conditioning, and learned helplessness. Psy-chon. Bull. Rev. 6, 412–423.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185184
Holland, P.C., 1992. Occasion setting in Pavlovian conditioning.In: Medin, D.L. (Ed.), The Psychology of Learning andMotivation, vol. 28. Academic Press, San Diego, CA, pp.69–125.
Holland, P.C., Forbes, D.T., 1982. Representation-mediatedextinction of conditioned flavor aversions. Learn. Motiv. 13,454–471.
Holland, P.C., 1990. Forms of memory in Pavlovian condition-ing. In: McGaugh, J.L., Weinberger, N.M., Lynch, G.(Eds.), Brain Organization and Memory: Cells, Systems andCircuits. Oxford University Press, New York, pp. 78–105.
Jung, J., 1963. Effects of response meaningfulness (m) ontransfer of training under two different paradigms. J. Exp.Psychol. 65, 377–384.
Kamin, L.J., 1968. Attention-like’ processes in classical condi-tioning. In: Jones, M.R. (Ed.), Miami Symposium on thePrediction of Behavior: Aversive Stimulation. University ofMiami Press, Miami, FL, pp. 9–33.
Kaufman, M.A., Bolles, R.C., 1981. A nonassociative aspect ofovershadowing. Bull. Psychon. Soc. 18, 318–320.
Keppel, G., Bonge, D., Strand, B.Z., Paker, J., 1971. Direct andindirect interference in the recall of paired associates. J. Exp.Psychol. 88, 414–422.
Kraemer, P.J., Lariviere, N.A., Spear, N.E., 1988. Expressionof a taste aversion conditioned with an odor– taste com-pound: overshadowing is relatively weak in weanlings anddecreases over a retention interval in adults. Anim. Learn.Behav. 16, 164–168.
Lashley, K.S., 1942. An examination of the continuity theoryas applied to discriminative learning. J. Gen. Psychol. 26,241–265.
Mackintosh, N.J., 1975. A theory of attention: variations in theassociability of stimuli with reinforcement. Psychol. Rev. 82,276–298.
Matute, H., Pineno, O., 1998. Stimulus competition in theabsence of compound conditioning. Anim. Learn. Behav. 26,3–14.
Matzel, L.D., Schachtman, T.R., Miller, R.R., 1985. Recoveryof an overshadowed association achieved by extinction ofthe overshadowed stimulus. Learn. Motiv. 16, 398–412.
Matzel, L.D., Held, F.P., Miller, R.R., 1988. Information andexpression of simultaneous and backward associations:implications for contiguity theory. Learn. Motiv. 19, 317–344.
McGeoch, J.A., 1932. Forgetting and the law of disuse. Psychol.Rev. 39, 352–370.
McLaren, I.P.L., Mackintosh, N.J., 2000. An elemental modelof associative learning. I. Latent inhibition and perceptuallearning. Anim. Learn. Behav. 28, 211–246.
Miller, R.R., Matute, H., 1998. Biological significance inforward and backward blocking: resolution of a discrepancybetween animal conditioning and human causal judgment.J. Exp. Psychol. Gen. 125, 370–386.
Miller, R.R., Matzel, L.D., 1988. The comparator hypothesis:A response rule for the expression of associations. In: Bower,G.H. (Ed.), The Psychology of Learning and Motivation,vol. 22. Academic Press, San Diego, CA, pp. 51–92.
Miller, R.R., Oberling, P., 1998. Analogies between occasion
setting and Pavlovian conditioning. In: Schmajuk, N.A.,Holland, P.C. (Eds.), Occasion Setting: Associative Learn-ing and Cognition in Animals. American PsychologicalAssociation, Washington, DC, pp. 3–35.
Miller, R.R., Schachtman, T.R., 1985. Conditioning context asan associative baseline: implications for response generationand the nature of conditioned inhibition. In: Miller, R.R.,Spear, N.E. (Eds.), Information Processing in Animals:Conditioned Inhibition. Lawrence Erlbaum, Hillsdale, NJ,pp. 51–88.
Miller, R.R., Kasprow, W.J., Schachtman, T.R., 1986. Retrievalvariability: sources and consequences. Am. J. Psychol. 99,145–218.
Neely, J.H., 1977. Semantic priming and retrieval from lexicalmemory: roles of inhibitionless spreading activation andlimited-capacity attention. J. Exp. Psychol. Gen. 106, 226–254.
Oberling, P., Bristol, A.S., Matute, H., Miller, R.R., 2000.Biological significance attenuates overshadowing, relativevalidity, and degraded contingency effects. Anim. Learn.Behav. 28, 172–186.
Osgood, C.E., 1949. The similarity paradox in human learning:a resolution. Psychol. Rev. 56, 132–143.
Pavlov, I.P., 1927. Conditioned Reflexes. Oxford UniversityPress, London.
Pearce, J.M., 1987. A model for stimulus generalization inPavlovian conditioning. Psychol. Rev. 94, 61–73.
Pearce, J.M., Hall, G., 1980. A model for Pavlovian learning:variations in the effectiveness of conditioned but not ofunconditioned stimuli. Psychol. Rev. 87, 532–552.
Pineno, O., Matute, H., 2000. Interference in human predictivelearning when associations share a common element. Int. J.Comp. Psychol. 13, 16–33.
Postman, L., 1962. Transfer of training as a function ofexperimental paradigm and degree of first-list learning. J.Verbal Learn. Verbal Behav. 1, 109–118.
Postman, L., Stark, K., Fraser, J., 1968. Temporal changes ininterference. J. Verbal Learn. Verbal Behav. 7, 672–694.
Rescorla, R.A., 1970. Reduction in the effectiveness of rein-forcement after prior excitatory conditioning. Learn. Motiv.1, 372–381.
Rescorla, R.A., 1980. Pavlovian Second-Order Conditioning.Lawrence Erlbaum, Hillsdale, NJ.
Rescorla, R.A., 1991. Associations of multiple outcomes withan instrumental response. J. Exp. Psychol. Anim. Behav.Process. 17, 465–474.
Rescorla, R.A., Wagner, A.R., 1972. A theory of Pavlovianconditioning: variations in the effectiveness of reinforcementand non-reinforcement. In: Black, A.H., Prokasy, W.F.(Eds.), Classical Conditioning II: Current Theory and Re-search. Appleton-Century Crofts, New York, pp. 64–99.
Rusiniak, K.W., Hankins, W.G., Garcia, J., Brett, L.P., 1979.Flavor-illness aversions: potentiation of odor by taste in rats.Behav. Neural Biol. 25, 1–17.
Savastano, H.I., Miller, R.R., 1998. Time as content in Pavlo-vian conditioning. Behav. Process. 44, 147–162.

R.R. Miller, M. Escobar / Beha�ioural Processes 57 (2002) 163–185 185
Slamecka, N.J., Ceraso, J., 1960. Retroactive and proactiveinhibition of verbal learning. Psychol. Bull. 57, 449–475.
Spear, N.E., 1976. Retrieval of memories: a psychobiologicalapproach. In: Estes, W.K. (Ed.), Handbook of Learning andCognitive Processes: Attention and Memory, vol. 4. Erl-baum, Hillsdale, NJ, pp. 17–90.
Van Hamme, L.J., Wasserman, E.A., 1994. Cue competition incausality judgments: the role of nonpresentation of com-
pound stimulus elements. Learn. Motiv. 25, 127–151.Wagner, A.R., Logan, F.A., Haberlandt, K., Price, T., 1968.
Stimulus selection in animal discrimination learning. J. Exp.Psychol. 76, 171–180.
Yin, H., Barnet, R.C., Miller, R.R., 1994. Second-order condi-tioning and Pavlovian conditioned inhibition: operationalsimilarities and differences. J. Exp. Psychol. Anim. Behav.Process. 20, 419–428.




















