
Excavations at Utica by the Tunisian-British Utica Project 2014
Upload
independentCategory
view
4download
0
1 23
Molecular Biology ReportsAn International Journal on Molecularand Cellular Biology ISSN 0301-4851 Mol Biol RepDOI 10.1007/s11033-014-3117-6
Association of CYP1A1 and CYP2D6 genepolymorphisms with head and neck cancerin Tunisian patients
Rim Khlifi, Amine Chakroun, AmelHamza-Chaffai & Ahmed Rebai

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Association of CYP1A1 and CYP2D6 gene polymorphismswith head and neck cancer in Tunisian patients
Rim Khlifi • Amine Chakroun • Amel Hamza-Chaffai •
Ahmed Rebai
Received: 8 March 2013 / Accepted: 10 January 2014
� Springer Science+Business Media Dordrecht 2014
Abstract The purpose of this study was to investigate the
relationship between head and neck cancer (HNC) and
environmental agents and polymorphisms in CYP1A1,
CYP2D6, NAT1 and NAT2 metabolic enzymes genes. To
the best of our knowledge, this is the first report on poly-
morphisms in CYP1A1 6310C[T, CYP2D6 Arg365His,
NAT1 52936A[T and NAT2 Arg268Lys (NAT2*12A)
genes and susceptibility to HNC in Tunisian population.
We study the prevalence of these polymorphisms in 169
patients with HNC and 261 control subjects using poly-
merase chain reaction based methods in a Tunisian popu-
lation. We detected an association between HNC and
CYP1A1 6310C[T (TT) and CYP2D6 Arg365His (His/His)
variant carriers (OR 1.75, P = 0.008 and OR 1.66,
P = 0.016, respectively). No association was found
between the polymorphisms genotypes of NAT1
52936T[A and NAT2 Arg268Lys and risk of HNC. An
association between HNC and CYP1A1 (TT) genotype was
found among patients with smoking (P = 0.011) and
drinking habit (P = 0.009). The combinations of NAT1
(AT or AA) and NAT2 (AA) at-risk genotypes increased
HNC risk (OR 4.23, P = 0.005 and OR 3.60, P = 0.048,
respectively). However, the combinations of CYP1A1 (AA)
and CYP2D6 (CC) genotypes decreased risk of HNC (OR
0.20; P = 0.006). Genetic polymorphisms in CYP1A1 and
CYP2D6 may significantly associate with HNC in the
Tunisian population. The results of this study suggest a
possible gene–environment interaction for certain carcin-
ogen metabolizing enzymes, but larger studies that fully
evaluate the interaction are needed.
Keywords Head and neck cancer � CYP1A1 � CYP2D6 �NAT1 � NAT2 � Genetic polymorphism � Risk factors
Abbreviations
CYP450 Cytochrome P450
CI Confidence interval
CY Consumption-year
HNC Head and neck cancer
LC Laryngeal cancer
NPC Nasopharyngeal
cancer
NAT N-Acetyl-transferase
OR Odds ratio
PY Pack-years
PCR Polymerase chain
reaction
RFLP Restriction fragment
length polymorphism
Introduction
Head and neck cancer (HNC) is a complex disease resulting
from both gene–gene and gene–environment interactions
[1]. Increasing epidemiological studies indicate that these
cancers are strongly linked to environmental factors such as
chemical carcinogens present in tobacco and alcohol [2, 3].
R. Khlifi (&) � A. Hamza-Chaffai
Unit of Marine and Environmental Toxicology, UR 09-03,
IPEIS, Sfax University, BP 1172, 3018 Sfax, Tunisia
e-mail: [email protected]
R. Khlifi � A. Rebai
Bioinformatics and Signalling Group, Centre de Biotechnologie
de Sfax, PO Box 1177, 3018 Sfax, Tunisia
A. Chakroun
Department of Otorhinolaryngology, Habib BOURGUIBA
Hospital, Sfax, Tunisia
123
Mol Biol Rep
DOI 10.1007/s11033-014-3117-6
Author's personal copy

Metabolic enzymes that are potentially involved in either the
activation or detoxication of chemical carcinogens have
received a great deal of attention recently as possible genetic
susceptibility factors for a variety of cancers [4–7].
Cytochrome P450s (CYP450) enzymes play a key role in
the metabolism of drugs and environmental chemicals [8–
11]. Among these cytochrome P450s, two important bio-
transformation enzymes, cytochrome P450 1A1 (CYP1A1)
and cytochrome P450 2D6 (CYP2D6), involved in the
metabolism of carcinogens. The CYP2D6 and CYP1A1
genes are localized on chromosome 22q13.1 and 15q22-24,
respectively. A large numbers of epidemiological studies
have since been performed examining the potential associ-
ation between HNC and the genetic variants of CYP1A1 [12–
18] and CYP2D6 [19–21]. However, no data were available
in the literature regarding the association between CYP2D6
Arg365His (rs1058172) variant and cancer risk. In the case
of CYP1A1 6310C[T (rs4646421) variant, there are a very
limited number of studies available in the literature, which
have investigated the role of CYP1A1 6310C[T polymor-
phism in the gene–environment interaction and the risk of
cancer development [11, 22].
The NAT genes are located on chromosome 8p21.3-23.1
and express two highly polymorphic isoenzymes, NAT1
and NAT2 with distinct functional roles. Thirty-six NAT2
genetic variants have been identified in human, of which
NAT2*12A is classified as rapid allele [23]. Four relatively
common polymorphic alleles exist for NAT1 with NAT1*4
being the most common allele [23]. Genetic variants of
both genes have been associated with increased risk of
cancer by either reduced capacity to detoxify aromatic
amine carcinogens or by increased capacity to produce
higher levels of reactive metabolites [24–29]. To date there
has been limited investigation of the role of the NAT gene
variants and conflicting data address their association with
HNC [20, 30–35].
In this case–control study, we examined the associa-
tion of CYP1A1 6310C[T, CYP2D6 Arg365His, NAT1
A52936T and NAT2 Arg268Lys (NAT2*12A) polymor-
phisms with risk of HNC in Tunisian population. Potential
gene–environment interactions were analyzed between
these polymorphisms and the risk factors in the etiology of
HNC. We also studied the effects of gene–gene interactions
of the carcinogen metabolizing-related genes in association
with HNC risk.
Materials and methods
Study subjects
Patients with pathologically proven HNC selected (n = 169)
were recruited from the department of Otorhinolaryngology
of the Sfax, University Hospital Habib Bourguiba between
January 2007 and December 2010. All cases were newly
diagnosed and previously untreated. Clinical characteristics
including basic medical data were obtained from medical
records. The control group consisted of 261 subjects. The
controls were recruited simultaneously from residents living
in the similar geographic area (South Tunisia) and matched
with patients for age and social conditions.
All individuals voluntarily participated in the study and
gave their written informed consent. Cases and controls
subjects were interviewed using a standard questionnaire
including demographic characteristics (e.g. age and sex),
history of smoking, chewing and drinking. Lifetime con-
sumption of tobacco smoking and chewing and alcohol
drinking were also collected. Tobacco smoking dose was
estimated as ‘pack-years’ (CY = number of packs of 20
cigarettes per days for 1 year). Likewise, tobacco chewing
dose was estimated as ‘consumption-year’ (CY = fre-
quency of tobacco chewing 9 kept/day 9 duration of
year). Similarly, subjects who had drunk alcoholic bever-
ages at least once a week for more than 1 year previously
were defined as drinkers, and non-drinkers were those who
had never drunk alcohol.
DNA extraction and SNPs genotyping
DNA was extracted from peripheral blood samples of
patients and controls using the conventional phenol/chlo-
roform extraction method. Four SNP CYP1A1 6310C[T,
CYP2D6 8356G[A, NAT1 52936T[A, NAT2 803A[G
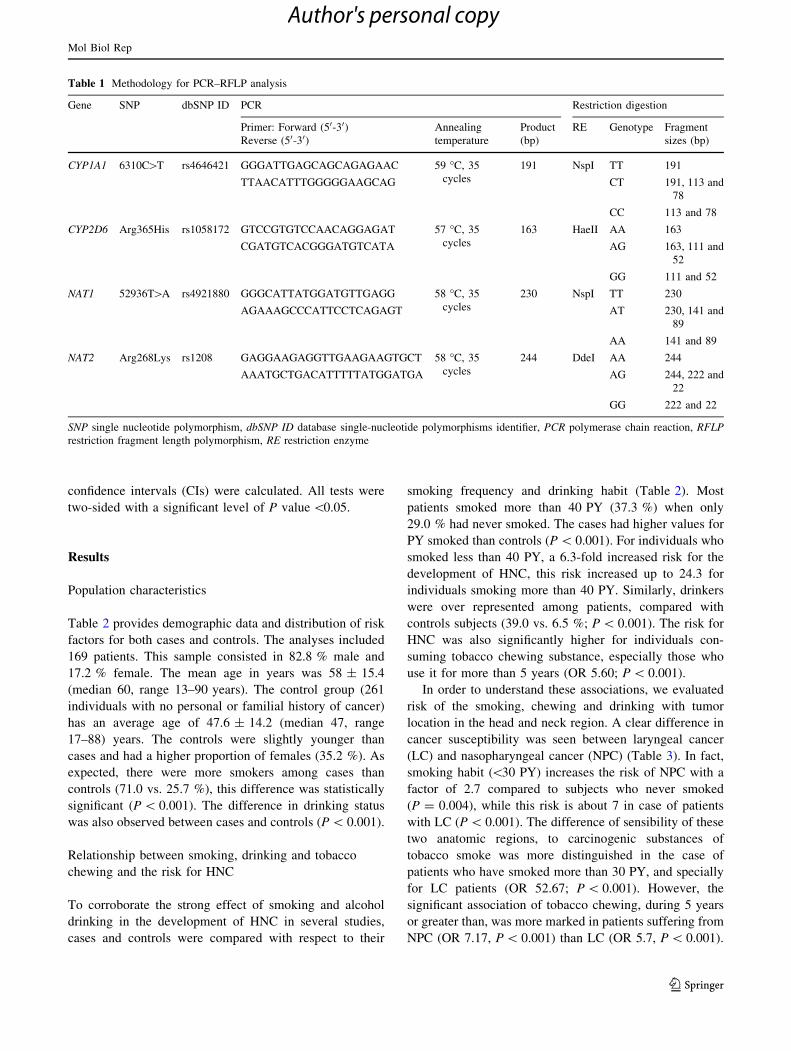
were analyzed in this study (Table 1). All these polymor-
phisms were analyzed by polymerase chain reaction (PCR)
combined with restriction fragment length polymorphism
(RFLP). The PCR fragment of the four investigated poly-
morphisms’ was subsequently digested with their specific
restriction enzyme. Digestion products were separated by
electrophoresis on ethidium bromide stained agarose gel
and visualized under UV light. All primers and PCR con-
ditions used are described in Table 1.
Statistical methods
Hardy–Weinberg equilibrium was examined using a Chi
square test with one degree of freedom. The other statis-
tical analyses were done using the SPSS statistical package
software, version 13.0. Means of quantitative variables
were compared between groups using Student t test and
distributions of categorical variables were compared by
Pearson Chi square test. In case of inequality of variances,
Welch test was then used. Multiple logistic regression was
conducted to estimate the relative risk of each SNP
adjusted for age, gender, smoking, chewing as well as
alcohol drinking. Odds ratios (ORs) and their 95 %
Mol Biol Rep
123
Author's personal copy
confidence intervals (CIs) were calculated. All tests were
two-sided with a significant level of P value \0.05.
Results
Population characteristics
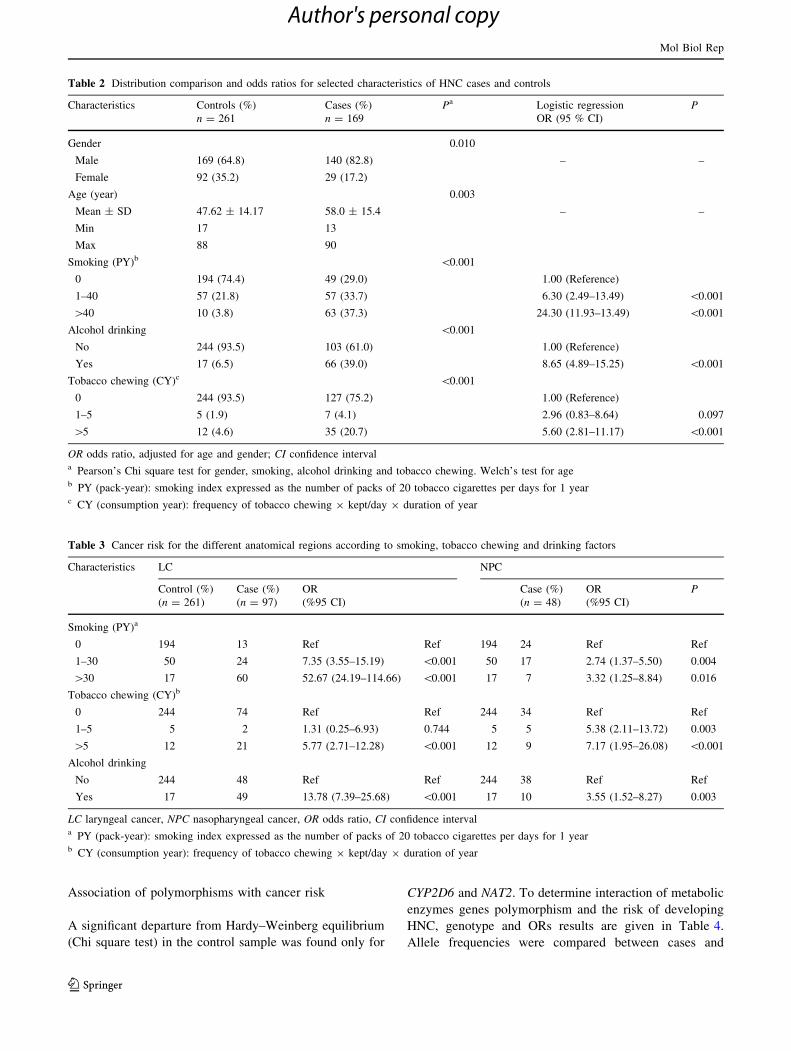
Table 2 provides demographic data and distribution of risk
factors for both cases and controls. The analyses included
169 patients. This sample consisted in 82.8 % male and
17.2 % female. The mean age in years was 58 ± 15.4
(median 60, range 13–90 years). The control group (261
individuals with no personal or familial history of cancer)
has an average age of 47.6 ± 14.2 (median 47, range
17–88) years. The controls were slightly younger than
cases and had a higher proportion of females (35.2 %). As
expected, there were more smokers among cases than
controls (71.0 vs. 25.7 %), this difference was statistically
significant (P \ 0.001). The difference in drinking status
was also observed between cases and controls (P \ 0.001).
Relationship between smoking, drinking and tobacco
chewing and the risk for HNC
To corroborate the strong effect of smoking and alcohol
drinking in the development of HNC in several studies,
cases and controls were compared with respect to their
smoking frequency and drinking habit (Table 2). Most
patients smoked more than 40 PY (37.3 %) when only
29.0 % had never smoked. The cases had higher values for
PY smoked than controls (P \ 0.001). For individuals who
smoked less than 40 PY, a 6.3-fold increased risk for the
development of HNC, this risk increased up to 24.3 for
individuals smoking more than 40 PY. Similarly, drinkers
were over represented among patients, compared with
controls subjects (39.0 vs. 6.5 %; P \ 0.001). The risk for
HNC was also significantly higher for individuals con-
suming tobacco chewing substance, especially those who
use it for more than 5 years (OR 5.60; P \ 0.001).
In order to understand these associations, we evaluated
risk of the smoking, chewing and drinking with tumor
location in the head and neck region. A clear difference in
cancer susceptibility was seen between laryngeal cancer
(LC) and nasopharyngeal cancer (NPC) (Table 3). In fact,
smoking habit (\30 PY) increases the risk of NPC with a
factor of 2.7 compared to subjects who never smoked
(P = 0.004), while this risk is about 7 in case of patients
with LC (P \ 0.001). The difference of sensibility of these
two anatomic regions, to carcinogenic substances of
tobacco smoke was more distinguished in the case of
patients who have smoked more than 30 PY, and specially
for LC patients (OR 52.67; P \ 0.001). However, the
significant association of tobacco chewing, during 5 years
or greater than, was more marked in patients suffering from
NPC (OR 7.17, P \ 0.001) than LC (OR 5.7, P \ 0.001).
Table 1 Methodology for PCR–RFLP analysis
Gene SNP dbSNP ID PCR Restriction digestion
Primer: Forward (50-30)Reverse (50-30)
Annealing
temperature
Product
(bp)
RE Genotype Fragment
sizes (bp)
CYP1A1 6310C[T rs4646421 GGGATTGAGCAGCAGAGAAC 59 �C, 35
cycles
191 NspI TT 191
TTAACATTTGGGGGAAGCAG CT 191, 113 and
78
CC 113 and 78
CYP2D6 Arg365His rs1058172 GTCCGTGTCCAACAGGAGAT 57 �C, 35
cycles
163 HaeII AA 163
CGATGTCACGGGATGTCATA AG 163, 111 and
52
GG 111 and 52
NAT1 52936T[A rs4921880 GGGCATTATGGATGTTGAGG 58 �C, 35
cycles
230 NspI TT 230
AGAAAGCCCATTCCTCAGAGT AT 230, 141 and
89
AA 141 and 89
NAT2 Arg268Lys rs1208 GAGGAAGAGGTTGAAGAAGTGCT 58 �C, 35
cycles
244 DdeI AA 244
AAATGCTGACATTTTTATGGATGA AG 244, 222 and
22
GG 222 and 22
SNP single nucleotide polymorphism, dbSNP ID database single-nucleotide polymorphisms identifier, PCR polymerase chain reaction, RFLP
restriction fragment length polymorphism, RE restriction enzyme
Mol Biol Rep
123
Author's personal copy
Association of polymorphisms with cancer risk
A significant departure from Hardy–Weinberg equilibrium
(Chi square test) in the control sample was found only for
CYP2D6 and NAT2. To determine interaction of metabolic
enzymes genes polymorphism and the risk of developing
HNC, genotype and ORs results are given in Table 4.
Allele frequencies were compared between cases and
Table 2 Distribution comparison and odds ratios for selected characteristics of HNC cases and controls
Characteristics Controls (%)
n = 261
Cases (%)
n = 169
Pa Logistic regression
OR (95 % CI)
P
Gender 0.010
Male 169 (64.8) 140 (82.8) – –
Female 92 (35.2) 29 (17.2)
Age (year) 0.003
Mean ± SD 47.62 ± 14.17 58.0 ± 15.4 – –
Min 17 13
Max 88 90
Smoking (PY)b \0.001
0 194 (74.4) 49 (29.0) 1.00 (Reference)
1–40 57 (21.8) 57 (33.7) 6.30 (2.49–13.49) \0.001
[40 10 (3.8) 63 (37.3) 24.30 (11.93–13.49) \0.001
Alcohol drinking \0.001
No 244 (93.5) 103 (61.0) 1.00 (Reference)
Yes 17 (6.5) 66 (39.0) 8.65 (4.89–15.25) \0.001
Tobacco chewing (CY)c \0.001
0 244 (93.5) 127 (75.2) 1.00 (Reference)
1–5 5 (1.9) 7 (4.1) 2.96 (0.83–8.64) 0.097
[5 12 (4.6) 35 (20.7) 5.60 (2.81–11.17) \0.001
OR odds ratio, adjusted for age and gender; CI confidence intervala Pearson’s Chi square test for gender, smoking, alcohol drinking and tobacco chewing. Welch’s test for ageb PY (pack-year): smoking index expressed as the number of packs of 20 tobacco cigarettes per days for 1 yearc CY (consumption year): frequency of tobacco chewing 9 kept/day 9 duration of year
Table 3 Cancer risk for the different anatomical regions according to smoking, tobacco chewing and drinking factors
Characteristics LC NPC
Control (%)
(n = 261)
Case (%)
(n = 97)
OR
(%95 CI)
Case (%)
(n = 48)
OR
(%95 CI)
P
Smoking (PY)a
0 194 13 Ref Ref 194 24 Ref Ref
1–30 50 24 7.35 (3.55–15.19) \0.001 50 17 2.74 (1.37–5.50) 0.004
[30 17 60 52.67 (24.19–114.66) \0.001 17 7 3.32 (1.25–8.84) 0.016
Tobacco chewing (CY)b
0 244 74 Ref Ref 244 34 Ref Ref
1–5 5 2 1.31 (0.25–6.93) 0.744 5 5 5.38 (2.11–13.72) 0.003
[5 12 21 5.77 (2.71–12.28) \0.001 12 9 7.17 (1.95–26.08) \0.001
Alcohol drinking
No 244 48 Ref Ref 244 38 Ref Ref
Yes 17 49 13.78 (7.39–25.68) \0.001 17 10 3.55 (1.52–8.27) 0.003
LC laryngeal cancer, NPC nasopharyngeal cancer, OR odds ratio, CI confidence intervala PY (pack-year): smoking index expressed as the number of packs of 20 tobacco cigarettes per days for 1 yearb CY (consumption year): frequency of tobacco chewing 9 kept/day 9 duration of year
Mol Biol Rep
123
Author's personal copy
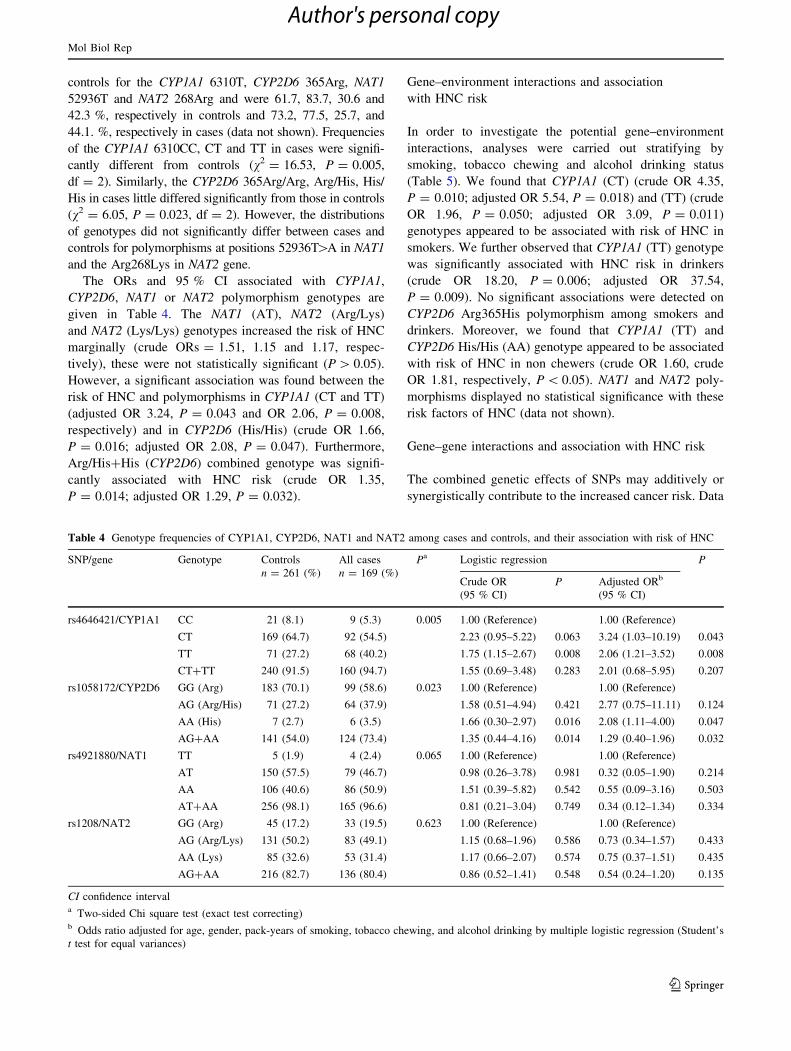
controls for the CYP1A1 6310T, CYP2D6 365Arg, NAT1
52936T and NAT2 268Arg and were 61.7, 83.7, 30.6 and
42.3 %, respectively in controls and 73.2, 77.5, 25.7, and
44.1. %, respectively in cases (data not shown). Frequencies
of the CYP1A1 6310CC, CT and TT in cases were signifi-
cantly different from controls (v2 = 16.53, P = 0.005,
df = 2). Similarly, the CYP2D6 365Arg/Arg, Arg/His, His/
His in cases little differed significantly from those in controls
(v2 = 6.05, P = 0.023, df = 2). However, the distributions
of genotypes did not significantly differ between cases and
controls for polymorphisms at positions 52936T[A in NAT1
and the Arg268Lys in NAT2 gene.
The ORs and 95 % CI associated with CYP1A1,
CYP2D6, NAT1 or NAT2 polymorphism genotypes are
given in Table 4. The NAT1 (AT), NAT2 (Arg/Lys)
and NAT2 (Lys/Lys) genotypes increased the risk of HNC
marginally (crude ORs = 1.51, 1.15 and 1.17, respec-
tively), these were not statistically significant (P [ 0.05).
However, a significant association was found between the
risk of HNC and polymorphisms in CYP1A1 (CT and TT)
(adjusted OR 3.24, P = 0.043 and OR 2.06, P = 0.008,
respectively) and in CYP2D6 (His/His) (crude OR 1.66,
P = 0.016; adjusted OR 2.08, P = 0.047). Furthermore,
Arg/His?His (CYP2D6) combined genotype was signifi-
cantly associated with HNC risk (crude OR 1.35,
P = 0.014; adjusted OR 1.29, P = 0.032).
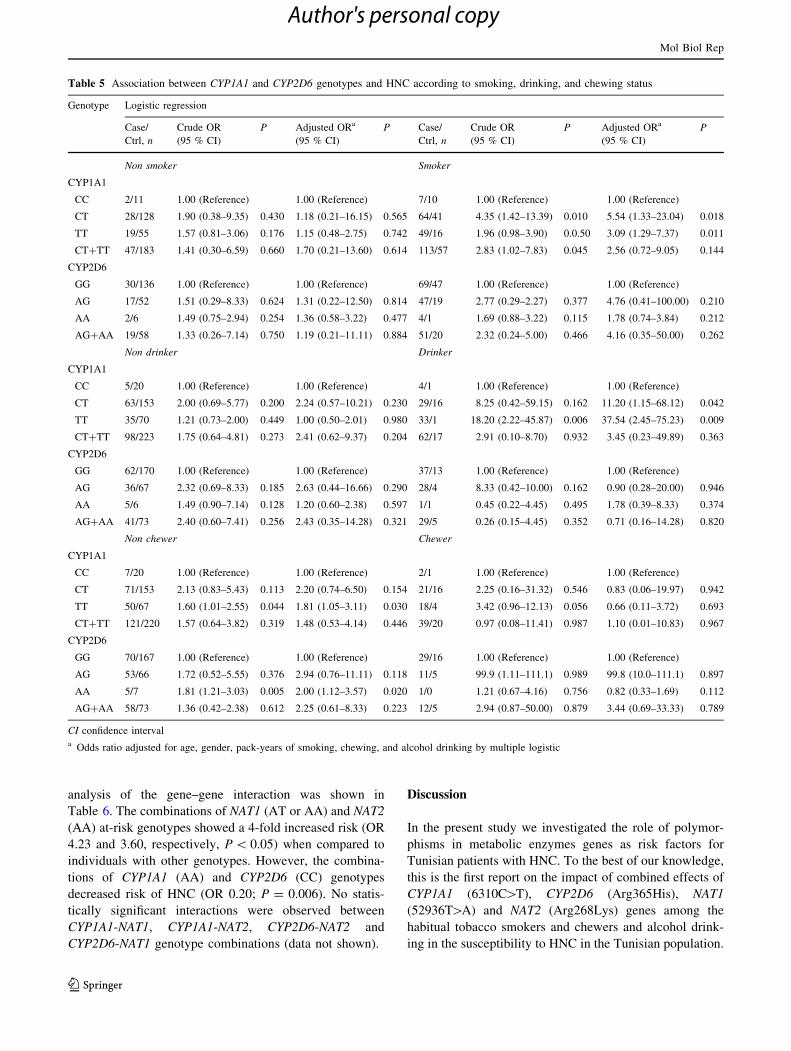
Gene–environment interactions and association
with HNC risk
In order to investigate the potential gene–environment
interactions, analyses were carried out stratifying by
smoking, tobacco chewing and alcohol drinking status
(Table 5). We found that CYP1A1 (CT) (crude OR 4.35,
P = 0.010; adjusted OR 5.54, P = 0.018) and (TT) (crude
OR 1.96, P = 0.050; adjusted OR 3.09, P = 0.011)
genotypes appeared to be associated with risk of HNC in
smokers. We further observed that CYP1A1 (TT) genotype
was significantly associated with HNC risk in drinkers
(crude OR 18.20, P = 0.006; adjusted OR 37.54,
P = 0.009). No significant associations were detected on
CYP2D6 Arg365His polymorphism among smokers and
drinkers. Moreover, we found that CYP1A1 (TT) and
CYP2D6 His/His (AA) genotype appeared to be associated
with risk of HNC in non chewers (crude OR 1.60, crude
OR 1.81, respectively, P \ 0.05). NAT1 and NAT2 poly-
morphisms displayed no statistical significance with these
risk factors of HNC (data not shown).
Gene–gene interactions and association with HNC risk
The combined genetic effects of SNPs may additively or
synergistically contribute to the increased cancer risk. Data
Table 4 Genotype frequencies of CYP1A1, CYP2D6, NAT1 and NAT2 among cases and controls, and their association with risk of HNC
SNP/gene Genotype Controls
n = 261 (%)
All cases
n = 169 (%)
Pa Logistic regression P
Crude OR
(95 % CI)
P Adjusted ORb
(95 % CI)
rs4646421/CYP1A1 CC 21 (8.1) 9 (5.3) 0.005 1.00 (Reference) 1.00 (Reference)
CT 169 (64.7) 92 (54.5) 2.23 (0.95–5.22) 0.063 3.24 (1.03–10.19) 0.043
TT 71 (27.2) 68 (40.2) 1.75 (1.15–2.67) 0.008 2.06 (1.21–3.52) 0.008
CT?TT 240 (91.5) 160 (94.7) 1.55 (0.69–3.48) 0.283 2.01 (0.68–5.95) 0.207
rs1058172/CYP2D6 GG (Arg) 183 (70.1) 99 (58.6) 0.023 1.00 (Reference) 1.00 (Reference)
AG (Arg/His) 71 (27.2) 64 (37.9) 1.58 (0.51–4.94) 0.421 2.77 (0.75–11.11) 0.124
AA (His) 7 (2.7) 6 (3.5) 1.66 (0.30–2.97) 0.016 2.08 (1.11–4.00) 0.047
AG?AA 141 (54.0) 124 (73.4) 1.35 (0.44–4.16) 0.014 1.29 (0.40–1.96) 0.032
rs4921880/NAT1 TT 5 (1.9) 4 (2.4) 0.065 1.00 (Reference) 1.00 (Reference)
AT 150 (57.5) 79 (46.7) 0.98 (0.26–3.78) 0.981 0.32 (0.05–1.90) 0.214
AA 106 (40.6) 86 (50.9) 1.51 (0.39–5.82) 0.542 0.55 (0.09–3.16) 0.503
AT?AA 256 (98.1) 165 (96.6) 0.81 (0.21–3.04) 0.749 0.34 (0.12–1.34) 0.334
rs1208/NAT2 GG (Arg) 45 (17.2) 33 (19.5) 0.623 1.00 (Reference) 1.00 (Reference)
AG (Arg/Lys) 131 (50.2) 83 (49.1) 1.15 (0.68–1.96) 0.586 0.73 (0.34–1.57) 0.433
AA (Lys) 85 (32.6) 53 (31.4) 1.17 (0.66–2.07) 0.574 0.75 (0.37–1.51) 0.435
AG?AA 216 (82.7) 136 (80.4) 0.86 (0.52–1.41) 0.548 0.54 (0.24–1.20) 0.135
CI confidence intervala Two-sided Chi square test (exact test correcting)b Odds ratio adjusted for age, gender, pack-years of smoking, tobacco chewing, and alcohol drinking by multiple logistic regression (Student’s
t test for equal variances)
Mol Biol Rep
123
Author's personal copy
analysis of the gene–gene interaction was shown in
Table 6. The combinations of NAT1 (AT or AA) and NAT2
(AA) at-risk genotypes showed a 4-fold increased risk (OR
4.23 and 3.60, respectively, P \ 0.05) when compared to
individuals with other genotypes. However, the combina-
tions of CYP1A1 (AA) and CYP2D6 (CC) genotypes
decreased risk of HNC (OR 0.20; P = 0.006). No statis-
tically significant interactions were observed between
CYP1A1-NAT1, CYP1A1-NAT2, CYP2D6-NAT2 and
CYP2D6-NAT1 genotype combinations (data not shown).
Discussion
In the present study we investigated the role of polymor-
phisms in metabolic enzymes genes as risk factors for
Tunisian patients with HNC. To the best of our knowledge,
this is the first report on the impact of combined effects of
CYP1A1 (6310C[T), CYP2D6 (Arg365His), NAT1
(52936T[A) and NAT2 (Arg268Lys) genes among the
habitual tobacco smokers and chewers and alcohol drink-
ing in the susceptibility to HNC in the Tunisian population.
Table 5 Association between CYP1A1 and CYP2D6 genotypes and HNC according to smoking, drinking, and chewing status
Genotype Logistic regression
Case/
Ctrl, n
Crude OR
(95 % CI)
P Adjusted ORa
(95 % CI)
P Case/
Ctrl, n
Crude OR
(95 % CI)
P Adjusted ORa
(95 % CI)
P
Non smoker Smoker
CYP1A1
CC 2/11 1.00 (Reference) 1.00 (Reference) 7/10 1.00 (Reference) 1.00 (Reference)
CT 28/128 1.90 (0.38–9.35) 0.430 1.18 (0.21–16.15) 0.565 64/41 4.35 (1.42–13.39) 0.010 5.54 (1.33–23.04) 0.018
TT 19/55 1.57 (0.81–3.06) 0.176 1.15 (0.48–2.75) 0.742 49/16 1.96 (0.98–3.90) 0.0.50 3.09 (1.29–7.37) 0.011
CT?TT 47/183 1.41 (0.30–6.59) 0.660 1.70 (0.21–13.60) 0.614 113/57 2.83 (1.02–7.83) 0.045 2.56 (0.72–9.05) 0.144
CYP2D6
GG 30/136 1.00 (Reference) 1.00 (Reference) 69/47 1.00 (Reference) 1.00 (Reference)
AG 17/52 1.51 (0.29–8.33) 0.624 1.31 (0.22–12.50) 0.814 47/19 2.77 (0.29–2.27) 0.377 4.76 (0.41–100.00) 0.210
AA 2/6 1.49 (0.75–2.94) 0.254 1.36 (0.58–3.22) 0.477 4/1 1.69 (0.88–3.22) 0.115 1.78 (0.74–3.84) 0.212
AG?AA 19/58 1.33 (0.26–7.14) 0.750 1.19 (0.21–11.11) 0.884 51/20 2.32 (0.24–5.00) 0.466 4.16 (0.35–50.00) 0.262
Non drinker Drinker
CYP1A1
CC 5/20 1.00 (Reference) 1.00 (Reference) 4/1 1.00 (Reference) 1.00 (Reference)
CT 63/153 2.00 (0.69–5.77) 0.200 2.24 (0.57–10.21) 0.230 29/16 8.25 (0.42–59.15) 0.162 11.20 (1.15–68.12) 0.042
TT 35/70 1.21 (0.73–2.00) 0.449 1.00 (0.50–2.01) 0.980 33/1 18.20 (2.22–45.87) 0.006 37.54 (2.45–75.23) 0.009
CT?TT 98/223 1.75 (0.64–4.81) 0.273 2.41 (0.62–9.37) 0.204 62/17 2.91 (0.10–8.70) 0.932 3.45 (0.23–49.89) 0.363
CYP2D6
GG 62/170 1.00 (Reference) 1.00 (Reference) 37/13 1.00 (Reference) 1.00 (Reference)
AG 36/67 2.32 (0.69–8.33) 0.185 2.63 (0.44–16.66) 0.290 28/4 8.33 (0.42–10.00) 0.162 0.90 (0.28–20.00) 0.946
AA 5/6 1.49 (0.90–7.14) 0.128 1.20 (0.60–2.38) 0.597 1/1 0.45 (0.22–4.45) 0.495 1.78 (0.39–8.33) 0.374
AG?AA 41/73 2.40 (0.60–7.41) 0.256 2.43 (0.35–14.28) 0.321 29/5 0.26 (0.15–4.45) 0.352 0.71 (0.16–14.28) 0.820
Non chewer Chewer
CYP1A1
CC 7/20 1.00 (Reference) 1.00 (Reference) 2/1 1.00 (Reference) 1.00 (Reference)
CT 71/153 2.13 (0.83–5.43) 0.113 2.20 (0.74–6.50) 0.154 21/16 2.25 (0.16–31.32) 0.546 0.83 (0.06–19.97) 0.942
TT 50/67 1.60 (1.01–2.55) 0.044 1.81 (1.05–3.11) 0.030 18/4 3.42 (0.96–12.13) 0.056 0.66 (0.11–3.72) 0.693
CT?TT 121/220 1.57 (0.64–3.82) 0.319 1.48 (0.53–4.14) 0.446 39/20 0.97 (0.08–11.41) 0.987 1.10 (0.01–10.83) 0.967
CYP2D6
GG 70/167 1.00 (Reference) 1.00 (Reference) 29/16 1.00 (Reference) 1.00 (Reference)
AG 53/66 1.72 (0.52–5.55) 0.376 2.94 (0.76–11.11) 0.118 11/5 99.9 (1.11–111.1) 0.989 99.8 (10.0–111.1) 0.897
AA 5/7 1.81 (1.21–3.03) 0.005 2.00 (1.12–3.57) 0.020 1/0 1.21 (0.67–4.16) 0.756 0.82 (0.33–1.69) 0.112
AG?AA 58/73 1.36 (0.42–2.38) 0.612 2.25 (0.61–8.33) 0.223 12/5 2.94 (0.87–50.00) 0.879 3.44 (0.69–33.33) 0.789
CI confidence intervala Odds ratio adjusted for age, gender, pack-years of smoking, chewing, and alcohol drinking by multiple logistic
Mol Biol Rep
123
Author's personal copy

Environmental factors and HNC
Tobacco smoking is the strongest risk factor for HNC
besides lung cancer. Similarly; alcohol consumption is also
linked to an increase in HNC [36]. Our data show a sig-
nificant relation between tobacco smoking and the risk of
HNC. These results are compatible with previous epide-
miological data that show a strong correlation between oral
squamous cell carcinoma and smoking [37–40]. Moreover,
it was seen that tobacco chewing habits were not a negli-
gible risk factor, although by far not as important as
smoking. The use of tobacco chewing is considered as an
old-fashioned practice and the number of tobacco chewers
is diminishing gradually as younger generations of Tuni-
sians no longer take on this habit [41]. Many epidemio-
logical studies have shown a risk link between smokeless
tobacco (chewing) use and HNC [12, 21, 41–44].
Polymorphisms and HNC
Several studies that have been conducted in various ethnic
populations to examine the impact of putatively functional
polymorphisms in CYP1A1 and CYP2D6 in risk of HNC [12,
16, 19, 21, 36, 45] have yielded conflicting results. However, in
this case–control study in Tunisian population, we found a
significant association between CYP1A1 6310C[T and the
CYP2D6 Arg365His polymorphisms and risk of developing
HNC (Table 4). There were no previous report on association
of CYP2D6 Arg365His and CYP1A1 6310C[T SNPs with risk
of HNC. On the other hand, Li et al. [22] analyzed the geno-
types of CYP1A1 6310C[T polymorphism in Chinese patients
with hepato-cellular carcinoma and reported a significantly
higher risk for subjects with the CT or TT genotype. They
suggested that rs4646421 variant cause impaired inhibition of
the enzyme by altering the sequence specificity adequate for
the upstream stimulatory factor recognition, which may
repress CYP1A1 transcription, resulting in high inducibility of
CYP1A1 and carcinogenesis. In our study, we can support this
hypothesis in the case of HNC especially that the higher risk
appears to be dependent to allelic dose. In fact, Sakata et al.
[46] reported that CYP1A1 enzyme has a low level of tran-
scription in normal conditions, without induction elements as
carcinogen substances. Then, the presence of negative regu-
lator elements has been suggested.
The present finding suggests that no association was
found between the polymorphism genotypes of NAT1
52936T[A (rs4921880) and NAT2 Arg268Lys (rs1208)
and risk of HNC in our population. To our knowledge,
there have been only three studies examining NATs poly-
morphisms (rs4921880 and rs1208) and cancer [47–49]. In
fact, Majumder et al. [48] found that NAT2 Arg268Lys
genetic variant could not modulate the risk of oral cancer in
Indian population. Furthermore, Hou et al. [47] reported
that no association was observed between NAT2 Arg268-
Lys and oral and pharyngeal squamous cell carcinoma in
Taiwan population. Moreover, Heck et al. [49] concluded
that no clear associations were observed between renal
cancer and these two NATs polymorphisms in Countries of
Central and Eastern Europe.
Gene–environment interactions and association with HNC
risk
In the present study, the confounding effect of tobacco
smoking on the HNC association with CYP1A1 C6310T
and CYP2D6 Arg365His polymorphisms shows that only
CYP1A1 (CT) and (TT) genotypes appeared to be associ-
ated with risk of HNC in smokers (Table 5). Similar results
were observed in studies of other cancers in which the risk
of smokers was modulated by CYP1A1 (rs4646421) genetic
variations [22, 50–52]. CYP1A1 is expressed in many
epithelial tissues especially in buccal mucosa, suggesting
the in situ activation of tobacco carcinogens [53]. Cigarette
smoke has been shown to up regulate CYP1A1 under
in vitro conditions as well as in smokers [54–56]. A sig-
nificant association between the genetic variability affect-
ing detoxification of carcinogens derived from tobacco
smoke and the risk of HNC has been reported previously
[18, 36, 41, 57–60]. However, few studies [61–63] did not
find a relationship between smoking and risk of HNC
among cases with the CYP1A1 polymorphisms.
Table 6 HNC risk associated with genotypic combinations of
CYP1A1–CYP2D6 and NAT1–NAT2 polymorphisms in cases and
controls
Gene–gene Control, n Case, n Crude OR (95 % CI) P
CYP2D6–CYP1A1
AA–TT 1 3 Reference
AA–CT 22 30 0.093 (0.01–1.09) 0.050
AA–CC 48 35 0.20 (0.06–0.63) 0.006
AG–TT 6 3 0.38 (0.12–1.12) 0.061
AG–CT 46 30 0.55 (0.10–3.05) 0.499
AG/GG–CC 117 59 0.42 (0.14–1.27) 0.126
GG–TT 3 4 0.55 (0.19–1.55) 0.261
GG–CT 18 5 0.20 (0.03–1.25) 0.049
NAT1–NAT2
TT–GG 31 23 Reference
TT–AG 57 45 2.69 (0.45–16.00) 0.275
TT–AA 18 18 2.53 (0.44–14.45) 0.296
AT–GG 51 30 2.00 (0.32–12.33) 0.445
AT–AG 72 34 3.40 (0.58–14.68) 0.172
AT/AA–AA 27 15 4.23 (0.73–15.26) 0.005
AA–GG 3 0 3.60 (0.58–12.01) 0.048
AA–AG 2 4 0.00 (0.00–0.00) 0.999
OR odds ratio, CI confidence interval
Mol Biol Rep
123
Author's personal copy

In addition, CYP1A1 (TT) and CYP2D6 His/His (AA)
genotypes appeared to be associated with risk of HNC in
non-chewers (P \ 0.05) (Table 5). However, no interac-
tion was observed between these two polymorphisms and
risk of HNC among chewers. In contrast, recent case–
control studies found a significant association between
CYP1A1 and CYP2D6 polymorphism and head and neck
and oral cancer among chewers [12, 19, 64, 65].
No interaction between the NATs polymorphisms and
risk of HNC was observed in our population (among
smokers, chewers and drinkers) (data not shown). HNC is
strongly associated with smoking, and a few studies have
explored the role of NAT1 polymorphisms in the risk of
developing HNC in smokers. However, overall findings are
inconsistent and associations if present are weak, and
indicate either a decreased risk in carriers of NAT1 poly-
morphism [35], an increased risk [59] or a lack of associ-
ation [10, 31, 44, 66].
Gene–gene interactions and association with HNC risk
The combined genetic effects of single nucleotide poly-
morphisms may additively or synergistically contribute to
the increased cancer risk. To date, no studies have examined
the risk conferred by the combination of CYP1A1
(6310C[T), CYP2D6 (Arg365His), NAT1 (52936T[A) and
NAT2 (Arg268Ly) polymorphisms. Gene–gene interactions
in the present study reflected a pattern of susceptibility
similar to independent gene effects. We observed that the
presence of a variant either at NAT1 (AT or AA) or NAT2
(AA) locus increased HNC risk by 4-fold (P = 0.005)
(Table 6). However, single gene suggested no significant
association between the polymorphisms of these two genes
(NAT1 and NAT2) and risk of HNC. The combinations of
NAT1 (AT or AA) is linked to NAT2 (AA), so the influence of
NAT1 on susceptibility can be partially determined by
interactions with NAT2 and vice versa. However, functional
interactions are still obscure, and other combinations of
particular genetic variants of NAT enzymes may affect
cancer risk. This finding supports the hypothesis that com-
bined gene variants can lead to increased HNC risk by acting
synergistically. It is becoming clearly evident that single
gene cannot explain susceptibility to diseases with complex
etiology such as HNC. The lack of other gene–gene inter-
actions could be due to low statistical power because of the
small number of individuals in some genotypic classes.
Conclusion
In the present study, increased risk of HNC was been
observed with CYP1A1 6310C[T (TT) and CYP2D6
Arg365His (His/His) variant carriers. No association was
found between the polymorphisms genotypes of NAT1
52936T[A and NAT2 Arg268Lys and risk of HNC.
However, an increased risk of HNC was been observed
with the combination of both the NAT1 (AT or AA) or
NAT2 (AA) genotypes. The combinations of CYP1A1
(AA) and CYP2D6 (CC) genotypes decreased risk of HNC.
An association between HNC and CYP1A1 (TT) genotype
was found among patients with drinking habit and smok-
ing. This study is first such in the Tunisian population and
provides a baseline for further studies on metabolic genes
(CYP1A1 6310C[T, CYP2D6 Arg365His, NAT1
52936A[T and NAT2 Arg268Lys) polymorphisms and risk
of HNC in the Tunisian population.
Acknowledgments This work was supported by the Ministry of
Higher Education and Scientific Research, Tunisia. We thank the
Department of Otorhinolaryngology, Habib Borguiba Hospital, Sfax,
Tunisia for the recruitment of patients. We thank all members of this
Department of and specially Dr Adel Chakroun and Dr Bouthaina
Hammami for their efforts and assistance in recruiting patients.
Conflict of interest The authors declare that they have no conflict
of interest.
References
1. Koifman S, Koifman RJ (2003) Environment and cancer in
Brazil: an overview from a public health perspective. Mutat Res
544(2–3):305–311
2. Cloos J, Spitz MR, Schaniz SP et al (1996) Genetic susceptibility
to head and neck squamous cell carcinoma. J Natl Cancer Inst
88:530–535
3. Risch A, Ramroth H, Raedts V et al (2003) Laryngeal cancer risk
in Caucasians is associated with alcohol and tobacco consump-
tion but not modified by genetic polymorphisms in class I alcohol
dehydrogenases ADIIIB and ADIIIC, and glutathione S-trans-
ferases GSTM1 and GSTT1. Pharmacogenetics 13:225–230
4. Smith G, Stanley LA, Sim E et al (1995) Metabolic polymor-
phisms and cancer susceptibility. Cancer Surv 25:27–65
5. Rebbeck TR (1997) Molecular epidemiology of the human glu-
tathione S-transferase genotypes GSTM1 and GSTT1 in cancer
susceptibility. Cancer Epidemiol Biomark Prev 6:733–743
6. Boffetta P (2010) Biomarkers in cancer epidemiology: an inte-
grative approach. Carcinogenesis 31:121–126
7. Klaunig JE, Kamendulis LM, Hocevar BA (2010) Oxidative
stress and oxidative damage in carcinogenesis. Toxicol Pathol
38:96–109
8. Kawajiri K, Nakachi K, Imai K, Hayashi S, Watanabe J (1990)
Individual differences in lung cancer susceptibility in relation to
polymorphisms of P-450IA1 gene and cigarette dose. Princess
Takamatsu Symp 21:55–61
9. Bennett WP, Alavanja MCR, Blomeke B et al (1999) Envi-
ronmental tobacco smoke, genetic susceptibility, and risk of
lung cancer in never-smoking women. J Natl Cancer Inst
91:2009–2014
10. Agundez JAG (2008) Polymorphisms of human N-acetyltrans-
ferases and cancer risk. Curr Drug Metab 9:520–531
11. Wang S, Chanock S, Tang D, Li Z, Jedrychowski W, Perera FP
(2008) Assessment of interactions between PAH exposure and
genetic polymorphisms on PAH–DNA adducts in African-
Mol Biol Rep
123
Author's personal copy

American, Dominican, and Caucasian mothers and newborns.
Cancer Epidemiol Biomarkers Prev 17(2):405–413
12. Chaudhuri SR, Mukherjee S, Paul RR, Haldar A, Chaudhuri K
(2013) CYP1AI and CYP2E1 gene polymorphisms may increase
susceptibility to oral submucous fibrosis among betel quid
chewers of Eastern India. Gene 513:268–271
13. Tanimoto K, Hayashi S, Yoshiga K, Ichikawa T (1999) Poly-
morphisms of CYP1A1 and GSTM1 gene involved in oral
squamous cell carcinoma in association with a cigarette dose.
Oral Oncol 35:191–196
14. Gajecka M, Rydzanicz M, Jaskula-Sztul R et al (2005) CYP1A1,
CYP2D6, CYP2E1, NAT2, GSTM1 and GSTT1 polymorphisms
or their combinations are associated with the increased risk of the
laryngeal squamous cell carcinoma. Mutat Res 574:112–113
15. Singh AP, Shah PP, Ruwali M, Mathur N, Pant MC, Parmar D
(2009) Polymorphism in cytochrome P4501A1 is significantly
associated with head and neck cancer risk. Cancer Invest 27:
869–876
16. Olivieri HER, Silva SD, Mendonca FF et al (2009) CYP1A2*1C,
CYP2E1*5B and GSTM1 polymorphisms are predictors of risk
and poor outcome in head and neck squamous cell carcinoma
patients. Oral Oncol 45:e73–e79
17. Gattas GJ, de Carvalh MB, Siraque MS et al (2006) Genetic
polymorphisms of CYP1A1, CYP2E1, GSTM1 and GSTT1
associated with head and neck cancer. Head Neck 28:819–826
18. Tai J, Yang M, Ni X, Yu D et al (2010) Genetic polymorphisms
in cytochrome P450 genes are associated with an increased risk
of squamous cell carcinoma of the larynx and hypopharynx in a
Chinese population. Cancer Genet Cytogenet 196:76–82
19. Shukla P, Gupta D, Pant MC, Parmar D (2012) CYP 2D6 poly-
morphism: a predictor of susceptibility and response to chemo-
radiotherapy in head and neck cancer. J Cancer Res Ther 8(1):
40–45
20. Gonzalez MV, Alvarez V, Pello MF, Meneandez MJ, Suaarez C,
Coto E (1998) Genetic polymorphism of N-acetyltransferase-2
glutathione S-transferase-M1 and cytochromes P450IIE1 and
P450IID6 in the susceptibility to head and neck cancer. Clin
Pathol 51:294–298
21. Matthias C, Jahnke V, Jones PW, Hoban PR, Alldersea JE,
Worrall SF, Fryer AA, Strange RC (1999) Cyclin D1 glutathione
S-transferase and cytochrome P450 genotypes and outcome in
patients with upper aerodigestive tract cancers: assessment of the
importance of individual genes using multivariate analysis.
Cancer Epidemiol Biomarkers Prev 8:815–823
22. Li R, Shugart YY, Zhou W et al (2009) Common genetic vari-
ation of the cytochrome P450 1A1 gene and risk of hepatocellular
carcinoma in a chinese population. Eur J Cancer 45:1239–1247
23. Hirvonen A (1999) Polymorphic NATs and cancer predisposi-
tion. IARC Sci Publ 148:251–270
24. Suzuki H, Morris JS, Li Y et al (2008) Interaction of the cyto-
chrome P4501A2, SULT1A1 and NAT gene polymorphisms with
smoking and dietary mutagen intake in modification of the risk of
pancreatic cancer. Carcinogenesis 29(6):1184–1191
25. Ambrosone CB, Kropp S, Yang J, Yao S, Shields PG, Chang-
Claude J (2008) Cigarette smoking, N-acetyltransferase 2 geno-
types, and breast cancer risk: pooled analysis and meta-analysis.
Cancer Epidemiol Biomarkers Prev 17:15–26
26. Rouissi K, Ouerhani S, Marrakchi R et al (2009) Combined effect of
smoking and inherited polymorphisms in arylamine N-acetyltrans-
ferase 2, glutathione S-transferases M1 and T1 on bladder cancer in a
Tunisian population. Cancer Genet Cytogenet 190(2):101–107
27. Cui X, Lu X, Hiura M, Omori H, Miyazaki W, Katoh T (2012)
Association of genotypes of carcinogen-metabolizing enzymes
and smoking status with bladder cancer in a Japanese population.
Environ Health Prev Med [Epub ahead of print]
28. de Lima Junior MM, Reis LO, Guilhen AC, Granja F et al (2012)
N-Acetyltransferase-2 gene polymorphisms and prostate cancer
susceptibility in Latin American patients. Med Oncol 29(4):
2889–2894
29. Zabost A, Roszkowska-Sli _z B, Wiatr E et al (2012) Polymor-
phism in the N-acetyltransferase 2 gene in patients with lung
cancer. Short communication. Pneumonol Alergol Pol 80(4):
323–328
30. Katoh T, Kaneko S, Boissy R, Watson M, Ikemura K, Bell AD
(1998) A pilot study testing the association between N-acetyl-
transferases 1 and 2 and risk of oral squamous cell carcinoma in
Japanese people. Carcinogenesis 19:1803–1807
31. Henning S, Cascorbi I, Munchow B, Kahnke V, Roots I (1999)
Association of arylamine N-acetyltransferases NAT1 and NAT2
genotypes to laryngeal cancer risk. Pharmacogenetics 9:103–111
32. Garte S, Gaspari L, Alexandrie AK, Ambrosone C, Autrup H,
Autrup JL et al (2001) Metabolic gene polymorphism frequencies
in control populations. Cancer Epidemiol Biomarkers Prev 10:
1239–1248
33. Marques CFS, Koifman S, Koifman RJ, Boffetta P, Brennan P,
Hatagima A (2006) Influence of CYP1A1, CYP2E1, GSTM3 and
NAT2 genetic polymorphisms in oral cancer susceptibility:
results from a case-control study in Rio de Janeiro. Oral Oncol
42:632–637
34. Gara S, Abdennebli M, Chatti S, Touati S, Ladgham A, Guemira
F (2007) Association of NAT2 gene substitution mutation T341C
with increased risk for head and neck cancer in Tunisia. Acta
Oncol 46:834–837
35. McKay JD, Hashibe M, Hung RJ, Wakefield J et al (2008)
Sequence variants of NAT1 and NAT2 and other xenometabolic
genes and risk of lung and aerodigestive tract cancers in Central
Europe. Cancer Epidemiol Biomarkers Prev 17:141–147
36. Sabitha K, Vishnuvardhan Reddy M, Jamil K (2010) Smoking
related risk involved in individuals carrying genetic variants of
CYP1A1 gene in head and neck cancer. Cancer Epidemiol 34:
587–592
37. Wynder EL, Stellman SD (1977) Comparative epidemiology of
tobacco-related cancers. Cancer Res 37:4608–4622
38. Blot WJ, McLaughlin JK, Winn DM (1988) Smoking and
drinking in relation to oral and pharyngeal cancer. Cancer Res
48:3282–3287
39. Reibel J (2003) Tobacco and oral diseases. Update on the evi-
dence, with recommendations. Med Princ Pract 12(1):22–32
40. Werbrouck J, De Ruyck K, Duprez F et al (2008) Single-nucle-
otide polymorphisms in DNA double strand break repair genes:
association with head and neck cancer and interaction with
tobacco use and alcohol consumption. Mutat Res 656:74–81
41. Sam SS, Thomas V, Reddy KS, Surianarayanan G, Chandr-
asekaran A (2010) Gene–environment interactions associated
with CYP1A1 MspI and GST polymorphisms and the risk of
upper aerodigestive tract cancers in an Indian population. J Can-
cer Res Clin Oncol 136:945–951
42. Muscat JE, Richie JP Jr, Thompson S, Wynder EL (1996) Gender
differences in smoking and risk for oral cancer. Cancer Res
56(22):5192–5197
43. Schildt EB, Eriksson M, Hardell L, Magnuson A (1998) Oral
snuff, smoking habits and alcohol consumption in relation to oral
cancer in a Swedish case–control study. Int J Cancer 77(3):
341–346
44. Fronhoees S, Bruning T, Ortiz-Pallado E, Brode P et al (2001)
Real-time PCR analysis of N-acetyltransferase NAT1 allele *3,
*4, *10, *11, *14 and *17 polymorphism in squamous cell cancer
of head and neck. Carcinogenesis 22:1405–1412
45. Bartsch H, Nair U, Risch A, Rojas M, Wikman H, Alexandrov K
(2000) Genetic polymorphism of CYP genes alone or in
Mol Biol Rep
123
Author's personal copy

combination as a risk modifier of tobacco-related cancers. Cancer
Epidemiol Biomarkers Prev 9:3–28
46. Sakata Y, Yoshioka W, Tohyama C et al (2007) Internal genomic
sequence of human CYP1A1 gene is involved in superinduction
of dioxin-induced CYP1A1 transcription by cycloheximide.
Biochem Biophys Res Commun 355:687–692
47. Hou YY, Ou HL, Chu ST et al (2011) NAT2 slow acetylation
haplotypes are associated with the increased risk of betel quid-
related oral and pharyngeal squamous cell carcinoma. Oral Surg
Oral Med Oral Pathol Oral Radiol Endod 112(4):484–492
48. Majumder M, Ghosh S, Roy B (2012) Association between
polymorphisms at N-acetyltransferase 1 (NAT1) and risk of oral
leukoplakia and cancer. Indian J Med Res 136(4):605–613
49. Heck JE, Moore LE, Lee Y-CA, McKay JD et al (2012) Xeno-
biotic metabolizing gene variants and renal cell cancer: a multi-
center study. Cancer Epidemiol Prev 2:16
50. Taioli E, Ford J, Trachman J, Li Y, Demopoulos R, Garte S
(1998) Lung cancer risk and CYP1A1 genotype in African
Americans. Carcinogenesis 19(5):813–817
51. Ishibe N, Wiencke JK, Zuo ZF, McMillan A, Spitz M, Kelsey KT
(1997) Susceptibility to lung cancer in light smokers associated
with CYP1A1 polymorphisms in Mexican- and African-Ameri-
cans. Cancer Epidemiol Biomarkers Prev 6(12):1075–1080
52. Vineis P, Kogevinas M, Simonato L et al (2000) Levelling-off of
the risk of lung and bladder cancer in heavy smokers: an analysis
based on multicentric case–control studies and a metabolic
interpretation. Mutat Res 463(1):103–110
53. Vondracek M, Xi Z, Larsson P et al (2001) Cytochrome P450
expression and related metabolism in human buccal mucosa.
Carcinogenesis 22:481–488
54. Nagaraj NS, Beckers S, Mensah JK, Waigel S, Vigneswaran N,
Zacharias W (2006) Cigarette smoke condensate induces cyto-
chromes P450 and aldo–keto reductases in oral cancer cells.
Toxicol Lett 165:182–194
55. Chi AC, Appleton K, Henriod JB, Krayer JW et al (2009) Dif-
ferential induction of CYP1A1 and CYP1B1 by benzoapyrene in
oral squamous cell carcinoma cell lines and by tobacco smoking
in oral mucosa. Oral Oncol 45:980–985
56. Spivack SD, Hurteau GJ, Jain R, Kumar SV, Aldous KM, Gierthy
JF et al (2004) Gene–environment interaction signatures by
quantitative mRNA profiling in exfoliated buccal mucosal cells.
Cancer Res 64:6805–6813
57. Cha IH, Park JY, Chung WY, Choi MA, Kim HJ, Park KK (2007)
Polymorphisms of CYP1A1 and GSTM1 gene and susceptibility
to oral cancer. Yonsei Med J 48:233–239
58. Szyfter K, Szmeja Z, Szyfter W et al (1999) Molecular and
cellular alterations in tobacco smoke-associated larynx cancer.
Mutat Res 445:259–274
59. Olshan AF, Weissler M, Watson MA, Bell D (2000) GSTM1,
GSTT1, GSTP1, CYP1A1, and NAT1 polymorphisms tobacco
use and risk of head and neck cancers. Cancer Epidemiol Bio-
markers Prev 9:185–191
60. Basham VM, Pharoah PD, Healey CS et al (2001) Polymor-
phisms in CYP1A1 and smoking: no association with breast
cancer risk. Carcinogenesis 22:1797–1800
61. Matthias C, Bockmuehl U, Jahnke V et al (1998) Polymorphism
in cytochrome P450 CYP2D6, CYP1A1, CYP2E1 and glutathi-
one S-transferase, GSTM1, GSTM3, GSTT1 and susceptibility to
tobacco-related cancers: studies in upper aerodigestive tract
cancers. Pharmacogenetics 8:91–100
62. Oude Ophuis MB, Van Lieshout EM, Roelofs HM, Peters WH,
Manni JJ (1998) Glutathione S-transferase M1 and T1 and
cytochrome P4501A1 polymorphism in relation to the risk for
benign and malignant head and neck lesions. Cancer 82:936–943
63. Varel a-Lema L, Taioli E, Ruano-Ravina A, Barros-Dios JM,
Anantharaman D, Benhamou S et al (2008) Meta-analysis and
pooled analysis of GSTM1 and CYP1A1 polymorphisms and oral
and pharyngeal cancers: a HuGE-GSEC review. Genet Med
10:369–384
64. Anantharaman D, Chaubal PM, Kannan S, Bhisey RA, Mahimkar
MB (2007) Susceptibility to oral cancer by genetic polymor-
phisms at CYP1A1, GSTM1 and GSTT1 loci among Indians:
tobacco exposure as a risk modulator. Carcinogenesis
28:1455–1462
65. Hernando-Rodriguez M, Rey-Barja N, Marichalar-Mendia X et al
(2012) Role of cytochrome P-450 genetic polymorphisms in oral
carcinogenesis. J Oral Pathol Med 41:1–8
66. Demokan S, Suoglu Y, Gozeler M, Demir D, Dalay N (2010) N-
Acetyltransferase 1 and 2 gene sequence variants and risk of head
and neck cancer. Mol Biol Rep 37:3217–3226
Mol Biol Rep
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















