
Asplenium ceterach and A. octoploideum on the Canary Islands (Aspleniaceae, Pteridophyta)
31
American Fern /ournal 94(2):81_111 (2oo4) Asplenium ceterach and A. octoploideum on the canary Islands (Aspleniaceae, pteridophyta) Canolrrun J. VnN DEN HEEDE Pteridological section, Department of Biology, Ghent Universitv, K.L. Ledeganckstraat 3S, 8-9000 Ghent, Belgium Snrurraco Pnlanórl and Evrr.ra parucua Departamento de Biología Vegetal I, Facultad de Biología, Universidad Complutense, E-2g040 Madrid, Spain Roxnln L.L. VrnNrl Pteridological Section, Department of Biology, Ghent University, K.L. Ledeganckstraat 3S, 8-9000 Ghent, Belgium AsstRact'-Isozyme and plastid DNA analysis prove that true A. ceterach occurs on the canary Islands' in addition to á. aureum and an óctoploid taxon. combining morphological and cytological observations leads to correct determination, but the exospore lerïgth alone also allows reliable identification of these canarian species. our allozyme data suggestihat the canarian á. ceteroch population is not completely genetically isolated frtm the n,rroiË"r, ones. The holotype of ceterach oureum var. paruifofium' formerly rágarded as an octoploid taxon, proved to be á. ceterach,leaving the octoploid without a correct name. The recently described A. octoploideum shows monomorphic, presumably fixed heterozygosity for a combinaiion of the patterns seen in á. ceteroch and á' oureum at four loci (áoÍ, skdh, ti", uÁd Pgi-2)confirming its allo-octoploid nature. It most probably originated by chromosome doubling in a Ltraploid hybrid between A. aureum and. A' ceterach or via the union of their unreduced gu-u1"r. Pgi-2indicatËs multiple origins of the allo- octoploid, implicating recurrent gene flow froÀ tetraploiás to octoploids. Asplenium subgenusCeÍerach(Willd.) Bir eÍ o1.is a small group of about nine fern taxa within the large (729 species), subcosmopohàn genus Asplenium L. (Kramer and Viane, 1990). rËis subgenus contains xerophytic rock ferns with the dorsal side of the lamina denóly covered with reddish- brown scales (: paleae). van den heed e et ol. (2003) háve shown that the grorrp must be restricted to the Eurasianand Macaronesian species. Ever since the description of Asplenium aureum Cav. from Tenerife by Cavanilles [taor), it has beenuncleai how many " ceterach., species are extant in the canarianArchipelago, and whetherthe ,,Éuropean,, A. àeterachL.(syn.: Ceterach officinarum Willd.) occurs in Macaronesia (Table 1). This confusion was caused by the lack of distinctive characters to disti.rgr,ish both ,pu"i"r. cavanilles (raor) and Bory de st. Vincent (1802) mentioied only the much larger size of A. aureum comparedto that of A. ceteracrl. Willdáo* (1810) introduced the concept of "toothed scales" as a diagnostic feature, whereas Milde (1865) claimed that "Cuticularstreifen" (cuticul-ar lines or ridges on the periclinal cell walls of the scales) could be used to distinguish A. oureum from t ar,tro. for correspondence.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Asplenium ceterach and A. octoploideum on the Canary Islands (Aspleniaceae, Pteridophyta)

American Fern /ournal 94(2):81_111 (2oo4)
Asplenium ceterach and A. octoploideum on thecanary Islands (Aspleniaceae, pteridophyta)
Canolrrun J. VnN DEN HEEDEPteridological section, Department of Biology, Ghent Universitv,
K.L. Ledeganckstraat 3S, 8-9000 Ghent, Belgium
Snrurraco Pnlanórl and Evrr.ra parucuaDepartamento de Biología Vegetal I, Facultad de Biología,
Universidad Complutense, E-2g040 Madrid, Spain
Roxnln L.L. VrnNrlPteridological Section, Department of Biology, Ghent University,
K.L. Ledeganckstraat 3S, 8-9000 Ghent, Belgium
AsstRact'-Isozyme and plastid DNA analysis prove that true A. ceterach occurs on the canaryIslands' in addition to á. aureum and an óctoploid taxon. combining morphological andcytological observations leads to correct determination, but the exospore lerïgth alone also allowsreliable identification of these canarian species. our allozyme data suggestihat the canarian á.ceteroch population is not completely genetically isolated frtm the n,rroiË"r, ones. The holotype ofceterach oureum var. paruifofium' formerly rágarded as an octoploid taxon, proved to be á.ceterach,leaving the octoploid without a correct name. The recently described A. octoploideumshows monomorphic, presumably fixed heterozygosity for a combinaiion of the patterns seen in á.ceteroch and á' oureum at four loci (áoÍ, skdh, ti", uÁd Pgi-2)confirming its allo-octoploid nature.It most probably originated by chromosome doubling in a Ltraploid hybrid between A. aureum and.A' ceterach or via the union of their unreduced gu-u1"r. Pgi-2indicatËs multiple origins of the allo-octoploid, implicating recurrent gene flow froÀ tetraploiás to octoploids.
Asplenium subgenus CeÍerach (Willd.) Bir eÍ o1. is a small group of aboutnine fern taxa within the large (729 species), subcosmopohàn genusAsplenium L. (Kramer and Viane, 1990). rËis subgenus contains xerophyticrock ferns with the dorsal side of the lamina denóly covered with reddish-brown scales (: paleae). van den heed e et ol. (2003) háve shown that the grorrpmust be restricted to the Eurasian and Macaronesian species.
Ever since the description of Asplenium aureum Cav. from Tenerife byCavanilles [taor), it has been uncleai how many " ceterach., species are extantin the canarian Archipelago, and whether the ,,Éuropean,, A. àeterachL.(syn.:Ceterach officinarum Willd.) occurs in Macaronesia (Table 1). This confusionwas caused by the lack of distinctive characters to disti.rgr,ish both ,pu"i"r.cavanilles (raor) and Bory de st. Vincent (1802) mentioied only the muchlarger size of A. aureum compared to that of A. ceteracrl. Willdáo* (1810)introduced the concept of "toothed scales" as a diagnostic feature, whereasMilde (1865) claimed that "Cuticularstreifen" (cuticul-ar lines or ridges on thepericlinal cell walls of the scales) could be used to distinguish A. oureum from
t ar,tro. for correspondence.

82 AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
TasLE 1. Taxa covered by names found in the literature. Abbreviations used: A. cet.:A. ceterach,A. aur.: A. aureum, A. lol.: A. lolegnamense, A. par.:" A. parvifofium sensu Vida and Reichst," :A. octoploideum. I: the filled symbol indicates that the taxon was included in this author'sconcept of the species mentioned in column 2; tr: symbol indicates that this taxon was included inthe author's concept of the species mentioned in column 2 prior to its formal description.
Taxa included in this name
Literature reference Name used or published A. cet. A. aur. A.Iol. "A. paL"
Borv de St. Vincent 1802 A. ceterach
Linnaeus 1753Cavanilles 1801
Desvaux 1827
Moore 1857
Lowe, E. I . 1858
Kuhn 1868Sauer 1.880
Luerssen 1889Schneider 1892
Christ 1897
Burchard 1929
Chevalier 1935
Copeland to+z
Manton 1950
Romariz L953
Dansereau 1961Fabbri 1965
Kunkel tgos
A. ceterachA. aureum
A. IatifoliumC. aurea
C. au rcumC. officinarumC. off icinorum
A. ceterach var. oureumC. aureumC. officinarum
C. aureumC. officinarumC. auteumC. officinarumC . a u r c u m
C. officinarumC. aureumC. aureumC. officianarumC. aureum
C. officinarumC, aureumC. aureumC. officinarumC. officinarumA. ceterachA. ceterach var. oureumC. aureumC. officinarumC. aureumC. officinarumC. officinarumC. officinarum var. aureum
TI T D t rI
I !I
trI
I t rt r !
tt
r ! !I
IT I
It
TTTTT
!TIT
von Buch 1.828 ["1825"]* C. aureum
Hooker 1860, 1861 A. ceterach
Bol le 1864
Milde 1866b. 1867a,b C. aurcum
r f lT
r t nT !
MI N
l
I
Tardieu-Blot 1946 C. officinarum
I t rI
I !I T t r
T !I
It
T !l !Lems 1958. 1960 C. aureum
Benl and Kunkel tg0z C. aureumvar. ouÍeumC. aureum var. parvifolium I
Lid 1e67Hansen 1969
C. aureumC. aureum tr
Benl and Sventenius 1970 C. aureumvaÍ. oureumC. aureum var. parvifolium I
II

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP
Tnels 1. Continued.
Taxa included in this name
Literature reference Name used or published
Kunkel tgzt
Hansen and Sunding 1979
Reichstein 1984
Bir et a l . tggs
Manton et al. tgg6
Gibby and Lovis 1989Ormonde 1990
Viane and Reichstein 1992
Griffiths 1997
Hoshizaki and Moran 2001
C. aureum var. oureumC. oureum var. parvifoliumC. oureum var. oureumC. aureum var. parvifolium
C. aureum var. ou.reumC. aureum var. parvifoliumC. offinarumA. aureumA. ceterach ssp. ceterachC. aureum var. aureumC. aureum var. parvifoliumC. lolegnamenseC. aureum vat. aureumC. aureum var. madeirenseC. aureum var. parvifoliumA. parvifoliumA. lolegnamenseA. aureumA. ceterachC. aureumC. officinarum
* According to Stafleu and Cowan (1976) this book was only published after 28 May 1828, it is notclear whether Link or von Buch made the combination "C. aureum", which is, in anv case,antedated by Desvaux (1.827).
i ts continental counterpart, though he soon (Milde 1866a, 1866b, L867a,1867b) cast doubt on the utility of this character. As early as May 1,866, Mildeadmitted that "die Cuticularstreifen, welche die Spreuschuppen von C.oureum stets zeigen, fand ich nun auch an exemplaren, die sich von C.officinarum nicht unterscheiden liessen." Finally, he came to the conclusionthat the character was useless to discriminate Á. ceterach from A. aureum
[Milde, 1867b). Nevertheless, Bornmiil ler (Plantae exsicc. Canarienses-l901),and Benl and Kunkel (rgOz) heavily relied on this character to recognize taxa.Since 1.970, chromosome numbers were used to distinguish species in thisgroup (T. Reichstein, pers. comm.), and morphological characters became lessimportant (e.9. , Bir et aL. ,1985; Manton et aL. ,1986; Gibby and Lovis, 19Bg).
In 1967, Benl and Kunkel considered all Canarian plants that looked likeA. ceterach to be a dwarfed variety of A. aureum. Unfortunately, their varietyCeterach oureum (Cav.) Desv. var. parvifolium Benl and G.Kunkel waspublished without cytological information. In March '1,967, T. Reichsteincollected living " A. ceterach' ' on Gran Canaria, and sent material forchromosome counts to G. Vida. In 1970, these plants were found to beoctoploid, but because good cytological photographs were lacking the resultswere not published (T. Reichstein, pers. comm.). From then onwards, butwithout studying the type of A. parvifolium (Benl and G.Kunkel) Vida and
!
I
T
!
I
II
tt t

AMERICAN FERN JOURNAL: VOLUME 94 NUMBER 2 (2004)
Reichst., octoploid status was attributed to it. To clarify the origin of Á.parvifoliltm, a hybridization program was started by G. Vida in Budapest;results were partly published in Manton et al. (1986). Meanwhile, T.Reichstein had informed many pteridologists about the putative allo-octoploidnature of. A. parvifolium and briefly mentioned it in Hegi (1984).
To date two cytologically different endemic species are generally accepted tooccur on the Canary Islands: A. aureum and Á. parvifolium. Aspleniumaureum was found to be tetraploid by Manton (1950). Vida and Reichstein(Vida, 1.972; Viane and Reichstein, 1,992) suggested A. oureum to beallotetraploid, which was confirmed by ITS analysis (Van den heede et al.,2003). The name A. parvifolium was used for the allo-octoploid that prob-ably formed by chromosome doubling of the tetraploid hybrid between Á.oureum and Á. ceterach (Vida, 1,972; Viane and Reichstein, 1.992). After 1.970,all small Canarian plants that looked like A. ceteracft were considered to bea) A.parvifolium and b) octoploid. According to Manton ef a1. (rge0) "C.officinarum is not positively recorded from Macaronesia, but its formerpresence, at least in the Canaries, is suggested by the morphology of somerepresentatives of C. oureum sens. IaÍ. from these islands."
Within A. ceterach sensu 1oÍo three cytotypes are known, and according tothe Biological Species Concept (Mayr, '1.942,2000; see review in King, 1993),autopolyploids should be considered separate species because they producesterile hybrids with their parents from which they are reproductively isolated.Diploid A. javorkeanum Vida [Syn.: Á. ceterach ssp. biva]ens (D.E. M"y.)Greuter and Burdet; C. officinarum Willd. ssp. bivalens D.E. Mey.l is knownfrom Albania, Bulgaria, Croatia, Greece, Hungary, Italy, Romania, and Slovenia(Vida, 1963; Reichstein, 1984), and should be looked for in northern Algeriaand Turkey, because the triploid hybrid A. xmantoniaeYáróczy and Vida wasfound there (Greuter, 1980; Viane et aL.,1996). Tetraploid (Manton, L950; Vida,1963) A. ceteracft [Syn.: C. officinorum Willd. ssp. officinarum] is supposed tohave originated via chromosome doubling in Á. javorkeanum; its autopoly-ploid status was confirmed cytologically by Rasbach et aI. (1987). Theautotetraploid is the more common species, occurring throughout Europe(see maps in falas and Suominen, '1,972; Pichi Sermolli, '1.979; Reichstein,1984), southwestern Asia and the western Himalayas. Asplenium ceterach ismore rare in northern Africa (Jahandiez and Maire, 1931; Maire, 1.952; Quezeland Santa,1962; Siddiqi, 19Bg), but extends into Eritrea and Somalia (Viane eÍo/., 1996), the Arabian Peninsula (Collenette, 19Bb), and Yemen (Christ, 1900;Wood, 1997). The autohexaploid A. cyprium Viane and Van den heede (Syn.:
A. ceteracft ssp. cyprium Viane) was described from Cyprus (Van den heedeand Viane, 2OO2; Viane and Van den heede, 2OO2), and is also known fromGreece and Sicily (Viane et aL.,1996; Van den heede et al., 2oo2).
For the biosystematic revision of ï}ire Ceterac.h group (Van den heede, 2003),field trips were organized to study the Macaronesian representatives. Plantsthat we could not distinguish from the European A. ceterach were tentativelycalled A. parvifolium, and assumed to be octoploid. To our great surprise manyof them turned out to be tetraploid.

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP
In order to clear up the A. ceterach-A. parvifolium muddle on the CanarianArchipelago, we studied type material, and cytologically checked 145 samplesfrom Gran Canaria, La Palma, and Tenerife. Because the type of A. parvifoliumturned out to be Á. ceterach, the octoploid taxon needed a new name and wasdescribed as Á. octoploideum Viane and Van den heede (Van den heede andViane,2OO2).
Because electrophoretic analysis of isozymes has been successfully used instudies of reticulate complexes of Pteridophyta (Werth et aI., 1985a, L985b;Werth, 1991; Haufler et aI., 1995) and has been applied at population andspecies levels (see Haufler, 1985b, 1997), we tried this method together withDNA sequencing, to determine whether true A. ceteracft grows on the CanaryIslands. A combination of morphological, cytological, and biogeographicaldata and isozyme markers can determine whether taxa are auto- or allo-polyploid (Crawford, 1985; Haufler, 1985b; Bryan and Soltis, 1987; Weedenand Wendel, tg8g; Crawford, 1990; Pryer and Haufler, 1993). An overviewof the literature about DNA sequencing in Pteridophyta is given in Van denheede et aI . (2003).
Marnzunl RNr MnrHous
Between April 1995 and May 1999, field trips were organized to three CanaryIslands from which A. parvifolium was known in the literature: Gran Canaria,La Palma and Tenerife. From 145 specimens, fronds with ripe spores werecollected by C.V. and R.V, and ecological notes were made. Voucherinformation (Appendix 1, 2, and g) is given only for specimens from whichwe were able to raise progeny and obtain cytological data.
The following localities are shown in Fig. 1:
1) Gran Canaria, S of Moya, "Los Tilos" Reserve, W exposed slopes ofBarranco del Laurel, degraded laurel forest, in fissures of volcanic rocks;28 '05 '03 'N, L5o35 '28"w,6oo m a l t .
2) Gran Canaria, 4 km from junction Tejeda-San Mateo-Las Mesas, Eexposed slopes of "El Nieblo" Nature Reserve, in fissures of volcanicrocks; 2B"O'1,'03"N, 15'36',07"W, L550 m alt.
3) Gran Canaria, lava field near Cueva Corcho, along road GC110 fromArtenara to Valleseco, 4 km NW of junction Artenara-Valleseco-Tejeda, infissures of volcanic rocks; 1350 m alt.
4) Gran Canaria, 900 m S of Valsendero, W exposed cliff sides of narrow gullywith laurel forest remnants;28"O2'48"N, '1,5"34'27"W, 900 m alt.
5) La Palma, c. 3 km E of Tijarafe, Pinar Lomo del Hornoi 28"42'N, 1,7o55'W,1.140 m alt.
6) La Palma, S of Gallegos, Barranco Lomo de los Machines, Laurel forest Wof tunnel El Envetadero, E exposed slope; 2B.4B'N, 17o50'W, 390 m alt.
7) La Palma, volcanic rocks above roadside to Fuencaliente, S of Monte deLuna; 28"3l',N, L7o49',W, 71,O m alt.
B) La Palma, footpath to Monte de Luna in Pinar S of Flores, in fissures ofvolcanic rocks; 28"31,'N, 17"49'W, 810 m alt.

t
Tenerife
25 km-
La Palma
Gran Canaria
AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
Frc. 1. Map of the western Canary Islands with localities of voucher specimens (see alsoAppendix 1, 2, and 3).
9) La Palma, along track to Pinar de la Virgen at junction with track to CalderaLos Arreboles, in fissures of volcanic rocks; 28"30'N, 17o50'W, g2O m alt.
10) La Palma, along track to Refugio de Tigalate from Zona Recreativa Fuentede los Roques, above "Malpais" W of Monte de Luna, in fissures ofvolcanic rocks; 28"3'1,'N, L7o49'W, 'Lo7o m alt.
L1) La Palma, Caldera de Taburiente, track from La Cumbrecita to Hoyo de losPinos, Pinar in Barranco de la Faya, in fi.ssures of volcanic rocks; 28o43'N,17050'w, 1.200 m alt.
1,2) La Palma, Iava field 2.5 km E of EI Paso church, in fissures of volcanicrocks; 28"38'N, 17"51,'W, 800 m alt.
13) La Palma, Iava field E of El Paso, NE of Montaía Las Moraditas, in fissuresof volcanic rocks; 28o39'N, 17o50'W, 800 m alt.
14) Tenerife, along road from Vilaflor to Pico del Teide, ca. 8.1 km fromjunction Vilaflor-Santiago del Teide-La Orotava, under disc-like, SWexposed volcanic rocks; 2B'10'55'N, 16o39'L7uW,1850 m al t .
15) Tenerife, Barranco de las Gambuesas above Arafo, N exposed slopes;2Bo2O'29"N, L6o26'Oz"w, 710 m al t .
16) Tenerife, Barranco del Espigon de Tea, NE exposed slopes; 28"20'44"N,16"26'33"W, 825 m alt.
17) Tenerife, Montafla de la Hoya, ridge S of Las Manchas, above Ermita de laSanta Angel del Guardo, in fissures of volcanic rocks; 28o16'34'N,16"48'05"w, L', l-20 m alt.
18) Tenerife, Teno, Barranco head between Tierra del Trigo and Ruigomez,along track above Tierra del Trigo, 1.8 km NW of Ruigomez, NW exposedbasaltic slopes; 29o2o'48'N, L6"48'28"W,800 m alt.

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP
Tenerife, Pinar above Vilaflor, Bandes de Chasna, c. 2.5 km NNW ofVilaflor, E exposed, in fissures of volcanic (phonolite) rocks; z8oL0'48"N,16"38'36"w, L880 m al t .Tenerife, Pinar above Vilaflor, W exposed slopes of small Barranco, infissures of volcanic rocks; 28"1,0'48'N, L6o38'39"W, 1900 m alt.Tenerife, Barranco de Ia Piedra Cumplida above (NW) Arafo; 28"2'1.'L7"N,1,6 '26'05'W. 900 m al t .Tenerife, volcanic outcrop along footpath between Santiago del Teide andArguavo, SE of El Retamar, between Montana de la Hoya and La Hoya;28oL6'N, L6o48"w, 920 m al t .Tenerife, lava field NW of Montafra de las Flores, "Vuelta Grande", alongtrack from El Porti l lo del Rastrojo to Llanos del Hospital; 2B.LB"N,16o45'W, \41.o m alt.
24) Tenerife, Chio Street, direction Cafradas, Restaurant "De Evora".
Vouchers l isted in Appendix 1, 2 and 3, are deposited in the personalherbarium of Viane and Van den heede (including the T. Reichsteinherbarium), with duplicates in GENT.
Between 1,992 and 2001, R.V., C.V., and W. Bennert gathered additionalmaterial in Europe, Madeira, and Turkey (Appendix ). Voucher informationabout 108 Cypriot samples is published in Van den heede et al. (zOOZ). OurIiving European and Macaronesian Asplenium subg. Ceterach collectioncontained rp to 550 specimens.
AII material for this study has been cultivated in Ghent University BotanicalGarden (Belgium). Spores were sown on agar-solidified medium containinga nutrient solution recommended by Dyer (1979). The cultures were stored incontinuous light at room temperature. After formation of mature gametophytes,distilled water was added to achieve fertilization. If necessary, prothallia weretransplanted onto fresh agar. Young sporophytes were planted individually inpots kept in a temperate greenhouse (minimum temperature 1.2"C). An air- andwater-permeable soil mixture was required for these xerophytic rock ferns.Full-grown maturity was reached after approximately two years.
For chromosome counts, immature spore mother cells were fixed in the field,or in the greenhouse, using freshly prepared 3:1 absolute ethanol:glacial aceticacid, and stored at freezing-temperature until required. Acetocarmine squashpreparations were made as described by Heitz ( '1.925,1950) and Manton (rgso).
Photographs were taken with an Olympus BH2 phase contrast microscope.Preparations were made permanent by dehydrating cover slip and slide ingraded mixtures of acetic acid and absolute ethanol, followed by mounting inEuparal (T. Walker and H. Rasbach, pers. comm.). All permanent preparationsare kept in the Pteridological Section of the Department of Biology at GhentUniversity. Sixteen cytologically checked plants (five tetraploids identified asA. aureum, seven tetraploids identif ied as "A, parvifolium sensu Benl," andfour octoploids, (Appendix L, 2, 3) were used as standards to compare thenuclear DNA content of the remaining specimens by a flow cytometer(Partec PA-1), using the manufacturer's protocol (Partec GmbH, Mrinster,
8 7
1e)
20)
21)
22)
23)

AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
Nordrein-Westfalen, Germany). Both nuclei extraction solution and DAPIstaining dilution were provided by Partec (Germany).
Methods for making permanent epidermis preparations and for measuringstomatal guard cells and spores, are described by Viane (1990, 1992). Forexospore measurements untreated, fresh spores mounted in DePeX were used.Spore size is unaffected by DePeX, whereas in some other mounting media,e.g., glycerin-gelatin (Ormonde, 1990), spores expand by 5-L5 %. The values ofmicrocharacters are extracted from our regularly updated database, presentlycontaining 110 different specimens of the European "A, ceteracft" group, and56 specimens of the Macaronesian " A. aurelrm" group (raw data availableupon request).
Only vigorously growing plants were included in the allozyme study. Freshleaves from 105 Canarian (Appendix '1., 2, 3) and 22O European and Turkishspecimens (Appendix a) were collected in the greenhouse, where the fernswere growing under the same conditions. Sporulating fronds of similar agewere wrapped in wet tissue, stored in plastic bags, and kept refrigerated at 4'Cfor maximum 0.5-2 days (until extraction). Polyacrylamide gel electrophoresis(PAGE) was performed by C.V. at the laboratory of "General Botany and NatureManagement" of the Free University Brussels (Belgium). Starch gel electro-phoresis (SGE) was done by S.P. and E.P. in the "Departamento de BiologiaVegetal I" of the Universidad Complutense in Madrid (Spain). All specimensfrom the Canarian Archipelago were analysed by starch gel electrophoresis.
Equal amounts of tissue and extraction buffer were used to obtain uniformconcentrations of extracts. Cooling (+'C) was applied during both homogeni-zation and electrophoresis. Acquaah (fgg2) was consulted for the EnzymeCommission (E.C.) numbers.
PAGE procedures mentioned in Triest (1989) and Van den heede et al, (zOoz)were used, whereas SGE protocols followed Soltis et al. (1983) and Haufler(1985a). In a preliminary survey 19 enzyme systems (G-3PDH, G-6PD, GDH,IDH, MDH, ME, 6-PGD, SKDH, SOD, XDH, ACO, AAT, HK, PGM, B-EST, LAP,ALD, PGI, and TPI) were checked for polymorphism. Because the primary goalof this isozyme research was to test the hypothesis that tetraploid A. ceterachoccurs on the Canary Islands in addition to A. aureum and related taxa, it wasnecessary to identify unique "marker" alleles characterizing each species or itsprogenitors. Finally, only five enzyme systems were suitable: aspartateaminotransferase (AAT: GOT, E.C. 2.6.L.1), shikimate dehydrogenase (SkDH,E.C. 1.1.'J-..25), malic enzyme (ME, E.C. L.1.1.4o), phosphoglucose isomerase(PGI, E.C. 5.1.3.9), and tr iosephosphate isomerase (TPI, E.C. 5.3.1.1). AIIpictures and dried gels are kept in the Pteridological Section of the Departmentof Biology at Ghent University.
Band homologies were determined by running samples side-by-side on thesame gel (see Haufler et oI., 1995). Allelic variants within loci weredistinguished from the products of different loci by assuming that Áspleniumenzymes conformed to established models of organellar compartmentalization(Gottlieb,'1.982; Gastony and Darrow, L9B3; Soltis, 1986; Weeden and Wendel,19Bg). Presumed loci were numbered sequentially, with the most anodally

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP
(i.e., the fastest band) migrating one designated "1." Similarly, different allelesof the same gene locus (i.e., allozymes, Crawford, 1990) were denotedalphabetically with the most anodal being "o."
Sequencing work was done by C.V. at the Jodrell Laboratory in Kew (UnitedKingdom). Nineteen cytologically and isozymically interesting, vigorouslygrowing plants, including three A. aureum specimens (CVí 64, CV670, CV712)from Gran Canaria, Tenerife, and La Palma, one putative Á. ceterach fromTenerife (CVl87), and one octoploid from La Palma (CV709), were selected togenerate DNA sequences from the plastid trnL-trnFintergenic spacer. Europeanmaterial for comparison included two Á. javorkeanum specimens from Italyand Slovenia (CV14 and CVBSb), two Á. ceterach samples from Italy and Cyprus(CV494 and CV225), and a hexaploid Á. cyprium plant (CV249) from Cyprus(see Appendix 4). Sequences of the closely related A. Iolegnamense (Gibby andLovis) Viane from Madeira (CV9Bí and CV993), of the less related A.dalhousiae Hook. from Ethiopia and Pakistan (CV3LB and Tn7ffi4), and ofthe more distantly related A. nidusL. (AF425118), and Á. scolopendrium L. andA. unilaterale Lam. (R. Cranfill, University of California, Berkeley, California,USA, unpublished data) were included as outgroups. The trnL-F sequence ofa species of Dennstaedtia (R. Cranfill, unpublished data) was used to representa group basal to the Aspleniaceae (e.g., Bower, '1,928; Christensen, 1938;Copeland, '1.947; Pichi Sermolli, 1,977; Kramer and Green, Lgg0; Hasebe et aL,1995; Pryer et aL. ,1995). Methods are explained in Van den heede et a l . (2003).
Rnsur,rs nrun PnrlrMrNARy DrscussroN
To avoid prolixity, we have combined both the results and the interpretationof the isozyme phenotypes.
Chromosomes were counted for 16 specimens collected on Gran Canaria, LaPalma, and Tenerife. In addition to five tetraploid A. aureum (n --72") plants,eleven small specimens that we could not distinguish from European Á.ceterach, were examined. Seven of them turned out to be tetraploid (n -- 72tt)and four were octoploid, having a meiotic chromosome number of n -- 1,44rr(Fig. 2). Meiosis in all cells examined was regular, showing only bivalents, andgiving no indication about the polyploid status of the species. This agrees withLovis' (tgzz) statement that most autopolyploid ferns possess diploidizedmeiosis (only bivalent formation), and "that the absence of multivalents is novalid evidence of allopolyploidy."
The counted samples were used as standards to determine the ploidy level ofthe remaining 146 specimens using a flow cytometer. Results are given inAppendix 1., 2 and 3; localit ies of cytologically checked material are indicatedon the map of the Canarian Archipelago (Fig. 1).
In May 1995 (nvilSs) and 1997 (CV165-L70; CVLB3-1B7), we discoveredtetraploi d A. ceterach specimens on both Gran Canaria and Tenerife (seeAppendix 1 and 3).
The three species (A. aureum, the "small tetraploid", and the octoploid)cannot always be distinguished macromorphologically, but can be identified

AMERICAN FERN |OURNAL: VOLUME e4 NUMBER 2 (2004)
a
I tart
*'ïl itrr .ltt '
3.a t
1ïL
Io
I
I t ooa e l l l
i,r"D'
A t
Éfuàf:ii{.'ga'g *
t . t t4 $Jtk^.*:dltr
lt&ffit$:Í.tï
É'trjf
t ',
BtB
Ftc.2. Cytology showing spore mother cel ls in f i rs t meiot ic d iv is ion. A, B: photographs; A' , B ' :
explanatory diagrams with bivalents in black.A, A' : A. ceterach (CV 170b), metaphase I showing n:72t t . B, B' : A. octoplo ideum (CV 188,
holotype), cel l showingn:1,44t t . Scale bar: L0 pm. (preparat ions, photos and diagrams: C.V.) .
by measuring exospore length (Table Z). Stomatal guard cell length can only beused to distinguish the "small tetraploid" (+s * 3.8 pm) from the octoploid(sz * 4.8 pm). We found no differences in perispore morphology, stomatal type,or epidermal cell pattern. Perispores have costato-cristate folds with fewperforations, stomates are mesopolocytic, and epidermal cells mostly sinuous.Polyploidy factors (Viane, 1986, 1990) in Á. ceterach are P,"1, exo: t.ZS (for theexospore) and Pcet, sro: 1.L6 (for the stomates), and P"r., u*o:1.L8 and Pu,r., ,1o:1.07 in A. aureum. Using these P-values, the theoretical spore and stomate sizescalculated for the octoploid are less than 1 s.d. different from their actual means(Table 2), thus supporting the proposed ancestry (Viane, 1990).
We stress the presence of a small indusium in all taxa; it can best be observed inepidermis preparations (Viane, rggO). Our observations show that the (toothed)margin and the "cuticular lines" of the scales, are unreliable characters to

VAN DEN HEEDE ET AL.: CANARIAN ÁSPTEN/UM CETERACH GROUP
Taslr 2. Microcharacters differentiating taxa within the á. auteum-ceterach group on the CanaryIslands. All measurements are based on cytologically checked material. Additional informationabout material and the number of measurements is available from the authors.
Taxon PloidyMean exosporelength + s.d.
Mean guard celllength + s.d.
A. auremA. ceterachA. octoploideum
4x4x8x
32 + 1 .9 pm
39 + 2.6 pm
44 + 3.1 pm
46 + 4.4 pm
45 + 3.8 pm
52 + 4.8 pm
discriminate Á. oureum and relatives from A. ceteracà. AII taxa have scales withmore or less dentate margins, and periclinal cell walls with or without "cuticularIines". These "cuticular stripes" are folds in the periclinal cell wall (FiS. 3), andthe bigger the cell the more folds seem to be present. However, A. aureum scalesusually show numerous folds, whereas A. ceteracft (from its entire range ofdistribution) paleae possess only few. In the octoploid the number of folds isusually intermediate between that in Á. ouÍeum and Á. ceterach.
Isozyme analysis can be used to determine whether taxa are auto- orallopolyploid. The electrophoretic phenotype of an autopolyploid shouldshow a subset of the isozymes present in its progenitor, assuming no mutationsubsequent to the origin of the polyploid (Weeden and Wendel, 1989;Crawford, 1990; Pryer and Haufler, 1993). An allopolyploid should displayfixed heterozygous (i.e., nonsegregating) banding patterns for many loci,resulting from the combination of different parental genomes (Gottlieb, 1982;Werth, et aI. 1985b; Pryer and Haufler, 1993; Soltis and Soltis, 2000). Fixedheterozygous banding patterns differ from normal heterozygous zymograms,because the bands do not segregate among progeny and remain fixed in allspecimens.
Nineteen enzyme systems were tested in a preliminary survey. The lowresolution of ALD, and the smeared patterns of XDH made both unusable. IDH,
FIc. 3. Folds ("cuticular l ines")micrographs of laminal scales (CV
B a r : A : 5 0 p m , B : 1 0 p m .
in periclinal cell walls of A. aurcum157). A: scale margin with several cells.
paleae. Phase contrastB: single cell with folds.

AMERICAN FERN fOURNAL: VOLUME e4 NUMBER 2 (2004)
I u m l v v
+ a
b - r = : --
C - -
genotypcs:bbb bbbb aaanhbbb atrcc aou
tere:"srnall tctraploif 'srnall tetraploif 'octoploid' A. atrann Á, awanr
Á. ceterach A- ceterach
distribution:Canry Islands Canry lslands Cmry Islands Canry lslsrds Canry lslands
Europe EuopeTurlccy
Frc. 4. Diagrams of electrophoretic AAT phenotypes, showing 3 alleles and the correspondinggenotypes, as observed in the European-Canarian Asplenium ceterach-aureum group. Zymotype II
was also found in all samples of A. javorkeanum (bb) and Á. cyprium (bbbbbb) examined.
MDH, ACO, and SOD yielded unclear patterns with limited variation. G-6PDand G-3PDH gave inconsistent zymograms. Consequently, these enzymesystems were not retained.
To test our hypothesis that in addition to A. aureum and the octoploid, trueA. ceterocft occurs on the Canary Islands, the following four enzyme systemswere suitable: AAT, SkDH, ME, and PGI. These enzyme systems yieldedreproducible, well resolved banding patterns discriminating specimensrepresenting Á. oureum, A. ceterach and the allo-octoploid hybrid. Theseenzymes, encoded by four putative loci, were also used to get an idea of thevariation of the species.
AAT or GOT: this dimeric enzyme was studied using both PAGE and SGE(system B of Haufler, 1985a).We observed only one activity zone, which agreeswith Gastony and Darrow (rggg), who proved that this single enzyme activityis chloroplastic.
Most of the specimens studied are homozygous, showing a single well-resolved AAT band (Fig. 4, zymotype II). This applies to all 50 Á. javorkeanumspecimens (Italy and Slovenia), most (1.o2) A. ceterach plants (Belgium,Croatia, Cyprus, France, Italy, Slovenia, Spain, Turkey, and the UnitedKingdom), 23 "small tetraploids" from the Canary Islands, and all (aa) A.cyprium samples tested (Van den heede et ol. 2OO2). Ten A. ceterachindividuals from Croatia, France, Italy, and Slovenia, and two "smalltetraploids" from the Canary Islands, showed the rarer three-banded zymotypeI with skewed staining intensities, interpretable as heterozygous for a dimer.

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP
Corresponding genotypes are bb, bbbb, or bbbbbb for zymotype II of thehomozygous (diploid to hexaploid) plants, versus bbbc for zymotype I of theheterozygous tetraploid specimens.
Identical banding patterns in the Canarian "small tetraploid" and inEuropean A. ceterach plants indicate that true Á. ceterach is growing on theCanary Islands. Because neither European nor Canarian samples reveal uniqueelectrophoretic phenotypes, the Canarian populations do not seem to be ge-netically isolated. Though more sampling of A. iavorkeanum is needed, thesepreliminary results (Fig. 4, zymotype II) seem to confirm the autotetra-ploidy of A. ceterach.
The a allele (Fig. 4, zymotype V) can be used as a "marker" allelecharacterizing A. oureum. Some plants, showing a single band correspondingto genotype ooao, are homozygous, whereas others, with a balanced three-banded zymogram corresponding to genotype aecc, are heterozygous. Toexplain zymotypes I and IV, an extra genotype cc is postulated and expectedin Á. javorkeanum, which was studied only on the basis of Italian and Slovenianmaterial.
All 54 octoploid specimens (from Gran Canaria, La Palma, and Tenerife)show a monomorphic, presumably fixed heterozygous banding pattern ofgenotype aaaabbbb (Fig. 4, zymotype III), which we postulate to be derivedfrom a combination of zymotypes II and V (Fig. ). This would agree with thesuggestions of Reichstein (rge+) and Viane and Reichstein (rggz), that theCanarian octoploid is an allo-octoploid, which originated either by chromo-some doubling in an unknown tetraploid hybrid between A. oureum (withzymotype V) and A. ceteracft (with zymotype II), or via unreduced gametes ofeach species (Fig. 9). The fact that, in all the octoploids (from 15 differentlocalities) only one AAT zymotype was detected can be explained by thepreponderance of the "small tetraploid" with zymotype II. It may also reflectincomplete sampling of the variation present in the octoploid.
SkDH: resolution for this monomeric enzyme was superior on SGE (system 2of Weeden and Wendel, 19Bg). In our study, the enzyme was represented bya single locus, which agrees with Gastony and Darrow (tgAg). As expected fora monomeric enzyme, homozygotes had a typical one-banded pattern whereasheterozygotes showed two or more bands.
We detected four alleles in the European-Macaronesian Aspleniumceterach-oureum group. Two of these, o and b, were observed in Á. eureum,whereas c and d characterized the "Á. ceterach" group. Although SkDH waspolymorphic in Á. ceterach (Fig. 5), only zymotyp e V (cccd) was found on theCanary Islands. This two-banded pattern with unequal staining intensitiesforms also part of zymotype VI found for all 54 octoploid specimens. Thus theoctoploid is monomorphic and presumably heterozygous for this locus,showing a four-banded zymogram corresponding to genotype aabbcccd. Theo and b alleles are unique "marker" alleles for A. eureum, one of theprogenitors of the octoploid. This monomorphic pattern showing presumedfixed heterozygosity seems to confirm the putative allo-octoploid origin of thisspecies (Reichstein, 1984; Viane and Reichstein, 1.gg?). The cccd SkDH

AMERICAN FERN |OURNAL: VOLUME e4 NUMBER 2 (2004)
| i l I I I l v v v l v l l
+ a i ' -b i - - ' - -
iC - - - : - -.l - - -
| -
Seootypes: :M cccc ccdd cMd i. cccd oahlrccd ubh
itrn (óbreviated): i
i -srnall4x' -oooploid- .4. our.A. jo. Á. cet. A. cet. A. cet. i .4. cet.
:distribution: i
I t a l y C y p r u s l t a l y C r o t i a i C a n a r i a n A r c h i p e l a g oSlovcnia France Spain Slovenia
'-"Ó;á,iË"
ttaty Trrkey CyprusSlovcnia Italy
Ftc. 5. Diagrams of electrophoretic SkDH phenotypes, with corresponding genotypes, as observedin the European{anarian Asplenium ceterach-auÍeum group. Zymotype II was also found insome Á. javorkeanum (genotype: cc); zymotype V was present in all Á. cyprium (genotype: ccccdd)samples checked. The Canarian small tetraploid is abbreviated as: "small 4x."
genotype (zymotype V) of the "small tetraploids" is not limited to the Canaries,but was also found in Á. ceterach from Croatia, Cyprus, and ltaly. These resultsagain both prove the occurrence of A. ceterach on the Canary Islands, and thefact that the populations in this Archipelago are not genetically isolated. Threeadditional zymotypes were detected in continental Á. ceterach: a single-banded (cccc), a balanced two-banded (ccdd), and an unbalanced two-bandedpattern (cddd). The presence of the unbalanced patterns (cccd, cddd) intetraploid A. ceteroch at Skdà can be explained by tetrasomic inheritance (seediscussion). Diploid Á. javorkeanum from Italy and Slovenia showed a single-banded pattern of either genotype cc (zymotype II) or dd (zymotype I).
ME: this tetrameric enzyme was studied only by PAGE. The single enzymeactivity visible was shown to be cytosolic by Gottlieb (rggZ), Gástony andDarrow (rgag), and Soltis (rgao).
ME was monomorphic in each of the three species, and thus can be used todistinguish them from each other (FiS. 6). All Á. ceterach specimens (Europe)and "small tetraploids" (Canary Islands) were heterozygous showing anidentical five-banded zymogram, typical for a tetrameric enzyme controlled byone locus with two alleles, o and d. Heterozygous A. aureum was characterizedby
" five-banded pattern controlled by the same locus, but with two different
alleles, b and c, and corresponding to genotype bbcc. All octoploid plants

- aadd
c - addd
d - dddd
getrotypcxr:addd
tere:"gnall4x"A. caerach
distribution:C a n a r i a n
EuropeTrukcy
-
---
.I
---
A r c h i p e l a g o
ellaazebffiB
::::- bbbbtÉc
- aail- bcCC- bbbd- agr!,
- bbcc- a,dld- accc + bbdd- bCCC
r addd + cccc- bddd- Eod
- oodd
- cddd
- dddd
VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP
FIc. 6. Diagrams explaining the ME zymotypes showing 4 alleles, with corresponding genotypes,as observed in the European-Canarian Asplenium ceterach-aureum group. For each band fourIetters represent the association of subunits (coded by alleles) joined to form this tetramericenzyme. The homotetramers in the "hybrid" pattern are in boldface. Zymotype III confirms theallopolyploid status of the octoploid. Dotted lines indicate very faint bands. The Canarian smalltetraploid is abbreviated as "small 4x."
showed a complex zymogram, and conform to the expected hybrid phenotyperesulting from the cross between A. ceteracft and A. aureum. The "hybrid" hadthe four parental alleles, and since ME is a tetramer, each of the six pairs ofalleles (ax b, ex c, ax d, bx c, bx d, cx d) formed three heterotetramers ofintermediate mobility. Theoretically this results in a 22-banded pattern (4homotetramers plus 6 X 3 - 18 heterotetramers, makes 22 bands), but becausetwice two bands have the same mobility, a maximum of 20 bands was visible(Fig. 6). The monomorphic and presumably fixed banding pattern of the"hybrid" zymogram is in agreement with the putative allopolyploid origin ofthe octoploid (Reichstein, 1984; Viane and Reichstein, 1992).
PGI: this dimeric enzyme was studied by both PAGE and SGE. Because theresolution was much better with SGE, all results shown were obtained usingstarch gel electrophoresis (system 6 of Soltis ef o/., tgag).
Two loci were present: Pgi-I, most probably chloroplastic, and Pgi-2, cyto-

Pgi-2
+ a
I
aa
- a C
b
c - c . c
d
e
gcnotypes:aacc
texe:
Á. ceterrch
distribution:
Italy
IVNIu
- c c - c c - c c- C d - C d r C d
dd -csFdd - dd- : :
ccdd cdde ccccccdd
A. aureun A. auream "octoploid"
V V I V I I(rr x ru) (r x IIr) 2
a a _ a a
- a d
- d d
aaccccdd qaqadddd
"octoploid" "octoploid"
- a A
ad
- C C
- c d- d d
- @
cccc
"small 4x"Á. ceterrch
C a n a r i a nEurope
A r c h i p e l a g o
AMERICAN FERN JOURNAL: VOLUME e4 NUMBER 2 (2004)
FIc. 7. Diagrams of electrophoretic Pgi-2 phenotypes, showing 5 alleles, with correspondinggenotypes and their distribution, as observed in the European{anarian Asplenium ceterach-aureum group. Zymolype II was also found in most A. javorkeanum (cc) samples. Each band isindicated by two letters representing the association of subunits, joined to form this dimericenzyme. The Canarian small tetraploid is abbreviated as "small 4x."
solic (Gastony and Darrow, L983; Soltis, 1986). Consistent with observationson other ferns (Gastony and Gottlieb, tg85; Werth, 1991; Haufler et al.,19gb;Hauk and Haufler, 1999), resolution of the more anodal locus Pgi-1was inferiorto that of Pgi-2. Because Pgi-l appears invariant across all taxa, it will not bediscussed.
Among the European and Macaronesian samples studied, five allozymeswere observed at Pgi-2 (Fig. 7). Although the continental Á. ceterach, with itssix different zymotypes, was highly polymorphic for this locus (Van den heedeet al., 2OOz), only a single banding pattern was detected for the 22 "smalltetraploids" from the Canary Islands, corresponding to genotype cccc. Thiswidely distributed zymotype was also found in Á. ceterach specimens fromBelgium, Croatia, France, Italy, Slovenia, Spain, and the United Kingdom. Weobtained two electrophoretic phenotypes for the 28 A. oureum plants, withcorresponding genotypes ccdd (ZS specimens) and cdde (3 specimens). Theoctoploid was the most variable taxon in the Canarian Archipelago, showingthree different zymotypes translated into genotypes (Fig. 7) ccccccdd(zymotype v), oaccccdd (zymotype vI), and aaaadddd (zymotype vII). zymo-type V (found only on La Palma) most probably resulted from hybridization
VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP
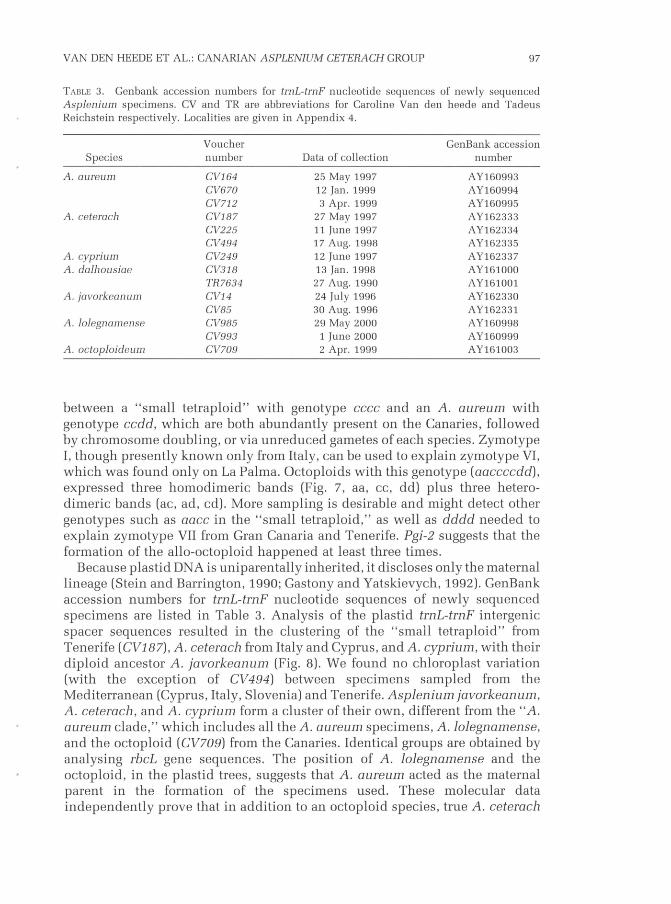
Tasl-e 3. Genbank accession numbers for trnL-trnF nucleotide sequences of newly sequencedAsplenium specimens. CV and TR are abbreviations for Caroline Van den heede and TadeusReichstein respectively. Localit ies are given in Appendix 4.
SpeciesVouchernumber Data of collection
GenBank accessionnumber
A. aureum
A. ceterach
A. cypriumA. dalhousiae
A. javorkeanum
A. Iolegnamense
A. octoploideum
25 IluÍay 1.99712 fan. 19993 Apr. 1999
27 May 1.99711 fune 199717 Aug. 19981.2ltne '1.997
13 |an. 199827 Aug. 19902a fu ly 199630 Aug. 199629 May 2000
1 fune 20002 Apr. 1999
4Y1609934Y1609944Y160995AY1623334Y1623344Y162335l'Y1.623374Y1610004Y1610014Y162330AY162331AY1609984Y1609994Y161003
cv164CV67OCV712cv187CV225CV494CV249CVS1BTn7ffi4CV14CVBsCVgB5cv993CV7O9
between a "small tetraploid" with genotype cccc and an Á. oureum withgenotype ccdd, which are both abundantly present on the Canaries, followedby chromosome doubling, or via unreduced gametes of each species. ZymotypeI, though presently known only from ltaly, can be used to explain zymotype VI,which was found only on La Palma. Octoploids with this genotype (aaccccdd),
expressed three homodimeric bands (Fig. 7, aa, cc, dd) plus three hetero-dimeric bands (ac, ad, cd). More sampling is desirable and might detect othergenotypes such as aocc in the "small tetraploid," as well as dddd needed toexplain zymotype VII from Gran Canaria and Tenerife. Pgi-2 suggests that theformation of the allo-octoploid happened at least three times.
Because plastid DNA is uniparentally inherited, it discloses only the maternalIineage (Stein and Barrington, 1990; Gastony and Yatskievych, 1.992). GenBankaccession numbers for trnL-trnF nucleotide sequences of newly sequencedspecimens are listed in Table 3. Analysis of the plastid trnL-trnF intergenicspacer sequences resulted in the clustering of the "small tetraploid" fromTenerife (CV187), A. ceterach from Italy and Cyprus, and A. cyprium, with theirdiploid ancestor A. javorkeanum (Fig. B). We found no chloroplast variation(with the exception of CV494) between specimens sampled from theMediterranean (Cyprus, Italy, Slovenia) and Tenerife. Asplenium javorkeenum,A. ceteracir, and A. cyprium form a cluster of their own, different from the "Á.oureum clade," which includes all the A. aureum specimens, Á. lolegnamense,and the octoploid (CV709) from the Canaries. Identical groups are obtained byanalysing rbcL gene sequences. The position of A. Iolegnamense and theoctoploid, in the plastid trees, suggests that Á. oureum acted as the maternalparent in the formation of the specimens used. These molecular dataindependently prove that in addition to an octoploid species, true Á. ceterach

AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
A. aurewn CVl64 Gran CanariaA. aureun CV670 TenerifeA. aurewnCVT|Z La PalmaA. Iolegnamense CV985 MadeiraA. I ole gnamen se CY 993 Madeiraoctoploid CV709 Le Pelma
Á. dolhousiae CV318 EthiopiaA. dalhousiae TR7634 Pakistan
A. i anrkestwn CY | 4 SloveniaA.j anr ke onan CV85 ItalyteÍa ploid- C_V l-87_Te ner ifeA, ceterachCVns Cyprus'A. celerach CV49a l-talyA. cypriumcYz4g Cyprus
A. scolopendriumA. nidus
A. unilaerale
hrutstaedlia - 5 changes
FIc. B. Tree randomly selected from the 73 shortest trees of European-Canarian Aspleniumceterach-aureum taxa and A. dalhousiae, resulting from parsimony analysis of our 1.4 trnL-trnFintergenic spacer sequences (Table 3) and 4 trnL-trnF sequences of other species available inGenBank; length :227 steps, CI :0.92, and RI:0.91. Based on rbcL evidence (Hasebe et a l . , 1995;Pryer et al., 1995), the sequence of the more distantly related DennstaedÍio was specified asoutgroup. Fitch branch lengths (ACCTRAN optimized) are shown above and bootstrap percentages(1000 replicates) below the branches. Other sequencing results are described extensively in Vanden heede et a l . (2003).
(: the "small tetraploid") is growing on the Canary Islands. Other sequencingresults are described extensively in Van den heede et al. (2003).
Because Gran Canaria, La Palma, and Tenerife are of volcanic origin, theseepilithic ferns mainly grow on rocks of basaltic types, like phonolites,rhyolites, trachytes and olivine basalts (Page, 1979). Asplenium oureumprefers moister, shady habitats at lower altitudes, whereas the "smalltetraploid" and octoploid plants share more exposed, drier habitats. However,on both Gran Canaria and Tenerife, only single localities were found where"small tetraploids" and octoploids grew together (loc. 2 and 19; see Appendix1 and 3). Though forty-five specimens from nine different localities on LaPalma were cytologically checked (see Appendix 2 and Van den heede andViane, unpublished data), we could not detect any "small tetraploid"specimen. We intensively looked for it in the field, especially in the higherregions of La Palma. Whereas we discovered "small tetraploids" between 1500and tg00 m altitude on Gran Canaria and Tenerife, the highest altitude wefound ferns of the Ceterach group on La Palma was near 1200 m.
We found Á. oureum between 300 and 1000 m altitude in valleys andsheltered ravines ("barrancos") with remnants of (degraded) evergreen laurel

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP
forest dominated by broad-leaved trees: Laurus azorica (Seub.) Franco, Perseaindica (L.) Spreng., Ocotea foetens (Aiton) Berthel., Apollonias barbujana(Cav.) Bornm., IIex canariensis Poir. , IIex platyphyllo Webb and Berthel., andArbutus canariensis Vieill. (Bramwell and Bramwell, 1.974). Aspleniumoureum usually grows in humus-rich soils, often together with Adiantumcapillus-veneris L., A. reniforme L., Anogramma leptophylla (L.) Link,Asplenium aethiopicum (Burm.f) Bech., A. hemionitis L., Cheilanthespulchella Bory ex Willd., Davollia canariensis (L.) Sm., PolypodiumcambricumL. ssp. mocaronesicum (A.E. Bobrov) Fraser-Jenk., and Selaginelladenticulafa (L.) Spring.
Asplenium ceteracft and A. octoploideum were found in the natural pineforests ("Pinar") at 900-2000 m on Tenerife, and at 1200-1600 m on GranCanaria. The octoploid was observed on La Palma at 700-1200 m. The opensavannah-like vegetation is dominated by Prnus canariensis C. Sm. and a fewshrubs, such as Adenocorpus foliolosus (Aiton) DC., Cistus symphytifoliusLam., Daphne gnidium L., Micromeria species, and Rumex lunaria L.(Bramwell and Bramwell, tgz+). Asplenium ceterach and the octoploid growin rock fissures, often together with Monanthes laxiflora (DC.) Bolle, Aeoniumspecies, Asplenium aethiopicum, A. trichomones L., Anogrammo leptophylla,Cheilanthes guanchico Bolle, C. pulchella, Cosentinia vellea (Aiton) Tod.,Notholaeno morontae (L.) Desv. subsp. subcordata (Cav.) G. Kunkel, andPolypodium cambricum ssp. mocoronesicum. Where A. ceterach and A.octoploideum grow together abundantly, we discovered their sterile hexaploidhybrid, A. Xchasmophilum Van den heede and Viane (Van den heede andViane, 2OO2)
DrscussroN
In combination with morphological, cytological, and biogeographical data,isozyme markers can determine whether taxa are auto- or allopolvploid(Crawford, 1985; Haufler, 1985b; Bryan and Soltis, 19B7; Weeden and Wendel,1989; Crawford, 1990; Gastony, 1990; Pryer and Haufler, 1993). Electrophoreticanalysis of isozymes is an ideal way to investigate the origin of allopolyploidtaxa because parental loci are expressed as stable marker bands in the progeny(Haufler, 1985b; Werth et al.,19B5b; Gastony, 1986). The potential of isozymedata to clarify relationships in fern complexes is dependent upon the degree ofdifferentiation among the ancestral genomes.
In the present study, four loci (Aat, Skdh, Me, and Pgi-2) showing a uniqueset of bands characterizing A. aureum and different from the banding patternsfound in European Á. ceterach, proved adequate to disentangle the"Ceterech"complex in the Canarian Archipelago.
AII zymograms present in the Canarian "small tetraploid", were alsoobserved in A. ceterach, and confirm that true Á. ceterach is growing on theCanary Islands. Moreover, this suggests an occasional spore flow from Europetowards the Canaries. A flow in the opposite direction is less likely because thewestern islands are dominated bv the northeast trade wind svstem. Conse-

100 AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
quently, the Canarian Á. ceterach population cannot be considered geneticallyisolated. No local zymotypes seem to have originated in this taxon in the CanaryIslands, contrary to the situation on Cyprus, which is much older than theCanary Islands (Van den heede et al., 2OO2). For example, the unique Tpi-2zymogram, present in all Á. ceterach and Á. cyprium specimens from Cyprus,suggests the local origin of the Cypriot taxa (Van den heede et al.,2OO2).
All four loci (Áof , Skdh, Me, and Pgi-2) of the Canarian octoploid showmonomorphic heterozygosity for a combination of the patterns seen in Á.ceterach and Á. aureum. Our allozyme data confirm the allo-octoploid natureof this species, which most probably originated by chromosome doubling ina tetraploid hybrid between A. aureum and A. ceteracft (Viane and Reichstein,1992). Theoretically, though less parsimoniously, the formation of this taxoncould also happen directly via the union of unreduced (ax) gametes (ongametophytes resulting from unreduced spores) of both species.
All allozymes observed in the allo-octoploid were electrophoreticallyidentical to those found in the parental tetraploids. However, in someoctoploid samples from Gran Canaria and Tenerrfe, Pgi-2 expressed a zymotype(corresponding to genotype aaaadddd) resulting from the combination of twoundetected genotypes (aaaa) in Á. ceterach and (dddd) in A. aureum. Theoccurrence of this putative "orphan" genotype may reflect incompletesampling of the variation present in the tetraploids, or alternatively thesegenotypes may no longer be present in extant A. ceteraclr and A. aurcumspecimens.
The variation in the allo-octoploid seems to be related to the mono- orpolymorphism (and its abundance) in the parental tetraploids. Thus, at the twoIoci (Skdft and Me) showing a single octoploid genotype, only one genotypewas observed in each of the Canarian parents. At Aat two different genotypeswere found for the Canarian A. ceteracft, though only a single zymotype wasdetected for the octoploid. However, the A. ceteracft genotype not detected inany octoploid, was found in only ca. 1.Oo/o of the population. On the otherhand, this may also be the result of limited sampling of the variation present inthe octoploid.
The allo-octoploid species showed three different isozyme profiles at Pgi-2,a locus that is polymorphic in its tetraploid progenitors, indicating that theoctoploid probably originated at least three times. According to Werlh et aI.(1985a) such patterns of variation in allopolyploids are almost certainly theresult of repeated allopolyploidizations involving pairs of different genotypes.Thus, each of the octoploid zymotypes may have arisen from a separatehybridization event. The present observations, demonstrating multiple originsof allopolyploids, are similar to those of Werth et al. (1985a, b) for Asplenium,Soltis et aI. (1987) for Polystichum, and Ranker et al. (rgag) for Hemionitis.Recurrent origins of the allo-octoploid species implicate a repeated gene flowfrom tetraploids to octoploids, and mean a continued gain of genetic diversityby the allopolyploid.
Our electrophoretic data also provide evidence for the operation oftetrasomic inheritance in natural populations of autotetraploid (Rasbach ef

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP
oI., '1.987) A. ceteroch. At Skdft, for which only two allozymes were observed,three types of heterozygotes were present: balanced heterozygotes (ccdd) andtwo types of unbalanced heterozygotes (cccd, cddd). The presence of thesethree types in tetraploid Á. ceterach at Skdft is suggestive of the three possibleclasses of heterozygotes expected in an autotetraploid at a locus having twoalleles (Weeden and Wendel rggg). Unbalanced staining activities indicatemultiple doses of individual alleles. Tetrasomic inheritance with chromatidalsegregation explains the arrays of homozygous, balanced heterozygous, andunbalanced heterozygous banding patterns observed in Á. ceterach (Weedenand Wendel tggg). Tetrasomic inheritance implies that a chromosome can pairwith any of its three homologous chromosomes (e.9., Soltis and Rieseberg,1986; Weeden and Wendel, tg8g; Crawford, 1990), and that there is apparentlyno strict preferential chromosome pairing. Consequently, the present isozymeanalysis confirms the autotetraploid status of A. ceterach, which wascytologically proven by Rasbach et al. (1982). The fact that isozyme studiespoint to tetrasomic inheritance and that we found only bivalents duringmeiosis in autotetraploid A. ceterach, suggests that both processes arecontrolled by different (sets ofl genes. Similar unbalanced patterns found inallotetraploid ferns have been explained also by segregating intralocusheterozygosity and fixed interlocus heterozygosity (Gastony, 1990).
We were able to prove, by isozyme and plastid DNA analysis, that inaddition to A. oureum and the octoploid, true A. ceteracft occurs on GranCanaria and Tenerife. A combination of morphological and cytologicalanalysis leads to correct determination, but even the exospore length aloneallows reliable identification of the three Canarian species: A. aureum (32 -+
1.9 pm), A. ceteraclr (gg -r 2.6 pm), and the octoploid (qq * 3.1 pm).As mentioned in the introduction, Benl and Kunkel (rg0z) published C.
aureumvar. paruifolium without cytological investigation. Plants collected in1.967 by T. Reichstein and G. Kunkel were found to be octoploid, leading T.Reichstein and other European pteridologists to attribute octoploid status to Á.parvifolium (including all small Canarian "Ceterach" specimens), but withouthaving checked the holotype.
We repeatedly visited the type locality of A. parvifolium, the Pinar aboveVilaflor (Tenerife), and found several taxa (see Appendix 3) growing together.As soon as we were convinced that two kinds of "small Ceterach" species weregrowing at the locus classicus (and on Gran Canaria), we decided to study theholotype (BenI s.n., 2611.211.966, M) of C. eureum var. parvifolium. Thisholotype consists of one single plant. We studied its microcharacters (see alsoTable 4) and found a mean exospore length (gg * 2.8 pm) and very few folds("cuticular lines") in the scales characteristic for true Á. ceterach. The valuesfor the exospore and stomate length (38 -r 3.7 pm) prove that the holotype wasnot an octoploid, but a tetraploid plant! Consequently, C. eureum var.parvifolium Benl and G.Kunkel and A. parvifolium are synonyms of Á.ceterach, and the octoploid had no correct name and was described as Á.octoploideum (Van den heede and Viane, 2OO2). Asplenium octoploideum ismorphologically intermediate between Á. eureum and Á. ceterach, from which
101

1.O2 AMERICAN FERN IOURNAL: VOLUME e4 NUMBER 2 (2004)
Tneln 4. Comparison of mean exospore length (LEXO) of various types to that of cytologicallychecked vouchers (see Table 2).
Taxon Voucher, status, and herbariumLEXO + s.d.
(types)LEXO + s.d.
(cytol. checked)
A. aureumA. ceterachA. parvifolium
A. octoploideum
Broussonet s. n., iso-:PHort. Cliff. Aspl. 4,lecto-: BMBenl s. n., holo-: MCV 188. holo-: GENT
32 + 1 .9 pm
39 + 2 .6 pm
44 + 3 .1 pm
it can be distinguished by its different mean exospore length (++ pm) andmean stomate length (SZ pm), and its octoploid chromosome number n :'l.44rr(Fig. 28 +B'). It is endemic to the Canarian Archipelago but presently confirmedonly (cytology) for Gran Canaria, La Palma, and Tenerife, and to be expected onLa Gomera and El Hierro. In addition to the holotype from Gran Canaria [avafield near Cueva Corcho, in fissures of volcanic rocks, 1350 m alt, 28th M"y1997, Ieg. Van den heede qnd Viane CV lSS (Holo-: GENT, iso-: personalherbarium of Viane and Van den heede)1, the following collections (paratypes)were also made (for localit ies see Appendix 1, 2, and 3): CV 171, CV 172, CV 173,cv 1.75, cv 176, cv 177, cv 179, cv 672A+8, cv 674, cv 686, cV 687, cV 695,cv 708, cv 709, cv 715, cv 716, cv 717, cv 718, cv 719A+8, cV 720, cV 72L,cv 723, cv 724, cv 725, cv 726, cv 727, cV 729, cV 730A+8, CV 731, CV 732,cv 733, cv 734, cv 740, cv 741, cv 742, cv 743, cv 744, cv 745, cV 746, cV747, cv 748, cv 749, cv 750, cv 751A+8, cV 752, CV 754A+B+C,WB 22/93.
Many herbarium specimens still need to be inspected before the ranges ofthe "small Canarian Ceteracft" taxa can be established. Literature referencesand information on herbarium labels are often unreliable, e.g., Bornmtiller3094 (P), labeled as C. officinorum f. typica (cellulis polearum non striatis!)collected on Gran Canaria, has a mean exospore length of q+ + 3.0 pm andnumerous folds in the scale cells: it is without any doubt an octoploid.
As far as is known, the octoploid is endemic to the Canary Islands, butbecause Madeira could also harbor this species (climatologically andtopographically), we studied 30 specimens from five Madeiran localit ies.However, all of them turned out to be hexaploid and were identified as Á.lolegnamense. Both in the Madeiran and the Canarian Archipelago thenortheast trade wind prevails during the year. This phenomenon may helpto explain the restricted range of several Macaronesian taxa, because mostpropagules fall into the Atlantic Ocean. Even when spores occasionally reachthe African continent, the Western Sahara and Mauritania offer no appropriatehabitats for these ferns, because of their ultra dry climate. The derivation wehypothesize for the allo-octoploid species is presented in Fig. 9.
It is generally accepted (e.g., Burchard, '1.929; Lems, 1960; Page, 1.923;Bramwell and Bramwell, '1,974; Page, '1.977, 1,979) that many of the Canarianferns are endemic relicts of the Tertiary fern flora that existed in southernEurope during the Miocene and Pliocene. These ferns form an important partof the original vegetation of the Canary Islands, especially of the evergreen
31 - f 1.6 pm
39 + 2 .8 pm
38 + 2 .8 pm
42 + 3.4 pm

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP 103
X
hybnidizationfollowed by
clromosome doublingOR
via unreduced gametes
-+
A. ceterachJJJJ
A. aureumJIXX
A. octoploideumJJJJJJXX
FIc. 9. Scheme of relationships explaining the origin of the Canarian Asplenium octoploideumbased on (micro)morphological, cytological and molecular data. Each capital represents one set of36 ancestral chromosomes. The fJ genome represents A. javorkeanum. Because molecular studies(Van den heede et al., 2003) suggest that Á. aureum is an allotetraploid, involving A. javorkeanum
0]) and "A. semi-alrreum" (XX, unknown) as ancestors, its genome formula is given as JfXX.
forests (Page, 1,977). Unfortunately, these habitats, if not totally destroyedtoday, are greatly endangered by modern tourism (building, water supply).Several mountain areas need further research, and new species and hybridsawait description. Undoubtedly, this Tertiary (fern) flora forms an irreplace-able genetic resource that should be conserved.
AcruowlnocMENTS
We are very grateful to L. Triest (Free University, Brussels), M. Chase (lodrell Laboratory, RoyalBotanic Gardens Kew, UK), and R. Johns (K) for providing laboratory facilities, and to the curatorsof various herbaria (BM, M, P) for permission to study type material. We thank R. Cranfill forproviding trnL-F sequences of some outgroups; G. Van der Kinderen for his technical assistance(PAGE) and for cultivating sporophytes; D. Rosseel for his help with photography and forcultivating sporophytes. We thank W. Bennert for extra material from Turkey, and P.S. and D.E.Soltis for discussions and encouragements, T. Ranker and an anonymous reviewer for constructivecomments on the manuscript. The management of Ghent Botanical Garden (Belgium) kindlyprovided greenhouse facilities.
Lrrrnarunn Clrno
AcquaaH, G. 1992. Pructical protein electrophoresis for genetic research. Dioscorides Press,Portland, Oregon.
Bnrqr, G. and G. KuNrsL. 'J.967. Ztn Taxonomie der Gattung Ceterach auf den Kanarischen Inseln.Ber. Schweiz. Bot. Ges. 77 :257-265.
Bnul, G. and E. R. SvnNrnrutus. 1970. Beitráge zur Kenntnis der Pteridophyten-Vegetation und -Flora

to4 AMERICAN FERN JOURNAL: VOLUME 94 NUMBER 2 (2004)
in der Kanarischen Westprovinz (Tenerife, La Palma, Gomera, Hierro). Nova Hedwigia2O:41,3462.
BtR, S. S., C. R. FnasEn-|ENKINS, and J. D. Lovts. 1,985. Asplenium punjabense sp. nov. and itssignificance for the status of Ceterach and Ceterachopsis. Fern Gaz. 13:54-63.
BoLLn, C. 1864. Die Standorte der Farrn auf den canarischen Inseln. lL Z. AIlg. Erdk. tz:z+9-282.BoRv os SnrNr-VINcaNr, f. B. G. M. '1.8O2. Essais sur les Isles Fortunées et I'antique Atlantide, ou
précis de I'histoire générale de l'archipel des Canaries. Baudouin, Paris.BowER, F. O. 1928. The Ferns (Filícales). Vol. il. The leptosporangiate ferns. Cambridge University
Press, Cambridge, England.BRavwELL, D. and Z. Bna\awstt, 1.974. Wild flowers of the Canary Islands. Stanley Thornes, Ltd.,
London.BRyaN, F. A. and D. E. Solrts. 1987. Electrophoretic evidence for allopolyploidy in the fern
Polypodium vírginianum. Syst. Bot. 12:553-561.BuncHRnn, O. 1929. Beitráge zur Ókologie und Biologie der Kanarenpflanzen. Bibliot. Bot. 98:
236-243.CavRNII-Les, A. I. 1801. De las plantas que el ciudadano Augusto Broussonet colectó en las costas
septentrionales de la Africa y en las Islas Canarias. Anales Ci. Nat. 4:52-'1.O9.CHnvRlrER, A. 1935. Les Iles du Cap Vert. Flore de I'Archipel. Rev. Bot. Appl. 15:733-1090.CHRtst, H. L897. Die Farnkra'uter der Erde. G. Fisher, Jena.CHRIst, H. 1900. Die Farnkrciuter der Schweiz. K.f. Wyss, Bern.CHntsrrNseru, C. 1938. Filicinae. Pp. 522-550, in F. Verdoorn, ed. Manual of Pteridology. Martinus
Nijhoff, The Hague.CoLI-ENntrn, S. 1985. An Íllustrated guide to the flowers of Soudi Arabia. MEPA: Meteorology and
Environmental Protection Administration, Kingdom of Saudi Arabia Flora Publication 1.Scorpion Publishing Ltd., London.
CopnlaNo. E. B. '1,947. Genera Fil icum. Chronica Botanica. Waltham. Massachusetts.CRawFonn, D. J. 1985. Electrophoretic data and plant speciation. Syst. Bot. 9:21.9-225.CRRwpoRn, D. J. 1990. Plant molecular systematics. Macromolecular approaches. fohn Wiley and
Sons, New York, New York.DarusnRrRu, P. 1961. Étrrdes Macaronésiennes. I. Géographie des cryptogames vasculaires. Agron.
Lusit. 23 :1.5'1.-1.81.DesvRux, N. A. 1827. Prodrome de la famille des Fougères. Mém. Soc. Linn. Paris 6:171-21.2.DvnR, A. F. 1979. The culture of fern gametophytes for experimental investigation. Pp. 253-305, rn
A. F. Dven, ed. Tfte experimental biology of ferns. Academic Press, London.Faenru, F. 1965. Secondo supplemento alle tavole cromosomiche delle Pteridophyta di Alberto
Chiarugi. Caryologia'l,8:675-7 31,GRSroNy, G. I. 1986. Electrophoretic evidence for the origin of fern species by unreduced spores.
Amer. I . Bot . 73:1563-1569.Gastoruv, G. J. 1988. The Pellaea glabella complex: electrophoretic evidence for the derivations of
the agamosporous taxa and a revised taxonomy. Amer. Fern J. 78:44-67.GRstoNv, G. J. L990. Electrophoretic evidence for allotetraploidy with segregating heterozygosity
in South African Pellaea rufa A. F. Tryon (Adiantaceae). Ann. Missouri Bot. Gard. 77:3o6-3 1 3 .
GRSroNy, G. J. and D. C. Dannow. 1983. Chloroplastic and cytosolic isozymes of the homosporousfern Athyrium filix-femina L. Amer. f . Bot. 70:1.4O9-1.41.5.
Gastoruy, G. f. and L. D. Gorrlme. 1985. Genetic variation in the homosporous fern Pellaeaandromedifolio. Amer. f. Bot. 72:257-267.
GRStoNy, G. f. and M. D. Wtnouau. 1989. Species Concepts in Pteridophytes: The treatment anddefinition of agamosporous ferns. Amer. Fern J. 79:65-77.
Gastottv, G. f. and G. YarsrrawcH. 1992. Maternal inheritance of the chloroplast and mitochondrialgenomes in Cheilanthoid ferns. Amer. f. Bot. 79:71,6-722.
GIBsy, M. and f. D. Lovrs. 1989. New ferns of Madeira. Fern Gaz. 1.3:285-29O.GottLIEn, L. D. 1982. Conservation and duplication of isozymes in plants. Science 216:373-38O.GRsutsR, W. 1980. Med-Checklist Notulae, 1. Willdenowia ' l.O:']-.3-21..
GRIRFrtus, M. 1997. Index of garden plants. MacMillan, London.

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETENACH GROUP 10s
HaruseN, A. 1969. Checklist of the vascular plants of the archipelago of Madeira. Bol. Mus. Munic.Funchal 24:1.-71.
HaNsEtt, A. and P. SuwoINc.1.g7g. Flora of Mocaronesia. ChecklÍst of vasculor plants,2. Revisededition. Port I. Botanical Garden and Museum, university of oslo, oslo.
Hasssn, M., P. G. wolr, K. M. PRvER, K. usnn, M. Iro, R. saxo, G. f. GasroNv, J. yorovava, |. R.MaNHaRt, N. Munarnm, E. H. CRaruE, C. H. HaurLen, and W. D. Haur. 19g5. Fern phylogenybased on rbcl- nucleotide sequences. Amer. Fern ]. 8b:134-181.
HAUFLER, C. H. 1985a. Enzyme variabil ity and modes of evolution in Bommeria (Pteridaceae). Syst.Bot . 10:92-104.
HRUFLER, C. H. 1985b. Pteridophyte evolutionary biology: the electrophoretic approach. Proc. Roy.Soc. Edinburgh 86:sts-323.
Haurran, C. H. 1997. Modes and mechanisms of speciation in Pteridophytes. Pp. 291-307, inK.Iwatsuki and P. Raven, eds. Evolution and diversification of land plants. Springer-Verlag,Tokyo.
HAUFLER, C. H., M. D. WIrunnau, and E. W. Rasn. 1995. Reticulate evolution in the Polypodiumvulgare complex. Syst. Bot. 20:89-109.
Haur, W. D., and C. H. Hnurr,rn. 1999. Isozyme variabil ity among cryptic species of Botrychiumsubgenus Botrychium (Ophioglossaceae). Amer. J. Bot. 86:614-633.
HErrz, E. 1925. Der Nachweis der Chromosomen. Vergleichende Studien iiber ihre Zahl, GróBe undForm im Pflanzenreich I. Z. Bot.18:625-681.
HBrrz, E. 1950. Elemente der BotanÍk. Eine Anleitung zum Studium der Pllanze durchBeobachtungen und Versuche an Crepis capillaris (L.) Wallr. Springer-Verlag, Wien.
HooKER, W. ]. 1860. Species Fil icum. Vol. l i l . W. Pamplin, London.Hooxrn, W. ]. 1861. The British Ferns; or, coloured figures and descriptions, with the needful
analyses of the fructification and venation, of the ferns of Great Britain and lreland,systematically ananged. Lovell Reeve, London.
HosHrzAKI, B. J. and R. C. Monar.r. 2001. Fern growers manual. Revised and expanded edition.Timber Press, Inc., Portland, Oregon.
|auaNomz, E. and R. Marne . 1,931.. Catalogue des plontes du Maroc (spermatophytes et Ptéridophytes).I. Ptéridophytes, Gymnospermes et Monocotylédones. Minerva, Alger, Algeria.
faras, |. and f. SuoutlEtt. 1.972. Atlas Floroe Europaeae 1. Pteridophyza. Comm. Mapping Fl. Europe& Soc. Biol. Fennica Vanamo. Helsinki.
KINc, M. 1993. Species evolution, the role of chromosome change. Cambridge University Press,Cambridge, England.
KRatranR, K. U. and P. S. GneEN. 1990. Pteridophytes and Gymnosperms. Pp. 1404 in K. Kubitzki,ed. Tàe families and genera of vascular plants. springer-Verlag, Berlin.
KRAMER, K. U. and R. L. L. Vtaur. 1990. Aspleniaceae. Pp. 52-56In K.U. Kramer and P.S. Green, eds.Pteridophytes and Gymnosperms. Springer-Verlag, Berlin.
KUHN, M. 1868. Filices Africanae. W. Engelmann, Leipzig.KururnL, G. 1965. Enumeración de los helechos (Pteridofítos) de Lanzarote y notas sobre su
distribución geográfica. Revista Mus. Canafio 26:Z-17.KuNrrL, G.1971. Lista revisada de los pteridofitos de las Islas Canarias. Cuad. Bot. Canarial,3:2146.LsNas, K. 1's58. Phytogeographic study of the Conary Islands. University of Michigan, Ann Arbor,
Michigan.Lrrras, K. 1960. Floristic botany of the canary Islands. sarracenia 5:1-94.Ltn, f. 1.967. Contributions to the flora of the Canary Islands. Universitatsforlaget, Oslo.LTtINAEUS, C. 1,753. Species Plantarum. L. Salvius, Stockholm.Lovrs, I.D. 1'977. Evolutionary patterns and processes in ferns. Pp.229415 in R.D. Preston and
H.W. Woolhouse, eds. Advances in botanical research. Academic Press, London.Lown, E. J. 1858. Ferns: British and Exotic. vol. v. Groombridge & Sons, London.LuEnssnN, C. 1889. Die Farnpflanzen oder Gefássbiindelkryptogamen (Pteridopyta). Pp. 1-906 in A.
Grunow, A. Fischer, F. Hauck, G. Limpricht, Ch. Luerssen, W. Migula, H. Rehm, P. Richterand G. Winter, eds. Dr. L. Rabenhorst's Kryptogamen-Flora von Deutschland, Oesterreich undder Schweiz. Eduard Kummer, Leipzig.
MaInn, R. 1952. Flore de l'Afrique du Nord (Maroc, Algérie, Tunisie, Tripolitaine, CyrénaiQue et

1 0 6 AMERICAN FERN fOURNAL: VOLUME e4 NUMBER 2 (2004)
Sahara). Volume l. Pteridophyto - Gymnospermae - Monocotyledonae: Pandanales, Fluviales,Glumiflorae. Paul Lechevalier, Paris.
MRNroN, I. 1950. Problems of cytology and evolution in the PteridophlZo. Cambridge UniversityPress, Cambridge, England.
MaNroN, L, I .D.Lovrs, G. Vrna and M. Grsev. 1986. Cytology of the fern f lora of Madeira. Bul l . Br i t .Mus. (Nat. Hist . ) Bot . 15:123-161.
MAvR, E. '1.542. Systematics and the origin of species. Columbia University Press, New York,
New York.Mayn, E. 2000. The biological species concept. Pp.'1,7-29 in Q. D. Wheeler and R. Meier, eds. Species
concepts and phylogenetic theory. A debate. Columbia University Press, New York, New York.MIron, J. 1865. Die hciheren Sporenpflonzen Deutschland's und der Schweiz. A. Felix, Leipzig.Mtma, |. 1866a. Die hóheren Sporenpflanzen Europa's und der Atlantis. Bot. Zeitung (Berlin)
24|1.37-1,41,.MtLon, J. 1866b. Materialien zur Beurtheilung der Darwinschen Theorie. Bot. Zeitung (Berlin)
51,:39741,1,.Mnoa, l. 1,867a. Das wesen der Farn-Flora der Atlantis. Bot. Zeitung (Berlin) 25:41,7423.
Mtma, I. 1867b. Fil ices europae et atlantÍdis, Asiae minoris et Sibiriae. A. Felix, Leipzig.MooRE, T. 1857. Index Filicum: a synopsis, with characters, of the genera, and an enumeration of
the species of ferns. W. Pamplin, London.OnvoNns, J. 1990. O genero Ceterach Willd. nas Ilhas Macaronésicas. Pp. 157-170 in f. Rita, ed.
Taxonomia, Biogeografia y Conservacion de Pteridofitos. IME, Palma de Mallorca, Spain.Pncs, C. N. 1973. Ferns, polyploids, and their bearing on the evolution of the Canarian flora. Pp.
83-88 in G. Kunkel, ed. Monographiae Biologicae Canarienses 4: Proceedings of the 1stinternational congress pro Flora Macaronesica. Excmo. Cabildo Insular de Gran Canaria, LasPalmas, Spain.
Pacs, C. N. 1977. An ecological survey of the ferns of the Canary Islands. Fern Gaz. 1,1.:297-31,1,.Pncn, C. N. 1979. Macaronesian heathlands. Pp. '1,1,7-1,23 in R.L. Specht, ed. Heathlands and
related shrublands of the world. Descriptive studies. Elsevier, Amsterdam, the Netherlands.Ptcst Sanuollr, R. E. G. 1.977. Tentamen Pteridophytorum genera in taxonomicum ordinem
redigendi . Webbia 31:313-512.PrcHr Snnuolr-r, R. E. G. 1979. A survey of the pteridological flora of the Meditenanean Region.
Webbia 34:L75-242.PRvER, K. M. and C. H. Hnupr-En. 1993. Isozymic and chromosomal evidence for the allotetraploid
origin of Gymnocarpium dryopteris (Dryopteridaceae). Syst. Bot. 1,8:1,5O-1,72.PnveR, K. M., A. R. Sr'arru, and |. E. Srcoc. 1995. Phylogenetic relationships of extant ferns based on
evidence from morphology and rbcl sequences. Amer. Fern |. 85:2o5-282.
QuÉzrr, P. and S. Snrura. 1.962. Nouvelle flore de l'Algérie et des régions désertiques méridionoles.Tome.I. Centre Nat. Recherches Sci., Paris.
RANKER, T. A., C. H. HAUFLER, P. S. Solrrs, and D. E. Solrrs. 1989. Genetic evidence for
allopolyploidy in the neotropical fern Hemionitis pinnatifida (Adiantaceae) and thereconstruction of an ancestral genome. Syst. Bot. 1,4:439447.
RasBACH, H., K. RRSBncH, and H. W. BENNEnT. 1987. X Asplenoceterach banancense Bennert etMeyer (Aspleniaceae, Pteridophyta) - Neufunde und cytologische Untersuchungen. Farn-blátter 1.7:3-1.6.
RstcHstnN, T. 1984. Aspleniaceae. Pp. 21,1,-275 in H.f . Conert, U. Hamann, W. Schultze-Motel andG. Wagenitz, eds. Gustav Hegi lllustrierte Flora von Mitteleuropo. Band 1, Pteridophya. P.Parey, Berlin.
RoMRRIZ, C. 1953. Flora da Ilha da Madeira. Pteridófitos. Revista Fac. Ci. Univ. Lisboa, Sér. 2, C, Ci.Na t . 3 :53 -112 .
SAUER, F. 1880. Catalogus plantarum in canariensibus insu/is sponte et subsponte crescentium.Hal le Univers i ty , Hal le 4.S. , Germany. - '
ScHruEtnER, G. 1892. The book of choice ferns for the garden, coltservatory, and stove. Vol. L L.Upcott Gill, London.
Stooret, M. A. 1989. Aspleniaceae. Pp. 19-26 in A.A. El-Gadi and A. El-Taife, eds. Floro of Libya.Al-Faateh University, Tripoli.

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP
SoLtts, D. E. 1986. Genetic evidence for diploidy in Equisetum. Amer. f. Bot. 73:908-913.SoLrrs, D. E., C. H. Haurlan, D. C. Dnnnow and G. f. GasroNv. 1983. Starch gel electrophoresis of
ferns: a compilation of grinding buffers, gel and electrode buffers, and staining schedules.Amer. Fern f. 73:9-27.
SoLrrs, D. E. and L. H. Rrnsnsanc. 1986. Autopolyploidy in Tolmiea menziesii (Saxifragaceae):
genetic insights from enzyme electrophoresis. Amer. f. Bot. 73:310-318.SoLrrs, P. S. and D. E. Sor-rrs. 2000. The role of genetic and genomic attributes in the success of
polyplo ids. Proc. Nat l . Acad. Sci . U.S.A.97:7O51.-5O57.SoLrrs, D. E., D. E. Solrrs and E. R. AlvnRsolr. 1,987. Electrophoretic and morphological
conformation of interspecific hybridization between Polystichum kruckebergii and P.
munitum. Amer. Fern f. 77:4249.STRFLEU, F. A. and R. S. CowaN. 1976. Taxonomic Literature-Volume I: A-G. Bohn, Scheltema &
Holkema. Utrecht.SrErN, D. B. and D. S. BannrNcroN. 1990. Recurring hybrid formation in a population of Polystichum
xpotteri: evidence from chloroplast DNA comparisons. Ann. Missouri Bot. Gard. zz:334-339.
Tnnntru-Bt-or, M.-L. 1946. Sur la flore ptéridologique des Iles Atlantiques. Mém. Soc. Biogéogr.
8 :325-347 .TRresr, L. 1989. Electrophoretic polymorphism and divergence in Noias marina L. (Najadaceae):
molecular markers for individuals, hybrids, cytodemes, lower taxa, ecodemes and
conservation of genetic diversity. Aquatic Bot. 33:301-380.VRN oeN HEEDE, C. J. 2003. A biosystematic study of Asplenium subgenus Ceterach (Aspleniaceae,
Pteridophyta) based on cytology, morphology, anatomy, isozyme analysis, and DNA
sequencing. PhD dissertation. Ghent University, Ghent, Belgium.
VaNoeNHEEDE, C.] . , S.Palanoru, E. PaNcun and R. L. L. VIaNE. 2OO2. A new species and a new hybr id
of Asplenium (Aspleniaceae) from Cyprus and evidence of their origin. Belg. |. Bot. 135:
92- ' t1.6.VRN nnN HEEDE, C. f. and R. L. L. VraNn. 2002. New species and new hybrids in Asplenium subgenus
Ceterach (Aspleniaceae). GEP News 9:1,4.VaN nnN HEEDE, C. I., R. L. L. VtaNr, and M. W. CHnse. 2003. Phylogenetic analysis of Asplenium
subgenus Ceterach (Pteridophyta: Aspleniaceae) based on plastid and nuclear ribosomal ITS
DNA sequences. Amer. J. Bot. 90:481-495.VTANE, R. L. L. 1.986. Taxonomical Significance of the leaf indument in Dryopteris (Pteridophyta): I.
Some North American, Macaronesian and European taxa. Pl. Syst. Evol. 1,53:77-1,O5.VraNn, R. L. L. L990. Epidermology of European ferns. Pp. 69-89 in f. Rita, ed. Taxonomia,
Biogeografia y Conservacion de Pteridofitos. IME, Palma de Mallorca, Spain.Vrarun, R. L. L. 1992. A multivariate morphological-anatomical analysis of the perispore in
Aspleniaceae. PhD thesis. Ghent University, Ghent, Belgium.Vrnrur, R. L. L., H. RaseacH, K. Rassacu and T. RnrcusrnrN. 1996. Observations on some ferns of Poros
and the adjacent parts of the Peloponnesus (Greece). Bocconea 5:279-3OO.VTANE, R. L. L. and T. RnrcusrnrN. 1992. Notes about Asplenium II: Some new names and
combinations in Ásplenium L. (Aspleniaceae, Pteridophyta). Biol. faarb. 59:157-165.
VTANE, R. L. L. and C. |. VnN DEN HEEDE. 2002. Subspecific names for the recently described new
Aspleniums from Cyprus. GEP News 10:5-6.VrnR, G. 1963. A new Asplenium (Sectio Ceterach) species and the problem of the origin oï Phyllitis
hybrido (Milde) C. Christ. Acta Bot. Acad. Sci. Hung. 9:1,97-21.5.Vroa, G. 1,972.Cytotaxonomy and genome analysis of the European ferns. Sy-p. Biol. Hung. 12:51-60.voN Bucu, L. 1828. Physicalische Beschreibung der Canarischen Inseln. Kon. Akademie der
Wissenschaften, Berlin.WEeonN, N. F. and |. F. WnNnu,. 1989. Genetics of plant isozymes. Pp. 46-72 rn D.E. Soltis and P.S.
Soltis, eds. lsozymes in Plant Biology. (Advances in Plant Sciences Series, Vol. +).Dioscorides Press, Portland, Oregon.
WnRrH, C. R. 1991. Isozyme studies on the DryopterÍs"spinulosa" complex,I: the origin of the Log
Fern Dryopteris celsa. Syst. Bot. 1,6:44646'1,.WERTH, C. R., S. E. Gurruaru, and W. H. EsnsaucH. 1985a. Recurring origins of allopolyploid species
in Asplenium. Science 228:731,-733.
'1,o7

108 AMERICAN FERN JOURNAL: VOLUME e4 NUMBER 2 (2004)
WERTH, C. R., S. E. GurruRN, and W. H. EsHsRucH. 1985b. Electrophoretic evidence of reticulateevolution in the Appalachian Asplenium complex. Syst. Bot. i.0:184-792.
WtrLonttow, C. L. 1810. Species Plantarum. Editio quarta. Tomus V, 1. G.C. Nauk, Berlin.WooD, I. R. L 'l,gg7. A handbook of the Yemen /oro, Royal Botanic Gardens, Kew.
Appnruux 1. Vouchers from Gran Canaria, with corresponding taxa, locality numbers, andchromosome numbers. CV is the abbreviation for Caroline Van de heede. The description of thelocalities and their number is given in Material and Methods. For samples with chromosomescounted, the number of bivalents is given. Counted specimens served as standards to determine theploidy level (indicated by 4x and 8x) by flow cytometry. All specimens were used for isozymeanalvsis.
Vouchernumber Taxon Localitv Date of collection
Meiotic chromosome number(n), or ploidy
CV 157 A. aureumCV 158 A. aureumCV 159 A. aureumCV 160 A. aureumCV 161 A. aureumCV 162 A. aurcumCV 163 A. aureumCV 164 A. aureumCV 165 A. ceterachCV 166 A. ceterachCV 167 A. ceterachCV L68 A. ceteruchCV 169 A. ceterachCV 17Oa A. ceterachCV 17Ob A. ceterachCV 171 A. octoploideumCV 172 A. octoploideumCV 173 A. octoploideumCV 175 A. octoploideumCV 176 A. octoploideumCV 177 A. octoploideumCV 179 A. octoploideumCV 188 A. octoploideumCV 180 A. aureumCV 181 A. aureumCV 182 A. aureum
25 May 199725 May 199725 May 'I.997
25 May 199725 May 799725 May 199725 May 199725 May 199725 May 199725 May 199725 May 199725 May 199725 May 199725 May 199725 May 199725 May 1.99725 May 1.99726 May 199726 May 199726 May 199726 May 199726 May 199728 May 199726 May 199726 May 199726 Mav 1997
7ztr4x4x72rr72II
4x4x72rl72rr4x4x
4x4x72rr8x8x8x8x8x8x8x
744rr4x4x

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP 109
Appnuux 2. Vouchers from La Palma, with corresponding taxa, locality numbers, andchromosome numbers. CV is the abbreviation for Caroline Van den heede. The description ofthe localities and their number is given in Material and Methods. For samples with chromosomescounted, the number of bivalents is given. Counted specimens served as standards to determine theploidy level (indicated by 4x and 8x) by flow cytometry. All specimens were used for isozymeanalvsis.
Vouchernumber Taxon Localitv Date of collection
Meiotic chromosome number(n), or ploidy
CV 708 A. octoploideumCV 709 A. octoploideumCV 711 A. aureumCV 712 A. aureumCV 713 A. aureumCV 715 A. octoploideumCV 716 A. octoploideumCV 717 A. octoploideumCV, 7L8 A. octoploideumCV 719A A. octoploideumCV 7198 A. octoploideumCV 720 A. octoploideumCV 721 A. octoploideumCV 723 A. octoploideumCV 724 A. octoploideumCV 725 A. octoploideumCV 726 A. octoploideumCV 727 A. octoploideumCV 729 A. octoploideumCV 730A A. octoploideumCV 7308 A. octoploideumCV 731 A. octoploideumCV 732 A, octoploideumCV 733 A. octoploideumCV 734 A. octoploideumCV 740 A. octoploideumCV 741 A. octoploideumCV 742 A. octoploideumCV 743 A. octoploideumCV 744 A. octoploideumCV 745 A. octoploideumCV 746 A. octoploideumCV 747 A. octoploideumCV 748 A. octoploideumCV 749 A. octoploideumCV 750 A. octoploideum
c
56667788B8IIII
1010101.1.'1.1.
1.1.1.21.21.21.21 31,31 313L313131313131 3
2 Apr. 19992 Apr. 19993 Apr. 19993 Apr. 19993 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19994 Apr. 19995 Apr. 19995 Apr. 19995 Apr. 19995 Apr. 19995 Apr. 19995 Apr. 19995 Apr. 1999I Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. 19999 Apr. L999
8x'l,44rr
4x4x
8x8x8x8x8x8xBx8x8x8x8x8x8x8x8x8x
1.44tr8x8x8x8x8xBx8xBx8x8x8x8x8x8x
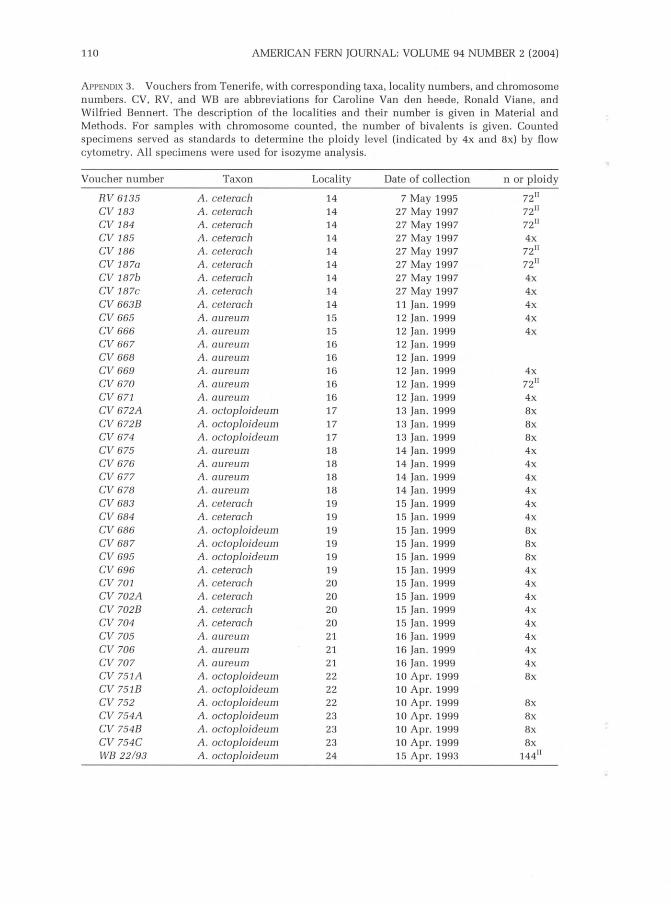
1 1 0 AMERICAN FERN fOURNAL: VOLUME e4 NUMBER 2 (2004)
AppsNInIx 3. Vouchers from Tenerife, with corresponding taxa, locality numbers, and chromosomenumbers. CV, RV, and WB are abbreviations for Caroline Van den heede, Ronald Viane, andWilfried Bennert. The description of the localities and their number is given in Material andMethods. For samples with chromosome counted, the number of bivalents is given. Countedspecimens served as standards to determine the ploidy level (indicated by 4x and 8x) by flowcytometry. All specimens were used for isozyme analysis.
Voucher number Taxon Locality Date of collection n or ploidy
RV 613scv 183cv 184cv 185cv 186CV 187acv 187bCV 1B7ccv 6638cv 665cv 666cv 667cv 668cv 669cv 670cv 671cv 672ACV 6728cv 674cv 675cv 676cv 677cv 678cv 68scv 684cv 686cv 687cv 69scv 696cv 701cv 702Acv 7028cv 704cv 705cv 706cv 707cv 7s1ACV 7518CV 752cv 754Acv 7s4Bcv 754cwB 22/9s
A. ceterachA. ceterachA. ceterachA. ceterachA. ceterachA. ceteruchA. ceterachA. ceterachA. ceterachA. aureumA. auteumA. aureumA. aureumA. aureumA. auteumA. aureumA. octoploideumA. octoploideumA. octoploideumA. aureumA. aureumA. auteumA. aureumA. ceterachA. ceterachA. octoploideumA. octoploideumA. octoploideumA. ceterachA. ceterachA. ceterachA. ceterachA. ceterachA. aureumA. aureumA. aureumA. octoploideumA. octoploideumA. octoploideumA. octoploideumA. octoploideumA. octoploideumA. octoploideum
1.41.4141.4741.47474741 51 516161 61 61 6' t 7
7 71,71 81 81 81 81 91 91 91 91 91 9202020202't2',t2 72222222 3232 324
7 May 199527 May 799727 May 799727 May 1.99727 IN.4.ay 799727 May 1.99727 May 799727 May 1.99711 Jan. 199912 fan. 199912 Jan. 199912 fan. L99912 fan. 199912 fan. 199912 )an. 199912 fan. 199913 fan. 199913 fan. 199913 fan. 199914 fan. 199914 fan. L99914 Jan. 1.99914 fan. 199915 fan. 199915 fan. 199915 fan. 199915 fan. 199915 fan. 199915 fan. 199915 lan. 199915 Jan. 199915 Jan. 199915 Jan. 1.99916 Jan. 199916 Jan. 199916 Jan. 199910 Apr. 199910 Apr. 199910 Apr. 199910 Apr. 199910 Apr. 199910 Apr. 199915 Apr. 1993
72TI72TT72TT4x72rr72IT4x4x4x4x4x
4x72rr4x8x8x8x4x4x4x4x4x4x8x8x8x4x4x4x4x4x4x4x4x8x
8x8x8x8x
'l.44rr

VAN DEN HEEDE ET AL.: CANARIAN ASPLENIUM CETERACH GROUP 771
Appexox 4. Alphabetical list of material for comparison used in this study. CV, RV, TR, and WB areabbreviations of C. Van den heede, R. Viane, T. Reichstein, and W. Bennert. Vouchers are depositedin GENT and in our personal herbarium at Ghent University. Additional information about locali-ties is available from the first and the last author ([email protected]; [email protected]). Voucher information about 108 Cypriot samples is given in Van den heede et al. (Zooz).
Asplenium ceterach:
CV25b+cCV25b, CVS0a+b, CV31CV36a+b, CV37cv78bcv41,cv4 2b, cv44,cv48, cv49bcv64, cv65, cv66, cv67, cV494cv225CV275a+b, CV276, CV277cv27g, cv279
cv429, cv430, cv431cv445, cv448, cV449cv450, cv451.cv657, cv658, cV659
cv77scv774, cv775, cv776nv'900WBlb+e/97wB10d/s7WBsc/97WB12c+d/97
Asplenium cyprium:
cv21.3cv24e
Asplenium dalhousiae
cv718TR7634
Asplenium jovorkeanum :cv7, cv4, cvíCVTa+b+c, CVSa+b+c
cv86, cv87, cv88cv89, cv90, cv91, cv92, cv93CVgqa+b, CV95cv404, cv405cv410cv480cv483, cv4B4cv504, cv506
Asplenium lolegnamense :
CV985cv993
CV767, CV768, CV769, CV770, CV771 Spain, Alava
Slovenia, KalCroatia, RoóCroatia, BassaniaSlovenia, KorteItaly, Valle della MarossaItaly, Termine di RoveranoCyprus, Troodos Mts., ChandriaBelgium, MarcourtItaly, Cannero Riviera
Crotia, LosinjItaly, BercetoItaly, BoioSpain, Torrelodones
France, CoulgensFrance, PaulmyFrance, NNE of Montpellier, La PeneTurkey, KaraobaTurkey, ManisaTurkey, OkEularTurkey, Mugla
Cyprus, Troodos Mts., Tsakistra-Vroiska roadCyprus, Kyrenia Mts., Kyrenia-Kythrea road
Ethiopia, Harerge Province, Asbe TeferiPakistan, Swat Province, Ambela
Italy, StupizzaSlovenia, 1 km E of Baóa towards Podbrdo
Italy, Monte Freddone, 730mItaly, Monte Freddone, 1320mItaly, Monte Freddone, 970mSlovenia, Bovec-Kobarid roadSlovenia, LjubinjItaly, N slope of Pania SeccaItaly, Fosso di AntonaItaly, E slope of Monte Corchia
Madeira, SW slope of Pico RuivoMadeira, N of Serra de Agua
CV2S1a*b, CV282, CV283, CV285, CV286 United Kingdom, Wales, Snowdonia
cv10, cv11, Cv12, Cv74, Cv472, Cv414 Slovenia, Baóa-valley, KneÍa-Klavze roadCV 2 0a+ b+ c+ d, CV2 1 a+b+ c+ d+ e Slovenia. MatavunCV81, CV82a+b, CV83, CVB4, CVsSalb ltaly, Arni




















