
As, Pb, Sb, and Zn transfer from soil to root of wild rosemary: do native symbionts matter?
18
REGULAR ARTICLE As, Pb, Sb, and Zn transfer from soil to root of wild rosemary: do native symbionts matter? Marie-Cécile Affholder & Anca-Diana Pricop & Isabelle Laffont-Schwob & Bruno Coulomb & Jacques Rabier & Andreea Borla & Carine Demelas & Pascale Prudent Received: 23 January 2014 /Accepted: 1 May 2014 /Published online: 28 May 2014 # Springer International Publishing Switzerland 2014 Abstract Background and aims This is an in natura study aimed to determine the potential of Rosmarinus officinalis for phytostabilization of trace metal and metalloid (TMM)-contaminated soils in the Calanques National Park (Marseille, southeast of France). The link be- tween rosemary tolerance/accumulation of As, Pb, Sb, and Zn and root symbioses with arbuscular my- corrhizal (AM) fungi and/or dark septate endophytes (DSE) was examined. Methods Eight sites along a gradient of contamination were selected for soil and root collections. TMM con- centrations were analyzed in all the samples and root symbioses were observed. Moreover, in the roots of various diameters collected in the most contaminated site, X-ray microfluorescence methods were used to determine TMM localization in tissues. Results Rosemary accumulated, in its roots, the most labile TMM fraction in the soil. The positive linear correlation between TMM concentrations in soil and endophyte root colonization rates suggests the involve- ment of AM fungi and DSE in rosemary tolerance to TMM. Moreover, a typical TMM localization in root peripheral tissues of thin roots containing endophytes forming AM and DSE development was observed using X-ray microfluorescence. Conclusions Rosemary and its root symbioses appeared as a potential candidate for a phytostabilization process of metal-contaminated soils in Mediterranean area. Keywords Trace metals and metalloid multicontamination . Arbuscular mycorrhizal fungi . Dark septate endophytes . Phytostabilization . μXRF analyses Abbreviations TMM Trace metals and metalloids AM Arbuscular mycorrhizal DSE Dark septate endophytes BCF Bioconcentration factor CF Contamination factor PLI Pollution load index Plant Soil (2014) 382:219–236 DOI 10.1007/s11104-014-2135-4 Responsible Editor: Peter Christie. Electronic supplementary material The online version of this article (doi:10.1007/s11104-014-2135-4) contains supplementary material, which is available to authorized users. M.<C. Affholder : A.<D. Pricop : B. Coulomb : C. Demelas : P. Prudent (*) Aix Marseille Université, CNRS, Laboratoire de Chimie de l’Environnement (LCE), FRE 3416, 3 place Victor Hugo, Case 29, 13331 Marseille cedex 03, France e-mail: [email protected] A.<D. Pricop : I. Laffont-Schwob : J. Rabier : A. Borla Aix Marseille Université, Avignon Université, CNRS, IRD, Institut Méditerranéen de Biodiversité et d’Ecologie marine et continentale (IMBE), 3 place Victor Hugo, Case 4, 13331 Marseille cedex 03, France
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of As, Pb, Sb, and Zn transfer from soil to root of wild rosemary: do native symbionts matter?

REGULAR ARTICLE
As, Pb, Sb, and Zn transfer from soil to root of wild rosemary:do native symbionts matter?
Marie-Cécile Affholder & Anca-Diana Pricop & Isabelle Laffont-Schwob &
Bruno Coulomb & Jacques Rabier & Andreea Borla & Carine Demelas &
Pascale Prudent
Received: 23 January 2014 /Accepted: 1 May 2014 /Published online: 28 May 2014# Springer International Publishing Switzerland 2014
AbstractBackground and aims This is an in natura study aimedto determine the potential of Rosmarinus officinalisfor phytostabilization of trace metal and metalloid(TMM)-contaminated soils in the Calanques NationalPark (Marseille, southeast of France). The link be-tween rosemary tolerance/accumulation of As, Pb,Sb, and Zn and root symbioses with arbuscular my-corrhizal (AM) fungi and/or dark septate endophytes(DSE) was examined.Methods Eight sites along a gradient of contaminationwere selected for soil and root collections. TMM con-centrations were analyzed in all the samples and root
symbioses were observed. Moreover, in the roots ofvarious diameters collected in the most contaminatedsite, X-ray microfluorescence methods were used todetermine TMM localization in tissues.Results Rosemary accumulated, in its roots, the mostlabile TMM fraction in the soil. The positive linearcorrelation between TMM concentrations in soil andendophyte root colonization rates suggests the involve-ment of AM fungi and DSE in rosemary tolerance toTMM. Moreover, a typical TMM localization in rootperipheral tissues of thin roots containing endophytesforming AM and DSE development was observed usingX-ray microfluorescence.Conclusions Rosemary and its root symbioses appearedas a potential candidate for a phytostabilization processof metal-contaminated soils in Mediterranean area.
Keywords Tracemetals andmetalloidmulticontamination . Arbuscular mycorrhizal fungi .
Dark septate endophytes . Phytostabilization .μXRFanalyses
AbbreviationsTMM Trace metals and metalloidsAM Arbuscular mycorrhizalDSE Dark septate endophytesBCF Bioconcentration factorCF Contamination factorPLI Pollution load index
Plant Soil (2014) 382:219–236DOI 10.1007/s11104-014-2135-4
Responsible Editor: Peter Christie.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11104-014-2135-4) contains supplementarymaterial, which is available to authorized users.
M.<C. Affholder :A.<D. Pricop : B. Coulomb : C. Demelas :P. Prudent (*)Aix Marseille Université, CNRS, Laboratoirede Chimie de l’Environnement (LCE),FRE 3416, 3 place Victor Hugo, Case 29,13331 Marseille cedex 03, Francee-mail: [email protected]
A.<D. Pricop : I. Laffont-Schwob : J. Rabier :A. BorlaAix Marseille Université, Avignon Université,CNRS, IRD, Institut Méditerranéen de Biodiversitéet d’Ecologie marine et continentale (IMBE),3 place Victor Hugo, Case 4, 13331Marseille cedex 03, France

Introduction
Trace metals and metalloids (TMM) are naturally pres-ent in the environment due to their occurrence in theEarth’s crust. However, for several years, high concen-trations (in comparison with background levels) havebeen detected, mainly in area utilized for agriculturaland/or industrial activities (He et al. 2005; Wuana andOkieimen 2011), and these may lead to major environ-mental issues. Depending on their speciation whichinfluences their mobility and bioavailability (Kabata-Pendias 2004), TMM can be disseminated in the variousecosystem compartments including plants. Indeed,plants are able to assimilate and, for some species, tohyperaccumulate TMM (Zheng et al. 2007), makingthem interesting to manage contaminated sites. Thus,in phytoremediation processes, plant and their associat-ed microorganisms can be used for TMM stabilizationor extraction from soil (Pilon-Smits 2005). Amongstassociated microbes involved in phytoremediation pro-cesses, root symbioses, especially those with arbuscularmycorrhizal (AM) fungi, have a significant effect ontolerance and absorption of TMM in plants (Gohre andPaszkowski 2006). In addition, these symbioses pro-mote plant nutrition, particularly phosphorus, and there-fore plant growth and health (Harrison 1999; Barea et al.2002). Plants appear as well to form symbiotic associa-tions with fungal endophytes, and among diverse groupsof endophytic fungi, dark septate endophytes (DSE)have received much attention in the recent years (Liet al. 2011). DSE are ubiquitous root-associated fungithat form inter- and intracellular melanized hyphaewhich can aggregate and form groups of thick-wallednodules (called microsclerotia) in epidermis and cortexof roots. They can form mutualistic, mycorrhiza-likerelationship though their relationships with their hostare not clear and seem to go from parasitic to mutualisticaccording to the species involved and the environmentalconditions (Jumpponen 2001). However, their presencein disturbed environments suggests a role in plant pro-tection in case of abiotic stress, such as TMM stress(Regvar et al. 2010).
Phytoremediation may be part of the solution fordispersed pollution as a non-invasive and ecologicalalternative method (Moreno-Jimenez et al. 2011). Itseems to be the most adapted choice for the particularcase of the contamination from the past industrial activ-ity of the former smelter factory of l’Escalette (southeastof Marseille, France), located in the Calanques National
Park. In this smelter factory, which ceased its ac-tivity in 1925, galena was converted into lead andsilver. As a consequence, this former industrial siteis still polluted by TMM, especially As, Pb, Sb,and Zn (Testiati et al. 2013). Moreover, the occur-rence of these trace elements was detected quite allover Marseilleveyre hills (Laffont-Schwob et al.2011). Today, neither the area affected by its activ-ity nor even a part of this area has been reclaimed,and in this context, the containment of pollution ofthis site must be a first priority. Since the pollutedarea is located on a protected zone for its plantspecies richness, non-intrusive solutions have tobe considered. Autochthonous plant species maybe evaluated for heavy metal stabilization or extrac-tion (Rabier et al. 2007). Previous studies (Calaet al. 2005; Sekeroglu et al. 2008; El-Rjoob et al.2008; Testiati et al. 2013) showed that Rosmarinusofficinalis is tolerant to very high TMM concentra-tions and could be used for phytostabilization ofamended soils polluted with trace elements(Madejon et al. 2009). Moreover, Affholder et al.(2013) indicated a limited health risk of rosemaryconsumption as medicinal (herbal tea) or aromatic(dry leaves) herbs, from plants grown on a highlycontaminated soil in the Calanques hills, since thisplant species does not accumulate high concentra-tions of As, Pb, Sb, and Zn in its aerial parts.These criteria are interesting for a phytostabilizationprocess.
Besides, as more than 80% of plant species (Harrison1999), it has been proven that rosemary forms rootsymbioses with AM fungi (Estaún et al. 1997;Sánchez-Blanco et al. 2004; Turrini et al. 2010;Sánchez-Castro et al. 2012), particularly with the genusGlomus (Turrini et al. 2010; Sánchez-Castro et al.2012).
Thus, this study aimed at determining the innatura ability of rosemary to transfer TMM in itsroots, namely As, Pb, Sb, and Zn which are majorcontaminants from past activity of the smelting fac-tory of l'Escalette, and to study the involvement ofroot symbiosis in tolerance and accumulation abili-ties of this species. Therefore, TMM accumulation inrosemary roots spontaneously grown in soils present-ing a gradient of contamination, localization of TMMin root, and pattern of adsorption was determined.Moreover, root colonization rate by AM fungi and/orDSE was estimated.
220 Plant Soil (2014) 382:219–236

Materials and methods
Study area
The study area, located in the Calanques hills, in theperi-urban area of Marseille (southeast of France), wascharacterized by a matorral vegetation dominated byR. officinalis, Cistus albidus, Quercus coccifera, andPistacia lentiscus under Mediterranean climaticconditions.
Eight sites were selected for this study, with similarsoil composition, vegetation, and climatic conditions(Fig. 1). The sampling areas were chosen along a doubletransect from the former smelter factory of l'Escalette tothe Garenne valley following the direction of the pre-vailing wind and from the latter to the sea through theMounine valley. Seven sites were selected along asuspected contamination gradient: G0 on the site of the
factory (close to the horizontal creeping chimney exit);G1, G2, and G3 in the Garenne valley; G4 and G5 in theMounine valley; and G6 in Sormiou cove which is faraway from the Escalette and the urban area and could beconsidered as low contaminated according to the map-ping of soil element concentrations conducted in anextended area around the factory site (unpublished re-sults). The last site S3, in a never industrialized part ofthe Calanques hills, was considered as a reference sitesince the soil was least contaminated on the mappingpreviously conducted cited above.
Activities of the factory of l’Escalette lasted from1851 to 1925, and during this period, around 30,000 t/year of silver-galena was treated by pyrometallurgicalprocesses (Raveux 2002). During this industrial activity,approximately 20,000 m3 of slag was deposited on theformer factory site, but slags were also scattered alongthe coast as several main deposits and as road fill. The
Fig. 1 Geographic localization of the eight sampling sites (plain dots) and the former smelting factory (star-shaped dot) on theMarseilleveyre massif in the National park of Calanques (Southeast France)
Plant Soil (2014) 382:219–236 221

study area is submitted to the Mediterranean climatewith average minimal and maximal temperatures of7.1 and 24.1 °C, respectively, and a mean annual rainfallof 554 mm on 56 days (reference period 1971–2000).Dominant winds are north and northeast and can reach120 km/h. The geological underground is limestone andbare rocks dominate. The soils of the selected sites arestony, and their thicknesses vary from place to place butare generally less than 50 cm. Soils are alkaline with anaverage pH (ISO 10390 2005) between 7.8 and 8.1,belonging to the typical pH range of soils from calcar-eous areas. Soil fertility is low with total organic carboncontents (ISO 10694 1995) varying from 3.6 to 14.2 %,total Kjeldahl nitrogen (ISO 11261 1995) contents from0.28 to 0.72 %, assimilable phosphorus (ISO 112631994) from 0.010 to 0.057 g P/kg, and cation exchangecapacity (CEC, ISO 22036 2008) from 15 to 42 cmol+/kg (Table 1).
Plant and soil sampling
Sample collection was done as previously reported byAffholder et al. (2013) on five rosemary individuals ofthe same population on each site. In order to obtainrepresentative samples from each site, an area of100 m2 (10×10 m) was delimited on the eight selectedsites, i.e., G0, G1, G2, G3, G4, G5, G6, and S3. Theplant cover was over 60 % on all the selected areas. Thefive rosemary individuals were selected according to across pattern inside the delimited area, spaced by around
2 m, and were with similar sizes, i.e., heights and collardiameters, and same phenological stage. Therefore, ineach of the eight sites, a total of five plant/soil coupleswere taken. Roots were collected for TMM analysis andsymbiosis detection. Soil samples were collected fromthe top 15 cm (after removal of the litter) in therhizospheric area of the plants. In addition, on the con-taminated site G0 (as previously defined by Affholderet al. 2013), the whole root network of one rosemaryindividual was sampled in order to determine TMMadsorption and localization in primary and lateral roots.G0 site was the best candidate to ensure TMM localiza-tion and quantification in rosemary since it is the mostpolluted one.
Fresh plant and soil samples were stored in cleanplastic bags for transport to the laboratory.
Soil TMM analysis
Soil samples were sieved on site to 2-mm mesh, air-dried at room temperature in laboratory, and then ground(RETSCH zm 1000 with tungsten blades) to passthrough a 0.2-mm titanium sieve, before analyses.
Pseudo-total TMM
Soils were mineralized (in triplicates) in a microwavemineralizer (Milestone Start D) using aqua regia (1/3HNO3 + 2/3 HCl). A ratio of dry soil/aqua regia solutioncorresponding to 1/20 w/v was used. The mineralization
Table 1 Soils characteristics of the studied sites
Sites Soil physicochemical parametersa
CEC (cmol+/kg) N % C % P (g/kg) pH
G0 31.6±6.6 ac 0.72±0.18a 9.6±5.7ab 0.057±0.014 7.9±0.2ab
G1 15.7±1.1bd 0.29±0.04bd 9.4±1.9a 0.014±0.041 8.1±0.08ab
G2 41.8±8.5c 0.55±0.10 ac 12.6±3.1a 0.030±0.010 7.8±0.3a
G3 24.6±6.2acd 0.44±0.05c 11.1±1.5a 0.012±0.002 8.1±0.06b
G4 41.7±15.7 ac 0.66±0.26 ac 14.2±6.1a 0.013±0.006 7.9±0.1a
G5 42.4±13.9c 0.54±0.15 ac 11.2±1.9a 0.013±0.006 7.9±0.1ab
G6 27.3±1.6a 0.43±0.13 cd 9.8±2.9a 0.010±0.002 7.9±0.1a
S3 14.0±8.4d 0.28±0.09d 3.6±2.8b 0.013±0.005 8.1±0.1ab
In the same column, values followed by the same letter are not significantly different (Mann–Whitney–Wilcoxon test, p<0.05)
CEC cationic exchange capacity (in cmol+ /kg), N% total Kjeldahl nitrogen (in percentage), C% total organic carbon (in percentage), Passimilable phosphorus (in g P/kg)aMean±standard deviation (n=5).
222 Plant Soil (2014) 382:219–236

products were filtered with a 0.45-μm mesh, and theTMM levels were determined by inductively coupledplasma-atomic emission spectroscopy (ICP-AES) andgraphite furnace-atomic absorption spectroscopy (GF-AAS) (Thermo Scientific ICE 3000) for As and Sb.Quality assurance–quality controls and accuracy werechecked using standard soil reference materials(CRM049–050, from RTC, USA) with accuracies with-in 100±10 %.
Exchangeable TMM
A 0.05 M Ca(NO3)2 solution was used as extractant andwas prepared following the protocol of the CommunityBureau of Reference (BCR) and Quevauviller (1998). Aratio of dry soil/Ca(NO3)2 solution corresponding to1/10 w/v was used. The mixture was prepared into aPTFE (Teflon) tube (triplicates per soil sample) andstirred at room temperature on an orbital shaker (FisherScientific Bioblock SM30B) at 125 rpm for 2 h. Thetubes were then centrifuged for 10 min at 8,000 rpm (JPSELECTA, Médifriger BL-S), and the supernatantswere collected and filtered to 0.45 μm. The resultingsolutions were acidified to 1 % with supra-pure nitricacid and stored at 4 °C until analysis by ICP-AES or GF-AAS.
Plant TMM analysis
Root samples were thoroughly washed, three times un-der tap water then three times with a dental spray usingMilli-Q water. Samples were dried at 40 °C during1 week and afterwards ground at 0.2 mm (RETSCHzm 1000 blender).
Pseudo-total TMM
About 0.5 g dry matter of each root sample (triplicates)was digested in microwave mineralizer system (Mile-stone Start D) with an acidic mixture (2/3 HNO3 + 1/3HCl). A ratio of dry plant/acid mixture solution corre-sponding to 1/18 w/v was used. After filtration(0.45 μm), mineralisates were analyzed for Pb and Zncontents by ICP-AES and for As and Sb by GF-AAS.Standard plant reference materials (DC 73349) fromChina National Analysis Centre for Iron and Steel(NCS) were analyzed as a part of the quality assur-ance–quality control protocol (accuracies within 100±10 %). Bioconcentration factors (BCFs), i.e., ratios of
root concentration vs. soil pseudo-total (BCFtotal) orexchangeable concentrations (BCFexch), werecalculated.
Adsorbed TMM at the root surface
Adsorbed TMM extraction was carried on the selectedrosemary roots, sampled on the site G0, consideringdifferent root diameters (Φ) gathered as follows: taproot(Φ ranging from 10 to 15 mm), thick primary root (Φranging from 5 to 10 mm), medium secondary root (Φranging from 2.5 to 5 mm), and thin secondary roots (Φranging from 1 to 2.5 mm). The dried root samples wereimmersed in a 20-mmol/L solution of NaEDTA andstirred for 15 min at 125 rpm, following the protocolproposed by Xiong et al. (2011). A ratio of dryroot/NaEDTA solution corresponding to ca. 1/20 w/vwas used. After stirring, the solution was recovered,filtered to 0.45 μm, and then stored at 4 °C until anal-ysis. In order to determine the percentage of adsorbedTMM, the treated roots were rinsed, dried, and groundfor remaining (i.e., absorbed) TMM quantification.
Symbiosis colonization rate
The selected roots have been prepared and stained byadapting the technique of Phillips and Hayman (1970)on rosemary. Previously, the roots were thoroughlycleaned with distilled water in order to remove soilparticles. The washed roots were stored at room temper-ature in ethanol 60 % until proceeding. Then, roots werecleared in KOH 20 % for 50 min at 90 °C. After threerinses with distilled water, roots were soaked in 2 %hydrochloric acid for 5 min. Fungal structures arehighlighted by staining in 0.05 % trypan blue inlactophenol. The roots were impregnated with dye andplaced in a water bath at 80 °C for 5 min. The excess ofdye was removed, and the roots were soaked in glycerolbefore the slide preparation. The myceliummorphologyof DSE was observed at ×400 magnification with anoptical microscope. The location of DSE structuresincluding parti t ioned melanized hyphae andmicrosclerotia was observed and their colonizationpercentage estimated. AM structures such asarbuscules, vesicles, and hyphae were examined. Thepercentages of colonization of AM fungi and DSE wereestimated independently according to the methodproposed by Zhang et al. (2010). On each microscopeslide, 10 fragments of stained roots were placed to
Plant Soil (2014) 382:219–236 223

estimate the proportion of roots colonized by AM fungiand/or DSE. For each rosemary individual, 10 slideswere prepared. Thus, the current results correspond tothe examination of 100 root fragments per individual.
TMM localization in roots
Fragments of dried root of various diameters were takenfrom the whole rosemary sampled on site G0, i.e., onecollected on the primary root (root 1), one on a second-ary root (root 2), and one on a thinner secondary root(root 3). These fragments were included in an epoxyresin (EPON), using De Jong et al. (2013) technique,and cross sections of 200-μm thickness were cut with awire saw.
TMM localization in roots was performed by X-raymicrofluorescence spectroscopy (μXRF) with aXGT7000 spectrometer (Horiba Jobin Yvon) equippedwith an X-ray tube constituted by a rhodium source,running at an accelerating voltage of 30 kV, a current of1 mA, and in partial vacuum. X-rays emitted from theirradiated sample were detected by an energy dispersiveX-ray spectrometry (EDXS) detector equipped with ahigh-purity silicon, cooled by liquid nitrogen. Elementalmapping (128 px2 meaning a pixel size of 4μm) showedthe distribution of Ca, Fe, K, S, Pb, and Zn in the rootsections with a counting time of 20×1,000 s. Maps weredone on whole root section for both secondary andthinner root sections. For the primary root, only onequarter was mapped since the section was too large fora complete observation. Then, to confirm mapping ob-servations, some X-ray fluorescence spectra were doneon selected areas with a counting time of 1,000 s. Forprimary root, spectra were obtained in the whole sectionmapped. Concerning secondary and thinner roots, twospectra were performed: one on a part where TMMweredetected and the other one beside the root in order toexclude a background noise.
Statistical analysis
Statistical analyses were performed using the R soft-ware. Comparisons of means were performed usingthe nonparametric Mann–Whitney–Wilcoxon test witha p≤0.05.
Significant linear correlations were determined withthe same level by the Pearson test available from theHmisc software.
Results
TMM in soil
The results of the TMM pseudo-total concentrations insoils showed clearly the influence of the former indus-trial site of l'Escalette on diffuse pollution in its sur-roundings (Table 2). Indeed, the furthest sites (G4, G5,G6, and S3) presented a significantly lower contamina-tion than those localized closer to the factory ruins (G0,G1, G2, and G3). In the case of important contamina-tion (G0 to G3), relative standard deviation exceeding50 % was observed between samples from the samesite. Awide range of pseudo-total TMM concentrationswas covered by whole sites (Table 2) since the mini-mum and maximum concentrations measured were4 and 2,738 for As, 21 and 19,646 for Pb, 1.5 and759 for Sb, and 46 and 5,843 for Zn. S3 could beconsidered as the reference area for this study sinceTMM soil concentrations in this site were very closeto the local background of contamination in theCalanques hills, which was estimated at <10, 40,<50, and 53 mg/kg for As, Pb, Sb, and Zn, respectively(Affholder et al. 2013). Thus, TMM concentrationvalues from S3 were used to calculate contamina-tion factors (CF=average TMM soil concentrationon a site/average TMM concentration of the localbackground). These factors gave a contaminationlevel per element and allowed to estimate a multi-contamination level by calculating pollution load index( PLI ¼4
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
CFAs � CFPb � CFSb � CFZnp� �
) (Rashed2010) (Table 3). Concerning G0, CF values showedcontamination above 200 for As and Pb, >100 forSb, and >40 for Zn. These factors decreased slightly insite G2 for all TMM, and then, CF values were dividedby a factor higher than 2 on G1 and G3. Sites G4 andG5 presented similar CF values between 10 and 15for As and Pb and <5 for Sb and Zn. Finally, G6appeared slightly contaminated with CF <2 for Pb,Sb, and Zn but still ca. 10 for As. Concerning PLIvalues, results showed an important decreasingmulticontamination following this order: G0 > G2 >G3 > G1 > G4 ≥ G5 > G6.
Exchangeable TMM in soils were low as the maxi-mum concentrations analyzed were 1.42, 1.80, 2.75, and10.8 mg/kg for As, Pb, Sb, and Zn, respectively(Table 2). Thus, there were low percentages of ex-changeable TMM, considered as the bioavailablefraction (Meers et al. 2007), since ratio between
224 Plant Soil (2014) 382:219–236
exchangeable and pseudo-total fractions was generallybelow 1 %, whatever the element (Table 2). On anotherhand, a significant and positive correlation (Pearson test,p≤0.05) of 0.66 was observed between the exchange-able concentrations and pseudo-total concentrations ofAs and Pb, 0.84 for Zn, and 0.93 for Sb. Thus, theaverage exchangeable fractions seemed to be correlatedwith concentrations of pseudo-total TMM.
TMM in roots
TMM concentrations in studied roots were between0.22 and 85, 0.8 and 1,983, 0.02 and 35, and 5.4 and
Table 2 As, Pb, Sb, and Zn average (in bold, ±standard deviation, n=5), minimal, and maximal concentrations in soils of the eight studiedsites, for pseudo-total (total) and exchangeable (Ca(NO3)2) TMM fractions in milligrams per kilogram
site Trace elements (mg/kg)
As Pb Sb Zn
Total Ca(NO3)2 Total Ca(NO3)2 Total Ca(NO3)2 Total Ca(NO3)2
G0 Min 461 0.28 4,492 0.23 133 0.51 1,278 0.90
Aver 1,134±934 0.66±0.44 9,210±6,193 0.77±0.59 319±256 1.47±0.82 2,723±1,874 3.10±2.95
Max 2,738 1.42 19,646 1.54 759 2.75 5,843 6.38
G1 Min 116 0.05 858 0.03 38 0.13 328 0.23
Aver 306±156 0.09±0.05 3,031±1,835 0.56±0.73 112±60.9 0.21±0.08 1,017±539 0.91±1.01
Max 517 0.17 5,437 1.80 190 0.34 1,669 2.63
G2 Min 379 0.15 4,163 0.37 118 0.25 974 0.44
Aver 949±543 0.34±0.17 8,939±4,475 0.73±0.32 284±158 0.79±0.55 2,821±1,802 3.83±3.78
Max 1,560 0.57 19,646 1.02 759 1.56 5,843 9.96
G3 Min 89.1 0.08 593 0.01 23.3 0.06 231 0.19
Aver 390±503 0.16±0.06 3,445±4,367 0.29±0.51 119±159 0.30±0.37 1,523±2,182 2.52±4.65
Max 1,277 0.26 11,110 1.20 400 0.92 5,397 10.81
G4 Min 20.3 0.04 593 0.13 9.1 0.02 231 0.14
Aver 53.1±37.5 0.08±0.05 605±250 0.73±0.55 10.5±4.2 0.03±0.01 295±87.2 0.57±0.40
Max 116 0.16 896 1.6 15.4 0.05 382 1.2
G5 Min 33.4 0.02 278 0.004 8.1 0.002 186 0.20
Aver 53.3±14.2 0.08±0.07 480±247 1.0±0.15 9.6±2.8 0.01±0.01 293±114 1.4±2.2
Max 72.1 0.18 867 0.35 14.5 0.03 473 1.2
G6 Min 34.5 <QL 37.4 <QL 1.7 0.0005 70.1 0.07
Aver 48.1±9.9 0.009±0.016 62.0±15.0 0.0005±0.0011 4.0±3.1 0.003±0.003 100±18.4 0.20±0.14
Max 56.5 0.04 73.0 0.003 9.4 0.01 119 0.40
S3 Min 4.0 0.01 21.4 0.003 1.5 0.003 45.6 <QL
Aver 4.9±0.69 0.06±0.04 42.9±17.3 0.008±0.004 3.1±1.8 0.009±0.011 66.0±23.3 0.16±0.29
Max 5.8 0.11 65.9 0.01 5.9 0.03 102 0.67
Min minimal,Max maximal, Aver average, QL quantification limit (≤0.010, ≤0.0015, and ≤0.016 mg/kg for exchangeable As, Pb, and Zn,respectively)
Table 3 As, Pb, Sb, and Zn contamination factors (CF) andpollution load index (PLI in bold) calculated for all studied sites
Sites Contamination factors PLI
As Pb Sb Zn
G0 231 215 102 41 120
G1 62 71 36 15 40
G2 193 208 91 43 112
G3 79 80 38 23 49
G4 11 14 3.4 4.5 6.9
G5 11 11 3.1 4.4 6.4
G6 10 1.4 1,3 1.5 2.3
Plant Soil (2014) 382:219–236 225
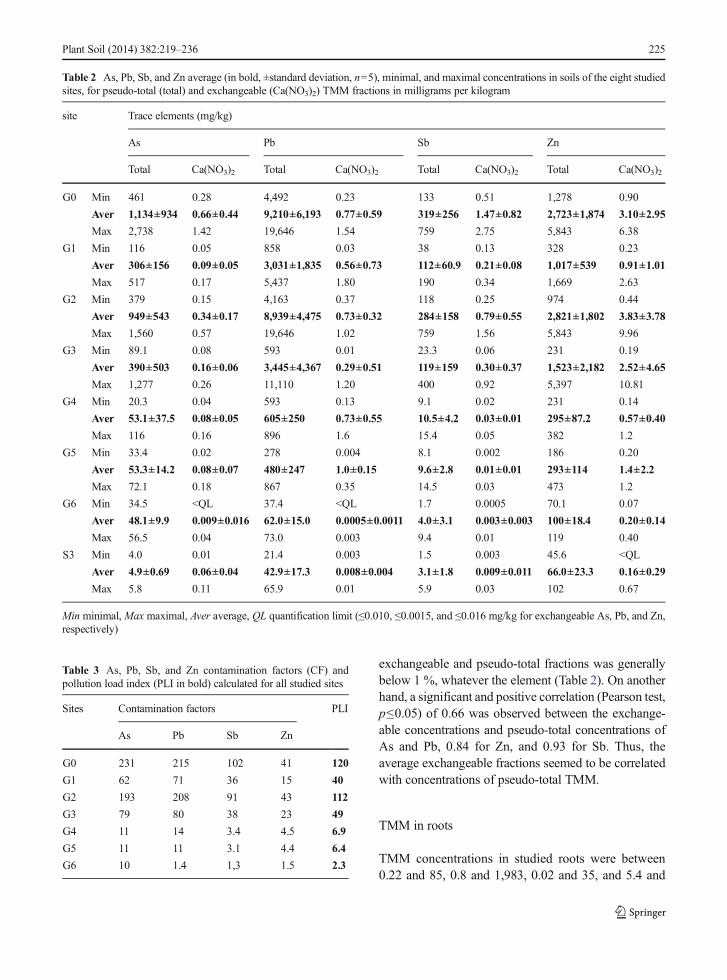
803 mg/kg of dry weight (DW) for As, Pb, Sb, and Zn,respectively (Fig. 2). The highest TMM concentrationswere found in the roots of rosemary individuals fromhighly contaminated sites (G0 to G3). Indeed, for Pb,Sb, and Zn in the four most contaminated sites (G0, G1,G2, and G3), the average root concentrations were sig-nificantly higher (Mann–Whitney–Wilcoxon test, p≤0.05) than those observed in roots collected in the lesscontaminated sites (G4, G5, G6, and S3). For As, onlythe average TMM root concentrations of G0, G1, andG2 sites were significantly higher than those of the fiveothers (Table in Supplementary Material). Thus, TMMcontents in roots were influenced by soil contaminationlevel. Figure 2 shows TMM concentrations in roots infunction of TMM pseudo-total concentrations in soil,and in this figure, each point represents one rosemaryindividual. There is a high discrepancy between eachsoil sample from the same site, especially in the mostcontaminated sites. However, a significant linear
correlation (Pearson's test, p≤0.05) was demonstratedfor As, Pb, Sb, and Zn with a correlation coefficient of0.52, 0.56, 0.52, and 0.7, respectively. Thus, TMMconcentrations in the roots of rosemary individuals werelinearly correlated with the pseudo-total concentrationsof soils.
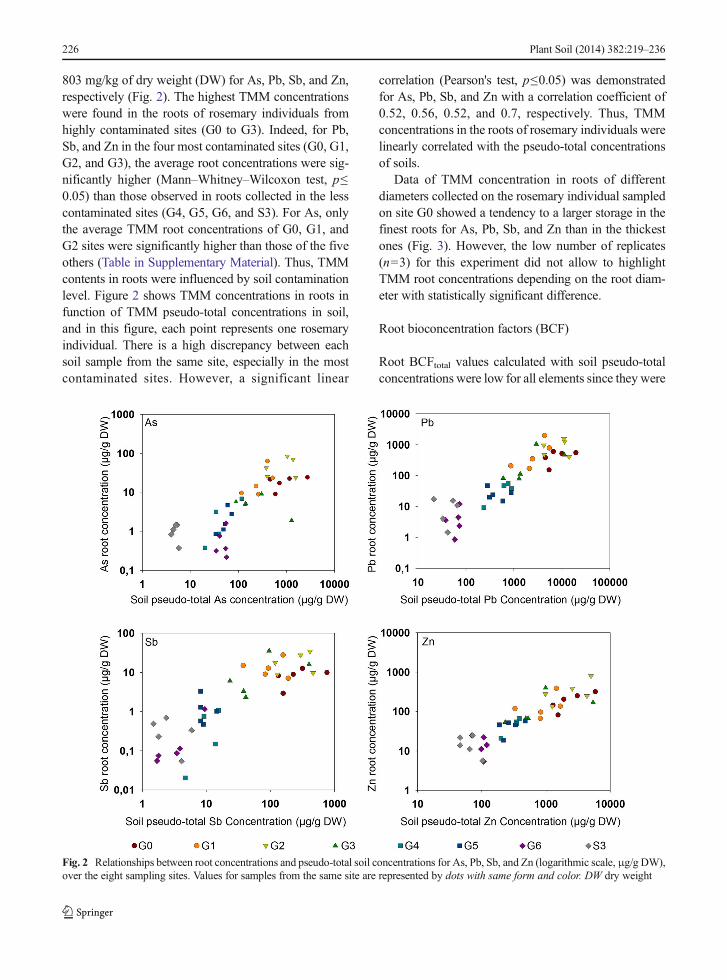
Data of TMM concentration in roots of differentdiameters collected on the rosemary individual sampledon site G0 showed a tendency to a larger storage in thefinest roots for As, Pb, Sb, and Zn than in the thickestones (Fig. 3). However, the low number of replicates(n=3) for this experiment did not allow to highlightTMM root concentrations depending on the root diam-eter with statistically significant difference.
Root bioconcentration factors (BCF)
Root BCFtotal values calculated with soil pseudo-totalconcentrations were low for all elements since they were
Fig. 2 Relationships between root concentrations and pseudo-total soil concentrations for As, Pb, Sb, and Zn (logarithmic scale, μg/g DW),over the eight sampling sites. Values for samples from the same site are represented by dots with same form and color. DW dry weight
226 Plant Soil (2014) 382:219–236
far below 1. Indeed, the average values of root BCFtotalfor all sites were between 0.02 and 0.23, 0.04 and 0.26,0.03 and 0.17, and 0.08 and 0.25 for As, Pb, Sb, and Zn,respectively. For these TMM, the maximal averagevalues were obtained on the reference site, S3. However,despite the low variability of the soil contamination onthis site, the root BCFtotal values were widely dispersed(between 30 to 77 % of variation). No significant differ-ences were found for the average values of root BCFtotalbetween the studied sites and also among the variouselements.
Root BCFexch values calculated from soil ex-changeable concentrations showed average values be-tween 24 and 246, 49 and 17,632, 6.3 and 115, and110 and 350 for As, Pb, Sb, and Zn, respectively. Theaverage values of root BCFexch were all distinctlyabove 1, with some particularly high values for Pbin contaminated sites (G3). Yet, as for BCFtotal, nosignificant differences were found for the averagevalues of root BCFexch between the studied sites andalso among the various elements.
Ab- and adsorption of TMM at the root level
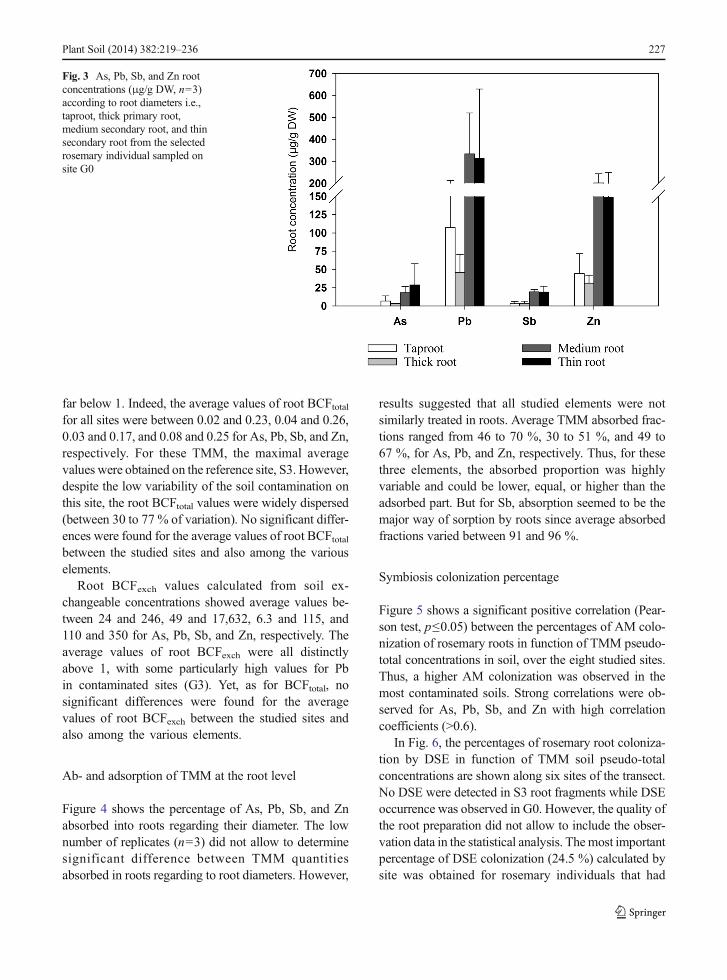
Figure 4 shows the percentage of As, Pb, Sb, and Znabsorbed into roots regarding their diameter. The lownumber of replicates (n=3) did not allow to determinesignificant difference between TMM quantitiesabsorbed in roots regarding to root diameters. However,
results suggested that all studied elements were notsimilarly treated in roots. Average TMM absorbed frac-tions ranged from 46 to 70 %, 30 to 51 %, and 49 to67 %, for As, Pb, and Zn, respectively. Thus, for thesethree elements, the absorbed proportion was highlyvariable and could be lower, equal, or higher than theadsorbed part. But for Sb, absorption seemed to be themajor way of sorption by roots since average absorbedfractions varied between 91 and 96 %.
Symbiosis colonization percentage
Figure 5 shows a significant positive correlation (Pear-son test, p≤0.05) between the percentages of AM colo-nization of rosemary roots in function of TMM pseudo-total concentrations in soil, over the eight studied sites.Thus, a higher AM colonization was observed in themost contaminated soils. Strong correlations were ob-served for As, Pb, Sb, and Zn with high correlationcoefficients (>0.6).
In Fig. 6, the percentages of rosemary root coloniza-tion by DSE in function of TMM soil pseudo-totalconcentrations are shown along six sites of the transect.No DSE were detected in S3 root fragments while DSEoccurrence was observed in G0. However, the quality ofthe root preparation did not allow to include the obser-vation data in the statistical analysis. Themost importantpercentage of DSE colonization (24.5 %) calculated bysite was obtained for rosemary individuals that had
Fig. 3 As, Pb, Sb, and Zn rootconcentrations (μg/g DW, n=3)according to root diameters i.e.,taproot, thick primary root,medium secondary root, and thinsecondary root from the selectedrosemary individual sampled onsite G0
Plant Soil (2014) 382:219–236 227
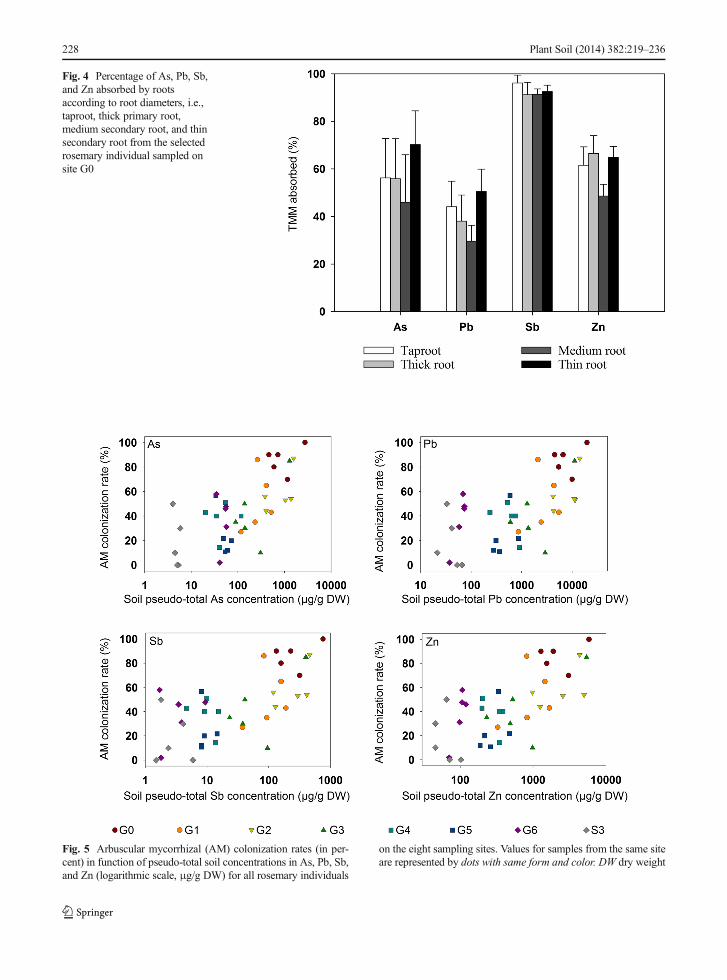
Fig. 4 Percentage of As, Pb, Sb,and Zn absorbed by rootsaccording to root diameters, i.e.,taproot, thick primary root,medium secondary root, and thinsecondary root from the selectedrosemary individual sampled onsite G0
Fig. 5 Arbuscular mycorrhizal (AM) colonization rates (in per-cent) in function of pseudo-total soil concentrations in As, Pb, Sb,and Zn (logarithmic scale, μg/g DW) for all rosemary individuals
on the eight sampling sites. Values for samples from the same siteare represented by dots with same form and color. DW dry weight
228 Plant Soil (2014) 382:219–236

grown on the site G2, the most contaminated site.On the other sites, except on G4 (average rate of5 %), DSE colonization was similar with an averagepercentage ca. 10 %. In this case also, a significantpositive correlation (Pearson test, p≤0.05) betweenDSE colonization and TMM soil contamination washighlighted with correlation coefficients higher than0.5 for As, Pb, Sb, and Zn.
TMM localization in roots
Localization and pictures of the analyzed root samplescollected on the selected rosemary individual on the siteG0, a section of primary root (root 1) and two sections ofsecondary roots, with different diameter (root 2 for thelargest and root 3 for the thinnest), were presented inFig. 7. It also shows μXRFmapping results for the threestudied root samples, where elements detected byμXRFare represented by white dots. Only Pb and Zn were
detected in this analysis, the other elements beingprobably below detection limits of the device. Themaps produced by X-ray microfluorescence spec-troscopy highlighted the presence of Pb and Zn inthe peripheral tissues of the lateral roots (roots 2and 3). On the primary root, none of these elementswere detected.
In order to confirm μXRF mapping observation, X-ray fluorescence spectra were recorded on each of thethree root samples by analyzing the whole mappedsection for root 1 and only the selected areas locatedinside the marked squares for roots 2 and 3 (Fig. 8). Thespectrum of the root 1 showed the presence of Zn andconfirmed the absence of Pb (Fig. 8), while the presenceof both elements was confirmed by spectra from theselected areas in roots 2 and 3. A background effectcould be rejected since spectra obtained after analysis ofareas outside the roots did not show peaks correspond-ing to Pb nor Zn (Figure in Supplementary Material).
Fig. 6 Dark septate endophyte (DSE) colonization rates (in per-cent) in function of pseudo-total soil concentrations in As, Pb, Sb,and Zn (logarithmic scale, μg/g DW) for all rosemary individuals
on the six sites along the transect. Values for samples from thesame site are represented by dots with same form and color. DWdry weight
Plant Soil (2014) 382:219–236 229

Discussion
Diffuse soil contamination
The results of the TMM pseudo-total concentrations insoils clearly showed the influence of the former indus-trial site of l’Escalette on diffuse contamination inCalanques Hills. Indeed, contaminations of the farthestsites (G4, G5, G6, and S3) were significantly lower thanthose located close to the former smelter factory (G0,G1, G2, and G3) (Tables 2 and 3). Furthermore, in thecase of highly contaminated areas, the high heterogene-ity of TMM soil concentrations was characteristic ofanthropogenic contamination, particularly contamina-tion related to atmospheric deposits from industrial ac-tivities (Yaylali-Abanuz 2011; Testiati et al. 2013).
PLI order of values (similar to CF for As, Pb, Sb, andZn) showed that, for the four most contaminated sites,the distance was not the only criterion affecting the levelof contamination. Indeed, a higher contamination levelwas found on the site G2 than on the site G1, while thelatter was closer to the former factory. Two phe-nomena might be at the origin of the very highcontamination measured on G2 (similar to thatmeasured in G0, located close to the old creepingchimney exit). On one hand, this site could beexposed to vortex phenomena promoting contami-nated particle deposition as G2 was located in theGarenne valley directly in the axis of the prevailingwind from the former factory and the chimney exitand where the valley presented a narrowing and asharp altitude increase. In addition, soil of G2 was
Fig. 7 Localization and pictures of the three different root sam-ples collected on a rosemary individual from site G0 and treatedfor micro-X-ray fluorescence spectrometry analyses and μXRF
maps obtained on each root sample for two elements: Pb and Zn(white dots represent detected elements)
230 Plant Soil (2014) 382:219–236

a colluvial soil formed by erosion and leaching ofupstream sites, where contamination particles werelikely to accumulate.
In soils from each site, TMM exchangeable fractionwas low (Table 2). Exchangeable TMM correspond tothe weakly adsorbed part on solid phase of soil, mainly
Fig. 8 Micro-X-ray fluorescence analysis spectra from wholemapped area of a primary root (root 1 and spectrum 1) and fromselected areas on secondary root, located inside marked squares
(roots 2 and 3 and spectra 2 and 3, respectively). Spectra 2 wererecorded in a zone of the roots 2 and 3 where Pb and Zb weredetected during mapping
Plant Soil (2014) 382:219–236 231

constituted of exchangeable ions within soil solution(Morel 1997) , which can be considered asphytoavailable TMM. An important result in our studywas that a significant positive linear correlation wasobserved between the exchangeable and pseudo-totalconcentrations of As, Pb, Sb, and Zn in soils, meaningthat TMM exchangeable concentrations increased withpseudo-total TMM concentrations (results not shown).Thus, the risk of transfer of these elements to the bioce-nosis seemed to be more important in the heavily con-taminated sites.
Rosemary as a good candidate for phytostabilization
Linear correlation between TMM concentrations in soiland concentrations in root (Fig. 2) matched the defini-tion of an indicator species according to Baker (1981). Italso suggested a major passive root absorption for Znand at least a partial passive absorption for As, Pb, andSb. The maxima of TMM concentration in roots were100, 50, 10, and 10 times higher for Pb, As, Sb, and Zn,respectively, than those found in aerial parts (results notshown). Moreover, the TMM concentrations measuredin rosemary roots reached high values, highlighting thatthis species accumulated preferentially TMM in rootsthan in aerial parts. The ability of rosemary root toaccumulate TMM was determined using root BCF.Following Baker and Brooks (1989), a BCF higher than1 for one element means that the plant species is able tohyperaccumulate this element. Thus, as BCFtot calculat-ed with soil pseudo-total concentrations were lower than1 for all TMM and all sites, rosemary could not beconsidered as a hyperaccumulator species. However,the nonsignificant differences between BCFtot obtainedfrom all sites whatever the element, except for S3,showed that TMM accumulation in rosemary rootswas not dependent on contamination level. Moreover,BCF values were of the same order of magnitude for thefour elements, highlighting that rosemary did not accu-mulate preferentially a TMM than another. Moreno-Jimenez et al. (2011) obtained similar BCF values forthe same species. For soil pseudo-total concentrations inthe same range of values, they calculated root BCFtot of0.025 and 0.1 against 0.06 to 0.03 and 0.13 to 0.17 in thepresent case for As and Zn, respectively. Unfortunately,to our knowledge, there is no data on Pb and Sb analysisin rosemary roots in the literature. However, rootBCFexch were largely above 1 for all TMM, and thishighlighted that rosemary was able to accumulate in its
root the most labile fraction of TMM in soil, whichpresented the largest risks for environment.
Insight in the mechanism involved in TMM storagein rosemary root
TMM partition in rosemary root system
The maps and spec t r a ob ta ined by X- raymicrofluorescence spectroscopy analyses highlightedthat Pb and Zn were stored preferentially in peripheraltissues and in thin roots (Figs. 7 and 8). These resultsseemed to be confirmed by the results of TMM concen-trations in roots regarding the different root diameters(Fig. 3). Indeed, a global trend was observed, showinghigher total TMM concentrations in both fine and me-dium roots (corresponding to secondary roots) than inthick roots (primary root) and in taproots. Vamerali et al.(2009) showed similar results in poplar and willow, i.e.,a higher concentration of As, Cu, Pb, and Zn in fine thanin thicker roots of these species. These observationscould be related to a higher concentration of tannins infine roots as it has been highlighted by Chin et al. (2009)in Symphytum officinale. Indeed, the tannins are able tobind TMM and thus increase their adsorption on thesurface of roots. However, as shown in Fig. 4, in the caseof rosemary, the absorbed TMM fraction seemed not tobe affected by root diameter. Despite this, the resultsshowed that an important part of As, Pb, and Zn wasadsorbed since, for instance, until 70 % of Pb could beadsorbed on rosemary roots growing in contaminatedsite. This low absorption could be related to a mecha-nism of resistance by rosemary, promoting the adsorp-tion of these elements on the surface of roots. Indeed,previous studies have shown that the presence of TMMin soil, including Pb, resulted in an increase of phenoliccompound production (Kovacik and Klejdus 2008;Pawlak-Sprada et al. 2011) which could be able to bindTMMon root surfaces (Michalak 2006). To confirm thishypothesis, a similar study with rosemary roots from anon-contaminated site would have been required tocompare the patterns of absorption and adsorption ofthese elements in both cases, i.e., in low and heavilycontaminated environments, if the low level of elementsin non-contaminated soils is compatible with the level ofdetection of the method.
Antimony seemed to have a different behavior. In-deed, this element was mainly absorbed (over 90 %),which was also observed in this study for Cu and Fe
232 Plant Soil (2014) 382:219–236

(results not shown). Usually, Sb and As behaviors aresimilar (Wilson et al. 2010). However, in this case, bothelements did not seem to be absorbed by rosemary in thesame way. Little is known about the absorption pathwayof Sb in plant roots, but it is likely that both forms of Sbare not absorbed in the same way. Sb(III) can passthrough the membrane aquaporins passively. ForSb(V), which is an anion, this kind of passive transportis excluded, and its absorption requires a carrier (Tschanet al. 2009). However, in discharges of lead smelters, thedominant form of Sb in soil seems to be Sb(V) asdescribed by Ettler et al. (2010), suggesting an activeabsorption of this element. Nevertheless, the hypothesisthat this observation is related to a not well-adaptedextractant for lixiviation of Sb bond on root surfacescould not be dismissed.
A contribution of root symbionts
Results highlighted that root colonization rates by AMfungi and DSE increased with soil contamination. Itseemed that the fungi strains present in soils in thesurroundings of l’Escalette former factory were adaptedand resistant to high levels of TMM contamination(Figs. 5 and 6). Although similar results have beenidentified in the literature for AM fungi (Whitfieldet al. 2004), contradictory data exist. Thus, several stud-ies showed a decrease in mycorrhizal colonization withincreasing TMM concentrations in soil (Gildon andTinker 1983; Del Val et al. 1999; Andrade et al. 2004;Yang et al. 2008; Turrini et al. 2010). Mechanisms usedbyAM fungi to increase plant tolerance to TMMdependon the studied species and the strategy response of plantsto tolerate TMM. It seems that for accumulator plantswith a high TMM translocation potential towards theaerial parts, the presence of AM fungi increases thisphenomenon, whereas in plants that exclude TMM,the opposite phenomenon is observed and amplifiedby the presence of AM fungi (Rabie 2005), allowingimproving plant capacity to extract or stabilize TMM(Gohre and Paszkowski 2006). Concerning DSE, simi-lar results have also been highlighted on Salix caprea, inwhich a positive correlation between DSE colonizationrates and concentrations of Pb in soil was shown(Regvar et al. 2010). In addition, Li et al. (2011) showeda significant increase of Pb, Zn, and Cd concentrationsin the roots of Zea mays and a decrease in the translo-cation of these elements to aerial parts in the presence ofDSE.
In the case of rosemary, we hypothesized that itsassociation with fungal species involved in root symbi-oses could promote accumulation in roots and limit thetransfer into the aerial parts, playing the role of a filter.This might be due to adsorption and/or absorption infungal compartments. AM and DSE hyphae may adsorbsignificant amounts of TMM. However, X-raymicrofluorescence methods require thick cross sectionsof non-stained roots on which the microscopicobservation of fungal structures is not easy. Thus, itwas not possible to distinguish if TMM were or notinside fungal structures. According to Gohre andPaszkowski (2006), metal-tolerant fungi may have agreater affinity for TMM than roots have (2.4 times).Hyphal cell walls are constituted by compounds able toadsorb TMM on their surface, like melanin for DSE(Gadd 1993) and chitin (Gohre and Paszkowski 2006)and glomalin (Driver et al. 2005) for AM fungi. Forexample, glomalin, which is a glycoprotein, is able toextract and sequester large amounts of TMM includingCu, Pb, and Zn (Gonzalez-Chavez et al. 2004; Chernet al. 2007; Vodnik et al. 2008; Cornejo et al. 2008).Results concerning the sequestration capacity of TMMby glomalin are contrasting. However, Cornejo et al.(2008) and Chern et al. (2007) agree on the fact thatup to 4.8 mg Cu and 188 mg of Pb could be sequesteredby 1 g of glomalin. Concerning AM vesicles, they alsoplay an important role since these fungal structuresallow the detoxification and storage of toxic elements.Their involvement in the immobilization of Pb has beendemonstrated by Chen et al. (2005), who showed acorrelation between the number of fungal vesicles andPb root uptake. It is noteworthy that a considerableamount of fungal vesicles was found in the roots ofrosemary exposed to high contamination and that asignificant correlation of 0.34 was highlighted betweenPb concentration in rosemary roots and AM vesiclepercentage (Pearson test, p≤0.05).
AM and DSE structures are located in the cortex ofthin roots, where TMM and particularly Pb and Zn wereheavily concentrated in rosemary root. Thus, the in-volvement of these fungal structures in rosemary rootaccumulation ability seems significant. Moreover, AMfungi and DSE could also be involved in rosemarytolerance to high TMM contamination levels. Indeed,previous studies highlighted that, in the case of rose-mary, these symbioses improved water status of theplant, promoting resistance to water stress (Sánchez-Blanco et al. 2004), and increased the survival and
Plant Soil (2014) 382:219–236 233

growth of rosemary through a replanting process(Estaún et al. 1997). More generally, AM fungi andDSE are able to improve plant growth, health, andtolerance to contamination promoting phosphorus nutri-tion (Haselwandter and Read 1982; Harrison 1999;Barea et al. 2002). Concerning AM fungi, glomalinexcretion in soil, following hyphae turnover (Driveret al. 2005) but also past fungal growth and slow turn-over of this protein once in soil (Purin and Rillig 2007),helps to stabilize soil aggregates (Wright andUpadhyaya 1998; Bedini et al. 2009). This enhancessoil structure favoring soil aeration and water drainagewhich optimize plant development (Oades 1984).
Conclusions
Developing non-intrusive ecotechnologies for TMMcontainment is a challenging target with an increasingdemand in protected areas like those of the NationalPark of Calanques. The use of native plant species withtheir native symbionts offers a promising perspective tomanage TMM phytostabilization objectives and ecolog-ical and biological conservation constraints. We demon-strated the ability of rosemary, a perennial with anextensive root network, to accumulate TMM in its roots.Indeed, accumulations of As, Pb, Sb, and Zn in rose-mary roots were linearly correlated to their pseudo-totalconcentrations in soil. Despite concentrations could beimportant in the roots of rosemary individuals subjectedto high levels of contamination, the root BCFtotal werelow (<0.3) for these four elements, which highlightedthat rosemary was not able to hyperaccumulate TMM inits root system. However, although this type of extrac-tion (i.e., pseudo-total soil concentration) was question-able because it is not necessarily representative of thephytoavailable fraction, BCFexch values, calculated withexchangeable fractions in soils, were greater than 1meaning that rosemary was able to significantly accu-mulate the most labile TMM in soils. Indeed, exchange-able TMM were representative of the fraction whichwas the most likely to leach during rainfall events and/or to be dissolved in case of physicochemical changes inthe soil (pH, Eh, complexing conditions, etc.), promot-ing expansion of TMM contamination into the environ-ment (groundwater, biocoenosis, etc.).
Among the TMM detected in roots, a significantproportion of As, Pb, and Zn seemed to be mainlyadsorbed on root surfaces and non-absorbed into roots.
This was not the case for Sb, which seemed mainlyabsorbed. For As, Pb, and Zn, strong adsorption onroots could lead to the limited accumulation in aerialparts, previously highlighted by Affholder et al. (2013),and which is reassuring since rosemary aerial parts areedible. From an ecological point of view, the contain-ment of TMM in root parts is also of interest loweringthe risk of transfer to the food web. Root symbiotic rolein rosemary tolerance to TMM and in TMM accumula-tion increase in root was difficult to prove with this insitu study. However, the positive correlation observedbetween the root colonization rates and the TMM con-tamination levels, as well as the storage of TMM inperipheral tissues of secondary roots, fully agrees withthis hypothesis. Endomycorrhizal fungi developed inthe cortical parenchyma and rhizoderm of rosemarysecondary roots. Thus, this study suggested that rose-mary in association with its symbiotic microorganismscould be an interesting candidate for a phytostabilizationprocess of TMM-contaminated soil in the surroundingsof l'Escalette former smelter factory or for other equiv-alent contaminated sites in the Mediterranean area.However, further studies are necessary (i) to elucidateTMM tolerance mechanisms in rosemary, (ii) to deter-mine more precisely symbiont role in rosemarytolerance/accumulation and, (iii) to characterize geneti-cal diversity of tolerant indigenous fungal strains.
Acknowledgments The authors thank Laetitia De Jong for heradvice for root sample inclusion, Perrine Chaurand for her techni-cal assistance in μXRF analyses, Laurent Vassalo for his analyticalassistance in TMMmeasurements, and Estelle Dumas for her helpat designing Fig. 1. This study was funded by the French ResearchNational Agency (ANRMarséco 2008 CESA 018) and financiallyamended by the National Innovative Cluster on RisksManagement.
References
Affholder MC, Prudent P, Masotti V, Coulomb B, Rabier J,Nguyen-The B, Laffont-Schwob I (2013) Transfer of metalsand metalloids from soil to shoots in wild rosemary(Rosmarinus officinalis L.) growing on a former lead smeltersite: Human exposure risk. Sci Total Environ 454–455:219–229
Andrade SA, Abreu C, de Abreu M, Silveira AP (2004) Influenceof lead additions on arbuscular mycorrhiza and Rhizobiumsymbioses under soybean plants. Appl Soil Ecol 26:123–131
Baker A (1981) Accumulators and excluders - strategies in theresponse of plants to heavy-metals. J Plant Nutr 3:643–654
234 Plant Soil (2014) 382:219–236

Baker AJM, Brooks RR (1989) Terrestrial higher plants whichhyperaccumulate metallic elements - a review of their distri-bution, ecology and phytochemistry. Biorecovery 1:81–126
Barea JM, Azcón R, Azcón-Aguilar C (2002) Mycorrhizosphereinteractions to improve plant fitness and soil quality. AntonieVan Leeuwenhoek 81:343–351
Bedini S, Pellegrino E, Avio L, Pellegrini S, Bazzoffi E, Argese E,Giovannetti M (2009) Changes in soil aggregation andglomalin-related soil protein content as affected by thearbuscular mycorrhizal fungal species Glomus mosseae andGlomus intraradices. Soil Biol Biochem 41:1491–1496
Cala V, Cases MA,Walter I (2005) Biomass production and heavymetal content of Rosmarinus officinalis grown on organicwaste-amended soil. J Arid Environ 62:401–412
Chen X, Wu CH, Tang JJ, Hu SJ (2005) Arbuscular mycorrhizaeenhance metal lead uptake and growth of host plants under asand culture experiment. Chemosphere 60:665–671
Chern EC, Tsai DW, Ogunseitan OA (2007) Deposition ofGlomalin-Related Soil Protein and Sequestered ToxicMetals into Watersheds. Environ Sci Technol 41:3566–3572
Chin L, Leung DWM, Taylor HH (2009) Lead chelation toimmobilised Symphytum officinale L. (comfrey) root tannins.Chemosphere 76:711–715
Cornejo P, Meiera S, Borie G, Rillig MC, Borie F (2008)Glomalin-related soil protein in a Mediterranean ecosystemaffected by a copper smelter and its contribution to Cu and Znsequestration. Sci Total Environ 406:154–160
De Jong L, Moreau X, Bestel I, Beaudoin E, Aimé A, Dolain C,Gladys Saez G, Tonetto A, Barthélémy P, Thiéry A (2013)Uptake of quantum dots into a freshwater flatworm : intra-cellular accumulation and transmission from parents to off-spring. J Nanosci Lett 3:28
Del Val C, Barea JM, Azcon-Aguilar C (1999) Assessing thetolerance to heavy metals of arbuscular mycorrhizal fungiisolated from sewage sludge-contaminated soils. Appl SoilEcol 11:261–269
Driver JD, Holben WE, Rillig MC (2005) Characterization ofglomalin as a hyphal wall component of arbuscular mycor-rhizal fungi. Soil Biol Biochem 37:101–106
El-Rjoob A-WO,Massadeh AM, Omari MN (2008) Evaluation ofPb, Cu, Zn, Cd, Ni and Fe levels in Rosmarinusofficinalis labaiatae (Rosemary) medicinal plant andsoils in selected zones in Jordan. Environ Monit Assess140:61–68
Estaún V, Savé R, Biel C (1997) AM inoculation as a biologicaltool to improve plant revegetation of a disturbed soil withRosmarinus officinalis under semi-arid conditions. Appl SoilEcol 6:223–229
Ettler V, Tejnecky V, Mihaljevic M, Sebek O, Zuna M, Vanek A(2010) Antimony mobility in lead smelter-polluted soils.Geoderma 155:409–418
Gadd GM (1993) Interactions of fungi with toxic metals. NewPhytol 124:25–60
Gildon A, Tinker PB (1983) Interactions of vesicular-arbuscularmycorrhizal infection and heavy metals in plants. NewPhytol 95:247–261
Gohre V, Paszkowski U (2006) Contribution of the arbuscularmycorrhizal symbiosis to heavy metal phytoremediation.Planta 223:1115–1122
Gonzalez-Chavez MC, Carrillo-Gonzalez R, Wright SF, NicholsKA (2004) The role of glomalin, a protein produced by
arbuscular mycorrhizal fungi, in sequestering potentially tox-ic elements. Environ Pollut 130:317–323
Harrison MJ (1999) Molecular and cellular aspects of thearbuscular mycorrhizal symbiosis. Annu Rev Plant PhysiolPlant Mol Biol 50:361–389
Haselwandter K, Read DJ (1982) The significance of a root-fungus association in two Carex species of high-alpine plantcommunities. Oecologia 53:352–354
He ZL, Yang XE, Stoffella PJ (2005) Trace elements inagroecosystems and impacts on the environment. J TraceElem Med Biol 19:125–140
ISO 10390 (2005) Soil quality—determination of pH. Frenchversion EN ISO 10390, AFNOR Paris
ISO 10694 (1995) Soil quality—determination of organic and totalcarbon after dry combustion (elementary analysis). Frenchversion EN ISO 10694, AFNOR Paris
ISO 11261 (1995) Soil quality—determination of total nitrogen—modified Kjeldhahl method. French version EN ISO 11261,AFNOR Paris
ISO 11263 (1994) Soil quality—determination of phosphorus—spectrometric determination of phosphorus soluble in sodiumhydrogen carbonate solution. French version EN ISO 11263,AFNOR Paris
ISO 22036 (2008) Soil quality—determination of trace elementsin extracts of soil by inductively coupled plasma-atomicemission spectrometry (ICP-AES). French version EN ISO22036, AFNOR Paris
Jumpponen A (2001) Dark septate endophytes—are they mycor-rhizal? Mycorrhiza 11:207–211
Kabata-Pendias A (2004) Soil–plant transfer of trace elements—an environmental issue. Biogeochem Process Role HeavyMet Soil Environ 122:143–149
Kovacik J, Klejdus B (2008) Dynamics of phenolic acids andlignin accumulation in metal-treated Matricaria chamomillaroots. Plant Cell Rep 27:605–615
Laffont-Schwob I, Dumas PJ, Pricop A, Rabier J, Miché L, AffreL, Masotti V, Prudent P, Tatoni T (2011) Insights on metal-tolerance and symbionts of the rare species Astragalustragacantha aiming at phytostabilization of polluted soilsand plant conservation. Ecol Mediterr Rev Int EcolMéditerr Int J Mediterr Ecol 37:57–62
Li T, Liu MJ, Zhang XT, Zhang HB, Sha T, Zhao ZW (2011)Improved tolerance of maize (Zea mays L.) to heavy metalsby colonization of a dark septate endophyte (DSE) Exophialapisciphila. Sci Total Environ 409:1069–1074
Madejon P, Burgos P, Cabrera F, Madejon E (2009)Phytostabilization of amended soils polluted with trace ele-ments using theMediterranean shrub: Rosmarinus officinalis.Int J Phytoremediat 11:542–557
Meers E, Samson R, Tack F, Ruttens A, Vandegehuchte M,Vangronsveld J, Verloo MG (2007) Phytoavailability assess-ment of heavy metals in soil by single extractions and accu-mulation by Phaseolus vulgaris. Environ Exp Bot 60:385–396
Michalak A (2006) Phenolic compounds and their antioxidantactivity in plants growing under heavy metal stress. Pol JEnviron Stud 15:523–530
Morel JL (1997) Assessment of phytoavailability of trace elementsin soils. Analusis 25:M70–M72
Moreno-Jimenez E, Vazquez S, Carpena-Ruiz RO, Esteban E,Penalosa JM (2011) Using Mediterranean shrubs for thephytoremediation of a soil impacted by pyritic wastes in
Plant Soil (2014) 382:219–236 235

Southern Spain: a field experiment. J Environ Manag 92:1584–1590
Oades JM (1984) Soil organic matter and structural stability:mechanisms and implications for management. Plant Soil76:319–337
Pawlak-Sprada S, Stobiecki M, Deckert J (2011) Activation ofphenylpropanoid pathway in legume plants exposed to heavymetals. Part II. Profiling of isoflavonoids and theirglycoconjugates induced in roots of lupine (Lupinus luteus)seedlings treated with cadmium and lead. Acta Biochim Pol58:217–223
Phillips J, Hayman D (1970) Improved procedures for clearingroots and staining parasitics and arbuscular mycorrhizal fungifor rapid assessment of infection. Trans Br Mycol Soc 55:158–161
Pilon-Smits EAH (2005) Phytoremediation. Annu Rev Plant Biol56:15–39
Purin S, Rillig MC (2007) The arbuscular mycorrhizal fungalprotein glomalin: limitations, progress, and a new hypothesisfor its function. Pedobiologia 51:123–130
Quevauviller P (1998) Operationally defined extraction proce-dures for soil and sediment analysis I. Standardization.TrAC Trends Anal Chem 17:289–298
Rabie GH (2005) Contribution of arbuscular mycorrhizal fungusto red kidney and wheat plants tolerance grown in heavymetal-polluted soil. Afr J Biotechnol 4:332–345
Rabier J, Laffont-Schwob I, Bouraima-Madjebi S, Léon V,Prudent P, Viano J, Nabors MW, Pilon-Smits EA (2007)Characterization of metal tolerance and accumulation inGrevillea exul var exul. Int J Phytoremediat 9:419–435
Rashed MN (2010) Monitoring of contaminated toxic and heavymetals, from mine tailings through age accumulation, in soiland some wild plants at Southeast Egypt. J Hazard Mater178:739–746
Raveux O (2002) L’usine à plomb de l’Escalette: le dernier grandvestige de la métallurgie marseillaise du plomb. Industries enProvence 10:7–9
Regvar M, Likar M, Piltaver A, Kugonic N, Smith JE (2010)Fungal community structure under goat willows (Salixcaprea L.) growing at metal polluted site: the potential ofscreening in a model phytostabilisation study. Plant Soil 330:345–356
Sánchez-BlancoMJ, Ferrández T, Morales MA,Morte A, AlarconJJ (2004) Variations in water status, gas exchange, andgrowth in Rosmarinus officinalis plants infected withGlomus deserticola under drought conditions. J PlantPhysiol 161:675–682
Sánchez-Castro I, Ferrol N, Cornejo P, Barea JM (2012) Temporaldynamics of arbuscular mycorrhizal fungi colonizing roots ofrepresentative shrub species in a semi-arid Mediterraneanecosystem. Mycorrhiza 22:449–460
Sekeroglu N, Ozkutlu F, Kara SM, Ozguven M (2008)Determination of cadmium and selected micronutrients in
commonly used and traded medicinal plants in Turkey. JSci Food Agric 88:86–90
Testiati E, Parinet J, Massiani C, Laffont-Schwob I, Rabier J,Pfeifer HR, Lenoble V, Masotti V, Prudent P (2013) Tracemetal and metalloid contamination levels in soils and in twonative plant species of a former industrial site: evaluation ofthe phytostabilization potential. J Hazard Mater 248–249:131–141
Tschan M, Robinson BH, Schulin R (2009) Antimony in the soil–plant system—a review. Environ Chem 6:106–115
Turrini A, Sbrana C, Strani P, Pezzarossa B, Risaliti R, GiovannettiM (2010) Arbuscular mycorrhizal fungi of a Mediterraneanisland (Pianosa), within a UNESCOBiosphere Reserve. BiolFertil Soils 46:511–520
Vamerali T, Bandiera M, Coletto L, Zanetti F, Dickinson NM,Mosca G (2009) Phytoremediation trials on metal- andarsenic-contaminated pyrite wastes (Torviscosa, Italy).Environ Pollut 157:887–894
Vodnik D, Grčman H, Maček I, van Elteren JT, Kovacevic M(2008) The contribution of glomalin-related soil protein to Pband Zn sequestration in polluted soil. Sci Total Environ 392:130–136
Whitfield L, Richards AJ, Rimmer DL (2004) Relationships be-tween soil heavy metal concentration and mycorrhizal colo-nisation in Thymus polytrichus in northern England.Mycorrhiza 14:55–62
Wilson SC, Lockwood PV, Ashley PM, Tighe M (2010) Thechemistry and behaviour of antimony in the soil environmentwith comparisons to arsenic: a critical review. Environ Pollut158:1169–1181
Wright SF, Upadhyaya A (1998) A survey of soils for aggregatestability and glomalin, a glycoprotein produced by hyphae ofarbuscular mycorrhizal fungi. Plant Soil 198:97–107
Wuana RA, Okieimen FE (2011) Heavy metals in contaminatedsoils: a review of sources, chemistry, risks and best availablestrategies for remediation. Int Sch Res Netw Ecol 2011:1–20
Xiong JB, Mahmood Q, Yue M (2011) The potential of Sedumalfredii Hance for the biosorption of some metals from syn-thetic wastewater. Desalination 267:154–159
Yang R, Yu G, Tang J, Chen X (2008) Effects of metal lead ongrowth and mycorrhizae of an invasive plant species(Solidago canadensis L.). J Environ Sci-China 20:739–744
Yaylali-AbanuzG (2011) Heavymetal contamination of surface soilaround Gebze industrial area, Turkey. Microchem J 99:82–92
Zhang HH, Tang M, Chen H, Zheng CL, Niu ZC (2010) Effect ofinoculation with AM fungi on lead uptake, translocation andstress alleviation of Zea mays L. seedlings planting in soilwith increasing lead concentrations. Eur J Soil Biol 46:306–311
Zheng N, Wang Q, Zheng D (2007) Health risk of Hg, Pb,Cd, Zn, and Cu to the inhabitants around Huludao ZincPlant in China via consumption of vegetables. Sci TotalEnviron 383:81–89
236 Plant Soil (2014) 382:219–236




















