
Artificial selection of biotypes of green rice leafhopper, Nephotettix cincticeps Uhler (Homoptera:...
11
INTRODUCTION The use of crop resistance is an effective method of integrated pest management. It offers the advan- tages of a relatively small effect on the environ- ment, reduced labor, and lower costs. However, a serious problem is that the resistance of crops has broken down with the occurrence of biotypes of pest insects that are virulent to previously resistant crop varieties. To use plant resistance for insect pest control, we should clarify the potential of pests to overcome resistance and predict the occur- rence of such biotypes. The existence of resistance-breaking biotypes has been reported in many crops (Sogawa, 1983a, b; Diehl and Bush, 1984). Such a biotype of the brown planthopper, Nilaparvata lugens, was ob- served after resistant varieties of rice were culti- vated for a few years (Sogawa, 1982; Gallagher et al., 1994). Several studies showed that N. lugens populations have genetic variation in virulence to resistant rice varieties using artificial selection in the laboratory (den Hollander and Pathak, 1981; Ito and Kisimoto, 1981; Claridge and den Hol- lander, 1982) and quantitative genetic analysis (Tanaka, 1999). The artificial selection of biotypes was also carried out in the green leafhopper, Nephotettix virescens (Kobayashi et al., 1983; Takita and Habibuddin, 1985). The green rice leafhopper (GRH), Nephotettix Artificial selection of biotypes of green rice leafhopper, Nephotettix cincticeps Uhler (Homoptera: Cidadellidae), and virulence to resistant rice varieties Masahiro HIRAE, 1, * Yoshimichi FUKUTA, 2 Katsunori TAMURA 3 and Shingo OYA 1 1 Hokuriku Research Center, National Agricultural Research Center, National Agriculture and Food Research Organization; Joetsu 943–0193, Japan 2 Japan International Research Center for Agricultural Sciences; Tsukuba 305–8686, Japan 3 National Agricultural Research Center for Kyushu Okinawa Region, National Agriculture and Food Research Organization; Chikugo 833–0041, Japan (Received 10 July 2006; Accepted 17 October 2006) Abstract A population of green rice leafhopper, Nephotettix cincticeps Uhler (GRH), was artificially selected on 5 resistant rice varieties in the laboratory. The GRH lines selected on Saikai 164, Saikai 182, and Kanto-PL 6 were able to survive and reproduce on their respective varieties. In these lines, the developmental period of nymphs was shortened by con- tinuous selection, although in the first generation it was longer than that of the line reared on Nipponbare carrying no resistance gene. We could not establish GRH lines virulent to Norin-PL 5 or Norin-PL 6. Six lines, reared on IR 24, Chugoku 105, Saikai 164, Saikai 182, Kanto-PL 6, and Aichi 80, were assessed for virulence among different resis- tant varieties by conducting a seedling test. All lines were highly virulent to the varieties on which they were selected. Similar virulence was obtained for combinations of IR 24 and Chugoku 105 lines, Saikai 164 and Saikai 182 lines, and Kanto-PL 6 and Aichi 80 lines. The Kanto-PL 6 and Aichi 80 lines were moderately virulent to Tadukan and Rantaj-emas 2. No lines were virulent to Norin-PL 5, Norin-PL 6, and Pe-bi-hun. The results of the leaf blade test were similar to those of the seedling test. The allele test confirmed that the GRH-resistance genes of Kanto-PL 6 and Aichi 80 are the same, and the resistance gene of Norin-PL 2 is located at a different locus from that of Kanto-PL 6 and Aichi 80. These results suggest that the virulence of GRH biotypes corresponds to the resistance genes of rice va- rieties and therefore, the use of biotypes allows the identification of groups of rice varieties that have similar resis- tance genes. We propose that the biotypes virulent to Grh 1-, Grh 2-, and Grh 3 (t)-carrying varieties be designated “Biotype 1”, “Biotype 2”, and “Biotype 3”, respectively. Key words: Nephotettix cincticeps; rice; resistance; biotypes; selection * To whom correspondence should be addressed at: E-mail: [email protected] DOI: 10.1303/aez.2007.97
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Artificial selection of biotypes of green rice leafhopper, Nephotettix cincticeps Uhler (Homoptera:...

INTRODUCTION
The use of crop resistance is an effective methodof integrated pest management. It offers the advan-tages of a relatively small effect on the environ-ment, reduced labor, and lower costs. However, aserious problem is that the resistance of crops hasbroken down with the occurrence of biotypes ofpest insects that are virulent to previously resistantcrop varieties. To use plant resistance for insectpest control, we should clarify the potential ofpests to overcome resistance and predict the occur-rence of such biotypes.
The existence of resistance-breaking biotypeshas been reported in many crops (Sogawa, 1983a,
b; Diehl and Bush, 1984). Such a biotype of thebrown planthopper, Nilaparvata lugens, was ob-served after resistant varieties of rice were culti-vated for a few years (Sogawa, 1982; Gallagher etal., 1994). Several studies showed that N. lugenspopulations have genetic variation in virulence toresistant rice varieties using artificial selection inthe laboratory (den Hollander and Pathak, 1981;Ito and Kisimoto, 1981; Claridge and den Hol-lander, 1982) and quantitative genetic analysis(Tanaka, 1999). The artificial selection of biotypeswas also carried out in the green leafhopper,Nephotettix virescens (Kobayashi et al., 1983;Takita and Habibuddin, 1985).
The green rice leafhopper (GRH), Nephotettix
Artificial selection of biotypes of green rice leafhopper, Nephotettix cincticepsUhler (Homoptera: Cidadellidae), and virulence to resistant rice varieties
Masahiro HIRAE,1,* Yoshimichi FUKUTA,2 Katsunori TAMURA3 and Shingo OYA1
1 Hokuriku Research Center, National Agricultural Research Center, National Agriculture and Food Research Organization; Joetsu943–0193, Japan
2 Japan International Research Center for Agricultural Sciences; Tsukuba 305–8686, Japan3 National Agricultural Research Center for Kyushu Okinawa Region, National Agriculture and Food Research Organization;Chikugo 833–0041, Japan
(Received 10 July 2006; Accepted 17 October 2006)
AbstractA population of green rice leafhopper, Nephotettix cincticeps Uhler (GRH), was artificially selected on 5 resistant ricevarieties in the laboratory. The GRH lines selected on Saikai 164, Saikai 182, and Kanto-PL 6 were able to surviveand reproduce on their respective varieties. In these lines, the developmental period of nymphs was shortened by con-tinuous selection, although in the first generation it was longer than that of the line reared on Nipponbare carrying noresistance gene. We could not establish GRH lines virulent to Norin-PL 5 or Norin-PL 6. Six lines, reared on IR 24,Chugoku 105, Saikai 164, Saikai 182, Kanto-PL 6, and Aichi 80, were assessed for virulence among different resis-tant varieties by conducting a seedling test. All lines were highly virulent to the varieties on which they were selected.Similar virulence was obtained for combinations of IR 24 and Chugoku 105 lines, Saikai 164 and Saikai 182 lines,and Kanto-PL 6 and Aichi 80 lines. The Kanto-PL 6 and Aichi 80 lines were moderately virulent to Tadukan andRantaj-emas 2. No lines were virulent to Norin-PL 5, Norin-PL 6, and Pe-bi-hun. The results of the leaf blade testwere similar to those of the seedling test. The allele test confirmed that the GRH-resistance genes of Kanto-PL 6 andAichi 80 are the same, and the resistance gene of Norin-PL 2 is located at a different locus from that of Kanto-PL 6and Aichi 80. These results suggest that the virulence of GRH biotypes corresponds to the resistance genes of rice va-rieties and therefore, the use of biotypes allows the identification of groups of rice varieties that have similar resis-tance genes. We propose that the biotypes virulent to Grh 1-, Grh 2-, and Grh 3 (t)-carrying varieties be designated“Biotype 1”, “Biotype 2”, and “Biotype 3”, respectively.
Key words: Nephotettix cincticeps; rice; resistance; biotypes; selection
* To whom correspondence should be addressed at: E-mail: [email protected]: 10.1303/aez.2007.97

cincticeps Uhler, is a serious pest in temperateAsia. Populations of this insect, which multiplyrapidly around the rice heading stage, can reachhigh levels and cause considerable losses of riceyield in northern Japan, though their peak densityis relatively low in southwestern Japan (Kidokoro,1979; Itô and Johraku, 1982; Satomi, 1993; Hiranoand Fujii, 1995). GRH also causes damage to riceindirectly by transmitting viral and phytoplasmaldiseases such as rice dwarf disease, waika disease,and yellow dwarf disease (Fukushi, 1934; Shinkai,1962; Hirao et al., 1974). As some indica varietiesare resistant to GRH (Inoue, 1966), resistancegenes from these varieties have been introducedinto japonica rice for breeding resistant varietiesand lines (Shumiya et al., 1984; Kaneda et al.,1985; Ono et al., 1986; Kishino et al., 1987;Tokura et al., 1995; Nakajima et al., 1998). In thefield, no resistance-breaking biotypes of GRH werefound after cultivation with the resistant varieties.In the laboratory, however, biotypes virulent to the rice breeding lines, Saikai 182 and Aichi 42,were selected using a GRH population collectedfrom Chikugo, Fukuoka Prefecture (Takita andNishiyama, 1989). At present, many GRH-resistantvarieties and lines have different genetic sources(Table 1); therefore, it is necessary to examine thepossibility that biotypes occur on those varieties.Virulence to IR 24 was different between popu-lations of GRH from the southern district(Kagoshima, Chikugo and Kainan) and northerndistrict (Mattou, Toyama, Joetsu and Teradomari)in Japan (Sato and Sogawa, 1981; Sogawa andSato, 1981). Therefore, the possibility of biotypeoccurrence in the Joetsu (northern Japan) popula-
tion also needs to be estimated as well as theChikugo (southern Japan) population. In this paper,we examine the development of virulence in GRHby subjecting the Joetsu population to artificial se-lection on several resistant varieties and lines. Wealso evaluate the virulence of selected GRH linesto clarify the differences of their virulence amongresistant varieties and between rice growth stages.In addition, we analyze the relationship betweenresistance genes by conducting allele tests amongsome resistant varieties. Furthermore, we classifybiotypes of GRH by examining the relationship be-tween the resistance genes of rice varieties and theselected GRH lines.
MATERIALS AND METHODS
Insects. The GRH used in this study was col-lected in paddy fields cultivated with a japonicarice variety, Nipponbare, at the Hokuriku ResearchCenter in October 1993. Nipponbare has no knownGRH-resistance genes. The insects were main-tained on Nipponbare seedlings by the method ofSugimoto (1969). The rearing of insects and theexperiments described below were carried out in anair-conditioned insectary at 25°C with a 16L : 8Dphotoperiod.
Plants. The rice varieties used in this study arelisted in Table 1. Saikai 164, Saikai 182, Kanto-PL6, Norin-PL 5 (former name: Saikai-PL 2), andNorin-PL 6 (former name: Ou-PL 1) carryingGRH-resistance genes were used for the selectionof biotypes and Nipponbare was used as a controlvariety. In addition to these six varieties, IR 24,Norin-PL 2 (former name: Kanto-PL 3), Chugoku
Table 1. List of rice varieties used in this study and their genetic sources resistant to N. cincticeps
VarietyDonor parent for
Resistance gene ReferenceN. cincticeps resistance
Nipponbare No resistance geneIR 24 Grh 1 International Rice Research Institute (1972)Norin-PL 2 Pe-bi-hun Grh 1 Kaneda et al. (1985)Chugoku 105 Norin-PL 2 Grh 1 National Agricultural Research Center (1995)Saikai 164 C203-1 Unknown National Agricultural Research Center (1995)Saikai 182 Norin-PL 5 Grh 2 Takita and Nishiyama (1989)Kanto-PL 6 Tadukan Unknown Kobayashi et al. (1980)Aichi 80 Rantaj-emas 2 Grh 3 (t) Nakagome et al. (1989)Norin-PL 5 C203-1 Grh 2�Grh 4 Ono et al. (1986)Norin-PL 6 Lepe dumai Grh 2�Grh 4 Kishino et al. (1987)

105, and Aichi 80, and three of their geneticsources, i.e. Pe-bi-hun, Tadukan, and Rantaj-emas2, were used for testing the virulence of the se-lected GRH lines among different resistant vari-eties.
Selection of GRH lines virulent to resistantvarieties. About 600 rice seeds of each varietywere sown in a plastic petri dish (9 cm in diameter,2.2 cm high) containing 60 ml of moist soil. Afterfour days, during which the rice seedlings grew tothe second leaf stage, 150 to 200 first-instarnymphs of GRH were released within 8 h of hatch-ing on rice seedlings that were covered by a cylin-drical vinyl chloride cage (8.5 cm in diameter,15 cm high) equipped with three Tetoron® gauzewindows. As a rule, seedlings were exchangedevery five days. Emerged adults were moved to an-other cage that contained rice seedlings of the re-spective variety used for selection and allowed tooviposit to produce the next generation. These se-lection procedures were continued for several gen-erations. In this paper, the name of the selected lineof GRH, e.g. the “IR 24 line”, is based on the vari-ety from which the line was selected.
Development and survival. Developmental pe-riods and survival rates of GRH nymphs were ob-served to examine the changes in these traits by ar-tificial selection. Fifty first-instar nymphs were re-leased within 8 h of hatching into a cylindrical cagecontaining about 150 rice seedlings of the varietyused for selection. The seedlings were exchangedevery five days. Each selected line was tested inthree cages (150 nymphs). The emerging adultswere removed from the cage and the number ofemerging adults and the nymphal period to emer-gence were recorded daily. The survival rate ofnymphs was calculated as the average per cage inthree replications.
Virulence of selected GRH lines among resis-tant varieties. The virulence of each GRH line se-lected for eight generations was evaluated by aseedling test (Hirae et al., 2002) and a leaf-bladetest (Kishino and Ando, 1978) to clarify the differ-ences in virulence among various resistant varietiesand between rice growth stages. In the seedlingtest, one seedling at the second leaf stage (fourdays after seeding) was placed into a test tube(1.8 cm in diameter, 18 cm high) with 0.5 ml ofwater. Five first-instar nymphs were released within8 h of hatching into the test tube, which was
plugged with cotton covered with Tetoron gauze.Ten seedlings of each variety were tested. Afterfour days, nymphs that had developed into secondinstars were counted. Virulence was evaluated asthe percentage of second-instar nymphs among the50 nymphs released. In addition to the Saikai 164,Saikai 182, and Kanto-PL 6 lines of GRH, selectedin the present study, three more lines were estab-lished by selection on IR 24, Chugoku 105, andAichi 80 for more than 11 generations. These sixlines were tested for their virulence to the 13 resis-tant varieties described above (Table 1).
In the leaf-blade test, test leaves were collectedfrom rice plants cultivated in paddy fields at theHokuriku Research Center. Six rice varieties, Nip-ponbare, Chugoku 105, Saikai 182, Kanto-PL 6,Aichi 80, and Norin-PL 5, were transplanted on 16May 1997 and the base of the most recently fullyexpanded leaf was cut at a length of 15 cm on 17August (panicle formation stage to heading stage).The detached leaf was placed into a test tube(1.8 cm in diameter, 18 cm high) with 3 ml ofwater. Ten first-instar nymphs of each GRH line se-lected over more than 14 generations were releasedand the nymphs that had developed into the secondinstar were counted after 4 days. Five replicationswere made for each variety.
Allelic relationships between resistance genes.As the virulence test of the selected GRH linessuggests that the relationship of resistance betweenNorin-PL 2, Kanto-PL 6, and Aichi 80 is differentfrom a previous report (Takita and Nishiyama,1989) (see Results), allele tests between resistancegenes were carried out by crossing the three vari-eties. F2 progeny from each cross of the three vari-eties was sown in a seedling cup (9 cm in diameter,4.5 cm high) containing 60 ml of moist soil. Resis-tance of the individual seedlings was examined bythe seedling test described above, using a non-se-lected line. We defined seedlings on which the pro-portion of nymphs developing into second instarswas �40% and �60% as resistant and susceptible,respectively, according to Tamura et al. (1999) andHirae et al. (2002). The relationship is expected tobe non-allelic if the segregation ratio of resistanceto susceptible progeny is consistent with 15 : 1, andallelic if all progeny shows resistance (Kobayashi,1983; Ikeda, 1985).
Statistical analysis. The developmental periodof nymphs was analyzed using ANOVA and the
Tukey-Kramer method. The survival rate ofnymphs and the percentage of nymphs developinginto second instars were analyzed using ANOVAand the Tukey method after arcsine transformationof the data. In the allele test, chi-square tests wereperformed using the expected segregation ratio of15 : 1 or 1 : 0.
RESULTS
Selection of GRH lines virulent to resistant vari-ety
The results of artificial selection experiments areshown in Table 2. In the GRH lines reared continu-ously on Saikai 164 and Saikai 182, a great numberof nymphs survived to adulthood from the firstgeneration. In the line reared on Kanto-PL 6, thenumber of emerging adults was not large in thefirst two generations, but became large from thethird generation; however, the number of adultswas small in the first generation reared on Norin-PL 5 and third generation raised on Norin-PL 6.Accordingly, we did not continue selection usingthese two varieties.
Development and survivalThe developmental period and survival rate of
nymphs for each generation of selection are shownin Table 3. In the Saikai 164 line and Saikai 182line, the developmental period was longer by 2–4 dthan that of the Nipponbare line in the first genera-tion. However, the developmental period was short-ened by continuous selection, and was not signifi-cantly different from that of the Nipponbare line inthe sixth generation. In these two lines, the survival
rate was almost the same as that of the Nipponbareline in the first generation. In the Kanto-PL 6 line,the developmental period was prolonged in the firstgeneration but did not differ significantly from thatof the Nipponbare line in the fourth generation. Inthis line, although the survival rate was lower thanthat of the Nipponbare line in the first generation, itrose from the second generation. The Norin-PL 5and Norin-PL 6 lines showed low survival rates inthe first generation.
Virulence of selected GRH lines among resistantvarieties
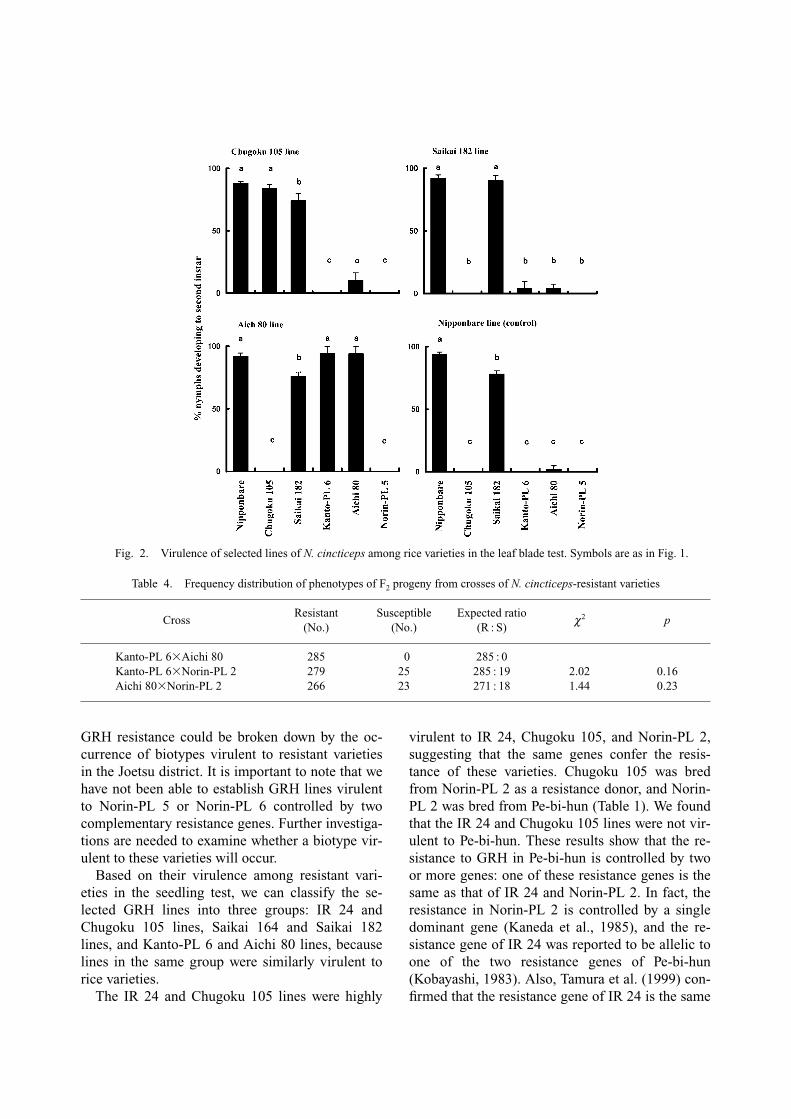
In the seedling test, the virulence of the selectedGRH lines among resistant varieties was evaluatedbased on the percentage of nymphs developing intosecond instars (Fig. 1). All 6 GRH lines (IR 24,Chugoku 105, Saikai 164, Saikai 182, Kanto-PL 6and Aichi 80) were highly virulent to the varietieson which they were selected. The IR 24 andChugoku 105 lines were also highly virulent toother varieties, i.e. IR 24, Norin-PL 2, andChugoku 105, but were not virulent to Pe-bi-hunwhich is the source of the resistance of Norin-PL 2.The Saikai 164 and Saikai 182 lines were highlyvirulent to Saikai 164 and Saikai 182; however,these lines had low virulence in Norin-PL 5, whichis the source of the resistance of Saikai 182. Thelines selected on Kanto-PL 6 and Aichi 80 werevirulent to both Kanto-PL 6 and Aichi 80, andmoderately virulent to Tadukan and Rantaj-emas 2which are the sources of the resistance of Kanto-PL 6 and Aichi 80, respectively. No lines were vir-ulent to Norin-PL 5, Norin-PL 6, and Pe-bi-hun.
The results of the leaf blade test are shown in
Table 2. The number of N. cincticeps nymphs released and the percentage of adults emerged on resistant varieties and a control (Nipponbare)
GenerationRice variety
1 2 3 4 5 6 7 8
Nipponbare 95.0 (300) 95.7 (300) 89.3 (300) 94.0 (300) 96.7 (150) 97.3 (150) 93.3 (150) 90.0 (150)Saikai 164 90.3 (300) 93.3 (300) 93.3 (300) 88.3 (300) 90.0 (150) 87.3 (150) 89.3 (150) 97.3 (150)Saikai 182 88.3 (300) 83.7 (300) 85.7 (300) 84.0 (300) 84.7 (150) 90.7 (150) 88.0 (150) 97.3 (150)Kanto-PL 6 37.8 (1,300) 69.6 (500) 74.2 (589) 74.0 (300) 88.0 (150) 88.0 (150) 89.3 (150) 94.0 (150)Norin-PL 5 0.4 (1,900)Norin-PL 6 7.9 (1,900) 41.3 (189) 26.1 (69)
The value shows the percentage of nymphs developing into adults. The numerical value in parentheses shows the number of firstinstar nymphs released on each variety.
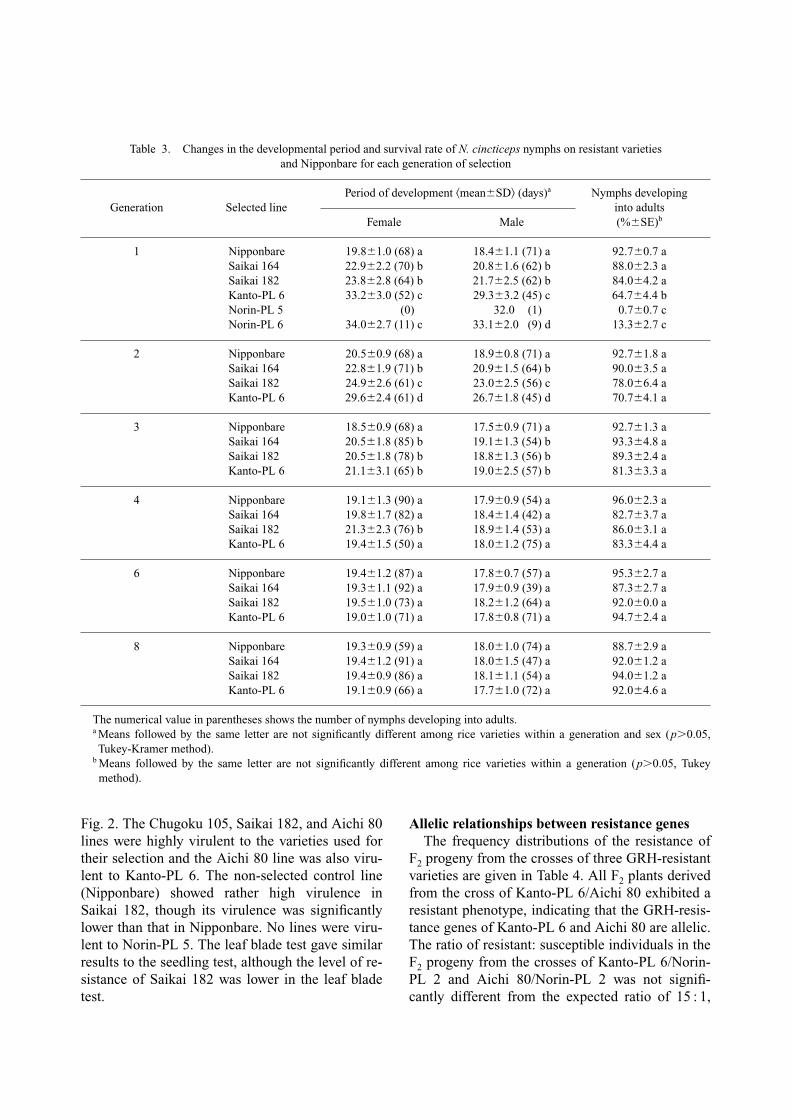
Fig. 2. The Chugoku 105, Saikai 182, and Aichi 80lines were highly virulent to the varieties used fortheir selection and the Aichi 80 line was also viru-lent to Kanto-PL 6. The non-selected control line(Nipponbare) showed rather high virulence inSaikai 182, though its virulence was significantlylower than that in Nipponbare. No lines were viru-lent to Norin-PL 5. The leaf blade test gave similarresults to the seedling test, although the level of re-sistance of Saikai 182 was lower in the leaf bladetest.
Allelic relationships between resistance genesThe frequency distributions of the resistance of
F2 progeny from the crosses of three GRH-resistantvarieties are given in Table 4. All F2 plants derivedfrom the cross of Kanto-PL 6/Aichi 80 exhibited aresistant phenotype, indicating that the GRH-resis-tance genes of Kanto-PL 6 and Aichi 80 are allelic.The ratio of resistant: susceptible individuals in theF2 progeny from the crosses of Kanto-PL 6/Norin-PL 2 and Aichi 80/Norin-PL 2 was not signifi-cantly different from the expected ratio of 15 : 1,
Table 3. Changes in the developmental period and survival rate of N. cincticeps nymphs on resistant varieties and Nipponbare for each generation of selection
Period of development �mean�SD� (days)a Nymphs developing Generation Selected line into adults
Female Male (%�SE)b
1 Nipponbare 19.8�1.0 (68) a 18.4�1.1 (71) a 92.7�0.7 aSaikai 164 22.9�2.2 (70) b 20.8�1.6 (62) b 88.0�2.3 aSaikai 182 23.8�2.8 (64) b 21.7�2.5 (62) b 84.0�4.2 aKanto-PL 6 33.2�3.0 (52) c 29.3�3.2 (45) c 64.7�4.4 bNorin-PL 5 (0) 32.0 (1) 0.7�0.7 cNorin-PL 6 34.0�2.7 (11) c 33.1�2.0 (9) d 13.3�2.7 c
2 Nipponbare 20.5�0.9 (68) a 18.9�0.8 (71) a 92.7�1.8 aSaikai 164 22.8�1.9 (71) b 20.9�1.5 (64) b 90.0�3.5 aSaikai 182 24.9�2.6 (61) c 23.0�2.5 (56) c 78.0�6.4 aKanto-PL 6 29.6�2.4 (61) d 26.7�1.8 (45) d 70.7�4.1 a
3 Nipponbare 18.5�0.9 (68) a 17.5�0.9 (71) a 92.7�1.3 aSaikai 164 20.5�1.8 (85) b 19.1�1.3 (54) b 93.3�4.8 aSaikai 182 20.5�1.8 (78) b 18.8�1.3 (56) b 89.3�2.4 aKanto-PL 6 21.1�3.1 (65) b 19.0�2.5 (57) b 81.3�3.3 a
4 Nipponbare 19.1�1.3 (90) a 17.9�0.9 (54) a 96.0�2.3 aSaikai 164 19.8�1.7 (82) a 18.4�1.4 (42) a 82.7�3.7 aSaikai 182 21.3�2.3 (76) b 18.9�1.4 (53) a 86.0�3.1 aKanto-PL 6 19.4�1.5 (50) a 18.0�1.2 (75) a 83.3�4.4 a
6 Nipponbare 19.4�1.2 (87) a 17.8�0.7 (57) a 95.3�2.7 aSaikai 164 19.3�1.1 (92) a 17.9�0.9 (39) a 87.3�2.7 aSaikai 182 19.5�1.0 (73) a 18.2�1.2 (64) a 92.0�0.0 aKanto-PL 6 19.0�1.0 (71) a 17.8�0.8 (71) a 94.7�2.4 a
8 Nipponbare 19.3�0.9 (59) a 18.0�1.0 (74) a 88.7�2.9 aSaikai 164 19.4�1.2 (91) a 18.0�1.5 (47) a 92.0�1.2 aSaikai 182 19.4�0.9 (86) a 18.1�1.1 (54) a 94.0�1.2 aKanto-PL 6 19.1�0.9 (66) a 17.7�1.0 (72) a 92.0�4.6 a
The numerical value in parentheses shows the number of nymphs developing into adults.a Means followed by the same letter are not significantly different among rice varieties within a generation and sex (p�0.05,Tukey-Kramer method).
b Means followed by the same letter are not significantly different among rice varieties within a generation (p�0.05, Tukeymethod).
confirming independent segregation of the twodominant genes. These results show that the GRH-resistance gene of Norin-PL 2 is certainly locatedon a different locus from that of Kanto-PL 6 andAichi 80.
DISCUSSION
It was reported that GRH populations fromKagoshima, Chikugo, and Kainan (Tokushima Pre-fecture) were easily reared for successive genera-tions on IR 24, whereas populations from Mattou
(Ishikawa Pref.), Toyama, Joetsu, and Teradomari(Niigata Pref.) could not be raised on IR 24 (Sogawa and Sato, 1981). Takita and Nishiyama(1989) established two selected lines of GRH, onevirulent to Aichi 42 and the other virulent to Saikai182, using the Chikugo population. In the presentstudy, GRH lines which could survive and repro-duce on several resistant varieties controlled bysingle resistance gene were established using theJoetsu population. This result shows that the GRHpopulation from Joetsu has genetic variations invirulence to resistant varieties. It also suggests that
Fig. 1. Virulence of selected lines of N. cincticeps among rice varieties in the seedling test. Columns and vertical bars showthe means and standard errors of the percentage of nymphs which developed to second instars. For each line, columns with thesame letter are not significantly different in virulence among the rice varieties at 0.05 (Tukey method).
GRH resistance could be broken down by the oc-currence of biotypes virulent to resistant varietiesin the Joetsu district. It is important to note that wehave not been able to establish GRH lines virulentto Norin-PL 5 or Norin-PL 6 controlled by twocomplementary resistance genes. Further investiga-tions are needed to examine whether a biotype vir-ulent to these varieties will occur.
Based on their virulence among resistant vari-eties in the seedling test, we can classify the se-lected GRH lines into three groups: IR 24 andChugoku 105 lines, Saikai 164 and Saikai 182lines, and Kanto-PL 6 and Aichi 80 lines, becauselines in the same group were similarly virulent torice varieties.
The IR 24 and Chugoku 105 lines were highly
virulent to IR 24, Chugoku 105, and Norin-PL 2,suggesting that the same genes confer the resis-tance of these varieties. Chugoku 105 was bredfrom Norin-PL 2 as a resistance donor, and Norin-PL 2 was bred from Pe-bi-hun (Table 1). We foundthat the IR 24 and Chugoku 105 lines were not vir-ulent to Pe-bi-hun. These results show that the re-sistance to GRH in Pe-bi-hun is controlled by twoor more genes: one of these resistance genes is thesame as that of IR 24 and Norin-PL 2. In fact, theresistance in Norin-PL 2 is controlled by a singledominant gene (Kaneda et al., 1985), and the re-sistance gene of IR 24 was reported to be allelic toone of the two resistance genes of Pe-bi-hun(Kobayashi, 1983). Also, Tamura et al. (1999) con-firmed that the resistance gene of IR 24 is the same
Fig. 2. Virulence of selected lines of N. cincticeps among rice varieties in the leaf blade test. Symbols are as in Fig. 1.
Table 4. Frequency distribution of phenotypes of F2 progeny from crosses of N. cincticeps-resistant varieties
Resistant Susceptible Expected ratioCross
(No.) (No.) (R : S)c 2 p
Kanto-PL 6�Aichi 80 285 0 285 : 0Kanto-PL 6�Norin-PL 2 279 25 285 : 19 2.02 0.16Aichi 80�Norin-PL 2 266 23 271 : 18 1.44 0.23

as that of Norin-PL 2. The results of this study areconsistent with these reports.
We also found that the Saikai 164 and Saikai182 lines were virulent to Saikai 164 and Saikai182 but not to Norin-PL 5. Saikai 182 was bredfrom Norin-PL 5, and Saikai 164 and Norin-PL 5were bred from C203-1 (Table 1). The resistance inNorin-PL 5 is controlled by two complementarygenes (Imbe and Iwasaki, 1987). These genes, des-ignated Grh 2 and Grh 4, are located on chromo-some 11 and chromosome 3, respectively, of therice plant (Tamura et al., 1997; Fukuta et al.,1998). Resistance in Saikai 182 is controlled by asingle gene (Grh 2) (Tamura et al., 1998). Saikai164 probably has the single resistance gene Grh 2the same as Saikai 182. Thus, the Saikai 182 andSaikai 164 lines of GRH may adapt to the resis-tance conferred by Grh 2 but not to the resistanceof Norin-PL 5, which is controlled by a combina-tion of Grh 2 and Grh 4. It is necessary to confirmthis tentative conclusion by conducting allele tests.
The Kanto-PL 6 line exhibited almost the samevirulence to the rice varieties tested as the Aichi 80line, suggesting that Kanto-PL 6 and Aichi 80 havethe same resistance genes. Takita and Nishiyama(1989) reported that a GRH line selected on Aichi42, which is a parent of Aichi 80, was highly viru-lent to both Kanto-PL 6 and Norin-PL 2, and clas-sified Aichi 42 and Norin-PL 2 into the samegroup. This result was different from that of ourexamination of the virulence of selected GRH
lines. Allelic analysis showed that the GRH-resis-tance genes of Kanto-PL 6 and Aichi 80 are allelicand differ from the gene of Norin-PL 2. Our con-clusion is consistent with the fact that the resis-tance gene (Grh 3 (t)) of Aichi 80 is located onchromosome 6 (Saka et al., 1997), while that ofNorin-PL 2 is located on chromosome 5 (Tamuraet al., 1999). The Chikugo population used byTakita and Nishiyama (1989) was certainly virulentto Grh 1-carrying varieties, e.g. IR 24 and Norin-PL 2 (Sato and Sogawa, 1981; Sogawa and Sato,1981), and therefore, the line selected on Aichi 42was able to adapt to Grh 1-carrying varieties aswell as Grh 3 (t)-carrying varieties.
In the case of the brown planthopper (BPH), Ni-laparvata lugens, resistance genes of rice varietieshave been identified on the basis of the relationshipbetween resistance gene and BPH biotypes (Inter-national Rice Research Institute, 1976; Kaneda et al., 1981; Ikeda, 1985; Nemoto et al., 1989). Asimilar method can be applied in the case of thecharacteristics of GRH biotypes and resistant ricevarieties. We propose that biotypes that have viru-lence to Grh 1-, Grh 2-, and Grh 3 (t)-carrying va-rieties be designated “Biotype 1”, “Biotype 2”, and“Biotype 3”, respectively (Table 5).
The resistance to GRH fluctuated with thegrowth stage of rice plants; it was highest at theyoung seedling stage and approximately 20 daysbefore rice heading (Kishino and Ando, 1979;Takeda and Nagata, 1987; Ando, 1998). In the
Table 5. Reactions of biotypes of N. cincticeps to resistant varieties
Selected line
Rice varietySource of Resistance
Biotype 1 Biotype 2 Biotype 3resistance gene
IR 24 Chugoku 105 Saikai 164 Saikai 182 Kanto-PL 6 Aichi 80
Nipponbare None � � � � � �IR 24 Grh 1 � � � � � �Norin-PL 2 Pe-bi-hun Grh 1 � � � � � �Chugoku 105 Pe-bi-hun Grh 1 � � � � � �Pe-bi-hun Grh 1�a � � � � � �Saikai 164 C203-1 Unknown � � � � � �Saikai 182 C203-1 Grh 2 � � � � � �Norin-PL 5 C203-1 Grh 2�Grh 4 � � � � � �Norin-PL 6 Lepe dumai Grh 2�Grh 4 � � � � � �Kanto-PL 6 Tadukan Grh 3 (t) � � � � � �Aichi 80 Rantaj-emas 2 Grh 3 (t) � � � � � �
�: Virulent, �: Avirulent.

present study, varietal resistance at the riceseedling stage was similar to that at the reproduc-tive stage (panicle formation stage to headingstage). This result indicates that a GRH biotype se-lected on resistant rice seedlings is able to adapt tothat variety from the seedling stage to maturity.
This study revealed that artificial selection couldshift the GRH to a biotype virulent to a previouslyresistant rice variety, and hence, suggests the oc-currence of such biotypes in the field. To predictand prevent the occurrence of resistance-breakingbiotypes, there is a need for further examination ofthe mode of inheritance of the biotypes, genetic re-lationships between biotypes, and relationships be-tween the virulence and life-history traits of GRH.
ACKNOWLEDGEMENTS
We thank Dr. H. Higuchi (National Agricultural ResearchCenter) and Dr. K. Tanaka (National Institute for Agro-Envi-ronmental Sciences) for critical reading and improvement ofthe manuscript; Dr. M. Matsumura (National Agricultural Re-search Center for Kyushu Okinawa Region) and Dr. M. Hattori(National Institute of Agrobiological Sciences) for helpfulsuggestions. We are grateful to the staff of the rice breedinglaboratory of the National Institute of Crop Science, the dis-ease and insect pest control laboratory of the National Agri-cultural Research Center for the Tohoku Region, the ricebreeding laboratory of the National Agricultural ResearchCenter for the Western Region, and the crop group of theAichi-ken Agricultural Research Center for kindly supplyingseeds of the rice varieties.
REFERENCES
Ando, Y. (1998) Changes in the resistance to green riceleafhopper (Nephotettix cincticeps) at various growing-stages of rice cultivars. Jpn. J. Trop. Agric. 42: 54–56(in Japanese).
Claridge, M. F. and J. den Hollander (1982) Virulence to ricecultivars and selection for virulence in populations of thebrown planthopper Nilaparvata lugens. Entomol. Exp.Appl. 32: 213–221.
den Hollander, J. and P. K. Pathak (1981) The genetics of the‘biotypes’ of the brown planthopper, Nilaparvata lugens.Entomol. Exp. Appl. 29: 76–86.
Diehl, S. R. and G. L. Bush (1984) An evolutionary and ap-plied perspective of insect biotypes. Annu. Rev. Ento-mol. 29: 471–504.
Fukushi, T. (1934) Studies on the dwarf disease of rice plant.J. Fac. Agric. Hokkaido Imp. Univ. 37: 41–164.
Fukuta, Y., K. Tamura, M. Hirae and S. Oya (1998) Geneticanalysis of resistance to green rice leafhopper (Nephotet-tix cincticeps Uhler) in rice parental line, Norin-PL 6,using RFLP markers. Breed. Sci. 48: 243–249.
Gallagher, K. D., P. E. Kenmore and K. Sogawa (1994) Judi-cial use of insecticides deter planthopper outbreaks and
extend the life of resistant varieties in Southeast Asianrice. In Planthoppers: Their Ecology and Management(R. F. Denno and T. J. Perfect eds.). Chapman & Hall,New York, pp. 599–614.
Hirae, M., Y. Fukuta, K. Tamura and S. Oya (2002) Amethod for evaluation of rice resistance to green riceleafhopper, Nephotettix cincticeps (Uhler), based onnymphal growth. Jpn. J. Appl. Entomol. Zool. 46: 178–181 (in Japanese with English summary).
Hirano, K. and K. Fujii (1995) The outbreak mechanisms ofthe green rice leafhopper, Nephotettix cincticeps Uhler, innorthern Japan. Res. Popul. Ecol. 37: 259–267.
Hirao, J., H. Satomi and T. Okada (1974) Transmission ofthe “Waika” disease of rice plant by the green riceleafhopper, Nephotettix cincticeps Uhler. Kyushu PlantProt. Res. 20: 128–139 (in Japanese with English sum-mary).
Ikeda, R. (1985) Studies on the inheritance of resistance tothe rice brown planthopper (Nilaparvata lugens Stål) andthe breeding of resistant rice cultivars. Bull. Natl. Agric.Res. Cent. 3: 1–54 (in Japanese with English summary).
Imbe, T. and M. Iwasaki (1987) Inheritance of resistance tothe green rice leafhopper Nephotettix cincticeps Uhlerand dwarf disease in rice Norin-PL 5. Jpn. J. Breed. 37:177–184 (in Japanese with English summary).
Inoue, H. (1966) Larval development of the green riceleafhopper, Nephotettix cincticeps Uhler, and the whiteback planthopper, Sogatella furcifera Horváth, on japon-ica and indica rice varieties. Jpn. J. Appl. Entomol.Zool. Chugoku Branch 8: 17–19 (in Japanese).
International Rice Research Institute (1972) IRRI AnnualReport for 1971. International Rice Research Institute,Los Baños. 238 pp.
International Rice Research Institute (1976) IRRI AnnualReport for 1975. International Rice Research Institute,Los Baños. 479 pp.
Ito, K. and R. Kisimoto (1981) Selection of new biotypes ofthe brown planthopper, Nilaparvata lugens Stål, capableof surviving on resistant rice cultivars. J. Cent. Agric.Exp. Stn. 35: 139–154 (in Japanese).
Itô, Y. and T. Johraku (1982) Differences in population dy-namics of the green rice leafhopper, Nephotettix cincti-ceps Uhler (Hemiptera: Deltocephalidae), in two districtsof Japan. Appl. Entomol. Zool. 17: 337–349.
Kaneda, C., K. Ito and R. Ikeda (1981) Screening of rice cul-tivars for resistance to the brown planthopper, Nila-parvata lugens Stål, by three biotypes. Jpn. J. Breed.31: 141–151.
Kaneda, C., M. Yokoo, H. Ikehashi, A. Kobayashi, R. Ikedaand H. Nemoto (1985) Breeding of rice Norin-PL 2, anew germplasm with resistance to green rice leafhopperand dwarf virus disease. Bull. Natl. Agric. Res. Cent. 5:81–91 (in Japanese with English summary).
Kidokoro, T. (1979) Geographic trend in the annual popula-tion fluctuation of the green rice leafhopper, Nephotettixcincticeps Uhler (Hemiptera: Deltcephalidae). Appl.Entomol. Zool. 14: 127–129.
Kishino, K. and Y. Ando (1978) Insect resistance of the riceplant to the green rice leafhopper Nephotettix cincticeps

Uhler 1. Laboratory technique for testing the antibiosis.Jpn. J. Appl. Entomol. Zool. 22: 169–177 (in Japanese).
Kishino, K. and Y. Ando (1979) Resistance of rice plant tothe green rice leafhopper, Nephotettix cincticeps Uhler 2.Fluctuation of antibiosis with the growing stages of theresistant rice varieties. Jpn. J. Appl. Entomol. Zool. 23:129–133 (in Japanese with English summary).
Kishino, K., Y. Ando, T. Suzuki, S. Kawabe, M. Takeda, R.Ikeda and S. Saito (1987) Breeding of rice “Norin-PL6”, a new germplasm with resistance to the green riceleafhopper, Nephotettix cincticeps Uhler (Homoptera:Deltocephalidae). Bull. Tohoku Natl. Agric. Exp. Stn.76: 1–11 (in Japanese).
Kobayashi, A. (1983) Inheritance of resistance to green riceleafhopper (Nephotettix cincticeps Uhler) and dwarf virusdisease. In Crop Improvement Research: Proceedings ofFourth International SABRAO Congress (T. C. Yap, K. M.Graham and J. Sukaimi eds.). SABRAO, Malaysia, pp.157–165.
Kobayashi, A., C. Kaneda, R. Ikeda and H. Ikehashi (1980)“Kanto-PL 6”, a new parental line of rice resistant todwarf virus. Jpn. J. Breed. 30 (suppl. 2): 102–103 (inJapanese).
Kobayashi, A., M. A. Supaad and O. Othman (1983) Inheri-tance of resistance of rice to tungro and biotype selectionof green leafhopper in Malaysia. Jpn. Agric. Res. Q. 16:306–311.
Nakagome, M., M. Takimoto and Y. Uebayashi (1989) In-secticide susceptibility of the green rice leafhopper,Nephotettix cincticeps Uhler and resistance to insect ofthe rice breeding line Aichi 80. Res. Bull. Aichi Agric.Res. Cent. 21: 78–84 (in Japanese with English sum-mary).
Nakajima, Y., N. Sugiura, N. Saka, T. Kato, T. Toyama, K.Fujii, S. Kudo, T. Tsuji, T. Izawa and A. Shumiya (1998)Development of an isogenic line of Koshihikari withgreen rice leafhopper resistance gene. Res. Bull. AichiAgric. Res. Cent. 30: 57–61 (in Japanese with Englishsummary).
National Agricultural Research Center (1995) Passport Dataof Rice Varieties and Breeding Lines (1995 edition).Agriculture, Forestry and Fisheries Technical InformationSociety, Tokyo. 247 pp. (in Japanese).
Nemoto, H., R. Ikeda and C. Kaneda (1989) New genes forresistance to brown planthopper, Nilaparvata lugens Stål,in rice. Jpn. J. Breed. 39: 23–28.
Ono, T., M. Okada, S. Watanabe, H. Nishiyama, H. Moto-mura, T. Imbe, E. Shimura, K. Wasano and Y. Akama(1986) A new breeding cultivar “Rice Norin-PL 5” withresistance to green rice leafhopper and dwarf virus dis-ease. Kyushu Agric. Res. 48: 27 (in Japanese).
Saka, N., T. Toyama, T. Tsuji, H. Nakamae and T. Izawa(1997) Fine mapping of green rice leafhopper resis-tance gene Grh-3 (t) and screening of Grh-3 (t) amonggreen rice leafhopper resistant and green rice leafhopperresistant cultivars in rice. Breed. Sci. 47 (suppl. 1): 55(in Japanese).
Sato, A. and K. Sogawa (1981) Biotypic variations in thegreen rice leafhopper, Nephotettix cincticeps Uhler (Ho-
moptera: Deltocephalidae), in relation to rice varieties.Appl. Entomol. Zool. 16: 55–57.
Satomi, H. (1993) Factors responsible for the difference indensities between southwestern and northern Japanesepopulations of the green rice leafhopper, Nephotettixcincticeps (Uhler) (Hemiptera: Deltocephalidae), in theirlater generations I. Effects of the heading time of rice va-rieties on the oviposition and propagation of the secondgeneration. Appl. Entomol. Zool. 28: 207–216.
Shinkai, A. (1962) Studies on insect transmissions of ricevirus diseases in Japan. Bull. Natl. Inst. Agric. Sci., Ser.C. 14: 1–112 (in Japanese with English summary).
Shumiya, A., T. Koumura, I. Shaku, M. Takamatsu, S. Kudo,Y. Nakajima and T. Kato (1984) Rice breeding for re-sistance to rice dwarf disease and its mechanism (VIII).Breeding of Aichi 42, resistant to rice dwarf disease andgreen leafhopper. Res. Bull. Aichi Agric. Res. Cent. 16:1–14 (in Japanese with English summary).
Sogawa, K. (1982) The rice brown planthopper: Feedingphysiology and host plant interactions. Annu. Rev. Ento-mol. 27: 49–73.
Sogawa, K. (1983a) Biotypes of phytophagous insects: in-traspecific variations in host affinity and infestivity (1).Plant Prot. 37: 7–10 (in Japanese).
Sogawa, K. (1983b) Biotypes of phytophagous insects: in-traspecific variations in host affinity and infestivity (2).Plant Prot. 37: 63–68 (in Japanese).
Sogawa, K. and A. Sato (1981) The green rice leafhopper,Nephotettix cincticeps Uhler (Hemiptera: Deltocephali-dae), populations with differential reactions to rice vari-eties. Jpn. J. Appl. Entomol. Zool. 25: 280–285 (inJapanese with English summary).
Sugimoto, A. (1969) Mass rearing systems of green riceleafhopper. Bull. Agric. Chem. Insp. Stn. 9: 19–24 (inJapanese).
Takeda, M. and T. Nagata (1987) Seasonal fluctuation of an-tibiosis to the green rice leafhopper, Nephotettix cincti-ceps, in the resistant parent lines of rice. Annu. Rep.Soc. Plant Prot. North Jpn. 38: 103–106 (in Japanese).
Takita, T. and H. Habibuddin (1985) Relationship betweenlaboratory-developed biotypes of green leafhopper andresistant varieties of rice in Malaysia. Jpn. Agric. Res.Q. 19: 219–223.
Takita, T. and H. Nishiyama (1989) Selection of biotypes ofgreen rice leafhopper and genetic analysis for the resis-tance in rice. Bull. Kyushu Agric. Exp. Stn. 25: 251–259.
Tamura, K., Y. Fukuta, M. Hirae, S. Oya, I. Ashikawa and T.Yagi (1997) Genetic analysis of the green rice leafhop-per (Nephotettix cincticeps Uhler) resistance IV. Localiza-tion of the chromosomal regions resistant to green riceleafhopper in Norin PL 5. Breed. Sci. 47 (suppl. 2): 167(in Japanese).
Tamura, K., Y. Fukuta, M. Hirae, S. Oya, I. Ashikawa and T.Yagi (1998) Genetic analysis of the green rice leafhop-per (Nephotettix cincticeps Uhler) resistance V. RFLPmapping of the resistance gene in Saikai 182. Breed.Sci. 48 (suppl. 2): 108 (in Japanese).
Tamura, K., Y. Fukuta, M. Hirae, S. Oya, I. Ashikawa and T.Yagi (1999) Mapping of the Grh 1 locus for green rice

leafhopper resistance in rice using RFLP markers.Breed. Sci. 49: 11–14.
Tanaka, K. (1999) Quantitative genetic analysis of biotypesof the brown planthopper Nilaparvata lugens: heritabilityof virulence to resistant rice varieties. Entomol. Exp.
Appl. 90: 279–287.Tokura, K., K. Yagasaki, M. Arakawa, S. Kato, T. Niwayama
and K. Otsuka (1995) A new rice variety “Sainoyume”with the resistance to green rice leafhopper. Breed. Sci.45 (suppl. 2): 278 (in Japanese).




















