
Altered neuronal nitric oxide synthase expression in the cerebellum of calcium channel mutant mice
12
Brain Research 977 (2003) 129–140 www.elsevier.com / locate / brainres Research report Altered neuronal nitric oxide synthase expression in the cerebellum of calcium channel mutant mice a b ,1 c ,1 a a Im Joo Rhyu , Sang-Soep Nahm , Seung Jun Hwang , Hyun Kim , Young-Suk Suh , d b b, * Sen-Ich Oda , Tamy C. Frank , Louise C. Abbott a Institute of Human Genetics and Department of Anatomy, Korea University College of Medicine, 126-1 Anam-Dong 5-Ga, Seongbuk-Ku, Seoul 136-705, South Korea b Department of Veterinary Anatomy and Public Health, College of Veterinary Medicine, Texas A&M University, 4458 TAMU, College Station, TX 77843-4458, USA c Department of Anatomy, University of Ulsan, College of Medicine, 388-1 PoongNap-Dong, Songpa-Ku, Seoul 138-736, South Korea d Laboratory of Animal Management, School of Agricultural Science, Nagoya University, Nagoya 464-8601, Japan Accepted 29 January 2003 Abstract Tottering, rolling Nagoya, and leaner mutant mice all exhibit cerebellar ataxia to varying degrees, from mild (tottering mice) to severe (leaner mice). Collectively, these mice are regarded as tottering locus mutants because each of these mutant mice expresses a different autosomal recessive mutation in the gene coding for the a calcium ion channel protein, which is the pore forming subunit for P/ Q-type 1A high voltage activated calcium ion channels. These mutant mice all exhibit varying degrees of cerebellar dysfunction and neuronal cell death. Nitric oxide (NO) is an important messenger molecule in the central nervous system, especially in the cerebellum, and it is produced via the enzyme, nitric oxide synthase (NOS). We investigated expression of neuronal-NOS (n-NOS) in the cerebella of all three mutant mice, as revealed by NADPH-diaphorase (NADPH-d) histochemical staining, quantitation of n-NOS protein using Western blotting and quantitation of n-NOS mRNA using in situ hybridization. The expression of n-NOS mRNA and protein as well as the NADPH-d histochemical reaction were elevated in tottering and rolling Nagoya cerebella. n-NOS mRNA and the NADPH-d histochemical reaction were decreased in the leaner cerebellum, but the leaner mouse n-NOS protein concentration was not significantly different compared to age- and gender-matched controls. These findings suggest that NO may act as an important mediator in the production of the neuropathology observed in these mutant mice. 2003 Elsevier Science B.V. All rights reserved. Theme: Motor systems and sensorimotor integration Topic: Cerebellum Keywords: Nitric oxide; Epilepsy; Ataxia; NADPH-d; In situ hybridization histochemistry; P/ Q-type calcium channels 1. Introduction system (CNS), including: synaptogenesis, modulation of neurotransmitter release, synaptic plasticity and cell death Nitric oxide (NO) is a cell-derived, diffusible, biological [19,20,27,28,44,58,63]. Nitric oxide synthase (NOS) cata- messenger molecule with multiple functions in the im- lyzes the conversion of L-arginine to NO and L-citrulline, mune, vascular and nervous systems [3,63]. NO is thought utilizing NADPH and O as substrates [12,40]. Cells 2 to be involved in various processes in the central nervous cannot sequester NO in order to regulate the amount of NO that is released. Instead, NO is synthesized on demand using NOS enzyme to produce the amount of NO needed *Corresponding author. Tel.: 11-979-845-2269; fax: 11-979-847- [13]. Therefore, the key to regulating NO activity resides 8981. at the level of controlling its synthesis through NOS [62]. E-mail address: [email protected] (L.C. Abbott). 1 NOS exists in three major isoforms: (1) neuronal NOS These two authors contributed equally as second author on this manu- script. (n-NOS); (2) endothelial NOS (e-NOS); and (3) inducible 0006-8993 / 03 / $ – see front matter 2003 Elsevier Science B.V. All rights reserved. doi:10.1016 / S0006-8993(03)02403-X
Transcript of Altered neuronal nitric oxide synthase expression in the cerebellum of calcium channel mutant mice

Brain Research 977 (2003) 129–140www.elsevier.com/ locate/brainres
Research report
A ltered neuronal nitric oxide synthase expression in the cerebellum ofcalcium channel mutant mice
a b ,1 c ,1 a aIm Joo Rhyu , Sang-Soep Nahm , Seung Jun Hwang , Hyun Kim , Young-Suk Suh ,d b b ,*Sen-Ich Oda , Tamy C. Frank , Louise C. Abbott
aInstitute of Human Genetics and Department of Anatomy, Korea University College of Medicine, 126-1 Anam-Dong 5-Ga, Seongbuk-Ku,Seoul 136-705, South Korea
bDepartment of Veterinary Anatomy and Public Health, College of Veterinary Medicine, Texas A& M University, 4458 TAMU, College Station,TX 77843-4458,USA
cDepartment of Anatomy, University of Ulsan, College of Medicine, 388-1 PoongNap-Dong, Songpa-Ku, Seoul 138-736, South KoreadLaboratory of Animal Management, School of Agricultural Science, Nagoya University, Nagoya 464-8601, Japan
Accepted 29 January 2003
Abstract
Tottering, rolling Nagoya, and leaner mutant mice all exhibit cerebellar ataxia to varying degrees, from mild (tottering mice) to severe(leaner mice). Collectively, these mice are regarded as tottering locus mutants because each of these mutant mice expresses a differentautosomal recessive mutation in the gene coding for thea calcium ion channel protein, which is the pore forming subunit for P/Q-type1A
high voltage activated calcium ion channels. These mutant mice all exhibit varying degrees of cerebellar dysfunction and neuronal celldeath. Nitric oxide (NO) is an important messenger molecule in the central nervous system, especially in the cerebellum, and it isproduced via the enzyme, nitric oxide synthase (NOS). We investigated expression of neuronal-NOS (n-NOS) in the cerebella of all threemutant mice, as revealed by NADPH-diaphorase (NADPH-d) histochemical staining, quantitation of n-NOS protein using Westernblotting and quantitation of n-NOS mRNA using in situ hybridization. The expression of n-NOS mRNA and protein as well as theNADPH-d histochemical reaction were elevated in tottering and rolling Nagoya cerebella. n-NOS mRNA and the NADPH-dhistochemical reaction were decreased in the leaner cerebellum, but the leaner mouse n-NOS protein concentration was not significantlydifferent compared to age- and gender-matched controls. These findings suggest that NO may act as an important mediator in theproduction of the neuropathology observed in these mutant mice. 2003 Elsevier Science B.V. All rights reserved.
Theme: Motor systems and sensorimotor integration
Topic: Cerebellum
Keywords: Nitric oxide; Epilepsy; Ataxia; NADPH-d; In situ hybridization histochemistry; P/Q-type calcium channels
1 . Introduction system (CNS), including: synaptogenesis, modulation ofneurotransmitter release, synaptic plasticity and cell death
Nitric oxide (NO) is a cell-derived, diffusible, biological [19,20,27,28,44,58,63].Nitric oxide synthase (NOS) cata-messenger molecule with multiple functions in the im- lyzes the conversion ofL-arginine to NO andL-citrulline,mune, vascular and nervous systems[3,63]. NO is thought utilizing NADPH and O as substrates[12,40]. Cells2
to be involved in various processes in the central nervous cannot sequester NO in order to regulate the amount of NOthat is released. Instead, NO is synthesized on demandusing NOS enzyme to produce the amount of NO needed
*Corresponding author. Tel.:11-979-845-2269; fax:11-979-847- [13]. Therefore, the key to regulating NO activity resides8981.
at the level of controlling its synthesis through NOS[62].E-mail address: [email protected](L.C. Abbott).1 NOS exists in three major isoforms: (1) neuronal NOSThese two authors contributed equally as second author on this manu-script. (n-NOS); (2) endothelial NOS (e-NOS); and (3) inducible
0006-8993/03/$ – see front matter 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S0006-8993(03)02403-X

130 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
NOS (i-NOS). Both n-NOS and e-NOS are considered to postnatal day 20[35] while leaner Purkinje cell deathbe constitutively expressed and dependent on calcium ions peaks at postnatal days 40–50[17]. It is known thatand calmodulin to function, while i-NOS is classified as neuronal intracellular calcium ion homeostasis is affectednon-constitutive (or inducible) and calcium-independent by these mutations in the gene coding for thea calcium1A
[2,8,48].n-NOS is the most important NOS isoform in the ion channel protein[15]. Thus, it is possible that changesCNS and it is highly expressed in the cerebellum in intracellular calcium homeostasis could affect NOS[20,28,47,58,63,65].NO itself is known to influence expression in these mutant mice.voltage-dependent calcium channel function[46]. While the full range of biological effects of NO pro-
It is likely that those neurons that normally express NOS duction continues to unfold, important roles of NO haveand that also have altered intracellular calcium homeosta- been implicated in the pathophysiology of different ner-sis, might have altered NOS expression. To investigate this vous system disorders such as, ischemic brain damagequestion we used tottering, rolling Nagoya and leaner [36,55], spinal cord trauma[67], Alzheimer’s disease[13]mutant mice, which all carry autosomal recessive muta- and Parkinson’s disease[21,51]. Inappropriate NO pro-tions in the gene coding for thea calcium ion channel duction also appears to play an important role in mediation1A
protein. Thea protein is the pore forming subunit for of some forms of neurotoxicity associated with excessive1A
P/Q-type voltage dependent calcium channels[16,42].The glutamate production[62]. On the other hand, NO maytottering mutation is a point mutation located in the also promote neuronal survival[6,7]. Finally, recent workextracellular region that links the IIS5 and IIS6 domains also points to a role of NO in refinement of experience-and results in a proline-to-leucine amino acid substitution dependent consolidation of cortical synaptic contacts[11][16]. The rolling Nagoya mutation also is a point mutation and long-term potentiation and depression[9].that is located in the voltage sensor-forming segment in Based on the possible roles of NO in synaptic circuitryrepeat IIIS4 and results in an arginine-to-glycine amino and adult brain function as well as neuronal cell death andacid substitution[42]. The leaner mutation occurs in a survival, and the fact that neuronal-NOS (n-NOS) expres-splice donor consensus sequence in the C-terminal region sion is regulated by calcium, it was intriguing to investi-and produces an out-of-frame splicing event[16,42]. P- gate whether alterations in functional capabilities of neuro-type calcium channels comprise greater than 90% of the nal P/Q-type calcium channels had any effect on nNOScalcium channels found on Purkinje cell somata[39]. expression in adult mice. To address this question, wePurkinje cell dendritic processes also highly express P-type examined n-NOS expression in the cerebellum of threechannels [66]. Calcium channel subtype expression in different P/Q-type calcium channel mutant mice. The threecerebellar granule cells is more diverse; P-type represent mutant genotypes included in this study, tottering, rolling11% and Q-type represent 35% of calcium channels[52], Nagoya and leaner mice, exhibit different degrees ofwith a higher P-type channel density (50%) reported for cerebellar dysfunction as well as different degrees ofgranule cell axon (parallel fiber) terminals in rat cerebellar postnatal cerebellar neuronal death. The goal of this studymolecular layer[39]. Taken together, mutations in thea was to gain insight into whether NO, as revealed by1A
calcium ion channel protein gene are likely to have very expression of n-NOS, shows different degrees of expres-profound effects on the function of cerebellar neurons in sion in the adult cerebellum of these three differentthese mutant mice. calcium ion channel mutant mice relative to the degree of
rolHomozygous tottering (tg), rolling Nagoya (tg ) and cerebellar dysfunction exhibited by these mutant mice. Welaleaner (tg ) mice express absence seizures, paroxysmal were interested in assessing correlations that might be
dyskinesia, cerebellar ataxia and postnatal cerebellar neu- present between n-NOS expression and the survivingronal cell death to varying degrees[22,37,53,60,68].The cerebellar neurons. We examined n-NOS mRNA andphenotypes for these mutant mice are discussed in detail protein expression and NADPH-diaphorase histochemistry,by Zwingman et al.[68]. In summary, the paroxysmal and we show that expression of n-NOS mRNA and proteindyskinesia is most obvious in tottering mice, while all as well as the NADPH-d histochemical reaction werethree mutants clearly exhibit ataxia and absence seizures, elevated in tottering and Nagoya rolling mouse cerebella.which both progress from mild in tottering mice, to more n-NOS mRNA and the NADPH-d histochemical reactionsevere in rolling Nagoya mice to most severe in leaner were decreased in the leaner mouse cerebellum, but themice. Tottering mice do not show any excessive cerebellar n-NOS protein concentration was not significantly differentneuronal cell death, at least up to 90 days of age[30], compared to age- and gender-matched controls.while modest cerebellar granule cell death is evident inrolling Nagoya mice[60]. In contrast, the leaner mousewill lose up to 50% of granule cells, Purkinje cells and 2 . Materials and methodsGolgi cells of the cerebellum[26]. Leaner mice firstdisplay excessive loss of cerebellar granule cells followed 2 .1. Animalsby loss of Purkinje cells during postnatal months one to six[24,26,35]. Peak leaner granule cell death occurs at Mice with the control genotype C57BL/6J:1 /1 (wild

I.J. Rhyu et al. / Brain Research 977 (2003) 129–140 131
type), or mutant genotypes, C57BL/6J:tg /tg (homozygous dated in chloroform for 5 min, rinsed in ethanol, thenla latottering) and C57BL/6J:tg /tg (homozygous leaner) air-dried. Dr. Tsutomu Ogura (Research Institute, East,
were housed in the Laboratory Animal Research and National Cancer Center, Japan) kindly provided the fullResource building at Texas A&M University. Male and cDNA plasmid of mouse n-NOS[45]. An antisense
lafemale heterozygous leaner (tg /1) and heterozygous riboprobe was constructed from the full cDNA and wastottering (tg /1) mice were bred to produce homozygous complementary to base pairs 1866–2406 of the mouse
la latg /tg and tg /tg offspring, respectively. Mice with the n-NOS sequence. The antisense probe was prepared usingcontrol genotype, PROD:1 /1 (wild type) or mutant a Riboprobe System kit (Promega, Madison, WI, USA) and
rol 35phenotypes PROD:tg /1 (heterozygous) or PROD: the incorporation of [ S]UTP (1000–1500 Ci /mmol,rol roltg /tg (homozygous rolling Nagoya), were bred at Amersham Biosciences, Piscataway, NJ, USA). A total of
6Nagoya University, then shipped to and maintained at 5310 cpm of labeled RNA probe was added to eachKorea University. Mice were housed and bred in a slide, the sections were coverslipped with a plastic film andconstant-temperature room (21–228C) with 12-h light / hybridization took place overnight at 538C in a humidifieddark cycle and allowed access to food and water ad chamber. After 16–20 h, the sections were washed in 23
libitum. All mice were weaned between 30 and 40 days of saline sodium citrate (SSC), treated with RNAase A (20la laage. Male and female homozygoustg /tg, tg /tg and mg/ml, Boehringer-Mannheim, Indianapolis, IN, USA) for
rol roltg /tg mice between the ages of 2.5–6 months were 30 min at 378C, then washed sequentially for 60 min inused in this study. Due to the expression of the ataxic 23 SSC at 508C, 60 min in 0.23 SSC at 558C, and 60phenotype, all adult homozygous mutant mice could easily min in 0.23 SSC at 608C. The slides were briefly rinsedbe distinguished from their heterozygous littermates, which in deionized water then 70% ethanol and allowed to dry.do not exhibit ataxia. After drying, the slides were exposed to Hyperfilm radiog-
Homozygous leaner pups often die of hypothermia and raphic film (Amersham Biosciences) using an exposure14dehydration at approximately 3 weeks of age due to their time of 7 days. A C standard (American Radiolabeled
severe ataxia, epileptiform seizures and paroxysmal Chemicals, St. Louis, MO, USA) was co-exposed to thedyskinesia, which limits their ability to move about the radiographic film along with the experimental tissuecage and adequately access food and water. Consequently,sections[34]. After film exposure, the slides were dippedhomozygous leaner mice used in this study were kept alive in NTB2 photographic emulsion (Eastman Kodak, Roches-into adulthood by fostering newborn homozygous leaner ter, NY, USA) diluted 1:1 with deionized water, dried forpups to lactating Swiss White Webster female mice. In 2–3 h in the dark and then stored at 48C. After 4–5 weeksaddition, moistened rodent chow was placed into the cagesof exposure, the emulsion-coated slides were developed,starting when leaner pups were approximately 2.5 weeksfixed, counterstained with cresyl violet, and observed withold and changed daily. Adult leaner mice were weaned at dark field microscopy using a Zeiss Axioscope.approximately 40 days of age, but their diet of dry rodent Autoradiographic images were captured with a Sonychow and water, given ad libitum, continued to be sup- XC-77 CCD camera using NIH image 1.61 softwareplemented with moistened rodent chow, which was (developed at the US National Institute of Health andchanged daily. All experimental procedures were carried available on the internet athttp: / / rsb.info.nih.gpv/nih-out in accordance with theNational Institutes of Health image/) and a Power Macintosh 7200 (Apple, Cupertino,Guide for the Care and Use of Laboratory Animals (NIH CA, USA). The gray values of autoradiographs werePublication No. 85-23, revised, 1996) and the minimum converted to nominal dpm (disintegrations/minute) with anumber of animals necessary for each experiment werethird-degree polynomial curve based on the co-exposed
14used. C-radiolabeled standard. The whole cerebellum wasdivided into anterior and posterior regions and the density
2 .2. In situ hybridization histochemistry (ISHH) of labeling was measured in each region. Five to sixanimals were used for each experimental group and the
ISHH was performed as described by Kim et al.[33] mean of three sections from each animal was used forwith minor modifications. Mice were killed by decapita- analysis. Care was taken to ensure that the sections weretion, the brains rapidly removed, then frozen with pow- taken from equivalent horizontal levels from each animaldered dry ice. Horizontal tissue sections (12mm) of the analyzed. The data were analyzed using a one-way analysiscerebella were cut on a cryostat, mounted onto gelatin- of variance (ANOVA), followed by the Student–Newman–coated slides and stored at270 8C until used. When ISHH Keuls post-hoc test.was carried out, the sections were allowed to warm toroom temperature, fixed in 4% formaldehyde, rinsed andtreated with 0.25% acetic anhydride in 0.1 M 2 .3. NADPH-diaphorase (NADPH-d) histochemistrytriethanolamine–0.9% NaCl (pH 8.0) for 10 min to reducenonspecific hybridization of the probe. The sections were Nicotinamide adenine dinucleotide phosphate (NADPH)-dehydrated in increasing concentrations of ethanol, delipi- diaphorase histochemistry is used as a marker for NOS

132 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
distribution in the brain since it is a component of the NO ame, CA, USA) in diluted in 2% skim milk /TBST. Thesynthesis pathway[23]. Mice were anesthetized and per- membrane was again washed five times in TBST. n-NOSfused as described. The cerebella were removed, postfixed immunopositive bands were visualized using enhancedin 4% paraformaldehyde over night, and cryoprotected by chemiluminescence (ECL) substrate (Amersham Biosci-immersion in 30% sucrose solution in phosphate buffer ences). The membrane was subsequently exposed to Hy-(PB) at 48C. Serial sections were processed in one of two perfilm ECL� (Amersham Biosciences), and developedways. In the first method 30-mm sections were cut in a using an automatic autoprocessor. The density of immuno-cryostat and collected in cold PB. The sections were positive bands was calculated, in arbitrary units, to com-treated as floating sections with 0.1 M Tris buffer (pH 8.0) pare the relative intensity of protein bands as describedcontaining 0.3% Triton X-100 for 30 min and transferred previously by Brager et al.[1]. Optical densities of n-NOSto a solution of 0.1 M Tris buffer (pH 8.0) containing 1 immunoreactive bands from mutant mice were normalizedmM b-nicotinamide adenine dinucleotide phosphate (b- by the average optical density of appropriate wild-typeNADPH) (Sigma–Aldrich, St. Louis, MO. USA), 0.5 mM controls. Statistical significance of the normalized opticalnitro blue tetrazolium (NBT, Sigma–Aldrich) and 0.3% densities was determined by using the Mann–Whitney testTriton X-100. After pre-incubation at room temperature for and Prism 3 software (GraphPad, San Diego, CA, USA).5 min with continuous shaking, the sections were incu-bated at 378C for 60–90 min. The reaction was stopped bythe addition of excess 0.1 M Tris buffer (pH 8.0), the 3 . Resultssections were mounted onto gelatin-coated slides andprocessed for dehydration and coverslipped. In the second Autoradiographic labeling of cerebellar tissue sectionsmethod, 25-mm sections were processed using the same indicated the presence of n-NOS mRNA in cerebella fromprotocol for NADPH-d staining but the sections were cut all mutant and wild-type mice examined. Under dark-fieldusing a cryostat mounted on to gelatin-coated slides and microscopic observation, dense accumulations of silverthen stained on slide at room temperature without shaking. grains were present over both the cerebellar granule cellBoth methods worked equally well to carry out the and molecular layers (Fig. 1A). In the molecular layer,NADPH-d histochemical reaction. silver grains primarily accumulated over the somata of
basket cells and satellite cells (bright field image,Fig. 1B).2 .4. Western blotting Basket cell somata were intensely labeled while satellite
cells typically were only moderately labeled, which isMice were killed by decapitation, the brains rapidly consistent with previous reports concerning the rat cere-
removed and immediately frozen with powdered dry ice, bellum[31,54]. Silver grains located over cerebellarthen stored at270 8C until used. At the time of analysis, granule cells formed patch-like clusters of more dense andthe cerebella were thawed individually and diced on a less dense accumulations (Fig. 1A), which also is inchilled glass plate. Total protein from each cerebellum was agreement with previous reports of molecular compart-extracted by sonication in 0.5 ml of tissue lysis buffer (10 mentalization of NOS expression with in the mousemM Tris, 1% SDS and 10% volume of a proteinase cerebellar granule cell layer[57]. Silver grain accumula-inhibitor cocktail (Roche, Mannheim, Germany)). Extracts tion over Purkinje cell somata was not different fromwere boiled (5 min), centrifuged at 12 0003g (15 min) background levels, indicating that no detectable n-NOSand the supernatant was collected into a new tube. Protein mRNA was present in Purkinje cells in any of the miceconcentration was determined using Bradford reagent examined.(BioRad, Hercules, CA)[1]. All extracts were stored at Densitometric analysis of in situ hybridization histoch-220 8C. Thirty mg of each protein extract were mixed with emistry (ISHH) autoradiographic films revealed a signifi-23 loading buffer, boiled at 958C (5 min), cooled, then cant increase in expression of n-NOS mRNA in theloaded onto an 8% SDS–polyacrylamide gel. After sepa- cerebella of both tottering and rolling mice (n55, P,
ration the proteins were transferred onto PVDF membrane 0.01), while n-NOS message expression was decreased inusing a Mini V8-10 system (Life Technologies, Gaithers- leaner mouse cerebella (n55, P,0.05) (Figs. 2 and 3).burg, MD, USA). After transfer, the gel was stained with This increase expression of n-NOS mRNA was observedCoomassie blue stain to determine the efficiency of protein in the granule cells and basket cells of the cerebellum.transfer. The membrane was blocked 2 h at room tempera- Although the total density of signal in the leaner cere-ture with 5% skim milk in Tris-buffered saline plus 0.1% bellum was lower than that of wild-type mice, the posteriorTween-20 (TBST). The membrane was then incubated lobe of the leaner cerebellum showed greater n-NOSovernight at 48C in 0.25mg/ml n-NOS antibody (Trans- mRNA expression than in the anterior lobe (Fig. 2C). Noduction Laboratories, Lexington, KY, USA) diluted in 2% significant differences in anterior versus posterior cere-skim milk /TBST. The membrane was washed five times in bellum signal intensity were observed in either the totter-TBST then incubated at room temperature for 2 h in 0.06 ing mouse or the rolling Nagoya mouse (n55, P.0.05).mg/ml peroxidase-labeled anti-rabbit IgG (Vector, Burling- Similar to previous reports[23,57,59] the NADPH-d

I.J. Rhyu et al. / Brain Research 977 (2003) 129–140 133
Fig. 1. Darkfield (A) and brightfield (B) photomicrographs of sections of the rolling Nagoya mouse cerebellum illustrating expression of nNOS mRNAusing in situ hybridization histochemistry. Clusters of silver grains in the molecular layer (Mol) indicate the location of basket and stellate interneurons.Clusters of silver grains in the granule cell layer (Gr) indicate the location of cerebellar granule cell somata. Note the lack of silver grains localized overPurkinje cell somata in the cerebellar Purkinje cell layer (Pur). Magnifications: (A)3250; (B) 3400.
histochemical reaction was observed in both the cerebellar stained parallel fibers, which also were present in thegranule cell and molecular layers, but not in Purkinje cells molecular layer. In general, cerebellar granule cell somataof either the mutant or wild-type mice (Fig. 4). Basket were labeled strongly, but numerous patches of more andcells and satellite cells in the molecular layer of all mice less intensely stained granule cell somata were observed,examined were moderately stained, revealing a ‘granular’ which also is consistent with previous reports[10,23,57].staining appearance found in primarily in the nuclei, which The patch-like spatial distribution pattern of NADPH-dalso was observed by Cork et al.[10] in neurons in the staining observed in the granule cells of all three differentmouse cerebellum. Staining of interneurons in the cerebel- mutant mice was similar to the pattern observed in wild-lar molecular layer tended to be obscured by positively type mice (Fig. 4C–E). More intense NADPH-d staining
Fig. 2. Photomicrographs of autoradiographic films of frontal sections of wild type (A,D), tottering (B), leaner (C) and rolling Nagoya mouse (E)cerebellum illustrating expression of n-NOS mRNA using in situ hybridization histochemistry. Note the increased intensity of labeling in the totteringcerebellum (B) and the decreased intensity of labeling in the leaner cerebellum (C) compared to the wild-type cerebellum (A). The rolling Nagoya mousecerebellum also shows increased intensity of labeling (E) when compared to the wild-type control mouse (D) of the same genotype. Magnification for allphotomicrographs,34.

134 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
Fig. 3. Average level of nNOS mRNA in control (wild type) and mutant mouse cerebella (tottering, leaner and rolling Nagoya). The amount of mRNApresent is indicated using relative DPM and the wild-type levels are used for baseline comparison with the mutant mice. nNOS mRNA expression wassignificantly higher in the tottering and rolling mouse cerebellum (indicated by the *) when compared to their respective wild-type controls. nNOS mRNAexpression was significantly lower in the leaner mouse cerebellum (indicated by the *) when compared to the C57 wild-type control.n55 for all fivegroups of mice. The error bars indicate standard error of the mean.
of granule cells was observed in cerebella of tottering and 4 . Discussionrolling Nagoya mice when compared to wild-type mice(Fig. 4). And in general, cerebella of leaner mice showed We observed significant increases in expression levels ofmuch less intense NADPH-d staining when compared to n-NOS mRNA and protein in cerebella from both totteringwild-type mice (Fig. 4E,C). These differences in NADPH- and rolling Nagoya mice, when compared to age-matchedd staining intensity of the mutant mouse cerebella corres- wild-type mouse cerebella. In addition, the increased n-ponded to the differences in levels of n-NOS mRNA NOS mRNA expression was accompanied by increasedexpression observed for these mice. histochemical staining for NADPH-d in the cerebellar
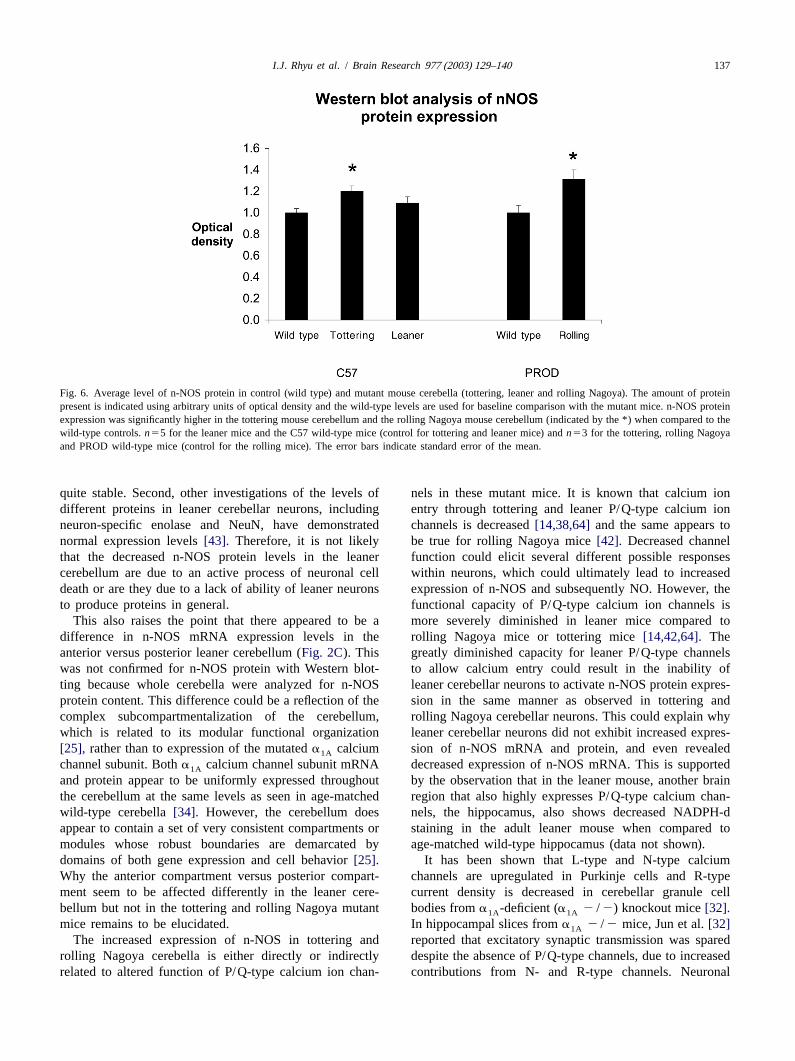
We utilized Western blotting to quantitate the amount of granule cell and molecular layers of tottering and rollingn-NOS protein present in whole cerebella (Fig. 5A). n- Nagoya mutant mice. Conversely, the leaner mouse cere-NOS protein expression was determined based on the bellum revealed only a modest decrease in n-NOS mRNAoptical density of n-NOS-positive bands on the film. It was expression compared to control mice, which was accom-difficult to find an appropriate control ‘housekeeping’ panied by a relatively large decrease in staining forprotein, such as actin, to use for comparison between the NADPH-d. However, the level of n-NOS protein expres-mutant mice and control mice because many proteins sion was not significantly decreased in the leaner cere-utilize cytosolic calcium ion concentration to regulate bellum when compared to age-matched wild-type mouseexpression, and calcium homeostasis is altered in these cerebella. In general, NADPH-d staining correlated wellmutant mice [15]. Therefore, we used Coomassie blue with the observations for in situ hybridization histoch-staining to ensure that exactly the same amount of protein emistry of n-NOS mRNA for all three mutant mice whenper sample was added to each lane of the gel (Fig. 5B). compared to the appropriate wild-type control mice. Corre-The optical densities of n-NOS-immunopositive bands lation between the intensity of neuronal NADPH-d histo-were calculated based on the density of the entire corre- chemical staining and the level of NOS expression hassponding lane on the Coomassie blue stained gel. We also been observed in numerous studies. In particular, de-used Coomassie blue staining of each gel after protein creased expression of NOS protein can be correlated withtransfer to the membranes, to determine that there were no decreased NADPH-d histochemistry staining under severaldifferences in transfer quality in the different lanes of each different conditions[49,63]. The patch-like spatial dis-gel. We then compared the relative concentration of n-NOS tribution pattern of NADPH-d staining observed in theprotein in the lanes from mutant mouse cerebella with the granule cell layer of all mice examined was very similar toappropriate control mouse cerebella. The n-NOS protein the report by Hawkes and Turner[23]. Patches were seenexpression was significantly increased in tottering and in all lobules. Examination of the NADPH-d stainingrolling Nagoya cerebella (n53, P,0.05), but was not pattern in these mutant mice did not demonstrate observ-changed in the leaner mouse cerebella (n55) (Fig. 6). able differences in patterning between mutant mice and

I.J. Rhyu et al. / Brain Research 977 (2003) 129–140 135
Fig. 4. Photomicrographs of NADPH-d histochemical staining of sagittal sections of wild-type (A,C), rolling mouse (B) tottering (D) and leaner (E)cerebellum. Note the lack of staining of the Purkinje somata in all sections. The granule cell layer and the molecular layers both are positively stained in allsections, though the intensity of staining that is observed varies with the genotype. The NADPH-d staining is more intense in the rolling mouse (B) andthetottering mouse (D) cerebellum than in the respective control cerebella (A,C). The leaner mouse cerebellum (E) expresses less intense NADPH-d stainingwhen compared to the control mouse (C). Magnifications: (A,B)3160; (C–E)3500.
wild-type mice, only differences in overall staining inten- meostasis also is known to be altered in leaner neuronssities. Hawkes and Turner[23] suggest that NADPH-d [15]. Differences in calcium homeostasis could affectstaining of granule cells, which is a reflection of n-NOS mRNA transcription rates or mRNA degradation rates inexpression, is related to the well-documented Purkinje cell leaner neurons, either of which could result in more proteincompartments[24]. expression per mRNA transcript in the leaner cerebellum
Increased n-NOS mRNA expression and NADPH-d when compared to neurons from tottering or rollingexpression both have been observed in situations in which Nagoya mice. Alternatively, n-NOS protein could be morethe CNS was lesioned or had undergone stress associated stable in leaner neurons, which also would effectivelywith injury such as trauma or toxic insult[4,5,7,59].While result in more protein expression per mRNA transcript inthe specific reason for the difference between n-NOS leaner mice. It is possible that decreased n-NOS mRNAmRNA expression and protein expression in the leaner expression levels in the leaner cerebellum could be due tocerebellum is not known, it is possible that the disparity decreased numbers of granule cells resulting from thebetween mRNA levels and protein levels is due to differ- granule cell loss that occurs primarily during the firstences in post-transcriptional and/or post-translational postnatal month of life in these mutant mice. However, thismodifications of proteins[61]. Intracellular calcium ho- is not likely since we recently reported that the density per

136 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
Fig. 5. Western blot (A) comparing amount of n-NOS protein in whole cerebella of control (C57 and PROD) and mutant mice (tottering (tg), leaner (la)and rolling Nagoya (rol)). Coomassie blue staining for protein (B) indicates that equal amounts of protein were loaded into each of the lanes. Note in (A)that the n-NOS bands from the rolling mouse cerebellum and the tottering mouse cerebellum are darker than the corresponding bands from the respectivecontrol cerebella (C57 and PROD).
unit area for granule cells is not different in the leaner is altered due to the fact that the neurons are in the processcerebellum when compared to age-matched wild-type of dying. However, two facts argue against this possibility.cerebella[43]. All mRNA quantification was carried out First, the majority of cerebellar neuronal cell loss hason a per unit area basis in this study, so mere loss of already taken place in leaner mice at the time these studiesgranule cells in the leaner cerebellum cannot explain the were conducted[17,26,35]. Leaner mice actually live adecreased n-NOS message present in these mice. longer than normal life span and their ataxia does not get
It also must be considered that the viability of leaner progressively worse (L. Abbott; personal observation), soneurons might be compromised and that protein expression the surviving neuronal population that is being studied is
I.J. Rhyu et al. / Brain Research 977 (2003) 129–140 137
Fig. 6. Average level of n-NOS protein in control (wild type) and mutant mouse cerebella (tottering, leaner and rolling Nagoya). The amount of proteinpresent is indicated using arbitrary units of optical density and the wild-type levels are used for baseline comparison with the mutant mice. n-NOS proteinexpression was significantly higher in the tottering mouse cerebellum and the rolling Nagoya mouse cerebellum (indicated by the *) when compared to thewild-type controls.n55 for the leaner mice and the C57 wild-type mice (control for tottering and leaner mice) andn53 for the tottering, rolling Nagoyaand PROD wild-type mice (control for the rolling mice). The error bars indicate standard error of the mean.
quite stable. Second, other investigations of the levels of nels in these mutant mice. It is known that calcium iondifferent proteins in leaner cerebellar neurons, including entry through tottering and leaner P/Q-type calcium ionneuron-specific enolase and NeuN, have demonstrated channels is decreased[14,38,64]and the same appears tonormal expression levels[43]. Therefore, it is not likely be true for rolling Nagoya mice[42]. Decreased channelthat the decreased n-NOS protein levels in the leaner function could elicit several different possible responsescerebellum are due to an active process of neuronal cell within neurons, which could ultimately lead to increaseddeath or are they due to a lack of ability of leaner neurons expression of n-NOS and subsequently NO. However, theto produce proteins in general. functional capacity of P/Q-type calcium ion channels is
This also raises the point that there appeared to be a more severely diminished in leaner mice compared todifference in n-NOS mRNA expression levels in the rolling Nagoya mice or tottering mice[14,42,64]. Theanterior versus posterior leaner cerebellum (Fig. 2C). This greatly diminished capacity for leaner P/Q-type channelswas not confirmed for n-NOS protein with Western blot- to allow calcium entry could result in the inability ofting because whole cerebella were analyzed for n-NOS leaner cerebellar neurons to activate n-NOS protein expres-protein content. This difference could be a reflection of the sion in the same manner as observed in tottering andcomplex subcompartmentalization of the cerebellum, rolling Nagoya cerebellar neurons. This could explain whywhich is related to its modular functional organization leaner cerebellar neurons did not exhibit increased expres-[25], rather than to expression of the mutateda calcium sion of n-NOS mRNA and protein, and even revealed1A
channel subunit. Botha calcium channel subunit mRNA decreased expression of n-NOS mRNA. This is supported1A
and protein appear to be uniformly expressed throughout by the observation that in the leaner mouse, another brainthe cerebellum at the same levels as seen in age-matched region that also highly expresses P/Q-type calcium chan-wild-type cerebella[34]. However, the cerebellum does nels, the hippocamus, also shows decreased NADPH-dappear to contain a set of very consistent compartments or staining in the adult leaner mouse when compared tomodules whose robust boundaries are demarcated by age-matched wild-type hippocamus (data not shown).domains of both gene expression and cell behavior[25]. It has been shown that L-type and N-type calciumWhy the anterior compartment versus posterior compart- channels are upregulated in Purkinje cells and R-typement seem to be affected differently in the leaner cere- current density is decreased in cerebellar granule cellbellum but not in the tottering and rolling Nagoya mutant bodies froma -deficient (a 2 /2) knockout mice[32].1A 1A
mice remains to be elucidated. In hippocampal slices froma 2 /2 mice, Jun et al.[32]1A
The increased expression of n-NOS in tottering and reported that excitatory synaptic transmission was sparedrolling Nagoya cerebella is either directly or indirectly despite the absence of P/Q-type channels, due to increasedrelated to altered function of P/Q-type calcium ion chan- contributions from N- and R-type channels. Neuronal

138 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
activity controls n-NOS expression through influx of possible that leaner mice cannot compensate for deficits incalcium ions[56] primarily through L-type voltage-gated neuronal function resulting from their severely decreasedcalcium channels[61]. It is largely unknown whether the P/Q-type channel function, and thus, cannot increase NOSdifferences in channel function associated with complete production and subsequent NO levels. This is consistentloss of P/Q-type channel function, as seen ina 2 /2 with a possible trophic role for NO in the mouse cere-1A
mice, are present with the partial loss of P/Q-type channel bellum that might explain the lack of Purkinje cell loss infunction in the three spontaneous mutant mice that are the tottering and rolling Nagoya mice, which overexpress NOSsubject of this report. It has been suggested that L-type in satellite and basket cells.calcium channels are upregulated in tottering mouse Alternatively, it has been shown that exposure of thecerebellar Purkinje cells[18], but it is not known how developing rat cerebellum toN-nitro-L-arginine methyles-granule cells or cerebellar interneurons respond with ter (L-NAME), a NOS inhibitor, does not appear to resultrespect to expression of non-P/Q-type channels in any of in significant alterations in cerebellar morphology[63]. Inthese spontaneous mutant mice. Clearly, neuronal re- fact, NOS inhibition in some instances has been observedsponses to partial or complete loss of P/Q-type neuronal to partially protect cerebellar granule cells from excitotoxiccalcium channels are complex and the signals leading to damage[6]. Such different observations can be due toincreased n-NOS expression in tottering and rolling several factors. One consideration is that the use of non-Nagoya mice still need to be elucidated. selective or partially selective NOS inhibitors result in a
Evidence for both a protective role and a toxic role for wide range of side effects that can obscure the precise roleNO has been reported in the literature. If NOS levels, and of n-NOS-derived NO in the CNS[41]. For example, thesubsequent NO levels, are elevated, this is thought to result use ofL-NAME affects both n-NOS and e-NOS andin the death of some populations of neurons[29], but the reports of increased, decreased or unaltered neuronalspecific mechanisms that bring about cell death are not effects have been reported following focal ischemic dam-understood. Alternatively, moderate elevations in NOS and age in experimental animal pretreated withL-NAME [41].thus NO, may actually function in a protective manner in Given the interest in uncovering the functions of NOsome neuronal systems. NO has been reported to partici- produced by n-NOS in the CNS, the differences in n-NOSpate in neuronal differentiation in conjunction with nerve expression observed in these neurological mutant mice aregrowth factor[50]. Additional evidence for a trophic role intriguing and these mutant mice offer an important modelfor NO in the cerebellum has been provided by Wang et al. system in which to explore the possible trophic role of NO
G[65]. They reported that in vivoN -nitro-L-arginine inhibi- in the mammalian cerebellum.tion of NOS activity in the developing rat cerebellumcaused disturbances in normal cerebellar cortical layerformation. Wang et al.[65] postulated that NO may A cknowledgementsfunction as a trophic factor in either a paracrine orautocrine manner during postnatal cerebellar development. Supported in part by BK21 Project support to I.J. RhyuA similar explanation could be put forth for the observed and H. Kim and CERH NIEHS grant, P30-EF09106, todifferences in n-NOS expression in the cerebellum of these L.C. Abbott.three calcium channel mutant mice. The decreased func-tion of P/Q type calcium ion channels leads to alteredneuronal intracellular calcium homeostasis[15], which
R eferencescould result in reduced viability of cerebellar neurons thathighly express P/Q-type calcium channels. What the
[1] D .H. Brager, M. Sickel, M. McCarthy, Developmental sex differ-specific downstream consequences of decreased calciumences in calbindin-D and calretinin immunoreactivity in the28Kion entry actually are in neurons of these mutant mice areneonatal rat hypothalamus, J. Neurobiol. 42 (3) (2000) 315–322.
not yet known, but reduced calcium-dependent production [2] D .S. Bredt, S.H. Snyder, Nitric oxide mediates glutamate-linkedand/or processing of trophic factors is one possibility. It is enhancement of cGMP levels in the cerebellum, Proc. Natl. Acad.quite likely that neuronal function in tottering and rolling Sci. USA 87 (1990) 682–685.
[3] D .S. Bredt, S.H. Snyder, Nitric oxide, a physiological messengerNagoya mice is not as severely compromised as in leanermolecule, Annu. Rev. Biochem. 63 (1994) 175–195.mice. As a result, tottering and rolling Nagoya neurons
[4] M . Chalimoniuk, J. Strosznajder, NMDA receptor-dependent nitricmight be able to compensate for trophic deficits that result oxide and cGMP synthesis in brain hemispheres and cerebellumfrom the abnormal function of P- and Q-type calcium ion during reperfusion after transient forebrain ischemia in gerbils:channels. These two mutant mice exhibit little (rolling effect of 7-Nitroindazole, J. Neurosci. Res. 54 (5) (1998) 681–690.
[5] S . Chen, G. Aston-Jones, Cerebellar injury induces NADPHNagoya) to no (tottering) excessive death of cerebellardiaphorase in Purkinje and inferior olivary neurons in the rat, Exp.neurons, perhaps through a mechanism that causes in-Neurol. 126 (2) (1994) 270–276.
creased n-NOS production. However, the leaner mouse [6] E . Ciani, L. Groneng, M. Voltattorni, V. Rolseth, A. Contestabile,first displays excessive loss of cerebellar granule cells R.E. Paulsen, Inhibition of free radical production or free radicalfollowed by loss of leaner Purkinje cells[24,26]. It is scavenging protects from the excitotoxic cell death mediated by

I.J. Rhyu et al. / Brain Research 977 (2003) 129–140 139
glutamate in cultures of cerebellar granule neurons, Brain Res. 728 [29] Z . Huang, P.L. Huang, N. Panahian, T. Dalkara, M.C. Fishman,(1) (1996) 1–6. M.A. Moskowitz, Effects of cerebral ischemia in mice deficient in
neuronal nitric oxide synthase, Science 265 (5180) (1994) 1883–[7] E . Ciani, M. Virgili, A. Contestabile, Akt pathway mediates a1885.cGMP-dependent survival role of nitric oxide in cerebellar granule
neurones, J. Neurochem. 81 (2002) 218–228. [30] K .R. Isaacs, L.C. Abbott, Development of the paramedian lobule ofthe cerebellum in wild type and tottering mice, Dev. Neurosci. 14[8] H .J. Cho, Q.W. Xie, J. Calaycay, R.A. Mumford, K.M. Swiderek,(1992) 386–393.T.D. Lee, C. Nathan, Calmodulin is a subunit of nitric oxide
synthase from macrophages, J. Exp. Med. 176 (2) (1992) 599–604. [31] K . Iwase, M. Takemura, T. Shimada, S. Wakisaka, T. Nokubi, Y.Shigenaga, Ontogeny of NADPH-diaphorase in rat forebrain and[9] A . Contestabile, Roles of NMDA receptor activity and nitric oxidemidbrain, Anat. Embryol. 197 (3) (1998) 229–247.production in brain development, Brain Res. Rev. 32 (2000) 476–
509. [32] J . Jun, E.S. Piedras-Renteria, S.M. Smith, D.B. Wheeler, S.B. Lee,T.G. Lee, H. Chin, M.E. Adams, R.H. Scheller, R.W. Tsien, H.-S.[10] R .J. Cork, M.L. Perrone, J. Wandell, C.A. Scheiner, R.R. Mize, A
21Shin, Ablation of P/Q-type Ca channel currents, altered synapticweb-accessible digital atlas of the distribution of nitric oxidetransmission, and progressive ataxia in mice lacking thea -synthase (http: / /nadph.anatomy.lsumc.edu/default.htm), Prog. Brain 1A
subunit, Proc. Natl. Acad. Sci. USA 96 (26) (1999) 15245–15250.Res. 118 (1998) 37–50.[33] H . Kim, J.P. Ko, U.G. Kang, J.B. Park, H.L. Kim, Y.H. Lee, Y.S.[11] K .S. Cramer, A. Angelucci, J.-O. Hahm, M.B. Bogdanov, M. Sur, A
Kim, Electroconvulsive shock reduces inositol 1,4,5-trisphosphaterole for nitric oxide in the development of the ferret retinogeniculate3-kinase mRNA expression in rat dentate gyrus, J. Neurochem. 63projection, J. Neurosci. 16 (1996) 7995–8004.(5) (1994) 1991–1994.[12] T .M. Dawson, D.S. Bredt, S.H. Synder, A novel neuronal messenger
[34] F . Lau, L.C. Abbott, I.J. Rhyu, D. Kim, H. Chin, Expression ofmolecule in brain: the free radical, nitric oxide, Ann. Neurol. 32calcium channela mRNA and protein in the leaner mouse(1992) 297–311. 1A
la la(tg /tg ) cerebellum, Mol. Brain Res. 59 (1998) 93–99.[13] V .L. Dawson, T.M. Dawson, Nitric oxide neurotoxicity, J, Chem.[35] F .C. Lau, Apoptosis, reduced intracellular free calcium level andNeuroanat. 10 (1996) 179–190.
altered gene expression in the cerebellum of the leaner mutant[14] L .S. Dove, L.C. Abbott, W.H. Griffith, Whole-cell and single-mouse, Ph.D. Dissertation, Texas A&M University, College Station,channel analysis of P-type calcium currents in cerebellar PurkinjeTexas, 1999.cells of leaner mutant mice, J. Neurosci. 18 (19) (1998) 7687–7699.
[36] R .R. Leker, A. Teichner, H. Ovadia, E. Keshet, E. Reinherz, T.[15] L .S. Dove, S.S. Nahm, D. Murchison, L.C. Abbott, W.H. Griffith,Ben-Hur, Expression of endothelial nitric oxide synthase in theAltered calcium homeostasis in cerebellar Purkinje cells of leanerischemic penumbra: relationship to expression of neuronal nitricmutant mice, J. Neurophysiol. 84 (1) (2000) 513–524.oxide synthase and vascular endothelial growth factor, Brain Res.[16] C .F. Fletcher, C.M. Lutz, T.N. O’Sullivan, J.D. Shaughnessy Jr., R.909 (1-2) (2001) 1–7.Hawkes, W.N. Frankel, N.G. Copeland, N.A. Jenkins, Absence
epilepsy in tottering mutant mice is associated with calcium channel [37] P . Levitt, J.L. Noebels, Mutant mouse tottering: selective increase ofdefects, Cell 87 (4) (1996) 607–617. locus coeruleus axons in a defined single-locus mutation, Proc. Natl.
[17] T .C. Frank, M.C. Nunley, H.D. Sons, L.C. Abbott, Fluoro-Jade Acad. Sci. USA 78 (1981) 4630–4634.identification of the cerebellar Purkinje cell death pattern in the [38] N .M. Lorenzon, C.M. Lutz, W.N. Frankel, K.G. Beam, Alteredleaner mouse, FASEB 16 (Abstract Program) (2002) 529.6. calcium channel currents in Purkinje cells of the neurological mutant
[18] B .E. Fureman, D.B. Campbell, E.J. Hess, L-type calcium channel mouse leaner, J. Neurosci. 18 (12) (1998) 4482–4489.regulation of abnormal tyrosine hydroxylase expression in cerebella [39] I .M. Mintz, B.L. Sabatini, W.G. Regehr, Calcium control of trans-of tottering mice, Ann. NY Acad. Sci. 868 (1999) 217–219. mitter release at a cerebellar synapse, Neuron 15 (1995) 675–688.
[19] J .A. Gally, R.P. Montague, G.N. Reeke, G.E. Edelman, The NO [40] S . Moncada, A. Higgs, TheL-arginine-nitric oxide pathway, Newhypothesis: possible effects of a short-lived, rapidly diffusible signal Engl. J. Med. 27 (1993) 2002–2012.in the development and function of the nervous system, Proc. Natl. [41] P .K. Moore, R.L. Handy, Selective inhibitors of neuronal nitricAcad. Sci. USA 87 (1990) 3547–3551. oxide synthase: is no NOS really good NOS for the nervous
[20] J . Garthwaite, C.L. Boulton, Nitric oxide signaling in the central system?, Trends Pharmacol. Sci. 18 (6) (1997) 204–211.nervous system, Annu. Rev. Physiol. 57 (1995) 683–706. [42] Y . Mori, M. Wakamori, S. Oda, C.F. Fletcher, N. Sekiguchi, E. Mori,
[21] E .M. Gatto, N.A. Riobo, M.C. Carreras, A. Chernavsky, A. Rubio, N.G. Copeland, N.A. Jenkins, K. Matsushita, Z. Matsuyama, K.21M.L. Satz, J.J. Poderoso, Overexpression of neurtrophil neuronal Imoto, Reduced voltage sensitivity of activation of P/Q-type Ca
nitric oxide synthatase in Parkinson’s disease, Nitric Oxide: Biol. channels is associated with the ataxic mouse mutation rollingrolChem. 4 (5) (2000) 534–539. Nagoya (tg ), J. Neurosci. 20 (15) (2000) 5654–5662.
[22] M .C. Green, R.L. Sidman, Tottering, a neuromuscular mutation in [43] S . Nahm, D. Tomlinson, L.C. Abbott, Decreased calretinin expres-the mouse and its linkage with oligosyndactylism, J. Hered. 53 sion in cerebellar granule cells in the leaner mouse, J. Neurobiol. 51(1962) 233–237. (2002) 313–322.
[23] R . Hawkes, R.W. Turner, Compartmentation of NADPH-diaphorase [44] Y . Noda, K. Yamada, Y. Komori, H. Sugihara, H. Furukawa, T.activity in the mouse cerebellar cortex, J. Comp. Neurol. 346 (4) Nabeshima, Role of nitric oxide in the development of tolerance and(1994) 499–516. sensitization to behavioural effects of phencyclidine in mice, Br. J.
[24] J .A. Heckroth, L.C. Abbott, Purkinje cell loss from alternating Pharmacol. 117 (7) (1996) 1579–1585.sagittal zones in the cerebellum of leaner mutant mice, Brain Res. [45] T . Ogura, T. Yokoyama, H. Fujisawa, Y. Kurashima, H. Esumi,658 (1-2) (1994) 93–104. Structural diversity of neuronal nitric oxide synthase mRNA in the
[25] K . Herrup, B. Kuemerele, The compartmentalization of the cere- nervous system, Biochem. Biophys. Res. Commun. 193 (3) (1993)bellum, Annu. Rev. Neurosci. 20 (1997) 61–90. 1014–1022.
[26] K . Herrup, S.L. Wilczynski, Cerebellar cell degeneration in the [46] S . Ohkuma, M. Katsura, Y. Hibino, J. Xu, K. Shirotani, K.leaner mutant mouse, Neuroscience 7 (9) (1982) 2185–2196. Kuriyama, Multiple actions of nitric oxide on voltage-dependent
21[27] T . Himi, M. Ikeda, I. Sato, T. Yuasa, S. Murota, Purkinje cells Ca channels in mouse cerebral cortical neurons, Mol. Brain Res.express neuronal nitric oxide synthase after methylmercury adminis- 54 (1998) 133–140.tration, Brain Res. 718 (1-2) (1996) 189–192. [47] J . Ohyu, S. Takashima, Developmental characteristics of neuronal
[28] C . Holscher, Nitric oxide, the enigmatic neuronal messenger: its role nitric oxide synthase (n-NOS) immunoreactive neurons in fetal toin synaptic plasticity, Trends Neurosci. 20 (1997) 298–303. adolescent human brains, Dev. Brain Res. 110 (1998) 193–202.

140 I.J. Rhyu et al. / Brain Research 977 (2003) 129–140
[48] D . Okada, Differential effects of protein kinase C on neuronal nitric suggests a role for mossy fibers in their development, Neuroscienceoxide synthase activity in rat cerebellar slices and in vitro, J. Chem. 59 (4) (1994) 893–903.Neuroanat. 10 (1996) 213–220. [58] E .M. Schuman, D.V. Madison, Nitric oxide and synaptic function,
[49] R . Paes de Carvalho, M.H. de Faria, J.L.M. Nascimento, J.N. Annu. Rev. Neurosci. 17 (1994) 153–183.Hokoc, Development of NADPH-diaphorase in the avian retina: [59] Y . Shigeyoshi, T. Ogura, H. Esumi, K. Chihara, Y. Ibata, H.regulation by calcium ions and relation to nitric oxide synthase, J. Okamura, Lesion-induced neuronal nitric oxide synthase in PurkinjeNeurochem. 67 (3) (1996) 1063–1071. cells of the rat cerebellar cortex: histochemical and in situ hybridiza-
[50] W . Poluha, C.M. Schonhoff, K.S. Harrington, M.B. Lachyankar, tion study, Brain Res. Mol. Brain Res. 44 (2) (1997) 229–237.N.E. Crosbie, D.A. Bulseco, A.H. Ross, A novel, nerve growth [60] Y .S. Suh, S. Oda, Y.H. Kang, H. Kim, I.J. Rhyu, Apoptotic cell deathfactor-activated pathway involving nitric oxide, p53, and p21WAF1 of cerebellar granule cells in rolling mouse Nagoya, Neurosci. Lett.regulates neuronal differentiation of PC12 cells, J. Biol. Chem. 272 325 (1) (2002) 1–4.(38) (1997) 24002–24007. [61] C .M. Thompson, A.E. Bernhard, H.W. Strobel, Barbiturate-induced
[51] S . Przedborski, V. Jackson-Lewis, R. Yokoyama, T. Shibata, V.L. expression of neuronal nitric oxide synthase in the rat cerebellum,Dawson, T.T. Dawson, Role of neuronal nitric oxide synthase in Brain Res. 754 (1-2) (1997) 142–146.l-methyl-phenyl-1,2,3,6-tetrahydrophyridine (MPTP)-induced dopa- [62] M . Trenago, M. Virgili, B. Monti, T. Guarnieri, A. Contestabile,minergic neurotoxicity, Proc. Natl. Acad. Sci. USA 93 (1996) Alteration of neuronal nitric oxide synthase activity and expression4565–4571. in the cerebellum and the forebrain of microencephalic rats, Brain
[52] A . Randall, R.W. Tsien, Pharmacological dissection of multiple Res. 793 (1998) 54–60.21types of Ca channel currents in rat cerebellar granule neurons, J. [63] M . Virgili, B. Monti, A. LoRusso, M. Bentivogli, A. Contestabile,
Neurosci. 15 (1995) 2995–3012. Developmental effects of in vivo and in vitro inhibition of nitric[53] I .J. Rhyu, L.C. Abbott, D.B. Walker, C. Sotelo, An ultrastructural oxide synthase in neurons, Brain Res. 839 (1999) 164–172.
study of granule cell /Purkinje cell synapses in tottering (tg /tg), [64] M . Wakamori, K. Yamazaki, H. Matsunodaira, T. Teramoto, I.la laleaner (tg /tg ) and compound heterozygous, tottering/ leaner (tg / Tanaka, T. Niidome, K. Sawada, Y. Nishizawa, N. Sekiguchi, E.
latg ) mice, Neuroscience 90 (3) (1999) 717–728. Mori, Y. Mori, K. Imoto, Single tottering mutations responsible for[54] J . Rodrigo, D.R. Springall, O. Uttenthal, M.L. Bentura, F. Abadia- the neuropathic phenotype of the P-type calcium channel, J. Biol.
Molina, V. Riveros-Moreno, R. Martinez-Murillo, J.M. Polak, S. Chem. 273 (52) (1998) 34857–34867.Moncada, Localization of nitric oxide synthase in the adult rat brain, [65] W . Wang, T. Nakayama, N. Inoue, T. Kato, Quantitative analysis ofPhil. Trans. R. Soc. London Ser. B: Biol. Sci. 345 (1312) (1994) nitric oxide synthase expressed in developing and differentiating rat175–221. cerebellum, Dev. Brain Res. 111 (1998) 65–75.
[55] J . Rodrigo, D. Alonso, A.P. Fernandez, J. Serrano, A. Richart, J.C. [66] R .E. Westenbroek, T. Sakurai, E.M. Elliott, J.W. Hell, T.V.B. Starr,Lopez, M. Santacana, R. Martinez-Murillo, S. Moncada, M.L. T.P. Snutch, W.A. Catterall, Immunochemical identification andBentura, M. Ghiglione, L.O. Uttenthal, Neuronal and inducible nitric subcellular distribution of thea subunits of brain calcium1A
oxide synthase expression and protein nitration in rat cerebellum channels, J. Neurosci. 15 (1995) 6403–6418.after oxygen and glucose deprivation, Brain Res. 909 (2001) 20–45. [67] W . Wu, F.J. Liuzzi, F.P. Schinco, A.S. Depto, Y. Li, J.A. Mong, T.M.
[56] M . Sasaki, M. Gonzalez-Zulueta, W.J. Hering, S. Ahn, D.D. Ginty, Dawson, S.H. Snyder, Neuronal nitric oxide synthase is induced inV.L. Dawson, T.M. Dawson, Dynamic regulation of neuronal NO spinal neurons by traumatic injury, Neuroscience 61 (1994) 719–synthase transcription by calcium influx through a CREB family 726.transcription factor-dependent mechanism, Proc. Natl. Acad. Sci. [68] T .A. Zwingman, P.E. Neumann, J.L. Noebels, K. Herrup, Rocker isUSA 97 (15) (2000) 8617–8622. a new variant of the voltage-dependent calcium channel gene
[57] K . Schilling, H.H.H.W. Schmidt, S.L. Baader, Nitric oxide synthase Cacna1a, J. Neurosci. 21 (4) (2001) 1169–1178.expression reveals compartments of cerebellar granule cells and




















