
On the equivalence of upward and downward inheritance reasoners

Available online at www.sciencedirect.com
Environmental and Experimental Botany 63 (2008) 207–215
Adaptive longitudinal growth of first-order lateral roots of a woodyspecies (Spartium junceum) to slope and different soil
conditions—upward growth of surface roots
Antonino Di Iorio a,∗, Bruno Lasserre b, Livio Petrozzi b,Gabriella S. Scippa b, Donato Chiatante a
a Dipartimento di Scienze Chimiche e Ambientali, Universita dell’Insubria, Via Valleggio 11, 22100 Como, Italyb Dipartimento di Scienze e Tecnologie per l’Ambiente ed il Territorio, Universita del Molise,
C.da Fonte Lappone, 86090 Pesche(IS), Italy
Received 26 February 2007; received in revised form 4 October 2007; accepted 8 October 2007
Abstract
First-order lateral roots originating in the upper part of the taproot of a woody species, usually termed surface roots, grow close beneath the soilsurface, even on irregular or sloping ground. In slope condition, in fact, the surface roots can assume upward as well as downward growth. Existingknowledge on the controls over root direction does not fully explain these field observations.
Two different soil types and sloping conditions were selected in field condition to explore the behaviour of the surface roots in the woody speciesSpartium junceum L. The root system 3D architecture was measured with a 3D digitizer and the angle of growth (0◦ = vertically downwards) andthe radial direction (0◦ = horizontally downslope or northwards) of all root segments measured.
Surface roots were more numerous in clay soil than in loam soil, independently from the slope inclination. They had initial angles larger than90◦, i.e. they grew upwards only in clay soil. The subsequent angles of growth maintained this value only in steep-slope condition, showing a clearsoil type x slope inclination interaction. The initial angle of all first-order lateral roots decreased linearly with depth of origin on the taproot alwaysin relation to the soil type, with this relationship being stronger in clay soil.
These findings showed that the liminal angle (the preferred angle of growth) of surface roots was mainly affected by the soil type rather than thesoil surface inclination. Thus, upward growth must stand in the plasticity of the plagiotropic response of these secondary laterals rather than in astrong internal control.© 2007 Elsevier B.V. All rights reserved.
Keywords: Plagiogravitropism; Secondary roots; Slope condition; Spartium junceum; Three-dimensional (3-D) coarse root architecture; Upward growth
1. Introduction
First-order lateral roots of woody species, shrubs and trees,develop in acropetal sequence from the positively gravitropicseedling taproot. Some of the first-order laterals originating inthe upper part of the taproot extend for several metres withinabout 20 cm of the ground surface (references in Fayle, 1975).According to Coutts and Nicoll (1991), they are here termedthe “surface roots”. They are the first ones to explore the upper,nutrient-rich layers of the soil and because of their rapid rate ofsecondary thickening, they become the major structural mem-
∗ Corresponding author. Tel.: +39 031 326213; fax: +39 031 326230.E-mail address: [email protected] (A. Di Iorio).
bers of the root system for support and anchorage (see Coutts,1987). Moreover, all first-order laterals form the main frame-work from which rise lateral roots of higher order. Some ofthese higher order roots growing beneath the soil surface havea similar growth rate and morphology to the parent roots, andmay also become surface roots.
First-order lateral roots are typically plagiogravitropic andgraviresponsive (Rufelt, 1965) and in slope condition thereis evidence of negative gravitropic growth in those growingupslope (Marler and Discekici, 1997; Di Iorio et al., 2005). Pla-giogravitropism is a widespread tropic behaviour of axial plantorgans growing longitudinally in a direction different from theplumb line (Rufelt, 1965). In fact, the first-order lateral roots typ-ically attain a preferred, or liminal, angle of growth that deviatesfrom the vertical. This angle tends to remain even after grow-
0098-8472/$ – see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.envexpbot.2007.10.022

208 A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215
ing around an obstacle (Wilson, 1967; Nakamoto, 1994). Otherfeatures of the soil environment such as moisture, aeration, tem-perature and mechanical impedance have also been found toinfluence the angle of growth of first-order laterals in a varietyof woody plants (see Coutts, 1989). However, few observationsrefer specifically to the surface roots of trees and shrubs (Couttsand Nicoll, 1991).
Nevertheless, existing knowledge on the controls over thelongitudinal growth direction of first-order laterals (Nakamoto,1994; Nakamoto and Oyanagi, 1994; Leopold and Wettlaufer,1988) does not fully explain field observation on the behaviourof the surface roots (Rufelt, 1965; Coutts, 1989). For example,such roots have some ability to retain their distance from thesoil surface, to track beneath it on uneven ground. Moreover, inslope condition, experimental studies (Coutts and Nicoll, 1991;Marler and Discekici, 1997; Shrestha et al., 2000; Chiatanteet al., 2003a,b; Di Iorio et al., 2005) and one modelling study(Tsutsumi et al., 2003, 2004) show that surface roots growupward as well as downward.
The slope is a complex environmental condition that sub-jects plants to several mechanical stresses, such as the turningmoment induced by the combination of the inclination and theweights of the stem and the soil (Di Iorio et al., 2005). More-over, soil nutrients, water and well aerated pores are distributedasymmetrically around the orthotropic taproot. Therefore, a rootapex growing on the upslope side of the plant with a diagrav-itropic direction (perpendicular to gravity) meets different soilconditions and increases its distance from the soil surface dur-ing its elongation. Thus, the slope represents the most suitablecondition to address the control of the plagiotropic growth andthe distance from the soil surface.
A tendency for upward growth is considered apparently inher-ent in some of the uppermost first-order laterals and that if soilcondition (nutrients, moisture, compaction) are favourable, suchinternal tendency is manifested up to the root tip approaches thesoil surface. As consequence, on sloping ground, it was expectedthat these roots could exploit the surface soil layer on the upslopeand down-slope sides of the plant (Coutts and Nicoll, 1991).
We hypothesized that the soil condition plays a key role inthe upward growth of the uppermost first-order laterals (surfaceroots) and that in slope condition, this growth response resultsfrom the interaction between the soil type and the slope factorsrather than from a specific response only to the slope condition.Considerations on the specific soil traits involved are discussed.
The way to test this hypothesis was to analyse the angle ofgrowth of the first-order laterals at increasing distances from thetaproot junction in two different soil types and two differentlyinclined slope conditions, with particular attention to the sur-face roots. Spanish broom (Spartium junceum), a pioneer shrubspecies naturally growing on hilly slope of the Mediterraneanbasin and frequently used for stabilizing the urban embankmentand motorway slopes, was an adequate plant for this study.Although we studied samples growing in natural field con-ditions, the selected study sites shared similar environmentalparameters (climate, temperature, precipitation, slope exposure,inclination, altitude), with the soil type as the only variablefactor.
2. Materials and methods
2.1. Field site
Two adjacent study sites located in loamy and clayey soilswith similar topographic (altitude, exposition) and climatic(rainfall, temperature) conditions were selected in the Trignoriver basin near Trivento (Molise, Italy), on the east side of thecentral Italian Apennine. The sites themselves are located on aNNW facing slope (640 and 620 m a.s.l., respectively, latitude41◦42′N, longitude 14◦32′E, mean slope of 40◦). The climate ismontane Mediterranean with dry summers and cool, wet win-ters. The average yearly rainfall is 701 mm, falling mostly asrain. The mean summer and winter temperatures are estimatedto be 22.5 and 7.1 ◦C, respectively. On both sites, two differentlyinclined slope conditions were selected: a steep-slope condition(40◦–45◦) and an adjacent shallow-slope condition (4◦–5◦), thelatter considered as control.
2.2. Selection and extraction of plant samples
In order to collect comparable samples, spanish broom plants(S. junceum L.) with the root-collar diameter averaged between 9and 12 mm, were selected. Twenty samples were collected fromeach soil type for a total of 40 plants. Within each soil type,half were located in shallow and the other half in steep slopeconditions. Only samples that had no neighbours in a radius ofabout 1 m were excavated. For all the considered samples, beforeexcavation, the references to the magnetic north and south, andthe horizontal plane were detected and marked at the stem basesurface with a permanent pen. The latter reference was detectedwith two small levels perpendicularly positioned on a calliper.Only for plants growing in slope, the down-slope and upslopedirections were recorded and always marked at the stem-basesurface. Each plant was then carefully dug out by hand to anextension of 0.4 m in radial distance and to 0.4 m depth, and to aroot diameter less than 1 mm. During excavations, three dimen-sional photos and drawings of the root systems were also madein situ to preserve the original position of the spanish broomroots after excavation. Such information was indeed useful forthe subsequent digitation phase (Fig. 1a).
2.3. Root system architecture measurements and analysis
The X, Y, Z position coordinates of roots together with thediameter and the topology (i.e. the branching hierarchic struc-ture) were measured using a 3D digitiser with a Long Rangertransmitter (3 SPACE Fastrak, Polhemus Inc., Colchester, VT)(Sinoquet and Rivet, 1997). The 3D spatial distribution of theroot system had been carefully restored within a large box (50 cmper side) with the vertical walls made of a transparent plasticsheet drilled at 5 cm intervals (Fig. 1a). Such holes allowed theuser to easily fix the orientation of a root axis by mean of thinstrings. Most of the first-order laterals preserved their orienta-tion within a maximum estimated distance of 20 cm from thetaproot junction, due to the rigidity of Spartium root wood. Asa consequence, their orientation was easy to restore. The ori-
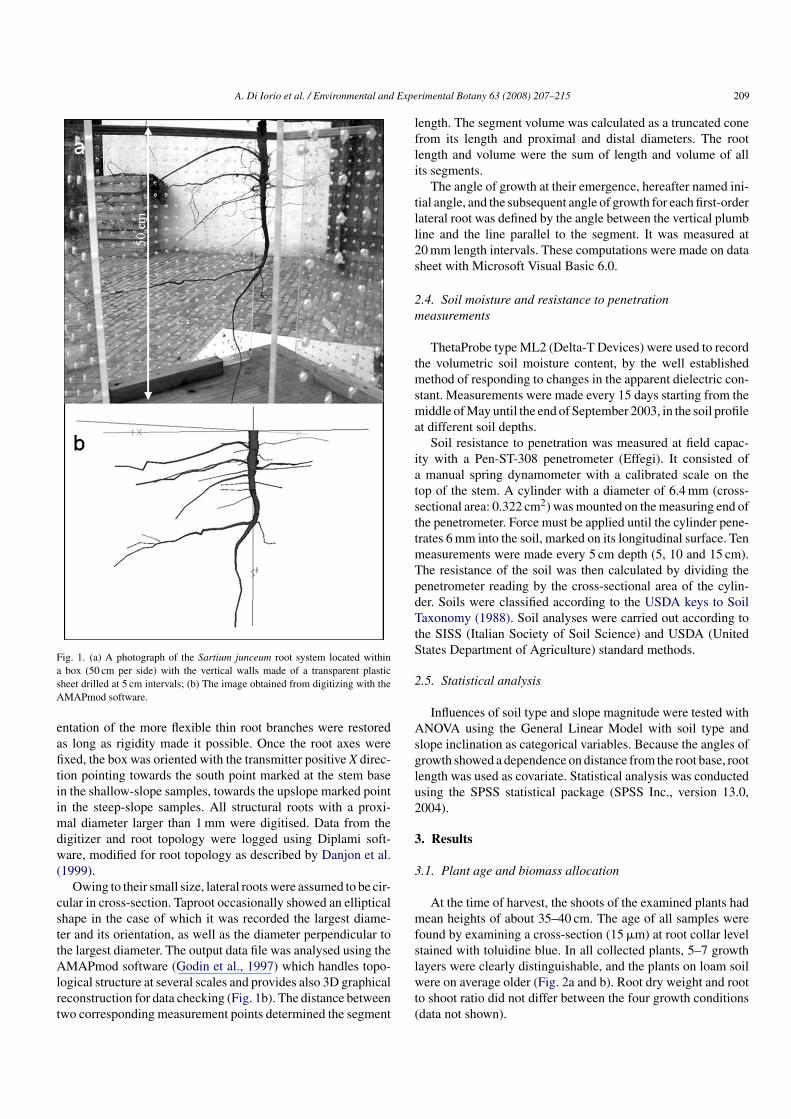
A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215 209
Fig. 1. (a) A photograph of the Sartium junceum root system located withina box (50 cm per side) with the vertical walls made of a transparent plasticsheet drilled at 5 cm intervals; (b) The image obtained from digitizing with theAMAPmod software.
entation of the more flexible thin root branches were restoredas long as rigidity made it possible. Once the root axes werefixed, the box was oriented with the transmitter positive X direc-tion pointing towards the south point marked at the stem basein the shallow-slope samples, towards the upslope marked pointin the steep-slope samples. All structural roots with a proxi-mal diameter larger than 1 mm were digitised. Data from thedigitizer and root topology were logged using Diplami soft-ware, modified for root topology as described by Danjon et al.(1999).
Owing to their small size, lateral roots were assumed to be cir-cular in cross-section. Taproot occasionally showed an ellipticalshape in the case of which it was recorded the largest diame-ter and its orientation, as well as the diameter perpendicular tothe largest diameter. The output data file was analysed using theAMAPmod software (Godin et al., 1997) which handles topo-logical structure at several scales and provides also 3D graphicalreconstruction for data checking (Fig. 1b). The distance betweentwo corresponding measurement points determined the segment
length. The segment volume was calculated as a truncated conefrom its length and proximal and distal diameters. The rootlength and volume were the sum of length and volume of allits segments.
The angle of growth at their emergence, hereafter named ini-tial angle, and the subsequent angle of growth for each first-orderlateral root was defined by the angle between the vertical plumbline and the line parallel to the segment. It was measured at20 mm length intervals. These computations were made on datasheet with Microsoft Visual Basic 6.0.
2.4. Soil moisture and resistance to penetrationmeasurements
ThetaProbe type ML2 (Delta-T Devices) were used to recordthe volumetric soil moisture content, by the well establishedmethod of responding to changes in the apparent dielectric con-stant. Measurements were made every 15 days starting from themiddle of May until the end of September 2003, in the soil profileat different soil depths.
Soil resistance to penetration was measured at field capac-ity with a Pen-ST-308 penetrometer (Effegi). It consisted ofa manual spring dynamometer with a calibrated scale on thetop of the stem. A cylinder with a diameter of 6.4 mm (cross-sectional area: 0.322 cm2) was mounted on the measuring end ofthe penetrometer. Force must be applied until the cylinder pene-trates 6 mm into the soil, marked on its longitudinal surface. Tenmeasurements were made every 5 cm depth (5, 10 and 15 cm).The resistance of the soil was then calculated by dividing thepenetrometer reading by the cross-sectional area of the cylin-der. Soils were classified according to the USDA keys to SoilTaxonomy (1988). Soil analyses were carried out according tothe SISS (Italian Society of Soil Science) and USDA (UnitedStates Department of Agriculture) standard methods.
2.5. Statistical analysis
Influences of soil type and slope magnitude were tested withANOVA using the General Linear Model with soil type andslope inclination as categorical variables. Because the angles ofgrowth showed a dependence on distance from the root base, rootlength was used as covariate. Statistical analysis was conductedusing the SPSS statistical package (SPSS Inc., version 13.0,2004).
3. Results
3.1. Plant age and biomass allocation
At the time of harvest, the shoots of the examined plants hadmean heights of about 35–40 cm. The age of all samples werefound by examining a cross-section (15 �m) at root collar levelstained with toluidine blue. In all collected plants, 5–7 growthlayers were clearly distinguishable, and the plants on loam soilwere on average older (Fig. 2a and b). Root dry weight and rootto shoot ratio did not differ between the four growth conditions(data not shown).
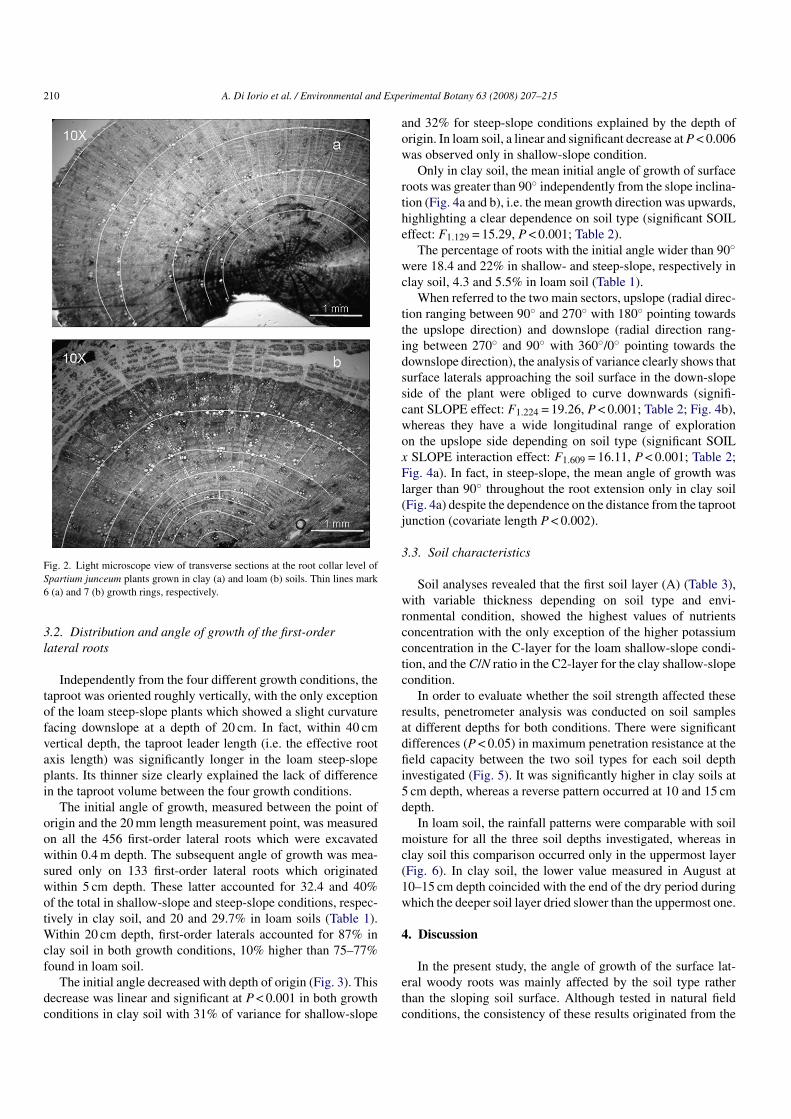
210 A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215
Fig. 2. Light microscope view of transverse sections at the root collar level ofSpartium junceum plants grown in clay (a) and loam (b) soils. Thin lines mark6 (a) and 7 (b) growth rings, respectively.
3.2. Distribution and angle of growth of the first-orderlateral roots
Independently from the four different growth conditions, thetaproot was oriented roughly vertically, with the only exceptionof the loam steep-slope plants which showed a slight curvaturefacing downslope at a depth of 20 cm. In fact, within 40 cmvertical depth, the taproot leader length (i.e. the effective rootaxis length) was significantly longer in the loam steep-slopeplants. Its thinner size clearly explained the lack of differencein the taproot volume between the four growth conditions.
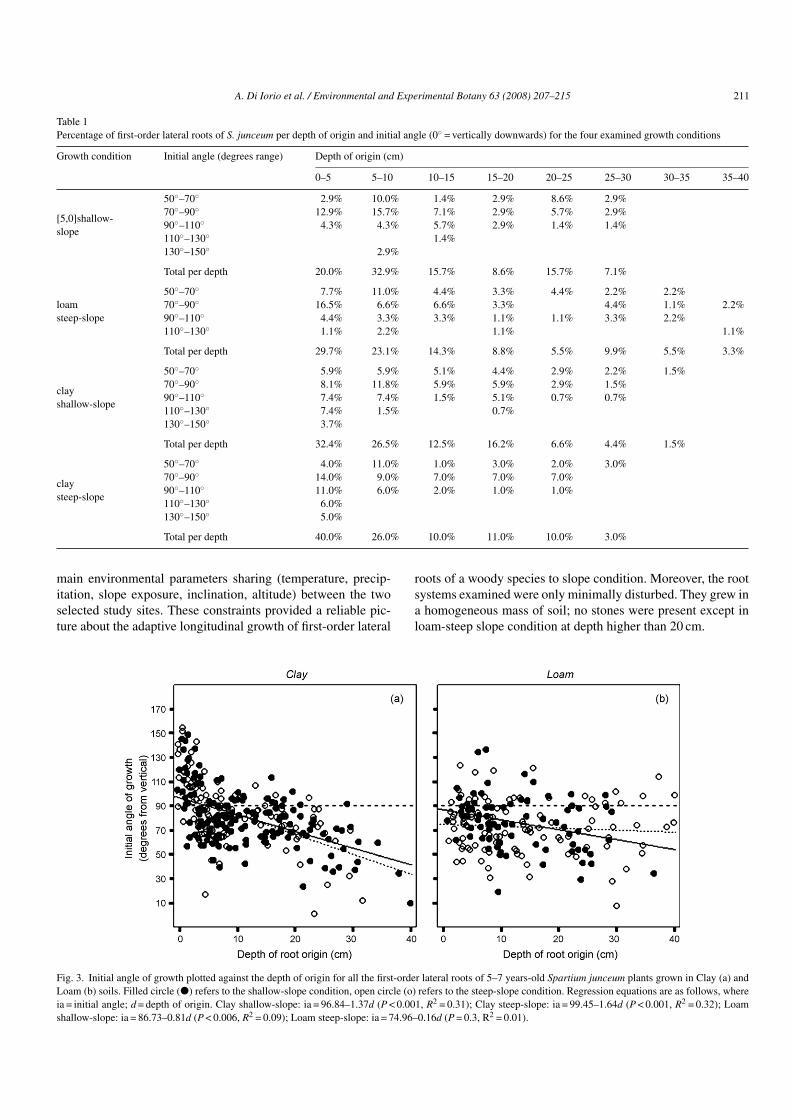
The initial angle of growth, measured between the point oforigin and the 20 mm length measurement point, was measuredon all the 456 first-order lateral roots which were excavatedwithin 0.4 m depth. The subsequent angle of growth was mea-sured only on 133 first-order lateral roots which originatedwithin 5 cm depth. These latter accounted for 32.4 and 40%of the total in shallow-slope and steep-slope conditions, respec-tively in clay soil, and 20 and 29.7% in loam soils (Table 1).Within 20 cm depth, first-order laterals accounted for 87% inclay soil in both growth conditions, 10% higher than 75–77%found in loam soil.
The initial angle decreased with depth of origin (Fig. 3). Thisdecrease was linear and significant at P < 0.001 in both growthconditions in clay soil with 31% of variance for shallow-slope
and 32% for steep-slope conditions explained by the depth oforigin. In loam soil, a linear and significant decrease at P < 0.006was observed only in shallow-slope condition.
Only in clay soil, the mean initial angle of growth of surfaceroots was greater than 90◦ independently from the slope inclina-tion (Fig. 4a and b), i.e. the mean growth direction was upwards,highlighting a clear dependence on soil type (significant SOILeffect: F1.129 = 15.29, P < 0.001; Table 2).
The percentage of roots with the initial angle wider than 90◦were 18.4 and 22% in shallow- and steep-slope, respectively inclay soil, 4.3 and 5.5% in loam soil (Table 1).
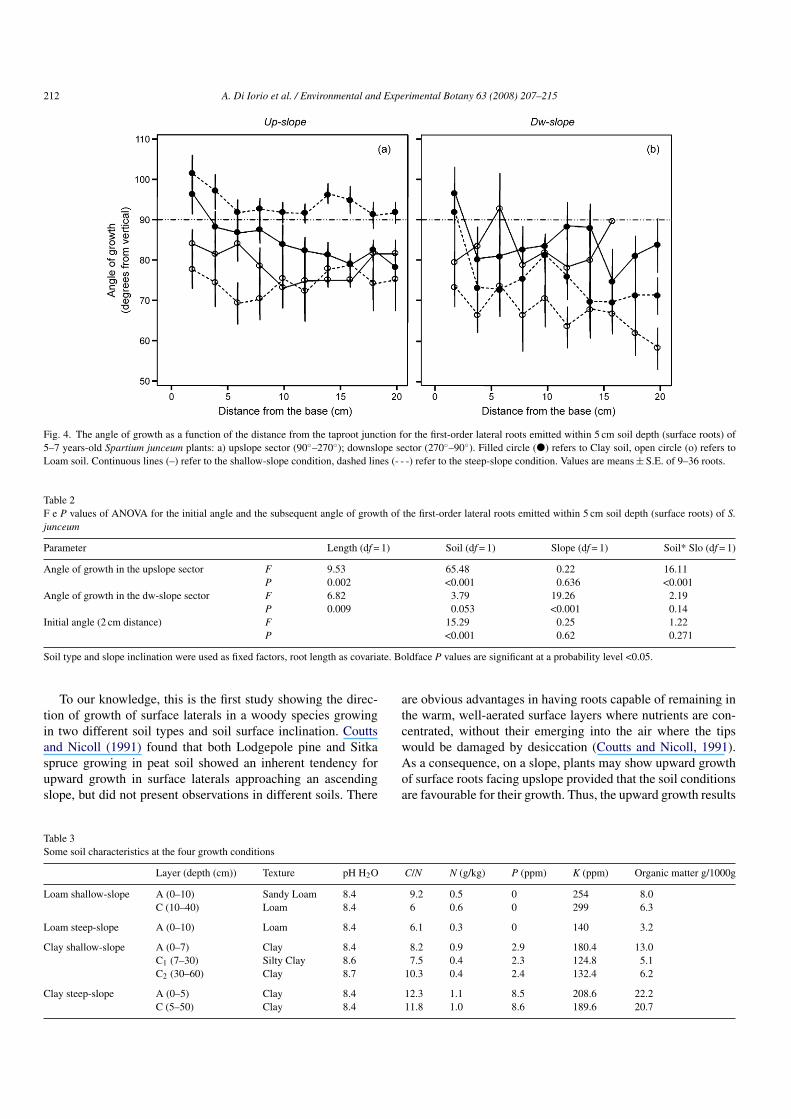
When referred to the two main sectors, upslope (radial direc-tion ranging between 90◦ and 270◦ with 180◦ pointing towardsthe upslope direction) and downslope (radial direction rang-ing between 270◦ and 90◦ with 360◦/0◦ pointing towards thedownslope direction), the analysis of variance clearly shows thatsurface laterals approaching the soil surface in the down-slopeside of the plant were obliged to curve downwards (signifi-cant SLOPE effect: F1.224 = 19.26, P < 0.001; Table 2; Fig. 4b),whereas they have a wide longitudinal range of explorationon the upslope side depending on soil type (significant SOILx SLOPE interaction effect: F1.609 = 16.11, P < 0.001; Table 2;Fig. 4a). In fact, in steep-slope, the mean angle of growth waslarger than 90◦ throughout the root extension only in clay soil(Fig. 4a) despite the dependence on the distance from the taprootjunction (covariate length P < 0.002).
3.3. Soil characteristics
Soil analyses revealed that the first soil layer (A) (Table 3),with variable thickness depending on soil type and envi-ronmental condition, showed the highest values of nutrientsconcentration with the only exception of the higher potassiumconcentration in the C-layer for the loam shallow-slope condi-tion, and the C/N ratio in the C2-layer for the clay shallow-slopecondition.
In order to evaluate whether the soil strength affected theseresults, penetrometer analysis was conducted on soil samplesat different depths for both conditions. There were significantdifferences (P < 0.05) in maximum penetration resistance at thefield capacity between the two soil types for each soil depthinvestigated (Fig. 5). It was significantly higher in clay soils at5 cm depth, whereas a reverse pattern occurred at 10 and 15 cmdepth.
In loam soil, the rainfall patterns were comparable with soilmoisture for all the three soil depths investigated, whereas inclay soil this comparison occurred only in the uppermost layer(Fig. 6). In clay soil, the lower value measured in August at10–15 cm depth coincided with the end of the dry period duringwhich the deeper soil layer dried slower than the uppermost one.
4. Discussion
In the present study, the angle of growth of the surface lat-eral woody roots was mainly affected by the soil type ratherthan the sloping soil surface. Although tested in natural fieldconditions, the consistency of these results originated from the
A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215 211
Table 1Percentage of first-order lateral roots of S. junceum per depth of origin and initial angle (0◦ = vertically downwards) for the four examined growth conditions
Growth condition Initial angle (degrees range) Depth of origin (cm)
0–5 5–10 10–15 15–20 20–25 25–30 30–35 35–40
[5,0]shallow-slope
50◦–70◦ 2.9% 10.0% 1.4% 2.9% 8.6% 2.9%70◦–90◦ 12.9% 15.7% 7.1% 2.9% 5.7% 2.9%90◦–110◦ 4.3% 4.3% 5.7% 2.9% 1.4% 1.4%110◦–130◦ 1.4%130◦–150◦ 2.9%
Total per depth 20.0% 32.9% 15.7% 8.6% 15.7% 7.1%
loamsteep-slope
50◦–70◦ 7.7% 11.0% 4.4% 3.3% 4.4% 2.2% 2.2%70◦–90◦ 16.5% 6.6% 6.6% 3.3% 4.4% 1.1% 2.2%90◦–110◦ 4.4% 3.3% 3.3% 1.1% 1.1% 3.3% 2.2%110◦–130◦ 1.1% 2.2% 1.1% 1.1%
Total per depth 29.7% 23.1% 14.3% 8.8% 5.5% 9.9% 5.5% 3.3%
clayshallow-slope
50◦–70◦ 5.9% 5.9% 5.1% 4.4% 2.9% 2.2% 1.5%70◦–90◦ 8.1% 11.8% 5.9% 5.9% 2.9% 1.5%90◦–110◦ 7.4% 7.4% 1.5% 5.1% 0.7% 0.7%110◦–130◦ 7.4% 1.5% 0.7%130◦–150◦ 3.7%
Total per depth 32.4% 26.5% 12.5% 16.2% 6.6% 4.4% 1.5%
claysteep-slope
50◦–70◦ 4.0% 11.0% 1.0% 3.0% 2.0% 3.0%70◦–90◦ 14.0% 9.0% 7.0% 7.0% 7.0%90◦–110◦ 11.0% 6.0% 2.0% 1.0% 1.0%110◦–130◦ 6.0%130◦–150◦ 5.0%
Total per depth 40.0% 26.0% 10.0% 11.0% 10.0% 3.0%
main environmental parameters sharing (temperature, precip-itation, slope exposure, inclination, altitude) between the twoselected study sites. These constraints provided a reliable pic-ture about the adaptive longitudinal growth of first-order lateral
roots of a woody species to slope condition. Moreover, the rootsystems examined were only minimally disturbed. They grew ina homogeneous mass of soil; no stones were present except inloam-steep slope condition at depth higher than 20 cm.
Fig. 3. Initial angle of growth plotted against the depth of origin for all the first-order lateral roots of 5–7 years-old Spartium junceum plants grown in Clay (a) andLoam (b) soils. Filled circle (�) refers to the shallow-slope condition, open circle (o) refers to the steep-slope condition. Regression equations are as follows, whereia = initial angle; d = depth of origin. Clay shallow-slope: ia = 96.84–1.37d (P < 0.001, R2 = 0.31); Clay steep-slope: ia = 99.45–1.64d (P < 0.001, R2 = 0.32); Loamshallow-slope: ia = 86.73–0.81d (P < 0.006, R2 = 0.09); Loam steep-slope: ia = 74.96–0.16d (P = 0.3, R2 = 0.01).
212 A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215
Fig. 4. The angle of growth as a function of the distance from the taproot junction for the first-order lateral roots emitted within 5 cm soil depth (surface roots) of5–7 years-old Spartium junceum plants: a) upslope sector (90◦–270◦); downslope sector (270◦–90◦). Filled circle (�) refers to Clay soil, open circle (o) refers toLoam soil. Continuous lines (–) refer to the shallow-slope condition, dashed lines (- - -) refer to the steep-slope condition. Values are means ± S.E. of 9–36 roots.
Table 2F e P values of ANOVA for the initial angle and the subsequent angle of growth of the first-order lateral roots emitted within 5 cm soil depth (surface roots) of S.junceum
Parameter Length (df = 1) Soil (df = 1) Slope (df = 1) Soil* Slo (df = 1)
Angle of growth in the upslope sector F 9.53 65.48 0.22 16.11P 0.002 <0.001 0.636 <0.001
Angle of growth in the dw-slope sector F 6.82 3.79 19.26 2.19P 0.009 0.053 <0.001 0.14
Initial angle (2 cm distance) F 15.29 0.25 1.22P <0.001 0.62 0.271
Soil type and slope inclination were used as fixed factors, root length as covariate. Boldface P values are significant at a probability level <0.05.
To our knowledge, this is the first study showing the direc-tion of growth of surface laterals in a woody species growingin two different soil types and soil surface inclination. Couttsand Nicoll (1991) found that both Lodgepole pine and Sitkaspruce growing in peat soil showed an inherent tendency forupward growth in surface laterals approaching an ascendingslope, but did not present observations in different soils. There
are obvious advantages in having roots capable of remaining inthe warm, well-aerated surface layers where nutrients are con-centrated, without their emerging into the air where the tipswould be damaged by desiccation (Coutts and Nicoll, 1991).As a consequence, on a slope, plants may show upward growthof surface roots facing upslope provided that the soil conditionsare favourable for their growth. Thus, the upward growth results
Table 3Some soil characteristics at the four growth conditions
Layer (depth (cm)) Texture pH H2O C/N N (g/kg) P (ppm) K (ppm) Organic matter g/1000g
Loam shallow-slope A (0–10) Sandy Loam 8.4 9.2 0.5 0 254 8.0C (10–40) Loam 8.4 6 0.6 0 299 6.3
Loam steep-slope A (0–10) Loam 8.4 6.1 0.3 0 140 3.2
Clay shallow-slope A (0–7) Clay 8.4 8.2 0.9 2.9 180.4 13.0C1 (7–30) Silty Clay 8.6 7.5 0.4 2.3 124.8 5.1C2 (30–60) Clay 8.7 10.3 0.4 2.4 132.4 6.2
Clay steep-slope A (0–5) Clay 8.4 12.3 1.1 8.5 208.6 22.2C (5–50) Clay 8.4 11.8 1.0 8.6 189.6 20.7
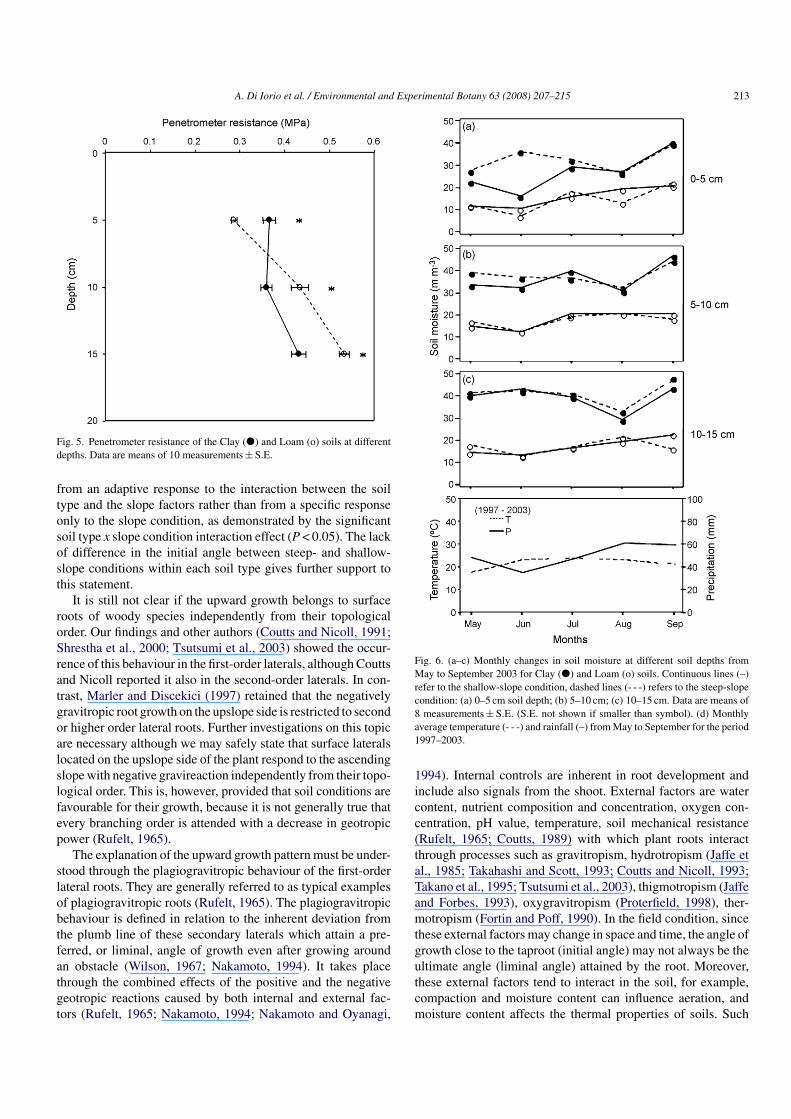
A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215 213
Fig. 5. Penetrometer resistance of the Clay (�) and Loam (o) soils at differentdepths. Data are means of 10 measurements ± S.E.
from an adaptive response to the interaction between the soiltype and the slope factors rather than from a specific responseonly to the slope condition, as demonstrated by the significantsoil type x slope condition interaction effect (P < 0.05). The lackof difference in the initial angle between steep- and shallow-slope conditions within each soil type gives further support tothis statement.
It is still not clear if the upward growth belongs to surfaceroots of woody species independently from their topologicalorder. Our findings and other authors (Coutts and Nicoll, 1991;Shrestha et al., 2000; Tsutsumi et al., 2003) showed the occur-rence of this behaviour in the first-order laterals, although Couttsand Nicoll reported it also in the second-order laterals. In con-trast, Marler and Discekici (1997) retained that the negativelygravitropic root growth on the upslope side is restricted to secondor higher order lateral roots. Further investigations on this topicare necessary although we may safely state that surface lateralslocated on the upslope side of the plant respond to the ascendingslope with negative gravireaction independently from their topo-logical order. This is, however, provided that soil conditions arefavourable for their growth, because it is not generally true thatevery branching order is attended with a decrease in geotropicpower (Rufelt, 1965).
The explanation of the upward growth pattern must be under-stood through the plagiogravitropic behaviour of the first-orderlateral roots. They are generally referred to as typical examplesof plagiogravitropic roots (Rufelt, 1965). The plagiogravitropicbehaviour is defined in relation to the inherent deviation fromthe plumb line of these secondary laterals which attain a pre-ferred, or liminal, angle of growth even after growing aroundan obstacle (Wilson, 1967; Nakamoto, 1994). It takes placethrough the combined effects of the positive and the negativegeotropic reactions caused by both internal and external fac-tors (Rufelt, 1965; Nakamoto, 1994; Nakamoto and Oyanagi,
Fig. 6. (a–c) Monthly changes in soil moisture at different soil depths fromMay to September 2003 for Clay (�) and Loam (o) soils. Continuous lines (–)refer to the shallow-slope condition, dashed lines (- - -) refers to the steep-slopecondition: (a) 0–5 cm soil depth; (b) 5–10 cm; (c) 10–15 cm. Data are means of8 measurements ± S.E. (S.E. not shown if smaller than symbol). (d) Monthlyaverage temperature (- - -) and rainfall (–) from May to September for the period1997–2003.
1994). Internal controls are inherent in root development andinclude also signals from the shoot. External factors are watercontent, nutrient composition and concentration, oxygen con-centration, pH value, temperature, soil mechanical resistance(Rufelt, 1965; Coutts, 1989) with which plant roots interactthrough processes such as gravitropism, hydrotropism (Jaffe etal., 1985; Takahashi and Scott, 1993; Coutts and Nicoll, 1993;Takano et al., 1995; Tsutsumi et al., 2003), thigmotropism (Jaffeand Forbes, 1993), oxygravitropism (Proterfield, 1998), ther-motropism (Fortin and Poff, 1990). In the field condition, sincethese external factors may change in space and time, the angle ofgrowth close to the taproot (initial angle) may not always be theultimate angle (liminal angle) attained by the root. Moreover,these external factors tend to interact in the soil, for example,compaction and moisture content can influence aeration, andmoisture content affects the thermal properties of soils. Such

214 A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215
interactions can make field and laboratory observations difficultto interpret. Since in this study, in slope condition, moisture con-tent and nutrient concentration were significantly higher in theupper layer of the clay soil (Table 3), the root apex facing ups-lope might had sensed such favourable conditions respondingwith an upward growth during its plagiogeotropic elongation.Thus, there must be plasticity within the plagiotropic response ofthese secondary laterals rather than there being a strong internalcontrol of upward growth.
It still remains unclear which of the many factors that varywith soil depth was responsible of this plagiotropic plasticity.Distance from the soil surface is believed to be more control-ling of root orientation than is gravitropism (Coutts and Nicoll,1991). Our findings partly support this hypothesis because inloam soil the initial and subsequent angles of growth did notshow any relationship with the soil depth (Fig. 2). In contrast,the good relationship occurring in clay soil is consistent withobservations on Pinus taeda L. (Bilan et al., 1978) and Pinuscontorta Douglas ex Loudon and Picea sitchensis Bong. Carr.(Coutts and Nicoll, 1991). Thus, our findings would be furtherevidence of the plasticity of the plagiotropic response to the soilenvironment.
For nodal roots of maize, it is reported that a slight increasein penetration resistance suppresses downward curvature, thusmaintaining a larger liminal angle (Nakamoto, 1994), whereas alow water content induces downward growth (Nakamoto, 1993).According to these findings, the occurrence of a low penetrationresistance along with low water content should be expected toinduce a marked downward growth. In this study, both bulk den-sity and soil moisture in the loam soil were lower than in the claysoil both in the upper layers in shallow-slope and through the20 cm soil vertical profile extent because of the surface incli-nation in steep-slope. The percentage of root branches with awide initial angle (90◦–110◦; Table 1) was lower than in claysoil. These findings give support to our hypothesis and to theconclusions of Nakamoto (1993, 1994). Moreover, always inloam soil, the strong penetration resistance increase at higherdepth than 15 cm on shallow-slope might had suppressed rootgrowth whereas, wherever possible, the low water content mighthad induced downward growth, as highlighted by the high per-centage of root branches with the narrow initial angle (50◦–70◦degrees; Table 1).
In clay soil, in the upper layers, the significantly higher pene-tration resistance explains the higher percentage of roots with awider initial angle of growth (90◦–110◦ degrees; Table 1). More-over, the higher compaction along with the oxytropic response(Proterfield, 1998) towards the more aerated surface layer mayexplain the upward growth observed in the root branches foundwithin 5 cm depth.
Because in clay soil the penetration resistance does notchange within 20 cm depth and there was no marked differencesbetween the two growth conditions (Section 2, Table 1), hypoxiacondition may explain the lack of root branches at higher depththan 30 cm.
The detailed mechanism on how the roots sense and respondto the loading is not yet fully understood. It is current opin-ion and of the authors to state that the root apex of the surface
laterals facing upslope elongates forward into the soil respond-ing mainly to their water and nutrients concentrations status.The lack of differences in the shoot dry weight among thefour growth conditions and the lack of upward oriented surfaceroots in the loam soil give further support to this hypothesis.It is only at a later stage, when the weight of the above-ground part of the plant assumes higher values that surfaceroots respond to the loading but at cambium level, distributingresource between laterals in function of their anchorage contri-bution (Chiatante et al., 2003b; Di Iorio et al., 2005; Scippa etal., 2006).
In conclusion, the soil type rather than the soil surface incli-nation affected the liminal angle of first-order lateral roots of ashrub species and promoted, at least to an extent, the upwardgrowth on surface roots. This behaviour must stand in theplasticity of the plagiotropic response of these secondary lat-erals rather than in a strong internal control. Moreover, thisbehaviour appears to be an inherent tendency expressed when aplagiotropic woody lateral root grows in favourable soil patcheson the upslope side of a plant.
Acknowledgments
Some of the results reported herein were obtained when work-ing on the European project “Eco-slopes” (QLK5-2001-00289).The authors also acknowledge the Italian MIUR (Italian Min-istry for University and Research) and the European project“COST Action E38”. The authors would like to express specialthanks to Maria Sarnataro for the soil classification.
References
Bilan, M.V., Leach, J.H. Davis, G., 1978. Root development in loblolly pine(Pinus taeda L.) from two Texas seed sources. In: van Eerden E., Kighorn,J.M. (Eds.), Root Form of Planted Trees. Canadian Forest Service, JointReport no. 8, pp 17–22.
Chiatante, D., Sarnataro, M., Fusco, S., Di Iorio, A., Scippa, G.S., 2003a. Modi-fication of root morphological parameters and root architecture in seedlingsof Fraxinus ornus L. and Spartium junceum L. growing on slopes. PlantBiosyst. 137, 47–56.
Chiatante, D., Scippa, G.S., Di Iorio, A., Sarnataro, M., 2003b. The influence ofsteep slope on root system development. J. Plant Growth Regul. 21, 247–260.
Coutts, M.P., 1987. Developmental processes in tree root systems. Can. J. Res.17, 761–767.
Coutts, M.P., 1989. Factors affecting the direction of growth of tree roots. In:Dreyer, E., Aussenac, G., BonnetMasimbert, M., Dizengremel, P., Favre,J.M., Garrec, J.P., Le Tacon, F., Martin, F. (Eds.), Annales des SciencesForestieres, 46 Supplement on Forest Tree Physiology. Elsevier/INRA, Paris,pp. 277–287.
Coutts, M.P., Nicoll, B.C., 1991. Orientation of the lateral roots of trees, I.Upward growth of surface roots and deflection near the soil surface. NewPhytol. 119, 227–234.
Coutts, M.P., Nicoll, B.C., 1993. Orientation of the lateral roots of trees, II.Hydrotropic and gravitropic responses of lateral roots of Sitka spruce grownin air at different humidities. New Phytol. 124, 277–281.
Danjon, F., Sinoquet, H., Godin, C., Colin, F., Drexhage, M., 1999. Characteri-zation of structural tree root architecture using 3D digitizing and AMAPmodsoftware. Plant Soil 211, 241–258.
Di Iorio, A., Lasserre, B., Scippa, G.S., Chiatante, D., 2005. Root system archi-tecture of Quercus pubescens trees growing on different sloping conditions.Ann. Bot. (Lond.) 95, 351–361.

A. Di Iorio et al. / Environmental and Experimental Botany 63 (2008) 207–215 215
Fayle, D.C.F., 1975. Extension and longitudinal growth during the developmentof red pine root systems. Can. J. Res. 5, 109–121.
Fortin, M.A., Poff, K.L., 1990. Temperature sensing by primary roots of maize.Plant Physiol. (Rockv.) 9, 637–639.
Godin, C., Guedon, Y., Costes, E., Caraglio, Y., 1997. Measuring and analyzingplants with theAMAPmod software. In: Michalewicz, M. (Ed.), Advances inComputational Life Science. Vol I: Plants to Ecosystems. CISRO, Australia,pp. 63–94.
Jaffe, M.J., Takahashi, H., Biro, R.L., 1985. A pea mutant for the study ofhydrotropism in roots. Science 230, 445–447.
Jaffe, M.J., Forbes, S., 1993. Thigmomorphogenesis, the effect of mechanicalperturbation on plants. Plant Growth Regul. 12, 313–324.
Leopold, A.C., Wettlaufer, S.H., 1988. Diagravitropism in corn roots. PlantPhysiol. (Rockv.) 87, 803–805.
Marler, T.E., Discekici, H.M., 1997. Root development of ‘Red Lady’ papayaplants grown on a hillside. Plant Soil 195, 37–42.
Nakamoto, T., 1993. Effect of soil water content on the gravitropic behavior ofnodal roots in maize. Plant Soil 152 (2), 261–267.
Nakamoto, T., 1994. Plagiogravitropism of maize roots. Plant Soil 165, 327–332.Nakamoto, T., Oyanagi, A., 1994. The direction of growth of seminal roots
of Triticum aestivum L. and experimental modification thereof. Ann. Bot.(Lond.) 73, 363–367.
Proterfield, D.M., 1998. The tropic response of plant roots to oxygen: oxytropismin Pisum sativum L. Planta 206, 1–6.
Rufelt, H., 1965. Plagiogeotropism in roots. Encyclopedia of Plant Physiology,vol. 17. Springer Verlag, Berlin, pp. (2): 322–343.
Scippa, G.S., Di Michele, M., Di Iorio, A., Costa, A., Lasserre, B., Chiatante,D., 2006. Root response to slope: anchorage and gene factors involved. Ann.Bot. (Lond.) 97, 857–866.
Shrestha, M.B., Horiuchi, M., Yamadera, Y., Miyazaki, T., 2000. A study onthe adaptability mechanism of tree roots on steep slopes. In: Stokes, A.(Ed.), The Supporting Roots of Trees and Woody Plants: Form, Functionand Physiology. Kluwer Academis Press, pp. 51–57.
Sinoquet, H., Rivet, P., 1997. Measurement and visualization of the architectureof an adult tree based on a three-dimensional digitising device. Trees (Berl.)11, 265–270.
Takahashi, H., Scott, T.K., 1993. Intensity of hydrostimulation for the inductionof root hydrotropism and its sensing by the root cap. Plant Cell Environ. 16,99–103.
Takano, M., Takahashi, H., Hirasawa, T., Suge, H., 1995. Hydrotropism in roots:sensing of a gradient in water potential by the root cap. Planta 197, 410–413.
Tsutsumi, D., Kosugi, K., Mizuyama, T., 2003. Root-system development andwater-extraction model considering hydrotropism. Soil Sci. Soc. Am. J. 67,387–401.
Tsutsumi, D., Kosugi, K., Mizuyama, T., 2004. Three-dimensional modelingof hydrotropism effects on plant root architecture along a hillslope. VadoseZone J. 3, 1017–1030.
USDA, 1998. Keys to Soil Taxonomy, eighth ed. USDA, Natural ResourcesConservation Service (NRCS), Washington, D.C.
Wilson, B.F., 1967. Root growth around barriers. Botanical Gazette 128 (2),79–82.
Copyright © 2022 FDOKUMEN




















