
"Privilege" and "Immunity" as Used in the Property Restatement
Upload
independentCategory
view
0download
0
Activated Monocytes in Peritumoral Stroma ofHepatocellular Carcinoma Promote Expansion of
Memory T Helper 17 CellsDong-Ming Kuang,1,2 Chen Peng,2 Qiyi Zhao,2 Yan Wu,2 Min-Shan Chen,1 and Limin Zheng1,2
Although cancer patients exhibit a generalized immunosuppressive status, substantial evi-dence indicates that the inflammatory reaction at a tumor site can promote tumor growthand progression. Hepatocellular carcinoma (HCC) is usually derived from inflamed cir-rhotic liver with extensive leukocyte infiltration. We recently found that proinflammatory Thelper (Th)17 cells are accumulated in HCC tissue, where they promote disease progressionby fostering angiogenesis. Here we show that interleukin (IL)-17-producing cells were en-riched predominantly in peritumoral stroma of HCC tissues, and their levels were wellcorrelated with monocyte/macrophage density in the same area. Most peritumoral CD68�
cells exhibited an activated phenotype. Accordingly, tumor-activated monocytes were sig-nificantly superior to the suppressive tumor macrophages in inducing expansion of Th17cells from circulating memory T cells in vitro with phenotypic features similar to thoseisolated from HCCs. Moreover, we found that tumor-activated monocytes secreted a set ofkey proinflammatory cytokines that triggered proliferation of functional Th17 cells. Inhi-bition of monocytes/macrophages inflammation in liver markedly reduced the level of tu-mor-infiltrating Th17 cells and tumor growth in vivo. Conclusion: The proinflammatoryTh17 cells are generated and regulated by a fine-tuned collaborative action between differenttypes of immune cells in distinct HCC microenvironments, and allows the inflammatoryresponse of activated monocytes to be rerouted in a tumor-promoting direction. Selectivelymodulating the “context” of inflammatory response in tumors might provide a novel strat-egy for anticancer therapy. (HEPATOLOGY 2010;51:154-164.)
Hepatocellular carcinoma (HCC) is characterizedby progressive development, high postsurgicalrecurrence, and extremely poor prognosis. The
dismal outcome has been attributed to the highly vascularnature of HCC, which increases the propensity to spreadand invade into neighboring or distant sites.1
Tumor progression is now recognized as the product ofevolving crosstalk between different cell types within tu-
mors.2,3 HCC is usually present in inflamed fibroticand/or cirrhotic liver with extensive leukocyte infiltration.Thus, the immune status at a tumor site can largely influ-ence the biologic behavior of HCC.1,4 Recent studies haveshown that high infiltration of intratumoral regulatory Tcells is associated with reduced survival and increased in-vasiveness in HCC.5 These findings are in accordancewith the general view that the tumor microenvironment
Abbreviations: Ab, antibody; APCs, antigen-presenting cells; CCM, conditioned medium from control (untreated) monocytes; HCC, hepatocellular carcinoma; IFN,interferon; IL, interleukin; M�, macrophage(s); TAM, tumor-associated M�; TCM, conditioned medium from TSN-exposed monocytes; TGF, transforming growth factor;Th, T helper; TIL, tumor-infiltrating leukocyte; TNF, tumor necrosis factor; TSN, tumor culture supernatant.
From the 1State Key Laboratory of Oncology in Southern China, Cancer Center, Guangzhou, P.R. China; 2State Key Laboratory of Biocontrol, School of Life Sciences,Sun Yat-Sen University, Guangzhou, P.R. China.
Received April 12, 2009; accepted August 12, 2009.Supported by the Outstanding Young Scientist Fund and project grants from the NSFC, China (30425025 and 30730086), the “973” Program (2004CB518801 and
2010CB529904), and the Ministry of Health of P.R. China (2008ZX10002-019).Address reprint requests to: Limin Zheng, School of Life Sciences, Sun Yat-Sen University, Guangzhou 510 275, P.R. China. E-mail: [email protected]; fax:
86-20-84112169.Copyright © 2009 by the American Association for the Study of Liver Diseases.Published online in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/hep.23291Potential conflict of interest: Nothing to report.Additional Supporting Information may be found in the online version of this article.
154

induces tolerance. However, there is substantial evidencethat the inflammatory response associated with cancerscan also promote tumor progression by stimulating an-giogenesis and tissue remodeling.4,6
Macrophages (M�) constitute a major component ofthe leukocyte infiltrate in tumors. These cells are derivedfrom circulating monocytes, and, in response to environ-mental signals, they acquire special phenotypic character-istics with diverse functions.7-9 In a previous study, wefound that tumor environments can alter the normal de-velopment of M� that is intended to trigger transientearly activation of monocytes in the peritumoral region,which in turn induces formation of suppressive M� incancer nests.8 Notably, the density of these activatedmonocytes is selectively associated with vascular invasionand poor prognosis in HCC patients.10 These resultsstrongly indicate that, besides inducing immune toler-ance, tumors may also reroute the proinflammatory im-mune response into a tumor-promoting direction,although the relative mechanism remains largely un-known.
A subset of interleukin (IL)-17-producing CD4� Thelper 17 (Th17) cells with potent proinflammatoryproperties has recently been detected in human tu-mors.11,12 Studies in other systems have found that severalkey cytokines, including IL-1�, IL-6, IL-23, tumor ne-crosis factor alpha (TNF-�), and transforming growthfactor beta (TGF-�), can create a cytokine milieu thatregulates the expansion of human Th17 cells.13-17 Thesecytokines are often present in environments that have thepotential to promote the incidence and growth of tu-mors.18-20 Despite the implications of Th17 cells indi-cated by the mentioned findings, very little is knownabout the nature and regulation of Th17 in human tu-mors.
In a recent study,21 we observed that Th17 cells werehighly enriched in HCCs and their levels were positivelycorrelated with microvessel density in tissues and poorsurvival in HCC patients. In contrast to the classical Th17cells that hardly express interferon (IFN)-�, almost half ofthe IL-17-producing CD4� T cells we isolated fromHCC tissues were able to simultaneously produce IFN-�,suggesting that the tumor microenvironment can pro-foundly determine the phenotype of such cells. Inasmuchas monocytes/M� represent an abundant population ofantigen-presenting cells (APCs) in solid tumors and theirdensity is inversely associated with the prognosis inHCC,8,10 we investigated whether monocytes/M� canregulate Th17 and, if so, how they exert that influence,paying particular attention to the tissue microlocalizationand phenotype of these cells in HCC.
Patients and Methods
Patients and Specimens. Detailed informationabout the patients and specimens is described in the Sup-porting Materials and Methods and Supporting Table 1.
Isolation of Mononuclear Cells from PeripheralBlood and Tissues. Peripheral leukocytes were isolatedby Ficoll density gradient centrifugation.8,22 Isolation oftumor-infiltrating leukocytes (TILs) are detailed in theSupporting Materials and Methods.
Cell Lines and Preparation of Tumor Culture Su-pernatant (TSN). Detailed information about cell linesand preparation of TSNs is described in the SupportingMaterials and Methods.
Isolation of Monocytes and Preparation of Condi-tioned Media. Monocytes were selected from peripheralblood mononuclear cells using anti-CD14 magnetic beads(Miltenyi Biotec). To generate conditioned media, mono-cytes were left untreated or cultured for 1 hour with 20%TSN from HepG2 cells and then washed and cultured inRPMI 1640 containing 10% human AB serum for 16 hours.Thereafter, the supernatants were harvested, centrifuged,and stored in aliquots at �80°C.
Enzyme-Linked Immunosorbent Assay (ELISA).Concentrations of cytokines were determined usingELISA kits (eBioscience, San Diego, CA).
In Vitro T Cell Culture System. In 2-day incubation,purified CD3 T cells, naive T cells, and memory T cells(Miltenyi Biotec) were left untreated or were coculturedwith autologous monocytes or M� or were exposed to50% conditioned medium or medium supplementedwith recombinant IL-6, IL-23, and/or IL-1� (Peprotech)in the presence of 2 �g/mL anti-CD3 and 1 �g/mL anti-CD28. Thereafter, cells were washed and maintained inRPMI medium supplemented with 20 IU/mL IL-2 for in-dicated times with conditioned medium or different cyto-kines. In some experiments, cells were labeled withcarboxyfluorescein succinimidyl ester (CFSE) or pretreatedwith a neutralizing Ab against IL-6, IL-23, or IL-1�, or acontrol immunoglobulin G (IgG) (R&D Systems), and sub-sequently exposed to conditioned media.
Flow Cytometry. Detailed information about flowcytometry is described in the Supporting Materials andMethods and Supporting Table 2.
Immunohistochemistry and Immunofluorescence.Paraffin-embedded or frozen HCC samples were pro-cessed for immunohistochemistry or immunofluores-cence, respectively, as described in the SupportingMaterials and Methods.
Evaluation of Immunohistochemical Variables.The evaluation of immunohistochemical variables is de-tailed in the Supporting Materials and Methods.
HEPATOLOGY, Vol. 51, No. 1, 2010 KUANG ET AL. 155

Immunoblotting. The proteins were extracted as de-scribed10 and as detailed in the Supporting Materials andMethods.
In Vivo M� Inhibition. All mouse procedures wereperformed as described in the Supporting Materials andMethods.
Statistical Analysis. The statistical analysis is detailedin the Supporting Materials and Methods.
Results
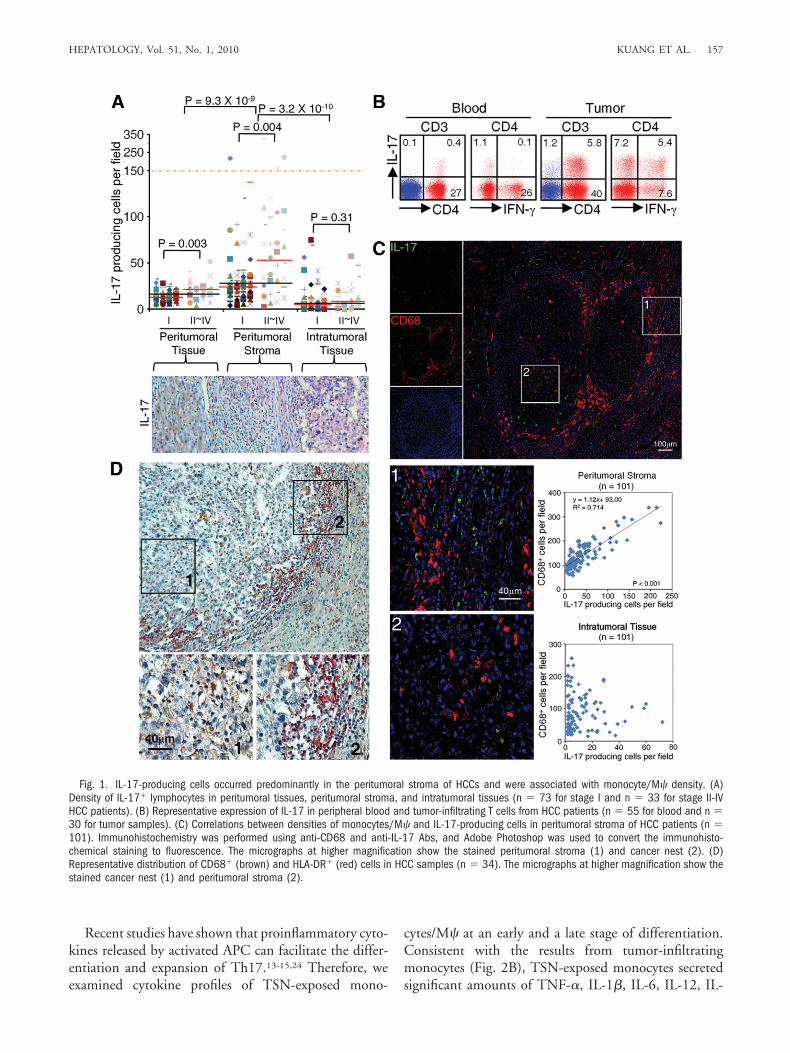
Correlations Between Monocyte/M� and IL-17-Producing Cell Density in Peritumoral Stroma ofHCC Patients. APCs are critical for initiating and main-taining tumor-specific T-cell responses. Because M�markedly outnumber other APCs in tumors,8,23 we firstinvestigated the association between monocytes/M� andIL-17-producing cells in human HCCs, paying particularattention to the tissue microlocalization of the cells. Thepresence of IL-17� cells was visualized by immunohisto-chemical staining of IL-17 in paraffin-embedded tissuesfrom 106 untreated HCC patients. As shown in Fig. 1A,such cells were present throughout the tissue, but oftenpredominantly in the peritumoral stroma rather than inthe cancer nests (43.2 � 4.9 and 10.5 � 1.2 cells/field,respectively; n � 106 for both). The numbers of IL-17�
cells in both peritumoral tissue and stroma were signifi-cantly increased and correlated with disease progression inHCC patients.
To identify the phenotypic features of tumor IL-17�
cells, we used flow cytometry to analyze leukocytes freshlyisolated from tissues obtained from 30 HCC patients un-dergoing surgery. The results showed that the levels ofIL-17� cells were significantly higher in tumors (7.6% �1.6%) than in nontumoral liver tissues (2.8% � 0.7%)and peripheral blood (0.7% � 0.1%; n � 55; P � 0.0001for both). Most tissue of IL-17� T cells (81.7% � 8.8%)were CD4� and appeared to be Th17 cells. Interestingly,over 40% of tumor Th17 cells were able to produce bothIL-17 and IFN-� (Fig. 1B). By contrast, most of the cir-culating Th17 cells did not express IFN-� (Fig. 1B). Thedifferences in phenotypes between circulating and tumorTh17 cells indicate that the tumor environment can pro-mote the expansion/differentiation of Th17 cells in situ.
M� are also predominantly found in the peritumoralstroma,8,23 and hence we examined the correlation be-tween densities of M� and Th17 cells in serial sections ofHCCs stained for CD68 (marker for monocytes/M�)and IL-17. In peritumoral stroma we found a significantcorrelation between the levels of CD68� cells and IL-17�
lymphocytes (r � 0.845, P � 0.001). However, there wasno such correlation in intratumoral tissue (Fig. 1C and
Supporting Fig. 1), suggesting that M� in different partsof a tumor play disparate roles in Th17 cell expansion.Furthermore, most of the CD68� cells in peritumoralstroma had a smaller volume and showed marked expres-sion of human leukocyte antigen DR (HLA-DR) (Fig.1D), which implies that they were newly recruited andactivated monocytes. In contrast, most M� in the cancernests were negative for HLA-DR. Confocal microscopicanalysis showed that most CD68� cells in HCC tumorswere also stained positive for CD14, but negative for CD3(Supporting Fig. 2).
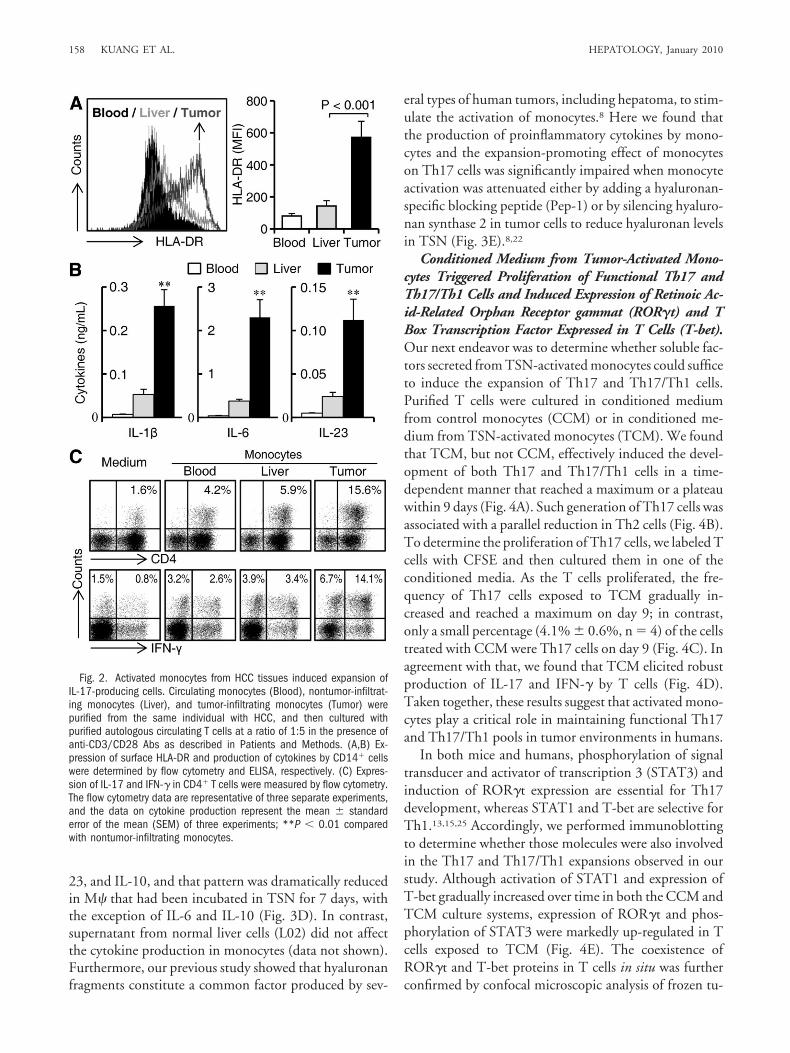
Tumor-Activated Monocytes Promote Expansion ofIL-17-Producing Cells from Memory T Cell Popula-tion. We recently observed that tumor environments canalter the normal development of M� that is intended totrigger transient early activation of monocytes in the peri-tumoral region.8 To investigate whether such a mecha-nism is also responsible for the selective accumulation ofTh17 in peritumoral stroma, we purified circulatingmonocytes (CD14high cells), nontumor-and tumor-infil-trating monocytes from the same HCC patient, and thencultured those cells with purified autologous T cells. Theresult showed that tumor monocytes expressed higher lev-els of HLA-DR, secreted larger amounts of inflammatorycytokines IL-1�, IL-6, and IL-23 (Fig. 2A,B), and weremore potent in promoting the expansion of Th17 cellswith phenotypic features similar to those isolated fromHCCs (Fig. 2C).
To further elucidate the effect of tumor mono-cytes/M� on Th17 expansion, we incubated monocyteswith TSN from hepatoma cells to generate tumor-acti-vated monocytes or immunosuppressive tumor-associ-ated M� (TAMs) (Fig. 3A),8 and then cultured those cellswith purified autologous T cells. The results showed thattumor-activated monocytes were significantly superior toTAMs in inducing Th17 expansion, whereas the effects ofcontrol monocytes or M� were negligible. Notably, al-most half of the Th17 cells generated from tumor-acti-vated monocytes were able to produce both IL-17 andIFN-� (Th17/Th1), whereas most of the TAM-inducedTh17 cells were negative for IFN-� (Fig. 3B).
We used commercial kits to further purify naive andmemory T cells. Tumor-activated monocytes effectivelypromoted the expansion of Th17 cells (45.4% � 7.2%,n � 4) from memory CD4� T cells, and most of thesememory IL-17� cells (65.7% � 11%, n � 4) were alsoable to produce IFN-� (Fig. 3C). These results indicatethat, compared to immunosuppressive TAMs, monocytesactivated by a short exposure to a tumor environment playa more prominent role in the development of memory Tcells into IL-17-producing T cells that have phenotypicfeatures similar to those of tumor-infiltrating Th17 cells.
156 KUANG ET AL. HEPATOLOGY, January 2010
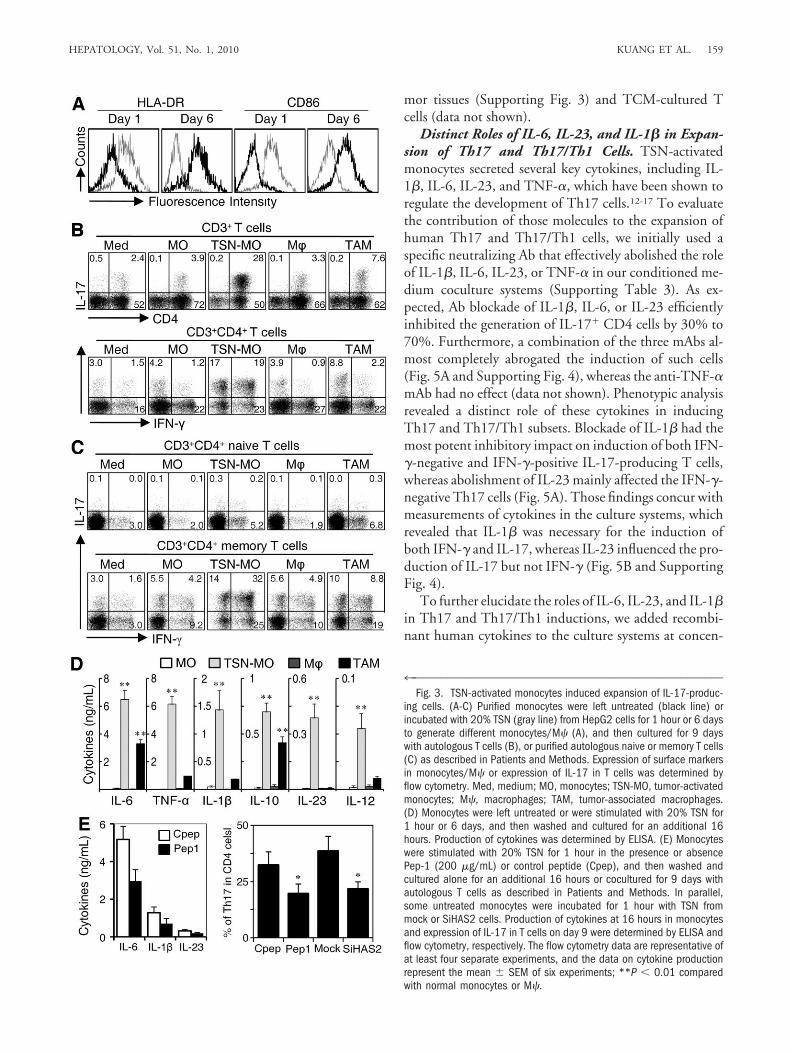
Recent studies have shown that proinflammatory cyto-kines released by activated APC can facilitate the differ-entiation and expansion of Th17.13-15,24 Therefore, weexamined cytokine profiles of TSN-exposed mono-
cytes/M� at an early and a late stage of differentiation.Consistent with the results from tumor-infiltratingmonocytes (Fig. 2B), TSN-exposed monocytes secretedsignificant amounts of TNF-�, IL-1�, IL-6, IL-12, IL-
Fig. 1. IL-17-producing cells occurred predominantly in the peritumoral stroma of HCCs and were associated with monocyte/M� density. (A)Density of IL-17� lymphocytes in peritumoral tissues, peritumoral stroma, and intratumoral tissues (n � 73 for stage I and n � 33 for stage II-IVHCC patients). (B) Representative expression of IL-17 in peripheral blood and tumor-infiltrating T cells from HCC patients (n � 55 for blood and n �30 for tumor samples). (C) Correlations between densities of monocytes/M� and IL-17-producing cells in peritumoral stroma of HCC patients (n �101). Immunohistochemistry was performed using anti-CD68 and anti-IL-17 Abs, and Adobe Photoshop was used to convert the immunohisto-chemical staining to fluorescence. The micrographs at higher magnification show the stained peritumoral stroma (1) and cancer nest (2). (D)Representative distribution of CD68� (brown) and HLA-DR� (red) cells in HCC samples (n � 34). The micrographs at higher magnification show thestained cancer nest (1) and peritumoral stroma (2).
HEPATOLOGY, Vol. 51, No. 1, 2010 KUANG ET AL. 157
23, and IL-10, and that pattern was dramatically reducedin M� that had been incubated in TSN for 7 days, withthe exception of IL-6 and IL-10 (Fig. 3D). In contrast,supernatant from normal liver cells (L02) did not affectthe cytokine production in monocytes (data not shown).Furthermore, our previous study showed that hyaluronanfragments constitute a common factor produced by sev-
eral types of human tumors, including hepatoma, to stim-ulate the activation of monocytes.8 Here we found thatthe production of proinflammatory cytokines by mono-cytes and the expansion-promoting effect of monocyteson Th17 cells was significantly impaired when monocyteactivation was attenuated either by adding a hyaluronan-specific blocking peptide (Pep-1) or by silencing hyaluro-nan synthase 2 in tumor cells to reduce hyaluronan levelsin TSN (Fig. 3E).8,22
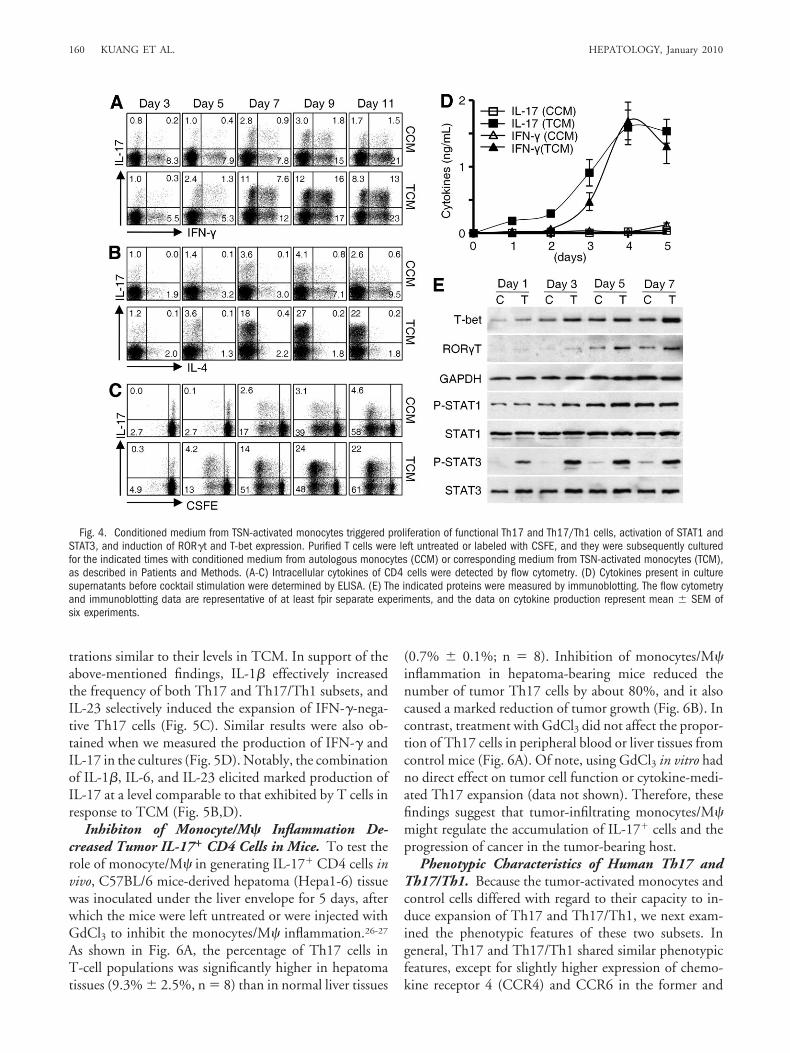
Conditioned Medium from Tumor-Activated Mono-cytes Triggered Proliferation of Functional Th17 andTh17/Th1 Cells and Induced Expression of Retinoic Ac-id-Related Orphan Receptor gammat (ROR�t) and TBox Transcription Factor Expressed in T Cells (T-bet).Our next endeavor was to determine whether soluble fac-tors secreted from TSN-activated monocytes could sufficeto induce the expansion of Th17 and Th17/Th1 cells.Purified T cells were cultured in conditioned mediumfrom control monocytes (CCM) or in conditioned me-dium from TSN-activated monocytes (TCM). We foundthat TCM, but not CCM, effectively induced the devel-opment of both Th17 and Th17/Th1 cells in a time-dependent manner that reached a maximum or a plateauwithin 9 days (Fig. 4A). Such generation of Th17 cells wasassociated with a parallel reduction in Th2 cells (Fig. 4B).To determine the proliferation of Th17 cells, we labeled Tcells with CFSE and then cultured them in one of theconditioned media. As the T cells proliferated, the fre-quency of Th17 cells exposed to TCM gradually in-creased and reached a maximum on day 9; in contrast,only a small percentage (4.1% � 0.6%, n � 4) of the cellstreated with CCM were Th17 cells on day 9 (Fig. 4C). Inagreement with that, we found that TCM elicited robustproduction of IL-17 and IFN-� by T cells (Fig. 4D).Taken together, these results suggest that activated mono-cytes play a critical role in maintaining functional Th17and Th17/Th1 pools in tumor environments in humans.
In both mice and humans, phosphorylation of signaltransducer and activator of transcription 3 (STAT3) andinduction of ROR�t expression are essential for Th17development, whereas STAT1 and T-bet are selective forTh1.13,15,25 Accordingly, we performed immunoblottingto determine whether those molecules were also involvedin the Th17 and Th17/Th1 expansions observed in ourstudy. Although activation of STAT1 and expression ofT-bet gradually increased over time in both the CCM andTCM culture systems, expression of ROR�t and phos-phorylation of STAT3 were markedly up-regulated in Tcells exposed to TCM (Fig. 4E). The coexistence ofROR�t and T-bet proteins in T cells in situ was furtherconfirmed by confocal microscopic analysis of frozen tu-
Fig. 2. Activated monocytes from HCC tissues induced expansion ofIL-17-producing cells. Circulating monocytes (Blood), nontumor-infiltrat-ing monocytes (Liver), and tumor-infiltrating monocytes (Tumor) werepurified from the same individual with HCC, and then cultured withpurified autologous circulating T cells at a ratio of 1:5 in the presence ofanti-CD3/CD28 Abs as described in Patients and Methods. (A,B) Ex-pression of surface HLA-DR and production of cytokines by CD14� cellswere determined by flow cytometry and ELISA, respectively. (C) Expres-sion of IL-17 and IFN-� in CD4� T cells were measured by flow cytometry.The flow cytometry data are representative of three separate experiments,and the data on cytokine production represent the mean � standarderror of the mean (SEM) of three experiments; **P � 0.01 comparedwith nontumor-infiltrating monocytes.
158 KUANG ET AL. HEPATOLOGY, January 2010
mor tissues (Supporting Fig. 3) and TCM-cultured Tcells (data not shown).
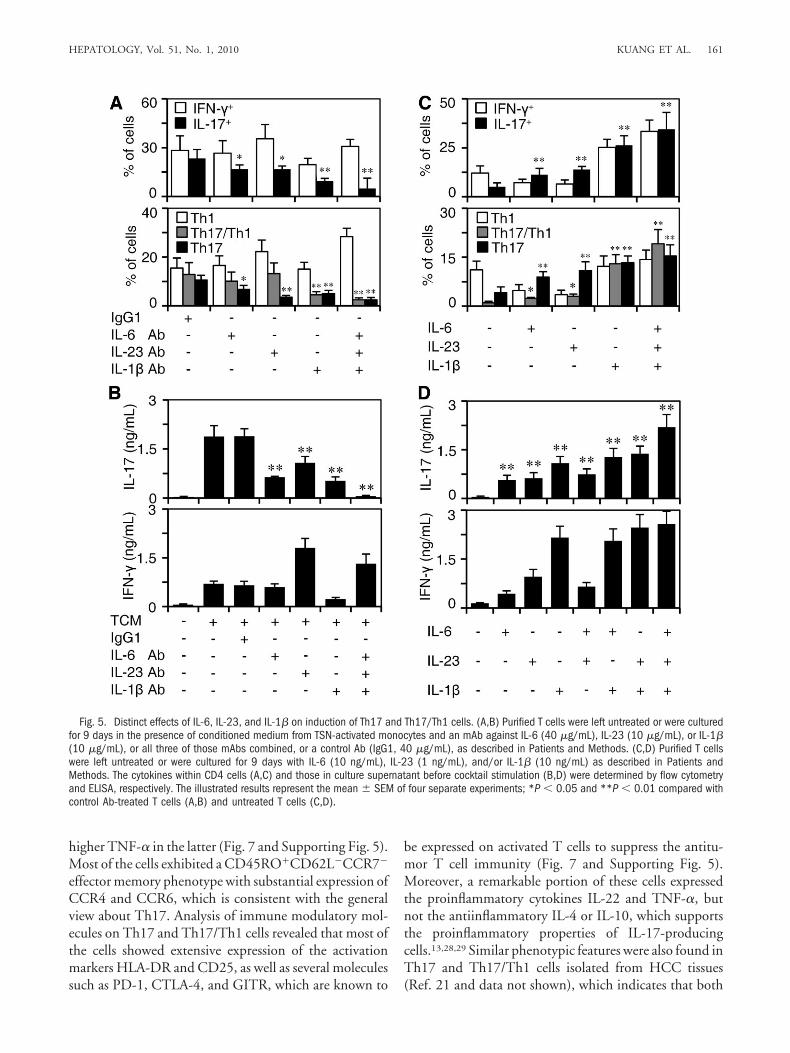
Distinct Roles of IL-6, IL-23, and IL-1� in Expan-sion of Th17 and Th17/Th1 Cells. TSN-activatedmonocytes secreted several key cytokines, including IL-1�, IL-6, IL-23, and TNF-�, which have been shown toregulate the development of Th17 cells.12-17 To evaluatethe contribution of those molecules to the expansion ofhuman Th17 and Th17/Th1 cells, we initially used aspecific neutralizing Ab that effectively abolished the roleof IL-1�, IL-6, IL-23, or TNF-� in our conditioned me-dium coculture systems (Supporting Table 3). As ex-pected, Ab blockade of IL-1�, IL-6, or IL-23 efficientlyinhibited the generation of IL-17� CD4 cells by 30% to70%. Furthermore, a combination of the three mAbs al-most completely abrogated the induction of such cells(Fig. 5A and Supporting Fig. 4), whereas the anti-TNF-�mAb had no effect (data not shown). Phenotypic analysisrevealed a distinct role of these cytokines in inducingTh17 and Th17/Th1 subsets. Blockade of IL-1� had themost potent inhibitory impact on induction of both IFN-�-negative and IFN-�-positive IL-17-producing T cells,whereas abolishment of IL-23 mainly affected the IFN-�-negative Th17 cells (Fig. 5A). Those findings concur withmeasurements of cytokines in the culture systems, whichrevealed that IL-1� was necessary for the induction ofboth IFN-� and IL-17, whereas IL-23 influenced the pro-duction of IL-17 but not IFN-� (Fig. 5B and SupportingFig. 4).
To further elucidate the roles of IL-6, IL-23, and IL-1�in Th17 and Th17/Th1 inductions, we added recombi-nant human cytokines to the culture systems at concen-
4™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™Fig. 3. TSN-activated monocytes induced expansion of IL-17-produc-
ing cells. (A-C) Purified monocytes were left untreated (black line) orincubated with 20% TSN (gray line) from HepG2 cells for 1 hour or 6 daysto generate different monocytes/M� (A), and then cultured for 9 dayswith autologous T cells (B), or purified autologous naive or memory T cells(C) as described in Patients and Methods. Expression of surface markersin monocytes/M� or expression of IL-17 in T cells was determined byflow cytometry. Med, medium; MO, monocytes; TSN-MO, tumor-activatedmonocytes; M�, macrophages; TAM, tumor-associated macrophages.(D) Monocytes were left untreated or were stimulated with 20% TSN for1 hour or 6 days, and then washed and cultured for an additional 16hours. Production of cytokines was determined by ELISA. (E) Monocyteswere stimulated with 20% TSN for 1 hour in the presence or absencePep-1 (200 �g/mL) or control peptide (Cpep), and then washed andcultured alone for an additional 16 hours or cocultured for 9 days withautologous T cells as described in Patients and Methods. In parallel,some untreated monocytes were incubated for 1 hour with TSN frommock or SiHAS2 cells. Production of cytokines at 16 hours in monocytesand expression of IL-17 in T cells on day 9 were determined by ELISA andflow cytometry, respectively. The flow cytometry data are representative ofat least four separate experiments, and the data on cytokine productionrepresent the mean � SEM of six experiments; **P � 0.01 comparedwith normal monocytes or M�.
HEPATOLOGY, Vol. 51, No. 1, 2010 KUANG ET AL. 159
trations similar to their levels in TCM. In support of theabove-mentioned findings, IL-1� effectively increasedthe frequency of both Th17 and Th17/Th1 subsets, andIL-23 selectively induced the expansion of IFN-�-nega-tive Th17 cells (Fig. 5C). Similar results were also ob-tained when we measured the production of IFN-� andIL-17 in the cultures (Fig. 5D). Notably, the combinationof IL-1�, IL-6, and IL-23 elicited marked production ofIL-17 at a level comparable to that exhibited by T cells inresponse to TCM (Fig. 5B,D).
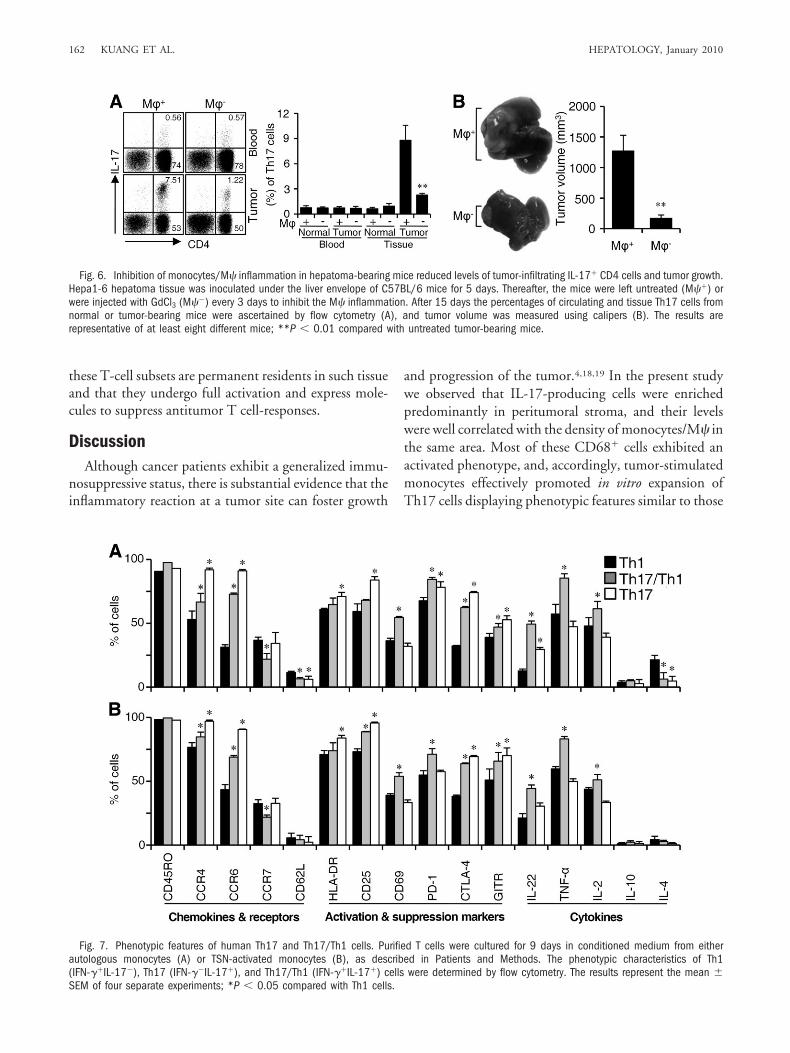
Inhibiton of Monocyte/M� Inflammation De-creased Tumor IL-17� CD4 Cells in Mice. To test therole of monocyte/M� in generating IL-17� CD4 cells invivo, C57BL/6 mice-derived hepatoma (Hepa1-6) tissuewas inoculated under the liver envelope for 5 days, afterwhich the mice were left untreated or were injected withGdCl3 to inhibit the monocytes/M� inflammation.26-27
As shown in Fig. 6A, the percentage of Th17 cells inT-cell populations was significantly higher in hepatomatissues (9.3% � 2.5%, n � 8) than in normal liver tissues
(0.7% � 0.1%; n � 8). Inhibition of monocytes/M�inflammation in hepatoma-bearing mice reduced thenumber of tumor Th17 cells by about 80%, and it alsocaused a marked reduction of tumor growth (Fig. 6B). Incontrast, treatment with GdCl3 did not affect the propor-tion of Th17 cells in peripheral blood or liver tissues fromcontrol mice (Fig. 6A). Of note, using GdCl3 in vitro hadno direct effect on tumor cell function or cytokine-medi-ated Th17 expansion (data not shown). Therefore, thesefindings suggest that tumor-infiltrating monocytes/M�might regulate the accumulation of IL-17� cells and theprogression of cancer in the tumor-bearing host.
Phenotypic Characteristics of Human Th17 andTh17/Th1. Because the tumor-activated monocytes andcontrol cells differed with regard to their capacity to in-duce expansion of Th17 and Th17/Th1, we next exam-ined the phenotypic features of these two subsets. Ingeneral, Th17 and Th17/Th1 shared similar phenotypicfeatures, except for slightly higher expression of chemo-kine receptor 4 (CCR4) and CCR6 in the former and
Fig. 4. Conditioned medium from TSN-activated monocytes triggered proliferation of functional Th17 and Th17/Th1 cells, activation of STAT1 andSTAT3, and induction of ROR�t and T-bet expression. Purified T cells were left untreated or labeled with CSFE, and they were subsequently culturedfor the indicated times with conditioned medium from autologous monocytes (CCM) or corresponding medium from TSN-activated monocytes (TCM),as described in Patients and Methods. (A-C) Intracellular cytokines of CD4 cells were detected by flow cytometry. (D) Cytokines present in culturesupernatants before cocktail stimulation were determined by ELISA. (E) The indicated proteins were measured by immunoblotting. The flow cytometryand immunoblotting data are representative of at least fpir separate experiments, and the data on cytokine production represent mean � SEM ofsix experiments.
160 KUANG ET AL. HEPATOLOGY, January 2010
higher TNF-� in the latter (Fig. 7 and Supporting Fig. 5).Most of the cells exhibited a CD45RO�CD62L�CCR7�
effector memory phenotype with substantial expression ofCCR4 and CCR6, which is consistent with the generalview about Th17. Analysis of immune modulatory mol-ecules on Th17 and Th17/Th1 cells revealed that most ofthe cells showed extensive expression of the activationmarkers HLA-DR and CD25, as well as several moleculessuch as PD-1, CTLA-4, and GITR, which are known to
be expressed on activated T cells to suppress the antitu-mor T cell immunity (Fig. 7 and Supporting Fig. 5).Moreover, a remarkable portion of these cells expressedthe proinflammatory cytokines IL-22 and TNF-�, butnot the antiinflammatory IL-4 or IL-10, which supportsthe proinflammatory properties of IL-17-producingcells.13,28,29 Similar phenotypic features were also found inTh17 and Th17/Th1 cells isolated from HCC tissues(Ref. 21 and data not shown), which indicates that both
Fig. 5. Distinct effects of IL-6, IL-23, and IL-1� on induction of Th17 and Th17/Th1 cells. (A,B) Purified T cells were left untreated or were culturedfor 9 days in the presence of conditioned medium from TSN-activated monocytes and an mAb against IL-6 (40 �g/mL), IL-23 (10 �g/mL), or IL-1�(10 �g/mL), or all three of those mAbs combined, or a control Ab (IgG1, 40 �g/mL), as described in Patients and Methods. (C,D) Purified T cellswere left untreated or were cultured for 9 days with IL-6 (10 ng/mL), IL-23 (1 ng/mL), and/or IL-1� (10 ng/mL) as described in Patients andMethods. The cytokines within CD4 cells (A,C) and those in culture supernatant before cocktail stimulation (B,D) were determined by flow cytometryand ELISA, respectively. The illustrated results represent the mean � SEM of four separate experiments; *P � 0.05 and **P � 0.01 compared withcontrol Ab-treated T cells (A,B) and untreated T cells (C,D).
HEPATOLOGY, Vol. 51, No. 1, 2010 KUANG ET AL. 161
these T-cell subsets are permanent residents in such tissueand that they undergo full activation and express mole-cules to suppress antitumor T cell-responses.
DiscussionAlthough cancer patients exhibit a generalized immu-
nosuppressive status, there is substantial evidence that theinflammatory reaction at a tumor site can foster growth
and progression of the tumor.4,18,19 In the present studywe observed that IL-17-producing cells were enrichedpredominantly in peritumoral stroma, and their levelswere well correlated with the density of monocytes/M� inthe same area. Most of these CD68� cells exhibited anactivated phenotype, and, accordingly, tumor-stimulatedmonocytes effectively promoted in vitro expansion ofTh17 cells displaying phenotypic features similar to those
Fig. 6. Inhibition of monocytes/M� inflammation in hepatoma-bearing mice reduced levels of tumor-infiltrating IL-17� CD4 cells and tumor growth.Hepa1-6 hepatoma tissue was inoculated under the liver envelope of C57BL/6 mice for 5 days. Thereafter, the mice were left untreated (M��) orwere injected with GdCl3 (M��) every 3 days to inhibit the M� inflammation. After 15 days the percentages of circulating and tissue Th17 cells fromnormal or tumor-bearing mice were ascertained by flow cytometry (A), and tumor volume was measured using calipers (B). The results arerepresentative of at least eight different mice; **P � 0.01 compared with untreated tumor-bearing mice.
Fig. 7. Phenotypic features of human Th17 and Th17/Th1 cells. Purified T cells were cultured for 9 days in conditioned medium from eitherautologous monocytes (A) or TSN-activated monocytes (B), as described in Patients and Methods. The phenotypic characteristics of Th1(IFN-��IL-17�), Th17 (IFN-��IL-17�), and Th17/Th1 (IFN-��IL-17�) cells were determined by flow cytometry. The results represent the mean �SEM of four separate experiments; *P � 0.05 compared with Th1 cells.
162 KUANG ET AL. HEPATOLOGY, January 2010

seen in such cells isolated from HCCs. These findingssuggest an intricate mechanism in which Th17 cells inhumans are generated and regulated by a fine-tuned col-laborative action between different types of immune cellsin distinct tumor microenvironments.
Human tumor tissues can be classified anatomicallyinto areas of intratumoral and peritumoral stroma, eachwith distinct compositions and functional properties.4,8,30
Intratumoral environments usually contain abundant im-munosuppressive molecules and cells to evade immunerecognition.31 In contrast, peritumoral stroma contains asignificant number of infiltrated leukocytes, which arethereby situated close to the advancing edge of a tu-mor.8,9,22 In the current study we observed that Th17 cellswere present primarily in the peritumoral stroma, andthey were colocalized with monocytes/M� that exhibitedan activated phenotype. However, the number of theseperitumoral Th17 cells was correlated with disease pro-gression, and we found that these tumor-activated mono-cytes could not stimulate antitumor T-cell responses inHCC patients.32 These observations suggest that theproinflammatory responses in peritumoral stroma maynot represent the host reaction to the malignancy, but thatthey instead constitute effects that are rerouted in a tu-mor-promoting direction to induce tissue remodelingand angiogenesis. This notion is supported by our recentinvestigations in which we found that the frequency oftissue Th17 cells was positively correlated with microves-sel density in tumors, and high numbers of monocytes inperitumoral stroma were selectively associated with vascu-lar invasion and poor prognosis in HCC patients.10,21
Consistent with our observations, recent studies haveshown that IL-17 could recruit neutrophils, which in turnstimulate angiogenesis and tissue remodeling.33-34
Despite recent advances in understanding the differenti-ation of Th17 cells in humans,15-19 little is known about themechanisms underlying the regulation of Th17 cells in tu-mors. The present investigation provides evidence thatproinflammatory cytokines released by tumor-activatedmonocytes/M� play a dominant role in the development ofTh17 cells in HCCs, as indicated by the results of four sets ofexperiments. First, we observed that the level of Th17 cellswas about 4 times higher in peritumoral stroma than in can-cer nests, and there were significant correlations between thedensities of Th17 and HLA-DRhighCD68� cells in peritu-moral stroma, which was not the case in cancer nests, wheremost of the CD68� cells were negative for HLA-DR. Sec-ond, tumor-activated monocytes were significantly superiorto the suppressive TAMs in inducing expansion of Th17cells exhibiting phenotypic features more similar to those oftumor-infiltrating Th17 cells (e.g., a remarkable proportionof Th17/Th1). Third, blocking a set of cytokines released
from tumor-activated monocytes clearly inhibited the gen-eration of Th17 cells, and relatively low concentrations of therecombinant cytokines could mimic the stimulatory effect ofTCM culturing in this regard. Fourth, inhibition of mono-cytes/M� inflammation in hepatoma-bearing mice mark-edly reduced the number of tumor Th17 cells and tumorgrowth. Therefore, activation of monocytes in tumors mayrepresent a novel route to promote Th17 expansion in hu-man cancer. This concept is supported by studies showingthat activated APCs are involved in the differentiation andexpansion of Th17 cells and thereby also in Th17-mediatedchronic inflammation.16,35,36 It should be noted that, in ad-dition to the local expansion of Th17 cells, migration fromblood is also a potential source for the increased Th17 cells intumors. In this context, we have recently found that CCR6 isexpressed in the majority of Th17 cells and that CCL20, theligand for CCR6, is significantly increased in HCCs.21
In one of our latest studies21 we observed that most of theTh17 isolated from HCCs exhibited a CD45RO�
CD62L�CCR7� effector memory phenotype. Consistentwith that finding, in the present experiments we found thattumor-activated monocytes promoted the development ofIL-17-producing cells from memory T cells. Moreover, weidentified several key cytokines secreted by tumor-activatedmonocytes that appeared to be responsible for the expansionof Th17. One of those molecules, IL-1�, played a dominantrole in the induction of both Th17 and Th17/Th1, whereasIL-23 stimulated essentially only Th17. IL-6 was also in-volved in this process, although to a lesser extent. Theseresults agree with the general view that a proinflammatorycytokine milieu facilitates the development of Th17.12-17,37
Interestingly, we noted that comparatively low concentra-tions of these proinflammatory cytokines (i.e., about 1/5 to1/10 of the levels commonly used by other researchers) ef-fectively induced the generation of both Th17 and Th17/Th1. Other studies have also demonstrated a critical role ofTGF-� in the development of human Th17,14,15 but we didnot find such a correlation in HCC tissues (data not shown).In one of the mentioned studies, it was observed that TGF-�up-regulated ROR�t expression but simultaneously inhib-ited the ability of that molecule to induce IL-17 expression,and that such inhibition can be relieved by proinflammatorycytokines.15 Thus, the proinflammatory cytokines might bethe critical determinant in triggering Th17 expansion inHCC tissues, which usually contain TGF-� produced byboth tumor and stroma cells.
There is substantial evidence that it is not inflammationper se, but rather the inflammatory “context” that determinesthe ability of proinflammatory factors to facilitate or preventtumor growth.4,18,19 Our results provide important new in-sights into the role of monocytes/M� in human tumor pro-gression. Soluble factors derived from cancer cells can trigger
HEPATOLOGY, Vol. 51, No. 1, 2010 KUANG ET AL. 163

transient activation of newly recruited monocytes in the peri-tumoral stroma8 and thereby induce the monocytes to pro-duce a significant amount of cytokines. The IL-1�, IL-6, andIL-23 promote Th17-mediated inflammation in peritu-moral stroma, whereas TNF-� and IL-10 released from tu-mor-activated monocytes up-regulate PD-L1 on the surfaceof those cells to inhibit tumor-specific T-cell immunity.32 Inthat way, these activated monocytes repurpose the inflam-matory response away from antitumor immunity (thesword) and toward tissue remodeling and proangiogenicpathways (a plowshare). Therefore, studying the mecha-nisms that can selectively modulate the functional activitiesof monocytes/M� might provide a novel strategy for anti-cancer therapy.
References1. Semela D, Dufour JF. Angiogenesis and hepatocellular carcinoma. J Hepa-
tol 2004;41:864-880.2. Mueller MM, Fusenig NE. Friends or foes — bipolar effects of the tumor
stroma in cancer. Nat Rev Cancer 2004;4:839-849.3. Tlsty TD, Coussens LM. Tumor stroma and regulation of cancer devel-
opment. Annu Rev Pathol 2006;1:119-150.4. Budhu A, Forgues M, Ye QH, Jia HL, He P, Zanetti KA, et al. Prediction
of venous metastases, recurrence, and prognosis in hepatocellular carci-noma based on a unique immune response signature of the liver microen-vironment. Cancer Cell 2006;10:99-111.
5. Fu J, Xu D, Liu Z, Shi M, Zhao P, Fu B, et al. Increased regulatory T cellscorrelate with CD8 T-cell impairment and poor survival in hepatocellularcarcinoma patients. Gastroenterology 2007;132:2328-2339.
6. Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation.Nature 2008;454:436-444.
7. Mosser DM, Edwards JP. Exploring the full spectrum of macrophageactivation. Nat Rev Immunol 2008;8:958-969.
8. Kuang DM, Wu Y, Chen N, Cheng J, Zhuang SM, Zheng L. Tumor-derivedhyaluronan induces formation of immunosuppressive macrophages throughtransient early activation of monocytes. Blood 2007;110:587-595.
9. Lewis CE, Pollard JW. Distinct role of macrophages in different tumormicroenvironments. Cancer Res 2006;66:605-612.
10. Ding, T, Xu J, Wang F, Shi M, Zhang Y, Li SP, et al. High tumor-infiltrating macrophage density predicts poor prognosis in patients withprimary hepatocellular carcinoma after resection. Hum Pathol 2009;40:381-389.
11. Kryczek I, Wei S, Zou L, Altuwaijri S, Szeliga W, Kolls J, et al. Cuttingedge: Th17 and regulatory T cell dynamics and the regulation by IL-2 inthe tumor microenvironment. J Immunol 2007;178:6730-6733.
12. Miyahara Y, Odunsi K, Chen W, Peng G, Matsuzaki J, Wang RF. Gen-eration and regulation of human CD4� IL-17-producing T cells in ovariancancer. Proc Natl Acad Sci U S A 2008;105:15505-15510.
13. Dong C. TH17 cells in development: an updated view of their molecularidentity and genetic programming. Nat Rev Immunol 2008;8:337-348.
14. Volpe E, Servant N, Zollinger R, Bogiatzi SI, Hupe P, Barillot E, et al. Acritical function for transforming growth factor-beta, interleukin 23 andproinflammatory cytokines in driving and modulating human T(H)-17responses. Nat Immunol 2008;9:650-657.
15. Manel N, Unutmaz D, Littman DR. The differentiation of humanT(H)-17 cells requires transforming growth factor-beta and induction ofthe nuclear receptor RORgammat. Nat Immunol 2008;9:641-649.
16. Kryczek I, Wei S, Gong W, et al. Cutting edge: IFN-gamma enables APCto promote memory Th17 and abate Th1 cell development. J Immunol2008;181:5842-5846.
17. Notley CA, Inglis JJ, Alzabin S, Shu X, Szeliga W, Vatan L, et al. Blockadeof tumor necrosis factor in collagen-induced arthritis reveals a novel im-munoregulatory pathway for Th1 and Th17 cells. J Exp Med 2008;205:2491-2497.
18. Kim S, Takahashi H, Lin WW, Descargues P, Grivennikov S, Kim Y, et al.Carcinoma-produced factors activate myeloid cells through TLR2 to stim-ulate metastasis. Nature 2009;457:102-106.
19. Langowski JL, Kastelein RA, Oft M. Swords into plowshares: IL-23 repur-poses tumor immune surveillance. Trends Immunol 2007;28:207-212.
20. Biswas S, Guix M, Rinehart C, Dugger TC, Chytil A, Moses HL, et al.Inhibition of TGF-beta with neutralizing antibodies prevents radiation-induced acceleration of metastatic cancer progression. J Clin Invest 2007;117:1305-1313.
21. Zhang JP, Yan J, Xu J, Pang XH, Chen MS, Li L, et al. Increased intratu-moral IL-17-producing cells correlate with poor survival in hepatocellularcarcinoma patients. J Hepatol 2009;50:980-989.
22. Kuang DM, Zhao Q, Xu J, Yun JP, Wu C, Zheng L. Tumor-educatedtolerogenic dendritic cells induce CD3epsilon down-regulation and apo-ptosis of T cells through oxygen-dependent pathways. J Immunol 2008;181:3089-3098.
23. Kryczek I, Zou L, Rodriguez P, Zhu G, Wei S, Mottram P, et al. B7–H4expression identifies a novel suppressive macrophage population in humanovarian carcinoma. J Exp Med 2006;203:871-881.
24. McGeachy MJ, Cua DJ. Th17 cell differentiation: the long and windingroad. Immunity 2008;28:445-453.
25. Zhang F, Meng G, Strober W. Interactions among the transcription fac-tors Runx1, RORgammat and Foxp3 regulate the differentiation of inter-leukin 17-producing T cells. Nat Immunol 2008;9:1297-1306.
26. Jiang W, Sun R, Wei H, Tian Z. Toll-like receptor 3 ligand attenuatesLPS-induced liver injury by down-regulation of toll-like receptor 4 expres-sion on macrophages. Proc Natl Acad Sci U S A 2005;102:17077-17082.
27. Pendino KJ, Meidhof TM, Heck DE, Laskin JD, Laskin DL. Inhibition ofmacrophages with gadolinium chloride abrogates ozone-induced pulmo-nary injury and inflammatory mediator production. Am J Respir Cell MolBiol 1995;13:125-132.
28. Kolls JK, Linden A. Interleukin-17 family members and inflammation.Immunity 2004;21:467-476.
29. Ma HL, Liang S, Li J, Napierata L, Brown T, Benoit S, et al. IL-22 isrequired for Th17 cell-mediated pathology in a mouse model of psoriasis-like skin inflammation. J Clin Invest 2008;118:597-607.
30. Finak G, Bertos N, Pepin F, Sadekova S, Souleimanova M, Zhao H, et al.Stromal gene expression predicts clinical outcome in breast cancer. NatMed 2008;14:518-527.
31. Rabinovich GA, Gabrilovich D, Sotomayor EM. Immunosuppressivestrategies that are mediated by tumor cells. Annu Rev Immunol 2007;25:267-296.
32. Kuang DM, Zhao Q, Peng C, Xu J, Zhang JP, et al. Activated monocytesin peritumoral stroma of hepatocellular carcinoma foster immune privilegeand disease progression through PD-L1. J Exp Med 2009;206:1327-1337.
33. Laan M, Cui ZH, Hoshino H, Lotvall J, Sjostrand M, Gruenert DC, et al.Neutrophil recruitment by human IL-17 via C-X-C chemokine release inthe airways. J Immunol 1999;162:2347-2352.
34. Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells inthe promotion of tumour angiogenesis. Nat Rev Cancer 2008;8:618-631.
35. Evans HG, Suddason T, Jackson I, Taams LS, Lord GM. Optimal induc-tion of T helper 17 cells in humans requires T cell receptor ligation in thecontext of Toll-like receptor-activated monocytes. Proc Natl Acad Sci U SA 2007;104:17034-17039.
36. van Beelen AJ, Zelinkova Z, Taanman-Kueter EW, Muller FJ, HommesDW, Zaat SA, et al. Stimulation of the intracellular bacterial sensor NOD2programs dendritic cells to promote interleukin-17 production in humanmemory T cells. Immunity 2007;27:660-669.
37. Annunziato F, Cosmi L, Santarlasci V, Maggi L, Liotta F, Mazzinghi B, etal. Phenotypic and functional features of human Th17 cells. J Exp Med2008;204:1849-1861.
164 KUANG ET AL. HEPATOLOGY, January 2010
Copyright © 2022 FDOKUMEN




















![Detection of Pancreatic Carcinomas by Imaging Lactose-Binding Protein Expression in Peritumoral Pancreas Using [18F]Fluoroethyl-Deoxylactose PET/CT](https://static.fdokumen.com/doc/165x107/631be6bc93f371de19012dfd/detection-of-pancreatic-carcinomas-by-imaging-lactose-binding-protein-expression.jpg)