
Newly hired lighting technician electrocuted while working ...
Upload
rwth-aachenCategory
view
0download
0
PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Sutter, C.]On: 27 September 2010Access details: Access Details: [subscription number 926896716]Publisher Psychology PressInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
The Quarterly Journal of Experimental PsychologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t716100704
Action control while seeing mirror images of one's own movements:Effects of perspective on spatial compatibilityChristine Suttera; Jochen Müsselera
a Department of Work and Cognitive Psychology, RWTH Aachen University, Aachen, Germany
First published on: 19 February 2010
To cite this Article Sutter, Christine and Müsseler, Jochen(2010) 'Action control while seeing mirror images of one's ownmovements: Effects of perspective on spatial compatibility', The Quarterly Journal of Experimental Psychology, 63: 9,1757 — 1769, First published on: 19 February 2010 (iFirst)To link to this Article: DOI: 10.1080/17470210903511244URL: http://dx.doi.org/10.1080/17470210903511244
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Action control while seeing mirror images of one’s ownmovements: Effects of perspective on spatial
compatibility
Christine Sutter and Jochen MusselerDepartment of Work and Cognitive Psychology, RWTH Aachen University, Aachen, Germany
It is examined whether and how the perspective of seeing one’s own movements exerted an influenceon action control. Such change in the perspective challenges an information processor, as she or he hasto handle successfully the discrepancies between the tactile/proprioceptive feedback and the visualfeedback on a projection screen. In the experiments participants responded to visual stimuli, butsaw their responses either from the top or with an x-axis reflection, a y-axis reflection, or a reflectionabout both axes. The results showed that a change in perspective did not impair performance as long asthe left–right relations corresponded with the body space (e.g., with visual feedback reflected aboutthe x-axis). By contrast, performance was drastically reduced when visual feedback was reflectedabout the y-axis or about both axes, so that performed left-side (right-side) movements were seenas right-side (left-side) movements. It is concluded that an actor preferably relies on the informationof the visual system, but refers to the tactile/proprioceptive information when it facilitates the taskdemands.
Keywords: Sensorimotor transformation; Multimodal processing; Action effect; Imitation; Stimulus–response compatibility.
The human tendency to mimic actions per-formed by others is a major precondition formotor skill learning. For example, in an aerobicsession with a face-to-face formation the imita-tor easily copies an instructor’s movement oflifting the right hand by lifting his or her lefthand (spatial congruency between instructorand imitator). However, difficulties occur whenthe imitator mimics the left movement of theinstructor anatomically—that is, by lifting his
or her right hand (anatomic congruencybetween instructor and imitator). The ability torepresent objects as they appear from viewpointsother than one’s own and the coordinationbetween these perspectives with one’s owndevelops in childhood. With increasing age thechildren’s ability to imitate a model’s behaviourchanges from simply mirroring to anatomicallycorrect transposition. However, in adulthoodwe still prefer mirroring actions of others
Correspondence should be addressed to Christine Sutter, RWTH Aachen University, Department of Work and Cognitive
Psychology, Jagerstraße 17–19, 52056, Aachen, Germany. E-mail: [email protected]
We are grateful to two anonymous reviewers for their constructive comments and suggestions on a previous version of this article.
We also wish to thank Bernd Stich for technical assistance and Lina Aurelia Wirth for carrying out the experiments. This research
was supported by a grant from the German Science Foundation to both authors (DFG Su 494/4).
# 2010 The Experimental Psychology Society 1757http://www.psypress.com/qjep DOI:10.1080/17470210903511244
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY
2010, 63 (9), 1757– 1769
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

(e.g., Bekkering, Wohlschlager, & Gattis, 2000;Wapner & Cirillo, 1968).
Various brain imaging studies have shown thatthe observation of another person’s actions canactivate the same motor circuit as that activatedwhen performing the action by oneself (e.g.,Buccino et al., 2004; Iacoboni, Woods, Brass,Bekkering, Mazziotta, & Rizzolatti, 1999; foroverviews see, e.g., Iacoboni & Dapretto, 2006;Rizzolatti & Craighero, 2004). Koski, Iacoboni,Dubeau, Woods, and Mazziotta (2003) found intheir study different patterns of activity withinthe underlying neural system associated with theperspective of imitation. Mirroring was found toengage the mirror system to a greater degreethan anatomic imitation. They concluded thatthe mirror system allows a direct match of the rep-resentation of observed and executed action andthus provides a relatively simple neural precursormechanism for the human ability to imitate.Results from brain imaging studies on the mirrorsystem in humans are consistent with predictionsproposed by common coding theories (for over-views see, e.g., Hommel, Musseler, Aschersleben,& Prinz, 2001; Musseler, 1999). They claim acommon framework for the recognition and plan-ning of motor actions. Perceived events andactions are represented in commensurable featurecodes, which implies facilitation in performancewhen anticipated and performed action effects cor-respond compared with when they do not.
Several studies have already shown the impor-tance of perspective in action planning (e.g.,Bertenthal, Longo, & Kosobud, 2006; Brass,Bekkering, Wohlschlager, & Prinz, 2000;Bruzzo, Borghi, & Ghirlanda, 2008). For instance,Craighero and colleagues (Craighero, Bello,Fadiga, & Rizzolatti, 2002) presented pictures ofthe right hand as seen in a mirror. The stimuli rep-resented the hand’s final posture as achieved ingrasping a bar oriented either clockwise or antic-lockwise. Participants responded to the stimuliwith their right thumb and index finger, andgrasped a plastic bar oriented either clockwise oranticlockwise. The authors found faster (slower)reaction times if the bar orientation betweenstimuli and the participant’s hand end position
corresponded (did not correspond). This resultwas also found for visual stimulation with anadditional 908 rotation of the mirror image ofthe hand. Still, the facilitation in reaction timeswas stronger for unrotated stimuli. Obviously,what counts is the correspondence between pre-sented and to-be-executed hand end positions.
Brass and colleagues (2000) compared theimpact of observing a symbolic, spatial, and imita-tive cue on movement execution. In their exper-iments pictures of a hand were presented thatmatched the posture of the observer’s own handas seen in a mirror. Pictures included an imitativecue (lifting movement of index or middle finger)and a symbolic cue (the digits “1” and “2”). Thetask was to execute a finger movement to eitherthe presented finger movement or the symboliccue (“1” required a movement of the indexfinger, “2” a movement of the middle finger) andto neglect the other dimension. The authorsassumed—similar to the Simon effect—facili-tation of responses in the symbolic cue conditionwith the (irrelevant) congruent imitative cue andan interference effect with the (irrelevant) incon-gruent imitative cue. If participants were askedto respond only to the imitative cue congruencyof the (irrelevant) symbolic cue was expected tohave no effect on the response. Results confirmedthese assumptions: Congruency affected reactiontimes in the symbolic cue condition but not inthe imitative cue condition. Results also showedgenerally faster responses to the imitative cuethan to the symbolic cue.
In further experiments Brass and colleagues(2000) showed that the strong impact of imitativecues did not overrule only symbolic but also spatialcues. In a follow-up study Bertenthal et al. (2006)discussed two competitive processes in actionplanning: automatic imitation and spatial compat-ibility. If participants responded to a mirrored imi-tation task (e.g., observing a left finger movementand responding with a right finger movement), thesame results were observed as in the study of Brassand colleagues. For the anatomic imitation task(e.g., observing a right finger movement andresponding with a right finger movement) reactiontimes to the moving finger were still distinctly
1758 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

faster than those to symbolic cues. However, thepriming effect in the symbolic cue condition wasreversed compared to previous findings: If the irre-levant finger movement was spatially congruent,responses were facilitated, but no interferenceeffect occurred if the irrelevant finger movementwas spatially incongruent. The authors assumedthat both factors contribute to priming and,when placed in opposition, tend to interfere witheach other.
All these studies demonstrated that mirrorimitation is a basic mechanism underlying motorskill learning. Imagine again the aerobic instructorperforming dance sequences in a face-to-face for-mation. This type of mirroring supports thespatial congruency between instructor and imita-tor, while anatomic imitation necessitates left–right transformations. What counts seems to bethe similarity between the observed and executedmovement, and obviously in this respect mirrorimitation is advantageous. This finding is consist-ent with predictions proposed by the ideomotorprinciple (Greenwald, 1970; James, 1890; forrecent overviews and empirical evidence see, e.g.,Hommel et al., 2001; Nattkemper & Ziessler,2004). It holds that actors select, initiate, andexecute an intended movement by activating theanticipatory codes of the action’s effects. Thesemay be representations of body-related effects,like the anticipated tactile feedback from themoving hand (proximal action effect) or the antici-pated visual postures of the moving hand when themovement is executed (distal action effect).Thereby the terms “proximal” and “distal” do notrefer to body-related effects alone (as, e.g., usedin medical context), but include the effects indistant space. Typically, in everyday-life situations,tactile feedback and visual feedback correspond,but they do not correspond any more when thedirect view of one’s own hands is covered, and par-ticipants receive a mirrored visual feedback of theirown movement. In a sense, this situation mergesthe situation of the aerobic instructor and imitatorin one person.
The present study examined whether and howseeing mirror images of one’s own movementsaffects action control. In three experiments we
systematically varied the similarity and the spatialstimulus–response (S–R) compatibility betweenproximal and distal hand movement by disruptingthe visual feedback. We assumed that the simi-larity between actual hand movement and themovement on the screen facilitates action control(egocentric top-down view), while the discrepancybetween movements interfere performance (e.g.,through transformation of performed upwardinto seen downward movements and/or trans-formation of performed left-side movements intoseen right-side movements). Further, weassumed that spatial relationships between per-formed and seen actions play a prominent role.With regard to research in S–R compatibility,responses can be expected to be faster and moreaccurate when stimulus and response position docorrespond than when they do not (for a recentoverview see Proctor & Vu, 2006). However,several studies found that the intended distalaction effects often superpose proximal S–Rrelationships (e.g., Hommel, 1993; Kunde,Musseler, & Heuer, 2007; Massen & Prinz,2007; Musseler, Kunde, Gausepohl, & Heuer,2008).
General procedure
The perspective of seeing one’s own movementswas manipulated in three experiments.Participants sat in front of a fibreboard with fourrectangular arranged response buttons and twogreen/red light-emitting diodes (LEDs). Thefibreboard was covered by an occluder, and partici-pants observed their own movements on the fibre-board via a projection screen in front of them(Figure 1). The location of the flashing LEDdetermined the response finger (left/right LEDmeant to move the index finger of the left/righthand), and its colour indicated to move thefinger to the ipsilateral or contralateral targetbutton. Two aspects are considered: the perspec-tive of the visual feedback presented on the projec-tion screen and the laterality of the movements(Figure 2a).
First, we manipulated the similarity betweenactual hand movement and the movement on the
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1759
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

screen by using four types of visual feedback: (a) Inthe basic condition of all experiments visual feed-back was consistent with looking at one’s ownhand from the top (egocentric top-down view;Figure 2a, lower left). In this condition theactual hand movement and the movement on thescreen are similar. In the following experimentalconditions they are dissimilar. (b) In Experiment1 we distorted the original visual feedback byreflection about the x-axis (Figure 2a, upper left).This perspective is consistent with observingone’s own hand in a mirror (nonegocentric view).That means that performing upward movementsresulted in seen downward movements. (c) InExperiment 2 we reflected the visual feedbackabout the y-axis (Figure 2a, lower right). Thisresulted in an egocentric view of the hands butleft–right relations were inverted—that is, per-forming a left-side movement was seen as a
right-side movement. (d) Consequently, wevaried both the perspective and the left–rightrelations in Experiment 3 by reflecting the originalvisual feedback about both the x- and the y-axes(¼ point reflection; Figure 2a, upper right). Thisperspective is consistent with looking at someoneelse’s hand copying the same action by anatomicimitation. In summary, we varied egocentric(basic condition) and nonegocentric view (exper-imental condition) in Experiment 1, while weretained left–right relations in both conditions.In Experiment 2 the egocentric view was held con-stant, while we either retained (basic condition) orreversed (experimental condition) left–rightrelations, and in Experiment 3 we varied both.
Secondly, although in all experiments the prox-imal hand movements—that is, the movements tothe ipsilateral or contralateral response buttons—were kept constant, distal effects on the projection
Figure 1. The experimental set-up in the experiments. Participants sat in front of a fibreboard with response buttons and light-emitting
diodes (LEDs, bottom left). An occluder prevented direct vision from top, but visual feedback from different perspectives was presented
via a camcorder and a projector on a screen in front of participants. To view a colour version of this figure, please see the online issue of
the Journal.
1760 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010
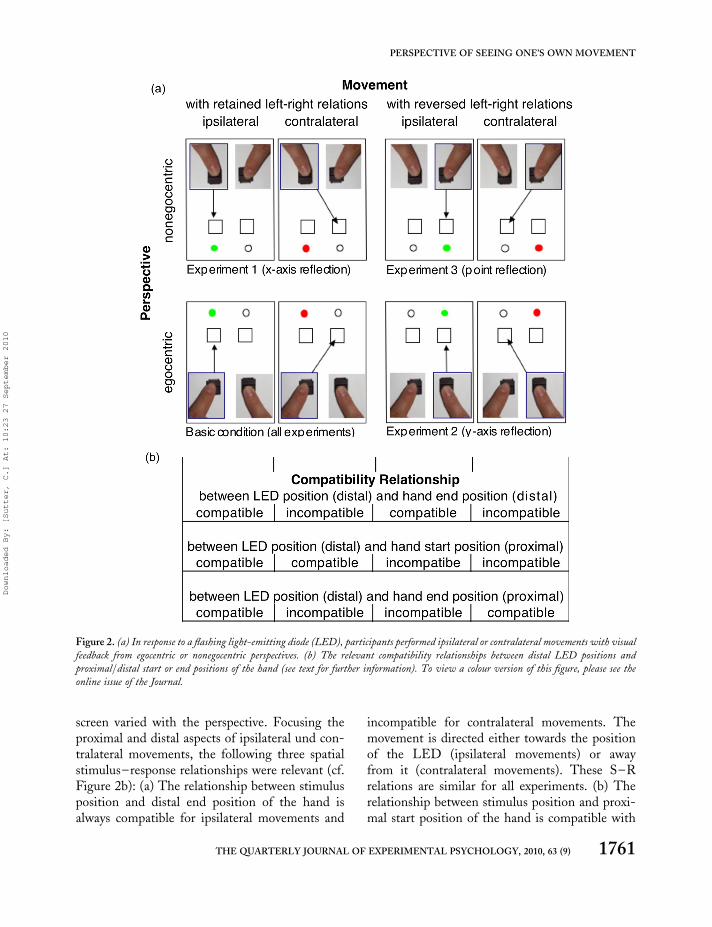
screen varied with the perspective. Focusing theproximal and distal aspects of ipsilateral und con-tralateral movements, the following three spatialstimulus–response relationships were relevant (cf.Figure 2b): (a) The relationship between stimulusposition and distal end position of the hand isalways compatible for ipsilateral movements and
incompatible for contralateral movements. Themovement is directed either towards the positionof the LED (ipsilateral movements) or awayfrom it (contralateral movements). These S–Rrelations are similar for all experiments. (b) Therelationship between stimulus position and proxi-mal start position of the hand is compatible with
Figure 2. (a) In response to a flashing light-emitting diode (LED), participants performed ipsilateral or contralateral movements with visual
feedback from egocentric or nonegocentric perspectives. (b) The relevant compatibility relationships between distal LED positions and
proximal/distal start or end positions of the hand (see text for further information). To view a colour version of this figure, please see the
online issue of the Journal.
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1761
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

retained left–right relations in Experiment 1 (leftpart of Figure 2), but it is incompatible with theadditional left–right transformation introducedin the Experiments 2 and 3 (right part of Figure2). Here, participants responded with their lefthand, but saw a movement of the screen’s righthand and vice versa. (c) The compatibility relation-ship between stimulus position and proximal endposition is compatible/incompatible for ipsilat-eral/contralateral movements in Experiment 1,but reversed with the additional left–right trans-formation in Experiments 2 and 3. The proximalipsilateral movement is directed to the responsebutton located contralateral to the stimulus, andtherefore the stimulus–response relationship isincompatible. In contrast, the proximal contralat-eral movement is directed toward the stimulus pos-ition yielding a compatible stimulus–responserelationship.1
EXPERIMENT 1
Two conditions were compared: the egocentricperspective with view from top (basic condition)and the nonegocentric perspective with a mirroredview (Figure 2a, left part). As mentioned above weexpected superior performance with the egocentricview than with the nonegocentric view, in whichupward movements resulted in seen downwardmovements. Further, ipsilateral movements shouldbe faster and more accurate than contralateral move-ments as ipsilateral movements were markedby approaching the LEDs and contralateral move-ments by avoiding them (e.g., Lavender &Hommel, 2007; Musseler, Aschersleben, Arning,& Proctor, 2009; Yamaguchi & Proctor, 2006).
Method
ParticipantsA total of 8 students (7 female) of the RWTHAachen University, aged between 19 and 24
years (mean age 23 years) participated in the exper-iment for credit in a psychology course or for 10 E.
Apparatus and stimuliThe experiment was carried out in a dimly lit roomand was controlled by an Apple Macintoshcomputer connected with a multi-I/O USB-Box(PMD-1208FS, Measurement ComputingCorporation). The software was written inMatlab using the Psychophysics Toolbox exten-sion (Brainard, 1997; Pelli, 1997).
Participants sat at a table in front of a screen (83� 43 cm) with a distance of approximately 80 cm.A fibreboard (14.5 � 20 cm) with four buttons(1.7 � 1.7 cm) and two green/red LEDs (diam-eter 0.4 cm) was placed in front of them (Figure1). The two lower buttons were used as startkeys, the two upper buttons as target keys, andthe green/red flashing LEDs as stimuli. Twostimulus dimensions were critical: the location ofthe flashing stimulus, which determined theresponse finger (left LED meant to move the leftfinger, and vice versa); and the colour of the stimu-lus, which determined whether the ipsilateral orcontralateral target button was pressed. An occlu-der prevented direct vision of fibreboard andhands. Fibreboard and hands were recorded at30 Hz with a colour camcorder (Canon MV 901,placed 128 cm above the fibreboard), which washooked directly to the projector (Sanyo ProxtraX PLC-XU48, recording at 60 Hz). Thevisual feedback was displayed on the projectionscreen in front of participants with a minimaland irreducible delay of approximately 110 ms.Generally, delays up to 150 ms are noncritical,since the viewed actions are still perceived asself-generated (Franck et al., 2001).
DesignFigure 2a (left part) depicted the main conditionsin this experiment. Participants worked through ablock with an egocentric perspective of visualfeedback (basic condition in all experiments)and through a block with a nonegocentric
1 There is a further, but less informative, compatibility relationship: As the position of the flashing LED determined the response
finger, the relationship between stimulus position and the fingers’ distal start position on the projection screen was always compatible.
1762 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

perspective (Experiment 1). The point of viewwith the egocentric perspective was as lookingat one’s own movement from top. In the nonego-centric condition of this experiment the visualfeedback was reflected about the x-axis—that is,stimuli and hand movements were seen as in amirror, vis-a-vis of the actual performed handmovement. The order of blocks was counterba-lanced across participants. Each block consistedof two laterality conditions (ipsilateral vs. contral-ateral) with 100 repetitions each. Additionally, 40trials were presented in advance of each blockfor training.
ProcedureAt the beginning of a trial, participants had theirindex fingers on the left and right start buttons(Figure 1, bottom left). They were instructed torespond with the left index finger to the leftLED and with the right index finger to theright LED. A green LED indicated that theyshould move the finger to the ipsilateral targetbutton, and the red LED indicated that theyshould move the finger to the contralateraltarget button. Colour of LEDs was counterba-lanced across participants: Half the participantswere asked to press the ipsilateral target buttonto the green LED and the contralateral targetbutton to the red LED. The colour–targetmapping was reversed for the other half ofparticipants. After pressing the correct targetbutton, participants pressed again the startbuttons, and a new trial was initiated. Participantswere instructed to react as fast and accuratelyas possible. In total, the experiment lasted about1 hour.
Results
Mean reaction times (the interval between stimu-lus onset and the release of the start buttons) andmean movement times (the interval betweenstart-button release and target keypress) were cal-culated for error-free trials. Additionally, meanerror percentages were determined on the basisof trials with incorrect target keypresses or whenresponse times exceeded 1,500 ms.
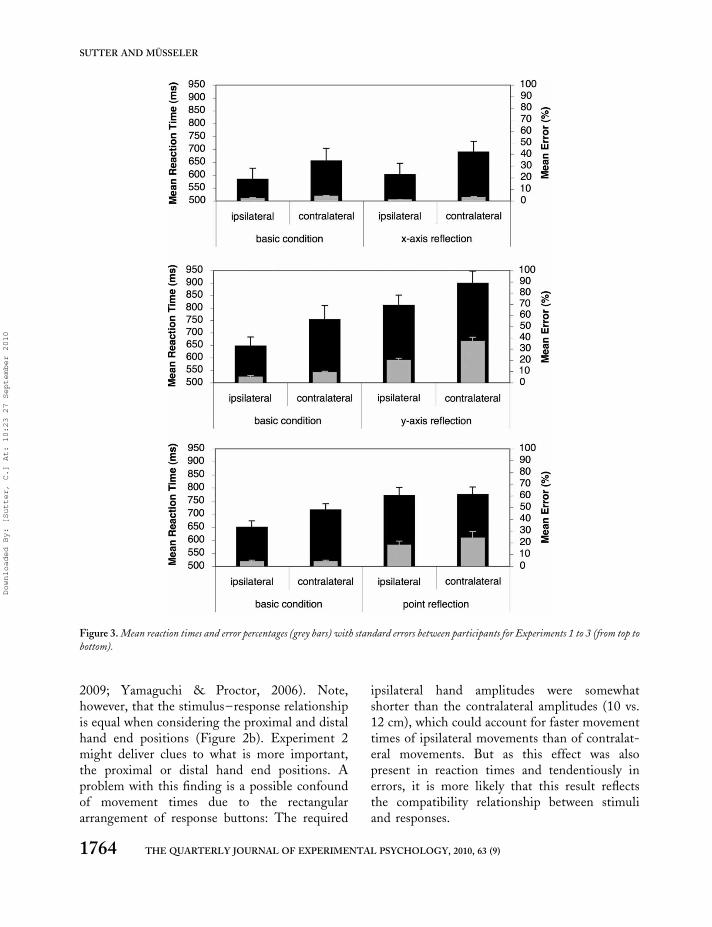
Data were analysed for each of the dependentvariables with 2 � 2 analyses of variances(ANOVAs) with the within-subject factors per-spective (egocentric vs. nonegocentric perspective)and laterality (ipsilateral vs. contralateral). TheANOVAs revealed significant main effects of thefactor laterality in the analyses of reaction times,F(1, 7) ¼ 14.05, MSE ¼ 3,565.87, p , .01,movement times, F(1, 7) ¼ 51.21, MSE ¼823.09, p , .01, and a corresponding trend ofthe errors, F(1, 7) ¼ 5.18, MSE ¼ 5.06, p ¼ .06.Reaction times were 79 ms faster with ipsilateralmovements than with contralateral movements(592 vs. 671 ms; Figure 3, top). This differencewas also observed in movement times (351 vs.423 ms). Additionally, participants tended to bemore accurate in the ipsilateral condition than inthe contralateral condition (2.0 vs. 3.8%). Othereffects were not observed (all ps . .15).
Discussion
The experiment revealed two main findings: Thefirst finding was that—although in the mirroredview upward movements were transformed intoseen downward movements—the results revealedno evidence for superior performance of the ego-centric view over the nonegocentric view. Itseemed that both conditions were equally simpleto perform. Maybe the equalization of both per-spectives originated from the fact that the ego-centric view also contained a transformation: thetransformation of the hand movement from thetable plane to the vertical screen plane. Anotherpossibility is to consider mirroring as a basicmechanism of imitating behaviour in socialsituations, which is important from early child-hood (Bekkering et al., 2000; Wapner &Cirillo, 1968).
The second finding was that action controlwas faster and more accurate for ipsilateral move-ments than for contralateral movements. Thisreflects the compatibility relationship betweenstimuli and responses: Stimulus position andhand end position corresponded in the ipsilateralcondition while they did not correspond in thecontralateral condition (cf. Musseler et al.,
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1763
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010
2009; Yamaguchi & Proctor, 2006). Note,however, that the stimulus–response relationshipis equal when considering the proximal and distalhand end positions (Figure 2b). Experiment 2might deliver clues to what is more important,the proximal or distal hand end positions. Aproblem with this finding is a possible confoundof movement times due to the rectangulararrangement of response buttons: The required
ipsilateral hand amplitudes were somewhatshorter than the contralateral amplitudes (10 vs.12 cm), which could account for faster movementtimes of ipsilateral movements than of contralat-eral movements. But as this effect was alsopresent in reaction times and tendentiously inerrors, it is more likely that this result reflectsthe compatibility relationship between stimuliand responses.
Figure 3. Mean reaction times and error percentages (grey bars) with standard errors between participants for Experiments 1 to 3 (from top to
bottom).
1764 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

EXPERIMENT 2
In this experiment the basic condition was com-pared with a perspective in which feedback wasreflected about the y-axis (Figure 2a, bottomright). This perspective comprised the view fromtop, but the distal visual feedback was now left–right inverted compared to the proximal handmovement. As a consequence the stimulus–response mappings established different compat-ibility relationships (Figure 2b). For instance,when the right LED flashed red on the projectionscreen, participants moved the proximal left handto the contralateral right target button (incompati-ble relationship between LED position and prox-imal hand start position, but compatiblerelationship between LED position and proximalhand end position), but actually saw their handmoving from the right start button to the lefttarget button (incompatible relationship betweenLED position and distal hand end position).
Method
ParticipantsAnother 8 students (5 female), aged between 19and 45 years (mean age 26 years) participated inthe experiment.
Stimuli, procedure, and designThese were the same as those in Experiment1. The basic condition (Figure 2a, bottom left)of the previous experiment was now combinedwith an egocentric condition in which the y-axisis the mirroring axis (Figure 2a, bottom right).
Results
The ANOVA of reaction times revealed signifi-cant main effects for only perspective, F(1, 7) ¼48.10, MSE ¼ 3,972.79, p , .01, and laterality,F(1, 7) ¼ 23.65, MSE ¼ 3,146.35, p , .01.Reaction times were 154 ms faster in the basiccondition than in the condition with the mirroredperspective (698 vs. 853 ms). Ipsilateral
movements were initiated 96 ms faster than con-tralateral movements (727 vs. 824 ms).
Significant main effects were also observed inthe ANOVA of movement times: perspective,F(1, 7) ¼ 6.65, MSE ¼ 12,648.83, p , .05, andlaterality, F(1, 7) ¼ 33.83, MSE ¼ 2,473.35, p, .01. Again the interaction was not significant(p . .40). Ipsilateral movements were executed102 ms faster than contralateral movements (429vs. 531 ms; cf. Figure 3, middle). Movementtimes were 102 ms faster in the basic conditionthan in the condition with the mirrored perspec-tive (429 vs. 531 ms).
The ANOVA on error percentages revealedalso significant main effects of perspective, F(1,7) ¼ 108.76, MSE ¼ 34.0, p , .01, and laterality,F(1, 7) ¼ 27.58, MSE ¼ 29.0, p , .01. Errorpercentages increased from 7% in the basic con-dition to 29% in the condition with the mirroredperspective. The laterality effect was present inboth conditions; however, it was more pronouncedin the mirrored condition, yielding a significantinteraction (5% and 9% for ipsilateral and contral-ateral movements in the basics condition vs. 20%and 37% for ipsilateral and contralateral move-ments in the mirrored condition), F(1, 7) ¼17.69, MSE ¼ 19.83, p , .01.
Discussion
Three main findings were observed: The firstfinding was that the pattern of results in the con-dition with the egocentric view from the top (basiccondition) replicated successfully the correspondingpattern of results of the previous experiment. Thesecond finding was that performance decreased dras-tically with the left–right reversed egocentric feed-back. The decrease was mainly observed inreaction times and error percentages indicatingthat in this condition the selection of the correctresponse finger tended to be a source of interference.The selection might be impaired by the instance thatthe proximal movement of the left (right) finger ispresented as a movement of the right (left) fingeron the projection screen. But also movement timeswere substantially higher than those observed inExperiment 1. Obviously, this inversion of the
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1765
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

left–right dimension hampered both action plan-ning and action control.
The third finding was the presence of the later-ality effect in the condition with left–rightreversed feedback. Remember, different S–Rmappings compete in this condition, and compat-ibility relationships between distal S–R mappingsare contrary to proximal–distal S–R mappings.However, again ipsilateral movements were fasterand more accurate than contralateral movements.The distal compatibility effect was even pro-nounced in the present experiment especiallywhen taking into account the differences in errorpercentages.
EXPERIMENT 3
In this experiment the egocentric perspectivefrom the top (basic condition in the experiments,Figure 2a, bottom left) was compared with thenonegocentric perspective of anatomical mirror-ing—that is, with the perspective at looking atsomeone else’s hand copying the same action byanatomic imitation (Figure 2a, upper right). Thisperspective resulted from point-reflection—thatis, the reflection of the original hand movementabout both axes.
As in Experiment 2, the left–right relationswere inversed. For instance, when the right LEDflashed green on the projection screen, participantsmoved the proximal left hand ipsilaterally, but themovement was presented on the right side of theprojection screen and with a nonegocentric view.
Method
ParticipantsA total of 8 fresh students (5 female) of theRWTH Aachen University, aged between 20and 44 years (mean age 28 years) participated inthe experiment.
Stimuli, procedure, and designThese were essentially the same as those inExperiment 1. The basic condition was now com-bined with the nonegocentric condition in which
the perspective is reflected about both the x- andy-axes (Figure 2a, upper right).
Results and discussion
The ANOVA of reaction times revealed signifi-cant main effects of the factors perspective,F(1, 7) ¼ 28.13, MSE ¼ 2,271.44, p , .01, andlaterality, F(1, 7) ¼ 9.56, MSE ¼ 1,038.95, p ,
.05. Reaction times were 89 ms faster in the ego-centric than in the nonegocentric condition (682vs. 771 ms), and they were 35 ms faster for ipsilat-eral than contralateral movements (709 vs. 744 ms;cf. Figure 3, bottom). However, the laterality effectwas based exclusively on a reaction-time differenceof 67 ms in the egocentric condition and was notobserved in the nonegocentric condition yieldinga significant interaction between the factors,F(1, 7) ¼ 76.46, MSE ¼ 102.85, p , .01.
The ANOVA of movement times revealed asignificance of the factor laterality, F(1, 7) ¼33.07, MSE ¼ 1,011.90, p , .01, and a marginalsignificance of the factor perspective, F(1, 7) ¼4.37, MSE ¼ 16,850.59, p ¼ .07, but no inter-action (p . .54). Ipsilateral movements were exe-cuted 64 ms faster than contralateral movements(360 vs. 424 ms), and mean movement timeswere 344 ms and 440 ms in the egocentric andnonegocentric conditions.
The ANOVA of the error percentages revealeda significant main effect of the factor perspectivewith F(1, 7) ¼ 21.76, MSE ¼ 106.21, p , .01.Laterality was marginally significant with F(1, 7)¼ 4.45, MSE ¼ 13.57, p ¼ .07, but the inter-action was not (p . .15). In the egocentriccondition participants responded with a meanerror percentage of 4.3% (4.2% in the ipsilateralcondition and 4.3% in the contralateral condition).Errors increased drastically to 21.3% in the none-gocentric condition. Here, mean error percentageswere 18.6% and 24.0% for ipsilateral and contral-ateral movements.
In sum, the present experiment replicatedsuccessfully the previous experiments in twoaspects. First, the pattern of results in the basic con-dition corresponded with those of Experiments 1and 2. Second, performance drastically decreased
1766 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

with a reversion of the left–right relationshipbetween stimuli and responses (in Experiment 2with reflection about the y-axis and in Experiment3 with reflection about both axes). The thirdfinding was the absence of the laterality effect inthe condition with anatomical mirroring. It mightbe that the laterality effect was cancelled out bythe left–right inversion of the response finger orsimply by a ceiling effect caused by the left–rightinversion. Another possibility was that the absenceof the laterality effect originated from changedcompatibility relationships. We discuss thisfinding in detail in the following section.
GENERAL DISCUSSION
The present study examined whether and how theperspective of seeing one’s own movements exertedan influence on action control. As outlined in theintroduction, changes in the perspective are oftenrequired when behaviour is imitated. But alsomodern technical environments often introducechanges in perspective, especially when the useracts by technical equipment in a distant space—for instance, in laparoscopic surgery, teleoperation,or virtual reality. When taking the viewpoint of arobotic avatar with a rotated sight (e.g., invirtual-reality applications), the actor has toaccount for the perspective, which sometimeseven requires incompatible responding (e.g., in aface-to-face perspective with an avatar, movingthe left hand means that from the viewpoint ofthe observer, the avatar moves its right hand).From a psychological point of view, these situ-ations are challenging as changes in the perspectivelead to discrepancies between the (anticipated)proximal tactile/proprioceptive feedback and thedistal visual feedback on a projection screen. Thequestion is whether and how an actor is able tointegrate the different sources of information.
In Experiment 1 the egocentric perspectivewith a view from the top (basic condition) wascompared with a perspective similar to seeingone’s own actions in a mirror (x-axis reflection).Although in the latter condition upward move-ments were transformed in seen downward
movements (nonegocentric view but remainingleft–right relations), participants had no difficul-ties in performing the task, and movementexecution was as fast and accurate as with theview from top. However, ipsilateral movementswere performed faster and more accurate than con-tralateral movements. This finding is not unusualas ipsilateral movements were marked byapproaching the stimuli and contralateral move-ments by avoiding them (e.g., Lavender &Hommel, 2007; Musseler et al., 2009;Yamaguchi & Proctor, 2006).
In Experiments 2 and 3 performances substan-tially decreased when visual feedback was reflectedabout the y-axis (egocentric view plus left–rightinversion) and about both the x- and y-axes (none-gocentric view plus left–right inversion). Bothconditions were marked by reversed left–rightrelations between stimuli and hand movementson the projection screen and the actual hand move-ments. Obviously, action control became muchmore difficult when the perspective comprehendedreversed left–right relations, and it can be assumedthat the reversion reduced the similarity betweenstimuli and movements and thus hampered per-formance (cf. Hommel et al., 2001). We can sum-marize that the human preference to imitateactions of others by mirroring is also observedwhen seeing mirror images of one’s own move-ments. If visual feedback inverted left–rightrelations the same difficulties occurred as thosethat can be observed with anatomic imitation.
Differences between ipsilateral and contralat-eral movements appeared in Experiment 2, too,but not in Experiment 3. To shed light on theabsence of the laterality effect in Experiment 3,it might be worth analysing the compatibilityrelationships between distal stimulus positionsand proximal/distal hand start and end positionsin detail (Figure 2b). At least two explanationsare possible. First, the compatibility relationshipbetween stimulus position and proximal handstart position accounts for the disappearance ofthe laterality effect. This relationship was incom-patible for both ipsilateral and contralateral move-ments. In other words, performance was equallybad in both conditions.
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1767
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

Second, it is also possible that in Experiment 3two tendencies in the opposite direction cancelledeach other out. The compatibility relationshipbetween stimulus position and distal hand end pos-ition was compatible for the ipsilateral condition,but incompatible for the contralateral condition,and, vice versa, the compatibility relationshipbetween stimulus position and proximal hand endposition was incompatible for the ipsilateralcondition, but compatible for the contralateral con-dition. If these relationships were equally importantfor generating the response, no differences betweenconditions should be observed in Experiment 3, andthis is what the results showed.
There is another argument favouring thesecond explanation. For example, when the redLED flashed at the right side, the observers’ lefthand on the fibreboard moved to the right targetbutton. As a consequence, the perceived positionof the LED corresponded with the proximalhand end position. Thus, the contralateral move-ment was not marked any more by avoiding thestimulus; it was rather marked as an approach be-haviour at the proximal plane cancelling out anydifferences between ipsilateral and contralateralmovements.
The main difference between the explanationsis that the first explanation emphasizes the proxi-mal hand start position, while the second expla-nation is goal oriented—that is, the proximaland/or distal hand end position determined theresponse. Theoretical and empirical evidencefavours the latter explanation (e.g., Bekkeringet al., 2000; Hommel, 1993; Kunde et al., 2007;Musseler et al., 2008). However, what remains tobe explained is, why the ipsilateral/contralateraldifference was present in all experiments exceptin Experiment 3. Most likely is that the actor pre-ferably rely on the distal information of the visualsystem, as she or he is sometimes not very aware ofher or his own actions at a proximal level (e.g.,Knoblich & Kircher, 2004; Mechsner, Kerzel,Knoblich, & Prinz, 2001; Musseler & Sutter,2009; Sulzenbruck & Heuer, 2009). But thisdoes not exclude reference to the proximal infor-mation when it facilitates action execution(Wohlschlager, Gattis, & Bekkering, 2003). A
recent study (Longo & Haggard, 2009) suggeststhat vision of one’s own hand movements, andspecifically the sense of agency over that hand,primes motor responses as well. Participantsperformed either active or passive hand move-ments, which they saw with either real-time ordelayed video feedback. Participants respondedfastest with real-time video feedback of one’sown active hand movements. This facilitationeffect corresponded with the sense of agency overthe seen hand. Results of the present experimentscould also be affected by representations of thebodily self. This should be addressed in futureexperiments.
Original manuscript received 20 July 2009
Accepted revision received 26 October 2009
First published online 19 February 2010
REFERENCES
Bekkering, H., Wohlschlager, A., & Gattis, M. (2000).Imitation of gestures in children is goal-directed.Quarterly Journal of Experimental Psychology A, 53,
153–164.Bertenthal, B. I., Longo, M. R., & Kosobud, A. (2006).
Imitative response tendencies following observationof intransitive actions. Journal of Experimental
Psychology: Human Perception and Performance, 32,
210–225.Brainard, D. H. (1997). The psychophysics toolbox.
Spatial Vision, 10, 433–436.Brass, M., Bekkering, H., Wohlschlager, A., & Prinz,
W. (2000). Compatibility between observed and exe-cuted finger movements: Comparing symbolic,spatial, and imitative cues. Brain and Cognition, 44,
124–143.Bruzzo, A., Borghi, A. M., & Ghirlanda, S. (2008).
Hand–object interaction in perspective.Neuroscience Letters, 441, 61–65.
Buccino, G., Vogt, S., Ritzl, A., Fink, G. R., Zilles,K., Freund, H.-J., et al. (2004). Neural circuitsunderlying imitation learning of hand actions:An event-related fMRI study. Neuron, 42,
323–334.Craighero, L., Bello, A., Fadiga, L., & Rizzolatti, G.
(2002). Hand action preparation influences the
1768 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9)
SUTTER AND MUSSELER
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010

responses to hand pictures. Neuropsychologia, 40,
492–502.Franck, N., Farrer, C., Georgieff, N., Marie-Cardine, M.,
Dalery, J., d’Amato, T., et al. (2001). Defective recog-nition of one’s own actions in patients with schizo-phrenia. American Journal of Psychiatry, 158, 454–459.
Greenwald, A. G. (1970). Sensory feedback mechanismsin performance control: With special reference to theideo-motor mechanism. Psychological Review, 77,
73–99.Hommel, B. (1993). Inverting the Simon effect by
intention. Psychological Research, 55, 270–279.Hommel, B., Musseler, J., Aschersleben, G., & Prinz,
W. (2001). The theory of event coding (TEC): Aframework for perception and action. Behavioral
and Brain Sciences, 24, 869–937.Iacoboni, M., & Dapretto, M. (2006). The mirror
neuron system and the consequence of its dysfunc-tion. Nature Reviews Neuroscience, 7, 942–951.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H.,Mazziotta, J. C., & Rizzolatti, G. (1999). Corticalmechanisms of human imitation. Science, 286,
2526–2528.James, W. (1890). The principles of psychology. New York:
Dover.Knoblich, G., & Kircher, T. T. J. (2004). Deceiving
oneself about being in control: Conscious detectionof changes in visuomotor coupling. Journal of
Experimental Psychology: Human Perception and
Performance, 30, 657–666.Koski, L., Iacoboni, M., Dubeau, M.-C., Woods, R. P.,
& Mazziotta, J. C. (2003). Modulation of corticalactivity during different imitative behaviors. Journal
of Neurophysiology, 89, 460–471.Kunde, W., Musseler, J., & Heuer, H. (2007). Spatial
compatibility effects with tool use. Human Factors,49, 661–670.
Lavender, T., & Hommel, B. (2007). Affect and action:Towards an event-coding account. Cognition &
Emotion, 21, 1270–1296.Longo, M. R., & Haggard, P. (2009). Sense of agency
primes manual motor responses. Perception, 38, 69–78.Massen, C., & Prinz, W. (2007). Programming tool-use
actions. Journal of Experimental Psychology: Human
Perception and Performance, 33, 692–704.Mechsner, F., Kerzel, D., Knoblich, G., & Prinz, W.
(2001). Perceptual basis of bimanual coordination.Nature, 414, 69–72.
Musseler, J. (1999). How independent from actioncontrol is perception? An event-coding account formore equally-ranked crosstalks. InG. Aschersleben, T. Bachmann, & J. Musseler(Eds.), Advances in psychology: Vol. 129. Cognitive
contributions to the perception of spatial and temporal
events (pp. 121–147). Amsterdam: Elsevier.Musseler, J., Aschersleben, G., Arning, K., & Proctor,
R. W. (2009). Reversed effects of spatial compatibil-ity in natural scenes. American Journal of Psychology,122, 325–336.
Musseler, J., Kunde, W., Gausepohl, D., & Heuer, H.(2008). Does a tool eliminate spatial compatibilityeffects? European Journal of Cognitive Psychology, 20,
211–231.Musseler, J., & Sutter, C. (2009). Perceiving one’s own
movements when using a tool. Consciousness and
Cognition, 18, 359–365.Nattkemper, D., & Ziessler, M. (2004). Cognitive
control of action: The role of action effects.Psychological Research, 68, 71–73.
Pelli, D. G. (1997). The videotoolbox software for visualpsychophysics: Transforming numbers into movies.Spatial Vision, 10, 437–442.
Proctor, R. W., & Vu, K.-P.L. (2006). Stimulus–
response-compatibility principles: Data, theory, and
application. New York: CRC Press, Taylor &Francis Group.
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27,
169–192.Sulzenbruck, S., & Heuer, H. (2009). Functional
independence of explicit and implicit motoradjustments. Consciousness and Cognition, 18,
145–159.Wapner, S., & Cirillo, L. (1968). Imitation of a
model’s hand movements: Age changes in transpo-sition of left–right relations. Child Development, 39,
887–894.Wohlschlager, A., Gattis, M., & Bekkering, H. (2003).
Action generation and action perception inimitation: An instance of the ideomotor principle.Philosophical Transactions of the Royal Society London
B, 358, 501–515. doi: 10.1098/rstb.2002.1257.Yamaguchi, M., & Proctor, R. W. (2006). Stimulus–
response compatibility with pure and mixedmappings in a flight task environment. Journal of
Experimental Psychology: Applied, 12, 207–222.
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2010, 63 (9) 1769
PERSPECTIVE OF SEEING ONE’S OWN MOVEMENT
Downloaded By: [Sutter, C.] At: 10:23 27 September 2010
Copyright © 2022 FDOKUMEN




















