Abdulrazzak et al. - 2010 - Biological characteristics of stem cells from foetal, cord blood and...
19
doi: 10.1098/rsif.2010.0347.focus published online 25 August 2010 J. R. Soc. Interface Hassan Abdulrazzak, Dafni Moschidou, Gemma Jones and Pascale V. Guillot and extraembryonic tissues Biological characteristics of stem cells from foetal, cord blood References html#ref-list-1 http://rsif.royalsocietypublishing.org/content/early/2010/08/24/rsif.2010.0347.focus.full. This article cites 205 articles, 39 of which can be accessed free P<P Published online 25 August 2010 in advance of the print journal. Rapid response http://rsif.royalsocietypublishing.org/letters/submit/royinterface;rsif.2010.0347.focusv1 Respond to this article Subject collections (188 articles) biomedical engineering (85 articles) bioengineering Articles on similar topics can be found in the following collections Email alerting service here right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top publication. Citations to Advance online articles must include the digital object identifier (DOIs) and date of initial online articles are citable and establish publication priority; they are indexed by PubMed from initial publication. the paper journal (edited, typeset versions may be posted when available prior to final publication). Advance Advance online articles have been peer reviewed and accepted for publication but have not yet appeared in http://rsif.royalsocietypublishing.org/subscriptions go to: J. R. Soc. Interface To subscribe to This journal is © 2010 The Royal Society on August 28, 2010 rsif.royalsocietypublishing.org Downloaded from
Transcript of Abdulrazzak et al. - 2010 - Biological characteristics of stem cells from foetal, cord blood and...

doi: 10.1098/rsif.2010.0347.focus published online 25 August 2010J. R. Soc. Interface
Hassan Abdulrazzak, Dafni Moschidou, Gemma Jones and Pascale V. Guillot and extraembryonic tissuesBiological characteristics of stem cells from foetal, cord blood
Referenceshtml#ref-list-1http://rsif.royalsocietypublishing.org/content/early/2010/08/24/rsif.2010.0347.focus.full.
This article cites 205 articles, 39 of which can be accessed free
P<P Published online 25 August 2010 in advance of the print journal.
Rapid responsehttp://rsif.royalsocietypublishing.org/letters/submit/royinterface;rsif.2010.0347.focusv1
Respond to this article
Subject collections
(188 articles)biomedical engineering � (85 articles)bioengineering �
Articles on similar topics can be found in the following collections
Email alerting service hereright-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top
publication. Citations to Advance online articles must include the digital object identifier (DOIs) and date of initial online articles are citable and establish publication priority; they are indexed by PubMed from initial publication.the paper journal (edited, typeset versions may be posted when available prior to final publication). Advance Advance online articles have been peer reviewed and accepted for publication but have not yet appeared in
http://rsif.royalsocietypublishing.org/subscriptions go to: J. R. Soc. InterfaceTo subscribe to
This journal is © 2010 The Royal Society
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from

J. R. Soc. Interface
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
doi:10.1098/rsif.2010.0347.focusPublished online
REVIEW
*Author for c
One contribcommercializa
Received 30 JAccepted 5 A
Biological characteristics of stem cellsfrom foetal, cord blood and
extraembryonic tissuesHassan Abdulrazzak, Dafni Moschidou, Gemma Jones
and Pascale V. Guillot*
Institute of Reproductive and Developmental Biology, Imperial College London,London W12 0NN, UK
Foetal stem cells (FSCs) can be isolated during gestation from many different tissues such asblood, liver and bone marrow as well as from a variety of extraembryonic tissues such asamniotic fluid and placenta. Strong evidence suggests that these cells differ on many biolo-gical aspects such as growth kinetics, morphology, immunophenotype, differentiationpotential and engraftment capacity in vivo. Despite these differences, FSCs appear to bemore primitive and have greater multi-potentiality than their adult counterparts. Forexample, foetal blood haemopoietic stem cells proliferate more rapidly than those found incord blood or adult bone marrow. These features have led to FSCs being investigated forpre- and post-natal cell therapy and regenerative medicine applications. The cells havebeen used in pre-clinical studies to treat a wide range of diseases such as skeletal dysplasia,diaphragmatic hernia and respiratory failure, white matter damage, renal pathologies aswell as cancers. Their intermediate state between adult and embryonic stem cells alsomakes them an ideal candidate for reprogramming to the pluripotent status.
Keywords: stem cell; foetal; regenerative medicine; induced pluripotent stem
1. INTRODUCTION
Stem cells are undifferentiated cells with the capacity toself-renew, differentiate and repopulate a host in vivo(Weissman et al. 2001). Their plasticity or potency ishierarchical ranging from totipotent (differentiatinginto all cell types including placenta), pluripotent (dif-ferentiating into cells of the three germ layers,ectoderm, mesoderm and endoderm, but not tropho-blastic cells), multipotent (differentiating into cells ofmore than one type but not necessarily into all thecells of a given germ layer) to unipotent (differentiatinginto one type of cell only, e.g. muscle or neuron).
Pluripotent stem cells can be derived from the innercell mass of the pre-implantation embryo (i.e. embryo-nic stem (ES) cells) or isolated from the foetalprimordial germ cell pool (PGC) above the allantois(i.e. embryonic germ (EG) cells and embryonic carci-noma (EC) cells; Thomson et al. 1998; Andrews et al.2005). The destruction of the blastocyst or earlyfoetus necessary for their derivation/isolation raisesethical concerns (Lo & Parham 2009), although recent
orrespondence ([email protected]).
ution to a Theme Supplement ‘Translation andtion of regenerative medicines’.
une 2010ugust 2010 1
work has shown that ES cells can be derived fromsingle blastomeres isolated using procedures similar tothose routinely used for pre-implantation genetic diag-nosis (Klimanskaya et al. 2007). However, safetyconcerns still remain because of the tumorigenicity ofES cells. Alternatively, adult stem cells can be foundin almost all tissues examined including brain, dentalpulp, muscle, bone marrow, skin and pancreas andhave been extensively characterized for their thera-peutic potential. The adult stem cell could be multi-potent (e.g. haematopoietic stem cells (HSCs) givingrise to all blood cells and adherent stromal/mesenchy-mal stem cells (MSCs) that give rise to bone, fat,cartilage and muscle) or unipotent (e.g. progenitorcells). Adult MSCs have the problem of being difficultto extract in sufficient numbers for therapy and/or pre-senting restricted plasticity and limited proliferativecapacity compared with ES cells.
In recent years, foetal stem cells (FSCs) and stemcells isolated from cord blood or extraembryonic tissueshave emerged as a potential ‘half way house’ betweenES cells and adult stem cells. FSCs can be foundin foetal tissues such as blood, liver, bone marrow,pancreas, spleen and kidney, and stem cells are alsofound in cord blood and extraembryonic tissues suchas amniotic fluid, placenta and amnion (Marcus &
This journal is # 2010 The Royal Society

2 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Woodbury 2008). Their primitive properties, expansionpotential and lack of tumorogenicity make them anattractive option for regenerative medicine in celltherapy and tissue engineering settings. While extraem-bryonic tissues could be used with few ethicalreservations, the isolation of FSCs from abortuses issubject to significant public unease. We review herethe phenotypic characteristics of stem cells fromfoetal, cord blood and extraembryonic tissues, theirapplication in cell therapy and their potential for repro-gramming towards pluripotentiality.
2. PHENOTYPIC CHARACTERISTICS OFFOETAL STEM CELLS
Stem cells are collected from abortal foetal tissue,surplus of pre-natal diagnostic tissues or tissues atdelivery, subject to informed consent, institutionalethics approval and compliance with national guidelinescovering foetal tissue research. Until recently theywere thought to be multi-potent (O’Donoghue & Fisk2004), but this picture is now changing as evidence ismounting regarding the existence of pluripotent sub-populations in some foetal and extraembryonic tissues(Cananzi et al. 2009). Some of the properties of thecells are summarized in table 1.
2.1. Foetal tissues
2.1.1. Foetal MSCs. MSCs are multi-potent stem cellsthat can differentiate towards mesoderm-derivedlineages (i.e. osteogenic, adipogenic, chondrogenic andmyogenic; Pittenger et al. 1999). Their potential torepair damaged tissue and their immunomodulatoryproperties make them very attractive for a wide rangeof regenerative medicine applications such as celltherapy and tissue engineering for hollow and solidorgans (Horwitz et al. 2002; Abdallah & Kassem2008; Le Blanc et al. 2008). They were first identifiedin adult bone marrow where they represent 0.001–0.01% of total nucleated cells (Owen & Friedenstein1988). Adherent stromal cells with similar characteristicswere subsequently isolated from other tissues such as adi-pose, dental pulp, muscle, liver and brain (Porada et al.2006; Huang et al. 2009). They are also found in foetaland extraembryonic tissues, where they seem to possessgreater proliferation capacity and differentiation poten-tial than their adult counterparts (Campagnoli et al.2001; Guillot et al. 2007b; Pappa & Anagnou 2009).
2.1.1.1. Common characteristics. MSCs isolated fromfoetal tissues such as blood, liver, bone marrow, lungand pancreas all share common characteristics. Forexample, they are spindle-shaped cells with the capacityto differentiate into the standard mesenchymallineages, i.e. bone, fat and cartilage. They do notexpress haematopoietic or endothelial markers (e.g.they are CD452/342/142 and von Willebrand factornegative; Gucciardo et al. 2009), but express stroma-associated markers CD29 (b1-integrin), CD73 (SH3and SH4), CD105 (SH2), CD44 (HCAM1), the earlybone marrow progenitor cell marker CD90 (thy-1) andthe extracellular matrix proteins vimentin, laminin and
J. R. Soc. Interface
fibronectin (Guillot et al. 2006, 2007a). Contrary toadult bone marrow MSCs, first-trimester foetal blood,liver and bone marrow MSCs express baseline levels ofthe pluripotency stem cell markers Oct-4, Nanog, Rex-1,SSEA-3, SSEA-4, Tra-1-60 and Tra-1-81 (Guillot et al.2007b; Zhang et al. 2009). Regardless of their tissue oforigin, first-trimester foetal MSCs self-renew faster in cul-ture than adult MSCs (30–35 h versus 80–100 h) andsenesce later while retaining a stable phenotype (Guillotet al. 2007b). Foetal MSCs are therefore more readilyexpandable to therapeutic scales for either pre- or post-natal ex vivo gene or cell therapy and tissue engineering.Having greater multi-potentiality and differentiatingmore readily than adult MSCs into cells of mesodermalorigin such as bone and muscle, they can also differentiateinto cells from other lineages such as oligodendrocytesand haematopoietic cells (MacKenzie et al. 2001; Chanet al. 2006; Kennea et al. 2009). In terms of engraftment,they have a competitive advantage compared with adultcells (Rebel et al. 1996; Harrison et al. 1997; Taylor et al.2002). Other advantageous characteristics relevant to celltherapy include having active telomerase (Guillot et al.2007b), expressing low levels of HLA I and lacking intra-cellular HLA II, and taking longer to express this uponinterferon g stimulation than adult MSCs (Gotherstromet al. 2004). Foetal MSCs also express a shared a2, a4and a5b1 integrin profile with first-trimester HSCs, impli-cating them in homing and engraftment, and, consistentwith this, they have significantly greater binding totheir respective extracellular matrix ligands than adultMSCs (de la Fuente et al. 2003). Finally, human foetalMSCs are readily transducible with transduction efficien-cies of greater than 95 per cent using lentiviral vectorswith stable gene expression at both short- and long-timepoints without affecting self-renewal or multi-potentiality(Chan et al. 2005).
2.1.1.2. Specific characteristics. Although the abovecharacteristics are shared among the various types offoetal stem, they also have specific traits that are depen-dent on tissue of origin and gestational age. For example,foetal MSCs are present in foetal blood from the earliestgestation tested, seven weeks, where they account forapproximately 0.4 per cent of foetal nucleated cells.Their numbers decline to very low levels after 13weeks’ gestation (Campagnoli et al. 2001). MSC derivedfrom foetal bone marrow also decline with age with oneMSC found among 10 000 in mid-trimester as comparedwith one MSC per 250 000 cells in adult marrow.
Significant variations have been found among foetaltissue cells in terms of cell surface markers. Forexample, foetal lung has a higher proportion ofCD34þ/CD452 cells (44%) than bone marrow (4.8%),spleen (12.6%) and liver (7.5%) (in’t Anker et al.2003a,b). Also foetal metanephric MSCs do not expressCD45, CD34 or other haematopoietic markers; instead,they express high levels of mesenchymal markers suchas vimentin, laminin and type I collagen.
There is also variation among the cells in terms ofdifferentiation capacity. MSCs derived from foetalliver during the first and second trimester have areduced osteogenic differentiation potential in

Tab
le1.
Cha
ract
eristi
csof
stem
cells
.E
,em
bryo
nic;
MSC
,m
esen
chym
al;
H,
haem
atop
oiet
ic;
EP,
epit
helia
l;V
SEL,
very
smal
lem
bryo
nic
like;
P,
plur
ipot
ent;
M,
mul
tipo
tent
;U
,un
ipot
ent;
n.d.
,no
tde
term
ined
.A
ster
isk
deno
tes
that
the
char
acte
rist
ics
ofth
ese
cells
are
not
liste
din
the
tabl
e(K
yoet
al.19
97;C
ampa
gnol
iet
al.20
01;G
amm
aito
niet
al.20
04;M
iki
etal
.20
05,20
07;P
rank
eet
al.20
05;D
anet
al.20
06;H
abic
het
al.20
06;M
iki&
Stro
m20
06;P
ortm
ann-
Lan
zet
al.2
006;
Zha
oet
al.2
006;
De
Cop
piet
al.20
07a;
Fon
get
al.20
07;G
uillo
tet
al.
2007
b;Ilan
cher
anet
al.
2007
;K
arah
usey
inog
luet
al.
2007
;K
imet
al.
2007
;Li
etal
.20
07;
Mus
ina
etal
.20
07;
Pra
t-V
idal
etal
.20
07;
Deg
isti
rici
etal
.20
08;
Moo
net
al.
2008
;M
ount
ford
2008
;P
olon
iet
al.20
08;Se
ssar
ego
etal
.20
08;T
roye
r&
Wei
ss20
08;B
rook
eet
al.20
09;C
anan
ziet
al.20
09;R
ieks
tina
etal
.20
09;Zha
nget
al.20
09;A
lam
inos
etal
.20
10;
Anz
alon
eet
al.2
010;
Cas
trec
hini
etal
.201
0;H
sieh
etal
.20
10;I
nsau
stiet
al.2
010;
Vaz
iriet
al.20
10).
embr
yoni
cst
emce
lls
foet
alti
ssue
sex
trae
mbr
yoni
c
adul
tB
MM
SCbl
ood
liver
bone
mar
row
cord
bloo
dW
hart
on’s
jelly
amni
otic
fluid
amni
on-d
eriv
edep
ithe
lialc
ells
amni
on-
deri
ved
MSC
chor
ion-
deri
ved
MSC
cell
phen
otyp
e(s
)E
MSC
/H*
MSC
/H*
MSC
/H*
MSC
/H*
MSC
MSC
/A
FS/
VSE
L*
EP
MSC
MSC
MSC
doub
ling
tim
e(D
T)/
popu
lati
ondo
ublin
gs(P
D)
48–24
h(D
T)/
inde
finit
e(P
D)
21.9+
1.1
h(fi
rst-
trim
este
rM
SC)
(DT
)/28
.4a
(PD
)
46h
(sec
ond-
trim
este
rM
SC)
(DT
)
32h
(sec
ond-
trim
este
rM
SC)
(DT
)
39–43
h(D
T)/
20b
(PD
)
85–11
h(D
T)
25–38
h(s
econ
d-tr
imes
ter
MSC
)36
h(A
FS)
(DT
)/66
(MSC
)/ov
er25
0A
FS
(PD
)
38.4
h(D
T)
30c(P
D)
21.2
5+
3.2
h(D
T)/
30d
54–11
1h
(DT
)/7.
1a(P
D)
feed
er/m
atri
gel
requ
irem
ent
þ2
22
22
22
22
2
pote
ncy
PP
/MP
/MP
/MP
/MP
/MP
/M
P/M
P/M
P/M
MO
ct-4
þþþ
þþ
þþ
þþ
þþ
þþ
þ+
Sox2
þþþ
n.d.
n.d.
n.d.
þ2
þn.
d.n.
d.+
Nan
ogþþþ
þþ
þþ
þþ
þþ
+þ
n.d.
þ+
c-M
ycþþþ
n.d.
n.d.
n.d.
n.d.
n.d.
þþ
n.d.
n.d.
n.d.
Klf-4
þþþ
n.d.
n.d.
n.d.
n.d.
n.d.
n.d.
n.d.
n.d.
n.d.
n.d.
TR
Fle
ngth
/tel
omer
ase
acti
vity
þþþ
þþ
þþ
þþ
þþ
þn.
d.2
n.d.
þ2
alka
line
phos
phat
ase
þþþ
n.d.
n.d.
þþ
þþ
þ2
n.d.
+c-
Kit
(CD
117)
þþþ
þþ
þ+
þþ
++
22
Rex
-1þþþ
þþ
þþþ
þn.
d.þ
þ2
þ+
SSE
A-4
þþþ
þþ
þþ
þþ
þþ
þþ
2þ
2
SSE
A-3
þþþ
þþ
þþ
þþ
þ2
2þ
(low
)2
þ2
Tra
-1-8
1þþþ
þþ
þþ
þþ
n.d.
þ2
þ2
þ2
Tra
-1-6
1þþþ
þþ
þþ
þþ
n.d.
þ+
þ2
þ2
MH
Ccl
ass
Iþ
(low
)þ
(low
)þ
(low
)þ
(low
)þ
þþ
þ(l
ow)
þ(l
ow)
þþ
MH
Ccl
ass
II(H
LA
-DR
)2
22
2+
2+
22
22
tera
tom
afo
rmat
ion
inim
mun
eco
mpr
omis
edm
ice
yes
n.d.
n.d.
n.d.
n.d.
n.d.
nono
n.d.
n.d.
no
a Ove
ra
peri
odof
50da
ys.
bO
ver
eigh
tpa
ssag
es.
c Ove
r14
pass
ages
.dO
ver
five
pass
ages
.
Review. Characteristics of stem cells H. Abdulrazzak et al. 3
J. R. Soc. Interface
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from

4 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
comparison with first-trimester foetal blood and withsecond-trimester spleen, lung and bone marrow MSCs,possibly because of a reduction in osteogenic progenitornumbers (in‘t Anker et al. 2003a,b). Foetal pancreaticMSCs, shown to be capable of differentiating into chon-drogenic, osteogenic or adipogenic lineages (Hu et al.2003), can also engraft, differentiate and secretehuman insulin in a sheep model (Ersek et al. 2010).Moreover, foetal pancreatic ductal stem cells candifferentiate into insulin-producing cells in vitro (Yaoet al. 2004).
Foetal metanephric MSCs can be induced in vitro toadopt an osteogenic or myogenic phenotype and, whentransplanted in a foetal lamb model, they show long-term persistence and site-specific differentiation(Almeida-Porada et al. 2002).
Altogether these data indicate a high level ofheterogeneity within the foetal stem cell pool.
2.1.2. Foetal HSCs. HSCs are multi-potent stem cellsthat maintain functional haematopoiesis by generationof all haematopoietic lineages throughout foetal andadult life (Weissman & Shizuru 2008). They are charac-terized by the expression of CD34 and CD45 antigens,and the absence of markers such as CD38 and humanleucocyte antigen (HLA)/DRE (Huss 2000; Tayloret al. 2002). During ontogeny the site of haematopoiesisis modified several times (Cumano et al. 2001). An areaalong the dorsal embryonic aorta termed the aorta–gonad–mesonephros (AGM) is a rich source of HSCs,which then migrate to the embryonic liver and laterseed other haematopoietic tissues such as the bonemarrow (McGrath & Palis 2008). HSCs are usuallyassayed in animal models based on their capacity torepopulate the entire haematopoietic system in con-ditioned recipients after transplantation. Similar toMSCs, it is possible to transplant human cells into xeno-geneic recipients such as foetal sheep that act as modelsof human haematopoiesis (O’Donoghue & Fisk 2004).
First-trimester foetal blood contains more CD34þ
cells than term gestation blood, yet CD34þ cellsaccount for only 5 per cent of CD45þ cells in theblood at that stage (Campagnoli et al. 2001). Thenumber of circulating HSCs increases from the first tri-mester to peak in the second trimester in utero,probably because of cells migrating from the foetalliver to establish haematopoiesis in the foetal bonemarrow (Clapp et al. 1995). Some HSCs remain in theumbilical cord at delivery, where they can be collectedfor allogeneic or occasionally autologous cell transplan-tation (see §2.2.1.).
In the second trimester, CD34þ cells constitute 4 percent of cells in blood, 16.5 per cent in bone marrow,6 per cent in liver, 5 per cent in spleen and 1.1 percent in the thymus (Lim et al. 2005). This frequencyof CD34þ cells in the blood gradually diminishesduring the third trimester, probably reflecting establish-ment of the marrow as the primary site ofhaematopoiesis and the declining role of the foetalliver in that regard (Jones et al. 1994; Wagers et al.2002). The number of cells negative for CD38 withinthe CD34þ population is also higher in early foetal
J. R. Soc. Interface
blood, suggesting that these cells are more primitiveand have greater potential than HSCs circulating laterin ontogeny (O’Donoghue & Fisk 2004). Even relativelydifferentiated erythroblasts are more primitive in firstcompared with later trimester foetal blood. Foetalblood HSCs proliferate more rapidly than those incord blood or adult bone marrow, and produce allhaematopoietic lineages (Campagnoli et al. 2001).
Foetal liver is also a source of HSCs, and, despitetheir relatively small number, it is possible to generatesufficient numbers of cells for transplantation in fourweeks (Rollini et al. 2004), although, to our knowledge,it has not been proved that foetal liver HSCs can suc-cessfully transplant a human recipient. The phenotypeof foetal liver HSCs changes with gestation age. Forexample, their differentiation potential decreases mark-edly with gestational age, and cells that expressed bothCD34 and c-kit (CD117) were identified in the first-trimester foetal liver whereas the cells expressing morecommitted hepatic markers only appeared during thesecond trimester (Nava et al. 2005).
2.2. Extraembryonic tissue
2.2.1. Umbilical cord blood. Umbilical cord blood is nowan established source of transplantable HSCs that havea greater proliferative capacity, lower immunologicalreactivity and lower risk of graft-versus-host disease(GVHD) than those derived from adult bone marrow(Broxmeyer et al. 1989; Yu et al. 2001; Ballen 2005;Schoemans et al. 2006; Brunstein et al. 2007; Hwanget al. 2007; Broxmeyer 2010). These cells are capableof repopulating bone following intra-bone injection ofsevere combined immunodeficiency mice (Mazurieret al. 2003; Wang et al. 2003) and are used clinicallyas an alternative to adult bone marrow HSCs in somecases (Delaney et al. 2010). The cells are CD34þ andCD382 and their frequency is greater than that ofbone marrow or peripheral blood following cytokinemobilization (Pappa & Anagnou 2009). It has alsobeen demonstrated that cord blood contains MSCs(Weiss & Troyer 2006; Secco et al. 2008) that can sup-port the in vivo expansion of HSCs and function as anaccessory cell population for engraftment (Javazonet al. 2004; Wang, J. et al. 2004). The frequency ofMSCs in umbilical cord blood is low however, withMSCs successfully isolated from only a third of allsamples collected (Bieback et al. 2004).
Cord blood stem cells expressing baseline levels of EScell markers such as Oct-4, Nanog, SSEA-3 and SSEA-4have also been described (Zhao et al. 2006). Using atwo-stage isolation approach whereby the erythrocytesin cord blood are lysed and the remaining cells aresorted using flow cytometry, it has been possible to iso-late and enrich for a subpopulation of cells that areCXCR4þ, CD133þ, CD34þ, Lin2 and CD452.
These cells have been described as very smallembryonic-like (VSEL) stem cells because they areonly 3–5 mm in diameter and express Oct-4, Nanogand SSEA-4 (Kucia et al. 2007; Zuba-Surma et al.2010). Compared with MSCs, VSEL cells are smaller,have a large nucleus to cytoplasm ratio and open-typechromatin. As a similar population has also been

Review. Characteristics of stem cells H. Abdulrazzak et al. 5
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
found in adult bone marrow, where they also express thesame primitive markers, it has been hypothesized thatVSEL cells are related to a population of early PGCsthat are retained during development (Kucia et al.2006).
2.2.2. Wharton’s jelly. Wharton’s jelly is the connectivetissue surrounding the umbilical vessels and includesthe perivascular, intervascular and subamnion regions(Troyer & Weiss 2008). MSCs have been isolated fromall three regions and it remains unclear whether theyrepresent distinct cell populations (Karahuseyinogluet al. 2007). The cells have been given different namesby different groups (Troyer & Weiss 2008). MSCs iso-lated from Wharton’s jelly share similar propertieswith other cord blood MSCs as well as adult bonemarrow MSCs on the expression of markers, differen-tiation potential and cytokine production (McElreaveyet al. 1991; Wang, H.-S. et al. 2004; Anzalone et al.2010). Wharton’s jelly MSCs can also be induced todifferentiate into cells similar to neural, expressingneuron-specific enolase and other neural markers(Mitchell et al. 2003; Anzalone et al. 2010).
2.2.3. Amniotic fluid. In recent years, amniotic fluid hasemerged as a major source of putative pluripotent stemcells that avoid many of the problems associated withES cells such as their non-suitability for autologoususe, their capacity for tumour formation and the ethicalconcerns they raise.
In humans, the amniotic fluid starts to appear at thebeginning of week 2 of gestation as a small film ofliquid between the cells of the epiblast. The fluid expandsseparating the epiblast (i.e. the future embryo) from theamnioblasts (i.e. the future amnion), thus forming theamniotic cavity (Miki & Strom 2006). The origin ofamniotic fluid cells is still very much debatable(Kunisaki et al. 2007; Cananzi et al. 2009). However,what is known is that the majority of cells present areterminally differentiated and have limited proliferativecapacity (Gosden & Brock 1978; Siegel et al. 2007). How-ever, a number of studies have demonstrated the presenceof a subset of cells with a proliferative and differentiationpotential (Torricelli et al. 1993; Streubel et al. 1996).
A variety of different types of stem cells have been iso-lated and characterized from amniotic fluid. Theseinclude cells found in mid-gestation expressing the haema-topoietic marker CD34 (Da Sacco et al. 2010) as well ascells with mesenchymal features, able to proliferate invitro more rapidly than comparable foetal and adultcells (Kaviani et al. 2001; in‘t Anker et al. 2003a,b;Nadri & Soleimani 2007; Roubelakis et al. 2007; Sessaregoet al. 2008). Amniotic fluid MSCs are negative forhaematopoietic markers such as CD45, CD34 and CD14(Prusa & Hengstschlager 2002; in‘t Anker et al. 2003a,b;Prusa et al. 2003; Tsai et al. 2004). Despite their high pro-liferation rate, these cells display a normal karyotypewhen expanded in vitro and do not form tumours invivo (Sessarego et al. 2008). They exhibit a broad differen-tiation potential towards mesenchymal lineages (in‘tAnker et al. 2003a,b; Kolambkar et al. 2007; Nadri &Soleimani 2007; Tsai et al. 2007).
J. R. Soc. Interface
De Coppi et al. isolated c-kit-positive (CD117) cellsthat represent about 1 per cent of cells present insecond-trimester amniotic fluid. These cells werenamed amniotic fluid stem (AFS) cells. They can becultured without feeders, double in 36 h, are nottumorigenic, have long telomeres and retain a normalkaryotype for over 250 population doublings (DeCoppi et al. 2007a). Cultured human AFS cells are posi-tive for ES cell (e.g. Oct-4, Nanog and SSEA-4) andmesenchymal cell markers such as CD90, CD105(SH2), CD73 (SH3/4) and several adhesion molecules(e.g. CD29 and CD44; Tsai et al. 2006; Chambers et al.2007; De Coppi et al. 2007a). Furthermore, it was poss-ible to generate clonal human lines from these cells,verified by retroviral marking, which were capable of dif-ferentiating into lineages representative of all three EGlayers. Almost all clonal AFS cell lines express Oct-4and Nanog, markers of a pluripotent undifferentiatedstate (De Coppi et al. 2007a). c-kitþ Lin2 cells fromhuman and mouse amniotic fluid display a multi-lineagehaematopoietic potential in vitro and in vivo, despitehaving low or negative CD34 expression (Ditadi et al.2009). c-kit-positive human AFS cell lines can alsoform embryoid bodies (EB) with an incidence of18–82%. EB formation was accompanied by a decreasein Oct-4 and Nodal and the induction of differentiationmarkers such as Pax 6, Nestin (ectodermal), GATA 4and HBE1 (mesodermal) (Valli et al. 2010). The poten-tial for a wide range of regenerative medicine and tissueengineering applications as well as their relatively fewethical concerns makes them an attractive sourcefor cell therapy, particularly considering that a bank of100 000 amniotic fluid specimens could potentiallysupply 99 per cent of the US population with a perfectmatch for transplantation (Atala 2009).
2.2.4. Placenta. The placenta is a fetomaternal organinvolved in maintaining foetal tolerance and allowsnutrient uptake and gas exchange with the mother,but also contains a high number of progenitor cells orstem cells (Parolini et al. 2010). It has two sides: onefoetal (amnion and chorion) and one maternal (decid-uas). The amnion membrane contains two cell types:the amniotic epithelial cells (AECs) derived from theepiblast; and amniotic mesenchymal cells derived fromthe hypoblast. The chorion layer consists of cytotropho-blasts and syncytiotrophoblasts that are derived fromthe outer layer of the blastocyst (trophectoderm)(Ilancheran et al. 2009; Insausti et al. 2010).
The availability, phenotypic plasticity and immuno-modulatory properties of placenta-derived progenitor/stem cells are useful characteristics for cell therapyand tissue engineering. Cells can be isolated duringongoing pregnancy using minimally invasive techniquessuch as chorionic villus sampling (CVS) and placentaltissues are readily available at delivery for allogeneicor autologous use. Cells that have been isolated fromplacenta include the human AECs, human amnionmesenchymal stromal stem cells (AMSCs), humanchorionic mesenchymal/stromal stem cells (CMSCs),human chorionic trophoblastic cells and HSCs (Paroliniet al. 2008).

6 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Mesenchymal placenta cells are plastic adherent,share a similar immunophenotype and have lineagedifferentiation potential. They express stromal markerssuch as CD166, CD105, CD73, CD90 and others, whilethey are negative for the haematopoietic markers CD14,CD34 and CD45 (Igura et al. 2004; Sudo et al. 2007).Additionally, they express pluripotency markers such asSSEA3, SSEA4, Oct-4, Nanog, Tra-1-60 and Tra-1-81(Yen et al. 2005; Battula et al. 2007). They lack or havevery low expression of MHC class I antigens and do notexpress MHC class II antigens or the T-cell co-stimulatorymolecule B7, giving the cells immunomodulatoryproperties (Bailo et al. 2004; Miao et al. 2006).
2.2.4.1. Amniotic epithelial cells. AECs are commonlyisolated from the amniotic membrane using digestiveenzymes. The cells are plastic adherent and grow underMSC conditions, i.e. Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 10–20% FBS,with the addition of growth factors such as epidermalgrowth factor (EGF) (Miki et al. 2010). c-Kit and CD90(Thy-1) expression is either negative or at a low level(Miki et al. 2005; Miki & Strom 2006). However, evidencesuggests that they express pluripotency markers and havethe ability to form xenogeneic chimera with mouse EScells in vitro (Tamagawa et al. 2004). The cells have sub-sequently been differentiated in vitro into cell types fromall three germ layers (Miki et al. 2005; Miki & Strom2006; Ilancheran et al. 2007; Parolini et al. 2008).However, in contrast with ES cells, Miki et al. (2005)demonstrated that human AE cells did not form grossor histological tumours up to seven months post-transplantation in SCID/Beige mice.
2.2.4.2. Amnion-derived mesenchymal stromal cells.Both amnion and chorion MSCs have been extensivelycharacterized and can be isolated throughout gestationfrom first trimester to delivery. Both cells sharecommon characteristics, such as plastic adherence,and, despite limited proliferation capacity, show invitro differentiation down the osteogenic, adipogenic,chrondrogenic and neurogenic lineages, although somereport and show in vivo multi-organ engraftmentcapacity. In addition, the volume of term placentamakes it an attractive source of stem cells, as on averagehuman term placenta weighs more than 590 g(Bolisetty et al. 2002). Human placenta contains var-ious types of cells of different developmental origin.Cells of the chorion and decidua are derived from tro-phectoderm, while amnion is derived from the epiblastof the developing embryo (Crane & Cheung 1988).
MSCs have been successfully isolated from first-,second- and third-trimester placental compartments,including the amnion, chorion, decidua parietalis anddecidua basalis (in‘t Anker et al. 2004; Portmann-Lanz et al. 2006; Soncini et al. 2007; Poloni et al.2008), and represent less than 1 per cent of cells presentin the human placenta (Zhang et al. 2004; Alviano et al.2007). First-trimester placenta stem cells generally growfaster than those found in the third trimester(Portmann-Lanz et al. 2006), and the majority of
J. R. Soc. Interface
term placenta-derived stem cells are found at thequiescent G0/G1 phase of the cell cycle.
2.2.4.3. Chorion-derived stem cells. CMSCs have beenisolated during pregnancy (CVS) or from the term pla-centa at delivery. The immunophenotype of termplacenta cells is similar to that of adult bone marrowMSCs, although renin and flt-1 have been shown tobe expressed uniquely in placenta MSCs (Fukuchiet al. 2004). Similarly to human amnion isolated fromterm placenta, term chorion MSCs successfully engraftin neonatal swine and rats and failed to induce a xeno-geneic response (Bailo et al. 2004), indicating that thesecells may have an immunoprivileged status consistentwith their low level of HLA I and absence of HLA IIexpression (Kubo et al. 2001). Of relevance for pre-natal autologous and allogeneic therapy, chorionMSCs can be successfully isolated from chorionic villiduring first-trimester gestation. The chorionic villi donot express typical MSC cell surface antigens andhave the capacity to differentiate into lineages of thethree germ layers, with a subset of cells expressing thepluripotency markers Oct-4, ALP, Nanog and Sox2(Spitalieri et al. 2009). In addition to differentiatinginto adipogenic, chondrogenic and osteogenic cells invitro (Portmann-Lanz et al. 2006; Ilancheran et al.2007), they can also differentiate into cells with somecharacteristics of hepatocytes in vitro and have the abil-ity to store glycogen (Chien et al. 2006; Huang 2007;Tamagawa et al. 2007). Differentiation into other celltypes such as endothelial cells (Alviano et al. 2007),cardiomyocytes (Zhao et al. 2005), neurons, oligoden-drocytes and glial cells (Miao et al. 2006; Yen et al.2008; Portmann-Lanz et al. 2010) has also beenreported, but these findings have mostly relied on upre-gulation of tissue-specific markers without providingrobust functionality tests.
3. THERAPUTIC APPLICATIONS OFFOETAL STEM CELLS
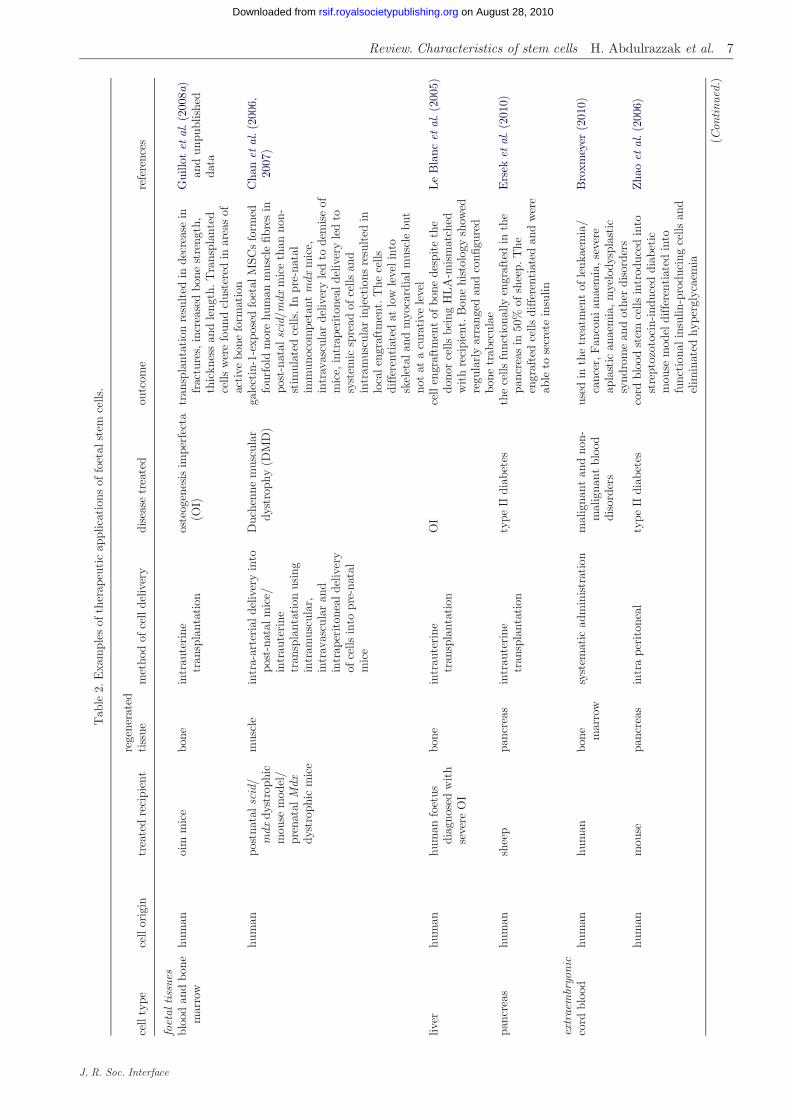
Regenerative medicine aims to restore diseased ordamaged tissue using the body’s own cells in order toovercome the shortage of donated organs and the riskassociated with their rejection (Atala 2009). The useof stem cells from foetal, cord blood and extraembryonictissues for regenerative medicine applications hasincreased over the past few years. However, the surpris-ing picture that has emerged is that cell replacement isprobably not the major mechanism by which celltherapy confers functional benefit. Rather, stem cellsact beneficially by exerting trophic effects on host tis-sues. Moreover, because they can be expanded to verylarge numbers compared with adult stem cells, theyare ideal candidates for seeding scaffolds and matricesused for tissue engineering applications such as therepair of hollow and solid organs. In this section, wewill consider some of the therapeutic applicationsFSCs have been put to in pre-clinical studies for repair-ing diseased or damaged tissue as well as in cancertreatment (summarized examples are listed in table 2).
Tab
le2.
Exa
mpl
esof
ther
apeu
tic
appl
icat
ions
offo
etal
stem
cells
.
cell
type
cell
orig
intr
eate
dre
cipi
ent
rege
nera
ted
tiss
uem
etho
dof
cell
deliv
ery
dise
ase
trea
ted
outc
ome
refe
renc
es
foet
altiss
ues
bloo
dan
dbo
nem
arro
whu
man
oim
mic
ebo
nein
trau
teri
netr
ansp
lant
atio
nos
teog
enes
isim
perf
ecta
(OI)
tran
spla
ntat
ion
resu
lted
inde
crea
sein
frac
ture
s,in
crea
sed
bone
stre
ngth
,th
ickn
ess
and
leng
th.T
rans
plan
ted
cells
wer
efo
und
clus
tere
din
area
sof
acti
vebo
nefo
rmat
ion
Gui
llot
etal
.(2
008a
)an
dun
publ
ishe
dda
ta
hum
anpo
stna
talsc
id/
mdx
dyst
roph
icm
ouse
mod
el/
pren
atal
Mdx
dyst
roph
icm
ice
mus
cle
intr
a-ar
teri
alde
liver
yin
topo
st-n
atal
mic
e/in
trau
teri
netr
ansp
lant
atio
nus
ing
intr
amus
cula
r,in
trav
ascu
lar
and
intr
aper
iton
ealde
liver
yof
cells
into
pre-
nata
lm
ice
Duc
henn
em
uscu
lar
dyst
roph
y(D
MD
)ga
lect
in-1
-exp
osed
foet
alM
SCs
form
edfo
urfo
ldm
ore
hum
anm
uscl
efib
res
inpo
st-n
atal
scid
/mdx
mic
eth
anno
n-st
imul
ated
cells
.In
pre-
nata
lim
mun
ocom
peta
ntm
dxm
ice,
intr
avas
cula
rde
liver
yle
dto
dem
ise
ofm
ice,
intr
aper
iton
ealde
liver
yle
dto
syst
emic
spre
adof
cells
and
intr
amus
cula
rin
ject
ions
resu
lted
inlo
calen
graf
tmen
t.T
hece
llsdi
ffer
enti
ated
atlo
wle
velin
tosk
elet
alan
dm
yoca
rdia
lm
uscl
ebu
tno
tat
acu
rati
vele
vel
Cha
net
al.(2
006,
2007
)
liver
hum
anhu
man
foet
usdi
agno
sed
wit
hse
vere
OI
bone
intr
aute
rine
tran
spla
ntat
ion
OI
cell
engr
aftm
ent
ofbo
nede
spit
eth
edo
nor
cells
bein
gH
LA
-mis
mat
ched
wit
hre
cipi
ent.
Bon
ehi
stol
ogy
show
edre
gula
rly
arra
nged
and
confi
gure
dbo
netr
abec
ulae
Le
Bla
ncet
al.(
2005
)
panc
reas
hum
ansh
eep
panc
reas
intr
aute
rine
tran
spla
ntat
ion
type
IIdi
abet
esth
ece
llsfu
ncti
onal
lyen
graf
ted
inth
epa
ncre
asin
50%
ofsh
eep.
The
engr
afte
dce
llsdi
ffer
enti
ated
and
wer
eab
leto
secr
ete
insu
lin
Ers
eket
al.(2
010)
extr
aem
bryo
nic
cord
bloo
dhu
man
hum
anbo
ne mar
row
syst
emat
icad
min
istr
atio
nm
alig
nant
and
non-
mal
igna
ntbl
ood
diso
rder
s
used
inth
etr
eatm
ent
ofle
ukae
mia
/ca
ncer
,Fan
coni
anae
mia
,se
vere
apla
stic
anae
mia
,m
yelo
dysp
last
icsy
ndro
me
and
othe
rdi
sord
ers
Bro
xmey
er(2
010)
hum
anm
ouse
panc
reas
intr
ape
rito
neal
type
IIdi
abet
esco
rdbl
ood
stem
cells
intr
oduc
edin
tost
rept
ozot
ocin
-ind
uced
diab
etic
mou
sem
odel
differ
enti
ated
into
func
tion
alin
sulin
-pro
duci
ngce
llsan
del
imin
ated
hype
rgly
caem
ia
Zha
oet
al.(
2006
)
(Con
tinu
ed.)
Review. Characteristics of stem cells H. Abdulrazzak et al. 7
J. R. Soc. Interface
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from

Tab
le2.
(Con
tinu
ed.)
cell
type
cell
orig
intr
eate
dre
cipi
ent
rege
nera
ted
tiss
uem
etho
dof
cell
deliv
ery
dise
ase
trea
ted
outc
ome
refe
renc
es
Wha
rton
’sJe
llyhu
man
rat
brai
nin
ject
ion
into
the
brai
nst
riat
umP
arki
nson
’sdi
seas
eth
ece
llsam
elio
rate
dap
omor
phin
e-in
duce
dro
tati
ons
ina
rat
mod
elof
Par
kins
on’s
dise
ase
Wei
sset
al.(2
006)
amni
otic
fluid
shee
psh
eep
mus
cle
cells
seed
edon
colla
gen
hydr
ogel
and
plac
edon
inte
stin
alce
llula
rgr
aft
part
ialdi
aphr
agm
atic
repl
acem
ent
diap
hrag
mat
iche
rnia
recu
rren
cew
assi
gnifi
cant
lyhi
gher
inan
imal
sth
atdi
dno
tre
ceiv
eth
ece
llgr
afts
Fuc
hset
al.(2
004)
rat
rat
smoo
thm
uscl
ece
llsin
ject
edin
tosi
teof
inju
ryw
ound
heal
ing
ofin
jure
dbl
adde
rce
llspr
even
ted
cryo
-inj
ury-
indu
ced
hype
rtro
phy
ofsu
rviv
ing
blad
der
smoo
thm
uscl
ece
llsbu
tfa
iled
todi
ffer
enti
ate
spec
ifica
llyto
smoo
thm
uscl
e
De
Cop
piet
al.
(200
7a)
mou
sem
ouse
brai
nin
tra
vent
ricu
lar
inje
ctio
nfo
calce
rebr
alis
chae
mia
repe
rfus
ion
inju
ry
the
cells
sign
ifica
ntly
reve
rsed
neur
olog
ical
defic
its
inth
etr
eate
dan
imal
s
Reh
niet
al.(2
007)
hum
anm
ouse
lung
syst
emat
icad
min
istr
atio
nlu
ngin
jury
AF
Sce
llsex
hibi
ted
ast
rong
tiss
ueen
graf
tmen
tin
am
ouse
lung
inju
rym
odel
and
expr
esse
dsp
ecifi
cal
veol
aran
dbr
onch
iola
rep
ithe
lialm
arke
rs
Car
raro
etal
.(2
008)
hum
anra
the
art
intr
amyo
card
ialin
ject
ion
myo
card
ialin
farc
tion
trea
ted
anim
als
show
eda
pres
erva
tion
ofth
ein
farc
ted
thic
knes
s,at
tenu
atio
nof
left
vent
ricl
ere
mod
ellin
g,a
high
erva
scul
arde
nsit
yan
dge
nera
lim
prov
emen
tof
card
iac
func
tion
Yeh
etal
.(2
010)
plac
enta
hum
anam
nion
AE
Cm
ouse
brai
nin
trac
rani
alin
ject
ion
Par
kins
on’s
dise
ase
cells
tran
spla
nted
into
am
ouse
mod
elof
Par
kins
on’s
dise
ase
differ
enti
ated
into
neur
on,as
troc
yte
and
olig
oden
droc
yte
and
prom
oted
endo
geno
usne
urog
enes
is
Kon
get
al.(2
008)
hum
anam
nion
MSC
rat
hear
tin
tram
yoca
rdia
lin
ject
ion
myo
card
ialin
farc
tion
cells
inte
grat
edin
tono
rmal
and
infa
rcte
dra
tca
rdia
cti
ssue
whe
reth
eydi
ffer
enti
ated
into
card
iom
yocy
te-lik
ece
llsan
dre
sult
edin
cons
ider
able
impr
ovem
ents
tove
ntri
cula
rfu
ncti
on,ca
pilla
ryde
nsit
yan
dsc
arti
ssue
Zha
oet
al.(
2005
),V
entu
raet
al.
(200
7)
8 Review. Characteristics of stem cells H. Abdulrazzak et al.
J. R. Soc. Interface
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from

Review. Characteristics of stem cells H. Abdulrazzak et al. 9
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
3.1. Bone
Osteogenesis imperfecta (OI) is characterized by osteo-penia and bone fragility due to abnormal collagenproduction caused by mutations in the a chains of col-lagen type I. First-trimester allogeneic HLA-mismatched male foetal liver MSCs were transplantedin utero into a female foetus diagnosed with severeOI. Bone biopsy showed regularly arranged and config-ured bone trabeculae and no adverse immune reactionwas observed (Le Blanc et al. 2005). Using a mousemodel of OI, our group has shown that in utero trans-plantation of foetal blood MSCs ameliorated thedisease phenotype, producing a clinically relevanttwo-thirds reduction in fracture incidence along withan improvement in bone structure and mechanicalproperties (Guillot et al. 2008a). Foetal bonemarrow MSCs loaded on a highly porous scaffold wereable to reach confluence within the scaffold morequickly than umbilical cord and adult MSCs. Further-more, they demonstrated higher in vitro and in vivoosteogenic differentiation capacity, highlighting theirsuitability for bone tissue engineering applications(Zhang et al. 2009).
3.2. Muscle
Duchenne muscular dystrophy (DMD) is an X-linkedmyopathy affecting one in 3500 boys. The main geneticdefect leads to absence of dystrophin, resulting inmuscle damage and wasting. The affected boysbecome wheelchair-bound by 12 years of age and suc-cumb to respiratory failure or cardiopmyopathy bythe third or forth decade of life (Manzur & Muntoni2009). The main animal model for studying DMD isthe mdx mouse that has a premature stop codon result-ing in a termination in exon 23 of the dystrophin gene(Grounds et al. 2008). Galectin-1-treated foetal bloodand bone marrow MSCs, transplanted into postnatalsevere combined immunodeficiency/dystrophic mice(scid/mdx) as well immunodeficient mice whosemuscle was induced to regenerate by cryodamage,formed fourfold more human muscle fibres than non-stimulated MSCs (Chan et al. 2006). Intrauterinetransplantation of foetal MSCs into immunocompetentE14-16 mdx mice resulted in widespread long-termengraftment in multiple organs with a predilection formuscle compared with non-muscle tissues. However,the engraftment level observed (0.5–1%) falls signifi-cantly below the levels required for functionalimprovement in DMD. The low engraftment might belinked to an absence of muscle pathology at the timeof transplantation (Chan et al. 2007).
AFS cells have been used in an ovine model ofdiaphragmatic hernia: repair of the muscle deficitusing grafts engineered with autologous mesenchymalamniocytes leads to better structural and functionalresults than equivalent foetal myoblast-based and acel-lular grafts (Fuchs et al. 2004; Kunisaki et al. 2006). Ina rat model of bladder cryo-injury, AFS cells also showthe ability to differentiate into smooth muscle and toprevent the compensatory hypertrophy of survivingsmooth muscle cells (De Coppi et al. 2007b).
J. R. Soc. Interface
Human AFS cells, transplanted into an immune-competent and ciclosporin immune-suppressed ratmodel of myocardial infarction, were rejected probablybecause of the expression of B7 co-stimulatorymolecules as well as macrophage marker CD68.Furthermore, chondro-osteogenic cell masses wereobserved in the infarcted hearts of some of the trans-planted animals (Chiavegato et al. 2007). However, inanother study it was found that these masses occurredindependently of AFS cell injection and that the cellsdid not increase the presence of these masses (Deloet al. 2010).
Human AMSCs are able not only to express cardiac-specific genes under specific culture conditions, but alsoto integrate into normal and infarcted rat cardiac tissuewhere they differentiate into cardiomyocyte-like cells(Zhao et al. 2005). The cells result in considerableimprovements to ventricular function, capillary densityand scar tissue (Ventura et al. 2007) and similar find-ings have been reported using native rat AMSCs(Fujimoto et al. 2009). Stem cells derived from pre-natal chorionic villi were also successfully engineeredinto a living autologous heart valve, which could havepostnatal applications for repairing congenital cardiacmalformations (Schmidt et al. 2006).
3.3. Kidney
In a mouse model of collagen deficiency characterizedby abnormal collagen deposition in renal glomeruli(Phillips et al. 2002; Brodeur et al. 2007), we recentlyshowed that intrauterine transplantation of foetalblood MSCs led to a reduction of abnormal homotri-meric collagen type I deposition in the glomeruli of4–12 week old col1a2-deficient mice. Furthermore, weshowed that the damaged kidneys preferentiallyrecruited donor cells in the glomeruli. The study sup-ports the feasibility of pre-natal treatment for renaldiseases such as Alport syndrome and the polycystickidney diseases (Guillot et al. 2008b).
Acute tubular necrosis could also be potentially trea-ted with foetal cells. Human AFS cells injected into animmunodeficient mouse model of the disease decreasedthe number of damaged tubules and reduced apoptosis.The cells also promoted the proliferation of tubular epi-thelial cells and appeared to have a beneficialimmunomodulatory effect (Perin et al. 2010).
3.4. Lung
Human AFS cells injected into the tail vein of nudemice following hyperoxia injury show the capacity tohome to the lung and engraft. They also expressedspecific alveolar and bronchiolar epithelial markers(e.g. TFF1, SPC and CC10; Carraro et al. 2008).
AECs and AMSCs from either human or mouse havebeen used in a mouse model of bleomycin-induced lunginjury and shown to cause a reduction in severity oflung fibrosis. This occurs regardless of cell source (allo-geneic or xenogeneic) or administration route (systemic,intravenous or intraperitoneal; local, intratracheal;Cargnoni et al. 2009).

10 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
3.5. Liver
Second-trimester foetal liver cells, enriched for CD326(hepatocyte progenitor marker) and labelled with Tc-d,l-hexamethyl-propylene-amine oxime (Tc-HMPAO)were infused into the hepatic artery of 25 patientswith end stage liver cirrhosis. The cells restored thelost liver function in the patients without provokingan immune reaction, probably because of the absenceof HLA class II expression of the infused cells (Khanet al. 2010).
Human AECs can secrete albumin in culture and,when b-galactosidase-tagged AECs were transplantedinto immunodeficient mice, the cells integrated intothe liver parenchyma and could be detected until day7 post transplant (duration of study; Sakuragawaet al. 2000). In another study, the engrafted cellsresulted in the detection of human a-1 antitrypsin inthe serum of recipient animals, confirming that AECsare capable of performing this important hepaticfunction in vivo (Miki & Strom 2006).
3.6. Brain
Human Wharton’s jelly MSCs ameliorate apomorphine-induced behavioural deficits in a hemiparkinsonian ratmodel (Weiss et al. 2006). There was a significantdecrease in apomorphine-induced rotations at fourweeks continuing up to 12 weeks post transplantationin Parkinson’s disease (PD) rats that received humanWharton’s jelly MSCs transplants compared with thePD rats that received a sham transplant. The behav-ioural findings correlated with the numbers of tyrosinehydroxylase-positive cell bodies observed in the mid-brain following sacrifice at 12 weeks, indicating a‘rescue from a distance’ phenomenon. One explanationfor this effect may be that Wharton’s jelly MSCssynthesize glial cell-derived neurotrophic factor(GDNF), a potent survival factor for dopaminergicneurons, as well as other trophic factors such asvascular endothelial growth factor (VEGF) and ciliaryneurotrophic factor. In another report, Wharton’sjelly MSCs were first induced towards dopaminergicneurons using neuron-conditioned media, sonic hedge-hog and fibroblast growth factor 8, and thentransplanted into hemiparkinsonian rats (Fu et al.2006). Despite the fact that the rats were not immunesuppressed, Wharton’s jelly MSCs were identified fivemonths later and prevented the progressive degener-ation/behavioural deterioration seen in control ratswith unilateral lesions. Similarly, rat Wharton’s jellycells transplanted into the brains of rats with global cer-ebral ischaemia caused by cardiac arrest andresuscitation significantly reduced neuronal loss, appar-ently owing to a rescue phenomenon (Jomura et al.2007). Lund et al. administered Wharton’s jelly MSCsinto the eyes of a rodent model of retinal disease.Here, Wharton’s jelly MSCs were compared with bonemarrow-derived mesenchymal stem cells (BMSCs) andplacental stem cells. They reported that the Wharton’sjelly MSCs exhibited the best histological evidence ofphotoreceptor rescue with increased production oftrophic factors such as brain-derived neurotrophic
J. R. Soc. Interface
factor and fibroblast growth factor 2 (Lund et al.2007). Interestingly, Wharton’s jelly MSCs trans-planted into the vitreous demonstrated a rescue effect,indicating that they could enhance the survival ofphotoreceptor cells without being in close proximityto them. This effect was presumably the result ofdiffusible growth factors.
Very similar findings have also been reported forAECs that confer neuroprotection and functional recov-ery in animal models of Parkinson’s disease (Bankiewiczet al. 1994; Kakishita et al. 2000, 2003; Kong et al.2008) and ischaemia (Liu et al. 2008). The observedtherapeutic effects are probably mediated by secretionof diffusible factors, including neurotransmitters(Elwan & Sakuragawa 1997; Sakuragawa et al. 1997,2001) and neurotrophic and growth factors (Koizumiet al. 2000; Uchida et al. 2000).
AFS cells can harbour trophic and protective effectsin the central and the peripheral nervous systems. Panet al. (2006, 2007, 2009) showed that AFS cells facilitateperipheral nerve regeneration after injury, particularlyif accompanied with suppressors of inflammatory cyto-kines such as fermented soya bean extracts. Aftertransplantation into the striatum, AFS cells are capableof surviving and integrating in the rat adult brain andcan migrate towards areas of ischaemic damage(Cipriani et al. 2007). Moreover, the intra-ventricularadministration of AFS cells in mice with focal cerebralischaemia–reperfusion injuries significantly reversesneurological deficits in the treated animals (Rehniet al. 2007).
3.7. Cancer therapy
HSCs from bone marrow or umbilical cord blood havelong been used to re-establish the haematopoieticsystem following radiation and/or chemotherapy(Osawa et al. 1996; Bhatia et al. 1997). More recently,research has shown that MSCs are attracted to tumoursbecause the latter often act as unresolved wounds pro-ducing a continuous source of chemoattractantinflammatory mediators. This property is beingexploited by using exogenously delivered MSCs asvehicles for delivering anti-cancer molecules to tumours(Loebinger & Janes 2010). Wharton’s jelly cells, likeneural stem cells and MSCs (Aboody et al. 2000;Studeny et al. 2004), appear to migrate to areas oftumour growth. Human breast carcinoma cells wereintravenously injected into SCID mice, followed byintravenous transplantation of fluorescently labelledWharton’s jelly MSCs. One week after transplantWharton’s jelly MSCs were found near or within lungtumours and not in other tissues. They were engineeredto express human interferon beta and were administeredintravenously into SCID mice bearing tumours. Thistreatment significantly reduced the tumour burden(Rachakatla et al. 2007). Similarly, human umbilicalcord blood-derived MSCs (UCB–MSCs) have beenused to deliver a secretable trimeric form of tumournecrosis factor-related apoptosis-inducing ligand(stTRAIL), via adenoviral transduction mediated bycell-permeable peptides, to human glioma cells innude mice. The genetically modified cells showed

Review. Characteristics of stem cells H. Abdulrazzak et al. 11
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
greater efficacy in terms of inhibiting tumour growthand prolonging the survival of glioma-bearing micecompared with direct injection of adenovirus encodingthe stTRAIL gene into the tumour mass (Kim et al.2008).
4. USE OF FSCs FOR REPROGRAMMING
The major advantage of human ES cells over other stemcell types is their capacity to differentiate into lineagesfrom the three germ layers. However, there are twomajor objections to their use. The moral objection isthat their derivation requires the destruction ofembryos. The practical objection is that they have alimited clinical application as they can only be used inan allogeneic fashion. This is also true for some foetaltissues such as liver and bone marrow. However, extra-embryonic tissues can be used autologously.
Thus, it was seen as advantageous to develop amethod of creating ES-like cells from somatic cells.Early attempts at achieving this were nuclear trans-plantation and fusion of somatic cells and ES cells(Jaenisch 2009). However, a considerable step forwardwas the generation of the induced pluripotent stem(iPS) cells. The production of iPS cells with quasi-identical genetic and functional properties offers thepossibility to bypass both moral conflicts and differentgenetic background inherent to the technologiesmentioned above. iPS are developed from a non-pluripotent cell, usually an adult somatic cell, bycausing a forced expression of several genetic sequencesand were first produced in 2006 by Takahashi &Yamanaka from mouse somatic cells. The key genesOct-4, the transcription factor Sox2, c-Myc protoonco-gene protein and Klf4 (Krueppel-like factor 4) weresufficient to reprogramme fibroblasts to cells closelyresembling ES cells (Takahashi & Yamanaka 2006;Takahashi et al. 2007).
Despite the high similarity between iPS and ES cells,tumour formation in iPS cell chimeric mice was high,presumably because of the expression of c-Myc in iPScell-derived somatic cells (Maherali et al. 2007).Thomson et al. showed that c-Myc was not necessaryas they were able to generate iPS cells using Oct-4,Sox2, Nanog and Lin28 using a lentiviral system (Yuet al. 2007). More recently, it was shown that humanfibroblasts cells can be used to generate iPS cells iftransduced with Oct-4 and Sox2 only in the presenceof valproic acid (Huangfu et al. 2008).
Terminally differentiated cells might not be the idealcandidate for iPS cell generation because a greaternumber of steps could be required to ‘reprogramme’their genome than somatic stem cells. Kim et al.(2009a,b) have shown that it is possible to generateiPS cells by transducing adult mouse neural stem cells(NSCs) and human foetal NSCs with Oct-4 only. Fur-thermore, when human adipose MSCs were used, theywere reprogrammed more efficiently than fibroblasts(Sun et al. 2009).
Few reports have also emerged on the use of cord andextra ES cells for iPS generation. Galende et al. usedterminally differentiated amniotic fluid cells as
J. R. Soc. Interface
candidates for reprogramming and found that the iPScell colonies were generated twice as fast, yieldingnearly a 200 per cent increase in number comparedwith cultured adult skin cells. However, they did notprovide absolute efficiency data (Galende et al. 2009).Li et al. reprogrammed human amniotic fluid-derivedcells (hAFDCs) with efficiencies varying from 0.059 to1.525 per cent. The iPS colonies generated expressedpluripotency markers such as Oct-4, Sox2 and SSEA 4and they maintained a normal karyotype (Li et al.2009). Wharton’s jelly cells and term placenta cellshave also been used, giving reprogramming efficienciesof 0.4 per cent and 0.1 per cent, respectively, which com-pare rather well with efficiencies reported for adultfibroblasts (less than 0.01%). The generated iPS cellcolonies expressed several pluripotency markers, formedEB and, when injected into nude mice, they generatedteratoma-containing derivates of the three germ layers(Cai et al. 2010). Giorgetti et al. selected CD133þ (hae-matopoietic stem and progenitor cell marker) cord bloodcells and managed to reprogramme them by retroviraltransduction with Oct-4 and Sox2 only without needingadditional chemical compounds. iPS cell colonies derivedfrom CD133þ cord blood cells (expressing pluripotencymarkers and capable of forming EB and teratomas)were generated in two weeks whereas no colonies wereobtained with control keratinocytes or fibroblasts usingthe two factors only (Giorgetti et al. 2009, 2010).
CD133þ cord blood cells and foetal NSCs alreadyhave a baseline level expression of Oct-4 and/or Sox2and that, along with a generally more permissive chro-matin organization, might be the key to their greaterreprogramming efficiency compared with adult cells.
5. CONCLUSION
Human FSCs can be isolated from foetal organs orextraembryonic sources. They have the advantage ofrapid proliferation, stable karyotype and low or negli-gible immunogenicity. Unlike ES cells, they do notform teratomas in vivo and have far fewer ethical con-cerns as they are mostly obtained from tissues thatwould otherwise be discarded. Many of these cellsseem to express some of the same pluipotency markersfound in ES cells, a feature largely absent from mostadult-derived stem cells. They also have the furtheradvantage over adult stem cells of senescing muchlater and being more readily amenable to geneticmodification.
All of these features make them valuable for poten-tial therapy applications. Thus far they have beenused in pre-clinical settings to treat a variety of diseasessuch as osteogenesis imperfecta, congenital diaphrag-matic hernia, Parkinson’s disease and cancer withencouraging results. Finally, their usefulness for iPSgeneration is very likely to expand their future clinicaluse even further.
REFERENCES
Abdallah, B. M. & Kassem, M. 2008 Human mesenchymalstem cells: from basic biology to clinical applications.Gene Ther. 15, 109–116. (doi:10.1038/sj.gt.3303067)

12 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Aboody, K. S. et al. 2000 Neural stem cells display extensivetropism for pathology in adult brain: evidence fromintracranial gliomas. Proc. Natl Acad. Sci. USA 97,12 846–12 851. (doi:10.1073/pnas.97.23.12846)
Alaminos, M., Perez-Kohler, B., Garzon, I., Garcia-Honduvilla,N., Romero, B., Campos, A. & Bujan, J. 2010 Transdiffer-entiation potentiality of human Wharton’s jelly stem cellstowards vascular endothelial cells. J. Cell. Physiol. 223,640–647.
Almeida-Porada, G., El Shabrawy, D., Porada, C. & Zanjani,E. 2002 Differentiative potential of human metanephricmesenchymal cells. Exp. Hematol. 30, 1454–1462.(doi:10.1016/S0301-472X(02)00967-0)
Alviano, F. et al. 2007 Term amniotic membrane is a highthroughput source for multipotent mesenchymal stemcells with the ability to differentiate into endothelial cellsin vitro. BMC Dev. Biol. 7, 11–11. (doi:10.1186/1471-213X-7-11)
Andrews, P. W., Matin, M. M., Bahrami, A. R., Damjanov, I.,Gokhale, P. & Draper, J. S. 2005 Embryonic stem (ES)cells and embryonal carcinoma (EC) cells: opposite sidesof the same coin. Biochem. Soc. Trans. 33, 1526–1530.
Anzalone, R., Iacono, M. L., Corrao, S., Magno, F., Loria, T.,Cappello, F., Zummo, G., Farina, F. & La Rocca, G. 2010New emerging potentials for human Wharton’s jellymesenchymal stem cells: immunological features and hep-atocyte-like differentiative capacity. Stem Cells Dev. 19,423–438. (doi:10.1089/scd.2009.0299)
Atala, A. 2009 Engineering organs. Curr. Opin. Biotechnol.20, 575–592. (doi:10.1016/j.copbio.2009.10.003)
Bailo, M. et al. 2004 Engraftment potential of human amnionand chorion cells derived from term placenta. Transplan-tation 78, 1439–1448. (doi:10.1097/01.TP.0000144606.84234.49)
Ballen, K. K. 2005 New trends in umbilical cord bloodtransplantation. Blood 105, 3786–3792. (doi:10.1182/blood-2004-10-4125)
Bankiewicz, K. S., Palmatier, M., Plunkett, R. J., Cummins,A. & Oldfield, E. H. 1994 Reversal of hemiparkinsoniansyndrome in nonhuman primates by amnion implantationinto caudate nucleus. J. Neurosurg. 81, 869–876.
Battula, V. L. et al. 2007 Human placenta and bone marrowderived MSC cultured in serum-free, b-FGF-containingmedium express cell surface frizzled-9 and SSEA-4 andgive rise to multilineage differentiation. Differentiation75, 279–291. (doi:10.1111/j.1432-0436.2006.00139.x)
Bhatia, M., Wang, J., Kapp, U., Bonnet, D. & Dick, J. 1997Purification of primitive human hematopoietic cellscapable of repopulating immune-deficient mice. Proc.Natl Acad. Sci. USA 94, 5320–5325. (doi:10.1073/pnas.94.10.5320)
Bieback, K., Kern, S., Kluter, H. & Eichler, H. 2004 Criticalparameters for the isolation of mesenchymal stem cellsfrom umbilical cord blood. Stem Cells 22, 625–634.(doi:10.1634/stemcells.22-4-625)
Bolisetty, S., Koh, T. H. H. G., Hammond, S., Panaretto, K. &Whitehall, J. 2002 Correlation of umbilical cord weight withbirth weight. Arch. Dis. Child Fetal Neonatal Ed. 86, F140.(doi:10.1136/fn.86.2.F140-a)
Brodeur, A., Wirth, D., Franklin, C., Reneker, L., Miner, J. &Phillips, C. 2007 Type I collagen glomerulopathy: post-natal collagen deposition follows glomerular maturation.Kidney Int. 71, 985–993. (doi:10.1038/sj.ki.5002173)
Brooke, G. et al. 2009 Manufacturing of human placenta-derived mesenchymal stem cells for clinical trials.Br. J. Haematol. 144, 571–579. (doi:10.1111/j.1365-2141.2008.07492.x)
J. R. Soc. Interface
Broxmeyer, H. 2010 Umbilical cord transplantation: epilogue.Semin. Hematol. 47, 97–103. (doi:10.1053/j.seminhema-tol.2009.10.002)
Broxmeyer, H. E., Douglas, G. W., Hangoc, G., Cooper, S.,Bard, J., English, D., Arny, M., Thomas, L. & Boyse,E. A. 1989 Human umbilical cord blood as a potentialsource of transplantable hematopoietic stem/progenitorcells. Proc. Natl Acad. Sci. USA 86, 3828–3832. (doi:10.1073/pnas.86.10.3828)
Brunstein, C. G., Setubal, D. C. & Wagner, J. E. 2007Expanding the role of umbilical cord blood transplan-tation. Br. J. Haematol. 137, 20–35.
Cai, J. et al. 2010 Generation of human induced pluripotentstem cells from umbilical cord matrix and amnioticmembrane mesenchymal cells. J. Biol. Chem. 285,11227–11 234. (doi:10.1074/jbc.M109.086389)
Campagnoli, C., Roberts, I., Kumar, S., Bennett, P.,Bellantuono, I. & Fisk, N. 2001 Identification of mesench-ymal stem/progenitor cells in human first-trimester fetalblood, liver, and bone marrow. Blood 98, 2396–2402.(doi:10.1182/blood.V98.8.2396)
Cananzi, M., Atala, A. & De Coppi, P. 2009 Stem cellsderived from amniotic fluid: new potentials in regenerativemedicine. Reprod. Biomed. Online 18(Suppl. 1), 17–27.
Cargnoni, A. et al. 2009 Transplantation of allogeneic andxenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant. 18, 405–422.(doi:10.3727/096368909788809857)
Carraro, G. et al. 2008 Human amniotic fluid stem cells canintegrate and differentiate into epithelial lung lineages.Stem Cells 26, 2902–2911. (doi:10.1634/stemcells.2008-0090)
Castrechini, N. M., Murthi, P., Gude, N. M., Erwich, J. J.,Gronthos, S., Zannettino, A., Brennecke, S. P. & Kalionis,B. 2010 Mesenchymal stem cells in human placental chor-ionic villi reside in a vascular niche. Placenta 31, 203–212.(doi:10.1016/j.placenta.2009.12.006)
Chambers, I. et al. 2007 Nanog safeguards pluripotency andmediates germline development. Nature 450, 1230–1234.(doi:10.1038/nature06403)
Chan, J., O’Donoghue, K., de la Fuente, J., Roberts, I. A.,Kumar, S., Morgan, J. E. & Fisk, N. M. 2005 Humanfetal mesenchymal stem cells as vehicles for gene delivery.Stem Cells 23, 93–102. (doi:10.1634/stemcells.2004-0138)
Chan, J. et al. 2006 Galectin-1 induces skeletal muscle differ-entiation in human fetal mesenchymal stem cells andincreases muscle regeneration. Stem Cells 24, 1879–1891.(doi:10.1634/stemcells.2005-0564)
Chan, J., Waddington, S., O’Donoghue, K., Kurata, H.,Guillot, P., Gotherstrom, C., Themis, M., Morgan, J. &Fisk, N. 2007 Widespread distribution and muscle differen-tiation of human fetal mesenchymal stem cells afterintrauterine transplantation in dystrophic mdx mouse.Stem Cells 25, 875–884. (doi:10.1634/stemcells.2006-0694)
Chiavegato, A. et al. 2007 Human amniotic fluid-derived stemcells are rejected after transplantation in the myocardiumof normal, ischemic, immuno-suppressed or immuno-deficient rat. J. Mol. Cell. Cardiol. 42, 746–759. (doi:10.1016/j.yjmcc.2006.12.008)
Chien, C., Yen, B., Lee, F., Lai, T., Chen, Y., Chan, S. &Huang, H. 2006 In vitro differentiation of humanplacenta-derived multipotent cells into hepatocyte-likecells. Stem Cells 24, 1759–1768. (doi:10.1634/stemcells.2005-0521)
Cipriani, S., Bonini, D., Marchina, E., Balgkouranidou, I.,Caimi, L., Grassi Zucconi, G. & Barlati, S. 2007 Mesench-ymal cells from human amniotic fluid survive and migrate

Review. Characteristics of stem cells H. Abdulrazzak et al. 13
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
after transplantation into adult rat brain. Cell Biol. Int.31, 845–850. (doi:10.1016/j.cellbi.2007.01.037)
Clapp, D. W., Freie, B., Lee, W. H. & Zhang, Y. Y. 1995 Mol-ecular evidence that in situ-transduced fetal liverhematopoietic stem/progenitor cells give rise to medullaryhematopoiesis in adult rats. Blood 86, 2113–2122.
Crane, J. & Cheung, S. 1988 An embryogenic model toexplain cytogenetic inconsistencies observed in chorionicvillus versus fetal tissue. Prenat. Diagn. 8, 119–129.(doi:10.1002/pd.1970080206)
Cumano, A., Ferraz, J. C., Klaine, M., Di Santo, J. P. &Godin, I. 2001 Intraembryonic, but not yolk sac hemato-poietic precursors, isolated before circulation, providelong-term multilineage reconstitution. Immunity 15,477–485. (doi:10.1016/S1074-7613(01)00190-X)
Dan, Y., Riehle, K., Lazaro, C., Teoh, N., Haque, J.,Campbell, J. & Fausto, N. 2006 Isolation of multipotentprogenitor cells from human fetal liver capable of differen-tiating into liver and mesenchymal lineages. Proc. NatlAcad. Sci. USA 103, 9912–9917. (doi:10.1073/pnas.0603824103)
Da Sacco, S., Sedrakyan, S., Boldrin, F., Giuliani, S.,Parnigotto, P., Habibian, R., Warburton, D., De Filippo,R. E. & Perin, L. 2010 Human amniotic fluid as a potentialnew source of organ specific precursor cells for future regen-erative medicine applications. J. Urol. 183, 1193–1200.(doi:10.1016/j.juro.2009.11.006)
De Coppi, P. et al. 2007a Isolation of amniotic stem cell lineswith potential for therapy. Nat. Biotechnol. 25, 100–106.(doi:10.1038/nbt1274)
De Coppi, P. et al. 2007b Amniotic fluid and bone marrowderived mesenchymal stem cells can be converted tosmooth muscle cells in the cryo-injured rat bladder andprevent compensatory hypertrophy of surviving smoothmuscle cells. J. Urol. 177, 369–376.
Degistirici, O., Jager, M. & Knipper, A. 2008 Applicability ofcord blood-derived unrestricted somatic stem cells in tissueengineering concepts. Cell Prolif. 41, 421–440. (doi:10.1111/j.1365-2184.2008.00536.x)
de la Fuente, J., Fisk, N., O’Donoghue, K., Chan, J., Kumar,S. & Roberts, I. 2003 a2b1 and a4b1 integrins mediate thehoming of mesenchymal stem/progenitor cells during fetallife. Haematol. J. 4, 13.
Delaney, C., Heimfeld, S., Brashem-Stein, C., Voorhies, H.,Manger, R. & Bernstein, I. 2010 Notch-mediated expan-sion of human cord blood progenitor cells capable ofrapid myeloid reconstitution. Nat. Med. 16, 232–236.(doi:10.1038/nm.2080)
Delo, D. et al. 2010 Calcification after myocardial infarction isindependent of amniotic fluid stem cell injection.Cardiovasc. Pathol. (doi:10.1089/scd.2008.0028)
Ditadi, A. et al. 2009 Human and murine amniotic fluidc-Kitþ Lin2 cells display hematopoietic activity. Blood113, 3953–3960. (doi:10.1182/blood-2008-10-182105)
Elwan, M. A. & Sakuragawa, N. 1997 Evidence for synthesisand release of catecholamines by human amniotic epi-thelial cells. Neuroreport 8, 3435–3438. (doi:10.1097/00001756-199711100-00004)
Ersek, A., Pixley, J., Goodrich, A., Porada, C.,Almeida-Porada, G., Thain, D. & Zanjani, E. 2010 Persist-ent circulating human insulin in sheep transplanted inutero with human mesenchymal stem cells. Exp. Hematol.38, 311–320. (doi:10.1016/j.exphem.2010.02.005)
Fong, C. Y., Richards, M., Manasi, N., Biswas, A. & Bongso,A. 2007 Comparative growth behaviour and characteriz-ation of stem cells from human Wharton’s jelly. Reprod.Biomed. Online 15, 708–718. (doi:10.1016/S1472-6483(10)60539-1)
J. R. Soc. Interface
Fu, Y.-S., Cheng, Y.-C., Lin, M.-Y. A., Cheng, H., Chu, P.-M.,Chou, S.-C., Shih, Y.-H., Ko, M.-H. & Sung, S. 2006 Con-version of human umbilical cord mesenchymal stem cells inWharton’s jelly to dopaminergic neurons in vitro: potentialtherapeutic application for Parkinsonism. Stem Cells 24,115–124. (doi:10.1634/stemcells.2005-0053)
Fuchs, J. R., Kaviani, A., Oh, J.-T., LaVan, D., Udagawa, T.,Jennings, R. W., Wilson, J. M. & Fauza, D. O. 2004 Dia-phragmatic reconstruction with autologous tendonengineered from mesenchymal amniocytes. J. Pediatr.Surg. 39, 834–838. (doi:10.1016/j.jpedsurg.2004.02.014)
Fujimoto, K. L., Miki, T., Liu, L. J., Hashizume, R., Strom,S. C., Wagner, W. R., Keller, B. B. & Tobita, K. 2009Naive rat amnion-derived cell transplantation improvedleft ventricular function and reduced myocardial scar ofpostinfarcted heart. Cell Transplant. 18, 477–486.(doi:10.3727/096368909788809785)
Fukuchi, Y., Nakajima, H., Sugiyama, D., Hirose, I.,Kitamura, T. & Tsuji, K. 2004 Human placenta-derivedcells have mesenchymal stem/progenitor cell potential.Stem Cells 22, 649–658. (doi:10.1634/stemcells.22-5-649)
Galende, E. et al. 2009 Amniotic fluid cells are more efficientlyreprogrammed to pluripotency than adult cells. CellReprogram 12, 117–125. (doi:10.1089/cell.2009.0077)
Gammaitoni, L. et al. 2004 Elevated telomerase activity andminimal telomere loss in cord blood long-term cultureswith extensive stem cell replication. Blood 103, 4440–4448. (doi:10.1182/blood-2003-09-3079)
Giorgetti, A. et al. 2009 Generation of induced pluripotentstem cells from human cord blood using OCT4 andSOX2. Cell Stem Cell 5, 353–357. (doi:10.1016/j.stem.2009.09.008)
Giorgetti, A., Montserrat, N., Rodriguez-Piza, I., Azqueta, C.,Veiga, A. & Izpisua Belmonte, J. C. 2010 Generation ofinduced pluripotent stem cells from human cord bloodcells with only two factors: Oct4 and Sox2. Nat. Protoc.5, 811–820. (doi:10.1038/nprot.2010.16)
Gosden, C. & Brock, D. J. 1978 Combined use of alphafeto-protein and amniotic fluid cell morphology in earlyprenatal diagnosis of fetal abnormalities. J. Med. Genet.15, 262–270. (doi:10.1136/jmg.15.4.262)
Gotherstrom, C., Ringden, O., Tammik, C., Zetterberg, E.,Westgren, M. & Le Blanc, K. 2004 Immunologic propertiesof human fetal mesenchymal stem cells. Am. J. Obstet.Gynecol. 190, 239–245. (doi:10.1016/j.ajog.2003.07.022)
Grounds, M. D., Radley, H. G., Lynch, G. S., Nagaraju, K. &De Luca, A. 2008 Towards developing standard operatingprocedures for pre-clinical testing in the mdx mouse modelof Duchenne muscular dystrophy. Neurobiol. Dis. 31,1–19. (doi:10.1016/j.nbd.2008.03.008)
Gucciardo, L., Lories, R., Ochsenbein-Kolble, N., Done, E.,Zwijsen, A. & Deprest, J. 2009 Fetal mesenchymal stemcells: isolation, properties and potential use in perinatologyand regenerative medicine. Br. J. Obstet. Gynaecol. 116,166–172.
Guillot, P. V., O’Donoghue, K., Kurata, H. & Fisk, N. M.2006 Fetal stem cells: betwixt and between.Semin. Reprod. Med. 24, 340–347. (doi:10.1055/s-2006-952149)
Guillot, P. V., Cui, W., Fisk, N. M. & Polak, D. J. 2007a Stemcell differentiation and expansion for clinical applicationsof tissue engineering. J. Cell. Mol. Med. 11, 935–944.(doi:10.1111/j.1582-4934.2007.00106.x)
Guillot, P. V., Gotherstrom, C., Chan, J., Kurata, H. & Fisk,N. M. 2007b Human first-trimester fetal MSC expresspluripotency markers and grow faster and have longertelomeres than adult MSC. Stem Cells 25, 646–654.(doi:10.1634/stemcells.2006-0208)

14 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Guillot, P. et al. 2008a Intrauterine transplantation of humanfetal mesenchymal stem cells from first-trimester bloodrepairs bone and reduces fractures in osteogenesisimperfecta mice. Blood 111, 1717–1725. (doi:10.1182/blood-2007-08-105809)
Guillot, P., Cook, H., Pusey, C., Fisk, N., Harten, S., Moss, J.,Shore, I. & Bou-Gharios, G. 2008b Transplantation ofhuman fetal mesenchymal stem cells improves glomerulo-pathy in a collagen type I alpha 2-deficient mouse.J. Pathol. 214, 627–636. (doi:10.1002/path.2325)
Habich, A., Jurga, M., Markiewicz, I., Lukomska, B.,Bany-Laszewicz, U. & Domanska-Janik, K. 2006 Earlyappearance of stem/progenitor cells with neural-likecharacteristics in human cord blood mononuclear fractioncultured in vitro. Exp. Hematol. 34, 914–925. (doi:10.1016/j.exphem.2006.03.010)
Harrison, D., Zhong, R., Jordan, C., Lemischka, I. & Astle, C.1997 Relative to adult marrow, fetal liver repopulatesnearly five times more effectively long-term thanshort-term. Exp. Hematol. 25, 293–297.
Horwitz, E. M., Gordon, P. L., Koo, W. K. K., Marx, J. C.,Neel, M. D., McNall, R. Y., Muul, L. & Hofmann, T.2002 Isolated allogeneic bone marrow-derived mesenchy-mal cells engraft and stimulate growth in children withosteogenesis imperfecta: implications for cell therapy ofbone. Proc. Natl Acad. Sci. USA 99, 8932–8937. (doi:10.1073/pnas.132252399)
Hsieh, J. Y., Fu, Y. S., Chang, S. J., Tsuang, Y. H. & Wang,H. W. 2010 Functional module analysis reveals differentialosteogenic and stemness potentials in human mesenchymalstem cells from bone marrow and Wharton’s jelly of umbi-lical cord. Stem Cells Dev. (doi:10.1089/scd.2009.0485)
Hu, Y., Liao, L., Wang, Q., Ma, L., Ma, G., Jiang, X. & Zhao,R. C. 2003 Isolation and identification of mesenchymalstem cells from human fetal pancreas. J. Lab. Clin. Med.141, 342–349. (doi:10.1016/S0022-2143(03)00022-2)
Huang, H. 2007 Isolation of human placenta-derived multi-potent cells and in vitro differentiation into hepatocyte-like cells. Curr. Protoc. Stem Cell Biol. 1, 1E.1. (doi:10.1002/9780470151808.sc01e01s1)
Huang, G. T., Gronthos, S. & Shi, S. 2009 Mesenchymal stemcells derived from dental tissues vs. those from othersources: their biology and role in regenerative medicine.J. Dent. Res. 88, 792–806.
Huangfu, D., Osafune, K., Maehr, R., Guo, W., Eijkelenboom,A., Chen, S., Muhlestein, W. & Melton, D. 2008 Inductionof pluripotent stem cells from primary human fibroblastswith only Oct4 and Sox2. Nat. Biotechnol. 26,1269–1275. (doi:10.1038/nbt.1502)
Huss, R. 2000 Isolation of primary and immortalized CD34-hematopoietic and mesenchymal stem cells from varioussources. Stem Cells 18, 1–9. (doi:10.1634/stemcells.18-1-1)
Hwang, W. Y. K., Samuel, M., Tan, D., Koh, L. P., Lim, W. &Linn, Y. C. 2007 A meta-analysis of unrelated donor umbi-lical cord blood transplantation versus unrelated donorbone marrow transplantation in adult and pediatricpatients. Biol. Blood Marrow Transplant. 13, 444–453.(doi:10.1016/j.bbmt.2006.11.005)
Igura, K., Zhang, X., Takahashi, K., Mitsuru, A., Yamaguchi,S. & Takashi, T. A. 2004 Isolation and characterization ofmesenchymal progenitor cells from chorionic villi of humanplacenta. Cytotherapy 6, 543–553. (doi:10.1080/14653240410005366-1)
Ilancheran, S., Michalska, A., Peh, G., Wallace, E. M., Pera,M. & Manuelpillai, U. 2007 Stem cells derived fromhuman fetal membranes display multilineage differen-tiation potential. Biol. Reprod. 77, 577–588. (doi:10.1095/biolreprod.106.055244)
J. R. Soc. Interface
Ilancheran, S., Moodley, Y. & Manuelpillai, U. 2009 Humanfetal membranes: a source of stem cells for tissue regener-ation and repair? Placenta 30, 2–10. (doi:10.1016/j.placenta.2008.09.009)
Insausti, C., Blanquer, M., Bleda, P., Iniesta, P., Majado, M.,Castellanos, G. & Moraleda, J. 2010 The amnioticmembrane as a source of stem cells. Histol. Histopathol. 25,91–98.
in‘t Anker, P. S. et al. 2003a Mesenchymal stem cells inhuman second-trimester bone marrow, liver, lung, andspleen exhibit a similar immunophenotype but a hetero-geneous multilineage differentiation potential.Haematologica 88, 845–852.
in‘t Anker, P. S., Scherjon, S. A., Kleijburg-van der Keur, C.,Noort, W. A., Claas, F. H., Willemze, R., Fibbe, W. E. &Kanhai, H. H. 2003b Amniotic fluid as a novel source ofmesenchymal stem cells for therapeutic transplantation.Blood 102, 1548–1549.
in‘t Anker, P. S., Scherjon, S. A., Kleijburg-van der Keur, C.,de Groot-Swings, G. M., Claas, F. H., Fibbe, W. E. &Kanhai, H. H. 2004 Isolation of mesenchymal stemcells of fetal or maternal origin from human placenta.Stem Cells 22, 1338–1345. (doi:10.1634/stemcells.2004-0058)
Jaenisch, R. 2009 Stem cells, pluripotency and nuclearreprogramming. J. Thromb. Haemost. 7(Suppl. 1), 21–23.
Javazon, E., Beggs, K. & Flake, A. 2004 Mesenchymal stemcells: paradoxes of passaging. Exp. Hematol. 32,414–425. (doi:10.1016/j.exphem.2004.02.004)
Jomura, S., Uy, M., Mitchell, K., Dallasen, R., Bode, C. & Xu,Y. 2007 Potential treatment of cerebral global ischemiawith Oct-4þ umbilical cord matrix cells. Stem Cells 25,98–106. (doi:10.1634/stemcells.2006-0055)
Jones, H. M., Nathrath, M., Thomas, R. M., Edelman, P.,Rodeck, C. H. & Linch, D. C. 1994 The effects of gestationon circulating progenitor cells. Br. J. Haematol. 87,637–639. (doi:10.1111/j.1365-2141.1994.tb08329.x)
Kakishita, K., Elwan, M. A., Nakao, N., Itakura, T. &Sakuragawa, N. 2000 Human amniotic epithelial cells pro-duce dopamine and survive after implantation into thestriatum of a rat model of Parkinson’s disease: a potentialsource of donor for transplantation therapy. Exp. Neurol.165, 27–34. (doi:10.1006/exnr.2000.7449)
Kakishita, K., Nakao, N., Sakuragawa, N. & Itakura, T. 2003Implantation of human amniotic epithelial cells preventsthe degeneration of nigral dopamine neurons in rats with6-hydroxydopamine lesions. Brain Res. 980, 48–56.(doi:10.1016/S0006-8993(03)02875-0)
Karahuseyinoglu, S., Cinar, O., Kilic, E., Kara, F., Akay,G. G., Demiralp, D. O., Tukun, A., Uckan, D. & Can, A.2007 Biology of stem cells in human umbilical cordstroma: in situ and in vitro surveys. Stem Cells 25,319–331. (doi:10.1634/stemcells.2006-0286)
Kaviani, A., Perry, T. E., Dzakovic, A., Jennings, R. W.,Ziegler, M. M. & Fauza, D. O. 2001 The amniotic fluidas a source of cells for fetal tissue engineering. J. Pediatr.Surg. 36, 1662–1665. (doi:10.1053/jpsu.2001.27945)
Kennea, N. et al. 2009 Differentiation of human fetal mesench-ymal stem cells into cells with an oligodendrocytephenotype. Cell Cycle 8, 1069–1079.
Khan, A. A. et al. 2010 Human fetal liver derived stem celltransplantation as supportive modality in the managementof end stage decompensated liver cirrhosis. Cell Transplant.19, 409–418.
Kim, J., Lee, Y., Kim, H., Hwang, K. J., Kwon, H. C., Kim,S. K., Cho, D. J., Kang, S. G. & You, J. 2007 Humanamniotic fluid-derived stem cells have characteristics ofmultipotent stem cells. Cell Prolif. 40, 75–90.

Review. Characteristics of stem cells H. Abdulrazzak et al. 15
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Kim, S. M. et al. 2008 Gene therapy using TRAIL-secretinghuman umbilical cord blood-derived mesenchymal stemcells against intracranial glioma. Cancer Res. 68,9614–9623. (doi:10.1158/0008-5472.CAN-08-0451)
Kim, J. et al. 2009a Oct4-induced pluripotency in adultneural stem cells. Cell 136, 411–419. (doi:10.1016/j.cell.2009.01.023)
Kim, J. B., Greber, B., Arauzo-Bravo, M. J., Meyer, J., Park,K. I., Zaehres, H. & Scholer, H. R. 2009b Direct reprogram-ming of human neural stem cells by OCT4. Nature 461,643–649. (doi:10.1038/nature08436)
Klimanskaya, I., Chung, Y., Becker, S., Lu, S. J. & Lanza, R.2007 Derivation of human embryonic stem cells from singleblastomeres. Nat. Protoc. 2, 1963–1972. (doi:10.1038/nprot.2007.274)
Koizumi, N. J., Inatomi, T. J., Sotozono, C. J., Fullwood,N. J., Quantock, A. J. & Kinoshita, S. 2000 Growthfactor mRNA and protein in preserved human amnioticmembrane. Curr. Eye. Res. 20, 173–177.
Kolambkar, Y. M., Peister, A., Soker, S., Atala, A. &Guldberg, R. E. 2007 Chondrogenic differentiation ofamniotic fluid-derived stem cells. J. Mol. Histol. 38,405–413. (doi:10.1007/s10735-007-9118-1)
Kong, X.-Y. et al. 2008 Transplantation of human amnioticcells exerts neuroprotection in MPTP-induced Parkinsondisease mice. Brain Res. 1205, 108–115. (doi:10.1016/j.brainres.2008.02.040)
Kubo, M., Sonoda, Y., Muramatsu, R. & Usui, M. 2001Immunogenicity of human amniotic membrane in exper-imental xenotransplantation. Invest. Ophthalmol. Vis.Sci. 42, 1539–1546.
Kucia, M., Zuba-Surma, E., Wysoczynski, M., Dobrowolska, H.,Reca, R., Ratajczak, J. & Ratajczak, M. 2006 Physiologicaland pathological consequences of identification of verysmall embryonic like (VSEL) stem cells in adult bonemarrow. J. Physiol. Pharmacol. 57(Suppl. 5), 5–18.
Kucia, M. et al. 2007 Morphological and molecular character-ization of novel population of CXCR4þ SSEA-4þ Oct-4þvery small embryonic-like cells purified from human cordblood: preliminary report. Leukemia 21, 297–303.(doi:10.1038/sj.leu.2404470)
Kunisaki, S. M., Jennings, R. W. & Fauza, D. O. 2006 Fetalcartilage engineering from amniotic mesenchymal progeni-tor cells. Stem Cells Dev. 15, 245–253. (doi:10.1089/scd.2006.15.245)
Kunisaki, S. M., Armant, M., Kao, G. S., Stevenson, K., Kim,H. & Fauza, D. O. 2007 Tissue engineering from humanmesenchymal amniocytes: a prelude to clinical trials.J. Pediatr. Surg. 42, 974–979. (doi:10.1016/j.jpedsurg.2007.01.031)
Kyo, S. et al. 1997 Expression of telomerase activity in humanchorion. Biochem. Biophys. Res. Commun. 241, 498–503.(doi:10.1006/bbrc.1997.7767)
Le Blanc, K. et al. 2005 Fetal mesenchymal stem-cell engraft-ment in bone after in utero transplantation in a patientwith severe osteogenesis imperfecta. Transplantation 79,1607–1614. (doi:10.1097/01.TP.0000159029.48678.93)
Le Blanc, K. et al. 2008 Mesenchymal stem cells for treatmentof steroid-resistant, severe, acute graft-versus-host disease:a phase II study. Lancet 371, 1579–1586. (doi:10.1016/S0140-6736(08)60690-X)
Li, C., Zhang, W., Jiang, X. & Mao, N. 2007 Human-placenta-derived mesenchymal stem cells inhibit proliferation andfunction of allogeneic immune cells. Cell Tissue Res.330, 437–446. (doi:10.1007/s00441-007-0504-5)
Li, C. et al. 2009 Pluripotency can be rapidly and efficientlyinduced in human amniotic fluid-derived cells. Hum. Mol.Genet. 18, 4340–4349. (doi:10.1093/hmg/ddp386)
J. R. Soc. Interface
Lim, F. T. H., Kanhai, H. H. H. & Falkenburg, J. H. F. 2005Characterization of the human CD34þ hematopoietic pro-genitor cell compartment during the second trimester ofpregnancy. Haematologica 90, 173–179.
Liu, T., Wu, J., Huang, Q., Hou, Y., Jiang, Z., Zang, S. &Guo, L. 2008 Human amniotic epithelial cells amelioratebehavioral dysfunction and reduce infarct size in therat middle cerebral artery occlusion model. Shock 29,603–611.
Lo, B. & Parham, L. 2009 Ethical issues in stem cell research.Endocr. Rev. 30, 204–213. (doi:10.1210/er.2008-0031)
Loebinger, M. & Janes, S. 2010 Stem cells as vectors for anti-tumour therapy. Thorax 65, 362–369. (doi:10.1136/thx.2009.128025)
Lund, R. D. et al. 2007 Cells isolated from umbilical cordtissue rescue photoreceptors and visual functions in arodent model of retinal disease. Stem Cells 25, 602–611.(doi:10.1634/stemcells.2006-0308)
MacKenzie, T. C., Campagnoli, C., Almeida-Porada, G., Fisk,N. M. & Flake, A. W. 2001 Circulating human fetal stro-mal cells engraft and differentiate in multiple tissuesfollowing transplantation into pre-immune fetal lambs.Blood 98, 798.
Maherali, N. et al. 2007 Directly reprogrammed fibroblastsshow global epigenetic remodeling and widespread tissuecontribution. Cell Stem Cell 1, 55–70. (doi:10.1016/j.stem.2007.05.014)
Manzur, A. Y. & Muntoni, F. 2009 Diagnosis and new treat-ments in muscular dystrophies. J. Neurol. Neurosurg.Psychiatry 80, 706–714.
Marcus, A. & Woodbury, D. 2008 Fetal stem cells from extra-embryonic tissues: do not discard. J. Cell. Mol. Med. 12,730–742. (doi:10.1111/j.1582-4934.2008.00221.x)
Mazurier, F., Doedens, M., Gan, O. & Dick, J. 2003Rapid myeloerythroid repopulation after intrafemoraltransplantation of NOD–SCID mice reveals a new classof human stem cells. Nat. Med. 9, 959–963. (doi:10.1038/nm886)
McElreavey, K., Irvine, A., Ennis, K. & McLean, W. 1991 Iso-lation, culture and characterisation of fibroblast-like cellsderived from the Wharton’s jelly portion of human umbi-lical cord. Biochem. Soc. Trans. 19, 29S.
McGrath, K. & Palis, J. 2008 Ontogeny of erythropoiesis inthe mammalian embryo. Curr. Top. Dev. Biol. 82, 1–22.(doi:10.1016/S0070-2153(07)00001-4)
Miao, Z., Jin, J., Chen, L., Zhu, J., Huang, W., Zhao, J., Qian,H. & Zhang, X. 2006 Isolation of mesenchymal stem cellsfrom human placenta: comparison with human bonemarrow mesenchymal stem cells. Cell Biol. Int. 30, 681–687. (doi:10.1016/j.cellbi.2006.03.009)
Miki, T. & Strom, S. 2006 Amnion-derived pluripotent/multi-potent stem cells. Stem Cell Rev. 2, 133–142. (doi:10.1007/s12015-006-0020-0)
Miki, T., Lehmann, T., Cai, H., Stolz, D. & Strom, S. 2005Stem cell characteristics of amniotic epithelial cells. StemCells 23, 1549–1559. (doi:10.1634/stemcells.2004-0357)
Miki, T., Mitamura, K., Ross, M., Stolz, D. & Strom, S. 2007Identification of stem cell marker-positive cells by immuno-fluorescence in term human amnion. J. Reprod. Immunol.75, 91–96. (doi:10.1016/j.jri.2007.03.017)
Miki, T., Marongiu, F., Dorko, K., Ellis, E. & Strom, S. 2010Isolation of amniotic epithelial stem cells. Curr. Protoc.Stem Cell Biol. 12, 1E.3.1–1E.3.10. (doi:10.1002/9780470151808.sc01e03s12)
Mitchell, K. E. et al. 2003 Matrix cells from Wharton’s jellyform neurons and glia. Stem Cells 21, 50–60.
Moon, J. H., Lee, J. R., Jee, B. C., Suh, C. S., Kim, S. H., Lim,H. J. & Kim, H. K. 2008 Successful vitrification of human

16 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
amnion-derived mesenchymal stem cells. Hum. Reprod.23, 1760–1770. (doi:10.1093/humrep/den202)
Mountford, J. 2008 Human embryonic stem cells: origins,characteristics and potential for regenerative therapy.Transfus. Med. 18, 1–12. (doi:10.1111/j.1365-3148.2007.00807.x)
Musina, R., Bekchanova, E., Belyavskii, A., Grinenko, T. &Sukhikh, G. 2007 Umbilical cord blood mesenchymalstem cells. Bull. Exp. Biol. Med. 143, 127–131. (doi:10.1007/s10517-007-0032-z)
Nadri, S. & Soleimani, M. 2007 Comparative analysis ofmesenchymal stromal cells from murine bone marrow andamniotic fluid. Cytotherapy 9, 729–737. (doi:10.1080/14653240701656061)
Nava, S., Westgren, M., Jaksch, M., Tibell, A., Broome, U.,Ericzon, B. & Sumitran-Holgersson, S. 2005 Characterizationof cells in the developing human liver.Differentiation73, 249–260. (doi:10.1111/j.1432-0436.2005.00019.x)
O’Donoghue, K. & Fisk, N. M. 2004 Fetal stem cells. BestPract. Res. Clin. Obstet. Gynaecol. 18, 853–875.
Osawa, M., Hanada, K., Hamada, H. & Nakauchi, H. 1996Long-term lymphohematopoietic reconstitution by asingle CD34-low/negative hematopoietic stem cell. Science273, 242–245. (doi:10.1126/science.273.5272.242)
Owen, M. & Friedenstein, A. J. 1988 Stromal stem cells:marrow-derived osteogenic precursors. Ciba Found.Symp. 136, 42–60.
Pan, H., Yang, D., Chiu, Y., Lai, S., Wang, Y., Chang, M. &Cheng, F. 2006 Enhanced regeneration in injured sciaticnerve by human amniotic mesenchymal stem cell. J. Clin.Neurosci. 13, 570–575. (doi:10.1016/j.jocn.2005.06.007)
Pan, H., Cheng, F., Chen, C., Lai, S., Lee, C., Yang, D.,Chang, M. & Ho, S. 2007 Post-injury regeneration in ratsciatic nerve facilitated by neurotrophic factors secretedby amniotic fluid mesenchymal stem cells. J. Clin. Neuro-sci. 14, 1089–1098. (doi:10.1016/j.jocn.2006.08.008)
Pan, H. C., Yang, D. Y., Ho, S. P., Sheu, M. L., Chen, C. J.,Hwang, S. M., Chang, M. H. & Cheng, F. C. 2009Escalated regeneration in sciatic nerve crush injuryby the combined therapy of human amniotic fluidmesenchymal stem cells and fermented soybean extracts,Natto. J. Biomed. Sci. 16, 75. (doi:10.1186/1423-0127-16-75)
Pappa, K. & Anagnou, N. 2009 Novel sources of fetal stemcells: where do they fit on the developmental continuum?Regen. Med. 4, 423–433. (doi:10.2217/rme.09.12)
Parolini, O. et al. 2008 Concise review: isolation and charac-terization of cells from human term placenta: outcome ofthe first international Workshop on Placenta DerivedStem Cells. Stem Cells 26, 300–311. (doi:10.1634/stem-cells.2007-0594)
Parolini, O. et al. 2010 Toward cell therapy using placenta-derived cells: disease mechanisms, cell biology, preclinicalstudies, and regulatory aspects at the round table. StemCells Dev. 19, 143–154. (doi:10.1089/scd.2009.0404)
Perin, L. et al. 2010 Protective effect of human amniotic fluidstem cells in an immunodeficient mouse model of acutetubular necrosis. PLoS ONE 5, e9357. (doi:10.1371/jour-nal.pone.0009357)
Phillips, C., Pfeiffer, B., Luger, A. & Franklin, C. 2002 Novelcollagen glomerulopathy in a homotrimeric type I collagenmouse (oim). Kidney Int. 62, 383–391. (doi:10.1046/j.1523-1755.2002.00451.x)
Pittenger, M. F. et al. 1999 Multilineage potential of adulthuman mesenchymal stem cells. Science 284, 143–147.(doi:10.1126/science.284.5411.143)
Poloni, A. et al. 2008 Characterization and expansion ofmesenchymal progenitor cells from first-trimester chorionic
J. R. Soc. Interface
villi of human placenta. Cytotherapy 10, 690–697. (doi:10.1080/14653240802419310)
Porada, C. D., Zanjani, E. D. & Almeida-Porad, G. 2006Adult mesenchymal stem cells: a pluripotent populationwith multiple applications. Curr. Stem Cell Res. Ther. 1,365–369.
Portmann-Lanz, C. B., Schoeberlein, A., Huber, A., Sager, R.,Malek, A., Holzgreve, W. & Surbek, D. V. 2006 Placentalmesenchymal stem cells as potential autologous graft forpre- and perinatal neuroregeneration. Am. J. Obstet.Gynecol. 194, 664–673. (doi:10.1016/j.ajog.2006.01.101)
Portmann-Lanz, C. B., Schoeberlein, A., Portmann, R.,Mohr, S., Rollini, P., Sager, R. & Surbek, D. 2010 Turningplacenta into brain: placental mesenchymal stem cellsdifferentiate into neurons and oligodendrocytes.Am. J. Obstet. Gynecol. 202, 294.e1–294.e11. (doi:10.1016/j.ajog.2009.10.893)
Pranke, P., Hendrikx, J., Debnath, G., Alespeiti, G.,Rubinstein, P., Nardi, N. & Visser, J. 2005 Immunopheno-type of hematopoietic stem cells from placental/umbilicalcord blood after culture. Braz. J. Med. Biol. Res. 38,1775–1789.
Prat-Vidal, C. et al. 2007 Umbilical cord blood-derived stemcells spontaneously express cardiomyogenic traits. Trans-plant. Proc. 39, 2434–2437.
Prusa, A. R. & Hengstschlager, M. 2002 Amniotic fluid cellsand human stem cell research: a new connection. Med.Sci. Monit. 8, RA253–RA257.
Prusa, A. R., Marton, E., Rosner, M., Bernaschek, G. &Hengstschlager, M. 2003 Oct-4-expressing cells inhuman amniotic fluid: a new source for stem cellresearch? Hum. Reprod. 18, 1489–1493. (doi:10.1093/humrep/deg279)
Rachakatla, R. S., Marini, F., Weiss, M. L., Tamura, M. &Troyer, D. 2007 Development of human umbilical cordmatrix stem cell-based gene therapy for experimentallung tumors. Cancer Gene Ther. 14, 828–835. (doi:10.1038/sj.cgt.7701077)
Rebel, V., Miller, C., Eaves, C. & Lansdorp, P. 1996 The repo-pulation potential of fetal liver hematopoietic stem cellsin mice exceeds that of their liver adult bone marrowcounterparts. Blood 87, 3500–3507.
Rehni, A. K., Singh, N., Jaggi, A. S. & Singh, M. 2007 Amnio-tic fluid derived stem cells ameliorate focal cerebralischaemia-reperfusion injury induced behavioural deficitsin mice. Behav. Brain Res. 183, 95–100. (doi:10.1016/j.bbr.2007.05.028)
Riekstina, U., Cakstina, I., Parfejevs, V., Hoogduijn, M.,Jankovskis, G., Muiznieks, I., Muceniece, R. & Ancans, J.2009 Embryonic stem cell marker expression pattern inhuman mesenchymal stem cells derived from bonemarrow, adipose tissue, heart and dermis. Stem Cell Rev.5, 378–386. (doi:10.1007/s12015-009-9094-9)
Rollini, P., Kaiser, S., Faes-van’t Hull, E., Kapp, U. &Leyvraz, S. 2004 Long-term expansion of transplantablehuman fetal liver hematopoietic stem cells. Blood 103,1166–1170. (doi:10.1182/blood-2003-06-1815)
Roubelakis, M. G., Pappa, K. I., Bitsika, V., Zagoura, D.,Vlahou, A., Papadaki, H. A., Antsaklis, A. & Anagnou,N. P. 2007 Molecular and proteomic characterization ofhuman mesenchymal stem cells derived from amnioticfluid: comparison to bone marrow mesenchymal stemcells. Stem Cells Dev. 16, 931–952. (doi:10.1089/scd.2007.0036)
Sakuragawa, N. et al. 1997 Evidence for active acetylcholinemetabolism in human amniotic epithelial cells: applicableto intracerebral allografting for neurologic disease. Neurosci.Lett. 232, 53–56. (doi:10.1016/S0304-3940(97)00570-3)

Review. Characteristics of stem cells H. Abdulrazzak et al. 17
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
Sakuragawa, N., Enosawa, S., Ishii, T., Thangavel, R.,Tashiro, T., Okuyama, T. & Suzuki, S. 2000 Humanamniotic epithelial cells are promising transgene carriersfor allogeneic cell transplantation into liver. J. Hum.Genet. 45, 171–176. (doi:10.1007/s100380050205)
Sakuragawa, N., Elwan, M. A., Uchida, S., Fujii, T. &Kawashima, K. 2001 Non-neuronal neurotransmittersand neurotrophic factors in amniotic epithelial cells:expression and function in humans and monkey. JpnJ. Pharmacol. 85, 20–23. (doi:10.1254/jjp.85.20)
Schmidt, D. et al. 2006 Living autologous heart valves engin-eered from human prenatally harvested progenitors.Circulation 114, I125–I131. (doi:10.1161/CIRCULATIO-NAHA.105.001040)
Schoemans, H., Theunissen, K., Maertens, J., Boogaerts, M.,Verfaillie, C. & Wagner, J. 2006 Adult umbilical cordblood transplantation: a comprehensive review. BoneMarrow Transplant. 38, 83–93. (doi:10.1038/sj.bmt.1705403)
Secco, M. et al. 2008 Multipotent stem cells from umbilicalcord: cord is richer than blood! Stem Cells 26, 146–150.(doi:10.1634/stemcells.2007-0381)
Sessarego, N. et al. 2008 Multipotent mesenchymal stromalcells from amniotic fluid: solid perspectives for clinicalapplication. Haematologica 93, 339–346. (doi:10.3324/haematol.11869)
Siegel, N., Rosner, M., Hanneder, M., Valli, A. & Hengstschlager,M. 2007 Stem cells in amniotic fluid as new tools to studyhuman genetic diseases. Stem Cell Rev. 3, 256–264. (doi:10.1007/s12015-007-9003-z)
Soncini, M., Vertua, E., Gibelli, L., Zorzi, F., Denegri, M.,Albertini, A., Wengler, G. S. & Parolini, O. 2007Isolation and characterization of mesenchymal cells fromhuman fetal membranes. J. Tissue Eng. Regen. Med. 1,296–305. (doi:10.1002/term.40)
Spitalieri, P. et al. 2009 Identification of multipotent cyto-trophoblast cells from human first trimester chorionicvilli. Cloning Stem Cells 11, 535–556. (doi:10.1089/clo.2009.0046)
Streubel, B., Martucci-Ivessa, G., Fleck, T. & Bittner, R. E.1996 In vitro transformation of amniotic cells to musclecells–background and outlook. Wien. Med. Wochenschr.146, 216–217.
Studeny, M., Marini, F. C., Dembinski, J. L., Zompetta, C.,Cabreira-Hansen, M., Bekele, B. N., Champlin, R. E. &Andreeff, M. 2004 Mesenchymal stem cells: potential pre-cursors for tumor stroma and targeted-delivery vehiclesfor anticancer agents. J. Natl Cancer Inst. 96, 1593–1603.
Sudo, K., Kanno, M., Miharada, K., Ogawa, S., Hiroyama, T.,Saijo, K. & Nakamura, Y. 2007 Mesenchymal progenitorsable to differentiate into osteogenic, chondrogenic,and/or adipogenic cells in vitro are present in mostprimary fibroblast-like cell populations. Stem Cells 25,1610–1617. (doi:10.1634/stemcells.2006-0504)
Sun, N. et al. 2009 Feeder-free derivation of induced pluri-potent stem cells from adult human adipose stem cells.Proc. Natl Acad. Sci. USA 106, 15 720–15 725. (doi:10.1073/pnas.0908450106)
Takahashi, K. & Yamanaka, S. 2006 Induction of pluripotentstem cells from mouse embryonic and adult fibroblast cul-tures by defined factors. Cell 126, 663–676. (doi:10.1016/j.cell.2006.07.024)
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka,T., Tomoda, K. & Yamanaka, S. 2007 Induction of pluripo-tent stem cells from adult human fibroblasts by definedfactors. Cell 131, 861–872. (doi:10.1016/j.cell.2007.11.019)
Tamagawa, T., Ishiwata, I. & Saito, S. 2004 Establishmentand characterization of a pluripotent stem cell line derived
J. R. Soc. Interface
from human amniotic membranes and initiation of germlayers in vitro. Hum. Cell. 17, 125–130.
Tamagawa, T., Oi, S., Ishiwata, I., Ishikawa, H. & Nakamura,Y. 2007 Differentiation of mesenchymal cells derived fromhuman amniotic membranes into hepatocyte-like cellsin vitro. Hum. Cell. 20, 77–84. (doi:10.1111/j.1749-0774.2007.00032.x)
Taylor, P. A., McElmurry, R. T., Lees, C. J., Harrison, D. E. &Blazar, B. R. 2002 Allogenic fetal liver cells have a distinctcompetitive engraftment advantage over adult bone marrowcells when infused into fetal as compared with adult severecombined immunodeficient recipients. Blood 99, 1870–1872. (doi:10.1182/blood.V99.5.1870)
Thomson, J. A., Itskovitz-Eldor, J., Shapiro, S. S., Waknitz,M. A., Swiergiel, J. J., Marshall, V. S. & Jones, J. M.1998 Embryonic stem cell lines derived from human blasto-cysts. Science 282, 1145–1147. (doi:10.1126/science.282.5391.1145)
Torricelli, F., Brizzi, L., Bernabei, P. A., Gheri, G., Di Lollo,S., Nutini, L., Lisi, E., Di Tommaso, M. & Cariati, E. 1993Identification of hematopoietic progenitor cells in humanamniotic fluid before the 12th week of gestation.Ital. J. Anat. Embryol. 98, 119–126.
Troyer, D. L. & Weiss, M. L. 2008 Wharton’s jelly-derivedcells are a primitive stromal cell population. Stem Cells26, 591–599. (doi:10.1634/stemcells.2007-0439)
Tsai, M. S., Lee, J. L., Chang, Y. J. & Hwang, S. M. 2004 Iso-lation of human multipotent mesenchymal stem cells fromsecond-trimester amniotic fluid using a novel two-stageculture protocol. Hum. Reprod. 19, 1450–1456. (doi:10.1093/humrep/deh279)
Tsai, M.-S., Hwang, S.-M., Tsai, Y.-L., Cheng, F.-C., Lee,J.-L. & Chang, J. 2006 Clonal amniotic fluid-derivedstem cells express characteristics of both mesenchymaland neural stem cells. Biol. Reprod. 74, 545–551.(doi:10.1095/biolreprod.105.046029)
Tsai, M. et al. 2007 Functional network analysis of the tran-scriptomes of mesenchymal stem cells derived fromamniotic fluid, amniotic membrane, cord blood, andbone marrow. Stem Cells 25, 2511–2523. (doi:10.1634/stemcells.2007-0023)
Uchida, S., Inanaga, Y., Kobayashi, M., Hurukawa, S., Araie,M. & Sakuragawa, N. 2000 Neurotrophic function of con-ditioned medium from human amniotic epithelial cells.J. Neurosci. Res. 62, 585–590. (doi:10.1002/1097-4547(20001115)62:4,585::AID-JNR13.3.0.CO;2-U)
Valli, A. et al. 2010 Embryoid body formation of humanamniotic fluid stem cells depends on mTOR. Oncogene29, 966–977. (doi:10.1038/onc.2009.405)
Vaziri, H. et al. 2010 Spontaneous reversal of the developmen-tal aging of normal human cells following transcriptionalreprogramming. Regen. Med. 5, 345–363. (doi:10.2217/rme.10.21)
Ventura, C. et al. 2007 Hyaluronan mixed esters of butyricand retinoic acid drive cardiac and endothelial fate interm placenta human mesenchymal stem cells and enhancecardiac repair in infarcted rat hearts. J. Biol. Chem. 282,14 243–14 252. (doi:10.1074/jbc.M609350200)
Wagers, A. J., Christensen, J. L. & Weissman, I. L. 2002 Cellfate determination from stem cells. Gene Ther. 9, 606–612.(doi:10.1038/sj.gt.3301717)
Wang, H.-S., Hung, S.-C., Peng, S.-T., Huang, C.-C., Wei,H.-M., Guo, Y.-J., Fu, Y.-S., Lai, M.-C. & Chen, C. 2004Mesenchymal stem cells in the Wharton’s jelly of thehuman umbilical cord. Stem Cells 22, 1330–1337.(doi:10.1634/stemcells.2004-0013)
Wang, J. et al. 2003 SCID-repopulating cell activity of humancord blood-derived CD34- cells assured by intra-bone

18 Review. Characteristics of stem cells H. Abdulrazzak et al.
on August 28, 2010rsif.royalsocietypublishing.orgDownloaded from
marrow injection. Blood 101, 2924–2931. (doi:10.1182/blood-2002-09-2782)
Wang, J., Wang, L., Wu, Y., Xiang, Y., Xie, C., Jia, B.,Harrington, J. & McNiece, I. 2004 Mesenchymal stem/progenitor cells in human umbilical cord blood as supportfor ex vivo expansion of CD34(þ) hematopoietic stem cellsand for chondrogenic differentiation. Haematologica 89,837–844.
Weiss, M. L. & Troyer, D. L. 2006 Stem cells in the umbilicalcord. Stem Cell Rev. 2, 155–162. (doi:10.1007/s12015-006-0022-y)
Weiss, M. L. et al. 2006 Human umbilical cord matrixstem cells: preliminary characterization and effect oftransplantation in a rodent model of Parkinson’sdisease. Stem Cells 24, 781–792. (doi:10.1634/stemcells.2005-0330)
Weissman, I. & Shizuru, J. 2008 The origins of theidentification and isolation of hematopoietic stemcells, and their capability to induce donor-specific trans-plantation tolerance and treat autoimmune diseases.Blood 112, 3543–3553. (doi:10.1182/blood-2008-08-078220)
Weissman, I., Anderson, D. & Gage, F. 2001 Stem andprogenitor cells: origins, phenotypes, lineage commitments,and transdifferentiations. Annu. Rev. Cell. Dev. Biol. 17,387–403. (doi:10.1146/annurev.cellbio.17.1.387)
Yao, Z., Qin, M., Liu, J., Chen, X. & Zhou, D. 2004 In vitrocultivation of human fetal pancreatic ductal stem cellsand their differentiation into insulin-producing cells.World J. Gastroenterol. 10, 1452–1456.
Yeh, Y. C. et al. 2010 Cellular cardiomyoplasty withhuman amniotic fluid stem cells: in vitro in vivo studies.Tissue Eng. Part A 16, 1925–1936. (doi:10.1089/ten.tea.2009.0728)
Yen, B. L. et al. 2005 Isolation of multipotent cells fromhuman term placenta. Stem Cells 23, 3–9. (doi:10.1634/stemcells.2004-0098)
J. R. Soc. Interface
Yen, B. L., Chien, C.-C., Chen, Y.-C., Chen, J.-T., Huang,J.-S., Lee, F.-K. & Huang, I. 2008 Placenta-derived multi-potent cells differentiate into neuronal and glial cells invitro. Tissue Eng. Part A 14, 9–17. (doi:10.1089/ten.a.2006.0352)
Yu, L. C., Wall, D. A., Sandler, E., Chan, K. W., Grayson, G. &Kletzel, M. 2001 Unrelated cord blood transplant experienceby the pediatric blood and marrow transplant consortium.Pediatr. Hematol. Oncol. 18, 235–245. (doi:10.1080/088800101750238531)
Yu, J. et al. 2007 Induced pluripotent stem cell lines derivedfrom human somatic cells. Science 318, 1917–1920.(doi:10.1126/science.1151526)
Zhang, Y., Li, C. D., Jiang, X. X., Li, H. L., Tang, P. H. &Mao, N. 2004 Comparison of mesenchymal stem cellsfrom human placenta and bone marrow. Chin. Med. J.(Engl) 117, 882–887.
Zhang, Z. Y., Teoh, S. H., Chong, M. S., Schantz, J. T., Fisk,N. M., Choolani, M. A. & Chan, J. 2009 Superior osteo-genic capacity for bone tissue engineering of fetalcompared with perinatal and adult mesenchymal stemcells. Stem Cells 27, 126–137. (doi:10.1634/stemcells.2008-0456)
Zhao, P., Ise, H., Hongo, M., Ota, M., Konishi, I. & Nikaido,T. 2005 Human amniotic mesenchymal cells have somecharacteristics of cardiomyocytes. Transplantation 79,528–535. (doi:10.1097/01.TP.0000149503.92433.39)
Zhao, Y., Wang, H. & Mazzone, T. 2006 Identification of stemcells from human umbilical cord blood with embryonic andhematopoietic characteristics. Exp. Cell Res. 312, 2454–2464. (doi:10.1016/j.yexcr.2006.04.008)
Zuba-Surma, E. K., Klich, I., Greco, N., Laughlin, M. J.,Ratajczak, J. & Ratajczak, M. Z. 2010 Optimization ofisolation and further characterization of umbilical-cord-blood-derived very small embryonic/ epiblast-like stemcells (VSELs). Eur. J. Haematol. 84, 34–46. (doi:10.1111/j.1600-0609.2009.01352.x)