
A Structural and Immunological Comparison of Rickettsial HSP60 Antigens with Those of Other Species,...
18
A Structural and Immunological Comparison of Rickettsial HSP60 Antigens with Those of Other Speciesayb GREGORY A. DASCH,',d WEI-ME1 CHING,' PETER Y. KIM,' HUONG PHAM,' CHARLES K. STOVER,'.' EDWIN V. OAKS,' MICHAEL E. DOBSON,' AND EMILIO WEISS' 'Rickettsia1 Diseases Division Naval Medical Research Institute Bethesda, Maryland 20814-5055 and 'Department of Enteric Infections Walter Reed Army Institute of Research Washington, D.C. 20307 INTRODUCTION At a cellular level, the physiological responses of plants, animals, and both eukaryotic and prokaryotic microorganisms to a rapid increase in temperature (heat shock) are remarkably universal.' Generally, a select subset of proteins is produced in greatly increased amounts while the majority of other proteins de- cline in content. The proteins which are hyperinduced are known as heat-shock proteins (HSPS).~ Heat-shock proteins are thought to have an important role in protection of the cell against heat as well as the physiological stresses, including exposure to heavy metal ions or other toxins, ultraviolet light, or ethanol, which also induce synthesis of heat-shock proteins. Seventeen HSP have been identified in Escherichia cok3 Under heat-shock conditions, a number of very different bacteria have been shown to synthesize a set of proteins with molecular weights remarkably similar to those of the HSP initially found in E. ~oli.~-* Strikingly, by DNA sequence analysis of genes encoding HSP, by comparison of physical prop- erties of the proteins themselves, and, indirectly, by demonstration of conserved antigenic domains with monoclonal and polyclonal antibodies, a number of these proteins have also been shown to be highly conserved between eukaryotes and prokaryotes. Such conservation has most lnotably been observed for proteins related to the E. coli DnaK protein {equivalent to the eukaryotic HSP70 family of 'This work was supported in part by the Naval Medical Research and Develop- ment Command, Department of the Navy, work units 3M161102.BS13.AK.lll and 3M162770.A870.AN. 121. $he opinions and statements contained herein are the private ones of the authors and are not to be construed as official or reflecting the views of the Navy Department or the naval service at large. dAddress correspondence to Dr. Gregory A. Dasch, Naval Medical Research Institute, Bethesda, Maryland 20814-5055. fPresent address: Molecular Vaccines, Inc., 19 Firstfield Road, Gaithersburg, MD 20878. 352
Transcript of A Structural and Immunological Comparison of Rickettsial HSP60 Antigens with Those of Other Species,...

A Structural and Immunological Comparison of Rickettsial HSP60
Antigens with Those of Other Speciesayb GREGORY A. DASCH,',d WEI-ME1 CHING,'
PETER Y. KIM,' HUONG PHAM,' CHARLES K. STOVER,'.' EDWIN V. OAKS,' MICHAEL E. DOBSON,' AND
EMILIO WEISS' 'Rickettsia1 Diseases Division
Naval Medical Research Institute Bethesda, Maryland 20814-5055
and 'Department of Enteric Infections
Walter Reed Army Institute of Research Washington, D.C. 20307
INTRODUCTION
At a cellular level, the physiological responses of plants, animals, and both eukaryotic and prokaryotic microorganisms to a rapid increase in temperature (heat shock) are remarkably universal.' Generally, a select subset of proteins is produced in greatly increased amounts while the majority of other proteins de- cline in content. The proteins which are hyperinduced are known as heat-shock proteins (HSPS).~ Heat-shock proteins are thought to have an important role in protection of the cell against heat as well as the physiological stresses, including exposure to heavy metal ions or other toxins, ultraviolet light, or ethanol, which also induce synthesis of heat-shock proteins. Seventeen HSP have been identified in Escherichia cok3 Under heat-shock conditions, a number of very different bacteria have been shown to synthesize a set of proteins with molecular weights remarkably similar to those of the HSP initially found in E. ~ o l i . ~ - * Strikingly, by DNA sequence analysis of genes encoding HSP, by comparison of physical prop- erties of the proteins themselves, and, indirectly, by demonstration of conserved antigenic domains with monoclonal and polyclonal antibodies, a number of these proteins have also been shown to be highly conserved between eukaryotes and prokaryotes. Such conservation has most lnotably been observed for proteins related to the E. coli DnaK protein {equivalent to the eukaryotic HSP70 family of
'This work was supported in part by the Naval Medical Research and Develop- ment Command, Department of the Navy, work units 3M161102.BS13.AK.lll and 3M162770.A870.AN. 121.
$he opinions and statements contained herein are the private ones of the authors and are not to be construed as official or reflecting the views of the Navy Department or the naval service at large.
dAddress correspondence to Dr. Gregory A. Dasch, Naval Medical Research Institute, Bethesda, Maryland 20814-5055.
fPresent address: Molecular Vaccines, Inc., 19 Firstfield Road, Gaithersburg, MD 20878. 352

DASCH el al.: RICKETTSIAL HSP60 ANTIGENS 353
70-78-kDa proteins), the GroEL protein (equivalent to the eukaryotic HSP60 family of 58-65-kDa proteins), and the HtpG (or C62.5) protein (equivalent to the eukaryotic HSP90 family of 83-108-kDa protein^).^,^ In addition, although a re- lated E. coli protein has not been described, Mycobacterium leprae was shown recently to have a 19-kDa protein homologous to the small HSP family (16-40 kDa) of e u k a r y ~ t e s . ~ Finally, although related eukaryotic sequences have not yet been described, genes with significant DNA sequence homology to that of the E. coli GroES HSP have been described for prokaryotes as diverse as Mycobacte- rium tuberculosis, Chlamydia psittaci, Rickettsia tsutsugamushi, and Coxiella burnetii. It may be surmised that some of the other 13 E. coli HSPs will prove to he highly conserved among most prokaryotes, if not also in eukaryotic organ- isms.
Although the role of the HSP in responses to stressful environmental condi- tions has stimulated their investigation, it has become clear that many HSP also have important roles in normal cell physiology. A number of the E. coli HSP were originally discovered b‘ecause mutations in those genes of the E. coli host inter- fered with the growth or morphogenesis of bacteriophage.14 The finding that both HSP60 and HSP70 molecules participated in a wide range of assembly processes in both prokaryotic and eukaryotic cells lead to their description as “molecular chaperones.”15 Molecular chaperones interact with other proteins to prevent their aggregation and precipitation until their proper assembly can be effected but are not components of the final structure. This concept has been both broadened and extended to include for molecular chaperones a catalytic role as specific “protein unfoldases” that hydrolyze ATP during the assembly of oligomeric proteins and a role in maintaining an unfolded transition state in proteins, allowing them to be translocated across membranes. 16-18 In this context, the increase in synthesis of these proteins under conditions of heat shock or other stress may be seen as a mechanism to facilitate proper assembly or translocation of proteins under condi- tions which would otherwise lead to a great increase in the rate of protein dena- turation and a resulting loss of cell function.
A number of HSP have been shown to be essential (GroEL, GroES, rpoD) or conditionally essential (DnaK, DnaJ) for growth of E. coli under a variety of condition^.'^ The central role of the E. coli GroEL (HSP60) and GroES proteins is apparent in the pleiotropic effects of mutations in their genes on translocation of some proteins,20 on phage morphogenesi~,’~ on folding of both oligomeric and monomeric proteins ?1*22 and on proteolysis .23 The strong conservation of func- tions of the HSP60s is obvious in the heat inducibility and oligomeric structure of the Tetrahymena mitochondrial 58-kDa protein,24 the essential role of the yeast HSP60 protein in assembly of proteins imported into the m i t o c h ~ n d r i o n , ~ ~ ~ ~ ~ and the selective mutagenesis of the mammalian mitochondrial protein P1 in response to microtubule inhibitor^.^^,^^ In chloroplasts, HSP60 functions as a binding pro- tein which catalyzes the assembly of the oligomeric enzyme ribulose bisphosphate carboxylaseloxygenase, the key enzyme in the photosynthetic fixation of COz in plant^.^^,^^ No cytoplasmic homologues of HSP60s have been described in plants, animals, or lower eukaryotes. DNA sequence homologies of these proteins are in agreement with the endosymbiotic theory that mitochondria originated from pur- ple bacteria and chloroplasts from ~yanobacteria.~’
Although the existence of cross-reactive polysaccharide antigens has been known for many years, it was only in 1975 that K a i j ~ e r ~ ~ and H ~ i b y ~ ~ (antigen 10 of Pseudomonas) each described the widespread existence of a common major protein antigen in numerous bacterial genera. By means of crossed immunoelec- trophoretic techniques, Hoiby and his c o - ~ o r k e r s ~ ~ , ~ ~ extended these findings to

354 ANNALS NEW YORK ACADEMY OF SCIENCES
many other genera, including some gram-positive bacteria. In 1980, Sompolinsky et ~ 1 . ~ ~ 9 ~ ~ isolated the antigen from Pseudomonas aeruginosa and demonstrated that it had an apparent monomeric molecular weight of 62 kDa by SDS-polyacryl- amide gel electrophoresis but that it formed aggregates of 600-900 kDa. How- ever, it was not until 1987 that three groups of investigator^^*-^^ characterizing the genes encoding the 65-kDa protein antigens of Mycobacterium species realized that this 65-kDa protein was related to the E. coli GroEL protein and that each protein belonged to the ubiquitous common protein antigen family. The two pro- teins were highly immunoreactive molecules; they contained broadly reactive epitopes, as well as genus and species-specific epitopes, had similar amino acid compositions, formed oligomers of greater than 240 kDa; and their genes had very extensive regions of sequence homology.
The strong sequence similarity of bacterial HSP60s to the mitochondria1 HSP60s has stimulated considerable speculation about the potential of these anti- gens to trigger autoimmune responses, particularly rheumatoid a r th r i t i~ .~ I -~~ Al- ternatively, host cell HSPs may have important roles in immune surveillance and
Both T cell recognition of and antibody binding to epitopes on the 65-kDa protein of Mycobacterium leprae and M. tuberculosis have been in- vestigated extensively .47-50 The great antibody cross-reactivity among the bacte- rial common antigens has long stifled interest in these proteins for diagnostic purposes and has complicated contemporary efforts to employ them as protective vaccine antigens. However, monoclonal antibody and DNA sequence analyses of the HSP60s of bacteria have suggested that genus-specific, and occasionally spe- cies-specific, monoclonal antibodies can be obtained which recognize the variant regions of these protein^.^'-'^
The HSP60s of two Rickettsiaceae, Rickettsia tsutsugamushi and Coxiella burnetii, have been cloned and sequenced. 12~13 In addition, a number of rickettsial HSP60s have been purified and partially ~haracter ized.~~ Immune responses to these proteins have also been examined with both polyclonal and monoclonal a n t i b o d i e ~ . ' ~ . ~ ~ , ~ ~ Here we review these findings and speculate on the importance of the host immune responses to the rickettsial HSP60s during rickettsial infection and the function of these proteins in rickettsial physiology.
HSP60 ISOLATION AND CHEMICAL CHARACTERIZATION
Since all Rickettsiaceae except Rochalimaea are obligate intracellular para- sites, it is difficult and expensive to produce sufficient mass to isolate and chemi- cally characterize individual proteins from these species. For investigating the properties of the rickettsial HSP60s, we felt that prior experience in characteriz- ing the Rochalimaea proteins would be valuable, particularly since they are the closest free-living relatives known for the rickettsiae. Moreover, we had previ- ously noted the presence of a major protein band of 60 kDa on SDS-polyacryl- amide gels that was present in high concentration in the wash fluid of cell enve- lopes prepared from Rochalimaea quintana and R . vinsonii following French pressure-cell disruption of the organism^.^* Rochalimaea whole cells analyzed by Western blotting with polyclonal sera obtained from rabbits immunized with ty- phus group rickettsiae, Proteus, or Legionella often exhibited strong reactivity with a protein of this molecular weight. However, occasionally even normal rabbit or guinea pig sera also reacted with this band. In addition, similar molecular weight proteins were prominent in the SDS-polyacrylamide gel protein profiles of

DASCH et af.: RICKElTSIAL HSP60 ANTIGENS 355
both typhus and spotted fever group rickettsiae, and similar less abundant anti- gens could also be detected by Western blotting in three Proteus and eight Le- gionella strains. These observations strongly suggested that the 60-kDa protein of each of these species was its common bacterial antigen or HSP60.
To prove this hypothesis, we then applied the procedure described by Pau et aLS6 for the Legionella pneumophila common antigen to the purification of the Rochalimaea 60-kDa proteins (FIG. 1) and to similar proteins from a number of control species. The fractionation of the cells was monitored by dot blotting of crude fractions and by Coomassie blue staining and Western blotting of proteins analyzed by polyacrylamide gel electrophoresis. Yields of 60-kDa protein in each fraction and for each bacterium varied greatly. The wash of the cell envelope fraction generally contained the greatest portion of the 60-kDa antigen, although a significant portion was also present in the cytoplasmic and washed envelope fractions. A similar 60-kDa protein was also obtained in the envelope wash from Escherichia, Shigella, Proteus, Plesiomonas, Aeromonas, Pseudomonas, Rick- ettsia typhi, and R . prowazekii. Each of these proteins was further purified by gel filtration on a Pharmacia Superose-12 column. Some proteins were further puri- fied by ion exchange on a Mono Q column. The native molecular weights of proteins obtained in sufficient quantity and purity were determined on a Dupont GF-450 gel filtration column.
The native GroEL protein of E. coli was shown previously by electron micros- copy to consist of two rings of seven monomers each, one ring stacked above the other, with a nominal molecular mass of 840 kDa.57 A similar symmetry had also been observed for the pea chloroplast protein, which had a molecular mass of 900-950 kDa by gel filtration and equilibrium ~edimentat ion.~~ Values of 665 kDa were obtained for the Pseudomonas HSP60 by gel filtration, while higher values were obtained by electrophoresis and ultracentrifugation (800-900 kDa).37 By HPLC gel filtration, we observed native values of 680-750 kDa for the Profeus, Pseudomonas, Plesiomonas, Shigella, and Escherichia 60-kDa proteins. By HPLC gel filtration, Pau et ~ 1 . ~ ~ observed that the Legionella 60-kDa protein eluted at 650 kDa. However, we found that Aeromonas hydrophila 60-kDa protein eluted in two peaks of 620 and 840 kDa, respectively, while both Rochalimaea proteins had values of only 450 kDa, a value consistent with oligomers of 8 rather than 14 monomers. The Mycobacteriurn bovis HSP60 protein has a molecular weight of 246 kDa by gel filtration suggesting it is a tetramer.58 All of the bacterial species with 60-kDa proteins whose native sizes are compatible with the presence of tetradecamers are gamma-subdivision purple bacteria, while Rochalimaea be- longs to the alpha s u b d i ~ i s i o n . ~ ' , ~ ~ Consequently, the aggregation behavior of HSP6O proteins does not appear to be completely conserved.
To confirm the identity of the Rochaiimaea 60-kDa protein antigens as mem- bers of the HSP60 family, the amino acid compositions of R . quintana and R . uinsonii were compared to the compositions deduced from DNA sequences of HSP60 genes (TABLE 1). The compositions of the Rochalimaea proteins were very similar, even though consistent differences existed in the pattern of HSP60 fragments (presumably derived from protease activity during growth or purifica- tion) detected in each species by Western blotting. The amino acid compositions of the Rochalimaea 60-kDa proteins were entirely comparable to those of the HSP60 standards, with the exception of the amounts of tyrosine, phenylalanine, and arginine, which were somewhat higher. To provide more definitive identifica- tion of the 60-kDa proteins we have purified, automated amino-terminal amino acid sequence analyses were also done (TABLE 2). Three conclusions are apparent from these data. First, each of the proteins had sufficient homology to the E. coli
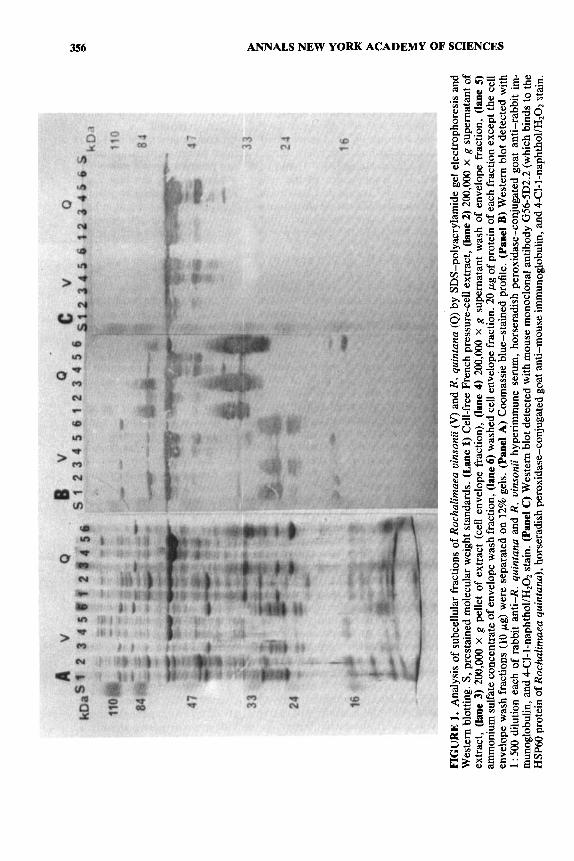
b
m e 0 * 0 E b
FIGURE 1. A
naly
sis
of s
ubce
llula
r fra
ctio
ns o
f R
ocha
limae
a vi
nson
ii (V) a
nd R
. qui
ntan
a (Q
) by
SDS-
poly
acry
lam
ide
gel e
lect
roph
ores
is a
nd
Wes
tern
blo
tting
. S, p
rest
aine
d m
olec
ular
wei
ght s
tand
ards
. (La
ne 1
) Cel
l-fr
ee F
renc
h pr
essu
re-c
ell
extr
act,
(lane
2)
200,
000
x g
supe
rnat
ant
of
extr
act,
(lane
3)
200,
000
x g
pelle
t of
ext
ract
(ce
ll en
velo
pe f
ract
ion)
, (la
ne 4
) 20
0,00
0 x
g su
pern
atan
t w
ash
of e
nvel
ope
frac
tion,
(la
ne 5
) am
mon
ium
sulf
ate
conc
entr
ate
of e
nvel
ope
was
h fr
actio
n, (l
ane 6
) was
hed
cell
enve
lope
frac
tion.
20
pg o
f pr
otei
n of
eac
h fr
actio
n ex
cept
the
cell
enve
lope
was
h fr
actio
ns (1
0 pg)
wer
e se
para
ted
on 1
2% g
els.
(Pa
nel A
) C
oom
assi
e bl
ue-s
tain
ed
prof
ile. (
Pane
l B)
Wes
tern
blo
t de
tect
ed w
ith
1 : 5
00 d
ilutio
n ea
ch o
f ra
bbit
anti-
R.
quin
tana
and
R.
vins
onii
hype
rim
mun
e se
rum
, ho
rser
adis
h pe
roxi
dase
-con
juga
ted
goat
ant
i-ra
bbit
im-
mun
oglo
bulin
, and
4-C
1-l-
naph
thol
/H20
2 sta
in. (
Pane
l C) W
este
rn b
lot d
etec
ted
with
mou
se m
onoc
lona
l ant
ibod
y G
56-5
D2.
2 (w
hich
bin
ds t
o th
e HSFW p
rote
in o
f Roc
halim
aea
qu
inta
na)
, hor
sera
dish
per
oxid
ase-
conj
ugat
ed g
oat a
nti-
mou
se i
mm
unog
lobu
lin, a
nd 4
43-1
-nap
htho
l/H
202 s
tain
.
rn '
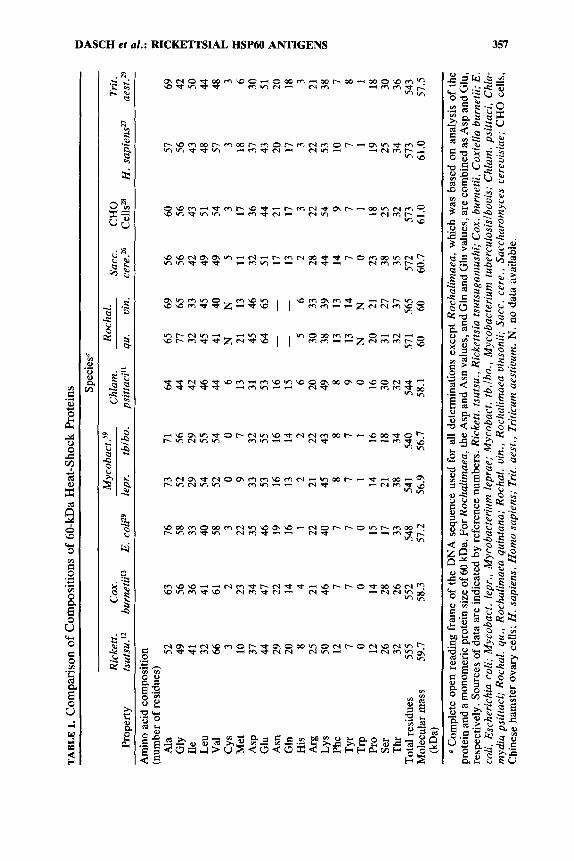
TABLE 1.
Com
paris
on o
f C
ompo
sitio
ns o
f 60
-kD
a H
eat-S
hock
Pro
tein
s Spec
ies"
Roc
ha'.
Sacc
. C
HO
Tr
it.
Myc
ob
a~
t.~
~ C
hlam
. R
icke
tt.
Cox
. Pr
ooer
tv
rsut
su.'2
bu
rnet
ii"
E. c
0P9
leor
. tb
lbo.
D
sitta
ci"
QU
. wi
n.
cere
.26
Cells
28
H. s
aDien
s2'
aest.
29
Am
ino
acid
com
posi
tion
(num
ber o
f re
sidu
es)
Ala
52
Ile
41
Leu
32
Val
66
CY
S 3
Met
10
AS
P 37
G
lu
44
Asn
29
G
ln
20
His
8
Arg
25
LY
S 50
Ph
e 12
TY
7
Trp
0
Pro
12
Ser
26
Thr
32
Tota
l res
idue
s 55
5 M
olec
ular
mas
s 59
.7
GlY
49
(kD
a)
63
56
36
41
61 2 23
34
47
22
14 4 21
46 7 7 0 14
28
26
552
58.3
76
58
33
40
58 3 22
35
46
19
16 1 22
40 7 7 0 15
17
33
548
57.2
73
52
29
54
52 0 9 33
53
16
13 2 21
45 8 7 1 14
21
38
54 1
56.9
71
56
29
55
54 0 7 32
55
16
14 2 22
43 8 7 1 16
18
34
540
56.7
64
44
42
46
44 6 13
31
53
16
15 6 20
49 8 9 0 16
30
32
544
58.1
65
77
32
45
41
N 21
45
64
-
- 5 30
38
13
13
N 20
31
32
57 1
60
69
65
33
45
40
N 13
46
65
-
- 6 33
39
13
14
N 21
27
37
565
60
56
56
42
49
49 5 11
32
51
17
13 2 28
44
14 7 0 23
38
35
572
60.7
60
56
43
51
54 3 17
36
44
21
17 3 22
54 9 7 1 18
25
32
573
61.0
57
56
43
48
57 3 18
37
43
20
17 3 22
53
10 7 1 19
25
34
573
61 .O
69
42
50
44
48 3 6 30
51
20
18 3 21
38 7 8 1 18
30
36
543
57.5
a C
ompl
ete
open
rea
ding
fra
me
of t
he D
NA
seq
uenc
e us
ed f
or a
ll de
term
inat
ions
exc
ept
Roc
halim
aea,
whi
ch w
as b
ased
on
anal
ysis
of
the
prot
ein
and
a m
onom
eric
pro
tein
siz
e of
60
kDa.
For
Roc
halim
aea,
the
Asp
and
Asn
val
ues,
and
Gln
and
Glu
val
ues,
are
com
bine
d as
Asp
and
Glu
, re
spec
tivel
y. S
ourc
es o
f da
ta a
re in
dica
ted
by re
fere
nce
num
bers
. Ric
kett.
tsut
su.,
Ric
ketts
ia t
suts
ugam
ushi
; C
ox. b
urne
tii, C
oxie
lla b
urne
tii; E
. co
li, E
sche
rich
ia c
oli;
Myc
obac
t. le
pr.,
Myc
obar
teri
um le
prae
; M
ycob
act.
tb.l
bo.,
Myc
obac
teri
um tu
berc
ulos
islb
ouis
; C
hlam
. psi
ttaci
, C
hla-
m
ydia
psi
ttaci
; R
ocha
l. q
u.,
Roc
halim
aea
quin
tana
; R
ocha
l. ui
n., R
ocha
limae
a vi
nson
ii; S
acc.
cer
e.,
Sacc
haro
myc
es c
ereu
isia
e; C
HO
cel
ls,
Chi
nese
ham
ster
ova
ry c
ells
; H. s
apie
ns, H
omo
sapi
ens;
Tri
t. ae
st.,
Triti
cum
aes
tiuur
n. N
, no
data
ava
ilabl
e.

TABL
E 2.
Com
paris
on o
f th
e A
min
o-T
erm
inal
Am
ino
Aci
d Se
quen
ces
of P
roka
ryot
ic 6
0-kD
a H
eat-
Shoc
k Pr
otei
ns
Bac
teriu
m
Sour
ce
Sequ
ence
" D
iff.
Esc
heri
chia
col
lz9
DN
A
MA
AK
DV
KF
GN
DA
RV
KM
LR
GV
NV
L
Ples
iom
onas
shi
gello
ides
Pr
otei
n ??
?DV
KF
GN
DA
RV
KM
LR
GV
NV
L
0 Sh
igel
la Jl
exne
ri
Prot
ein
???T
VK
FG
ND
AR
VK
ML
RG
VN
VL
1
Pseu
dom
onas
aer
ugin
osa
Prot
ein
AA
KE
VK
FG
MA
R&
KM
LV
?V
NV
L
5 C
hine
se h
amst
er o
vary
cell
s28
DN
A
-
YA
KD
VK
FG
AD
AR
UM
LQ
GV
DL
L
I H
omo
sapi
ensz
7 D
NA
Y
AK
DV
KF
GA
DA
RU
ML
QG
VD
LL
I
Roc
halim
aea
quin
tana
Pr
otei
n A
AK
EV
KF
Gm
AR
mL
RG
VW
L
8 R
ocha
limae
a ui
nson
ii Pr
otei
n A
AK
EV
KF
Gm
AR
mL
RG
VU
L
8
Ric
ketts
ia t
yphi
Pr
otei
n ?
? K
UK
HQ
?_K
? R
NM
>
I C
hlam
ydia
psi
ttaci
" D
NA
M
AA
KU
KY
NE
DA
R&
KE
KG
VK
TL
11
C
oxie
lla b
urne
tii"
DN
A
MA
AK
VL
KF
SH
EV
LH
AM
SR
GV
BV
L
11
Pisu
m s
ativ
um a
lpha
sub
unit6
1 Pr
otei
n A
AK
DU
FD
QH
SR
UM
QA
GD
12
Pi
sum
sat
ivum
bet
a su
buni
t61
Prot
ein
AK
UF
NK
DA
R.
KL
QN
GV
NE
L
12
Ric
ketts
ia p
row
azek
ii Pr
otei
n ?
? K
UK
HQ
? &A
R _E
? M
L
>6
Myc
obac
teri
um le
prae
" D
NA
M
.AK
TIA
YD
EE
AR
RG
LE
RG
LN
SL
13
M
ycob
acte
rium
tb.
lbov
iss9
D
NA
M
.AK
TIA
YD
EE
AR
RG
LE
RG
LN
AL
13
Sa
ccha
rom
yces
cer
euisi
ae26
D
NA
S
HK
BK
FG
VE
GR
AS
LL
KG
VX
L
13
Ric
ketts
ia t
suts
ugam
ushi
'2
DN
A
M.S
KQ
IVH
GD
QC
RK
K~
GIN
V~
14
Tr
iticu
m a
estiu
uma
DN
A
DA
KE
IAF
DQ
KS
RA
AL
QA
GV
EK
L
15
Syne
choc
occu
s 63
Ol6O
D
NA
M
TA
KR
IIY
NE
NA
RR
AL
EK
GU
L
15
Aer
orno
nas
hydr
ophi
la
Prot
ein
??
GA
GS
GK
VT
FN
GE
IIN
AP
?E
19
A
min
o ac
ids
diff
erin
g fr
om th
ose
foun
d at
the
corr
espo
ndin
g po
sitio
n in
the
E. c
oli
Gro
EL p
rote
in a
re u
nder
lined
and
enu
mer
ated
in
the
diff
eren
ce (
Diff
.) co
lum
n. T
hree
am
ino
acid
s re
mov
ed f
rom
the
P. s
ativ
um b
eta
subu
nit
sequ
ence
wer
e co
unte
d as
mis
mat
ches
. Th
e le
ader
se
quen
ces o
f se
vera
l euk
aryo
tic p
rote
ins
are
not d
ispl
ayed
.
.c 0 E

DASCH ef al.: RICKE’ITSIAL HSPQ ANTIGENS 359
HSP60 to establish its identity as an HSP60, with the exception of the Aeromonas hydrophila 60-kDa protein. The conservation of certain residues, such as the lysines at positions 4 and 7, the phenylalanine at 8, the arginine at 13, the glycine- valine pair at 19-20 and the leucine at 23, are particularly helpful in aligning and comparing sequences. Second, although HSP60s are believed to be among the most conserved proteins known, the amino terminus has undergone considerable alteration in evolution. Although the limited sequence available from this ap- proach only comprises about 4% of the protein, it is interesting that certain clusterings can be deduced from this information. A number of bacteria clustered by 16s rRNA sequence analysis in the gamma and alpha branches of purple b a ~ t e r i a ~ ~ ~ ~ ~ have very similar HSP60 amino-terminal sequences that diverge little from the mitochondrial HSP60 sequences. On the other hand, Coxiella burnetii is a gamma group member, but its HSP60 appears to have diverged to the same extent as Chlamydia HSP60, approaching the divergence seen in the Mycobacte- rium, cyanobacteria (Synechococcus), yeast mitochondrial, and chloroplast HSP60s. The close relationship of Rochalimaea quintana and R . uinsonii is con- firmed by the identity and uniqueness of their HSP60 sequences, as was also observed for the R . typhi and R . prowazekii HSP60s, although these data were less complete. Finally, the Rickettsia tsutsugamushi HSP60 sequence is quite divergent from that of all the other sequences, supporting its exclusion from the genus Rickettsia and the likelihood that it represents a distinct lineage of bacteria. These conclusions will be reexamined below by means of epitope analysis with monoclonal antibodies and the very limited set of DNA sequences of cloned HSP genes.
IMMUNOLOGICAL RELATIONSHIPS AMONG HSP60 ANTIGENS
Hindersson et summarized the results of crossed immunoelectrophoretic detection of the bacterial common antigen in 25 different genera of gram-negative, gram-positive, and spirochete bacteria. They suggested that the’ antigen might contain a number of phylogenetically stable epitopes and that normal human immune responses to bacterial flora might elicit antibodies and immune responses against this antigen that could comprise a type of “natural immunity” against pathogens sharing this antigen. The very extensive analyses conducted on both T cell- and antibody-mediated immune responses to the 65-kDa antigens of Myco- bacterium leprae and M . tuberculosis provide some support for at least the first of these ~iews.~’-~O At least 14 spatially separate epitopes have been defined by competition assays among monoclonal antibodies recognizing the HSP6O of M . l e p r ~ e . ~ ’ The reactivity of these antibodies for 23 species of mycobacteria was further determined. One antibody recognized only M. leprae, three recognized all species to varying degrees, while the others recognized at least eight of the spe- cies. A number of these antibodies have further been shown to react with a number of genera of Archaebacteria and both gram-positive and gram-negative genera, including Rickettsia r i~ke t t s i i .~~-~O Monospecific serum prepared against the Chlamydia psittaci HSP60 also cross-reacted with 57-60-kDa proteins from E. coli, Salmonella, Neisseria, Coxiella, Rickettsia rickettsii, Borrelia burgdor- feri, and Mycobacterium tuberculosis.” More than 80% of the sera from culture- positive or serologically confirmed human infections with Legionella pneumophila contained antibodies against the Legionella HSP60.* The Le- gionella HSP60 was shown to contain Legionella-specific and non-specific epi-

360 ANNALS NEW YORK ACADEMY OF SCIENCES
topes recognized by both human and rabbit antisera.65 Helsel et al.S2 found a monoclonal antibody against Legionella HSP60 that recognized 22 species of Legionella but only Acinetobacter IwofJii and Bordetella among 27 other bacterial species. Both conserved and specific epitopes on the HSP60 of Borrelia burgdor- feri were recognized by human serum from cases of acrodermatitis chronica atrophicans.%
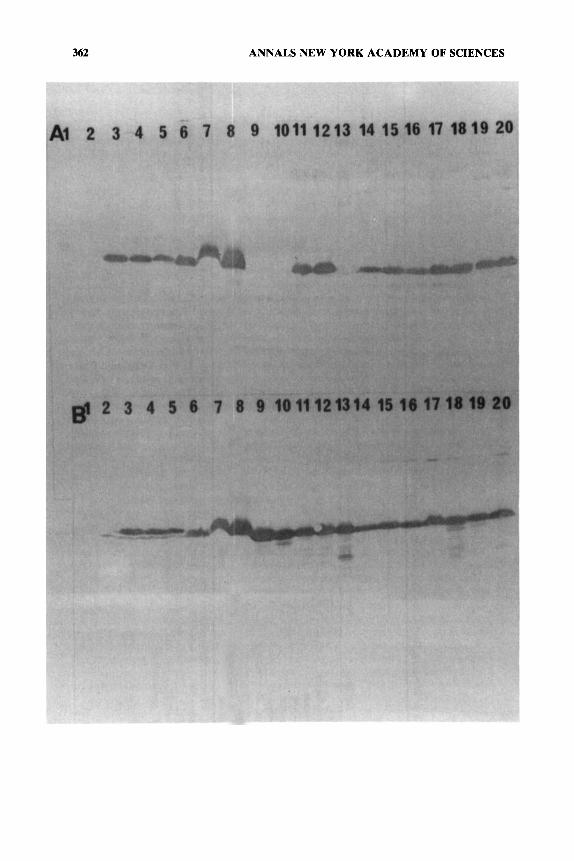
Monoclonal antibodies against Mycobacterium and those prepared against several species of Rickettsiales were used to demonstrate the existence of both cross-reactive and specific epitopes on the HSP60s of several genera (TABLE 3, FIG. 2).'2353 Although all typhus and spotted fever group rickettsiae appeared to react very similarly with polyclonal sera and most monoclonal antibodies, R . Canada HSP60 and Rochalimaea appeared to lack the particular conserved epi- tope recognized by Mycobacterium antibody Y 1.2 (FIG. 2A), while a second, more conserved epitope was recognized by an antibody against R . typhi HSP60 (FIG. 2B). Other epitopes that were Ehrlichia-specific and typhus and spotted fever group rickettsiae-specific were also detected. Most interesting, however, were antibodies which detected Ehrlichia, Rickettsia, and Rochalimaea, or did not react with these genera but did detect Wolbachia and other gamma-subdivi- sion purple bacteria such as Escherichia, Legionella, and Proteus. These group- ings were consistent with phylogenetic relationships deduced from 16s rRNA sequencing.62 On the other hand, neither the polyclonal sera nor broadly reactive typhus monoclonal antibodies exhibited much cross-reactivity against the HSP60 of Rickettsia tsutsugamushi.12 The 16s rRNA sequence of scrub typhus is not known, but these data suggest that scrub typhus may be a distinct eubacterial lineage, lying outside the purple bacteria, possibly diverging from this group more than do the Chlamydia, Mycobacterium, or even the spirochete branches.63
STRUCTURAL CONSERVATION OF HSP60 GENES
The complete sequences of the HSP60 genes of only two rickettsiae, Rickett- sia tsutsugamushi and Coxiella burnetii, are known at present.I2J3 These genes share with E. coli, Chlamydia psittaci, and Synechococcus 6301 genes11p29,60 a common organization in an operon consisting of a heat-shock-like promoter re- gion followed by two open reading frames (ORFs), the first ORF with homology to the GroES gene of E. coli and the second with the GroEL gene. The two genes appear to be cotranscribed. The cloned genes of Coxiella appeared to be heat- regulated in E. coli, while those of R . tsutsugamushi were not. In Mycobacte- rium, the two genes are not present in the same operon.I0 Gly-Gly-Met motifs in the carboxyl-terminal region have been observed for each of the known HSP60 sequences except for Chlamydia and for wheat, Triticum aestivum. This hydro- philic domain may function as a membrane anchor, as it appears to follow a hydrophobic, putative transmembrane domain. Hydrophobicity plots of the indi- vidual HSP60s are remarkably superimposable. j2 Pairwise comparisons of de- duced amino acid sequences indicated homologies of 40-50%. Since many of the non-identical amino acids were conservative replacements, overall HSP60 se- quence similarities were approximately 70%. No one region of the protein con- tains the majority of the conserved amino acids; rather, regions of no more than ten amino acids of identical sequence among the various HSP60 are scattered throughout the protein. Similarly, regions showing low similarity in sequence were no greater than ten amino acids in length and were also scattered through the

i? Y;
X
2 a
5 ..
TABL
E 3.
W
este
rn B
lot C
ross
-Rea
ctiv
ity o
f Ant
i-KSP
60 M
onoc
lona
l Ant
ibod
ies
E fi !
Chl
amyd
ia
- N
N
f
-
1 1
N
-
2 q
Mon
oclo
nal A
ntib
ody
Prep
ared
Aga
inst
a
Ehr
lichi
a R
ocha
limae
a Ty
phus
Ric
ketts
iae
senn
,, ri
st.
Myc
obac
teri
um
Ant
igen
N
o. 1
N
o. 2
N
o. 3
N
o. 1
N
o. 2
N
o. 3
N
o. 4
N
o. 5
N
o. 1
N
o. 1
N
o.2
5:
f
2 f
3 -
-
2 3
m
-
-
Prot
eus
-t
3 C
ampy
loba
cter
t
N
N
Aer
omon
as
- N
N
3
* 3
1 1
4 Le
gion
ella
R
ocha
limae
a 4
4 4
4 4
4 Ty
phus
, SF
rick
etts
iab
3 2
1 4
Ric
k. ts
utsu
gam
ushi
N
N
N
E
hrlic
hia
rist
., se
nn.
2 N
N
1
2 W
olba
chia
-
N
N
-3
-g
4
4 4
4 1
:s
-
-
-
-
4 -
3 2 3
-
-
-
- -
-
-
-
-
-
-
-
-
-
-
N
N
4 3
-
-
-
-
-
1 -
-
- -
-
-
-
- -
3 -
0 N
, not
don
e. S
ome o
rigin
al d
ata
from
Ref
s. 1
2 and
53.
The
stre
ngth
of r
eact
ion
was
sco
red
from
neg
ativ
e (-
) to
ver
y st
rong
(4).
Roc
halim
aea
antib
odie
s No.
1-3
: G
56-5
D2.
2, G
43-1
H12
.6, V
55-8
D10
.4. T
yphu
s ric
ketts
ial a
ntib
odie
s No.
1-5
: TC
30-2
F2.2
, T5-
3H10
.1, T
C30
-4H
8.1,
T5-
1F1.
5,
T5-4
B11
.2.2
. Ehr
lichi
a m
onoc
lona
l ant
ibod
y (N
o. 1
): m
ixtu
re o
f E1
34-4
B3
and
E133
-2A
8. M
ycob
acre
rium
ant
ibod
ies
No.
1-2
: II
H9
and
Y 1.
2.
Ehr
lichi
a se
nn.,
rist
.: E
hrlic
hia
senn
etsu
, ris
ticii.
SF
, spo
tted
feve
r.
362 ANNALS NEW YORK ACADEMY OF SCIENCES

DASCH el al.: RICKETTSIAL HSP60 ANTIGENS 363
protein sequence, notably at both the carboxyl and amino termini. As might be guessed from the differences among HSP60s in amino acid composition, cysteine and histidine appear rather variable in location, while a number of tyrosine, phenylalanine, proline, arginine, and lysine residues may have important struc- tural roles, as deduced from their invariance in location. As anticipated from the amino-terminal sequence comparisons (TABLE 2), the sequence data suggests that Coxiella burnetii and E. coli HSP60s are relatively closely related, while the Rickettsia tsutsugamushi sequence is rather different from the other known se- quences.
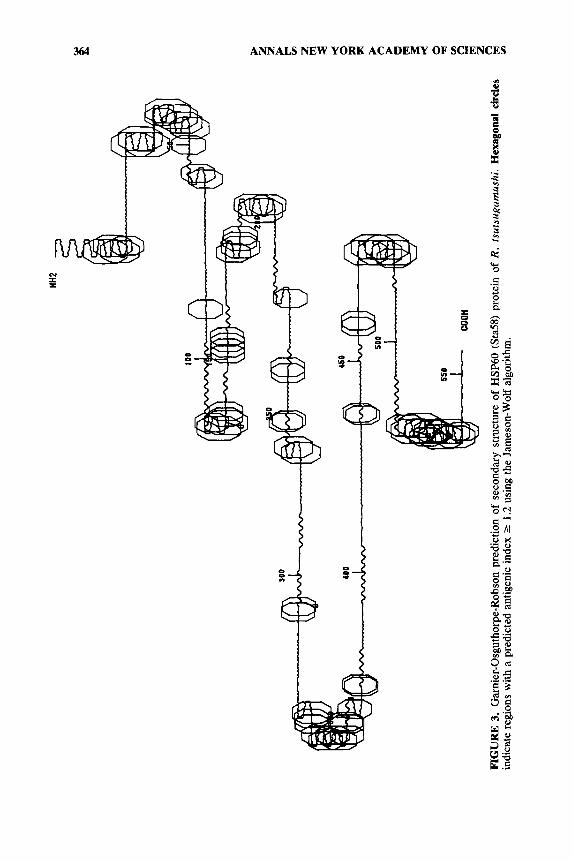
In order to compare structural aspects of different HSP60s, Garnier-Os- guthorpe-R~bson~~ predictions of the secondary structure of the five known com- plete bacterial proteins were plotted (shown in FIG. 3 for R. tsutsugamushi HSP60). Those regions predicted to have a high likelihood of antigenicity were also determined by the algorithm of Jameson and Wolf.68 None of the secondary structure plots was very similar. Several regions of high antigenic index appeared to be conserved (residues 25-60,190-220,350-380,460-480) among the proteins, while the scrub typhus HSP60 had relatively unusual antigenic domains at both the amino and carboxyl ends of the protein. Except for domain 25-60, none of these regions appeared to be much more strongly conserved than other domains. Although two of these domains are in regions of the M . leprae protein sequence where monoclonal antibodies are known to bind, the other predicted antigenic domains do not apparently bind murine an t ibodie~ .~~
DISCUSSION AND CONCLUSIONS
The analysis of stress proteins in pathogenic bacteria is most advanced for the agents of leprosy and tuberculosis. At least three mycobacterial antigens are known to be homologues of well-characterized stress proteins from E. coli or euka ryo te~ .~~ The implications of this observation for other pathogenic bacteria are obvious. In retrospect, it appears logical to expect that immune responses would cause some stress in pathogenic microorganisms and that elevations in the amounts of such stress proteins would be reflected in antibody- and T cell-medi- ated responses to such proteins. However, as regards the HSP60s of C. burnetii, Rochalimaea, Chlamydia, Ehrlichia, and all three groups in the genus Rickettsia, the apparent constitutive levels of these proteins appear remarkably high. In the absence of any direct information on the temperature regulation of any of these proteins, it may be asked whether the process of growth at their usual temperature of cultivation (34°C) constitutes sufficient inductive stress relative to the ambient environment they would encounter in a host arthropod. Alternatively, intracellu-
4 FIGURE 2. Western blot analysis of bacterial antigens with monoclonal antibodies against the HSP60 proteins. Antigens were separated on 8-16% SDS-polyacrylamide gradient gels. Protein loading: (lanes 1, 2) 5 pg, (lanes 3-10) 40 pg, (lanes 11-20) 20 pg. (Lanes 1, 2) Mycobacterium bouis, M. phlei cell walls; (lanes 3-5) Proteus 0x19, 0x2, and OXK cells; (lanes 6-8) Legionella pneumophila, L. micdadei, and L. bozernanii cells; (lanes 9-10) Rochalimaea quintana, R . uinsonii cells; (lanes 11-20) rickettsiae: R. typhi, R. prowazekii, R. Canada, R. rickettsii, R. conorii, Israeli tick typhus, R . sibirica, R . akari, R . australis, and Thai tick typhus. Blots were developed as in FIGURE 1 but with (Panel A) monoclonal antibody Y1.2, against Mycobacterium leprae 65-kDa protein or (Panel B) monoclonal antibody T5-1F1.5, which recognizes R. typhi HSP60.
364 ANNALS NEW YORK ACADEMY OF SCIENCES

DASCH et af.: RICKE’ITSIAL HSP60 ANTIGENS 365
lar growth may constitute significant stress in itself to induce synthesis of these proteins. Synthesis of stress proteins may be particularly essential for adaptive growth in professional phagocytes such as macrophages. Cross-reactive 70-kDa antigens are present in Rochalirnaea and both the typhus and spotted fever group rickettsiae (Dasch, unpublished observations), but their relationship to the DnaK proteins of E. coli and Mycobacteriurn has not been proven.
Structural, immunologic, and DNA sequence information is now available about the HSP60 proteins in Rickettsiaceae and a few other bacteria. Although the broad immunological cross-reactivity of this common bacterial antigen was ini- tially discouraging for its use as a diagnostic antigen, the existence of at least genus-specific epitopes and the distribution of sequence substitutions suggest that this protein has undergone considerable evolution that may be exploited. For example, even with the limited sequences available, there are remarkable paral- lels that can be drawn to phylogenetic relationships deduced previously by 16s rRNA sequencing. If monoclonal antibodies can be selected for binding to phylo- genetically important specific amino acid sequence changes, a more rapid method of typing novel bacteria for phylogenetic purposes may be possible than can be achieved by nucleic acid sequencing. At least such HSP60 studies should indepen- dently validate conclusions about phylogeny based on the ribosomal RNA se- quences. The relatively large amount of the HSP60 antigen present in pathogenic bacteria may permit new approaches to rapid detection of bacteremia utilizing the cross-reactive epitopes and antigen-detection technologies. Alternatively, poly- merase chain reaction amplification of conserved sequences may also permit development of simple broad methods for enumeration of slowly growing or non- cultivable bacteria.
Although the 65-kDa antigen of Mycobacteriurn has been proposed as a vac- cine candidate, this seems inappropriate due to the evidence for its involvement in rheumatoid arthritis and the recent demonstration that the Chlamydia HSP60 antigen may be the cause of ocular hypersensitivity and blindness in trachoma infections.” Similarly, the presence of immune responses to the HSP6O of Bor- relia burgdorferi suggests that autoimmune responses to specific bacterial HSP60 antigens may be significant in the pathology of certain diseases. At present, autoimmunity appears not to be a significant aspect of disease caused by the genus Rickettsia, but it should not be ruled out for Coxiella, particularly in cases of chronic endocarditis. Indeed, even if the rickettsial HSP60 antigens do not elicit autoimmune problems following vaccination, evidence for a protective response against these highly conserved proteins may be difficult to obtain. Despite their conservation, the proteins are clearly immunogenic. Many individuals have a variety of preexisting antibodies to HSP60 proteins and may eliminate vaccines containing these proteins rapidly due to their “natural immunity.” Furthermore, with the rickettsiae, long-lasting protection seems to be more dependent on spe- cies-specific antigens. Following infection, these antigens elicit stronger antibody- and cell-mediated responses in man than do the HSP60 antigens.”
The most important aspect of the HSP60 antigens will probably relate to their normal physiological role in the rickettsiae. Discovery of therapeutic agents which affect the function of these proteins might be used in inhibiting rickettsial replication. On the other hand, the likely role of these proteins in promoting proper translocation, folding, and assembly of proteins suggests that cloned rick- ettsial antigens may be produced more efficiently in E. coli expression systems if, rather than having these processes rely solely on constitutive levels of the E. coli GroEL protein, the appropriate overexpressed rickettsial HSP60 is also present.

366 ANNALS NEW YORK ACADEMY OF SCIENCES
ACKNOWLEDGMENTS
We thank David Marana, Walter G. Sewell, Rick Jaffe, Dzung Pham, Janet Miller, and Richard Grays for their technical assistance on different phases of this project.
1. 2.
3.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
REFERENCES
LINDQUIST, S. 1986. The heat-shock response. Annu. Rev. Biochem. 5 5 1151-1191. LINDQUIST, S. & E. A. CRAIG. 1988. The heat-shock proteins. Annu. Rev. Genet. 22:
NEIDHARDT, F. C. & R. A. VANBOGELEN. 1987. Heat shock response. In Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. F. C. Neidhardt, J. L. Ingraham, K. B. Low, B. Magasanik, M. Schaechter, and H. E. Umbarger, eds.: 1334-1345. American Society for Microbiology. Washington, D.C.
ALLAN, B., M. LINSEMAN, L. A. MACDONALD, J. S. LAM & A. M. KROPINSKI. 1988. Heat shock response of Pseudornonas aeruginosa. J. Bacteriol. 170 3668-3674.
LEMA, M. W., A. BROWN, C. A. BUTLER & P. S. HOFFMAN. 1988. Heat-shock response in Legionella pneumophila. Can. J. Microbiol. 34: 1148-1153.
DANIELS, C. J., A. H. Z. MCKEE & W. F. DOOLITTLE. 1984. Archaebactenal heat- shock proteins. EMBO J. 3: 745-749.
TODD, J . A., T. J. P. HUBBARD, A. A. TRAVERS & D. J. ELLAR. 1985. Heat-shock proteins during growth and sporulation of Bacillus subrilis. FEBS Lett. 188: 209- 214.
KLIMPEL, K. W. & V. L. CLARK. 1989. The heat shock response of type 1 type 4 gonococci. Sex. Transm. Dis. 1 6 141-147.
NERLUND, A. H., A. S. MUSTAFA, D. SWEETSER, T. GODAL & R. A. YOUNG. 1988. A protein antigen of Mycobacrerium leprae is related to a family of small heat shock proteins. J. Bacteriol. 170: 5919-5921.
BAIRD, P. N., L. M. C. HALL & A. R. M. COATES. 1989. Cloning and sequence analysis of the 10 kDa antigen gene of Mycobacterium tuberculosis. J. Gen. Micro- biol. 135: 931-939.
MORRISON, R. P., R. J. BELLAND, K. LYNG & H. D. CALDWELL. 1989. Chlamydia1 disease pathogenesis. The 57-kD chlamydial hypersensitivity antigen is a stress pro- tein. J. Exp. Med. 170 1271-1283.
STOVER, C. K., D. P. MARANA, G. A. DASCH & E. V. OAKS. 1990. Molecular cloning and sequence analysis of the Sta58 major antigen of Rickettsia tsutsugamushi: Se- quence homology and antigenic comparison to the 60-kilodalton family of stress proteins. Infect. Immun. 58. In press.
VODKIN, M. H. & J . C. WILLIAMS. 1988. A heat shock operon in Coxiella burnetii produces a major antigen homologous to a protein in both mycobacteria and Esche- richia coli. J. Bacteriol. 170: 1227-1234.
FRIEDMAN, D. I., E. R. OLSON, C. GEORGOPOULOS, K. TILLY, I. HERSKOWITZ & F. BANUETT. 1984. Interactions of bacteriophage and host macromolecules in the growth of bacteriophage A. Microbiol. Rev. 48: 299-325.
ELLIS, R. J . & S. M. HEMMINGSEN. 1989. Molecular chaperones: Proteins essential for the biogenesis of some macromolecular structures. Trends Biochem. Sci. 14: 339- 342.
ROTHMAN, J. E. 1989. Polypeptide chain binding proteins: Catalysts of protein folding and related processes in cells. Cell 59: 591-601.
FLYNN, G. C., T. G. CHAPPELL & J . E. ROTHMAN. 1989. Peptide binding and release by proteins implicated as catalysts of protein assembly. Science 245: 385-390.
PELHAM, H. R. B. 1989. Heat shock and the sorting of luminal ER proteins. EMBO J.
FAYET, O., T. ZIEGELHOFFER & C. GEORGOPOULOS. 1989. The groES and groEL heat
631-677.
8: 3171-3176.

DASCH ef al.: RICKE’ITSIAL HSP60 ANTIGENS 367
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
shock gene products of Escherichia coli are essential for bacterial growth at all temperatures. J. Bacteriol. 171: 1379-1385.
KUSUKAWA, N., T. YURA, C. UEGUCHI, Y. AKIYAMA & K. ITO. 1989. Effects of mutations in heat-shock genes groES and groEL on protein export in Escherichia coli. EMBO J. 8 3517-3521.
GOLOUBINOFF, P., A. A. GATENBY & G. H. LORIMER. 1989. GroE heat-shock proteins promote assembly of foreign prokaryotic ribulose bisphosphate carboxylase oligo- mers in Escherichia coli. Nature 337: 44-47.
VAN DYK, T. K., A. A. GATENBY & R. A. LAROSSA. 1989. Demonstration by genetic suppression of interaction of GroE products with many proteins. Nature 342: 451- 453.
STRAUS, D. B., W. A. WALTER & C. A. GROSS. 1988. Escherichia coli heat shock gene mutants are defective in proteolysis. Genes & Dev. 2 1851-1858.
MCMULLIN, T. W. & R. L. HALLBERG. 1988. A highly evolutionarily conserved mito- chondrial protein is structurally related to the protein encoded by the Escherichia coli groEL gene. Mol. Cell. Biol. 8: 371-380.
CHENG, M. Y., F.-U. HARTL, J. MARTIN, R. A. POLLOCK, F. KALOUSEK, W. NEUPERT, E. M. HALLBERG, R. L. HALLBERC & A. L. HORWICH. 1989. Mitochon- drial heat-shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature 337: 620-625.
READING, D. S., R. L. HALLBERC &A. M. MYERS. 1989. Characterization of the yeast HSP60 gene coding for a mitochondrial assembly factor. Nature 337: 655-659.
JINDAL, S., A. K. DUDANI, B. SINGH, C. B. HARLEY & R. S. GUPTA. 1989. Primary structure of a human mitochondrial protein homologous to the bacterial and plant chaperonins and to the 65-kilodalton mycobacterial antigen. Mol. Cell. Biol. 9
PICKETTS, D. J., C. S. K. MAYANIL & R. S. GUPTA. 1989. Molecular cloning of a Chinese hamster mitochondrial protein related to the “chaperonin” family of bacte- rial and plant proteins. J. Biol. Chem. 264: 12001-12008.
HEMMINGSEN, S. M., C. WOOLFORD, S. M. VAN DER VIES, K. TILLY, D. T. DENNIS, C. P. GEORCOPOULOS, R. W. HENDRIX & R. J. ELLIS. 1988. Homologous plant and bacterial proteins chaperone oligomeric protein assembly. Nature 333: 330-334.
PUSHKIN, A. V., V. L. TSUPRUN, N. A. SOLOVJEVA, V. V. SHUBIN, Z. G. EVSTIG- NEEVA & W. L. KRETOVICH. 1982. High molecular weight pea leaf protein similar to the groE protein of Escherichia coli. Biochim. Biophys. Acta 704: 379-384.
GUPTA, R. S. , D. J. PICKETTS & S. AHMAD. 1989. A novel ubiquitous protein ‘cha- peronin’ supports the endosymbiotic origin of mitochondrion and plant chloroplast. Biochem. Biophys. Res. Commun. 163: 780-787.
KAUSER, B. 1975. Immunological studies of an antigen common to many Gram-nega- tive bacteria with special reference to E . coli. Int. Arch. Allergy Appl. Immunol. 48:
HOIBY, N. 1975. Cross-reactions between Pseudomonas aeruginosa and thirty-six other bacterial species. Scand. J. Immunol. 4(Suppl. 2): 187-196.
COLLINS, M. T., F. ESPERSEN, N. HOIBY, S-N. Cno, A. FRIIS-MOLLER & J. S. REIF. 1983. Cross-reactions between Legionella pneumophila (serogroup 1) and twenty- eight other bacterial species, including other members of the family Legionellaceae. Infect. Immun. 39: 1441-1456.
HINDERSSON, P., C. S. PETERSEN, N. S. PEDERSEN, N. HOIBY & N. H. AXELSEN. 1984. Immunological cross-reaction between antigen Tp-4 of Treponema pallidurn and an antigen common to a wide range of bacteria. Acta Pathol. Microbiol. Im- munol. Scand. B 92: 183-188.
SOMPOLINSKY, D., J. B. HERTZ, N. HOIBY, K. JENSEN, B. MANSA & Z. SAMRA. 1980. An antigen common to a wide range of bacteria: 1. A biochemical study of a ‘com- mon antigen’ from Pseudomonas aeruginosa. Acta Pathol. Microbiol. Scand. B 88:
SOMPOLINSKY, D., J. B. HERTZ, N. HOIBY, K. JENSEN, B. MANSA, V. B. PEDERSEN & Z. SAMRA. 1980. An antigen common to a wide range of bacteria: 2. A biochemical
2279-2283.
72-81.
143- 149.

368 ANNALS NEW YORK ACADEMY OF SCIENCES
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
study of a 'common antigen' from Pseudomonas aeruginosa. Acta Pathol. Micro- biol. Scand. B 88: 253-260.
YOUNG, D. B., J. IVANYI, J . H. Cox & J. R. LAMB. 1987. The 65kDa antigen of mycobacteria-a common bacterial protein? Immunol. Today 8 215-219.
SHINNICK, T. M., M. H. VODKIN & J. C. WILLIAMS. 1988. The Mycobacferiurn tuberculosis 65-kilodalton antigen is a heat shock protein which corresponds to common antigen and to the Escherichia coli GroEL protein. Infect. Immun. 56: 446- 451.
THOLE, J. E. R., P. HINDERSSON, J. DE BRUYN, F. CREMERS, J. VAN DER ZEE, H. DE COCK, J. TOMMASSEN, W. VAN EDEN & J. D. A. VAN EMBDEN. 1988. Antigenic relatedness of a strongly immunogenic 65 kDa mycobacterial protein antigen with a similarly sized ubiquitous bacterial common antigen. Microb. Pathog. 4: 71-83.
VAN EDEN, W., J. E. R. THOLE, R. VAN DER ZEE, A. NOORDZIJ, J. D. A. VAN EMBDEN, E. J. HENSEN & I. R. COHEN. 1988. Cloning of the mycobacterial epitope recognized by T lymphocytes in adjuvant arthritis. Nature 331: 171-173.
TSOULFA. G., G. A. W. ROOK, G. M. BAHR, M. A. SATTAR, K. BEHBEHANI, D. B. YOUNG, A. MEHLERT, J. D. A. VAN EMBDEN, F. C. HAY, D. A. ISENBERG & P. M. LYDYARD. 1989. Elevated IgG antibody levels to the mycobacterial 65-kDa heat shock protein are characteristic of patients with rheumatoid arthritis. Scand. J. Immunol. 30 519-527.
VAN DEN BROEK, M. F., E. J. M. HOGERVORST, M. C. J. VAN BRUGGEN, W. VAN EDEN, R. VAN DER ZEE & W. B. VAN DEN BERG. 1989. Protection against strepto- coccal cell wall-induced arthritis by pretreatment with the 65-kD mycobacterial heat shock protein. J. Exp. Med. 170: 449-466.
POLLA, B. S. 1988. A role for heat shock proteins in inflammation? Immunol. Today 9
KOGA, T., A. WAND-WURTTENBERGER, J. DEBRUYN, M. E. MUNK, B. SCHOEL & S. H. E. KAUFMANN. 1989. T cells against a bacterial heat shock protein recognize stressed macrophages. Science 245: 11 12-1 115.
YOUNG, R. A. & T. J . ELLIOT. 1989. Stress proteins, infection, and immune surveil- lance. Cell 59: 5-8.
THOLE, J. E. R., W. C. A. VAN SCHOOTEN, W. J. KEULEN, P. W. M. HERMANS, A. A. M. JANSON, R. R. P. DE VRIES, A. H. J. KOLK & J . D. A. VAN EMBDEN. 1988. Use of recombinant antigens expressed in Escherichia coli K-12 to map B-cell and T- cell epitopes on the immunodominant 65-kilodalton protein of Mycobacferiurn bouis BCG. Infect. Immun. 56: 1633-1640.
BRETT, S. J., J. R. LAMB, J. H. Cox, J. B. ROTHBARD, A. MEHLERT & J. IVANYI. 1989. Differential pattern of T cell recognition of the 65-kDa mycobacterial antigen following immunization with the whole protein or peptides. Eur J. Immunol. 19
ANDERSON, D. C., M. E. BARRY & T. M. BUCHANAN. 1988. Exact definition of species-specific and cross-reactive epitopes of the 65-kilodalton protein of Mycobac- ferium leprae using synthetic peptides. J . Immunol. 141: 607-613.
MEEKER, H. C., D. L. WILLIAMS, D. C. ANDERSON, T. P. GILLIS, G. SCHULLER- LEVIS & W. R. LEVIS. 1989. Analysis of human antibody epitopes on the 65-kilo- dalton protein of Mycobacterium leprae by using synthetic peptides. Infect. Immun. 57: 3689-3694.
BUCHANAN, T. M., H. NOMAGUCHI, D. C. ANDERSON, R. A. YOUNG, T. P. GILLIS, W. J. BRITTON, J. IVANYI, A. H. J. KOLK, 0. CLOSS, B. R. BLOOM & V. MEHRA. 1987. Characterization of antibody-reactive epitopes on the 65-kilodalton protein of Mycobacterium leprae. Infect. Immun. 55: 1000-1003.
HELSEL, L. O . , W. F. BIBB, c. A. BUTLER, P. S. HOFFMAN & R. M. MCKINNEY. 1988. Recognition of a genus-wide antigen of Legionella by a monoclonal antibody. Curr. Microbiol. 16: 201-208.
DASCH, G. A., E. WEISS & J. C. WILLIAMS. 1990. Antigenic properties of the Ehrli- chiae and other Rickettsiaceae. Kluwer Academic Publishers of the Netherlands. Amsterdam, Netherlands. In press.
134-1 37.
1303-1310.

DASCH et al.: RICKETTSIAL HSP60 ANTIGENS 369
54.
55.
56.
57.
58.
59.
60.
61.
62.
63. 64.
65.
66.
67.
68.
69.
. 70.
71.
CHING, W-M., F. L. A. BUCKMIRE, P. Y. KIM & G. A. DASCH. 1988. Comparison of properties of the 60 kDa heat shock proteins of Rochalimaea uinsonii and R. quin- tuna with those of other bacteria. J. Cell Biol. 107: 539a.
WEISS, E. & G. A. DASCH. 1982. Differential characteristics of strains ofRochalirnaea: Rochalimaea uinsonii sp. nov., the Canadian vole agent. Int. J. Syst. Bacteriol. 32:
PAW, C-P., B. B. PLIKAYTIS, G. M. CARLONE & I. M. WARNER. 1988. Purification, partial characterization, and seroreactivity of a genuswide 60-kilodalton Legionella protein antigen. J. Clin. Microbiol. 26: 67-71.
HENDRIX, R. W. 1979. Purification and properties of groE, a host protein involved in bacteriophage assembly. J. Mol. Biol. 129 375-392.
DE BRUYN, J., R. BOSMANS, M. TURNEER, M. WECKX, J. NYABENDA, J-P. VAN VOOREN, P. FALMAGNE, H. G. WIKER & M. HARBOE. 1987. Purification, partial characterization, and identification of a skin-reactive protein antigen of Mycobacte- rium bouis BCG. Infect. Immun. 55: 245-252.
SHINNICK, T. M., D. SWEETSER, J. THOLE, J. VAN EMBDEN & R. A. YOUNG. 1987. The etiologic agents of leprosy and tuberculosis share an immunoreactive protein antigen with the vaccine strain Mycobacteriurn bouis BCG. Infect. Immun. 55:
COZENS, A. L. & J. E. WALKER. 1987. The organization and sequence of the genes for ATP synthase subunits in the cyanobacterium Synechococcus 6301. Support for an endosymbiotic origin of chloroplasts. J. Mol. Biol. 194: 359-383.
MUSGROVE, J. E., R. A. JOHNSON & R. J. ELLIS. 1987. Dissociation of the ribulosebis- phosphate-carboxylase large-subunit binding protein into dissimilar subunits. Eur. J. Biochem. 163: 529-534.
WEISBURG, W. G., M. E. DOBSON, J. E. SAMUEL, G. A. DASCH, L. P. MALLAVIA, 0. BACA, L. MANDELCO, J. E. SECHREST, E. WEISS & C. R. WOESE. 1989. Phyloge- netic diversity of the rickettsiae. J. Bacteriol. 171: 4202-4206.
305-3 14.
1932-1935.
WOESE, C. R. 1987. Bacterial evolution. Microbiol. Rev. 51: 221-271. SAMPSON, J. S. , B. B. PLIKAYTIS & H. W. WILKINSON. 1986. Immunologic response of
patients with legionellosis against major protein-containing antigens of Legionella pneumophila serogroup 1 as shown by immunoblot analysis. J. Clin. Microbiol. 23:
PLIKAYTIS, B. B., G. M. CARLONE, C-P. PAU & H. W. WILKINSON. 1987. Purified 60- kilodalton Legionella protein antigen with Legionella-specific and nonspecific epi- topes. J. Clin. Microbiol. 25: 2080-2084.
HANSEN, K., J. M. BANGSBORG, H. FJORDVANG, N. S. PEDERSEN & P. HINDERSSON. 1988. Immunochemical characterization of and isolation of the gene for a Borrelin burgdorferi immunodominant 60-kilodalton antigen common to a wide range of bac- teria. Infect. Immun. 5 6 2047-2053.
GARNIER, J., D. J. OSGUTHORPE & B. ROBSON. 1978. Analysis of the accuracy and implications of simple methods for predicting the secondary structure of globular proteins. J. Mol. Biol. 120 97-120.
JAMESON, B. A. & H. WOLF. 1988. The antigenic index: A novel algorithm for predict- ing antigenic determinants. Comput. Appl. Biosci. 4: 181-186.
YOUNG, D., R. LATHIGRA, R. HENDRIX, D. SWEETSER & R. A. YOUNG. 1988. Stress proteins are immune targets in leprosy and tuberculosis. Proc. Natl. Acad. Sci. USA
MORRISON, R. P., K. LYNG & H. D. CALDWELL. 1989. Chlamydia1 disease pathogene- sis: Ocular hypersensitivity elicited by a genus-specific 57-kD protein. J. Exp. Med.
OHASHI, N., A. TAMURA & T. SUTO. 1988. Immunoblotting analysis of anti-rickettsial antibodies produced in patients of tsutsugamushi disease. Microbiol. Immunol. 32:
92-99.
85: 4261-4270.
169 663-675.
1085- 1092.




















