
The Use of Polliniferous Resources by Melipona capixaba , an Endangered Stingless Bee Species

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
Journal of Chemical Ecology, Vol. 30, No. 4, April 2004 (©C 2004)
A STINGLESS BEE (Melipona seminigra) MARKS FOODSOURCES WITH A PHEROMONE FROM ITS CLAW
RETRACTOR TENDONS
STEFAN JARAU,1,2,∗ MICHAEL HRNCIR,1 MANFRED AYASSE,2
CLAUDIA SCHULZ,3 WITTKO FRANCKE,3 RONALDO ZUCCHI,4
and FRIEDRICH G. BARTH1
1Institute of Zoology, University of Vienna, Althanstrasse 141090 Vienna, Austria
2Department of Experimental EcologyUniversity of Ulm, Albert-Einstein-Allee 11
89069 Ulm, Germany3Institute of Organic Chemistry, University of Hamburg
Martin-Luther-King Platz 6, 20146 HamburgGermany
4Department of Biology, FFCLRP, University of Sao PauloAv. Bandeirantes 3900, 14040-901 Ribeirao Preto
SP, Brazil
(Received October 9, 2003; accepted December 19, 2003)
Abstract—By depositing scent marks on flowers, bees reduce both the searchtime and the time spent with the handling of nonrewarding flowers. They therebyimprove the efficiency of foraging. Whereas in honey bees the source of thesescent marks is unknown, it is assumed to be the tarsal glands in bumble bees.According to histological studies, however, the tarsal glands lack any openingsto the outside. Foragers of the stingless beeMelipona seminigrahave previouslybeen shown to deposit an attractant pheromone at sugar solution feeders, whichis secreted at the tips of their tarsi. Here we show that the claw retractor tendonshave specialized glandular epithelia within the femur and tibia of all legs thatproduce this pheromone. The secretion accumulates within the hollow tendon,which also serves as the duct to the outside, and is released from an openingat the base of the unguitractor plate. In choice experiments,M. seminigrawasattracted by feeders baited with pentane extracts of the claw retractor tendons inthe same way as it was attracted by feeders previously scent marked by foragers.Our results resolve the seeming contradiction between the importance of footprint secretions and the lack of openings of the tarsal glands.
∗To whom correspondence should be addressed. E-mail: [email protected]
793
0098-0331/04/0400-0793/0C© 2004 Plenum Publishing Corporation

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
794 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
Key Words—Hymenoptera, Apidae, Meliponini, stingless bee,Melipona, tendongland, food source marking, pheromone, attractant, hydrocarbons.
INTRODUCTION
Honey bees (Apidae, Apini), bumble bees (Apidae, Bombini), and stingless bees(Apidae, Meliponini) use repellent scent marks to keep foragers from visitingrecently depleted flowers (N´unez, 1967; Giurfa and N´unez, 1992; Giurfa, 1993;Goulson et al., 1998, 2001; Stout et al., 1998; Williams, 1998; Stout and Goulson,2001) as well as attractant substances to mark rich food sources (Ferguson and Free,1979; Cameron, 1981; Free and Williams, 1983; Schmitt and Bertsch, 1990; Villaand Weiss, 1990; Nieh, 1998; Aguilar and Sommeijer, 2001; Hrncir et al., 2004;Nieh et al., 2003; Schmidt et al., 2003). The bees thereby improve the efficiency offood gathering by reducing both the time spent with nonrewarding flowers and thesearch for rewarding flowers (Wetherwax, 1986; Kato, 1988; Schmitt and Bertsch,1990; Giurfa and N´unez, 1992).
The stingless beeMelipona seminigramarks rich food sources with attractantscent marks (Hrncir et al., 2004). After about 40 visits of one or more foragers ata sugar solution feeder, newly arriving bees prefer this food source to a clean butotherwise identical control feeder. The scent marks remain effective for at least2 hr and were shown to be left on the substrate as “foot prints.” When the bees’ legtips were covered with nail polish, thereby sealing the openings of excretory ductsof glands within the legs,Meliponawas no longer able to mark a feeder even after80 visits (Hrncir et al., 2004).
In honey bees, the origin of the scent marks left at food sources by foragersremains unresolved (Free, 1987; Winston, 1987). Foot print secretions are knownto be used to mark the entrance of the hive to help returning bees find their wayhome (Butler et al., 1969). Bumble bees are assumed to deposit both their repellentand their attractant scent marks at food sources as foot prints. The tarsal glandswithin the fifth tarsomeres of their legs were considered the most obvious candidateglands producing the foot print substances (Schmitt et al., 1991; Stout et al., 1998;Goulson et al., 2000). In histological studies, however, openings of the tarsal glandsto the outside could not be found in bumble bees (Pouvreau, 1991) or honey bees(Lensky et al., 1985; Federle et al., 2001), nor inM. seminigra(Jarau, unpublished)or other stingless bees (Cruz-Landim et al., 1998).
The present study was undertaken (i) to look for other possible glandu-lar origins of the foot print substances used byM. seminigra to scent markfood sources, (ii) to examine the effect of gland extracts on the bees’ behav-ior, and (iii) to analyze the chemical composition of scent marks and of glandextracts.

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
STINGLESS BEE MARKS FOOD SOURCES WITH A PHEROMONE 795
METHODS AND MATERIALS
Bee Colonies. We used three colonies ofM. seminigraFRIESE 1903 (Hy-menoptera, Apidae, Meliponini) that were collected in Nova Xavantina–MT, Brazil,and housed in wooden boxes inside the laboratory, connected to the outside byplastic tubes in the wall of the building. The experiments were carried out on theRibeirao Preto—Campus of the University of S˜ao Paulo.
Histology and SEM. Legs of foraging workers were fixed in Dubosq-Brasilsolution for 72 hr, washed in 70% ethanol, dehydrated in a series of alcohols withincreasing concentration, and embedded in Spurr’s resin. Serial sections 5–8µmthick were stained with toluidine blue. For scanning electron microscopy, legswere stored in 70% ethanol, dehydrated in a series of alcohols with increasingconcentration, placed in acetone, and finally dried using hexamethyldisilazane.Preparations were mounted on stubs, sputter coated (Agar 103) with gold for300 sec, and examined in a Jeol JSM-35CF scanning electron microscope.
Bioassays. Gland extracts were prepared by dissecting the entire claw retractortendons with their glandular tissues from the legs of foraging workers. To avoidcontamination with material from the tarsal glands, the fifth tarsal segments wereremoved prior to cutting the legs and taking out the tendons. Two extracts wereprepared by placing the 18 tendons from three bees in 1200µl pentane for 24hr each. Four hundred microliters of solution therefore corresponded to one beeequivalent (six tendons). For testing the bees’ behavioral reaction to differentconcentrations of the gland extracts, 8, 40, and 200µl of the stock solutionscorresponding to 0.02, 0.1, and 0.5 bee equivalents were used, respectively.
Bees were trained to collect unscented sugar solution (50% weight on weight)ad libitumat a training feeder 20 m northwest of the hives between 10:00 and 11:30on days when no choice experiments were performed. The feeders used for thebioassays were small plastic cups containing 50% unscented sugar solution onplastic discs (65 mm in diam). The distance between the test feeder (baited withgland extract at the beginning of the experiment) and the control feeder (sameamount of pure pentane) measured 20 cm consistently. To avoid side bias, thepositions of the two feeders were exchanged every 5 min. When the setup wasmounted at 10:00 at the training site, a few drops of sugar solution were injectedinto the nest entrances at the same time in order to stimulate the foragers. Thebees quickly started to visit the known feeding site. For 40 min, the number ofbees that landed and extended their proboscises into the sugar solution on eitherthe test feeder or the control feeder was recorded. All bees were captured on theirfirst visit to one of the feeders, marked, and released at the end of the respectiveexperiment only. In order to avoid individual bees being tested more than once,only those captured during a given experiment and not already marked in precedingexperiments were included in the analysis. Control experiments followed the sameprocedure, but the bees had to choose between two identical clean feeders. For

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
796 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
each bee equivalent tested and for the control series, six experiments were carriedout. During all 24 experiments (N = 24), a total of 465 individual bees (control:N = 137; 0.02 bee equivalents:N = 115; 0.1 bee equivalents:N = 131; 0.5 beeequivalents:N = 82) were tested.
Data Analyses. We compared the median percentage of bees choosing the testfeeder in the choice experiments and the percentage of bees choosing one (definedprior to the experiment) of the two identical control feeders (during the controlexperiments) using Mann–WhitneyU tests followed by the sequential Bonferroniprocedure (overallP < 0.05) in order to control for tablewide type I errors (Rice,1989).
Chemical Analyses. Scent marks left on artificial flowers were collected usingfour cleaned (rinsed with ethanol, acetone, and pentane; heated 3 hr in an oven at230˚C) Teflon discs (Ø = 18 mm) as “petals.” The bees landed on these discs andtook up sugar solution (50% weight on weight) from a small plastic cup that wascontinuously refilled by means of a syringe connected to its bottom. For chemicalanalyses, washings of 12 Teflon discs (pooled), which contained the substancesleft by a total of 2000 bee visits, were used. Extracts of the fifth tarsomeres withthe tarsal glands and of the dissected claw retractor tendons with the glandularepithelium (dissected as described for the bioassays) were prepared by extractingthe respective parts of the legs (150 tarsomeres and 150 tendons) in 1 ml pentane for24 hr. All samples were analyzed using a combined Fisons Instruments GC 8000series/MD 800 mass spectrometer (carrier gas: helium; column: 30 m× 0.25 mmfused silica coated with DB5-MS, 0.25µm thickness; electron impact 70 eV). Thetemperature was initially kept at 60˚C for 5 min, then increased by 10˚C min−1 to300◦C, and kept at this temperature for 30 min. Double bond positions in alkeneswere determined by investigation of the corresponding dimethyl disulfide adducts(Buser et al., 1983), while geometries of double bonds were assigned on the basisof reference substances, synthesized by standard laboratory methods.
RESULTS
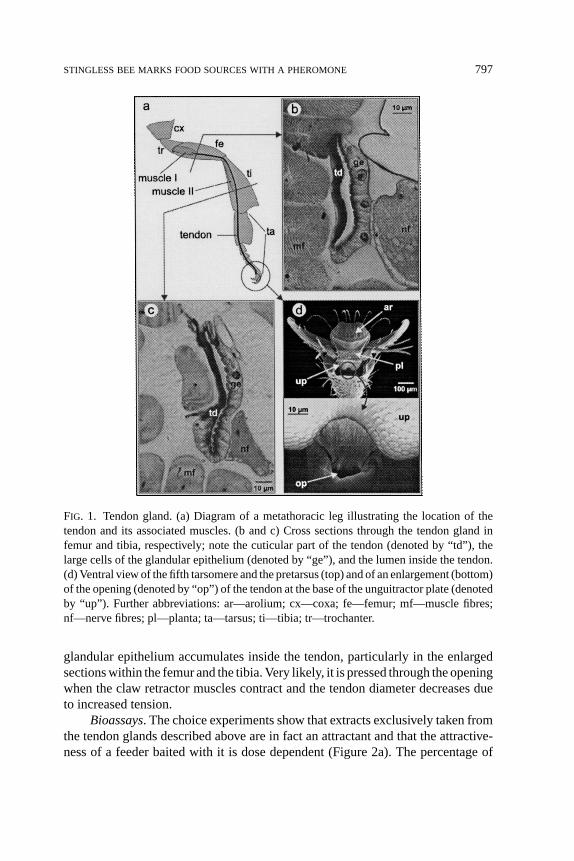
Glandular Structures. In the legs ofMelipona, a claw retractor tendon runsfrom the femur through the tibia and tarsus and connects to the unguitractor plateat the base of the pretarsus. The claw retractor muscle is divided into a femoraland a tibial part (Figure 1a). The epidermis covering the tendon partly formsa glandular epithelium within the femur and the tibia (Figure 1b and c). Thisepithelium is well developed in all legs. The tendon itself forms a hollow cuticulartube along its entire length and opens to the outside at the base of the unguitractorplate (Figure 1d). Inside the fifth tarsomere, the claw retractor tendon crosses theepithelium of the tarsal gland, but there is no connection between the lumina ofthe tarsal gland and the tendon gland, respectively. The secretion produced by the
P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
STINGLESS BEE MARKS FOOD SOURCES WITH A PHEROMONE 797
FIG. 1. Tendon gland. (a) Diagram of a metathoracic leg illustrating the location of thetendon and its associated muscles. (b and c) Cross sections through the tendon gland infemur and tibia, respectively; note the cuticular part of the tendon (denoted by “td”), thelarge cells of the glandular epithelium (denoted by “ge”), and the lumen inside the tendon.(d) Ventral view of the fifth tarsomere and the pretarsus (top) and of an enlargement (bottom)of the opening (denoted by “op”) of the tendon at the base of the unguitractor plate (denotedby “up”). Further abbreviations: ar—arolium; cx—coxa; fe—femur; mf—muscle fibres;nf—nerve fibres; pl—planta; ta—tarsus; ti—tibia; tr—trochanter.
glandular epithelium accumulates inside the tendon, particularly in the enlargedsections within the femur and the tibia. Very likely, it is pressed through the openingwhen the claw retractor muscles contract and the tendon diameter decreases dueto increased tension.
Bioassays. The choice experiments show that extracts exclusively taken fromthe tendon glands described above are in fact an attractant and that the attractive-ness of a feeder baited with it is dose dependent (Figure 2a). The percentage of

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
798 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
FIG. 2. Percentage of bees choosing a test feeder. (a) Choice experiments testing the effectof tendon gland extracts. (b) For comparison, results from Hrncir et al. (2004) for feedersvisited by 20, 40, or 100 foragers prior to testing are shown. The number of bees choosingthe test feeder depends on the concentration of the attractant pheromone deposited on it(i.e., bee equivalents in (a) and accumulation of scent marks in (b)). The boxes represent themedians with 1st and 3rd quartiles, the bars below and above a box indicate the 10th and90th percentile, respectively. Asterisks (*) indicate experiments with significantly more beeschoosing the test feeder than one of the two feeders in the respective control experiments(Mann–WhitneyU tests and sequential Bonferroni method;P = 0.002 in (a) and P =0.004 in (b)).
bees choosing the feeders baited with 0.02 bee equivalents was not significantlydifferent from the control experiments (U = 7.5,N1 = N2 = 6, P = 0.093). Whenfeeders were baited with 0.1 and 0.5 bee equivalents, however, a significantly largerpercentage of bees landed on the test feeder as compared to the control experiments(for both comparisons:U = 0, N1 = N2 = 6, P = 0.002). The percentages of beeschoosing the baited feeder in the experiments with 0.1 and 0.5 bee equivalents,respectively, did not differ significantly from each other (U = 14, N1 = N2 = 6,P = 0.589). To compare these results with data for bees choosing between a clean

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
STINGLESS BEE MARKS FOOD SOURCES WITH A PHEROMONE 799
feeder and a feeder visited by 20, 40, or 100 foragers prior to testing, results fromHrncir et al. (2004) are shown in Figure 2b.
Chemical Analyses. According to chemical analyses, the foot prints left byM. seminigraforagers at food sources contain a complex mixture mainly of hy-drocarbons (Table 1). The same substances are also found in the tendon glandsand in the extracts of the last tarsal segments, which contain the tarsal glands.In addition to these substances, another 41 compounds could be identified in thetendon gland and the tarsal extracts (a number of methylalkanes and wax-typeesters in both extracts; additional fatty acids and fatty acid ethyl esters in the tarsalglands; Table 1).
DISCUSSION
Despite the many studies demonstrating the importance of repellent and at-tractive scent marks at food sources (see Introduction), knowledge about the ori-gin of these scent marks is scarce. Foot print substances are at least used bybumble bees (Schmitt et al., 1991; Stout et al., 1998; Goulson et al., 2000) andby M. seminigra(Hrncir et al., 2004). Considering the lack of openings of thetarsal glands to the outside (Lensky et al., 1985; Pouvreau, 1991; Cruz-Landimet al., 1998; Federle et al., 2001; Jarau, unpublished), secretion of the respectivepheromones by them is unlikely. The tendon glands ofM. seminigradescribed inthe present study resolve the contradiction between the known importance of footprint secretions on the one hand and the lack of openings of the tarsal glands on theother.
Attractiveness of Tendon Gland Extracts. According to our choice experi-ments, the attractive pheromone left by the foragers ofM. seminigraat the foodsource is produced by the glandular epithelium of their claw retractor tendons. In-terestingly, the dose dependence of the attractiveness now found in the bioassayssupports results of our earlier study (Hrncir et al., 2004). In this study, the abilityto choose a sugar solution feeder scent marked by foragers was shown to dependon the number of the preceding bee visits (Figure 2b). It reached a maximum after40 forager landings and decreased at still higher numbers. Likewise, the attrac-tiveness of the scent marks reached a maximum at a pheromone concentrationcorresponding to 0.1 bee equivalent and decreased again at a higher concentration.Thus, at still higher concentrations, the pheromone may also function as a repellentsignal indicating flowers that have already been depleted by many visits (manyscent marks accumulated). Experiments with bumble bees showing correspond-ing effects of synthetic mixtures of hydrocarbons previously identified in scentmarks (Schmitt et al., 1991) support this assumption. It has long been known thatin social insects the same pheromone can release different behaviors at differentconcentrations (Wilson, 1965).

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
800 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
TABLE 1. SUBSTANCESIDENTIFIED FROMFOOTPRINTS LEFT AT FOOD
SOURCES AND FROMEXTRACTS OF THETENDON GLANDS AND THE FIFTH
TARSAL SEGMENTS, RESPECTIVELY, OF FORAGERS OFMelipona seminigraa
Substance Foot prints Tendon glands 5th tarsal segment
AlkanesHeptadecane ++ + +Nonadecane + ++Eicosane +Heneicosane ++ + +Docosane ++ ++Tricosane +++ ++ +Tetracosane ++ ++ ++Pentacosane ++++ ++++ +++Hexacosane +++ ++ +Heptacosane ++++ +++ +++Octacosane +++ + +Nonacosane +++ ++ ++Triacontane +++ +Hentriacontane +++ ++ ++Tritriacontane +
Alkenes8-(Z)-Heptadecene +9-(Z)-Nonadecene + + +7-(Z)-Nonadecene +9-(Z)-Tricosene ++ + +7-(Z)-Tricosene ++ ++ ++7-(Z)-Tetracosene ++ ++9-(Z)-Pentacosene ++ ++ ++7-(Z)-Pentacosene ++++ ++++ ++++9-(Z)-Hexacosene +7-(Z)-Hexacosene ++ ++9-(Z)-Heptacosene +++ ++ +++7-(Z)-Heptacosene ++++ ++++ +++9-(Z)-Nonacosene ++7-(Z)-Nonacosene +++ +++ +++
Methyl alkanes11-Methyltricosane + +9-Methyltricosane + +7-Methyltricosane + +5-Methyltricosane + ++3-Methyltricosane ++ ++ ++13-Methylpentacosane + ++11-Methylpentacosane + ++9-Methylpentacosane + ++7-Methylpentacosane + +5-Methylpentacosane ++ +3-Methylpentacosane ++ ++
Continued

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
STINGLESS BEE MARKS FOOD SOURCES WITH A PHEROMONE 801
TABLE 1. CONTINUED
Substance Foot prints Tendon glands 5th tarsal segment
13-Methylheptacosane + ++11-Methylheptacosane + ++5-Methylheptacosane + +3-Methylheptacosane + +3-Methylnonacosane +
AldehydeOctadecanal +++ ++
AcidsTetradecanoic acid ++Hexadecenoic acid ++Hexadecanoic acid ++Octadecanoic acid ++
EstersEthyl hexadecanoate ++Ethyl linoleate ++Ethyl oleate +++Ethyl linolenoate +++Tetradecyl tetradecanoate + +Dodecyl hexadecanoate + +Dodecyl octadecenoate +Tetradecenyl hexadecenoate + ++Tetradecyl hexadecanoate ++ ++Tetradecenyl octadecenoate ++ ++Hexadecenyl hexadecenoate +Hexadecyl hexadecanoate + ++Hexadecenyl octadecenoate ++ ++
a (+) <0.1%; (++) 0.1–0.9%; (+++) 1.0–9.9%; (++++)≥10%.
Tendon Glands. The development of parts of the epidermis of the claw re-tractor tendons into pheromone producing glands is an intriguing example of theadaptation of a structure already serving a particular function to a different one.To the best of our knowledge, our study is the first to demonstrate the behav-ioral significance of this gland in a bee. Importantly, however, ants of the genusCrematogasterlay scent trails using a pheromone produced in their metathoraciclegs (Fletcher and Brand, 1968). The glands involved were described as a reservoirlined by cuticle and surrounded by a monolayered epithelium inside the tibia, witha hollow tendon guiding the secretions to the pretarsal opening (Billen, 1984). Al-though not explicitly stated by the author, the tibial gland inCrematogastermostlikely represents a specialized region of the claw retractor tendon similar to thatnow found in the stingless bee. In contrast to the glands ofM. seminigra, how-ever, the glands ofCrematogasterwere only found in the tibiae and were far less

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
802 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
developed in the first and second legs than in the hind legs. In the queens of fivespecies of stingless bees, sacculiform glands formed by a fold of the epidermiscovering the apodeme of the femoral muscles have been described recently (Cruz-Landim et al., 1998). Openings of these glands could not be found, however, andtheir function remains unknown. The question of how widespread (in terms of bothphylogeny and behavioral context) the use of tendon glands is within the socialHymenoptera (or in insects in general) can only be answered by extending thestudy to other groups. Our assumption that the foot print substances are secretedby tendon glands instead of tarsal glands is strongly supported by recent findingsin honey bees and weaver ants, which demonstrate that the liquid content of thetarsal gland reservoir is not released to the outside but serves to fill the arolium inorder to increase adhesion to the substrate during walking (Federle et al., 2001).
Scent Marks and Gland Contents. The scent marks left at food sources byM. seminigramainly consist of hydrocarbons. Most of these hydrocarbons arealso contained in tarsal gland extracts and foot print substances of the bumble bee,Bombus terrestris(Schmitt et al., 1991), where they serve as attractant scent marksat sugar solution feeders like inMelipona. Goulson et al. (2000) found some of thesehydrocarbons in tarsal extracts ofB. terrestris, B. pascuorum, andB. lapidariusas well. These authors, however, reported a repellent effect of tarsal extracts onforaging bumble bees when applied to flowers. ForM. seminigra, no data on theeffect of foot prints or of tendon gland extracts on bees foraging at natural foodsources are available yet. Also, it is not known yet whether the hydrocarbonsthemselves are the active compounds in the foot prints ofM. seminigraor ratherserve as solvents for other substances.
As our analyses show, the chemical composition of the tendon gland ex-tracts is similar to that of the last tarsal segments containing the tarsal glands inM. seminigra. Dahl (1885) interpreted the tarsal glands of the honey bee as saclikefoldings of the epidermis of the most distal parts of the claw retractor tendons. Acommon ontogenetic origin of tendon glands and tarsal glands may well explainthe chemical similarity of their products. It has also to be taken into account, how-ever, that it is difficult to completely exclude small pieces of the tendons and theirsecretions when preparing extracts of the tarsal glands.
Acknowledgments—We are grateful to Sidnei Mateus for letting us participate in his rich practicalexperience with stingless bees. This study was supported by grant P 14328 of the Austrian ScienceFoundation (FWF) to FGB. The work in Brazil was conducted under permit Ibama 074/00–Difas–Direc.
REFERENCES
AGUILAR, I. and SOMMEIJER, M. 2001. The deposition of anal excretions byMelipona favosafor-agers (Apidae: Meliponinae): Behavioural observations concerning the location of food sources.Apidologie32:37–48.

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
STINGLESS BEE MARKS FOOD SOURCES WITH A PHEROMONE 803
BILLEN, J. P. J. 1984. Morphology of the tibial gland in the antCrematogaster scutellaris. Naturwis-senschaften71:324–325.
BUSER, H.-R., ARN, H., GUERIN, P., and RAUSCHER, S. 1983. Determination of double bond positionin mono-unsaturated acetates by mass spectrometry of dimethyl disulfide adducts.Anal. Chem.55:818–822.
BUTLER, C. G., FLETCHER, D. J. C., and WATLER, D. 1969. Nest-entrance marking with pheromonesby the honeybee—Apis mellifera L., and by a wasp,Vespula vulgarisL. Anim. Behav.17:142–147.
CAMERON, S. A. 1981. Chemical signals in bumble bee foraging.Behav. Ecol. Sociobiol.9:257–260.CRUZ-LANDIM , C. DA, SILVA DE MORAES, R. L. M., SALLES, H. C., and REGINATO, R. D. 1998. Note
on glands present in Meliponinae (Hymenoptera, Apidae) bees legs.Rev. Bras. Zool.15:159–165.DAHL, F. 1885. Die Fussdr¨usen der Insekten.Arch. Mikroskop. Anat.25:236–262.FEDERLE, W., BRAINERD, E. L., MCMAHON, T. A., and HOLLDOBLER, B. 2001. Biomechanics of the
movable pretarsal adhesive organ in ants and bees.Proc. Natl. Acad. Sci. USA98:6215–6220.FERGUSON, A. W. and FREE, J. B. 1979. Production of forage-marking pheromone by the honeybee.J.
Apic. Res.18:128–135.FLETCHER, D. J. C. and BRAND, J. M. 1968. Source of the trail pheromone and method of trail laying
in the antCrematogaster peringueyi. J. Insect Physiol.14:783–788.FREE, J. B. 1987. Pheromones of Social Bees. Cornell University Press, Ithaca, NY.FREE, J. B. and WILLIAMS , I. H. 1983. Scent-marking of flowers by honeybees.J. Apic. Res.22:86–90.GIURFA, M. 1993. The repellent scent-mark of the honeybeeApis mellifera ligusticaand its role as
communication cue during foraging.Insectes Soc.40:59–67.GIURFA, M. and NUNEZ, J. A. 1992. Honeybees mark with scent and reject recently visited flowers.
Oecologia89:113–117.GOULSON, D., CHAPMAN, J. W., and HUGHES, W. O. H. 2001. Discrimination of unrewarding flowers
by bees; direct detection of rewards and use of repellent scent marks.J. Insect Behav.14:669–678.GOULSON, D., HAWSON, S. A., and STOUT, J. C. 1998. Foraging bumblebees avoid flowers already
visited by conspecifics or by other bumblebee species.Anim. Behav.55:199–206.GOULSON, D., STOUT, J. C., LANGLEY, J., and HUGHES, W. O. H. 2000. Identity and function of scent
marks deposited by foraging bumblebees.J. Chem. Ecol.26:2897–2911.HRNCIR, M., JARAU, S., ZUCCHI, R., and BARTH, F. G. 2004. On the origin and properties of scent
marks deposited at the food source by a stingless bee,Melipona seminigraFRIESE1903.Apidologie35:3–13.
KATO, M. 1988. Bumblebee visits toImpatiensspp.: Pattern and efficiency.Oecologia76:364–370.LENSKY, Y., CASSIER, P., FINKEL, A., DELORME-JOULIE, C., and LEVINSOHN, M. 1985. The fine structure
of the tarsal glands of the honeybeeApis melliferaL. (Hymenoptera).Cell Tissue Res.240:153–158.
NIEH, J. C. 1998. The role of a scent beacon in the communication of food location by the stinglessbee,Melipona panamica. Behav. Ecol. Sociobiol.43:47–58.
NIEH, J. C., RAMıREZ, S., and NOGUEIRA-NETO, P. 2003. Multi-source odor-marking of food by astingless bee,Melipona mandacaia. Behav. Ecol. Sociobiol.54:578–586.
NUNEZ, J. A. 1967. Sammelbienen markieren versiegte Futterquellen durch Duft.Naturwissenschaften54:322–323.
POUVREAU, A. 1991. Morphology and histology of tarsal glands in bumble bees of the generaBombus,Pyrobombus, andMegabombus. Can. J. Zool.69:866–872.
RICE, W. R. 1989. Analyzing tables of statistical tests.Evolution43:223–225.SCHMIDT, V. M., ZUCCHI, R., and BARTH, F. G. 2003. A stingless bee marks the feeding site in addition
to the scent path (Scaptotrigonaaff. depilisMoure 1942). Apidologie34:237–248.SCHMITT, U. and BERTSCH, A. 1990. Do foraging bumblebees scent-mark food sources and does it
matter?Oecologia82:137–144.

P1: IAZ
Journal of Chemical Ecology [joec] pp1165-joec-484405 May 14, 2004 13:48 Style file version June 28th, 2002
804 JARAU, HRNCIR, AYASSE, SCHULZ, FRANCKE, ZUCCHI, AND BARTH
SCHMITT, U., LUBKE, G., and FRANCKE, W. 1991. Tarsal secretion marks food sources in bumblebees(Hymenoptera: Apidae).Chemoecology2:35–40.
STOUT, J. C. and GOULSON, D. 2001. The use of conspecific and interspecific scent marks by foragingbumblebees and honeybees.Anim. Behav.62:183–189.
STOUT, J. C., GOULSON, D., and ALLEN, J. A. 1998. Repellent scent-marking of flowers by a guild offoraging bumblebees (Bombusspp.).Behav. Ecol. Sociobiol.43:317–326.
VILLA , J. D. and WEISS, M. R. 1990. Observations of the use of visual and olfactory cues byTrigonaspp. foragers.Apidologie21:541–545.
WETHERWAX, P. B. 1986. Why do honeybees reject certain flowers?Oecologia69:567–570.WILLIAMS , C. S. 1998. The identity of the previous visitor influences flower rejection by nectar-
collecting bees.Anim. Behav.56:673–681.WILSON, E. O. 1965. Chemical communication in the social insects.Science149:1064–1071.WINSTON, M. L. 1987. The Biology of the Honey Bee. Harvard University Press, Cambridge, MA.
Copyright © 2022 FDOKUMEN















