
Management of Astrocaryum standleyanum (Arecaceae) for handicraft production in Colombia
Upload
independentCategory
view
0download
0
A revision of Asterogyne (Arecaceae: Arecoideae: Geonomeae)
FRED W . STAUFFER, CONNY B . ASMUSSEN,
ANDREW HENDERSON, AND PETER K . ENDRESS
Stauffer, E W. (Institute of Systematic Botany, University of Zurich, Zolliker- strasse 107, CH-8008, Zurich, Switzerland; email: [email protected]), C. B. Asmussen (Department of Ecology, Royal Veterinary and Agricultural Univer- sity, Rolighedsvej 21, DK-1958, Frederiksberg C, Denmark; email: [email protected]), A. Henderson (Institute of Systematic Botany, The New York Botanical Garden, Bronx, NY 10458-5126, U.S.A., email: [email protected]) & P. K. Endress (Institute of Systematic Botany, University of Zurich, Zollikerstrasse 107, CH-8008, Zurich, Switzerland; email: [email protected]). A revision of Asterogyne (Arecaceae: Arecoideae: Geonomeae). Brittonia 55: 326-356. 2003.--A taxonomic revision of the neotropical genus Asterogyne Hook. (Arecaceae: Arecoideae: Geon- omeae) is presented. The genus is characterized by one autoapomorphic character, the separation of anther thecae by a bifid connective, and the combination of small to medium-sized stems, bifid simple leaf blades, floral pits in bud covered by a rounded upper lip that overlaps a lower lip, and pistillate flowers with staminodial lobes that are displayed in a starlike shape. Five species are recognized in this treatment: one of them (Asterogyne martiana) is widely distributed from Belize in Central America to northern Ecuador, three species (A. ramosa, A. L~picata, and A. yaracuyense) are endemic to small areas in the Venezuelan Coastal Range, and one species (A. guianensis) is endemic to eastern French Guiana. The taxonomic history, morphology, reproductive biology, distribution and ecology, intrageneric relation- ships, and conservation status are reviewed.
Key words" Arecaceae, Palmae, Geonomeae, Asterogyne, Neotropics.
Stauffer, E W. (Institute of Systematic Botany, University of Zurich, Zolliker- strasse 107, CH-8008, Zurich, Switzerland; email: [email protected]), C. B. Asmussen (Department of Ecology, Royal Veterinary and Agricultural Uni- versity, Rolighedsvej 21, DK-1958, Frederiksberg C, Denmark; email: [email protected]), A. Henderson (Institute of Systematic Botany, The New York Bo- tanical Garden, Bronx, NY 10458-5126, U.S.A.; email: [email protected]) & R K. Endress (Institute of Systematic Botany, University of Zurich, Zollikerstrasse 107, CH-8008, Zurich, Switzerland; email: [email protected]). A revi- sion of Asterogyne (Arecaceae: Arecoideae: Geonomeae). Brittonia 55: 326--356. 2003.--Se presenta una revisi6n taxon6mica del gdnero neotropical Asterogyne Hook. (Arecaceae: Arecoideae: Geonomeae). El g6nero se caracteriza por un car- ficter autoapom6rfico, tecas separadas pot un conectivo bffido, ademfis de tenet tallos de pequefio a mediano tamafio, lfiminas foliares enteras y bffidas, f6veas florales cubiertas por un labio superior redondeado que sobrelapa el labio interior, flores pistiladas con 16bulos estaminodiales dispuestos de manera estrellada. Para este tratamiento se reconocen cinco especies: una de ellas (Asterogyne martiana) ampliamente distribuida desde Belice en Amdrica Central hasta el norte de Ec- uador, tres especies (A. ramosa, A. spicata, and A. yaracuyense) enddmicas de pequefias fireas en la Cordillera de la Costa de Venezuela y una especie (A. gui- anensis) enddmica del oriente de la Guyana Francesa. Se revisa la historia tax- ondmica, morfolog/a, biologfa reproductiva, distribuci6n, ecologfa, relaciones in- tragendricas y estado de conservaci6n.
Brittonia. 55(4), 2003, pp. 326 356. ISSUED: 07 October 2003 �9 2003, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A.

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 327
1
1 P
41 i
D
A I B
i j
' : . ' : :7c '=~ 22o~2,2~'::z'=~ . _ . c c. ' : : ; ~ - ' ' : :x . . . . . . . . . . - -
F I G . 1. Type specimens of Asterogyne. A, B. Isotype of A. guianensis (de Granville 7124, K); C. Isotype of A. martiana (Wendland 56, K/; D-F. Isotype of A. ramosa (Steyermark et al. 96088, VEN). Scale bars: 8 cm.
The small genus A s t e r o g y n e H. Wendl. ex Hook. f. is one of six genera in the neo- t ropical tribe Geonomeae (Dransfield & Uhl, 1998; Henderson et al., 1995) and consists of five species in Central Amer i ca and northern South Amer i ca (Figs. 1 A - E 2 A - F , 4). In a let ter to J. D. Hooker in 1883, Wendland p roposed A s t e r o g y n e as a new genus to be inc luded in the monu- men ta l t r e a t m e n t o f G e n e r a P l a n t a r u m (Bentham & Hooker, 1883). The genus was descr ibed with several character states
a l ready known in other genera of Geono- meae, e.g., bifid s imple leaf blades and sta- minodia basa l ly united into a tube. Al - though modern t reatments of the genus have proposed several characters to differ- entiate A s t e r o g y n e f rom other genera of Geonomeae , floral pits in bud covered by a rounded upper l ip that overlaps the lower l ip (Fig. 9D) and free thecae on a bifid connect ive (Figs. 8B, 10D, 12C, 14C), have a lways been regarded as the two most r e l e v a n t f e a t u r e s (Bur re t , 1930; Hen -

328 BRITTONIA [VOL. 55
': . < i j
i FIG. 2.
Holotype of A. yaracuyense (Steyermark et al. 100311, VEN). Scale bars: 8 cm. Type specimens of Asterogyne. A-C. lsotype of A. ~picata (J. Steyermark 90010, VEN); D-F.
derson et al., 1995, Moore, 1966; Uhl & Dransfield, 1987; Wessels Boer, 1968).
A s t e r o g y n e was published without a type species in the original description in 1883. The typification of the new genus was made two years later in 1885 by Wendland, with the transfer of G e o n o m a mar t iana to the ge- nus A s t e r o g y n e . The type specimen selected by H. Wendland for this taxon was obtained from a plant cultivated at the gardens of Herrenhausen (Fig. 1C). This plant at Her- renhausen originated from plants cultivated by L. van Houtte in Gent, apparently from seeds sent by Warscewicz during his trip to Central America (Wendland, 1856). AI-
though there was no original explanation of the generic name, it was probably suggested because of the starlike shape of the stami- nodial lobes of the pisti l late f lowers (Moore, 1978) (Figs. 7E 8D). Moore (1966) described the genus Ar i s t eyera , separating it from A s t e r o g y n e on the basis of differ- ences in the number of stamens and stam- inodes. Wessels Boer (1968) observed great plasticity of these characters, especially in flowers of Ar i s t eyera spicata, and consid- ered that they were inconsistent for a delim- itation at the generic level. He merged both genera and argued that the number of sta- mens and staminodes reported by Moore

2003] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 3 2 9
|
~ ~.,~/. ~ "* , , " .~ ~''~ ~
y - -
FIG, 3. Pollen of A s t e r o g y n e . A. Pollen grain of A. mar t iana . B. Detail of exine in A. mar t iana . C. Pollen grain of A. ramosa . D. Detail of exine in A. ramosa . E. Pollen grain of A. ,spicata. F. Detail of exine in A. spicata . G. Pollen grain of A. y a r a c u y e n s e . H. Detail of exine in A. y a r a c u y e n s e . Scale bars: A, C, G = 5 p~m; B , D , E H = 1 btm;E = 6btm.

330 BRITTONIA [VOL. 55
2 0 ~ 90~ J / ~ 80~ , '-~_ ~ 70~ 60 ~
)
10 o - A ~ ~
5 o
Asterogyne
r162 A. guianensis "~ �9 A. martiana
FIG. 4. Distribution of Asterogyne in Central and South America.
78 t Aslerogyne guianensis
A. spicata
A. martiana
97 90
A. YOOTOSa
A. .vs
Geonoma interrupta
C~/vptron~mu~ I'ivalis
FIG. 5. Strict consensus tree of the two most parsimonious trees based on a phylogenetic analysis of mor- phological data. Bootstrap values are given on the branches.
(1967) for Asterogyne ramosa filled the gap between the two genera. Due to the vari- ability of stamens and staminodes of Aster- ogyne, Wessels Boer (1968, 1988) did not recognize the two populations in the states of Sucre and Yaracuy of the Venezuelan Coastal Cordillera (Fig. 4) as representing two different species, but included them in a broadly defined A. ramosa. After the de-
scription of the widespread type species, A. mart iana (Fig. 4), in 1856, four additional taxa were added to the genus within a rel- atively short period, from 1967 to 1988. These recently discovered species show re- markably limited distributions for the neo- tropical palms, and compared to A. marti- ana, they are known from few herbarium specimens (limited to two for A. guianen-

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 331
FIG. 6. Habit ofAsterogyne guianesis. (Photo: J. J. de Granville, Camopi River, French Guiana). Scale bar: 80 cm.
sis); thus their morphological and anatom- ical features are incompletely known. This revision aims to improve the taxonomic knowledge of the genus and to compile and reevaluate the published information on its systematics and natural history.
Materials and Methods
Morphology and distribution of the spe- cies were studied based on 334 specimens (see Exsiccatae) deposited at AAU, B, BH, BM, C, CAY, CR, G, K, MER, MO, M Y E NY, R U, US, VEN, WlS, and Z. A review of critical literature for the genus (Burret, 1930; de Granville & Henderson, 1988; Henderson et al., 1995; Uhl & Dransfield, 1987; Wessels Boer, 1968) was made, Ad- ditional information on the distribution of A. martiana was obtained from the curators at BRH, PMA, and SCZ, and of A. gui- anensis f rom J. J. de Granville (CAY). Specimens were collected in Venezuela by the first author in 1996, 1997, 2001, and 2002. For scanning electron microscopy, the dissected specimens were dehydrated,
critical-point dried, and sputter-coated with gold. Micrographs were obtained with a Cambridge $4 scanning electron micro- scope.
For anatomical investigations, flowers in bud or at anthesis were dehydrated and em- bedded in Kulzer 's Technovit 7100 (2-hy- droxyethyl methacrylate, HEMA). The ma- terial was sectioned at 7 microns using a rotary microtome, then stained with ruthe- nium red and toluidine blue and embedded in Histomount.
A prel iminary phylogenetic analysis o f all species of Asterogyne was made for this revision. Asterogyne was recently shown to be sister to a clade formed by Geonoma and Calyptronoma (Asmussen & Chase, 2001), therefore, we have irlcluded Geonoma in- terrupta and Calyptronoma rivalis as out- groups. Nineteen morphological characters (Table I), 10 of them parsimony-uninfor- mative, were included in the data matrix (Table II) constructed in McClade 4.0b10 (Maddison and Maddison, 2000). Clear gaps between the character states of quan- titative features were observed. A parsi- mony analysis was performed using Paup 4.0 (Swofford, 1998). All the characters were treated as "unordered" and were equally weighted. Support for the clades was calculated by doing a 10,000 replicate bootstrap analysis using the branch and bound option.
Morphology and Anatomy
STEMS AND ROOTS
Stems of Asterogyne are solitary and mostly erect; only A. martiana and A. yar- acuyense may be slightly decumbent at the base. Basal vegetat ive branching was de- scr ibed by H e n d e r s o n and S t e y e r m a r k (1986) for A. yaracuyense but the branches do not fully develop into mature stems. Lat- eral vegetative branches are normally pre- sent in A. yaracuyense and are especially numerous in A. ramosa, in which 5 to 30 branches were observed, attached to the stem, 2 0 - 4 0 cm below the crownshaft (Fig. 9A). Each of these lateral branches had 4 to 7 small bifid simple leaf blades, and were richly rooted. Our observations showed that these lateral branches do not develop into

332 BRITTONIA [VOL. 55
FIG. 7. Habit, inflorescence, flowers, and fruits of Asterogyne martiana. A, B. Habit. C. Inflorescence. D. Detail of rachilla at staminate anthesis. E & F. Detail of rachilla at pistillate anthesis. G. Rachilla with fruits. (Photos: R. Schmid, La Selva, Costa Rica). Scale bars: A, B = 1.2 m: C = 5 cm: D = 0.4 cm: E, E G 0.3 cm.
new plants. S tem ana tomy remains un- k n o w n for the genus.
Adven t i t i ous roots that form a basal cone are present in several species. In Asterogyne guianensis, the aerial roots are r emarkab ly long, to 90 cm, and conta in we l l -deve loped white pneuma todes (Fig. 6). Seuber t (1998) studied the root ana tomy of A. martiana and found that it is s imilar to that of other Geonomeae . The rh izodermis cells are thin- wal led and disappear in mature roots. In t ransverse section, the exodermis and the outer cortex, which is reduced to a few lay- ers, are not dist inct . In longi tud ina l section, the exodermis and outer cortex are distinct. The cells of the one- layered exodermis are stretched with b lun t ends; those of the outer cortex are long stretched with po in ted ends.
The i nne r cortex is usual ly homogeneous , and the ae renchyma, only weakly devel- oped, ma y occur in the center. In contrast to all other genera of the Arecoideae , the cells o f the inner cortex of the G e o n o m e a e are m o d e r a t e l y t h i c k - w a l l e d , a nd th in - wal led cells are lacking. Narrow fibers are scattered throughout the inner cortex. En- d o d e r m i s ce l l s are t a n n i f e r o u s , r a d i a l l y stretched, and u-shaped, with thin outer walls and thick inner walls. The per icycle is one- layered and the vascular cy l inder shows the no rma l pattern observed in the fami ly (Seubert , 1998).
LEAVES
The leaves, at least the young ones, in Asterogyne are spirally arranged and rather

2003] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 3 3 3
i\i:l 7 (
FIG. 8. Flowers in Asterogyne martiana. A. Staminate flower. B. Stamen. C. Epidermis of the filament. D. Pistillate flower. E. Ovary in abaxial view. F. Ovary in adaxial view. G. Protuberances at the base of the styles. H. Stigmatic branches at anthesis. I. Cross section of the trilocular ovary, arrow points to the septal nectary. J . Cross section of the secretory epi thel ium of the nectary. (From Knudsen & Asmussen 604, AAU). Scale bars: A, D, H = 1 mm; B = 300 ~m; C = 25 ixm: E, E 1 - 200 ttm; G 12 I~m: J 50 ~rn.

334 BRITTONIA [VOL. 55
FIG. 9. Habit, leaf, and inflorescence of Asterogyne ramosa. A. Habit. B. Detail of blade. C. Inflorescence at anthesis. D. Detail of rachillae in bud. E. Detail of rachilla at pistillate anthesis. F. Detail of rachilla at staminate anthesis. (Photos: E J. Stauffer, Cerro Humo, Venezuela]. Scale bars: A = 28 cm: B = 18 cm; C = 5 cm; D 7 ram; E 2 ram; F 3 ram.
erect and often purplish. The b lade is nor- mal ly bifid and simple but it may be lon- g i tud ina l ly split with age or damaged by the wind (Figs. 6, 7A). Longi tud ina l splits were especia l ly consp icuous in some col lec t ions of A. martiana (Stevens 13505; Quesada 43) f rom Costa Rica, and A. ramosa (Stey- ermark & Liesner 120915) and A. yaracu- yense (Stauffer et al. 267) f rom Venezuela . Accord ing to Wessels Boer (1968) the blades o f A. martiana and A, spicata show a hypodermis (sensu Toml in son 1961, i.e.,
a colour less hypodermal cell layer) on ly on the abaxia l side. The mesophyl l lacks a dis- t inct pa l isade layer and the cells are some- what t ransverse ly extended. The fibers are numerous , of ten solitary, each with a rather wide lumen , scattered th roughout the me- sophyll but c o m m o n l y found in the adaxial layer. Some of the vascular bund les are very small , not p roducing r ibb ing of the lamina . Wessels Boer (1968) noted that the leaf ana tomy of Asterogyne shared most o f the characters already obse rved in other

2003] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 335
: .~c'-'~ H ~ , ' r ~ ' ~ ~ ; ~ j , ~ ~,~ ~--'~
FIG. 10. Flowers of Asterogyne ramosa. A. Staminate flower in bud. B. Staminate flower at anthesis. C. Pistillate flower in young bud. D. Stamen. E. Ovary in abaxial view. F. Gynoecium shortly before anthesis. G. Epidermis of the filament. H. Longitudinal section of the ovule, arrow points to the micropyle. I. Cross section of the trilocular ovary. . I . Cross section of the style and the staminodes, arrow points to the raphide idioblasts in the mesophyll of the staminodes. K. Cross section of the style showing the ventral slits. (From Stauffer et al. 824, Z). Scale bars: A, D = 450 ixm; B, C 1 ram: E, H - K = 400 p~m: F 500 txm; G = 25 l, zm.

336 BRITTONIA [VOL. 55
FIG. l l . Habit, leaf, inflorescence, and fruits of Asterogyne spicata. A. Habit. B. Detail of blade. C. Inflo- rescence and infructescence. D. Detail of rachilla at staminate anthesis. E. Detail of rachilla with fruits. (Photos: E J. Stauffer, Parque Nacional Guatopo, Venezuela). Scale bars: A = 1.2 m: B = 20 cm: C = 8 cm; D, E - 4 mm.
Geonomeae , but pointed out that the one- c e l l - l a y e r e d abax i a l h y p o d e r m i s of the blades could be interpreted as less devel- oped wi th in the tribe. Vegetat ive ana tomy of A. spicata (as Aristeyera spicata) was studied by Toml inson (1966). His obser- vat ion of two wide vessels per bund le in the petiole ofA. spicata was also found in other species of Geonomeae by Klotz (1978).
INFLORESCENCES AND FLORAL TRIADS
Inflorescences of Asterogyne are always interfoliar, solitary, and pro tandrous (Figs.
7A, 9A, 11A). Once -b ranched inf lorescenc- es are presen t in A. martiana, A. ramosa, and A. yaracuyense (Figs. 7C, 9C, 13B), while A. guianensis and A. spicata show wel l -deve loped spikes, which are thyrses in a compara t ive morpholog ica l sense (Figs. 1B, 2C, 11C). A remarkable dev ia t ion f rom the b ranched inf lorescences was obse rved in some col lec t ions of A. martiana restrict- ed to the Coma r c a of San Blas in P a n a m a (e.g., Elias 1769; de Nevers et al. 5209, 5881, 7914", McDonagh, J. et al. 246) and C o l o m b i a (Duke 11338), in which the in-
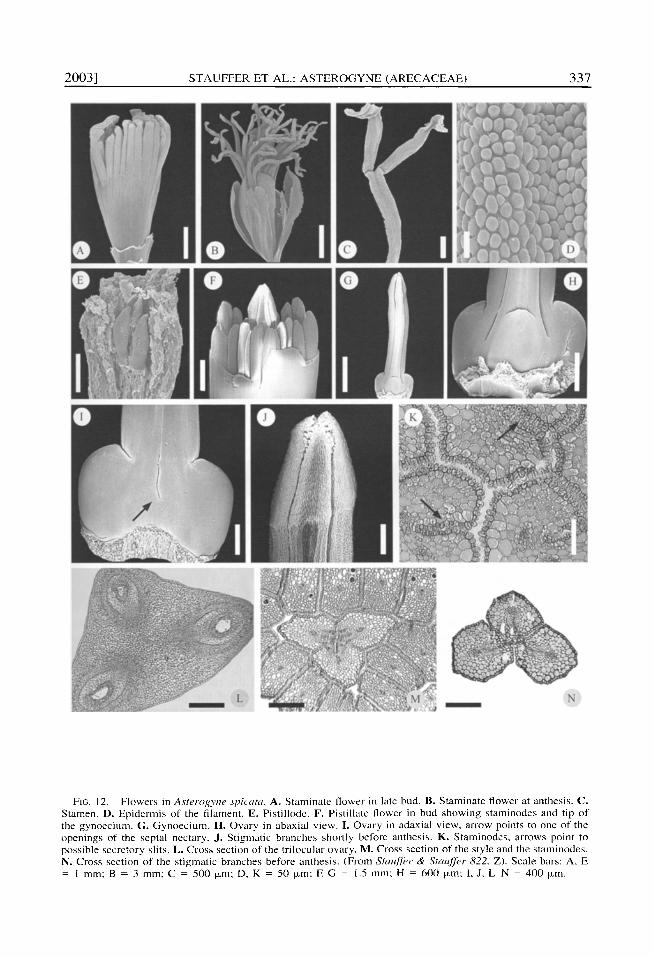
2 0 0 3 ] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 3 3 7
D I
l i ~i �84 i .... ~i:" . . . . ,
N m
FIG. 12. Flowers in Asterogyne .v~icata. A, Staminate flower in late bud. B. Staminate flower at anthesis. C. Stamen. D. Epidermis of the filament. E, Pistillode. F. Pistillate flower in bud showing staminodes and tip of the gynoecium. G. Gynoecium. H. Ovary in abaxial view. I. Ovary in adaxial view, arrow points to one of the openings of the septal nectary. J. Stigmatic branches shortly before anthesis. K. Staminodes, arrows point to possible secretory slits. L. Cross section of the tr i locular ovary. M. Cross section of the style and the staminodes. N. Cross section of the s t igmatic branches before anthesis. ~From Stm~ler & Stauffer 822, Z). Scale bars: A, E
1 mm: B = 3 ram; C 500 ixm: D, K = 50 p,m: E G = 1.5 l n m : H = 600 p,m; I , J , L - N = 400 p,m.

338 BRITTON|A [VOL. 55
FIG. 13. Habit and inflorescence of Asterogyne yaracto,ense. A. Habit. B. Inflorescence. C. Detail of rachilla at staminate anthesis. (Photos: E J. Stauffer, Cerro Humo, Venezuela). Scale bars: A = 30 cm; B 8 cm; C 3 mm.
florescences are spicate (Fig. 7B). In one of these collections (de Nevers et al. 5881) the spike is rather thick.
Morphology and anatomy of the inflores- cence axis and especially the floral vascu- lature of Asterogyne spicata (as Aristeyera spicata) was examined by Ubl (1966). The vascular bundles of the flowers of this spe- cies are small because the xylem does not contain large vessels and has a few helical elements. The number of vascular traces to perianth parts is variable. In the petal tube of staminate flowers and sepals of pistillate flowers, some of the traces appear to dif- ferentiate over a long period of time and as a result the number of traces increases with the age of the flower. The traces developing later do not become connected to the vas- cular strands in the floral receptacles.
The peduncle of Asterogyne is long and slender. The prophyll is fibrous to membra- nous and normally inserted at the base of the peduncle or just above it. The inflores- cences are enclosed by one or two tubular, char taceous to membranous , peduncular bracts, Asterogyne ramosa and A. yaracu- yense always have two peduncular bracts, whereas the species with spicate inflores- cences and A. martiana (with the exception of one collection from Panama [Pittier 4226], which has two peduncular bracts),
show a reduction to only one. Moore (1967) suggested that the reduction in the number of bracts observed in A. spicam (as Aris- teyera spicata) represented a derived char- acter within the genus. As first reported by Wessels Boer (1968) for A. martiana, all the species studied in the field showed that the small unexpanded inflorescence remains straight and unfolded within closed bracts; during expansion it elongates rapidly and emerges through a terminal split of the per- sistent bracts. The inflorescences are always erect at anthesis and become pendent in fruit (Fig. 7B). In branched inflorescences the rachis is normally well developed, but in A. martiana (Cuatrecasas 17375; Knapp 3777; Moore 6530) it may be almost absent and then all the rachillae appear to emerge from the apex of the peduncle. The inflo- rescence raches are normally salmon to pink at anthesis and become brown to red- dish brown in fruit. Floral pits are spirally arranged on the rachillae. As described by Uhl and Dransfield (1987) for Welfia, Ca- lyptrogyne, and PholMostachys, the foral pits in bud of Asterogyne are covered by an immersed, rounded, lower lip, overlapped by a short upper lip (Fig. 9D). Wessels Boer (1968) interpreted the immersed lower lips as less specialized in a morphological sense and therefore as phylogenetically basal

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 339
FIG. 14. Flowers of Asterogyne yaracuyense. A. Staminate flower in late bud. B. Staminate flower at anthesis. C. Detail of stamen. D. Epidermis of the filament. E. Pistillate flower in young bud. F. Gynoecium in late bud. G. Abaxial view of the ovary. H. Adaxial view of the ovary. I. Stigmatic branches before anthesis. J. Cross section of the trilocular ovary. (From Staz~fJer &Smuffcr 823, Z). Scale bars: A 900 Ixm: B, E l ram: C
400 ~Lm: D = 50 ixm; F 450 gtm : G, H, J = 200 ~xm; I = 300 Ixm.
wi th in the tribe. The l ower lip rolls back when the f lowers b e c o m e exserted. The un- c o m m o n spicate in f lo rescence o b s e r v e d in one co l lec t ion o f A. mart iana (ide Nevers et al. 58811) also has the lip margins dense ly cove red by short whi t i sh hairs. The hairs are lacking in the no rma l -b r anched inflo- rescences on other plants o f this species. As is c o m m o n in the Areco ideae , the pits con- tain triads c o m p o s e d o f one pist i l late and two s taminate f lowers . Each triad is sunken in the floral pi t and represents a short c in-
cinnus. Based on the vascula ture , however , Uhl (1966) sugges ted that the triads in As- terogyne (as Aris teyera) or ig inated f rom a more ramif ied type o f partial inf lorescence . As also repor ted by Stauffer et al. (2002) for Geonoma interrupta, the first s taminate f lower o f the c inc innus , and the c inc innus as a whole , is sub tended by the p i t -c los ing bract, which represents the l ower lip o f the pit. A c c o r d i n g to the interpreta t ion o f Wes- sels B o e r (1968), the largest bract ins ide the pit subtends the second s taminate flower, a

340 BRITTONIA [VOL. 55
TABLE I CHARACTERS AND UNORDERED CHARACTER STATES
1. Leaf blade: (0) entire-bifid: (1) divided 2. Inflorescence branching: (0) branched; (1) spicate 3. Upper lip of the pit: (0) absent: (1) present 4. Peduncle (length): (0) <80 cm; (1) -->85 cm 5. Peduncular bracts (number): (0) one: (1) two 6. First peduncular bract (length): (0) --<60 cm; (1) >64 cm 7. Number of stamens: (0) 6: (1) 8-10; (2) 11-12; (3) 21-27 8. Filament color: (0) white; (1) yellow 9. Filament surface: (0) smooth; (1) papillate
10. Anther shape: (0) divided: (1) with a bifid connective: (2) sagittate 11. Complete fusion of the staminodial tube with the corolla: (0) absent; (1) present 12. Upper part of the sterile androecium: (0) calyptrate: (1) lobed; (2) crenulate 13. Complete fusion of the petals of the pistillate flowers in a tube: (0) absent; (1) present 14. Gynoecium: (0) with 3 more or less equally developed ovaries: (1) pseudomonomerous 15. Central protrusion between the three carpels: (0) absent: I l) present 16. Position of the ovule: (0) at midlength of the locule: (1) basal 17. Lateral vascular bundles in the styles at anthesis: (0) absent: (1) present 18. Upper part of the style curved: (0) absent: (1) present 19. Fruit length: {0) 0.3-7 cm; (1) 1.2 cm: (2) 1.5-1.8 cm: (3) 2.5 cm
m e d i u m - s i z e d bract subtends the pist i l late flower, and a smal l bract represents the pro- phyl l of the pist i l late flower.
STAMINATE FLOWERS
Two s tamina te flowers occur la teral ly to the pist i l late flower. The per ianth is com- posed of three basal ly uni ted and apical ly free sepals and three basal ly un i ted and api- cally valvate petals (Figs. 8A, 10B, 12B, 14B). The n u m b e r of s tamens ranges from six (Asterogyne martiana, Fig. 8A) to 27 (A. guianensis); the higher n u m b e r of sta- mens be ing present in species with spicate inf lorescences . Taxa of G e o n o m e a e with u n b r a n c h e d inf lorescences and flowers with more than six s tamens are rather u n c o m - mon. They are present in only two species of Asterogyne (A. guianensis and A. spica- ta', Fig. 12B) and three species of Geonoma (G. camana Trail, G. chococola Wess. Boer, and G. polyandra Skov). Wessels Boer (1968) cons ide red the increased n u m b e r of s tamens in some species of Asterogyne (as Aristeyera) as a der ived character state wi thin the tribe. Based on the increased n u m b e r of s tamens in A. spicata, Moore (1966) sugges ted a closer re la t ionship be- tween Asterogyne (as Aristeyera) and Wel- fia, than with the other G e o n o m e a e . The s tamens are un i ted into a tube up to mid- length and free near the top (Figs. 8A, 10A,
10B, 12B, 14A). Anthers of A. yaracu- yense, and especial ly A. martiana, are ver- satile because of the th inner apical regions of the f i laments (Fig. 8B). Wessels Boer (1968) found that the j u n c t i o n be t w e e n the f i lament and the connec t ive in Asterogyne is less r igid than in Geonoma. The separa- t ion of an ther thecae on a bifid connec t ive in Asterogyne (Figs. 8B, 10D, 12C, 14C) has been cons idered un ique wi th in the G e o n o m e a e (Uhl & Dransfield, 1987). The connec t ive is consp icuous ly swol l en in A. martiana. The anthers are inf lexed toward the center of the flower and introrse in bud but spread and extrorse at anthesis. A three- lobed pis t i l lode is normal ly fused with the s taminodia l tube to its middle (Fig. 12E). Morpho logy and vasculature of the stami- nate f lowers of A. spicata (as Aristeyera spicata) were studied by Uhl (1966) and Moore (1966L and A. martiana by Uhl and Moore (1977).
P O L L E N M O R P H O L O G Y
Pol len in Asterogyne has been descr ibed by Uhl and Dransf ie ld (1987) as elliptic, monosu lca te , with tectate exine, which is vermicula te or reticulate. In a detai led study of the G e o n o m e a e , Punt and Wessels Boer (1966) stated that Asterogyne species show two dif ferent po l len types, the "'Asterogyne type ," occur r ing in A. martiana and the

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 341
T A B L E I I
M A T R I X OF 19 M O R P H O L O G I C A L C H A R A C T E R S ,SCORED FOR THE FIVE SPECIES OF Asterog)'tze AND T W O O U T G R O U P
T A X A (Geonomct interrw~ta A N D CLltVtgtotloltlCt rivalis). T H E Q U E S T I O N MARK ( ' ? ) INDICATES DATA N O T A V A I L A B L E
Character
Species 1 2 3 4 5 6 7 8 9 If) 11 12 13 E4 15 16 17 18 19
A. gu ianens i s 0 t 1 0 0 0 3 0 0 1 1 1 0 0 "? 0 ? 0 3 A. mar t iana 0 0 1 0 0 0 0 0 0 1 1 1 0 0 1 0 0 0 1 A. ramosa 0 0 1 1 1 0 2 0 0 l 1 1 0 0 1 0 0 0 2 A. spicata 0 1 1 0 0 0 3 l 1 1 1 1 0 0 1 0 1 0 2 A. ya racuy 0 0 1 1 1 0 1 0 0 1 1 1 0 0 1 0 0 0 9 G. interrupta 1 0 0 0 0 0 0 0 0 0 0 2 0 1 1 1 0 0 0 C. rivalis 1 0 0 0 0 1 0 1 0 2 0 0 1 0 0 0 0 1 0
"Calyptronoma occidentalis type," occur- ring in A. spicata (as Aristeyera spicata).
Pollen of four of the five species of As- terogyne was studied for this revision (,4. martiana, Knudsen & Asmussen 604; A, ra- mosa, Smt~'er et al. 824; A. spicata, Stauf- fer et al. 822; A. yaracuyense, Stauffer & Stauffer 823). The pollen (Fig. 3 A - H ) is asymmetr ic and monosulcate with a sulcus length as long as the main axis, The exine is tectate and finely perforate-rugulate (per- forate fossulate in A. spicam). This agrees with the study of Harley (unpubl.) of A. martiana (Schipp 392). Punt and Wessels Boer (1966) described the pollen of A. mar- tiana (Archer 1978, Schipp 392) as easily recognized by its fine, but distinct reticu- lum, and the distinctly visible muri (bacu- la), which support the reticulum. The pollen ofA. spicata (Tamayo 4177) was described by Punt and Wessels Boer (1966) as having a distinctly vermiculate ornamentation. Re- sults of our study could not differentiate the two groups proposed by Punt and Wessels Boer (1966) and show that the pollen is rather morphological ly uniform within the genus.
PISTILLATE FLOWERS
The perianth of the pistillate flowers is composed of three free, imbricate sepals and three petals, which are united to their middle (Fig. 8D). The carpels alternate with the petals. The ovary is superior and triloc- ular (Figs. 8E, 8I, 10E, 10I, 12H, 12L, 14G, 14J). Each carpel bears a pendent, anatro- pous, crassinucellate ovule (Fig. 10H). One of these per gynoec inm develops fully to maturity. Only one well-differentiated in-
tegument could be identified. It is five to six cell layers thick at midlength and six to ten cell layers thick toward the micropyle, The nucellus protrudes into the micropyle, which faces the ventral side of the locule. A well-developed septal nectary occurs be- tween the carpel flanks (Fig. 8I). It becomes a triradiate cavity at about midlength of the ovary and extends upward and gradually outward to the apical region of the ovaries. The epithelium, especially differentiated in the gynoecium ofA. rnartiana, is formed by large columnar, uninucleate cells (Fig. 8J). The three openings of the nectary are to- ward the top o f the ovary and are nonsec- retory ducts lined by isodiametric epider- mal cells. Even though Asterogyne was de- scribed (Uhl and Dransfield, 1987; Wessels Boer, 1968) as having apical styles, our study shows they are basal to lateral, which is probably a common pattern within Geon- omeae (Stauffer & Endress, unpubl, data). Epidermal protuberances at the basal and midregion of the styles were observed in anthetic flowers of A. martiana (Schmid, 1970a) and A. yaracuyense (Stauffer & En- dress, unpubl, data) (Fig. 8G). These pro- tuberances are absent in young buds and seem to develop shortly before anthesis. The three stigmatic branches are papillate and reflexed at anthesis (Fig. 8H). The staminodes are united to their middle into a tube and free above this level (Figs. 8D, 10C, 14E). The number of staminodial lobes ranges f rom five in A. ramosa to 22 in A. guianensis. A well-developed papil- late epidermis in the staminodial lobes is present in A. spicata (Fig. 12K). Reduction of the number of staminodes with respect

342 BRITTONIA [VOL. 55
to the number of fertile stamens in the same species is common within the genus; how- ever, in Asterogyne martiana the number of fertile and sterile stamens is a lways six (Fig. 8A, D). The morphology and vascu- lature of the pistillate flowers of A. spicata were studied by Uhl (1966) and for A. mar- tiana by Uhl and Moore (1977). The struc- ture of the gynoecia in four species of As- terogyne is part of a current study including representatives of all Geonomeae (Stauffer & Endress, unpubl, data). The chromosome number in the genus is unknown (M. R6ser, pets. comm.) .
FRUITS
Fruits o f Asterogyne are ellipsoid to el- lipsoid-ovoid, purplish, black, or garnet red drupes, with a smooth surface and a single seed (Figs. 7G, 11E). They are conspicu- ously keeled at the apex. The remains of the st igma are basal. The seed is ellipsoid and slightly dorsiventrally compressed. The endosperm is homogeneous (not ruminate), and the embryo is basal (Uhl & Dransfield, 1987). The morphology and ana tomy of the fruits in A. martiana and A. spicata were studied by Wessels Boer (1968). Fruits of A. martiana resemble those of A. spicata, but the arrangement of the vascular bundles in the pa renchyma of the mesocarp is less regular. In A. spicata, bundles of fibers without conductive elements are present in the outer layer and the endocarp is not ad- herent to the seed (Wessels Boer, 1968).
Germinat ion is adjacent-ligular (Uhl & Dransfield, 1987). Seeds ofA. spicata have been reported to germinate within three and four months (Braun, 1970; Nehlin, 1994).
Reproductive Biology What we know about the reproductive bi-
ology of Asterogyne is limited almost en- t i re ly to A. mar t iana (Schmid , 1970a, 1970b; Uhl & Moore, 1977). Asterogyne is m o n o e c i o u s and the in f lo re scences are markedly protandrous. Male and female flowers on the same inflorescence of A. martiana are rarely open at the same time, thus maximizing pollen transfer among dif- ferent plants (Schmid, 1970a). In the spe- cies in Venezuela usually just one inflores-
cence of a plant is in flower at a time, while the other inflorescences are still in bud or in fruit. This pattern was already observed in A. martiana by Schmid (1970a) and may be common for the other species in the ge- nus. Staminate flowers emerge first, one af- ter another, and when both of them have fallen, the pistillate flower emerges. Two staminate flowers at anthesis in the same pit were occasionally observed in A. yaracu- yense and A. martiana (Schmid, 1970a). In A. martiana the anthesis of individual sta- minate flowers lasts from two to six days and the anthesis of the pistillate flowers then follows and lasts one to two days (Schmid, 1970a; Uhl & Moore, 1977). Young fruits were observed to occur si- multaneously with pistillate flowers on the rachillae of A. martiana (Bonifaz & Cor- nejo 3758).
Several structural features of Asterogyne f lowers sugges t insect po l l ina t ion . The well-developed septal nectary (Fig. 8I) ob- served in the pistillate flowers of four of the five species in the genus (pistillate flowers of A. guianensis were not available for the structural study), and the rudimentary septal nectary observed by Schmid (1970a, 1983) in the pistillode of the staminate flowers of A. martiana, may provide nectar for polli- nators. Schmid (1970b) found that the translucent protuberances at the base of the styles and the fleshy staminodes in A. mar- tiana readily exude fluid when slightly damaged. The production of fragrance is well known in both staminate and pistillate flowers of Asterogyne but its histological basis is unknown. Floral fragrances were found in the male flowers of all of the Ve- nezue l an species . Scent was e spec i a l l y strong in A. spicata, and has also been re- ported in A. martiana (Knudsen, 1999; Schmid, 1970a, 1983; Knudsen & Asmus- sen 620; Moore 6535). The presence of os- mophores was preliminarily tested on a vis- it by the first author (between 2001 and 2003) to the Venezuelan populations of As- terogyne. The staminate flowers of A. spi- cata that were submerged for five minutes in a water solution containing neutral red showed traces of the dye at the base of the petals and filaments. Floral scent composi- tion in A. martiana and its potential impor-

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 343
tance in maintaining reproductive isolation were studied by Knudsen (1999).
Insect pollination of Asterogyne marti- ana was discussed by Schmid (1970b). In two populations in Costa Rica, he found in- sects of five different orders visiting the flowers; syrphid flies probing for nectar were considered effective pollinators for the species. The flies also visited staminate flowers to eat pollen. Syrphid flies have also been reported by de Nevers & Hen- derson 6323 to visit flowers of A. martiana in Panama. Ants (Formicidae, Hymenop- tera) were reported by Schmid (1970b) to visit inflorescences ofA. martiana (Fig. 7E) and were observed by the first author in sta- minate anthesis in A. spicata. Asterogyne spicam has bright orange staminate flowers, which emit a very strong, sweet scent and are visited by bees (Henderson, 1986). Dis- persal of the fruits has never been studied, but the ripe, dark purplish, black, or garnet red fruits contrasting with the brown, red- dish brown, reddish, or salmon to pink rachillae suggest bird dispersal.
Distribution and Ecology
The five species of Asterogyne are dis- tributed in Central America and northern South America (Fig. 4). They are understo- ry palms; A. martiana and A. ramosa oc- casionally are dominant in that stratum of the forest. The only widespread species is A. martiana: the other four species are nar- row ecological endemics, restricted to small areas in Venezuela and French Guiana. As- terogyne martiana grows from Belize to northern Ecuador, usually forming large stands in the understory of the lowland non- flooded rain forest, but sometimes also along streamlets and seasonally flooded patches of forest. It has also been collected in the cloud forests of Costa Rica and Co- lombia, sometimes to 1400 m.
Asterogyne ramosa, A. spicata, and A. yaracuyense are restricted to the Venezue- lan Coastal Cordillera (Fig. 4). Asterogyne ramosa is known only from the cloud for- ests of the Peninsula de Paria, state of Su- cre, at 730 to 1250 m elevation. There it forms large stands in particularly moist ar- eas of the Cerro Humo and grows in asso-
ciation with Euterpe precatoria var. longev- aginata (Mart.) Henderson, Geonoma inter- rttpta (Ruiz & Pay.) Mart. var. interrupta, and especially Prestoea pubigera (Griseb. & H. Wendl.) Hook. f. ex Benth. & Hook. f. The general habit and the bifid simple leaf blades of P. pubigera surprisingly re- semble A. ramosa and this might have ob- scured the recognition of the latter in the relatively nearby cloud forest of Trinidad. As pointed out by Henderson and Steyer- mark (1986), all the Venezuelan popula- tions of Asterogyne are located in three plant refugia of the Coastal Cordillera (Nir- gua, Guatopo, Paria) mentioned by Steyer- mark (1979). Asterogyne spicam is endemic to the Guatopo National Park and was re- cently collected in the forests NW of Gua- tire, in the state of Miranda, where it grows at 600 to 1100 m and is restricted to the ecotone between the lowland rain forest and the low altitude cloud forest. In the locali- ties that the first author has visited, Aster- ogyne spicata is rare and associated with Bactris setulosa H. Karst., Euterpe preca- toria var. Iongevaginata, and Geonoma un- data Klotzsch. Asterogyne yaracuyense was once thought to be endemic to the cloud forest on the Cerro La Chapa (Yaracuy State), normally above 1200 m, but recent field work of W. Meier (pers. comm.) con- firmed its presence in the nearby cloud for- est on Cerro Zapatero. In Cerro La Chapa, A. yaracuyense is rare and usually associ- ated with DicO, ocarvum fuscum (H. Karst.) H. Wendl., Geonoma interrupta var. inter- rupta, G. spinescens var. braunii E W. Stauffer, Socratea altissima Balslev & E W. Stauffer (spec. ined.), and Wettinia prae- morsa (Willd) Wess. Boer. Appun (1871) provided a short description of a palm in the cloud forest of the Pico Hilaria (central Venezuelan Coastal Cordillera) that match- es an Asterogyne species, probably A. yar- acuyense.
After 20 years of intensive field work in French Guiana, J. J. de Granville discov- ered a filth species of Asterogyne, A. gui- attensis, from a single locality in the eastern part of the country, on the Brazilian border (Fig. 4). The genus had never been reported for the Guianas: its presence in French Gui- ana suggests it may occur in other regions

344 BRITTONIA [VOL. 55
of northeastern South America. The very limited range of A. guianensis may be re- lated to low adaptive and reproductive ca- pacity (de Granville and Henderson, 1988). Asterogyne guianensis is found in the Oya- pock refugium described by Brown (1982), also known as the East Guiana refugium (Prance, 1982). De Granville and Hender- son (1988) suggest that A. guianensis and A. spicata may have once been a single population that became isolated as a result of climate changes.
In a study at La Selva Biological Station, Costa Rica, Braker and Chazdon (1993) as- sessed the levels of damage produced by the neotropical forest grasshoper Micro~- lopte~x hebardi (Acrididae: Ommatolam- pinae) that feeds on leaves of A. martiana and two species of Geonoma. Asterogyne martiana was the least preferred species, probably due to lower water content, lower fresh/dry mass per unit of leaf area, and higher crude lignin content.
The Placement of Asterogyne Based on Molecular Phylogenetic Analyses
The taxonomic position of Asterogyne within the tribe Geonomeae is corroborated by phylogenetic analyses of DNA sequenc- es. A clade of Geonoma congesta, Calyp- tronoma occidentalis, and Asterogyne mar- tiana is resolved as monophyletic in the analysis of three plastid DNA sequences, rbcL, rpsl6, and trnL-trnF by Asmussen and Chase (2001). Several other phyloge- netic analyses of palms have included A. martiana. Some of these studies (Hahn, 2002; Lewis & Doyle, 2001; Uhl et al., 1995) had A. martiana as the only repre- sentative for tribe Geonomeae and therefore did not add information about tribal rela- tionships of Asterogyne. In other studies (Asmussen et al., 2000; Baker et al., 1999), A. martiana and other representatives of Geonomeae were part of a poorly resolved clade of Arecoideae: these studies are also uninformative with respect to the position of Asterogyne.
In the family level study by Asmussen and Chase (2001), Asterogyne martiana is resolved as sister to a poorly supported clade of Geonoma congesta and Calyptron-
oma occidentalis. More taxa and more char- acters are needed to obtain a robust hypoth- esis of the relationships within Geonomeae. Two studies, one based on rpll6 and the other on rpsl6 plastid DNA sequences, spe- cifically addressed the relationships of the six genera o f G e o n o m e a e (Asmussen , 1999a, 1999b). However, Asterogyne was not included in the study based on rpll6 because it was not possible to get a rpll6 sequence for A. martiana at the time (As- mussen, 1999a). In the study based on rpsl6, Asterogyne was part of a poorly re- solved monophyletic group containing spe- cies of Geonoma, Calyptrogyne, and Calyp- tronoma, with Welfia and Pholidostachvs as sisters to this clade (Asmussen, 1999b). A number of studies of Arecoideae and Geon- omeae include Asterogyne, and a robust hy- pothesis of the position of Asterogyne can be expected to be published in the near fu- ture. A study (Lewis, unpubl, data) of DNA sequences of the nuclear gene phosphori- bulokinase from selected genera within sub- family Arecoideae resolves species of As- terogyne as distinct and Geonoma as a monophyletic group with Asterogyne as sis- ter to Geonoma. An unpublished study (Borchsenius et al., unpubl, data) based on morphological characters corroborates the result from the nuclear phosphoribulokinase DNA sequences. In conclusion, most of the data available, preliminary as they may be, point to a sister group relationship of Geon- oma and Asterogyne, with the remaining genera of Geonomeae outside this mono- phyletic clade.
Relationships within Asterogyne Based on Molecular Phylogenetic Analyses
Asterogyne martiana has a wide natural distribution and is widely cultivated in bo- tanical and private gardens. It is more dif- ficult to obtain material of the remaining four species of Asterogyne, which are re- stricted in their natural habitats and are not cultivated. This is the main explanation for the lack of phylogenies of Asterogyne spe- cies. Attempts made to extract DNA from herbarium material ofA. spicata and A. yar- acuyensis have failed, but succeeded only for A. ramosa (Asmussen & Deichmann,

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 345
pers. comm.). Based on preliminary data of plastid DNA, A. ramosa forms a clade with A. martiana that is sister to Geonorna (As- mussen et al,, unpubl, data),
Fresh leaf material for DNA analyses from Asterogyne guianensis, A. ramosa, A. spicata, and A. yaracuyensis is critical for examining the infraspecific relationships of Asterogyne. The combination of data from morphology, a number of variable DNA se- quences, and AFLP characters for all spe- cies of Asterogyne is likely to provide a ro- bust interpretation of the interspecific rela- tionships for the genus.
Relationships within Asterogyne Based on a Morphological Phylogenetic
Analysis
The strict consensus tree (Fig. 5, length: 26, CI: 0.962, RI: 0.889) based on mor- phology resolved Asterogyne as a well-sup- ported monophyletic group with a bootstrap value of 97%. However, it is clear that more outgroup taxa should be included in the analysis to test this hypothesis. Asterogyne guianensis and A. spicata form a clade with a bootstrap support of 78%: this group is characterized by spicate inflorescences and male flowers with more than 21 stamens. Asterogyne ramosa and A. yaracuyense form a clade with a bootstrap support of 90%; the species of this group grow in the cloud forest of the Venezuelan Coastal Cor- dillera and have a peduncle longer than 85 cm. The relationships between these two clades and A. martiana were unresolved (Fig. 5). The parsimony search detected two most parsimonious trees. One of the two most parsimonious trees had the same to- pology as the strict consensus tree, whereas the other showed A. martiana as sister to the remaining species of Asterogyne. Our phylogeny supports the close relationship between A. guianensis and A. spicata, al- ready suggested by de Granville and Hen- derson (1988). A clarification of the exact position of the widespread A. martiana will have to await additional data.
Conservation Status
Asterogyne martiana is a widespread species considered to be nonthreatened in
Central America and Colombia (Bernal, 1989). Most of the populations of A, spi- cam grow in the Guatopo National Park and therefore are protected by the rules of the national park, Asterogyne spicata is list- ed as vulnerable in the IUCN red list of threatened species (Johnson, 1996). The difficult access to the populations of A. ra- mosa in Venezuela and A. guianensis in French Guiana is a major factor contribut- ing to the conservation of these species. As- terogyne yaracuyense was also listed as vulnerable by Johnson (1996). A recent vis- it to its natural habitat on Cen'o La Chapa has confirmed more extensive alteration and destruction of the forest than originally re- ported by de Granville and Henderson (1988) and Stauffer 'and Duno de Stefano (1998). Small agricultural areas for the cul- tivation of bananas, coffee, and tubers have gradually taken possession of the cloud for- est atop Cerro La Chapa. Cattle raising and construction associated with agricultural ac- tivities have also altered the species content and structure of the cloud forest. Therefore, Asterogyne yaracuyense should be listed as critically endangered in the IUCN red list of threatened species.
Systematic Treatment
ASTEROGYNE H. Wendl. ex Hook. f.
Asterogyne H. Wendl. ex Hook. f. in Ben- tham & Hook. f., Gen. P1. 3: 914. 1883; Burret, Bot. Jahrb. Syst. 63: 140. 1930; Wessels Boer, Verh. Kon. Ned. Akad. Wetensch., Afd. Nat., 2 Reeks, 58: 79. 1968; Uhl & Dransfield, Genera Palma- ruin, 539. 1987; Dransfield & Uhl, Pal- mae, in Kubitzki (editor), Faro. gen. vasc. pl., ft. pl., 4: 385. 1998. TYPE: Asterogyne mart iana (H. Wendl.) H. Wendl. ex Hemsl. (Geonoma martiana H. Wendl.), typified by H. Wendl. ex Hemsl. in God- man & Salvin, Biol. Centr. Am. Bot. 3: 409. 1885.
Aristeyera H. E. Moore, J. Arnold Arbor. 47: 3. 1966. T'cPE: A. ~v)icata H. E. Moore.
Small to med ium-s i zed , sol i tary, un- armed, pleonanthic, monoecious palm. Stem smooth, brown to brownish cream, erect, sometimes basally decumbent, occasionally

346 BRITTONIA [VOL. 55
with basal and/or lateral vegetative branch- es, unarmed; internodes short; adventitious roots forming a basal cone. Leaves spirally arranged, 6-31, erect to arching, marces- cent; sheath brown or cream colored, short, tubular, splitting opposite petiole, the mar- gins stiff fibrous, the tomentum brown to reddish brown; petiole slender, adaxially flat or channeled, glabrous, abaxially round- ed or angled, brown tomentose, blade bifid simple, somet imes becoming i r regular ly split with age or damage caused by wind, narrowly cuneate at base; veins 25-70, emerging at 20-35 ~ angle from midvein (rachis), prominent at one or both surfaces of blade. Inflorescence interfoliar, solitary, branched to one order, rarely spicate or forked, erect at anthesis, becoming pendent in fruit, protandrous; peduncle long, slen- der, brown to whitish tomentose; prophyll tubular, fibrous, membranous or charta- ceous, bicarinate, brown tomentose, insert- ed near base of peduncle; peduncular bracts t or 2, tubular, chartaceous or membranous, inserted 1 to several cm above base of pe- duncle ; in f lo rescence rachis with short, ovate, pointed bracts, subtending rachillae, salmon to pink at anthesis, brown to reddish brown in fruit; rachillae 1-16, ca. equal in length, ending in a short, sterile apex, same color as inflorescence rachis. Flower pits containing triads comprising two staminate flowers and one pistillate flower, the pits spirally arranged on rachis, covered by a rounded upper lip that overlaps the lower, immersed lip when in bud; floral bracts 3, the largest bract subtending the second sta- minate flower, a medium-sized, 2-keeled bract subtending the pistillate flower, and a small bract representing the prophyll of the pistillate flower. Staminate flowers oblong to oblong-obovoid, borne abaxially to pis- tillate flowers; sepals 3, unequal, united ba- sally, narrow, elongate, keeled dorsally; petals 3, ca. as long as or longer than sepals, united into a tube for ca. 2,4 their length, distally valvate; stamens 6-27; filaments united into a tube up to midlength, white or yellow, with or without a papillate epider- mis; connective bifid, tanniferous, bearing separated thecae, the anthers inflexed to- ward center of flower and introrse in bud, spread and extrorse at anthesis; pollen
asymmetric in shape, monosulcate, the sul- cus as long as main axis, the exine tectate and finely perforate-rugulate or perforate- fossulate (Fig. 3A-H); pistillode irregularly 3-lobed (Fig. 12E). Pistillate flowers ob- long to oblong-ovoid; sepals 3, unequal, imbricate in bud, dorsally keeled; petals 3, united up to midlength, distally valvate; staminodes 5-22, fused to corolla and unit- ed basally, fleshy, with tanniferous tips; gy- noecium syncarpous, trilocular, triovulate, the nectary septal; style basal to lateral, elongate; stigmatic branches 3, papillate, re- flexed at anthesis; ovule anatropous, insert- ed at midlength of locule, pendent. Fruit a drupe, ellipsoid to ellipsoid-ovoid, 1-seed- ed, dorsiventrally compressed, slightly to conspicuously keeled at apex, the remains of the stigma basal; epicarp smooth, pur- plish black or garnet red at maturity; me- socarp fleshy and juicy, with an inner layer of closely appressed, longitudinal fibers; endocarp thin, crustaceous, and shiny (Uhl and Dransfield, I987). Eophyll with a bifid simple blade.
Distribution.--Five species distributed in Central America (Belize, Guatemala, Hon- duras, Nicaragua, Costa Rica, and Panama) and northern South America (Colombia, Ecuador, Venezuela, and French Guiana). (Fig. 4)
Key to the species of Asterogyne
Stamens 21-27; inflorescences spicate. 2. Inflorescence rachis and rachil lae pale
green at anthesis; filaments white and smooth; fruits 2.5 cm long, 1.5 cm diam . . . . . . . . . . . . . . A . g u i a n e n s i s
2. Inf lorescence rachis and rach i l l ae brown at anthesis; fi laments conspic- uously yel low orange and papillate; fruits to 1.8 cm long, 0.8 cm diam. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . s p i c a t a
Stamens 6 -12 ; inf lorescences branched, rarely spicate.
3. Peduncular bract 1, rarely 2: stamens 6; staminodial lobes 6 A . m a r t i a n a
3. Peduncular bracts 2: stamens 8-12: s taminodial lobes 5-9.
4. Inflorescence raehis 0.5-2.5 cm long; rachillae 4 - 6 ; s tamens 11- 12 ...... A . r a m o s a
4. Inflorescence rachis 5 -10 cm long; rachillae 9 16; stamens 8 -10 --- . . . . . . . A . y a r a c u y e n s e

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 347
1. ASTEROGYNE GUIANENSIS Granv. & A. J. Hend. (Fig. 1A, B)
A s t e r o g y n e g u i a n e n s i s G r a n v . & A. J. Hend. , Br i t ton ia 40: 76. 1988. TYPE: F R E N C H G U I A N A . C a m o p i River, ca. 1.5 km N E o f M o n t Belv6dbre , in s w a m p forest , 150 m, 4 Dec 1984, J. J. de Gran- ville 7124 [HOLOTYPE: P; ISOTYPES: B, BR(n.v.) , CAY, K, NY, US]
Stem sol i ta ry , erect , 1 .5-2 m tall, 3 - 6 c m diam. , b rown, not b ranched ; in t e rnodes short: aer ia l roots b ranched , f o rming a basa l cone, each roo t 3 8 - 9 0 c m long, 0 . 2 -0 .3 c m d i a m . , y e l l o w i s h b r o w n ; p n e u m a t o d e s white , conica l , to 0.2 c m long. Leaves 1 5 - 18, the o lde r ones not pers is tent ; sheath 7 • 4 - 5 cm, b rown; pet io le 5 3 - 6 0 X 0 .8-1 cm, channe l ed adax ia l ly , rounded abax ia l ly , green, the t o m e n t u m redd i sh b rown; mid - ve in 8 0 - 9 0 • 0.5 cm, green, the i n d u m e n - turn same as on pe t io le , channe led adax i - al ly, r ounded abax ia l ly ; blade 1-1.1 x 0 . 3 - 0.4 m, d e e p l y bi f id at apex for ca. 35 cm; p r i m a r y ve ins 29 pe r side, e m e r g i n g at 2 0 - 30 ~ angle at midve in , p r o m i n e n t on abax ia l side, a lmos t y e l l o w when dry, the l ep ido te t o m e n t u m s l igh t ly r edd i sh brown, hlflores- cence spicate ; p e d u n c l e 4 0 - 5 5 cm long, 0 . 4 - 0 . 5 c m d i a m . , d o r s i v e n t r a l l y c o m - pressed , the t o m e n t u m b rown to whi t i sh ; p rophy l l 20 c m long, 1.5 c m diam. , m e m - branous , green , inse r t ed at base o f p e d u n - c l e ; p e d u n c u l a r b r a c t l , m e m b r a n o u s , green, 4 0 - 4 2 • 1 .3-1 .5 cm, inser ted 1 c m a b o v e the base o f the pedunc le : inf lores- cence rach is 2 0 - 3 0 c m long, 0 . 9 - 1 . 5 c m d iam. , terete, pa l e g reen at anthesis , r edd i sh b r o w n in fruit , the t o m e n t u m whi t i sh to b rownish , the s ter i le apex acute, 1-2 m m long: f lower pi ts sp i r a l ly a r ranged , 3 - 5 m m apar t f rom each other, cove red with an in- f lexed lower l ip, 3 - 4 • 2 - 3 mm. Staminate f lowers obovo id , 9 - 1 0 m m long, 3 - 4 m m diam. , whi te at an thes is , the ped ice l to 1 -2 m m long; sepa ls ob long , 9 - 9 . 2 • 2 - 3 mm; pe ta ls o b l o n g - o b o v a t e , 8 X 2 . 5 - 3 ram; sta- mens 2 5 - 2 7 ; f i l aments un i ted into a tube up to 4 - 5 m m long, the free parts 2 . 5 - 3 m m long, 1 m m diam. , whi te ; connec t ive 0.8 m m long; thecae 1 .2-1 .5 m m long; p is t i l - lode 1-2 m m long. Pistil late f lowers o b l o n g
in bud, 5 m m long, 2 . 5 - 3 m m diam. , the ped ice l to 1 m m long; sepals o b l o n g - o b - ovoid , 4 - 5 • 1 ram; pe ta l s o b l o n g - o b o v o i d , 4 X 1 .4 -1 .6 m m ; o v a r y 3 m m long, 2 m m diam. ; s tyle 3 m m long; s t igmat ic b r a n c h e s 1 m m long; s t a m i n o d i a l tube 4 m m long, the lobes 2 0 - 2 2 , 1 -1 .5 m m long. Fruit nar- r o w l y e l l ipso id , s t rong ly kee led ap ica l ly , 2.5 cm long, 1.5 c m d iam. ; ep i ca rp ga rne t red and sh iny at ma tur i ty : seed n a r r o w l y el - l ipsoid , 2 cm long , 0.8 c m diam.
Local names and u se s . - - None repor ted . Distribution and ecology. K n o w n o n l y
f rom two co l l ec t ions o f eas tern F r e n c h Gu i - ana, r e m a r k a b l y m o r e than 200 k m f rom each other. H e n d e r s o n and Ba l i ck (1987) sugges ted that this spec ies , at that t ime not ye t descr ibed , m a y occu r also in nor thern Brazi l .
Additional specimens examined: FRENCH GUI- ANA. Crique Rapatea, Bassin de 1Approuague, jonc- tion de la Kouroua'/e et de la RN2, 4~ 52~ 24 Oct 1996, Blanc 189 (CAY); plantlets cultivated at Cayenne, 22 Oct 1985, de Granville 7124-A (CAY).
2. ASTEROGYNE MARTIANA (H. Wendl . ) H. Wendl . ex Hems l .
Asterogyne mart iana (H. Wendl . ) H. Wendl . ex Hemsl . in G o d m a n & Salvin , Biol . Centr. Am. Bot . 3: 409. 1885; Geonoma mart iana H. Wendl . , L i n n a e a 28: 342. 1856. TYPE: cult . hort . Her renhausen , H. Wendland 56 [HOLOTYPE: GOET; ISOTYPE: K ( F neg. 20765, B H neg. 20368)] (Fig . 1C)
Geonoma trifi~rcata Oerst., Kjoeb. Vidensk. Med- del. 1858: 34. 1859. TYPE: Nicaragua, ad flumen San Juan, A. S. Oersted 6566 [HOLOTYPE: C(F neg. 21 O69)].
Asterogyne minor Burret, Bot. Jahrb. Syst. 63: 143. 1930. TYPE: SW Colombia, above Barbacoas, 500 m, F. C. Lehnlann 708 [HOLOTYPE: B(destroyed)].
Stem soli tary, erect, somet imes basa l ly de- cumbent , 0 . 4 - 1 . 6 ( - 3 ) m tall, 2 .5 -5 diam. , dull b rown to orange , not branched; inter- nodes ( 1 - ) 2 . 5 - 3 c m long; aerial roots fo rm- ing an i r regular basa l cone, each root 4 - 8 c m long, 0 . 3 -0 .4 c m diam.; p n e u m a t o d e s b rownish c r eam-co lo red , conical , to 1 m m long. Leaves ( 6 - ) 1 2 - 2 0 , ye l lowish red when young, green when mature , the o ld ones somet imes pers is tent ; sheath ( 8 - ) 1 1 - 1 6 ( - 3 8 )

348 BRITTONIA [VOL. 55
X 3 cm, with reddish brown tomentum, lon- gitudinally splitting, the margins slightly fi- brous; petiole (4- )20-68( -75) • 0.5-1.5 cm, keeled to channeled adaxially, rounded to angled abaxially, a lmost glabrous to densely brown tomentose, toward the base slightly fibrous at margins; midvein (33-)55- 791-108) X 0.2-0.7 cm, greenish yellow, glabrous or with pinkish brown tomentum, slightly keeled adaxially, rounded abaxial- ly; blade 0.4-1.3 • 0.15-0.3 m, light to dark green, deeply bifid at apex for 18-32 cm: primary veins 28-40 per side, emerg- ing at 20-35 ~ angle at midvein, prominent at both surfaces of blade, yellow when dry. h ~ o r e s c e n c e branched to 1 order or very rarely spicate: peduncle (20 - )34-70( -125) cm long, 0 .4-0.7 cm diam., dorsiventrally compressed: prophyll 11-26(-36) cm long, (/ .6-1.5(-2) cm diam., inserted at base of peduncle: peduncular bract 1, rarely 2, chartaceous, brownish cream-colored, the indumentum same as on prophyll; first pe- duncular bract 20 -31( -40) X 0.5-1.5 cm, inserted 0.5-2 cm above base of peduncle: second peduncular bract, when present, 15 X 0.5-0.9 cm, inserted 25-27 cm above base of peduncle; inflorescence rachis ab- sent or 0 .5-3(-7) cm long, 0.3-0.5(-1.2) cm diam., dorsiventral to irregularly com- pressed, salmon to pink at anthesis and in fruit, glabrous or with indumentum same as on peduncle; rachillae (2- )3-5( -14) , terete, erect in bud, slightly pendent in fruit, the color same as inflorescence rachis, the to- mentum white, farinose or short and ap- pressed: basal rachillae (9 - )14 .5-22( -27) cm long, 0 .4-0.6(-1.1) cm diam.: middle rachillae (8.5-) 14-21 (-23.5) cm long, 0 .4 - 0.6(-1) cm diam.; apical rachillae ( 9 - ) 1 5 - 22 cm long, 0.4-0.6(-1.2) cm diam., the sterile apex acute, 0.2-1.1(-5.5) cm long; flower pits spirally arranged, ca. 2-3 mm apart from each other, covered with a lower semicircular lip, 1-2 • 1-2 ram, the lip be- coming reflexed in fruit. Staminate flowetw oblong-obovoid in bud, 6-9 mm long, 2-3 mm diam., white at anthesis, the pedicel to 0.7 mm long; sepals oblong-lanceolate, 3.6-4.8 • 1 ram: petals oblong-lanceolate, 5-7 • 1-1.3 ram: stamens 6, the filaments united into a tube up to 5 .4-7 mm long, the free parts 2 .4-3 mm long, 0.3-0.5 mm
diam., r emarkab ly thinner at apex and therefore the anther versatile, white, with- out a papillate epidermis; connective slight- ly swollen, 0.7-1.1 mm long; thecae 0 .7- 1.6 mm long: pistillode 2-5 mm long, 1 mm diam., basally fused with staminodial tube. Pistillate f lowers oblong in bud, 6-8 mm long, 2,7-3.1 mm diam.; sepals oblong, 4 -5 • 1-l ,5 mm; petals oblong, 4,4-5.5 • 1,1-1.8 ram; ovary 1 mm long, 0.8 mm diam.; style 6 - 8 mm long: s t igmat ic branches 1-2 mm long; staminodial tube 3.9-5 mm long, the lobes 6, 1.5-2.1 mm long. Fruit ellipsoid to ellipsoid-ovoid, 0 .8- 1.2 cm long, 0.4-0.7 cm diam.; epicarp or- ange to red when young, purplish black at maturity; seed ellipsoid, 0.7-0.8 cm long, 0.4-0.5 cm diam.
Local names and uses.--capoca (Guate- mala); pacuquilla (Honduras); pata de gallo, suita (Nicaragua) ; pacaya, suita (Costa Rica), dobo-gaw (Guaymi of Panama); cor- tadera, panda~ panga, rabihorcao, rabo de zorro (Colombia): guacamayo, pico (Ecua- dor). The use of the leaves for thatching houses has been reported in Nicaragua (Neill 1723, Salick 8106), in Costa Rica (Chazdon, 1986), in Panama (Gordon 70), in Colombia (Bernal & Galeano 887), and in Ecuador (Aulestia & Aulestia 1306).
Distribution and ecology. Widespread in Central America: Belize (El Cayo, Stann Creek, Toledo), Guatemala (Izabal), Hon- duras (Atl~intida, Col6n, Cort6s, Gracias a Dios, Santa B~irbara), Nicaragua (Chonta- les, Jinotega, Matagalpa, Rfo San Juan, Ze- laya). Costa Rica (Alajuela, Cartago, He- redia, Lim6n, Puntarenas, San Jos6), Pana- ma (Bocas del Toro, Chiriquf, Coc16, Col6n, Dari6n, Panama, San Blas, Veraguas), and in northwestern South America: Colombia (Antioquia, Choc6, C6rdoba, Narifio, Valle del Cauca), Ecuador (Esmeraldas) (Fig. 4). Common in the understory of the lowland and mountain rain forests, in flooded and nonflooded areas; from sea level to 1400 m, but normally below 600 m.
Selected specimens examined: BELIZE. El Cayo District: 47 mile section, Humming Bird Hwy.. 28 Apr 1956, Gentle 9072 (NY). 60 m, 6 Sept 1929. Schipp 392 (G, K, MO, NY, Z); W of Isla Rivas, 29 Jan 1903, Wilson 224 (NY, US). Toledo: Monkey Rivel; in high

2003] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 349
"Monkey Tail," 1 Nov, 1941, Gentle 3743 (MO, NY, US1.
GUATEMALA. Izabal: Wet wooded slopes nr. Km 824 past Villa Jesfis on rd. from Puerto Barrios to Gua- temala, 100 m, 8 Nov 1959, Moore & Cetto 8213 (BH): valley of tributary of Rio San Francisco del Mar, 2 miles NE of Hopi, 12 miles from Entre Rfos, 10-20 m, 21 Apr 1940, Stevermark 39785 (US), E portion of Vera Paz and Chiquimula, 1885, Watson 238 (BH/.
HONDURAS. Atl~intida: Vic. Lancetilla, 45 m, 8 Oct 1934, Ynncker 5002 (MO, NY). Graeias a Dios: Laimos Creek, 15 km SW of Waspan, 7 Mar 1961, 15-30 m, Bunting & Licht 401 (NY): Rio Plfitano, 4 hours upriver from village of Ras, nr. sea level, 23 May 1973, Gent O, et al. 7542 (MO).
NICARAGUA. Chontales: Chontales, 600 m, Jun 1870, Levv 505 (G, P): 1870, Seemann s.n. (BMI. J in- otega: Salto Kayaska, Rio Bocaz, ca. 13~ 85~ 190-340 m, 7 Mar 1980, Beach et al. 16545 (US); Comarca de Bocaycito, Cordillera Isabelia, nr. Rio Bote, 6 May 1976, Neill 7151 (MO, US/. Mata- galpa: E1 Comej6n, 1 km W de la carretera a Waslala, 13~ 85~ 600 m, Moreno & Robleto 20602 (MO). Rio San Juan: Sabalo, 1 k m a l N de Rio San Juan, l l~ 84~ 100 m, 9-10 Jul 1985, Mo- reno 26083 (MO): "Los Filos,'" nr. Loma Los Filos, Rio Santa Cruz, 1 l~ 84~ 6 Sept 1993, Salick 8106 (MO). Zelaya: Ca. 1 km N of El Zapote, 6 km S of Colonia Verdfin, 11~ 84~ 250-300 m, 8 Sept 1983, Nee & Vega 27905 (NY, US); El Hor- miguero, carlo above camp and on W slope at Loma, 13~ 84~ 800-1000 m, 17 Mar 1980, Pipoly 6143 & 6168 (MO, US/: ca. 6.3 km S of bridge at Colonia Yolania and ca. 0.8 km S of ridge of Serranias de Yolania on rd. to Colonia Manantiales (Colonia So- moza/, 11~ 37'N, 84~ 200-300 m, 29-31 Oct 1977, Stevens 4810 (BH, MO, NY).
COSTA RICA. Alajuela: Vic. Guatuso de San Ra- fael (on Rio Frfo), 10~ 84~ 80-100 m, 4 Aug 1949, Hohn & lltis 914 (BH, BM, MO, US): 25 km NNW of San Ram6n by rd. on way to San Lor- enzo, 10~ 84~ 500 m, 24 Apr 1983 Liesner & Judziewicz 14825 (MO, WIS). Cartago: Bois de Peralta, 300 m, Jun 1903, Pittier 16658 (US). Heredia: Finca La Selva, 12 Dec 1984, Henderson 53 (NY): along "Starkey Rd.,'" 4.5 km SE of bridge at Puerto Viejo, 10~ 83~ 50 m, 7 Aug 1979, Stevens 13505 (K, MO, US/. Lim6n: Finca Castilla, 30 m, 24 Jul 1936, Donee & Goerger 9284 (MO/: 7 km SW of Bribrf, 100 250 m, G6mez et al. 20341 (MO, NY, W1S): woodlands south of La Lola on the railroad, 120 m, 15 Apr 1953, Moore 6717 (BH). Puntarenas : Gol- fito, 15 m, 22 Oct 1950, Hari s.n. /G); hills above Palmar Norte, 550 m, 6 Mar 1953, Moore 6530 (AAU, BH, K): Osa Peninsula, Agabuena, 4 km W of Rinc6n, at cleared path dividing lands of Don Fortunato and Don Contreras, SSW of 4-ha sample plot, 8~ 83~ 'W, 250 m, 18 Nov 1992, Thomsen 187 (C). San Jos& Rio Negro, Cerro La Cangreja, Santa Rosa de Puriscal, 400-700 m, 20 Jun 1986, Chac6n & ChacDn 1958 ICR/: Tarrazfi, San Lorenzo, Llanos de Santa Maria, 9~ 84~ 500 600 m, 28 Jan 1998, Valverde 665 (K).
PANAMA. Boeas del Toro: Top of -Fi la" above
Almirante, 30 Nov 1971, Gentry 2803 (MO); Rio San Pedrito, 16 Jul 1978, Gordon 70 (MO); without local- ity, 24 Sept 1940, yon Wedel 952 (BH). Chiriquf: Bur- ica Peninsula, 19 Feb 1973, Croat 21997 (K, MO); Fortuna Darn region, hr. Chiriquf Grande, 8~ 82~ 200 m, 18 Jan 1986, McPherson 8080 (MO): vic, Chiriquf Lagoon, 9 Oct 1940, yon Wedel 1108 (BH, MO/. Coc16: Rd. to Coclesitn, logging camp, 12 miles from Llano Grande, 8~ 80~ 200 m, 16 Dec 1983, Churchill et al. 4122 (K, MO), Boca del Toabrd at confluence of Rio Toabr6 and Rio Coc16 del Norte, 11 Apr 1969, Lewis et al. 5558 (BH, MO/. Co- 16n: Santa Rita Ridge, 11 May 1999, Henderson & Ferreira 3048 (NY); Santa Rita Ridge, end rd. from Transisthmian Hwy., ca. 10 miles from Hwy., 29 Mar 1969, Porter et al. 4736 (BH, MO, NY); Santa Rita Lumber Rd., ca. 3.1 km E of Agua Clara Rain Gauge, 9 Sep 1975, Witherspoon et al. 8322 (MO), Dari6n: Parque Nacional del Dari6n, ridge between Rio Topal- isa and Rio Pacuro, ca. 13 km E of Pacuro, Quebrada Pobre to Mi Casita, 8~ 77~ 450 600 m, 14 Oct 1987, de Nevers et al. 8320 (MO, US). Panama: Canal Zone, Apr 1925, Cook s.n. (US); Cerro Jefe roadside, 2 km N of turnoff to radio tower on rd. [from?] Alto de Pacora, 790 m, 30 Sep 1978, Hammel 4863 (MO). San Bias: 10 km from Pan-American Hwy. on El Llano Cartf Rd., 9~ 79~ 250 rn, 6 Mar 1987, BarJ~)d & MePherson 13 (AAU/: 44 km N of Penonome on rd. to Coclecito, 90 150 m, 21 Feb 1978, Hammel 1682 (BH, MO/: Nusagandi, E1 Llano- Cart/ Rd., 19 km from Inter-American Hwy. 9~ 78~ 300 400 m, 9 Oct 1996, Knudsen & Asmus- sen 604, 620 (AAU): E1 Llano-Cart /Rd. , Km 10 15, 300-400 m, 3 Sept 1977, Maas et al. 2803 (U). Ver- aguas: Atlantic slope, Rio Concepci6n to Rio Barrera, ca. 60 m, 16 Oct 1978, Hammel 5153 (MO).
COLOMBIA. Antioquia: Mun. Frontino, Corregi- miento de Murrf, rd. from Nutibara to La Blanquita, 950 m, 18 Mar 1982, Bernal & Galeano 256 (NY), Mun. de San Rafael, 15 k m a l E del pueblo en la carretera a San Carlos, 960 m, 19 Sept 1987, Bernal & Tobdn 1380 (AAU, BH, NY); Villa Arteaga, Mu- tata, ca. 1000 m, 5 Apr 1976, Dran~sfield et al. 4859 (BH, K). Choe6: Area of Baud6, on left bank of river Baud6, ca. 11.5 km upstream of estuary, slightly downstream of estuary of Quebrada Anguerad6, 50 m, 7 Feb 1967, Fuchs & Zanella 21808 (U): north ridge of Alto de Buey, above Dos Bocas del Rio Mutat~i, tributary of Rio El Valle, ESE of E1 Valle, 200 500 m, 8 Aug 1976, Gentry & Fallen 17445 (MO). Cor- doba: Junction of Rio Tigre and Rio Manso, Paramillo National Park, transect 1, 7~ 76~ 200 m, 25 Jul 1988, Gentry & Cnadros 63735 (MO). Narifio: Mun. de Tumaco, 3 5 km al E de Guayacana, 100 m, 3 Oct 1985, Bernal & Galeano 887 (AAU, NY). Vaile del Cauca: Buenaventura, 6 May 1926, Cook 84 (US); Bajo Calima, Juanchaco Palmeras, ca. 10 km NW of Buenaventura, 3~ 77~ 13 Apr 1987, Gentry et al. 56871 (MO, NY/.
ECUADOR. Esmera ldas : Reserva Etnica Awfi, Parroquia Alto Tambo, Centro de la Uni6n, carl6n del Rio Mira, 00~ 78~ 250 m, 22 Mar 1993, Aulestia & Aulestia 1306 (MO): Playa de Oro on the Rio Santiago, ~ hour walk S of village, 250 m, 25 Feb

350 BRITTONIA [VOL. 55
1994. Borchsenius 242 (AAU, NY); San Lorenzo Can- t6n, Reserva Etnica Awli, Centro Ricaurle, a orillas del Rio PalabL ~~ 78~ 300 m, 28 Oc~ 1992, Tipaz et aL 2250 (AAU. MO).
Cult. Holt. Herrenhausen, Wendland 56 (GOET, HO- LOTYPE; K, ISOTYPE). Probably originated from seeds sent by Warscewicz from Central America.
3. ASTEROGYNE RAMOSA (H. E. Moore) Wess. Boer. (Fig. 1D-F)
Asterogyne ramosa (H. E. Moore ) Wess. Boer, Verb. Kon. Ned. Akad, Wetensch. , Aid. Nat,, 2 Reeks, 58: 81, 1968. Aris- tevera ramosa H. E. Moore~ J, Arnold Arbor. 48: 144-146. 1967. TYPE: VE- N E Z U E L A . Sucre: Peninsula de Paria, Cerro Espejo, en la cumbre entre Man- tecal y Paujf, arriba de M u n d o Nuevo y Rio Seco de Irapa, 7 5 0 - 8 5 0 m, 6 Aug 1966, J. Steyermark & M. Rabe 96088 (HOLOTYPE: BH; ISOTYPES: NY, US, VEN)
Stem solitary, erect, 3 - 6 m tall, 6 - 1 0 cm diam., c reamy brown, occas iona l ly with lat- eral vegetat ive branches; in ternodes 5 - 1 0 cm long; aerial roots fo rming a short basal cone, each up to 15 cm long, 0 .5-1 cm diam., brown, conical; pneumatodes very short to a lmost absent. Leaves 15-20, the older ones briefly persistent; sheath 9 - 1 0 ( - 17) X 7 cm, densely brown tomentose , the margins slightly fibrous; pet io le 3 0 - 3 5 • 0.6-1 cm, channeled adaxially, rounded abaxially, b rown tomentose; midve in 39 • 0.5 cm, the indumentum brown, slightly channeled adaxially, a lmost flat abaxially; blade 0.5-0 .7 • 0 .18-0 .27 m, green, deeply bifid at apex for ca. 33 cm; p r imary veins 3 4 - 4 5 per side, emerging at 2 5 - 3 0 ~ angle at midvein, prominent on both surfaces o f blade, pale green abaxially. Inflorescence branched to 1 order; peduncle 9 0 - 1 2 0 cm long, 0 .4 -0 .5 cm diam., dorsiventral ly com- pressed, green; prophyll 15 -20 c m long, 1.8 cm diam., chartaceous, inserted at base o f peduncle; peduncular bracts 2, char taceous, brown, the indumentum same as on pro- phyll; first peduncular bract 3 1 - 4 9 • 1.4 cm, inserted 0.5 cm above base o f pedun- cle; second peduncular bract 4 1 - 5 3 • 1 - 1.2 cm, inserted 1 cm above base o f pedun- cle; inflorescence racbis 0 . 5 - 1 . 2 ( - 2 . 5 ) cm long, 0 .4 -0 .5 cm diam., dorsiventraI to ir-
regular ly compressed , pale green at anthe- sis, reddish in fruit, g labrous; rachil lae 4 - 6, terete, erect, same color as inf lorescence rachis, each rachilla subtended by a curved bract up to 0,5 cm long, covered with a whitish far inose indumentum; basal, medi- al, and apical rachillae 17-19 cm long, 0.5 cm diam., the sterile apex acute, 0 .2 -0 .4 c m long; f lower pits spirally arranged, ca. 2 -3 m m apart f rom each other, covered with an inflexed lower lip, 1.5-2 • 1 .5-2 mm. Sta- minate f lowers ob long-obovo id in bud, 6 -8 m m long, 3 - 4 m m diam., sl ightly fragrant, white at anthesis, the pedicel to 1.5 m m long: sepals ob long-obovate , 5 - 6 • 2 -3 mm; petals oblong, 4 - 6 • 2 mm; s tamens 1 1 - 1 2 , o c c a s i o n a l l y 1 s ter i le ; f i l aments united into a tube to 3 m m long, the free parts 3 m m long, 0.5 m m diam., white, not r emarkab ly thinner at apex, the epidermis not papillate; connect ive 1 m m long; thecae 1.2 m m long; pistil lode 2 m m long. Pistil- late f l owers ob long-ovo id in bud, 5 - 7 m m long, 3 - 4 m m diam., the pedicel to 1 m m long; sepals oblong to ovoid, 4 - 5 • 2 -2 .5 mm; petals ovate, 5 - 6 x 2 ram; o v a r y 1.5 m m long, 1.5 m m diam.; style 1 .5-2 m m long; s t igmatic branches l m m long; stam- inodial tube 4 m m long, the lobes ( 5 - ) 7 - 9 , 2 .8 -3 .4 m m long. Fruit ellipsoid, 1.5-1.7 cm long, 0 .5 -0 .6 cm diam.; epicarp dark purplish red when young, purpl ish black at maturi ty; seed ellipsoid, 1.3 cm long, 0.4 cm diam.
Local name and uses.--anare. Uses not known.
Distr ibut ion and ecology. K n o w n only f rom the unders tory o f the mon tane cloud forest in the northeastern Coastal Cordil le- ra, state o f Sucre, Venezuela (Fig. 4); f rom 7 3 0 - 1 2 5 0 m. Al though the voucher for col- l e c t i on S t e y e r m a r k 9 4 8 0 3 m e n t i o n s the palm g rowing at 1600 m on the Cerro Humo, this must be an error because the summit o f this mounta in has been reported to be 1 2 5 0 m .
Additional specimens examined: VENEZUELA. Sucre: Peninsula de Paria National Park, above Las Melenas, N of Rio Grande Arriba, trail to Cerro de Humo, 10~ 62~ 900-1000 m, 3 Jan 2002, Stauffer et al. 824 (MY, MYE VEN. Z); Peninsula de Paria, Cerro Hnmo, a 2o largo de uno de los affuer~tes del Rio Santa Isabel arriba de Siparo, 1600 m (prob-

2003] S T A U F F E R ET AL.: A S T E R O G Y N E ( A R E C A C E A E ) 3 5 1
able erroneous elevation), l Mar 1966, Steyermark 94803 (BH, NY, U, VEN); Peninsula de Paria, above Las Melenas, N of Rio Grande Arriba, SE of Cerro de Humo, 10~ 62~ 730 1050 m, 1 Dec 1979, Steyetwlark & Liesner 120915 (MO, NY, VEN); Pen- fnsula de Paria, Distrito Marino and Distrito Ar ismen- dL trail between crossing of Rio Tacarigua to summi t of slopes E of Cerro Humo, descending to Las Mele- nas, N of Rio Grande Arriba, 10~ 62o36 - 37'W, 760-1000 m, 24 Feb 1980, Steyermark et al. 121713 (MO).
4. ASTBROaYNE SPICATA (H. E. Moore) Wess. Boer. (Fig. 2 A - C )
Asterogyne spicata (H. E. Moore) Wess. Boer, Verh. Kon. Ned. Akad. Wetensch., Aid. Nat., 2 Reeks, 58: 82. 1968; Aris- tevera spicata H. E. Moore, J. Arnold Arbor. 47: 5-9. 1966. TYPE: VENEZUE- LA. Miranda, Parque Nacional Guatopo, moist rich mixed evergreen forest on steep slopes bordering Rio Santa Cruz and Altagracia de Orituco, 14.5 km f rom Los Alpes, 12 km f rom Rancherfa Mi Querencia, 520 m, 23 Nov 1961, J. Stey- ermark 90010 (HOLOTYPE: BH; ISOTYPES: NY, VEN)
Stem solitary, erect, 2 - 4 ( - 8 ) m tall, 4 - 7 cm diam., brown, occasionally with basal v e g e t a t i v e b ranches ; in te rnodes 0.5 cm long; aerial roots forming a short basal cone; pneumatodes very short. Leaves 15- 19(-25), somet imes reddish brown when young, green when mature, the older ones not persistent; sheath 10-15 X 9 cm, red- dish brown tomentose, the margins slightly fibrous; petiole 3 5 - 4 0 X 0.7-1.5 cm, chan- neled adaxially, rounded to angled ab- axially, brown tomentose, slightly fibrous at margins toward base; midvein 68-84 • 0.5-0.7 cm, green, with same indumentum as on petiole, ridged adaxially, flat abaxi- ally; rachis with a filiform extension of 1.5- 5 X 0.1 cm; blade 0.7-1 x 0.2-0.3 m, dull g reen adaxia l ly , s i lve r g reen abax ia l ly , deeply bifid at apex for ca. 45 cm; pr imary veins 25-27 per side, emerging at 25 -30 ~ angle at midvein, prominent abaxially, yel- lowish, slightly brown tomentose. Inflores- cence spicate; peduncle 4 0 - 5 0 cm long, 0 . 5 - 0 . 6 cm diam. , do r s iven t r a l ly c o m - pressed; prophyll 14-15( -25) cm long, 1.5 cm diam., fibrous, inserted at base of pe-
duncle; peduncular bract 1, chartaceous, brown tomentose, 2 2 - 4 0 • 1.3-2 cm, in- serted 1.4 cm above base of peduncle; in- florescence rachis 2 3 - 4 0 cm long, 1-1.5 cm diam., terete, pale brown at anthesis, brown in fruit, with same indumentum as on pe- duncle, the sterile apex acute, ca. 0 .6-3 cm long; flower pits spirally arranged, ca. 2 - 4 m m apart f rom each other, covered with an inflexed lower lip, 3 X 2 - 4 mm. Staminate flowers obovoid, 0.8-1.3 cm long, 0 .4-0 .5 cm diam., fragrant, the pedicel almost ab- sent; sepals oblong-obovoid, 5 - 6 • 1 .4-2 mm; petals obovoid, 7 -8 x 2-3 ram; sta- mens 21-26, occasionally 1 or 2 partially or totally sterile; filaments united into a tube to 6-7 m m long, conspicuously yel low orange, not remarkably thinner at apex, the epidermis papillate, the free parts 3 - 4 m m long, 0.4 m m diam.; connective 1.2-1.5 m m long; thecae 0 .5-1.2 m m long; pistil- lode 4 - 5 m m long. Pistillate flowers oblong in bud, 8 m m long, 4 m m diam., the pedicel to 1 m m long; sepals oblong, 5 -7 X 2-3 ram; petals oblong, 7 -8 m m • 2-3 mm; ovary 0.9 m m long, I -1 .2 m m diam.; style 2.5-3 m m long; stigmatic branches 1-1.5 m m long; staminodial tube 2 m m long, the lobes 15-21, 2-2 .2 m m long. Fruit ellip- soid-ovoid, 1.5-1.8 cm long, 0.5-0.8 cm diam.; epicarp purplish red in young fruits, purplish black at maturity; seed ellipsoid, 0.8-1 cm long, 0.5 cm diam.
Local name and uses.~palma, palmito, palmiche. Moore (1966) noted that the fruits are said to be edible and the leaves used for a durable thatch supposed to last ten years. Nehlin (1994) reported that the endocarp has a sweet acidic taste.
Distribution and ecology.--Known only f rom the understory of the lowland and m o u n t a n e c loud fo res t in the nor the rn Coastal Cordillera, state of Miranda, Ve- nezuela (Fig. 4); normally between 6 0 0 - 1000 m.
Addi t ional s p e c i m e n s examined : V E N E Z U E L A . M i r a n d a : Parque Nacional Guatopo, 20 Dec 1961, Agostini 10 (VEN): Carretera Santa Teresa-Altagracia de Orituco, 600 -700 m, Jun 1953, Aristeguieta 1772 (NY, VEN); selvas de Guatopo, 500 m, Jun 1958, Ar- isteguieta 3185 (NY, VEN); selvas pluviales de Gua- topo, 4 0 0 - 6 0 0 m~ 13 Nov 1956, Bernardi 5633 (NY); Parque Nacional de Guatopo, N of Altagracia de Ori- tuco, 400 m, 16 Feb 1979, Gent~ & Troth 24823 (BH,

352 BRITTONIA [VOL. 55
MO/; Dist. Paez, Fila La Tigra, Quebrada San Juan, 18 km al SW de C0pira, 10~ 65~ 47'W, 600 m, 2-7 Sep 1977, Gonzdlez & Ortega 1296 (BH, MO); NW de Guatire, excursidn Fila Juan Torres Fila Las Perdices, por el Rfo Guayabal hacia el pueblo Guayabal, 10~ 66~ 900 1100 m, 19-22 Feb 1993, Meier 3386 (NY, VEN); Parque Nacional de Guatopo, steep slopes bordering Rio Santa Cruz, be- tween Santa Teresa and Altagracia de Orituco, 14.5 km from Los Alpes, 12 km from Rancherfa Mi Querencia, 520 m, 6 Aug 1970, Moore et al. 9601 (BH); Guatopo National Park, along S end of La Macanilla trail, 32 km (by air/ NW of Altagracia de Orituco, 27 Aug 1979, Nee 17789 (NY): Guatopo National Park, region of "La Macanilla,'" 10~ 66~ 570 m, 18 Dec 2001, SmuffOr et al. 822 (MY. MYE VEN, Z): Parque Nacional Guatopo. along Rfo Santa Cruz. be- tween Santa Teresa and Altagracia de Orituco, 450 m, 27 Oct 1966, Steyermark 97_573 (G, US); Cerros del Bachiller, nr. E end, above Quebrada Corozal, S of Santa Cruz, 10 km (by air) W of Cfpira, 10~ 65~ 20-700 m, 20-26 Mar 1978, Stevermark & Davidse 116530 (BH, MO); Cerros del Bachiller, W sector, between base and summit, above Quebrada Bachiller, S of Carlo, 10~ 65~ 20-690 m, Steyermark & Davidse 116565 IBH. MO, NY, VEN); Parque Nacional Guatopo, entre Santa Teresa y Alta- gracia de Orituco, 14.5 km de Los Alpes. 520 m, 29 Nov 1970. Steyermark & Braun. 104041 (BH, US, VEN): montafias de Guatopo, 22 Mar 1956, Tamayo 4177 (US); Altagracia de Orituco. Santa Teresa, Parque Nacional Guatopo, 11 May 1969, van Rooden & Akkermans 9_5 (U).
5. ASTEROGYNE YARACUYENSE A. J. H e n d . &
Steyerm. (Fig. 2D-F)
Asterogyne yaracuyense A. J. Hend. & Steyerm., Brittonia 38: 309-313. 1986. TYPE: VENEZUELA. Yaracuy: Cerro La Chapa, selva nublada al norte de Nirgua, 1200-1400 m, 9 -10 Nov 1967, J. A. Steyermark, G. Bunting & J. G. Wessels Boer 100311. (HOLOTYPE: VEN: ISOTYPE: BH)
S t e m s o l i t a r y , e rec t , s o m e t i m e s b a s a l l y decumbent, 3 -6 m tall, 5-8 cm diam., brown, occasionally with basal and lateral vegetative branches; internodes 2-3 cm long; aerial roots forming a short basal cone; pneumatodes very short. Leaves 18- 25(-31), reddish brown when young, green when mature, the older ones briefly persis- tent; sheath 12-30 X 7-9 cm, reddish brown tomentose, the margins slightly fi- brous; petiole (18-)25-35 X 1-1.5 cm, flat to channeled adaxially, angled to rounded abaxially, the indumentum same as on
sheath, the margin slightly fibrous near base; midvein 56-95 cm • 0.3-0.6 cm, loosely brownish gray tomentose, flat to channeled adaxially, rounded to flat ab- axially; blade 0.70-1.20 X 0.23-0.30 m, dull green adaxially, grayish green and cov- ered with a whitish farinose indumentum abaxially, deeply bifid at apex for ca. 42 cm; primary veins 40-70 per side, emerg- ing at 25-30 ~ angle at midvein, mostly prominent abaxially, loosely brown tomen- tose. b!florescence branched to 1 order; pe- duncle 85-95 cm long, 1.5-2 cm diam., dorsiventrally compressed; prophyll 27-35 cm long, 2-3 .5(-5) cm diam., chartaceous, inserted at base of peduncle or 1 cm above it: peduncular bracts 2, with indumentum same as on prophyll, chartaceous, brown; first peduncular bract 45-58 • 2 -2 .5( -4) cm, inserted 3 cm above base of peduncle, second peduncular bract 58-60 • 2.5-3.5 cm, inserted 4 cm above base of peduncle: inflorescence rachis 5-10 cm long, 0.5 cm diam., dorsiventrally to irregularly com- pressed, pale green at anthesis, almost gla- brous; rachillae 9-16, slightly pendent, the same color as inflorescence rachis, gla- brous; basal rachillae 28-32 cm long, 0.5 cm diam.: middle rachillae 27-29 cm long, 0.5 cm diam.: apical rachillae 24-28 cm long, 0.5 cm diam., the sterile apex acute, 0.2-0.6 cm: flower pits spirally arranged, 2-3 mm apart from each other, covered with an inflexed lower lip, 2.5-3 • 2-5 ram. Staminate flowers obovoid, 6-8 mm long, 2 mm diam., slightly fragrant, white at anthesis, the pedicel to 1 mm long: sepals oblong-obovoid, 4 X 1-2 mm: petals ob- long-obovoid, 6-7 X 1.5-2 ram; stamens 8-10: filaments united into a tube to 4 mm long, the free parts 3-3.5 mm long, 0.5 mm diam., white, conspicuously thinner at apex, lacking a papillate epidermis; connective 1 mm long; thecae 1.1-1.2 mm long: pistillode 2 mm long. Pistillate flowers oblong-ovoid in bud, 4 mm long, 3 mm diam., the pedicel to 1 mm long; sepals oblong-ovate, 3 X 2 - 2.5 mm; petals oblong, 3 x 2 mm; ovary 1 mm long, 1.2 mm diam.: style 1.5 mm long: stigmatic branches 1 mm long; staminodial tube 2 mm long, the lobes 7-8 or some- times fewer by congenital union of 2 of them, 2 mm long. Fruits not known.

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 353
L o c a l n a m e s a n d u s e s . - - n o t reported. D i s t r i b u t i o n a n d eco logy . K n o w n only
f rom the unders to ry o f the Cerro La Chapa c loud forest, in the nor thern Coasta l Cor- dil lera, state Yaracuy, Venezuela ; r ecen t ly repor ted on the Cer ro E1 Zapatero o f the same state (W. Meier, pets . comm. ) (Fig. 4); be tween 1 2 0 0 - 1 4 0 0 m.
Additional specimens examined: VENEZUELA. Yaracuy: Dist. Nirgua, selva nublada del Cerro La Chapa, N of Nirgua, 12 Nov 1985, Henderson 502 (MYE NY): 5 km N of Nirgua. 10~ 68~ 28 Nov-1 Dec 1996, Stauffer et al. 265, 267 (VEN- Z), 5 km N of Nirgua. 10~ 68~ 4 Apr 1997. Stauffer et al. 319 (VEN/, 5 km N of Nirgua, 10~ 13'N, 68~ 27 Dec 2001, Stauffer & Stauf-
fer 823 (MY, MYE VEN, Z), 9-10 Nov 1967, Stey- ermark et al. 100307 (BH, VEN), 100311 (VEN, HO- LOT'~'PE; BH, ISOTYPE), 10~ 68~ 9-10 Nov 1967, Wessels Boer et al. 2032 (BH, MER, NY, U), 2036 (BH, MER, NY, U); Cerro Picacho, selva nub- lada en la fila, N de Nirgua, 1200-1360 m, 11 Mar 1967, Steyermark & Bunting 97661 (BH, US, VEN).
A c k n o w l e d g m e n t s
Specia l thanks to the curators o f the her- baria A A U , B, BH, B M , C, CAY, CR, (3, K, M E R , M O , M Y E NY, P, U, US, V E N , W I S , and Z. Hec to r M a y kindly p rov ided i n f o r m a t i o n on s p e c i m e n s d e p o s i t e d at B R H and M i r e y a Cor rea p rov ided databas- es conta in ing i n fo rma t ion on the spec imens depos i ted at P M A and SCZ. Thanks to Ur- sula H o f m a n n for p r o v i d i n g in fo rmat ion on the spec imens at GOET, and to M a r y En- dress and J ean - Jacques de Granvi l l e for cons t ruc t ive c o m m e n t s on the manuscr ip t . F red J. Stauffer, L. Pappaterra , R ica rda Ri- ina, Rodr igo Duno , and W. Me ie r he lped dur ing field w o r k in Venezue la , and R u d o l f S c h m i d kindly p r o v i d e d photos and infor- mat ion on A s t e r o g y n e m a r t i a n a in Cent ra l Amer ica . M i k e Th iv is great ly thanked for he lp ing with the phy logene t i c analysis. Ma- d e l i n e H a r l e y p r o v i d e d i m p o r t a n t c o m - ments on the po l len morpho logy . Urs Jauch p rov ided gu idance wi th the S E M work. Visits to A A U , B M , C, K, NY, and US by F. W. Stauffer were granted by the Inter- nat ional Pa lm Soc ie ty (IPS), P rog ram for Shor t Term Visits o f the Smi thson ian Insti- tut ion ( N M N H ) , and the K e w Latin A m e r - ica Research Fe l l owsh ips Program. The Min is te r io del A m b i e n t e y de los Recur sos
Naturales R e n o v a b l e s ( M A R N R ) and the Inst i tuto Nac iona l de Parques ( Inparques) k indly p rov ided the permi ts to co l lec t in Venezuela .
L i t e r a t u r e C i t e d
Appun, K, F. 1871. Unter den Tropen. Hermann Cos- tenoble, Jena.
Asmussen, C�9 B. 1999a. Toward a chloroplast DNA phylogeny of the tribe Geonomeae (PalmaeI. Pages 121-129. bz: A. Henderson & E Borchsenius, ed- itors. Evolution, variation, and classification of pahns. Mere. New York Bot. Gard., vol. 83. Bronx, New York.
�9 1999b. Relationships of the tribe Geonomeae (Arecaceae) based on plastid rpsl6 DNA sequenc- es. Acta Bot. Venez. 22: 65-76.
, W. J. Baker & J. Dransfield. 2000. Phylog- eny of the palm family (Arecaceae) based on rpsl6 intron and trnL-trnF plastid DNA sequences. Pages 525-537. In: K. L. Wilson & D. A. Morrison, ed- itors. Systematics and evolution of monocots. CSI- RO, East Melbourne, Victoria, Australia.
- - & M. Chase�9 2001. Coding and noncoding plastid DNA in palm systematics. Amer. J. Bot. 88: 1103-1117.
Baker, W. J., C. B. Asmussen, S. Barrow, J. Drans- field & T. A. Hedderson. 1999. A phylogenetic study of the palm family (Palmae) based on chlo- roplast DNA sequences from the trnL-trnF region. P1. Syst. Evol. 219: 111-126.
Bentham, G. & J. D. Hooker. 1883. Genera Plantar- um. Vol. 3(2). Reeve & Co., London.
Berna], R. 1989. Endangerment of Colombian palms. Principes 33:113-128.
Braker, E. & R. Chazdon. 1993. Ecological, behav- ioral and nutritional factors influencing use of palms as host plants by a neotropical forest grass- hopper. J. Trop. Ecol. 9: 183-197.
Braun, A. 1970. Palmas cultivadas de Venezuela. Acta Bot. Venez. 5: 1-94.
Brown, K. S. 1982. Paleoecology and regional pat- terns of evolution in neotropical forest butterflies. Pages 255-308. In: G. T. Prance, editor. Biological diversification in the tropics. Columbia University Press, New York.
Burret, M. 1930. Geonomeae Americanae. Bot. Jahrb. Syst. 63: 123-270.
Chazdon, R. L. 1986. Physiological and morpholog- ical basis of shade tolerance in rain forest under- story palms. Principes 30: 92-99.
de Granville, J. J. & A. Henderson, 1988. A new species of Asterogyne (Palmae) from French Gui- ana. Brittonia 40:76 80.
Dransfield, J. & N. Uhl. 1998. Palmae. Pages 306- 389. hi: K. Kubitzki, editor. Families and genera of vascular plants, flowering plants: monocotyledons. Vol. 4. Springer, Berlin.
Hahn, W. J. 2002. A molecular phylogenetic study of the Palmae (Arecaceael based on atpB, rbcL, and 18S nrDNA sequences, Syst. Biol. 51: 92-112.

3 5 4 B R I T T O N I A [ V O L . 5 5
H e n d e r s o n , A. 1986. A review of pollination studies in the Palmae. Bot. Rev. 52: 221-259.
- - & M. Bal iek. 1987. Notes on the palms of the Amazon ia Legal. Principes 31 : 116-122.
- - , G. G a l e a n o & R. Berna l . 1995. Field guide to the palms of the Americas. Princeton Universi ty Press, Princeton.
- - & J. S t e y e r m a r k . 1986. New palms from Ve- nezuela. Brittonia 38: 309-313.
John son , D., editor. 1996. Palms, their conservat ion and sustained utilization: status survey and conser- vation action plan. IUCN/SSC Palm Specialist Group, Gland, Switzerland.
Klotz, L. 1978. The number of wide vessels in petiolar vascular bundles of palms: an anatomical feature of systematic significance. Principes 22: 64 -69 .
Knudsen , J. 1999. Floral scent chemis t ry in geono- moid pa lms (Palmae: Geonomeae) and its impor- tance in maintaining reproductive isolation. Pages 141-168. In: A. Henderson & E Borchsenius , ed- itors. Evolution, variation, and classification of palms. Mere. New York Bot. Gard., vol. 83. Bronx, New York.
Lewis, C. E. & J. J . Doyle�9 2001. Phylogenet ic utility of the nuclear gene malate synthase in the palm family (Arecaceae). Molec. Phyl. Evol. 19: 4 0 9 - 420.
Maddison , W. P. & D. R. M a d d l s o n . 2000. Mac- Clade. Version 4.0. Sinauer Associates , Sunder- land, Massachuset ts .
Moore , H. E. 1966. Aristeyera, a new genus of geon- omoid palms. J. Arnold Arbor. 47: 1-8.
. 1967. Two new American palms. J. Arnold Arb. 48: 143-151.
- - . . 1978. Wha t ' s in a name. Principes 22: 112- 113.
Nehl in , S. 1994. A gem for tissue culture: Asterogyne spicata of Venezuela. Principes 38: 211-213.
Prance , G. T. 1982. Forest refuges: evidence from woody angiosperms. Pages 137-158. hi: G. T. Prance. editor. Biological diversification in the tropics. Columbia University Press, New York.
Punt , W. & J. G. Wesse ls Boer. 1966. A palynolog- ical s tudy in geonomoid palms. Acta Bot. Neerl. 15: 266-275.
S e h m i d , R. 1970a. Notes on the reproduct ive biology of Asterogyne martiana (Palmae). I. Inflorescence and floral morphology; phenology. Pr incipes 14: 3-9.
- - . 1970b. Notes on the reproductive biology of Asterogyne martiana (Palmae). II. Pollination by syrphid flies. Principes 14: 39-49 .
- - . 1983. Septal nectaries of Asterogyne martiana and other Palmae. Principes 27: 168-174.
Seuber t , E. 1998. Root anatomy of palms. IV. Are- coideae, part 1, general remarks and descriptions of the roots. Fedde Rep. 109: 89-127.
S tauf fe r , F. W. & R. D u n o de Stefano. 1998. Notes on the conservat ion status of Asterogyne yaracu- yense in Venezuela. Principes 42: 57-58.
- - , R. R u t i s h a u s e r & P. K. E n d r e s s . 2002. Mor- phology and development of the female flowers in Geonoma interrupta (Arecaceae). Amer. J. Bot. 89: 220-229 .
S t e y e r m a r k , J . 1979. Plant refuge and dispersal cen- ters in Venezuela: their relict and endemic element. Pages 185-221. In: K. Larsen & L. B. Holm-Niel- sen, editors. Tropical botany. Academic Press, New York.
Swofford , D. L. 1998. PAUP*. Phylogenetic analysis us ing pars imony (and other methods). Version 4. Sinauer Associates, Sunderland, Massachuset ts .
T o m l i n s o n , P. B. 1961. Palmae. Ana tomy of the monocotyledons . Clarendon Press, Oxford.
�9 1966. Notes on the vegetative ana tomy of Ar- isteyera spicata (Palmae). J. Arnold Arbor. 47: 23 - 29.
Uhl , N. 1966. Morphology and ana tomy of the inflo- rescence axis and flowers of a new palm, Aristeyera ~picata. J. Arnold Arbor. 47: 9 -22 .
- - & J. Dransf ie ld . 1987. Genera Palmarum: a classification o f palms based on the work of H. E. Moore, Jr., L. H. Bailey Hortorium, and the Inter- national Palm Society. Allen Press, Lawrence, Kan- sas.
, - - , J. I. Davis , M. A. L u c k o w , K. S. H a n s e n & J. J . Doyle. 1995. Phylogenet ic rela- t ionships among palms: cladistic analyses of mor- phological and chloroplast D N A restriction site variation. Pages 623-661. In: R J. Rudall, R J. Cribb, D. E Cutler & C. J. Humphries , editors. Monocotyledons: systematics and evolution. Royal Botanic Gardens, Kew.
- - & H. E. Moore . 1977. Correlations of inflo- rescence, flower structure and floral ana tomy with pollination in some palms. Biotropica 9: 170-190.
W e n d l a n d , H. 1856. Einige neue Palmen Amerikas. Linnaea 28: 333-352.
Wessels Boer , J. G. 1968. The geonomoid palms. Verh. Kon. Ned. Akad. Wetensch, Afd. Natuurk., Tweede Sect. ser. 2, 58: 1-202.
- - . 1988. Palmas indfgenas de Venezuela. Pittier- ia 17: 1-332.
N u m e r i c a l L i s t o f S p e c i e s
1. As t e rogyne guianensis G r a n v . & A. J. 3. A. ramosa ( H . E . M o o r e ) W e s s . B o e r
H e n & 4. A. spicata ( H . E . M o o r e ) W e s s . B o e r
2. A. mar t iana ( H . W e n d l . ) H . W e n d l . e x 5. A. yaracuyense A . J. H e n d . & S t e y -
H e m s l . e r m .

2003] STAUFFER ET AL.: ASTEROGYNE (ARECACEAE) 355
Exsiccatae
Only the first collector has been mentioned.
Agostini, G. l0 (4) Aguilar, R. 2140, 2318 (2) Alverson, W. 94 (2) Anderson, R. 126 (2) Archer, W. 1978 (2) Aristeguieta, L. 1772, 3185 (4) Aulestia, C. 350, 455, 508, 1306 (2) Baker, R. 89021 (2) Barfod, A. 13 (2) Beach, J. H, 16545 (2) Bernal, R. 256, 259, 887, 1327, 1380 (2) Bernardi, A. 5633 (4) Blanc, M. 189 (1) Bonifaz, C. 3758 (2) Borchsenius, E 234, 242 (2) Bunting, G. 401 (2) Burger, W. 5599, 8076, 10616, 11173 (2) Callejas, R. 2759, 4220, 8528, 9202 (2) Chac6n, A. 973 (2) Chac6n, E. 1958 (2) Chac6n, I. 1311, 1615 (2) Chazdon, R. 16 (2) Churchill, H. 4122, 4186 (2) Cogollo, A. 4290 (2) Cook, O. E 61, 84, s.n. (2) Correa, M. 624 (2) Croat, T. 9933, 13202, 13884, 16834,
21997, 24524, 26052, 26161, 27201, 33683, 35353, 35740, 36459, 36461, 59889, 67689, 68156, 68405 (2)
Cuatrecasas, J. 17375, 21300, 24195 (2) Danin, A. 77-5-2, 77-14-12 (2) Davidse, G. 23727, 31109, 31267 (2) de Granville, J. J. 7124, 7124-A (1) de Nevers, G. 3700, 3795, 3974, 4460,
5209, 5556, 5881, 5896, 5941, 5942, 6111, 6167, 6282, 6323, 7914, 8320 (2)
Denslow, J. 2745 (2) Dodge, C. 9284 (2) Dransfield, J. 4859 (2) Duke, J. 11334, 11336 (2) Elias, T. 1769 (2) Folsom, J. 5901, 6179 (2) Fonnegra, R. 2839, 3024, 4756 (2) Forero, E. 8975, 9039 (2) Fuchs, H. R 21808, 21994 (2) Gentle, R 3743, 8635, 9072, 9074, 9669 (2) Gentry, A. 2803, 7542, 8818, 8868, 17445,
40972, 41092, 48546, 56871, 63735 (2); 24823 (4).
G6mez, L. D. 19506, 19655, 20341 (2) Gonzfilez, A. 1296 (4) Gordon, B. 70 (2) Grayum, M. 4052 (2) Grijalva, A. 262, 380, 3481 (2) Gudifio, E. 1283 (2) Haber, W. 11580 (2) Hamilton, C. 1087, 2927, 2945 (2) Hammel, B. 1682, 1797, 4863, 5153,
12377(2), 12648 (2) Hari, R s.n. (2)] Henderson, A. 53, 714, 3048 (2); 502 (5) Herrera, H. 1188, 1332 (2) Holdridge, L. 5122 (2) Holm, R. 914, 970 (2) Hoover, W. 1241 (2) Janzen, D. 11098 (2) Juncosa, A. 633, 732, 1311, 1604, 1720,
2532 (2) Killip, E. 35293 (2) Knapp, S. 1417, 1772, 3777 (2) Knudsen, J. 604, 620, 623 (2) Koch, R. 5048 (2) Kress, W. J. 3778 (2) Lehmann, E C. 708 (2) Lems, K. 5199 (2) Lent, R. 80 (2) Levy, P. 505 (2) Lewis, W. 5558 (2) Liesner, R. 2924, 3223, 14825 (2) Maas, R J. M. 2803 (2) McDonagh, J. 246, 322 (2) McDowell, T. 886 (2) McPherson, G. 9716, 8080 (2) Meier, W. 3386 (4) Miller, J. 1095 (2) Molina, A. 17559 (2) Mondrag6n, M. 119 (2) Monsalve, M. 671, 927 (2) Moore, H. E. 6530, 6535, 6717, 8213, 9420
(2); 9601 (4) Morales, J. 1118 (2) Moreno, R 12280, 12580, 12771, 12979,
12985, 13032, 13151, 13287, 14821, 15013, 19142, 20602, 20640, 20661, 23621, 23890, 24015, 26083 (2)

356 BRITTONIA [VOL. 55
Mori, S. 2312, 2319, 4095, 4589, 4905, 6383 (2)
Navarro, E. 76 (2) Nee, M. 17789 (4), 27905 (2) Neill, D. 1723, 4442, 7151 (2) Oersted, A. 6566 (2) Ortiz, E 297, 733 (2) Picado, A. 123 (2) Pipoly, J. 5111, 5323, 6112, 6143, 6168,
6223, 6290 (2) Pittier, H. 4226, 16658, 16717, 16922 (2) Porter, D. 4736 (2) Quesada, E 43 (2) Ramirez, J. 46, 167, 1131, 1364, 1770,
1860 (2) Ram/rez, V. 183 (2) Read, R. 640 (2) Rfos, D. 187 (2) Robles, R. 1148, 1192 (2) Rowlee, W. 1010 (2) Rueda, R. 2670, 3110, 3459 (2) Salick, J. 8075, 8106 (2) Sandino, J. 4782 (2) Schipp, W. 392 (2) Schmalzel, R. 1156 (2) Seemann, B. 163, s.n (2) Seymour. E 5905 (2) Shepherd, J. 863 (2) Soza, D. 347 (2) Sperry, J. 606 (2) Standley, R 25015, 53156 (2)
Stauffer, E W. 824 (3); 822 (4): 265, 267, 319, 823 (5)
Stevens, W. D. 4810, 4943, 6336, 6795, 6820, 8322, 12080, 12246~ 12686, 13065, 13505, 14927, 19343, 20743, 20936, 23489, 23651, 24026, 24331, 24451 (2)
Stevenson, D. 1204 (2) Steyermark, J. 94803, 96088, 120915,
121713 (3); 39785, 90010, 97573, 104041, 116530, 116565 (4); 97661, 100307, 100311 (5)
Sudgen, A. 351 (2) Sullivan, G. 159 (2) Sytsma, K. 1660 (2) Tamayo, F. 4177 (4) Tessene, M. 1329 (2) Thomsen, K. 187 (2) Tipaz, G, 2250~ 2693 (2) Tomlinsom R B. s.n. (2) Utley, K. 1093, 5975 (2) Valverde, O. 665 (2) van Rooden, J. 95 (4) Vincelli, R 164, 228 (2) von Wedel, H. 952, 1108, 2186 (2) Watson, S. 238 (2) Wendland, H. 56, s.n. (2) Wessels Boer~ J. G. 2032, 2036 (5) Wilbur, R. L. 15071 (2) Wilson, R 224 (2) Witherspoon, J. 8322 (2) Yuncker, T. 5002 (2) Zarucchi, J. 5056 (2)
Copyright © 2022 FDOKUMEN




















